
Movement of the multicellular slug stage of Dictyostelium discoideum: an analytical approach

Cellular and functional specificity among ferritin-likeproteins in the multicellular cyanobacteriumNostoc punctiforme
Martin Ekman,1† Gustaf Sandh,1 Anja Nenninger,1
Paulo Oliveira1,2 and Karin Stensjö1*1Department of Chemistry – Ångström Laboratory,Uppsala University, SE-751 20 Uppsala, Sweden.2Bioengineering and Synthetic Microbiology Group,IBMC – Instituto de Biologia Molecular e Celular, R. doCampo Alegre, 823, 4150-180 Porto, Portugal.
Summary
Ferritin-like proteins constitute a remarkablyheterogeneous protein family, including ferritins,bacterioferritins and Dps proteins. The genome ofthe filamentous heterocyst-forming cyanobacteriumNostoc punctiforme encodes five ferritin-like pro-teins. In the present paper, we report a multi-dimensional characterization of these proteins. Ourphylogenetic and bioinformatics analyses sug-gest both structural and physiological differencesamong the ferritin-like proteins. The expression ofthese five genes responded differently to hydrogenperoxide treatment, with a significantly higher risein transcript level for Npun_F3730 as compared withthe other four genes. A specific role for Npun_F3730in the cells tolerance against hydrogen peroxidewas also supported by the inactivation of Npun_F3730, Npun_R5701 and Npun_R6212; among these,only the ΔNpun_F3730 strain showed an increasedsensitivity to hydrogen peroxide compared withwild type. Analysis of promoter-GFP reporter fusionsof the ferritin-like genes indicated that Npun_F3730and Npun_R5701 were expressed in all cell typesof a diazotrophic culture, while Npun_F6212 wasexpressed specifically in heterocysts. Our study pro-vides the first comprehensive analysis combiningfunctional differentiation and cellular specificity
within this important group of proteins in amulticellular cyanobacterium.
Introduction
To stay alive and to be competitive, all organisms needto adapt to changes in their surrounding environment,such as changes in photon flux and temperature overdiurnal or seasonal cycles and/or changes in nutrientavailability. These changes will alter the cellular redoxmilieu and will in oxygenic environments lead to anincreased production of reactive oxygen species (ROS).In cyanobacteria, the responses to rising levels of ROSand iron metabolism are closely interconnected (Latifiet al., 2005) with multiple systems involved in thedefence against ROS and control of iron homeostasis.Parts of these systems are enzymes such as the ferritin-like proteins. In cyanobacteria this protein familyincludes both Dps (DNA-binding proteins from starvedcells), bacterioferritins (Bfr) and in a few cases alsoferritins (Ftn) (Andrews, 2010). Common for ferritin-likeproteins is their ability to catalyse the oxidation of Fe2+
to Fe3+ via ferroxidase centres (FOCs). The oxidizedferrous iron may subsequently be stored in the form of aferric oxo product, assigning this group of proteins acentral role in iron storage and homeostasis (Andrews,2010). While Ftns and Bfrs primarily use O2 to oxidizethe ferrous iron (Bou-Abdallah, 2010), the FOCs of Dpsproteins have a preference for H2O2 (over 100-foldhigher rate; Zhao et al., 2002; Franceschini et al., 2006;Chiancone and Ceci, 2010). This characteristic givesDps proteins a specific function related to oxidativestress protection, e.g. by catching and sequestering freeFe2+ while at the same time reducing H2O2 to water andthereby preventing the Fenton reaction to occur. Dpsproteins may bind to DNA (Almiron et al., 1992), indicat-ing a function of these proteins in specifically protectingDNA from oxidative damage. They are also known toaccumulate under nutrient starvation (Almiron et al.,1992; Azam and Ishihama, 1999) and under suchconditions provide resistance towards H2O2 and oxida-tive stress (Martinez and Kolter, 1997). While Ftnsare found in all kingdoms of life, Bfrs are restricted to
Received 14 May, 2013; accepted 28 July, 2013. *For correspond-ence. E-mail [email protected]; Tel. (+46) 0 18471 6586;Fax (+46) 0 18471 6844. †Present address: Department of Ecology,Environment and Plant Sciences, Stockholm University, SE-106 91Stockholm, Sweden.
bs_bs_banner
Environmental Microbiology (2014) 16(3), 829–844 doi:10.1111/1462-2920.12233
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd

bacteria and archaea, and Dps proteins are only foundin bacteria, with some archaea containing non-typicalBfrs that have some characteristics of the Dps proteins.Bacterial genomes typically encode for one Dps protein,usually in combination with a Bfr and/or Ftns. A limitednumber of bacteria have multiple Dps, e.g. Bacillusanthracis, Deinococcus radiodurans and Mycobacteriumsmegmatis (Schwartz et al., 2010; Ghatak et al., 2011;Reon et al., 2012). On the basis of sequenced bacterialgenomes (see, i.e. http://img.jgi.doe.gov/cgi-bin/w/main.cgi), cyanobacteria are clearly overrepresented amongbacteria with multiple Dps proteins.
Cyanobacteria perform oxygenic photosynthesis andexist in a wide range of morphotypes from unicellular tocolony forming and filamentous species (Rippka et al.,1979). Certain filamentous strains undergo cell differen-tiation under conditions of nitrogen limitation, andheterocysts, cells specialized for fixing atmospheric nitro-gen (N2) via the enzyme nitrogenase, develop at regularintervals along the filaments (Kumar et al., 2010). Thereare several published studies on ferritin-like proteins incyanobacteria (see, e.g. Peña & Bullerjahn, 1995;Hernández et al., 2007; Wei et al., 2007; Shcolnick et al.,2009; Narayan et al., 2010; Sato et al., 2012). However,none of these have addressed functional differences orcell specificity for multiple Dps proteins when present inthe same organism.
Nostoc punctiforme belongs to the multicellularheterocyst-forming category of cyanobacteria and wasoriginally isolated from symbiosis with a cycad (Rippkaet al., 1979). In addition to heterocyst differentiation,they are also capable of differentiating spore-likeakinetes or motile hormogonia during conditions of star-vation (Meeks et al., 2002). Given the complex responseof N. punctifome to stressful conditions, we hypothesizethat the five ferritin-like proteins encoded in the genomeof this bacterium may have different functions,which may in part be related to cell-specific processes.Indeed, previous shotgun proteomics data, whereprotein abundances were compared between isolatedheterocysts and N2-fixing filaments, show a differentialabundance of several of the ferritin-like proteins inheterocysts compared with entire filaments (Ow et al.,2009; Fig. S1).
In this study, we present the phylogenetic relationshipbetween the five annotated ferritin-like proteins inN. punctiforme and compare their sequences in relationto conserved amino acid residues with known functionalproperties. We further analyse the cellular specificity andfunctional differences of several of the ferritin-like pro-teins in N. punctiforme by characterizing gene deletionstrains under different environmental conditions and byfollowing the transcriptional regulation of the ferritin-likegenes by reverse transcription quantitative polymerase
chain reaction (RT-qPCR) and in promoter-GFP-reporterstrains.
Results
Phylogenetic analysis
Phylogeny. Given the high prevalence of multicopyferritin-like proteins within the genomes of cyanobacteria,we wanted to assess how these different proteins areevolutionarily related. We therefore constructed aphylogenetic tree of protein sequences belonging to theferritin-like protein family (Pfam00210) from all availablesequenced cyanobacterial genomes [downloaded fromhttp://img.jgi.doe.gov/cgi-bin/w/main.cgi in March 2013,thus also including sequences from the new CyanoGEBAphylogenetic expansion of available cyanobacterialgenomes (Shih et al., 2013)]. In addition, a number ofreference sequences from other organisms (bacteria,archaea and eukaryotes) were included (supplementalFASTA file). The overall clustering is in line with previousfindings (Andrews, 2010), e.g. that Ftn, Bfr and Dps pro-teins form separate clusters. Four of the ferritin-like pro-teins of N. punctiforme (Npun_R3258, Npun_R5799,Npun_R5701 and Npun_F3730) cluster within the Dpsclade while the fifth (Npun_F6212) clusters in the Bfrclade, indicating that N. punctiforme has four Dps proteinsand one Bfr.
Within the Dps cluster, four distinct cyanobacterial-specific subclusters can be distinguished, each ofwhich contain one of the N. punctiforme Dps proteins.These four cyanobacterial Dps clusters do not form amonophyletic group, as Dps sequences from other bac-teria are found interspersed among the cyanobacterialsubclusters. This indicates evolutionary and possiblyfunctional differences among these paralogous genes inN. punctiforme. One of the Dps subclusters, containingNpun_R5799, forms an out-group to the other three Dpsclusters. The nearest relatives to these sequences arefound in a cluster of archaeal sequences. When lookingmore closely into the cyanobacterial strains found withineach Dps subcluster, it is evident that the Npun_R3258,Npun_R5799 and Npun_R5701 subclusters are domi-nated by N2-fixing strains while the Npun_F3730 clustercontains also many Dps proteins from non-N2-fixingcyanobacteria (Fig. 1; Fig. S2).
The Bfr cluster containing Npun_F6212 is cyano-bacteria specific. Furthermore, the cyanobacterialstrains within this cluster are mostly filamentous and N2
fixing. Certain cyanobacterial Bfrs, primarily sequencesfrom unicellular Synechocystis and Synechococcusstrains, cluster with bacterial (non-cyanobacterial)Bfr sequences. A few cyanobacterial Bfr sequences(Gloeocapsa sp. PCC 7428, Fischerella sp. PCC 9605,Gloeobacter violaceus PCC 7421, Pleurocapsa sp. PCC
830 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

