
Different growth strategies in the tropical pioneer tree Trema micrantha during succession on a...
18
Changes in the Herbaceous Communities on the Landslide of the Casita Volcano, Nicaragua, during Early Succession Eduardo Velázquez & Antonio Gómez-Sal # Institute of Botany, Academy of Sciences of the Czech Republic 2009 Abstract This study examined the main changes in the herbaceous communities during the first four years of succession in a large landslide on Casita Volcano, Nicaragua, located in a densely populated area that has a tropical dry climate. Our main objective was to determine the major pathways of change in community features, such as richness, biovolume, species composition, and abundances of plant traits and to verify if they varied between the different landslide areas based on abiotic heterogeneity and landscape context. Number, percent cover, mean height and traits of herbaceous species, and several abiotic factors related to fertility and geomorphological stability of substrates were sampled in 28 permanent plots. Environmental heterogeneity strongly influenced early successional changes in the herbaceous communities during the four years of the study. Biovolume increased in the unstable and infertile areas and decreased in the stable and fertile landslide areas. In most zones, species richness decreased significantly because of the expansion of a few dominant species that developed a large cover and excluded other species. Those dominant species were mainly responsible for changes in species composition and included annual forbs, e.g., Calopogonium mucunoides and Stizolobium pruriens, graminoids that have rhizomes or stolons, e.g., Hyparrhenia rufa, and perennial nitrogen-fixing forbs, e.g., Clitoria ternatea. They might be important in determining future successional patterns on the Casita Volcano landslide. Keywords Biovolume . Environmental heterogeneity . Plant traits . Richness . Species composition Folia Geobot (2009) 44:1–18 DOI 10.1007/s12224-009-9031-3 Plant nomenclature Stevens et al. (2001) E. Velázquez (*) : A. Gómez-Sal Departamento de Ecología, Universidad de Alcalá de Henares, Alcalá de Henares 28871, Madrid, Spain e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Different growth strategies in the tropical pioneer tree Trema micrantha during succession on a...

Changes in the Herbaceous Communitieson the Landslide of the Casita Volcano, Nicaragua,during Early Succession
Eduardo Velázquez & Antonio Gómez-Sal
# Institute of Botany, Academy of Sciences of the Czech Republic 2009
Abstract This study examined the main changes in the herbaceous communitiesduring the first four years of succession in a large landslide on Casita Volcano,Nicaragua, located in a densely populated area that has a tropical dry climate. Ourmain objective was to determine the major pathways of change in communityfeatures, such as richness, biovolume, species composition, and abundances of planttraits and to verify if they varied between the different landslide areas based onabiotic heterogeneity and landscape context. Number, percent cover, mean heightand traits of herbaceous species, and several abiotic factors related to fertility andgeomorphological stability of substrates were sampled in 28 permanent plots.Environmental heterogeneity strongly influenced early successional changes in theherbaceous communities during the four years of the study. Biovolume increased inthe unstable and infertile areas and decreased in the stable and fertile landslide areas.In most zones, species richness decreased significantly because of the expansion of afew dominant species that developed a large cover and excluded other species.Those dominant species were mainly responsible for changes in species compositionand included annual forbs, e.g., Calopogonium mucunoides and Stizolobiumpruriens, graminoids that have rhizomes or stolons, e.g., Hyparrhenia rufa, andperennial nitrogen-fixing forbs, e.g., Clitoria ternatea. They might be important indetermining future successional patterns on the Casita Volcano landslide.
Keywords Biovolume . Environmental heterogeneity . Plant traits . Richness .
Species composition
Folia Geobot (2009) 44:1–18DOI 10.1007/s12224-009-9031-3
Plant nomenclature Stevens et al. (2001)
E. Velázquez (*) :A. Gómez-SalDepartamento de Ecología, Universidad de Alcalá de Henares, Alcalá de Henares 28871, Madrid,Spaine-mail: [email protected]

Introduction
Herbaceous communities play an important role in a wide range of ecosystemfunctions, such as energy flow and nutrient cycling (Frelich et al. 2003) and, duringearly succession, experience significant changes in features, such as richness,biomass, species composition, and dominance of species with certain plant traits(Roberts and Zhu 2002). Typically, herbaceous communities that colonize a recentlydisturbed site show high species richness (Roberts 2004). This high richness maypersist during the first years of succession due to high rates of species turnover(Walker and Willig 1999), but it can decline rapidly if competition for soil resourcesoccurs among dominant species (Pausas and Austin 2001). In a newly opened site,herbaceous communities can generate a large biomass in a few years (Silvertown etal. 1994; Grace 1999), and marked differences in species composition can occurwithin those communities depending on the abiotic heterogeneity of the area beingcolonized (Gómez-Sal et al. 1986; Pineda et al. 1987). In addition, herbaceouscommunities usually exhibit a wide variety of plant traits (McIntyre et al. 1995). Theenvironmental conditions at a site influence the abundances of species with certainplant traits within communities (Montalvo et al. 1991; Rodríguez et al. 1997) andthese abundances can change considerably even in the years immediately afterdisturbance (Bonet and Pausas 2004). Such change is particularly true of plant traitsassociated with vegetation type, life history, and the geographic origin of the species(McIntyre et al. 1995; Tsuyuzaki and del Moral 1995). The species, however, oftenshow some associated traits that are rarely considered individually; rather, they aretreated collectively as a “plant strategy” (Leishman and Westoby 1992).
In many studies of vegetation recovery on disturbed ground, herbaceouscommunities have received limited attention because of difficulties when assessingsuccessional patterns due to their high species turnover (Small and McCarthy 2003).In the tropics, herbaceous communities have been considered in successional studiescarried out in old fields (Aide et al. 1995; Rivera et al. 2000), restored forests (deSouza and Batista 2004), areas affected by hurricanes (Chinea 1999), and volcaniceruptions (Whittaker et al. 1989; Bush and Whittaker 1995). Herbaceouscommunities have also been considered in landslides (Lundgren 1978; Walker1994; Restrepo and Vitousek 2001). However, specific studies addressing changes inherbaceous communities during early succession in this type of habitat areparticularly rare.
Landslides are severe disturbances (Guariguata 1990) with high abioticheterogeneity (Walker et al. 1996). Successional patterns in the highly unstableand infertile upper or “erosional” zones are completely different from the patterns inthe lower or “depositional” zones that are stable and fertile (Myster and Fernández1995). For that reason, landslides are considered “patch-dynamics systems” (Pickettand White 1985) and may be very useful for studying the effect of contingenciesassociated with geomorphological stability, fertility of substrates and with landscapecontext on patterns of change of community features (Myster and Walker 1997).
In this study, we examined the early successional changes in the herbaceouscommunities of a large landslide that presents a high abiotic heterogeneity and islocated in the Pacific region of Central America (Casita Volcano, Nicaragua).Human disturbance has dramatically reduced the formerly abundant tropical dry
2 E. Velázquez, A. Gómez-Sal

forests in the area (Gillespie et al. 2000). These forests are among the mostendangered and least known ecosystems in the world (Janzen 1988).
The study’s principal objective was to determine the major patterns of change inthe herbaceous communities on the landslide during the first four years following thedisturbance. We sought to determine whether herbaceous communities vary amongthe different landslide areas, and to identify the main factors driving such change.
Specifically, we addressed the following questions: i) How do richness,biovolume, turnover rate, and species composition vary among the early years ofsuccession and among the landslide areas? ii) How does the abundance of plant traitsor plant strategies change over time and between landslide areas?
