
Blood-based biomarkers for the optimization of anti-angiogenic therapies
Upload
vanderbiltCategory
view
5download
0
Differential Expression of Adenosine Receptors in HumanEndothelial Cells
Role of A2B Receptors in Angiogenic Factor Regulation
Igor Feoktistov, Anna E. Goldstein, Sergey Ryzhov, Dewan Zeng, Luiz Belardinelli,Tatyana Voyno-Yasenetskaya, Italo Biaggioni
Abstract—Adenosine has been reported to stimulate or inhibit the release of angiogenic factors depending on the cell typeexamined. To test the hypothesis that differential expression of adenosine receptor subtypes contributes to endothelialcell heterogeneity, we studied microvascular (HMEC-1) and umbilical vein (HUVEC) human endothelial cells. Basedon mRNA level and stimulation of adenylate cyclase, we found that HUVECs preferentially express A2A adenosinereceptors and HMEC-1 preferentially express A2B receptors. Neither cells expressed A1 or A3 receptors. The nonselectiveadenosine agonist 5�-N-ethylcarboxamidoadenosine (NECA) increased expression of interleukin-8 (IL-8), basicfibroblast growth factor (bFGF), and vascular endothelial growth factor (VEGF) in HMEC-1, but had no effect inHUVECs. In contrast, the selective A2A agonist 2-p-(2-carboxyethyl)phenylethylamino-NECA (CGS 21680) had noeffect on expression of these angiogenic factors. Cotransfection of each type of adenosine receptors with a luciferasereporter in HMEC-1 showed that A2B receptors, but not A1, A2A, or A3, activated IL-8 and VEGF promoters. Theseeffects were mimicked by constitutively active �Gq, �G12, and �G13, but not �Gs or �Gi1-3. Furthermore, stimulation ofphospholipase C indicated coupling of A2B receptors to Gq proteins in HMEC-1. Thus, differential expression ofadenosine receptor subtypes contributes to functional heterogeneity of human endothelial cells. A2B receptors,predominantly expressed in human microvascular cells, modulate expression of angiogenic factors via coupling to Gq,and possibly via G12/13. (Circ Res. 2002;90:531-538.)
Key Words: adenosine receptors � vascular endothelium � angiogenesis � vascular endothelial growth factor� interleukin-8
The purine nucleoside adenosine is an intermediate catab-olite of adenine nucleotides. Adenosine serves as an
autocoid in situations when oxygen supply is decreased orenergy consumption is increased. Under these conditions,adenosine is released into the extracellular space and signalsto restore the balance between local energy requirements andenergy supply. Endothelial cells interact with adenosinemechanisms in many different ways. Endothelial cells areknown to have a very active adenosine metabolism, charac-terized by a large capacity for uptake and release of thenucleoside,1,2 and can be an important source of adenosinereleased during ischemia.3 Conversely, adenosine may mod-ulate endothelial function via activation of cell membranereceptors. The precise nature of the interaction betweenadenosine receptor subtypes and endothelial cells and theirrole in the regulation of endothelial function is not completelyunderstood.
Adenosine receptors belong to the G protein–coupled 7transmembrane superfamily of cell surface receptors andinclude A1, A2A, A2B, and A3 subtypes. Endothelial cells areknown to express adenosine receptors, but there are conflict-ing reports on the presence and the role of specific adenosinereceptor subtypes. For example, human umbilical vein endo-thelial cells (HUVECs) were reported to express either A1,4
A2A,5,6 A2B,7 or A36 adenosine receptors, depending on the
functional end-point studied and pharmacological tools used.Coexpression of more than one adenosine receptor subtypehas been reported also in endothelial cells8,9; it is not clear,however, if and how coexpressed receptors interact. Further-more, endothelial cells from different blood vessels areheterogenous, and it is possible that diverse endothelial cellsshow differential expression of adenosine receptor subtypes.
The functional role of adenosine receptors in endothelialcells also remains unclear. Adenosine-induced vasodilation
Original received December 12, 2001; revision received January 22, 2002; accepted January 22, 2002.From the Divisions of Cardiology (I.F., A.E.G.) and Clinical Pharmacology (S.R., I.B.), Departments of Medicine (I.F., A.E.G., S.R., I.B.) and
Pharmacology (I.F., I.B.), Vanderbilt University, Nashville, Tenn; CV Therapeutics (D.Z., L.B.), Palo Alto, Calif; and the Department of Pharmacology(T.V.-Y.), University of Illinois, Chicago, Ill.
This work was supported in part by a research grant from CV Therapeutics, Inc, Palo Alto, Calif. Dewan Zeng and Luiz Belardinelli, coauthors of thisarticle, are employees of CV Therapeutics.
Correspondence to Igor Feoktistov, PhD, 315 PRB, Vanderbilt University, Nashville, TN 37232-6300. E-mail [email protected]© 2002 American Heart Association, Inc.
Circulation Research is available at http://www.circresaha.org DOI: 10.1161/01.RES.0000012203.21416.14
531
Molecular Medicine

has been attributed, at least in part, to activation of endothe-lial-derived factors in some vascular beds.10,11 Adenosine isthought to promote endothelial barrier function and to main-tain vascular integrity.4,7 Adenosine has also been found tostimulate the proliferation of capillary endothelial cells12–14
and to promote neovascularization.15
Regulation of neovascularization depends on a delicatebalance of proangiogenic and antiangiogenic factors, pro-teases and their inhibitors, and adhesion molecules. Vascularendothelial growth factor (VEGF), interleukin-8 (IL-8), andbasic fibroblast growth factor (bFGF) occupy a particularplace among positive modulators of angiogenesis due to theirpotency and the reported essential role each of them plays inpromoting angiogenesis.16–18 Extracellular adenosine, gener-ated in high concentrations in hypoxic tissues, has been foundto modulate secretion of VEGF.12–14,19–22 However, depend-ing on the cell type studied, adenosine has been shown toeither stimulate or inhibit secretion of proangiogenic factors.Recent studies demonstrated that adenosine stimulates VEGFproduction in human retinal endothelial cells,13,14,20 and thiseffect was attributed to activation of A2B adenosine recep-tors.13,14 In contrast, adenosine inhibited hypoxia-inducedVEGF production in rat pheochromocytoma P12 cells, andthis effect was attributed to activation of A2A adenosinereceptors.21,22
In this study, we characterized the adenosine receptorsubtypes present in 2 different human endothelial cells types:one obtained from large venous conduit vessels, HUVECs,and the other derived from skin microvasculature, HMEC-1.Our purpose was to test the hypothesis that diverse endothe-lial cells would show differential expression of adenosinereceptor subtypes and that this will translate into distinctphenotypes in regards to expression of angiogenic factors.