7327 and Chroococcidiopsis thermalis PCC 7203) clusterwith archaeal sequences.
Nostoc punctiforme does not contain any ferritin-likeprotein that clusters with the Ftn subclade. However, acyanobacteria-specific subcluster is found within the Ftncluster, with sequences belonging to species from theecologically important marine genera Synechococcusand Prochlorococcus. The observation that Ftns incyanobacteria are often found in Prochlorococcus andSynechococcus could indicate that these proteins play animportant role in the environmental success of thesegenera in their natural iron-limited habitat, in the oceans(see, e.g. Partensky and Garczarek, 2010).
When further investigating the distribution of ferritin-likegenes in all the sequenced genomes of cyanobacteria(Table S1), it appears that as mentioned above, Ftns areabsent in most cyanobacterial species. One or two Bfrscan be found in most cyanobacterial genomes with themajor exceptions being all Prochlorococcus species and amajority of all Synechococcus species. The most wide-spread group is the Dps proteins that is represented inalmost all genomes with the exception of Cylindro-spermopsis raciborskii, the reduced genome of the newlydiscovered non-oxygenic phototroph CyanobacteriumUCYN-A (Zehr et al., 2008), Arthrospira sp. PCC8005,Leptolyngbya valderiana BDU 20041, Prochlorothrixhollandica PCC 9006, Synechococcus sp. BL107 andCC9616 and the majority of all Prochlorococcusspecies. Overall, many cyanobacterial genomes containmultiple Dps-encoding genes, but their presence is notlinked to a monophyletic subgroup within cyanobacteriaas there are representatives of unicellular, filamentousnon-heterocyst-forming and filamentous heterocyst-
forming species lacking Dps-encoding genes in theirgenomes.
Sequence analysis and structure predictions. To furtherexplore the findings from the phylogenetic analysis, weinvestigated the presence of conserved sequence fea-tures of the different ferritin-like proteins of N. punctiformethat were cluster/subcluster specific. Sequence alignmentof N. punctiforme ferritin-like proteins together with repre-sentatives of different ferritin-like proteins from otherbacteria reveals strong predicted structural homologybetween the proteins Npun_F3730, Npun_R3258,Npun_R5701 and the Dps family prototype of Escherichiacoli and DpsA of Synechocysis sp. PCC 6803 (Fig. 2).Their tertiary structure as predicted by Phyre 2 (Kelleyand Sternberg, 2009; http://www.sbg.bio.ic.ac.uk/phyre2/html/) showed the classical Dps protein fold with the four-helix bundle including an additional smaller helix (helixBC) in between the major helices B and C. These Dpsproteins contain highly conserved amino acid residuesthat coordinate or are associated to the FOCs, whichare located in-between two subunits. The conservedamino acid ligands forming each di-iron centre (followingthe numbering in E. coli) are His51, His63, Asp78 andGlu82. Theses amino acids together with the conservedtryptophan (Trp52) associated to the FOCs form the Dpsspecific motif HW (helixA) and H-14-DxxxE (helix B)(Fig. 2). None of the three putative Dps proteins(Npun_F3730, Npun_R3258 and Npun_R5701) inN. punctiforme has a long lysine-rich N-terminus that hasbeen shown to be important for the DNA binding andDNA condensation ability of E. coli Dps (Ceci et al.,2004). However, both Npun_R3258 and Npun_R5701
Fig. 1. Phylogenetic approximatelymaximum-likelihood tree of ferritin-likeproteins family consisting of three maingroups: ferritins (Ftn), bacterioferritins (Bfr)and Dps proteins (Dps). Sequences usedinclude all the ones from genome-sequencedcyanobacteria and annotated to theferritin-like protein family (Pfam00210), andalso a selection of Ftn family sequences fromeukaryotes, bacteria and archaea. Nostocpunctiforme sequences are found in fivedifferent cyanobacteria-dominated clusters(dashed ovals) and indicated by theN. punctiforme gene locus tag. All sequencesused for constructing the tree can be found assupplemental data (supplemental FASTA file).
The role of Dps proteins in Nostoc punctiforme 831
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

possess long C-terminal extensions and Npun_R5701’sC-terminal extension contains several positively chargedlysines.
The fourth protein predicted from the phylogeneticstudy to be a Dps protein, Npun_R5799, showed a pre-dicted structure that did not resemble the typical Dps fold.Npun_R5799 showed the highest structural similarity tothe atypical DpsA from Thermosynechococcus elongatus(DpsA-Te; Alaleona et al., 2010) with the Dps canonicalaspartate (Asp78 in E. coli) replaced with a histidine(His78), the Trp52 replaced with a phenylalanine (Phe52)and the Phe161 replaced with a histidine (His164; Fig. 2).The predicted structure indicates that as in T. elongatus,the FOC may be formed by the interplay of three adjacentsubunits; His51 and His63 from one subunit, His78 andGlu82 from a second and the third subunit providesHis164. Another similarity to DpsA-Te is the longN-terminal extension, which lacks the DNA-binding fea-tures of the E. coli N-terminus. Npun_R5799 differs from
DpsA-Te by the fact that the unusual feature of DpsA-Te,with five cysteines within the sequence, is not shared byNpun_R5799, in which only one of the five cysteines ofDpsA-Te is conserved.
The fifth N. punctiforme’s ferritin-like protein,Npun_F6212, clusters with the Bfr clade (Fig. 1) and alsoshows a predicted structure with similarities to Bfrs suchas the lack of the small helix (BC) between helices B andC in the Dps 4-helix bundle (data not shown, for con-served FOC ligands see Fig. 2). The amino acids, fourglutamates and two histidines, coordinating the FOCwithin the four-helix bundle of Bfrs are all present inNpun_F6212. However, the extended Bfr-specific FOCligand binding motif, E-5-QY (helixA) EMxH (helixB) E-6-Y(helix C) and ExxH (helix D) (Andrews, 2010), is lessconserved in Npun_F6212, giving the motif E-6-Y in helixA and the motif ExxxH in helix B. Furthermore, theextended C-terminus forming the E-helix of typical Bfrs ispredicted to be absent in Npun_F6212.
SYNE- ANp3730Np3258Np5701EC- DPSTHEE- ANp5799THEE- BSYNE- BEC- BFRNp6212EC- FRT
Helix A
Helix B Helix C
Helix D
Helix BC
* *
* *
*
SYNE- ANp3730Np3258Np5701EC- DPSTHEE- ANp5799THEE- BSYNE- BEC- BFRNp6212EC- FRT
SYNE- ANp3730Np3258Np5701EC- DPSTHEE- ANp5799THEE- BSYNE- BEC- BFRNp6212EC- FRT
Fig. 2. Alignment of the five ferritin-like proteins encoded in the genome of Nostoc punctiforme and selected reference sequences [DpsA(SYNE-A) and Synpcc7942_0109 (SYNE-B) from Synechococcus elongatus PCC 7942, Dps-Te (THEE-A) and DpsA-Te (THEE-B) fromThermosynechococcus elongatus, and the Dps protein (EC-DPS), bacterioferritin (EC-BFR) and ferritin (EC-FRT) from E. coli]. Positivelycharged amino acids near the C- and N-termini of the sequences are indicated by filled grey boxes. Helices are indicated by solid lines abovethe sequences and are named according to Alaleona and colleagues (2010). Ferroxidase-binding amino acids are light grey shaded and alsoindicated by an asterisk. Conserved sequence differences between Dps of the Npun_R5799 type (Np5799; also THEE-B and SYNE-B) and allother Dps are boxed. An approximately maximum-likelihood phylogenetic tree indicates the sequence similarity of the different sequences.
832 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844
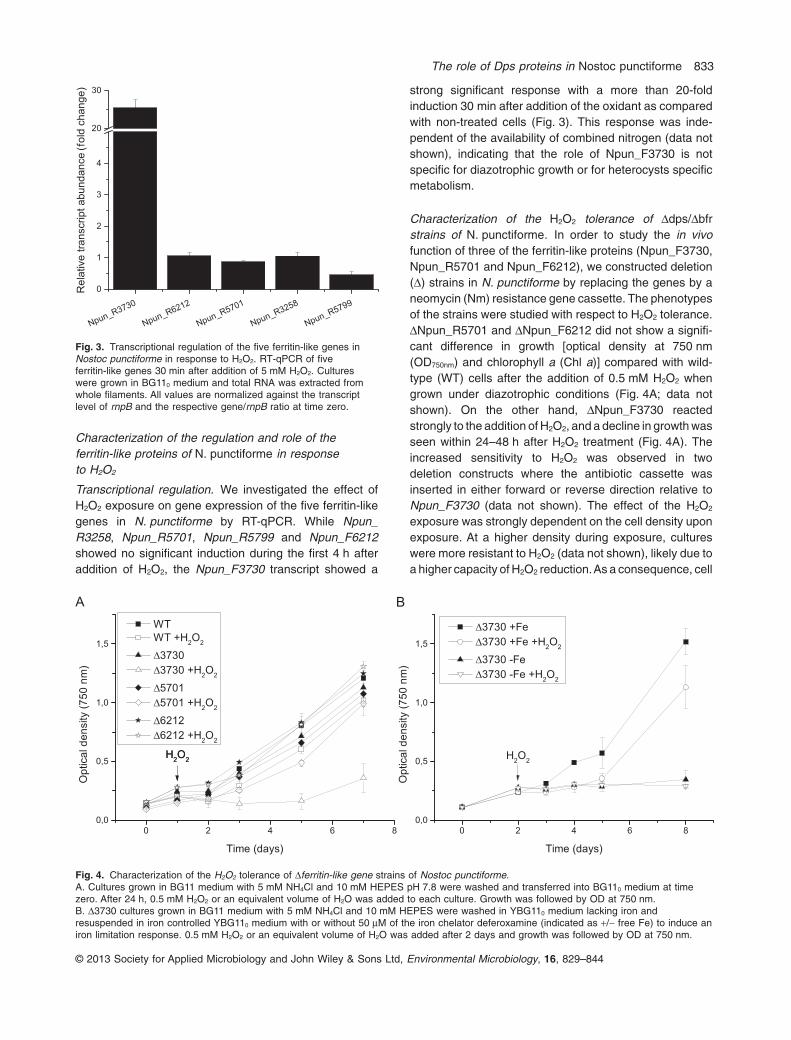
Characterization of the regulation and role of theferritin-like proteins of N. punctiforme in responseto H2O2
Transcriptional regulation. We investigated the effect ofH2O2 exposure on gene expression of the five ferritin-likegenes in N. punctiforme by RT-qPCR. While Npun_R3258, Npun_R5701, Npun_R5799 and Npun_F6212showed no significant induction during the first 4 h afteraddition of H2O2, the Npun_F3730 transcript showed a
strong significant response with a more than 20-foldinduction 30 min after addition of the oxidant as comparedwith non-treated cells (Fig. 3). This response was inde-pendent of the availability of combined nitrogen (data notshown), indicating that the role of Npun_F3730 is notspecific for diazotrophic growth or for heterocysts specificmetabolism.
Characterization of the H2O2 tolerance of Δdps/Δbfrstrains of N. punctiforme. In order to study the in vivofunction of three of the ferritin-like proteins (Npun_F3730,Npun_R5701 and Npun_F6212), we constructed deletion(Δ) strains in N. punctiforme by replacing the genes by aneomycin (Nm) resistance gene cassette. The phenotypesof the strains were studied with respect to H2O2 tolerance.ΔNpun_R5701 and ΔNpun_F6212 did not show a signifi-cant difference in growth [optical density at 750 nm(OD750nm) and chlorophyll a (Chl a)] compared with wild-type (WT) cells after the addition of 0.5 mM H2O2 whengrown under diazotrophic conditions (Fig. 4A; data notshown). On the other hand, ΔNpun_F3730 reactedstrongly to the addition of H2O2, and a decline in growth wasseen within 24–48 h after H2O2 treatment (Fig. 4A). Theincreased sensitivity to H2O2 was observed in twodeletion constructs where the antibiotic cassette wasinserted in either forward or reverse direction relative toNpun_F3730 (data not shown). The effect of the H2O2
exposure was strongly dependent on the cell density uponexposure. At a higher density during exposure, cultureswere more resistant to H2O2 (data not shown), likely due toa higher capacity of H2O2 reduction.As a consequence, cell
f
Fig. 3. Transcriptional regulation of the five ferritin-like genes inNostoc punctiforme in response to H2O2. RT-qPCR of fiveferritin-like genes 30 min after addition of 5 mM H2O2. Cultureswere grown in BG110 medium and total RNA was extracted fromwhole filaments. All values are normalized against the transcriptlevel of rnpB and the respective gene/rnpB ratio at time zero.
Fig. 4. Characterization of the H2O2 tolerance of Δferritin-like gene strains of Nostoc punctiforme.A. Cultures grown in BG11 medium with 5 mM NH4Cl and 10 mM HEPES pH 7.8 were washed and transferred into BG110 medium at timezero. After 24 h, 0.5 mM H2O2 or an equivalent volume of H2O was added to each culture. Growth was followed by OD at 750 nm.B. Δ3730 cultures grown in BG11 medium with 5 mM NH4Cl and 10 mM HEPES were washed in YBG110 medium lacking iron andresuspended in iron controlled YBG110 medium with or without 50 μM of the iron chelator deferoxamine (indicated as +/− free Fe) to induce aniron limitation response. 0.5 mM H2O2 or an equivalent volume of H2O was added after 2 days and growth was followed by OD at 750 nm.
The role of Dps proteins in Nostoc punctiforme 833
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