Material and Methods
Study Area
The study area was a landslide on the Casita Volcano (12°41’ N; 85°57’ W), whichis part of the “Maribios” volcanic range in western Nicaragua (Fig. 1a). Thelandslide, which is mainly composed of blocks (highly fractured dacitic rocks,volcanic and tectonic breccias), debris material and saprolite (Marquínez et al. 2002),was triggered by an exceptional rainfall event (500 mm in 24 h) on 30 October 1998,during Hurricane Mitch (Sheridan 1998). It formed an enormous lahar at mid-slope(Kerle et al. 2003; Scott et al. 2005) that killed more than 2,000 people and displacedanother 8,000 (CEPAL 1999). The landslide destroyed the pre-existing vegetation
Fig. 1 Study area. a Location inNicaragua. b Detailed map ofthe landslide indicating the dif-ferent subzones based on abioticheterogeneity (fertility and geo-morphologic stability) and land-scape context; erosional (E1, E2,E3), transitional (T1 and T2) anddepositional (D1 and D2). Thelocations of sampling plots areindicated
Herbaceous communities during early succession on a landslide 3

almost completely, leaving only some broken trees and a small “vegetation island” inthe upper part of the affected area. With an altitude range of 150–1,350 m a.s.l., andan area of 11.21 km2, the Casita Volcano landslide is much larger than others studiedin Central America and the Caribbean region. In addition, the ecological andsocioeconomic characteristics of the surrounding area are rather different (Velázquezand Gómez-Sal 2007). Mean annual rainfall (1,250 mm/yr) is considerably lowerand the dry season (November to April) is longer in the dry forests of westernNicaragua than they are in the rain and cloud forests of eastern Central America andon the Caribbean islands. In addition, the frequency of hurricanes (long-term averageof hurricane strikes per year equals 0.2; Pielke et al. (2003)) and heavy rainfallevents that trigger landslides is lower in western Nicaragua than it is in easternCentral America and on the Caribbean islands. In western Nicaragua, the populationdensity (172 people/km2, for 2005, Instituto Nacional de Estadísticas y Censos,Nicaragua, http://www.inec.gob.ni/) and poverty levels [0.690 HDI (UNDP 2005)] arehigh, and local ecosystems have experienced substantial exploitation and reduction(Corrales-Rodríguez 1983), however, large patches of relatively well-preserved forestsremain. The structure and composition of the plant communities near the crater of theCasita Volcano are fairly similar to those in cloud forests (Salas-Estrada 1999), but thevolcano is covered mostly by tropical dry forests that have been partially convertedinto “shaded” coffee plantations, and are interspersed with scattered huts and small-holding lands at mid-slope. At the base of the volcano, cultivated lands dominate thelandscape. Human disturbances, such as the clearcutting of forests and fire arecommon in the area (Corrales-Rodríguez 1983; Salas-Estrada 1999).
Field Sampling
In 1999, three major “zones” (Erosional, Transitional, and Depositional) wereidentified in the landslide. They reflect, respectively, the “initiation,” “transport,”and “deposition” zones in landslides, based on geomorphological criteria (Martin etal. 2002). The erosional zones (E), where the landslide began, were the steepestareas. The transitional zones (T) were the moderately steep areas where the landslidemass flowed and possibly also settled, and the depositional zones (D) were almostflat areas where the landslide mass settled. In the erosional zones, the agriculturaland forest soils were completely removed by the landslide, but they remained intactin the depositional zones, and in small patches within the transitional zones. Basedon the adjacent vegetation of the landslide we identified three “subzones” in theerosional zones (E1, E2 and E3) and two in the transitional (T1 and T2) anddepositional zones (D1 and D2) (Fig. 1b). E1 and D2 had shaded coffee plantationsalong their eastern border; E2 and T1 had tropical dry forests along their edges; E3was enclosed by tropical dry forests that had plant species typical of cloud forests;and cultivated lands were dominant beyond the western border of T2 and along theedges of D1. A group of remnant trees was present in the middle of D2, adepositional landing zone at mid-slope.
To document the early successional changes in the landslide, the vegetation andsoil were sampled in 1999, 2000, 2001 and 2002 combining different methods(Table 1). In 1999, the vegetation was surveyed in 10×10 m2 non-permanent plots,randomly located within each of the seven subzones (three plots in E1, E2, and D2,
4 E. Velázquez, A. Gómez-Sal

five plots in E3, T2 and T1, and six plots in D1). These plots were not re-sampledduring the study period. In 2000, we established one permanent 10×10 m2 plot ineach subzone (seven permanent plots sampled in total). In 2001, these sevenpermanent plots were re-sampled, and three additional permanent plots wereestablished in each subzone (28 permanent plots sampled in total). In 2002, all ofthe 28 permanent plots were re-sampled again. Within each subzone, each permanentplot was assigned a letter (“a” through “d”), depending on its N-S location.
In each of the four years (1999, 2000, 2001, and 2002), at the end of the rainy season(November to January), the herbaceous vegetation was surveyed within four 1-m2
quadrants positioned along the diagonal of each plot. We recorded the number ofspecies of herbs, grasses, ferns, and seedlings of woody plants (“low vegetation”(Benitez-Malvido 2006)), and measured the cover (%) and average height (cm) of eachspecies. In addition, species’ characteristics, such as shape, size and number of fruit,and the presence of nodules with nitrogen-fixing bacteria were noted. That information,coupled with local knowledge and expert opinion, allowed us to identify a set of planttraits or attributes for each species based on the most important categories of plant traitsduring early succession; vegetation type, life history, and geographic origin (Tsuyuzakiand del Moral 1995). Each trait was coded to “nominal multistate” and “binary”(presence/absence) classes (Leishman and Westoby 1992) (Table 2).
Within each plot, we measured abiotic factors associated with stability [elevation,m a.s.l.), slope (%), stone cover (%), and mean diameter of stones (cm)] and fertility
Table 1 Plots (10×10 m2) sampled in the different subzones of the Casita Volcano landslide in thedifferent years of the study period (1999, 2000, 2001 and 2002)
Zones Subzones Years
1999 2000 2001 2002
Erosional E1 E11, E12, E13 E1d E1a, E1b, E1c, E1d E1a, E1b, E1c, E1dE2 E21, E22, E23 E2d E2a, E2b, E2c, E2d E2a, E2b, E2c, E2dE3 E31, E32, E33,
E34, E35E3c E3a, E3b, E3c, E3d E3a, E3b, E3c, E3d
Number of plots 11 3 12 12Transitional T1 T11, T12, T13,
T14, T15T1a T1a, T1b, T1c, T1d T1a, T1b, T1c, T1d
T2 T21, T22, T23,T24, T25
T2d T2a, T2b, T2c, T2d T2a, T2b, T2c, T2d
Number of plots 10 2 8 8Depositional D1 D11, D12, D13,
D14, D15, D16D1b D1a, D1b, D1c, D1d D1a, D1b, D1c, D1d
D2 D21, D22, D23 D2b D2a, D2b, D2c, D2d D2a, D2b, D2c, D2dNumber of plots 9 2 8 8Total numberof plots
30 7 28 28
Non-permanent plots (Np), in italics, were established and sampled only in 1999. Within each subzone,each one of them was assigned a number. Permanent plots were established in a stepwise fashion between2000 and 2001, and sampled in 2000, 2001 and 2002. Within each subzone, each one of them wasassigned a letter “a” through “d”, depending on its N-S location. Permanent plots that were re-sampled arein bold letters.
Herbaceous communities during early succession on a landslide 5

(amounts of NO3−, P, and K (ppm) and the proportions of sand, silt, and clay) of
substrates. These last factors could only be measured in 2002. Stone cover wasdefined as the percentage of total plot area covered by stones: rock fragments over25-cm diameter, according to the USDA classification system of particle sizes (Portaet al. 1999). The mean diameter of stones was calculated from a sample of 50 rockfragments per plot using callipers and a tape measure. The amount of NO3
−, P, andK, was determined using a LaMotte soil test kit (La Motte, Chesterton, Maryland,USA, 1997) in three 200-g soil samples collected in each plot. Sand, silt and claywere defined as particles with diameters ranging from 2 to 0.05 mm, from 0.05 to0.002 mm and less than 0.002 mm, respectively, according to the USDAclassification system of particle sizes (Porta et al. 1999). To determine theirproportions, 200-g soil samples were collected in each plot, dried and sieved. Wealso estimated the cover (%) of the woody canopy (trees and shrubs) per plot.
