
Differential expression of SAPK isoforms in the rat brain. An in situ hybridisation study in the...
12
Ž . Molecular Brain Research 60 1998 57–68 Research report Differential expression of SAPK isoforms in the rat brain. An in situ hybridisation study in the adult rat brain and during post-natal development Lucia Carboni ) , Renzo Carletti, Stefano Tacconi, Corrado Corti, Francesco Ferraguti Department of Pharmacology, Glaxo Wellcome Medicines Research Centre, Verona, Italy Accepted 7 July 1998 Abstract MAPK pathways transduce a broad variety of extracellular signals into cellular responses. Despite their pleiotropic effects and their ubiquitous distribution, surprisingly little is known about their involvement in the communication network of nerve cells. As a first step to elucidate the role of MAPK pathways in neuronal signalling, we studied the distribution of SAPK arJNK2, SAPK brJNK3, and SAPK grJNK1, three isoforms of SAPKrJNK, a stress-activated MAPK subfamily. We compared the mRNA localisation of the three main isoforms in the adult and developing rat brain using in situ hybridisation. In the adult brain, SAPK a and b were widely but heterogeneously distributed, reproducing the pattern of a probe that does not discriminate the isoforms. Differently, high labelling for the SAPK g probe was exclusively localised in the endopiriform nucleus and medial habenula. Intermediate staining was detected in the hippocampus. During post-natal development, SAPK b showed the same localisation as in the adult. Nevertheless, the semi-quantitative analysis of optical densities showed significantly different mRNA levels. In the adult, SAPK g signal was weak, whereas in newborn rats the labelling was intense and widely distributed. SAPK g mRNA levels decreased during development, to reach the low signals detected in the adult. These results suggest that in the central nervous system SAPK-type MAP kinases perform significant physiological functions which are particularly relevant during post-natal development. The distinct distribution patterns of SAPK isoforms in the adult rat brain support the hypothesis that separate functions are performed by the products of the three SAPK genes. q 1998 Elsevier Science B.V. All rights reserved. Keywords: Stress-activated protein kinase; JNK; MAPK; Central nervous system 1. Introduction Ž . Mitogen-activated protein kinase MAPK pathways transduce extracellular stimuli into cellular responses both by direct phosphorylation of target proteins and by trans- activation of transcription factors, which consequently modulate gene expression. Typical MAPK cascades are composed of three enzymes, which are sequentially acti- vated by phosphorylation: a MAPK kinase kinase Ž . Ž . MAPKKK ; a MAPK kinase MAPKK ; and a MAPK Ž . Fig. 1 . Several parallel pathways have been described in w x mammalian cells 24 . Stress-activated protein kinasesrc-Jun NH -terminal ki- 2 Ž . nases SAPKsrJNKs belong to a more recently discov- ) Corresponding author. Molecular Pharmacology, Glaxo Wellcome Medicines Research Centre, via A. Fleming, 4, 37135 Verona, Italy. Fax: q39-45-9218174; E-mail: [email protected] ered MAPK pathway as compared to extracellular-signal Ž . regulated kinases ERKs . SAPKrJNK cascade is stimu- lated by stressful signals, such as osmotic stress, ultraviolet and ionising radiation, heat shock, and reperfusion after w x ischemia 22,17 . It is also activated by physiological w x stimuli, such as hematopoietic cytokines 11 , and G pro- wx tein-coupled receptors 5 . The activation of SAPKrJNK pathway has been associ- ated with many final cellular responses, some of which generate opposite outcomes. Depending on cell or tissue type and experimental conditions, SAPKrJNK stimulation has been reported to induce as different effects as apopto- w x w x w x sis 32 , rescue from apoptosis 21 , proliferation 19 , and w x differentiation 33 . How such a wide range of specific responses can be obtained by the activation of MAPK is not presently clear. The existence of multiple kinases which belong to the stress-activated cascade may contribute to these diverse 0169-328Xr98r$ - see front matter q 1998 Elsevier Science B.V. All rights reserved. Ž . PII: S0169-328X 98 00166-1
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Differential expression of SAPK isoforms in the rat brain. An in situ hybridisation study in the...

Ž .Molecular Brain Research 60 1998 57–68
Research report
Differential expression of SAPK isoforms in the rat brain. An in situhybridisation study in the adult rat brain and during post-natal
development
Lucia Carboni ), Renzo Carletti, Stefano Tacconi, Corrado Corti, Francesco FerragutiDepartment of Pharmacology, Glaxo Wellcome Medicines Research Centre, Verona, Italy
Accepted 7 July 1998
Abstract
MAPK pathways transduce a broad variety of extracellular signals into cellular responses. Despite their pleiotropic effects and theirubiquitous distribution, surprisingly little is known about their involvement in the communication network of nerve cells. As a first step toelucidate the role of MAPK pathways in neuronal signalling, we studied the distribution of SAPK arJNK2, SAPK brJNK3, and SAPKgrJNK1, three isoforms of SAPKrJNK, a stress-activated MAPK subfamily. We compared the mRNA localisation of the three mainisoforms in the adult and developing rat brain using in situ hybridisation. In the adult brain, SAPK a and b were widely butheterogeneously distributed, reproducing the pattern of a probe that does not discriminate the isoforms. Differently, high labelling for theSAPK g probe was exclusively localised in the endopiriform nucleus and medial habenula. Intermediate staining was detected in thehippocampus. During post-natal development, SAPK b showed the same localisation as in the adult. Nevertheless, the semi-quantitativeanalysis of optical densities showed significantly different mRNA levels. In the adult, SAPK g signal was weak, whereas in newborn ratsthe labelling was intense and widely distributed. SAPK g mRNA levels decreased during development, to reach the low signals detectedin the adult. These results suggest that in the central nervous system SAPK-type MAP kinases perform significant physiological functionswhich are particularly relevant during post-natal development. The distinct distribution patterns of SAPK isoforms in the adult rat brainsupport the hypothesis that separate functions are performed by the products of the three SAPK genes. q 1998 Elsevier Science B.V. Allrights reserved.
