
DIE GEDRAGSEKOLOGIE VAN DIE OORBIETJIE OUREBIA ...
302
DIE GEDRAGSEKOLOGIE VAN DIE OORBIETJIE OUREBIA OUREBI OUREBI (ZIMMERMANN, 1783) IN TRANSVAAL deur PETRUS CORNELIUS VILJOEN Voorgele ter vervulling van 'n deel van die vereistes vir die graad M.Sc. (Dierkunde) in die Fakulteit Wis- en Natuurkunde Departement Dierkunde Universiteit van Pretoria PRETORIA APRIL 1982 © University of Pretoria
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of DIE GEDRAGSEKOLOGIE VAN DIE OORBIETJIE OUREBIA ...

DIE GEDRAGSEKOLOGIE VAN DIE OORBIETJIE OUREBIA OUREBI
OUREBI (ZIMMERMANN, 1783) IN TRANSVAAL
deur
PETRUS CORNELIUS VILJOEN
Voorgele ter vervulling van 'n deel
van die vereistes vir die graad
M.Sc. (Dierkunde)
in die
Fakulteit Wis- en Natuurkunde
Departement Dierkunde
Universiteit van Pretoria
PRETORIA
APRIL 1982
©© UUnniivveerrssiittyy ooff PPrreettoorriiaa

DIE GEDRAGSEKOLOGIE VAN DIE OORBIETJIE
OUREBIA OUREBI OUREBI (ZIMMERMANN, 1783)
IN TRANSVAAL
deur
PETRUS CORNELIUS VILJOEN
Leiers Prof. J.D. Skinner
Departement Dierkunde, Universiteit van Pretoria
Prof. G.K. Theron
Departement Plantkunde, Universiteit van Pretoria
UITTREKSEL
Die oorbietjie se basiese aktiwiteitspatroon word deur faktore soos reen
en veeversteurings bernvloed. Oorbietjies is gemiddeld 34,2% van
dagligperiodes aktief terwyl 83,6% van die aktiewe periodes aan wei
gespandeer word.
Oorbietjies kom in familiegroepe voor. Die ramme verdedig reg deur die
jaar n territorium wat met die tuisgebied oorvleuel en n gemiddelde
grootte van 34 ha in die Amsterdam-studiegebied gehad het.

Die lamseisoenpiek duur van November tot Januarie. Oorbietjielammers
beweeg eers na n periode van drie tot vier rnaande saam met die familie
groep.
Oorbietjies is hoofsaaklik grasvretend en n hoe voorkeur word vir veral
Eulalia villosa en Monocymbium ceresiiforrne getoon.
Oorbietjiedigthede van tot 4,56 oorbietjies/km2 is gevind. Mens like
besetting, plantasies en landerye bernvloed oorbietjieverspreiding
terwyl voorkeure vir plato-uitlopers getoon word. Kleinskaalse
bewegings kom voor wat veral deur brande veroorsaak word.

(iii)
DANKBETUIGINGS
Graag wil ek my dank teenoor die volgende persone en instansies uitspreek:
Prof. J.D. Skinner, Departement Dierkunde, en dr. G.K. Theron, Departement
Plantkunde aan die Universiteit van Pretoria wat as projekleiers opgetree
het. Hulle belangstelling, behulpsaamheid en aanmoediging word opreg
waardeer.
Die Afdeling Natuurbewaring van die Transvaalse Provinsiale Administrasie
vir die geleentheid om die navorsingsresultate te kon gebruik. Dr. S.M.
Hirst, voorheen Assistent-direkteur (Navorsing), se persoonlike belang
stelling en aanmoediging word waardeer. Alle beamptes van die Afdeling
het waardevolle bydraes gemaak; in besonder:
Mnr. N.HoGo Jacobson, Senior Vakkundige Beampte, vir die identifisering
van vele planbmonsters.
Mnre. N. Zambatis, N.o.s. van Rensburg en J. van Jaarsveld, Natuurbewarings
beamptes, vir hulp tydens die tellings en sommige ander fases van die
veldwerk.
Mnr. H.Jo Meyer, Senior Natuurbewaringsbeampte, wat behulpsaam was met die
oprigting van kampe en vele ander aspekte.
Mnr. J.W.H. Ferguson, Senior Vakkundige Beampte, vir hulp en advies met
die telemetriese opsporings en verwerking van sekere data.
Wyle mej. CoCoGo van der Walt, Bibliotekaresse, vir haar persoonlike
belangstelling, aanmoediging en spesiale hulp wat verleen is met vertaal
werk asook die verkryging van literatuur.
Alle eienaars vir hul samewerking en in besonder:
Mnr. en mev. D. Forbes van Forbes Athole, Amsterdam, vir al bulle belang
stelling en ga&vryheid tydens my veldwerk, mnr. en mev. M. Devenish van

(iv)
Mooihoek, mnr$ en mev. J.Ao Labuschagne van Utopia en mnr. en mev. S.F.
Klingenberg van Rooi Kraal, mnr. G. Khors van Uitgevallen, mnr. R. Klingen
berg van Driehoek en Rooi Kraal, mnr. W. de V. Scheepers van Sandbach en
mnr. NoJo Devenish van Wakkerstroom vir sy bydrae tot sekere plantopnames.
Mnr. R. Kotze, Senior Tegnikus, Nooitgedacht-navorsingstasie vir sy belang
stelling en hulp tydens die plantopnames.
Mnr. J.Ho Roos, voorheen van die Nooitgedacht-navorsingstasie, vir die
beskikbaarstelling van ongepubliseerde data.
Mnr. J. Booysen, Lektor, Departement Aardrykskunde, Universiteit van Pretoria,
vir hulp en belangstelling met die geomorfologiese gebiedsindelings.
Die personeel van die Nasionale Herbarium, Pretoria, wat met die identifi
sering van verskeie plantmonsters behulpsaam was.
Dr. R.HoN. Smithers, Senior Navorsingsbeampte Soogdiernavorsingsinstituut,
vir die beskikbaarstelling van verspreidingsdata oor oorbietjies in Zimbabwe.
Mnr. P.Ho Lloyd, Departement Natuur- en Omgewingsbewaring, Kaapse Provin
siale Administrasie, vir inligting oor die verspreiding van oorbietjies in
die Kaapprovinsie.
Mnr. Keith Joubert, Hoedspruit,vir sy bydrae met die voorbereiding· van
sekere figure.
Mnr. William Massyn, voorheen fotograaf van die Afdeling Natuurbewaring,
tans verbonde aan die Nasionale Parkeraad te Pretoria, vir raad en aan
moediging met veral die fotografie.

INHOUDSOPGAWE
HOOFSTUK
1.
2.
3.
4.
UITTREKSEL
DANKBETUIGINGS
INHOUDSOPGAWE
LYS VAN TABELLE
LYS VAN FIGURE
INLEIDING
TAKSONOMIE EN MORFOLOGIE
Taksonomie
Morfo1ogie
GEOGRAFIESE VERSPREIDING
A1gemeen
Vroee Verspreiding
Huidige Verspreiding in Suide1ike Afrika
TRANSVAAL
NATAL
ORANJE-VRYSTAAT
KAAPPROVINSIE
TRANSKEI
ZIMBABWE
MOSAMBIEK
BOTSWANA
SUIDWES-AFRIKA
SWAZILAND, LESOTHO EN ANGOLA
STUDIEGEBIEDE
In1eiding
Ligging en Grootte van Studiegebiede
AMSTERDAM-STUDIEGEBIED
(v)
BLADSY
(i)
(iii)
(v)
(xii)
(xvii)
1
4
4
4
6
6
6
7
7
7
9
9
9
9
10
10
10
10
12
12
12
12

(vi)
BLADSY
PIET RETIEF-STUDIEGEBIED 16
Fisiografie 16
AMSTERDAM-STUDIEGEBIED 16
PIET RETIEF-STUDIEGEBIED 19
K1imaa t 19
ALGEMEEN 19
SONSKYNDUUR EN WOLKBEDEKKING 21
TEMPERATUUR 21
RELATIEWE LUGVOGTIGHEID 25
WIND 25
REENVAL 25
P1antegroei 28
AMSTERDAM- EN PIET RETIEF~STUDIEGEBIEDE 28
Vroee Geskiedenis en Bestuur 29
AMSTERDAM-STUDIEGEBIED 29
PIET RETIEF-STUDIEGEBIED 30
5. GEOMORFOLOGIESE GEBIEDSINDELING 32
In1eiding 32
Metodes 32
Resu1 tate 33
AMSTERDAM-STUDIEGEBIED 33
Algemeen 33
Plato-uit1oper 37
P1atoberggebied 37
Ta11ushang 37
Pediment 39
Rivierva11ei 39
PIET RETIEF-STUDIEGEBIED 40

(vii)
BLADSY
Algemeen 40
Plato-uitloper 40
Eskarp 44
Platotraphang 44
Tallushang 44
Pediment 44
Riviervallei 46
Bespreking 46
6. DIE PLANTGEMEENSKAPPE 48
Inleiding 48
Metodes 48
ALGEMEEN 48
MONSTERNEMING 49
WAARNEMINGS IN EN OM MONSTERPERSELE 52
DATAVERWERKING 55
Resultate 56
AMSTERDAM-STUDIEGEBIED 56
Die Berkheya setifera - Vernonia natalensis -Eulalia villosa - grasveld 56
Die Andropogon eucomus - Helichrysurn sp. -oulandgemeenskap 63
Die Paspalrn comrnersonii - Hyparrhenia hirta -Helichrysurn sp. - grasveld 64
Die Hyparrhenia hirta - Eragrostis racernosa -Cyperaceae - grasveld 65
Die Trachypogon spicatus - Alloteropsis sernialata - Monocymbium ceresiiforme - grasveld 66
Die Loudetia simplex - Indigofera sp. -Berkheya setifera - grasveld 70
Die Eragrostis racemosa - Erioserna pauciflorum -Heteropogon contortus - grasveld 71

(viii)
Bladsy
Die Tristachya hispida - Berkheya setifera -Panicum natalense - grasveld 73
Die Vlei- en Riviergemeenskappe 74
Die Plantasies en ander Uitheemse Plante 76
PIET RETIEF-STUDIEGEBIED 77
Die Xerophyta sp. - Oldenlandia herbacea -klipplaatplantegroei 77
Die Aloe marlothii - Crassula sp. - struikveld 83
Die Loudetia simplex - Trachypogon spicatus -Rendlia altera - grasveld 88
Die Monocymbium ceresiiforme - Diheteropogon amplectens - Tristachya hispida - grasveld 89
Die Alloteropsis semialata - Trachypogon spica tus - grasveld 90
Die Berkheya sp. - Helichrysum sp. -Tristachya hispida - grasveld 93
Die Gerbera sp. - Eulalia villosa -Eragrostis plana - grasveld 96
Die Kloofbosgemeenskap 99
Die Vleigemeenskap 101
Die Plantasies en ander Uitheemse Plante 101
Bespreking en Gevolgtrekking : Amsterdam- en Piet Retief-studiegebiede 102
MONSTERNEMING 102
DATAVERWERKING 103
PLANTEGROEI 103
7. VOEDING EN WATERBEHOEFTES 106
Inleiding 106
Metodes 106
VOEDSELVOORKEURE 106
VOEDINGSGEDRAG 107

8.
9.
WATERBEHOEFTES
Resu1tate en Bespreking
VOEDSELVOORKEURE
VOEDINGSGEDRAG
WATERBEHOEFTES
AKTIWITEITSPATRONE
In1eiding
Metodes
Resu1tate en Bespreking
BASIESE AKTIWITEITSPATROON
WEI-AKTIWITEITE
LYFKOSINGSAKTIWITEITE
URINERING, ONTLASTING EN MERK
LE SOSIALE AKTIWITEITE
AKTIWITEITE GEDURENDE DIE NAG ..
FAKTORE WAT AKTIWITEITE BEINVLOED
Versteurings
Inv1oed van weersomstandighede
Gevo1gtrekking
SOSIALE ORGANISASIE
In1eiding
Metodes
GROEPSAMESTELLING
GEDRAG
Resu1late en Bespreking
BEVOLKINGSTRUKTUUR EN GROEPSAMESTELLING
(ix)
B1adsy
108
108
108
113
116
122
122
122
124
124
128
131
131
131
136
136
140
140
140
143
144
144
145
145
146
147
147

Amsterdam-studiegebied
Piet Retief-studiegebied
A1gemeen
SOSIALE GEDRAG
Groepsbande
Dominansie
Submisiwiteit
TERRITORIALE GEDRAG
Territoria1e Ste1se1 en Territoriumgrootte
Afbakening
Verdediging
VOORTPLANTING EN REPRODUKSIE
Voortp1antingsgedrag
Reproduksie
Lamseisoen
10. INTERSPESIEVERHOUDINGS
In1eiding
Metodes
Resu1tate en Bespreking
ALGEMENE INTERAKSIES
VLUGREAKSIES EN ANTIROOFDIERVERHOUDINGS
Groepgrootte
Primere Verdedigingsmeganismes
Sekondere Verdedigingsmeganismes
V1ugafstande
11. HABITATVOORKEURE
In1eiding
Metodes
(x)
B1adsy
147
153
155
160
160
161
165
166
166
170
185
188
188
194
194
203
203
203
204
204
207
207
208
210
219
223
223
224

(xi)
Bladsy
KARTERING 224
OORBIETJIEDIGTHEDE 224
VELDTOESTAND 225
Resultate en Bespreking 226
AMSTERDAM-STUDIEGEBIED 226
Oorbietjieverspreiding en Voorkeurgebiede 226
Seisoenale Veranderinge in Oorbietjiedigt-hede 230
PIET RETIEF-STUDIEGEBIED 236
Oorbietjieverspreiding en Voorkeurgebiede 236
Seisoenale Veranderinge in Oorbietjiedigt- 238 he de
Gevolgtrekking 245
OPSOMMING 249
SUMMARY 251
LITERATUURLYS 253
BYLAES

TABELNR.
1.
LYS VAN TABELLE
Besonderhede van die weerstasies waarvan temperatuur-
en reengegewens verkry is. (Weerburo 1954, 1965, 1967a,
1967b)
2. Temperatuurstatistieke vir die Amsterdam- en Piet
Retief-weerstasies in die omgewing van die Amsterdam
en Piet Retief-studiegebiede. (Weerburo 1954)
3. Die gemidde1de maande1ikse en jaar1ikse reenva1 vir n
aanta1 weerstasies in die omgewing van die Amsterdam
en Piet Retief-studiegebiede.
4. Geomorfo1ogiese eenhede en opperv1aktes wat bes1aan word
in die Amsterdam-studiegebied.
5. Geomorfo1ogiese eenhede en opperv1aktes wat bes1aan word
in die Piet Retief-studiegebied.
6. Die fisiografiese-fisionomiese eenhede in die Amsterdam
studiegebied waarin monsterperse1e versprei is.
7. Die fisiografiese-fisionomiese eenhede in die Piet
Retief-studiegebied waarin monsterperse1e versprei is.
8. Die p1antgemeenskappe van die Amsterdam-studiegebied.
9. n P1antsosio1ogiese tabe1 van die p1antgemeenskappe wat
in die Amsterdam-studiegebied aangetref word.
10. n Uiteensetting van die gemidde1de hoogte en kroon
bedekking van die Berkheya setifera-Vernonia nata1ensis
Eu1a1ia vi11osa -grasveld.
11. n Uiteensetting van die gemidde1de hoogte en kroon
bedekking van die Hyparrhenia hirta-Eragrostis racemosa
Cyperaceae -grasve1d.
(xii)
Bls.
22
23
25
34
41
50
51
57
58
61
67

12. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Trachypogon spicatus-Alloteropsis
semialata-Monocymbium ceresiiforme -grasveld.
13. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Loudetia simplex-Indigofera sp.
Berkheya setifera -grasveld.
14. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Tristachya hispida-Berkheya setifera
Panicum natalense -grasveld.
15. Die plantgemeenskappe van die Piet Retief-studiegebied.
16. n Plantsosiologiese tabel van die plantgemeenskappe wat
in die Piet Retief-studiegebied aangetref word.
17. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Xerophyta sp.-Oldenlandia herbacea
klipplaatplantegroei.
18. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Aloe marlothii-Crassula sp.
struikveld.
19. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Monocymbium ceresiiforme-Diheteropogon
amplectens-Tristachya hispida -grasveld.
20. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Alloteropsis semialata-Trachypogon
spicatus -grasveld.
21. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Berkheya sp.-Helichrysum sp.
Tristachya hispida -grasveld.
22. n Uiteensetting van die gemiddelde hoogte en kroon
bedekking van die Gerbera sp.-Eulalia villosa
Eragrostis plana -grasveld.
(xiii)
69
72
75
78
79
82
85
91
94
97
100

23. Geskatte persentasie benutting deur oorbietjies van
die vernaamste plantspesies in die Eulalia villosa~
Hypoxis rigidula-Berkheya setifera- en die Eulalia
villosa-Pentanisia angustifolia-Setaria splendida -grasveld,
Amsterdam-studiegebied. 112
24. Verskille in sekere voedingsaspekte tussen die oorbiet
jie, blesbok en springbok.
25. Die persentasie verspreiding, met die standaard fout vir
e1ke katagorie, van verskil1ende aktiwiteite van oorbiet
jies gedurende daglig met ~ gemiddelde waarnemingsperiode
van 11,18 h in die Amsterdam-studiegebied.
26. Die verhouding van verskil1ende aktiwiteite tussen oor
bietjie ~~ en ~~ in die Amsterdam-studiegebied met~
gemidde1de en totale waarnemingsperiode van 11,18 hen
113,0 h respektiewelik.
27. Ontlastings- en urineringstempo's van oorbietjies, ~~
sowel as ~~' met~ gemidde1de waarnemingsperiode van
10,48 h in die Amsterdam-studiegebied.
28. Veranderinge in die merktempo's van oorbietjieramme, beide
preorbitaa1 en interdigitaal, gedurende daglig met~
gemidde1de waarnemingsperiode van 11,05 h in die Amsterdam
studiegebied.
29. Resu1tate van vier seisoenale oorbietjietel1ings (1974-
1975) in die Amsterdam-studiegebied.
30. Geslags- en ouderdomsgroepe van oorbietjies in die
Amsterdam-studiegebied gedurende vier seisoenale tellings
vanaf November 1974 tot Oktober 1975.
31. Seisoenale veranderinge van spesifieke oorbietjiegroepe
in verge1yking met die ander groepe in die Amsterdam-studie
gebied : November 1974 tot Oktober 1975.
117
129
130
134
135
148
150
151
(xiv)

32. Resu1tate van vier seisoena1e oorbietjiete11ings (1974-
1975) in die Piet Retief-studiegebied.
33. Ges1ags- en ouderdomsgroepe van oorbietjies in die Piet
Retief-studiegebied gedurende vier seisoena1e te11ings
vanaf November 1974 tot Oktober 1975.
34.
35.
Seisoena1e veranderinge van spesifieke oorbietjiegroepe
in verge1yking met die ander groepe in die Piet Retief
studiegebied : November 1974 tot Oktober 1975.
Besonderhede van oorbietjiefetusse.
36. n Verge1yking van die tyd gespandeer deur verski11ende
oorbietjiegroepe met hu11e koppe om1aag gedurende n weiperiode (Amsterdam-studiegebied).
37. Oorbietjiedigthede in die verski1lende kampe van die
Amsterdam-studiegebied (November 1974 tot Oktober 1975).
38. Opsomming van veekonsentrasies en ve1dtoestande in die
verski1lende kampe van die Amsterdam-studiegebied
gedurende oorbietjiesensusse (November 1974 - Oktober
1975).
39.
40.
41.
42.
Die gemiddelde oorbietjiebiomassas in die verskillende
kampe van die Amsterdam-studiegebied (November 1974 tot
Oktober 1975).
Veranderinge in oorbietjiedigthede in die verskillende
kampe van die Amsterdam-studiegebied (November 1974 tot
Oktober 1975).
Oorbietjiedigthede in die verski1lende kampe van die
Piet Retief-studiegebied (November 1974 tot Oktober 1975).
Opsomming van veekonsentrasies en veldtoestande in die
verskillende kampe van die Piet Retief-studiegebied
gedurende oorbietjiesensusse (November 1974- Oktober 1975).
154
156
157
195
209
231
232
234
235
240
241

43.
44.
Die gemiddelde oorbietjiebiomassas in die verskillende kampe
van die Piet Retief-studiegebied (November 1974 tot Oktober
1975).
Veranderinge in oorbietjiedigthede in die verskillende kampe
van die Piet Retief-studiegebied (November 1974 tot Oktober
1975).
(xvi)
242
243

LYS VAN FIGURE
FIGUURNR.
1. n Oorbietjieram.
2. Geografiese verspreiding van oorbietjies in Suide1ike Afrika.
3. Loka1iteite van die Amsterdam- en Piet Retief-studie
gebiede.
4. Panoramiese foto's van die Amsterdam- en Piet Retief
studiegebiede.
5.
6.
7.
8.
9.
10.
11.
'n Kaart van die Ams terdam-s tudiegebied.
n Kaart van die Piet Retief-studiegebied.
n Geo1ogiese kaart van die Amsterdam-studiegebied.
n Geo1ogiese kaart van die Piet Retief-studiegebied.
Gewysigde k1imaatsdiagram vir die Amsterdam-weerstasie
444/277, 1239 m bo seespiee1.
Gewysigde klimaatsdiagram vir die Piet Retief-weer
stasie 444/540, 1260 m bo seespiee1.
n Geomorfologiese kaart van die Amsterdam-studiegebied.
12. Skema tiese voors telling van 'n deursni t ( noord-suid)
deur die Amsterdam-studiegebied met n afbakening in
geomorfologiese eenhede.
13. n Fami1iegroep oorbietjies wat aan die kant van n
doleriet-k1ipgedee1te (plato-uit1oper) wei~ Kamp 11,
Arnsterdam-studiegebied.
14. 'n Geomorfo1ogiese kaart van die Piet Retief-studie
gebied.
(xvii)
B ls.
2
8
13
14
15
17
18
20
26
27
35
36
38
42

15. Skematiese voorstelling van ~ deursnit (noord-suid) deur
die Piet Retief-studiegebied met ~ afbakening in
geomorfologiese eenhede.
16. Plato-uitloper met spitskruinreste (a), dagsome (b),
eskarp (c), tallushang (d), platovleikom (e) en
pediment (f) van die Piet Retief-studiegebied.
17.
18.
19.
~ Plantegroeikaart van die Amsterdam-studiegebied.
~ Plantegroeikaart van die Piet Retief-studiegebied.
Die Cyrnbopogon excavatus-~.eckloniana-Aloe
marlothii- struikveld (Piet Retief-studiegebied).
20. Die Alloteropsis semialata-Trachypogon spicatus
grasveld (a) met kenmerkende plantsoorte soos
Helichrysum nudifolium en Trachypogon spicatus.
Die Diospyros whyteana-Aloe marlothii-struikveld
(b) word op die hoer noordfrontglooiings aange
tref (Piet Retief-studiegebied).
21. Die Berkheya sp. - Helichrysum sp. - Tristachya
hispida- grasveld (a), kamp M 13 (Piet Retief
studiegebied).
22. Die Gerbera sp. - Eulalia villosa-Eragrostis plana
grasveld (a)Q ~ Ou wattelplantasie (b) is sedertdien
met Eragrostis curvula beplant~
23. ~ Oorbietjieooi wat in ~ kortgebrande en beweide
gedeelte van die Eulalia villosa-Hypoxis rigidula
Berkheya setifera- grasveld wei (Amsterdam-studie
gebied).
24. 'n Oorbietjie kan voedselplante in 'n wye halfsirkel
bereik sender om te beweeg.
25. Veranderinge in voginhoude van vier grasspesies ge
d.urende 'n 24-uur periode (Juliemaand) in die
Amsterdam-studiegebied.
(xviii)
43
45
59
80
87
92
95
98
114
115
120

26~
28.
29.
Daaglikse aktiwiteitspatrone van oorbietjies in die
Amsterdam-studiegebied.
Seisoenale aktiwiteitspatrone van oorbietjies in die
Amsterdam-studiegebied.
Oorbietjie-familiegroepe toon n groot mate van koordinasie
tydens alle aktiwiteite. Oorbietjies gedurende n
rusperiode, Amsterdam-studiegebied.
Tipiese lyfkosingsaktiwiteite van oorbietjies
(vanaf 35 rnm kleurskyfies).
30. n Opperv1aktediagram om die oorbietjie se verhouding
tussen die kosing van verskillende liggaamsdele aan
te toon.
31. n Oorbietjieram in n slaapposisie terwyl sy kop op
sy flank rus (vanaf n 35 mm kleurskyfie).
32. n Oorbietjieram wat gedurende die nag wei (foto geneem
met behulp van n nagbesigtigingsteleskoop.
33.
34.
Nagtelike aktiwiteitspatrone van oorbietjies in die
Amsterdam-studiegebied.
Die invloed van temperatuur, windspoed en reenval op
die daaglikse aktiwiteitspatrone van oorbietjies.
n Territoria1e oorbietjieram wat individuele afstand
afdwing op n ondergeskikte ram.
36. n Oorbietjieram in n aggressiewe/dreigende houding met
die ore en horings na vore gerig (vanaf n 35 mm kleur
skyfie).
37. n Territoriale oorbietjieram wat n onderskikte ram jaag
(vanaf n foto).
(xix)
125
126
127
132
133
137
138
139
141
163
164
164

38.
39.
40.
Lokaliteite van vyf bekende oorbietjie-territoriums in die
Amsterdam-studiegebied.
Die bewegings van twee volwasse oorbietjieramme in die
Piet Retief-studiegebied wat met behulp van te1emetrie
bepaal is (September 1975 tot Januarie 1976).
'n Oorbie tj ieram merk 'n grashalm (Hyparrhenia hirta) met
die preorbitale klier.
41. 'n Preorbitale klierafskeiding op die punt van 'n grashalm
(Eulalia vil1osa). Die afgebyte dee1 van die grasha1m
is in die agtergrond sigbaar.
42.
43.
44.
45.
'n Oorbietjieram wat 'n grasb1aar (Themeda triandra) laag
merk (vanaf 'n 35 mm kleurskyfie).
'n Oorbietjieram merk 'n ooi se stert met die preorbitale
klier direk na 'n naso-anale kontak (vanaf 'n 35 mm kleur
skyfie).
'n Oorbietjieram in 'n urineringsposisie.
'n Oorbietjieram in 'n ont1astingsposisie.
46. Frekwensie van verskillende reaksies van 'n territoriale ram
167
169
172
172
174
176
178
178
wat in volgorde plaasvind nadat 'n ooi urineer/ontlas het. 179
47. 'n Oorbietjieram lek aan vars urine en fekaliee van 'n ooi.
48. 'n Oorbietjieram krap 'n gesament1ike rniskol heftig met sy
voorpote.
49.
50.
Fekaliee van 'n oorbietjieram bo-op die van 'n ooi.
ram se krapmerke is op die grond sigbaar.
Die
Gesament1ike oorbietjiemiskol in 'n voetpad (Amsterdam
studiegebied).
180
180
182
183
(xx)

51. Skematiese voorstelling van gemeenskap1ike oorbietjie
miskolle in die Amsterdam-studiegebied.
52.
53.
54.
Twee oorbietjieooie in n aggressiewe houding (vanaf n 35 mm k1eurskyfie).
Naso-anale kontak wat kopulasie voorafgaan (vanaf n 35 mm kleurskyfie).
Kopulasie (vanaf n 35 rnm kleurskyfie).
55. Postkopulasiegedrag waar die oorbietjieram probeer om
die ooi op te jaag.
56. Postkopulasiegedrag : die oorbietjieram probeer om aan
die ooi se vulva te ruik terwyl sy n matige submissiewe
houding inneem.
57. Geboortes van oorbietjielammers in die Amsterdam- en
Piet Retief-studiegebiede, bereken vanaf observasies op
lammers jonger as een maand oud.
58.
59.
60.
61.
Swaar dragtige oorbietjieooi gedurende die lamseisoenpiek.
Oorbietjielam (ongeveer een maand oud) wat aan die ooi
drink. Die anale dele van die lam word heftig gelek.
Sebras wat in die direkte omgewing van n familiegroep
oorbietjies wei (Amsterdam-studiegebied).
Beeste wat n versteuring veroorsaak het.
62. Primere verdedigingsmeganisme by n jong oorbietjie
ooi waar sy agter n graspol verskuil le.
63.
64.
n Oorbietjie-ooi skuil tussen klippe na n versteuring.
n Oorbietjieram 1e in n waaksame/onsekere houding
(nek gestrek, en ore na agter gedraai).
(xxi)
184
187
190
192
193
193
197
198
200
205
206
211
211
212

65.
66.
67.
68.
69 ..
70.
n Skematiese voorste11ing van oorbietjie-v1uggedrag.
Norma1e 1oophouding van 'n oorbietj ie ( vanaf 'n 16 mm
ro1film).
Loophouding van n oorbietjie nadat potensie1e gevaar
opgemerk is (vanaf n 16 mm ro1fi1m).
V1ugbewegings van 'n oorbietj ieooi wa t in n wipsprong
eindig (vanaf n 16 rnm ro1fi1m).
n Oorbietjieooi in die 1aaste fase van n wipsprong.
Skematiese voorste1ling van oorbietjie-v1ugreaksies.
71. Oorbietjieooie neem die 1eiding in die meeste v1ug
reaksies.
72. V1ugafstande van oorbietjies (fami1iegroepe) in die
Amsterdam-studiegebied.
73. Verski11ende horisonta1e p1antdigtheidsk1asse soos
deur n digtheidsbord aangetoon.
74. Voorbee1de van die verski11ende horisonta1e p1antdigt
heidsk1asse.
75.
76.
77.
Oorbietjieverspreiding in die Amsterdam-studiegebied.
Oorbietjieverspreiding in die Piet Retief-studiegebied.
Mens1ike besetting, landerye en p1antasies in die Piet
Retief-studiegebied.
(xxii)
213
214
214
216
217
218
220
221
227
228
229
237
239

HOOFSTUK 1
INLEIDING
Die oorbietjie (Fig. 1), Ourebia ourebi ourebi (Zimmermann, 1783), is
die grootste van die Neotragini. Ten spyte van groot onderlinge
verskille sluit die Neotragini ook onder andere die klipspringer
Oreotragus oreotragus, dikdik Madogua kirki en steenbok Rhapicerus
campestris in.
In die verlede het oorbietjies wydverspreid in die grasveldgebiede van
Suidelike Afrika voorgekom (duPlessis 1969, Millar 1970). Die oorbietjie
se posisie het egter die laaste tyd aansienlik verswak regdeur die ver
spreidingsgebied in Suidelike Afrika. n Ernstige afname het in al vier
provinsies in Suid-Afrika voorgekom, maar veral in die Kaapprovinsie
(Millar 1970) en die Oranje-Vrystaat (Von Richter, Lynch en Wessels 1972).
Die oorbietjie se status in Suid-Afrika word deur verskeie werkers as
swak of selfs haglik beskou (Anon. 1952, Knobel 1958, Bigalke en Bateman
1962, Vincent 1962, Ansell 1971 en Von Richter~ al. 1972). Von Richter
(1974) het die oorbietjie as bedreig in Suid-Afrika beskryf in sy bespreking
van bewaarde gebiede in verhouding tot wildsoorte. Daarinteen beskou
Bothma (1975) die oorbietjie se bewaringstatus as skaars/veilig in Suid
Afrika en slegs in Swaziland as bedreig. Skinner, Fairall en Bothma
(1977) beskryf egter die oorbietjie se status tans wel as kwesbaar.
Met die aanvang van die studie was uiters min bekend oor die etologie en
ekologie van die oorbietjie, nie net plaaslik nie maar ook elders in die
boksoort se verspreidingsgebied. Afgesien van Monfort en Monfort(l974)
se studie op enkele aspekte van bevolkingsdigtheid en gedrag van die oor
bietjie in die Nasionale Akagera-wildtuin in Rwanda, het slegs Gosling
(1972) inligting oar die oorbietjie se preorbitale kliermerking gepubliseer.
Verskeie kort verwysings na sekere aspekte van die oorbietjie se ekologie
is egter beskikbaar,onder andere deur Hediger (1951), Edwards (1968, 1969a,
1969b) en Tait (1969). Leuthold (1977) wys egter op die gebrek aan inlig
ting oor die kleiner boksoorte en veral die Neotragini.

2
Fi guur 1 . 'n Oo r b i e t j i e r am.

3
Om doeltreffende bewaringsbeplanning vir skaars of bedreigde wildsoorte
te laat geskied, is basiese inligting oor hierdie diere se ekologie nood
saaklik. Die oorbietjie kom tans hoofsaaklik op privaat eiendom voor
waar die dier se voortbestaan grootliks onseker is weens menslike aktiwi
teite soos gewasverbouing en die aanle van plantasies. Ten einde n aan
duiding van die oorbietjie se voortbestaan op privaat eiendom te kry, is
n studie op die ekologie dringend noodsaaklik.
Met hierdie eto-ekologiese studie is daar klern gele op verskillende
aspekte van die oorbietjie se gedrag. Habitatvoorkeure is bestudeer deur
n studie van die geornorfologie en plantegroei van twee studiegebiede om
vergelykings te bewerkstellig. Inligting oor die verspreiding en getalle
van oorbietjies in Transvaal is gedurende 1973 versarnel terwyl na geskikte
studiegebiede gesoek is. Daar is finaal op twee studiegebiede, naby
Amsterdam en Piet Retief in die Suidoos-Transvaal, besluit en die veld
waarnernings het in Desember 1973 n aanvang geneem wat tot Julie 1976
geduur het.

4
HOOFSTUK 2
TAKSONOMIE EN MORFOLOGIE
Taksonomie
Zimmermann bet die oorbietjie se genotipe, Antilope ourebi Zimmermann,
wat afkomstig was uit die Uitenhage distrik, in 1783 beskryf (Roberts 1954).
Dje vroegste verwysing na oorbietjies was moontlik deur Thunberg (1795,
In: Millar 1970) wat n oorbietjie gedurende 1773 naby Humansdorp gesien
bet. Die klassifikasie van die oorbietjie is tans soos volg (Ansell
1971):
Orde:
Familie:
Sub-familie:
Tribus:
Genus:
Subspesie:
Artiodactyla
Bovidae
Antilopinae
Neotragini
Ourebia Laurillard, 1842
Ourebia ourebi ourebi (Zimmermann, 1783)
Slegs die een spesie Ourebia ourebi word aangetref met 13 subspesies wat
verspreid oor die kontinent van Afrika voorkom. Die verspreidingsgrense
en inskakeling van die onderskeie subspesies is egter glad nie duidelik
nie (Ansell ££• cit.). Die subspesie Q·~· ourebi (Zimmermann, 1783) word
in Suid-Afrika aangetref en waarskynlik ook in die suidelike deel van
Mosambiek.
Morfologie
Die oorbietjie is die grootste van die kleiner boksoorte met n gemiddelde
massa van 14 kg en n skouerhoogte van ongeveer 58 em (Bylae A). Die ooie
is soms groter as die ra~~e. Horings is slegs by die ramme aanwesig wat
gemiddeld 11,3 em lank word, maar kan volgens Best (1962) n maksimum
lengte van tot 18,41 em bereik.
Die haarkleed het bo-op die oorbietjie n kenmerkende rooi- tot geelbruin

5
kleur wat skerp afsteek teen die wit hare onderkant. n Duidelike
kontras word deur die kort, bossieagtige swart stert en die withare
van die binneboude gevorm. Gewoonlik vertoon die haarkleed syagtig
en effens krullerig.
Preorbitale kliere word by beide geslagte aangetref alhoewel dit
grater in die geval van ramme is. n Kleinerige kol haarlose vel
kom onder die ore voor terwyl klossies langerige hare op die knie
gedeeltes sigbaar is.

6
HOOFSTUK 3
GEOGRAFIESE VERSPREIDING
Algemeen
Die oorbietjie is prim;r ~ grasveldsoort wat in effens variante vorme in
beide die Afrogematigde/Bergagtige en Tropiese gebiede aangetref word
(Tinley 1969, Pienaar 1974). Volgens Ansell (1971) is die oorbietjie
afwesig in die Suidelike Savanne sone in gebiede aangrensend aan die
Suidwestelike Droe sone. Verder noord ko~ die oorbietjie in die Noorde
like Savanne sone vanaf Uganda en weswaarts tot by Senegal voor, terwyl
dit ook in die Sudanese Droe sone deur Kenya tot in sentraal en westelike
Ethiopie versprei is. Die oorbietjie word vanaf seevlak in die Kaap
provinsie (Ansell ££• cit.) tot op ~ hoogte van 2 500 m in Kenya (Stewart
en Stewart 1963) aangetref.
Vroee Verspreiding
In Suider-Afrika het die oorbietjie se oorspronklike verspreidingsgebeid
in baie gebiede aansienlik verander en verklein as gevolg van menslike
besetting met gepaardgaande plantegroeiveranderings en swaar jagdruk
(Von Richter 1971), alhoewe1 duPlessis (1969) van rnening is dat die oor
bietjie se getalle veel grater veranderinge ondergaan het as die versprei
dingsgebied self.
Vroeer het die oorbietjie wyd verspreid in die grasveldgebiede van die
Kaapprovinsie, Natal, Oranje-Vrystaat, Transvaal, Mosambiek en Zimbabwe
voorgekom (du Plessis 1969, Millar 1970, Ansell 1971). Oorbietjies het
egter uit baie gebiede verdwyn sodat die verspreiding tans meer gelokali
seerd is (Ansell 1971, Anon. 1952).
In Kenya het die oorbietjie oor ~ groat gebied verdwyn (Stewart en Stewart
1963) terwyl ~ groat verrnindering in getalle in Nigerie voorgekom het
(Petrides 1965, ~: Ansell ££• cit.). Geen inligting oor getalsveranderinge
in ander dele van Afrika is beskikbaar nie.

7
Huidige Verspreiding in Suidelike Afrika
TRANSVAAL
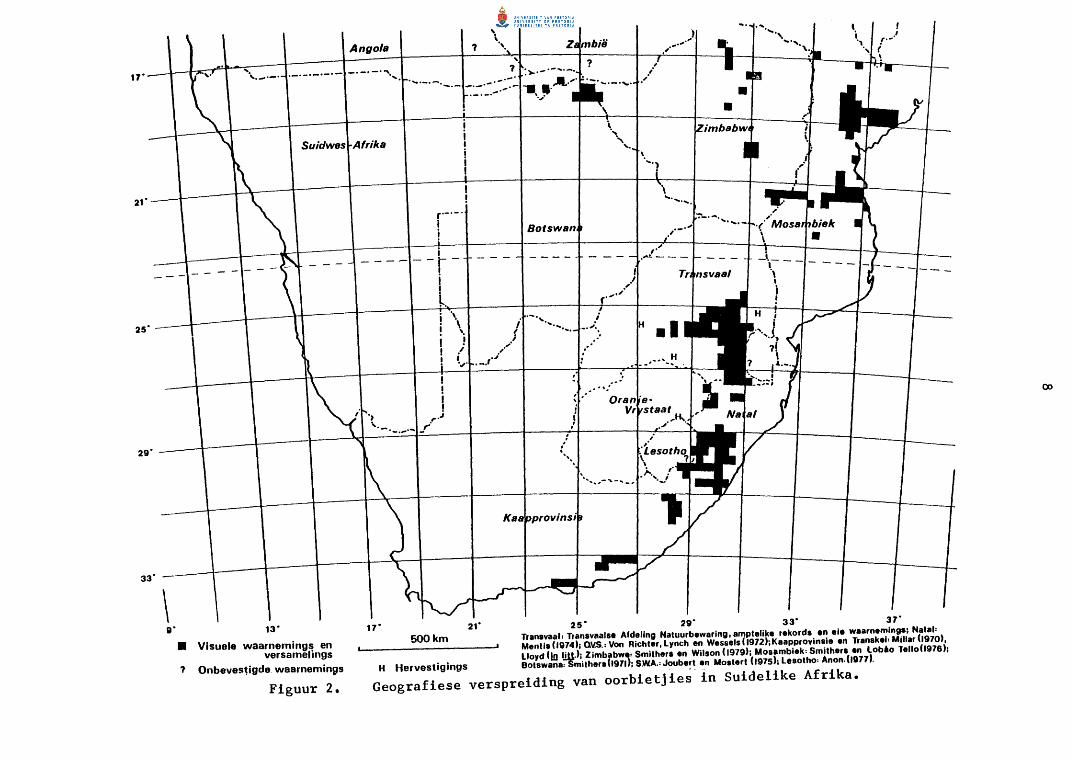
In Transvaal kom die grootste konsentrasies van oorbietjies in die gebied
tussen Ermelo, Piet Retief en Carolina voor (Fig. 2)o Kleiner groepies
word verspreid in die hoerliggende dele van Oos- en Sentraal-Transvaal
aangetref tot sever noord as Blyderivierspoort (25° 30' SB), en weswaarts
tot suidwes van Pretoria (27° 45' 01). Ooswaarts strek die verspreidings
gebied tot in die omgewing van Barberton in die noorde en Piet Retief in
die suidooste (31° 15 1 01). Vanaf die grootste konsentrasies neem die
getalle noordwaarts en weswaarts af terwyl die diere ook meer gelokaliseerd
voorkom, veral in die intensiewe boerdery- en plantasiegebiede. Alhoewel
Stevenson-Hamilton (1929) meld dat oorbietjies voorheen in die Laeveld
voorgekom bet, bestaan daar geen aanduidings dat hierdie gebied werklik
deel van die oorspronklike verspreidingsgebied was nie. Volgens Pienaar
(1963) is n oorbietjie gedurende 1925 naby Mlondozi in die Nasionale
Krugerwildtuin gesien.
Oorbietjies is op verskeie plekke in Transvaal hervestig. Die loslating
van oorbietjies in die Loskopdam-natuurreservaat en Ohrigstaddam-natuur
reservaat (Riney en Kettlitz 1964) was slegs gedeeltelik suksesvol (Anon.
1967, Von Richter 1971). Aanvanklik het die vestigingspoging in die
Nasionale Krugerwildtuin gefaal (Pienaar en van Niekerk 1963), maar latere
pogings was egter meer suksesvol (Anon. 1973). Oorbietjies is ook in die
Suikerbosrand- en Rustenburg-natuurreservate losgelaat (Transvaalse Afdeling
Natuurbewaring, amptelike leers). Oorspronklike kleiner groepe kom ook
in die Blyderivierspoort-natuurreservaat voor en munisipale reservate soos
die Van Riebeeck-natuurreservaat naby Pretoria en die Lydenburg-natuurreser-
vaat.
NATAL
In Natal is die oorbietjie se verspreidingsgebied hoofsaaklik tot die voet
heuwels van die Drakensberg beperk, alhoewel dit ook in die Natalse Midde-
w- \ ~-.. ··\ -·-.-·-·L-·-·-·-\-·-·-·,.l I -,-k-·--·-1? I /, 1 l i -,r ~r).
21"
25"
29"
33"
g• 13"
• Visuele waarnemings en versamelings
1 Onbeves~igde waarnemin9s
Figuur 2.
17" 21" 25" 29" 33" 37" 500 km Tranevaalt Transvaalse Afdellng Natuurbewarlng, amptelike rekorda en ele waarnemings; Natal•
Mentis (1974); Q'lS.: Von Richter, Lynch en Wessels ( 1972);Kaapprovinsie en Transkel: Millar (1970), Lloyd (lo !lllJ; Zimbabwe: Smithers en Wilson (1979); Mosambiek• Smithers en Loblo Telfo(l976);
H Hervestigings Botswana-;8mitherah971); S.WA.= Joube~t en Mostert (1975); Lesotho• Anon.(ID77).
Geografiese verspreiding van oorbietjies in Suidelike Afrika.
00

9
lande voorkom (Von Richter 2£e cit.). Mentis (1974) meld dat oorbietjies
in Natal veral in die intermediere dele en vlaktes aangetref word. n Groot aantal oorbietjies word in die Giants Castle-wildtuin beskerm
(Bourquin 1966).
ORANJE-VRYSTAAT
Alhoewel die oorbietjie vroeer wyd verspreid in die oostelike dele van
die o.v.s. voorgekom het (Edwards 1968), het n opname deur Von Richter,
Lynch en Wessels (1972) aangetoon dat oorbietjies moontlik nog in die
Harrismith, Bethlehem en Vrede distrikte aangetref word.
n Aantal oorbietjies is in die Golden Gate Hoogland Nasionale Park losge
laat (Penzhorn1971b, Strybis 1974). Die voortbestaan van die spesie in
hierdie provinsie word egter ernstig bedreig (Von Richter~ al. 1972).
KAAPPROVINSIE
n Ernstige afname in getalle het die oorspronklike wye verspreiding van
die oorbietjie in die Kaapprovinsie laat krimp tot enkele distrikte, onder
andere Bathurst, Humansdorp en Mount Currie (Millar 1970, Lloyd~ litt.).
Die verspreidingsgebied is beperk tot Oos-Kaapland, en veral die dele
aangrensend aan Transkei. Gedurende 1960 is n paar oorbietjies na die
Thomas Baines-reservaat by Grahamstown oorgeplaas (Millar ££• cit.).
TRANSKEI
Tans is geen akkurate inligting beskikbaar oor die verspreiding van oor
bietjies-in Transkei nie, maar volgens Millar (1970) is die oorbietjie in
hierdie gebied hoofsaaklik tot die suidwestelike deel beperk en word slegs
baie gelokaliseerde bevolkings aangetref.
ZIMBABWE
Die hoogste konsentrasies van oorbietjies in Zimbabwe kom in die intensiewe
landbougebiede van die noordelike en noordoostelike dele voor (Thomson 1973).
Lloyd, P~H., Privaatsak 5014, Stellenbosch 7600

10
Smithers en Wilson (1979) dui egter n kleiner verspreidingsgebied as
Thomson (2£.cit.) aan. n Aantal oorbietjies word ook in die Gonarezhou
wildtuin aangetref asook in die Nasionale Lake Kyle-wildtuin waar oor
bietjies gedurende 1965 losgelaat is (Ferrar 1973, In: Thomson 1973).
Groepies oorbietjies is ook in die westelike gedeelte naby Botswana op
gemerk (Smithers en Wilson 2£· cit.).
MOSAMBIEK
Oorbietjies is volop in die savanne en grasveld gedeeltes van die Nasionale
Gorongosa-wildtuin (Tinley 1977), terwyl hulle ook in die Nasionale Banhine~
wildtuin aangetref word. Volgens Smithers en Lobao Tello (1976) word
oorbietjies in die noordelike distrikte van Vila Pery en Beira, asook in
die suid-westelike dele soos Tete- en Zambezia distrikte gevind.
BOTSWANA
Volgens Smithers (1971) is die oorbietjies in Botswana beperk tot die
noord·Jostelike gedeeltes aangrensend aan Zimbabwe en die Caprivigebied.
Daar is geen aanduidings dat die oorbietjie se verspreidingsgebied in
Botswana enige drastiese veranderinge ondergaan het nie.
SUIDWES-AFRIKA
Alhoewel Joubert en Mostert (1975) geen aanduidings kon vind dat die oor
bietjie tans in hierdie gebied voorkom nie, het Shortridge (1934) en die
Carp-ekspedisie (Smithers 2£• cit.) die teenwoordigheid van oorbietjies
in die Caprivigebied gerapporteer. Volgens Von Wilhelm (1933) het oor
bietjies vroeer ook in Owamboland voorgekom.
SWAZILAND, LESOTHO EN ANGOLA
Roberts (1954) meld dat die oorbietjie in Swaziland voorkom, maar daar
bestaan geen betroubare inligting oor die huidige verspreiding nie.

11
Oorbietjies het waarskynlik vroeer ook oor n groo-t gebied in Lesotho
voorgekom (Sclater 1900), maar inligting oor die huidige verspreiding
en status is tans egter nie beskikbaar nie. Enkele oorbietjies is
wel in die Nasionale Sehlabathebe-wildtuin opgemerk (Anon. 1977)e
Volgens Shortridge (1934) was oorbietjies volop in Angola veral in die
suidoostelike gedeelte langs die Okavangorivier. Huntley (pers. rned.)
verklaar dat oorbietjies tans in die suid en suid-westelike asook noordelike
grasveld en oop savanne gebiede (ongeveer 900 tot 1 000 rn bo seevlak)
gevind word.
Huntley, B.R., WNNR, Posbus 395, Pretoria 0001

12
HOOFSTUK 4
STUDIEGEBIEDE
Inleiding
Die digste bevolkings van oorbietjies op private eiendom in Suid-Afrika
word waarskynlik in Transvaal aangetref met die middelpunt van die huidige
verspreidingsgebied in die suid-oostelike deel van die provinsie gelee.
Afgesien van die oorbietjies op privaat eiendom kom hulle ook in verskeie
provinsiale reservate asook die Nasionale Krugerwildtuin voor. Hierdie
reservate val almal binne die bree historiese verspreidingsgebied in
Transvaal (du Plessis 1969), maar die bokke het, behalwe in die geval van
Blyderivierspoort-natuurreservaat, heeltemal in die reservate uitgesterf
voordat daar met hervestigingspogings begin is.
Twee gebiede waar groo~ konsentrasies van oorbietjies in Transvaal voorkom
is as studiegebiede uitgesoek, naamlik die plaas Forbes Athole naby Amster
dam en gedeeltes van die plase Mooihoek, Rooi Kraal, Uitgevallen en Drie
hoek suidwes van Piet Retief (Fig. 3 en 4). Die studiegebiede is in die
gemengde boerderystreek oos van die platorand gelee. Aangesien hierdie
gebiede in verskeie opsigte verskil, beide ekologies en wat boerdery
aktiwiteite betref, het dit n geleentheid gebied om verskillende aspekte
van oorbietjie-ekologie vergelykend te ondersoek. Die Amsterdam-studie
gebied is vir die etologiese studies gebruik, terwyl aspekte van bevolkings
dinamika, bewegings en habitatsvoorkeure van die oorbietjie in beide gebiede
bestudeer is.
Ligging en Grootte van Studiegebiede
AMSTERDAM-STUDIEGEBIED
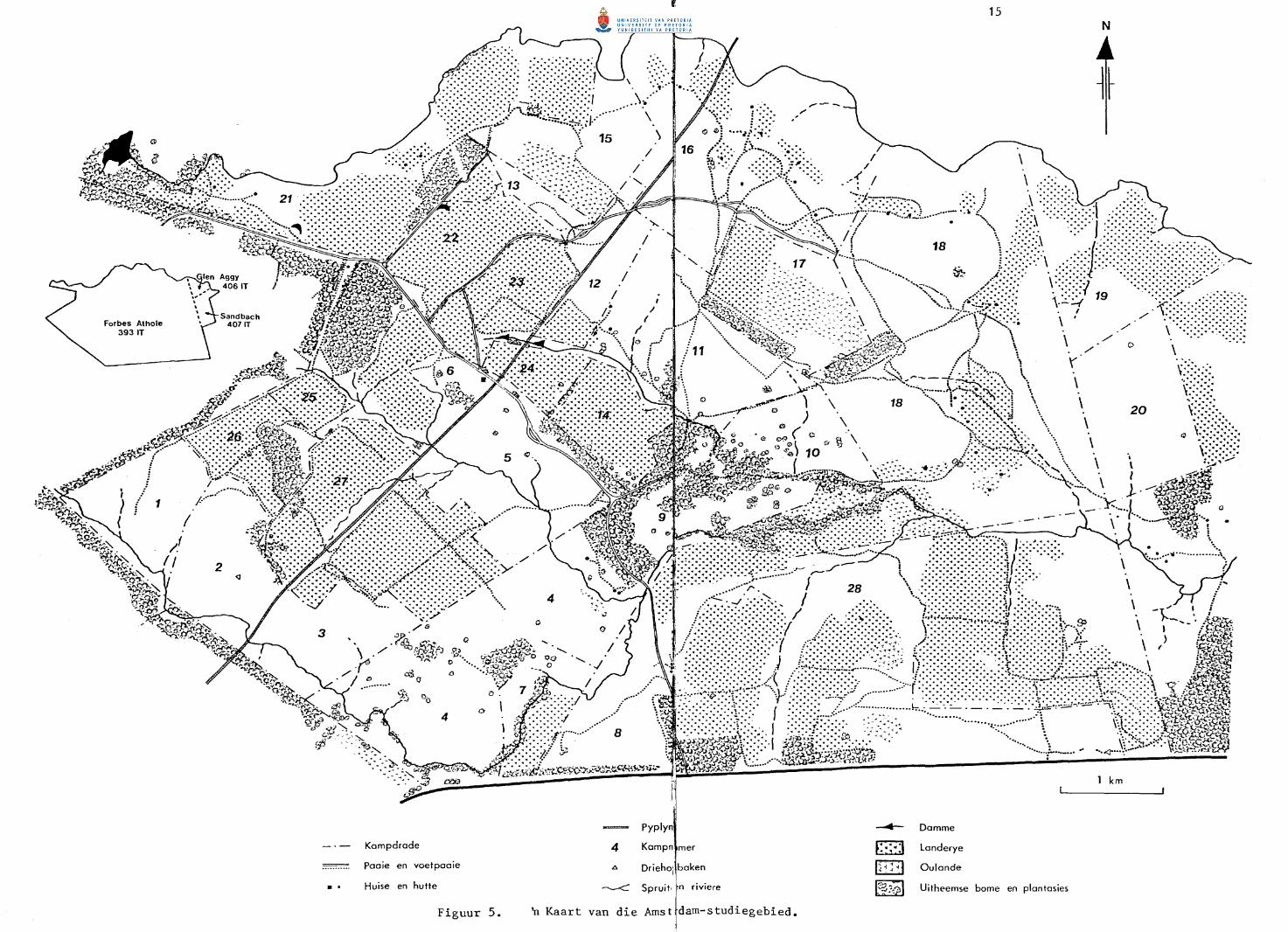
Die vernaamste studiegebied (Fig. 5), bestaande uit gedeeltes van Forbes
Athole 393 IT, Glen Aggy 406 IT en Sandbach 407 IT (Fig. 5) in die Ermelo
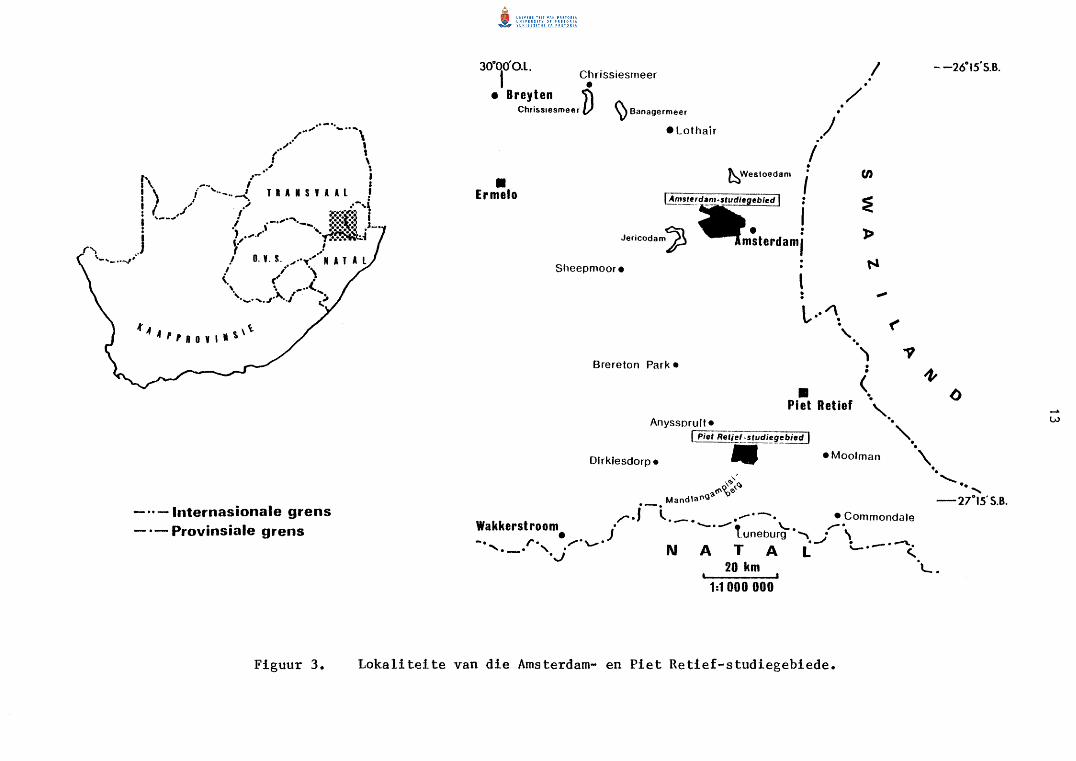
....... ,/··"' _ ..... ,
,··"· \ ' \ ··' ' f\ ~- •
~~ I f I \ l '··-··-! T • A I s ' A A l • I J ,/ ,; •• -,;· : ( ,.1 I ;-;•;•-'.•.., ,.,._.; . . ..... I I ·-·""' "'·- •••••• t:.g : . ·' -., :·:·:~~··· .J-
J }) ,·'·' "'rl~~ • I ..... . .6·
.. _ .. __ .__,,. I 0. V. S . ..,··"·' N AT A l I ,·· \ \. c' • \ .. ., .... <. ...
'·._. ,,J""··I :!
f ,f \\. .fl'riiOVII\
-··-lnternasionale grens - ·-Provinsiale grens
Jo•oo'o.L. Chrissiesrneer
I • • Breyten ~ (\. Banagermeer Chri~s1esmeer V V ,
• Ermelo
Sheeprnoor •
• Lothair
~Wesloedam
I /. . .
\., .. '\ '
en
~ )>
tol
.,.,.
.. "
- -26°15'5.8.
('
? Brereton Park • . •
• < .. ,
Piet Relief ' Anyssorult •
I Piet Retje~studi_!!l_!bled]
Dlrklesdorp • • ~' Q' ... ~ 'l)~'Qe
Mandlaf'<}
•Moolrnan
.. "· .'\
J·-l -·-. • Cornrnondale ,... ·-·'- _.,• '- ,.... Wakkerstroom J. · t.uneburg ·"""\ ; \ __ . ·- . -·~ ....
()
··,. ...... -27.15' 5.8.
-· r·, ,..'-'. N A T A L '-' <.. ' . . . ·- "...J 20 km L.
1:1000 000
Figuur 3. Lokaliteite van die Amsterdam- en Piet Retief-studiegebiede.
w

NOORDOOS
Westoedam
I
Arnst erda m-stud iegebied
SUID
Luneburg
I
Piet Retief - st ~diegebied
Fi guur 4. Panor amiese fo t o ' s van die Amsferdam- en Pi et Retief- s tudiegebiede .
14
oos Amsterdam
I
SUI OWES eMhlongamvula Mand langampi si
.......
Forbes Athole 393 IT
Aggy 4061T
Sand bach 4071T
4 0
Kampdrade
= Paaie en voetpaaie
.. Huise en hutte
Figuur 5.
4
riviere
n Kaart van die Arnst dam-studiegebied.
~ ~
Damme
Landerye
Oulande
15
Uitheemse borne en plantasies
r \ \ \ \
\
N
1 km
20

16
distrik, is privaat eiendom tussen 26° 34' en 26° 39' SB en 30° 30' en
30° 38' OL gelee. Amsterdam is direk oos van die gebied gelee met die
eskarpement ongeveer 20 km wes in die omgewing van Bankkop (Wellington
1955) met Ermelo nog 50 km verder wes.
Die totale oppervlakte van die Amsterdam-studiegebied is 5 920 ha
waarvan Forbes Athole 5 590 ha uitmaak. Sandbach en Glen Aggy beslaan
200 ha en 130 ha onderskeidelik.
PIET RETIEF-STUDIEGEBIED
Die Piet Retief-studiegebied bestaan uit gedeeltes van vier private plase,
naamlik Mooihoek 168 HT, Rooi Kraal 173 HT, Driehoek 174 HT en Uitgevallen
175 HT tussen 27° 06 1 en 27° 10' SB en 30° 34 1 en 30° 40' OL (Fig. 6).
Hierdie gebied is ongeveer 22 km suidoos van Piet Retief, en noordoos van
die Mandlangampisiberg gelee en sluit die 2 115 m hoe eMhlongamvulapiek in
(Fig. 4). Volgens Humphrey en Krige (1931) asook Wellington (1955) is
die gebied in n oorgangsarea tussen die Hoeveld en Laeveld gelee. Die
studiegebied beslaan 4 308 ha waarvan Mooihoek 1 773 ha, Rooi Kraal 2 085,
Driehoek 50 ha en Uitgevallen 400 ha uitrnaak.
Fisiografie
AMSTERDAM-STUDIEGEBIED
Geologies bestaan die studiegebied hoofsaaklik uit gesteentes van die
Supergroep Karoo met sandsteen en skalie van Ecca~formasie en tilliet
en fluvioglasiale sedimente van die Dwyka=formasie gebied. (Fig. 7).
Doleriet van die Karoo Supergroep bedek die hoer sentrale gedeelte van die
gebied. Ultrabasiese gesteentes van die Usushwana-kompleks en ska1ies
van die Archaiese-komp1eks kom ook in die gebied voor (Hammerbeck 1977).
n Klipve1d bestaande uit do1erietrotse is kenmerkend van die hoer gedee1tes.
Vo1gens Fitzpatrick (1974) sorteer die Arnsterdam-studiegebied onder die
sogenaamde Athole=pedosisteem waar die gronde hoofsaak1ik uit rooi en

Mooihoek 168HT
I
.. .. .# .& -..... & ..... .& •
I
1 I
I
Rc3
Rc 1
\ v .
'
'
••·••• •••• ·."\ Ra 7 ................ & ......... . . . ... . . ... .
• .a ... .& .... ..
1 km
Kampdrade
Rc4
·--·--. .
- Paaie en voetpaai ..
• • Huise en hutte
\ /
\/
Figuur 6.
•
Ra2
A
-..<:::
___,.__
'· ...... ·/'----'( J ·, M4
./l· /
....... , •
- .. .a ... .. ... ... ... . ... • • ... ·.a .......
•
M9
' \ / · ...... /· \
Ra 1
Rb4
l t---·-
Kampnommer
Drieh-:>eksbaken
Spruite en riviere
Damme
I
.I
• •
o o A . ...
·-f..
I
I
I
, M7
M11
/
I
·~ I /'·-·J.. -
~ landerye A . t@ J.. Oulande
M Uitheemse bome
'n Kaart van die Piet R ef-studiegebied.
N
en plantasies

llli1i1l :1'! Doleriet
~ Sandsteen, skalie
0 Tilliet, fluvioglasiale sediment
lk%4tl Miluograniet
rJ!J/IJJlJ Leucogabbro, l<wartsgabbro
[]]]] Pirol<lastiese rots, rioliet r+++""n . ~ Ultrabas1ese rots
f:-=:-1 Ysterhoudende sl<alie
~ Kwartsiet met sl<alie
Grens van studiegebied
lkm
Ecca formasie
Dwyl<a formasie
} Karoo Supergroep
Amsterd'lm formasie
Thole formasie
) Usushwana kompleks
Voorslag formasie . } Archa"iese kompleks
Sl<urwerant formas1e
Figuur 7. n Geologiese kaart van die Amsterdam-studiegebied.
N
t CXl

19
geel distrofiese klei bestaan. Litosoliese en grys klei en leemgronde
word ook in gedeeltes aangetref. Fitzpatrick (££• cit.) meld verder
dat die Griffin en Clovelly gronde in die gebied van die Dwyka tilliet
afkomstig is. Klam, suur v1eigronde kom in die 1aagliggende dele voor~
Topografies is die studiegebied n golwende laer plato ge1ee teen die kant
van die p1atorand verder wes. n Dendritiese dreineringspatroon kom voor
met die Thole en Kristalspruit wat ooswaarts na die Ngwempisirivier vloei.
Die hoogte bo seespieel wisse1 tussen 1 265 en 1 490 m.
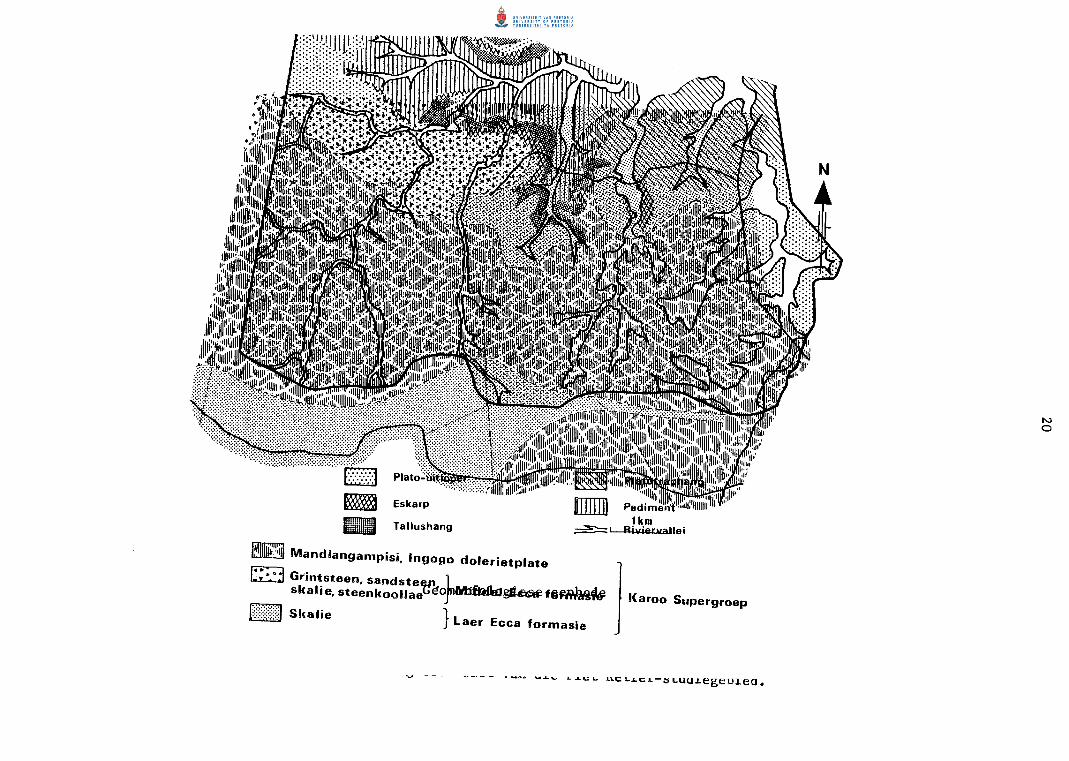
PIET RETIEF-STUDIEGEBIED
Geologies bestaan die gebied grotendeels uit Mandlangampisi- en Ingogo
do1erietp1ate. Onderliggende sandsteen en skalie van die Middelste Ecca=
formasie en Laer Ecca~formasie:ska1ie is in sommige dele blootge1e (Fig.
8). Steenkoollae maak dee1 van die Middel Ecca=formasie uit (Humphrey en
Krige 1931).
Die Piet Retief-studiegebied is gedee1telik in die Majuba- en Dirkiesdorp
pedosisteme gelee met geel en grys hidromorfiese leem en sandgronde in
die 1aagliggende gedee1tes (Fitzpatrick 1974). Sterk verweerde gronde
is aanwesig op die boer do1erietdele, terwyl matig verweerde tot onge
differensieerde en litosoliese gronde teen die g1ooiings gevind word.
Die bergagtige en golwende topografie is kenmerkend van die studiegebied.
Die hoe do 1erie tdekp la to met verkerfde eskarpemen t word deur 'n vlak en
vertakte dreineringspatroon ingekerf wat in die noordooste1ike deel van
die gebied saamvloei om die Boesmansprui t, 'n sytak van die Assegaairivier,
te vorm. n Relatiewe onge1yke topografie en betreklike steil g1ooiings
lei tot 'n meer onge lyke land skap as die van die Amsterdam- s tudiege hied.
Die hoogte bo seespiee1 wissel tussen 1 234 en 1 559 m.
Klimaat
ALGEMEEN
Beide studiegebiede is in n somerreenvalgebied gelee wat gekeTh~erk word
deur gematigde somers en koue winters wat volgens KBppen as n Cwa-klimaat
mrnn Eskarp
Tallushang ~· 'k.m Rtwervallei
E3 G•;ntstoen. sandsto'I!'~J-~ f8~ l<amo Sup.,gmop skahe, steenkoollae T'
~ Mandlangampisi, lngogo dolerietplate 1 [}td Sl<alie } laer Ecca formasie
--~--- .. -~ ... -~'-.&..a.'!§;;""' u.c\...Lc.L-::;LuuJ.e.geu~ea.
N
N 0