Materials and MethodsCell CultureHUVECs and HMEC-1 were kindly provided by Dr D.E. Vaughan(Vanderbilt University, Nashville, Tenn). Chinese hamster ovaryCHO-K1 cells were obtained from the American Type CultureCollection (Manassas, Va).
Chemicals5�-N-ethylcarboxamidoadenosine (NECA) and 2-p-(2-carboxyethyl)phenylethylamino-NECA (CGS21680) were pur-chased from Research Biochemicals, Inc. 3-Isobutyl-8-pyrrolidinoxanthine (IPDX) was synthesized as previouslydescribed.23
Gene Expression AssayTotal RNA was isolated using Quiagen RNeasy Mini Kit. Expressionof angiogenic factors was evaluated using gene expression arrays(Super Array, Inc). Human adenosine receptors gene expressionarray was custom designed by Super Array, Inc. The assay wasperformed according to manufacturer’s instructions. In brief, gene-specific [32P]-labeled cDNA probes were generated from 5 to 10 �gof total RNA using gene-specific set of primers for reverse transcrip-tion. The cDNA probes were then hybridized with gene-specificcDNA fragments spotted on nylon membrane. The relative expres-sion level of each gene was analyzed using a PhosphorImager andImage Quant software (Molecular Dynamics).
Reverse Transcription–Polymerase Chain ReactionTwo micrograms of total RNA from each sample were subjected toreverse transcription followed by 40 cycles of amplification usingPromega Access reverse transcription–polymerase chain reaction(RT-PCR) system in accordance with manufacturer’s instructions.PCR primers were synthesized to target mRNA sequence represent-ing parts of different exons in human A1, A2A, and A2B adenosinereceptors genes to yield PCR products of 500 to 800 base pairs. Thefollowing primer pairs were used for A1: 5�-TCTGGGCGGTGAAG-GTGAAC-3� (sense) and 5�-AGTTGCCGTGCGTGAGGAAG-3�(antisense); A2A: 5�-TGCTTCGTCCTGGTCCTCAC-3� (sense) and5�-GCTCTCCGTCACTGCCAT-3� (antisense); and A2B: 5�-CCCT-TTGCCATCACCATCAG-3� (sense) and 5�-CCTGACCATTC-CCACTCTTGA-3� (antisense). PCR primers for human A3 adeno-sine receptor were used as described by Mitchell et al.24 PurifiedPCR fragments were analyzed by restrictive digestion.
Real-time RT-PCR was performed on ABI Prism SequenceDetection System 5700 (PE Applied Biosystems) in accordance withmanufacturer’s recommendations.
Transfections and Luciferase Reporter AssayHMEC-1 and CHO-K1 cells were transfected using Fugene 6transfection reagent (Roche) with cDNA described in the Resultssection and luciferase reporters at a ratio of 10:1. The same ratio,10:1, was used for experimental firefly luciferase reporter:controlRenilla luciferase reporter combination. VEGF promoter–drivenluciferase reporter, a firefly luciferase reporter plasmid, comprising5� flanking �1005 to �379 base pairs of the human VEGF gene25
was kindly provided by Dr G.L. Semenza, Johns Hopkins Hospital,Baltimore, Md. IL-8 promoter-driven luciferase reporter �133-luc, afirefly luciferase reporter plasmid, comprising 5� flanking �133 to�44 base pairs of the human IL-8 gene26 was generously gifted byDr Naofumi Mukaida, Kanazawa University, Ishikawa, Japan. ThecDNAs encoding the human adenosine receptors in the pRc/CMVexpression vector (Invitrogen) were a generous gift from Drs P.R.Schofield and A. Townsend-Nicholson (Garvan Institute of MedicalResearch, Australia). Plasmids �sRCpcDNAI, �i1QLpcDNAI,�i2QLpcDNAI, �i3QLpcDNAI, �qRCpcDNAI, �12QLpcDNAI neo,and �13QLpcDNA3, encoding constitutively active � subunits ofG-proteins, were constructed as described previously.27–30 Reporteractivity was measured 30 hours after transfection using a Dual-Luciferase Reporter Assay System (Promega). Firefly luciferasereporter activities were normalized against Renilla luciferase activ-ities from the coexpressed pRL-TK (Promega) and expressed asrelative luciferase activities over basal (set as 1).
Measurement of Second Messengers andAngiogenic FactorscAMP concentrations and formation of [3H]inositol phosphates weredetermined as previously described.31 IL-8 and VEGF concentra-tions were measured using ELISA kits (R&D Systems) as previouslydescribed.31
An expanded Materials and Methods section can be found in theonline data supplement available at http://www.circresaha.org.