Keywords: Stress-activated protein kinase; JNK; MAPK; Central nervous system
1. Introduction
Ž .Mitogen-activated protein kinase MAPK pathwaystransduce extracellular stimuli into cellular responses bothby direct phosphorylation of target proteins and by trans-activation of transcription factors, which consequentlymodulate gene expression. Typical MAPK cascades arecomposed of three enzymes, which are sequentially acti-vated by phosphorylation: a MAPK kinase kinaseŽ . Ž .MAPKKK ; a MAPK kinase MAPKK ; and a MAPKŽ .Fig. 1 . Several parallel pathways have been described in
w xmammalian cells 24 .Stress-activated protein kinasesrc-Jun NH -terminal ki-2
Ž .nases SAPKsrJNKs belong to a more recently discov-
) Corresponding author. Molecular Pharmacology, Glaxo WellcomeMedicines Research Centre, via A. Fleming, 4, 37135 Verona, Italy. Fax:q39-45-9218174; E-mail: [email protected]
ered MAPK pathway as compared to extracellular-signalŽ .regulated kinases ERKs . SAPKrJNK cascade is stimu-
lated by stressful signals, such as osmotic stress, ultravioletand ionising radiation, heat shock, and reperfusion after
w xischemia 22,17 . It is also activated by physiologicalw xstimuli, such as hematopoietic cytokines 11 , and G pro-
w xtein-coupled receptors 5 .The activation of SAPKrJNK pathway has been associ-
ated with many final cellular responses, some of whichgenerate opposite outcomes. Depending on cell or tissuetype and experimental conditions, SAPKrJNK stimulationhas been reported to induce as different effects as apopto-
w x w x w xsis 32 , rescue from apoptosis 21 , proliferation 19 , andw xdifferentiation 33 . How such a wide range of specific
responses can be obtained by the activation of MAPK isnot presently clear.
The existence of multiple kinases which belong to thestress-activated cascade may contribute to these diverse
0169-328Xr98r$ - see front matter q 1998 Elsevier Science B.V. All rights reserved.Ž .PII: S0169-328X 98 00166-1

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–6858
Fig. 1. The MAPK pathway transduces extracellular stimuli through a cascade of activated kinases, ending in phosphorylation of transcription factors andcellular proteins. For the SAPK pathway, many different enzymes exist which can work as activators of MAPKKK; as MAPKKK; as MAPKK, and asMAPK. STE20-HOMs: mammalian kinases homologous to STE20, the activator of MAPKKK in S. cereÕisiae. 3pK: chromosome 3p kinase; ASK:apoptosis signal-regulated kinase; ATF2: activating transcription factor 2; DLK: dual leucine zipper bearing kinase; GCK: germinal centre kinase; GLK:GCK-like kinase; HPK: hematopoietic progenitor kinase; HSF: heat shock factor; JNKK: JNK kinase; KHS: kinase homologous to SPS1rSTE20; MEKK:MAPK Erk kinase kinase; MKK: MAPK kinase; MLK: mixed lineage kinase; MST: mammalian STE20-like protein kinase; MTK: MAP three kinase;MUK: MAPK-upstream kinase; NFAT: nuclear factor of activated T cells; NIK: Nck interacting kinase; PAK: p21-activated protein kinase; PTK: proteintyrosine kinase; SEK: SAPK ERK kinase; SPRK: Src-homology 3 domain-containing proline-rich Kinase; TAK: TGF b-activated kinase; Tpl-2: tumorprogression locus 2; ZPK: zipper protein kinase.
physiological responses. Many distinct enzymes have beendiscovered that diverge in selectivity and tissue distribu-
w x w xtion, and which act as MAPKKK 8 , or MAPKK 22,28Ž .in the SAPKrJNK pathway Fig. 1 . Many distinct iso-
forms of SAPKrJNK have been reported as well, whichexhibit different properties of affinity and substrate speci-
w xficity 7,13,15,16,20,26 . In the rat, three main SAP ki-nases, probably arising from three separate genes, havebeen described. They have been named SAPK a , b and g
w x6,16 . Each SAPK gene gives rise to two RNA variants ofdifferent length, which diverge at the 3X-end. The two
Ž .mRNA forms are translated into a short p46 or long
Ž .p55 protein. Moreover, SAPK a exhibits an additionalsite of alternative splicing in protein kinase subdomains IXand X, therefore producing four a isoforms. In humantissue, the three homologous gene products have been
Ž 3F12 .called JNK 2, 3 or p49 and 1, respectively. Shorterand longer mRNA forms were reported for each gene. Thecorresponding site of alternative splicing that was charac-terised for SAPK a has been described for both JNK2 and
Ž . w x1, therefore creating ten isoforms in all Fig. 1 7,13,15,20 .Available data show that the central nervous system
Ž .CNS is highly enriched in SAPK mRNA and protein,which are widely and heterogeneously distributed in brain

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–68 59
w x3,7,13,18,29 . As compared with other tissues, brain dis-plays a higher baseline activation of SAPKrJNK, activa-
w xtion that can be modulated by environmental stimuli 29 .However, no detailed description of the distribution of thedifferent isoforms has yet been reported. At least certainisoforms appear to mediate specific activities. Indeed,products of the Jnk3rSAPK brp49 3F12 gene, which are
w xselectively expressed in brain 20,18 , are reported tomediate glutamate signalling and excitotoxicity in hip-
w xpocampus 31 . Therefore, a detailed analysis of the rela-tive distribution of individual isoforms in brain was neededto provide a neuroanatomical basis on which to supporthypotheses as to their relative function.
The aim of this investigation was to study the localisa-tion of the three SAPK gene products both in the adultbrain and during post-natal development. To this end, weperformed in situ hybridisation with oligonucleotide probeswhich specifically recognised each isoform.
2. Materials and methods
2.1. Tissue preparation
Ž .Adult male 300 g and pregnant female Sprague–Daw-ley rats were purchased from Charles River, Calco, Italy.The research complied with national legislation and withthe company policy on the Care of Use of Animals and
Ž .with related codes of practice. Adult animals ns7 wereŽ .deeply anaesthetised with urethane 1.5 grkg, i.p. and
intracardially perfused with ice-cold saline. Brains wereimmediately removed, frozen in dry-ice cold isopentaneand cryostat-cut into coronal, parasagittal or horizontal 14
mm thick sections; groups of three consecutive slices werecut, to be hybridised to each probe. Sections were thaw-mounted on poly-L-lysine-coated slides and stored aty808C until the day of the experiment. Groups of three
Žrats at different post-natal days P: 1, 3, 6, 9, 12, 15, 18.and 21 were decapitated after urethane anaesthesia; brains
were removed, frozen and cut as previously described.