In the second (2000–2001) and third (2001–2002) periods of the study, human-induced disturbances occurred in the depositional and transitional zones. Theyinvolved widespread, extensive removal of pioneer trees for firewood and theexpansion of croplands (D1 and T1), and fires created by peasants living nearby thatexpanded into the landslide (T1 and T2). In each sampling period, plots wereidentified as “cleared” or “burned” if there were signs of clearcutting (e.g., presence
Table 2 Categories of plant traits and their classes, for the species recorded at the Casita Volcanolandslide during the study period
Categories/ Traits/ Classes Class type
VegetativeGrowth form1 NM4woody2 • graminoid3• forb4 • fern
Vegetative spreading mechanisms (rhizomes/stolons) BMorphology of the shoots5 NM3Erects • vines • creeping shoots
Life-historyLongevity6 NM2perennial • annual
Nitrogen fixation BResprouting ability7 B
Seed biologyDispersal mode8 NM3gravity • animal • wind
Geographic origin9
exotic ● native NM2
Class type and number indicate how each attribute is coded for the analysis. Traits are grouped into threemain categories: vegetative, life history, and geographic origin. Class types: B – binary (presence/absence); NM – nominal multistate1 Based on the classes of Klinka et al. (1989); 2 Seedlings of woody species <20 cm tall; 3 Grasses andgrass-like plants, including sedges and rushes; 4 Non-graminoid herbaceous plants; 5 Based on Orshan’s1982 “morphological characters”; 6 Based on Tsuyuzaki’s 1989 classes; 7 Based on Leishman andWestoby’s 1992 traits; 8 Based on Tsuyuzaki and del Moral’s 1995 classes; 9 Based on “Flora deNicaragua” (Stevens et al. 2001).
6 E. Velázquez, A. Gómez-Sal

of tailings) or burning (e.g., scorched individuals and stems), respectively. Speciestaxonomy and nomenclature follow the Flora de Nicaragua (Stevens et al. 2001).
Statistical Analysis
To determine the direction of change (Bakker et al. 1996) in species richness and inbiovolume, for each zone and year of study, the mean number of species and meantotal biovolume (height (m) × cover (m2)) were calculated. To determine rates ofchange in species turnover, we calculated the mean Sørensen’s Dissimilarity Index(S) (Kent and Coker 1992) for each zone, in each two-year period of study (1999–2000, 2000–2001 and 2001–2002). To assess changes in species composition,frequency distributions of the species present in the landslide were calculated, andordination analyses (DCA) were performed on cover matrices using only the mostabundant species in the landslide (those present in more than 20% of the plots).“Plant strategies” were defined based on those groups of species that had sets ofassociated traits (Leishman and Westoby 1992). Most species present in theherbaceous communities of the landslide were dispersed by wind and were ofexotic origin, therefore “Dispersal mode” and “Geographic origin” were not used indefining groups of plant strategies.
To test for the effects of zone on successional changes in community featureswithin the landslide, repeated-measures ANOVA tests were used. Mean values ofnumber of species, total biovolume, Sørensen’s Dissimilarity Index, plot scores inthe Axis I and II of the ordination space and total cover of each group of speciesbased on plant strategies, were calculated per subzone and year, and considered asdependent variables in the analysis. “Zone and “Time” were considered as between-and within-subjects factors in all the analyses with three (Erosional, Transitional,Depositional) and four (1999, 2000, 2001 and 2002) levels, respectively. The Box-M, Bartlett and Mauchley’s tests were performed to meet the homogeneity of thevariance-covariance matrix, the independence of the dependent variables, and thesphericity assumptions. When the sphericity assumption was not assumed, P-valuesfor between-subjects factors were based on the Huynh and Feldt (1976) epsilonadjustment (von Ende 2001). A similar approach was used to test for the effects ofsubzone on successional changes in community features within each landslide zone.In each of these analyses, however, values of number of species, total biovolume,Sørensen’s Dissimilarity Index, plot scores in the Axis I and II of the ordinationspace and total cover of each group of species based on plant strategies per plot andyear were considered as dependent variables. “Subzone” was considered as thebetween-subjects factor, with three levels in the erosional zone (E1, E2, E3) and twoin both the transitional (T1 and T2) and the depositional (D1 and D2) zones. Due tothe differences in the number of plots per year between 1999 and 2000 censuses and2001 and 2002 censuses (Table 1), only the last two years of the study period, 2001and 2002, with an equal number of plots, were used in these analyses. Thus, “Time”was considered as the within-subjects factors, but with two levels (2001 and 2002).
To determine the main causes of change in each zone, we calculated theSpearman rank correlation coefficients (r) between each of the biotic variables (meannumber of species, mean total biovolume, plot scores in the Axis I and II of the DCAand covers of each group of species based on plant strategies) and each of the
Herbaceous communities during early succession on a landslide 7

environmental factors (elevation, slope, stone cover, mean diameter of stones, amountsof NO3
−, P and K, proportions of sand, silt and clay, presence of clearcutting, andburning, and total woody cover) for each zone. When it was appropriate, variableswere log-, square root or box-cox transformed to achieve normality (Sokal and Rohlf1995). Clustering, repeated-measures ANOVAs, and Spearman rank correlations wereperformed using STATISTICA software (Statsoft 2001). Ordination Analysis wasperformed using CANOCO (ter Braak and Šmilauer 2002).
Results
Changes in Species Richness, Biovolume, and Turnover Rates
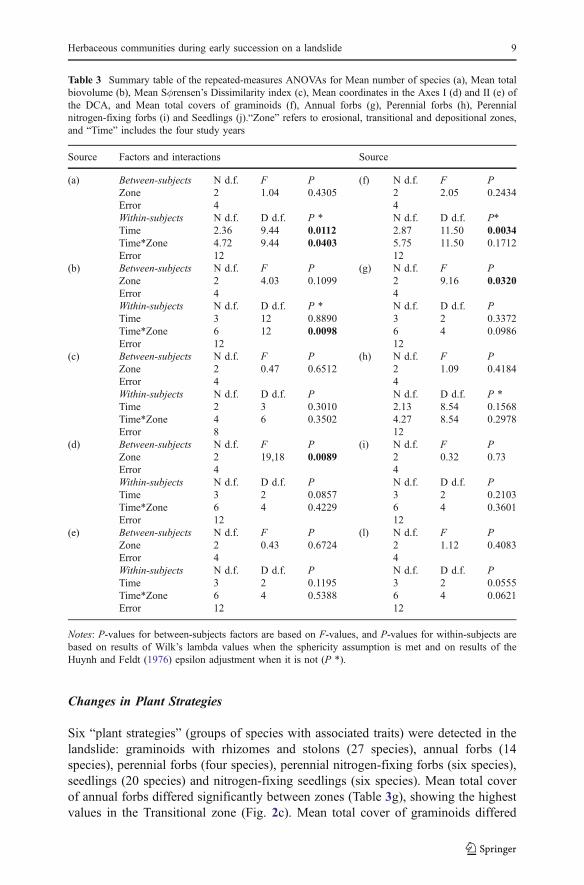
Species richness of the herbaceous communities within the landslide was high (83species were registered in total). Species richness clearly decreased in all zonesthroughout the study period, although patterns of decline differed significantlybetween zones (Table 3a, Fig. 2a). Within the depositional zone, changes in speciesrichness differed significantly between subzones D1 and D2 (Table 4a). During thefour years of the study, changes in biovolume also differed significantly betweenzones (Table 3b). Biovolume decreased in the depositional zone and increased in thetransitional and erosional zones (Fig. 2b). Within the erosional zone, values ofbiovolume differed significantly between subzones E1, E2 and E3 (Table 4f). Overthe course of the four-year study, species turnover rates increased slightly, but theydid not differ significantly between zones, between years of the study period, orbetween zones during years of the study period (Table 3c).