ResultsExpression of Adenosine Receptor mRNAGene expression array results indicate that HUVECs prefer-entially express mRNA encoding A2A receptors (Figure 1A).As a percentage of �-actin expression, HUVECs expressedlevels of A2A and A2B receptors of 3.2�0.09% and0.38�0.04%, respectively (Figure 1B). Therefore, the ratio ofexpression of A2A:A2B was approximately 10 to 1. In contrast,HMEC-1 preferentially express mRNA encoding A2B recep-tors (Figure 1A). HMEC-1 expressed mRNA levels for A2B
and A2A receptors of 0.55�0.07% and 0.13�0.01% of �-actinmRNA, respectively (Figure 1B). Therefore, the ratio of
532 Circulation Research March 22, 2002
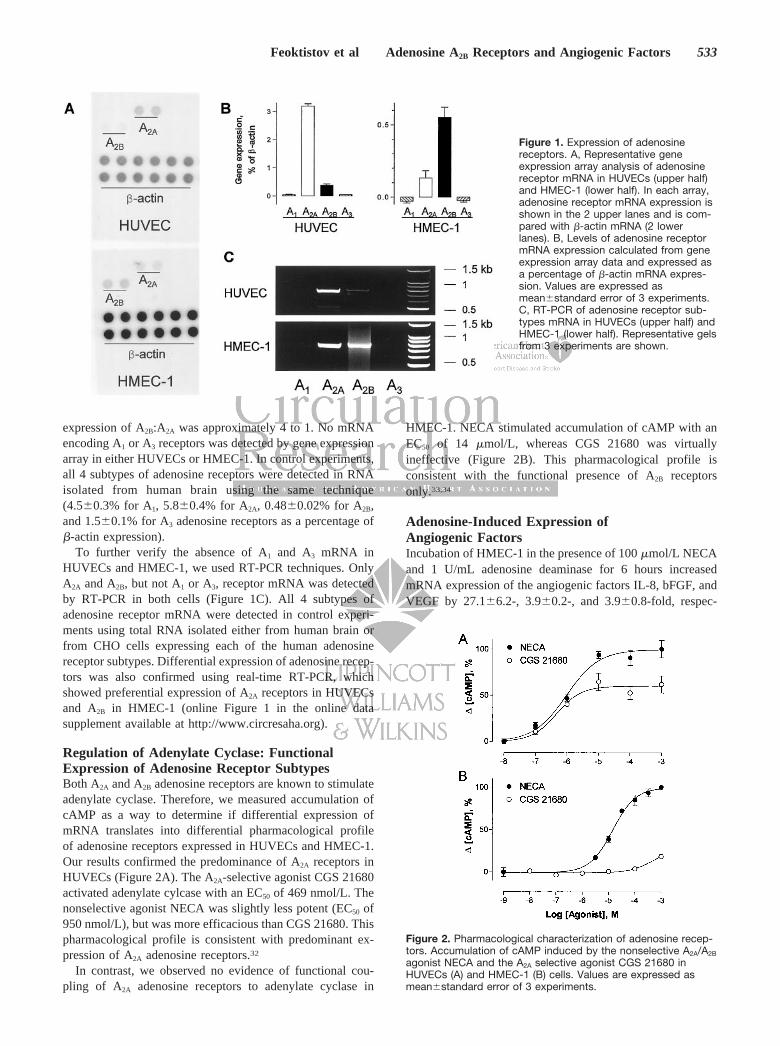
expression of A2B:A2A was approximately 4 to 1. No mRNAencoding A1 or A3 receptors was detected by gene expressionarray in either HUVECs or HMEC-1. In control experiments,all 4 subtypes of adenosine receptors were detected in RNAisolated from human brain using the same technique(4.5�0.3% for A1, 5.8�0.4% for A2A, 0.48�0.02% for A2B,and 1.5�0.1% for A3 adenosine receptors as a percentage of�-actin expression).
To further verify the absence of A1 and A3 mRNA inHUVECs and HMEC-1, we used RT-PCR techniques. OnlyA2A and A2B, but not A1 or A3, receptor mRNA was detectedby RT-PCR in both cells (Figure 1C). All 4 subtypes ofadenosine receptor mRNA were detected in control experi-ments using total RNA isolated either from human brain orfrom CHO cells expressing each of the human adenosinereceptor subtypes. Differential expression of adenosine recep-tors was also confirmed using real-time RT-PCR, whichshowed preferential expression of A2A receptors in HUVECsand A2B in HMEC-1 (online Figure 1 in the online datasupplement available at http://www.circresaha.org).
Regulation of Adenylate Cyclase: FunctionalExpression of Adenosine Receptor SubtypesBoth A2A and A2B adenosine receptors are known to stimulateadenylate cyclase. Therefore, we measured accumulation ofcAMP as a way to determine if differential expression ofmRNA translates into differential pharmacological profileof adenosine receptors expressed in HUVECs and HMEC-1.Our results confirmed the predominance of A2A receptors inHUVECs (Figure 2A). The A2A-selective agonist CGS 21680activated adenylate cylcase with an EC50 of 469 nmol/L. Thenonselective agonist NECA was slightly less potent (EC50 of950 nmol/L), but was more efficacious than CGS 21680. Thispharmacological profile is consistent with predominant ex-pression of A2A adenosine receptors.32
In contrast, we observed no evidence of functional cou-pling of A2A adenosine receptors to adenylate cyclase in
HMEC-1. NECA stimulated accumulation of cAMP with anEC50 of 14 �mol/L, whereas CGS 21680 was virtuallyineffective (Figure 2B). This pharmacological profile isconsistent with the functional presence of A2B receptorsonly.33,34
Adenosine-Induced Expression ofAngiogenic FactorsIncubation of HMEC-1 in the presence of 100 �mol/L NECAand 1 U/mL adenosine deaminase for 6 hours increasedmRNA expression of the angiogenic factors IL-8, bFGF, andVEGF by 27.1�6.2-, 3.9�0.2-, and 3.9�0.8-fold, respec-
Figure 1. Expression of adenosinereceptors. A, Representative geneexpression array analysis of adenosinereceptor mRNA in HUVECs (upper half)and HMEC-1 (lower half). In each array,adenosine receptor mRNA expression isshown in the 2 upper lanes and is com-pared with �-actin mRNA (2 lowerlanes). B, Levels of adenosine receptormRNA expression calculated from geneexpression array data and expressed asa percentage of �-actin mRNA expres-sion. Values are expressed asmean�standard error of 3 experiments.C, RT-PCR of adenosine receptor sub-types mRNA in HUVECs (upper half) andHMEC-1 (lower half). Representative gelsfrom 3 experiments are shown.
Figure 2. Pharmacological characterization of adenosine recep-tors. Accumulation of cAMP induced by the nonselective A2A/A2B
agonist NECA and the A2A selective agonist CGS 21680 inHUVECs (A) and HMEC-1 (B) cells. Values are expressed asmean�standard error of 3 experiments.
Feoktistov et al Adenosine A2B Receptors and Angiogenic Factors 533

tively, compared with cells incubated with vehicle and 1U/mL adenosine deaminase. In contrast, NECA did notproduce any significant increase in expression of these factorsin HUVECs (Figure 3). Expression levels of mRNA for othermembers of the VEGF family factors VEGF-B, VEGF-C, andVEGF-D were not significantly changed after incubation withNECA in either HMEC-1 or HUVECs (online Figure 2).