density for all experiments was diluted to a fixed startingOD (OD750 = 0.135). It was also clear that the effect of thissingle addition of H2O2 was transient; within 24 h theextracellular levels of H2O2 in the cultures were down to0 μM (data not shown), and within 6 days, H2O2-treatedcells recovered their growth. A second addition of 0.5 mMH2O2 at the start of the recovery phase (day 4) inducedanother negative growth response but with a shorter recov-ery phase than in the initial treatment. This responseindicates an induction of a general antioxidative defencesystem triggered upon the first H2O2 treatment, whichconfers a greater fitness level against a second H2O2
exposure (data not shown).The higher sensitivity of ΔNpun_F3730 to H2O2 was
observed in both ammonium-supplemented cultures aswell as in combined nitrogen-free cultures. As Npun_R5701 and Npun_F6212 have been shown to be moreabundant under N2-fixing conditions (Ow et al., 2009;Fig. S1), and Npun_F6212 is mainly expressed inheterocysts (see Fig. 5), we also expected these twodeletion strains to show reduced growth under N2-fixingconditions when stressed with H2O2. However, all culturesgrown without combined nitrogen were overall moreresistant to H2O2 (data not shown).
The H2O2 tolerance of Δdps/Δbfr strains under iron depri-vation. Ferritin-like proteins are not only involved inDNA protection, but also in peroxide and oxygen detoxi-fication and iron storage (Andrews, 2010). To test if theperoxide sensitivity observed in ΔNpun_F3730 was dueto Fenton chemistry, which is dependent on availableferrous iron, we analysed how the deletion strainsreacted to H2O2 treatment during iron limitation. Aclear reduction in Chl a levels was detected in all testedstrains approximately 48 h after addition of thesiderophore deferoxamine (data not shown). Iron limita-tion also inhibited growth monitored as OD750nm. In con-trast to the results in iron replete media, no additionalnegative effect on growth was observed when exposingthe iron deplete culture of ΔNpun_F3730 to 0.5 mMH2O2 (Fig. 4B). The slow growth of the iron-limited cul-tures could however mask the response. However, apositive increase in Chl a concentration at day 3 (theday after H2O2 addition) was observed for both H2O2-treated and non-treated WT and ΔNpun_F3730 in theiron deplete cultures (data not shown). No differenceto WT could as before be detected for ΔNpun_R5701and ΔNpun_F6212 (data not shown). Although severalputative iron storage proteins of N. punctiformewere knocked-out (ΔNpun_F3730, ΔNpun_R5701 andΔNpun_F6212), no further negative phenotypic effectson growth or pigmentation was apparent in the deletionstrains compared with WT under the tested conditions(Fig. 4A and B; data not shown).
Cellular localization of Dps gene expression
To visualize the cell type-specific gene expression offerritin-like proteins, promoter-gfp fusion strains weremade (Np6212-GFP, Np5701-GFP and Np3730-GFP).While Np6212-GFP and Np5701-GFP were prepared byrecombination of the promoter-gfp fusion into the genome,in Np3730-GFP the promoter-gfp construct was expressedfrom a replicative plasmid. Cells were grown for 6 daysunder diazotrophic or combined nitrogen conditions, andthe GFP fluorescence was detected using confocal scan-ning microscopy. The genomic construct Np6212-GFP didnot show any GFP signal in filaments of ammonium-supplemented cultures (Fig. 5A), and under the samegrowth conditions Np5701-GFP showed only a weak signal(Fig. 5B). However, the construct Np3730-GFP (Fig. 5C)showed strong GFP fluorescence along the ammoniumgrown filaments. The GFP expression from a self-replicating vector in the strain Np3730-GFP was strongerand less homogeneous along the filaments and betweenfilaments than from the two strains with genomic insertions.This elevated GFP expression has been seen for otherreplicating vectors in cyanobacteria (Ekman et al., 2013).When cultures were grown diazotrophically, the expressionof GFP was specific to heterocysts of Np6212-GFP(Fig. 5D) with no fluorescence visible from the vegetativecells. The majority of the heterocysts of Np5701-GFP(Fig. 5E) also show a high GFP fluorescence. However, thefluorescence from the heterocysts of Np5701-GFP has alarger variation in intensity as compared with the homog-enous fluorescence from the vegetative cells along thefilaments, and no significant difference (P-value < 0.05)could be determined between the two cell types. Thevariation seen might be due to the different age of theheterocysts in the culture. Under the Npun_F3730 pro-moter, the GFP fluorescence levels were similar betweenvegetative cells and heterocysts of the N2-fixing filament(Fig. 5F). A significant increase of almost 50% (pairedt-test, P-value < 0.05) and a more evenly distributed GFPfluorescence was observed in vegetative cells underdiazotrophic growth for Np5701-GFP as compared with inammonium-grown filaments, whereas no significant differ-ence in GFP fluorescence level of Np3730-GFP was foundin the two different cultivation regimes.
Discussion
Iron homeostasis and cellular defense against ROS arecrucial features in phototrophic organisms (see, i.e. Latifiet al., 2009; Kranzler et al., 2013). In diazotrophiccyanobacteria, an increased iron demand of thenitrogenase enzyme complex, in combination with theenzyme’s oxygen sensitive nature, lead to a need foran even tighter regulated defence mechanism. We havehere presented a broad approach to study the function,
834 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

Fig. 5. Confocal micrographs of N. punctiforme filaments grown with (A–C) and without (D–F) combined nitrogen. The four panels show GFPfluorescence in green, autofluorescence in magenta, a merged and bright field image (bars = 5 μm). Panel lines A and D give GFP distributionunder the Npun_F6212 promoter, B and E under the Npun_R5701 promoter, and C and F under the Npun_F3730 promoter. The first twostrains carry the promoter-GFP construct as a genomic insert, the latter shows gfp expression from the self-replicating vector pSUN119.Heterocysts are marked with a star. Wild-type filaments of N. punctiforme did not give a signal in the GFP channel under the chosen imagingconditions.
The role of Dps proteins in Nostoc punctiforme 835
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