2.2. Probe choice, synthesis and labelling
Forty-five-mer sequences were chosen in the SAPK a ,b and g cDNAs. SAPK a probe was complementary tonucleotides 1914–1958 in sequence L27111, SAPK b to189-233 in sequence L27128, SAPK g to 131-175 insequence L27129 in GenBank. The specificity of the probeswas evaluated with FastA in the Genetic Computer GroupŽ .GCG, 1991 package. Each sequence was specific for theselected isoform and did not display any significant simi-larity with the other isoforms or with other sequences indatabases. As several splice variants have been identifiedfor SAPK a , b and g, the probes were chosen in a regionthat do not distinguish between the different splice vari-ants.
Oligodeoxynucleotides were synthesised on a MilliporeExpedite 8909 DNA synthesiser using a standard-cyanoethyl phosphoramidite chemistry. Phosphoramiditeswere purchased from Millipore, Milan, Italy. Forty-five-mers were subsequently purified by reverse-phase chro-
Ž .matography on Cruachem cartridges Glasgow, UK fol-lowing the manufacturer’s instructions. The probes were
X w35 x Ž3 -end-labelled with S dATP specific activity 1100Cirmmol, from New England Nuclear, Frankfurt, Ger-
. Žmany using terminal transferase Amersham Italy, Milan,
Table 1Semi-quantitative analysis of in situ hybridisation signals at different stages of post-natal development
A
P1 P3 P6 P9 P12 P15 P18 P21 Adult
STR 113"4)) 89"4)) 65"3)) 27"2)) 33"4 36"3) 31"4)) 52"4 47"2CTX 162"6) 128"5 119"5)) 64"4)) 76"6)) 93"6)) 115"5)) 160"5 142"4DG 236"7 214"8 193"6 108"6)) 115"10)) 137"5)) 160"9)) 217"10 209"6CA1 240"7 203"10 206"6 128"7)) 130"10)) 176"8)) 207"9 256"10)) 217"7CA3 284"10 234"10)) 257"9)) 157"9)) 164"10)) 212"9)) 238"11)) 308"11 300"7
BP1 P3 P6 P9 P12 P15 P18 P21 Adult
SRR 37"4)) 31"3)) 32"3)) 14"7 7"3 10"2 14"1 9"2 8"1CTX 80"5)) 68"5)) 74"4)) 34"6)) 36"3)) 34"3)) 32"2)) 21"2 15"1DG 82"5)) 84"6)) 88"5)) 56"6)) 55"5)) 47"5) 49"4) 37"3 28"2CA1 92"4)) 81"5)) 103"5)) 67"8)) 58"5)) 50"5)) 41"2)) 29"3 19"2CA3 97"5)) 89"6)) 126"6)) 73"9)) 66"6)) 72"6)) 59"3)) 45"3 35"2MHB 122"7)) 114"8)) 148"11)) 103"19 109"20) 142"11)) 78"8 97"9 74"3
Table shows mean"S.E.M. of relative optical density values measured in film autoradiographs.Panel A displays values measured in slides treated with the SAPK b probe whereas panel B shows values for SAPK g.P: post-natal day; STR: striatum; CTX: cerebral cortex; DG: dentate gyrus; CA1 and CA3: hippocampal fields; MHB: medial habenula.
Ž .) p-0.05 and )) p-0.01 in Dunnett’s test vs. adult values SNK results are not shown in the table but are discussed in the text .

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–6860
.Italy . Radiolabelled oligonucleotides were separated fromŽunincorporated nucleotides using Nensorb columns New
.England Nuclear, Frankfurt, Germany according to manu-facturer’s instructions. Specific activity of the 35Sylabelled probes was routinely between 5=105 and 8=105
cpmrml probe.
2.3. In situ hybridisation histochemistry
Hybridisations were carried out as previously describedw x 33,4 . Sections were exposed to Amersham H yHyperfilms and developed after 18–25 days. Afterwardsthey were dipped in Kodak NTB2 photographic emulsion,exposed for 8 to 12 weeks at y208C, developed for 2 minin Kodak D19 developer, fixed and counterstained withCresyl violet. In order to assess the amount of non-specifichybridisation, adjacent sections were incubated with 100-fold excess unlabelled probe added together with the ra-dioactive oligodeoxynucleotide.
2.4. Analysis of results
Microdensitometric analyses of film autoradiographsŽwere performed using a computer imaging device Imag-
.ing Research, Canada . The specific optical density ofsampled areas was evaluated by subtracting the optical
Ždensity of the non-specific labelling as defined by themean optical density measured in sections treated with
.excess of unlabelled probe . The atlas of Paxinos andw xWatson 23 was used to identify brain structures. Each
experiment contained all experimental groups. Samplesbelonging to the various experimental groups of eachexperiment were treated under the very same conditionsand hybridisation carried out with the same radiolabelledprobe. The statistical analysis of optical density valuesmeasured in brain areas at different post-natal days wasperformed by ANOVA, followed by the Student–New-
Ž .man–Keuls SNK test or the Dunnett’s post-hoc compari-Žson. In Table 1 only the results of Dunnett’s test vs. adult
.levels are shown.
3. Results
3.1. Localisation of SAPK a , b , and g hybridisationsignals in adult rat brain
In order to analyse the expression pattern of SAPKisoforms in rat brain, three 45-mer probes were chosen.
Fig. 2. In situ hybridisation film autoradiographs of parasagittal sectionsof adult rat brain. Each section was hybridised with an oligonucleotide
Ž . Žprobe that specifically recognised SAPK a Panel A or SAPK b Panel. Ž .B or SAPK g Panel C . The hybridisation signals obtained with the
SAPK a and SAPK b probes are widely although heterogeneouslydistributed in the adult brain and appear to be localised in the very samebrain regions. On the other hand, SAPK g specific labelling is restrictedto fewer brain areas. Scale bar: 2.7 mm.