These results were confirmed with measurements of IL-8and VEGF protein levels in conditioned media. NECAproduced a dose-dependent increase in levels of VEGF(Figure 4A) and IL-8 (Figure 4B) in conditioned media after6 hours of incubation with HMEC-1. The selective A2B
antagonist IPDX23 competitively inhibited NECA-inducedIL-8 secretion with a KB of 1.5 �mol/L (online Figure 3). Incontrast, the A2A selective agonist CGS 21680 at a concen-tration of 10 �mol/L, at which it produces maximal activationof A2A adenosine receptors, had no significant effect on basal
levels of VEGF and IL-8. Incubation of HUVECs withNECA and CGS 21680 up to 24 hours did not change basallevels of VEGF and IL-8 in conditioned media, whereas 1U/mL thrombin, used as a positive control, increased IL-8concentrations from 90�11 pg/mL to 1.1�0.08 ng/mL (on-line Figure 4).
Effect of Adenosine Receptors on Activity of IL-8and VEGF PromotersTo examine the role of adenosine receptor subtypes intranscriptional regulation of IL-8 and VEGF mRNA expres-sion, plasmids encoding human A1, A2A, A2B, and A3 adeno-sine receptors, or an empty expression vector (mock trans-fection), were cotransfected either with IL-8 promoter–drivenluciferase reporter or with VEGF promoter–driven luciferasereporter plasmids in HMEC-1. Twenty-four hours after trans-fection, cells were incubated in the presence or absence of100 �mol/L NECA for 6 additional hours. Under the condi-tions of our experiments, NECA increased activity of VEGFand IL-8 promoters in mock-transfected cells by 1.2- and2-fold, respectively, compared with vehicle-treated cells.Cotransfection of VEGF or IL-8 reporters with A1 or A3
adenosine receptors had no significant effect, and cotransfec-tion with A2A adenosine receptors attenuated by 84�17% and73�8% NECA-induced stimulation of VEGF and IL-8 pro-moters, respectively. In contrast, cotransfection of VEGF orIL-8 reporters with adenosine A2B receptors in HMEC-1resulted in a 2.6�0.1- and 11.7�0.4-fold NECA-inducedstimulation of their activity, respectively (Figure 5). Theobserved effects were not due to variation of transfectionefficiency because such differences were overcome by nor-malization of data with the cotransfected control constitu-tively active Renilla luciferase plasmid pRL-TK. Similarresults were obtained also in CHO-K1 cells lacking endoge-nous adenosine receptors, where cotransfection of IL-8 re-porter with adenosine A2B receptors resulted in 5.1�0.7-foldNECA-induced stimulation, whereas cotransfection with A1,A2A, or A3 had no effect on reporter activity (online Figure 5).In ancillary experiments, the expression of recombinant A2A
and A2B receptors in CHO-K1 cells was verified by NECA-induced increase in intracellular cAMP levels. The expressionof recombinant A1 and A3 adenosine receptors was confirmedusing previously described radioligand binding technique.23
Effect of Constitutively Active � Subunits of GProteins on IL-8 and VEGF PromotersTo examine requirements of IL-8 and VEGF gene expressionfor specific G-protein activation, we transfected HMEC-1with vectors encoding mutationally activated � subunits of Gproteins together with either a IL-8 promoter–driven lucif-erase reporter or with a VEGF promoter–driven luciferasereporter. Cells assayed 30 hours after transfection withconstitutively active mutants of �q, �12, or �13 G proteinsubunits produced 7.6�0.1-, 6.1�0.2-, and 5.7�0.1-foldincrease in VEGF reporter luciferase activity and 23.2�1.6-,19.6�0.9-, and 14.1�0.6-fold increase in IL-8 reporter lucif-erase activity, respectively, when compared with mock-transfected cells. Luciferase activity in cells transfected with
Figure 3. Modulation of angiogenic factors expression by aden-osine. Increase in the levels of IL-8, bFGF, and VEGF mRNAexpression (normalized to �-actin mRNA expression) in HMEC-1and HUVECs induced by incubating with the stable adenosineagonist 100 �mol/L NECA for 6 hours. Data were calculatedfrom gene expression array analysis and expressed asmean�standard error of 3 experiments.
Figure 4. Effect of adenosine receptor activation on VEGF andIL-8 release. Levels of VEGF (A) and IL-8 (B) measured by ELISAin conditioned media of HMEC-1 cells incubated for 6 hourswith the nonselective A2A/A2B agonist NECA and the A2A selec-tive agonist CGS 21680 at indicated concentrations or with theirvehicle. Values are expressed as mean�standard error of 4experiments.
534 Circulation Research March 22, 2002
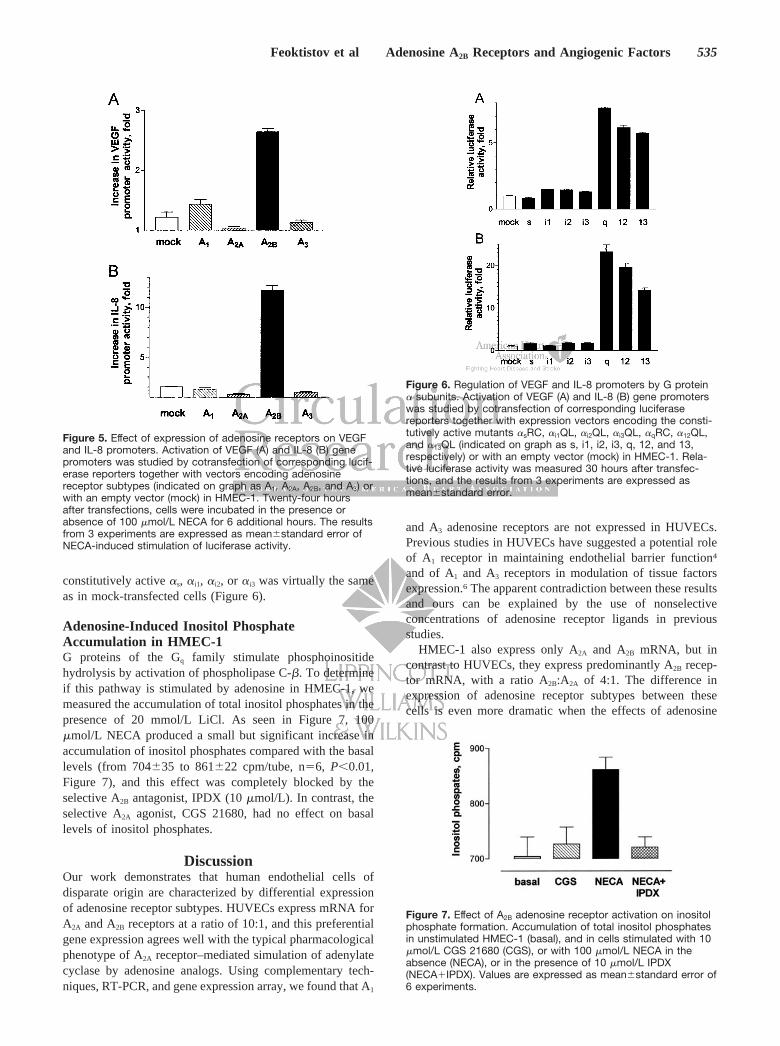
constitutively active �s, �i1, �i2, or �i3 was virtually the sameas in mock-transfected cells (Figure 6).