Changes in Species Composition
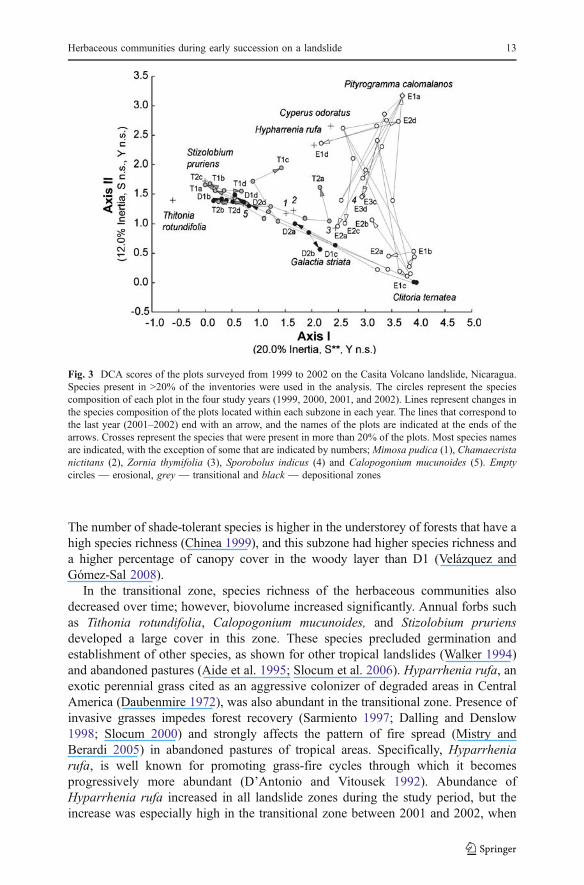
Twelve species were the most abundant in the landslide, appearing in more than 20%of the inventories. Most were wind-dispersed graminoids and forbs of exotic origin(Table 5). The DCA that included only these species (Fig. 3) had a reasonably highpercentage of inertia explained and, although the DCA excluded many species (71 of83), those excluded appeared mostly in the two first years and were not important indetermining the overall changes in species composition. Species compositionexhibited highly divergent trajectories of change during the study period in mostplots (Fig. 3), and values of plot scores on Axis I of the DCA differed significantlybetween zones (Table 3d). In the erosional zone, plots of E3 and E2 were dominatedby Sporobolus indicus and Zornia thymifolia, and those of E1 by Clitoria ternatea,Hyparrhenia rufa and Pityrogramma calomalanos. In the transitional zone at thebeginning of the study, Calopogonium mucunoides was dominant in all of the plotsbut over the study period, the cover of Stizolobium pruriens and Tithoniarotundifolia increased significantly. Two plots exhibited clear convergent trajectoriestoward a high abundance of Hyparrhenia rufa. In the depositional zone, species suchas Galactia striata, Mimosa pudica, Chamaecrista nictitans and Calopogoniummucunoides became dominant early in the study and remained so throughout theperiod. However, in two D1 plots (D1b and D1d) the cover of Tithonia rotundifoliaand Stizolobium pruriens increased in the last year.
8 E. Velázquez, A. Gómez-Sal
Changes in Plant Strategies
Six “plant strategies” (groups of species with associated traits) were detected in thelandslide: graminoids with rhizomes and stolons (27 species), annual forbs (14species), perennial forbs (four species), perennial nitrogen-fixing forbs (six species),seedlings (20 species) and nitrogen-fixing seedlings (six species). Mean total coverof annual forbs differed significantly between zones (Table 3g), showing the highestvalues in the Transitional zone (Fig. 2c). Mean total cover of graminoids differed
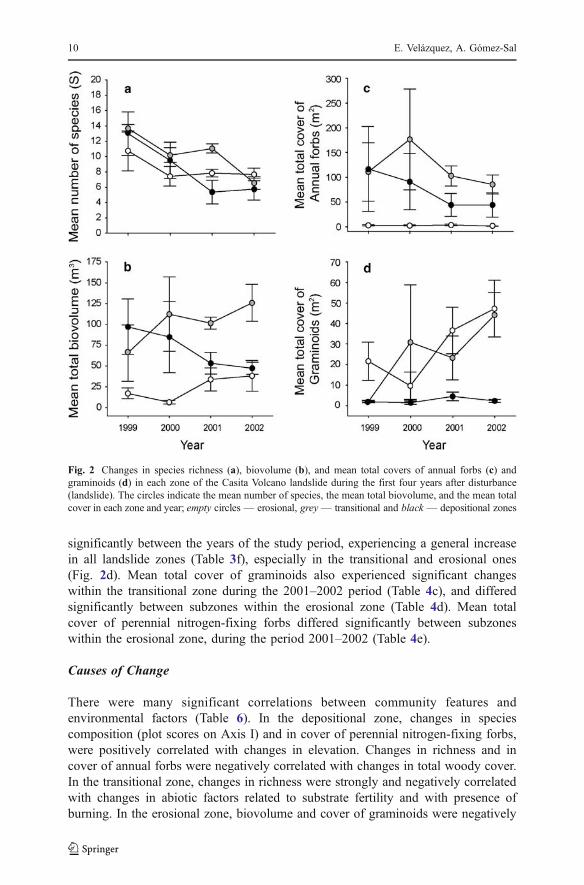
Table 3 Summary table of the repeated-measures ANOVAs for Mean number of species (a), Mean totalbiovolume (b), Mean S�rensen’s Dissimilarity index (c), Mean coordinates in the Axes I (d) and II (e) ofthe DCA, and Mean total covers of graminoids (f), Annual forbs (g), Perennial forbs (h), Perennialnitrogen-fixing forbs (i) and Seedlings (j).“Zone” refers to erosional, transitional and depositional zones,and “Time” includes the four study years
Source Factors and interactions Source
(a) Between-subjects N d.f. F P (f) N d.f. F PZone 2 1.04 0.4305 2 2.05 0.2434Error 4 4Within-subjects N d.f. D d.f. P * N d.f. D d.f. P*Time 2.36 9.44 0.0112 2.87 11.50 0.0034Time*Zone 4.72 9.44 0.0403 5.75 11.50 0.1712Error 12 12
(b) Between-subjects N d.f. F P (g) N d.f. F PZone 2 4.03 0.1099 2 9.16 0.0320Error 4 4Within-subjects N d.f. D d.f. P * N d.f. D d.f. PTime 3 12 0.8890 3 2 0.3372Time*Zone 6 12 0.0098 6 4 0.0986Error 12 12
(c) Between-subjects N d.f. F P (h) N d.f. F PZone 2 0.47 0.6512 2 1.09 0.4184Error 4 4Within-subjects N d.f. D d.f. P N d.f. D d.f. P *Time 2 3 0.3010 2.13 8.54 0.1568Time*Zone 4 6 0.3502 4.27 8.54 0.2978Error 8 12
(d) Between-subjects N d.f. F P (i) N d.f. F PZone 2 19,18 0.0089 2 0.32 0.73Error 4 4Within-subjects N d.f. D d.f. P N d.f. D d.f. PTime 3 2 0.0857 3 2 0.2103Time*Zone 6 4 0.4229 6 4 0.3601Error 12 12
(e) Between-subjects N d.f. F P (l) N d.f. F PZone 2 0.43 0.6724 2 1.12 0.4083Error 4 4Within-subjects N d.f. D d.f. P N d.f. D d.f. PTime 3 2 0.1195 3 2 0.0555Time*Zone 6 4 0.5388 6 4 0.0621Error 12 12
Notes: P-values for between-subjects factors are based on F-values, and P-values for within-subjects arebased on results of Wilk’s lambda values when the sphericity assumption is met and on results of theHuynh and Feldt (1976) epsilon adjustment when it is not (P *).