21
geklassifiseer kan word (Trewartha 1954). Vo1gens Schulze (196)) is die
gebiede in die "H-streek" (Hoeveld) gelee, met die Piet Retief-studiegebied
net noord van die "D-streek'' (Drakensberg).
K1imaatsgegewens vir die streek is beperk aangesien Piet Retief (eerste
orde-stasie) die enigste groat weerstasie in die omgewing is. Robbertse
(1972, In: Fitzpatrick 1974) verwys na die beperkte gegewens oor klimaat
in die Suidoos-Transvaal. Gegewens afkomstig van verskeie kleiner weer-
stasies (Tabel 1) word egter gebruik om die klimaat van die studiegebied
te beskryf (Weerburo 1954, Schulze 1965, Weerburo 1965, 1967a, 1967b).
Ten einde vergelyking te vergemaklik word die makrok1imaat van beide
studiegebiede saam bespreek.
SONSKYNDUUR EN WOLKBEDEKKING
Die maksimum sowel as die minimum gemiddelde maandelikse persentasies
sonskynduur vir Piet Retief is die hoogste gedurende Junie en die laagste
gedurende Oktober. Junie het gemiddeld die minste dae sonder sonskyn
(0,3 dae) in teenstelling met Oktober wat die meeste dae (4,0 dae) het
(Schulze 1965).
TEMPERATUUR
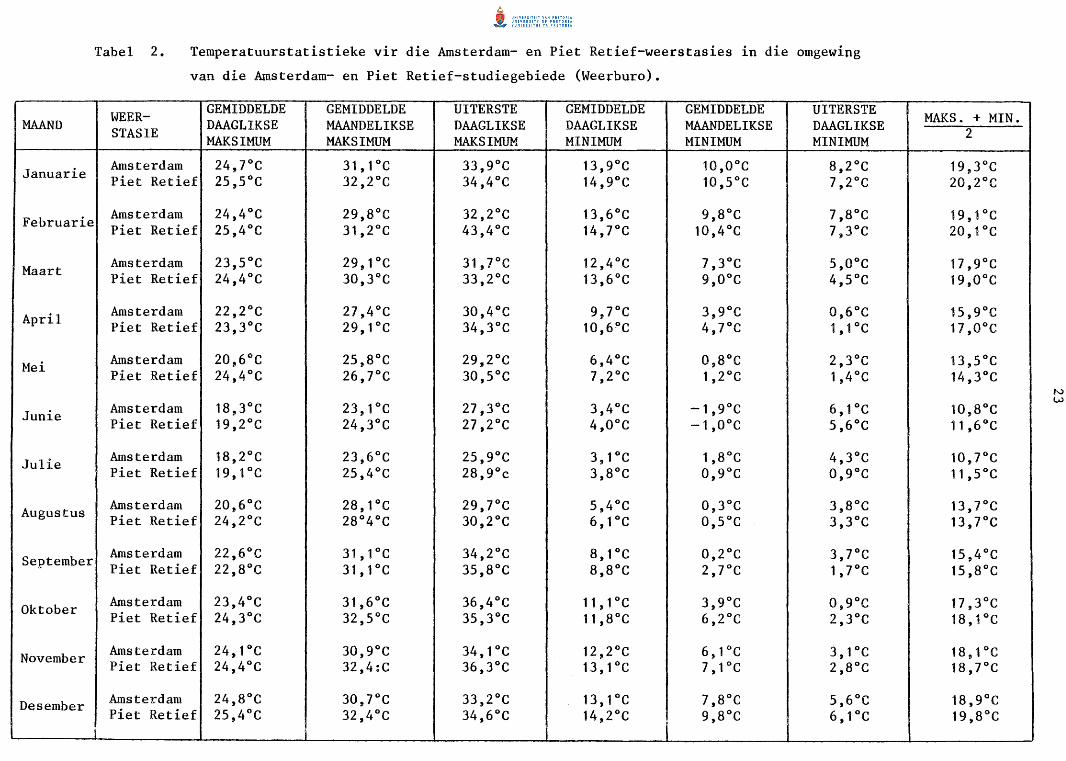
Temperatuurgegewens (Tabel 2) is afkomstig van Amsterdam en Piet Retief
(Weerburo 1954). Hierdie weerstasies, beide na aan die twee studiegebiede,
is op n hoogte bo seespieel gelee wat feitlik ooreenstem met die van die
studiegebiede.
Die hoogste daaglikse maksimum temperature kom vanaf Oktober tot Maart
voor (Tabel 2) met 'n gemiddelde maandelikse maksimum van 31,6°C en 32,5°C
vir Oktober by Amsterdam en Piet Retief respektiewelik. Junie tot Augus
tus is die koudste maande met n gemiddelde maandelikse minimum van -1,9°C
en -1,0°C gedurende Junie by Amsterdam en Piet Retief onderskeidelik.
Volgens die temperatuurgegewens blyk dit dat Piet Retief-omgewing effens
warmer is as die Amsterdam-omgewing.
Ryp kom gedurende die wintermaande voor. Die gemiddelde intreedatum van
ryp by Piet Retief is 16 Junie wat gemiddeld vir 43 dae duur tot die ge-

Tabel 1.
WEERSTASIE
At hole 444/126
Amsterdam 444/277
De Hoop 444/304
Piet Retief 444/540
Goede hoop 408/37
Tafel berg 408/464
Besonderhede van die weerstasies waarvan temperatuur- en reengegewens
verkry is (Weerburo 1954, 1965, 1967a, 1967b).
PERIODE VAN WAARNEMING SUIDER- OOSTER- HOOGTE BO (JAAR) BREEDTE LEN GTE SEESPIEEL (m) Temperatuur Reenval
26° 14' 30° 35' 1 620 - 2H:
30° 40' 1 239 9 50 26° 37' 1904-1913
26° 34' 30° 41' 1 525 - 11
30° 48' 1 263 46 54 27° OO' 1904-1950
27° 07' 30°32' 1 432 - 24
27° 14' 30° 46' 1 372 - 25
LIGGING VAN WEERSTASIES T.O.V.
Amsterdam- Piet Retief-studiegebied studiegebied
In studiegebied -
± 10 km. 0. -
± 13 kme 0. -
± 50 kmo S.O. ± 22 km. NeO.
- ± 2 km. We
- ± 16 kme S .0 e
I
N N
MAAND
Januarie
Februarie
Maart
April
Mei
Junie
Julie
Augustus
September
Oktober
November
Des ember
Tabel 2. Temperatuurstatistieke vir die Amsterdam- en Piet Retief-weerstasies in die omgewing
van die Amsterdam- en Piet Retief-studiegebiede (Weerburo).
WEER- GEMIDDELDE GEMIDDELDE UITERSTE GEMIDDELDE GEMIDDELDE UITERSTE
STASIE DAAGLIKSE MAANDELIKSE DAAGLIKSE DAAGLIKSE MAANDELIKSE DAAGLIKSE MAKSIMUM MAKSIMUM MAKSIMUM MINIMUM MINIMUM MINIMUM
Amsterdam 24,7°C 31,1°C 33,9°C 13,9°C 10,0°C 8,2°C Piet Retief 25,5°C 32,2°C 34,4°C 14,9°C 10,5°C 7,2°C
Amsterdam 24,4°C 29,8°C 32,2°C 13,6°C 9,8°C 7,8°C Piet Retief 25,4°C 31,2°C 43,4°C 14,7°C 10,4°C 7,3°C
Amsterdam 23,5°C 29,1°C 31,7°C 12,4°C 7,3°C 5,0°C Piet Retief 24,4°C 30,3°C 33,2°C 13,6°C 9,0°C 4,5°C
Amsterdam 22,2°C 27,4°C 30,4°C 9,7°C 3,9°C 0,6°C Piet Retief 23,3°C 29,1°C 34,3°C 10,6°C 4,7°C 1,1°C
Amsterdam 20,6°C 25,8°C 29,2°C 6,4°C 0,8°C 2,3°C Piet Retief 24,4°C 26,7°C 30,5°C 7,2°C 1,2°C 1,4°C
Amsterdam 18,3°C 23,1°C 27,3°C 3,4°C -1, 9°C 6,1°C Piet Retief 19,2°C 24,3°C 27,2°C 4,0°C -1 ,0°C 5,6°C
Amsterdam 18,2°C 23,6°C 25,9°C 3,1°C 1,8°C 4,3°C Piet Retief 19,1°C 25,4°C 28,9°C 3,8°C 0,9°C 0,9°C
Amsterdam 20,6°C 28' 1 °C 29,7°C 5,4°C 0,3°C 3,8°C Piet Retief 24,2°C 28°4°C 30,2°C 6,1°C 0,5°C 3,3°C
Amsterdam 22,6°C 31,1°C 34,2°C 8,1°C 0,2°C 3,7°C Piet Retief 22,8°C 31,1°C 35,8°C 8,8°C 2,7°C 1,7°C
Amsterdam 23,4°C 31,6°C 36,4°C 11 '1 °C 3,9°C 0,9°C Piet Retief 24,3°C 32,5°C 35,3°C 11,8°C 6,2°C 2,3°C
Amsterdam 24' 1 °C 30,9°C 34,1°C 12,2°C 6,1°C 3,1°C Piet Retief 24,4°C 32,4:C 36,3°C 13,1°C 7,1°C 2,8°C
Amsterdam 24,8°C 30,7°C 33,2°C 13,1°C 7,8°C 5,6°C Piet Retief 25,4°C 32,4°C 34,6°C 14,2°C 9,8°C 6,1°C
MAKS. + MIN. 2
19,3°C 20,2°C
19,1°C 20,1°C
17,9°C 19,0°C
15,9°C 17,0°C
13,5°C 14,3°C
10,8°C 11,6°C
10,7°C 11,5°C
13,7°C 13,7°C
15,4°C 15,8°C
17,3°C 18,1°C
18,1°C 18,7°C
18,9°C 19,8°C
~...__
I
I
N w

middelde uittreedatum van 29 Julie.
rypdae voor (Weerburo ££• cit.).
RELATIEWE LUGVOGTIGHEID
24
Jaarliks kom n gemiddeld van 3,2
Gegewens oor relatiewe lugvogtigheid vir Amsterdam is nie beskikbaar nie,
maar wel vir Piet Retief (Weerburo 1954). Die hoogste gemiddelde rela
tiewe lugvogtigheid (85%) en die laagste gerniddelde maandelikse minimum
(66%) om 08h00 is onderskeidelik gedurende Maart en September gemeet.
Uit die gegewens blyk dit dat die relatiewe lugvogtigheid gedurende die
somer en herfs hoog is, terwyl dit die laagste gedurende die winter en
lente is.
WIND
Windgegewens vir Amsterdam is nie beskikbaar nie, daar word gevolglik
slegs van die gegewens van die Piet Retief-weerstasie gebruik gemaak.
Volgens Schulze (1965) kom winde uit die noordoostelike sektor meer dik
wels in die somer voor in teenstelling met die noordwestelike winde ge-
durende die winter. Die sterkste winde word vanaf Augustus tot November
ondervind teenoor die herfsrnaande wanneer die minste wind waai •
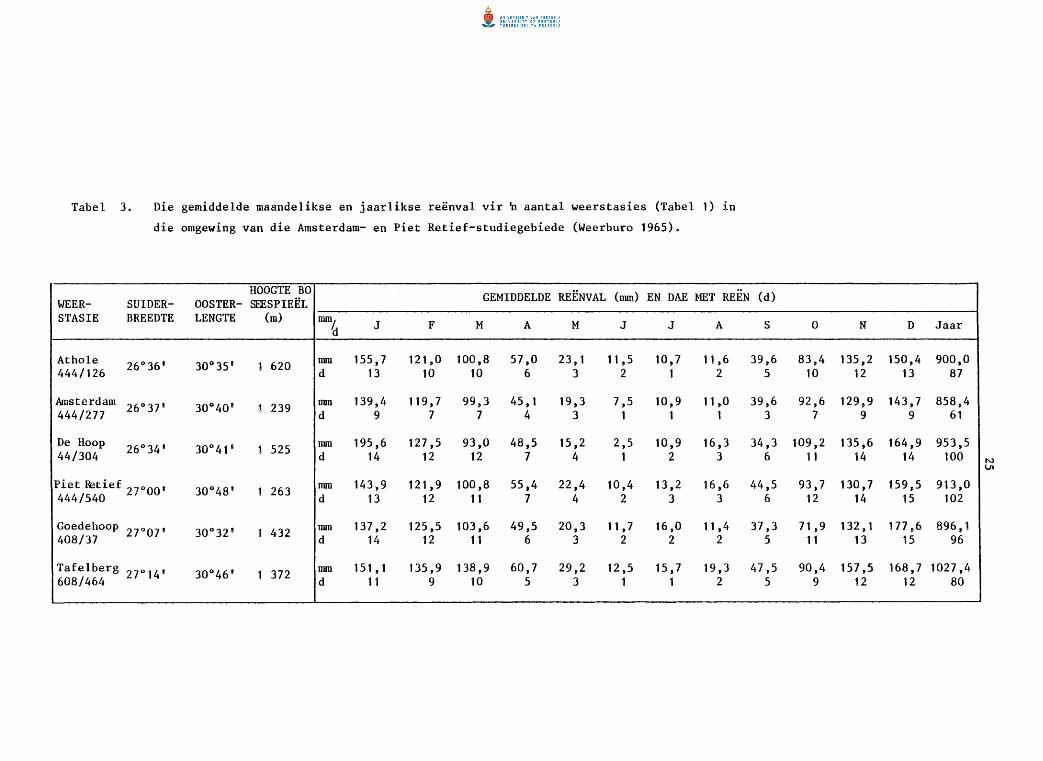
.. REENVAL
Volgens die reenvalrekords van die weerstasies in die omgewing (Tabel 3)
is die gemiddelde jaarlikse reenval vir die Amsterdam-studiegebied ongeveer
900 mrn, en vir die Piet Retief-studiegebied tussen 896 en 913 rnrn (Weer
buro 1965). By Amsterdam kom 84% van die reenval tussen Oktober en Maart
voor in vergelyking met 82% by Piet Retief. Uitgesonderd Tafelberg kom
die maksimum maandelikse neerslag gedurende Desember en Januarie voor, en
die minimum gedurende Junie en Julie (Tabel 3).
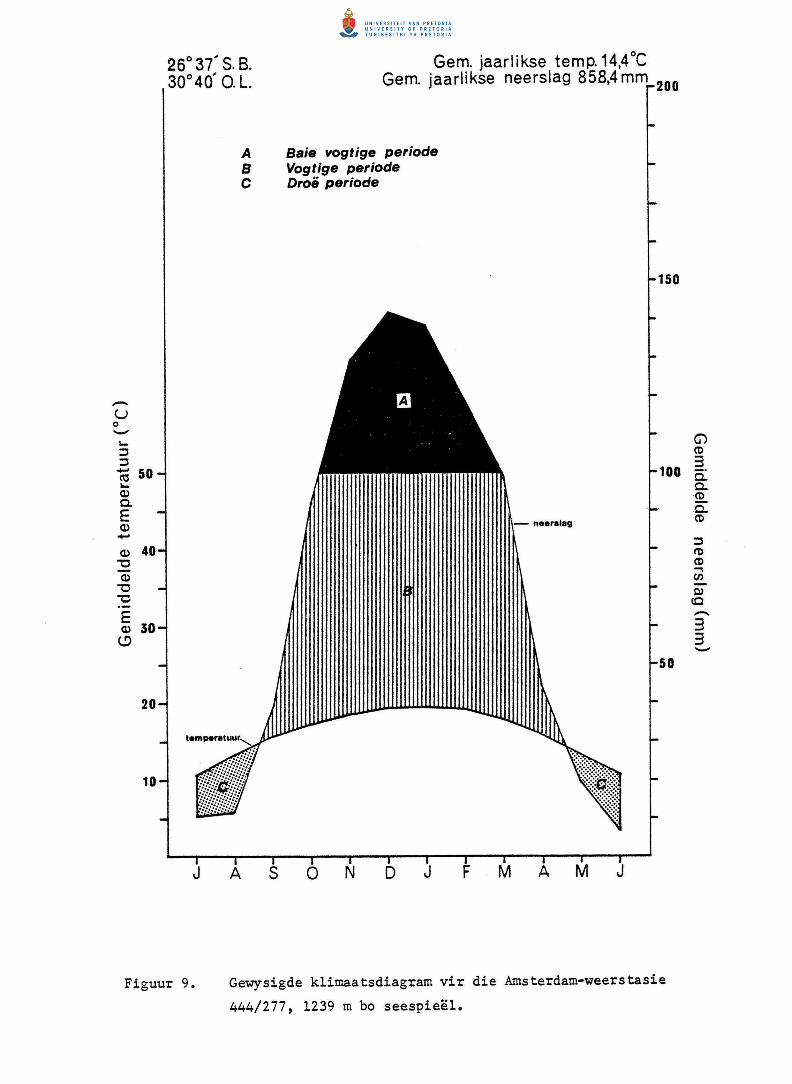
Klirnaatsdiagramme (Walter 1971, soos gewysig en gebruik deur Hall-Martin
1972, Theron 1973, Bredenkamp 1975 en Wilson 1975) is saamgestel uit reen
val en temperatuurgegewens (Fig. 9 en 10)~ Hierdie diagramme toon dat
die baie vogtige periode langer by Piet Retief as by Amsterdam duur.
Gedurende hierdie periode groei die meeste plante meestal aktief (Theron
Tabel 3. Die gemiddelde maandelikse en jaarlikse reenval vir n·aantal weerstasies (Tabel 1) in
die omgewing van die Amsterdam- en Piet Retief-studiegebiede (Weerburo 1965).
HOOGTE BO GEMIDDELDE REENVAL (mm) EN DAE MET REEN (d) WEER- SUIDER- OOSTER- ~SPIEEL STASIE BREEDTE LENGTE (m) mm/ F M A
d J M J J A s
At hole 26°36' 30°35' 1 620 mm 155,7 121 ,0 100,8 57,0 23,1 11,5 10,7 11,6 39,6 444/126 d 13 10 10 6 3 2 1 2 5
Amsterdam 26°37' 30°40' 1 239 nun 139,4 119,7 99,3 45' 1 19,3 7,5 10,9 11 ,o 39,6 444/277 d 9 7 7 4 3 1 1 1 3
De Hoop 26°34' 30° 41 i 1 525 mm 195,6 127,5 93,0 48,5 15,2 2,5 10,9 16,3 34,3
44/304 d 14 12 12 7 4 1 2 3 6
Piet futief 27000 , 30°48' 1 263 nun 143,9 121 '9 100,8 55,4 22,4 10,4 13,2 16,6 44,5
444/540 d 13 12 11 7 4 2 3 3 6
Goedehoop 27007 , 30°32 1 1 432 mm 137,2 125,5 103,6 49,5 20,3 11 '7 16,0 11 '4 37,3
408/37 d 14 12 11 6 3 2 2 2 5
Tafelberg 27014 , 30°46 1 1 372 mm 151 , 1 135,9 138,9 60,7 29,2 12,5 15,7 19,3 47,5 608/464 d 11 9 10 5 3 1 1 2 5
----- ---- -~-- ~-~~~
0 N
83,4 135,2 10 12
92,6 129,9 7 9
109,2 135,6 11 14
93,7 130,7 12 14
71 '9 132, 1 11 13
90,4 157,5 9 12
D Jaar
150,4 900,0 13 87
143,7 858,4 9 61
164,9 953,5 14 100
159,5 913,0 15 102
177,6 896,1 15 96
168,7 1027,4 12 80
N lll
-u 0 -'-
::::l ::::l - 50 ~ '-(1) a. E (1) -(1) 40
""0 (1)
""0 ""0
E (1) 30
<.!)
20
J
Figuur 9.
A
A 8 c
s
26
Gem. jaarlikse temp.14,4°C Gem. jaarlikse neerslag 858.4 mm 200
Baie vogtige periode Vogtige periode Droe periode
0 N D J F
150
100
neeralag
50
M A M J
Gewysigde klimaatsdiagram vir die Amsterdam-weerstasie
444/277, 1239 m bo seespieel.
(j) (!)
3 a: 0..
-~ .0.. (!)
::::l tb (!)
""' {/)
Q)
<C -3 3 -

-u 0 -'-~
= -ctS '-(1) a. E (1) -(1)
'"0 (1)
'"0 '"0
E (1)
(!)
30
20
27°00"S.B. 30°48" 0. L.
J
A 8 c
A
27
Gem. jaarli kse temp.16,6 oC Gem. jaarlikse neerslag 919,0mm
Btiie vogtige periode Vogtige periode Droe periode
0 N D J p M A M
200
150
100
50
Figuur 10. Gewysigde klimaatsdiagram vir die Piet Retief-weerstasie
444/540, 1260 m bo seespieel.
(j) CD 3 a. a. CD a. CD
:::1 CD CD ..., en Sll <0 -3 3 -

28
££· cit.). Die droe periode, wanneer die reenvalkurwe laer as die tempera-
tuurkurwe daal, strek vanaf einde April tot begin September. Volgens
Bredenkamp (££. cit.) verkeer die meeste plante inn russtadiurn gedurende
die droe periode.
Amsterdam-omgewinge
Die droe periode is korter in die Piet Retief- as
Donderstorrns kom gereeld in die gebied voor, veral in die vroee somer.
Geen gegewens is egter oor die voorkorns van sulke storms beskikbaar nie.
Plantegroei
AMSTERDAM- EN PIET RETIEF-STUDIEGEBIEDE
Beide studiegebiede is in die Piet Retief-suurveld (Veldtipe 63) van Acocks
(1975) gelee. Hierdie veldtipe word gekenmerk deur kolle bos in beskutte
dele en n oop, suur grasveld. Die Piet Retief-studiegebied is op die rand
van die Piet Retief-suurveld en die Noordelike Langgrasveld (Veldtipe 64),
wat n oorgangstipe tussen die Piet Retief-suurveld en die Suidelike Langgras
veld (Acocks op. cit.) is, gelee. Edwards (1974) bet bereken dat die Piet
Retief-suurveld 7 594 km2
beslaan wat 0,7% van die totale landsoppervlakte
insluit.
Volgens Acocks (££. £i!.)kom die volgende algemene gras en nie-grasagtige
kruidsoorte onder andere in die Piet Retief-suurveld voor:
Tristachya hispida
Themeda triandra
Rendlia altera
Andropogon schirensis
Monocymbiurn ceresiiforme
Trachypogon spicatus
Brachiaria serrata
Diheteropogon amplectens
Heteropogon contortus
Alloteropsis semialata
Loudetia simplex
Berkheya setifera
Helichrysum oreophilum
Vernonia natalensis
In die beskutte klowe korn daar byvoorbeeld borne soos Cussonia spicata,
Ficus capensis, Faurea speciosa en Rhus transvaalensis voor. Acocks (2£e
cit.) beweer dat hierdie veldtipe moontlik vroeer n doringveld of n oop,
suur;bosveld was.

29
Die Noordelike Langgrasveld stem in n groot mate ooreen met die Piet
Retief-suurveld, maar word hoofsaaklik deur Tristachya hispida oorheers.
Verskeie nie-grasagtige kruidsoorte kom voor, byvoorbeeld:
Gnidia microcephala
Hypoxis argentea
Berkheya echinacea
Acalypha angustata
Phyllanthus glaucophyllus
Gerbera aurantiaca
Verskeie uitheemse boomsoorte word in die twee studiegebiede aangetref.
Afgesien van wattels (Acacia spp.) kom bloekomplantasies (Eucalyptus spp.)
algemeen voore
Geen plantkundige opnames is voor die aanvang van hierdie studie in die
gebied onderneem nie, met die uitsondering van enkele wielpuntopnames
op die Athole Proefplaas (Roos, Rethman en Kotze 1973).
Vroee Geskiedenis en Bestuur
AMSTERDAM-STUDIEGEBIED
Forbes Athole word sedert 1860 as veeplaas benut, terwyl die eerste blankes
reeds vanaf 1866 permanent daar gevestig is (Forbes 1938). Die plaas
is reeds vroeg op n gereelde basis gebrand en kort daarna selektief bewei.
Een derde van die gebied het gemiddeld vir twee jaar gerus, terwyl dit
gedurende Januarie en Februarie gebrand is. Die vleigebiede is geduren
de die lente gebrand. Hierdie brandprogram is tot 1973 uitgevoer waarna
daar hoofsaaklik van lentebrande gebruik gemaak is (Forbes~·~.).
Alhoewel daar aanvanklik slegs met beeste geboer is, is skape ook later
ingebring sodat daar tans ongeveer 1 400 beeste, 3 000 skape, 30 perde en
100 boerbokke in die studiegebied aangetref word. Die eerste mielies is
ongeveer 20 jaar gelede gesaai. Vir die afgelope 10 jaar word mielies
en aartappels op groot skaal gesaai en geplant. Enkele ou lande is aan-
wesig asook aangeplante weidings. Uitheemse borne, hoofsaaklik Eucalyptus
spp. en Acacia sppe, kom verspreid in die gebied voor asook in kleiner
plantasies. Die studiegebied is in n groot aantal kampe verdeel terwyl
daar ook 'n netwerk van paaie is.
Forbes, D., Forbes Athole, Posbus 32, Amsterdam 2375

30
Volgens Forbes (pers. mede) het verskeie wildsoo~te vroeer in die omgewing
van Amsterdam voorgekom wat later uitgesterf het. Onder hierdie soorte
was daar die leeu Panthera leo (Linnaeus, 1758), sebra Equus burchelli
Gray, 1824, vlakvark Phacochoerus aethiopicus Lonnberg, 1908, eland
Taurotragus oryx (Pallas, 1766), en swartwildebees Connochaetes gnou
(Zimmermann, 1780)e Baie van hierdie diere het tot ongeveer 1900 in
die gebied voorgekom, terwyl die meeste groot herbivore net vir sekere
tye gedurende die jaar opgemerk is (Forbes 1938, Forbes pers. med.).
Afgesien van oorbietjies kom daar vandag nog die volgende wildsoorte voor:
rietbok Redunca arundinum (Boddaert, 1785), rooiribbok Redunca fulvorufula
(Afzelius, 1818), vaalribbok Pelea capreolus Forster 1790, blesbok
Damaliscus dorcas Harper, 1939, springbok Antidorcas marsupialus (Zimmermann,
1970), duiker Sylvicapra grimmia (L. 1758) en steenbok Rhapicerus campestris
(Thunberg, 1811). Enkele kleiner roofdiere kom voor, byvoorbeeld rooi-
jakkals Canis mesomelas Schreber, 1775, silverjakkals Vulpes chama (A.
Smith, 1833) en moontlik tierboskat Felis serval Schreber, 1776. Twaalf
sebras is gedurende 1972 op Forbes Athole hervestig, terwyl elande geduren
de 1976 losgelaat is.
PIET RETIEF-STUDIEGEBIED
Daar word reeds sedert 1890 in die omgewing van die Mandlangampisiberg met
beeste geboer, terwyl die gebied ook later vir skaapbeweiding gebruik is
(Klingenberg pers. med.). Gereelde laatsome~en herfsbrande is tot onge-
veer 1972 toegepas, maar vroee lentebrande is tans algemeen. Sekere
kampe word egter nog gedurende die somer gebrand. Alhoewel sommige kampe
uitsluitlik vir beesbeweiding gebruik word, word enkele kampe ook vir
skape gebruik. n Driejarige rotasiestelsel word in die meeste gevalle
gebruik, waar kampe elke drie jaar gebrand word. Tans is daar n totaal
van ongeveer 1 000 beeste en soveel as 1 200 skape gedurende die winter·
maande in die studiegebied.
Landerye beslaan slegs n klein gedeelte van hierdie studiegebied. Mielies
word hoofsaaklik verbou. Die bergagtige gebied het n relatiewe klein
oppervlakte wat geskik is vir gewasverbouing met die beste moontlikhede
Klingenberg, A.W., Von Brandisstraat, Piet Retief 2380.

31
vir beweiding (Fitzpatrick 1974). Plantasies van Eucalyptus spp. en Acacia spp5 kom veral in die suidelike gedeeltes van die studiegebied
voor (Fig. 6) Die hoe reenval en gereelde voorkoms van mis maak die
omgewing besonder geskik vir die aanplant van plantasies (Cole 1966).
Sekere van die wattelplantasies (Acacia spp.) is uitgehaal waarna Eragros
tis curvula gesaai is.
Min is bekend oor die wildsoorte wat vroeer in die gebied voorgekom het.
Volgens Klingenberg (pers. med.) het die volgende diersoorte in die studie
gebied uitgesterf: luiperd Panthera pardus (L. 1785) en bosbok Tragelaphus
scriptus Pallas,l766, terwyl buffels Syncerus caffer (Sparrman 1779) voor-
heen naby die Mandlangampisiberg aangetref is. Die vo1gende grotere soog-
diersoorte, afgesien van die oorbietjie, is tans teenwoordig: rooiribbok
Redunca fulvorufu1a (Afze1ius, 1818), vaa1ribbok Pelea capreolus Forster
1790, blesbok Damaliscus dorcas Harper, 1939, duiker Sylvicapra grimmia
(L. 1758) en steenbok Rhapicerus campestris (Thunberg, 1811). Klipspringer
Oreotragus oreotragus A. Smith, 1834 het k1aarblyklik onlangs verdwyn
(Devenish pers. med.), terwyl bobbejane Papio ursinus (Kerr, 1792) periodiek
in die studiegebied opgemerk word. Tierboskat Felis serval Schreber, 1776
kom moontlik voor, terwyl die rooijakka1s Canis mesomelas Schreber, 1775
en die si1werjakka1s Vulpes chama (A. Smith, 1833) wel soms gesien word.
Devenish, M., Hooihoek, Piet Retief 2380

32
HOOFSTUK 5
GEOMORFOLOGIESE GEBIEDSINDELING
Inleiding
Fisiografie, naamlik topografie en geologie, tesame met klimaat bernvloed
die plantegroei van ~ gebied. Die topografie van ~ streek word hoof
saaklik deur die geologie bepaal, terwyl geomorfologie die huidige land
vorme beskryf in die lig van die geologiese geskiedenis (King 1963).
Geomorfologiese kartering gee die geomorfologiese eienskappe van ~ gebied
in kaartvorm weer. Hierdie eienskappe sluit die oppervlaktevorm en die
aard van die materiaal in, sodat ~ geomorfologiese kaart volledig en om
vattende inligting oor landvorme bevat (Cooke en Doornkamp 1974).
Ten einde die voorkeurhabitat van oorbietjies in die twee studiegebiede
te beskryf, nie net sover dit die plantegroei betref nie, maar ook met
betrekking tot die fisiografie, is die gebiede geomorfologies gekarteer
en die verskillende eenhede beskryf. Alhoewel die gebiedsindeling ook
direk by die plantegroeistudie aansluit, is dit primer beplan om die
fisiese habitat te beskryf.
Metodes
~ Gebiedsafbakening word in Australie gebruik waar eenhede in terme van
plantegroei en grond beskryf word (Stewart, ·Perry, Traves, Patterson,
Slatyer, Dunn, Jones en Sleeman 1970, In: Fitzpatrick 1974) terwyl ~
"terrain system" in Kanada gebruik word (Valentine 1970, ~: Fitzpatrick
~· cit.). Plaaslik is n pedosisteemafbakening gebruik om oppervlakte
en grond in die Suidoos-Transvaal te karteer en te beskryf (Fitzpatrick
~· cit.). Vir die doel van hierdie studie was dit egter noodsaaklik om
beide studiegebiede in meer detail te beskryf en te karteer as wat Fitz
patrick (££. cit.) gedoen bet.
Die glooiingsmodel van King (1962) is gebruik vir die basiese beskrywing
van die geomorfologiese eenhede in die studiegebiede, naamlik plato,

33
eskarp, tallushang en pediment. Hierdie eenhede is egter gewysig of aan-
gepas waar nodig na gelang van die samestelling van die onder1iggende
gesteentes, geomorfo1ogiese geskiedenis en die topografie.
Lugfoto's (taak nr. 515/64 van 1964) aangevul met 1:10 000 infra
rooi 1ugfoto's vir die Amsterdam-studiegebied, en taak Nr. 482/63
van 1963 vir die Piet Retief-studiegebied is gebruik vir die kartering.
Met behulp van n stereoskoop is die verskillende geomorfologiese eenhede
op n ooreen1iggende vel geteken, waarna dit tot n skaal van 1:50 000 ver
klein is. Glooiings en hoogtepunte is vanaf die bestaande 1:50 000 Topo
Kaarte (2630 DA en 2730 BA, Staatsdrukker, Pretoria) verkry.
Alle oppervlaktes is met behulp van n outomatiese oppervlaktemeter1 bepaal.
Die betrokke apparaat het n berekende korreksiefaktor van 0,988533 wat in
berekening gebring is nadat die gemiddeld van drie lesings verkry is.
Resultate
AMSTERDAM-STUDIEGEBIED
Algemeen
Die studiegebied bestaan uit n golwende 1aer plato (Fig. 4) wat geleidelik
ooswaarts daal vanaf die groot eskarpement ongeveer 20 km verder wes. n Dendritiese dreineringspatroon wat wes-oos georienteer is, is aanwesig.
Fitzpatrick (1974), wat die Suidoos-Transvaal in pedosisteme afgebaken het
in terme van grand en fisiese struktuur, sluit die Amsterdam-studiegebied
onder die Athole pedosisteem in.
Op grand van die fisiese landvorme en geologie is die volgende geomorfo1o
giese eenhede in hierdie studiegebied onderskei. (Tabel 4, Fig. 11 en 12):
Plato-uitloper
Dagsome en klipperige gedeeltes
P1atoberggebied
1 Hayaski Automatic Areameter, Model AAC-400

Tabel 4. Geomorfologiese eenhede en oppervlaktes wat dit beslaan
in die Amsterdam-studiegebied.
EENHEID OPPERVLAKTE
Plato-uitloper 2 453,24 ha Dagsome 650,21 Platoberggebied 127,03
Kruin 50,66 Tallushang 76,37
Tallushang 385,53 Pediment 2 544,45
Dagsome 139,38 Riviervallei 536,78
Platorivier 27,5 Boloop Vlakteloop
% VAN TOTALE OPPERVLAKTE
41,44 ----
6,51 42,98
-9,07 ---
100,0%
w .p.

lj,l"''.c\:·.·.;,.:.:,·,~
lff{{l Plato-uitloper
~ Kruin
riiiflliiA Tallushang
- Tallushang
mnn Pediment
~ Dagsome
}
~AIIIIIIIIIUIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIIII/IIIIIIIIIIIIIIIIIIIIIIIII''''' 1 1 1
I 1km I
Platoberggebied
~ Platoriviet }
_!.__ Boloop Riviervallei
~ Vlal,teloop
- Grense van studiegebied
Figuur 11. n Geomorfologiese kaart van die Amsterdam-studiegebied.
N
t w V1

-SUID
c.. IU ~ IU
E
·= iQ; 1 402
.! :- 1 370 IU IU 1 33 9 fl)
0 .a IU
~ 0 0 ::c
A
Figuur 12.
... c
" E ] 0..
'
-:::> ... a. .... .E "' ~ a. 0 0
! m ~ c 0 0
- ...c > "'
1 i
m c 0
...c a. "' 0 ~ 0 -0 .!!cO
m .... c ... c 0 ., " ...c g- E "' - - ;:) .. "'t]-
'3 Q) 0 b 0.. ....
? l J ~
II II II
1kn1
NO ORO-
.!
.~ "" " 0
...c !::
m a. ... "
c
a. 0
0
0 ...c
0
Ill
:::> +-... c ;:)
2 ~ " 0
1 ,g
Q..
"'t]
I " 0. I
, ..... --------
Skematiese voorstelling van n deursnit (noord-suid) deur
8
die Arnsterdarn-studiegebied met n afbakening in geornorfologiese
eenhede.
w 0'\

Kruin
Ta1lushang
Tal1ushang
Pediment
37
Dagsome en klipperige gedeeltes
Riviervallei
Platorivier
Bo1oop
Vlakteloop
Somrnige van hierdie eenhede word om praktiese redes saam bespreek terwyl
alle geologiese inligting vanaf Hammerbeck (1977) verkry is.
Plato-Uitloper
~ Effens golwende plato, wat op ~ hoogte van tussen 1 323 en 1 521 m bo
seespieel voorkom beslaan die grootste gedeelte (Tabel 4) van die sentrale
studiegebied (41,44% van die totale studiegebied). Geologies bestaan die
plato-uit1oper uit do1eriet, sandsteen, skalie, tilliet en gesteentes van
die Usushwana kompleks. Klipperige gedeeltes (doleriet) met groot klippe
is kenmerkend van die sentrale plato-uitloper en bes1aan 26,5% van die
plato (Fig. 13).
P1atoberggebied
In die suidwestelike gedeelte van die plato-uitloper word ~ p1atoberg
gebied aangetref. Hierdie gedeelte bestaan uit onderliggende gesteentes
soos piroklastiese rots van die Usushwana kompleks. Die platobergge
bied se opperv1akte maak 5,2% van die p1ato-uitloper uit, en kom op ~
hoogte van tussen 1 417 en 1 521 m bo seespiee1 voor. Steil he11ings
kom feitlik deurgaans voor wat wissel tussen 1° en 40°.
Hierdie geomorfologiese eenheid kan in ~ kruin en ta1lushang verdeel word.
Die kruin is ~ gordel van ongeveer 1 tot 300 m wyd, terwyl die tallushang
slegs op sommige plekke onderskeibaar is.
Ta1lushang
~Ware eskarp is afwesig sodat die oorgang van die p1ato-uitloper na die

Figuur 13. n Fami1iegroep oorbietjies wat aan die kant van n do1eriet-
k1ipgedee1te (p1ato-uit1oper) wei .
studiegebied .
Kamp 11, Amste rdam-
w 00

39
1aerliggende gedeeltes in die meeste geval1e ge1eide1ik geskied (Fig. 12).
Die plato-uit1oper is vir groot gedee1tes omsoom deur n ta11ushang wat 6,5%
van die totale gebied uitmaak. Die ta11ushang kom tussen 1 339 en 1 401 m
bo seespiee1 voor met die he11ing van die g1ooiings van 15° tot 40°. n
Verskeidenheid van onder1iggende gesteentes is aanwesig waarvan sandsteen
en ska1ie die belangrikste is met baie kenmerkende 1itoso1iese gronde.
Pediment
n Pediment, ge1ee tussen 1 291 en 1 443 m bo seespieel, word onderkant
die tal1ushang aangetref. In sommige geva11e strek die pediment tot
direk teenaan die p1ato-uit1oper waar die ta11ushang ontbreek, soos
in die geva1 van die p1atoberggebied (Fig. 11). Die pediment bes1aan
die grootste gedee1te van die studiegebied wat 43,0% van die tota1e
area uitmaak. Die pedimenthel1ings is ge1eidelik tot stei1 (1° - 15°).
Ti11iet en fluviog1asia1e sediment van ~ie Dwyka formasie vorm die ver
naamste onder1iggende gesteentes. Die gronde is oor die a1gemeen diep
en bestaan onder andere uit rooi en gee1 apeda1e k1ei (Fitzpatrick 1974)
wat hierdie gebied die be1angrikste eenheid vir gewasverbouing maak.
Enke1e k1ipperige gedee1tes en dagsome word ook aangetref.
Rivierva11ei
n Dendritiese en hoekige dreineringspatroon, wat in n ooswaartse rigting
dreineer, kom voor (Fitzpatrick ££• £!!.) en bes1aan 9,1% van die tota1e
studiegebied. Ten einde die k1assifikasie te vergemak1ik is die rivier
val1ei aan die hand van die gebiede waardeur dit v1oei, in drie dele
onderskei (Fig. 11).
Die platoriviere is bo-op die p1ato-uit1oper ge1ee wat as oorsprongsgebied
vir twee van die spruite dien. n V1ak v1ei-agtige voorkoms is kenmerkend
van die boonste gedee1tes. Die p1atoriviere gaan in bo1ope oor wat hoof-
saak1ik oor die tal1ushange v1oei (Fig. 11)~ Kristalspruit en die Tho1e
rivier (Fig. 5 en 12) is v1akte1ope wat uits1uit1ik oor die pediment v1oei.

40
PIET RETIEF-STUDIEGEBIED
Algemeen
Hierdie studiegebied bestaan uit ~ bergagtige dolerietplato met ~ verkerfde
en swak gedefinieerdeeskarpement aan die noordekant (Fig. 4). Die hele
studiegebied het ~ bergagtige voorkoms en sluit volgens Fitzpatrick (1974)
gedeeltes van die Majuba- en Dirkiesdorp-pedosisteme in. Humphrey en
Krige (1931) gee~ bre~ geologiese uiteensetting van die streek.
Aan die hand van die fisiese landvorme en geologie is die volgende geomor
fologiese eenhede onderskei (Tabel 5, Fig. 14 en 15):
Plato-uitloper Spitskruinreste
Dagsome
Eskarp
Plato-traphang
Tallushung
Pediment
Riviervallei
Plato-Uitloper
Plato-vleikom
Intermediere platorivier
Geut
Vlakteloop
~ Golwende plato-uitlo~er beslaan 66,8% van die totale studiegebied op ~
hoogte van tussen 1 338 en 1 597 m bo seespieel~ Die plato-uitloper
beslaan feitlik die hele suidelike gedeelte van die studiegebied (Fig. 14).
Mandlangarnpisi- en Ingogo dolerietplate vorm tesame met ~ gedeelte van die
Ecca-skalie die onderliggende gesteentes. Blootgestelde dagsome kern
verspreid voor terwyl die grande hoofsaaklik litolies is. Die spitskruin
reste, wat ~ kenmerkende voorkoms aan die plato verleen, kern verspreid
voor (Fige 14) en vorm die hoogste gedeeltes van die onmiddellike landskap
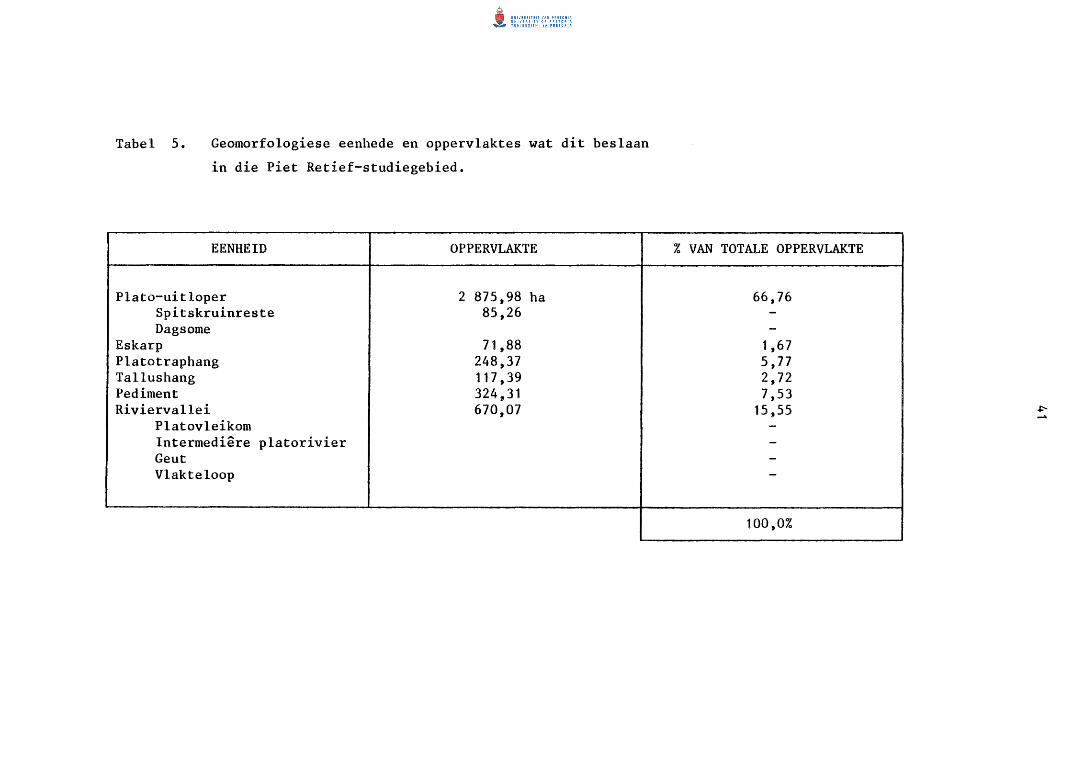
Tabel 5. Geomorfologiese eenhede en oppervlaktes wat dit beslaan
in die Piet Retief-studiegebied.
EENHEID OPPERVLAKTE
Plato-uitloper 2 875,98 ha Spitskruinreste 85,26 Dagsome
Eskarp 71 ,88 Platotraphang 248,37 Tallushang 117,39 Pediment 324,31 Riviervallei 670,07
Platovleikom Intermediere platorivier Geut Vlakteloop
% VAN TOTALE OPPERVLAKTE
66,76 --
1,67 5,77 2,72 7,53
15,55 .p.
----
100,0%
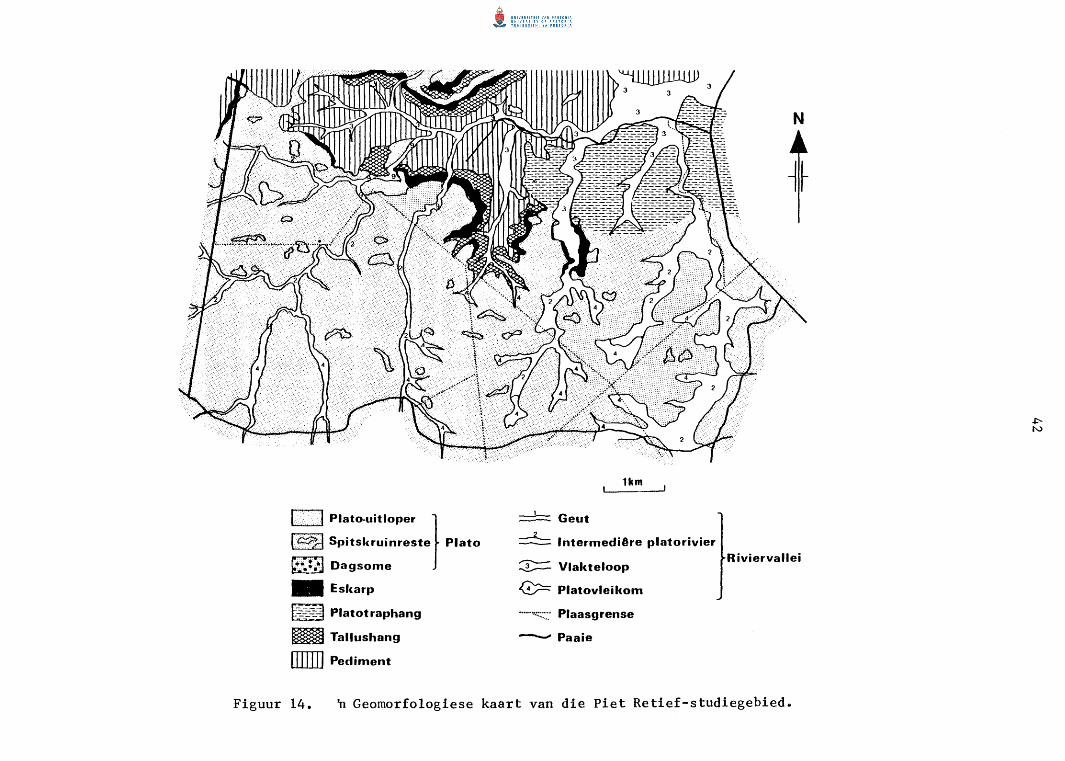
( . ····••J Plato-uitloper }
~~.: .•... : ...... •}) .....•. ·.· .. Spitskruinreste Plato
~:_~::-) Dagsome
- Esl<arp
f~j Platot raphang
mlm Tallushang
[I]]]] Pediment
I 1km I
__ , __ Geut
~ lntermedi@re platorivier
~ Vlakteloop
0== Platovlei I< om
·······,:~:·.:::· Plaasgrense
-- Paaie
N
il~
R iviervallei
Figuur 14. n Geomorfologiese kaart van die Piet Retief-studiegebied.
~ N
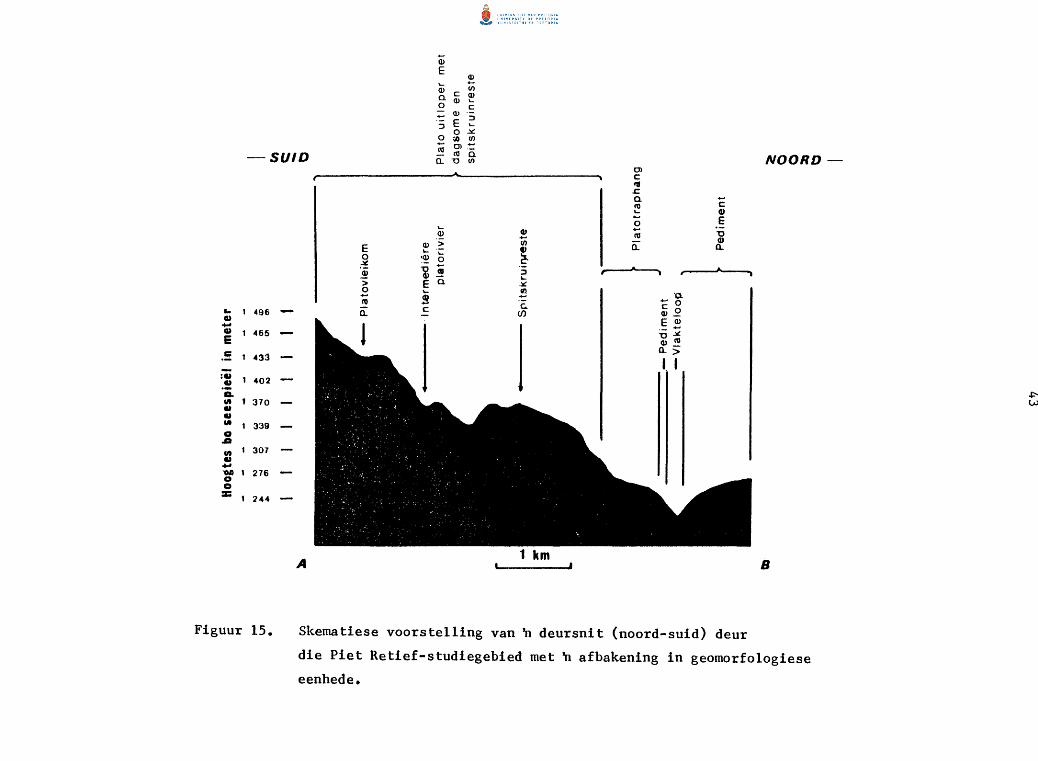
Q)
E OJ
Q; c ~ Q Q) ..... 0 c :: Q.l ·:; ·:; E .._
0 ~ .2 ~ ~
-SUID ro OJ ·- ro u a.. u !J) NOORD-
... G)
1 496 -.... G) 1 465 -E c: 1 433 -
:ea 1 402 -.!
CL ell ., 1 370
G) w 1 339 0 .a ell 1 307 ., .... ~ 1 276 0 0 = 1 244
Figuur 15.
Q; Q) > E ..... ·-
0 ~~ 0 .~
~.! Q)
> E a. 0 '-- ! cv a: c -
I
A
" u;
~ ;:, .... ~
! c..
(j)
I
1 km
Ol c;:
"' .c: Cl ~ 0
ro ([
c Q)
E "'0
~
r--~ ~ ~ ----,
·~ c 0 Q) 0 eQi :0~ Q) cv 0..-> I I
Skematlese voorstelling van n deursnit (noord-suid) deur
B
die Piet Retief-studlegebied met n afbakening in geomorfologiese
eenhede.
..J:'VJ

44
(Fig. 16). Verspreide groat klippe en rotsblokke kom teen die hange van
die spitskruinreste voor. S1egs 2,9% van die plato-uitloper word deur
die spitskruinreste beslaan. Die belling van die g1ooiings op die plato~
uitloper wisse1 van 1° tot 20°.
Eskarp
n Ware eskarp met wandkranse is tot n klein gedee1te van die noordelike
verkerfde rand van die plato-uitloper beperk wat s1egs 1,7% van die studie
gebied uitmaak (Fig. 14 en 16). Die hel1ing van die g1ooiings is deur
gaans baie stei1 (meer as 50°) met blootgestelde dagsome en rotsblokke.
Platotraphang
Aan die noordoostelike kant daal die plato-uitloper trapsgewys (Fig. 14 en
15) in die vorm van n p1atotraphang. Weens die afwesigheid van n eskarp
of tal1ushang geskied die oorgang van die plato na die 1aagliggende pedi
ment baie ge1eidelik.
Die p1atotraphang, ge1ei tussen 1 269 en 1 323 m bo seespiei1, bes1aan
5,8% van die totale studiegebied. Dagsome is opva11end en gee 1okaa1
aan vlak, swak gedreineerde grande oorsprong.
Ta11ushang
n Tallushang omsoom die grootste gedee1te van die plato-uit1oper en word
s1egs deur die eskarp en platotraphang onderbreek (Fig. 14 en 16). Die
tallushang maak slegs 2,7% van die totale studiegebied uit, en kom tussen
1 329 en 1 386 m bo seespieel voor. Skalie, sandsteen en grintsteen vorm
die onderliggende gesteentes. Die glooiings, wat lokaal met rotsblokke
en groat klippe bedek is, het hellings wat van 15° tot 40° wissel.
Pediment
n Pediment wat van wes na oos strek vorm n strook onderkant die tallushang.
Hierdie strook maak 7,6% van die totale studiegebied uit op n hoogte van

Figuur 16 . Pl ato-ui t l oper met spitskruinres t e (a), dagsome (b), eskarp
(c) , tallushang (d), pl atovleikom ( e) en pediment (f ) van die
Piet Retief - s tudieeebi ed.
~ Vl

46
1 276 tot 1 433 m bo seespieel. Die onderliggende gesteentes bestaan
hoofsaaklik uit skalie, sandsteen en doleriet. Volgens Fitzpatrick
(1974) kom hidromorfiese leem- en sandgronde voor.
Riviervallei
Die dreineringspatroon, wat hoofsaaklik inn noordelike en noordoostelike
rigting dreineer, beslaan soveel as 15,6% van die totale studiegebied.
Aan die hand van ligging en struktuur is vier tipes onderskei (Tabel 5).
Op die boer plato-uitloper is komme aanwesig wat oorsprongspunte vir
spruite is. Hierdie plato-vleikornme (Fig. 14, 15 en 16) het 'n vleiagtige
voorkoms.
Intermediere platoriviere (Fig. 14 en 15) strek oor die grootste deel van
die plato-uitloper. Hierdie riviere, met verskeie rotsblokke en groat
klippe, kerf in die dolerietplate in.
Die intermediere platoriviere gaan in sommige plekke in geute ocr (Fig. 14)
waar die waterlope diep in die tallus ingekerf het. Enkele kleiner water
valle is ook aanwesig. n Vlakteloop kom op die pediment voor (Fig. 14 en
15).
Bespreking
Die bree geomorfologiese gebiedsindelin.g van beide studiegebiede stem in
n groot mate ooreen, alhoewel die meer bergagtige Piet Retief-studiegebied
tot meer geomorfologiese eenhede aanleiding gee. 'n Golwende
hoer plato, gedreineer deur n vertakte dreineringspatroon, gaan met enkele
uitsonderings ocr in steil tallushange wat die plato-uitlopers grotendeels
omsoom. n Pediment vorm die laerliggende gedeeltes waar die dieper gronde
kontrasteer met die vlakker en dikwels litosoliese grande van die plato-uit
lopers.
Die Piet Retief-studiegebied verskil van die Amsterdam-studiegebied deurdat
eersgenoemde gebied n meer komplekse variasie tussen spitskruinreste, dag
some en platovleikornrne het. In teenstelling hiermee het die Amsterdam-

47
studiegebied ~ egalig-golwende plato-uitloper met ~ tafelvoorkoms (Fig. 11
en 12). Die oorgang van plato-uitloper na pediment geskied ook baie meer
geleidelik en met ~ kleiner verskil in die wisseling van hoogtes bo see
spieel. Alhoewel die twee studiegebiede dus in verskeie opsigte careen
stem, insluitend geologies, kan die Amsterdam- en Piet Retief-studiegebiede
as afsonderlike geomorfologiese gebiede beskou word.
Hierdie gebiedsindeling word nie alleen by die plantegroeistudie (kyk
HOOFSTUK 6) gebruik nie, maar ook by die bestudering van die oorbietjie
se spesifieke habitatsvoorkeure (kyk HOOFSTUK 11 : Gevolgtrekking).

48
HOOFSTUK 6
DIE PLANTGEMEENSKAPPE
Inleiding
n Dier se habitat word deur verskillende faktore bepaa1 waarvan voedsel,
sigbaarheid en skuiling veral belangrik is. Die verskillende plantsoorte
wa t in 'n bepaalde gebied teenwoordig is asook die hoeveelhede daarvan,
bepaa1 die beskikbare voedsel. Verder word die sigbaarheid en skuiling
in n gebied hoofsaaklik deur die verspreiding en digtheid van die plant
materiaal in verskillende hoogtes en die groeivorm van die plante bepaal.
Keast (1968) meld dan ook tereg dat plantegroei die vernaamste faktor in
herbivoorbiologie is.
Die hoofdoel van die plantegroeistudie in die twee studiegebiede is om die
plantegroei te klassifiseer, te beskryf en ekologies te interpreteer, en
waar moont1ik met omgewingsfaktore te korreleer. Hierdeur word daar
gepoog om die oorbietjie se voorkeurhabitat te bepaal en ook n aanduiding
te kry van die moontlike invloed van plantegroei op die verspreiding van
oorbietjies.
Metodes
ALGEMEEN
Vir die doel van hierdie studie is die Braun-Blanquet-metode (Werger 1974a)
gebruik om die plantegroei van die twee s tudiegebiede op 'n gedeel telik
subjektiewe basis te klassifiseer. Volgens Coetzee (1972) kan die nie
s,tatiese Braun-Blanquet-metode gebruik word om onder andere die interne
variasies en interverwantskappe van groepe releves op n eenvoudige wyse
uit te beeld, terwyl Bredenkamp (1975) daarop wys dat die metode ook
vinnige resu1 tate lewer en 'n betroubare klassifikasie gee.
Die ZUrich-Montpe11ier-benadering waarvan Braun-B1anquet die grondlegger
rvas (Braun-Blanquet 1928), is deur verskeie ander werkers uitgebou, onder
andere KUchler (1967) en Werger (1974a). Werger (1974b) meld dat die

49
Braun-Blanquet-metode waarskynlik in die toekorns baie meer in Suid-Afrika
gebruik sal word.
MONSTERNEMING
Die studiegebiede is voor die aanvang van die plantegroeistudie in Februarie
1976 stelselrnatig ondersoek om fisiografies-fisionomiese eenhede op 1: 30 000
skaal lugfoto's te kan onderskei. n Stereoskoop is gebruik om eenhede soos
plato's, klowe en glooiings (kyk HOOFSTUK 5 : Metodes) te onderskei.
Elke fisiografies-fisionomiese eenheid wat op die lugfoto's gekarteer is,
is nie noodwendig gernonster nie. Daar is egter wel gepoog om n verteen
woordigende aantal monsterpersele binne die verskillend·e fisiografies-
fisionorniese eenhede te plaas. Die presiese ligging van elke monster-
perseel is op lugfoto's aangebring. Werger (1973) meld dat monsterpersele
sorns inn gebied kom wat heterogeen is sever dit die habitat en floristiese
samestelling betref. In sulke gevalle is die betrokke monsterpersele
verskuif ten einde te verseker dat dit wel inn hornogene gebied geplaas is.
Die aantal monsterpersele hang eerstens af van die omvang van die opname,
tweedens die verskeidenheid in die gebied, en derdens die mate van presies
heid wat verlang word (Werger 1974a). Met inagnerning van bogenoemde is
daar besluit om die aantal monsterpersele te beperk. n Totaal van 49 en
28 monsterpersele is in die Amsterdam- en Piet Retief-studiegebiede
respektiewelik uitgeplaas (Tabel 6 en 7).
Elke monsterperseel moet n spesifieke minimum-grootte oorskrei wat deur die
aard van die plantegroei bepaal word, en elke monsterperseel moet die
struktuur van die omringende plantegroei voldoende verteenwoordig (Werger
1974b). Volgens Coetzee (1974) is monsterpersele met n oppervlakte van
16 m2
doeltreffend vir die Hoeveldse grasveld, terwyl monsterpersele van 2
100m deur Werger (pers. med.), In: Bredenkamp 1975) as voldoende vir
boomveld beskou word. Alhoewel monsterperseelgroottes nie tydens die
studie getoets is nie, is vierkantige monsterpersele van 16 m2 in grasveld
en 100 m2
in die boomveldgedeeltes van die Piet Retief-studiegebied gebruik.
Dieselfde monsterperseelgroottes is deur Bredenkamp (££. £!!.) in die
Suikerbosrand-natuurreservaat gebruik.
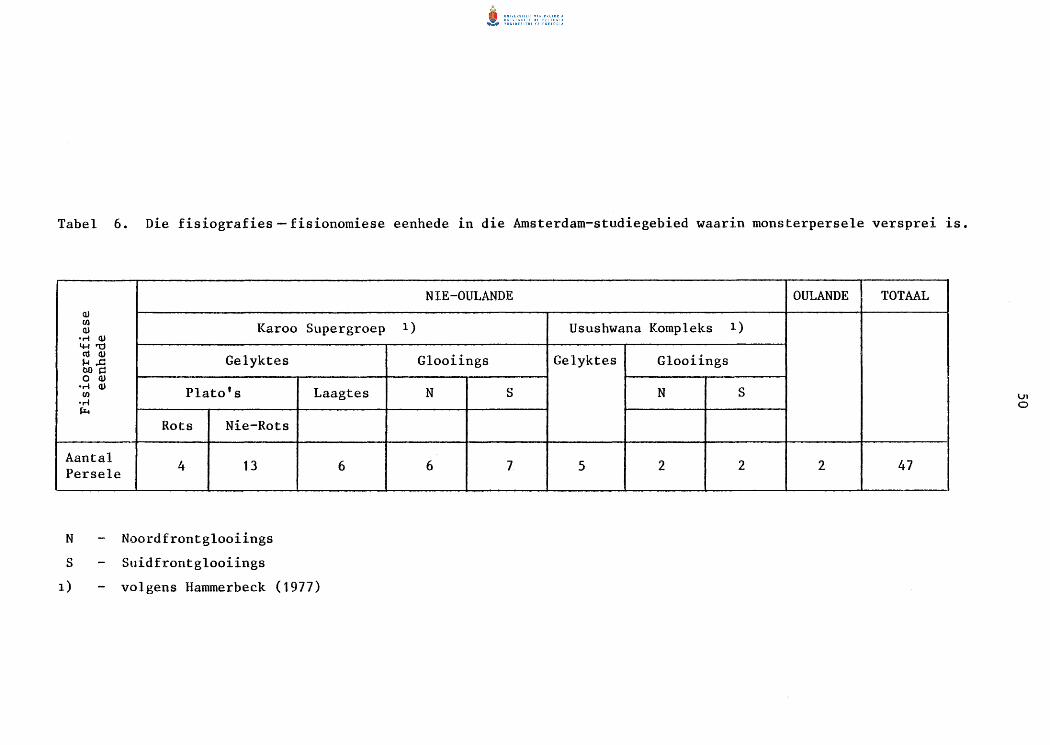
Tabel 6. Die fisiografies-fisionomiese eenhede in die Amsterdam-studiegebied waarin monsterpersele versprei is.
NIE-OULANDE OULANDE TOTAAL Q)
CJl Karoo Supergroep 1) Usushwana Kompleks 1) Q) •ri Q) 4-l"'d cd Q) Gelyktes Glooiings Gelyktes Glooiings H..C: bO~ 0 Q)
•ri Q) Plato's Laagtes N s N s CJl ·~ ~
Rots Nie-Rots
Aantal 4 13 6 6 7 5 2 2 2 47 Persele
N Noordfrontglooiings
S Suidfrontglooiings
1) volgens Hammerbeck (1977)
Vl 0
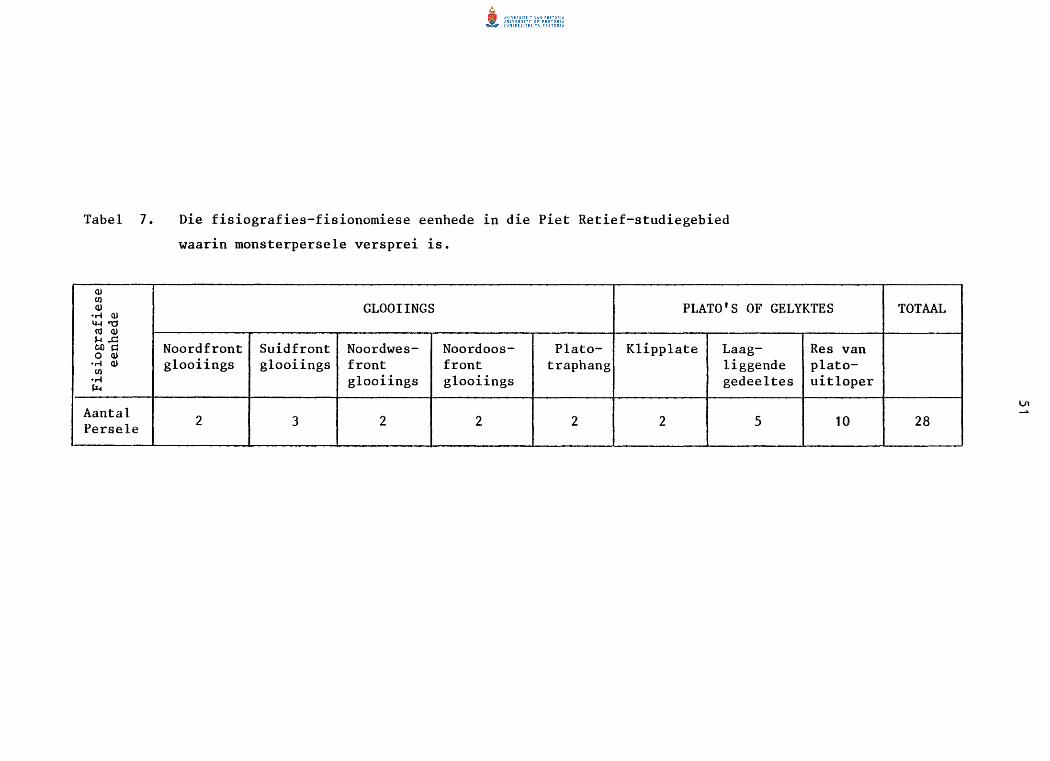
Tabel 7.
Q) (Jl Q)
•r-1 Q) 4-I'U (1j Q)
J...I..C: OOQ 0 Q)
·r-1 Q) (Jl
•r-1 r.z..
Aantal Persele
Die fisiografies-fisionomiese eenhede in die Piet Retief-studiegebied
waarin monsterpersele versprei is.
GLOOIINGS PLATO'S OF GELYKTES
Noordfront Suidfront Noordwes- Noordoos- Plato- Klipp late Laag- Res van glooiings glooiings front front trap hang liggende plato-
glooiings glooiings gedeeltes uitloper
2 3 2 2 2 2 5 10
--------- ---- -
TOTAAL
IJl
28

52
WAARNEMINGS IN EN OM MONSTERPERSELE
Floristiese waarnemings
Die aantekening van spesiesamestelling, fisionomie en sekere omgewings
faktore in en om elke monsterperseel het tot die opstel van releves gelei.
n Releve word deur Coetzee (1972) gedefinieer as n groep ekologiese en/of
plantsosiologiese waarnemings wat op n bepaalde lokaliteit betrekking het.
Die kruinbedekkingswaarde en/of getallesterkte waarde vir elke spesie wat
in die monsterperseel aangetref is, is geskat met behulp van die bedekking
skaal van die Zurich-Montpellier-benadering (Braun-Blanquet 1928, KUchler
1967 en Werger 1974b):
r: n enkele individu met n klein bedekking;
+: teenwoordig maar nie volop nie (kruinbedekking minder as 1% van
die monsterperseeloppervlakte);
1: volop (kruinbedekking minder as 1% van die monsterperseel
oppervlakte, of nie so volop nie maar met n kruinbedekking
van 1 - 5% van die monsterperseeloppervlakte),
2: baie volop (kruinbedekking minder as 5% van die monsterperseel
oppervlakte) of nie so volop nie maar met n kruinbedekking
van 6 - 25% van die monsterperseeloppervlakte;
3: kruinbedekking van 26 tot 50% van die monsterperseeloppervlakte
afgesien van die aantal individue;
4: kruinbedekking van 51 tot 75% van die monsterperseeloppervlakte
afgesien van die aantal individue;
5: kruinbedekking van meer as 75% van die monsterperseeloppervlakte
afgesien van die aantal individue.
Addisionele floristiese waarnemings is in die onrniddellike omgewing van
elke monsterperseel gedoen waartydens byvoorbeeld baie opvallende spesies
aangeteken is. Die verskillende stratums van g~stratifiseerde plantegroei
is ook volgens die Zurich-Montpellierskool aangeteken (Braun-Blanquet
~· cit.). In die huidige studie is die totale persentasie kruinbedekking
van die boom-, kruid- en grasstratum afsonderlik geskat tesame met die
hoogtes van die verskillende stratums in elke monsterperseel.