Adenosine-Induced Inositol PhosphateAccumulation in HMEC-1G proteins of the Gq family stimulate phosphoinositidehydrolysis by activation of phospholipase C-�. To determineif this pathway is stimulated by adenosine in HMEC-1, wemeasured the accumulation of total inositol phosphates in thepresence of 20 mmol/L LiCl. As seen in Figure 7, 100�mol/L NECA produced a small but significant increase inaccumulation of inositol phosphates compared with the basallevels (from 704�35 to 861�22 cpm/tube, n�6, P�0.01,Figure 7), and this effect was completely blocked by theselective A2B antagonist, IPDX (10 �mol/L). In contrast, theselective A2A agonist, CGS 21680, had no effect on basallevels of inositol phosphates.
DiscussionOur work demonstrates that human endothelial cells ofdisparate origin are characterized by differential expressionof adenosine receptor subtypes. HUVECs express mRNA forA2A and A2B receptors at a ratio of 10:1, and this preferentialgene expression agrees well with the typical pharmacologicalphenotype of A2A receptor–mediated simulation of adenylatecyclase by adenosine analogs. Using complementary tech-niques, RT-PCR, and gene expression array, we found that A1
and A3 adenosine receptors are not expressed in HUVECs.Previous studies in HUVECs have suggested a potential roleof A1 receptor in maintaining endothelial barrier function4
and of A1 and A3 receptors in modulation of tissue factorsexpression.6 The apparent contradiction between these resultsand ours can be explained by the use of nonselectiveconcentrations of adenosine receptor ligands in previousstudies.
HMEC-1 also express only A2A and A2B mRNA, but incontrast to HUVECs, they express predominantly A2B recep-tor mRNA, with a ratio A2B:A2A of 4:1. The difference inexpression of adenosine receptor subtypes between thesecells is even more dramatic when the effects of adenosine
Figure 5. Effect of expression of adenosine receptors on VEGFand IL-8 promoters. Activation of VEGF (A) and IL-8 (B) genepromoters was studied by cotransfection of corresponding lucif-erase reporters together with vectors encoding adenosinereceptor subtypes (indicated on graph as A1, A2A, A2B, and A3) orwith an empty vector (mock) in HMEC-1. Twenty-four hoursafter transfections, cells were incubated in the presence orabsence of 100 �mol/L NECA for 6 additional hours. The resultsfrom 3 experiments are expressed as mean�standard error ofNECA-induced stimulation of luciferase activity.
Figure 6. Regulation of VEGF and IL-8 promoters by G protein� subunits. Activation of VEGF (A) and IL-8 (B) gene promoterswas studied by cotransfection of corresponding luciferasereporters together with expression vectors encoding the consti-tutively active mutants �sRC, �i1QL, �i2QL, �i3QL, �qRC, �12QL,and �13QL (indicated on graph as s, i1, i2, i3, q, 12, and 13,respectively) or with an empty vector (mock) in HMEC-1. Rela-tive luciferase activity was measured 30 hours after transfec-tions, and the results from 3 experiments are expressed asmean�standard error.
Figure 7. Effect of A2B adenosine receptor activation on inositolphosphate formation. Accumulation of total inositol phosphatesin unstimulated HMEC-1 (basal), and in cells stimulated with 10�mol/L CGS 21680 (CGS), or with 100 �mol/L NECA in theabsence (NECA), or in the presence of 10 �mol/L IPDX(NECA�IPDX). Values are expressed as mean�standard error of6 experiments.
Feoktistov et al Adenosine A2B Receptors and Angiogenic Factors 535

analogs on cAMP accumulation are compared. In contrast tostimulation of adenylate cyclase in HUVECs, the selectiveA2A agonist CGS 21680 had virtually no effect on adenylatecyclase in HMEC-1. Thus, we found no evidence of func-tional coupling of A2A receptors in HMEC-1.
Interestingly, previously published data on expression ofadenosine receptors in human and porcine endothelial cellsderived from large coronary arteries also demonstrated thepredominance of A2A receptor–mediated simulation of ade-nylate cyclase,9 similar to that observed in this study inHUVECs derived from human umbilical vein. In contrast,adenosine actions were thought to be mediated by A2B
receptors in human retinal microvascular endothelialcells,13,14 a finding similar to that observed in this study inHMEC-1 derived from human skin microvasculature. It istempting to suggest that endothelial cells lining large conduitvessels express predominantly A2A receptors and endothelialcells lining small vessels or capillaries express predominantlyA2B receptors. However, an expanded comparative study ofvarious endothelial cells of different origin is needed to reacha definite conclusion.