Herbaceous communities during early succession on a landslide 9
significantly between the years of the study period, experiencing a general increasein all landslide zones (Table 3f), especially in the transitional and erosional ones(Fig. 2d). Mean total cover of graminoids also experienced significant changeswithin the transitional zone during the 2001–2002 period (Table 4c), and differedsignificantly between subzones within the erosional zone (Table 4d). Mean totalcover of perennial nitrogen-fixing forbs differed significantly between subzoneswithin the erosional zone, during the period 2001–2002 (Table 4e).
Causes of Change
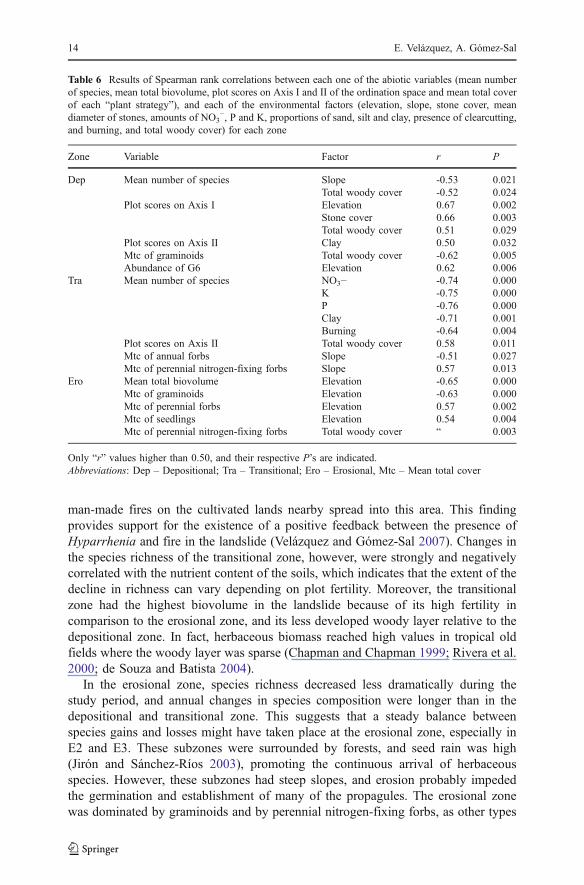
There were many significant correlations between community features andenvironmental factors (Table 6). In the depositional zone, changes in speciescomposition (plot scores on Axis I) and in cover of perennial nitrogen-fixing forbs,were positively correlated with changes in elevation. Changes in richness and incover of annual forbs were negatively correlated with changes in total woody cover.In the transitional zone, changes in richness were strongly and negatively correlatedwith changes in abiotic factors related to substrate fertility and with presence ofburning. In the erosional zone, biovolume and cover of graminoids were negatively
Fig. 2 Changes in species richness (a), biovolume (b), and mean total covers of annual forbs (c) andgraminoids (d) in each zone of the Casita Volcano landslide during the first four years after disturbance(landslide). The circles indicate the mean number of species, the mean total biovolume, and the mean totalcover in each zone and year; empty circles — erosional, grey — transitional and black — depositional zones
10 E. Velázquez, A. Gómez-Sal
correlated with elevation, but changes in covers of perennial forbs and seedlingswere positively correlated with this factor.
Discussion
During the first four years after disturbance, changes in species richness and biovolumein the herbaceous communities of the Casita Volcano landslide differed significantlyamong the areas within the landslide defined by abiotic heterogeneity and landscapecontext. Species composition and mean total cover of annual forbs did not changesignificantly throughout time, but they differed significantly among the differentlandslide zones. These results indicate that environmental heterogeneity played a majorrole in the early succession of herbaceous communities on the landslide, but alsohighlight the difficulties in identifying distinct patterns of change in species composition(Frelich et al. 2003). Species composition in the herbaceous communities of thelandslide was strongly influenced by a few species (Calopogonium mucunoides,Stizolobium pruriens, Clitoria ternatea, Tithonia rotundifolia, Galactia striata, Zorniathymifolia, Hyparrhenia rufa and Sporobolus indicus), which were characteristic of
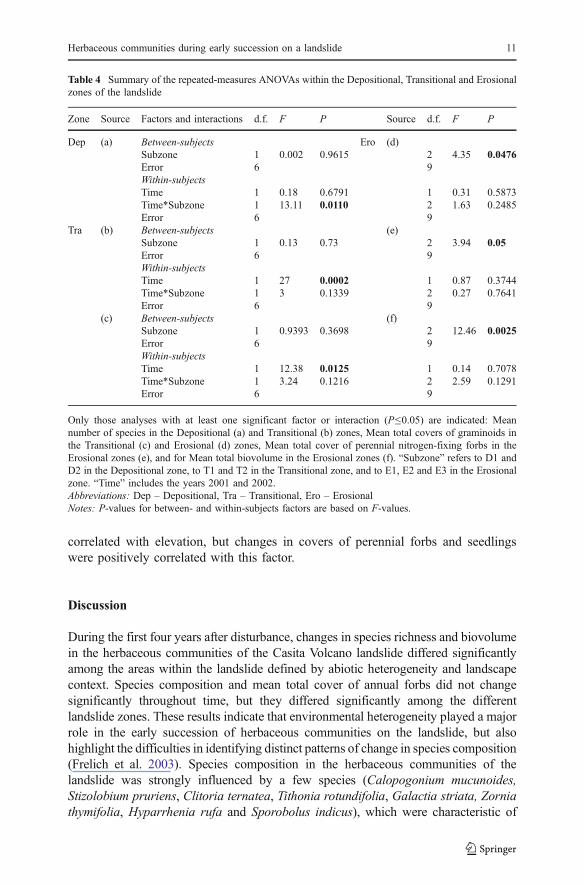
Table 4 Summary of the repeated-measures ANOVAs within the Depositional, Transitional and Erosionalzones of the landslide
Zone Source Factors and interactions d.f. F P Source d.f. F P
Dep (a) Between-subjects Ero (d)Subzone 1 0.002 0.9615 2 4.35 0.0476Error 6 9Within-subjectsTime 1 0.18 0.6791 1 0.31 0.5873Time*Subzone 1 13.11 0.0110 2 1.63 0.2485Error 6 9
Tra (b) Between-subjects (e)Subzone 1 0.13 0.73 2 3.94 0.05Error 6 9Within-subjectsTime 1 27 0.0002 1 0.87 0.3744Time*Subzone 1 3 0.1339 2 0.27 0.7641Error 6 9
(c) Between-subjects (f)Subzone 1 0.9393 0.3698 2 12.46 0.0025Error 6 9Within-subjectsTime 1 12.38 0.0125 1 0.14 0.7078Time*Subzone 1 3.24 0.1216 2 2.59 0.1291Error 6 9
Only those analyses with at least one significant factor or interaction (P≤0.05) are indicated: Meannumber of species in the Depositional (a) and Transitional (b) zones, Mean total covers of graminoids inthe Transitional (c) and Erosional (d) zones, Mean total cover of perennial nitrogen-fixing forbs in theErosional zones (e), and for Mean total biovolume in the Erosional zones (f). “Subzone” refers to D1 andD2 in the Depositional zone, to T1 and T2 in the Transitional zone, and to E1, E2 and E3 in the Erosionalzone. “Time” includes the years 2001 and 2002.Abbreviations: Dep – Depositional, Tra – Transitional, Ero – ErosionalNotes: P-values for between- and within-subjects factors are based on F-values.