Fig. 3. In situ hybridisation film autoradiographs showing a rostrocaudal series of adult rat brain coronal sections hybridised with oligonucleotide probesŽ . Ž .that recognise SAPK b Panels a, b, c, g, h, i, m, n, o or SAPK g Panels d, e, f, j, k, l, p, q, r . The hybridisation signal obtained with SAPK g probe
Ž . Ž .shows high levels only in the endopiriform nucleus En and in the medial habenula MHb . Panels m and p show sections hybridised with a 100-foldŽ .excess of unlabelled SAPK b or g oligonucleotide, respectively, used to evaluate non-specific binding NSB . Scale bar: 1.6 mm.

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–68 61

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–6862
Each probe was designed to specifically recognise onlyŽ . w xone of the three SAPK a , b or g isoforms 16 , as
Ž .evaluated with FastA see Section 2 . The probes wereunable to discriminate among the splice variants of everyisoform, since they were chosen in conserved regions.
Consecutive coronal and parasagittal slices were cutfrom adult rat brains and hybridised to the three labelledprobes. Non-specific labelling was measured in slices hy-bridised to each probe in the presence of 100-fold excessof the corresponding unlabelled oligonucleotide. The hy-bridisation patterns were compared between each other andwith the signal obtained with a probe which does not
Ždistinguish among the three isoforms pan-SAPK probe,w x.Ref. 3 .
The hybridisation signals obtained with the probes forSAPK a and b were generally localised in the same brainareas, although the signal intensity was slightly differentŽ .Fig. 2, Panels A, B . The hybridisation of the probes forSAPK a and b showed a similar pattern to that detected
w xwith the pan-SAPK probe 3 . On the other hand, SAPK g
labelling appeared markedly different, with only few brainŽ .areas intensely labelled Figs. 2 and 3 . High mRNA levels
were measured exclusively in the endopiriform nucleusŽ . ŽFig. 3, Panels f, j, k, l and in the medial habenula Fig. 3,
.Panels q, r . Intermediate to low staining was detected inCA1 and CA3 fields of the pyramidal layer of the hip-pocampus and in the granule cell layer of the dentate gyrusŽ . Ž .DG Fig. 3, Panels q, r . Intermediate to low levels were
Žalso observed in the piriform cortex Fig. 3, Panels d, e, f,.j, k . SAPK g hybridisation signal appeared weak in the
majority of the brain regions analysed, with low levels inthe cerebral cortex, diencephalon, basal ganglia, and brain-
Ž .stem Fig. 3 .
In order to obtain cellular resolution of the hybridisationsignal, selected sections were processed for emulsion auto-radiography. Given the overlap of hybridisation patternbetween SAPK b and SAPK a , and the selective expres-sion of SAPK b in the CNS, all subsequent experimentswere processed only with the SAPK b and SAPK g
probes.Strong SAPK b labelling was found on the vast major-
ity of neuronal-like cell bodies of the cerebral cortex,hippocampus and diencephalic areas. Grain density overstriatal cellular profiles was substantially lower. The la-belling pattern of the cerebral cortex followed the cellulararchitectural with no apparent differences between layersor cortical sub-areas. Although specific labelling appearedto be located mainly over neuronal cells, the presence ofmRNA in glial cells cannot be ruled out. The identificationof labelled small cell bodies was often difficult in sectionsprocessed for in situ hybridisation. Very intense labellingwas observed in the principal cells of the pyramidal celllayer of the hippocampal fields CA1 and CA3 and thesubiculum, as well as in the granule cells of the dentate
Ž .gyrus Fig. 4 . Marked staining was also detected innumerous interneuron-like cells, in particular in the stra-tum oriens of CA1 and CA3 fields. A large number ofcellular profiles in the polymorphic area of the dentate
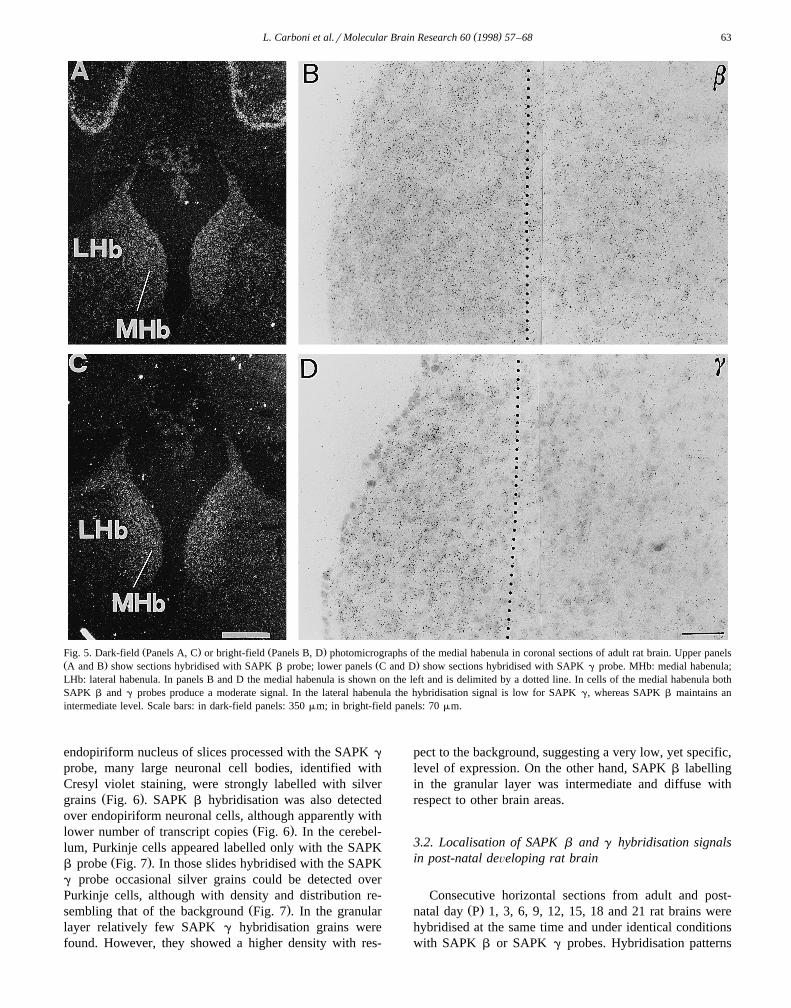
Ž .gyrus were also intensely labelled Fig. 4 . In the habenula,both SAPK b and SAPK g hybridisations produced amoderate labelling of all the medium-small sized cells in
Ž .the medial areas Fig. 5 . On the other hand, in the lateralŽ .part SAPK g signal appeared consistently lower Fig. 5 .