53
Fisiese omgewingsfaktore
Verskillende fisiese faktore van die omgewing het ~ invloed op die ver-
spreiding van plante (Daubenmire 1968). Die volgende fisiese omgewings-
faktore is vir elke monsterperseel bepaal, of waargeneem en aangeteken.
Geologie
Die geologiese forrnasie by elke monsterperseel is aangeteken na aanleiding
van bestaande geologiese kaarte. Vir die Arnsterdam-studiegebied is ~
kaart gebruik wat deur Hammerbeck (1977) opgestel is, en Humphrey en Krige
(1931) is vir die Piet Retief-studiegebied geraadpleeg.
Hoogte bo seespieel
Die hoogte bo seespieel van elke monsterperseel is naastenby bepaal
vanaf die bestaande 1: 50 000 Topo Kaarte (2630 DA en 2730 BA).
Aspek en belling
'n Prisma tiese kompas· is gebruik om die rig ting waarin die glooiings front
te bepaal, met noord as die magnetiese noorde.
gebruik:
N
NO
0
so
noord
noordoos
oos
suidoos
s sw w NW
Die volgende klasse is
suid
suidwes
wes
noordwes
Die belling van die glooiings is vanaf l
in die volgende klasse aangeteken:
50 000 Topo Kaarte bereken en
0/4 00 1,5/4 26 - 40°
0,5/4 1 - 15° 2/4 41 - 55°
l/4 16 - 25° 2,5/4 56 - 70°
3/4 >71°

54
Oppervlakerosie
Die volgende klasindeling is gebruik om die graad van oppervlakerosie
aan te dui (Loxton 1966, Bredenkamp 1975):
1
2
3
4
geen erosie waarneembaar nie
ligte erosie
strawwe erosie
baie strawwe erosie
Klipperigheid van grondoppervlak
'n Klasindeling, gebaseer op Theron (1973), is ·gebruik om die klipbedekking
van die grondoppervlakte te skat.
0 geen klippe op die grondoppervlakte nie
1 grondoppervlakte 1 tot 20io bedek
2 grondoppervlakte 21 tot 40io bedek
3 grondoppervlakte 41 tot 60io bedek
4 grondoppervlakte 61 tot 80i'o bedek
5 grondoppervlakte 81 tot lOOi'o bed ek
Die klipgroottes is in die volgende klasse verdeel na aanleiding van Loxton
(1966) soos deur Theron (1973) gewysig:
1 gruis met 'n deursnee van kleiner as 12 rnm
2 klein klippies met 'n deursnee van 12 tot 50 mm
3 mediumgrootte klippe met 'n deursnee van 51 tot 100 mm
4 groot klippe met n deursnee van 101 tot 200 mm
5 rotsblokke met n deursnee van groter as 200 mrn
Geomorfologie
Die betrokke geomorfologiese eenheid (kyk HOOFSTUK 5
elke monsterperseel aangeteken.
Resultate) is vir

55
Grond
Slegs die grondkonsistensie (Loxton 1966) is in die volgende klasse onder
skei:
1 los
2 sag
3 effens hard
4 hard
5 baie hard
DATAVERWERKING
n Tabel is op n skuiftafel (Muller, Werger, Coetzee, Edwards en Jarman 1972)
uit die floristiese gegewens opgestel sodat die kolomme releves en die rye
spesies verteenwoordig (Braun-Blanquet 1928, Kuchler 1967, Werger 1973
1974b). Die getal-sterktewaardes en/of kruinbedekkings van die spesies
wat in die monsterpersele aangetref is, is ook in die tabel aangetoon.
Hierdie sogenaamde "rou- tabel" is geherrangskik na 'n visuele s tudie van die
assosiasies tussen die spesies, sodat positief-geassosieerde spesies saam
gegroepeer is, waarna releves met ooreenstemmende spesies saamgegroepeer
is. Die mees algemene spesies is onderaan die tabel geplaas wanneer
geen duidelike floristiese assosiasies getoon is nie. Soortgelyke herrang
skikkings van rye ( spesies) en kolornme ( releves) is herhaal totdat 'n duide
like patroon van groepe releves met n kenmerkende floristiese samestelling
verkry is. Die tabel is gevolglik in nodums verdeel waar elke nodum
verteenwoordig word deur n aantal releves met n ooreensternmende, karakteris
tieke spesiesamestelling (Bredenkamp 1975).
Volgens Ellenberg (1956, In: Werger 1974b) is die herhaaldelike herrangskik
kings n objektiewe proses. Releves kan uit die tabel weggelaat word
wanneer dit n duidelike heterogene voorkoms bet (Werger 1974b), maar geen
releves is tydens die dataverwerking van die studie weggelaat nie.
Al die nodums wat verkry is, is bevestig deur korrelasies tussen die
plantgemeenskap wat deur die nodum verteenwocrdig word en spesifieke eien
skappe van die habitat wat tydens die opname ingewin is. Al die releves
van elke studiegebied is op twee plantsosiologiese tabelle aangetoon (Tabelle
9 en 16).

56
Sekere spesies het n hoe graad van beperktheid wat slegs tot een of enkele
nodums beperk is, en is min of meer getrou aan die een of ander gemeenskappe
waarin hulle voorkom. Alhoewel hierdie spesies wat as differensierende
spesies (Werger 1973) bekend staan wel in ander gemeenskappe teenwoordig
kan wees, kan hulle wel gebruik word om gemeenskappe te karakteriseer of
te omgrens.
Volgens Moravec (1969, In: Bredenkamp 1975) moet slegs sogenaamde karakter-
spesies gebruik word in die benaming van gemeenskappe. Differensierende
spesies wat meer getrou is aan een gemeenskap as aan ander gemeenskappe
word karakterspesies genoem (Bredenkamp ££• cit.). Dit is egter nie altyd
moontlik om hierdie karakterspesies in die praktyk te onderskei nie (Werger
1973, 1974b). Werger (1973) beveel aan dat die spesies wat inn gemeen
skap se naam genoem word ten minste teenwoordig moet wees al is dit nie
oorheersend nie.
Kruinbedekkings van beide die gras- en nie-grasagtige kruide word gesamerit
lik as n persentasie gegee. Die maksimum hoogte van die verskillende
stratums word in sommige gevalle in tabelle aangetoon.
Resultate
AMSTERDAM-STUDIEGEBIED
Alle plantegroei-eenhede wat in die Arnsterdam-studiegebied onderskei is
word in Tabel 8 weergegee, terwyl die floristiese en habitatresultate in
Tabel 9 uiteengesit word (Fig. 17).
Die Berkheya setifera - Vernonia natalensis - Eulalia villosa - grasveld
Die Berkheya setifera - Vernonia natalensis - Eulalia villosa - grasveld
kom op die sentrale plato-uitloper op n hoogte van tussen 1 370 en 1 418 m
bo seespieel voor. Doleritiese gesteentes kom algemeen in die gebied voor,
terwyl verspreide klipperige dele kenmerkend daarvan is. Die plato het
n effens golwende voorkoms met die hellings van glooiings tussen 1 tot 15°.
Die Vernonia natalensis - spesiegroep (Tabel 9) differensieer die Berkheya
setifera - Vernonia natalensis - Eulalia villosa - grasveld.

57
Tabel 8. Die plantgemeenskappe van die Mnsterdam-gebied.
Die Berkheya setifera - Vernonia natalensis - Eulalia villosa - grasveld
die Heteropogon contortus - Conyza bonariensis - Lobelia spo- grasveld
die Eulalia villosa - Hypoxis rigidula - Berkheya setifera - grasveld
die Eulalia villosa - Pentanisia angustifolia - Setaria splendida -
grasveld
die Andropogon eucomus - Helichrysum sp. - oulandgemeenskap
die Paspalum commersonii - Hyparrhenia hirta - Helichrysum sp. - grasveld
die Hyparrhenia hirta - Eragrostis racemosa - Cyperaceae - grasveld
die Trachypogon spicatus - Alloteropsis semialata - Monocymbium ceresiiforme -
grasveld
die Alloteropsis semialata - Pteridium aquilinum - grasveld
die Alloteropsis sernialata - Crassula mollus - grasveld
die Loudetia simplex - Indigofera sp. - Berkheya setifera - grasveld
die Eragrostis racemosa - Eriosema pauciflorum - Heteropogon contortus -
grasveld
die Tristachya hispida - Berkheya setifera - Panicum natalense - grasveld
die Vlei- en Riviergerneenskappe
die Eragrostis patentissirna - Echinochloa sp. - vleigemeenskap
die Riviervalleigerneenskap
die Plantasies en Uitheemse Plante

Tabel 9. 'n Plantsosiologiese tabel van die plantgemeenskappe wat in die Amsterdam-studiegebied aangetref word.
GEMlTNSKAPPI
Rclevenommer
Hoogte bo seespieCI (m)
Klipperigheid
Groottc van klippe
Geologie
Aspek
Helling
Vertrapping
"' "' @
"' ~ ~ 8
~
:: ~
> ~ ~
" ~~ Q "' <: ~ e3 ~ c.
" "' "' ~ :'5 ~ " f::: ~ r...:. '
"' " : ~ :...: ;.,.;
~ ~ "' ;.; ~ ~ "'<
.,
"" ~ . ~ ~ ~ ..... h :s ~ :"\ ~ ~
~ ;; :i '-':l ~ v < __; ~ Q ...... ::::J r_; ~ ~ ~ ~ :g 3 :::...< ._, >-. •,• ::t:: ~ \..) > v Q Q Vl ~ ...; < -< ~ .....: a ::r: N ~ ~ :.J
~-~ I ~ 'I I
· I , I I I I I I I ! I I ' ' "' I ~
'-' I o I ? P.. ~ I .....l
~~ ; ~ h I ~ ~~ ::5 ~ 0 !SE "!; ~ ~ :3 'l'; ;:: ~5 ~ t2 :5 . h :Y: ::..
c.::: !>- ~:t::: e:::..:..: ~~ C:: ~ .? ~< I V <~: ~o~~o ~ D:::> t;u; ~ V) ~o ~8 i ~~ ~ ~""""i'~...J ~ ~< o-< ~ ~ ~~ V)> , t"" z e::::;;o- ~.....; • >-.. ~:t::::::r: h . ~'-'-' o-~ ::;::;;;;: ;..:..l VJ<.r.i~> ~ .._~ '.:;:.) ~ v "'-&; ....::,9 '"""'-l w.J
~ ._,<r:,....-v.. ..... '""~ "' C...:: ~v :.:::.....: "';::;; h ~:::r:~~~ .... ~o. ~;:__ ~ h: ~x '2 i?i~ ~G o::r::o Cl...JI;;.>::..; ~ <::~ -<.;2 "'P-..; Qo :"t:O c:::"' '-':l-a....~ ::.... ~ ::,-~"- a"" v .....:;,...J ._, ~ 01 0:: ~ V)
!:--;.:.; :::~·Cl.."' a< V)~ ~< "'""':So "' ~~ "'~ ~; <> ,...., ,,:;.:: D:::VJ .....,p;:: ';;:!> ~....l ~0:: ~.. ::::::~ < c~ "'~ "'~ 5~ ~~ ~- ~~ '"~ '"~ ~~ t~ ~. V< VJ;::: VJC:. Z ';;f:c.:; ::r:~ kl...:r; V1> .....;'"'-. u....._ '-"'0 he.. <:::r: or-:: O> ->..~_ ~.._ :;::: VJ:::r: Vl Cl..~ "' ._,> "'-"" ar..:> --;:..:..: ~~<:..:..:esC :Sv ~v!~< ::."-l 0:::~ :::::~ Cl.."' c.:: '. ::::; V) ::::. ....: a ::;;;: v ""i >- V"l V) :::r: """ VJ V) :::::; "' "'" V) o a~~~;:,.~ t.;:~ ~.§ ~~ ~~So ~"'::: f:::O:::o
1
1
::_: h~ ~:S "'<::._; ~:S 2p ~..._ ~() o:;;> ~~ h~ 'ij~= I ()~ ~t; ::r::....; ~::r: ....:r:c; oz. .._ D:::~ ~::S h...:> ;...::::r:; r:::.::""-> I ~~ D:::~ ~:..:..: "':::.:: "'~ D:::< ~~ "'(~ a:::: o-....~ ::::::::.:: ~o<Zl ! h~.:.. (.).c: 1
Gj~ :3:3 st- ~s·~~:::: ::::~ "-i~ :3;s ~.(;( ~~~ i ~F: ~s:: 1, :J::...; c..;"C ~~.'io[~:t: :-:::r.r: ~~ ~~ 8~ ~~:.J 1 f::~ ~~ t·---+--: - t . ___ ____j____ I -r- ~ 21 2s 4 lR. 2019116171414249li 9 313250344533 2524! 274039267 1438443035484647 238 u/ 295 226 1::.1336371 2 ro[
I I I I I ~ ~ I ~ ~ I ~ M ~ 0 0 I 0 ~ ~ ~ ~ ~ 00 I ~ ~ i M ~· 0! M ~ ~ - ~ 0 0 M ~· ~ ~ ~ M ~ 'i - ~ ~ ~ c N ~ 00
11 ~ I ~ ~
~ 0 ~ . - 0
c C:l
;::: ~
~ ~ ~ oc
~-
0 0 0C ~ ~ M ~ ~ M ~ ~ ~ M M M M ~ 00 0 0 0 0 ~ ~ 0C ~ ~ ~ 0 0 ~ N ~ C ~ 7 - 00 00 ~ ~ M ~ N ~ 7- M M I M ~ M ~ M ~:: M M M: ~ ~ ~ ~ ~ ~ ~ - ~ M M M ~ ~ M ~ ~ ~ M ' ~ ~ M M M M M M I - ~I
000MM~IO
"!_::.!:_ •• - '<t ! -- - ---M-<--
1
c a !:. ! '"' '"' '"' @_ I '"' I 00 ClC: ~f-C :::::f-r..rJ;-f-VlVl 2.o.. 222c....o.. c...c...C:C:f-O:.t::..::IC:VJC:C:
I :3:. 00 2l::.S:3: 0 8 .o 0 o o:s= ~ o;::: oo;s:;s:o9;s:;:;: I o r./llv;<r. QZ VJVlVl <r. VlZVlZ VlVJ VJVJ ZVJ ZZ<Zl<ZlZL:VlVJ IVJ
7 ~7 ~ "d" "d" 7~-d"":t ~~ ~~ ~'<t 7 7 I ~
~~ ~~ ~~ 0~~ ~~~~~0~ ~~ ~~~~~ ~~~~~~~~ ~~~~ co oo co -- oooo o o- ooo- -NN -~oooo
I ('IM'-- <"..N ~. N I~
I ~~ --~ M---~~M NM NN--N N·-ONNNN -~--
'"' '"' ~VlOOl--f-1:-f--<f-
It;; ~ 2 ~ ~ ~
~:!-~:t~~:t:t"1"1"?.lr;"l"1 ooo-oooo
'="-
'"' ~
~
io
"'
~ 0
DIFFERENSIERENDE SPESIES VIR DIE HETEROPOGON CONTORTUS- CONYZA BONARIFNSIS- LOBELIA SP. - GKASVI-.LD
Berkheya ~P
Lohelia >p
Conyza bonariensis
+ + + + ;
+
DIFFERF.NSIERENDf SPLSILS VlR DIE EULALIA VILU}SA- HYPOXIS RIGIDULA- BERKHEYA Sl::TTFERA- GRASVELD
+
----------,--,---,--,--·-·--·-
I Q J_j_J Nidorel!a sp.
Borreria sp.
I =-l _ _L___l__==~ DIFFLRENSIERFNDF SPESIES VIR DIF EULALIA VILLOSA- PENTANISIA ANGUSTTFOLIA- SETARIA SPLENDIDA- GRASVLLD
~
. ,. I ---------Pentanisia angustifolia
Gnaphalium luteo-album
Setaria splendida
Vernonia natalensis
Rhynchelytrnm repens
+ + ; +
1 + + r I + + ; + ; + ; I ; ' 2 ++ I
DIITLRFNSll·.RENDE SPFSIFS VIR DIE ANDROPOGON EUCOMUS- HF.LICHRYSUM SP. -- OULANDGFMEENSKAP
I +
lfdichcy>UUC'P I I [] L ~-T I Andropogon euconws I + 1 1
Digitaria trichol!Jenoides I 1 I + j_ ! 1
DIHTRENSIERFNDI: SPI:SII':S VIR DIE PASPALUM COMMERSONII- HYPARRIJFNIA HIRTA- HELICHRYSUM SP.- GRASVLLD
Paspalum commersonii
Helichry~um ~p.
Stiburus conrathii
Eriosema salignum
1 + I I I I
+ + + 1
+
+ !
DIITTRLNS[J-.Rl-NDI· SPI:SIJ:S VIR DIL HYPARRHENIA HIRTA- ERAGROSTIS RACEMOSA- CYPF.RACFAL GRASVFI.D
Setariil neglecta
Gladiolus 'P-
Andropo,;on appendiculatus
Aristida sp.
Rhynchelytrnm setifolium
+ I
+ + + 2
+ 2 I I
DIH:ERENSIEIU:NDE SPLSII·:S \'IR Dll: ALLOTEROPSIS SEMIALATI\- PTERIDIUM AQUILINUM -GRASVELD
!
+
2 ;
+ +
Pte~idium aquilinum
Vernonia poskeana j -,------,-----,--------=t:l=-.~-.. -. -,--+ ~,,-------,~-1,------11
DllTERENSil·,RLNDl· SPLSIFS VIR DIE ALLOTEROPSIS SEMIALATA- CRASSULA MOLLUS- GRASVLLD
Crassula molfus
Gnidia capitata
Hypo:r:is roo peri
Alloteropsis semialata ;
+ 2 I
DIFFERIJ\SIJ·:RL~Dl SPLSILS VIR DIF LOUDETIA SIMPLEX- INDIGOFERA SP. BERKHEYA SETIFERA- GRASVJ·:LD
Aloe sp.
!ndixofera sp.
Loudetia simplex
Elyonurus muticus
Berkheya sp.
Pearsoniil se.~silzj'olia
Acalypha ~p.
Microchloa caffra
Conyza sp.
Pelarr:onium luridum
Rhus discolor
Dicoma zeyheri
Setaria nigrirostris
+ + +
--
+ + +
+ + + + ' +
+ ' 1 1 1
+
DIFFFRENSIERL·:NDL: SPI-.Sll·S VIR DIE ERAGROSTIS RACEMOSA- ERIOSEMA PAUCIPLORUM- HETEROPOGON CONTORTUS- GRASVELD
Eriosema paucijlornm
Gerhera ~p.
Ctenium concinnum
Senecio hieracioides
---...--.----+II n, ·.
DIFFERENSIERENDE SPESIFS VIR DIE ERAGROSTIS PATENTISSIMA- ECHINOCHLOA SP. - VLEIGI-_MELNSKAP
Eragrostis patentissima
Hyparrhenia aucta
Helichrysum sp.
Echinochloa sp.
Ledebouria sp.
ALGFMENE SPES!ES
Hyparrhenia hirta
Trachypogon spicatus
Themeda triandra
Berkheya setifera
Hypo xis dgidula
Eulalill villosa
Emgmstis plana
Eragrostis racemosa
Diheteropogon amplectens
Monocymbium ceresiifonne
Tristachya hispida
Heteropogon contortus
Rendlia altera
Panicum natalense
Eragrostis curvulil
Brachiaria serrata
Cyperaceae
Cymbopogon excavarus
Helichrysum nudiflorum
Hypoxis ~p
SPESIES MET 'N LAE KONSTANTIIEJD
Raphionacme ~p.
Eragrostis capensis
Acalypa sp.
Haplocarpha scaposa
Panicurn ecklonii
Justica sp.
Harpechloa falx
Thunbergia sp.
nicoma zeyheri
1/elichrysum swynnertonii
Cassia biensis
Raphionacme hirsuta
Eragrostis sp.
K oelerill cristata
Pellaea calomelanos
Eragrostis gummiflua
Acalypha schinzii
Eriosema sp.
Crotofaria sp.
Sporobolus pectinatus
Kedrostis sp.
Schistostephium crataegifolium Scilla sp.
Setaria sp.
;
; 2
I
2 I
I ; 2 '
2 2
2 I
2 2
I +
I
2
2 2
2 ;
2 2
I ;
I ;
2 3
2 1
1 ;
2 1
+
1 3
+ + I 1
2 2
1 I
1 2
I
; 2
+ 2
2 2
I
+ +
+ + + +
1
+
+
+ I
2 2
2 3
2 2 I
+
+
T
3 3 1
+ ; 1 1
1 2
2 + I
+ + I
I 1
+
+
+
32'22233
+ + 1 r +
2 2 2 + + l + r +
; + I
2 2 2
3
I
1
+ 2 3 I
I I
+ +
+ 2
1 2
I I + r !
+ + I I
+ +
2 3 2 I r I
I
+ I 1
1 2
1
1 ;
+ I
1
+ 2 + 2 2
2
I ;
+
I +
+ I +
+ +
+ + +
!
I + +
+ I
Geologie D (doloriet), SK (skalie), T (til!iet), M (mikrograniet), P (piroklastiese rots), U (ultrabasiese rots)
Die ander habitatsimbole \\ ord in die teks verk!aar
+ +
2 1
l 2 I 1 r
2 3 1 2 2
1 + 1 2 2
T + + 1 2 2 3 2 l
1 1 1 1
+ + 1
3 2 + 1 I
l 1 2 3
+ + 1 2 I 1 l 1
1 +
+
1 I
+
+
+; +
+ ! + I + + + + r +
+ +
2 1 + I
+ + + + + +
; I
+;
+ +
+ +I
+
+ + +
+
+ + +
'
+ +
+ +
I +
+ +
2 ;
+I + I I 2 I
I + I I 2 +
+
32 2212
; + I
+ + 2 2
2 2 1 +
3 2 3 2
+ I 2 2
1 I I
+I r I + + 1 2
+ I 3
I + r 2 2 + 1 I
1 1
2 1 + I + ;
2 ;
I + r + + I + I
I 1
2 ;
+
2 I
+ +
2 I 2
; ; 1
+ 2 1 2
I 2 2 r
+ ; 1
2 2 2 2 I I
r 1 2 2
2 2 + 3 3 + 3
+ +
2 + r +
+
+
2 2
I
+
33+r221
1 r r + I + 2 + 2
; 2
+ I I + 2 I I
I ;
2
+ 2
1 I I
2 I
I I 1 I
2 2 + 1 3 1 I + 2 2 I 2 3
222rl2 2
1 L 2 I
2 I I
1+1+2+ r
+
+
+ ;
+
2 2
+ I
+
I
2 I
+
+
+ +
+ +
'---
--
2 1
1 2
+ 2
+ 1
+ ' I--
+ ;
3 3

Berkheya setifera-Vernonia natalensisEulalia villosa- grasveld Andropogon el,(cOmus-Helichrysum sp.oulandgelneenskap PJl!ipalum commersonii -dHyparrhenia hirtaHel1chrysum sp.- grasvel
Hyparrhenia hirta- Eragrostis racemosaCyperaceae -grasveld
~ '
Ill ~ [IE]
Trachypogg,(l spicatu~-Alloteropsis semialata-Monocym tum ceresuforme-grasveld Loudetia simP..Iex-lnd~fera sp.-Berkheya setifera - gra eld Eragrostis racemosa- Eriosema eauciflorum-Heteropogon contortus-grasveli:l
Tristachya hiseida - Berkhej{l setifera -~niew11 nt~tll7f!n~tJ - grasv ltl
Plantgemeenskappe
~ Vlei- en Riviergemeenskappe
[I] Eragrostis curvula-Jand
t-= landerye
~ ' Uitheemse Borne

CJ Plato-uitloper
~ Platobt:rggebied
Geomorfologiese eenhede
1'8 Tallushang
liiiTI1 Pediment
==::= Riviervallei

Heteropogon contortus- Conyza bonariensisLobelia sp.-grasveld Eulalia vi/losa-HVDoxts rigidu/a- Berkheya setifera- grasverd Eulalia vilfosa-Pentanisia angustifoliaSetaria splendida-grasveld
Andropogon eucomus-Helichrysum sp.oulondgemeenskop Paspalum commersonii-
1·Hyparrhenia hirta
Heltchrysum sp.-grasve d
Hyparrhenia hirta- £ragrostis racemosaCyperaceoe-grasveld Alloteropsis semialata-Pteridium aquilinum-grasveld Alloteropsis semialata- Crassula mol/us -grosvelcf
IMiill E:3 nmrn ~ ~
Loudetia simp/Tcx-Jndigofera sp.- Berkheya setifer.t-grasve d Eragrostis racemosa-Eriosemf:. paucif/orum-Heteropogon contortus-grosve d Tris!achya hisfeida-Berkhe~a Pamcum nata ense- grasve d
setifera-
Vlei- en Riviergerneenskappe
Figuur 17. n Plantegroeikaart van die Arnsterdarn-studiegebied.
59
lkm
IL1I Eragrostis curvula-land
landerye
. li[3 Uitheemse Borne
1 tot 50 : Releyeposisies

60
Hierdie gemeenskap kan aan die hand van spesiesamestelling in die vol
gende variasies onderverdeel word:
die Heteropogon contortus - Conyza bonariensis - Lobelia sp.-grasveld,
die Eulalia villosa - Hypoxis rigidula - Berkheya setifera-grasveld, en
die Eulalia villosa - Pentanisia angustifolia - Setaria splendida
grasveld.
Die Heteropogon contortus - Conyza bonariensis - Lobelia sp. -grasveld
Hierdie grasveld word deur releves 21 en 28 in kampe 16 en 12 respektiewelik
verteenwoordig en is op die plato-uitloper op n hoogte van tussen 1 401 en
1 418 m bo seespieel gelee. Doleritiese gesteentes kom deurgaans in die
gebied voor. Die grondkonsistensie wissel van sag tot effens hard. Ligte
erosie kom in die omgewing van releve 28 voor. Die gebied is lokaal
klipperig (klein klippies) met n klipbedekking van ongeveer 20%. Die
suidfrontglooiing het n hell~ng van tussen 1 en 15°.
Verskeie spesies differensieer die grasveld, naamlik Conyza bonariensis,
Lobelia sp. en Berkheya sp. Plantsoorte met n relatiewe goeie bedekking
is Eragrostis plana, Berkheya setifera, Heteropogon contortus en
Monocymbium ceresiiforme. Ander opvallende spesies is Acalypha sp.,
Hyparrhenia hirta, Hypoxis rigidula en Panicum natalense.
Gedeeltes van die grasveld is aan vertrapping en oorbeweiding blootgestel,
soos aangetoon word deur die teenwoordigheid van Conyza bonariensis,
Eragrostis plana en Eragrostis curvula. Sekere gedeeltes van die grasveld
is ook vogtiger en word gekenmerk deur Eragrostis patentissima, verskeie
verteenwoordigers van die Cyperaceae en Andropogon eucomus.
Die kruinbedekking van die gras- en kruidstratum wissel van 65 tot 80%
(Tabel lO)o
Die Heteropogon contortus - Conyza bonariensis - Lobelia sp. - grasveld
toon nie aileen verwantskappe met die ander variasies van die Berkheya
setifera - Vernonia natalensis - Eulalia villosa - grasveld nie (uitgesonderd

Tabel 10c n Uiteensetting van die gemiddelde hoogte en kroonbedekking van die
Berkheya setifera - Vernonia natalensis - Eulalia villosa - grasveld.
Releve 21 28 4 18 20
Hoogte (m) : Kruidstratum 1 0,9 1 1 0,9
1) Kroonbedekking (%) . . 65 80 85 80 75 Kruidstratum .____
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
19
0,8 0"1
80

62
die Vernonia natalensis - spesiegroep~ maar ook met die Eragrostis
racemosa - Eriosema pauciflorum - Heteropogon contortus - grasveld aan
gesien Brachiaria serrata, Heteropogon contortus en Monocymbium ceresiiforme
hoe bedekkingswaardes in beide gemeenskappe het.
Die Eulalia villosa - Hypoxis rigidula - Berkheva setifera - grasveld.
Hierdie grasveld word deur releves 4 en 18 in kamp 11 verteenwoordig en
kom op n hoogte van tussen 1 386 en 1 408 m bo seespieel op die plato
uitloper voor. Doleriet is die enigste geologiese formasie in die ge
bied. Die grondkonsistensie wissel van sag tot hard. Klipperige
gede~ltes is kenmerkend in hierdie grasveld met n bedekking van tussen
1 en 60%. Die klipgrootte is medium tot groot. Al die suidfrontglooiings
het n helling van tot 15°.
Die Eulalia villosa - Hypoxis rigidula - Berkheya setifera - grasveld
word gedifferensieer deur Nidorella sp. en Borreria sp. Spesies wat
n hoe kruinbedekking het sluit Hyparrhenia hirta, Themeda triandra,
Eulalia villosa en Monocyrobium ceresiiforme in.
Die grasveld is matig oorbewei soos weerspieel word deur spesies soos
Nidorella sp. en Borreria sp. Op die klipperige dele is Cymbopogon
excavatus en n Aloe sp. kenmerkend. Die kruinbedekking in die releves.
wissel tussen 80 en 85% (Tabel 10).
Die Eulalia villosa - Hypoxis rigidula - Berkheya setifera - grasveld
too~ afgesien van die reeds vermelde verwantskappe met die variasies
van die Berkheya setifera - Vernonia natalensis - Eulalia villosa -
grasveld, ook n verwantskap met die Trachypogon spicatus - Alloteropsis
semialata - Monocymbium ceresiiforme - grasveld deur die teenwoordigheid
van Eulalia villosa, Eragrostis racemosa en Alloteropsis semialata.
Die Eulalia villosa - Pentanisia angustifolia - Setaria splendida - grasveld
Releves 20 en 19 in kamp 11 verteenwoordig hierdie grasveldtipe op die

63
suidelike gedeelte van die sentrale plato-uitloper op ~ hoogte van tussen
1 370 en 1 408 m bo seespieel. Soos in die geval van die vorige gras
veldtipes kom slegs doleriet in die gebied voor. Die grondkonsistensie
is deurgaans effens hard. Anders as in die Eulalia villosa - Hypoxis
rigidula - Berkheya setifera - grasveld kom hier minder klippe voor sodat
minder as 20% van die grondoppervlakte deur klippe bedek is. Net soos
in die vorige twee variasies het die suidoosfrontglooiings ~ belling van
tot 15°.
Hierdie grasveldtipe word gedifferensieer deur Pentanisia angustifolia
Gnaphalium luteo - album en Setaria splendida. Hyperrhenia hirta,
Berkheya setifera, Heteropogon contortus, Hypoxis rigidula en Eulalia
villosa het ook hoe kruinbedekkingswaardes in die gemeenskap.
Verskeie spesies se teenwoordigheid dui op versteuring, byvoorbeeld
Gnaphalium luteo - album, Paspalum cornmersonii, Eragrostis plana en
Helichrysum sp. Spesies wat kenmerkend is van vogtige toestande soos
verskeie verteenwoordigers van die Cyperaceae, dui op vogtige toestande.
Die kruinbedekking wissel tussen 75 en 80% (Tabel 10).
Hierdie grasveld toon ~ verwantskap met die Eragrostis racemosa -
Eriosema pauciflorum-Heteropogon contortus - grasveld deur die teenwoordig-
heid van Heteropogon contortus en Eragrostis racemosa. ~ Verdere ver-
wantskap word met die Eragrostis patentissima - Echinochloa sp. - vleigemeen
skap getoon deur Eragrostis patentissima, Hyparrhenia aucta, Helichrvsum
sp. en verskeie verteenwoordigers van die Cyperaceae.
Die Andropogon eucomus - Helichrysum sp.-oulandgemeenskap.
Enkele oulande kom in die studiegebied voor waarvan slegs twee ~ noemens
waardige oppervlakte beslaan (kampe 15 en 17). Hierdie oulandgemeenskap
word deur releves 16 en 17 in kampe 17 en 15 respektiewelik verteenwoordig.
Die oulande is op die plato-uitloper, met doleriet as onderliggende
gesteente, op ~ hoogte van tussen 1 386 en 1 439 m bo seespieel gelee.
Die grondkonsistensie is deurgaans effens hard met verspreide swak

64
gedreineerde gedeeltes, terwyl geen klippe op die grondoppervlakte
voorkom nie. Die noord- en oosfrontglooiings het ~ belling van tot 10°.
Verskeie spesies is kenmerkend van die oulandgemeenskap, veral Andropogon
eucomus, Digitaria tricholaenoides en Helichrysum sp. Eragrostis curvula,
Eragrostis plana en Eragrostis racemosa is ook baie opvallende komponente
van hierdie gemeenskap.
~ Relatiewe lae kruinbedekking van tussen 45 en 60% word aangetref.
~ Opvallende verskil tussen die oulandgemeenskap en die naasliggende
Eragrostis racemosa - Eriosema pauciflorum - Heteropogon contortus -
grasveld is die feitlik algehele afwesigheid van Trachypogon spicatus,
Eulalia villosa, Diheteropogon amplectens, Monocymbium ceresiiforme en
Heteropogon contortus. Die volgende spesies dui egter op ~ verwantskap
tussen die twee gemeenskappe : Eragrostis plana, Eragrostis racemosa en
Andropogon eucomus. Andropogon eucomus dui verder ook op nat toestande.
Die Paspalum commersonii - Hyparrhenia hirta- Helichrysum sprgrasveld.
Hierdie grasveld word deur releves 42 (kamp 5), 41 en 49 (kamp 9) verteen
woordig. Die grasveld word op die suidwesfron~tallushang, met sandsteen
en skalie as die vernaamste onderliggende gesteentes, op ~ hoogte van 1 326
tot 1 386 m bo seespieel aangetref. ~ Effens harde tot harde grondkonsis
tensie kom voor, terwyl tot 40% van die grondoppervlakte deur gruis en
klein klippies bedek is. Die suidwesfrontglooiings het ~ belling van
tussen 15 en 25°.
Die Paspalum commersonii - Hyparrhenia hirta - Helichrysum sp.- grasveld
word deur Paspalum commersonii sp., Stiburus conrathii en Eriosema salignum
differensieer. Ander spesies met~ hoe bedekking sluit Hyparrhenia hirta
en Eragrostis plana in terwyl Berkheya setifera, Eulalia villosa en Rendlia
altera ~ definitiewe aspek aan die gemeenskap verleen.
Oorbeweiding en vertrapping word weerspieel deur die teenwoordigheid van
spesies soos Eragrostis plana, Helichrysum sp., Paspalum commersonii en

65
Aristida sp. Die voorkoms van Stiburus conrathii en Paspalum cornmersonii
dui opvogtige grond. n Kruinbedekking van tussen 55 en 60% kom voor.
Hierdie gemeenskap toon slegs n verwantskap met die Hvparrhenia hirta -
Eragrostis racemosa - Cyperaceae grasveld deur die gesamentlike teen
woordigheid Eragrostis plana en Rendlia altera.
Die Hyparrhenia hirta - Eragrostis racemosa - Cyperaceae - grasveld
Die Hyparrhenia hirta - Eragrostis racemosa - Cyperaceae - grasveld word
deur releves 9, 31, 45 (kamp 18), 32, 50 (kamp 20), 34 en 33 (kamp 19)
verteenwoordig. Die meeste releves (9, 31, 32 en 50) is op die coste
like gedeelte van die plato-uitloper gelei (met onderliggende gesteentes
soos sandsteen en skalie, tilliet asook doleriet en ultrabasiese rots),
terwyl die ander releves (34, 45 en 33) op die pediment gelee is. Hierdie
gemeenskap kom op n hoogte tussen 1 307 en 1 373 m bo seespie~l voor.
Die gronde bo-op die plato-uitloper is meestal vlak en swak gedreineer
met n effens harde tot harde konsistensie in teenstelling met die
dieper gronde van die pediment waar die grondkonsistensie sag tot effens
hard is. Dagsome kom verspreid op die plato voor, maar die klipbedekking
is oor die algemeen laag terwyl gruis tot groot klippe tot 50% van die
pediment se grondoppervlakte bedek. Die noordoos- en suidoosfront
glooiings het n helling van tot 35°.
Hierdie gemeenskap word gedifferensieer deur Andropogon appendiculatus,
Setaria neglecta en n Gladiolus sp. asook verskeie verteenwoordigers van
die Cyperaceae. Hyparrhenia hirta het die hoogste kruinbedekking terwyl
Berkheya setifera ook n belangrike komponent van die gemeenskap is.
Ander algemene spesies is Eragrostis plana, Eragrostis racemosa, Rendlia
altera, Eragrostis curvula, Trachypogon spicatus en Hypoxis rigidula.
Alhoewel Eulalia villosa minder konstant voorkom verleen dit wel n definitiewe aspek aan die gemeenskap.

66
Gedeeltes van die gemeenskap is aan vertrapping en oorbeweiding bloot
gestel, soos aangetoon word deur die teenwoordigheid van spesies soos
Aristida sp., Rhynchelytrum repens, Eragrostis plana, Eragrostis curvula
en Haplocarpha scaposa. Verskeie spesies, kenmerkend van vlak en
vogtige grande, kom voor soos Andropogon appendiculatus en verteenwoordi
gers van die Cyperaceae. Die kruinbedekking wissel tussen 50 en 80%
met die hoogste bedekking by die laer-liggende releves (Tabel 11).
Die Hyparrhenia hirta - Eragrostis racemosa - Cyperaceae - grasveld toon
verwantskappe met die aangrensende gemeenskappe deur die teenwoordigheid
van verskeie spesies:
(a) Die Eragrostis racemosa - Eriosema pauciflorum - Heteropogon
contortus - grasveld deur Eragrostis racemosa en verteen
woordigers van die Cyperaceae.
(b) Die Tristachya hispida - Berkheya setifera - Panicum
natalense - grasveld deur Panicum natalense, Eragrostis
plana, Eragrostis curvula en ~ Hypoxis sp.
Die Trachypogon spicatus - Alloteropsis semialata - Monocymbium ceresiiforme -grasveld.
Hierdie gemeenskap word deur releves 25, 26 (kamp 1), 27, 40, 39, 26 (kamp 4)
en 7 (kamp 3) op die suid- en suidoosfrontglooiings van die platoberggebied
op ~ hoogte van tussen 1 386 en 1 521 m bo seespieel verteenwoordig.
Gesteentes van die Usushwana-kompleks kom in hierdie bergagtige gebied
voor. Die glooiings het ~belling van tussen 15 en 40°.
Die Trachypogon spicatus - Alloteropsis semialata - Monocymbium ceresiiform~
grasveld word hoofsaaklik deur Alloteropsis semialata gedifferensieer en
kan aan die hand van spesiesamestelling en fisionomie in twee variasies

Tabel 11. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van die
Hyparrhenia hirta- Eragrostis racemosa- Cyperaceae-grasveld.
Releve 9 31 32 50 34
Hoogte (m) : Kruidstratum 0,2 0,5 0,6 0,8 1
1) Kroonbedekking (%) : 50 55 65 60 80 Kruidstratum -~-----------~----- ------~--- --- ------- --~-- -- -------- -- --- ---
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
45
0,6
70 --- ---
33
0,9
60 -- ---
0\ -....,J

68
verdeel word, naamlik:
die Alloteropsis semialata - Pteridium aguilinum - grasveld en
die Alloteropsis semialata - Crassula mollus - grasveld.
Die Alloteropsis semialata - Pteridium aguilinum - grasveld
Hierdie grasveld word verteenwoordig deur releves 25 en 24 in kamp 1 en
kom op die platoberggebied tussen 1 449 en 1 516 m bo seespieel voor.
Piroklastiese rots, riolieten mikrograniet vorm die onderliggende gesteen
tes. Die grond het ~ sagte tot harde konsistensie. Tussen 21 en 40%
van die grondoppervlakte word deur klein tot mediumgrootte klippe bedek.
Steil glooiings van tot 40° wat in ~ suidwestelike rigting front is ken
merkend van die noordwestelike gedeelte van die platoberggebied.
Die grasveld word gedifferensieer deur Pteridium aguilinum en Vernonia
poskeana. Plantspesies met ~ relatiewe hoe kruinbedekking in die
gemeenskap sluit soorte soos Alloteropsis semialata, Themeda triandra,
Monocymbium ceresiiforme, Diheteropogon amplectens en Trachypogon spicatus
in.
Die veld bet ~ meer heterogene voorkoms teen die laer hange wat moontlik
aan vertrapping en matige oorbeweiding toegeskryf kan word. Eragrostis
racemosa, Rhynchelytrum repens en Helichrysum nudiflorum is opvallende
komponente in hierdie laagliggende grasvelddele. Hyparrhenia hirta wat
'n hoe konstantheid het, is hier opvallend swak verteenw·oordig. Die
kruinbedekking wissel tussen 50 en 70% (Tabel 12).
Die Alloteropsis semialata - Pteridium aguilinurn - grasveld toon geen
duidelike verwantskappe met enige ander plantgemeenskap nie, behalwe
met die naasliggende Alloteropsis semialata - Crassula mollus - grasveld
in die gesamentlike teenwoordigheid van veral Monocymbium ceresiiforrne
Heteropogon contortus en Eragrostis racemosa.
Die Alloteropsis semialata - Crassula mollus - grasveld
Die grasveld word.deur releves 27, 30, 39, 26 (kamp 4) en 7 (kamp 3) op

Tabel 12. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van die Trachypogon
spicatus - Alloteropsis semialata - Monocyrnbium ceresiiforme - grasveld.
Releve 24 25 26 27 39 40
Hoogte (m) : Kruidstratum 0,4 1 1,2 1 1 0,9
1) Kroonbedekking (%) . . 50 70 85 70 90 80 Kruidstratum
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
7
0,4
50
Q'\ 1..0

70
die suidoostelike gedeeltes van die platoberggebied en plato-uitloper,
met onderliggende rioliet en piroklastiese rots, op n hoogte van 1 386
en 1 453 m bo seespieel verteenwoordig. Groot en mediumgrootte klippe
bedek minder as 20% van die grondoppervlakte. Die glooiings, wat in
n suidwestelike, suidelike en suidoostelike rigting front, het n belling 0 van tot 40 •
Crassula mollus, Gnidia capitata en Hypoxis rooperi is differensierend
uit die grasveld. Die volgende spesies het ook relatiewe hoe kruin
bedekkingswaardes in die gemeenskap : Trachypogon spicatus, Themeda
triandra, Berkheya setifera Eulalia villosa, Eragrostis racemosa,
Monocymbium ceresiiforme en Rendlia altera.
Die grasveld is oor die algemeen in n goeie toestand en is slegs lokaal
gedeeltelik oorbewei, soos aangetoon word deur die teenwoordigheid van
spesies soos Rhynchelytrum repens, Helichrysum sp., en n Crotolaria sp.
Sekere spesies, tiperend van klipperige gedeeltes, word ook aangetref
onder andere Rendlia altera, Cvmbopogon excavatus en Hypoxis rooperi.
n Kruinbedekking van tussen 50 en 90% kom voor (Tabel 12).
Die Alloteropsis semia1ata - Crassula mollus - grasveld toon, afgesien
van die verwantskap met die naasliggende Alloteropsis semialata -
Pteridium aguilinum - grasveld, ook n verwantskap met die Eragrostis
racemosa - Eriosema pauciflorum - Heteropogon contortus - grasveld deur
die gesamentlike teenwoordigheid van Heteropogon contortus en Tristachya
hispida.
Algemeen
Fisionomies behoort releve 7 tot die Alloteropsis semialata - Pteridium
aguilinum - grasveld van die stei1 suidoosfrontglooiings van die plato
berggebied, maar is weens spesiesamestelling onder die Alloteropsis
semialata - Crassula mollus - grasveld geplaas.
Die Loudetia simplex - Indigofera sp. - Berkheya setifera - grasveld.
Hierdie grasveld word deur releves 35, 46, 47 (kamp 18), 14, 38 (kamp 2),
48 (kamp 15), 30 (kamp 11) en 44 (kamp 10) verteenwoordig. Die re1eves

71
is teen klipperige glooiings tussen 1 370 en 1 496 m bo seespieel op
die plato-uitloper en platoberggebied gelee. Doleriet en skalie is
die vernaamste onderliggende gesteentes. n Sagte tot harde grond
konsistensie kom voor, terwyl tussen 20 en 60% van die grondoppervlakte
deur klein of groot klippe bedek is. Die meeste releves kom teen noord
oosfrontglooiings (releves 14, 35, 38 en 46) en suidwesfrontglooiings
(releves 30, 44 en 47) voor. Hierdie glooiings het n belling van tussen
15° en 25°.
Die Loudetia simplex - Indigofera sp. - Berkheya setifera - grasveld
word deur verskeie spesies gedifferensieer waarvan Aloe sp., Elyonurus
muticus, Rhus discolor en Dicoma zeyheri die vernaamste is. Hyparrhenia
hirta, Trachypogon spicatus, Themeda triandra en Berkheya S€tifera het
relatiewe hoe bedekkingswaardes in die gemeenskap.
Gedeeltes van die gemeenskap is aan swaar beweiding en vertrapping
blootgestel (veral releves 14 en 38) soos aangetoon word deur die teen
woordigheid van spesies soos Eragrostis plana, Aristida spp. en
Rhynchelytrum repens. Ligte erosie kom lokaal voor. Die kruinbedekking
wissel tussen 40 en 80% (Tabel 13).
Die Loudetia simplex - Indigofera sp. - Berkheya setifera - grasveld toon
n verwantskap met die Eragrostis racemosa - Eriosema pauciflorum -
Heteropogon contortus - grasveld deur die teenwoordigheid van Loudetia
simplex, Conyza sp. en Ctenium concinnumm beide gemeenskappe. Elyonurus
muticus dui op n verdere verwantskap met die Tristachya hispida - Berkheya
setifera - Panicum natalense - grasveld.
Die Eragrostis racemosa - Eriosema pauciflorum - Heteropogon contortus grasveld.
Die Eragrostis racemosa - Eriosema pauciflorum - Heteropogon contortus -
grasveld word deur releves 23 (Kamp 15), 8, 1 (Kamp 16) en 11 (Kamp 11)
op die noordelike gedeelte van die sentrale plato-uitloper op n hoogte
van tussen 1 385 en 1 424 m bo seespieel verteenwoordig. Die gemeenskap
kom hoofsaaklik op doleriet en gedeeltelik op sandsteen en skalie voor.
Vlakkerige gronde kenmerk die gebied, terwyl die grondkonsistensie van
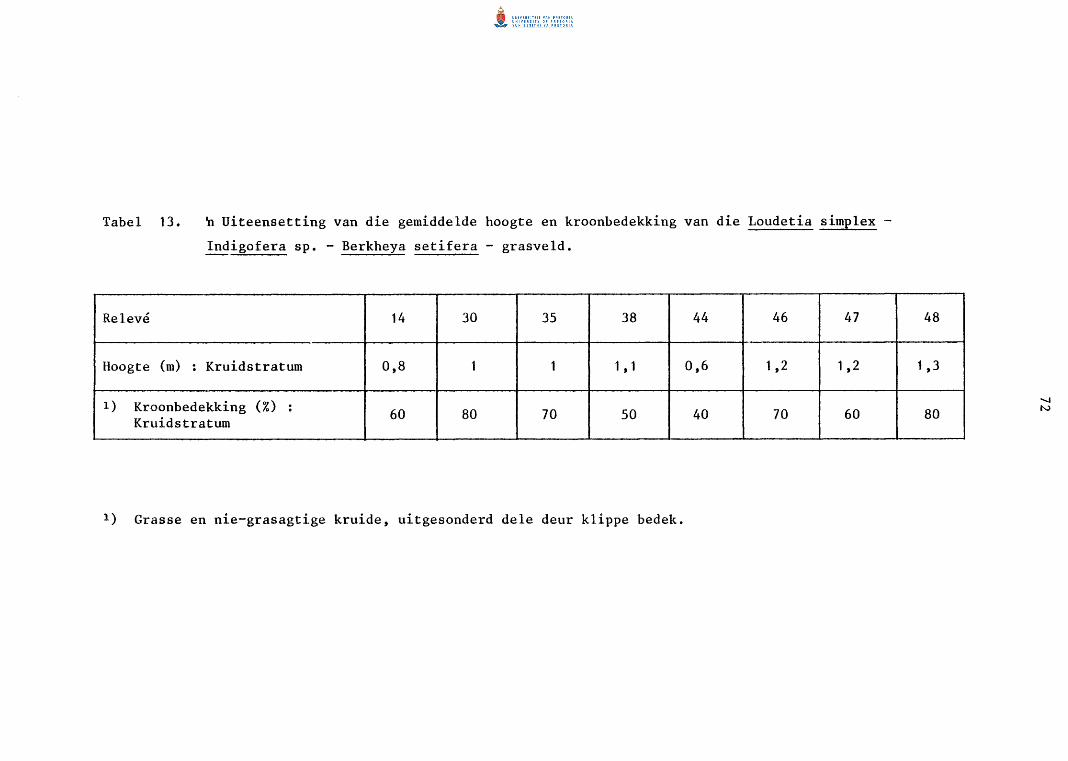
Tabel 13. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van die Loudetia simplex -
Indigofera sp. - Berkheya setifera - grasveld.
Releve 14 30 35 38 44 46 '• 7
Hoogte (m) : Kruidstratum 0,8 1 1 1 '1 0,6 1 '2 1 '2
1) Kroonbedekking (%) : 60 80 70 50 40 70 60 Kruidstratum
------- --- L.___ __ ----- - -------- --- --
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
48
1 '3
80 ...... N

73
effens hard tot hard wissel. Die hele gebied is klipperig en gruis en
klein klippies tot mediumgrootte klippe bedek in sommige gevalle tot
40% van die grondoppervlakte. Die suidoos- en noordoosfrontglooiings
het n helling van minder as 10°.
Eriosema pauciflorum, Senecio hieracioides en n Gerbera sp. differensieer
hierdie gemeenskap. Plantspesies met n hoe kruinbedekkingswaarde sluit
Eragrostis racemosa en Heteropogon contortus in. Die hele gebied, uitge
sonderd kampe 11 en 15 (releves 11 en 13 respektiewelikh word periodiek
aan hoe weidingsdruk onderwerp en toon tekens van uittrapping. Die teen
woordigheid van n Conyza sp. dui op versteuring. Andropogon eucomus en
Senecio hieracioides en verskeie teenwoordigers van die Cyperaceae is
kenmerkend van swak gedreineerde en vlakkerige grande. Die feitlik totale
afwesigheid van Berkheya setifera en Eulalia villosa, wat algemeen op
die res van die plato-uitloper in die Eulalia villosa - Hypoxis rigidula -
Berkheya setifera - grasveld en die Eulalia villosa - Hyparrhenia hirta -
Pentanisia angustifolia - grasveld aangetref word, is opvallend. n Kruinbedekking van tot 65% kom voor.
Verwantskappe met die Loudetia simplex - Indigofera sp. - Berkheya setifera -
grasveld en die Hyparrhenia hirta - Eragr~stis racemosa - Cyperaceae -
grasveld is reeds bespreek.
Die Tristachya hispida - Berkheya setifera - Panicum natalense - grasveld
Hierdie gemeenskap, verteenwoordig deur releves 29 (Karnp 21), 5 (Kamp 6),
22, 6, 13 (Karnp 5), 12 (Karnp 4), 36 en 37 (Kamp 8) korn op die suidelike
deel van die plato-uitloper (releves 29, 5 en 22), die tallushang (re1eve
6) sowe1 as die pediment (re1eves 13, 12, 36 en 37) op n hoogte van tussen
1 359 en 1 442 m bo seespieel voor. Verskeie geo1ogiese forrnasies vorrn
die onder1iggende gesteentes, waarvan ti1liet en f1uviog1asiale sediment
(re1eves 6, 12, 13, 36 en 37) die belangrikste is. U1trabasiese rots
(re1eve 29) en doleriet (re1eve 5) is ook aanwesig. Die grondkonsisten
sie wisse1 van sag tot hard, met die 1aer1iggende re1eves (re1eves 12, 13,
36 en 37) verteenwoordigend van die harder grande.

74
Slegs een releve (releve 5) was in u klipperige omgewing gelee met klein
klippies wat tot 20% van die grondoppervlakte bedek. Enkele dagsome,
wat nie gemonster was nie, kom op die pediment voor.
suidoosfrontglooiings het n belling van tot 25°.
Die noordoos- en
Alhoewel die gemeenskap deur negatiewe assosiasies gekenrnerk word aangesien
n differensierende spesiegroep ontbreek, het bepaalde spesies soos
Panicum natalense, Eragrostis plana en Tristachya hispida nogtans n hoe
bedekking in die gemeenskap. Berkheya setifera kom hier minder konstant
voor, maar verleen wel n definitiewe aspek aan die gemeenskap. Eulalia
villosa en Themeda triandra, wat n relatiewe hoe konstantheid in meeste
van die ander gemeenskappe het, is ook swak in die gemeenskap verteen
woordigc
n Groot deel van die gemeenskap is aan vertrapping blootgestel. Hierdie
word beklemtoon deur die teenwoordigheid van spesies soos Paspalum
conunersonii, Eragrostis curvula en Eragrostis plana. Die laerliggende
gedeeltes word gekenmerk deur vogtiger gronde waar die volgende spesies
opvallend is : Andropogon eucomus, Digitaria tricholaenoides en verskeie
verteenwoo~digers van die Cyperaceae. Die kruinbedekking wissel tussen
45 en 90% (Tabel 14).
Die Tristachya hispida - Berkheya setifera - Panicum natalense - grasveld
toon,afgesien van die reeds bespreekte verwantskap met die Loudetia simplex
Indigofera sp. - Berkheya setifera - grasveld, geen ander duidelike verwant
skappe nie.
Die Vlei- en Riviervalleigemeenskappe.
Die Eragrostis patentissima - Echinochloa sp. - vleigemeenskap·
Op die hoerliggende gedeeltes word vleigebiede in sekere gedeeltes
aangetref. Alhoewel hierdie gemeenskappe met 'n kenmerkende plan tegroei
n beperkte omvang het, is dit wel gemonster aangesien die vleigemeenskappe
dikwels 'n belangrike komponent van die oorbietjie-tuisgebiede uitmaak.

Tabel 14. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van die Tristachya hispida -
Berkhey~setifera- Panicum natalense- grasveld
Releve 29 5 22 6 12 13 36
Hoogte (m) : Kruidstratum 1 ,0 1,0 1 '2 1,0 0,9 0,8 0,8
1) Kroonbedekking (%) . . 80 85 90 75 Kruidstratum 70 70 45
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
37
0,6
60
I -....,J
L11

76
Die Eragrostis patentissima - Echinochloa sp. - vleigemeenskap word
deur releves 2 (kamp 12) en 10 (Kamp 11) op die sentrale plato-uitloper,
gelee op n hoogte van tussen 1 370 en 1 424 m bo seespieel, verteenwoordig.
Doleriet vorm deurgaans die vernaamste onderliggende gesteentes in die
gebied, terwyl die gronde, met n konsistensie van sag tot effens hard,
vir die grootste gedeelte van die reenseisoen vogtig is. Klippe is
afwesig. Hierdie vleigemeenskappe kom teen glooiings met n helling
· d 1- 0 van m1n er as ) voor.
Hierdie gemeenskap word gedifferensieer deur Eragrostis patentissima,
Eragrostis plana, Hyparrhenia aucta en n Ledebouria sp. asook n groot
aantal verteenwoordigers van die Cyperaceae. Die Eragrostis patentissima -
Echinochloa sp. - vleigemeenskap is n duidelike omgrensende gemeenskap
wat, afgesien van die reeds bespreekte verwantskap met die naasliggende
Eulalia villosa - Pentanisia angustifolia - Setaria splendida - grasveld,
geen ander duidelike verwantskappe toon nie.
Riviervallei-gemeenskappe
Die riviervalleie het n kenmerkende plantegroei, maar aangesien dit n baie beperkte omvang het en vir die doel van hierdie studie minder belang
rik is, is dit nie gemonster nie. Verskeie spesies soos Hyparrhenia
aucta en verteenwoordigers van die Cyperaceae vorm oral digte stande langs
die spruite en riviere.
Plantasies en ander Uitheemse Plante
n Aantal kleiner plantasies kom verspreid deur die studiegebied voor.
Die belangrikste uitheemse boom is die wattelboom (Acacia mearnsii) wat
in die sentrale en westelike gedeeltes van die gebied aangeplant is.
Wattelbome het vanaf die aangeplante dele, veral langs die spruite,
versprei. n Groot aantal jong boompies word in hierdie dele aangetref
asook verspreid in ander laagliggende gedeeltes.

77
Natuurlike plantegroei is feitlik heeltemal afwesig onder die wattelbome,
wat oral digte stande vorm. Die uitbreiding van die wattelboom se
verspreidingsgebied hou dan ook ~ ernstige bedreiging vir die natuurlike
plantegroei in.
~Ander belangrike uitheemse boom in die studiegebied is die bloekomboom
(Eucalyptus sp.) wat verspreid in lanings aangeplant is. Alhoewel
natuurlike plantegroei onder die borne ook feitlik afwesig is, is daar
geen aanduidings dat hierdie boom verder versprei en ~ onmiddelike gevaar
vir die natuurlike plantegroei inhou nie. Ander uitheemse borne wat in
beperkte getalle, hoofsaaklik in die omgewing van die woonhuis voorkom,
sluit dennebome (Pinus spp.) in.
DIE PIET RETIEF-STUDIEGEBIED
Die plantgemeenskappe wat in die Piet Retief-Studiegebied onderskei is,
word in Tabel 15 saamgevat, terwyl die floristiese en habitatgegewens
in Tabel 16 uiteengesit word (Fig. 18).
Die Xerophyta sp. - Oldenlandia herbacea - klipplaatplantegroei.
Hierdie gemeenskap word deur releves 17 (Kamp Ra8), 23 (Kamp D), 8 (Kamp M7)
en 3 (Kamp M5) op die plato-uitloper met onderliggende Mandlangampisi
dolerietplate, op ~ hoogte van tussen 1 933 en 1 575 m bo seespiegl ver-
teenwoordig. Verskeie dagsome kenmerk die gebied.
Die gemeenskap, wat deur die Xerophyta sp.- spesiegroep gedifferensieer word,
kan op grond van spesiesamestelling en fisionomie in twee variasies
verdeel word, naamlik
die Aristida sp. - Pellaea calomelanos - Xerophyta sp. -
klipplaatplantegroei, en
die Aristida junciformis - Euphorbia pulvinata - klipplaatplante
groei.

78
Tabel 15. Die Plantgemeenskappe van die Piet Retief-studiegebied.
Die Xerophyta sp. - Oldenlandia herbacea - klipplaatplantegroei
die Aristida sp. - Pellaea calornelanos - Xerophyta sp. - klipplaat
plantegroei
die Aristida junciformis - Euphorbia pulvinata - klipplaatplante
groei
Die Aloe rnarlothii - Crassula sp. - struikveld
die Diospyros whyteana - Aloe rnarlothii - struikveld
die Cymbopogon excavatus - Rhus eckloniana - Aloe rnarlothii -
struikveld
die Loudetia simplex - Trachypogon spicatus - Rendlia altera - grasveld
die Monocymbiurn ceresiiforme - Diheteropogon arnplectens - Tristachya
hispida - grasveld
die Alloteropsis sernialata - Trachypogon spicatus - grasveld
die Berkheya sp. - Helichrysurn sp. - Tristachya hispida - grasveld
die Gerbera sp. - Eulalia villosa - Eragrostis plana - grasveld
die Kloofbosgerneenskap
die Vleigemeenskap
die Plantasies en Uitheernse Plante


r::::::::::::l ~
~ 0 0
Aristida sp.- Pel/aea calomelanos- . Xerophyta sp.-klipplaatplaatplantegroeJ Aristida junciform~s-Euphorbia pu/vinata-klipplaatplantegroeJ .. Diosperos whyteana-Aioe marlothustruikveld Cymbopogon excavatus-Rhus ecklonianaAioe marlothii-struikveld Loudetia simplex-Trachypogon spicatusRendlia altera-grasveld
Monocymbium ceresiiforme -Diheteropogon amplectens-Tristachya hispida- grasveld Al~oteropsis semialata- Trachypogon sp1catus-grasveld Be_rkheya sp.- Helichrysum sp.Trtstachya hispida- g rasveld
·····EB ~ ·Ill
80
Gerbera sp.- Eulalia villosa- Eragrostis planagrasveld
Vlei- en Riviergemeenskappe
Kloofbosgemeenskap
Oulande
Figuur 18. n Plantegroeikaart van die Piet'Retief-studiegebied.
• • • • l d ... ·.··:· an erye
~ Uitheemse Borne
2 tot 29 : Refeveposisies
N
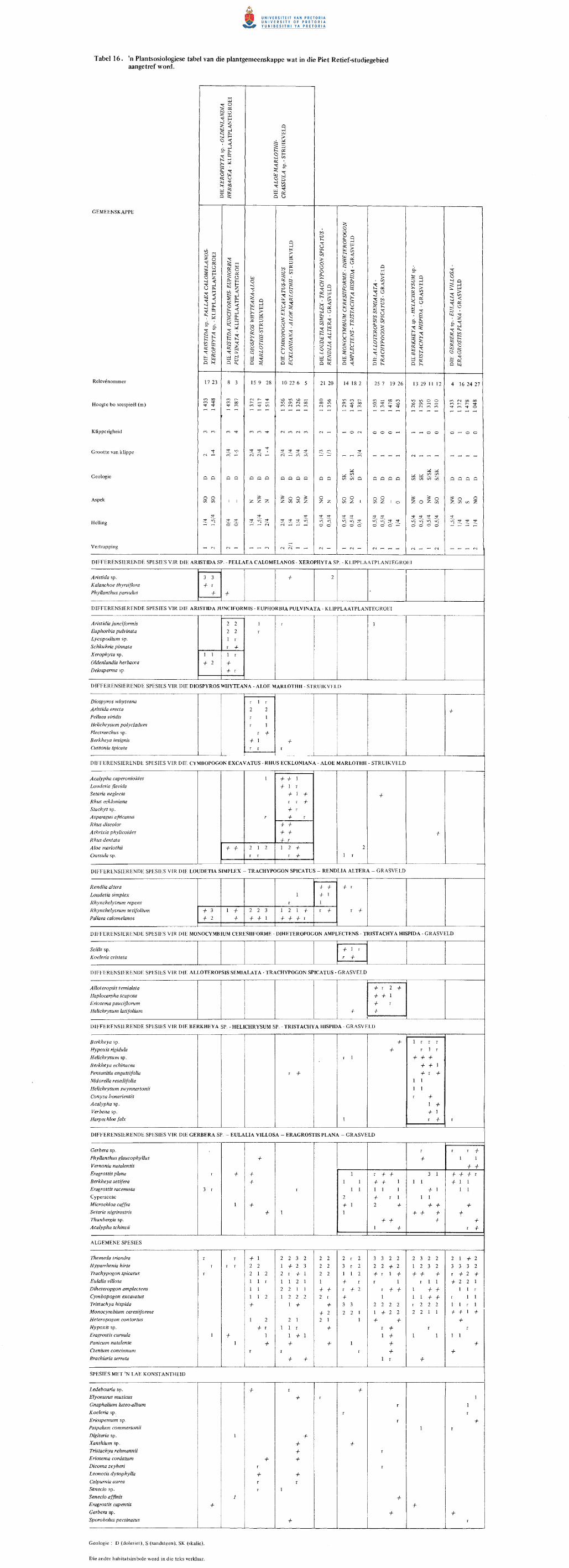
Tabcll6. 'n Plantsosiologiese tabel van die plantgemeenskappc wat in die Piet Retief·studiegebied aangetref word.
GEMFFNSKAPPE
Re\evenommcr 10 22 6 5 21 20 14 18 2 25 7
Hoo~Le bo scc.~picel (m)
19 26
KlippcJigheid 0 0 c ......
Groottc van klippe ~-~;g-~-1::
Geologie I ~ " a a " " p
a o
Aspek z ~ z 0 7, z I I
Helling _;t_ .'!_ 0 c
13 29 II 12
:t "1" :I: :"" ..;;~"'0 0 0 0 0
Vertrarping ! N - - - I N - - N
DH·'FJ::Rr_NSIERl:J\DI: SPLSWS VlR DIE ARIST[[)A SP.- PELLAEA CALOMELAKOS- XEROF'HYTA SP.- KT.IPPl AATPT.ANTEGROLI
4 16 24 27
::::I
Aristida sp.
Kalanchoe thyrsijlora
Phyllo.nthus pan•ulus
3 3
+I +
~ --~~- - ~~ -
+ I j + 2 I r-
-~- _ _____]_________~--------'------
DIFFI:.RENSIERL-:NDE SPESIES VJR DIF ARTSTIDA HINCTFORMTS- FUPHORHIA PULVINATA- KLJPPLAATPLANTt-:C_;RQU
---~~-------~----~~----- ~1-
Ari~·tidia juncif(mnis 2 2
Euphorbia pulvinata _
Lycopodium sp. l r
SchkuhrW pin.nara r + Xerophyta sp. I l 1 r
I
I
(JldenlmuliLJ herhacea + 2 + I Delosperma sp + r I l
- ---~-------L--~ ______ L_ ___ _
DIFFERENSILRENUE SPJ:SJES VIR DIF OIOSPYROS WHYTE ANA- ALOE MARL01'HII- STRUIK VELD
l
-----------~- -~- ~ -~ - --- ~---_-_~ _ _,. -.-----,-----~--------~-------,----
1 !
Diospyros whyteana
Aristida erect a
PeUaea viridis
Helichrys.ttm polycladum
Plectranthu.s ~p.
Berkheya insignis
Cussonia spicala
+ 1
I.
2 I
1
+ +
+
I 1 1 i __ l__ _ _l ___ _L ____ _j _____ _!_l ___ -1 DIFFERENSIERI::t'··."DL SPF:SIES VIR DIF CYMllOPOGON EXCAVATUS- RHUS ECKLONIA..KA- ALOE MARLOTifii- STRUJKVELD
--~---------~~~~----F====1---r--~----~----~-- ---Acalypha r:apemnioides I + + 1
Loudetia j7udJa + 1 r
Setaria neglecta + I + Rhus eckloniana r 1 + Stachy'l sp. + r
Asparagus afriranus
Rhus discolor
Alhrixia phylicoides
Rhus dentata
+ + + + + '
+
I
Aloe marlothii
Cra~~·uia ::;p. ____________ _j __
+ + 2 l 2
' ' I 2 +
' + 1 ' ____ j__ D!Ff'ERI.:NS!ERFNDF SPESII~S VIR DIF JJ)UDFTIA SI\IPLEX - TRACHYPOGON SPlCATCS- RFNDLTA AT TFRA- c;.RASVF.I D
Rendlia alter a
Loudetia 'Simplex
Rhynchelytrnm repens
Hhynchelytrum se t1jolium
Pallaea calomelanos
+ + +I I
+ '
' +
I
I
--
+3 1+ 223 J2lf- r+ 1
.
+2 + ++l +++r -.r===C==±====C===~===L--~----~----_L ____ _
DIFFFRfNSTERFNDr SPf7SWS VIR DTr MONOCYMTHU\1 Cl'RESHI'ORMF • nTHETFROPO(;QN A~1PLECTENS- TRISTACHYA HISPIDA · GRASVELD
Scilla sp.
Koeleria cristata lJJ Dll;l ERENSIERLNDE SPJ::Sll~S VIR OIL ALLOTEROPSlS SHUALATA- TRACH'r'POGON SPICATUS- GRASVELD
Alloterop~'is scmialala
I fapl1 warpha scaposa
F:riosema pm.u. .. ·i:tfomm
Hetichry.nlm latifolium
I
--------------------~----L---~----_L------~--l~
~--
+ r 2 + + + 1
+ ' +
DiFFLRFNSJLRENDE SYLS!I::S VIR DIE BERKHI:: YA SP.- HELICHRYSUM SP.- TRISTACUYA UISPIUA- GRASVELD
!
I
·~--~--~----~---,---~----~====~-----~ Berkhcya ~p.
Hypo:.,- is rigidula
Helichryrum sp.
Berkheya echinacea
Pentanisi.a angustifolia
.fllidorella resedij'olia
!Jelichrysum sw.vnnertvnii
Cvn.r·za honariettSi'l
A r:alypha ~p.
Verbena sp.
ffarpcchloa falx
' 1
' +
I Dlrl:ERENSIERENDE SPESIES VIR DIE GERBERA SP. -EULALIA VILLOSA- ERAGROSTIS PLANA- GRASVELD
Cerbera sp.
Phyllanthui glaucophyllus
Vernonia natalensis
~---T-. ------,---~ -------,~--- -
I
+
+ + 1 r r r
' 1 ' + + +
+ + 1
+ ' + I 1
I 1
+ 1 + 1- I
' 1-
- ~-----~
+
3 I Eragrostis pkma j 1
-/- r + + Berkheya setifera + ! I + + 1
Eragrmtifl racemosa 3 r r 1 I 1 1 I + I
Cypt~raccac 2 + r 1 1 1
' + 1 1
+ + + + + r
+ 1 I
I 1
+ Microchloa caffra 1 + 1 + 1 2 + + +
s_._'_"_"_a~_n_i_g'_'_'o_s_"_'_'-----------'----'----~'- + I ____ L __ _j'::::1===::::11::::=====±=/-=~~/~==~+-:i'l---.... ~+ ............ :::1 Thu.nbergia s.p + + 1 + Acalypha schinzii I + r +
ALGFMEI\E SP.ESIES
-----,------,--------,------.------,-------,----------,------~ -, -~~
Them.eda trian.dra
Hyparrhenia hirta
Trachypogon spicatus
Eulalia villosa
Diheteropgon amplectens
Cymbopogon excavatus
Tristachya hispida
Monocymbi.um ceresiiforme
Heterop11g01l contorf!l\'
Hypoxis sp.
Erawostis cun!ula
Pamcum natalense
Ctenium concirmum
Brachiaria sermta
SPESIES MET 'N LAE KONSTANTHEID
l,edehouria sp.
h'lyonurus muticus
Cnaphalium luteo-{1/bum
Koeleria '"P·. Friospemum sp.
Paspalum commersonii
nigitaria sp.
Xanthium s.p. 1ristachya rehmannii
Eriosema curdatum
Dicoma zeyheri
reorwtis dysophylla
tlllptunio. ourea
Senecio ~p.
Senecio affini~·
Erugrostis capensis
Cerbcra sp.
Sporof>olus pectinatu.~
Geologie: D (dolcri('1_l, S (sandsteen), SK (skalie).
Die ander habimtsimbole word in die tcks vcrklaar.
' '
+
+1 2 2
2 1 2
+
+' 1
+
+
+
+
2 2 3 2
I + 2 3
2 r + 1
2 2
2
+
+
2
+
+
+
+ + +
+
+
+
2 2 2 2
2 2
+ + 2 '
+ + 2
+
+~
2 ' 2 3 r 2
1 1 2
+ + 2
+
2 2
+
+
3 3 2 2
2 2 + 2
+ r 1 +
+ +
2 2
1 + 2 2
+ + + + + +
I '
2 3 2 2
1 2 3 2
+ + +
+ + + +
2 2 2
2
+
2 1 + 2
3 3 3 2
r + 2 + +
I 1
1- + +
+
2
I
+
+
+
, I ~_c __ • __ _L __ + ____ _L __ __l ____ __l _____ +_+__l_+ ______ ~+-----~

81
Die Aristida sp. Pellaea calomelanos - Xerophyta sp. - klipplaatplantegroeie
Die Aristida sp. - Pellaea calomelanos - Xerophyta sp. - klipplaatplante~
groei word verteenwoordig deur releves 17 (Kamp Ra8) en 23 (Kamp D), en
kom op die plato-uitloper met onderliggende dolerietplate voor op n
hoogte tussen 1 490 en 1 575 m bo seespieel. Klipplate bedek van 40 tot
60% van die grondoppervlakte. Afgesien van die klipplate kom gruis en
klein klippies ook voor. n Sagte tot harde grondkonsistensie is aanwesig.
Die gemeenskap kom hoofsaaklik teen suidoosfrontglooiings met n belling 0 van 5 - 25 voor.
Die klipplaatplantegroei.word gedifferensieer deur Kalanchoe thyrsiflora
Aristida sp. en.Phyllanthus parvulus. Spesies m~t n relatiewe hoe
bedekking sluit Oldenlandia herbacea, Rhynchelytrum setifolium, Pellaea
calomelanos, Eragrostis racemosa en~ Xerophyta sp. in. Die vogtige,
klipperige toestande word beklemtoon deur die teenwoordigheid van Pallaea
calomelamos, Eragrostis racemosa, Kalanchoe thyrsiflora en Rhynchelytrum
setifolium. Die opvallendste komponent van hierdie l<lipplaatplantegroei
is egter die Xerophyta sp. Themeda triandra en Trachypogon spicatus
is grassoorte wat relatief swak verteenwoordig is. Die kruinbedekking
is besonder laag en wissel van 45 tot 55% (Tabel 17).
Die Aristida sp. - Pellaea calomelanos - Xerophyta sp. - klipplaatplante
groei toon 'n verwantskap met die Aristida juncifomis - Euphorbia
pulvinata - klipplaatplantegroei deur die Xeroohyta sp. - spesiegroep.
Die Aristida junciformis - Euphorbia pulvinata - klipplaatplantegroei
Releves 8 (Kamp M7) en 3 (Kamp MS) verteenwoordig hierdie klipplaatplante
groei op die plato-uitloper gelee op n hoogte van tussen 1 464 en 1 575m
bo seespieen. Die onderliggende doleritiese gesteentes gee oorsprong
aan vlak, harde litosoliese grond wat hoofsaaklik swak gedreineer is.
Klipplate, gruis en klein klippies bedek tussen 40 en 80% van die grond
oppervlakte. Die klipplaatgedeeltes het hoofsaaklik geen belling nie
maar kom in sommige geva'lle teen glooiings wa t 'n noordoos tel ike aspek he t,
voor.