Herbaceous communities during early succession on a landslide 11
the different groups of plant strategies. These species became highly dominant in thedifferent zones of the landslide, which caused a general decline in species richnessover time in all zones. However, mainly exotic and wind-dispersed plants colonizedthe landslide. These findings coincide with those of Lundgren (1978) for a group ofsmall landslides in Tanzania that were also located in a densely populated area with atropical dry climate.
Despite the existence of these general trends, the landslide created many zones inwhich changes in the herbaceous communities were different. In the depositionalzone, the decline in species richness was highest. Species richness of herbaceouscommunities can decrease sharply in fertile habitats where strong competition forsoil resources occurs among species (Pausas and Austin 2001). However, in thedepositional zone, biovolume also decreased and total woody cover was an importantcause of change, which points to the competitive elimination of the opportunistic,shade-intolerant species that colonized the area just after the disturbance in 1998 asanother cause of the sharp richness decline. The establishment of woody species andan increase in canopy cover may have been driven this. Similar patterns are apparentin the herbaceous communities of other landslides (e.g. Nakashizuka et al. 1993),abandoned pastures (Holl et al. 2000) and reforested sites (Ruiz-Jaen and Aide 2005)throughout the Neotropics. Changes in species richness differed significantly betweensubzones D1 and D2 toward the end of the study period. This change happenedbecause in D2 species richness increased slightly due to the appearance of somespecies usually found in the understorey of local forests (e.g., Oplismenum burmanii).
Table 5 Species present in >20% of the inventories in the Casita Volcano landslide during the first fouryears after disturbance; number of plots in which they were recorded, total cover in the landslide (m2), andtheir plant traits
Species N° ofplots
Total cover(m2)
Growthform
Veget.spr.
Morph. Long. N2 fix. Res. Disp. Geog.
Calopogoniummucunoides
14 1249.2 forb no vine annual no yes wind exotic
Chamaecristanictitans
11 198 forb no erect annual no yes wind exotic
Clitoria ternatea 13 665.1 forb no creep. per. yes yes wind exoticCyperus odoratus 8 26.8 gram. no erect per. no no wind exoticGalactia striata 13 242.4 forb no vine annual yes no wind exoticHyparrhenia rufa 14 365.9 gram. yes erect per. no yes wind exoticMimosa pudica 8 47.3 forb no erect annual yes no wind exoticPityrogrammacalomalanos
10 131.4 fern yes erect per. no no wind native
Sporobolusindicus
11 66.7 gram. yes erect per. no no wind exotic
Stizolobiumpruriens
7 673.8 forb no vine annual yes yes wind exotic
Tithoniarotundifolia
10 284.4 forb no erect annual no yes wind exotic
Zornia thymifolia 12 380.2 forb yes erect per. yes yes wind native
Abbreviations: Veget. spr. – Vegetative spreading mechanisms; Morph. – Morphology of the shoots; Long.– Longevity; N2 fix. – Ability to fix N2; Res. – Resprouting ability; Disp. – Dispersal mode; Geog. –Geographic origin; gram. – graminoid; creep. – creeping-shoots; per. – perennial.
12 E. Velázquez, A. Gómez-Sal
The number of shade-tolerant species is higher in the understorey of forests that have ahigh species richness (Chinea 1999), and this subzone had higher species richness anda higher percentage of canopy cover in the woody layer than D1 (Velázquez andGómez-Sal 2008).
In the transitional zone, species richness of the herbaceous communities alsodecreased over time; however, biovolume increased significantly. Annual forbs suchas Tithonia rotundifolia, Calopogonium mucunoides, and Stizolobium pruriensdeveloped a large cover in this zone. These species precluded germination andestablishment of other species, as shown for other tropical landslides (Walker 1994)and abandoned pastures (Aide et al. 1995; Slocum et al. 2006). Hyparrhenia rufa, anexotic perennial grass cited as an aggressive colonizer of degraded areas in CentralAmerica (Daubenmire 1972), was also abundant in the transitional zone. Presence ofinvasive grasses impedes forest recovery (Sarmiento 1997; Dalling and Denslow1998; Slocum 2000) and strongly affects the pattern of fire spread (Mistry andBerardi 2005) in abandoned pastures of tropical areas. Specifically, Hyparrheniarufa, is well known for promoting grass-fire cycles through which it becomesprogressively more abundant (D’Antonio and Vitousek 1992). Abundance ofHyparrhenia rufa increased in all landslide zones during the study period, but theincrease was especially high in the transitional zone between 2001 and 2002, when
Fig. 3 DCA scores of the plots surveyed from 1999 to 2002 on the Casita Volcano landslide, Nicaragua.Species present in >20% of the inventories were used in the analysis. The circles represent the speciescomposition of each plot in the four study years (1999, 2000, 2001, and 2002). Lines represent changes inthe species composition of the plots located within each subzone in each year. The lines that correspond tothe last year (2001–2002) end with an arrow, and the names of the plots are indicated at the ends of thearrows. Crosses represent the species that were present in more than 20% of the plots. Most species namesare indicated, with the exception of some that are indicated by numbers; Mimosa pudica (1), Chamaecristanictitans (2), Zornia thymifolia (3), Sporobolus indicus (4) and Calopogonium mucunoides (5). Emptycircles — erosional, grey — transitional and black — depositional zones
Herbaceous communities during early succession on a landslide 13
man-made fires on the cultivated lands nearby spread into this area. This findingprovides support for the existence of a positive feedback between the presence ofHyparrhenia and fire in the landslide (Velázquez and Gómez-Sal 2007). Changes inthe species richness of the transitional zone, however, were strongly and negativelycorrelated with the nutrient content of the soils, which indicates that the extent of thedecline in richness can vary depending on plot fertility. Moreover, the transitionalzone had the highest biovolume in the landslide because of its high fertility incomparison to the erosional zone, and its less developed woody layer relative to thedepositional zone. In fact, herbaceous biomass reached high values in tropical oldfields where the woody layer was sparse (Chapman and Chapman 1999; Rivera et al.2000; de Souza and Batista 2004).
In the erosional zone, species richness decreased less dramatically during thestudy period, and annual changes in species composition were longer than in thedepositional and transitional zone. This suggests that a steady balance betweenspecies gains and losses might have taken place at the erosional zone, especially inE2 and E3. These subzones were surrounded by forests, and seed rain was high(Jirón and Sánchez-Ríos 2003), promoting the continuous arrival of herbaceousspecies. However, these subzones had steep slopes, and erosion probably impededthe germination and establishment of many of the propagules. The erosional zonewas dominated by graminoids and by perennial nitrogen-fixing forbs, as other types
Table 6 Results of Spearman rank correlations between each one of the abiotic variables (mean numberof species, mean total biovolume, plot scores on Axis I and II of the ordination space and mean total coverof each “plant strategy”), and each of the environmental factors (elevation, slope, stone cover, meandiameter of stones, amounts of NO3
−, P and K, proportions of sand, silt and clay, presence of clearcutting,and burning, and total woody cover) for each zone
Zone Variable Factor r P
Dep Mean number of species Slope -0.53 0.021Total woody cover -0.52 0.024
Plot scores on Axis I Elevation 0.67 0.002Stone cover 0.66 0.003Total woody cover 0.51 0.029
Plot scores on Axis II Clay 0.50 0.032Mtc of graminoids Total woody cover -0.62 0.005Abundance of G6 Elevation 0.62 0.006
Tra Mean number of species NO3− -0.74 0.000K -0.75 0.000P -0.76 0.000Clay -0.71 0.001Burning -0.64 0.004
Plot scores on Axis II Total woody cover 0.58 0.011Mtc of annual forbs Slope -0.51 0.027Mtc of perennial nitrogen-fixing forbs Slope 0.57 0.013
Ero Mean total biovolume Elevation -0.65 0.000Mtc of graminoids Elevation -0.63 0.000Mtc of perennial forbs Elevation 0.57 0.002Mtc of seedlings Elevation 0.54 0.004Mtc of perennial nitrogen-fixing forbs Total woody cover “ 0.003
Only “r” values higher than 0.50, and their respective P’s are indicated.Abbreviations: Dep – Depositional; Tra – Transitional; Ero – Erosional, Mtc – Mean total cover
14 E. Velázquez, A. Gómez-Sal

of unstable and infertile areas (Olander et al. 1998; Tsuyuzaki and Hase 2005),during early succession. Zornia thymifolia and Sporobolus indicus were the mostabundant graminoids. However, significant differences in mean total cover ofgraminoids between subzones E1, E2 and E3 during the period 2001–2002 wererelated to the large cover attained by Hyparrhenia rufa in E1. These differencessuggest that this species is migrating up from the transitional zone toward theerosional zone, in the uppermost part of the landslide. The existence of significantdifferences in mean total cover of perennial nitrogen-fixing forbs between erosionalsubzones was due to the high increase in cover of Clitoria ternatea. This species hascreeping shoots and can propagate vegetatively. It formed patches that rapidly grewand combined to develop dense ground-level thickets (E. Velázquez, pers.observation). Herbaceous thickets can arrest succession by impeding the establish-ment of woody seedlings (Zhou et al. 2003; Slocum et al. 2006), but they can beimportant for amelioration of fertility and stabilization of slopes, thereby promotinggermination of subsequent colonists (Walker 1994). Although Clitoria ternateaprobably precludes colonization by woody plants, it might have a long-term positiveeffect on succession not only by stabilizing the substrates but also by improving theirnutrient content.