Ependimal cells of the third ventricle clearly showed nohybridisation signal for SAPK g, whereas several silvergrains could be detected with the SAPK b probe. In the
Fig. 4. Dark-field photomicrograph of hippocampus in a coronal section of adult rat brain hybridised with SAPK b probe. Pyramidal cells in CA1 andŽ .CA3 fields and the granule cells in the dentate gyrus DG show a high hybridisation signal. 3V: third ventricle. Scale bar: 500 mm.
( )L. Carboni et al.rMolecular Brain Research 60 1998 57–68 63
Ž . Ž .Fig. 5. Dark-field Panels A, C or bright-field Panels B, D photomicrographs of the medial habenula in coronal sections of adult rat brain. Upper panelsŽ . Ž .A and B show sections hybridised with SAPK b probe; lower panels C and D show sections hybridised with SAPK g probe. MHb: medial habenula;LHb: lateral habenula. In panels B and D the medial habenula is shown on the left and is delimited by a dotted line. In cells of the medial habenula bothSAPK b and g probes produce a moderate signal. In the lateral habenula the hybridisation signal is low for SAPK g, whereas SAPK b maintains anintermediate level. Scale bars: in dark-field panels: 350 mm; in bright-field panels: 70 mm.
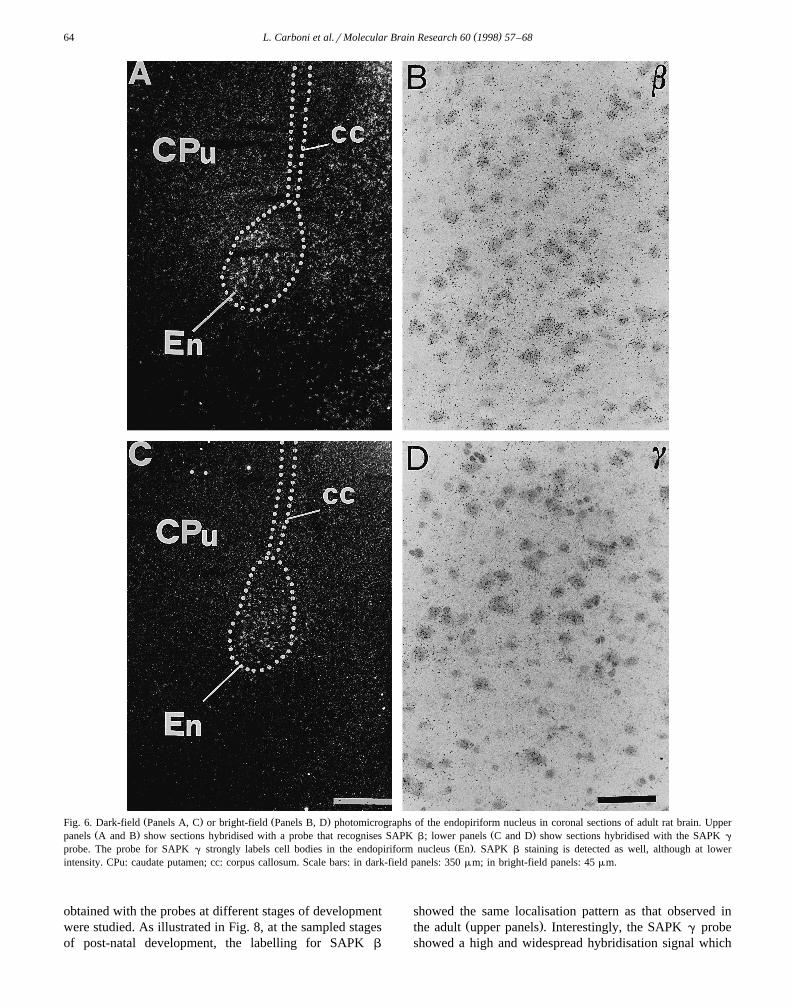
endopiriform nucleus of slices processed with the SAPK g
probe, many large neuronal cell bodies, identified withCresyl violet staining, were strongly labelled with silver
Ž .grains Fig. 6 . SAPK b hybridisation was also detectedover endopiriform neuronal cells, although apparently with
Ž .lower number of transcript copies Fig. 6 . In the cerebel-lum, Purkinje cells appeared labelled only with the SAPK
Ž .b probe Fig. 7 . In those slides hybridised with the SAPKg probe occasional silver grains could be detected overPurkinje cells, although with density and distribution re-
Ž .sembling that of the background Fig. 7 . In the granularlayer relatively few SAPK g hybridisation grains werefound. However, they showed a higher density with res-
pect to the background, suggesting a very low, yet specific,level of expression. On the other hand, SAPK b labellingin the granular layer was intermediate and diffuse withrespect to other brain areas.