localization and diversity of members of the putative iron-storing and ROS-detoxifying ferritin-like protein family in amulticellular diazotrophic cyanobacterium. Our initialhypothesis that different members of this protein familymay have distinct roles within cells found along the fila-ments was based on:
(i) the presence of multiple paralogous ferritin-like pro-teins in the genome of N. punctiforme
(ii) their differential abundance in filaments of Nostocspp. cultivated with or without combined nitrogen
(iii) as well as their difference in abundance in differentcell-types of N. punctiforme under diazotrophicgrowth.
Indeed, previous studies on bacteria-encoding multipleDps proteins have reported differential transcriptionalregulation, physiological activities and cellular expressionpatterns of these proteins (Facey et al., 2009; Reon et al.,2012; Tu et al., 2012). Investigations into the ferritin-likeprotein family in cyanobacteria have also revealed regu-latory and biochemical differences, i.e. in connection toiron storage, DNA protection, and their affinity and effi-ciency in detoxification of H2O2 (Keren et al., 2004;Narayan et al., 2010) as well as disclosed novel biochemi-cal activities (Alaleona et al., 2010).
Our bioinformatic study of the distribution of ferritin-likeproteins in all sequenced cyanobacterial genomes(Table S1) revealed a large variation in how common spe-cific members of the ferritin-like protein family (Ftns, Bfrsand Dps proteins) are in different groups and speciesof cyanobacteria. While N. punctiforme like most cyano-bacteria lack Ftn-encoding genes, these seem to be espe-cially abundant in the marine representatives within thegenera Synechococcus and Prochlorococcus. Thesecyanobacteria are naturally found in oligotrophic oceansthat often experience Fe limitation, and the Ftn couldplay an important role in their Fe storage mechanism aswas suggested for Dpstery in Trichodesmium (Castruitaet al., 2006; 2007), another ecologically important marinecyanobacterial genus (Bergman et al., 2013). Theabsence of Dps proteins in most species of the genusProchlorococcus could also be part of the explanationwhy previous reports have shown them unable to detoxifyH2O2 on their own (Morris et al., 2011).
In addition we have also noted the common feature ofmultiple Dps proteins in cyanobacteria. Most genomes ofheterocyst-forming cyanobacteria encode multiple Dpsproteins (with just a few exceptions) and thus the questionremains if this is due to the close evolutionary relationshipor to a metabolic/ecological advantage in this particularheterocyst-forming group. The phylogenetic analysis ofthe cyanobacterial ferritin-like protein family members(Fig. 1), including the recently released CyanoGEBAdataset of newly sequenced cyanobacterial genomes
(Shih et al., 2013), showed that, like their closely relatedorthologues in Anabaena sp. PCC 7120 (Sato et al.,2012), all five N. punctiforme protein sequences cluster inseparate subclusters. Four (Npun_F3730, Npun_R3258,Npun_R5701 and Npun_R5799) cluster within the Dpsclade and one (Npun_F6212) within the Bfr clade.
The clustering of Npun_F6212 with the Bfr clade wasnot surprising given the presence of amino acid motivessimilar to the canonical motives of the E. coli Bfr (Fig. 2)and the lack of the Dps-specific small helix and the con-served motif (HW; H-14-DxxxE) present in most ofthe Dps proteins identified to date (Andrews, 2010). Asseen in Fig. 4A, Npun_F6212 is not playing a primary rolein H2O2 detoxification, neither in actively growingdiazotrophic cells nor in cells cultivated with combinednitrogen. There is, however, a clear heterocyst-specificGFP fluorescence from the Npun_F6212 promoter(Fig. 5), indicating that Npun_F6212 is primarilyexpressed in heterocyst. Bfrs are mainly involved in ironstorage, as they store up to 10 times more iron as com-pared with Dps and uses O2 to oxidize the ferrous iron.Our results suggest that Npun_F6212 could be involved insupporting the high Fe demand and the regulation of theO2 level in the heterocyst. Npun_F6212 is in the genomeof N. punctiforme located directly upstream of a geneannotated as an iron permease (Fig. S3), suggesting alsoa putative coordination between the Bfr and iron uptake.In the phylogenetic analysis Npun_F6212 did not clustertogether with the E. coli Bfr (Fig. 1), and some of theconserved features of the typical Bfrs are missing. We arecurrently investigating its structure, Fe-binding capacityand O2/H2O2 selectivity because both Fe sequestrationand O2 removal are crucial for maintaining the main func-tion of the heterocyst, N2 fixation and adenosinetriphosphate production. A recently published quantitativeproteomics investigation (Ekman et al., 2011) showeda significantly higher abundance of Npun_F6212 inthe heterocysts of a modified H2-producing strain ofN. punctiforme (NHM5) as compared with WT. Theoxygen tension within the heterocysts of the H2-producingstrain was suggested to be increased because of the lackof H2 reoxidation as a result of an inactivated uptakehydrogenase (Lindberg et al., 2004). The higher relativeabundance of Npun_F6212 could therefore be interpretedas a response to overcome this imbalance. Its role in ironstorage, and in heterocysts in general, is, however,unclear as no significant difference was detected ingrowth between WT and ΔNpun_F6212 during the first 5days following Fe limitation (data not shown), nor duringgrowth under nitrogen-depleted (N2-fixing) conditions(Fig. 4A). This is surprising considering the strongheterocyst specificity (Fig. 5; Fig. S1) and the higherFe-binding capacity of Brf compared with Dps proteins(Andrews, 2010). Yet, because of the importance of iron
836 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

homeostasis in heterocysts, other proteins involved in ironregulation might well compensate for the lack ofNpun_F6212. Although we were unable to detect a rolefor Npun_F6212 in H2O2 detoxification, a recent study hasshown that heterologous over-expression of all3940 (theorthologue of Npun_F6212 in Anabaena sp. PCC 7120) inE. coli leads to increased tolerance against both nutrientlimitation and different abiotic stresses, including oxida-tive stress (Narayan et al., 2010). The physiologicalimportance of Npun_F6212 in the context of H2O2/O2
scavenging and iron homeostasis in the heterocystsneeds further clarification because also other enzymesinvolved in these processes are known (Valladares et al.,2003; Zhao et al., 2007; Ermakova et al., 2013).
The sequence and predicted structural similarities(Fig. 2) among Npun_F3730, Npun_R3258, Npun_R5701and Npun_R5799 indicate that they belong to the classicalDps protein family. However, our results show that theirindividual importance in response to H2O2 tolerance is verydifferent (Fig. 4A). The results presented here clearly indi-cate that under Fe replete conditions Npun_F3730 is amajor player in the initial response mechanism towardsH2O2 tolerance, which might include taking part in thedetoxification response as no other peroxide scavengingproteins are able to compensate for the negative growthphenotype caused by the lack of Npun_F3730 activity. Itshould be noted that Npun_F3730 is not the only proteininvolved in H2O2 detoxification, as the added H2O2 inΔNpun_F3730 is actively removed within 24 h (data notshown) but clearly plays an important role in the initialdefense response to prevent oxidative stress. Compensa-tory expression of genes encoding peroxiredoxins (Prxs)and catalases in the strains lacking the ferritin-like proteinsmight well take place. However, our results show that suchcompensatory mechanisms, if present, are not able toreplace the function of Npun_F3730. The importance ofNpun_F3730 was observed for cultures grown either withor without combined nitrogen (data not shown), and thepromoter-GFP localization studies (Fig. 5C and F) supporta role of Npun_F3730 in both ammonium-grown anddiazotrophic cultures. This is interesting as N-limitation hasbeen shown to induce Dps expression in othercyanobacteria (Peña et al., 1995; Sandh et al., 2011). Ourresults also indicate that Npun_F3730 primarily has a rolein vegetative cells, as this protein is the only one out of thefour Dps proteins in N. punctiforme that had a lower orequal relative abundance in heterocysts compared withvegetative cells (Fig. 5; Fig. S1). This finding is also in linewith previous proteomics studies where the relative abun-dance of Npun_F3730 was shown to be similar in N2-fixingcultures as compared with ammonium grown cultures (Owet al., 2009). Similar data were also obtained for theorthologue to Npun_F3730 in Anabaena sp. PCC 7120,All4145 (Stensjö et al., 2007). Our data furthermore show
that Npun_F3730 is of major physiological importance inactively growing cells, which is consistent with the expres-sion pattern of the orthologue to Npun_F3730 in Anabaenasp. PCC 7120, All4145 (Sato et al., 2012). This is differentto both the Dps protein in E. coli and for example the DpsAprotein in Synechococcus elongatus PCC 7942, whichhave protective roles during stationary phase (Almironet al., 1992; Sen et al., 2000).
Peroxidases and catalases are suggested to be themain antioxidants for H2O2 in bacteria (Mishra and Imlay,2012). The strong phenotype of ΔNpun_F3730 is there-fore unexpected considering that in N. punctiforme sevengenes are annotated as Prxs and three as catalases [twoMn-catalases, and one Fe-catalase, Cyanobase (http://genome.microbedb.jp/cyanobase/), 23 January 2013].Although H2O2 detoxification activity has been shown invitro and overexpression strains have shown to be moretolerant to H2O2 (Banerjee et al., 2011; 2012), the physi-ological importance of Prxs and catalases in Nostoc spp.is not resolved. However, when deleting the katG gene inthe unicellular cyanobacterium Synechocystis sp. PCC6803, encoding the only catalase-peroxidase in thisorganism, it was possible to observe that the strain had anormal phenotype and its doubling time was similar to theone measured for the WT (Tichy and Vermaas, 1999).Most interestingly, resistances to H2O2 and methylviologen were indistinguishable from those of the WT,even though the rate of H2O2 decomposition was approxi-mately 30 times lower in the mutant. This set of results ledthe authors to suggest that the catalase-peroxidase KatGin Synechocystis sp. PCC 6803 plays a protective role inH2O2 detoxification in the environment, where the mostpotentially damaging H2O2 is extracellular, generated byother bacteria and algae, and that a thiol-specificperoxidase should account for protecting Synechocystissp. PCC 6803 from H2O2 exposure under laboratory con-ditions (Tichy and Vermaas, 1999). In contrast, MrgA, aDps-like protein, seems to have a key role in protectingcells against exogenous oxidative and light stress inSynechocystis sp. PCC 6803 (Foster et al., 2009;Shcolnick et al., 2009). When ΔmrgA cells were exposedto H2O2 treatment, up to 60% of the cells were damagedafter only 30 min of exposure, while approximately 5% ofthe WT cells became damaged upon the same treatment.Similar results were observed when ΔmrgA cells wereexposed to high light (Foster et al., 2009). It also becameapparent from other studies that MrgA has a very activerole in the internal transport of Fe in this cyanobacterium(Shcolnick et al., 2007). Thus, it was suggested that MrgAhas a pivotal role in coordinating iron homeostasis and theoxidative stress response because its action, togetherwith a ferritin-type protein complex, would allow a suc-cessful and efficient mobilization of stored Fe within thecells, minimizing damage caused by the interaction of
The role of Dps proteins in Nostoc punctiforme 837
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