Adenosine is known to upregulate VEGF expression inendothelial cells of microvascular origin. Adenosine wasshown to play a role in the hypoxic induction of VEGF inporcine brain-derived microvascular endothelial cells.35
Adenosine A2B is the predominant receptor subtype in humanretinal microvascular endothelial cells.13 In these cells, sim-ilarly to HMEC-1, only A2B adenosine receptors are function-ally linked to activation of adenylate cyclase and stimulateexpression of VEGF.13,14 In the present study, we demon-strated that adenosine stimulates mRNA expression of severalproangiogenic factors, namely VEGF, IL-8, and bFGF, inHMEC-1 expressing predominantly A2B receptors, but not inHUVECs expressing predominantly A2A adenosine receptors.We found that adenosine receptor activation specificallyincreased levels of VEGF mRNA in HMEC-1, and had nosignificant effect on mRNA levels for other members ofVEGF family angiogenic factors including VEGF-B,VEGF-C, and VEGF-D. We have also confirmed that in-creases in VEGF and IL-8 mRNA expression in HMEC-1resulted in increased proteins levels, and these effects weremediated via A2B adenosine receptors because the nonselec-tive A2A/A2B agonist NECA, but not the selective A2A agonistCGS 21680, elevated VEGF and IL-8 levels in conditionedmedia.
We have previously reported that stimulation of A2B
adenosine receptors increased synthesis and secretion of IL-8in human mast cells.31 Adenosine-induced stimulation of IL-8production in endothelial cells is a novel finding with directrelevance to angiogenesis. There is growing evidence thatIL-8 plays an important and specific role in promotingangiogenesis.16–18,36,37 IL-8 is elevated in wounds and en-hances wound healing.38 IL-8, secreted by inflammatory andneoplastic cells, stimulates angiogenesis by paracrine mech-anism in various solid tumors.16–18,36,37 Hypoxia has beenshown to induce expression of both IL-839 and VEGF40 inendothelial cells, implying the existence of autocrine path-ways regulating their growth. Hypoxia can induce angiogen-esis by various mechanisms, including but not limited to
oxygen-dependent regulation of hypoxia-inducible factor 1,the activator of VEGF transcription,25 or stabilization ofVEGF mRNA.41 Our results raise the possibility that adeno-sine, elevated during hypoxia, can also contribute to hypoxia-induced angiogenesis via stimulation of A2B receptors.
In experimental models using cotransfection of IL-8 andVEGF reporters with adenosine receptors, we confirmed thatonly A2B receptors mediate adenosine-induced transcriptionof IL-8 and VEGF genes. In contrast, overexpression of A2A
adenosine receptors attenuated the stimulation of IL-8 andVEGF promoters mediated by native A2B receptors inHMEC-1. These data agree with the previously reporteddownregulation of IL-8 in HUVECs42 and VEGF in ratpheochromocytoma PC12 cells22 mediated via A2A adenosinereceptors. The exact mechanisms underlying the A2A-mediated downregulation of VEGF and IL-8 are currentlyunknown and will require further investigation. It is ofinterest, that selective blockade of A2A adenosine receptors inPC12 cells, which also express a small number of A2B
adenosine receptors,33 resulted in an increase in basal levelsof VEGF,22 presumably due to stimulation of A2B adenosinereceptors.
It is obvious that the opposing effects of A2A and A2B
adenosine receptors on the synthesis of angiogenic factorsimply their coupling to different G-proteins. In this study, wedemonstrate that constitutively active Gq, G12, and G13 �subunits stimulated VEGF and IL-8 promoters. These Gproteins are potential candidates for coupling of A2B receptorsto stimulation of proangiogenic factors. Remarkably, theconstitutively active � subunit of Gs protein, which is coupledto both A2A and A2B adenosine receptors, had virtually noeffect on VEGF and IL-8 reporters. Activation of G proteinsof the Gq subfamily is known to stimulate phosphoinositol-specific phospholipase C-�.43 We have previously demon-strated that A2B adenosine receptors not only stimulate ade-nylate cyclase via coupling to Gs, but can also stimulatephospholipase C-� in human mast cells by cholera toxin– andpertussis toxin–resistant mechanism, presumably via activa-tion of Gq protein.31 Here, we report the activation ofphospholipase C via A2B adenosine receptors in HMEC-1,suggesting that G proteins of the Gq subfamily may beinvolved in the signal transduction leading to increase of IL-8and VEGF production in these cells. In contrast to the Gq
subfamily, the signaling through G12/13 proteins is still poorlyunderstood. Recent evidence suggests that these G proteinsare involved in regulation of cell growth,44 differentiation,45
and apoptosis.46 They have been implicated in regulation ofNa�/H� exchange,27 MAPK pathways,47 and actin cytoskel-eton rearrangement.48 It should be noted, that many Gq-coupled receptors have been shown to couple also to G12
and/or G13 proteins (for review see Fields and Casey49). Ofinterest, while this article was in preparation, Shepard et al50
reported stimulation of IL-8 secretion by the receptor ofKaposi’s sarcoma-associated herpesvirus via coupling to G13
protein in HeLa cells. Potential coupling of G12 and G13
proteins to A2B adenosine receptors and their involvement inthe regulation of angiogenic factors in endothelial cellsdeserves further study.
536 Circulation Research March 22, 2002

In summary, several novel findings are derived from ourresults. We report differential expression of adenosine recep-tor subtypes in endothelial cells of disparate origin; A2A
receptors are the predominant subtype in HUVECs and A2B
receptors are the predominant subtype in the human micro-vascular cell line HMEC-1. Adenosine A2B receptors mediatethe expression of the angiogenic factors IL-8, VEGF, andbFGF. These effects are not linked to stimulation of Gs
proteins, but appear to be mediated by G proteins of the Gq,and possibly of the G12/13 subfamilies. These results confirmand expand those previously reported in other human micro-vascular endothelial cells. Our findings add to a growingevidence that angiogenesis is an important function of aden-osine A2B receptors.
AcknowledgmentsThis work was supported by grants HL55596 (to I.F.), GM56159 (toT.V.-Y.), and HL67232 (to I.B.) from NIH, and by a research grantfrom CV Therapeutics. The authors thank Tenning Maa (CVTherapeutics) for assistance with real-time RT-PCR experiments.
References1. Nees S, Herzog V, Becker BF, Bock M, Des Rosiers C, Gerlach E. The
coronary endothelium: a highly active metabolic barrier for adenosine.Basic Res Cardiol. 1985;80:515–529.
2. Gerlach E, Becker BF, Nees S. Formation of adenosine by vascularendothelium: a homeostatic and antithrombogenic mechanism? In:Gerlach E, Becker BF, eds. Topics and Perspectives in AdenosineResearch. Berlin, Germany: Springer-Verlag; 1987:309–320.