Tabel 17. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van
die Xerophyta sp. - Oldenlandia herbacea - klipplaatplantegroei.
Releve 17 23 8 3 I
I i
I
Hoogte (m) : Kruidstratum 0,5 0,7 0,7 1,5
1) Kroonbedekking (%) . . 45 55 60 55 Kruidstratum -------- ----------- ~--~--~---
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
00 N

83
Verskeie plantspesies differensieer hierdie klipplaatplantegroei, naamlik
Aristida junciformis , Euphorbia pulvinata, Lycopodium sp. en Schkuhria
pinnata. Spesies met 'n hoe kruinbedekking sluit ondermeer Rhynchelytrum
setifolium en Aloe marlothii in. Die mees opvallende komponent is Aloe
marlothii en Euphorbia pulvinata wat n skerp kontras vorm met die naas
liggende grasveldgemeenskappe. Sekere spesies, byvoorbeeld Eragrostis
plana, wat tiperend is van versteurde gebiede is ook aanwesig. Schkuhria
pinnata is n onkruid wat ook kenmerkend van versteuring is (Henderson
en Anderson 1966, Scheepers 1975).
Die Aristida junciformis - Euphorbia pulvinata - klipplaatplantegroei toon,
afgesien van die verwantskap met die Aristida sp. - Pellaea calomelanos -
Xerophyta sp. - klipplaatplantegroei, ook 'n verwantskap met die Aloe
marlothii - Crassula sp.-struikveld deur die Rhynchelytrum setifolium -
spesiegroep.
Algemeen
Klipplate is n belangrike deel van beide variante in die Xerophyta -
Oldenlandia herbacea - klipplaatplantegroei maar die Aristida sp. -
Pellaea calomelanos - Xerophyta - klipplaatplantegroei is tot die hoe
suidfrontglooiings met klipplate beperk teenoor die Aristida junciformis -
Euphorbia pulvinata - klipplaatplantegroei van die meer gelyke klipplaat-
gedeeltes van die noordfrontglooiings. Die mees opvallende verskil
tussen die twee variante is die afwesigheid van Aloe marlothii en Euphorbia
pulvinata teen die suidfrontglooiings.
Die Aloe marlothii - Crassula sp. - struikveld
Hierdie struikveldgemeenskap word deur releves 15 (Kamp Rb4), 9 (Kamp
M7), 28 (Kamp Ra3), 10 (Kamp M3), 22(Kamp M8), 6 (Kamp M6) en 5 (Kamp Ra2)
verteenwoordig, en is hoofsaaklik tot die plato-uitloper beperk wat
ondermeer die spitskruinreste en tallushange tussen 1 338 en 1 496 m
bo seespieel insluit.
Die gemeenskap word gedifferensieer deur die Aloe marlothii - spesiegroepc

84
Opvallende klipplate is in die Aloe marlothii - Crassula sp. - struikveld
afwesig. Die struikveld kan in twee variasies onderverdeel word :
die Diospyros whyteana - Aloe marlothii - struikveld, en
die Cymbopogon excavatus - Rhus eckloniana - Aloe marlothii -
struikveld.
Die Diospyros whyteana - Aloe marlothii - struikveld
Die Diospyros whyteana - Aloe marlothii - struikveld word deur releves
15 (Kamp Rb4), 9 (Kamp 7) en 28 (Kamp Ra3) op die spitskruinreste
(uitgesondered releve 9) van tussen 1 417 en 1 496 m bo seespieel, verteen
woordig. Die doleriet gee oorsprong aan litosoliese gronde met ~ konsis
tensie wat van sag tot hard wissel. Klein tot groot klippe bedek tussen
41 en 80% van die grondoppervlakte. Groo~ los klippe is ~ opvallende
kenmerk van hierdie plantegroeivariasie.
-~ helling van tot 55°.
Die noordfrontglooiings het
Die Diospyros whyteana - Aloe marlothii - struikveld word gedifferensieer
deur Diospyros whyteana, Cussonia spicata, Aristida sp., Pellaea
viridis, Helichrysum polycladum en Berkheya insignis.
Spesies met 'n hoe kruinbedekking sluit Aloe marlothii, Rhynchelytrum
setifolium, Trachypogon spicatus, Hyparrhenia hirta en Cymbopogon excavatus
in.
Die boom- en struikstratum het ~ hoogte van 1,9 tot 2,8 m (Tabel 18) en
is swak ontwikkel. Aloe marlothii is die belangrikste boom, alhoewel
Cussonia spicata een van die opvallendste komponente is. Al die borne en
struike wat aangetref word is karakteriserend van klipperige glooiings,
soos Diospyros whyteana, Cussonia spicata en Aloe marlothii asook Calpurnia
aurea.
Spesies wat kenmerkend van klipperige gedeeltes of glooiings i~ kom ook
in die grasstratum voor, waaronder Rhynchelytrum setifolium, Trachypogon

Tabel 18. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van
die Aloe marlothii - Crassula sp. - struikveld.
Releve 15 9 28 10 22
1) Hoogte (m) : Boomstratum 2,8 1 '9 2,4 2,3 3,2
Hoogte (m) : Kruidstratum 1 ' 1 1 '6 1 , 3 1 ,8 1 '3
Kroonbedekking (%) : 5 5 20 7 5 Boomstratum
Kroonbedekking (%) . . 70 60 50 60 70 Kruidstratum ---- ---
1) Insluitend Aloe marlothii
6
3
0,2
10
75
5
0,6
0,4
1
60
I
i
00 lJl

86
spicatus, Hyparrhenia hirta, Cymbopogon excavatus,Aloe marlothii en
Calpurnia aurea. Die grasstratum se kruinbedekking wissel van
SO tot 70io (Tabel 18).
Die Cymbopogon excavatus - Rhus eckloniana - Aloe marlothii - struikveld.
Hierdie struikveld word deur releves 10 (Kamp M3), 22 (Kamp M8), 6 (Kamp
M6) en 5 (Kamp Ra2) op die tallushange net onderkant die plato-uitloper
op n hoogte van 1 323 tot 1 433 m bo seespieel verteenwoordig (Fig. 19).
Die onderliggende gesteentes is hoofsaaklik doleriet, terwyl die litoso
liese grande n konsistensie van sag tot hard bet. Groot klippe en rots
blokke, kenmerkend van die tallushange, bedek tesame met klein klippies
tussen 21 en 60% van die grondoppervlakte. Die noordwesfront (releves
10 en 5) en suidoosfrontglooiings (releves 22 en 6) is steil (15 - 50°).
Differensierende spesies van die struikveld is onder andere Acalypha
caperonioides, Loudetia flavida, Setaria neglecta, Rhus eckloniana,
Stachys sp. en Asparagus africanus. n Variasie kan egter in die struik
veld onderskei word. Releves 10 en 22 word gekenmerk deur die teenwoor
digheid van Rhus discolor, Athrixia phylicoides en Rhus dentata wat in
releves 6 en 5 afwesig is. Spesies met 'n rela tiewe hoe kruinbedekking
is ondermeer Trachypogon spicatus, Eulalia villosa, Diheteropogon
amplectens, Cymbopogon excavatus, Heteropogon contortus, Hyparrhenia
hirta en Themeda triandra.
Die boom- en struikstratum van 0,6 tot 3 m hoog is swak ontwikkel en bet
'n lae kruinbedekking (Tabel 18). In die geval van releve 5 is die
struik- en boomstratum feitlik heeltemal afwesig. Die mees opvallende
komponent van hierdie stratum is Aloe marlothii (Fig. 19). Rhus dentata,
Cussonia spicata en Calpurnia aurea kom ook algemeen op rotsagtige
of klipperige glooiings voor.
Verskeie spesies wat tiperend van klipperige gedeeltes is is ook in die
grasstratum teenwoordig. Voorbeelde hiervan is Rhynchelytrum setifolium,
Cymbopogon :xcavatus en Sporobolus pectinatus. Athrixia phylicoides,
Rhus discolor (Scheepers 1975) en Tristachya rehmannii (Chippendall 1955)

Figuur 19.
87
Die ~bopogon excavatus-~hu~ eckl oniana-Aloe marlothii
struikveld (Piet Retief-studiegebied).

88
word ook met klipperige glooiings geassosieer. Die teenwoordigheid van
spesies soos Aristida sp., Rhvnchelytrum repens en Eragrostis curvula dui
op n mate van versteuring. n Kruinbedekking van 50 tot 75% kom in die
grasstratum voor (Tabel 18).
Die Cymbopogon excavatus - Rhus eckloniana - ~ marlothii - struikveld
toon, afgesien van die verwantskap met die Diospyros whyteana -
Aloe marlothii - struikveld, ook 'n verwantskap met:
die Loudetia simplex - Trachypogon spicatus - Rendlia altera -
grasveld deur die Rhynchelytrum setifolium - spesiegroep.
Algemeen
Releve 5 is aan die hand van spesiesamestelling by die Cymbopogon excavatus -
Rhus eckloniana - Aloe marlothii - struikveld geplaas, maar pas fisionomies
by die Alloteropsis semialata - Trachypogon spicatus - grasveld. Die
teenwoordigheid van onder andere ~ eckloniana, Setaria neglecta,
Rhynchelytrum setifolium en Pallaea calomelanos bevestig egter die plasing
van hierdie releve by die betrokke struikveld. Releve 9 is weens die
teenwoordigheid van Diospyros whyteana en Plectranthus sp. by die Diospvros
whyteana - Aloe marlothii - struikveld geplaas alhoewel dit fisionomies
onder die Cyrnbopogon excavatus - Rhus eckloniana - Aloe marlothii -
struikveld sou sorteer.
Die Loudetia simplex - Trachypogon spicatus - Rendlia altera - grasveld
Die Loudetia simplex - Trachypogon spicatus - Rendlia altera - grasveld
word deur releves 20 (Kamp M7) en 21 (Kamp Ml2) op die platotraphang
van tussen 1 307 en 1 402 m bo seespieel verteenwoordig. Alhoewel
~oleriet die grootste gedeelte van die gebied beslaan, word dit in die
laerliggende dele van die gemeenskap deur onderliggende skalies vervang.
Die lokaal swak gedreineerde gronde is deurgaans vlak met n klipbedekking
deur klein en groot klippe van tot 40% Klipplate is ook aanwesig.
Die platotraphang het noord- en noordoosfrontglooiings met 'n belling van
tot 5°.

89
Die gemeenskap het n homogene voorkoms en word deur Rendlia altera en
Loudetia simplex gedifferensieer. Themeda triandra, Trachypogon spicatus
en Hyparrhenia hirta is egter die oorheersende spesies met die hoogste
kruinbedekking. Ander spesies met n relatiewe hoe kruinbedekking is
Rhynchelytrum setifolium en Heteropogon contortus.
Die teenwoordigheid van Rhynchelytrum setifolium en Cymbopogon excavatus
in die gebied dui op die klipperige voorkoms van die oppervlakte terwyl
Rendlia altera algemeen op rotsriwwe (Killick 1963) aangetref word. Die
gemeenskap toon n mate van versteuring soos deur die teenwoordigheid van
Rhynchelytrum repens en n Aristida sp. - aangetoon word.
van tussen 55 en 60% kom voor.
n Kruinbedekking
Die Loudetia simplex - Trachypogon spicatus - Rendlia altera - grasveld
toon, afgesien van die reeds bespreekte verwantskappe met die Diospyros
whyteana - Aloe marlothii - struikveld en die Cymbopogon excavatus -
Rhus eckloniana - Aloe marlothii - struikveld, ook die volgende verwant
skappe:
(a) die Xerophyta sp. - Oldenlandia herbacea - klipplaatplante
groei en die Aloe marlothii - Crassula sp. - struikveld deur
die gesamentlike teenwoordigheid van die Rhynchelytrum
setifolium-spesiegroep.
(b) die Monocymbium ceresiiforme - Diheteropogon amplectens -
Tristachya hispida - grasveld deur die teenwoordigheid van
Rend1ia altera en Rhynche1ytrum setifolium.
Die Monocymbium ceresiiforme - Diheteropogon amp1ectens - Tristachya hispida - grasve1d.
Hierdie grasveldgemeenskap word deur releves 14 (kamp M6), 18 (Kamp Ra10)
en 2 (Kamp Ra3) op die plato-uitloper tussen 1 338 en 1 535 m bo seespiee1
verteenwoordig. Die gemeenskap kom hoofsaaklik op do1eritiese gesteentes
voor maar 1okaa1 word dit ook op ska1ie aangetref~ Diep grande met n
sag tot harde konsistensie word aangetref. Klippe kom slegs in n beperkte
gebied voor waar 21 tot 40% van die grondoppervlakte deur klein tot

mediumgrootte klippe bedek word.
en het n belling van tot 15°.
90
Die glooiings front suidoos of noordoos
Die gemeenskap word gedifferensieer deur Koeleria cristata en n Scilla sp.
Spesies met n hoe kruinbedekking sluit Themeda triandra, Hyparrhenia
hirta, Trachypogon spicatus, Tristachya hispida, Monocyrnbium ceresiiforme
en Diheteropogon amplectens in. Die teenwoordigheid van plantspesies
soos Aloe marlothii (Palgrave 1977), Rhynchelytrum setifolium, Rendlia
altera en Microchloa caffra dui op die voorkoms van klipperige gedeeltes.
Koeleria cristata, Microchloa caffra en Monocyrnbium ceresiiforme (Tainton,
Bransby en Booysen 1976) word algemeen op vlakker en dikwels vogtiger
gronde gevind. Enkele spesies word aangetref wat ook kenmerkend is
van laerliggende"vleigedeeltes, byvoorbee~d Senecio affinis, .Xanthium
sp. en Helichrysum nudifolium (Scheepers. 1975), terwyl die teenwoordigheid
van spesies soos Aristida junciformis en Eragrostis plana op n mate van
versteuring dui. n Kruinbedekking van 70 tot 90% kom voor (Tabel 19).
Die Monocyrnbium ceresiiforme - Diheteropogon amplectens - Tristachya
hispida - grasveld toon n verwantskap met die Loudetia simplex - Trachypogon
spicatus - Rendlia altera - grasveld wat reeds bespreek is. Deur die
Eragrostis plana - spesiegroep toon die Monocyrnbium ceresiiforme
Diheteropogon amplectens - Tristachya hispida - grasveld ook verwant-
skappe met die Alloteropsis semialata - Trachypogon spicatus - grasveld,
die Berkheya sp. - Helichrysum sp. - Tristachya hispida - grasveld en die
Gerbera sp. - Eulalia villosa - Eragrostis plana - grasveld.
Relev~ 2 sou fisionomies onder die Alloteropsis semialata - Trachypogon
spicatus - grasveld sorteer, maar is aan die hand van spesiesamestelling
by die Monocyrobium ceresiiforme - Diheteropogon amplectens - Tristachya
hispida - grasveld geplaas.
Die Alloteropsis semialata - Trachypogon spicatus - grasveld.
Die Alloteropsis semialata - Trachypogon spicatus - grasveld word deur
releves 25 (Kamp Rcl), 7, 19 (Kamp Ml) en 26 (Kamp Rcl) op die plato
uitloper met onderliggende doleritiese gesteentes op n hoogte van tussen
1 386 en 1 528 m bo seespieel verteenwoordig (Fig. 20). Die grondkonsis-
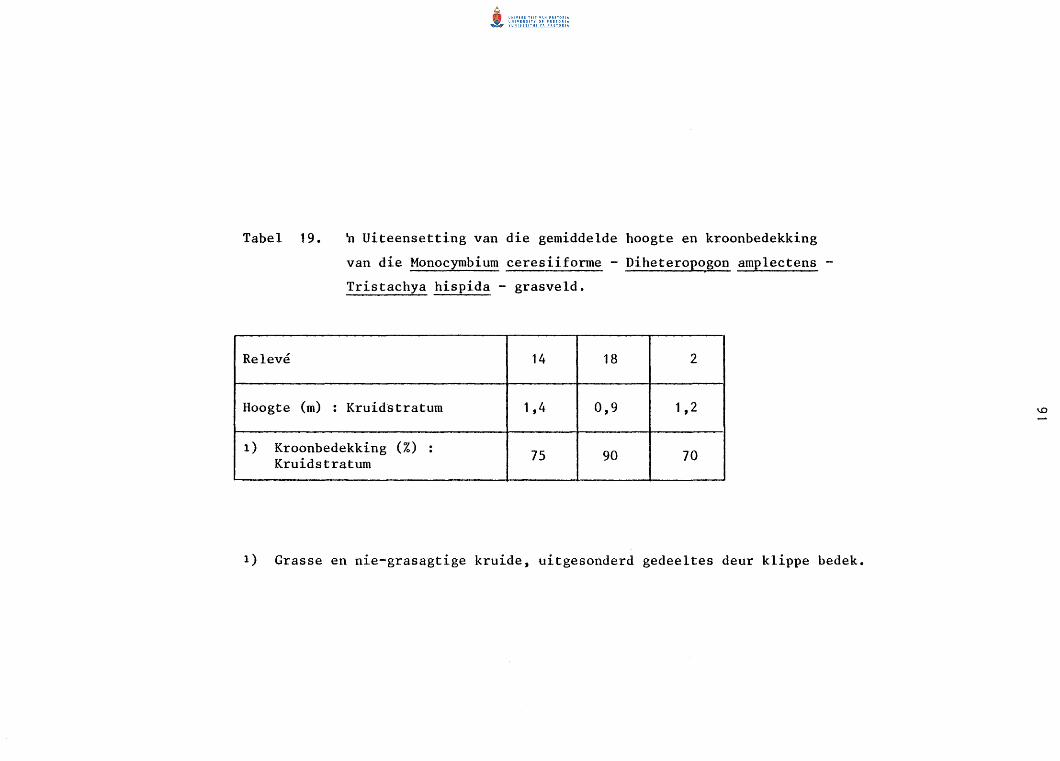
Tabel 19.
Releve
Hoogte (m)
n Uiteensetting van die gerniddelde hoogte en kroonbedekking
van die Monocyrnbiurn ceresiiforrne - Diheteropogon arnplectens -
Tristachya hispida - grasveld.
14 18 2
: Kruid·straturn 1 '4 0,9 1 '2
1) Kroonbedekking (%) . . 75 90 70 Kruidstraturn --- -~-----~-~~-- -- ----~---- --- --
1) Grasse en nie-grasagtige kruide, uitgesonderd gedeeltes deur klippe bedek.
\()

Figuur 20 .
92
Die Alloteropsis semialata- Trachypogon spicatus - gra sveld (a)
met kenmerkende plantsoorte soos Helichrysu~ nudifolium en
Trachypogon spicatus . Die Diospyros whyteana- Aloe marlothii-
struikveld (b) word op die hoer noordfrontglooiings aangetref
(Piet Retief- studiegebied) .

93
tensie wissel van sag tot hard. Klippe ontbreek feitlik heeltemal en
bedek lokaal minder as 20% van die grondoppervlakte. Suidoos- en
noordoosfrontglooiings bet n belling van minder as 25°.
Plantspesies soos Alloteropsis semialata, Haplocarhpa scaposa, Eriosema
pauciflorum en Helichrysum nudifolium is differensierend van die gemeen
skap terwyl Trachvpogon spicatus, Tristachya hispida, Monocymbium
ceresiiforme, Hyparrhenia hirta en Themeda triandra n relatiewe hoe
kruinbedekking bet. Ander opvallende spesies is ondermeer Diheteropogon
amplectens en Eragrostis curvula. Verskeie spesies is ook kenmerkend
van vogtige toestande, waaronder Haplocarhpa scaposa, Eriosema pauciflorum
(Scheepers 1975), Alloteropsis semialata (Tainton ~ al. 1976) en
verteenwoordigers van die Cyperaceae. Die voorkoms van spesies soos
Eragrostis plana en Helichrysum nudifolium (Scheepers 1975) dui op ver•
steuring en uittrapping.
(Tabel 20).
n Kruinbedekking van 70 tot 90% kom voor
Die Alloteropsis semialata - Trachypogon spicatus - grasveld toon ver
wantskappe deur die Eragrostis plana - spesiegroep met die Berkheya sp. -
Helichrysum sp. - Tristachya hispida - grasveld en die Gerbera sp. -
Eulalia villosa - Eragrostis plana - grasveld sowel as die reeds bespreekte
verwantskap met die Monocymbium ceresiiforme - Diheteropogon amplectens -
Tristachya hispida - grasveld.
Die Berkheya sp. - Helichrysum sp. - Tristachya hispida - grasveld.
Hierdie gemeenskap word deur releves 13 (Kamp MlO), 11 (Kamp M3), 12
(Kamp M4) en 29 (Kamp M6) op hoofsaaklik die pediment tussen 1 307 en
1 355m bo seespieel verteenwoordig (Fig. 21). Met die uitsondering
van lokale dolerietkolle word die gemeenskap op skalies aangetref.
n Grondkonsistensie wat van sag tot hard wissel kom voor, en met n klipbedekking van minder as 20%. Behalwe in die omgewing van releve
29 met n oostelike aspek word die gemeenskap hoofsaaklik teen noord
tot noordwesfrontglooiings met n belling van tot 15° aangetref.

Tabel 20. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van
die Alloteropsis semialata - Trachypogon spicatus - grasveld.
Releve 26 19 7 25
Hoogte (m) : Kruidstratum o.9 0,8 1 '4 0,9
1) Kroonbedekking (%) : 70 95 90 70 Kruids t·ratum
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
\0 p.

Fi guur 21 .
95
Die Berkheya s p. - He1ichrysum sp . - Tristachya hispida -
gr asve1d (a), kamp M 13 (Piet Retief-studiegebied) .

96
Die differensierende spesies van hierdie gemeenskap is onder andere
Hypoxis rigidula, Berkheya sp., Helichrysum sp. Berkheya echinacea,
Pentanisia angustifolia en Conyza bonariensis. Plantspesies met n relatiewe hoe bedekking in die gemeenskap is Tristachya hispida,
Monocvmbium ceresiiforme, Themeda triandra en Hyparrhenia hirta.
Eulalia villosa en Cymbopogon excavatus is baie opvallende plantspesies.
Tekens van vertrapping en versteuring word lokaal in hierdie gemeenskap
aangetref, soos aangedui word deur die teenwoordigheid van spesies soos
Conyza bonariensis en Verbena sp. wat ook as onkruide beskou word
(Henderson en Anderson 1966). Nidorella resedifolia (Scheepers 1975)
en Eragrostis plana is ook tiperend van sulke toestande, terwyl Setaria
nigrirostris, Paspalum comrnersonii en verteenwoordigers van die Cyperaceae
op lokale vogtige gronde dui. Die kruinbedekking wissel van 60 tot
90% (Tabel 21).
n Variasie is in die groep waarneembaar waar releves 11 en 12 en releves
13 en 29 respektiewelik bymekaar hoort. Nidorella resedifolia en
Helichrysum swynnertonii dui op n sterk verwantskap tussen releves 13
en 29, terwyl in die geval van releves 11 en 12 Acalypha sp., Verbena sp.
en Harpechloa falx op n verwantskap dui.
Die Berkheya sp. - Helichrysum sp. - Tristachya hispida - grasveld toon
verwantskappe met die Monocymbium ceresiiforme - Diheteropogon amplectens -
Tristachya hispida - grasveld, die Alloteropsis semialata - Trachypogon
spicatus - grasveld sowel as die Gerbera sp. - Eulalia villosa - Eragrostis
plana - grasveld deur die Eragrostis plana - spesiegroep.
Die Gerbera sp. - Eulalia villosa - Eragrostis plana - grasveld
Hierdie gemeenskap word deur releves 4 (Kamp Ra2), 16 (Kamp Rb3), 24 (Kamp
Ra6) en 27 (Kamp Rc4) verteenwoordig en is op die plato-uitloper gelee
tussen 1 417 en 1 527 m bo seespieel (Fig. 22)o Die onderliggende
doleritiese gesteentes gee oorsprong aan redelike vlak grond wat n
konsistensie van sag tot effens hard het. Klein klippies kom lokaal voor
en bedek minder as 20% van die grondoppervlakte. Die relatief steil

Tabel 21. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van
die Berkheya sp. - Helichrysum sp. - Tristachya hispida - grasveld.
Releve 13 11 12 29
Hoogte (m) : Kruidstratum 0,9 1, 3 1 , 2 1,0
1) Kroonbedekking (%) . . 60 90 80 60 Kruidstratum '--~------- --- -- ~------ --- ----- --
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
\0 ..........

Figuur 22.
98
Die Gerbera sp. - Eulali~ vi llosa - Eragrostis plana -
grasveld (a). h Ou wattelpl antasie (b) is sedertdien
met Eragrostis curvula beplant.

99
glooiings (15 tot 40°) front noordwes tot suid, suidoos en noordoos.
Die differensiirende spesies van die gemeenskap is~ Gerbera sp.,
Phyllanthus glaucophvllus en Vernonia natalensis. Verskeie spesies
het n hoe bedekkingswaarde, waaronder Hyparrhenia hirta, Themeda triandra,
Eragrostis plana, Trachypogon spicatus, Eulalia villosa en Tristachya
hispida. Die opvallendste plantspesies sluit onder andere Eragrostis
racemosa, Cymbopogon excavatus en Berkheya setifera in. Lokale oor
beweiding kom voor en daar is tekens van vertrapping sigbaar soos aange-
dui word deur Eragros tis plana, Aris tid a sp., Elyonurus muticus en
Gnaphalium luteo - album. Verskeie spesies wat met n klipperige omgewing
en vlakkerige grande geassosieer word is ook teenwoordig, byvoorbeeld
Eragrostis racemosa, Microchloa caffra, Diheteropogon amplectens,
Monocymbium ceresiiforme, Panicum natalense en Sporobolus pectinatus
(Chippendall 1955). Die kruinbedekking wissel van 65 tot 90% (Tabel 22).
Fisionomies sou releves 27 en 4 onder die Alloteropsis semialata -
Trachypogon spicatus - grasveld sorteer, maar is op grond van spesie
samestelling by die Gerbera sp. - Eulalia villosa - Eragrbstis plana -
grasveld geplaas. Hierdie gemeenskap se verwantskappe met die
Monocyrnbium ceresiiforme - Diheteropogon amplectens - Tristachya hispida -
grasveld, die Alloteropsis semialata - Trachypogon spicatus - grasveld
en die Berkheya sp. - Helichrysum sp. - Tristachya hispida - grasveld is
reeds· bespreek.
Die Kloofbosgemeenskap
Vier vername beskutte klowe wat noord tot noordoos front is in die studie
gebie'd aanwesig. Acocks ( 197 5) beskou die bos- en s truikp lan tegroei
wat in sulke beskutte gedeeltes gevind word as struik-woud relikte.
Hierdie relikte kan gevolglik as oorblyfsels beskou word van die daring
veld of bosveld wat oorspronklik in die Piet Retief-suurveld voorgekom
het.
Die belangrikste kloofbos word gedeeltelik in kamp M8 aangetref waar die
borne 'n dig te booms tra tum vonn.
seespieel gelee.
Die klowe is tussen 1 345 en 1 406 m bo

Tabel 22. n Uiteensetting van die gemiddelde hoogte en kroonbedekking van
die Gerbera sp. - Eulalia villosa - Eragrostis plana - grasveld.
Releve 4 16 24 27
Hoogte (m) : Kruidstratum 1 ' 1 0,9 1 '2 1 , 1
1) Kroonbedekking (%) . . 65 90 90 90 Kruidstratum
1) Grasse en nie-grasagtige kruide, uitgesonderd dele deur klippe bedek.
0 0

101
Verskeie algernene spesies is in die kloofbosgerneenskap aanwesig, waaronder
Cussonia spicata
Rhus transvaalensis
Faurea speciosa
Rhamnus prinoides
Dalbergia obovata
Die Vleigerneenskap
Ficus capensis
Rhus pyroides
Syzygiurn cordaturn
Buddleia sp.
Op die hoer=liggende gedeeltes van die plato-uitloper korn verskeie plato
vleikomrne (kyk HOOFSTUK 5 : PIET. RETIEF-STUDIEGEBIED) voor wat saarn met
die ander vleigedeeltes aangrensend aan die riviervalleie n kenrnerkende
plantegroei bet. Opvallende spesies wat verspreid in hierdie gerneenskap
voorkorn is :
Hyparrhenia aucta
Nidorella resedifolia
Koeleria cristata
Senecio affinis
Plantasies en ander Uitheemse Plante
Eragrostis plana
Erioserna pauciflorurn
Xanthium sp.
verskeie verteenwoordigers
van die Cyperaceae.
Verskeie kleiner plantasies korn verspreid deur die gebied voor, maar is
hoofsaaklik tot Rooi Kraal beperk (Fig. 6). Wattelborne (Acacia mearnsii)
is die belangrikste uitheernse boornspesie wat in die gebied aangeplant is,
en wat later vanaf hierdie gedeeltes versprei bet sodat individuele struike
of borne tans oral binne die studiegebied en veral langs die spruite aan
getref word.
Lokaal is wattelborne rneganies of per hand uitgedun waarna Eragrostis
curvula gesaai is om genoeg blaremateriaal te voorsien sodat die gedeeltes
geraeld gebrand kan word (Fig. 22). Net soos in die geval van die
Amsterdarn-studiegebied hou die uitbreiding van wattelborne n ernstige gevaar
in vir die natuurlike plantegroei veral langs die waterlope. Enkele
bloekornborne (Eucalyptus sp.) korn ook voor.

102
Bespreking en Gevolgtrekking: Amsterdam- en
Piet Retief-studiegebiede
MONSTERNEMING
n Standaard prosedure van die Zurich-Montpellierskool vir plantsosiologie
is die subjektiewe uitsoek van monsterpersele (Werger 1973). Volgens
Werger(l974b) kan hierdie metode wel moontlik met sukses in nie-intensiewe
opnames oor groot gebiede toegepas word. Gedurende hierdie studie is n
poging in beide studiegebiede aangewend om gestratifiseerde ewekansige
monsterperseelverspreiding te gebruik (Scheepers 1969, Coetzee 1972,
Theron 1973 en Bredenkamp 1975).
Weens die meer bergagtige aard van die Piet Retief-studiegebied word n
meer heterogene plantegroei in hierdie gebied aangetref in vergelyking
met die Arnsterdam-studiegebied. Omdat verskillende habitattipes tot
verskille in plantegroei kan lei (Dahl 1956, ~: Coetzee 1972), is
fisiografiese eenhede onderskei na gelang van geologiese formasie,
hoogte bo seespieil, aspek en klipperigheid. Deur die koppeling van
sommige van hierdie fisiografiese eenhede aan die fisionomie van die
plantegroei, is 13 en 9 fisionomiese eenhede in die Amsterdam- en Piet
Retief~studiegebiede onderskeidelik onderskei. Plantasies, kloofbos
en sekere vleigebiede is nie gemonster nie maar word slegs kortliks be-
skryf. Die plantegroeigemeenskappe wat gedurende die studie geLdentifi-
seer is, stem oor die algemeen ooreen met die voorafbepaalde fisiografies
fisionomiese eenhede, veral in die geval van die Arnsterdam-studiegebied.
Met die opstel van die plantsosiologiese tabelle is daar soms meer as een
gemeenskap in sommige van die fisiografies-fisionomiese eenhede onderskei.
Aan die hand van sekere omgewingsfaktore soos klipperigheid asook die
aanwesigheid van spesiegroepe kon gemeenskappe van mekaar onderskei word.
n Voorbeeld hiervan, in die Amsterdam-studiegebied, is die Eulalia villosa -
Hypoxis rigidula - Berkheya setifera - grasveld en die Eulalia villosa -
Pentanisia angustifolia -Setaria splendida- grasveld waar klipperigheid
op n verskil gedui het.

103
In sekere gevalle is omgewingsfaktore gebruik om indivuele releve-plasings
in sekere gemeenskappe te versterk waar n blote plasing op grond van
spesiegroepe aileen nie voldoende was nieQ In die Piet Retief-studie
gebied is n voorbeeld hiervan releve 2 by die Monocymbium ceresiiforme -
Diheteropogon amplectens - Tristachya hispida - grasveld waar doleriet
plate, behalwe met n enkele uitsondering, in al die releves gevind is.
Uit die resultate blyk dit dat daar wel n verband bestaan tussen die
plantegroei en die fisiografies-fisionomiese eenhede soos vir die
stratifisering van die monsterpersele aangewend is. Soos deur ander
werkers elders gevind is (Coetzee 1972, Bredenkamp 1975), kan gestrati
feseerde ewekansig-verspreide monsterpersele met sukses in sommige
streke gebruik word.
DATAVERWERKING
Die hoofdoel van die plantkundige studie was om die plantegroei van die
Amsterdam-en Piet Retief-studiegebiede in gemeenskappe te klassifiseer,
ekologies te interpreteer en te beskryf ten einde oorbietjievoorkeure
te bepaal. Die rou gegewens wat vir die Braun-Blanquet-tabuleertegnieke
gebruik is (Braun-Blanquet 1932, Kllckler 1967), het berus op spesies
wat in meer as een releve teenwoordig isQ
Aangesien te min bekend is oor die algemene plantegroei in die omgewing
van die reservaat, is die ge1dentifiseerde gerneenskappe nie in n hierar
giese stelsel geklassifiseer nie. Die differensierende plantspesies
wat in die studie gebruik is moet slegs ook voorlopig as sodanige spesies
beskou word. Geen plantspesies kan op die stadium as karakterspesies
aangewys word nie.
PLANTEGROEI
Verskille en variasies in die plantegroei word veral aan fisiografiese
verskille soos geologiese forrnasie, aspek, klipperigheid van die grond
oppervlakte en in n mindere mate hoogte bo seespieel, grondkonsistensie,
beskutte valleie en selfs oulande toegeskryf. Hierdie faktore is onder
ling of gesarnentlik verantwoordelik vir die plantegroeiverskille.

104
Die groat ooreenkoms tussen die geomorfologiese eenhede en verspreiding
van plantegroeigemeenskappe word in beide studiegebiede aangetoon.
Enkele voorbeelde hiervan is die Eulalia villosa - Hypoxis rigidula -
Berkheya setifera - grasveld op die klipveld van die plato-uitloper
in die Amsterdam-studiegebied, en die Aristida junciformis - Euphorbia
pulvinata - klipplaatplantegroei op die dolerietdagsome van die plato
uitloper in die Piet Retief•studiegebied.
Die twee studiegebiede se plantegroei toon n redelike sterk ooreenkoms,
maar is meer heterogeen in die Piet Retief-studiegebied wat meestal tot
n beter omgrensing van gemeenskappe leio In die Piet Retief-studie
gebied is die plantegroei-variasie oak meer prominent, van die bloot
gestelde Aristida spo - Pellaea calomelanos = Xerophyta spo = en die
Aristida junciformis - Euphorbia pulvinata - klipplaatplantegroei op
die hoer plato 0 s tot die struik-woud relikte in die beskutte kloweo
Die grootste ooreenkoms tussen die twee studiegebiede is in die Berkhyea
setifera - Vernonia natalensis - Eulalia villosa - grasveld en die
Tristachya hispida - Berkheya setifera - Panicum natalense - grasveld
op die plato-uitloper en gedeeltelik op die westelike pediment (Amsterdam
studiegebied) met die Alloteropsis semialata - Trachypogon spicatus -
grasveld en die Gerbera sp. - Eulalia villosa - Eragrostis plana -
grasveld op die Piet Retief-studiegebied se plato-uitloper. Die volgende
spesies is onder andere deurgaans in hierdie gemeenskappe, teenwoordig:
Trachypogon spicatus, Hyparrhenia hirta, Themeda triandra, Eulalia
villosa en Monocymbium ceresiiforme.
n Verdere ooreenkoms word tussen die vleigemeenskappe van die twee studie
gebiede aangetoon, veral deur die teenwoordigheid van spesies soos
Hyparrhenia aucta, Eragrostis plana en verskeie verteenwoordigers van
die Cyperaceae. Die opvallendste verskil in die plantegroei wat teen
glooiings in die studiegebiede aangetref word is die teenwoordigheid van
verskeie struike en borne in die Piet Retief-studiegebied. Voorbeelde
van boomspesies in die genoemde studiegebied is Diospyros whyteana,
Cussonia spicata, ~ marlothii en Calpurnia aurea.
n Opvallende eienskap van die Amsterdam-studiegebied is die verspreide
versteuring wat in sekere gedeeltes aangetref word soos ge~llustreer

105
deur die teenwoordigheid van Helichrysum spp., Rhynchelytrum repens,
Eragrostis plana en Conyza bonariensis. In die Piet Retief-studie
gebied is die vertrapping veel meer gelokaliseerd, soos byvoorbeeld
in die Berkheya sp. - Helichrysum sp. - Tristachya hispida - grasveld.
Acocks (1975) vermoed dat die Piet Retiefsuurveld oorspronklik n oop
daring- of bosveld met struikwoud in beskutte gedeeltes was. Weens
hierdie rede het hy die veldtipe onder die valsgrasveldtipes geklassi
fiseer alhoewel dit vandag prakties n grasveld is afgesien van die
kloofbos in beskutte gedeeltes. Acocks (££. cit.) maak ook melding
van die relatiewe belangrikheid van veral Tristachya hispida, Themeda
triandra en Rendlia altera. Die plantsosiologiese tabelle van beide
studiegebiede dui egter op n hog voorkoms van veral Hyparrhenia
hirta, Trachypogon spicatus asook Themeda triandra.

106
HOOFSTUK 7
VOEDING EN WATERBEHOEFTES
Inleiding
Die meeste hoefdiere toon ~ mate van selektiwiteit tydens voeding sodat
die chemiese en botaniese samestelling van die dieet verskil van die van
die beskikbare plantegroei in die algemeen (Novelli 1975). Leuthold (1977)
wys daarop dat die waarskynlike harmonie waarin Afrika-hoefdiere leef
moontlik veral te danke is aan ekologiese skeiding waar verskille in voed
selvoorkeure ~ belangrike funksi~ vervul. Ewer (1973) bet egter reeds na
die gebrek aan inligting hieroor verwys. ~ Studie op voeding en water
behoeftes is nie alleen belangrik om die ekologiese nis van ~ dier te
bestudeer nie, maar bet ook belangrike bestuurstoepassings.
Metodes
VOEDSELVOORKEURE
Direkte waarnemings is gebruik om die voedselkeure van oorbietjies in die
Amsterdam-studiegebied te bepaal. Twee familiegroepe oorbietjies bet
~ voertuig na ~ p~riode van ses jaar tot op ~ afstand van 20 m toegelaat
sender dat die gedrag merkbaar beinvloed is. Alle waarnemings is gevolg
lik vanaf ~ ~oertuig gedoen waar bekende plante met behulp van ~ 8 x 40
Leitz Trinovid verkyker ge1dentifiseer is. Alle onbekende plante is
later versamel en geidentifiseer.
voedselplante beskou.
Plantspesies wat gevreet is, is as
Daar is gepoog om ~ seisoenale indeling van voedselvoorkeure te maak.
Waarnemings is op ~ driemaandelikse basis gedoen waartydens die diere so
ver moontlik vir ononderbroke periodes gedurende bulle daaglikse wei-akti
witeite gevolg is. ~ Totaal van 16 waarnemingsure is gevolglik gebruik
om waarnemings gedurende die somer (Desember- Februarie), herfs (Maart tot
Mei), winter (Junie- Augustus) en lente (September- November) te doen.
~ Bree voorkeur indeling is gebruik:

Voorkeur a
Voorkeur b
Voorkeur c
107
meer as 15% benutting
11 - 15% benutting
minder as 11% benutting
Een van die grootste tekortkominge van bierdie metode was die versteuring
wat die voortdurende identifiserings in die veld tot gevolg gehad het, met
die gevolg dat die aantal waarnemings noodwendig beperk moes word. Die
periodieke teenwoordigheid van vee en ander boksoorte het indirekte waar
nemings op benutte plante onmoontlik gemaak.
Verskeie werkers bet direkte waarnemings gebruik om voedselvoorkeure te
bepaal, waaronder Lamprey (1963), van Zyl (1965) en Tinley (1969). Field
(1968) wys egter daarop dat dit moeilik is om grassoorte wat gevreet word
te identifiseer tensy die diere van naby bestudeer word. Verdere nadele
is die interpretasie van kwantitatiewe inname en ook die feit dat die
metode tydrowend is (Hofmeyr 1970). n Poging om direkte waarnemings op
mak oorbietjies te doen, gebaseer op Field (££. cit). se metode, het misluk
nadat drie studiediere dood is.
Aangesien die twee familiegroepe oorbietjies wat bestudeer is tot kamp 11
(Fig. 5) beperk was, is al die waarnemings in die Eulalia villosa - Hypoxis
rigidula - Berkheya setifera - grasveld en die Eulalia villosa -
Pentanisia angustifolia - Setaria splendida - grasveld gedoen.
VOEDINGSGEDRAG
Oorbietjies wat besig was om te wei is met n 8 x 40 verkyker dopgehou vir
periodes wat van 1 tot 5 minute gewissel bet. n Bandopnemer is gebruik
om alle aksies soos aantal tree gegee, lyfkosing, ens. op te neem. n Wisselende aantal waarnemings is van April tot Julie 1975 in die Eulalia
villosa - Hypoxis rigidula - Berkbeya setifera - grasveld van die Amster
dam-studiegebied gedoen.
Die volgende aspekte, gebaseer op Novelli (1975) se studie, is kwantitatief
ondersoek:

108
a die gemiddelde tydsin tervalle in sekondes wat aan
voeding gespandeer word,
b die aan tal byte per sekonde,
c die aantal byte per voedingstasie, en
d die aantal tree per sekonde geloop.
n Voedingsstasie is gedefinieer as die oppervlakte bedek deur n hipote
tiese halfsirkel voor die oorbietjie waartydens dit voedingsplante kan
bereik sender om die voorpote te beweeg (Goddard 1968). Die tyd gespan
deer aan lyfkosingsaktiwiteite, interaksies en urinering en ontlasting is
afgetrek.
WATERBEHOEFTES
Om die waterinhoud van vier voorkeurgrasse, naamlik Eulalia villosa,
Hyparrhenia hirta, Monocymbium ceresiiforme en Themeda triandra, as 'n
persentasie van droe gewig te bepaal, is die volgende metode gevolg:
Ongeveer 0,5 kg grasblare is van die genoemde grasspesies gedurende
Julie 1976 al om die vier ure in die Eulalia villosa - Hypoxis rigidula
Berkheya setifera - grasveld (Amsterdam-studiegebied) versamel. Die
monsters, wat verkry is terwyl die veld in n kort toestand was, is in
afsonderlike plastieksakke geplaas en onmiddelik lugdig verseel. Hier
die monsterneming is gedurende n periode van vier nagte herhaal waarna
die gemiddeldes bepaal is. Alle neerslag in die vorm van ryp en dou op
die grasblare is saam versamel.
Elke verseelde monster is, nada t di t geweeg is, in 'n droogoond gedroog
teen 105°C vir 48 ure. Die monsters is hierna herhaaldelik afgekoel en
weer geweeg totdat n konstante massa verkry is.
Resultate en Bespreking
VOEDSELVOORKEURE
Volgens die direkte waarnemings gedurende al vier seisoene toon oorbietjies
die volgende voedselvoorkeure in die Eualia villosa - Hypoxis rigidula -

109
Berkheya setifera - grasveld en die Eulalia villosa- Pentanisia
angustifolia - Setaria splendida - grasveld van die Amsterdarn-studiegebied
(kyk HOOFSTUK 6 : DIE AMSTERDAM-STUDIEGEBIED).
Alloteropsis sernialata
Andropogon sp.
Acalypha sp.
Brachiaria serrata
Corrunelina sp.
Diheteropogon arnplectens
Digitaria sp.
Elvonurus rnuticus
Eulalia villosa
Eragrostis capensis
§.. curvula
~· racernosa
Erioserna pauciflorurn
Heteropogon contortus
Hyparrhenia hirta
Hypoxis rigidula
Monocymbiurn ceresiiforrne
Pentanisia angustifolia
Rendlia altera
Rhynchelytrurn setifoliurn
Panicum sp.
Therneda triandra
Thunbergia sp.
Trachypogon spicatus
Tristachya hispida
Vernonia sp.
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
Voorkeur:
c
c
c
c
c
c
c
c
a
c
c
c
c
c
b
c
a
c
c
c
c
a
c
c
c
c
Volgens die resultate blyk dit dat oorbietjies deurgaans n hoe voorkeur vir
Eulalia villosa, Themeda triandra en Monocymbiurn ceresiiforrne toon, gevolg
deur Hvoarrhenia hirta.

110
Nie-grasagtige kruide vorm n opvallende komponent van die dieet (5,8 -
12,0%). Onge~dentifiseerde grasse en minder belangrike grasse (waar
onder Rendlia altera, Panicum sp. en Rhynchelytrum setifolium) vorm
4,6 - 19,0% van die dieet.
Hoefdiere word algemeen in grasvreters, blaarvreters en gemengde vreters
verdeel waar grasvreters feitlik uitsluitlik verteenwoordigers van die
Poaceae vreet en blaarvreters hoofsaaklik plante behorende tot die
Dicotyledoneae benut. Hofmann en Stewart (1972) beskou die oorbietjie as
'n grootmaat en ru-voedsel vreter ("bulk and roughage eater") wat verder
geklassifiseer word onder die subgroep van varsgrasvreters wat ook van
drinkwater afhanklik is. Volgens Ansell (1960) is die oorbietjie uit
sluitlik n grasvreter, maar Tinley (1969) vermoed dat verdere studies dit
moontlik as n gemengde vreter kan aantoon. Jarman (1974) beskryf die
oorbietjie as n selektiewe vreter wat feitlik uitsluitlik grasvretend is.
Anders as die tipiese grasvreters met bree ongedifferensieerde bekke soos
die wildebees en sebra, het die oorbietjie n skerp, srnal kop en bek wat
ook beweegbare lippe het. Hierdie kenmerke is karakteristiek van blaar
vreters soos die duiker wat hierdie diere in staat stel om selektief te
wees en voedseldele in moeilik bereikbare plekke te bekom. Na aanleiding
van die voedselvoorkeurstudie se resultate kan die oorbietjie as n gemengde
vreter geklassifiseer word alhoewel dit hoofsaaklik verteenwoordigers van
die Poaceae in die Amsterdam-studiegebied benut.
Gedurende die studie is daar waargeneem dat oorbietjies nie net verskil
lende grasspesies selekteer nie, maar ook sekere plantdele. Individuele
grasblare van byvoorbeeld Hyparrhenia hirta en Diheteropogon amplectens
word dikwels afgevreet. Wanneer groat graspolle van byvoorbeeld Eulalia
villosa benut word, buig oorbietjies hulle nekke om sekere plantdele in
moeilik bereikbare plekke te bekom. Hierdie selektiewe vreet is veral
opvallend in langgras en gebrande veld. Monfort en Monfort (1979) ver
klaar dat oorbietjies ook in Rwanda sekere plantdele selektief vreet,
byvoorbeeld jong gedeeltes van Themeda triandra en Hyparrhenia filipendula.
Volgens Leuthold (1977) vreet baie hoefdiere egter selektief wat bepaalde
plantdele betref.

111
n Studie op die spesievoorkeure van beeste sowat 10 krn wes van die Arnster
darn-studiegebied in soortgelyke veld deur Roos, Rethrnan en Kotze (1973)
het getoon dat beeste n voorkeur vir Diheteropogon amplectens, Eulalia
villosa, Themeda triandra, Trachypogon spicatus, Tristachya hispida en
Monocymbruim ceresiiforrne toon.
Volgens 'fabel 23 is daar n geringe seisoenale wisseling in die spesie
voorkeure. n Opvallende toenarne in die voorkeur van nie-grasagtige
kruide kern gedurende die lente voor. n Toename in die voorkeur vir
Eulalia villosa is gedurende die lente en sorner opgemerk vir Diheterooogon
arnplectens gedurende die herfs. Monfort en Monfort (1974) het verrnoed dat
oorbietjies in die Nasionale Akagera-wildtuin in Rwanda nie n baie duidelike
seisoenale wisseling in bulle voedselvoorkeure vir spesifieke plante toon
nie. Voorkeurplante volgens Monfort en Monfort (££• cit.) is onder andere
Themeda triandra, Hyparrhenia filipendula, Loudetia simplex en Cymbopogon
spQ In die Nasionale Gorongosa-wildtuin het oorbietjies ook net n geringe
seisoenale wisseling in spesievoorkeure. n Verskeidenheid plante van
grasse tot houtagtige plante soos Ziziphus mucronata word benut (Tinley 1977).
Roos et ~· (1975) se studie in soortgelyke veld het ook n seisoenale
wisseling in spesievoorkeur aangetoon waar Diheteropogon arnplectens veral
gedurende die vroee somer gevreet word, terwyl Themeda triandra en Trachvpogon
spicatus weer teen die einde van die sorner verkies word. Eulalia villosa
en Hyparrhenia hirta word veral gedurende die winter benut. Roos ~ ~·
(££. cit.) verklaar dat die voorkeure in die gebied nie deur die veldsame
stelling beinvloed word nie.
Hierdie plantvoorkeure en seisoenaie veranderinge in benutte plante van
bees te stern in 'n groo t rna te ooreen met die van die oorbie tj ie in die
Amsterdarn-st~diegebied. Of dit werklik op kornpetisie dui is onseker.
Anders as wat HofmannenStewart (1972) beweer blyk die oorbietjie nie 'n
grootrnaat ruvoedselvreter te wees nie, maar beeste en skape wel. Daar
bestaan moontlik n ekologiese skeiding tussen die betrokke spesies wat
die uitsoek van sekere plantdele betref. Die weibewegings van vee kan
egter 'n belangrike versteuringsfaktor wees (kyk HOOFSTHK 10 : ALGEMENE
INTERAKSIES).

Tabel 23. Geskatte persentasie benutting deur oorbietjies van die vernaamste plantspesies in die Eulalia villosa
Hypoxis ridigula - Berkheya setifera - grasveld en die Eulalia villosa - Pentanisia angustifolia -
Setaria splendida - grasveld, Amsterdam-studiegebied.
SPES IE SOMER HERFS WINTER LENTE
Frekwensie % Frekwensie % Frekwensie % Frekwensie %
Eulalia villosa 39 29,77 21 16,03 32 34,04 42 31 "11
Themeda triandra 21 15,96 20 51 ,27 18 19' 15 14 10,37
Monocyrnbium ceresiiforme 19 14,44 18 13,74 16 17,02 23 17,03
Trachypogon spicatus 3 2,28 6 4,58 1 1,06 2 1,48
Hyparrhenia hirta 12 9' 12 13 9,92 12 12.77 15 11 ' 11
Tristadya hispida 3 2,28 8 6' 11 1 1,06 4 2,96
Brachiaria serrata 3 2,28 4 3,05 2 2' 13 6 4,44
Diheteropogon amplectens 6 4,56 12 9 '16 3 3" 19 6 4,44
Ongeidentifiseerde grasse en ander grasse 16 12,21 22 16,79 4 4,26 12 8,89
Nie-grasagtige kruide 9 5,34 8 6' 11 5 5,32 15 11 " 11
Totaal 131 100% 131 100% 9l~ 100% 135 100%
I
~--------- -- ------ - - ----
N

113
VOEDINGSGEDRAG
Jarman (1974) klassifiseer al die Bovidae na aanleiding van voedingstyl
("feeding style"). Hierdie klassifikasie berus eerder op die manier
waarop voedse1 bekom word as watter p1antsoorte gevreet word. Volgens
Jarman (2£• cit.) se k1assifikasie sorteer die oorbietjie onder die soge
naamde groep b wat selektiewe vreters en hoofsaak1ik grasvretend is. Daar
by bly hierdie groep in een of meer plantegroeitipes en een tuisgebied reg
deur die jaar.
Se1ektiewe voeders soos blaarvreters beweeg baie rond om geskikte voedsel
te bekom (Jarman ££• £!!.) maar die voedingstyl is verwant aan verskeie
ander aspekte van n dier se biologie, veral wat sosiale organisasie be
tref. Jarman (££. £!!.) se hipotese wys daarop dat n weidende bok se
patroon van beweging onder andere die verspreiding van voedselitems
reflekteer.
Oorbietjies wei gemiddeld 83,6% van huile aktiewe periodes wat gemiddeld
28,6% van n 12-uur daglig periode beloop (kyk HOOFSTUK 8 : WEI-AKTIWITEITE)
en wat n totaal van 3,44h per 12 ure is. Gedurende die weiperiodes wei
die familiegroep saam. Gewoonlik wei n individuele oorbietjie voortdurend
om slegs af en toe rond te kyk, terwyl verskeie tree soms gegee word voordat
daar gevreet word. Verskeie byte word dan gegee terwyl die oorbietjie
op dieselfde plek bly staan (Fig. 23 en 24). Wanneer daar verder beweeg
word, word die nek dikwe1s in n horisonta1e posisie gehou behalwe wanneer
die wei-periode byvoorbeeld kortliks deur lyfkosingsaktiwiteite onderbreek
word. Novelli (1975) het gevind dat springbokke en blesbokke ook dikwels
vir lang periodes met hu11e koppe omlaag wei. Volgens Jarman (1974)
is die tyd wat gespandeer word op n spesifieke plek tydens voeding en die
mate van beweging na die vo1gende vreetplek belangrik om die mate van
se1ektiwiteit te bepaal.
Ten einde n beter aanduiding van die oorbietjie se voedingsnis te verkry
is n studie op die tempo- en bewegingspatrone, soortgelyk aan Novelli
(££. cit.) se studie op die springbok en blesbok, gedoen. Oorbietjies

Figuur 23. 'n Oorbietj ieooi wa t in 'n kortgebrande en beweide gedeel te
van die Eulalia villosa - Hypoxis rigidula - Berkheya setifera -
grasveld wei (Amsterdam-studiegebied).
-!:'-
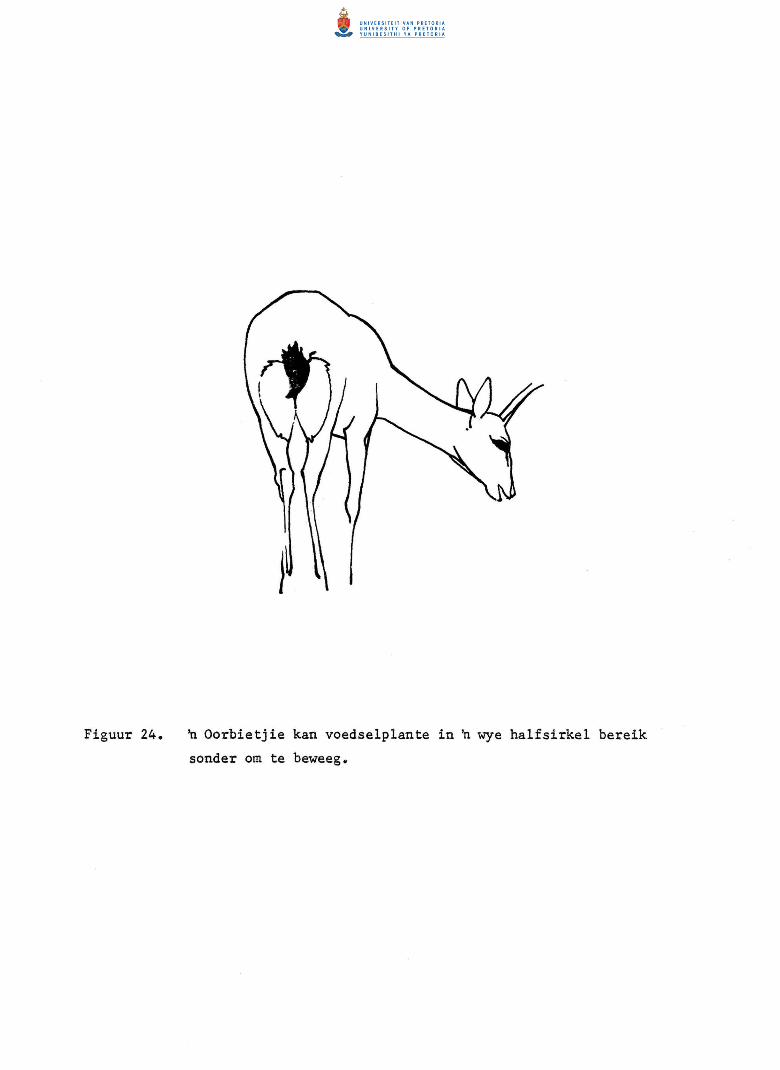
Figuur 24.
115
n Oorbietjie kan voedselplante inn wye halfsirkel bereik
sander om te beweeg.

116
gee gerniddeld 11,57 byte (n = 143; S = 14,82; reeks 1 - 109) per voeding
stasie. Die bokke is egter in staat om verskeie plante en los plantdele
te bereik sender om te beweeg (Fig.24) sodat selfs vyf of rneer verskillende
plante per voedingstasie benut kan word. Die tempo waarteen gevreet word
is gerniddeld een byt elke 1,37 sekondes, terwyl die voedingstasies gerniddeld
10,48 sekondes duur (S = 10,43; reeks 1 - 61). Die totale aantal tree
wat tydens n weiperiode gegee word beloop gerniddeld 1 tree per 3,14 sekondes
(S = 4,34) wat ook n relatiewe aanduiding gee van die bewegingsternpo tydens
n weiperiode, terwyl die aantal tree van een voedingstasie tot die ander
die verspreiding van voelseliterns reflekteer, waar rneer tree gegee.word
indien voorkeur plantspesies wyd verspreid is.
Volgens Jarman (1974) se hipotese sal n selektiewe boksoort se voedingsta
sie relatief van korte duur wees terwyl dit ook verder sal loop om voorkeur
plantspesies te bekorn. By vergelyking tussen die oorbietjie, blesbok en
springbok (Tabel 24) na aanleiding van Novelli (1975) se studie op die
laasgenoernde twee spesies, duur die voedingstasies langer in die oorbietjie
se geval terwyl dit ook rninder tree per rninuut ge~ (uitgesondered die bles
bok). Alhoewel hierdie vergelyking tussen die drie boksoorte daarop dui
dat die oorbietjie rnoonlik rninder selektief is, is dit nie noodwendig die
geval nie. Die oorbietjie kan in n wye halfsirkel voor horn vreet (Fig.
24) en wanneer n boksoort baie selektief is wat plantdele betref kan dit
lank per voedingstasie vertoef om sekere voorkeurplantdele van een of rneer
plantspesies te selekteer. Jarman (££• £!!.) se klassifikasie kan dus as
korrek aanvaar word.
WATERBEHOEFTES
Baie hoefdiere drink gereeld water terwyl die rneerderheid van n oop water
oppervlakte vir bulle waterbehoeftes afhanklik is. Enkele Afrika-boksoorte
kan egter rnoonlik oorleef sonder om enigsins water te drink, waaronder die
girafgasel Litocranius walleri (Leuthold 1977). Hierdie boksoorte kan ge
noeg water vanuit bulle voedsel verkry en/of bet spesiale water besparende
rneganismes. Daarteenoor bet somrnige ander boksoorte weer die vermoe om
vir verskeie dae sander water te bly, byvoorbeeld die rcoibok Aepyceros
rnelarnpus (Young 1970). Die rietbok kan maande in waterlose gebiede bly

Tabel 24e Verskille in sekere voedingsaspekte tussen die oorbietjie,
blesbok en springbok.
I
Oorbietjie l) Blesbok 2 ) Springbok 2 )! I
'
Duur van voedingstasie (sek.) 10,48 7,65 5,56
Gerniddelde tree per min. 13,51 11,25 23,71
1) Arnsterdarn-studiegebied
2) Jack Scott-natuurreservaat (Novelli 1975)
......