reduced iron and oxygen radicals, which are abundant incyanobacterial cells because of the function of the photo-synthetic apparatus (Shcolnick et al., 2009). Interestingly,the best homologue of MrgA in N. punctiforme isNpun_F3730, sharing approximately 62% of amino acidsequence identity. Therefore, it is possible thatNpun_F3730 plays a role in N. punctiforme similar to theone MrgA does in Synechocystis sp. PCC 6803, giventhat the gene-encoding Npun_F3730 was upregulatedupon H2O2 treatment (Fig. 3) as described for mrgA inSynechocystis sp. PCC 6803 (Li et al., 2004), and that aN. punctiforme strain lacking Npun_F3730 is hypersensi-tive to H2O2 (Fig. 4). What is remarkable in N. punctiformeis that it has three other Dps-like proteins (Npun_R5701,Npun_R3258 and Npun_R5799), none of them capable ofrescuing the ΔNpun_F3730 phenotype. The data pre-sented here support our initial hypothesis that differentmembers of the Dps protein family may have distinct rolesin N. punctiforme. What the other Dps proteins do and inwhat circumstances they are active remains unknown, butrepresent one of the areas where we have been focusingour research.
In addition to the mechanisms described, two proteinshave also clearly shown to have an in vivo role in render-ing Anabaena sp. PCC 7120 cultures more tolerant toH2O2: a sulforedoxin encoded by asl4146 (Boileau et al.,2011) and a rubrerythrin encoded by alr1174 (Zhao et al.,2007). Both these genes are located in direct geneticvicinity to dps genes either upstream and in the sameorientation of all4145 (orthologue of Npun_F3730), orupstream and in the reverse orientation of all1173 (theorthologue of Npun_R5701) (Fig. S3).
The H2O2 sensitivity of ΔNpun_F3730 is observed irre-spective of the antibiotic cassette orientation and is not anartefact of ectopic expression from the cassette’s strongpromoter. Given the lack of effect of H2O2 exposure to theiron-starved culture of ΔNpun_F3730 (Fig. 4B), it appearsthat the level of iron, and thus Fenton chemistry, is animportant factor of the decreased H2O2 tolerance inΔNpun_F3730.
Moreover, the capacity of simultaneously oxidizingFe2+ and reducing H2O2, e.g. to prevent Fenton chemistryand oxidative stress, is a capacity of Npun_F3730that cannot be replaced by other antioxidants inN. punctiforme. We are currently investigating the tran-scriptional regulation of the Npun_F3730 gene clusterand further characterizing Npun_F3730 using both invivo and in vitro biochemical techniques. Althoughheterocysts have a lower oxygen tension, ROS formationin heterocysts can be detected under stressful conditionsusing ROS dyes (Zhao et al., 2007). Both Npun_F3730(Fig. 5; Ow et al., 2009) and a majority of all annotatedPrxs in N. punctiforme are primarily located in the veg-etative cells (Ow et al., 2009; Ekman et al., 2011), raising
the question of which antioxidants are responsible forH2O2 detoxification in the heterocysts.
Based on the GFP expression, the two Dps genesNpun_F3730 and Npun_R5710 showed different expres-sion patterns, e.g. while Npun_F3730 (Fig. 5F) was highlyexpressed in nitrogen-supplemented media as well as inN2-fixing cultures, Npun_R5701 (Fig. 5E) expression wasincreased after transfer to nitrogen depleted media. Theseresults are also in line with previous proteomics data(Fig. S1, Ow et al., 2009) and support a role ofNpun_R5701 that is related to growth in the absence ofcombined nitrogen and/or to heterocyst function. That nosuch phenotype could be identified (i.e. growth rate underN-deplete conditions) could possibly be due to compen-sation effects of any of the other two Dps proteins ofN. punctiforme, Npun_R5799 or Npun_R3258, especiallygiven that both these proteins are also more abundant inheterocysts than in vegetative cells (Fig. S1; Ow et al.,2009). Indeed, the recent work on mapping transcriptionalstart sites (TSS) in Anabaena sp. PCC 7120 transcriptshas revealed two TSS regulating the expression ofall3808 (the orthologue of Npun_R5799), one of thembeing heterocyst specific (Mitschke et al., 2011). Further-more, Sato and colleagues (2012) recently showed thatthe inactivation of the Anabaena sp. PCC 7120orthologue to Npun_R3258 (all0458) was compensatedby increased transcript level of another dps gene, theorthologue of Npun_R5799 (alr3808).
Despite the findings described here regarding cell-typelocalization of the ferritin-like genes’ expression inN. punctiforme, the transcriptional network regulating oxi-dative stress response in cyanobacteria (particularly ofthe filamentous and heterocystous type) is largelyunknown. The response to an increase in superoxideproduction in E. coli is orchestrated by the SoxRS couple(Latifi et al., 2009). Alternatively, H2O2 sensing in E. coliand many other Gram-negative bacteria is achieved by aLysR-type transcription factor, OxyR, while a ferric uptakeregulator (Fur)-like peroxide-sensing repressor, PerR, firstidentified in Bacillus subtilis was shown to be conserved inGram-positive, but also Gram-negative bacteria lackingOxyR (Latifi et al., 2009). On one hand, no SoxRShomologues have been reported in cyanobacteria, andthe regulator sustaining the superoxide response is stillunclear in these phototrophs (Latifi et al., 2009) eventhough we were able to show in a recent study that CalA,an AbrB-like transcription factor, negatively regulates theexpression of FeSOD in Anabaena sp. PCC 7120(Agervald et al., 2010). On the other hand, a PerRorthologue has been identified in Synechocystis sp. PCC6803 (Li et al., 2004). Transcription of the perR gene wasinduced by peroxide, and its inactivation derepressed theexpression of a set of genes including the dps-homologuemrgA and ahpC alkylhydroperoxidase, even in the
838 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

absence of peroxide. However, a majority of the genesthat responded to oxidative stress were not regulated byPerR. This suggests that in contrast to its role inB. subtilis, PerR does not act as the master regulator ofthe peroxide response in Synechocystis sp. PCC 6803(Latifi et al., 2009). What is also interesting is that filamen-tous heterocystous cyanobacteria do not have a PerRorthologue, and the transcription factor with highesthomology is FurA (encoded by all1691 in Anabaena sp.PCC 7120 and Npun_F0997 in N. punctiforme) sharing31% of amino acid sequence identity. Regardless of thelow homology level, the Fur protein is one of the mostimportant switches for the regulation of iron metabolism inprokaryotes. FurA is a constitutive and relatively abundantprotein that seems to be essential for the in vitro growth ofAnabaena sp. PCC 7120 (Pellicer et al., 2012), becauseattempts at getting a fully segregated knockout mutant ofthis protein have invariably failed. FurA has been shown inAnabaena sp. PCC 7120 to be directly involved in regu-lation of several different targets belonging to differentfunctional categories including photosynthesis, energymetabolism, oxidative stress defenses, redox regulation,signal transduction mechanisms, DNA replication,thiamine biosynthesis and heterocyst differentiation(González et al., 2011). Interestingly, a strain of Anabaenasp. PCC 7120 overexpressing FurA has been shown topossess a decreased activity of both catalase andsuperoxide dismutase (González et al., 2010). Moreover,the Dps-like proteins All4145 (orthologue of Npun_F3730)and Alr3808 (orthologue of Npun_R5799) have beenshown to be under the control of the global iron regulatorFurA, working in both cases as a repressor (Hernándezet al., 2007; González et al., 2012). FurA-DNA-bindingactivity has been reported to be lost in the presence ofH2O2 and restored by thiol-reducing agents (Hernándezet al., 2006); thus, one could postulate that an excess ofintracellular H2O2 could impair binding of FurA to the pro-moter of all4145 and lead to a derepression of its tran-scription. It is thus tempting to assume that FurA inN. punctiforme may also regulate the transcription ofNpun_F3730, being one of the regulators mediating H2O2
sensing and transcription response. Nevertheless, thispossibility deserves to be further tested. Additionally, thepossible contribution of two other Fur-like transcriptionregulators, FurB (Npun_6581) and FurC (Npun_R5017)on the control of Npun_F3730 transcription, should not bediscarded either.
Finally, concerning Dps-like protein subcellular localiza-tion, studies performed in Anabaena sp. PCC 7120 haveshown that All1173 (the orthologue of Npun_R5701) andAll0458 (the orthologue of Npun_R3258) are localizedin the nucleoid of vegetative cells of a nitrogen-supplemented culture, suggesting a possible involvementin DNA protection (Sato et al., 2012). From work on the
same cyanobacterium, it is also known that the orthologueof Npun_R5701 (All1173) forms a native 12-mer Dpsstructure and protects DNA from H2O2 when expressed inE. coli (Wei et al., 2007). Additional studies definitelyneed to be performed to further dissect the physiologicalrole of each Dps-like protein in filamentous heterocyst-forming cyanobacteria.
An important task within the future oxidative stressresearch in cyanobacteria will be to resolve the regulationand interaction of the different players involved in H2O2
tolerance, i.e. ferritin-like proteins, Prxs and catalases ona systems level.
Taken together, we have demonstrated thatNpun_F3730 has a general role as being the majorprotein that renders cells tolerant to H2O2 in N.punctiforme independent on nitrogen status of thecultures, and the Bfr Npun_F6212 has a heterocyst-specific role, possibly connected to iron homeostasis andO2/H2O2 scavenging. The physiological roles of the threeadditional Dps proteins (Npun_R5701, Npun_R5799 andNpun_R3258) in N. punctiforme are currently underfurther investigation in our lab.
Conclusions
The work presented here is the first study that combinescell-type specific, phylogenetic, sequence-related, regula-tory and functional studies of members of the ferritin-likeprotein family in a multicellular cyanobacterium. We wereable to gather data in strong support of the multiple Dpsproteins encoded in the genome of N. punctiforme havingdifferent functions and being expressed in distinct cells, aswell as during different growth conditions. Furtherresearch is now ongoing to analyse the regulation andfunction of these five ferritin-like proteins both in vitroand in vivo under a wide range of environmentalconditions. We will also investigate their role inhormogonia and akinetes, which are vital cell types toNostoc’s symbiotic competence and to the ecologicalsuccess of the wide range of hosts that this cyanobacterialgenus interacts with.
Experimental procedures
Organisms and growth conditions
Nostoc punctiforme strain ATCC 29133-S (known as UCD153 Campbell et al., 2007; 2008) was grown in BG110
medium [BG11 medium (Rippka et al., 1979) free of com-bined nitrogen] or BG110 medium supplemented with 5 mMNH4Cl and 10 mM Hepes-NaOH buffer (pH 7.8) at 30°C inthe light (25–40 μmol photons m−2 s−1) in shaken (100 r.p.m.)in 6 or 50 ml of cultures or in 400 ml up to 1.4 l cultures withaeration and stirring, or grown on medium solidified with 1%Noble agar (BD, Difco, Franklin Lakes, NJ, USA), unless
The role of Dps proteins in Nostoc punctiforme 839
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