3. Costa F, Sulul P, Angel M, Cavalcante J, Haile V, Christman BW,Biaggioni I. Intravascular source of adenosine during forearm ischemia inhumans: implications for reactive hyperemia. Hypertension. 1999;33:1453–1457.
4. Richard LF, Dahms TE, Webster RO. Adenosine prevents permeabilityincrease in oxidant-injured endothelial monolayers. Am J Physiol. 1998;274:H35–H42.
5. Sexl V, Mancusi G, Holler C, Gloria-Maercker E, Schutz, Freissmuth M.Stimulation of the mitogen-activated protein kinase via the A2A-adenosinereceptor in primary human endothelial cells. J Biol Chem. 1997;272:5792–5799.
6. Deguchi H, Takeya H, Urano H, Gabazza EC, Zhou H, Suzuki K.Adenosine regulates tissue factor expression on endothelial cells. ThrombRes. 1998;91:57–64.
7. Lennon PF, Taylor CT, Stahl GL, Colgan SP. Neutrophil-derived5�-adenosine monophosphate promotes endothelial barrier function viaCD73-mediated conversion to adenosine and endothelial A2B receptoractivation. J Exp Med. 1998;188:1433–1443.
8. Iwamoto T, Umemura S, Toya Y, Uchibori T, Kogi K, Takagi N, Ishii M.Identification of adenosine A2 receptor-cAMP system in human aorticendothelial cells. Biochem Biophys Res Commun. 1994;199:905–910.
9. Olanrewaju HA, Qin W, Feoktistov I, Scemama JL, Mustafa SJ. Aden-osine A2A and A2B receptors in cultured human and porcine coronaryartery endothelial cells. Am J Physiol. 2000;279:H650–H656.
10. Li J, Fenton RA, Wheeler HB, Powell CC, Peyton BD, Cutler BS,Dobson JG Jr. Adenosine A2a receptors increase arterial endothelial cellnitric oxide. J Surg Res. 1998;80:357–364.
11. Smits P, Williams SB, Lipson DE, Banitt P, Rongen GA, Creager MA.Endothelial release of nitric oxide contributes to the vasodilator effect ofadenosine in humans. Circulation. 1995;92:2135–2141.
12. Fischer S, Sharma HS, Karliczek GF, Schaper W. Expression of vascularpermeability factor/vascular endothelial growth factor in pig cerebralmicrovascular endothelial cells and its upregulation by adenosine. BrainRes. 1995;28:141–148.
13. Grant MB, Tarnuzzer RW, Caballero S, Ozeck MJ, Davis MI, Spoerri PE,Feoktistov I, Biaggioni I, Shryock JC, Belardinelli L. Adenosine receptoractivation induces vascular endothelial growth factor in human retinalendothelial cells. Circ Res. 1999;85:699–706.
14. Grant MB, Davis MI, Caballero S, Feoktistov I, Biaggioni I, BelardinelliL. Proliferation, migration, and ERK activation in human retinal endo-thelial cells through A2B adenosine receptor stimulation. Invest Oph-thalmol Visual Sci. 2001;42:2068–2073.
15. Adair TH, Montani JP, Strick DM, Guyton AC. Vascular development inchick embryos: a possible role for adenosine. Am J Physiol. 1989;256:H240–H246.
16. Rofstad EK, Halsor EF. Vascular endothelial growth factor, interleukin-8,platelet-derived endothelial cell growth factor, and basic fibroblastgrowth factor promote angiogenesis and metastasis in human melanomaxenografts. Cancer Res. 2000;60:4932–4938.
17. Westphal JR, Van’t Hullenaar R, Peek R, Willems RW, Crickard K,Crickard U, Askaa J, Clemmensen I, Ruiter DJ, De Waal RM. Angiogenicbalance in human melanoma: expression of VEGF, bFGF, IL-8, PDGFand angiostatin in relation to vascular density of xenografts in vivo. Int JCancer. 2000;86:768–776.
18. Slaton JW, Inoue K, Perrotte P, El Naggar AK, Swanson DA, Fidler IJ,Dinney CP. Expression levels of genes that regulate metastasis andangiogenesis correlate with advanced pathological stage of renal cellcarcinoma. Am J Pathology. 2001;158:735–743.
19. Hashimoto E, Kage K, Ogita T, Nakaoka T, Matsuoka R, Kira Y.Adenosine as an endogenous mediator of hypoxia for induction ofvascular endothelial growth factor mRNA in U-937 cells. BiochemBiophys Res Commun. 1994;204:318–324.
20. Takagi H, King GL, Robinson GS, Ferrara N, Aiello LP. Adenosinemediates hypoxic induction of vascular endothelial growth factor inretinal pericytes and endothelial cells. Invest Ophthalmol Visual Sci.1996;37:2165–2176.
21. Kobayashi S, Millhorn DE. Stimulation of expression for the adenosineA2A receptor gene by hypoxia in PC12 cells: a potential role in cellprotection. J Biol Chem. 1999;274:20358–20365.
22. Olah ME, Roudabush FL. Down-regulation of vascular endothelialgrowth factor expression after A2A adenosine receptor activation in PC12pheochromocytoma cells. J Pharmacol Exp Ther. 2000;293:779–787.
23. Feoktistov I, Garland E, Goldstein AE, Zeng D, Belardinelli L, Wells JN,Biaggioni I. Inhibition of human mast cell activation with the novelselective adenosine A2B receptor antagonist 3-isobutyl-8-pyrrolidinoxanthine (IPDX). Biochem Pharmacol. 2001;62:1163–1173.
24. Mitchell CH, Peterson-Yantorno K, Carre DA, McGlinn AM, Coca-Prados M, Stone RA, Civan MM. A3 adenosine receptors regulateCl-channels of nonpigmented ciliary epithelial cells. Am J Physiol. 1999;276:C659–C666.
25. Forsythe JA, Jiang BH, Iyer NV, Agani F, Leung SW, Koos RD,Semenza GL. Activation of vascular endothelial growth factor genetranscription by hypoxia-inducible factor 1. Mol Cel Biol. 1996;16:4604–4613.