118
(Haltenorth 1963, In: Jungius 1971), maar dit is nie seker of dit wel
onafhanklik van water is nie. Hierdie beperking in fisiologiese water-
toleransie het ~ groat invloed op ~ spesie se keuse van habitat, bewegings
en daaglikse aktiwiteite.
In hulle klassifikasie van Afrika-hoefdiere na aanleiding van voedselge
woontes meld Hofmann en Stewart (1972) dat oorbietjies van cop wateropper
vlaktes afhanklik is. Stevenson-Hamilton (1929) verklaar ook dat oor
bietjies altyd naby water gevind word, alhoewel Shortridge (1934) twyfel
of oorbietjies ooit water drink. Volgens Roosevelt en Heller (1914) word
oorbietjies in gebiede gevind waar geen water voorkom nie. Ook in Rwanda
het Monfort en Monfort (1974) opgemerk dat oorbietjies k1aarblyklik nooit
water drink nie en het se1fs nie gedurende die droe maande migreer wanneer
daar geen water in hulle gebied was nie.
A1 die bekende oorbietjieterritoriums in die Amsterdam-studiegebied
sowel as die Piet Retief-studiegebied (kyk HOOFSTUK 9 : TERRITORIAL£
GEDRAG) het geen permanente water nie aangesien standhoudende water tot
die grater spruite of riviere in beide studiegebiede beperk is. Gedurende
die vogtige periode tydens die somermaande (Fig. 9 en 10) is water in beide
studiegebiede volop en staan dikwels dae lank in laer gedeeltes of konvekse
klippe (veral die klipperige- of klipp1aatgedeeltes). Enke1e v1eigedeel
tes kom wel voor maar het slegs water gedurende die somermaande, byvoor
beeld die Eragrostis patentissima - Echinochloa sp.- vleigemeenskap in die
Amsterdam-studiegebied (kyk HOOFSTUK 6 : DIE AMSTERDAM-STUDIEGEBIED).
Gedurende die studie is geen oorbietjie opgemerk wat water gedrink het
nie, selfs nie wanneer water vrylik in die veld gedurende die vogtige
periode beskikbaar was nie. Oorbietjies toon ~ gehegtheid aan hulle
territoriums (kyk HOOFSTUK 9 : TERRITORIAL£ GEDRAG) wat in die meeste
geva11e permanent blyk te wees, en aangesien die bekende oorbietjie-terri
toriums geen· permanente vrye water insluit nie, is hulle op ander bronne
vir waterbehoeftes aangewese.
Die enigste alternatiewe bron van water is die voedselplante en wel deur
die voginhoudbinne die plant en/of deur neeslag soos ryp en dou op die

119
plante Gevolglik is die voginhoude van die vier belangrikste voorkeur
grasse gedurende die kritiese periode wat water betref, naamlik die
droer maande wanneer die maandelikse gemiddelde neerslag tot onder 13 mm
(Julie) in beide studiegebiede daal (Tabel 3; Fig. 9 en 10), al om die
vier ure bepaal. Die volgende voorkeurgrasse is gebruik: Eulalia villosa,
Hyparrhenia hirta, Monocymbium ceresiiforme en Themeda triandra.
Die resultate (Fig. 25) toon dat die persentasie voginhoud van al vier
grasspesies gedurende n 24-uur periode wissel. Die plantmonsters het
die hoogste voginhoud om 06h00 getoon waarna dit vinnig gedaal het om die
laags te inhoud om 14h00 tot 18h00 te bereik. Hie rna he t daar 'n s tyging
in die voginhoude voorgekom sodat die hoogste voginhoude om 06h00 bepaal
is. Eulalia villosa het deurgaans n hoer voginhoud as die ander gras
spesies getoon. Weinmann (1955) meld dat die voginhoud van jong gras
so hoog soos 70 - 80% is teenoor die 0 - 10% van dooie gras.
Die voginhoude wat verkry is sluit neerslag in die vorm van dou en ryp in.
Ryp kern gereeld gedurende die wintermaande voor, naamlik gemiddeld 43 dae
per jaar by Piet Retief (kyk HOOFSTUK 9 : Klimaat).
Schmidt-Nielsen ( 1964) het voorgestel dat "higroskopiese plante" 'n belang
rike waterbron vir naglewende woestyndiere kan wees. Taylor (1968) het
aangetoon dat gemsbokke ~ gazella en Grantgasel Gazella granti uitsluit
lik van plan tegroei vir water afhanklik is. Volgens Taylor (££• cit.)
he t die plan te se voginhoude 'n dras tiese wisseling gedurende 'n 24-uur
periode getoon met die hoogste inhoud tussen 19h00 en 0700, en hy vermoed
dat die aktiwiteitsperiodes van die betrokke twee boksoorte aangepas word
om die maksimum vogbenutting te verkry.
Alhoewel oorbietjies voortdurend aktiewe met onaktiewe periodes afwissel
(kyk HOOFSTUK 8 : BASIESE AKTIWITEITSPATROON) toon oorbietjies wel n duidelike aktiewe periode vroegoggend wat meer opvallend is gedurende die
wintermaande, en veral tussen 06h00 en 07hOO. Hierdie aktiwiteitsperiode
stem ooreen met die periode waartydens die vier genoemde grasspesies die
hoogste voginhoude het. Ryp en dou kern ook hoofsaaklik gedurende die
laaste gedeelte van die nag voor wanneer die maksimum radiasie plaasvind.

70
60
"C = 0 50 .1: c -C)
~ •o C»
·c;; 30 IU .... c C» ~ 20 l
10
0
•
"'-- •- Eulalia villosa
Hyparrhenia hirta ~ , ' , '
Monocymbium ceresiiforme , ' , ' , ' , ' , ' , ' Themeda triandra
... - ,,' -- ... -... ,,' ._ ..... ., ,
-. . e••• .. ·, . . .. . . . . . . . . . . , ........ · ...... . -· -· -· -· -·
, ' , ' , ' , ' , ' , ' , ' , ' , ' ~---------- ' ,~ ' , .
,'
... ~ ·-·-·"" ~8-·- . ., , _,
·,. ' ·, ·. ·, ·.
·~:·. .,
14h00 18h00 22h00 02h00 06h00 10h00
Figuur 25.
Tyd
Veranderinge in voginhoude van vier grasspesies gedurende n 24-uur periode (Juliemaand) in die Amsterdam-studiegebied.
N 0

121
Oorbietjies is wel ook aktief wanneer die voginhoude die laagste is
(om 14h00). n Addisionele bron van water kan deur die oorbietjie se
haarkleed verskaf word tydens reen of na ryp- of douneerslag. Verskillende
lyfkosingsaktiwiteite (kyk HOOFSTulZ 8 : LYFKOSINGSAKTIWITEITE) kan dan
hier n belangrike funksie vervu1. Monfort en Monfort (1974) het ook ver-
moed dat oorbietjies moont1ik die dou benut wanneer hulle vroegoggend
in die Nasionale Akagera-wildtuin wei. Jungius (1971) noem die moontlikheid
dat rietbokke in die Nasiona1e Krugerwildtuin moontlik gedurende Junie en
Julie groot hoeveelhede water inkry deur middel van dou.
Min is bekend oor die waterbehoeftes van die ander Neotragini. Volgens
beskikbare inligting b1yk die klipspringer (Shortridge 1934, Norton 1980)
en steenbok (Eloff 1959) ook onafhanklik van drinkwater te wees. Schoen
(1972) en Ma1oiy (1973) het gevind dat die dikdik baie doeltreffende
waterbesparende meganismes ontwikkel het deur urine te konsentreer.
Die afhanklikheid van vrye water stel sekere vereistes aan die fisiese
struktuur van n boksoort se tuisgebied. By boksoorte soos die oorbietjie
kan dit n groot beperkende faktor wees wat die verspreiding van tuisge
biede betref aangesien n oorbietjie se tuisgebied relatief klein is en
die hele gebied verdedig word (kyk HOOFSTUK 9 : TERRITORIAL£ GEDRAG).
Afgesien hiervan kan addisionele bewegings om water te bekom die waarskyn
likheid van predasie verhoog veral in die geval van die oorbietjie wat
grootliks op primere verdedigingsmeganismes staat rnaak om potensiele roof
diere te ontwyk (kyk HOOFSTUK 10 :VLUGGEDRAG EN ANTIROOFDIERVERHOUDINGS).

122
HOOFSTUK 8
AKTIWITEITSPATRONE
Inleiding
Die verskeie aktiwiteite van diere is nie eweredig in tyd versprei nie
en duidelike fases kan voorkom waartydens sekere aktiwiteite kan plaas
vind, en die volgorde van hierdie fases word algemeen as die aktiwiteits
patroon beskryf (Leuthold 1977). Verskeie groat soogdiersoorte se daag
likse aktiwiteite is reeds.kwantitatief ondersoek deur Spinage (1968) op
die defassa waterbok Kobus ellipsipryrnnus derassa, Clough en Hassam (1970)
op die vlakvark, Joubert (1971, 1972) op die Harbmannsebra Equus zebra
hartmannae, Jarman en Jarman (1973) op die rooibok, terwyl Wyatt en
Eltringham (1974) asook Guy (1976) olifante Loxodonta africana bestudeer
het. Die aktiwiteite van klipspringers is deur Norton (1980) ondersoek.
Alhoewel baie werkers melding maak van daaglikse aktiwiteitspatrone by
Afrika-boksoorte bestaan daar egter min kwantitatiewe gegewens. Die
doel van hierdie studie was om die oorbietjie se aktiwiteitspatrone kwan
titatief te ondersoek terwyl aandag ook aan die faktore wat hierdie patrone
be~nvloed, gegee is.
Metodes
Spinage (1968) beklemtoon die belangrikheid om metodes by die studie van
Afrika-soogdiere te standaardiseer sodat direkte vergelykings tussen
verskillende spesies gedoen kan word. Verskeie werkers, waaronder Spinage
(££. £!!.) en Jarman en Jarman (1973), het van 4-minuut intervalle gebruik
gemaak om aktiwiteite te noteer soos aanbeveel deur Rollinson, Harker en
Taylor (1956). Ander werkers, onder andere Innis (1958) en Waser (1975)
het 5-minuut intervalle gebruik.
Vir alle dagligwaarnemings op aktiwiteitspatrone van oorbietjies is 4-
minuut intervaile gebruik waartydens die aktiwiteite in vyf hoof katagoriee

123
geplaas is, naamlik wei, lyfkosing, urinering/ontlasting/merk, le en
sosiale aktiwiteite. Aangesien kliermerking dikwels met urinering en
ontlasting gepaard gegaan het, is dit om praktiese redes as een aktiwi
teit beskryf. Dagligwaarnemeningsperiodes het van 06h00 tot 18h00
gestrek, maar in sommige gevalle kon oorbietjies nie voor 07h00 opgespoor
word nie. Vir die seisoenale waarnemings is vier seisoene
onderskei, nl. somer (Desember, Januarie en Februarie), herfs (Maart,
April en Mei), winter (Junie, Julie en Augustus) en lente (September,
Oktober en November).
Alle waarnemings is tot kamp 11 in die Amsterdam-studiegebied (Fig. 5)
beperk. Enkele familiegroepe in hierdie kamp is oor n periode van ver
skeie maande gekondisioneer om die teenwoordigheid van n voertuig te aan
vaar. Die waarnemings is gevolglik vanaf n voertuig oor afstande van 40
tot 300 m gedoen sender dat die oorbietjies se aktiwiteite befnvloed is.
n Leitz 8 x 40 verkyker is gebruik om die waarnemingsafstand prakties so
ver moontlik te verleng. Die voertuig is dikwels verskuif wanneer die
diere buite sig beweeg het.
Afgesien van die vyf reeds genoemde aktiwiteitskatagoriee is alle klier-
merkings, urinering en ontlasting afsonderlik genoteer. Daarby is data
ook oor alle ander gedragsaspekte versamel wat sosiale en territoriale
gedrag insluit.
Die invloed van sekere weersomstandighede, naamlik wind, temperatuur en
reenval op aktiwiteite is ook ondersoek. n Casella termohigrograaf is
in n Stevensonskerm in kamp 11 geplaas om inligting oor temperatuur
veranderinge in te win. Die temperatuurwaarnemings is ononderbroke ge
durende die studieperiodes gedoene Windspoed is met behulp van n bakkies
anenometer bepaal gedurende 15-minuut intervalle vir periodes van twee
minute elk gedurende die waarnemingsperiodes.
n Poging is ook aangewend om die aktiwiteite van oorbietjies gedurende
die nag te bestudeer. n Scotos 1 nagbesigtigingsteleskoop is gebruik
terwyl die oorbietjies aktief was, maar dit was meestal o~~oontlik om
hulle op te spoor tydens onaktiewe periodes wanneer die plantegroei sig
baarheid benadeel het. Onder hierdie omstandighede is n 12 volt kollig

124
gebruik om die posisie van die oorbietjies vas te stelo Om versteuring
deur die gebruik van die kollig te beperk, is die waarnemingsintervalle
tot 15 minute verleng. Nagstudies op ander Afrika-hoefdiere se aktiwi·
teite was hoofsaaklik tot maan1ignagte beperk, byvoorbee1d Spinage (1968).
Waser (1975) het egter ook met sukses van n nagbesigtingsteleskoop tydens
sy studie van die bosbok Tragelaphus scriptus gebruik gemaak.
Die aktiwiteite is as n persentasie vir elke uur uitgedruk. Aktiewe
periodes is gedefinieer as die periodes waartydens die oorbietjies beweeg
het en verskillende aktiwiteite soos lyfkosing, urineer en ontlas, wei
en sosiale interaksies vertoon het. Die periodes waartydens die oor
bietjies gele het is as onaktief beskou.
Resultate en Bespreking
BASIESE AKTIWITEITSPATROON
Aktiewe periodes wissel van dag tot dag waar periodes van rus met periodes
van aktiwiteit afgewissel word (Fig. 26). n Basiese aktiwiteitspatroon
kan, net soos in die geval van ander soogdiersoorte wat bestudeer is,
onderskei word wat vir alle seisoene geld. Die meeste oorbietjies is
vroegoggend, gedurende die middel van die dag en laatmiddag aktief (Fig.
26). Hierdie aktiewe periodes word met twee onaktiewe periodes afgewissel
waar gemiddeld 96,4% van die oorbietjies onafktief is tussen 07h00 en 08hOO,
en 80,78% tussen 15h00 en 16hOO.
Gedurende die somermaande is daar n relatiewe groot aantal oorbietjies
(gemidde1d 81,48%) tussen llhOO en 12h00 aktief terwyl so veel as ge-
middeld 99,4% onaktief is tussen 07h00 en 08h00 (Fig. 27). Die somer-
aktiwiteitspatrone verski1 in verskeie opsigte van die aktiwiteitspatrone
gedurende die wintermaande (Fige 27). n A1gemene toename in aktiwiteite
tussen 06h00 en 07h00 asook tussen 16h00 en 18h00 word aangetref. Die
aktiewe periode gedurende die middel van die dag verskuif na die periode
tussen 09h00 en lOhOO terwyl meer oorbietjies onaktief is gedurende die
warmste gedeelte van die dag (13h00 tot 16h00). Al1e oorbietjie-aktiwi
teit~ is goed gekoordineer, maar sinkronisering is beter in kleiner
familiegroepe as in die geval van groter groepe (Fig. 28). Dorst en

125
100
-N
~ ~
"'"" e ~ ..c
"' :~ 50 -; :.c 0 0
.E c 0 <
06h00 12h00 18h00
[il Onaktief (Je) D Aktief (wei) • Ander aktiwiteite
(532 woornemingsure, aile seisoene)
SKEMATIESE VOORSTELLING VAN VIER VERSKILLENDE VOORBEELDE VAN DAAGLIKSE
AKTIWITEITSPATRONE
2
3
4
--l -- I
I
I !
it I I I
-l I
I T .
I I -l
I i~ .. i
i IL.:J I
06h00 12h00 18h00
Figuur 26.
D Onaktiewe oorbietjies • Aktiewe oorbietjies
Daaglikse aktiwiteitspatrone van oorbietjies in die Amsterdam
studiegebied.

N
" -" -" e 4i ..a ~
.! ·; :a 0 0
a c 0
<(
06h00
06h00
Figuur 27.
12h00
Somer
12h00
Winter
il!l Onaktief (lei
126
50
18h00 06h00
18h00 06h00
0 Aktief {wei)
(418 waarnemingsure)
12h00
Herfs
12h00
Lente
18h00
18hOO
• Ander aktiwiteite
Seisoenale aktiwiteitspatrone van oorbietjies in die Amsterdam
studiegebied.

Figuur 28 . Oorbietjie- familiegroepe toon 'n groat mate van koordinasie
tydens alle aktiwiteite.
Amsterdam-studiegebied.
Oorbietjies gedurende n rusperiode,
N -...,J

128
Dandelot (1970) meld ook dat oorbietjies gedurende die warrnste deel van
die dag le.
Rooseveldt en Heller (1914) en Edwards (1969b) het ook opgemerk dat oor-
bietjies enige tyd van die dag of nag aktief kan wees. n Soortgelyke
basiese patroon waar aktiewe periodes onaktiewe periodes afwissel is
deur Tinley (1969) by die dikdik Madoqua kirki en Norton (1980) by die
klipspringer gerapporteer. Min rooibokke wei gedurende die periode
direk na sonop (Jarman en Jarman 1973). Esser (1973) meld dat vaalrib
bokke vroegoggend en laatmiddag n toename in aktiwiteite toon.
WEI-AKTIWITEITE
Oorbietjies wei gemiddeld 83,57% van hul aktiewe periodes waartydens geen
diere herkou nie. Gemiddeld word 28,64% gedurende n 12-uur dagligperiode
aan wei gespandeer.
Volgens duPlessis (1972), Lynch (1974) en Novelli (1975) wei blesbokke
minder gedurende die winterrnaande, maar soos in die geval van springbokke
(Novelli ££• £!l.) en klipspringers (Norton 1980) toon oorbietjies n toe-
name in wei-aktiwiteite gedurende die winter (Tabel 25) wanneer die tyd aan wei
gespandeer van gemiddeld 26,3% tot 31,6% toeneem. Jarman en Jarman (1973)
het gevind dat n toename in wei-aktiwiteite ook by rooibokke gedurende die
winter voorkom.
In vergelyking met ander groat soogdiere spandeer oorbietjies minder tyd
aan voeding. Waterbokke wei gemiddeld 42,1% van die dagligperiodes (Spi
nage 1968), rooibokke 39,9% (Jarman en Jarman~· cit.) en bosbokke 38%
(Waser 1975). Slegs 22,5% van n 24-uur periode word deur vlakvarke aan
voeding gespandeer (Clough en Hassam 1970).
Daar is geen betekenisvolle verskil in die tyd wat aan weiding gespandeer
word tussen oorbietjieramrne en ooie nie (Tabel 26; p>O,l). Volgens David
(1973) wei bontebokooie Darnaliscus dorcas dorcas meer as die ramme.
(1975) bevestig dit met sy studie op springbokke en blesbokke.
Novelli
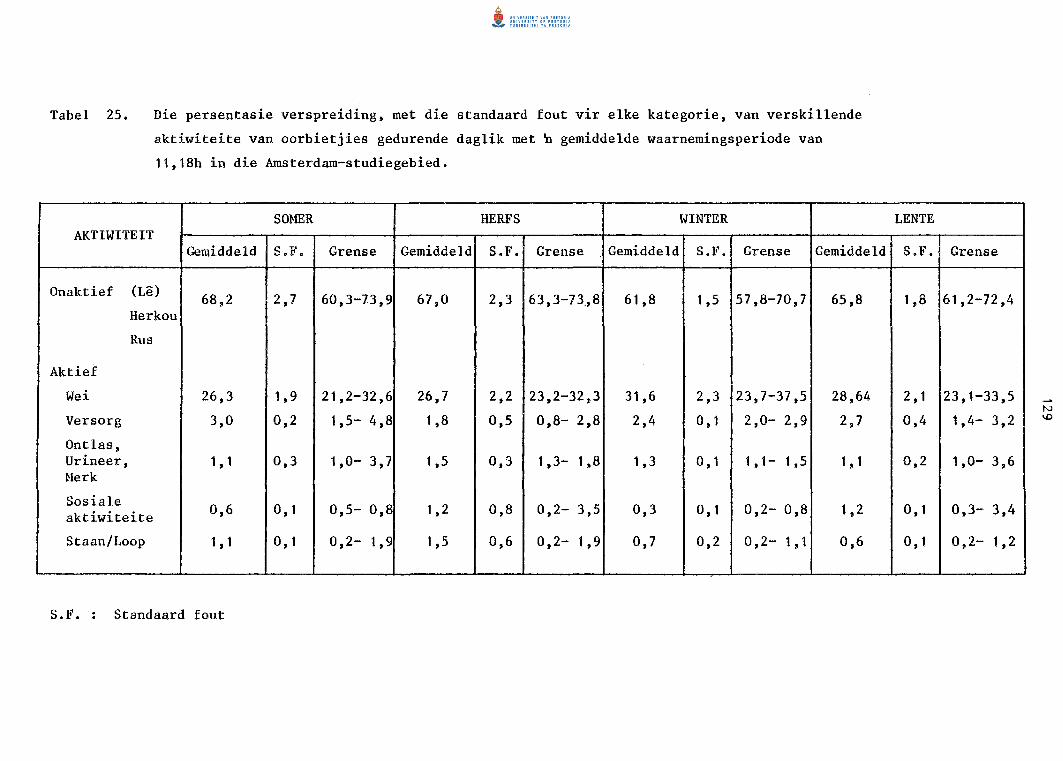
Tabel 25. Die persentasie verspreiding, met die standaard fout vir elke kategorie, van verskillende
aktiwiteite van oorbietjies gedurende daglik met n gemiddelde waarnemingsperiode van
11,18h in die Amsterdam-studiegebied.
SOMER HERFS WINTER AKTIWITEIT
Geniiddeld S.F. Grense Gemiddeld S.F. Grense Gemiddeld S.F. Grense Gemiddeld
Onaktief (U!) 68,2 2,7 60,3-73,9 67,0 2,3 63,3-73,,8 61 ,8 1 '5 57,8-70,7 65,8
Herkou
Rus
Aktief
Wei 26,3 1, 9 21,2-32,6 26,7 2,2 23,2-32,3 31,6 2,3 23,7-37,5 28,64
Versorg 3,0 0,2 1,5- 4,8 1 '8 0,5 0,8- 2,8 2,4 0' 1 2,0- 2,9 2,7
Ontlas, Urineer, 1 ' 1 0,3 1 ,o- 3,1 1 '5 0,3 1,3- 1,8 1 '3 0' 1 1,1-1,5 1 ' 1 Nerk
Sosiale 0,6 0' 1 0,5- 0,8 1 , 2 0,8 0,2- 3,5 0,3 0' 1 0,2- 0,8 1,2 aktiwiteite
Staan/Loop 1 ' 1 0,1 0,2- 1,9 1 '5 0,6 0,2- 1,9 0,7 0,2 0,2- 1,1 0,6
----------------- - ~~· ---------- -- -- - ---------- •~~ -~
S.F. : Standaard fout
LENTE
S.F.
1 '8
2' 1
0,4
0,2
0' 1
0, 1
Grense
61,2-72,4
23,1-33,5
1,4- 3,2
1,0- 3,6
0,3- 3,4
0,2- 1,2
I I
I I
I I
I I I
N \D

Tabel 26.
Aktiwiteit
Le
Wei
Versorg
Outlasting, Urinering
Sosiale aktiwiteite
Staan/loop
Die verhouding van verskillende aktiwiteite tussen oorbietjie dd en ~~ in die Amsterdam-studiegebied
met n gemiddelde en totale waarnemingsperiode van 11,18h en 113,0h respektiewelik.
FAMILIE-GROEPE : AANTAL WAARNEMINGS EN VERHOUDINGS TUSSEN dd EN ~~
d~ d~ d~~ d~~ dd~ dd~ d~ dd~':? dd~~ ddd~~':?
d ~ d ':? d ~ d ~ d ~ d ~ d ~ d ':? d ~ d ~
88 97 89 81 112 238 102 238 228 126 202 105 1511 1589 402 416 450 470 741 840
1 : 1 t 1 1 : 1 , 1 1 : 1 ,06 1 : 1 , 7 1 : 1,05 1 : 0 '96 1 : 1 '05 1 : 1,03 1 : 1 '04 1 : 1,13
50 46 37 57 47 87 57 95 106 46 81 45 686 664 142 140 214 214 465 393
1 : 1,08 1 : 1 ,5 1 : 1,08 1 : 1 '2 1 : 1,21 1 : 1 , 1 1 : 1 '03 1 : 1 ,o 1 1 : 1 1 : 1,17
5 3 5 3 3 8 6 9 6 2 5 1 69 56 16 12 30 22 57 48
1 : 1,67 1 : 1,67 1 : 1,33 1 : 1,33 1 : 1,15 1 : 2,5 1 : 1 ,06 1 : 1 • 33 1 : 1.36 1 : 1.19
4 1 3 1 4 2 7 3 5 0 11 1 50 8,2 18 2 10 4 18 3
1 : 4 1 : 33 1 : 4 1 : 4,67 1 : 5,5 1 : 9 1 : 2,5 1 : 6
1 1 1 0 1 2 1 2 2 1 1 1 19 18 22 20 0 0 9 6
1 : 1 1 : 1 'I : 1 1 : 1 1 : 2 1 : 1 '06 1 : 1 ' 1 1 : 1 ,5
3 3 4 1 0 1 2 6 3 0 2 2 24 24 12 22 12 6 3 6
1 : 1 1 : 4 1 : 1 '5 1 : 2 1 : 1 1 : 83 1 : 2 1 : 2 ----------~- ---
w 0

131
LYFKOSINGSAKTIWITEITE
Lyfkosingsaktiwiteite volg gewoonlik na lang rusperiodes en sluit ak
tiwiteite in wat te doen het met die versorging van die haarkleed deur
middel van krap en lek (Fig. 29). Gemiddeld word 2,7% van die tyd (12-
uur periode) aan versorging gespandeer, en dit wissel van 3,0% gedurende
die somer tot 2,4% gedurende die wintermaande (Tabel 25, Fig. 27).
Gedurende lyfkosingsaktiwiteite word die meeste tyd aan die kopgedeeltes
gespandeer, gevolg deur die lyf, flanke en rug (Fig. 30). Daar is geen
betekenisvolle verskil wat tydsbesteding aan lyfkosingsaktiwiteite tussen
ramme en ooie betref nie (Tabel 26; p > 0,1).
URINERING, ONTLASTING EN MERK
Urinering gaan meestal gepaard met ontlasting terwyl dit ook dikwels met
kliermerk by die ramme gekoppel is (kyk HOOFSTUK 9 : TERRITORIALE GEDRAG).
Hierdie ak tiwi tei te volg gewoonlik na 'n lang rusperiode maar daar is 'n
toename tussen lOhOO en llhOO asook tussen 14h00 en 15h00.
Seisoenale verskille wissel van 1,0% gedurende die somer tot 1,8% gedurende
die herfsmaande (Tabel 25).
Die urinering- en ontlastingstempo's verskil betekenisvol tussen ramme en
ooie (Tabel 27; p < 0,01). Ramme urineer en ontlas gemiddeld een keer
elke OlhOl terwyl die tempo by ooie gemiddeld een keer elke Olh54 geduren
de aktiewe periodes is, en 03h08 en 05h50 gedurende waarnemingsure respek
tiewelik (Tabel 27). Hierdie verskil kan aan die ramme se gedrag tydens
ooie se urinering en ontlasting toegeskryf word (kyk HOOFSTUK 9 : TERRITO
RIALE GEDRAG). Territoriale ramme merk voortdurend met hulle preorbitale
kliere gedurende aktiwiteitsperiode (kyk HOOFSTUK 9 : TERRITORIALE GEDRAG).
Die merk-tempo wissel van een merk elke 00h21 tot Olh24 gedurende aktiewe
periodes, en een merk tussen 00h55 en 04hl2 vir kombinasies van gesamentlike
preorbitale en interdigitale kliermerke (Tabel 28) •
.. LE
Aktiewe periodes word gevolg deur onaktiewe periodes waartydens oorbietjies
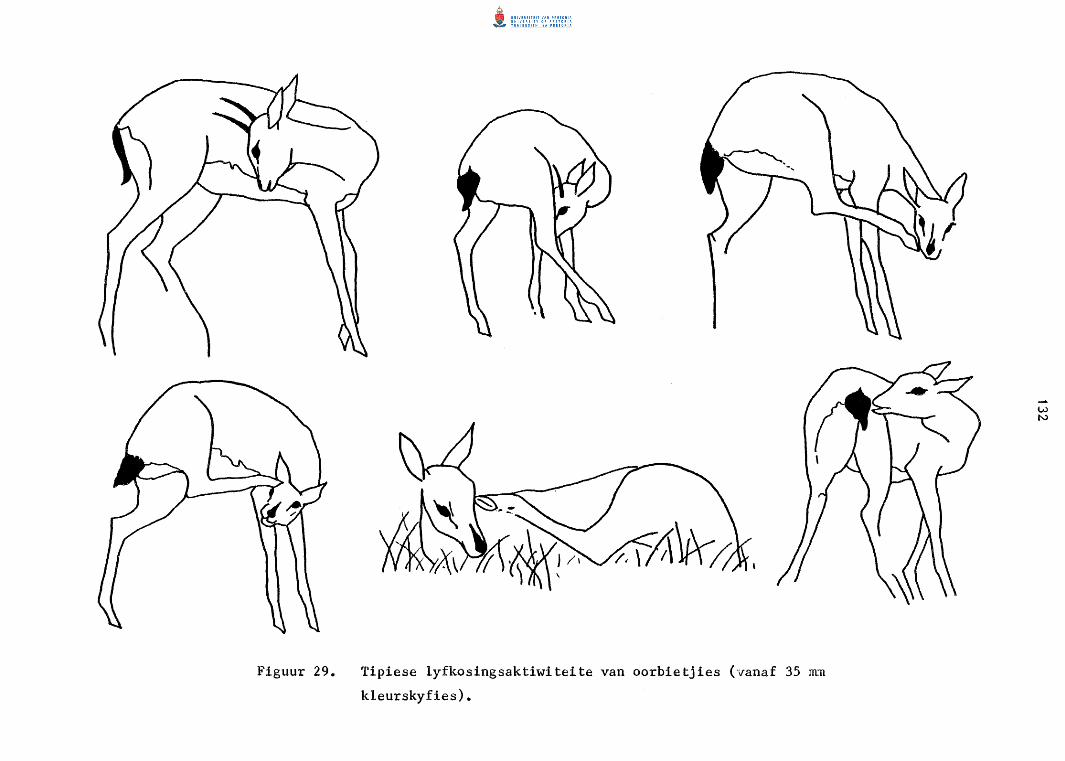
Figuur 29. Tipiese lyfkosingsaktiwi tei te van oorbie tj ies ( vanaf 35 m:n
kleurskyfies).
w N
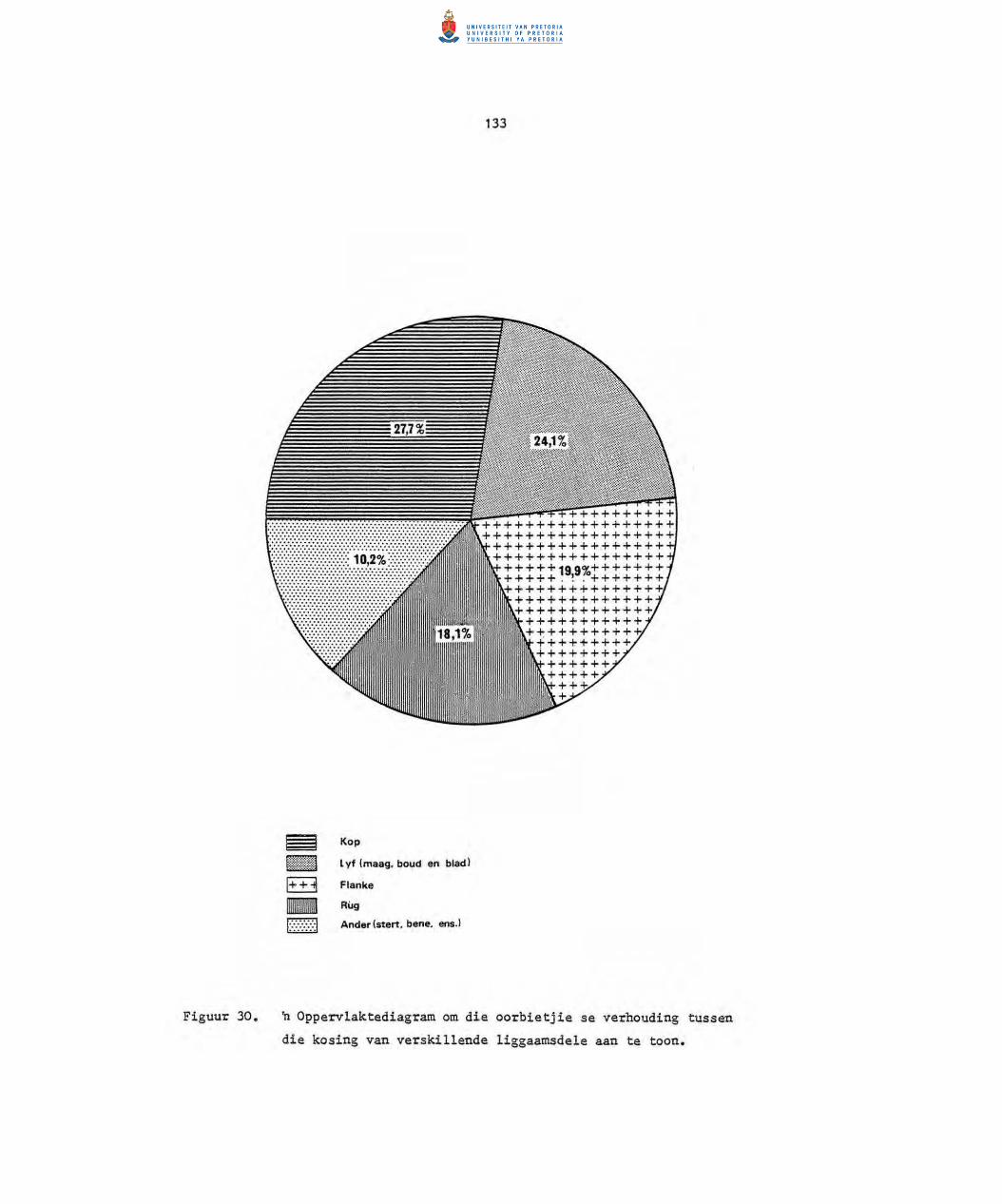
Figuur 30.
~ 1m 1++3 lillll [I;]
Kop
l yf ( maag. boud en blad l
Flanke
Rug Ander (stert. bene. ens.!
133
n Oppervlaktediagram om die oorbietjie se verhouding tussen
die kosing van verskillende liggaamsdele aan te toon.
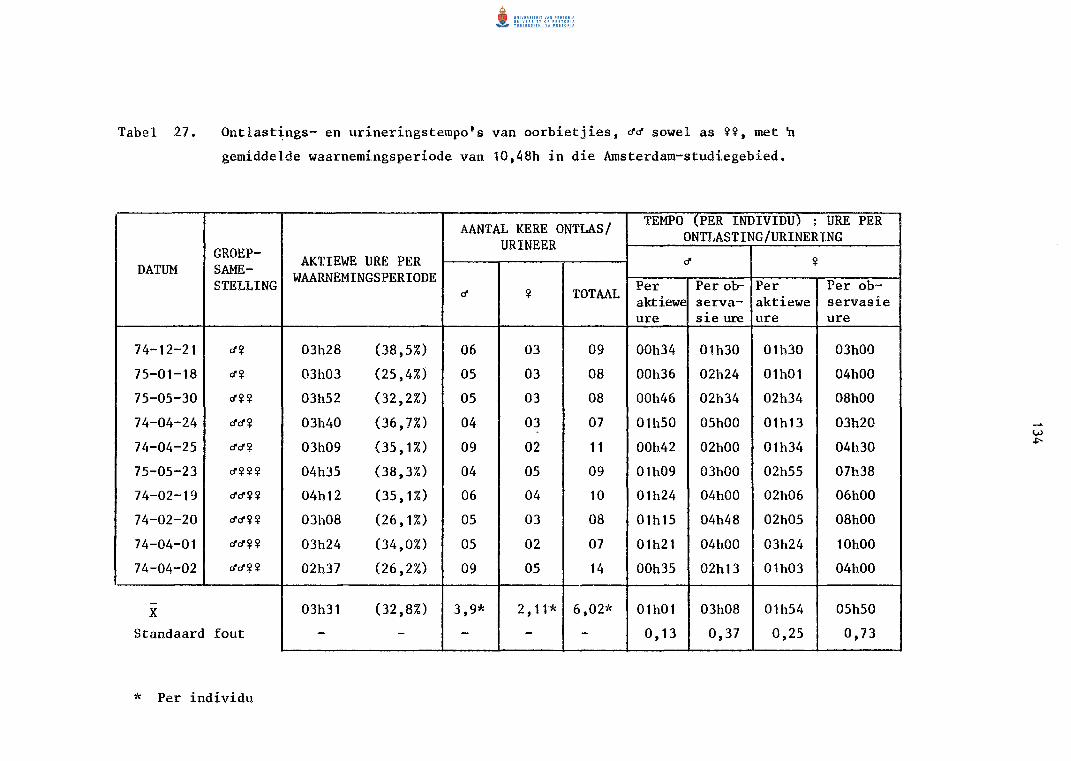
Tabel 27. Ontlast~ngs- en urineringstempo's van oorbietjies, dd sowel as ~~.met n gemiddelde waarnemingsperiode van 10,48h in die Amsterdam-studiegebied.
AANTAL KERE ONTLAS/ TEMPO (PER INDIVIDU) : URE PER ONTLASTING/URINERING
GROEP- URINEER
DATUM SAME-AKTIEWE URE PER d ~
STELLING WAARNEMINGSPERIODE Per Per ob- Per Per ob-d ~ TOTAAL aktiewe serva- aktiewe servasie
ure sie ure ure ure
74-12-21 d~ 03h28 (38,5%) 06 03 09 00h34 01h30 01h30 03h00
75-01-18 d~ 03h03 (25,4%) 05 03 08 00h36 02h24 01h01 04h00
75-05-30 d~~ 03h52 (32' 2%) 05 03 08 00h46 02h34 02h34 08h00
74-04-24 dd~ 03h40 (36' 7%) 04 03 07 01h50 05h00 01h13 03h20
74-04-25 dd~ 03h09 (35' 1 %) 09 02 11 00h42 02h00 01h34 04h30
75-05-23 d~~~ 04h35 (38,3%) 04 05 09 01h09 03h00 02h55 07h38
74-02-19 dd':/.':1. 04h12 (35, 1%) 06 04 10 01h24 04h00 02h06 06h00
74-02-20 dd~~ 03h08 (26, 1%) 05 03 08 01h15 04h48 02h05 08h00
74-04-01 dd~~ 03h24 (34,0%) 05 02 07 01h21 04h00 03h24 10h00
74-04-02 dd~~ 02h37 (26,2%) 09 05 14 00h35 02h13 01h03 04h00
- 03h31 (32,8%) 3,9* 2, 11* 6,02* 01h01 03h08 01h54 05h50 X
Standaard fout - - - - - o, 13 0,37 0,25 0,73
* Per individu
I
w .p.

Tabel 28. Veranderinge in merktempo's van oorbietjieramme, beide preorbitaal en interdigitaal,
gedurende daglig met n gemiddelde waarnemingsperiode van 11,0h in die Amsterdam
studiegebied.
AANTAL MERKINGS PER d TEMPO (PER INDIVIDUELE d) :
URE PER MERK GROEP- AK.TIEWE URE PER DATUM SAME- Per akt1.ewe Per totale STELLING W.\ARNEMINGSPERIODE ure waarnemingstyd
Preorbitaal Kombinasie* Preorbi- Kombi~ Preorbi- Kombi-* taal nasie taal nasie
74-12-21 cf~ 03h28 (38,5%) 07 03 00h29 01h09 01h17 03h00
75-01-06 cf~ 03h56 (32,8%) 11 02 00h21 01h59 01h05 06h00
75-01-18 cf~ 03h03 (25,4%) 02 02 00h36 Olh31 02h24 06h00
75-05-2.'\ cf~~ 04h35 (38 ,3%) 08 OS 00h34 OOhSS 01h30 02h24
75-05-30 cf~~ 03h52 (32,2%) 08 03 00h29 01h17 01h30 04h00
74-04-24 clef~ 03h40 (36 t 7%) 04 01 OOhSS 03h40 03h30 10h00
74-04-25 clef~ 03h09 (35, 1%) 04 01 ,s 00h47 02h06 02h15 06h00
74-02-19 cfcl'.f'.f 04h12 (35,1%) 03 01 01h24 04h12 04h00 12h00
74-02-20 clef~~ 03h08 (26, 1%) 05,5 01 ,5 00h34 02h05 02h11 08h00 I
74-04-01 cfcfn 03h24 (34,0%) 09,5 02,5 00h21 01h21 01h03 04h00
- 03h39 (33,43% 06,5 2,25 00h39 02h02 01h58 05h58 X
-~--~~--'- -
Standaard fout 0,79 0,36 0' 1 0,33 0~27 1 , 79 L___
~~-~ ~~ ~ ----~-~ --- -- -- ~- ----- ------- ~
* Preorbitale merking tesame met interdigitale merking
w lJ1

136
le. Oorbietjies le gemiddeld 65,8% van dagligperiodes (Fig 26,27 en 28)
maar minder gedurende die wintermaande (Tabel 25). Volgens Tabel 26
spandeer rarnme minder tyd om te le as ooie, maar die verskil is nie
betekenisvol nie (p > 0,1).
Anders as in die oorbietjie se geval staan verskeie ander boksoorte ook
onaktief vir lang periodes wanneer daar geherkou word, byvoorbeeld die
waterbok (Spinage 1968) en rooibok (Jarman en Jarman 1973). Oorbietjies
herkou slegs wanneer hulle le.
Ten spyte van vele beskrywings van aktiwiteite by boksoorte word daar
selde na slaap verwys. Spinage (££• cit.) beskryf diep slaap waar water
bokke met hulle koppe op die grond le (bulle) en op die flanke (koeie).
Vlak slaap word deur Spinage (2£• £!!.) beskryf waar die diere met toe oe
regop le. Balch (1955) wys daarop dat herkouers selde slaap omdat die
borskas regop gehou moet word wanneer daar geherkou word. Oorbietjies,
ramme sowel as ooie, le met tye met hulle koppe op hul flanke, soortgelyk
aan die posisie wat Spinage (2£. cit.) vir waterbokkoeie beskryf het.
Tydens vier waarnemings het oorbietjies gemiddeld 2 min. 56 sek. (2 min.
32 sek. tot 4 min. 20 sek.) in die betrokke posisie verkeer (Fig. 31).
Balch (1955) is van mening dat slaap, indien dit by herkouers voorkom,
baie kortstondig en lig is. By waterbokke is dit nooit langer as vier
minute nie (Spinage 2£• cit.).
SOSIALE AKTIWITEITE
Sosiale interaksies is beperk en vorm slegs gemiddeld 1,2% van alle akti
witeite (Fig. 26). n Toename in sosiale aktiwiteite kom gedurende die
herfs voor (Tabel 25). Geen betekenisvolle verskil (p > 0,1) kon tussen
die ramme en ooie gevind word wat tydsbesteding aan sosiale interaksies
betref nie (Tabel 26)e
AKTIWITEITE GEDURENDE DIE NAG
Oorbietjies is ook gedurende die nag aktief, ongeag of daar maanlig is of
nie (Fig. 32). Volgens die resultate blyk dit dat oorbietjies se aktiwi
teitspatrone gedurende die nag ooreenstem met die algemene daaglikse
aktiwiteite (Fig. 26 en 33) waar aktiewe periodes ook met onaktiewe periodes

137
Figuur 31. n Oorbietjieram in n slaapposisie terwyl sy kop op sy flank
rus (vanaf n 35 mm kleurskyfie) .

Figuur 32.
138
n Oorbietjieram wat gedurende die nag wei (foto geneem met
behulp van 'n nagbesigtingsteleskoop).
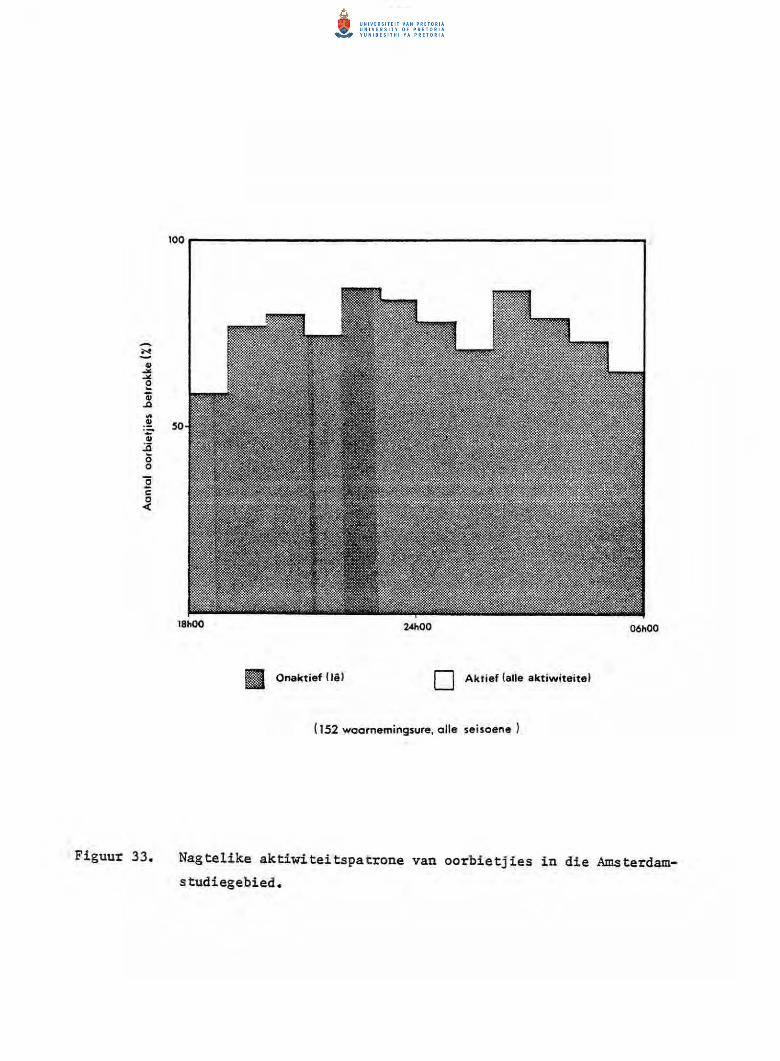
Figuur 33.
139
100~--------------------------------------------------,
l8h00 24h00 06h00
g Onaktief I I e) D Akrief {aile aktiwiteitel
( 152 woarnemingsure, aile seisoene )
Nagtelike aktiwiteitspa~rone van oorbietj i es in die Amsterdam
studiegebied.

140
afgewisse1 word.
Gernidde1d word 24,6% gedurende n 12-uur nagperiode aan die verski11ende
aktiwiteite bestee wat onder andere wei-aktiwiteite, lyfkosingsaktiwiteite
en sosia1e aktiwiteite insluit. Die aanvanklike aktiewe periode vanaf
danker word met n rede1ike onaktiewe periode later gedurende die nag af
gewisse1. Duidelike aktiewe periodes het in al die waargeneemde geva11e
weer vroegoggend voor sonop voorgekom.
FAKTORE WAT AKTIWITEITE BEINVLOED
Versteurings
Veebewegings en persone in die direkte orngewing van oorbietjies het n groot invloed op die aktiwiteitspatrone. Skape en beeste steur vera1
oorbietjies wat le (kyk HOOFSTUK 10 : ALGEMENE INTERAKSIES). Hood en
Inglis (1974) het gevind dat die bewegings van beeste ook die aktiwiteite
van herte Odocoileus virgianus beinvloed.
Indien oorbietjies 1e en deur n versteuring opgejaag word, bly die bokke
sorns vir n kort periode aktief waartydens bulle wei, lyfkoos of urineer
en ontlas. Voortdurende versteurings veroorsaak gevolglik n toenarne in
die totale tyd wat aan aktiwiteite gespandeer word.
Invloed van Weersomstandighede
Makroklimaatstoestande bernvloed die aktiwiteite van oorbietjies sodat n verskil tussen aktiwiteitspatrone tydens rnooiweersdae en koue, reenerige
dae opgemerk is (Fig. 34). Afgesien van reen is die invloed van ternpera
tuur en wind ook ondersoek.
Van al die faktore blyk reen die be1angrikste enke1e faktor te wees. Die
verhouding tussen aktiewe en onaktiewe oorbietjies wissel van 1.2,88(X = 74,25% onaktief; S = 0,35) gedurende mooiweersdae tot 1.1,56 (X= 60,8%
onaktief; S = 0,84) tydens reenerige dae. Op reenerige dae bly
oorbietjies dus vir langer periodes le.
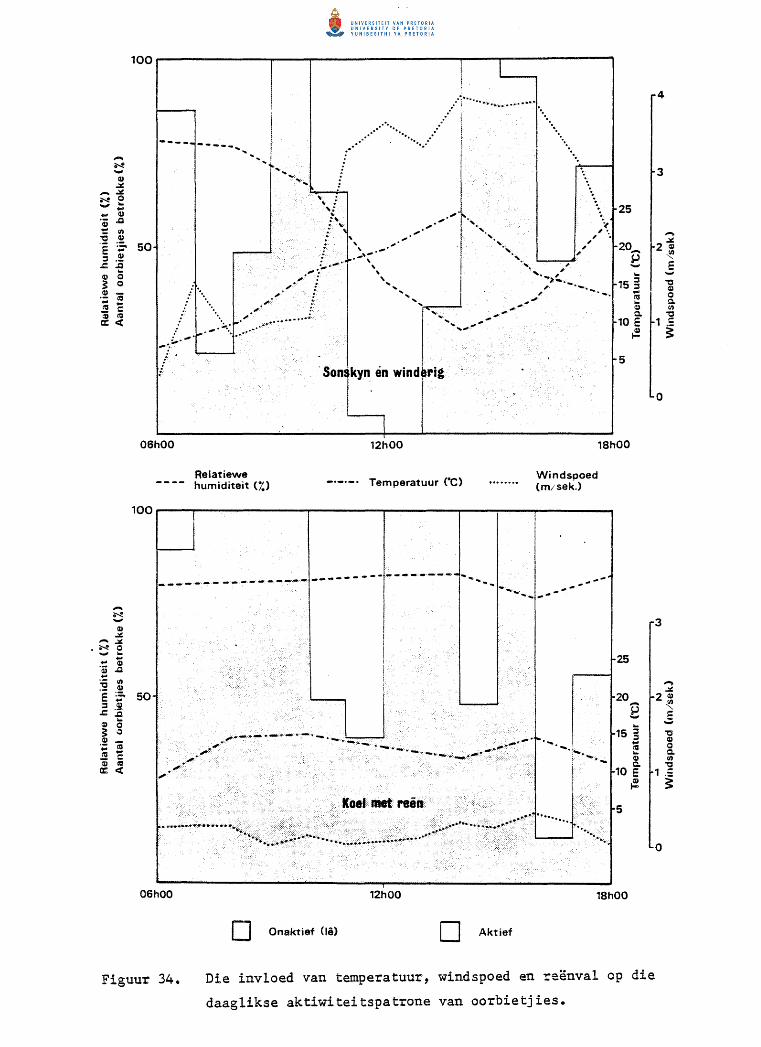
-N -Q) ...:.::
-...:.:: N 0 -;; 9"' Q) ';) .Q ... -c ·e :: .c:
-N -... ";) ... :.0 ·e :: .c: (I)
~ -~ ... ~ -a; a:
-:::; (I)
...:.::
.::t. 2 ... (I)
.Q
0
:~ ... (I)
:a 0 ~
-; ... c: ca <(
141
100~------------~--~----------------------~------------
50
---~-- ..... ,
'\. :· ·"-·i' :~
: "
,_..._:.-, .•. .,. .. ~· , . ·:
;.~-· :. ./ ·•·... ,.~
.. / ······.:...· .~·''' ~· _ ....... ····· ,. .. . ., tllfiP. • •••••• -·
4flll''··.i..· ,. ~,....-. _ __..
.·. . ........... ·;·~ ......... ·····•··
.. ······· ···· ... . ·· ·· .... ··
......... ·!'~..·,, ·, ·, ·, ·, ·, ·, , ' . , , I '· ... ,,
... ..... ... .... ' #I
... *"-" ... 41"
'._ ,..~'" ......
' ... , ~ ,' .............. _.
. Son ky.n_ en wind~rig
25
20_
e 15 ~ ..
~ Q) Q,
10 E t!
5
06h00 12h00 18h00
Relatiewe humiditeit (%) -·-·-· Temperatuur (°C)
Windspoed (m;sek.)
100r---~----~-----,--------~------~--~----~-------
-....... --------.-- .... ·---· ...................... ~·-.-.-:· ---------- _ _._. ...... : .... -----.- ...... ...... ............ _ ...... ----....... -- ..
25
50 20
E 15 ~ ::1 ...
'··~ ......... ~ ... Q) .... Q.
10 E ~
5
06h00 12h00 18h00
D Onaktief (le) D Aktief
Figuur 34. Die invloed van temperatuur, windspoed en reenval op die
daaglikse aktiwiteitspatrone van oorbietjies.
4
3
-.::t. 2 (I)
~ E -"0 Q) 0 Q, 0 "0
1 .= ~
0
3
..... ...:.::
2Q) '~ .§ ,
Q) 0 Q, 0
-g ~
0

142
Vo1gens Jarman en Jarman (1973) het reen ook n invloed op die aktiwiteite
van rooibokke wanneer hu1le ophou vreet en in n rigting weg vanaf die reen
staan en kyk. Die uitsak van skie1ike reenbuie veral gedurende die somer
maande het veroorsaak dat aktiewe oorbietjies gaan le het. Hierdie invloed
van reen is veral be1angrik as in ag geneem word dat daar gemiddeld 13 reen
dae per jaar gedurende Januarie en n totaal van 87 dae per jaar by Amsterdam
voorkom (Tabel 3).
n Sterk of koue wind het nie n merkbare invloed op die persentasie aktiwi
teite nie, behalwe dat oorbietjies neig om meer skrikkerig te wees en
gevolglik makliker vlug met steurings. Jarman en Jarman (££• cit.) het
ook opgemerk dat n sterk wind rooibokke meer skrikkerig maak, terwyl
Zagata en Haugen (1973) gevind het dat herte se bewegingsaktiwiteite
deur sterk winde beinv1oed word deurdat hulle uit die wind b1y beweeg.
Hartmansebras vermy koue winde (Joubert 1971). n Sterk, koue wind het
egter wel n daling in die tota1e tyd wat oorbietjies aan aktiwiteite span
deer tot gevolg. Norton (1980) het gevind dat weersomstandighede min
invloed op die aktiwiteite van k1ipspringers gehad het.
Ten spyte van temperature so 1aag as -6°C gedurende die wintermaande by
Amsterdam (Tabe1 2), is geen merkbare inv1oed op die aktiwiteite van oor
bietjies waargeneem nie. Se1fs gedurende die koudste gedee1te van n 24-uur periode in die winter is gemidde1d 45,93% aktiewe oorbietjies tot
voor sonop aangetref. Lae temperature laat rooibokke vroegoggend 1e
(Jarman en Jarman~. cit.), en Hartmannsebras spandeer minder tyd aan wei
aktiwiteite onder sulke toestande (Joubert ££• £!!.).
Tydens die warmste gedeelte van die dag (13h00 - 15h00) het oorbietjies n onaktiewe periode (Fig. 26 en 27), maar Jarman en Jarman (op. cit.) het
gevind dat die warmste periode nie met die minste aktiwiteite van rooibokke
saamval nie. In Oos-Afrika is aangetoon dat diere die meeste liggaams-
hitte verloor wanneer hulle onaktief in die skaduwee staan gedurende die
warmste gedeelte van die dag (Bligh en Harthoorn 1965). Diere soos die
Hartmannsebra se aktiwiteite word verder deur hoe dagtemperature betnvloed
deurdat hulle drinkplekke aktief besoek.

143
Gevolgtrekking
~ Basiese aktiwiteitspatroon kan, teo spyte van baie variasies , wel by cie
oorbietjie waargeneem word . Aktiwiteitspatr one kan deur verskeie faktore
be1nvloed wor d waar onder r eenval en versteurings deur vee en mense die
belangrikste is . Behalwe in die geval van uriner ing- en ontl astingsakti
witeite, is daar geen be tekenisvolle verskille in die aktiwiteite van ramme
en ooie nie.
Oor bietjies is aktief gedurende die dag en nag a lhoewel daar n defioi t i ewe
neiging is om meer aktief gedurende daglig te wees . Al hoewel groter bok
soor te soos die waterbok (Spioage 1968 ) eo r ooibok (Jarman en Jarman 1973)
hoofsaaklik dag l ewend is, is verskeie boksoor t e wat in klein gr oepies of
enkellewend voorkom soos die dikdik en rietbok (Leuthold 1977 ) meer nag-
lewend . Hierdie diere vertrou hoofsaaklik op primere verdedigiogsmeganis -
mes (kyk HOOFSTUK 10 : VLUGREAKSIES EN ANTIROOFDIERVERHOUDINGS) om poten-
siele roofdiere te ootwyk eo miode r op gesig. Die oor bie tjie maak van
primer e verdedigingsmeganismes sowel as gesig gebruik om mooot like pr edasie
te voorkom en die tydsver spreiding van aktiwiteitsper iodes is gevol glik
van minder be l ang . Oor bietjies se aktiwiteite is goed gesinkr oniseer
terwyl aktiwiteite soos sosiale inter aks i es tot n mi nimum beper k is en selfs
herkou gedureode leperiodes plaasvind. Pr imere verdedigingsmeganismes
wor d gevolglik maks i maal beout.

144
HOOFSTUK 9
SOSIALE ORGANISASIE
Inleiding
Sosiale organisasie is die gevolg van alle sosiale interaksies en ruim
telike verhoudings tussen die lede van n enkelspesiebevolking (Leuthold
1977). Die sosiale organisasie van n spesie sluit aspekte soos groep
tipes, groepgroottes, verboudings van individuele diere ten opsigte van
ander gelyksoortiges en verboudings tussen verskillende geslags- en ouder
domsgroepe in. Leuthold (££. cit.) wys ook daarop dat alhoewel die sosiale
organisasie meestal spesie-spesifiek is dit onderworpe is aan groat veran
deringe weens omgewingstoestande soos klimaat en plantegroeitipes. Wynne
Edwards (1962) bet voorgestel dat die sosiale organisasie n belangrike
funksie bet omdat optimum verspreiding en oorlewing bevorder word deur
kompetisie. Volgens Jarman (1974) se hipotese is daar n nou verband tussen
die ekologie en sosiale organisasie van Afrika-boksoorte.
Vir die doel van hierdie studie word sosiale gedrag beskou as daardie ge
dragspa trone- spesie-spesifieke aksies en kommunikasie- waarmee gelyk
soortiges in n groep inter-reageer en mekaar se aksies bernvloed (Morrison
en Menzel 1972). Min is bekend oar die sosiale gedrag van die Neotragini,
hoofsaaklik omdat die meerderbeid enkellewend is en slegs die oorbietjie
en klipspringer in pare of familiegroepe voorkom. Oorbietjies gebruik
verskeie gedragspatrone tydens sosiale interaksies, maar die intensiteit
en gebruik hang van die omstandigbede en sosiale posisie van die betrokke
diere af. Direkte sosiale interaksies is in die oorbietjie se geval be
perk en vorm slegs gemiddeld 1,2% van alle aktiwiteite (kyk HOOFSTUK 8 :
SOSIALE AKTIWITEITE).
Hediger (1951) en Gosling (1972) het voorgestel dat oorbietjies territoriaal
is, terwyl Monfort en Monfort (1974) verklaar bet dat oorbietjies wel in
Rwanda territoriaal is. Nie een van bierdie werkers gee egter n gede-
tailleerde beskrywing van die oorbietjie se territoriale stelsel nie.

145
Metodes
GROEPSAMESTELLING
Aangesien oorbietjies relatief moeilik uit die lug sigbaar is en weens
die hoe koste verbonde aan lugtellings, is vier seisoenale grondtellings
in beide die Amsterdam- en Piet Retief-studiegebiede gebruik om:
(a) die totale oorbietjiebevolking te bepaal,
(b) die bevolkingstruktuur en groepsamestelling vergelykend te ondersoek, en
(c) gegewens te versamel oor spesifieke habitatsvoorkeure en verspreiding.
Totale tellings is deurgaans gedoen wee~s die diversiteit van veral die
Piet Retief-studiegebied wat beide topografie en plantegroei betref, en
die mosaiek van landerye en plantasies in die Amsterdam-studiegebied.
Wildsoorte wat algemeen in oopveld of in groot groepe voorkom word vel
gens Robinette(l970) wel bevredigend deur totale tellings gesensus.
Volgens Robinette (££. £!!.) kan boksoorte soos steenbokke en oorbietjies
onder andere nie akkuraat deur middel van n monstermetode getel word nie.
Feitlik alle tellings is te voet onderneem aangesien die gebruik van voer
tuie uiters beperk was weens die klipperige en soms bergagtige voorkoms
van die grootste gedeeltes van die twee studiegebiede. In die Amsterdam
studiegebied is n voertuig wel in die oulande gebruik.
Die studiegebiede is vooraf in sensusblokke verdeel wat ooreenstem met die
bestaande kampe (Fig. 5 en 6). Robinette (££. cit.) meld dat totale
tellings slegs deur die gebruik van klein eenhede meet geskied, en hierdie
eenhede meet verkieslik begrens word deur uitkenbare gedeeltes soos paaie
en riviere. Norton-Griffiths (1975) wys ook op die belangrikheid van
klein sensusblokke tydens grondtellings.
Elke sensusblok is stelselmatig deursoek deur n ry waarnemers bestaande
uit agt tot tien persone. Hierdie ry waarnemers het in reguit, paralelle
telstroke beweeg. Die afstande tussen elke twee waarnemers het gewissel
na gelang van plantegroeidigtheid en topografie waar die waarnemers nader
na mekaar beweeg het in langgras en meer bergagtige gedeeltes weens die
korter vlugafstande van oorbietjies onder sulke omstandighede (kyk HOOFSTUK

146
10 : Primere Verdedigingsmeganismes). Radiokornmunikasie is gebruik om
kontinufteit te behou terwyl van verkykers gebruik gemaak is om geslagte
en waar moontlik ouderdomsklasse van die oorbietjies te bepaal. Slegs
volwasse en jong bokke (tot ongeveer vier maande oud) is onderskei.
Weersomstandighede is ook in aanmerking geneem aangesien oorbietjies by
voorbeeld gedurende koue, reenerige dae minder aktief is (kyk HOOFSTUK 8
FAKTORE WAT AKTIWITEITE BEINVLOED) en voorlopige waarnemings het getoon
dat die bokke moontlik ook korter vlugafstande onder hierdie weersomstandig-
hede kan vertoon.
weersdae beperk.
Alle tellings is gevolglik sover moontlik tot mooi-
Nadat elke telstrook voltooi is, is die aantekeninge van elke waarnemer
nagegaan. Die waarnemers op die verste punte van elke telstrook het
voortdurend gepoog om herhaalde tellings van dieselfde oorbietjiegroepe
te verhoed. Norton-Griffiths (1975) beklemtoon die belangrikheid om
elke groep diere wat tydens totale telling opgemerk word afsonderlik
aan te teken sodat foute tot n minimum beperk kan word. Gedurende die
tellings is ook aantekeninge gemaak van alle ander hoefdiere, nie net
wat die verspreiding betref nie, maar ook getalle. Probleme is in sornmige
gevalle ondervind om tussen onvolwasse en volwasse ooie te onderskei. In
die geval van jong ramme kan horings eers maklik op n ouderdom van ongeveer
agt maande gesien word.
GEDRAG
Verskillende gedragsaspekte, naamlik sosiale gedrag, territoriale gedrag
en voortplantingsgedrag, is hoofsaaklik bestudeer tydens die aktiwiteits
patroonstudie (kyk HOOFSTUK 8 : Metodes). Hierdie studie is in die
Amsterdam-studiegebied, veral in kampe 11 en 17 (Fig. 5), gedoen. Die
waarnemings is vanaf n voertuig gedoen oor afstande wat tussen 40 en 300 m
gewissel het waartydens n Leitz 8 x 40 verkyker en n enkellensreflekskamera
gebruik is. Enkele familiegroepe het na n periode van verskeie maande aan
die teenwoordigheid van n voertuig gewoond geraak, sender dat hul gedrag
enigsins be~nvloed is. Die uitkenning van individue inn bevolking is
noodsaaklik om die sosiale gedrag van n spesie te identifiseer (Kitchen
1974). Individuele oorbietjierarnme kon uitgeken word weens verskille
in die vorm en lengte van hulle horings. Die uitkenning van individuele
ooie het geskied na aanleiding van verskille in die kleur van die haar-

147
kleed en gesigsmerke. n Totaal van vier ramme en vyf ooie kon, deur
op die individuele verskille te let, onderskei word.
Die lokaliteite en grootte van enkele oorbietjie-territoriums in die
Amsterdam-studiegebied is deur midde1 van waarnemings op bekende familie
groepe gedurende die studietydperk (Desember 1973 - Julie 1976) bepaa1.
Ten einde n vergelyking met die Piet Retief-studiegebied te tref is van
telemetriese opsporing in die Piet Retief-studiegebied gebruik gernaak
weens die meer bergagtige geaardheid daarvan (kyk HOOFSTUK 5 : DIE PIET
RETIEF-STUDIEGEBIED).
Twee oorbietjierarnme is gedurende September 1975 met behulp van nette ge
vang. Na behandeling met 'n kalmeermidde1, gemidde1d 1,8 mg/kg Azaparone
(Janssen Pharmaceutica), is e1keen met 'n AVM-radiosender (AVM Instrument
Company, I1linois)gemerk. Die senders, met 'n potensiele 1eeftyd van 84
dae, het n massa van ongeveer 300 g gehad. Beide oorbietjies is onmidde1-
lik hierna weer in dieselfde gebied losgelaat waar hulle gevang is. Op
sporing het geskied met behu1p van AVM-toerusting,naamlik n radio-ontvanger
en draagbare Yagi-antenne. Vier observasiepunte is op geskikte hoer-
liggende dele in die omgewing uitgesoek waarvandaan die algemene rigting
van die twee oorbietjies bepaal is. Daarna is die bokke te voet gesoek
totdat hulle gesien kon word. Aantekeninge is ook gemaak van assosiasies,
vlugrigtings en aktiwiteite. n Totaa1 van 26 en 30 opsporings is van die
twee oorbietjies gedoen in die periode tussen September 1975 en Januarie
1976 waarna beide radiosenders opgehou het om te funksioneer.
vangsafstand het tot sover as 6 km gestrek.
Resultate en Bespreking
BEVOLKINGSTRUKTUUR EN GROEPSAMESTELLINGS
Die Amsterdam-studiegebied
Die ont-
Gedurende die seisoenale te11ings in die A~sterdam-studiegebied (1974 tot
1975) is n gemiddeld van 126,25 oorbietjies (S= 7,63; reeks 114-134) gete1
(Tabe1 29). n Gemiddeld van 122,5 oorbietjies (S= 9,36; reeks 114-134)
se ges1agte kon bepaal word wat 97,03% van die bevo1king was (Tabe1 29).
Dit s1uit vo1wasse en onvo1wasse diere in.