3.2. Localisation of SAPK b and g hybridisation signalsin post-natal deÕeloping rat brain
Consecutive horizontal sections from adult and post-Ž .natal day P 1, 3, 6, 9, 12, 15, 18 and 21 rat brains were
hybridised at the same time and under identical conditionswith SAPK b or SAPK g probes. Hybridisation patterns
( )L. Carboni et al.rMolecular Brain Research 60 1998 57–6864
Ž . Ž .Fig. 6. Dark-field Panels A, C or bright-field Panels B, D photomicrographs of the endopiriform nucleus in coronal sections of adult rat brain. UpperŽ . Ž .panels A and B show sections hybridised with a probe that recognises SAPK b; lower panels C and D show sections hybridised with the SAPK g
Ž .probe. The probe for SAPK g strongly labels cell bodies in the endopiriform nucleus En . SAPK b staining is detected as well, although at lowerintensity. CPu: caudate putamen; cc: corpus callosum. Scale bars: in dark-field panels: 350 mm; in bright-field panels: 45 mm.
obtained with the probes at different stages of developmentwere studied. As illustrated in Fig. 8, at the sampled stagesof post-natal development, the labelling for SAPK b
showed the same localisation pattern as that observed inŽ .the adult upper panels . Interestingly, the SAPK g probe
showed a high and widespread hybridisation signal which

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–68 65
Fig. 7. Bright-field photomicrographs of coronal sections of the adult rat cerebellar cortex. Panel A shows a section hybridised with the oligonucleotideŽ .probe that recognises SAPK b, whereas the section depicted in panel B was hybridised with the SAPK g probe. Purkinje cells P, pointed by arrows are
Ž .labelled only by the probe for SAPK b. In the granule cell layer g SAPK b shows an intermediate level of staining and SAPK g displays a low signal.ml: molecular layer. Scale bar: 50 mm.
decreased substantially during development to reach theŽhighly discrete staining observed in the adult Fig. 8, lower
.panels .
3.3. Semi-quantitatiÕe analysis of SAPK b and g labellingduring post-natal deÕelopment
Hybridisation signals of SAPK b and g were measuredin specific brain regions at different stages of post-nataldevelopment. Sampled regions were selected according tofeasibility of sampling and presence of high hybridisationsignal. The microdensitometric analysis of film autoradio-grams was performed by means of a computer-based im-
age analysis system. The optical densities were comparedin the sampled stages of development in order to evaluatewhether mRNA levels were modulated.
3.3.1. SAPK b
In the striatum, optical density values were significantlyŽhigher at P1 as compared with later days p-0.01 vs. any
.other group in the SNK test . The signal decreased slowlyat P3 and P6, maintaining levels that were both different to
Ž .any other group p-0.01, SNK test . Optical densityvalues reached a minimum at P9 and remained essentially
Žunmodified to match those measured in the adult Table 1,.Panel A . SAPK b labelling in the cerebral cortex and the

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–6866
Fig. 8. In situ hybridisation film autoradiographs of rat brain horizontal sections at different post-natal developmental stages hybridised withŽ . Ž .oligonucleotide probes which recognise SAPK b upper panels or SAPK g lower panels . SAPK b maintains throughout post-natal development the
Ž .same localisation observed in the adult brain. SAPK g hybridisation signal is high and widely distributed in the rat brain at early post-natal days P .During development signal intensity decreases substantially in the majority of brain areas remaining relatively high only in the endopiriform nucleus and in
Ž .the medial habenula MHb . A: adult. Scale bar: 2.7 mm.
hippocampal fields CA1, CA3, and DG showed a differenttemporal pattern, with a degree of hybridisation in theearly stages of post-natal development comparable to thatmeasured in the adult. Starting from P3, optical densityvalues started to decrease, reaching a minimum at P9 andP12. Signals measured at P9 and P12 were not signifi-cantly different, but were different from any other groupŽ .p-0.01, SNK test . At P15, the labelling increased
Ž .again, reaching adult levels at P21 Table 1, Panel A .
3.3.2. SAPK g
Statistically significant differences in optical densityvalues for SAPK g were observed during development. Inthe hippocampus, the pyramidal cell layer in CA3 andCA1 displayed high hybridisation signals at P1 and P3 that
Žslightly increased at P6 p-0.01 vs. any other group,.SNK test . Starting from P9, mRNA levels progressively
Ždecreased to reach those observed in the adult Table 1,.Panel B . In caudate-putamen, high mRNA levels were
also detected at P1 and remained high until P6. After-wards, optical density values steeply decreased. At P9,signals appeared as low as those detected in the adultŽ .Table 1, Panel B . The same temporal trend was observedin the cerebral cortex and DG, but optical density valuesfrom P9 to P18 were slightly higher than in the adultŽ .Table 1, Panel B . On the other hand, medial habenuladisplayed a high hybridisation signal from P1 and re-
mained high throughout the post-natal development. Aslight reduction was observed at P18 and P21, which
Ž .exhibited the same levels as in adult Table 1, Panel B .
4. Discussion
We studied by means of in situ hybridisation withspecific oligonucleotide probes the localisation of themRNA transcripts of the three SAPK genes in the adult ratbrain. In addition, we investigated the mRNA expressionof SAPK b and g during post-natal development. Ourresults show that SAPK a and b are the main isoformsexpressed in the adult brain. The probes that recognisethese isoforms marked the same brain regions and repro-duced the same hybridisation pattern observed with aprobe that does not distinguish among the different SAPKvariants. On the other hand, the SAPK g probe revealed aselective localisation in the adult brain, with high levelsrestricted to the endopiriform nucleus and the medialhabenula. In these regions, the presence of the otherisoforms could be detected as well. Hybridisation signalsfor SAPK a , b and g were all present in the hippocam-pus, with SAPK g showing the lowest signal.
We detected low labelling for all of the three isoformsin striatum. Nevertheless, SAPK protein has been detectedin this region, and its activity can be stimulated by gluta-
w xmate 25 .

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–68 67
In newborn rats, SAPK b appeared expressed in thesame brain areas as in adults. A generalised reduction inthe levels of expression was found between P9 and P12.Values from P15 to the adult returned gradually to thosemeasured in the early days after birth. On the other hand,SAPK g showed a completely different pattern, with highand diffuse expression in newborn rats. Labelling for thisisoform progressively declined during post-natal develop-ment, to become restricted to few areas in the adult rat.Interestingly, these findings are consistent with a previousobservation that JNK1 mRNA could be detected withNorthern blot analysis in human foetal but not adult brainw x7 , suggesting a specific role being played by this kinaseduring development.
A function for the stress-activated MAPKs during de-velopment is supported by studies in Drosophila. Kinasesbelonging to the fruitfly homologue of SAPK pathwayŽ .DJNKrBsk and Hep exert an essential role in embryonic
w xdevelopment 12,27 . This function is likely maintained inmammals, as targeted disruption of the MKK4 gene, a
w xMAPKK for SAPK, causes embryonic death in mice 30 .SAPK brJNK3 protein is highly expressed in mouse
w xembryonic brain 18 , and SAPK arJNK2 is localised inw xneuroepithelium of chick embryos 14 . In this study, we
report a regional regulation of SAPK b and g mRNAlevels during different post-natal developmental stages,thus suggesting that this cascade may exert a role in theformation of synaptic circuitry.