26. Ishikawa Y, Mukaida N, Kuno K, Rice N, Okamoto S, Matsushima K.Establishment of lipopolysaccharide-dependent nuclear factor �B acti-vation in a cell-free system. J Biol Chem. 1995;270:4158–4164.
27. Voyno-Yasenetskaya T, Conklin BR, Gilbert RL, Hooley R, Bourne HR,Barber DL. G�13 stimulates Na-H exchange. J Biol Chem. 1994;269:4721–4724.
28. Conklin BR, Chabre O, Wong YH, Federman AD, Bourne HR. Recom-binant Gq�: mutational activation and coupling to receptors and phospho-lipase C. J Biol Chem. 1992;267:31–34.
29. Wong YH, Federman A, Pace AM, Zachary I, Evans T, Pouyssegur J,Bourne HR. Mutant � subunits of Gi2 inhibit cyclic AMP accumulation.Nature. 1991;351:63–65.
30. Lyons J, Landis CA, Harsh G, Vallar L, Grunewald K, Feichtinger H,Duh QY, Clark OH, Kawasaki E, Bourne HR. Two G protein oncogenesin human endocrine tumors. Science. 1990;249:655–659.
31. Feoktistov I, Biaggioni I. Adenosine A2b receptors evoke interleukin-8secretion in human mast cells: an enprofylline-sensitive mechanism withimplications for asthma. J Clin Invest. 1995;96:1979–1986.
32. van der Ploeg I, Ahlberg S, Parkinson FE, Olsson RA, Fredholm BB.Functional characterization of adenosine A2 receptors in Jurkat cells andPC12 cells using adenosine receptor agonists. Naunyn-SchmiedebergsArch Pharmacol. 1996;353:250–260.
33. Feoktistov I, Biaggioni I. Adenosine A2B receptors. Pharmacol Rev.1997;49:381–402.
34. Feoktistov I, Biaggioni I. Characterization of adenosine receptors inhuman erythroleukemia cells: further evidence for heterogeneity of aden-osine A2 receptors. Mol Pharmacol. 1993;43:909–914.
35. Fischer S, Knoll R, Renz D, Karliczek GF, Schaper W. Role of adenosinein the hypoxic induction of vascular endothelial growth factor in porcine
Feoktistov et al Adenosine A2B Receptors and Angiogenic Factors 537

brain derived microvascular endothelial cells. Endothelium. 1997;5:155–165.
36. Inoue K, Slaton JW, Kim SJ, Perrotte P, Eve BY, Bar-Eli M, Radinsky R,Dinney CP. Interleukin-8 expression regulates tumorigenicity andmetastasis in human bladder cancer. Cancer Res. 2000;60:2290–2299.
37. Inoue K, Slaton JW, Eve BY, Kim SJ, Perrotte P, Balbay MD, Yano S,Bar-Eli M, Radinsky R, Pettaway CA, Dinney CP. Interleukin-8expression regulates tumorigenicity and metastases in androgen-independent prostate cancer. Clin Cancer Res. 2000;6:2104–2119.
38. Rennekampff HO, Hansbrough JF, Kiessig V, Dore C, Sticherling M,Schroder JM. Bioactive interleukin-8 is expressed in wounds andenhances wound healing. J Surg Res. 2000;93:41–54.
39. Karakurum M, Shreeniwas R, Chen J, Pinsky D, Yan SD, Anderson M,Sunouchi K, Major J, Hamilton T, Kuwabara K. Hypoxic induction ofinterleukin-8 gene expression in human endothelial cells. J Clin Invest.1994;93:1564–1570.
40. Namiki A, Brogi E, Kearney M, Kim EA, Wu T, Couffinhal T, VarticovskiL, Isner JM. Hypoxia induces vascular endothelial growth factor in culturedhuman endothelial cells. J Biol Chem. 1995;270:31189–31195.
41. Levy AP, Levy NS, Goldberg MA. Post-transcriptional regulation of vascularendothelial growth factor by hypoxia. J Biol Chem. 1996;271:2746–2753.
42. Bouma MG, van den Wildenberg FA, Buurman WA. Adenosine inhibitscytokine release and expression of adhesion molecules by activatedhuman endothelial cells. Am J Physiol. 1996;270:C522–C529.
43. Smrcka AV, Hepler JR, Brown KO, Sternweis PC. Regulation of poly-phosphoinositide-specific phospholipase C activity by purified Gq.Science. 1991;251:804–807.
44. Voyno-Yasenetskaya TA, Pace AM, Bourne HR. Mutant � subunits ofG12 and G13 proteins induce neoplastic transformation of Rat-1 fibroblasts.Oncogene. 1994;9:2559–2565.
45. Denis-Henriot D, de Mazancourt P, Morot M, Giudicelli Y. Mutant �-subunitof the G protein G12 activates proliferation and inhibits differentiation of3T3-F442A preadipocytes. Endocrinology. 1998;139:2892–2899.
46. Berestetskaya YV, Faure MP, Ichijo H, Voyno-Yasenetskaya TA. Reg-ulation of apoptosis by �-subunits of G12 and G13 proteins via apoptosissignal-regulating kinase-1. J Biol Chem. 1998;273:27816–27823.
47. Voyno-Yasenetskaya TA, Faure MP, Ahn NG, Bourne HR. G�12 andG�13 regulate extracellular signal-regulated kinase and c-Jun kinasepathways by different mechanisms in COS-7 cells. J Biol Chem. 1996;271:21081–21087.
48. Buhl AM, Johnson NL, Dhanasekaran N, Johnson GL. G�12 and G�13
stimulate Rho-dependent stress fiber formation and focal adhesionassembly. J Biol Chem. 1995;270:24631–24634.
49. Fields TA, Casey PJ. Signaling functions and biochemical properties ofpertussis toxin-resistant G-proteins. Biochem J. 1997;321:561–571.
50. Shepard JW, Yang M, Xie P, Browning DD, Voyno-Yasenetskaya T, Kozasa T,Ye RD. Constitutive activation of NF-�B and secretion of IL-8 induced by the Gprotein–coupled receptor of Kaposhi’s sarcoma-associated herpesvirus involvesG�13 and RhoA. J Biol Chem. 2001;276:45979–45987.
538 Circulation Research March 22, 2002
Copyright © 2022 FDOKUMEN




