stated otherwise. For the Δstrains described below, Nm wasused in liquid media at 12.5 μg ml−1 and in solid media at25 μg ml−1. Escherichia coli strain DH5α, used for plasmidconstructions, and strains HB101 and ED8654, used for con-jugations with N. punctiforme strains were grown in LB(Luria–Bertani) medium, supplemented when appropriatewith antibiotics at standard concentrations.
In silico analyses of ferritin-like genes in N. punctiforme
All cyanobacterial amino acid sequences belonging to theferritin-like protein family (Pfam00210) were downloaded inMarch 2013 from the Integrated Microbial Genomes data-base (http://img.jgi.doe.gov/cgi-bin/w/main.cgi). In additionto the cyanobacterial sequences, a selection of non-cyanobacterial sequences (eukaryotic, bacterial andarchaeal) belonging to the ferritin protein family were down-loaded from the Uniprot database (http://www.uniprot.org).These sequences were then aligned with MUSCLE software(http://www.drive5.com/muscle/) using default settings. Theresulting alignments were used for constructing approxi-mately maximum-likelihood phylogenetic trees with theFASTTREE software (http://www.microbesonline.org/fasttree/)using default settings. The sequences shown in Fig. 2 werealigned also with MUSCLE software, and this alignment wasused for constructing the approximately maximum-likelihoodphylogenetic tree shown in Fig. 2, with the FASTTREE softwareusing default settings. Secondary structure predictions wereperformed with PHYRE 2 software (Kelley and Sternberg,2009) (http://www.sbg.bio.ic.ac.uk/phyre2/html/).
Transcript level analysis
Duplicate cultures of N. punctiforme-S were grown in 1.4 lBG-110 medium at 30°C in light (40 μmol photons m−2 s−1).H2O2 was added to the cultures to a final concentration of5 mM, and growth was continued under the same conditions.Cells were harvested at 30 min after treatment. RNA wasisolated as previously described (Agervald et al., 2008). TheRNA quality and concentration was analysed with theExperion System (Bio-Rad, Hercules, CA, USA) accordingto the manufacturer’s instructions. Prior to RT reactions,RNA was treated with DNaseI (Fisher Scientific-Sweden,Gothenburg, Sweden), and the RNA was converted to cDNAusing iScriptTM cDNA Synthesis Kit (Bio-Rad) according tothe manufacturers’ protocol. Experimental procedures andcalculation of relative mRNA levels were carried out asdescribed (Ow et al., 2008). Samples with no added reversetranscriptase were used as a negative control. rnpB was usedas reference gene. All primers used are listed in Table S2. Twobiological and two technical replicates were used in theRT-qPCR analysis.
Construction of cyanobacterial mutants
Genomic DNA was isolated from N. punctiforme as describedpreviously (Tamagnini et al., 1997). Gene deletions were per-formed by replacing the ferritin-like gene by the Nm resis-tance encoding gene cassette C.K3 in both forward andreverse directions, according to Ekman et al. 2013 (Fig. S4;
primers are listed in Table S3). To produce genomic GFPreporter strains (Np5701-GFP and Np6212-GFP) for locali-zation of gene expression, a fragment including the six firstcodons of Npun_R5701 and Npun_F6212 and the upstreamregion of the same gene (3287 bp for Npun_F6212and 1689 bp for Npun_R5701; Fig. S4B) was amplifiedby PCR using primer pairs 6212-PstI/6208-SacIXhoI,NpR5701GFP_F/NpR5701GFP_R. The forward primer ineach primer pair contains a PstI restriction site, and thereverse primer contains both a XhoI and a SacI restrictionsite. Nostoc punctiforme DNA was used as template. Afterdigestion with PstI and SacI, these fragments were cloned toPstI/SacI-digested pUPO2 vector [pBluescript where thegfp-mut1 gene (amplified with oligonucleotides GFP-F andGFP-R, see Table S3) bearing PstI and XhoI on its 5′ and 3′ends respectively) had been inserted as blunt PCR fragmentin the EcoRV site] producing a fusion of the gfp gene to thedps/bfr upstream region. The promoter-gfp translationalfusion was subsequently transferred as a XhoI fragment toXhoI digested pUPO1 vector (a derivative of mobilizablevector pCSV3 (Pernil et al., 2008) in which the antibioticcassette C.S3 was replaced by the Nm resistance cassetteC.K3 in direct orientation (P. Oliveira, unpublished). Transferof the final deletion and genomic GFP reporter constructs(cargo plasmids) into the genome of N. punctiforme was per-formed by conjugation based on previously described proce-dures (Elhai et al., 1997). The genetic structure of theresulting N. punctiforme exconjugants was studied by PCRusing the primer pairs indicated in Fig. S4. Np3730-GFP wasunlike the previous constructs produced as a vector basedGFP reporter strain. The selected promoter region upstreamof Npun_F3730 was amplified by PCR (Table S3). Afterdigestion with PstI and KpnI, this fragment was cloned toPstI/KpnI-digested pSUN119 GFP reporter vector (Arguetaet al., 2004). The vector was subsequently transferred intoN. punctiforme strain ATCC 29133-S using electroporation asdescribed earlier (Holmqvist et al., 2009) to produce the GFPreporter strain Np3730-GFP.
Characterization of deletion strains using H2O2 andiron limitation
In both the H2O2 and iron limitation experiments, WTN. punctiforme and the three ferritin-like knock-out strains,ΔNpun_R5701, ΔNpun_F3730 and ΔNpun_F6212, weregrown in BG110/NH4Cl/Hepes and 12.5 μg ml−1 Nm (for themutant strains), in 400 ml cultures in light (40 μmol photonsm−2 s−1) until a sufficient cell concentration was reached. Forthe H2O2 treatment, cells were washed twice in BG110 beforebeing transferred into BG110 to a final volume of 400 ml anda final optical density at 750 nm (OD750nm) of 0.135. Cultureswere maintained under these conditions for 24h, after whichthe presence of mature heterocysts was determined by lightmicroscopy. H2O2 was then added to a final concentration of0.5 mM, while an equal volume of H2O was added to thecontrol cultures at time zero. OD750nm and Chl a concentrationwere monitored at times 0, 24, 48, 72, 120 and 168 h after thestart of the experiment.
For the iron limitation experiments, the cells were washedtwice in combined nitrogen and iron-free YBG11 media (con-taining 0.3 μm Fe, Shcolnick et al., 2007) before being trans-
840 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

ferred into YBG11 media containing Fe but no combinednitrogen source. Optical density at 750 nm and Chl a concen-tration were determined, and cultures were diluted to OD750
∼ 0.1. Cultures were divided into six well plates and 50 μM ofthe iron-binding siderophore deferoxamine (Sigma, Aldrich,Seelze, Germany) was added to triplicates of each culture toinduce iron limitation. Cells were then grown for 48 h until ironlimitation was detectable, as determined by OD750nm and Chla measurements. At this point, H2O2 was added to a finalconcentration of 0.5 mM, to both iron-limited (–Fe) culturesand control cultures (+ Fe). OD750nm and Chl a concentrationwere determined at regular time points. The external H2O2
concentration was determined by Ferrous ion oxidationxylenol orange 1 (FOX1) assay according to Wolff (1994).
Microscopy
Nostoc punctiforme WT and the promoter GFP strains weregrown in liquid cultures and for microscopy spotted ontoBG110 plates with or without ammonia. Samples were ana-lysed in a Leica DM6000CS confocal laser scanning micro-scope (Leica microsystems, Wetzlar, Germany) equippedwith a TCS SP5 confocal unit and a 100 mW argon laser.The samples were excited using a 488 nm laser, and emis-sion was collected for GFP between 500 and 525 nm andfor the photosynthetic pigments of the cells between 600and 700 nm. Images were taken with a 70 μm pinhole overa 512 × 512 pixel area, and every line was averaged sixtimes. Settings (laser intensities and gain) were the samefor all images shown. Images were analysed by the opensource software ImageJ (http://rsbweb.nih.gov/ij/index.html),measuring the mean gray value in 10 consecutive cellsalong a filament. To assess a significant difference of fluo-rescence, a paired t-test at P-value < 0.05 in MINITAB®
Release 14 (Minitab Inc. State College, PA, USA) was per-formed. The average values for 10 different filaments wereanalysed.
Acknowledgements
This work was supported by the Swedish Energy Agency, theKnut and Alice Wallenberg Foundation, the FP7-ENERGY-2012-1-2STAGE (Project number 308518) EU project Cyano-Factory, the Carl Tryggers Foundation, and the MagnusBergvall Foundation. PO acknowledges Fundação para aCiência e a Tecnologia (FCT) for financial support (fellowshipSFRH/BPD/74894/2010). We wish to express our gratitude toProf. Enrique Flores (University of Sevilla, Spain) for thegenerous gift of plasmid pCSV3. We also want to thank Dr. PiaLindberg, for rnpB primers used for the RT-qPCR, and Assoc.Prof. Fikret Mamedov for help with the iron experiments. Wealso acknowledge the undergraduate students Lei Chen,Ievgen Dzhygyr, Xin Li and Niklas Lundberg for skilled experi-mental assistance. The authors declare no conflict of interest.
References
Agervald, A., Stensjö, K., Holmqvist, M., and Lindblad, P.(2008) Transcription of the extended hyp-operon in Nostocsp. strain PCC 7120. BMC Microbiol 8: 69.
Agervald, A., Baebprasert, W., Zhang, X., Incharoensakdi, A.,Lindblad, P., and Stensjö, K. (2010) The CyAbrB transcrip-tion factor CalA regulates the iron superoxide dismutase inNostoc sp. strain PCC 7120. Environ Microbiol 12: 2826–2837.
Alaleona, F., Franceschini, S., Ceci, P., Ilari, A., andChiancone, E. (2010) Thermosynechococcus elongatusDpsA binds Zn(II) at a unique three histidine-containingferroxidase center and utilizes O2 as iron oxidant with veryhigh efficiency, unlike the typical Dps proteins. FEBS J 277:903–917.
Almiron, M., Link, A.J., Deirdre, F., and Kolter, R. (1992) Anovel DNA-binding protein with regulatory and protectiveroles in starved Escherichia coli. Genes Dev 6: 2646–2654.
Andrews, S.C. (2010) The Ferritin-like superfamily: evolutionof the biological iron storeman from a rubrerythrin-likeancestor. Biochim Biophys Acta 1800: 691–705.
Argueta, C., Yuksek, K., and Summers, M. (2004) Construc-tion and use of GFP reporter vectors for analysis of cell-type-specific gene expression in N. punctiforme. JMicrobiol Methods 59: 181–188.
Azam, T.A., and Ishihama, A. (1999) Twelve species of thenucleoid-associated protein from Escherichia coli.Sequence recognition specificity and DNA binding affinity.J Biol Chem 274: 33105–33113.
Banerjee, M., Ballal, A., and Apte, S.K. (2011) A novelglutaredoxin domain-containing peroxiredoxin ‘All1541’protects the N2-fixing cyanobacterium Anabaena PCC7120 from oxidative stress. Biochem J 442: 671–680.
Banerjee, M., Ballal, A., and Apte, S.K. (2012) Mn-catalase(Alr0998) protects the photosynthetic, nitrogen-fixingcyanobacterium Anabaena PCC7120 from oxidativestress. Environ Microbiol 14: 2891–2900.
Bergman, B., Sandh, G., Lin, S., Larsson, J., and Carpenter,E.J. (2013) Trichodesmium – a widespread marinecyanobacterium with unusual nitrogen fixation properties.FEMS Microbiol Rev 37: 286–302.
Boileau, C., Eme, L., Brochier-Armanet, C., Janicki, A.,Zhang, C.C., and Latifi, A. (2011) A eukaryotic-likesulfiredoxin involved in oxidative stress responses and inthe reduction of the sulfinic form of 2-Cys peroxiredoxin inthe cyanobacterium Anabaena PCC 7120. New Phytol191: 1108–1118.
Bou-Abdallah, F. (2010) The iron redox and hydrolysischemistry of the ferritins. Biochim Biophys Acta 1800: 719–731.
Campbell, E.L., Summers, M.L., Christman, H., Martin, M.E.,and Meeks, J.C. (2007) Global gene expression patterns ofNostoc punctiforme in steady-state dinitrogen-grownheterocyst-containing cultures and at single time pointsduring the differentiation of akinetes and hormogonia. JBacteriol 189: 5247–5256.
Campbell, E.L., Christman, H., and Meeks, J.C. (2008) DNAmicroarray comparisons of plant factor- and nitrogendeprivation-induced Hormogonia reveal decision-makingtranscriptional regulation patterns in Nostoc punctiforme. JBacteriol 190: 7382–7391.
Castruita, M., Saito, M., Schottel, P.C., Elmegreen, L.A.,Myneni, S., Stiefel, E.I., and Morel, F.M. (2006)Overexpression and characterization of an iron storage
The role of Dps proteins in Nostoc punctiforme 841
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