Our study suggests that in large landslides located in highly humanized areas witha tropical dry climate, changes in the herbaceous communities during the first yearsof succession are strongly influenced by abiotic heterogeneity and landscape context,and are mainly controlled by the cover of the woody layer, abiotic factors associatedwith fertility of substrates, and the incidence of human disturbances. Likewise,changes in community features are due to the expansion of a few dominant speciesthat developed a large cover and excluded other species. These species may arrestsuccession, contribute to the amelioration of soil fertility, or increase the frequencyof human-induced disturbances by promoting grass-fire cycles. Consequently,herbaceous communities are important in determining future successional patternson the Casita Volcano landslide.
Acknowledgements This research was part of the collaborative program between the Universidad deAlcalá and the Universidad Nacional Autónoma de Nicaragua-León, and supported by a grant from theThird Scientific Research and Technological Innovation Program of the Regional Authority of Madrid.Pedrarias Dávila and Aníbal Rodríguez of the Universidad Nacional Autónoma de Nicaragua-León wereextremely helpful with the fieldwork. Ricardo Rueda, Dania Paguaga, and Franklin Flores helped us toidentify and assign plant traits to the species. We are very grateful to the Sevilla family in the communityof Pikin Guerrero who provided accommodations and supported our fieldwork. We are very grateful toPilar Castro-Díez for her comments on the earlier versions of the manuscript and to Bruce McWhirther forimproving the English.
References
Aide TM, Zimmerman JK, Herrera L, Rosario M, Serrano M (1995) Forest recovery in abandoned tropicalpastures in Puerto-Rico. Forest Ecol Managem 77:77–86
Bakker JP, Olff H, Willems JH, Zobel M (1996) Why do we need permanent plots in the study of long-term vegetation dynamics? J Veg Sci 7:147–156
Benitez-Malvido J (2006) Effect of low vegetation on the recruitment of plants in successional habitattypes. Biotropica 38:171–182
Herbaceous communities during early succession on a landslide 15

Bonet A, Pausas JG (2004) Species richness and cover along a 60-year chronosequence in old-fields ofsoutheastern Spain. Pl Ecol 174:257–270
Bush MB, Whittaker RJ (1995) Colonization and succession on Krakatu: an analysis of the guild of viningplants. Biotropica 27:355–372
CEPAL (1999) Nicaragua, evaluación de los daños ocasionados por el huracán Mitch, 1998.Implicaciones para el desarrollo económico y social y el medio ambiente. Naciones Unidas,Comisión Económica para América Latina y el Caribe (CEPAL), México DF
Chapman CA, Chapman LJ (1999) Forest restoration in abandoned agricultural land: a case study fromEast Africa. Conservation Biol 13:1301–1311
Chinea JD (1999) Changes in the herbaceous and vine communities at the Bisley ExperimentalWatersheds, Puerto Rico, following Hurricane Hugo. Canad J Forest Res 29:1433–1437
Corrales-Rodríguez D (1983) Impacto ecológico sobre los recursos naturales renovables de Centroa-mérica. Instituto de los Recursos Naturales y del Ambiente (IRENA), Managua
Dalling JW, Denslow JS (1998) Soil seed bank composition along a forest chronosequence in seasonallymoist tropical forest, Panama. J Veg Sci 9:669–678
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass-fire cycle andglobal change. Annual Rev Ecol Syst 23:674–689
Daubenmire R (1972) Ecology of Hyparrhenia-rufa (Nees) in derived savanna in northwestern Costa-Rica. J Appl Ecol 9:11–23
de Souza FM, Batista JLF (2004) Restoration of seasonal semideciduous forests in Brazil: influence of ageand restoration design on forest structure. Forest Ecol Managem 191:185–200
Frelich LE, Machado JL, Reich PB (2003) Fine scale environmental variation and structure of understoryplant communities in two old-growth pine forests. J Ecol 91:283–293
Gillespie TW, Grijalva A, Farris CN (2000) Diversity, composition and structure of tropical dry forests inCentral America. Pl Ecol 147:37–47
Gómez-Sal A, de Miguel JM, Casado MA, Pineda FD (1986) Successional changes in the morphologyand ecological responses of a grazed pasture ecosystem in Central Spain. Vegetatio 67:33–44
Grace JB (1999) The factors controlling species density in herbaceous plant communities: an assessment.Perspect Pl Ecol Evol Syst 2:1–28
Guariguata MR (1990) Landslide disturbance and forest regeneration in the upper Luquillo Mountains ofPuerto Rico. J Ecol 78:814–832
Holl KD, Loik ME, Lin EHV, Samuels IA (2000) Tropical montane forest restoration in Costa Rica:Overcoming barriers to dispersal and establishment. Restoration Ecol 8:339–349
Huynh H, Feldt LS (1976) Estimation of the Box correction for degrees of freedom from sample data inthe randomized block and split-plot designs. J Educ Statist 1:69–82
Janzen DH (1988) Tropical dry forests, the most endangered major tropical ecosystem. In Wilson EO (ed)Biodiversity. National Academy Press, Washington DC, pp 130–137
Jirón FA, Sánchez-Ríos MR (2003) Caracterización de suelos y oferta de semillas en diferentes sectoresdel deslave del Volcán Casita. Thesis de Maestría, Universidad Nacional Autónoma de Nicaragua –León, León
Kent M, Coker P (1992) Vegetation description and analysis: a practical approach. Belhaven, LondonKerle N, de Vries BV, Oppenheimer C (2003) New insight into the factors leading to the 1998 flank
collapse and lahar disaster at Casita volcano, Nicaragua. Bull Volcanology 65:331–345Klinka K, Krajina VJ, Ceska A, Scagel AM (1989) Indicator plants of coastal British Columbia.