Tabel 29. Resultate van vier seisoenale oorbietjietellings
(1974-1975) in die Amsterdam-studiegebied.
Totale telling
Totaal geseks
Rannne
Ooie
Ges lagsverhouding ( cf: ~ )
x 2 (y = 1)
SOMER
114
114
52
62
1:1,19
0,88
----~--
HERFS WINTER
126 131
112 130
51 59
61 71
1 : 1 , 20 1 : 1 , 2
1,08 1 '11
---~--- ·--
LENTE
134
134
58
76
1:1,31
2,42
-X
126,25
122,5
55,0
67,5
1 : 1 , 23
1,28
+:--00

149
~ Gemidde1de ges1agsverhouding van 1,23 ooie tot ~ ram (reeks: 1 d :
1,19 ~~ - 1,31 Q~) is bepaal wat nie betekenisvol van~ l : l verhouding
op die 5% vlak verski1 nie. Die geslagsverhoudings het net betrekking
op vo1wasse oorbietjies aangesien dit moeilik was om die geslagte van
onvo1wasse of baie jong bokke te bepaal.
Tydens die te1lings is die groepsameste1lings ook bepaa1. Leuthold (1977)
definieer 'n groep as 'n aan tal ge1yksoortige diere wa t op 'n gegewe oomblik
saam is, terwyl 'n groep van 'n trop onderskei kan word weens die kleiner
aantal diere wat in eersgenoemde geval betrokke is. Die gemiddelde groep
grootte viral vier tellings was 1,84 (reeks l - 6), uitgesondered 1ammers
wat saam met groepe was, of a1leenlopende 1ammers, of sommige ander groep-
samestel1ings wat nie in Tabel 30 aangetoon word nie. Die meerderheid
oorbietjies het gedurende die tel1ings in ~ fami1iegroep bestaande uit ~
volwasse ram vergesel van ~ volwasse ooi (oorbietjiepaar) voorgekom wat
37,45% van alle groepe was (Tabe1 30), gevolg deur a1leenlopende ramme (on-
volwasse ramme ingesluit). Al1eenlopende ooie het gemidde1d 12,81% van
alle groepe gevorm, met groepe bestaande uit een volwasse ram en twee
volwasse ooie (gemiddeld 9,51% van a11e groepe). Pare vergese1 van~
onvolwasse ooi is selde opgemerk (0,74% van alle groepe), maar dit kan
toegeskryf word aan die feit dat onvolwasse ooie moeilik van volwasse ooie
onderskei kan word.
Ramgroepe, bestaande uit twee ramme, is tydens al die tel1ings waargeneem
maar het slegs 1,87% van aldie groepe gevorm teenoor die 3,77% van die
groepe bestaande uit twee ooie. Die meerderheid oorbietjies het in groepe
van twee of meer voorgekom (80,0% van al1e groepe) waarvan 39,6% in oor
bietjiepare was. Die meeste alleen1opende bokke was ramme (12,87%) met
3,96% onvolwasse ramme teenoor 6,73% van die ooie en 1,98% van ooie en
lamrners. A1hoewel enkele groepe bestaande uit meer as vier individue
(vo1wasse en onvolwasse oorbietjies) aangetref is, het die grootste groep
ui t ses individue bestaan waaronder twee onvolwasse diere ('n ram en 'n ooi).
Tabe1 30 toon ook die seisoena1e wisseling in groepsamestellings. Hierdie
veranderinge is statisties getoets om te bepaal of dit betekenisvol is (Tabel
31). Die hipotese wat getoets is, is dat die verhouding van elke waarge
neemde oorbietjiegroep tot die ander groepe saam onveranderd geb1y het
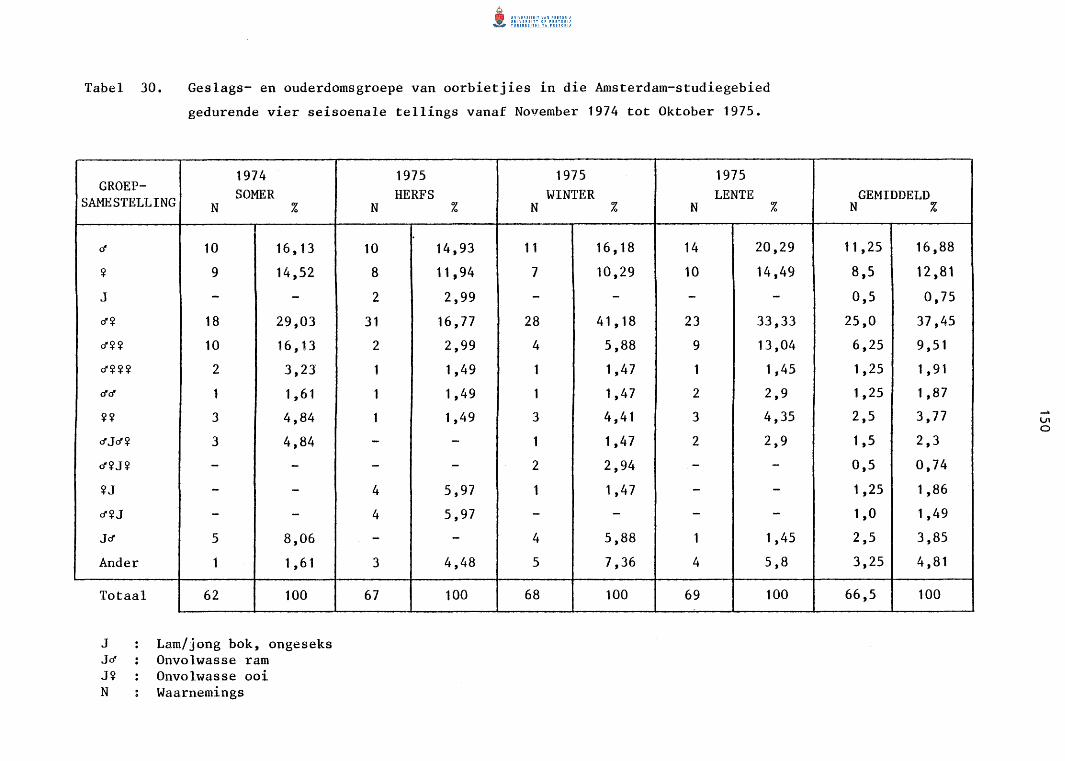
Tabel 30. Geslags- en ouderdomsgroepe van oorbietjies in die Amsterdam-studiegebied
gedurende vier seisoenale tellings vanaf November 1974 tot Oktober 1975.
1974 GROEP- SOMER SAME STELLING N %
cf 10 16' 13
~ 9 14,52
J - -d'~ 18 29,03
d'~~ 10 16' 13
d'~~~ 2 3,2J
d'd' 1 1,61
~~ 3 4,84
d'jd'~ 3 4,84
cf~j~ - -~J - -d'~j - -
jd' 5 8,06
Ander 1 1 '61
Totaal 62 100
J : Lam/jong bok, ongeseks Jd' : Onvolwasse ram J~ : Onvolwasse ooi N : Waarnemings
1975 HERFS
N
10
8
2
31
2
1
1
1
--
4
4
-3
67
1975 1975 WINTER LENTE
% N % N %
14,93 11 16' 18 14 20,29
11,94 7 10,29 10 14,49
2,99 - - - -16,77 28 41 '18 23 33,33
2,99 4 5,88 9 13,04
1,49 1 1 '47 1 1,45
1,49 1 1,47 2 2,9
1,49 3 4,41 3 4,35
- 1 1,47 2 2,9
- 2 2,94 - -
5,97 1 1,47 - -5,97 - - - -- 4 5,88 1 1 ,45
4,48 5 7,36 4 5,8
100 68 100 69 100
GEMIDDELD N %
11,25 16,88
8,5 12,81
0,5 0,75
25,0 37,45
6,25 9,51
1,25 1 '91
1,25 1,87
2,5 3,77
1,5 2,3
0,5 0,74
1,25 1,86
1 ,o 1,49
2,5 3,85
3,25 4,81
66,5 100
I
I
l/1 0

151
Tabel 31. Seisoenale veranderinge van spesifieke oorbietjiegroepe
in vergelyking met die ander groepe in die Amsterdam
studiegebied : November 1974 - Oktober 1975.
31.1 I II III
Groep c! (10,49)a (11,33)a (11,5)a (volwasse c!c! ) 10b 10b 11 b
1Nie-groep (51 ,51)a (55,67)a (56,5)a 52b 57b 57b
Totaal 62 67 68
x 2 = o,8007 (y = 3)
a = Verwagte frekwensie b = Waargeneemde frekwensie I : Somer; II : Herfs; III : Winter; IV Lente Nie-groep : res van groepe (scm van)
31.2 I II III
Groep ~ (7,92)a (8,56)a (8,69)a (volwasse ~~) 9b 8b 7b
1Nie-groep (54,08)a (58,44)a (59,31)a 53b 59b 61b
Totaal 62 67 68
x 2 = 0,9258 (y = 3)
31.3
I II III
Groep c!<f. (23,31)a (25, 19) a (25,56)a
18b 31b 28b
lN· (38,69)a (41 ,81)a (42,44)a J.e-groep 44b 36b 40b
Totaal 62 67 68
x 2 = 4,993 (y = 3)
IV N
(11,67)a 45 14b
(57,32)a 221 55b
69 266
IV N
(8,82)a 34 10b
(60, 18)a 232 59b
69 266
IV N
(25,94)a 100 23b
(43,06)a 166 46b
69 266
152
Tabel 31 (vervolg)
31.4 I II III IV N
Groep a-~~ (5,83)a (6,3)a (6,3)a (6,48)a 25 10b 2b 4b 9b
1 Nie-groep (56,17)a (60,7)a (61 ,61)a (62,52)a 241 52b 65b 64b 60b
Totaal 62 67 68 69 266
x 2 = 8,6 (y = 3)
31 .5 I & II III & IV N
Groep ~~ (4,85)a (5,15)a
10 4b 6b
1Nie-groep (124,15)a (131,85)a 256 125b 131b
Totaal 129 137 266
x2 = o,1s2

153
gedurende die seisoenale tellings (Marascuilo 1966). Aangesien ~
frekwensie van vyf of meer nodig is vir n Chi-kwadraatberekening (Sakal
en Rohlf 1969), kon aldie groepe nie met hierdie hipotese getoets word
nie. In die geval van ooigroepe (bestaande uit twee ooie elk) is somer
en herfs, en winter en lente saamgevoeg om n hoer frekwensie te verkry.
Volgens Tabel 31blyk dit dat alleenlopende ramme, alleenlopende ooie, pare
(x2
en ooigroepe geen betekenisvolle seisoenale wisseling getoon het nie <
7,815 Stoker 1977). Slegs in die geval van een groeptipe (een ram en
twee ooie) het betekenisvolle seisoenale veranderinge voorgekom waar
die hoogste frekwensie gedurende die lente en somer (13,04 en 16,13%
van alle groepe respektiewelik) voorgekom het (Tabel 31.4).
Gedurende die tellings is slegs twee lammers opgejaag wat versteek gele
het (herfs), en vyf ooie is gewaar wat van larnmers vergesel was (herfs:
5,97% van alle groepe; en winter: 1,47% van alle groepe). Hierdie lam-
mers was almal ouer as drie tot vier maande. Duidelike verskille is in
die geval van alleenlopende ramme waargeneem waar n toename gedurende
die somer- en wintermaande gekry is.
Die Piet Retief-Studiegebied
n Gemiddeld van 196,5 oorbietjies (S= 17,78; reeks: 175-216) is gedurende
die seisoenale tellings (1974 tot 1975) in die Piet Retief-studiegebied
getel (Tabel 32). Die geslagte van 192,5 oorbietjies (S= 9,36; reeks
168-212) kon bepaal word wat 97,96% van die bevolking was (Tabel 32).
Meer ooie as ramme is deurgaans getel sodat n gemiddelde geslagsverhouding
van 1,31 ooie tot een ram (reeks 1~ : 1,21 ii - 1~ : 1,38 ~~) gekry is.
Hierdie verhouding het egter nie betekenisvol van n 1:1 verhouding op die
5% vlak verskil nie. Soos in die geval van die Amsterdam-studiegebied
het die geslagsverhoudings net betrekking op volwasse en onvolwasse oor
bietjies aangesien die geslagte van baie jong bokke nie bepaal kon word
nie. In enkele gevalle was dit byvoorbeeld moeilik om tussen onvolwasse
ooie en volwasse ooie te onderskei.
Die gemiddelde groepgrootte viral vier tellings was 1,72 (reeks 1-5),
uitgesondered lammers (saam met familiegroepe of alleen), of sommige

Tabel 32. Resultate van vier seisoenale oorbietjietellings
(1974-1975) in die Piet Retief-studiegebied.
Totale telling
Totaal geseks
Rannne
Ooie
Geslagsverhouding (d : ~)
X2
(Y = 1)
SOMER
212
209
88
121
1 : 1 '38
2,62
HERFS WINTER
175 183
168 181
72 77
96 104
1 : 1 '33 1 : 1 , 35
1 , 71 2,01
~·-~·---- -------
LENTE
216
212
96
116
1:1,21
0,94
----
-X
196,5
192,5
83,25
109,25
1:1,31
1,76
----------------
I
\Jl -1>-

155
ander groepe wat nie in Tabel 33 aangetoon word nie. Die meerderheid
groepe gedurende die tellings was n volwasse ram vergesel van n volwasse
ooi (pare) wat 36,1% van al1e groepe gevorrn bet, gevo1g deur alleenlopende
ooie (20,63% van alle groepe) en rarnme, insluitend onvolwasse alleen1opende
ramme (20,18% van al1e groepe). Groepe bestaande uit n ram en twee ooie
bet gemiddeld 8,52% van a1le groepe gevorm. Ander opvallende groepe
bet uit n ram en drie ooie (2,02% van alle groepe) en groepe van twee ooie
elk (4,71% van a11e groepe) bestaan. Ramgroepe, bestaande uit twee ramme,
bet s1egs 0,45% van a1 die groepe gevorrn.
Die meerderheid oorbietjies bet in groepe van twee of meer individue voor
gekom ( 76,28io), terwyl 40,97io in pare opgemerk is-. Die .meeste alleen-
1opende bokke was ooie (20,63%) wat 0,45% meer was as die onvo1wasse en
volwasse ramme tesame (3,14% en 17,04% respektiewelik).
Seisoenale veranderinge in die groepsamestellings word in Tabel 33 aange
toon. Soos in die geval van die Amsterdam-studiegebied is al1e veranderinge
statisties getoets om te bepaal of dit betekenisvol is (Tabe1 34). Elke
oorbietjiegroep is met die res saam gedurende die vier seisoenale te11ings
verge1yk om te bepaal of die verhouding van e1ke waargeneemde oorbietjie
groep tot die ander groepe onveranderd gebly bet (Marascuilo 1966). Net
vyf verski11ende groepe is getoets omdat 'n frekwensie van vyf of meer
nodig is vir 'n Chi-kwadraattoets (Sokal en Rohlf 1969).
Volgens die resu1tate (Tabe1 34) b1yk dit dat alleenlopende ooie, pare,
'n ram en twee ooie, en twee ooie saam nie 'n betekenisvo11e seisoena1e
wisse1ing toon nie aangesien x2 7,815 moet oorskry (Stoker 1977). Slegs
die alleenlopende ramme bet 'n betekenisvo11e verandering getoon waar die
meeste gedurende die winter en lente voorgekom bet.
n Totaa1 van vier versteekte lamrners (0,9% van a11e groepe) is gedurende
die herfs gevind.
is ook opgemerk.
Algemeen
Duidelike toename in pare wa t vergese1 was van 'n lam
Alhoewel die geslagsverhouding in die Piet Retief-studiegebied meer ten

Tabel 33. Geslags- en ouderdomsgroepe van oorbietjies in die Piet Retief-studiegebied
gedurende vier seisoenale tellings vanaf November 1974 tot Oktober 1975.
GROEP- 1974
SAME STELLING SOMER N %
cf 9 8,57
~ 15 14,29
J 1 0,95
cf~ 40 38' 1
cf~~ 13 12,38
cf~~~ 6 5,71
clef 1 0,95
~~ 5 4,76
cfjcf~ 4 3,81
cf~j~ 3 2,86
~J 1 0,95
~cf 1 0,95
jcf 6 5,71
Ander - -
Totaal 105 100
J : Lam/jong bok, ongeseks Jcf : Onvolwasse lam J~ : Onvolwasse ooi N : Waarnemings
1975 HERFS
N %
16 16,33
17 17,35
3 3,06
37 37,76
8 8,16
1 1,02
- -5 5' 1
1 1,00
- -2 2,04
4 4,08
2 2,04
2 2,04
98 100
1975 1975 WINTER LENTE
N % N %
21 19,08 30 22,56
26 23,64 34 25,56
- - - -39 35,45 45 33,83
8 7,27 9 6,77
1 0,91 1 0,75
- - 1 0,75
6 5,45 5 3,76
- - 2 1 '5
2 1,82 - -1 0,91 - -1 0,91 - -3 2,73 3 2,26
2 1,82 3 2,26
110 100 133 100 ------ ----- ~------'---
GEMIDDELD N %
19 17,04
23 20,63
1 ,0 0,9
40,25 36' 1
9,5 8,52
2,25 2,02
0,5 0,45
5,25 4,71
1,75 1,57
1,25 1 '12
1,0 0,9
1 , 5 1, 35
3,5 3,14 , 1,75 1,57
111 '5 100
Ln 0'\

157
Tabel 34. Seisoenale veranderinge van spesifieke oorbietjiegroepe
in vergelyking met die ander groepe in die Piet Retief
studiegebied : November 1974 - Oktober 1975.
34. 1 I II III
Groep r:1 (17,89)a (16,7)a (18,74)a (volwasse r:!d') 9b 16b 21b
1 Nie-groep (87,11)a (81,3)a (91,26)a 96b 82b 89b
Totaal 105 98 110
x2 = 8,55 (y = 3)
a = Verwagte frekwensie b = Waargeneemde frekwensie I : Somer; II : Herfs; III : Winter; IV Lente Nie-groep : res van groepe (scm van)
34.2 I II III
Groep ~ (21,66)a (20,22)a (22,69)a (volwasse ~~) 15b 17b 26b
1 Nie-groep (83,34)a (77,78)a (87,31)a 90b 81b 84b
Totaal 105 98 110
x2 = 5,82 (y = 3)
34.3 I II III
Groep r:!~ (37,9)a (35,38)a (39,71)a
40b 37b 39b
1 Nie-groep (67,1)a (62,62)a (70,29)a 65b 61b 71b
Totaal 105 98 110
IV
(22,66)a 30b
(110,34)a 103b
133
IV
(27,43)a 34b
(105,57)a 99b
133
IV
(48,01)a 39b
(84,99)a 88b
133
x2 = 0,613 (y = 3)
N
76
370
446
N
92
354
446
N
161
285
446

158
Tabel 34 (vervolg)
34.4 I II III IV N
Groep r:!~~ (8,95)a (8,35)a (9,37)a (11,33)a 38 13b 8b 8b 9b
1 Nie-groep (96,05)a (89,65)a (100,63)a (121 ,67)a 408 92b 90b 102b 124b
Totaal 105 98 110 133 446
x 2 = 2,762 (y = 3)
34.5 I II III IV N
Groep ~~ (4,94)a (4,61)a (5,18)a (6,26)a
21 5b 5b 6b 5b
lN· (110,05)a (93,39)a (104,82)a (126,74)a 425 1e-groep 100b 93b 104b 128b
Totaal 105 98 110 133 446
x 2 = 0,4377 (y = 3)

159
gunste van ooie was (1~: 1,31 ~~) in verge1yking met die Amsterdam-studie=
gebied (1~~= 1,23 ~~),was daar in beide studiegebiede geen betekenisvo1le
verski1 van n 1:1 verhouding nie (Tabe11e 29 en 32). Min is bekend oor
die geslagsverhoudings van die ander Neotragini, alhoewel Penzhorn (197la)
en Oates (1972) gevind het dat die ges1agsverhouding van steenbokke ook
geen betekenisvo1le verski1 toon nie.
Oorbietjies kom tipies in pare of familiegroepe voor. Verskeie werkers
het reeds hierna verwys, onder andere Hediger (1951) in die eertydse
Kongo, Mason (1973) in die Jack Scott-natuurreservaat en Monfort en
Monfort (1974) in Rwanda. In beide die Amsterdam- en Piet Retief-studie
gebiede het die meeste oorbietjies in pare voorgekom (Tabelle 30 en 33),
gevo1g deur a11eenlopende ramme (Amsterdam-studiegebied) en ooie (Piet
Retief-studiegebied). Fami1iegroepe bestaande uit n ram en twee ooie
was naas die pare die belangrikste groep. Lammers is selde opgemerk
en was tot die somer (Piet Retief-studiegebied) en herfs (beide studie
gebiede) beperk. Ooigroepe het meer dikwels as ramgroepe in die twee
studiegebiede voorgekom.
Volgens Mason (1973) het die vernaamste oorbietjiegroep in die Jack Scott
natuurreservaat ook uit n ram en ooi (oorbietjiepare) bestaan wat verder
ooreenstem met Monfort en Monfort (1974) se bevindings in Rwanda. Al
hoewel Monfort en Monfort (££. cit.) ook n groot aanta1 alleenlopende
ramrne gevind het, het Mason(££. citQ) slegs enkeles aangetref. In die
eertydse Nasionale Queen Elizabeth-wildtuin (Uganda) het Gosling (1972)
gevind dat ramgroepe, pare en familiegroepe die mees algemene sosia1e
groepe is.
Enke1e groepe wat uit meer individue bestaan het as wat in Tabel1e 30 en
33 aangedui word, is ook tydens die te11ings opgemerk. Die grootste groep
het uit ses individue ( 2 ~cf 4 ~~) bestaan (Piet Retief-studiegebied) waar
van een ram en ooi onvolwasse was. In die Amsterdam-studiegebied het die
groo ts te groep ui t vyf indi vi due bes taan ( 2 ~~ 3 ~~ ; waaronder n onvo 1wasse
ram en 'n onvolwasse ooi). Grater groepe wat uit tot agt individue
bestaan het is periodiek opgemerk maar het tydens die v1ugreaksies, wat
weens die tellings veroorsaak is, in twee tot drie groepe verdeel. Hierdie

160
grater groepe kan as aggregasies (Eibl-Eibesfeldt 1970) beskou word waar
oorbietjies soms weens n omgewingsfaktor soos n kortgebrande veld of
voorbrand (kyk HOOFSTUK 11) tydelik konsentreer. In die Amsterdam-studie
gebied is n groep van twaalf oorbietjies op n land (Eragrostis curvula)
gedurende Maart (1974) waargeneem (Fig. 5 : kamp 17).
Hierdie groep bestaande uit twaalf oorbietjies, wat oor n gebied van
ongeveer 40 ha versprei was, het weens n versteuring in verskillende
rigtings gevlug. Slegs twee familiegroepe is daarna weer in dieselfde
gebied opgemerk gedurende die daaropvolgende twee dae. Roosevelt en
Heller (1914) het groepe van tien•tot twaalf oorbietjies in Oos-Afrika
opgemerk, maar maak ook melding van die feit dat hulle in kleiner groepe
verdeel het na n versteuring. Al die ander Neotragini, uitgesonderd die
klipspringer (Dunbar en Dunbar 1974, Norton 1980) blyk hoofsaaklik enkel
lewend te wees, of kom net in pare voor (Jarman 1974, Leuthold 1977).
Alhoewel die seisoenale tellings op seisoenale veranderinge in die groep
samestellings in beide studiegebiede wys, het n statistiese toets getoon
dat slegs enkele veranderinge betekenisvol is. In die Amsterdam-studie
gebied het een familiegroeptipe (1~ 2~~) die hoogste frekwensie tydens
die somer gehad wat ooreenstem met die Piet Retief-studiegebied alhoewel
dit nie n betekenisvolle verandering in laasgenoemde gebied getoon het
nie. Alleenlopende rannne het 'n betekenisvolle verskil in die Piet Retief
studiegebied getoon waar die meeste gedurende die lente getel is. Hierdie
verandering, alhoewel nie betekenisvol nie, is ook in die Amsterdam
studiegebied waargeneem. Daar kan egter geen betekenisvolle afleidings
van hierdie seisoenale verandering gemaak word nie.
SOSIALE GEDRAG
Groepsbande
Die groepsbande in n oorbietjiepaar of familiegroep is sterk terwyl daar
terselfdertyd n groat mate van koordinasie in alle aktiwiteite bestaan
(Fig. 28). Leuthold (1977) beskou koordinasie van aktiwiteite as ken
merkend van n sosiale groep terwyl Remane (1960, In: Jungius 1971) n nou verband tussen individue veral in pare "Ehe" noem.

161
Ten spyte van die sterk groepsbande loop individue van groepe bestaande
uit meer as twee oorbietjies soms vir n dag of langer alleen alboewel
bulle nog steeds in die territorium bly. Hierdie wegdwaal van indivi-
due bet veral betrekking op onvolwasse rarnme. Daar bestaan egter wel
aanduidings dat die kontak nooit heeltemal verbreek word nie weens die
relatiewe klein territoriums (kyk Territoriale Stelsel en Territorium
grootte) sodat die verskillende individue van n familiegroep mekaar
meestal kan opmerk gedurende aktiewe periodes. Olfaktoriese kommuni-
kasie vervul moontlik ook n funksie.
In die Amsterdam-studiegebied bet dieselfde volwasse ram en ooi in elk
van twee familiegroepe vir die volle duur van die veldwerk gebly. On
bekende oorbie tj ie- ooie he t soms vir kort periodes van enkele dae of
weke by hierdie bekende gro~pe aangesluit, terwyl onvolwasse ooie en
veral onvolwasse ramme die groepe periodiek verlaat bet. Dit wil dus
voorkom asof n volwasse ram en ooi die basiese permanente eenheid in
elke familiegroep vorm.
n Permanente paarbinding word in die sosiale eenbeid, wat boofsaaklik uit
'n volwasse ram met 'n ooi bestaan, gevorm. Monfort en Monfort ( 1974)
beweer dat oorbietjiepare nie permanent by mekaar bly nie maar slegs in
diese1fde omgewing. Daarenteen bet Dunbar en Dunbar (1974) opgemerk dat
klipspringers in Etbiopie nooit tydens aktiwiteite in k1einer groepe ver
dee1 raak nie.
Dominansie
Leuthold (1977) definieer dominansie as n eienskap wat die betrokke in
dividu toegang tot sekere bronne verleen ten koste van ander individue
sonder enige direkte wedywering. Aangesien territoriale gedrag in sover
dit interaksies tussen verskillende territoria1e diere nie bier van toe
passing is nie, kan dominansie binne-in die oorbietjie-familiegroep as
n absolute sosia1e bierargie beskou word waar die bierargiese status van
die betrokke individue nie deur eksterne faktore beinv1oed word nie
(Leyhausen 1965).
Soos in die geval van die k1ipspringer (Dunbar en Dunbar 1974, Norton 1980)
is sosia1e interaksies in die oorbietjiegroep tot die minimum beperk. Die

162
meerderheid interaksies vind tussen die ramme p1aas. 'n Duide1ike
dominansie is binne die sosia1e orde van die oorbietjie-fami1iegroep
waarneembaar waar die enke1e territoriale ram dominant is oor al die
ander ramme in die groep. Aan die ander kant is een van die ooie die
1eier, soos in die geval van rietbokke (Jungius 1971) en k1ipspringer
(Dunbar en Dunbar .2£.• cit.). Bewegings word hoofsaak1ik deur 'n spesi
fieke ooi bepaal wat veral duide1ik is by vlugreaksies (kyk HOOFSTUK 10
VLUGREAKSIES EN ANTIROOFDIERVERHOUDINGS). Aktiwiteite as sulks word
egter nie deur enige spesifieke individu be~nvloed nie.
Aggressiewe gedrag in die familiegroep word relatief selde waargeneem en
is feitlik uitsluitlik tot rarnrne beperk waar n dreigende houding meestal
voldoende is om die hierargie te handhaaf. Territoriale ra~~e dwing
dikwels n individue1e afstand af (gedurende n 10-uur waarnemingsperiode
het 'n territoriale ram tydens vier geleenthede individuele afstand afge
dwing.) Hediger (1949) definieer individuele afstand as "sfeer van in
vloed" om die betrokke individu. Hediger (1951) beskou die oorbietjie
dan ook .. as 'n "afstand tipe" waar fisiese kontak, behalwe tussen 'n ooi en
lam, afwesig is. Individuele afstand word afgedwing wanneer die dominan
te of territoria1e ram sy kop vinnig in die rigting van die ander ram laat
sak. Terselfdertyd word die ore in 'n vorentoe-posisie gehou terwyl die
horings ook na vore wys (Fig. 35 en 36). In sommige gevalle laat die
ondergeskikte ram ook sy kop sak as n teken van aggressie waarna die
territoria1e ram sy kop in die rigting van die ander swaai met n ver
hoogde intensiteit. Indien die ondergeskikte ram nie onmiddellik weg
beweeg nie, word di t in 'n aggressiewe houding genader. In somrnige ge
valle word die ondergeskikte ram se1fs gejaag (Fig. 37). Oorbietjie
ooie handhaaf ook individuele afstande in die sosiale groep, maar die af
dwinging daarvan word baie se1de gesien. Daar is nooit opgemerk dat n ram n individuele afstand met n ooi afdwing nie. Die aggressiewe of
dreigende houding van die ram, kop-omlaag met die ore en horings na vore
gerig (Fig. 36), stem ooreen met die houding tussen aggressiewe territoriale
rarnme. Volgens Walther (1958) kan die houding as "Horner-prasentieren"
beskryf word.
Direkte s taar is ook 'n aggressiewe houding maar is min waargeneem. 'n Op
vallende kenmerk van leende oorbietjies is dat hulle selde in so n posisie

Figuur 35 .
163
n Territoriale oorbietjieram wat individuele afstand
afdwing op 'n ondergeskikte ram.

Figuur 36.
Figuur 37.
164
n Oorbietjieram in n aggr essiewe/dr eigende houding met die
or e en horings na vor e gerig (vanaf n 35 mm kleurskyfie) .
n Territoriale oorbietjieram wat n onder skikte ram jaag
(vana£ 'n foto) .

165
1~ dat daar direk na mekaar gekyk word (Fig. 28).
Indien daar meer as een ooi in ~ familiegroep is, bestaan daar ook ~
hierargie tussen die ooie. Sosiale interaksies van hierdie aard is egter
tot ~ absolute minimum beperk. Daar is slegs in ~ enkele geval opgemerk
dat ~ ooi ~ ander jonger ooi oor ~ afstand van ongeveer 20 meter gejaag
het. Die betrokke ooie se houdings het grootliks met die van ramrne in
soortgelyke situasies ooreengestem.
Die urinering en outlasting van~ territoriale ram of die urine en feka
liee van 'n ondergeskikte ram hou moontlik direk verband met dominansie
(.kyk TERRITORIAL£ GEDRAG) en het 'n sterk visuele funksie. Die territo
riale ram dwing in alle gevalle enige ander ramme weg van ~ ooi wat uri
neer of ontlas. Sosiale kompetisie is gewoonlik verwant aan reproduk
tiewe sukses (Eibl-Eibesfeldt 1970). Ware verplasingsaktiwiteite (Tin
bergen 1940) wat by verskeie boksoorte gerapporteer is, is nooit waarge-
neem nie.
Submisiwiteit
Aggressiewe interaksies eindig gewoonlik wanneer een van die betrokke in
dividue ~ onderdanige houding inneem. In die oorbietjie se geval is ~
kop-omlaag-houding ook ~ submissiewe houding, maar dit verskil van die
aggressiewe houding deurdat die kop weggedraai word terwyl die ore na ag
ter gerig is. In alle gevalle word die neus meer gelig sodat die ramme
se horings meer na agter wys. ~ Soortgelyke submissiewe houding is deur
Walther (1974) vir somrnige gaselle beskryf, afgesien van die stert wat
by oorbietjies gelig word.
~ Submissiewe oorbietjie kan ook gaan le. Die kop word dan laag op die
grond gehou terwyl die nek gewoonlik effens krom getrek word. In een
geval is waargeneem dat ~ submissiewe ooi haar kop stadig heen en weer in
n horisontale posisie beweeg het terwyl haar ore terselfdertyd heeltemal
na agter gerig was. Jungius (1971) het ook by rietbokke opgemerk dat
'n ooi 'n submissiewe houding met haar kop op die grond ingeneem het terwyl
sy in~ le-posisie was. Volgens Leuthold (1977) kan die kop-omlaag-houding
as 'n submissiewe gedragspatroon ge:Lnterpreteer word. Burckhardt ( 1958,
In: Leuthold ££• £!!.) verrnoed dat die lehouding tydens submissiewe gedrag
sy oorsprong het by die legedrag van jong lammers.

166
Submissiewe gedrag is slegs by jong onvolwasse diere en vreemde individue
opgemerk. Ander gevalle van submissiewe houdings was waargeneem tydens
ram-ooi interaksies waar die ooi submissief was tydens sommige ontlasting
en urineringsgedragspatrone (kyk TERRITORIALE GEDRAG).
TERRITORIAL£ GEDRAG
Territoriale Stelsel en Territoriumgrootte
Verskeie definisies beskryf territorialiteit by verskillende diere onder
verskillende omstandighede (Burt 1943, Hediger 1949). Eibl-Eibesfeldt
(1970) definieer territorialiteit as enige ruimte-geassosieerde intoleran
sie, terwyl Ewer (1973) meld dat 'n territorium 'n gebied is waaruit gelyk
soortige diere uitgesluit word terwyl dit ook gelyktydig die tuisgebied
kan wees. Die tuisgebied is die gebied waardeur die dier gewoonlik
beweeg op soek na voedsel, terwyl die territorium die verdedigde deel
van die tuisgebied is (Burt 1943). Leuthold (1977) beskou ~ territo
rium as daardie deel van die tuisgebied waarvan alle ander individue met
'n gelyke sosiale status aktief uitgedryf word. Leuthold (.2£• cit.) wys
verder daarop dat aktiewe uitdrywing nie noodwendig moet plaasvind nie
maar da t afbakening of vorige ondervinding ook 'n funksie kan vervul, en
beklemtoon dat "territorium" en "tuisgebied" duidelik by die bespreking
van hoefdiere gedefinieer moet word. Aangesien die oorbietjie-territorium
en tuisgebied oorvleuel sal daar deurgaans na 'n "terri tori urn" verwys word.
Die groottes van vyf territoriums is in die Amsterdarn-studiegebied bepaal
(Fig. 38). Hierdie territoriums, wat ~ gemiddelde grootte van 34 ha ge
had het, het gedeeltelik oorvleuel. Geeneen van die bekende territoriums
het enige permanente water ingesluit nie, alhoewel 'n vleigebied in die
grootste territorium (groep A) teenwoordig was (Fig. 38). Die territorium
groottes en lokaliteite het konstant gebly gedurende die studie.
~ Opvallende kenmerk van hierdie territoriums is die teenwoordigheid van
n klipveld wat die grootste gedeelte beslaan (kyk HOOFSTUK 5 : DIE AMSTER
DAM-STUDIEGEBIED). Die belangrikste plantgemeenskappe in die gebied is
die Eulalia villosa - Hypoxis rigidula - Berkheya setifera -grasveld en
die Eulalia villosa - Pentanisia angustifolia - Setaria splendida - grasveld,
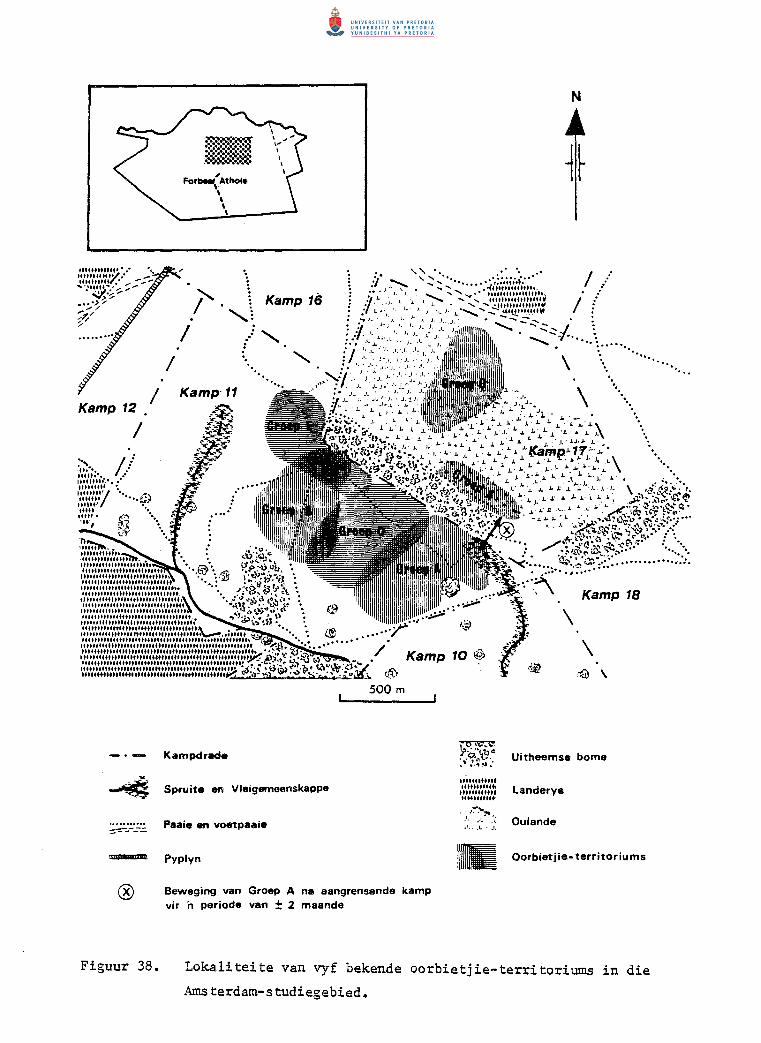
Kamp 12. I
I
~-=-----..:::.
®
Figuur 38.
I
/' I
Kamp-11
·.
167
Kamp 16
·.
Kamp 10 ~
500 m
Kampdrade
Spruite en Vleigemeenskappe
Paaie en voetpaaie
Pyplyn
Beweging van Groep A na aangrensende kamp vir h periode van ± 2 maande
ttttttUtft"t .......... ~ ............ ............ ·::.~~~-~: ·····'- . .).
N
·.
\
\ <it? \
Uitheemse bome
Landerye
Oulande
Oorbietjie- territoriums
Lokaliteite van vyf bekende oorbietjie-territoriums in die
Amsterdam-studiegebied.

168
asook die Eragrostis patentissima - Echinochloa sp.- vleigemeenskap in
Groep A se territorium (kyk HOOFSTUK 6 : DIE AMSTERDAM-STUDIEGEBIED).
In die Piet Retief-studiegebied het die twee territoriums, wat met
behulp van telemetrie bepaal is, groottes van 34,5 en 21,3 ha vanaf
September 1975 tot Januarie 1976 gehad (Fig. 39}. Geen verskille
kon in die groottes gedurende hierdie periode waargeneem word nie.
n Ooi wat saam met een van die ramme (nr. 27) in die Piet Retief-studie
gebied gevang is, is later na Suikerbosrand-natuurreservaat oorgeplaas.
Die twee ooie wat saam met die ander ram (nr. 29) opgemerk is, is nie
gevang nie. Binne twee dae na die vangs is nr. 29 met n ooi opgemerk en
in die daaropvolgende studieperiode soms met twee ooie. Nr. 27 is ge
durende die daaropvolgende periode (12 dae na die vangs) alleen opgemerk,
maar daarna saam met een volwasse ooi.
Die gebied waar die twee territoriums gelee was word gekenmerk deur n bergagtige voorkoms waar verskeie spitskruinreste bo-op die plato-uitloper
voorkom. Dagsome kom lokaal voor (kyk HOOFSTUK 5 : DIE PIET RETIEF
STUDIEGEBIED). Die belangrikste plantgemeenskappe in hierdie territoriums
is die Alloteropsis semialata - Trachypogon spicatus - Tristachya hispida
grasveld op die plato, terwyl die Diospvros whyteana - Aloe marlothii
struikveld op die spitskruinreste n opvallende kenmerk van die gebied is.
Op die dagsome kom die Xerophyta sp. - Oldenlandia herbacea - klipplaat
plantegroei voor (kyk HOOFSTUK 6 : DIE PIET RETIEF-STUDIEGEBIED). Soos
in die geval van die bekende territoriums in die Amsterdam-studiegebied,
was geen permanente water in die twee territoriums beskikbaar nie.
Die bekende oorbietjie-territoriums in beide studiegebiede was relatief
klein. Volgens Monfort en Monfort (1974) is oorbietjie-territoriums
in die Nasionale Akagera-wildtuin saam gegroepeer of versprei na gelang
van bevolkingsdigtheid. Norton (1980) het gevind dat klipspringers ook
klein territoriums in die Kaapprovinsie gehad het met gemiddelde groottes
wat van 15 ha tot 49 ha gewissel het.
Oorbietjies toon soms n voorkeur vir sekere gedeeltes van bulle territo
riurns. Klipperige gedeeltes of stande lang gras word vir skuiling benut
indien gevaar dreig (kyk HOOFSTUK 10 : VLUGREAKSIES EN ANTIROOFDIERVER-

/ \
/
/
\ ./ '/.
/
169
' N . ' ..... 4l:7\ / ....
• • • I . (~ ... ·. ··~~c27~ ,
/
~'\ c_,:od Ka~, ' . \ '
/ ' 48o
Figuur 39.
\ Kamp Ra3 /
\ /
Kampdrade
-< Spruite
/
Kontoerlyne (hoogte in m) ~~u;.~.
M:.il~ Uitheemse bome
~; 6 ~ ~ Ouland
0 Lokal iteit gevang
8 lokal iteit losgelaat
& Observasiepu nte
• Posisies m.b.v. telemetrie bepaal
Die bewegings van twee vo1wasse oorbietjieramme in die Piet
Retief-studiegebied wat met behu1p van te1emetrie bepaal is
(September 1975 tot Januarie 1976) .

170
HOUDINGS).
Omdat die oorbietjie se territorium en tuisgebied oorvleuelend is kan
dit gedeeltelik onder Leuthold (1977) se SO-tipe 3b en SO-tipe 4 terri
toriurn/tuisgebied geplaas word. Leuthold (££. cit.) meld dat die dikdik,
sommige duikers, die steenbok en klipspringer ook onder die SO-tipe 3b
geklassifiseer word, waar die ram en ooi saam deelneem aan aktiwiteite
soos territoriale merk en verdediging van die perrnanente territorium.
Die SO-tipe 4 stelsel, waaronder oorbietjies volgens Monfort en Monfort
(1974) se gegewens klassifiseer word, bestaan uit twee sosiale eenhede met
n territoriale ram en klein familiegroepe waar die twee groepe onafhank-
lik van mekaar is. Dit is nie die geval by die oorbietjie in die Amsterdam
en Piet Retief-studiegebiede nie. Volgens Leuthold (££. cit.) ·se
klassifikasie van territoriums sal die oorbietjie dus in die studiegebiede
onder die bree indeling van 'n "eksklusiewe tuisgebied vir beide geslagte"
sorteer.
Afbakening
Territoriale hoefdiere het verskeie aktiwiteite wat verband hou met die
afbakening en/of verdediging van bulle territoriums. n Verskeidenheid
stereotipe en geritualiseerde gedragspatrone kan as plaasvervangers vir
fisiese kontak gebruik word en die verskillende afbakeningsmetodes sluit
visuele en olfaktoriese tegnieke in.
Oorbietjieramme baken die hele bewegingsgebied van die betrokke familie-
groep regdeur die jaar op verskillende maniere af. Alhoewel slegs die
volwasse ram die territorium afbaken kan die blote teenwoordigheid van
een of meer ooie in 'n terri torium moontlik 'n funksie vervul by indirekte
afbakening.
S ta tie s-op ties
Volgens Hediger (1949) kan die blote rondstaan van territoriale diere
binne hulle territoriums as n tipe afbakening funksioneer - die soge-
naamde staties-optiese afbakening. Die blote teenwoordigheid van n terri toriale dier in sy terri torium toon da t die gebied "beset" is ( Kok
1975).

171
Oorbie tj ieramme s taan dikwels in 'n gespanne, kop-op houding wanneer 'n
gelyksoortige ram in die nabyheid opgernerk word. Hierdie houding gaan
dikwels oor in ander vorrne van afbakening soos preorbitale klierrnerking
of selfs aktiewe verdediging. Verskeie ander houdings of aksies van 'n
territoriale ram soos ontlasting en urinering, preorbitale klierrnerking
en selfs seksuele interaksies met ooie kan rnoontlik as sekondere staties-
optiese afbakening by die oorbietjie funksioneer. Die funksionele doel-
treffendheid van staties-optiese afbakening is beperk weens die feit
dat oorbietjies vir die grootste gedeelte van~ 24-uur periode onaktief
is (kyk HOOFSTUK 8 : BASIESE AKTIWITEITSPATROON).
Staties-optiese afbakening is by verskeie Afrika-hoefdiere beskryf,
onderrneer by die klipspringer (Norton 1980), rooibok (Schenkel 1966),
rietbok (Jungius 1971), blouwildebees (Talbot en Talbot 1963, Estes 1969)
en swartwildebees (von Richter 1972).
Preorbitale Klierrnerking
Ewer (1973) beskryf klierrnerking by soogdiere as die gebruik van gespe
sialiseerde rnotoriese gedragspatrone om cherniese stowwe op voorwerpe of
ander diere in die orngewing te plaas. Die verskillende funksies van
klierrnerking word deur Ralls (1971) beskryf, onder andere territoriale
mer king wa t Hediger ( 1949) definieer as rnerke wa t dien om 'n terri tori urn
te iden ti fiseer. Kliermerke is rnoontlik ook met seksuele aktiwiteite
geassosieer (Johnson 1973).
~ Opvallende aktiwiteit van oorbietjierarnrne is die gereelde gebruik van
die preorbitale kliere om klierafskeidings op plantegroei te plaas.
Hierdie aktiwiteit is deur beide Hediger (1951) en Gosling (1972) beskryf
en word ook deur Monfort en Monfort (1974) gemeld. Gosling (2£• cit.)
beskryf hoe oorbie tj ierarmne 'n rnerkplek kons trueer deur 'n grashalrn op 'n
gemiddelde hoogte van 55,8 ern af te byt met ~ draaibeweging van die kop
(Fig. 40). 'n Dik, swart sekresie word dan op die oorb lywende gedee 1 te
geplaas (Fig. 41) terwyl die afgebyte deel ongevreet gelaat word.
Die gerniddelde merkhoogte in die Arnsterdarn-studiegebied was 38,67 em (n = 25; S = 7,16) wat laer is as die gerniddelde hoogte deur Gosling (££. cit.)

172
Fi g uur 40 . 'n Oorbietjieram merk 'n grashalm (Hyparrhenia hir ta) met die
preorbitale klier .
Figuur 41. 'n Pr eorbi tale klierafskeiding op die punt van 'n grasha l m
(Eulalia villosa) . Die a f geby t e deel van die g rashal m i s
in die agtergr ond sigbaar .

173
gemeet. Wanneer 'n volwasse oorbietj ierarn met sy kop in 'n ui tgestrekte
horisontale posisie staan (Fig. 40) is die preorbitale klier ongeveer
14 ern laer as die gerniddelde skouerhoogte van 58 ern en is dus slegs 5,3
em hoer as die gerniddelde merkhoogte in die Amsterdam-studiegebied.
Oorbietjies raak dikwels die grashalrn vlugtig met hulle neuse aan al-
vorens dit afgebyt word. Daar is nooit opgemerk dat die sekresie ge-
vreet word nie, of dat reeds bestaande rnerkplekke weer opnuut gernerk
word nie. Gosling (££. cit.) beweer dat oorbietjies wel bestaande
rnerkplekke weer gebruik.
Monfort en Monfort (1974) verklaar dat die rnerkfrekwensie in kortgras-
veld laer is as in langgrasveld. In die Amsterdarn-studiegebied is
gesien hoe oorbietjierarnme in kortgrasveld op die grasblare van byvoor
beeld Eulalia villosa en Themeda triandra en selfs plante soos Hypoxis
rigidula rnerk nadat die distale blaargedeeltes afgebyt is (Fig. 42). Die
gemiddelde merkhoogte op grasblare was 19,33 em (n = 13; reeks 15 - 28 em;
S-= 4,76). In langgrasveld word grashalrns egter deurgaans gebruik en
die gebruik van blare in sulke veld is slegs in die geval van Hypoxis
rigidula opgemerk. Alhoewel die preorbitale klier ook by ooie aanwesig
is, is die gebruik daarvan nooit waargeneem nie.
Oorbietjierarnme rnerk regdeur die gebied waarin hulle beweeg, maar daar
is n toenarne in rnerkaktiwiteit na die kante van die territorium, of wan
neer n ander ram in die nabyheid gewaar word. Daar is nooit waargeneem
dat territoriale ramrne doelgerig na die territoriale grense loop en met
gereelde tussenposes rnerk soos Monfort en Monfort (1974) beweer nie.
Die ramme rnerk ook periodiek gedurende aktiwiteitsperiodes sonder enige
ooglopende rede terwyl dit ook geassosieer is met urinering en ontlasting
(kyk Urine en Fekaliee). As gevolg hiervan en die ander faktore wat
tot preorbitale kliermerking aanleiding kan gee (byvoorbeeld die teenwoor
digheid van ander ramme) veroorsaak da t die rnerkternpo in 'n groot rna te
wissel: van 0,21 uur tot 1,24 uur per merk, met n gerniddeld van 0,39h
(kyk AKTIWITEITSPATRONE: URINERING, ONTLASTING EN MERK). Volgens Mon
fort en Monfort (££. £i!.)rnerk oorbietjies ongeveer 16 keer per uur, maar
dit is nie duidelik of dit slegs op die aktiwiteitsperiodes betrekking
het nie.

Figuur 42.
174
n Oorbietjieram wat n grasblaar (Themeda triandra) laag merk
(vanaf n 35 mm kleurskyfie).

175
In enkele gevalle merk die territoriale ram ~ ooi in die familiegroep
deur met die preorbitale klier teen die ooi se stert te vryf nadat die
ooi ontlas en urineer het. Direk hierna loop die ooi dan met 'n swaai
beweging van die stert weg (Fig. 43). Die presiese funksie hiervan is
nie duidelik nie, maar kan moontlik dien om die paarbinding te versterk
soos in die geval van die Maxwellduiker Cephalophus maxwelli in gevangen
skap (Aeschlimann 1963, In: Leuthold 1977, Ralls 1971). Enkele ander
gevalle is by bokke gerapporteer waar die een individu die ander met
preorbitale kliere merk, byvoorbeeld die girafgasel (Leuthold 1977).
Wanneer jong ramme in dieselfde familiegroep aanwesig is, merk bulle
veel minder as die territoriale ram (10,25% van totale aantal preor
bitale kliermerke). Mykytowycz (1955, In: Eibl-Eibesfeldt 1970)
he t gevind da t 'n dominan te haas Oryc to lag us cuniciculus meer dikwels
merk as die ander individue.
"Klierwewing" soos deur Walther (1964b en 1965) vir Thomsongasel Gazella
thomsoni en Grantgasel Gazella granti beskryf waar die ram sy kop van
kant tot kant deur plantegroei swaai,is nooit opgemerk nie. Gedurende
die studie is wel waargeneem dat ~ oorbietjieram soms sy kop op en af
deur ~ graspol of ander plant beweeg (Hyparrhenia hirta en Eulalia villosa).
Daar is egter geen tekens van ~ klierafskeiding gevind nie en kon moont-
lik net ~ irritasie verlig het. By een geleentheid het ~ ram sy kop in
~ Hypoxis rigidula vir ongeveer 70 sekondes geskuur waarna dieselfde
plant twee keer met die preorbitale klier gemerk is.
Ander aktiwiteite wat dikwels by sommige hoefdiere aan territoriale ge
drag gekoppel word soos grondomdolwing is nooit waargeneem nie. Monfort
en Monfort (1974) meld dat oorbietjies soms hulle voorpote gebruik om
die grond los te woel. Hierdie gedrag kon egter met urinering en ant
lasting (kyk Urine en Fekaliee) geassosieerd gewees het.
Opsommend kan daar gemeld word dat slegs die oorbietjieramme merk en
dat verskeie omstandighede tot die merkaktiwiteit aanleiding kan gee.
Die merke het klaarblyklik nie net 'n terri toriale funksie nie, maar kan
ook dien om die gebied aan die ram "bekend" te maak. Die merkaktiwiteit

Figuur 43.
176
n Oorbietjieram merk n ooi se stert met die preorbitale
klier direk na n naso-anale kontak (vanaf n 35 mm kleur
skyfie).

178
Figuur 44 . ~ Oorbietjieram in ~ urineringsposisie .
Figuur 45. 'n Oo r bietjieram in 'n ontlastingsposisie .

177
kan moontlik ook n staties-optiese funksie ins1uit en kan onder sekere
omstandighede se1fs n verp1asingsaktiwiteit wees. So ver bekend is
die oorbietjie, uitgesonderd die basterhartebees Damaliscus lunatus
(Duncan 1975, In: Leuthold 1977) die enigste boksoort wat n merkplek
voorberei.
Urine en Fekaliee
Wanneer n dier inn sekere gebied bly beweeg sal fekaliee en urine uit
eindelik oral versprei word sodat die hele gebied later die betrokke
dier se reuk sal he (Ewer 1973). Volgens Leuthold (1977) is urinering
en ontlasting, verspreid of gelokaliseerd, die mees algemene vorm van
teukmerking, en wys verder daarop dat die funksies daarvan nie volkome
duidelik is nie. Monfort en Monfort (1974) beweer dat die gebruik van
urine en fekaliee n primitiewe vorm van merk is, terwyl Schenkel (pers.
med. In: Monfort en Monfort ££• cit.) van mening is dat urinering en
ontlasting net as n merkproses beskou kan word indien dit met n ritueel
gepaard gaan.
Die houding van oorbietjies wat urineer en/of ont1as is stereotiep.
Tydens urinering word die agterpote effens uitmekaar geplaas en terse1f
dertyd ook effens gebuig met n vergroting in die afstand tussen die
voor- en agterbene (Fig. 44). Daarna word die agterbene ingebring en
verder gebuig sodat die kop en nek boer gelig word (Fig. 45).
Oorbietjies urineer en ontlas onder verskillende omstandighede en dit
is soms gekoppel aan ander gedragspatrone. Die urinerings- en ontlas
tingstempo's verskil tussen ramme en ooie. Ramme urineer en ontlas
gemiddeld een keer elke 61 minute en ooie elke 114 minute gedurende
aktiewe periodes (kyk HOOFSTUK 8 : URINERING, ONTLASTING EN MERK).
Urinering en ontlasting kom 96,5% saam voor.
Sodra n ooi urineer of ont1as loop die ram (met sy nek in n horisontale
posisie) vinnig nader en probeer om aan die ooi se vulva te ruik. Indien
die ooi nog in die onmiddellike omgewing is loop sy in alle gevalle direk
hierna weg waarna die ram n stereotipe gedragspatroon, onderhewig aan
kleinere variasies, volg (Fig. 46, 47 en 48). Die algemene patroon
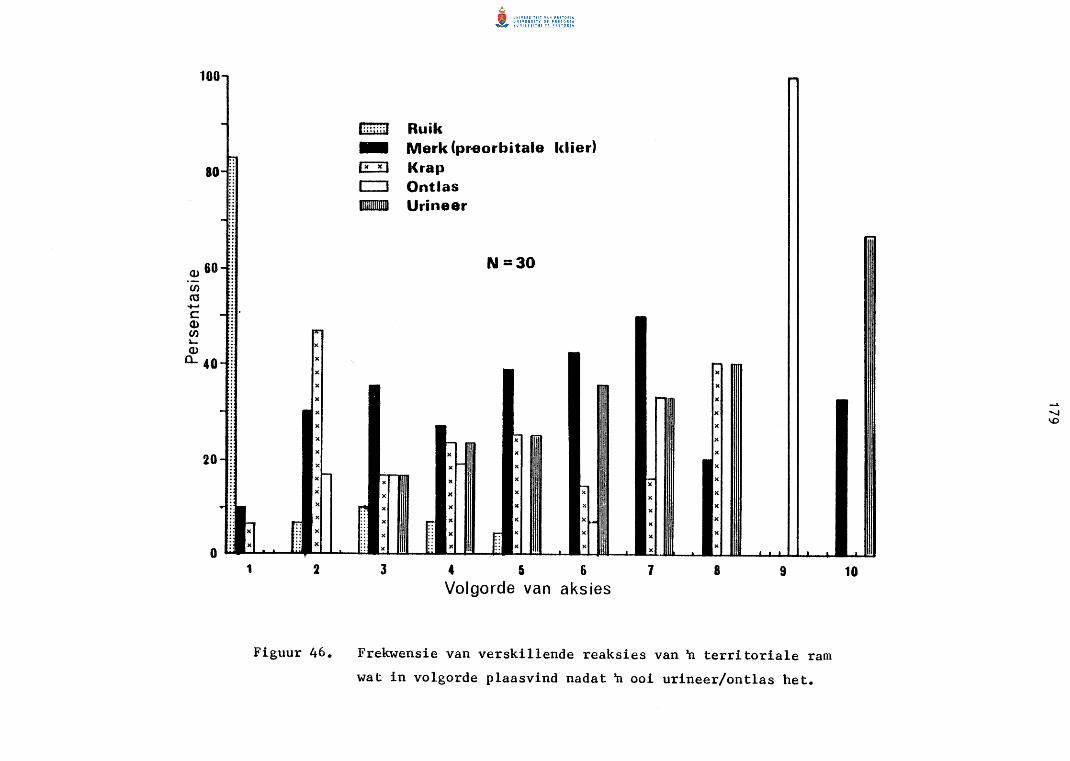
100]
ao-tl
(J) 6J en ccs +J
c -
~ Jl en :
f. L- : (J) :
Q.. 40 1 X
)(
X
-1;1 .Jx )(
X
20 -1;1 .X X
)(r-
x ~]. X
X
ru 0 ,,.~ X
X .
1 2
Figuur 46.
r ErrrJ . Ruik - Merk (pr-eorbitale ldier) [I3J Krap c::J Ontlas IBm Uri near
N=30
IC
)(
r- X
:1 IIIII II 1-
t- )( X
X l( X
xl- X X
r-.- X 7 X )( X
~ X X )( X )(
X
r.'!'! )( X IC X X X
)( r X X x .. ·X X X X
f X X
X X
X X X X XI IIIII II I . .
I I X
3 4 5 6 1 8 9 Volgorde van aksies
Frekwensie van verskillende reaksies van n territoriale ram
wat in volgorde plaasvind nadat n ooi urineer/ontlas het.
• 11m " \0
-10

180
Figuur 47 . 'n Oorbietj ieram l ek aan vars urine en fekaliee van 'n ooi.
Figuur 48 . 'n Oorbietjieram krap 'n gesamentlike miskol heftig met sy voo r pote .

181
is as volg: die ram ruik aan die ooi se urine en fekaliee, merk met
die preorbitale klier aan n grashalm, urineer en ontlas dan bo-op die
ooi se urine en fekaliee, en krap dan die kol aktief met die voorpote.
Alhoewel die verskillende aksies afgewissel word, is n basiese patroon
wel teenwoordig (Fig. 46). Inn oorbietjiepaar of familiegroep vind
hierdie gedragspatroon in 86,01% van alle gevalle plaas nadat n ooi
urineer en ontlas het. Dit kom ook voor wanneer n jonger ram in die
familiegroep urineer en ontlas.
Submissiewe ramme arriveer soms eerste by die ooi en begin dan dieselfde
basiese patroon van ruik, merk, urineer, ontlas en krap volg. Die
territoriale ram sal die miskol vinnig in n dreigende houding nader
(kyk SOSIALE GEDRAG) en dan self die merkpatroon begin volg nadat die
ander ram wegbeweeg het.
Oorbietjies van beide geslagte urineer en ontlas regdeur die gebied waar
in hulle beweeg gedurende aktiwiteitsperiodes en gee tot ewekansige mis
kolle aanleiding (Leuthold 1977), alhoewel die territoriale ram se akti
witeite in hierdie opsig hoofsaaklik deur die ooi(e) in die familiegroep
bepaal word. Gemeenskaplike miskolle wat ontstaan weens gelokaliseerde
ontlasting, word in sommige gevalle besoek. Hierdie miskolle word op
opvallende oop gedeeltes soos voetpaaie of uitgetrappe dele aangetref
(Fig. 49 en 50). Aangesien relatief min sulke miskolle in beide studie
gebiede opgemerk is, was dit moeilik om die posisies daarvan ten opsigte
van die liggings van territoriums te interpreteer, maar dit wil voorkom
asof die gemeenskaplike miskolle se verspreiding nie verband hou met
territoriale grense nie en eerder bepaal word deur geskikte oop gedeeltes
binne die territorium (Fig. 51). Oorbietjies loop nie doelgerig ver om
op hierdie kolle te urineer en ontlas nie. Slegs rarnme het gemeenskaplike
miskolle tydens die studie gebruik, en die gedragspatroon en volgorde van
aksies volg dieselfde basiese patroon soos reeds beskryf. Dit wil voor-
kom asof die oorbietjieramme slegs sulke miskolle gebruik wanneer hulle
in die onmiddellike omgewing is. Die gemeenskaplike miskolle het weens
hulle ligging moontlik ook n sterk optiese funksie.
Rooseveld en Heller (1914) het opgemerk dat oorbietjies oop gedeeltes vir
urinering en ontlasting gebruik - "accumulations of which characterise

Figuur 49. Fekaliee van n oorbietjieram bo-op die van n ooi.
se krapmerke is op die grond sigbaar.
Die ram
00 N

Figuur SO . Gesamentlike oo r bietjiemiskol in 'n voetpad (Amsterdam- studiegebied) .
(X) w
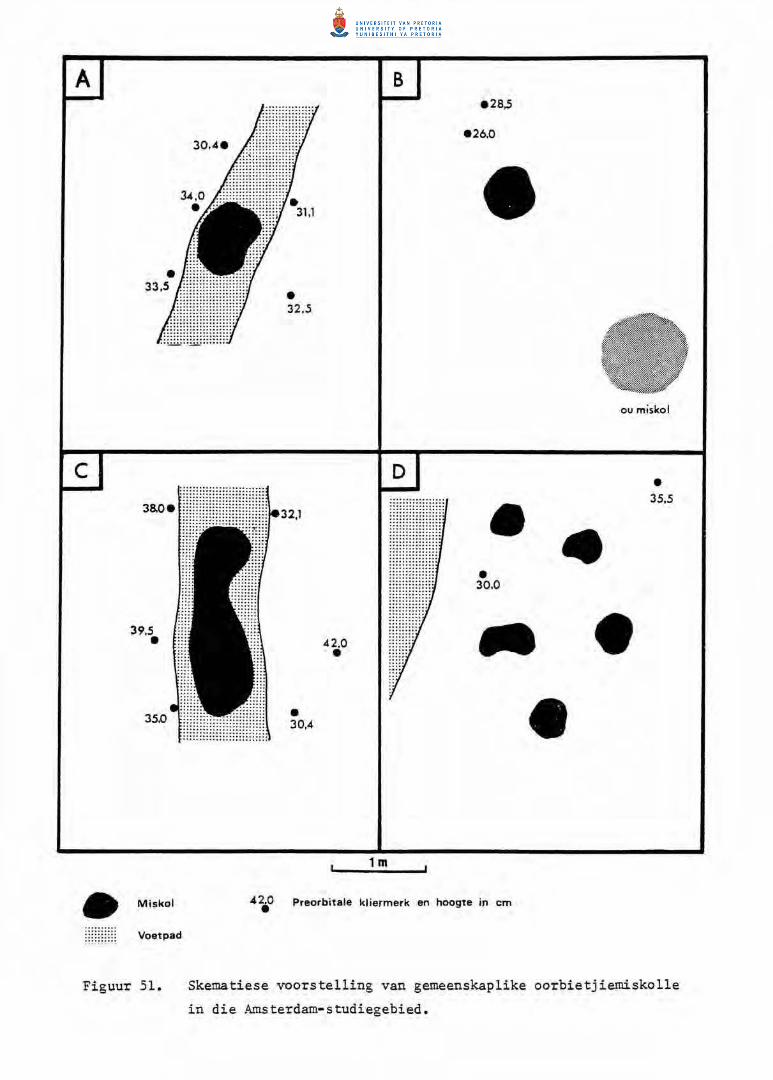
A
c
39.5 •
35.0
• Miskol
Voetpad
Figuur 51.
• 32 . .5
32,1
• 30,4
42,0 •
1m
184
B
D
~g~;~;g;gg ~ ~ .. .. ....... ..... n~ ~n~n ~ ~nn ............ ... ..... ........ .. ........... .... ::::::::::::::: ··············· ............... ············· ·· ............. .. ........ ...... ........ ...... ······· ······· .............. ;g~~g ~ g~ ~~= ............. : :: ::::::::::
e 28,5
e26.o
• • 30.0
..
42.0 Preorbitale kliermerk en hoogte in em
•
ou miskol
• 35,5
Skematiese voorstelling van gemeenskaplike oorbietjiemiskolle
in die Amsterdam- studiegebied .

185
their haunts". Tait (1969) daarenteen beweer dat oorbietjies op enige
plek ontlas en nie op spesifieke kolle nie, alhoewel Leuthold (1977)
ook meld dat gemeenskaplike miskolle gebruik word. Beide geslagte van
die dikdik urineer en ontlas op spesifieke gemeenskaplike miskolle (Tin
ley 1969). Gelokaliseerde ontlasting kom by verskeie ander boksoorte
voor, onder andere Thomsongasel (Leuthold ££• cit.) en klipspringer
(Dunbar en Dunbar 1974, Norton 1980).
Die funksie van die oorbietjieram se aktiewe krap by miskolle (Fig. 48)
is nie duidelik nie. Alhoewel interdigitale kliere (Pocock 1910) aan
wesig is, kan daar nie met sekerheid verklaar word of daar wel enige
afskeidings op hierdie manier vrygestel word nie.
Verdediging
n Opvallende en belangrike aktiwiteit vir die behoud van n territorium
is aktiewe verdediging. Aanvalsgedrag kan by diere waargeneem word wat
uitsluitlik veroorsaak word deur die verskyning van gelyksoortiges voor
dat fisiese kontak gemaak is (Eibl-Eibesfeldt 1970). Leuthold (1977)
definieer verdediging as n intraspesie intoleransie en gewoonlik n sukses
volle demonstrasie van dominansie deur die territoriale dier.
Sodra n territoriale oorbietjieram n teenstander in die onmiddellike om
gewing gewaar, neem hy n regop posisie in met die nek gestrek en kop om
laag terwyl die ore na vore gerig is. Hierdie houding funksioneer as
vertoon en gaan verdere fisiese verdediging soms vooraf tesame met preorbi
tale kliermerking.
Werklike fisiese kontak tussen twee territoriale rarnme onder natuurlike
omstandighede is tot n minimum beperk en is slegs tydens vier geleenthede
waargeneem. Die rarnme nader mekaar stadig en in gespanne houdings terwyl
die nekke in n horisontale posisie gehou word. Sodra die twee ramme na aan
mekaar is sak die koppe effens laer totdat die neuse vir n kort oomblik
teen mekaar druk. Beide ramme se ore is in n vorentoe posisie maar die
horings word steeds in n byna vertikale posisie gehou. In elke geval
het die een ram hierna vinnig weggebreek en tydens twe~ gevalle het die
een ram die ander oor n afstand van minder as 400 m in wye sirkels gevolg.

186
Die 1aasgenoemde ram het daarna verskeie kere met die preorbita1e k1ier
gemerk terwy1 die ander ver wegbeweeg het.
Daar is nooit waargeneem dat die horings en/of voorkop, soos in die geval
van baie ander boksoorte byvoorbeeld die gemsbok Oryx gazella bisa (Walther
1958) en k1ipspringer (Dunbar en Dunbar 1974), in territoria1e verdediging
gebruik word nie. Fisiese kontak was dus in e1ke waargeneemde geva1
tot~ uiterste minimum beperk (naso-nasa1e kontak). Territoria1e geveg
te van horingdraende hoefdiere is geritualiseerd en Eibl-Eibesfe1dt (1970)
wys daarop dat borings primer funksioneer tydens intraspesie konf1ikte.
Horings moet gevo1g1ik aangepas wees vir die spesifieke fisiese kontak
by die verskillende spesies. Monfort en Monfort (1974) is van mening
dat die beperkte fisiese kontak in dia oorbietjie se geva1 ~ aanpassing
is om beserings te voorkomo
In die meerderheid inter-territoria1e konf1ikte (84,6% in die Amsterdam
studiegebied) het fisiese kontak tussen die rarnme nooit plaasgevind nie.
Tydens hierdie geva11e het een van die ramme, of beide, a1tyd wegbeweeg.
Geen voka1isasie, soos byvoorbee1d by die rietbok gevind word (Jungius
1971), is tydens su1ke interaksies gehou nie.
E1ke territoria1e ram is meer aggressief naby die midde1punt van sy
territorium en is minder aggressief verder weg van die sentra1e gedee1te.
Preorbita1e k1iermerking word dikwe1s naby die territorium se randgedee1-
tes gebruik wat, soos reeds geme1d, ook ~ verp1asingsaktiwiteit in ~ kon
f1ik situasie kan wees. Manning (1970) beskryf gedrag onder hierdie
konf1iksituasies as agonistiese gedrag waar ~ kompromis tussen v1ug en
aanva1 gevind word.
Ware interaksies tydens territoria1e verdediging is tot die betrokke
territoria1e ramme beperk, maar in~ enke1e geva1 is aggressie tussen
ooie opgemerk wat ook ~ funksie by territoriale verdediging kon ver~ig
het. 'n Onbekende ooi het stadig in die omgewing van 'n territorium ver
skyn waarna een van die betrokke fami1iegroep se ooie vinnig nader ge
draf het. Beide ooie het ~ aggressiewe houding ingeneem met hu11e koppe
en nekke om1aag terwy1 hu11e stadig om mekaar beweeg het (Fig. 52).
Hierna het die onbekende ooi weggevlug met die ander ooi kort op haar

Figuur 52.
187
Twee oorbietjieooie in n aggressiewe houding (vanaf n 35 mm kleurskyfie).

188
hakke. Nadat hulle ongeveer 150 m gehardloop het, het beide ooie weer
begin loop alhoewel die onbekende ooi elke keer begin vlug het as die
invidiuele afstand korter as een meter geword het. Hierdie jaagtog het
later tot verby die territorium se randgebied gestrek waarna die bekende
ooi met n draai na die res van die familiegroep terug gehardloop het.
Die aktiewe verdediging van territoriums vind dikwels plaas tydens groot
skaalse versteurings in die gebied soos tydens veebewegings. Tydens
sulke gevalle vlug 'n terri toriale ram soms tot binne 'n aangrensende terri
torium om weer dadelik terug te beweeg wanneer die ander territoriale
ram gewaar word. In enkele gevalle het die tweede ram die ander selfs
vir verskeie meter buite sy territorium gevolg voordat hy self weer
terug beweeg het.
VOORTPLANTINGSGEDRAG EN REPRODUKSIE
Voortplantingsgedrag
Prekopulasiegedrag
Oorbietjieramme toon prekopulasiegedrag regdeur die jaar. Hierdie akti-
witeite sluit aktiewe belangstelling in urinering en outlasting van die
ooi, en met gepaardgaande preorbitale- en interdigitale kliermerking
(kyk TERRITORIAL£ GEDRAG).
Wanneer n ram die urine van n ooi op die grond of aan die ooi se vulva
ruik, word "Flehmen" (Schneider 1930, In: Leuthold 1977) vertoon. Die
tipiese houding is met die nek in n horisontale posisie of effens ho~r
terwyl die kop ook effens gelig word. Die bek word oopgehou terwyl
die lippe oopgetrek word. Volgens Leuthold (1977) kom "Flehmen" ook
by ander Neotragini-spesies soos die dikdik voor, maar Monfort en Mon
fort (1974) beweer dat "Flehmen" nooit tydens hul studie van die oor
bietjie opgemerk is nie. Die "Flehmen"-houding is in die oorbietjie
se geval wel nie so opvallend as by sommige ander hoefdiere soos byvoor-
beeld sebras (eie waarnemings) nie. Volgens die omstandighede waaronder
"Flehmen" voorkom, blyk di t 'n olfaktoriese toe ts van die ooi se reproduk
tiewe toestand te wees (Leuthold ££• cit.).