The mRNA levels of both SAPK b and g varied duringthe stages of post-natal development. SAP kinase activityis usually reported to be regulated through the control ofits phosphorylated state exerted by MAPKK, instead ofthrough the modulation of mRNA transcription or proteintranslation. Our data suggest that this level of control maybe important as well, at least in the CNS and in agreementwith Ferrer et al., who report a variation in SAPKimmunoreactivity in rat brain after excitotoxic stimuliw x9,10 .
The different distribution pattern of SAPK isoforms inthe adult rat brain suggests the existence of separate,non-redundant, functions being performed by the productsof the three SAPK genes. A large body of evidencesupports the hypothesis that different isoforms mediatedifferent outputs. Indeed, proteins deriving from each geneŽ .Jnk1, 2 or 3 exhibit diverse selectivity and affinity forthe transcription factors whose activity they modulatew x1,6,13,15,26 . It has been suggested that, at physiologicalconcentrations in cells, only one of the isoforms is able tophosphorylate a specific transcription factor, therefore in-
w xducing a distinct gene expression response 13,15 . Sluss etal. reported that JNK1, but not JNK2, complements adefect in the expression of the HOG MAPK in Saccar-omyces cereÕisiae, demonstrating that the isoforms have
w xdifferent properties 26 . Distinct SAPK forms have beensuggested to be responsible for the activation of protective
w xor apoptotic responses in small cell lung cancer cells 2
and only a p46 SAPK mediates the c-myc-dependent apop-w xtosis in rat fibroblasts 34 . Only one SAPK isoform,
which may correspond to SAPK brJNK3, or to a stillunknown subtype, has been involved in the induction of
w xstaurosporine-mediated differentiation in PC12 cells 33 .Furthermore, the generation of mice with targeted disrup-tion of individual Jnk genes has provided evidence forselectivity of function among isoforms, in particular in thebrain. Mice lacking the expression of the Jnk3 genedisplayed reduced sensitivity to kainic acid-induced toxic-
Ž .ity, which was not shared by the Jnk1 yry orŽ . w xJnk2 yry animals 31 .
In conclusion, this study shows that SAPK isoforms aredifferently localised in specific brain regions. The selectivelocalisation supports the hypothesis that specific functionsare performed by each isoform. This study also reports adifferent modulation of mRNA levels of SAPK isoforms inthe developing rat brain. These findings suggest that SAPKisoforms may be differently involved in signal transductionduring post-natal development.
Acknowledgements
The authors wish to thank Dr. Valerie Matarese and Dr.Francesco Belardetti for critical review of the manuscriptand helpful suggestions.
References
w x1 V. Adler, A. Schaffer, J. Kim, L. Dolan, Z. Ronai, UV irradiationand heat shock mediate JNK activation via alternate pathways, J.
Ž .Biol. Chem. 270 1995 26071–26077.w x2 L. Butterfield, B. Storey, L. Mas, L.E. Heasley, c-Jun NH -terminal2
kinase regulation of the apoptotic response of small cell lung cancerŽ .cells to ultraviolet radiation, J. Biol. Chem. 272 1997 10110–10116.
w x3 R. Carletti, S. Tacconi, E. Bettini, F. Ferraguti, Stress activatedprotein kinases, a novel family of mitogen-activated protein kinases,are heterogeneously expressed in the adult rat brain and differen-tially distributed from extracellular-signal-regulated protein kinases,
Ž .Neuroscience 69 1995 1103–1110.w x4 L. Carboni, S. Tacconi, R. Carletti, E. Bettini, F. Ferraguti, Localiza-
tion of the messenger RNA for the c-Jun NH -terminal kinase2
kinase in the adult and developing rat brain: an in situ hybridizationŽ .study, Neuroscience 80 1997 147–160.
w x5 O.A. Coso, M. Chiariello, G. Kalinec, J.M. Kyriakis, J. Woodgett,J.S. Gutkind, Transforming G protein-coupled receptors potently
Ž . Ž .activate JNK SAPK , J. Biol. Chem. 270 1995 5620–5624.w x6 T. Dai, E. Rubie, C.C. Franklin, A. Kraft, D.A.F. Gillespie, J.
Avruch, J.M. Kyriakis, J.R. Woodgett, Stress-activated protein ki-nases bind directly to the d domain of c-Jun in resting cells:
Ž .implications for repression of c-Jun function, Oncogene 10 1995849–855.
w x7 B. Derijard, B. Hibi, I.-H. Wu, T. Barrett, B. Su, T. Deng, M. Karin,R.J. Davis, JNK1: a protein kinase stimulated by UV light andHa-Ras that binds and phosphorylates the c-Jun activation domain,
Ž .Cell 76 1994 1025–1037.w x8 G.R. Fanger, P. Gerwins, C. Widmann, M.B. Jarpe, G.L. Johnson,
MEKKs, GCKs, MLKs, PAKs, TAKs, and Tpls: upstream regula-

( )L. Carboni et al.rMolecular Brain Research 60 1998 57–6868
tors of the c-Jun amino-terminal kinases?, Curr. Opin. Genet. Dev. 7Ž .1997 67–74.
w x9 I. Ferrer, A.M. Planas, E. Pozas, Radiation-induced apoptosis indeveloping rats and kainic acid-induced excitotoxicity in adult ratsare associated with distinctive morphological and biochemical c-
Ž . Ž .JunrAP-1 N expression, Neuroscience 80 1997 449–458.w x10 I. Ferrer, E. Pozas, J. Ballabriga, A.M. Planas, Strong c-JunrAP-1
immunoreactivity is restricted to apoptotic cells following intracere-Ž .bral ibotenic acid injection in developing rats, Neurosci. Res. 1997
21–31.w x11 I. Foltz, J.W. Schrader, Activation of the stress-activated protein
kinases by multiple hematopoietic growth factors with the exceptionŽ .of interleukin-4, Blood 89 1997 3092–3096.
w x12 B. Glise, H. Bourbon, S. Noselli, Hemipterous encodes a novelDrosophila MAP kinase kinase, required for epithelial cell sheet
Ž .movement, Cell 83 1995 451–461.w x13 S. Gupta, T. Barrett, A.J. Whitmarsh, J. Cavanagh, H.K. Sluss, B.