and DNA-binding Dps protein from Trichodesmiumerythraeum. Appl Environ Microbiol 72: 2918–2924.
Castruita, M., Elmegreen, L.A., Shaked, Y., Stiefel, E.I., andMorel, F.M. (2007) Comparison of the kinetics of ironrelease from a marine (Trichodesmium erythraeum) Dpsprotein and mammalian ferritin in the presence andabsence of ligands. J Inorg Biochem 101: 1686–1691.
Ceci, P., Cellai, S., Falvo, E., Rivetti, C., Rossi, G.L., andChiancone, E. (2004) DNA condensation and self-aggregation of Escherichia coli Dps are coupled phenom-ena related to the properties of the N-terminus. NucleicAcids Res 32: 5935–5944.
Chiancone, E., and Ceci, P. (2010) The multifaceted capacityof Dps proteins to combat bacterial stress conditions:detoxification of iron and hydrogen peroxide and DNAbinding. Biochim Biophys Acta 1800: 798–805.
Ekman, M., Ow, S.Y., Holmqvist, M., Zhang, X., vanWagenen, J., Wright, P.C., and Stensjö, K. (2011) Meta-bolic adaptations in a H2 producing heterocyst-formingcyanobacterium: potentials and implications for biologicalengineering. J Proteome Res 10: 1772–1784.
Ekman, M., Picossi, S., Campbell, E.L., Meeks, J.C., andFlores, E. (2013) A Nostoc punctiforme sugar transporternecessary to establish a Cyanobacterium-plant symbiosis.Plant Physiol 161: 1984–1992.
Elhai, J., Vepritskiy, A., Muro-Pastor, A.M., Flores, E., andWolk, C.P. (1997) Reduction of conjugal transfer efficiencyby three restriction activities of Anabaena sp. strain PCC7120. J Bacteriol 179: 1998–2005.
Ermakova, M., Battchikova, N., Allahverdiyeva, Y., andAro, E.M. (2013) Novel heterocyst-specific flavodiironproteins in Anabaena sp. PCC 7120. FEBS Lett 587:82–87.
Facey, P.D., Hitchings, M.D., Saavedra-Garcia, P.,Fernandez-Martinez, L., Dyson, P.J., and Del Sol, R.(2009) Streptomyces coelicolor Dps-like proteins: differen-tial dual roles in response to stress during vegetativegrowth and in nucleoid condensation during reproductivecell division. Mol Microbiol 73: 1186–1202.
Foster, J.S., Havemann, S.A., Singh, A.K., and Sherman,L.A. (2009) Role of mrgA in peroxide and light stress in thecyanobacterium Synechocystis sp. PCC 6803. FEMSMicrobiol Lett 293: 298–304.
Franceschini, S., Ceci, P., Alaleona, F., Chiancone, E.,and Ilari, A. (2006) Antioxidant Dps protein from thethermophilic cyanobacterium Thermosynechococcuselongatus. FEBS J 273: 4913–4928.
Ghatak, P., Karmakar, K., Kasetty, S., and Chatterji, D. (2011)Unveiling the role of Dps in the organization ofmycobacterial nucleoid. PLoS One 6: e16019.
González, A., Bes, M.T., Barja, F., Peleato, M.L., and Fillat,M.F. (2010) Overexpression of FurA in Anabaena sp. PCC7120 reveals new targets for this regulator involved inphotosynthesis, iron uptake and cellular morphology. PlantCell Physiol 51: 1900–1914.
González, A., Bes, M.T., Peleato, M.L., and Fillat, M.F. (2011)Unravelling the regulatory function of FurA in Anabaena sp.PCC 7120 through 2-D DIGE proteomic analysis. JProteomics 74: 660–671.
González, A., Bes, M.T., Valladares, A., Peleato, M.L., andFillat, M.F. (2012) FurA is the master regulator of iron
homeostasis and modulates the expression of tetrapyrrolebiosynthesis genes in Anabaena sp. PCC 7120. EnvironMicrobiol 14: 3175–3187.
Hernández, J.A., López-Gomollón, S., Muro-Pastor, A.,Valladares, A., Bes, M.T., Peleato, M.L., and Fillat, M.F.(2006) Interaction of FurA from Anabaena sp. PCC 7120with DNA: a reducing environment and the presence ofMn(2+) are positive effectors in the binding to isiB and furApromoters. Biometals 19: 259–268.
Hernández, J.A., Pellicer, S., Huang, L., Peleato, M.L., andFillat, M.F. (2007) FurA modulates gene expression ofalr3808, a DpsA homologue in Nostoc (Anabaena) sp.PCC7120. FEBS Lett 581: 1351–1356.
Holmqvist, M., Stensjö, K., Oliveira, P., Lindberg, P., andLindblad, P. (2009) Characterization of the hupSL promoteractivity in N. punctiforme ATCC 29133. BMC Microbiol 9:54.
Kelley, L.A., and Sternberg, M.J.E. (2009) Protein structureprediction on the web: a case study using the Phyre server.Nat Protoc 4: 363–371.
Keren, N., Aurora, R., and Pakrasi, H.B. (2004) Critical rolesof bacterioferritins in iron storage and proliferation ofcyanobacteria. Plant Physiol 135: 1666–1673.
Kranzler, C., Rudolf, M., Keren, N., and Schleiff, E. (2013)Iron in cyanobacteria. In Advances in Botanical Research.Chauvat, F., and Cassier-Chauvat, C. (eds). Amsterdam,the Netherlands: Elsevier, 65, pp. 57–105.
Kumar, K., Mella-Herrera, R.A., and Golden, J.W. (2010)Cyanobacterial heterocysts. Cold Spring Harb PerspectBiol 2: a000315.
Latifi, A., Jeanjean, R., Lemeille, S., Havaux, M., and Zhang,C.C. (2005) Iron starvation leads to oxidative stress inAnabaena sp. strain PCC 7120. J Bacteriol 187: 6596–6598.
Latifi, A., Ruiz, M., and Zhang, C.C. (2009) Oxidative stress incyanobacteria. FEMS Microbiol Rev 33: 258–278.
Li, H., Singh, A.K., McIntyre, L.M., and Sherman, L.A. (2004)Differential gene expression in response to hydrogen per-oxide and the putative PerR regulon of Synechocystis sp.strain PCC 6803. J Bacteriol 186: 3331–3345.
Lindberg, P., Lindblad, P., and Cournac, L. (2004) Gasexchange in the filamentous cyanobacterium Nostocpunctiforme strain ATCC 29133 and its hydrogenase-deficient mutant strain NHM5. Appl Environ Microbiol 70:2137–2145.
Martinez, A., and Kolter, R. (1997) Protection of DNA duringoxidative stress by the nonspecific DNA-binding proteinDps. J Bacteriol 179: 5188–5194.
Meeks, J.C., Campbell, E.L., Summers, M.L., and Wong, F.C.(2002) Cellular differentiation in the cyanobacteriumNostoc punctiforme. Arch Microbiol 178: 395–403.
Mishra, S., and Imlay, J. (2012) Why do bacteria use so manyenzymes to scavenge hydrogen peroxide? Arch BiochemBiophys 525: 145–160.
Mitschke, J., Vioque, A., Haas, F., Hess, W.R., andMuro-Pastor, A.M. (2011) Dynamics of transcriptional startsite selection during nitrogen stress-induced cell differen-tiation in Anabaena sp. PCC7120. Proc Natl Acad Sci USA108: 20130–20135.
Morris, J.J., Johnson, Z.I., Szul, M.J., Keller, M., and Zinser,E.R. (2011) Dependence of the cyanobacterium
842 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