University of British Columbia Press, VancouverLeishman MR, Westoby M (1992) Classifying plants into groups on the basis of associations of individual
traits- evidence from Australian semi-arid woodlands. J Ecol 80:417–424Lundgren L (1978) Studies of soil and vegetation development on fresh landslide scars in the Mgeta
Valley, Western Uluruga Mountains, Tanzania. Geogr Ann 60:91–120Martin Y, Rood K, Schwab JW, Church M (2002) Sediment transfer by shallow landsliding in the Queen
Charlotte Islands, British Columbia. Canad J Earth Sci 39:189–205Marquínez JM, Dévoli G, Menendes-Duarte R (2002) Deslave del Volcán Casita (Nicaragua). In Ayala-
Carcedo FJ, Olcina J (eds) Riesgos naturales. Ariel Ciencia, Barcelona, pp 307–312McIntyre S, Lavorel S, Tremont RM (1995) Plant life-history attributes – their relationship to disturbance
responses in herbaceous vegetation. J Ecol 83:31–44Mistry J, Berardi A (2005) Assessing fire potential in a Brazilian savanna nature reserve. Biotropica
37:439–451Montalvo J, Casado MA, Levassor C, Pineda FD (1991) Adaptation of ecological systems: compositional
patterns of species and morphological and functional traits. J Veg Sci 2:655–666
16 E. Velázquez, A. Gómez-Sal

Myster RW, Fernández DS (1995) Spatial gradients and patch structure on two Puerto Rican landslides.Biotropica 27:149–159
Myster RW, Walker LR (1997) Plant successional pathways on Puerto Rican landslides. J Trop Ecol13:165–173
Nakashizuka T, Lida S, Suzuki W, Tanimoto T (1993) Seed dispersal and vegetation development on adebris avalanche on the Ontake Volcano, Central Japan. J Veg Sci 4:537–542
Olander LP, Scatena FN, Silver WL (1998) Impacts of disturbance initiated by road construction in asubtropical cloud forest in the Luquillo Experimental Forest, Puerto Rico. Forest Ecol Managem109:33–49
Orshan G (1982) Monocharacter growth from types as a tool in an analytic-synthetic study of growthforms in mediterranean type ecosystems. A proposal for an inter-regional program. EcologíaMediterránea 8:159–171
Pausas JG, Austin MP (2001) Patterns of species richness in relation to different environments: anappraisal. J Veg Sci 12:153–166
Pickett STA, White TS (1985) The ecology of natural disturbance and patch dynamics. Academic Press,New York
Pielke RA, Rubiera J, Landsea C, Fernández ML, Klein R (2003) Hurricane vulnerability in LatinAmerica and the Caribbean: Normalized damage and loss potentials. Nat Hazards Rev 4:101–114
Pineda FD, Casado MA, Peco B, Olmeda C, Levassor C (1987) Temporal changes in therophyticcommunities across the boundary of disturbed-intact ecosystems. Vegetatio 71:33–39
Porta J, López-Acevedo M, Roquero C (1999) Edafología para la agricultura y el medio ambiente.Ediciones Mundi-Prensa, Madrid
Restrepo C, Vitousek P (2001) Landslides, alien species and the diversity of a Hawaiian montane mesicecosystem. Biotropica 33:409–420
Rivera LW, Zimmerman JK, Aide TM (2000) Forest recovery in abandoned agricultural lands in a karstregion of the Dominican Republic. Pl Ecol 148:115–125
Roberts MR (2004) Response of the herbaceous layer to natural disturbance in North American forests.Canad J Bot 82:1273–1283
Roberts MR, Zhu LX (2002) Early response of the herbaceous layer to harvesting in a mixed coniferous-deciduous forest in New Brunswick, Canada. Forest Ecol Managem 155:17–31
Rodríguez MA, Borges PAV, Gómez-Sal A (1997) Species and life-forms composition of Mediterraneanmountain pastures in two years of contrasting precipitation. Flora 192:231–240
Ruiz-Jaen MC, Aide TM (2005) Vegetation structure, species diversity, and ecosystem processes asmeasures of restoration success. Forest Ecol Managem 218:159–173
Salas-Estrada JB (1999) Biodiversidad en Nicaragua, un estudio de país. Ministerio de los RecursosNaturales y del Ambiente (MARENA) – Ministerio Agropecuario y Forestal (MAGFOR), Managua
Sarmiento FO (1997) Arrested succession in pastures hinders regeneration of Tropandean forests andshreds mountain landscapes. Environm Conservation 24:14–23
Scott KM, Vallance JW, Kerle N, Macías JL, Strauch W, Devoli G (2005) Catastrophic precipitation-triggered lahar at Casita volcano, Nicaragua: occurrence, bulking and transformation. Earth SurfaceProcesses and Landforms 30:59–79
Sheridan MF (1998) Report on the October 30, 1998 avalanche and breakout flow of Casita Volcano,Nicaragua, triggered by hurricane Mitch. United States Geological Survey (USGS)
Silvertown J, Dodd ME, McConway K, Potts J, Crawley M (1994) Rainfall, biomass variation andcommunity composition in the Park Grass Experiment. Ecology 75:2430–2437
Slocum MG (2000) Logs and fern patches as recruitment sites in a tropical pasture. Restoration Ecol8:408–413
Slocum MG, Aide TM, Zimmerman JK, Navarro L (2006) A strategy for restoration of montane forest inanthropogenic fern thickets in the Dominican Republic. Restoration Ecol 14:526–536
Small CJ, McCarthy BC (2003) Spatial and temporal variability of herbaceous vegetation in an easterndeciduous forest. Pl Ecol 164:37–48
Sokal RR, Rohlf FJ (1995) Biometry. W.H. Freeman, New YorkStatSoft (2001) STATISTICA. Data analysis software system 6. Tulsa, OklahomaStevens WD, Ulloa-Ulloa C, Pool A, Montiel OM (2001) Flora de Nicaragua. Missouri Botanical Garden
Press, St. Louister Braak CJF, Šmilauer P (2002) CANOCO Reference manual and CanoDraw for windows user’s guide:
Software for Canonical Community Ordination (version 4.5). Microcomputer Power, Ithaca NYTsuyuzaki S (1989) Analysis of revegetation dynamics on the volcano Usu, northern Japan, deforested by
1977–1978 eruptions. Amer J Bot 76:1468–1477
Herbaceous communities during early succession on a landslide 17

Tsuyuzaki S, del Moral R (1995) Species attributes in early primary succession on volcanoes. J Veg Sci6:517–522
Tsuyuzaki S, Hase A (2005) Plant community dynamics on the Volcano Mount Koma, northern Japan,after the 1996 eruption. Folia Geobot 40:319–330
UNDP (2005) Human development report 2005. United Nations Development Program (UNDP), NewYork
Velázquez E, Gómez-Sal A (2007) Environmental control of early succession on a large landslide in atropical dry ecosystem (Casita Volcano, Nicaragua). Biotropica 39:601–609
Velázquez E, Gómez-Sal A (2008) Landslide early succession in a neotropical dry forest. Pl Ecol199:295–308
von Ende CN (2001) Repeated-measures analysis, growth and other time-dependent measures. In ScheinerSM, Gurevitch J (eds) Design and analysis of ecological experiments. Oxford University Press,Oxford, pp 134–157
Walker LR (1994) Effects of fern thickets on woodland development on landslides in Puerto Rico. J VegSci 5:525–532
Walker LR, Willig MR (1999) An introduction to terrestrial disturbances. In Walker LR (ed) Ecosystems ofdisturbed ground. Elsevier, Amsterdam, pp 1–16
Walker LR, Zarin DJ, Fetcher N, Myster RW, Johnson AH (1996) Ecosystem development and plantsuccession on landslides in the Caribbean. Biotropica 28:566–576
Whittaker RJ, Bush MB, Richards K (1989) Plant recolonization and vegetation succession on the KrakatuIslands, Indonesia. Ecol Monogr 59:59–123
Zhou ZH, Miwa M, Nara K, Wu B, Nakaya H, Lian C, Miyashita N, Oishi R, Maruta E, Hogetsu T (2003)Patch establishment and development of a clonal plant, Polygonum cuspidatum, on Mount Fuji.Molec Ecol 12:1361–1373
Received: 19 March 2007 /Revised: 14 October 2008 /Accepted: 20 October 2008 /Published online: 4 March 2009
18 E. Velázquez, A. Gómez-Sal




