189
Alhoewel "Flehmen" nie altyd by die ruik van urine of aan die ooi, of
op die grond vertoon word nie, nader die ram altyd die ooi met n hori
sontaal-gestrekte nek tydens naso-anale kontak. In die meeste gevalle
neem die ooi n houding in wat in n mate met n submissiewe posisie oor
eenstem (kyk TERRITORIAL£ GEDRAG). Die nek word gewoonlik effens om
laag gehou met die ore in n neutrale posisie (Fig. 53) waarna die ooi
haar stert lig.
Na die naso-anale kontak loop die ooi dadelik weg waartydens die stert
heftig vir 'n kort periode beweeg word. Tydens "F1ehmen" word die ooi
nie dadelik deur die ram gevolg nie sodat die ooi dikwels n afstand
verwyder is voordat die ram weer begin vo1g, of sy ander aktiwiteite
soos wei hervat. Volgens Leuthold (1977) sal die ram slegs die ooi
vo1g wanneer laasgenoemde in of naby estrus is. In sornmige geva1le
sal die ram nie "Flehmen" vertoon nie maar wel die ooi b1y volg vir 'n
rede1ike afstand terwyl daar gepoog word om die naso-nasale kontak te
behou. Die ooi versnel dan uiteinde1ik haar stapsnelheid totdat dit
in 'n draf s tap oorgaan indien die ram b1y vo 1g. Di t kan moon t1ik 'n vorm
van "Harnfordern" (Backhaus 1961) wees waar die ram poog om urine.ring te
stimu1eer. Die ram volg die ooi in 61,1%, (n = 18) van waargeneemde
gevalle vir n sekere afstand.
Alhoewe1 nie-territoria1e ramme in die fami1iegroep sekere aspekte van
pre-kopulasiegedrag soos die ruik van urine toon, is "Flehmen" en die
doe1gerigte volg van die ooi nie opgemerk nie. Afgesien van die gestrek
te-nek-nadering en paging tot naso-anale kontak is alle ander pre-kopu1a
siegedrag tot die territoriale ram beperk.
Kopu1asie en Postkopu1asiegedrag
Kopulasie is slegs tydens twee geleenthede (gedurende Mei 1975) waargeneem.
Die territoriale ram bet een van die ooie vir ongeveer ses minute bly volgs
maar meer aktief as die gewone volging tydens urinering en ontlasting.
Hierdie gedrag stem grootliks ooreen met die "Treiben" wat deur Walther
( l964a) beskryf is. Die nek en kop van die ram was in 'n vorentoe-opwaarts
houding gestrek, terwyl die ram voortdurend probeer het om n naso-ana1e
kon tak te behou.

Figuur 53.
190
Naso-anale kontak wat kopulasie voorafgaan (vanaf n 35 mm
kleurskyfie).

191
Nadat die ooi in ~ submissiewe houding gaan staan het, bet die ram die
ooi 'n enkele keer met die voorpoot tussen die agterbene gestamp, ooreen
stemmend met die "Laufschlag" deur Walther (1958) beskryf. Na die
"Laufschlag" bet die ram twee keer probeer om die ooi te dek, maar was
beide kere onsuksesvol. Die ooi is, nadat sy weer gaan staan bet, wel
suksesvol gedek (Fig. 54) met 'n vinnige penetrasie wat slegs enkele sekon
des geduur bet terwyl die ram regop gestaan bet sander om met sy bars
op die ooi te druk. Opvallend was die neutrale kop-op houding van die
ooi tydens die penetrasie (Fig. 54) in teenstelling met die kop-nek
omlaag houding tydens die prekopulasie gedrag (Fig. 53). 'n Verdere
kopulasie waarby dieselfde individue betrokke was, is die daaropvolgende
dag opgemerk. Dit is moontlik dat verskeie kopulasies gedurende die
ooi se estrus-periode kon.plaasvind, maar Leuthold (1977) wys daarop
dat dit moeilik is om te bepaal hoeveel kopulasies kan plaasvind aangesien
dit ook gedurende die nag kan geskied.
Cade (1966) bet opgemerk dat kopulasie by oorbietjies in gevangenskap
nie kan plaasvind alvorens die ram sy kop tussen die ooi se agterbene
geplaas bet en haar van die grand afgelig het nie. Die naso-anale kon
tak, onsuksesvolle paring en "Laufschlag" kan dien om die ooi te kondi
sioneer vir kontak en om te bepaal of sy gereed is vir kopulasie (Leut
hold .£E.• cit.).
Na die suksesvolle kopulasie het die ooi gaan le terwyl die ram haar
probeer opjaag bet (Fig. 55). Die ram bet~ tipiese aggressiewe houding
ingeneem waartydens die kop omlaag gehou is met die borings vorentoe gerig.
Daarna bet die ram aan die ooi se vulva probeer ruik terwyl die ooi in 'n
submissiewe houding,waar die kop en nek effens omlaag en omgekrul gehou
is, bly le bet (Fig. 56). Na herhaalde onsuksesvolle opjaag-pogings het
die ram voortgegaan om in die onmiddellike omgewing te wei. Ander post-
kopulasie gedragspatrone soos "Laufschlag" en penislek wat soms by ander
boksoorte soos die rietbok (Jungius 1971) en Uganda kob Kobus kob (Leut
hold 1977) voorkom is nie waargeneem nie.
Soos reeds gemeld het die beskryfde kopulasie tussen ~ territoriale ram en
~ ooi van dieselfde familiegroep plaasgevind. Dit is nie duidelik of

192
Figuur 54 . Kopulasie (vanaf n 35 mm kleurskyfie).

Figuur 55.
Figuur 56.
193
Postkopulasiegedrag waar die oorbietjieram probeer om die
ooi op te jaag.
Postkopulasiegedrag : die oorbietjieram probeer om aan die
ooi se vulva te ruik terwyl sy n matige submissiewe houding
inneem.

194
slegs territoriale ramrne in hulle eie territoriums suksesvol paar nie
alhoewel die oorbietjie se territoriale stelsel wel daarop dui (kyk
TERRITORIAL£ GEDRAG). Forbes (pers. med.) het opgemerk dat n territo
riale ram n ooi inn aangrensende territorium gedek het terwyl daardie
ram buite sig was.
Relatief min is bekend oor voortplantingsgedrag by die Neotragini, waar
die enigste gegewens feitlik net oor die dikdik bekend is (Tinley 1969,
Hendrichs en Hendrichs 1971, In: Leuthold 1977). Hierdie beskrywings
van die dikdik stem ooreen met die oorbietjie se gedrag tydens die
verskillende fases van kopulasie.
Reproduksie
Draagtyd
Die draagtyd van oorbietjies word deur Von Wilhelm (1933) as 210 dae, deur
Asdell (1946) as 7 rnaande (± 210 dae) en deur le Riche (1970) as 192
dae aangegee. Na n suksesvolle kopulasie in die Amsterdam-studiegebied
(kyk Kopulasie en Postkopulasiegedrag) het die geboorte van n enkele lam
210 - 211 dae later plaasgevind (27 en 28 Mei tot 25 Desember 1975).
Besonderhede oor oorbietjiefetusse word in Tabel 35 weergegee. Sewe
oorbietjiefetusse wat gedurende die einde van Oktober 1971 by Loskopdarn
natuurreservaat versamel is nadat n aantal dragtige ooie tydens n relo
kasie dood is, het n gemiddelde massa van 1,2 kg gehad.
Min is bekend oor die algemene reproduksie van die ander kleiner boksoorte.
Mentis (1972) gee die draagtyd van n steenbok as 5~ rnaande, dikdik as
169 - 174 dae en klipspringer as 214 dae (Jennison 1927, ~: Mentis 1972).
Volgens Norton (1980) is dit onwaarskynlik dat die klipspringer se draag-
tyd meer as 6 rnaande is. Alle draagtye is egter baie lank as die dier-
groottes in ag geneem word.
Lamseisoen
Oorbietjies toon, soos verskeie ander boksoorte, n lampiek alhoewel Ansell
(1960) beweer dat geen vaste lamseisoen voorkom nie. In die Amsterdam-

Tabel 35. Besonderhede van oorbietjiefetusse
Versamel Lokaliteit Datum Massa Nr. versamel versamel (g)
* OR5 Maweni 1962-06-29 680,0
* F476 Nkawbwa 1961-07-20 129,2
* F478 Maweni 1962-08-07 43,9
* F477 Maweni 1962-06-27 71 '7
L 1 (~) Loskopdam 1971-10-30 1051 '2
L2(ct) Loskopdam 1971-10-30 1454,0
L3(~) Loskopdam 1971-10-30 1140,,5
L4(ct) Loskopdam 1971-10-30 1484,6
L5(~) Loskopdam 1971-10-31 1168' 1
L6(ct) Loskopdam 1971-10-31 908,6
L7(~) Loskopdam 1971-10-31 1238,0
* National Museum of Zimbabwe, Bulawayo
1) Volgens Ansell (1965) : HF: Lengte van agterpoot
L : Links HL: Koplengte
R : Regs FR: Voorkop/Kruislengte
DCL: Dorsale gebuigde lengte
HF 1 )
HL 1 ) L R (em) (em) (em)
9,0 11,9 12,0
5,3 5,3 5,3
3,5 3,4 3,4
4,5 4,2 4' 1
9,3 13,6 13,5
9,8 16' 1 16' 1
9,4 14,6 41 '7
9,6 16,2 16,0
9,0 14,6 14,7
8,8 13,6 13,4
9,5 14,9 14,8
FR 1 )
(em)
27,0
6,2
11 '8
13,0
30,2
33,7
31 ,6
32,6
30,0
28,5
31 ,s
DCL 1 )
(em)
29,3
23,3
16,8
22,0
46,5
52,5
48,6
51 ,5
46,6
44,6
49,5
\0 VI

196
en Piet Retief-studiegebiede is n piek in die lamseisoen waargeneem vanaf
November tot Januarie, maar volgens die waarnemings blyk dit asof die
lamseisoen effens vroeer in die laasgenoemde studiegebied is (Fig. 57).
Jong oorbietjielamrners is so laat as Mei in die Amsterdam-studiegebied
opgemerk. Die groot aantal swaar dragtige ooie gedurende die laat lente
of vroee somer is baie opvallend (Fig. 58). In beide studiegebiede val
die lampiek saam met die hoogste neerslag (Fig. 9 en 10).
Lamseisoen-gegewens van beide studiegebiede stem grootliks ooreen met
die gegewens wat deur Millar (1970) vir die Kaapprovinsie aangegee word,
alhoewel die lamseisoen klaarblyklik effens vroeer is (piek van September
tot Oktober). Tinley (1977) het pasgebore oorbietjiela~mers in die
Nasionale Gorongosa-wildtuin vanaf November tot so laat as April opgemerk.
In Zimbabwe toon oorbie tj ies 'n lampiek gedurende die begin van die reen
seisoen in Oktober, November en Desember (Thomson 1973). Volgens Norton
(1980) word n lampiek by klipspringers in die Kaapprovinsie vanaf Julie
tot September aangetref. Afgesien hiervan is min ander gegewens op hier
die stadium beskikbaar oor lampieke by die Neotragini.
Swaar dragtige ooie het n neiging getoon om met tye weg van die familie
groep te wei of te le en hierdie neiging was veral meer opvallend wan
neer daar meer as twee ooie in die familiegroep was. Leuthold (1977)
verklaar dat sulke neigings redelik algemeen by dragtige hoefdierooie
voorkom en kan moontlik die gevolg wees van die ooi se soeke na n spesi
fieke habitattipe. Alhoewel geen bewys hiervoor gevind kon word nie,
verklaar Scheepers (pers. med.) dat oorbietjielammers gewoonlik in laag
liggende gedeeltes of vleie aangetref word en dat die ooie moontlik in
hierdie gedeeltes geboorte gee. Dit wil gevolglik voorkom asof oorbiet-
jies in isolasie geboorte gee.
studiegebiede waargeneem nie.
n Geboorte is egter nooit in die twee
Le Riche (1970) beskryf die geboorte van
n oorbietjie in gevangenskap. Direk voor geboorte het die ooi gaan le,
en na geboorte het die ooi die lam sowel as die direkte omgewing gelek.
Na ongeveer vyf minute het die lam daarin geslaag om op te staan. Volgens
Le Riche (~. cit.) he t die lam 'n skouerhoog te van 330 rrnn gehad en 'n mass a
van 2,3 kg. Ongeveer drie ure na geboorte het die lam gesuip, en daarna
Scheepers, W. de V., Bergdal, Amsterdam 2375.
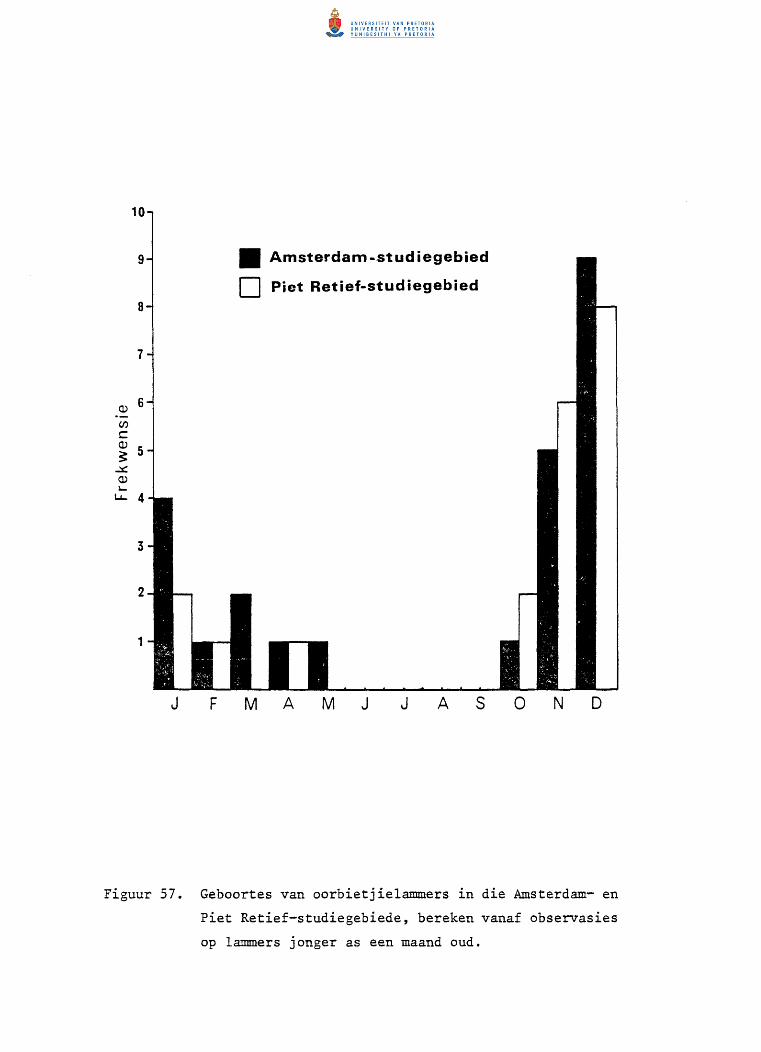
197
10
9 • Amsterdam-.studiegebied
0 Piet Retief-stud iegebied 8
7
Q) 6
(/)
c Q)
5 ~ ~ Q) 1...
w... 4
3
2
1
J F M A M J J A s 0 N D
Figuur 57. Geboortes van oorbietjielammers in die Amsterdam- en
Piet Retief-studiegebiede, bereken vanaf observasies
op lammers jonger as een maand oud.

198
Figuur 58 . Swaar dragtige oorbietjieooi gedurende die lamseisoenpiek .

199
ongeveer drie keer per dag vir periodes wat van een tot vyf minute ge
duur het.
In die Amsterdam-studiegebied is waargeneem dat oorbietjielammers na
geboorte versteek word en slegs vir kort periodes gedurende die dag te
voorskyn kom om te suip (Fig. 59). n Groot aantal hoefdiere se lammers
steek hulself weg (Leuthold 1977) wat deur Walther (1958) as "Ablieger"
beskryf is. Die wegkruipgedrag is n doelgerigte gedragspatroon van die
oorbietjielam. Leuthold (££. cit.) beweer dat hierdie wegkruip van
lammers na geboorte algemeen by nie-migrerende boksoorte voorkom wat
deur Gosling (1969) as "lying-out" gedefinieer is. Dunbar en Dunbar
(1974) en Norton (1980) het gevind dat klipspringerlamrners vir 'n lang
periode na geboorte ook nie saam met die res van die familiegroep opge
merk word nie.
Oorbietjielammers le gedurende die eerste drie tot vier maande in laag
liggende gedeel tes met 'n goeie grasbedekking of tussen ro tsblokke met
die gevolg dat hulle moeilik sigbaar is. Volgens Fick (pers. med.)
word jong lammers selfs in die ingang van erdvarkgate aangetref. Walther
(1964a) meld dat lammers in die algemeen wel op sekere stimuli reageer
soos onder andere in digte stande plantegroei. Die versteekte le van
die oorbietjielam kan as n primere verdedigingsmeganisme beskou word
(kyk HOOFSTUK 10 : Primere Verdedigingsmeganismes) waar 'n potensieHe
roofdier nie maklik die lam sal kan waarneem nie. Tydens hierdie "wegle"
van die oorbietjielam wei die ooi in wisselende afstande weg van die lam
af.
Soos reeds gemeld word die oorbietjielam slegs vir baie kort periodes
gedurende die dag tydens die eerste paar weke na geboorte opgemerk waar
tydens die ooi elke keer in die rigting van die versteekte lam gewei het,
terwyl die ram n ent verder terug gevolg het. Nadat ~ie ooi tot sowat
6 - 10 m vanaf die lam gewei het, het laasgenoemde skielik te voorskyn
gekom en direk na die betrokke ooi beweeg. Alhoewel geen vokalisasie
Fick, LoJ., Langberg, Wakkerstroom 2480.

Figuur 59 . Oorbietjielam (ongeveer een maand oud) wat aan die ooi drink.
Die anale dele van die lam word heftig ge lek.
N 0 0

201
tydens hierdie gedrag gehoor is nie, meld Leuthold (2£• cit)dat die ooie
van die meeste boksoorte onder hierdie omstandighede wel roep.
Na n vlugtige naso-anale kontak tussen die lam en ooi begin die lam dade
lik suip(Fig. 59). Die tipiese suiphouding is omgekeerd-parallel wat
die ooi terselfdertyd in staat stel om onder die lam se stert te lek
wat waarskynlik urinering en ontlasting stimuleer. Volgens Leuthold
(1977) vreet die ooie van boksoorte wat soortgelyke gedragspatrone ver
toon deurgaans die lammers se fekaliee terwyl die urine ook gedrink word.
Dit kon egter nie met sekerheid by die oorbietjie bepaal word nie. Wan
neer die ooi nie die lam gelyfkoos het nie, het sy net in n neutrale
posisie gestaan. Hierdie kontak tussen die ooi en lam was tydens alle
waargeneemde gevalle beperk tot vroegoggend (twee waarnemings), middel
van die dag (een waarneming) en laabmiddag (twee waarnemings). Dit wil
ook voorkom asof die ooi hierdie kontakperiodes bepaal en gevolglik ook
die lam se aktiwiteitspatroon.
Steurings soos voertuie laat beide die lam en ooi vlug. Lammers vertoon
deurgaans lang vlugaf~tande, en reageer op die snuif/fluit asook die
tipiese waaksame houding van die volwasse bokke (kyk HOOFSTUK 10 VLUG
REAKSIES EN ANTIROOFDIERVERHOUDINGS). Oorbietjielamrners word na onge
veer vyf rnaande gespeen wanneer die ooie n toenemende onverdraagsame
houding teenoor bulle lamrners wat wil suip, toon •
Oorbietjielarnrners is aanvanklik donker van kleur (donkerbruin in plaas
van die rooibruinkleur van die volwasse oorbietjies) en word later ligter
van kleur. Le Riche (1970) meld dat n oorbietjielam onmiddellik na
ge boor te 'n donker grys kleur gehad he t en eer s na vyf weke 'n lig ter kleur
begin vertoon het.
Die aanvanklike interaksies tussen die lam en ooi is so min as 1,89% van
die aktiewe periodes gedurende daglig (6 minute gedurende 8 ure) waarna
dit geleidelik toeneem. Die lam beweeg eers na ongeveer drie maande
voltyds saam met die familiegroep terwyl dit periodiek tot op n ouderdom
van ongeveer drie maande permanent saam met die familiegroep beweeg en
tot vyf rnaande oud probeer suip. Oorbietjielammers groei vinnig sodat

202
daar, veral in die geval van ooie, moeilik tussen n jaaroud individu
en n volwasse individu onderskei kan word. Le Riche (1970) bet gevind
dat n oorbietjielam in gevangenskap se skouerhoogte na ses weke meer as
50 em was (Die gemiddelde skouerhoogte van oorbietjie-ooie in die Ermelo
distrik is 58 em- Bylae A).

203
HOOFSTUK 10
INTERSPESIEVERHOUDINGS
Inleiding
In beide studiegebiede deel oorbietjies bulle omgew~ng met n verskeiden
heid van ander hoefdiere waaronder blesbokke en springbokke in baie
gevalle die vernaamste is, alhoewel vee ook dikwels aanwesig is (kyk
HOOFSTUK 4 : Vroee Geskiedenis en Bestuur). Estes (1967) onderskei
tussen natuurlike of toevallige assosiasies (waar veskillende spesies
b)~oorbeeld langs mekaar wei) en positiewe assosiasies (een dier van n spesie wat by n ander trap diere· aangesluit het). Leuthold (1977) maak
ook melding van negatiewe assosiasies (n assosiasie wat nadelige gevolge
vir die betrokke spesie kan inhou). Oor die algemeen is interaksies
tussen geassosieerde herbivore egter tot n minimum beperk (Estes op.
cit.). Mason (1973) wys op die min beskikbare inligting oor interaksies.
Metodes
Inligting oor vlugreaksies is hoofsaaklik ingewin tydens oorbietjie
tellings (kyk HOOFSTUK 9 : Metodes) waartydens gedrag wat op vlug
reaksies betrekking het, aangeteken is. Enkele ander aspekte van die
vluggedrag is egter later in die Amsterdam-studiegebied bestudeer,
naamlik die verhouding tussen groepgroottes wat blootstelling aan
potensiele gevaar en vlugafstande betref.
Om te bepaal of sommige groepgroottes meer blootgestel is aan naderende
gevaar tydens wei-aktiwiteite, is van n Leitz 8 x 40 verkyker en band
opnamemasjien gebruik gemaak. Die aktiwiteite van elke individu in die
verskillende studiegroepe (alleenlopende ~, oorbietjiepaar en n familie
groep van vier individue is waargeneem as kop-omlaag of kop-op tydens
bulle wei-aktiwiteite. Die resultate is later vanaf die bandopnames op
datavorms aangebring. Hierna is n t-toets op die resultate gedoen
(Sokal en Rohlf 1969) om vas te stel of daar betekenisvolle verskille
tussen die studiegroepe was wat die totale tyd wat een of meer individue
(van elke groep) aan rondkyk of kop-omlaag per waarnemingsperiode
gespandeer het.

204
Sekere vlugreaksies soos wipspronge is bestudeer deur van 16 mm rolprent
film gebruik te maak. n Paillard Bolex 16 mrn rolprentkamera is gebruik
om films teen 64 raampies/sekonde van vlugreaksies te neem. Hierdie
films is later raampie vir raampie omdersoek. Soortgelyke studies op
bewegings van sekere diere is onder andere deur Hildebrand (1963, 1964)
en Bullock (1974) gedoen.
Vlugafstande is bepaal in kortgrasveld met horisontale plantegroeidigt
heid C en grasveld met n matige plantegroeidigtheid (kyk HOOFSTUK 11 :
Metodes). n Enkele waarnemer het oorbietjie-familiegroepe te voet in
die twee verskillende veldtoestande genader. Vlugafstande, deur middel
van n Wild-afstandvinder bepaal, is geneem as die afstand waar oorbietjies
begin wegbeweeg het.
Resultate en Bespreking
ALGEMENE INTERAKSIES
Oorbietjies toon geen positiewe assosiasies met enige ander hoefdier in
die twee studiegebiede nie. Sebras en blesbokke (Amsterdam-studiegebied)
asook vaalribbokke en rooiribbokke (Piet Retief-studiegebied) is dikwels
in dieselfde omgewing opgemerk maar daar was geen tekens van enige
assosiasie nie (Fig. 60).
Interaksies met vee kom dikwels voor wanneer bees-en skaaptroppe in die
direkte omgewing van oorbietjies wei. Skaapbewegings laat oorbietjies
voortdurend van posisie verander wanneer hulle nader as gemiddeld 13,6 m
(n = 62; reeks 9 - 22 m; S = 4,28) van die skape kom. Die bokke beweeg
dan vir wisselende afstande weg. Beeste veroorsaak soortgelyke versteur
ings, maar tot n mindere mate (Fig. 61). Soms sal kalwers doelbewus na
n familiegroep oorbietjies loop en hulle opjaag (Gemiddelde afstand
totdat die oorbietjies wegbeweeg is 9,7 m; n = 24; reeks 8- 15m;
S = 3,4). Skape wei meestal in klein groepies na aan mekaar terw~l
beeste meer verspreid deur die veld beweeg, gevolglik is die moontlikheid
hoer dat n familiegroep oorbietjies meer dikwels deur skape as deur
be~ste versteur word. Die skaap- en beesbewegings lei dikwels tot
ernstige versteurings en be~nvloed sodoende aktiwiteitsperiodes (kyk

Figuur 60. Sebras wat in die direkte omgewing van n familiegroep oorbietjies
wei (~msterdam-studiegebied).
N 0 V>

Figuur 61. Beeste wat 'n versteuring veroorsaak het .
N 0 (J\

207
HOOFSTUK 8 : FAKTORE WAT AKTIWITEITE BEINVLOED) en kan gevolglik as
negatiewe interaksies beskou word.
Die enigste ander waargeneemde negatiewe interaksies, uitgesonderd
honde en mense (kyk VLUGREAKSIES EN ANTIROOFDIERVERHOUDINGS), was
weens die teenwoordigheid van n sekretarisvoel Sagittarius serpentarius.
Die betrokke sekretarisvoel bet weens sy loopbewegings n groep van
vier leende oorbietjies tydens drie geleenthede laat wegvlug wanneer dit
nader as ongeveer 10 m gekom bet.
VLUGREP~SIES EN ANTIROOFDIERVERHOUDINGS
Een van die belangrikste omgewingsfaktore wat die gedrag van hoefdiere
in die loop van ewolusie be1nvloed bet, is predasie (Leuthold 1977)~
Die voortdurende risiko om gevang te word bet gevolglik n belangrike
invloed op n hoefdier se aktiwiteitspatrone, groepgrootte en selfs
habitatvoorkeur.
Groepgrootte
Jarman (1974) het voorgestel dat die groepgrootte van n boksoort
tesame met ander faktore, die dier se gedrag teenoor roofdiere be~nvloed.
Groot groepe verminder die waarskynlikheid van n enkele individu om
roofdiere tee te kom, in teenstelling met verspreide individue, maar
terselfdertyd maak dit die waarneming van die hele groep makliker
(Brock en Riffenburgh 1960, In: Edmunds 1974). Walther (1969) bet
verklaar dat die eenvoudigste manier waarop n Thomsongasel die nadering
van n roofdier kan verhoed is deur bloat net rand te kyk. Diere wat in
n groep beweeg hoef nie konstant op die uitkyk vir gevaar te wees nie,
want in n groep is die waarskynlikheid hoog dat een lid van die groep
n roofdier sal gewaar terNYl die res met ander aktiwiteite besig is.
Oorbictjies berus uitsluitlik op bulle gesigsintuig om enige potensiele
gevaar betyds waar te neem. Hierdie feit is veral belangrik as in ag
geneem word dat oorbietjies meestal oop grasveld verkies (kyk HOOFSTUK 11:
Gevolgtrekking). Om te bepaal of dit enige voordeel vir oorbietjies
inhou om in groepe eerder as om alleenlopend voor te kom saver dit die
waarneming van gevaar betref, is n alleenlopende ram met n oorbietjiepaar

208
(d ~), en~ paar weer met~ farniliegroep bestaande uit vier individue
vergelyk.
Oorbietjies wei gerniddeld 28,64% van ~ 12-uur periode (daglig) wat die
vernaarnste aktiwiteit is (kyk HOOFSTUK 8 : WEI-AKTIWITEITE) en is gedurende
hierdie periodes aan bespeuring blootgestel terwyl bulle gesigsvermoe ook
benadeel word wanneer hulle kop-ornlaag in die grasstraturn wei. Volgens
Tabel 36 het 'n alleenlopende ram, terwyl hy gewei het, gedurende 28
minute 18 sekondes sy kop vir ~ totale periode van 22 minute 8 sekondes
ornlaag gehad (83,69% van die observasie tyd). In teenstelling hiermee
bet~ oorbietjiepaar vir 60,74% van die tyd nie opgekyk nie (een of
albei individue) wat betekenisvol verskil van die alleenlopende ram
(t = 2,427). Die hipotese dat die alleenlopende ram net so min rond
kyk as die oorbietjiepaar word gevolglik verwerp (p>0,05). Dit is ook
duidelik da t 'n farni liegroep bes taande ui t vier indi vi due meer tyd aan
rondkyk spandeer as 'n oorbietjiepaar (Tabel 36 : t = 18,828, p >0,05).
Uit hierdie resultate is dit gevolglik duidelik dat ~ groter groep
oorbie tj ies rnakliker gevaar sal kan waarneern as 'n oorbie tj iepaar of
alleenlopende individue, en oorbietjies bet dus ~ voordeel om in groter
groepe voor te kern wat die waarneming van potensiele gevaar betref.
~ Thomsongasel wei dikwels vir verskeie minute en selfs so lank as 15
minute sender om op te kyk (Walther 1969). Rooibokke kyk egter weer
dikwels rend terwyl bulle wei (Schenkel 1966).
Prirnere Verdedigingsrneganisrnes
Primere verdedigingsmeganisrnes funksioneer by diere selfs al is daar
geen roofdier in die omgewing nie. (Kruuk 1972, soos gewysig deur
Edmunds 1974). Die funksie van hierdie rneganisrne is om die waarskynlik
heid da t 'n roofdier sy prooi kan teekom, te verminder.
Oorbietjies is vir die grootste gedeelte van ~ 12-uur daglig periode
onaktief (kyk HOOFSTUK 8 : BASIESE AKTIWITEITSPATROON) waartydens die
bokke altyd le. Oorbietjies smelt diktvels saam met die agtergrond as
bulle onaktief is, nie net wat kleur in enkele gevalle betref nie (rooi
bruinkleur van die oorbietjie se haarkleed met roesbruin dolerietrotse),

Tabel 36. n Vergelyking van die tyd gespandeer deur verskillende oorbietjiegroepe
met hulle koppe omlaag gedurende n weiperiode (Amsterdam-studiegebied).
TOT ALE TOTALE TYD OBSERVASIE- MET
TYD KOP-OMLAAG
Alleenlopende 28 min. 22 min. c:f 18 sek. 08 sek.
12 min. 7 min. c:fc.t 34 sek. 38 sek.
22 min. 12 min. c:fc.tc.t Jd' 55 sek. 21 sek.
--- -~-- -~-- ---- ~~----~---
N : Totale aantal waarnemingsperiodes
S : Standaardafwyking
x % VAN TOTAAL: N s (sek.) KOP-OMLAAG
14,59 83,69 91 11 , 01
10,65 60,74 43 8,63
7,34 53,89 101 7,51
OMVANG (sek.)
1 - 71
1 - 50
1 - 51
N 0 \.0

210
maar ook met die fisiese vorm van klippe (Fig. 62 en 63). In die
Amsterdam-studiegebied (kamp 11) het 'n opname getoon dat oorbietjies
gedeeltes met klippe of graspolle benut vir skuilding indien dit
beskikbaar is (p> 0,05), of dat hulle na ~ versteuring na die meer klip
perige gedeeltes of lang grasveld van die territorium beweeg het (Fig. 63).
Wanneer oorbietjies le herkou hulle periodiek terwyl die omgewing voort
durend bespied word. Indien daar wel gevaar opgemerk word, kan ~
posisie ingeneem word met selfs die kop en nek omlaag en in somrnige
gevalle selfs op die grand uitgestrek. Hierdie houding is soortgelyk
aan die tydens 'n submissiewe houding ( kyk HOOFSTUK 9 : SOSIALE GEDRAG) o
Dit stem ooreen met die le-houdings wat byvoorbeeld deur steenbokke
duikers en rietbokke onder dieselfde omstandighede ingeneem word (Jungius
1971). Daar word dan stip in die rigting van die potensiele gevaar
gekyk. Indien die oorbietjie nie plat le nie sal dit wel in~ gespanne
regop houding le met~ gestrekte nek (Fig. 64). Hierdie houdings kan
direk in die sekondere verdedigingsmeganisme oorgaan (Fig. 65).
Sekondere Verdedigingsmeganismes
Die sekondere verdedigingsmeganismes van n dier word in werking gestel
sodra dit bewus word van n roofdier of potensiele gevaar (Edmunds 1974).
~ Hoefdier kan, nadat dit deur n roofdier opgemerk is, die volgende
reaksies toon (a) die roofaier ignoreer, (b) n waaksamer houding inneem,
(c) ontwyking toon, (d) vlug voor of na die aanval, en (e) die roofdier
dreig of selfs aanval (Walther 1969, Smythe 1970, Jarman 1974 en Leuthold
1977).
Sodra 'n aktiewe oorbietj ie potensiele gevaar sien, word 'n gespanne, regop
houding ingeneem waartydens die nek gestrek en die ore na vore geplaas
word (Fig. 66 en 67). Hierdie houding stem ooreen met die dominante
houding tydens sosiale interaksies (kyk HOOFSTUK 9 : SOSIALE GEDRAG).
In uitsonderlike gevalle word daar selfs met die voorpoot gestamp en kan
n fluit as alarmroep gebruik word. Hierna kan die oorbietjie gaan le
in die houding wat reeds beskryf is (Fig. 62 en 63) maar indien die
graad van versteuring groot genoeg is sal die oorbietjie vlug, maar nie
noodwendig dadelik hardloop nie. ~ Gallopbeweging volg wanneer daar

Fi guur 62 .
211
Primere verdedigingsmeganisme by 'n jong oorbietj ieooi waar sy
agter ~ graspol verskuil le.
Figuur 63 . 'n Oorbie tjie -ooi skuil tussen klippe na 'n ver s t euring .

212
Figuur 64. n Oorbietjieram le in n waaksame/onsekere houding (nek gestrek,
en ore na agter gedraai).
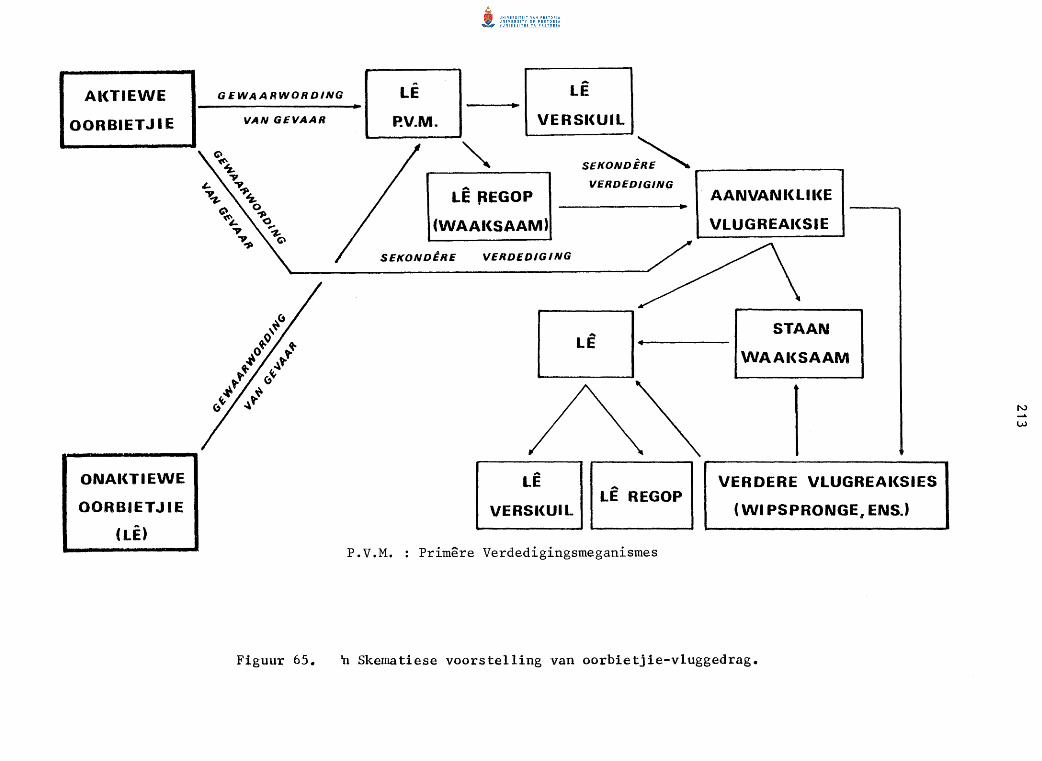
AI<TIEWE
OORBIET J IE
ONAI<TIEWE
OORBIETJIE
(LE)
GEWAARWORDING
VAN GEVAAR
Q~
" L ,..,. .,~
.z. Q('C\
L ., 0~
o_ ..,.(;\
.,")\
~(J o' ~ ~0 tt
h.t ~t t" u~ ~ ~
(J~ ~'t
LE
P.V.M.
LE VERSI<UIL
"' LE PEGOP
(WAAI<SAAM)
SEKONDi~ r-----------VERDEDIGING
.. AANVANI<LII<E
VLUGREAI<SIE
SEKONDERE VERDEDIGING
P.V.M.
~ J STAAN
L.:_j I WAAKSAAM I
LE VERSI<Uil
~ LE REGOP
Primere Verdedigingsmeganismes
I VERDERE VLUGREAI<SIES
(WI PSPRONGE, ENSJ
Figuur 65. u Skernatiese voorstelling van oorbietjie-vluggedrag.
N ~
w

214
1
4 5 6
Figuur 66 . Normale loophouding van n oorbietjie (vanaf n 16 mm rolfilm).
Figuur 67.
3 4
Loophouding van n oorbietjie nadat potensiel e gevaar opgemerk
is (vanaf n 16 mm ro1fi lm) .

215
met spronge vorentoe beweeg word waartydens die agterpote verby die
voorpote beweeg. ~ Baie kenmerkende sprang wat tydens vlugreaksies
opgemerk word is die sogenaarnde wipsprong ("stotting"). Die oorbietjie
spring regop in die lug terwyl die voorpote en agterpote teen mekaar
gehou word (Fig. 68 en 69) met die nek gestrek en stert in ~ horison-
tale posisie. Met elke wipsprong kan die withare van die binneboude
duidelik gesien word wat ~ skerp kontras met die swart stertjie vorm.
Die wipspronge word gewoonlik uitgevoer net voor of nadat teen volspoed
gevlug word.
Soortgelyke wipspronge word by ~ aantal boksoorte aangetref, onder andere by
al die Gazellinae (Walther 1964b), rooihartebeeste (Estes en Goddard 1967,
Kok 1975) en selfs gaffelbokke Antilocapra americana (Kitchen 1974). Leuthold
(1977) wys daarop dat wipspronge by meer sosiale boksoorte voorkorn en
~ seinfunksie kan vervul, maar die presiese funksie van wipspronge was
tot dusver die onderwerp van heelwat bespreking. Walther (1969)
beweer dat wipspronge 'n roofdier tydens agtervolging kan verwar en rnoontlik
'n alannfunksie het, terwyl Smythe (1970) vermoed dat dit 'n roofdier tot
~ aanval kan uitlok onder toestande wat gunstig vir die bok is. Stein-
hardt (1921, ~: Walther 1969) meen dat wipspronge help met optiese
orientering by die springbok. In die lig van hierdie menings en waar-
nemings op oorhietjie-vlugreaksies, kan die wipspronge (a) ~ alarmfunksie ver
rig of (b) die potensiele gevaar uitlok sodat daar nie onnodig ver hoef
gevlug te word nie, en indien di t wel tot 'n aanval oorgaan di t onder die
gunstigste toestande vir die oorbietjie sal geskied.
Indien die gevaar baie naby kom sal die oorbietjie met n vaart wegvlug
om op die aanvanklike speed staat te maak. Lang, plat spronge word
dan gegee.
ver nie.
Oorbietjies vlug selde in ~ reguit lyn (Fig. 70) en nooit
Die bokke sal dikwels weer gaan staan om in die rigting van
die potensiele gevaar te kyk, en dan weer verder te vlug of te gaan le
(Fig. 63). Oorbietjies sal binne ~ afsienbare periode weer na
bulle territoriums terugbeweeg indien bulle buite die gebiede gevlug
het.
Terwyl 'n rondloperhond 'n oorbietjie-ooi gejaag het, is opgemerk dat sy
dikwels skerp van rigting verander het. Die vlugreaksie is telkens met

Figuur 68.
216
9
Vlugbewegings van n oorbietjie-ooi wat in n wipsprong eindig
(vanaf n 16 mm rolfilm).