Derijard, R.J. Davis, Selective interaction of JNK protein kinaseŽ .isoforms with transcription factors, EMBO J. 15 1996 1670–2770.
w x14 T. Ishikawa, Y. Nakada-Moriya, C. Ando, N. Tanda, S. Nishida, Y.Minatogawa, T. Nohnon, Expression of the JNK2-a1 gene in thedeveloping chick brain, Biochem. Biophys. Res. Commun. 234Ž .1997 489–492.
w x15 T. Kallunki, B. Su, I. Tsigelny, H.K. Sluss, B. Derijard, G. Moore,R. Davis, M. Karin, JNK2 contains a specificity-determining regionresponsible for efficient c-Jun binding and phosphorylation, Genes
Ž .Dev. 8 1994 2996–3007.w x16 J.M. Kyriakis, P. Banerjee, E. Nikolakaki, T. Dai, E.A. Rubie, M.F.
Ahmad, J. Avruch, J.R. Woodgett, The stress-activated proteinŽ .kinase subfamily of c-Jun-kinases, Nature 369 1994 156–160.
w x17 J.M. Kyriakis, J. Avruch, Sounding the alarm: protein kinase cas-cades activated by stress and inflammation, J. Biol. Chem. 271Ž .1996 24313–24316.
w x18 J.H. Martin, A.A. Mohit, C.A. Miller, Developmental expression inthe mouse nervous system of the p493F12 SAP kinase, Mol. Brain
Ž .Res. 35 1996 47–57.w x19 H. Mitsui, N. Takuwa, K. Kurokawa, J.H. Exton, Y. Takuwa,
Dependence of activated Ga12-induced G to S phase cell cycle1
progression on both Rasrmitogen-activated protein kinase andRasrRac1rJun N-terminal kinase cascades in NIH3T3 fibroblasts,
Ž .J. Biol. Chem. 272 1997 4904–4910.w x 3F1220 A.A. Mohit, J.H. Martin, C.A. Miller, p49 kinase: a novel MAP
kinase expressed in a subset of neurons in the human nervousŽ .system, Neuron 14 1995 67–78.
w x21 H. Nishima, K.D. Fischer, L. Radvanyl, A. Shahinian, R. Hakem,E.A. Rubie, A. Bernstein, T.W. Mak, J.R. Woodgett, J.M. Pen-
ninger, Stress-signalling kinase Sek-1 protects thymocytes fromŽ .apoptosis mediated by CD95 and CD3, Nature 385 1997 350–353.
w x22 A. Paul, S. Wilson, C.M. Belham, C.J.M. Robinson, P.H. Scott,G.W. Gould, R. Plevin, Stress-activated protein kinases: activation,
Ž .regulation and function, Cell. Signal. 9 1997 403–410.w x23 G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates,
Academic Press, Orlando, FL, 1986.w x24 M.J. Robinson, M. Cobb, Mitogen-activated protein kinase path-
Ž .ways, Curr. Opin. Cell Biol. 9 1997 180–186.w x25 M.A. Schwarzschild, R. Cole, S.E. Hyman, Glutamate, but not
dopamine, stimulates stress-activated protein kinase and AP-1-medi-Ž .ated transcription in striatal neurons, J. Neurosci. 17 1997 3455–
3466.w x26 H.K. Sluss, T. Barrett, B. Derijard, R. Davis, Signal transduction by
tumor necrosis factor mediated by JNK protein kinases, Mol. Cell.Ž .Biol. 14 1994 8376–8384.
w x27 H.K. Sluss, Z. Han, T. Barrett, R.J. Davis, Y.T. Ip, A JNK signaltransduction pathway that mediates morphogenesis and an immune
Ž .response in Drosophila, Genes Dev. 10 1996 2745–2758.w x28 C. Tournier, A.J. Whitmarsh, J. Cavanagh, T. Barrett, R. Davis,
Mitogen-activated protein kinase kinase 7 is an activator of the c-JunŽ .NH -terminal kinase, Proc. Natl. Acad. Sci. U.S.A. 94 19972
7337–7342.w x29 X. Xu, J. Raber, D. Yang, B. Su, L. Mucke, Dynamic regulation of
c-Jun N-terminal kinase activity in mouse brain by environmentalŽ .stimuli, Proc. Natl. Acad. Sci. U.S.A. 94 1997 12655–12660.
w x30 D. Yang, C. Tournier, M. Wysk, H.-T. Lu, J. Xu, R.J. Davis, R.A.Flavell, Targeted disruption of the MKK4 gene causes embryonicdeath, inhibition of c-Jun NH -terminal kinase activation, and de-2
fects in AP-1 transcriptional activity, Proc. Natl. Acad. Sci. U.S.A.Ž .94 1997 3004–3009.
w x31 D.D. Yang, C.-Y. Kuan, A.J. Whitmarsh, M. Rincon, T.S. Zheng,R.J. Davis, P. Rakic, R.A. Flavell, Absence of excitotoxicity-in-duced apoptosis in the hippocampus of mice lacking the Jnk3 gene,
Ž .Nature 389 1997 865–870.w x32 X. Yang, R. Khosravi-Far, H.Y. Chang, D. Baltimore, Daxx, a novel
Ž .Fas-binding protein that activates JNK and apoptosis, Cell 89 19971067–1076.
w x33 R. Yao, M. Yoshihara, H. Osada, Specific activation of a c-JunNH -terminal kinase isoform and induction of neurite outgrowth in2
Ž .PC-12 cells by staurosporine, J. Biol. Chem. 272 1997 18261–18266.
w x34 K. Yu, C.P. Ravera, Y.-N.P. Chen, G. McMahon, Regulation ofc-myc-dependent apoptosis by p53, c-Jun N-terminalkinasesrstress-activated protein kinases, and Mdm-2, Cell Growth
Ž .Diff. 8 1997 731–742.




