Prochlorococcus on hydrogen peroxide scavengingmicrobes for growth at the ocean’s surface. PLoS One 6:e16805.
Narayan, O.P., Kumari, N., and Rai, L.C. (2010) Heterologousexpression of Anabaena PCC 7120 all3940 (a Dps familygene) protects Escherichia coli from nutrient limitation andabiotic stresses. Biochem Biophys Res Commun 394:163–169.
Ow, S.Y., Cardona, T., Taton, A., Magnuson, A., Lindblad, P.,Stensjö, K., and Wright, P.C. (2008) Quantitative shotgunproteomics of enriched heterocysts from Nostoc sp. PCC7120 using 8-plex isobaric peptide tags. J Proteome Res 7:1615–1628.
Ow, S.Y., Noirel, J., Cardona, T., Taton, A., Lindblad, P.,Stensjö, K., and Wright, P.C. (2009) Quantitative overviewof N2 fixation in N. punctiforme ATCC 29133 through cellu-lar enrichments and iTRAQ shotgun proteomics. JProteome Res 8: 187–198.
Partensky, F., and Garczarek, L. (2010) Prochlorococcus:advantages and limits of minimalism. Ann Rev Mar Sci 2:305–331.
Pellicer, S., González, A., Peleato, M.L., Martinez, J.I., Fillat,M.F., and Bes, M.T. (2012) Site-directed mutagenesis andspectral studies suggest a putative role of FurA fromAnabaena sp. PCC 7120 as a heme sensor protein. FEBSJ 279: 2231–2246.
Peña, M.M., and Bullerjahn, G.S. (1995) The DpsA protein ofSynechococcus sp. Strain PCC7942 is a DNA-bindinghemoprotein. Linkage of the Dps and bacterioferritinprotein families. J Biol Chem 22: 22478–22482.
Peña, M.M., Burkhart, W., and Bullerjahn, G.S. (1995) Puri-fication and characterization of a Synechococcus sp. strainPCC 7942 polypeptide structurally similar to the stress-induced Dps/PexB protein of Escherichia coli. ArchMicrobiol 163: 337–344.
Pernil, R., Picossi, S., Mariscal, V., Herrero, A., and Flores, E.(2008) ABC-type amino acid uptake transporters Bgt andN-II of Anabaena sp. strain PCC 7120 share an ATPasesubunit and are expressed in vegetative cells andheterocysts. Mol Microbiol 67: 1067–1080.
Reon, B.J., Nguyen, K.H., Bhattacharyya, G., and Grove, A.(2012) Functional comparison of Deinococcus radioduransDps proteins suggests distinct in vivo roles. Biochem J447: 381–391.
Rippka, R., Deruelles, J., Waterbury, J., Herdman, M., andStanier, R. (1979) Generic assignments, strain historiesand properties of pure cultures of cyanobacteria. J GenMicrobiol 111: 1–61.
Sandh, G., Ran, L., Xu, L., Sundqvist, G., Bulone, V., andBergman, B. (2011) Comparative proteomic profiles ofthe marine cyanobacterium Trichodesmium erythraeumIMS101 under different nitrogen regimes. Proteomics 11:406–419.
Sato, N., Moriyama, T., Toyoshima, M., Mizusawa, M., andTajima, N. (2012) The all0458/lti46.2 gene encodes a lowtemperature-induced Dps protein homologue in thecyanobacteria Anabaena sp. PCC 7120 and Anabaenavariabilis M3. Microbiology 158: 2527–2536.
Schwartz, J.K., Liu, X.S., Tosha, T., Diebold, A., Theil, E.C.,and Solomon, E.I. (2010) CD and MCD spectroscopicstudies of the two Dps miniferritin proteins from Bacillus
anthracis: role of O2 and H2O2 substrates in reactivity of thediiron catalytic centers. Biochemistry 49: 10516–10525.
Sen, A., Dwivedi, K., Rice, K.A., and Bullerjahn, G.S. (2000)Growth phase and metal-dependent regulation of the dpsAgene in Synechococcus sp. strain PCC 7942, USA. ArchMicrobiol 173: 352–357.
Shcolnick, S., Shaked, Y., and Keren, N. (2007) A role formrgA, a DPS family protein, in the internal transport of Fein the cyanobacterium Synechocystis sp. PCC6803.Biochim Biophys Acta 1767: 814–819.
Shcolnick, S., Summerfield, T.C., Reytman, L., Sherman,L.A., and Keren, N. (2009) The mechanism of iron homeo-stasis in the unicellular cyanobacterium Synechocystis sp.PCC 6803 and its relationship to oxidative stress. PlantPhysiol 150: 2045–2056.
Shih, P.M., Wu, D., Latifi, A., Axen, S.D., Fewer, D.P., Talla,E., et al. (2013) Improving the coverage of thecyanobacterial phylum using diversity-driven genomesequencing. Proc Natl Acad Sci USA 110: 1053–1058.
Stensjö, K., Ow, S.Y., Barrios-Llerena, M.E., Lindblad, P., andWright, P.C. (2007) An iTRAQ-based quantitative analysisto elaborate the proteomic response of Nostoc sp. PCC7120 under N2 fixing conditions. J Proteome Res 6: 621–635.
Tamagnini, P., Troshina, O., Oxelfelt, F., Salema, R., andLindblad, P. (1997) Hydrogenases in Nostoc sp. strain PCC73102, a strain lacking a bidirectional enzyme. ApplEnviron Microbiol 63: 1801–1807.
Tichy, M., and Vermaas, W. (1999) In vivo role of catalase-peroxidase in Synechocystis sp. strain PCC 6803. JBacteriol 181: 1875–1882.
Tu, W.Y., Pohl, S., Gizynski, K., and Harwood, C.R. (2012) Theiron-binding protein Dps2 confers peroxide stress resist-ance on Bacillus anthracis. J Bacteriol 194: 925–931.
Valladares, A., Herrero, A., Pils, D., Schmetterer, G., andFlores, E. (2003) Cytochrome c oxidase genes required fornitrogenase activity and diazotrophic growth in Anabaenasp. PCC 7120. Mol Microbiol 47: 1239–1249.
Wei, X., Mingjia, H., Xiufeng, L., Yang, G., and Qingyu, W.(2007) Identification and biochemical properties ofDps (starvation-induced DNA binding protein) fromcyanobacterium Anabaena sp. PCC 7120. IUBMB Life 59:675–681.
Wolff, S.P. (1994) Ferrous ion oxidation in presence of ferricion indicator xylenol orange for measurement ofhydroperoxides. Methods Enzymol 233: 182–189.
Zehr, J.P., Bench, S.R., Carter, B.J., Hewson, I., Niazi, F., Shi,T., et al. (2008) Globally distributed uncultivated oceanicN2-fixing cyanobacteria lack oxygenic photosystem II.Science 14: 1110–1112.
Zhao, G., Ceci, P., Ilari, A., Giangiacomo, L., Laue, T.M.,Chiancone, E., and Chasteen, N.D. (2002) Iron and hydro-gen peroxide detoxification properties of DNA-bindingprotein from starved cells. A ferritin-like DNA-bindingprotein of Escherichia coli. J Biol Chem 277: 27689–27696.
Zhao, W., Ye, Z., and Zhao, J. (2007) RbrA, a cyanobacterialrubrerythrin, functions as a FNR-dependent peroxidase inheterocysts in protection of nitrogenase from damage byhydrogen peroxide in Anabaena sp. PCC 7120. MolMicrobiol 66: 1219–1230.
The role of Dps proteins in Nostoc punctiforme 843
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844

Supporting information
Additional Supporting Information may be found in the onlineversion of this article at the publisher’s web-site:
Fig. S1. Previously reported protein abundance of ferritin-like proteins in N. punctiforme in heterocysts relative toN2-fixing filaments (Ow et al., 2009). The values are given asLog10-ratios, and dashed line indicates the cut-off value forsignificant changes used in the experiment. All values arestatistically significant, and the error factor (EF) is given aserror bar for all values to show the 95% confidence interval.The asterisk indicates that the value is in-between the cut-offvalues and not used by Ow et al. (2009).Fig. S2. Large-scale phylogenetic approximately maximum-likelihood tree of ferritin-like proteins family consisting ofthree main groups: ferritins (Ftn), bacterioferritins (Bfr) andDps proteins (Dps). Sequences used include all the onesfrom genome-sequenced cyanobacteria and annotated to theferritin-like protein family (Pfam00210), and also a selectionof Ftn family sequences from eukaryotes, bacteria andarchaea. Nostoc punctiforme sequences are found infive different cyanobacteria-dominated clusters, and theN. punctiforme sequences are indicated by a green tag. Allsequences used for constructing the tree can be found assupplemental data (Supplemental FASTA file).Fig. S3. Genomic context of the five ferritin-like proteins (redcolor) in N. punctiforme. Blue colour indicates that a similargene organization is found also in Anabaena sp. PCC 7120.Note that Npun_F6210 is in the IMG database incorrectly notidentified as a gene.
Fig. S4. Cloning construct design.A. Ferritin-like deletion strains of N. punctiforme ATCC29133-S were constructed by replacing the correspondingferritin-like gene with the neomycin resistance cassette(C.K3) in reverse orientation. For Npun_F3730, the cassettewas inserted both in reverse and forward orientation. Theprimers used for amplification of 1 kb flanking region on eachside of the genes to be deleted, later used for homologousrecombination into the genome, are indicated by arrows num-bered 1, 2, 3 and 4.B. The promoter-gfp vector constructs of the promoters ofNpun_R5701, Npun_F6212 and Npun_F3730 were made byfusing upstream regions of the respective gene including 18nt of the gene of interest to gfp-mut1. The primers that wereused to amplify the upstream DNA region of each gene ofinterest are indicated as arrows. The length (bp) of the ampli-fied region is also indicated.Table S1. The distribution of ferritin-like proteins insequenced cyanobacterial genomes. The annotatedmembers of the three COG groups, DPS proteins(COG0783), bacterioferritin (COG2193) and ferritin (diiron,COG1528), are listed below with the number in each cat-egory listed in parentheses after the strain name. Allgenomes containing at least one member of the relevantcategory is written in red, strains lacking representatives arewritten in black.Table S2. Oligonucleotides used for RT-qPCR.Table S3. Oligonucleotides used for deletion and promoter-GFP constructs.
844 M. Ekman et al.
© 2013 Society for Applied Microbiology and John Wiley & Sons Ltd, Environmental Microbiology, 16, 829–844
Copyright © 2022 FDOKUMEN















![[Prevalence and demographic factors associated with ferritin deficiency in Colombian children, 2010]](https://static.fdokumen.com/doc/165x107/63415b14189652a6680a7eb2/prevalence-and-demographic-factors-associated-with-ferritin-deficiency-in-colombian.jpg)