217
Figuur 69 . n Oorbietjie- ooi in die laaste fase van n wipspr ong .

....,._ . -, vlug verder butte sig
h •
2
'\ ,- - n rf~ - ... I \ ' ;;: ' I \ '
I I I \ I t \
!;! ~ ~ staan, 1 'J/ kyk terug :
' wipspronge I I I I
I I I I I
I I
I J
I I I
II\ I I I \ /
I I \ ~
• I I I.~
~ ~ ~ ~~ cf
v~
vlug buite sig
\ ~ c!----\
st••n ,, ' , \ en kyk \ \
1
wind
\ \ I \ I
I I I I I I f
J I
I
\
' I I
wipspronge I I
I I
I I
I I
• Klippe en rotse
218
~v
20m
vlug buite sig cf/ / . ~,( ./ •
1 wtpspronge 1.1. I
~~~ 1 staan, 1 kyk terug
I I I I l
' I I I I \
I \ I \ .~
\ • I \
cfv~ • q
~ _____ ..,._ _ . / -....: ~ ,;,
\ .. \
wipspronge
' ' ~ "staan en kyk
I I
' \ '
I I
I , , ~
~
', . ' ' .... ~~
• Vers teuring
•
Fi guur 70 . Skematiese voorstelling van oor bi etjie- vlugr eaksies .

219
~ paar wipspronge afgesluit. Oorbietjies toon soms n neiging om nader
aan 'n vreemde voorwerp of selfs potensiele gevaar te beweeg, moontlik
om meer inligting te bekom ("information behaviour" : Walther 1969).
n Gespanne en regop houding is kenmerkend van su1ke situasies terwyl
die stert prominent in ~ horisontale posisie gehou word. Soms vlug die
bok n paar meter terug om dan weer nader te beweeg. In a11e waargeneem
de geval1e het slegs een individu gereageer terwyl die res bloat in n
waaksame houding gestaan bet.
Ooie neem die 1eiding in die meerderheid vlugreaksies deur bloat voor
te hardloop (p > 0,05; Fig 71). Die reaksies van ooie met lammers
word onder voortplantingsgedrag bespreek (kyk HOOFSTUK 9 : VOORTPLANTINGS
GEDRAG EN REPRODUKSIE).
Vlugafstande
Hoefdiere v1ug nie dadelik as gevaar gesien word nie, maar wag totdat
die naderende gevaar binne 'n sekere afstand, die sogenaamde vlugafstand,
beweeg het (Hediger 1934, ~: Walther 1969). Volgens Hediger (££. cit.)
is die vlugafstand verskillend en tipies vir elke spesie. fiierdie feit
is deur verskillende werkers aangetoon vir ~ verskeidenheid van spesies,
onder andere by die Amerikaanse eland Alces alces en elk Cervus canadensis
(Altman 1958), Thomsongasel (Walther 1969), blesbok (Rowe-Rowe 1979) en
rooihartebees (Kok 1975).
Vir die doel van hierdie studie is die vlugafstand geneem as daardie
af stand waar oorbie tj ies 'n vlugreaksie begin toon he t deur weg te beweeg
(sekondere verdedigingsmeganisme).
Die vlugafstande van oorbietjies op bewegende persone in die veld wissel
na gelang van verskeie toestande maar die belangrikste blyk die habitat
self te wees. Fig.72 toon dat oorbietjies in die Amsterdam-studiegebied
gemiddeld n grater v1ugafstand in kort grasveld met min skuiling (X = 143,15 m; n = 18; omvang 98 - 236m, S = 45,21) het in verge1yking met lang
grasve1d of ~ gebied waar addisionele skuiling soos klippe of rotsblokke
(Fig. 63) beskikbaar is. (X= 98,55 m; n = 21; omvang 38 ~ 167,9 m;
S = 37,11) gebied (Fig. 72). Gedurende die telemetriese studie op die

Fi guur 71. Oorbie tjie - ooi e neem die l e iding in di e mees t e v lugr eaks i es .
N N 0

N =21
l(ort, gebrande veld I L. ___ > -·---------u-- --~
N =18
Langgrasveld
l__-------~~ _____ _ __ l__ I I I I
0 50
Figuur 72.
100 150 200
Vlugafstand ( m)
Horisontale lyn a omvang
Vertil<ale lyn : geaniddeld
Ball< : standaardafwyl<ing
Vlugafstande van oorbietjies (familiegroepe) in die Amsterdam
studiegebied.
250 N N

222
bewegings van twee oorbietjieramme (kyk HOOFSTUK 9 TERRITORIALE GEDRAG)
is gevind dat die gemiddelde vlugafstand van 46,5 rn na 62 m verander
het nadat ~ veldbrand voorgekom het.
Topografie het ook ~ belangrike invloed op vlugafstande. 'n Golwende
of bergagtige landskap kan bokke soos oorbietjies, wat hoofsaaklik op
visuele waarnemings staatmaak om gevaar waar te neem, in ~ groot mate
befnvloed. Wanneer 'n glooiing byvoorbee ld die oorbie tj ie van 'n
bewegende persoon skei, sal dit eers die persoon sien sodra daar geen
fisiese versperring meer is nie. In teenstelling hiermee kan n oor
bietjie ~ persoon reeds verskeie honderde meter ver aan die oorkant van
~ vallei waarneem.
Weersomstandighede.het klaarblyklik ook ~ invloed. Op reenerige dae
is bokke minder geneig om te vlug. Kok (1975) verklaar dat rooiharte
beeste ook op ~ soortgelyke manier be~nvloed word. Reuk dra ook nie
so goed in vogtige lug nie (Darling 1969), maar dit word betwyfel of
reuk enigsins ~ belangrike funksie hier vervul.
Vlugafstande is van groot belang by dryftellings. Indien die afstande
tussen die waarnemers nie klein genoeg is nie, sal verskeie individue
nooit opgejaag word nie.
Akkurate inligting oor die spoed waartoe oorbietjies in staat is, is
nie beskikbaar nie, maar gedurende ~ vangspoging het n volwasse ooi
tot so vinnig as 60 km/uur oor 'n kort afstand gehardloop (volgens 'n
voertuig se snelheidsmeter.

223
HOOFSTUK 11
HABITATVOORKEURE
Inleiding
Alhoewel slegs enkele hoefdiere gespesialiseerd is wat hulle habitat
betref, selekteer die meeste soorte tog n habitat met bepaalde eienskappe
en kom gewoonlik nie in gebiede voor waar hierdie eienskappe afwesig is
nie (Leuthold 1977). Jarman (1974) meld die moontlike verwantskappe tussen
die tipe habitat waarin n boksoort voorkom en die spesie se grootte,
voedselvoorkeure, antiroofdiergedrag en sosiale organisasie. Leuthold
(££• £!!.) wys ook daarop dat verskeie spesies in verskillende habitatte
gedurende verskillende tye van die jaar voorkom. So kan bewegings op n klein skaal tussen verskillende plantegroeitipes plaasvind of grootskaalse
migrasies kart oor lang afstande voorkom.
Groot gedeeltes van die Amsterdam- en Piet Retief-studiegebiede word deur
verskillende aspekte van menslike besetting en aktiwiteite beslaan waar
veral plantasies en landerye kenmerkend van die gebied is. Verder kom
n verskeidenheid van ander boerderyaktiwiteite voor wat nie alleen tussen
die verskillende studiegebiede wissel nie, maar ook in baie gevalle tussen
aangrensende kampe verskil. Verskille in die topografie en plantegroei
kom ook voor en daar kan gevolglik verwag word dat nie alleen die boerdery
aktiwiteite nie, maar ook die variasie in topografie en plantegroei moontlik
n invloed op die oorbietjie se gedrag en verspreiding sal uitoefen.
Die doel van hierdie ondersoek was om eerstens die oorbietjie se versprei
ding in beide studiegebiede te bepaal en vas te stel waar die hoogste
konsentrasies voorkom. Tweedens is probeer om die faktore wat die ver-
sprciding van oorbietjies be1nvloed te bepaal, asook die eienskappe van die
voorkeurgebiede. Die moontlike invloed van faktore soos menslike aktiwi
teite, geomorfologie, plantgemeenskappe en horisontale plantdigtheid is vir
hierdie doel ondersoek. Met die horisontale plantdigtheid word die mate

224
waarin die plantegroei die dier se uitsig beinvloed bedoel en nie die
aantal individue per eenheidsoppervlakte nie. Derdens is die moontlike
seisoenale wisseling in voorkeurgebiede ondersoek deur gegewens op n seisoenale basis in te win.
Metodes
KARTERING
Die bestaande 1:50 000 Topo Kaarte en lugfoto's alleen was nie akkuraat
genoeg vir hierdie studie nie aangesien plantasies, landerye, kampdrade,
hutte en paaie nie volledig op die kaarte aangedui is nie en veral nie
in die geval van die Amsterdam-studiegebied nie. Hierdie studiegebied is
gedurende 1974 met infrarooi film op n skaal van ongeveer 1:10 000 gefoto
grafeer. n Finale kaart is vanaf die 58 infrarooifoto's tesame met
bykomende inlig ting vanaf 'n 1: 50 000 Topo Kaart ( 2 630 DA, S taa tsdrukker,
Pretoria) en aanvullende veldwaarnemings opgestel.
Die Piet Retief-studiegebied is gekarteer met behulp van die bestaande
lugfoto's (Taak nr. 51536 van 1964) en n 1:50 000 Topo Kaart (2730 BA,
Staatsdrukker, Pretoria). Inligting oor onder andere kampdrade, plantasies
en butte is op n vergrote kaart (1:20 000) aangebring waar die presiese
posisies deur middel van triangulasie bepaal is.
is vir hierdie doel gebruik.
n Prismatiese kompas
Die voltooide kaarte van beide studiegebiede is tot op n presiese skaal
van 1:10 000 vergroot waarna alle oppervlaktes van kampe, lander1e en
plantasies deur middel van n Haff 317 planimeter bepaal is deur die
gemiddelde van drie lesings in elke geval te neem.
OORBIETJIEDIGTHEDE
Inligting oor die digtheid en verspreiding van oorbietjies is bepaal tydens
vier seisoenale tellings (Somer : Desember 1974; Herfs : April 1975; Winter
Augustus 1975; Lente : Oktober 1975) wat in beide studiegebiede gedoen

225
is (kyk HOOFSTUK 9 : Metodes). Alle oorbietjies wat gedurende hierdie
tellings opgemerk is, se posisies is op n kaart (1 : 50 000) waarop n 1 km2
ruitpatroon op skaal aangebring is, aangeteken. Die vaste oor
bietjieverspreidingsgebied in beide studiegebiede is na afloop van al die
seisoenale tellings op kaarte omgrens. Die oorbietjiebiomassas is bereken
na aanleiding van n gemiddelde oorbietjiemassa van 14 kg (kyk HOOFSTUK 2 :
Morfologie).
VELDTOESTAND
Direk na elke seisoenale oorbietjietelling is die veldtoestand van elke
kamp aangeteken. Die veldtoestand is ge-evalueer deur n opname van die
horisontale plantdigtheid te maak. Verskeie werkers het in die verlede
al van n digtheidsbord gebruik gemaak om horisontale plantdigtheid te
evalueer soos onder ander Wight (1938 In: De Vos en Mosby 1969) en
Wilson (1975). DeVos en Mosby (1969) meld egter die beperkinge van
hierdie metode en wys daarop dat dit slegs n aanduiding van digtheid gee.
n Metode, waar van foto's en digtheidsindekse gebruik gemaak word, is
deur Thomson (1975) gebruik om laterale bedekkingsdigtheid te kwantifiseer.
Hierdie metodes is hoofsaaklik van waarde wat die houtagtige plantegroei
(struike en borne) betref.
Om die toestand of horisontale plantdigtheid van die grasveldgedeeltes van
beide studiegebiede te evalueer is n spesiale digtheidsbord vervaardig.
Op n 1 x 1,5 m bord is n ooreenliggende ruit geverf met n blokgrootte van
5 cm2
• Die digtheidsbord is regop in die grasveld geplaas en op n afstand
en vanaf n hoogte van een meter met behulp van n 35 mm reflekskamera (28 rnm
lens) en swart en wit film gefotografeer. Die bord is subjektief op ver
teenwoordigende plekke in elke kamp geplaas ten einde n beeld van die
horisontale plantdigtheid te verkry.
Drie hoof horisontale plantdigtheidsklasse is onderskei:
(A) Hoe horisontale plantdigtheid
(B) Matige horisontale plantdigtheid
(C) Lae horisontale plantdigtheid

226
'n Kamp se horisontale plantdigtheid is as homogeen beskou wanneer meer
as 80% van die kampoppervlakte deur een horisontale plantdigtheidsklas
beslaan is, terwyl 'n heterogene toestand beskou is as 'n geval waar
minder as 80% van die kampoppervlakte deur een horisontale plantdigtheids-
klas bedek is. Fig. 73 en 74 illustreer die verskillende horisontale
plantdigtheidsklasse.
Resultate en Bespreking
AMSTERDAM-STUDIEGEBIED
Oorbietjieverspreiding en Voorkeurgebiede
Alhoewel oorbietjies feitlik regdeur die Amsterdam-studiegebied voorkom,
is die verspreiding van die oorbietjies tog in n groat mate gelokaliseerd
(Fig. 75). Die totale oppervlakte van die vaste oorbietjieverspreidings
gebied is 8,9 km2 wat 15,0% van die totale oppervlakte van die studie-
gebied uitmaak (Fig. 75). Gebiede waar klein groepies oorbietjies on
gereeld waargeneem is, is nie by die berekening van die vaste verspreidings
gebied se oppervlakte ingesluit nie.
n Gemiddelde oorbietjiedigtheid van 2,13 oorbietjies/km2 is in die Amster
dam-studiegebied gedurende die vier seisoenale tellings gevind. Die
hoogste gemiddelde oorbietjiedigtheid (11,25 oorbietjies/km2) het op die
noordelike gedeelte van die sentrale plato-uitloper (Fig. 75) in die
Eragrostis racemosa-Eriosema pauciflorum - Heteropogon contortis - gras
veld en die Andropogon eucomus - Helichrysum sp. - oulandgemeenskap voorge
kom. Ander hoe digthede ( > 4 oorbietjiesfkm2
) is op die res van die
sentrale plato-uitloper met verspreide dagsome en die suidwestelike
plato-uitloper met die platoberggebied aangetref. Die vernaamste plant-
gemeenskappe in hierdie gebied, afgesien van die reeds genoemde o~ee
gemeenskappe, is die Berkheya setifera - Vernonia natalensis - Eulalia
villosa - grasveld en die Hyparrhenia hirta - Eragrostis racemosa -
Cyperaceae - grasveld op die sentrale plato-uitloper, en die Trachypogon
spicatus - Alloteropsis semialata - Monocymbium ceresiiforme - grasveld

Hoe horisontale plantdigthe id
Figuur 73 .
Matige horisontale plantdigtheid. Lae horisontale plantdigtheid
Verskillende horisonta le plantdigtheidsklasse soos deur n digtheidsbord aangetoon.
N N .....,

Hoe horisontale plantdigtheid
Figuur 74.
Matige hori sontale plantdigtheid Lae hori sontale plantdigtheid
Voorbeelde van die verskillende horisontale plantdig theidsklasse
N N ex:>
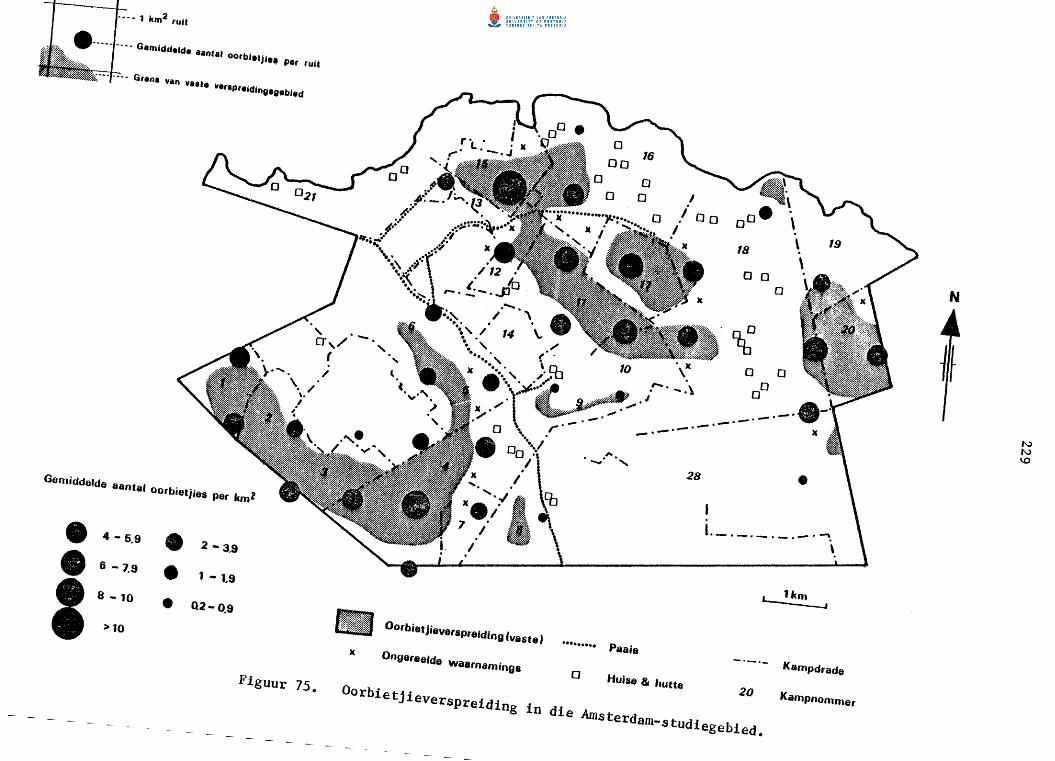
~-- 1 km2 ruit
·-----~---- Gamtddelda aontat •••btatJtao .,., ••it -·- Grena van vaaae verapreidingegebled
Gerniddelde aantal oorbietjies per krnl
• • a 4-5.9
6 -1.9
8 -10
>10
e • •
2-3.9
1 -1,9
Q2-0.9
' ·-o·,/ ·, (
' /
• Oorbietjieversprefding lvaste)
)(
Ongereelde waarnamings
_q/~# .r-. -.\ ~ _,·'
18
0 0 0
\: 0 0
00
-·- .-·-·-·-·-·-._.~.,
28 • l._ ·-·- ·- .-.-.
\
1 l<rn L... __,
Paaie
0 Kampdrade
Figuur 75. Huise & hutte
Oorbietjieverspreiding in die Amsterdam-studiegebied, 20 Kampnornmer
N
1/f
N N \0

230
op die suidwestelike plato-uitloper.
Met die uitsondering van n gedeelte van die Tristachya hispida - Berkheya
setifera - Panicum natalense - grasveld en die Riviervallei - gemeenskap
op n gedeelte van die pediment (kamp 4), het die res van die studiegebied
n gemiddelde oorbietjiedigtheid van minder as 4 oorbietjies/km2 gehad
(Fig. 75). Die gedeeltes met lae oorbietjiedigthede sluit die grootste
gedeeltes van die pediment en tallushange met die Tristachya hispida -
Berkheya setifera - Panicum natalense - grasveld~die Loudetia simplex
Indigofera sp. - Berkheya setifera - grasveld en gedeeltes van die
Hyparrhenia hirta - Eragrostis racemosa - Cyperaceae - grasveld in.
Geen oorbietjies is gedurende die tellings in plantasies of stande uit
heemse borne opgemerk nie. Behalwe in die geval van die aangeplante
Eragrostis curvula - land (kamp 17) is ook geen oorbietjies in die landerye
opgemerk nie. Afgesien van uitgebreide landerye op die pediment-gedeeltes
van die studiegebied, word die grootste gedeelte van die westelike plato
uitloper ook deur landerye beslaan (Fig. 75). Oorbietjies was ook afwesig
in die direkte omgewing van woonhuise en butte.
Die verspreide voorkoms van plantasies, landerye en bewoonde gedeeltes is
waarskynlik die vernaa~ste oorsaak vir die gefragmenteerde oorbietjie-
verspreidingsgebied. Die oorbietjieverspreiding word ook klaarblyklik
meer direk deur die uitgebreide plantasies, landerye en bewoonde gedeeltes
beinvloed as deur spesifieke plantgemeenskappe. Oorbietjies kom hoof-
saaklik op die plato-uitloper voor waar ook die minste versteuring gevind
word.
Seisoenale Veranderinge in Oorbietjiedigthede
Die seisoenale veranderinge in die oorbietjiedigthede volgens die
seisoentellings (November 1974 tot Oktober 1975) word in Tabel 37 aangetoon.
Besonderhede van veekonsentrasies, veldbrand en veldtoestand (horisontale
plantdigtheid) gedurende die studieperiode word in Tabel 38 uiteengesit.
Bylae B toon die oppervlaktes van die verskillende kampe aan asook die
gebiede wat deur geomorfologiese eenhede, plantasies, landerye, menslike
besetting en plantgemeenskappe beslaan word.
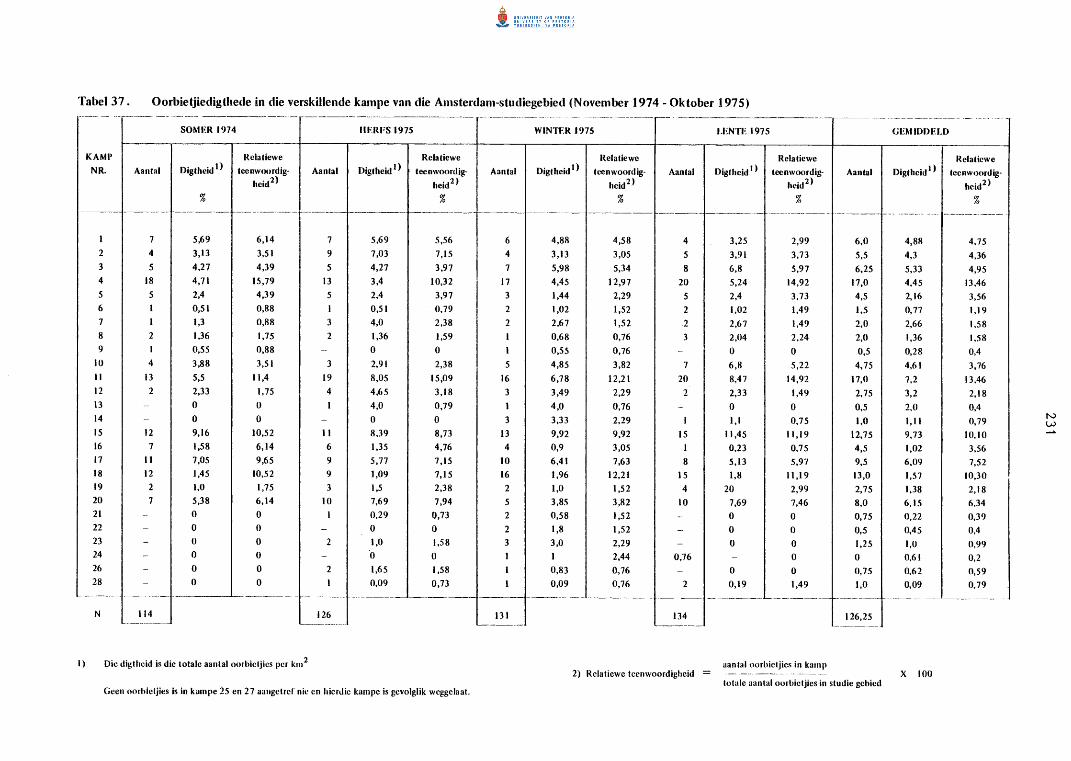
Tabel37. Oorbietjiedigthede in die verskillende kampe van die Amsterdam-studiegebied (November 1974- Oktober 1975)
I)
---- ---·----- -------------- ------------------------- ------------------------- ------------- r----- ----------------------
KAMI,
NR.
I
2 3
4
5
6
7
8
9
10
II
12
13
14
15
16
17
18
19
20
21
22 23
24
26
28
N
Aantal
-----
7
4
5
18
5
I
I
2
I
4
13
2
--
-
12
7
I 1
12
2
7
--
-
-
-
-
-----------
114 ----------
SOMER 1974
Rclatiewe
Digtheid 1) tecnwoordig-hcid2)
Aantal
% ------- -~--------- -----------
5,69 6,14 7
3,13 3,51 9
4,27 4,39 5 4,71 15,79 13
2,4 4,39 5
0,51 0,88 I
1,3 0,88 3
1,36 1,75 2
0,55 0,88 --
3,88 3,51 3
5,5 11,4 19
2,33 1,75 4
0 0 I
0 0 -
9,16 10,52 11 1,58 6,14 6
7,05 9,65 9
1,45 10,52 9
1,0 1,75 3
5,38 6,14 10
0 0 I 0 0 --
0 0 2 0 0 -
0 0 2
0 0 I -------- ----
126 -------
Die digtheid is die totale aantal oorbietjics per km 2
IIERFS 1975
Rclaticwe
Digtheid 1) tecnwoordig-
heid2 )
% -------- r---------
5,69 5,56
7,03 7,15
4,27 3,97
3,4 10,32
2,4 3,97
0,51 0,79
4,0 2,38
1,36 1,59
0 0
2,91 2,38
8,05 15,09
4,65 3,18
4,0 0,79
0 0
8,39 8,73
1,35 4,76
5,77 7,15
1,09 7,15
1,5 2,38
7,69 7,94
0,29 0,73
0 0
1,0 1,58 ·o 0
1,65 1,58
0,09 0,73 --------------
WINTER 1975 LENTE 1975
Relatiewc Relaticwe
Aantal Digtlteid 1) teenwoordig- Aantal Diglhcid 1) tecnwoordig-heid2 ) heid 2 )
% % -------- -------· __ .. _ ------- ------- -------- -------------
6 4,88 4,58 4 3,25 2,99
4 3,13 3,05 5 3,91 3,73
7 5,98 5,34 8 6,8 5,97
17 4,45 12,97 20 5,24 14,92
3 1,44 2,29 5 2,4 3,73
2 1,02 1,52 2 1,02 1,49
2 2,67 1,52 2 2,67 1,49
1 0,68 0,76 3 2,04 2,24
1 0,55 0,76 - 0 0
5 4,85 3,82 7 6,8 5,22
16 6,78 12,21 20 8,47 14,92
3 3,49 2,29 2 2,33 1,49
1 4,0 0,76 - 0 0
3 3,33 2,29 I 1,1 0,75
13 9,92 9,92 15 11,45 11,19
4 0,9 3,05 I 0,23 0,75
10 6,41 7,63 8 5,13 5,97
16 1,96 12,21 15 1,8 11,19
2 1,0 1,52 4 20 2,99
5 3,85 3,82 10 7,69 7,46
2 0,58 1,52 -- 0 0
2 1,8 1,52 - 0 0 3 3,0 2,29 - 0 0 I I 2,44 0,76 - 0 I 0,83 0,76 - 0 0 1 0,09 0,76 2 0,19 1,49
----------------- ----- -------<--------
131 134 ~------
.____ _______
aantal oorhictjics in karnp 2) Rclaticwe tcenwoordigheid
------------------- --·- --·-- -- -
GEMIDDELD
Relaticwe
Aantal Digthcid 1) tccnwoordig-
hcid2 )
% -------- --·- ------- -- -- ----------- -----·
6,0 4,88 4,75
5,5 4,3 4,36
6,25 5,33 4,95
17,0 4,45 13,46
4,5 2,16 3,56
1,5 0,77 1,19
2,0 2,66 1,58
2,0 1,36 1,58
0,5 0,28 0,4
4,75 4,61 3,76
17,0 7,2 13,46
2,75 3,2 2,18
0,5 2,0 0,4
1,0 1,11 0,79
12,75 9,73 10,10
4,5 1,02 3,56
9,5 6,09 7,52
13,0 1,57 10,30
2,75 1,38 2,18
8,0 6,15 6,34
0,75 0,22 0,39
0,5 0,45 0,4
1,25 1,0 0,99
0 0,61 0,2
0,75 0,62 0,59 ),0 0,09 0,79
r-------- --- -· ---~---- ·---- --
126,25 --------
X 100
Gccn oorhictjies is in kampe 25 en 27 aangctrcf nic en hierdie kampc is gcvolglik wcggclaat. to tale aantal oorbictjies in studie gcbied
N w
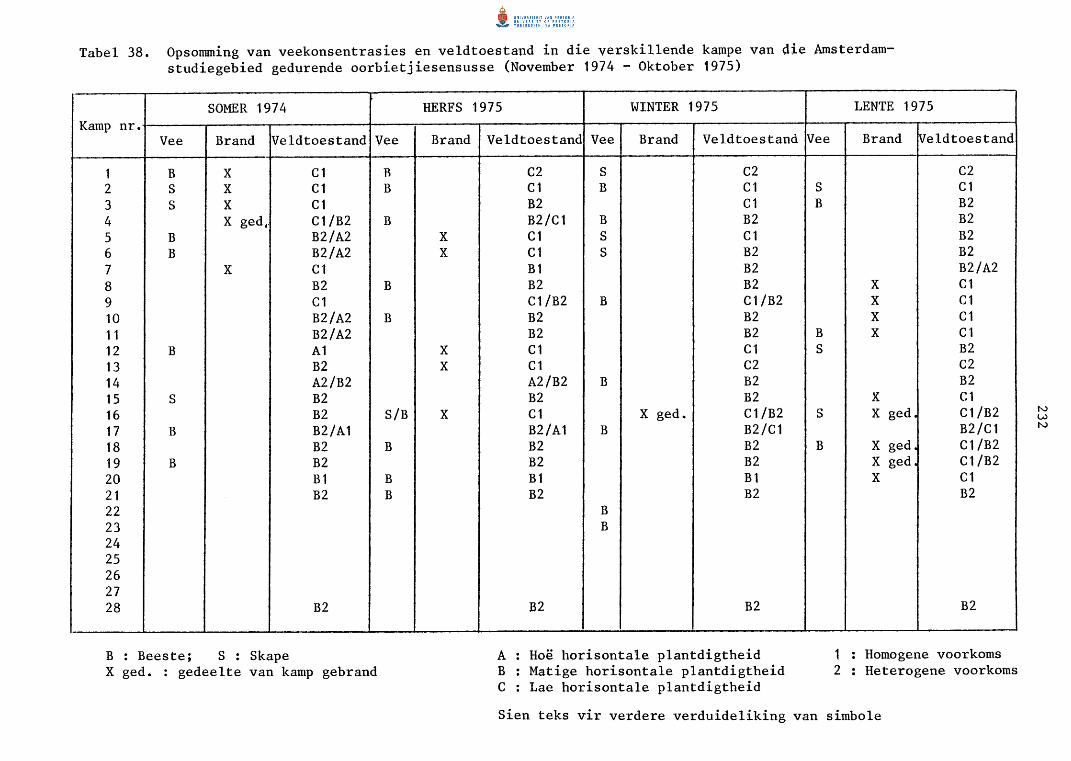
Tabel 38. Opsomming van veekonsentrasies en veldtoestand in die verskillende kampe van die Amsterdamstudiegebied gedurende oorbietjiesensusse (November 1974 - Oktober 1975)
SOMER 1974 Kamp nr.
Vee Brand Veldtoestand Vee
1 B X C1 R 2 s X C1 B 3 s X C1 4 X ged,. C1/B2 B 5 B B2/A2 6 B B2/A2 7 X C1 8 B2 B 9 C1 10 B2/A2 B 11 B2/A2 12 B A1 13 B2 14 A2/B2 15 s B2 16 B2 S/B 17 B B2/A1 18 B2 B 19 B B2 20 B1 B 21 B2 B 22 23 24 25 26 27 28 B2
B : Beeste; S : Skape X ged. : gedeelte van kamp gebrand
HERFS 1975 WINTER 1975
Brand
X X
X X
X
Veldtoestand Vee Brand Veldtoestand
C2 s C2 C1 B C1 B2 C1 B2/C1 B B2 C1 s C1 C1 s B2 B1 B2 B2 B2 C1/B2 B C1/B2 B2 B2 B2 B2 C1 C1 C1 C2 A2/B2 B B2 B2 B2 C1 X ged. C1/B2 B2/A1 B B2/C1 B2 B2 B2 B2 B1 Bl B2 B2
B B
B2 B2
A : Hoe horisontale plantdigtheid B : Matige horisontale plantdigtheid C : Lae horisontale plantdigtheid
LENTE 1975
Vee Brand IVeldtoestand
C2 s C1 B B2
B2 B2 B2 B2/A2
X C1 X C1 X C1
B X C1 s B2
C2 B2
X C1 s X ged C1/B2
B2/C1 B X ged C1/B2
X ged C1/B2 X C1
B2
B2
1 : Homogene voorkoms 2 : Heterogene voorkoms
Sien teks vir verdere verduideliking van simbole
N w N

233
In die meeste gevalle het die oorbietjiedigthede (en oorbietjiebiomassas)
in die verskillende kampe gedurende die studieperiode relatief konstant
gebly (Tabel 37, 39 en 40). Die grootste veranderinge in oorbietjie
digtheid het in kampe 4, 16, 18 en 20 voorgekom (Tabel 40). Kampe 4 en 18
bet lae horisontale plantdigthede gedurende die herfsmaande gehad. Die
grasveld in beide kampe het deurgaans n matig/heterogene horisontale
plantdigtheid gehad, terwyl beesbeweiding periodiek voorgekom het. Kamp
16 het n hoe somer-horisontale plantdigtheid gehad terwyl die kamp n lae
lente-horisontale plantdigtheid gehad het na n herfsbrand. Skape was
gedurende die lente aanwesig terwyl die veld gedeeltelik kortgebrand was.
n Hoe oorbietjiedigtheid is gedurende die lente in kamp 20 opgemerk na
'n len tebrand. Voor hierdie brand was die horisontale plantdigtheid hoog.
Belangrike vetanderinge in oorbietjiedigthede het ook in kampe 2, 11 en 17
voorgekom (Tabel 40). n Lae oorbietjiedigtheid is in kampe 2 en 11
gedurende die somer opgemerk waar die veld respektiewelike kortgebrand en
'n matigjheterogene horisontale plantdigtheid gehad het. Skape was egter
ook gedurende bierdie periode in kamp 2 aanwesig. Die oorbietjiedigtbeid
bet gedurende die lente in kamp 11 toegeneem na n lentebrand. Nadat die
veldtoestand van n rnatige tot hoe horisontale plantdigtheid (somer) in
kamp 17 tot 'n lae/rnatige horisontale plantdigtheid in die winter en lente
verander bet, het die oorbietjiedigtbeid ook gedaal.
die kamp word deur aangeplante weiding beslaan.
n Gedeelte van hier-
n Hoe oorbietjiedigtheid is in kamp 19 gedurende die lente waargeneem.
Hier het die veld 'n konstante, matig/heterogene borisontale plantdigtheid
gebad met die uitsondering van die lentemaande waartydens n gedeelte van
die kamp gebrand was. Gedurende die res van die jaar bet die veldtoestand
tussen n matige en n hoe horisontale plantdigtheid gewissel.
Met die uitsondering van kamp 3 en 15 het al die kampe wat belangrike
verandering in oorbietjiedigthede getoon het, hoe gemiddelde biomassas
gehad (Tabel 39). In al die kampe waar die berekende standaardafwyking
meer as 1,5 was (Tabel 40) bet n gemiddelde oorbietjiebiomassa van meer 2
as 60 kg/km voorgekom.
Geen bewyse van grootskaalse en/of langafstand seisoenale bewegings kon ge
vind word nie. Die veranderinge in oorbietjiedigthede het klaarblyklik as
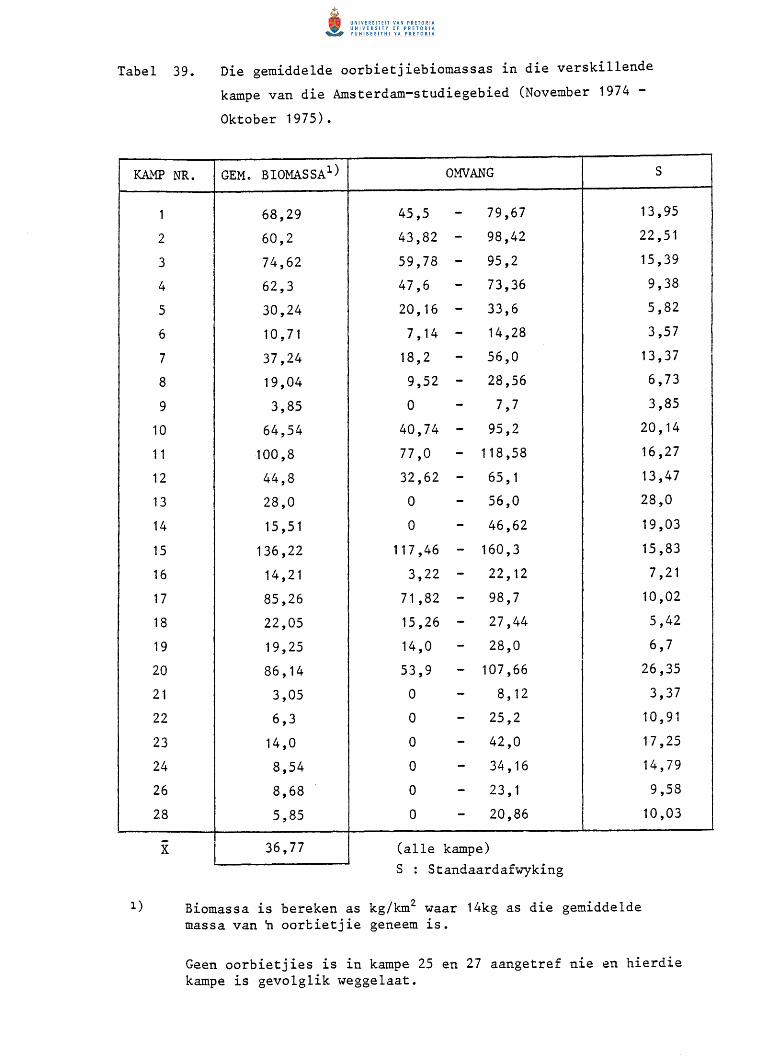
234
Tabel 39. Die gemiddelde oorbietjiebiomassas in die verskillende
kampe van die Amsterdam-studiegebied (November 1974 -
Oktober 1975).
KAMP NR. GEM. BIOMASSA1 ) OMVANG s
1 68,29 45,5 - 79,67 13,95
2 60,2 43,82 - 98,42 22,51
3 74,62 59,78 - 95,2 15,39
4 62,3 47,6 - 73,36 9,38
5 30,24 20' 16 - 33,6 5,82
6 10,71 7' 14 - 14,28 3,57
7 37,24 18,2 - 56,0 13,37
8 19,04 9,52 - 28,56 6,73
9 3,85 0 - 7,7 3,85
10 64,54 40,74 - 95,2 20' 14
11 100,8 77,0 - 118,58 16,27
12 44,8 32,62 - 65' 1 13,47
13 28,0 0 - 56,0 28,0
14 15,51 0 - 46,62 19,03
15 136,22 117,46 - 160,3 15,83
16 14,21 3,22 - 22' 12 7,21
17 85,26 71,82 - 98,7 10,02
18 22,05 15,26 - 27,44 5,42
19 19,25 14,0 - 28,0 6,7
20 86' 14 53,9 - 107,66 26,35
21 3,05 0 - 8' 12 3,37
22 6,3 0 - 25,2 10,91
23 14,0 0 - 42,0 17,25
24 8,54 0 - 34' 16 14,79
26 8,68 0 - 23' 1 9,58
28 5,85 0 - 20,86 10,03
X 36,77 (alle kampe) S : Standaardafwyk~ng
1 ) Biomassa is bereken as kg/km2 waar 14kg as die gemiddelde massa van n oorcietjie geneem is.
Geen oorbietjies is in kampe 25 en 27 aangetref nie en hierdie kampe is gevolglik weggelaat.

Tabel 40. Veranderinge in oorbietjiedigthede in die verskillende kampe van
die Amsterdam-studiegebied (Noveinber 1974- Oktober 1975).
VERANDERING KAMP NR. (STANDAARDAFWYKING)
0,1- 0,49 6' 9' 13' 24
0,5 - 0,99 3, 5, 7, 8, 12, 19, 21, 22, 26, 28
1,0- 1,49 1, 10, 14, 15, 23
1,5- 1,99 2, 11, 17
2,0 - 2,49 4' 16, 18' 20
Stanrlaardafwyking bereken op die relatiewe teenwoordigheid.
Geen oorbietjies is in kampe 25 en 26 aangetref nie en hierdie kampe is gevolglik weggelaat.
'
I
N w l/1

236
gevolg van heen en weer bewegings van die oorbietjies tussen aangrensende
kampe plaasgevind. n Voorbeeld hiervan is die bewegings tussen kampe 11,
16 en 17 waar kamp 11 se lae somerdigtheid gepaard gegaan het met hoe
digthede in kampe 16 en 17. Soortgelyke ooreenstemmende wisselings in
oorbietjiedigthede is onder andere ook tussen kampe 18, 19 en 20 waargeneem.
Familiegroep A (Fig. 38) in kamp 11 het na die aangrensende kamp 17 beweeg
nadat n gedeelte van laasgenoemde kamp gedurende die herfs van 1975 afgebrand
het. Hierdie familiegroep is vir n periode van ongeveer twee maande in kamp
17 opgemerk. Veertien uur nadat ongeveer 6 ha in kamp 15 as gevolg van
weerlig afgebrand het, is oorbietjies op die gebrande gedeelte gewaar.
Dit wil dus voorkom asof die seisoenale veranderinge in digthede van die
oorbietjies eerder aan menslike aktiwiteite as aan die teenwoordigheid
van skape en beeste toegeskryf kan word.
PIET RETIEF-STUDIEGEBIED
Oorbietjieverspreiding en Voorkeurgebiede
Die oorbietjies in die Piet Retief-studiegebied is hoofsaaklik tot die
sentrale gedeelte beperk (Fig. 76). Die totale vaste verspreidingsgebied
beslaan n oppervlakte van 17,6 km2
wat 40,9% van die studiegebied se opper-
vlakte uitmaak. Afgesien van ander ongereelde waarnemings van oorbietjies
het die verspreidingsgebied gedurende die studie onveranderd gebly.
n Gemiddeld van 4,56 oorbietjies/km2 is gedurende die vier seisoenale
tellings in die studiegebied getel. Die hoogste gemiddelde oorbietjie
digtheid (12 - 20 oorbietjies/km2) is op die oostelike gedeelte van die
plato-uitloper in die Alloteropsis semialata - Trachypogon spicatus - grasveld
en op die platotraphang in die Loudetia simplex - Trachypogon spicatus -
Rendlia altera - grasveld aangetref. Op die res van die sentrale plato
uitloper het n gemiddeld van tot 11,9 oorbietjies/km2 voorgekom. Afgesien
van die Alloteropsis semialata - Trachypogon spicatus - grasveld kom die
Gerbera sp. - Eulalia villosa - Eragrostis plana - grasveld ook in hierdie
gebied voor. Verskeie dagsome met die Xerophyta sp.- Oldenlandia herbacea-
klipplaatplantegroei en vleigemeenskappe in die omgewing van die riviervalleie
is ook in die gebied aanwesig maar oorbietjies is selde in die gebiede waar-
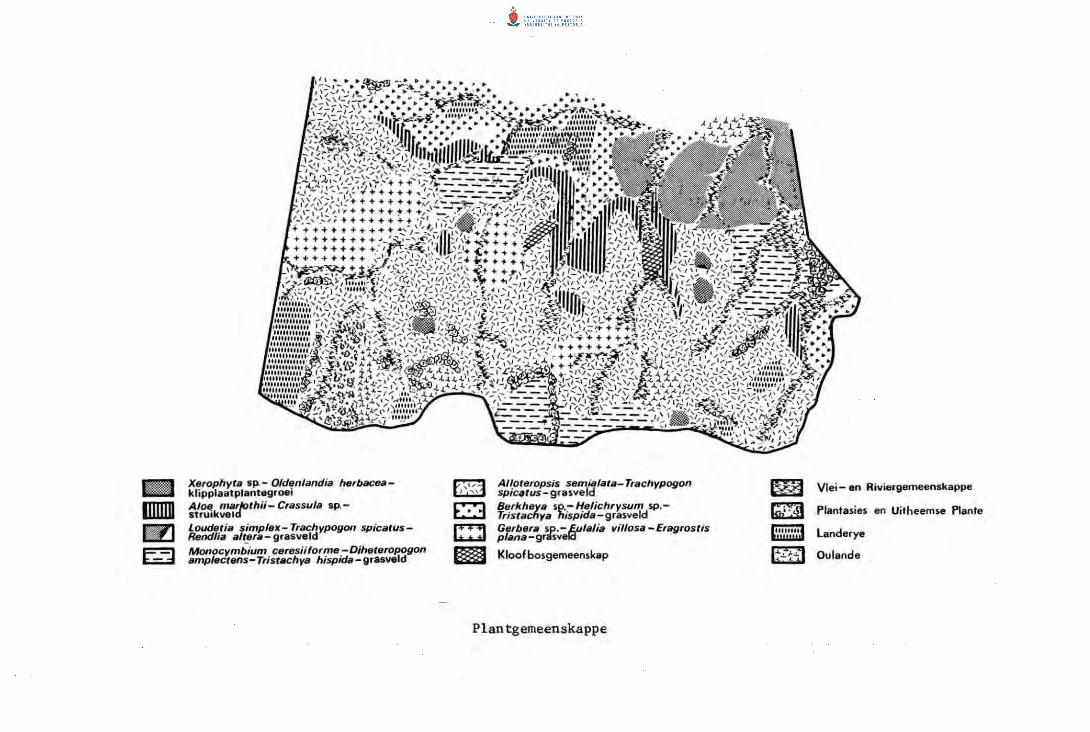
li!D Xerophyta sp. - Oldenlandia herbacea- [;§ Alloteropsis semialata-Trachypogon f-£ ... ;1 Vlei- en Riviergemeenskappe klipplaatplantegroei \ spic•tus-grasveld .
DlliiD Aloe ma';fthii- Crassula sp.-struikvea EZ3 Berkhex,a s~.-Helichrysum sp.-
Tristac ya ispida-grasveld ~ Plantasies en Uitheemse Plante
ll!l Loudetia simplex- Trachypogon spicatus- [!:!) Gerber11 Ts·- (Jul11lia villosa- Eragrostis ~ landerye Rendlia altilrll- grasveld pl;ma-gr sve
r;:=J Monocymbium ceresiiforme -Diheteropogon mml Kloofbosgemeenskap 1~{~\1 Oulande amplectens-Trist11chya hispida-grasveld
Plantgemeenskappe

••• 1 km2 ruit
--- Gemidctelde aantaf oorbietjiea per ruit
--- Grena van vaate verapreidingagebied
Gemiddelde aantal oorbietjiea per km2
• Q -7.9 • 4-5.9
• 8-9,9 • 2-3.9
~
10-11.9 • 1 -1.9
>12 • 02-0.9
M15
0 00
M15
• Oorbietjieverapreiding Cvaste t
X Ongereelde waarnemings D
1km
Paaie
Huise & hut te
IQ cP
Kampdrade
R a 5 Kampnommer
Figuur 76. Oorbietjieverspreiding in die Piet Retief-studiegebied.
N
11t
N w .........

238
gene em. Oorbietjies is nooit op die spitskruinreste in die ~ rnarlothii-
Crassula sp. - struikveld waargeneem nie.
Op die plato-uitloper bet minder as 4 oorbietjies/km2
in die direkte om
gewing van woonbuise, butte, landerye en plantasies voorgekom (Fig. 76).
Die grootste gedeelte van die suidwestelike en suidoostelike plato-uitloper
word deur klein plantasies en landerye beslaan (Fig. 77), en oorbietjies
is nie gedurende die studieperiode in bierdie plantasies of landerye waar
geneem nie.
Oorbietjies is nie op die pediment, eskarp en tallusbang asook in die
laerliggende gedeelte van die riviervalleie aangetre~ nie. Die tallushange
en eskarp is dig met kloofbosgemeenskappe begroei terwyl die Berkbeya sp. -
Helicbrysum sp. - Tristacbya bispida - grasveld kenmerkend van die pediment
is. Verspreide klein plantasies, landerye, paaie, woonbuise en butte is ook
in die pedimentgebied aanwesig.
Net soos in die geval van die Amsterdam-studiegebied kern die oorbietjies
meer dikwels op die plato-uitloper voor. Voorkeure vir spesifieke plant
gemeenskappe is nie duidelik waarneembaar nie, terwyl oorbietjies die
gebiede waar menslike aktiwiteite voorkom vermy.
Seisoenale Veranderinge in Oorbietjiedigtbede
Volgens die vier seisoenale tellings (November 1974 tot Oktober 1975) kern
daar seisoenale veranderinge in die oorbietjiedigtbede in die Piet Retief
studiegebied voor (Tabel 41). Die veranderinge in veekonsentrasies, veld
brand en veldtoestand (borisontale plantdigtbeid) gedurende die studieperiode
word in Tabel 42 aangetoon. Die oppervlaktes van die verskillende kampe
sowel as die gebiede wat deur geomorfologiese eenbede, plantasies, landerye,
menslike besetting en plantgemeenskappe beslaan word, word in Bylae C aangedui.
Soos in die geval van die ~~sterdam-studiegebied bet die oorbietjiedigtbede
(en oorbietjiebiomassas) meestal konstant gebly (Tabel 41, 43 en 44). Die
vernaamste veranderinge bet in kampe Ral en M6 plaasgevind. In die geval
van kamp Ral bet die digtbeid van 2,81 oorbietjies/km2 tot 11,8 oorbietjies/
km2 gewissel. n Ko~binasie van veekonsentrasies (skape gedurende die somer-

Figuur 77 . Men s like besetting , l anderye en plantas i es in die Pi et Re tie f
s tudiegebied.
N w
"'
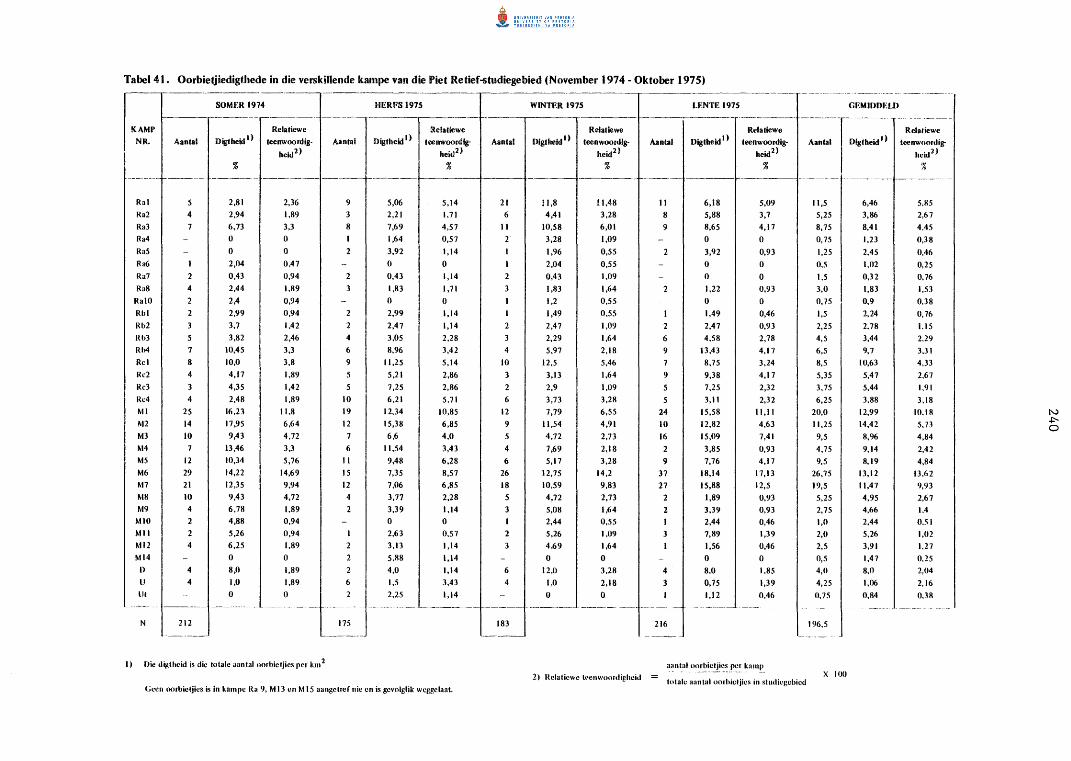
Tabel 41. Oorbietjiedigthede in die verskillende k~mpe van die Piet Retief-studiegebied (November 1974- Oktober 1975) ·-----4------ ------------------ --------------- --------------- -
SOMI~R 1974 HERFS 1975 WINTER 1975 LENTE 1975 GEMIDDEW ----------- ---- --- --- -------------
!CAMP Relatiewe Relaticwe Relatiewe Relatiewe NR. Aantal Digtheid I) teenwoordig- Digtheid 1) teenwoord ig- teenwoordig- Digtheid 1) teenwoordig-
heid2 ) lteid2} heid2 ) lteid2 )
% % % %
r --~--- ----Relatiewe
Aantal Digtheid 1) teenwoordig-
I hcid2
)
% -------- ------- --~~- ---- -------- ~ -----
Ral 5 2,81 2,36 9 5,06 5,14 21 u.s 11,48 II 6,18 5,09 11,5 6,46 5,85
Ra2 4 2,94 1,89 3 2,21 1.71 6 4,41 3,28 8 5,88 3,7 5,25 3,86 2,67
Ra3 7 6,73 3,3 8 7,69 4,57 11 10,58 6,01 9 8,65 4,17 8,75 8,41 4.45
Ra4 0 0 I 1,64 0,57 2 3,28 1,09 0 0 0,75 1,23 0,38
Ra5 0 0 2 3,92 1,14 I 1,96 0,55 2 3,92 0,93 1,25 2,45 0,46
Ra6 I 2,04 0,47 0 0 I 2,04 0,55 0 0 0,5 1,02 0,25 Ra7 2 0,43 0,94 2 0,43 1,14 2 0,43 1,09 0 0 1,5 0,32 0.76
Ra8 4 2,44 1,89 3 1,83 1,71 3 1,83 1,64 2 1,22 0,93 3,0 1,83 1,53
RalO 2 2,4 0,94 0 0 1 1,2 0,55 0 0 0,75 0,9 0,38
Rbl 2 2,99 0,94 2 2,99 1,14 I 1,49 0,55 1 1,49 0,46 1,5 2,24 0,76
Rb2 3 3,7 1,42 2 2,47 1,14 2 2,47 1,09 2 2,47 0,93 2,25 2.78 1.15 Rb3 5 3,82 2,46 4 3,05 2,28 3 2,29 1,64 6 4,58 2,78 4,5 3,44 2.29 Rh4 7 10,45 3,3 6 8,96 3,42 4 5,97 2,18 9 13,43 4,17 6,5 9,7 3.31
Rei 8 10,0 3,8 9 11,25 5,14 10 12.5 5,46 7 8,75 3,24 8,5 10,63 4,33
Rc2 4 4,17 1,89 5 5,21 2,86 3 3,13 1,64 9 9,38 4,17 5,35 5,47 2,67
Rc3 3 4,35 1,42 5 7,25 2,86 2 2,9 1,09 5 7,25 2,32 3,75 5,44 1,91
Rc4 4 2,48 1.89 10 6,21 5,71 6 3,73 3,28 5 3,11 2,32 6,25 3,88 3,18 Ml 25 16,23 11.8 19 12,34 10,85 12 7,79 6,55 24 15,58 11,11
M2 14 17,95 6,64 12 15,38 6,85 9 11,54 4,91 10 12,82 4,63
20,0 12,99 10,18
I N
11,25 14,42 5,73 ~ 0
M3 10 9,43 4,72 7 6,6 4,0 5 4,72 2,73 16 15,Q9 7,41 9,5 8,96 4,84 M4 7 13,46 3,3 6 11,54 3,43 4 7,69 2,18 2 3,85 0,93 4,75 9,14 2,42
MS 12 10,34 5,76 II 9,48 6,28 6 5,17 3,28 9 7,76 4,17 9,5 8,19 4,84 M6 29 14,22 14,69 IS 7,35 8,57 26 12,75 14,2 37 18,14 17,13 26,75 13,12 13,62 M7 21 12,35 9,94 12 7,06 6,85 18 10,59 9,83 27 15,88 12,5 19,5 11,47 9,93 M8 10 9,43 4,72 4 3,77 2,28 5 4,72 2,73 2 1,89 0,93 5,25 4,95 2,67 M9 4 6,78 1.89 2 3,39 1,14 3 5,08 1,64 2 3,39 0,93 2,75 4,66 1,4
MIO 2 4,88 0,94 0 0 I 2,44 0,55 I 2,44 0,46 1,0 2,44 0,51 Mil 2 5,26 0,94 I 2,63 0,57 2 5,26 1.09 3 7,89 1,39 2,0 5,26 1,02 Ml2 4 6,25 1,89 2 3,13 1,14 3 4,69 1,64 I 1,56 0,46 2,5 3,91 1.27 Ml4 0 0 2 5,88 1,14 0 0 0 0 0,5 1,47 0,25
() 4 8,0 1,89 2 4,0 1,14 6 12,0 3,28 4 8,0 1,85 4,0 8,0 2,04
lJ 4 1,0 1,89 6 1,5 3,43 4 1,0 2,18 3 0,75 1,39 4,25 1,06 2,16 lit 0 0 2 2,25 1,14 0 0 I 1,12 0,46 0,75 0,84 0,38
___ ......_ ___ - ------------ ---- -------- ---------- ---------- ---------- ---- --------- ------------- ------- -------------- ------------ - . - -~------
N 212 175 183 216 196,5 ----------- --
1) Die digtheid is die to tale aantal uorhietjies per km 2 a;~nt_al m~r-~i_e.!r~~-(l~! kam!'_
totalc aantal uorhictjies in studiegcbied 2) Rclaticwe tl~cnwuordigheid
Gcen oorbietjies is in kampc Ra 9, M 13 en M 15 aangclrcf nie en is gcvolglik weggelaat.
X 100

241
Tabel 42. Opsomming van veekonsentrasies en veldtoestand in die verskillende kampe van die Piet Retief-studiegebied gedurende oorbietjiesensusse (November 1974 - Oktober 197S)
SOMER 1974 HERFS / WINTER 197S LENTE 197S
Kam.p nr. Vee Brand Veld- Vee Brand Veld- Vee Brand toe stand toe stand
Ra1 80B/100S A1 3SB B2 BOB X Ra2 BOB/100S A1 3SB B2 BOB X Ra3 BOB/100S A1 3SB B1 BOB Ra4 40B/60S X C1 2SB B2 40B RaS 40B/60S X C1 2SB C2 40B Ra6 40B/60S X C1 2SB C2 40B Ra7 40B/60S B2 2SB B2 40B X RaB 40B/100S X C1 3SB C2 X Ra9 X C1 C2 Ra10 B2 B2 Rb1 X C1 C2 Rb2 A2 A2 X Rb3 X C1 140B C2 100B Rb4 X C1 B2 Rc1 A2 A2 100B X Rc2 60B B1 1408 B2 SOB X Rc3 SOB X C1 120S C2 Rc4 90B X C1 ~40S C2 M1 A2 BOB B2 100B X M2 SOB X C2 ·40B B2 M3 A2 60B B2 SOB X M4 40B X C1 B2 30B M5 110B X C1 B2 70B M6 A2 100B B2 tOOB X M7 A2 40B B2 SOB X MB 100B X C1 SOB C2 M9 A1 SOB B2 SOB X M10 B2 SOB B2 SOB X M11 A2 30B A2 40B X M12 A2 SOB B2 60B X M13 A2 A2 M14 B2 B2 M1S C2 C2 D X C1 C2 u C2 C2 Ut B2 B2
B : Beeste; S : Skape (getalle benaderd) Gereelde 14 dae wisselweiding; 3 uit 4 kampe per keer in gebruik
A Hoe horisontale plantdigtheid B Matige horisontale plantdigtheid C Lae horisontale plantdigtheid 1 Homogene voorkoms 2 Heterogene voorkoms
Sien teks vir verdere verduideliking van simbole
Veld-toe stand
C1 C1 C1 A2 B2 B2 C1 C1 C2 B2 B2
A2/C1 B2 C1 C1 C1 B2 B2 C1 B2 C1 B2 B2 C1 C1 B2 C1 C1 C1 C1 A2 B2 C2 C2 C2 B2

Tabel 43.
KAMP NR.,
Ra 1 Ra 2 Ra 3 Ra 4 Ra 5 Ra 6 Ra 7 Ra 8 Ra 10 Rb 1 Rb 2 Rb 3 Rb 4 Rc 1 Rc 2 Rc 3 Rc 4 M 1 M 2
M 3 M 4 M 5 M 6 M 7 M 8
M 9 M 10 M 11 M 12 M 14
D
u Ut
X
242
Die gemiddelde oorbietjiebiomassas in die verskillende kampe
van die Piet Retief-studiegebied (November 1974- Oktober 1975).
GEM., BIOMASSA1 ) OMVANG s
90,48 39,34 - 165,2 53,54 54,04 30,94 - 82,32 19,74
117,78 94,22 - 148,12 19,93 17,22 0 - 45,92 19,04 34,30 0 - 54,88 22,75 14,28 0 - 28,56 14,28
4,52 0 - 6,02 3,01 25,62 17,08 - 34,16 6,97 12,6 0 - 33,6 13,93 31,36 20,86 - 41,86 10,5 38,89 34,58 - 51,8 7,46 48,09 32,06 - 64., 12 13,81
135,84 83,58 - 188,02 37,65
148,75 122,5 - 175,0 19,57 76,62 43,82 - 131,32 38,36
76' 13 40,6 - 101 ,5 26,37 54,36 34,72 - 86,94 22,87
181,79 109,06 - 227,22 46,78
201,92 161,56 - 251,3 34,46
125,44 66,08 - 211,26 54,83
127,89 53,9 - 188,44 51 ,68
114' 7 72,66 - 144,76 27,53
183,61 102,9 - 253,96 54' 15 160,58 98,84 - 222,32 44,52
69,34 26,46 - 132,32 38,9 65,24 47,46 - 94,92 19,67
34,16 0 - 68,32 24' 15 73,64 36,82 - 110,46 26,04
54,71 21,84 - 87,5 24,46
20,58 0 - 82,32 35,65
112 '0 56,0 - 168,0 45,72
14,88 10,5 - 21,0 3,81
11 '8 0 - 31,5 13,05
66,67 (alle kampe)
S : Standaardafwyking
l) Biomassa is bereken as kg/km2 waar 14kg as die gemiddelde massa van n oorbietjie geneem is.
Geen oorbietjies is in kampe Ra 9, M 13 en M 15 aangetref nie en hierdie kampe is gevolglik weggelaat.

Tabel 44. Veranderinge in oorbietjiedigthede in die verskillende kampe van
die Piet Retief-studiegebied (November 1974- Oktober 1975).
VERANDERING KAMP NR. (STANDAARDAFWYKING)
0,1- 0,49 Ra 6, Ra 8, Ra 10, Rb 1, Rb 2, Rb 3, M 9, M 10, M 11
0,5 - 0,99 Ra 2, Ra 4, Ra 5, Ra 7, Rb 4, Rc 3, M 12, M 14, D, U, Ut
1,0- 1,49 Ra 3, Rc 1, Rc 2, M2, M4, M5
1,5- 1,99 P_c 4, M 3, M 8
2,0 - 2,49 M 1, M 7
2,5 - 2,99 -0 3,0 - 3,49 -3,5 - 3,99 Ra 1, M 6
Standaardafwyking bereken op die relatiewe teenwoordigheid.
N .p. w

244
maande) en veldtoestand (van n hoe heterogene horisontale plantdigtheid
tot n matige en lae homogene horisontale plantdigtheid) was moontlik die
vernaamste faktore. Die veldtoestand het ook in kamp M6 op n soortgelyke
wyse gewissel alhoewel vee gedurende somermaande afwesig was terwyl die kamp
"gerus" het. Hier het die oorbietjiedigtheid gewissel van 7,35 oorbietjies/
km2 (herfs) tot 18,14 oorbietjies/km2 in die lente nadat die veld gebrand
het.
Ander belangrike veranderinge in oorbietjiedigthede het in kampe Ml en M7
voorgekom. In beide kampe was daar n hoe oorbietjiedigtheid gedurende
die somer en lente waar die veld van n hoe/heterogene horisontale plant
digtheid sender vee (somer) tot n cop, gebrande veld (lente) met beeste
gewissel het. n Duidelike laer oorbietjiedigtheid is gedurende die herfs
en winter opgemerk waartydens die veld n matig/heterogene horisontale plant
digtheid met beesbeweiding gehad het.
In kamp Rc4 het n lae oorbietjiedigtheid gedurende die sorner voorgekom wat
afgewissel is met n hoer digtheid veral gedurende die herfs. Voor die
somertelling het die veld n hoe/heterogene horisontale plantdigtheid gehad.
Na n somerbrand is die kamp deur skape bewei.
n Lae/oorbietjiedigtheid het gedurende die winter in kamp M3 (matig/hetero
gene horisontale plantdigtheid) voorgekom, in teenstelling met die hoe
oorbietjiedigtheid gedurende die lente (oop, gebrande veld). Hier het
beesbeweiding voortdurend voorgekom met die uitsondering van die somermaande.
Na n somerbrand het n hoe oorbietjiedigtheid in karnpe M8 voorgekom wat
geleidelik verminder het tot n baie lae digtheid gedurende die lente.
Alhoewel die veld n matig/heterogene horisontale plantdigtheid gedurende
die lente gehad het, was geen vee aanwesig nie.
Verdere belangrike veranderinge in die oorbietjiedigthede het ook in kampe
Ra3, Rcl, M2, M4, MS en Rc2 voorgekom. n Hoe oorbietjiedigtheid gedurende
die somer is in kampe M2, M4 en M5 waargeneem waartydens die veld n lae/
homogene tot heterogene horisontale plantdigtheid gehad het. Lae oorbietjie
digthede het gedurende die winter (kampe M2 en M5) en lente (kamp M4) in
hierdie kampe voorgekom terwyl die horisontale plantdigtheid matig/heterogeen
was.

245
Kampe Ra3 en Rcl het n hoe oorbietjiedigtheid zedurende die winter gehad met
lae digthede gedurende die somer en lente respektiewelik. In die geval
van kamp Rcl het die hoogste oorbietjiedigtheid voorgekom terwyl die veld
n hoe/heterogene voorkoms gehad het. Tydens die laagste digtheid was die
horisontale plantdigtheid laag en homogeen. Met die uitsondering van
kampe Rcl en M5 (gedurende die winter) het al ses kampe vee (hoofsaaklik
beeste) gedurende die studieperiode in gehad.
Slegs klein veranderinge het in die ander oorblywende kampe voorgekom in
welke geval daar slegs tussen een en drie oorbietjies per kamp betrokke was.
Al die kampe wat belangrike veranderinge in oorbietjiedigthede getoon het,
het relatiewe hoe gemiddelde biomassas gehad (Tabel 41 en 43). n Gemiddelde
biomassa van meer as 50 kg/km2 het in die kampe voorgekom waar die verandering
in die berekende standaardafwyking in oorbietjiedigthede meer as 1,5 was (Tabel
43 en 44).
Geen grootskaalse migrasies oor lang afstande is waargeneem nie en dit blyk,
soos in die geval van die Amsterdam-studiegebied, dat bewegings ook hoofsaak
lik tussen aangrensende kampe voorgekom het. n Voorbeeld hiervan is kampe
Rc2 en Rc4 waar n hoe oorbietjiedigtheid gedurende die lente (kamp Rc2) met
n hoe digtheid gedurende die herfs (kamp Rc4) afgewissel het. Soortgelyke
veranderings in oorbietjiedigthede is onder andere tussen kampe M3 en M8,
en kampe Ml en M6 waargeneem.
Die veranderinge in oorbietjiedigthede is klaarblyklik hoofsaaklik deur
kort gebrande veld en in n mindere mate deur veebewegings veroorsaak.
Gevolgtrekking
Die oorbietjieverspreidingsgebied in die Piet Retief-studiegebied is groter
as die in die Amsterdam-studiegebied, terwyl die oppervlakte van die versprei
dingsgebied in verhouding tot die totale studiegebied ook groter is in die
geval van die Piet Retief-studiegebied. Die oorbietjie-verspreidingsgebied
in die Amsterdam-studiegebied is opvallend meer gefragmenteer as in die
Piet Retief-studiegebied.

246
Die gemiddelde oorbietjiedigtheid in die Piet Retief-studiegebied (4,56
oorbietjies/km2) is die hoogste digtheid bekend. Tinley (1977) het n digtheid van 3 oorbietjies/km2 in die Nasionale Gorongosa-wildtuin bepaal.
Volgens Van Lavieren en Esser (1979) wissel die oorbietjiedigtheid tussen
0,81 en 2,81 oorbietjies in die Nasionale Bouba Ndjida-wildtuin in die
Kameroen. n Digtheid van 2,68 oorbietjies/km2 is in die Nasionale
Akagera-wildtuin deur Monfort en Monfort (1974) aangeteken, alhoewel
Bourliere (1965) n digtheid van 0,6 oorbietjies/km2
en Spinage (1969, In:
Monfort en Monfort ££• cit.) 0,8 oorbietjies/km2
in die Akagera-wildtuin
gevind het. n Oorbietjiedigtheid van 65,5 ha/oorbietjie (1,53 oorbietjies/
km2) is deur Oliver, Short en Hanks (1978) in die Highmoor Staatsbos in
Natal opgemerk.
In beide studiegebiede word die hoogste oorbietjiedigthede op die plato
uitlopers met verspreide dagsome aangetref. Die plato-uitlopers het
hellings wat van 1° tot 20° wissel behalwe waar n platoberggebied (Amsterdam
studiegebied en spitskruinreste (Piet Retief-studiegebied) aangetref word
(kyk HOOFSTUK 5 : Resultate). Alhoewel oorbietjies nie op die spitskruin
reste gevind is nie, kom daar tussen 4 en 5,9 oorbietjies/km2 op die plato
berggebied voor waar die hellings tussen 1° en 4° wissel. Die plantegroei
van die twee studiegebiede se plato-uitlopers toon n sterk ooreenkoms, veral
wat die Berkheya setifera - Vernonia natalensis - Eulalia villosa - grasveld
(~~sterdam-studiegebied) en die Alloteropsis semialata - Trachypogon spicatus -
grasveld (Piet Retief-studiegebied) betref. Trachypogon spicatus,
Hyparrhenia birta, Tbemeda triandra, Eulalia villosa en Monocymbium ceresiiforme
is deurgaans in hierdie gemeenskappe teenwoordig (kyk HOOFSTUK 6 : Resultate).
Geen oorbietjies·is op die Piet Retief-studiegebied se eskarpe en tallusbange
aangetref nie, terwyl slegs lae konsentrasies op die tallusbange van die
Amsterdam-studiegebied gevind is. Die tallushange in beide studiegebiede
het n belling van 15° tot 40°. Die eskarpe, wat slegs in die Piet Retief
studiegebied teenwoordig is, het n belling van meer as 50°. In laasgenoemde
studiegebied kom struikveld- of kloofbosgemeenskappe teen die tallushange
voor.
In beide studiegebiede kom daar min oorbietjies op die pedimentgedeeltes voor
waar menslike besetting, plantasies, en landerye aangetref word. Plant-

247
gemeenskappe wat deur versteuring gekenmerk word, is ook in hierdie gedeeltes
aanwesig. Oorbietjies het plantasies en landerye deurgaans vermy.
Verskeie werkers (Dorst en Dandelot 1970, Mason 1973, Monfort en Monfort
1974 en Smithers en Lobao Tello 1976) maak van die oorbietjie se voorkeure
vir oop grasveld en glooiings met lae hellings melding. In die Highmoor
Staatsbos het slegs 9,68% van die oorbietjies op glooiings met n helling
van meer as 15° voorgekom (Oliver et. !!• 1978).
Oorbietjies se voorkeur vir gebrande veld is in beide studiegebiede gevind
waar oorbietjies selfs enkele ure na n brand in die gebrande veld opgemerk
is. Oorbietjies beweeg waarskynlik oor kart afstand sodat hulle op die
gebrande gedeeltes van of na aan hulle territoriums konsentreer. Irby
(1973) vermoed dat rooiribbokke soortgelyke kort-afstandbewegings na
gebrande gedeeltes doen. In die Jack Scott-privaatnatuurreservaat het
Mason (1973) ook waargeneem dat gebrande gedeeltes deur oorbietjies benut
word en hy verklaar dat dit moontlik op n voorkeur vir kart gras dui.
Monfort en Monfort (1974) het nie gevind dat brande die verspreiding van
oorbietjies inn groot mate beinvloed nie. Volgens Oliver~· !l· (1978)
het oorbietjies n hoe voorkeur vir n Septemberbrand in die Highrnoor
Staatsbos getoon. Seisoenale oorbietjiebewegings vind soms in die Gorongosa
wildtuin plaas, maar alhoewel Tinley (1977) nie die oorsaak kon bepaal nie,
meen hy dat bytvliee moontlik tot hierdie bewegings aanleiding kan gee.
Die bewegings van ander kleiner boksoorte soos klipspringer en steenbok
word klaarblyklik nie deur brande beinvloed nie, alhoewel verskeie ander
grater hoefdiere wel soms op brande konsentreer, soos onder andere blou
wildebeeste (Talbot en Talbot 1963), waterbokke (Hanks, Price en Wrangham
1969) en blesbokke (Lynch 1971, Mason 1973).
Millar (1970) vermoed dat brande n beperkende faktor by oorbietjiebevolkings
is weens die gehegtheid aan spesifieke territoriums. Volgens Millar
(££• £!!.) bly oorbietjies weens hulle territoriale gehegtheid op gebrande
veld waar nie voldoende voedsel is nie. Gedurende 1975 is vier oorbietjies
in die Arnsterdam-studiegebied gevind wat weens n ongeluksvuur doodgebrand
het. Brande kan wel mortaliteite veroorsaak wanneer onoordeelkundige of
ongeluksvure oorbietjies oorval. Brande kan ook n indirekte beperkende

248
faktor wees wanneer die oorbietjies op die kort veld duideliker sigbaar is.
Tensy klippe aanwesig is kan die oorbietjies dan nie op primere verdedigings
patrone staatmaak om moontlike predasie te ontwyk nie. Indien oorbietjies
op kortgebrande veld versteur word, vlug bulle selfs na aangrensende kampe
waar langer gras voorkom (kyk HOOFSTUK 10 : VLUGREAKSIES EN ANTIROOFDIER
VERHOUDINGS).
Kleinskaalse bewegings tussen kampe is in beide studiegebiede aangetref wat
grootliks aan die veldtoestand en in n mindere mate aan veebewegings toe
geskryf kan word. Die bestaande kampdrade is bedoel om veebewegings te
beperk, en bierdie kampdrade laat oorbietjiebewegings ongebinderd toe,
alboewel ander boksoorte soos blesbokke wel gekeer word. Min is bekend
oor die invloed van boerderyaktiwiteite op wildsoorte. Hood en Inglis
(1974) bet aangetoon dat takbokke Odocoileus virginianus wel tydelik
bul tuisgebied verskuif bet weens die invloed van beesbewegings. Bees
bewegings het klaarblyklik min invloed op die bewegings van oorbietjies
ten spyte daarvan dat aktiwiteitsperiodes wel geraak word (kyk HOOFSTUK
8 : FAKTORE WAT AKTIW~TEITE BEINVLOED). Die teenwoordigheid van groot
getalle skape in n kamp het in somnige gevalle moontlik wel daartoe bygedra
dat oorbietjies die betrokke kamp verlaat het.

249
OPSOMMING
Oorbietjies het vroeer regdeur die grasveldgebiede van Suider-Afrika
voorgekom. Die getalle van oorbietjies het egter drasties gedurende
die afgelope tyd afgeneem. Uin is bekend oor hierdie spesie se ekologie en
etologie. Die doelstelling van die huidige studie was om die oorbietjie G
se e~o-etologie te ondersoek met klem op habitatsvereistes en verspreiding.
Die studiegebiede naby Amsterdam en Piet Retief in Suidoos-Transvaal is
uitgesoek na n opname regdeur die provinsie en veldwerk het gedurende
Desember 1973 in aanvang geneem wat tot Julie 1976 geduur het.
Die oorbietjie toon spesifieke aanpassings wat aktiwiteitspatrone, sosiale
organisasie, vo.edingsgedrag en verdedigingsmeganismes betref soda t di t
in staat gestel word om in oop grasveldgebiede te bestaan.
n Basiese aktiwiteitspatroon kom voor waartydens oorbietjies gemiddeld
34,2% van dagligperiodes aktief is. Faktore soos reen en veeversteurings
beinvloed die aktiwiteitspatroon. Aktiwiteite in familiegroepe is
grootliks gesinkroniseer. Seisoenale verskille wat die tydsverspreiding
van aktiwiteitsperiode betref is waargeneem.
daar n toename in wei-aktiwiteite.
Gedurende die winter is
Oorbietjies kom in familiegroepe voor bestaande uit veral pare en groepe wat
deur n ram en twee ooie gevorrn word. Die rarnrne verdedig reg deur die jaar
n territorium wat met die tuisgebied oorvleuel. Hierdie territorium word
veral deur preorbitale tiiermerking en fekaliee afgebaken. In die Amsterdam
studiegebied is n gemiddelde territoriumgrootte van 34 ha gevind.
n Enkele lam word na n draagtyd van ongeveer 210 dae gebore. Die lam-
seisoen se piek duur van November tot Januarie, en die lammers beweeg vir
n periode van drie tot vier maande nie saam met die familiegroep nie.
Tydens voeding toon oorbietjies selektiwiteit wat plantspesies sowel
as verskillende plantdele betref. Oorbietjies is hoofsaaklik grasvretend
alhoewel verskeie nie-grasagtige kruide ook benut word. Oorbietjie
territoriums sluit een of meer plantegroeigemeenskappe in. n Hoe voedsel-

250
voorkeur word vir veral Eulalia villosa, Monocymbium ceresiiforme en
Hyparrhenia hirta getoon5
Oorbietjies is nie van oop wateroppervlaktes vir drinkwater afhanklik nie.
Die enigste bron van water is die voedselplante en neerslag soos dou of
ryp op hierdie plante.
Interaksies met ander wilde hoefdierspesies is beperk en geen positiewe
assosiasies kom voor nie. Oorbietjies toon aanpassings wat beide primere
en sekondere verdedigingsmeganismes betref, en vlugafstande wissel na
gelang van topografie en horisontale plantdigtheid. n Kleiner vlug
afstand word in lang grasveld aangetref as in kort grasveld.
Oorbietjiedigthede van 4,56 en 2,68 oorbietjies/km2 is in die Piet Retief
en Amsterdam-studiegebiede respektiewelik gevind. Voorkeure word vir
plato-uitlopers getoon terwyl oorbietjies selde op tallushange aangetref
word. Menslike besetting, plantasies en landerye word grootliks vermy.
Kleinskaalse bewegings kom soms voor wat deur veebewegings, algemene veld
toestand en brande beinvloed word. Duidelike voorkeure word vir gebrande
veld getoon.

251
SUMMARY
In the past, oribi's were widespread throughout the grassland areas
of Southern Africa. Numbers have, however, declined drastically
in recent years. Little is known about the ecology and ethology
of this species and the objectives of the present study were to investi
gate these particularly with regard to habitat requirements and distribu
tion. The study areas near Amsterdam and Piet Retief in the south
eastern Transvaal were only selected following a survey throughout the
Province and fieldwork commenced in December 1973 lasting until July
1976.
The oribi has developed specific adaptations relating to activity patterns,
social organisation, feeding behaviour and defence mechanisms which have
enabled the species to survive in open grassveld.
There is a basic daylight activity pattern during which oribis are
active for 34,2% of daylight hours. Factors such as rain or domestic
stock movements influence this pattern. Activities in family groups
are largely synchronised and there are seasonal changes in activity
patterns with an increase in grazing during the winter.
Oribis occur in family groups consisting particularly of pairs and groups
of a ram and two ewes. The rams defend a territory throughout the year
which overlaps with the home range. Each territory is marked out using
pre-orbital and faecal marking. Mean territory size in the Amsterdam
study area was 34 ha.
A single lamb is born following a gestation length of approximately 210
days. The peak in lambing occurs from November to January and, following
birth, the lambs do not move around with the group for three to four months.
Oribis select for both plant species and plant parts during feeding. They
are primarily grazers although forbs are also utilised. Territories may
include one or more plant community. A high nutritional content is
selected for in particularly Eulalia villosa, Monocymbium ceresiiforme and

253
LITERATUURLYS
+ Literatuur nie gesien nie
ACOCKS, J.P.H. 1975. Veld types of South Africa. Mem. bot.
Surv. s. Afr. 40: 1 - 128. Government Printer, Pretoria.
+AESCHLIMANN, A. 1963. Observations sur Philantomba maxwelli
(Hamilton-Smith), une antilope de la foret eburnee.
Acta trop. 20: 341 - 368.
ALTMAN, M. 1958. The flight distance in free-ranging big game.
J. Wildl. Mgmt. 22(2): 207 -· 209.
ANONIEM. 1952. The vanishing oribi - the position reviewed.
Afr. wild Life 6(1): 8 - 10.
ANONIEM. 1967. Transfer of oribi to nature reserve.
Afr. wild Life 21(1): 81
ANONIEM. 1973. Die herinvoer van oorbietjies in die Nasionale
Krugerwildtuin. Custos 2(11): 25 - 29.
ANONIEM. 1977. Sehlabathebe -Lesotho's National Park.
Afr. wild Life 31(4): 26 - 28.
ANSELL, W.F.Ho 1960. Mammals of Northern Rhodesia. The
Government Printer, Lusaka.
ANSELL, W.F.H. 1965. Standardisation of field data on mammals.
Zool. Afr. 1(1): 97 - 113.
ANSELL, W.F.H. 1971. Order Artiodactyla. In: The Mammals of
Africa: an identification manual. Eds. J. Meester and
H.W. Setzer. Smithsonian Institute, Washington D.C.
ASDELL, S.A. 1946. Patterns of mammalian reproduction. Comstock
Publishing Associates, Cornell University, Itaca, New York.

254
BACKHAUS, D. 1961. Beobachtungen an Giraffen in zoologischen Garten
und in freier Wildbahn. Inst. Pares Nat. Congo, Brussels.
BALCH, c.c. 1955. Sleep in ruminants. Nature, Lond. 175:
940 - 941.
BEST, G.A. 1962. Rowland Ward's records of big game. Ninth edition
(Africa). Rowland Ward, London.
BIGALKE, R.C. & J.Ao BATEMAN. 1962. On the status and distribution
of ungulate mammals in the Cape Province, South Africa. Ann. Cape
Prov. Mus. 2: 85 - 109.
BLIGH, J. & HARTHOORN, A.M. 1965. Continuous radiotelemetric records
on the deep body temperature of some unrestrained African mammals
under near-natural conditions. J. Physiol., Lond. 176: 145 - 162.
BOURQUIN, 0. 1966. The larger mammals occuring in the Natal Game and
Nature Reserves. Unpublished report of the Natal Parks, Game and
Fish Preservation Board, Pieterrnaritzburg.
BOTHMA, J. DU P. 1975. Conservation status of the larger mammals of
Southern Africa. Biol. Conserv. 7: 87 - 95.
' BOURLIERE, F. 1965. Densities and biomass of some ungulate populations
in Eastern Congo and Rwanda, with notes on population structure and
lion/ungulate ratios. Zool. Afr. 1(1): 199 - 207.
BRAUN-BLANQUET, J. 1928. Pflazensoziologie. 1. Aufl. Springer-Verlag,
Wien.
BREDENKAMP, G.J. 1975.
randnatuurreservaat.
~ Plantsosiologiese studie van die Suikerbos
M.Sc. -verhandeling. Universiteit van Pretoria.
+BROCK, V.Eo & R.H. RIFFENBURGH. 1960. Fish schooling: a possible factor
in reducing predation. J. Cons. perm. int. Explor. Med. 25: 307 - 317.
BULLOCK, R.E. 1974. Functional analysis of locomotion in pronghorn antelope.
Pp. 274 - 305. In: The behaviour of ungulates and its relation to
management. Eds. V. Geist and F. Walther. IUCN Publo New Series 24,
Morges, Switzerland.

255
+BURCKHARDT, D. 1958. Kindliches Verhalten als Ausdrucks-bewegung im
Fortpelanzungszeremoniell einiger Wiederkauer. Rev. suisse Zool.
65: 311 - 316.
BURT, W.H. 1943. Territoriality and home range concepts as applied to
mammals. J. Mammal. 24: 346 - 352.
CADE, LoE. 1966. A note on the mating behaviour of Kenya oribi
Ourebia ourebi in captivity. Int. Zoo Yb. 6: 205.
CHIPPENDALL, L.K.A. 1955. A guide to the identification of grasses
in South Africa. In: The grasses and pastures of South Africa.
Ed. D. Meredith. Central News Agency, Johannesburg.
CLOUGH, G. & AoG. HASSAM. 1970. A quantitative study of the warthog
in the Queen Elizabeth National Park, Uganda. E. Afr. Wildl. J.
8: 19 - 24.
COETZEE, B.J. 1972. n Plantsosiologiese studie van die Jack Scott-
natuurreservaat. M.Sc. -verhandeling, Universiteit van Pretoria.
COETZEE, B.J. 1974. A phytosociological classification of the Jack
Scott Nature Reserve. Bothalia 11: 329 - 374.
COLE, M. 1966. South Africa. Methuen and Co. Ltd., London.
COOKE, RoU. & J.C. DOORNKAMP. 1974. Geomorphology in environmental
management. Clarendon Press, Oxford.
+DAHL, E. 1956. Rondane mountain vegetation in South Norway and its
relation to the environment. Skr. norske Vidensk-Akad. 3: 1 - 374.
DARLING, F.F. 1969. A herd of red deer. Oxford University Press, London.
DAUBENMIRE, R.F. 1968. Plant communities. A textbook of plant synecology.
Harper & Row, New York.
DAVID, JoH.M. 1973. The behaviour of the bontebok, Damaliscus dorcas
dorcas (Pallas, 1766), with special reference to territorial behaviour.
Z. Tierpsychol. 33: 38 - 107.

256
DEVOS, A. & H.S. MOSBY. 1971. Habitat analysis and evaluation.
Pp. 135 - 172. In: Wildlife management techniques. Third
edition. Ed. R.H. Giles. The Wildlife Society, Washington.
DORST, J. £ P. DANDELOT. 1970. A field guide to the larger mammals
of Africa. Collins, London.
DUNBAR, R.I.M. & E.P. DUNBAR. 1974. Social organization and ecology
of the klipspringer (Oreotragus oreotragus) in Ethiopia. z. Tierpsychol.
35: 481 - 493.
+DUNCAN, Po 1975. Topi and their food supply. Ph.D. Thesis. University
of Nairobi.
DU PLESSIS, S.F. 1969. Past and present geographical distribution of the
Perissodactyla and Artiodactyla in Southern Africa. MoSc. Thesis.
University of Pretoria.
DU PLESSIS, s.s. 1972. Ecology of blesbok with special reference to
productivity. Wildl. Monogr. Chestertown 30.
EDMUNDS, M. 1974. Defence in animals. Longman, Essex.
EDWARDS, D. 1974. Survey to determine the adequacy of existing conserved
areas in relation to vegetation types. A preliminary report. Koedoe
17: 2 - 37.
EDWARDS, Ko 1968. The varnishing oribi. The Eastern Cape Naturalist
32: 20 - 23.
EDWARDS, K. 1969a. The oribi - a review. The Eastern Cape Naturalist
36: 17 - 18.
EDWARDS, K. 1969b. Vanishing oribi. Inyala News 16: 9 - 10.

257
EIBL-EIBESFELDT, I. 1970. Ethology. The biology of behavior. Holt,
Rinehart and Winston, New York~
+ELLENBERG, Ho 1956. Aufgaben und Methoden der Vegetationskunde.
In: Einfuhrung in die Phytologie. 4: 1 - 136. Ed. H. Walter.
Eugen Ulmer, Stuttgart.
ELOFF, F.C. 1959. Observations on the migration and habits of the
antelopes of the Kalahari Gemsbok Park. Parts 1 and 2. Koedoe
2: 1 - 51.
ESSER, J. 1973. Beitrage zur Biologie des afrikanischen Rhebockes
(Pelea capreolus Forster 1790). Doktorsdissertation. Christian
Albrechts Universitat, Kiel.
ESTES, R.Do 1967. The comparative behavior of Grant's and Thomson's
gazelles. J. Mammal. 48: 189 - 209.
ESTES, R.D. 1969. Territorial behaviour of the wildebeest (Connochaetes
taurinus Burchell, 1823). Z. Tierpsychol. 26: 284 - 370.
ESTES, R.D. & J. GODDARDo 1967. Prey selection and hunting behavior of
the African wild dog. Jo Wildl. Mgmt. 31: 52 - 70.
EWER, RoF. 1973. Ethology of mammals. Elek Science, London.
+FERRAR, AoAo 1973. Niche quantification: an approach to the analysis
of habitat relationships in large herbivore communities. M.Sc.
Thesis. University of Rhodesia.
FIELD, CoR. 1968. Methods of studying the food habits of some wild
ungulates in Uganda. Proc. Nutr. Soc. 27: 172 - 173.
FITZPATRICK, RoWo 1974. Mineralo-chemical studies on soils and related
material from pedosystems in the south-eastern Transvaal.
Thesis. University of Natal, Pietermaritzburg.
M.Sc.(Agric.)
FORBES, D. 1938. My life in South Africa. H.F. & G. Witherby Ltd.,
London.
GODDARD, J. 1968. Food preferences of two black rhinoceros populations.
E. Afr. Wildl. J. 6: 1 - 18.

258
GOSLING, L.Mo 1969. Parturition and related behaviour in Coke's harte-
beest, Alcelaphus buselaphus cokei GUnther. J. Reprod. Fert., Suppl.
6: 265 - 286.
GOSLING, L.Mo 1972. The construction of antorbital gland marking sites
by male oribi (Ourebia ourebi Zimmerman 1783). z. Tierpsychol. 30
(3): 271 - 276.
1976.
Area, Rhodesia.
Diurnal activity patterns of elephant in the Sengwa
E. Afr. Wildl. J. 14: 285 - 295.
HALL-MARTIN, AoJo 1972. Aspects of the plant ecology of the Lengwe
National Park, Malawi. M.Sc. Thesis. University of Pretoria.
HAMMERBECK, EoCoio 1977. Ushuswana complex in South-eastern Transvaal,
with special reference to its economic potential. DoSe. Thesis.
University of Pretoria.
HANKS, Jo, M.So PRICE & R.W. WRANGHAM. 1969. Some aspects of the defassa
waterbuck (Kobus defassa) in Zambia. Mammalia 33: 471 - 494.
+HALTENORTH, To 1963. Klassifikation der Saugetiere. Artiodactyla Handb.
Zool. 8 (13): 1 - 167.
+HEDIGER, H. 1934. Zur Biologie und Psychologie der Flucht bei Tieren.
Biol. Zbl. "Bd. 54 (1 & 2).
HEDIGER, Ho 1949. Saugetier-Territorien und ihre Markierungen. Bijdr.
Dierk. 28: 171 - 184.
HEDIGER, H. 1951. Observations sur la psychologie animale dans les
Pares Nationaux du Congo Belge. Institut des Pares Nationaux du Congo
Belge, Bruxelles.
HENDERSON, Mo & J.G. ANDERSON. 1966. Common weeds in South Africa.
Mem. bot. Surv. s. Afr. 37: 1 - 440. Government Printer, Pretoria.
+HENDRICHS, H. & Uo HENDRICHS. 1971. Freilanduntersuchungen zur Okologie
und Ethologie der Zwerg-Antilope Madoqua (Rhynchotragus) kirki (GUnther
1880). Pp. 9 - 75. In: Dikdik und Elefanten. Eds. H. Hendrichs & U.
Hendrichs. Piper, Munich.

259
HILDEBRAND, M. 1963. The use of motion pictures for the functional
analysis of vertebrate locomotion. Proc. XVl, Int. Cong. Zool.
3: 263 - 268.
HILDEBRAND, M. 1964. Cinematography for research on vertebrate
locomotion. Res. Film 5(1): 1 - 4.
HOFMANN, R.R. & D.RoM. STEWART. 1972. Grazer or browser: a classifica-
tion based on the stomach structure and feeding habits of East African
ruminants. Mammalia 36: 226 - 240.
HOFMEYR, JoM. 1970. A review on the food preferences and feeding habits
of some indigenous herbivores in the Ethiopian faunal region and some
studies on animal: plant relationships. Proc. s. Afr. Soc. Anim.
Prod. 9: 89 - 99.
HOOD, R.E. & J.M. INGLIS. 1974. Behavioral responses of white-tailed
deer to intensive ranching operations. J. Wildl. Mgmt. 38: 488 - 498.
HUMPHREY, W.A. & L.J. KRIGE. 1931. The geology of the country south
of Piet Retief. Explan. Sheet 68. Geol. Surv. s. Afr. Department of
Mining. Government Printer, Pretoria.
INNIS, A.C. 1958. The behaviour of the giraffe, Giraffa camelopardalis,
in the Eastern Transvaal. Proc. zool. Soc. Lond. 131: 245 - 278.
IRBY, L.R. 1973. A preliminary report on the mountain reedbuck (Redunca
fulvorufula) in the Loskop Dam Nature Reserve. J. South. Afr. Wildl.
Manage. Assoc. 3: 53 - 58.
JARMAN, P.Jo 1974.
their ecology.
The social organisation of antelope in relation to
Behaviour 48: 215 - 267.
JARMAN, M. V. & P .J. JARMAN.
11: 75 - 92.
1973. Daily activity of impala. E. Afr. Wildl. J.
+JENNISON, G. 1927. Table of gestation periods and number of young. Ao
and C. Black Ltd., London.
JOHNSON, R.P. 1973. Scent marking in animals. Anim. Behav. 21: 521 - 535o

260
JOUBERT, Ee 1971. Ecology, behaviour and population dynamics of the
Hartmann zebra Eguus zebra hartmannae Matschie, 1898 in South West
Africa. D.Sc. Thesis. University of Pretoria.
JOUBERT, E. 1972. Activity patterns shown by mountain zebra Eguus
zebra hartmannae in South West Africa with reference to climatic
factors. Zool. Afr. 7: 309 - 331.
JOUBERT, E. & P.K.N. MOSTERT. 1975. Distribution patterns and status
of some mammals in South West Africa. Madogua 9: 5 - 44.
JUNGIUS, H. 1971. The biology and behaviour of the reedbuck (Redunca
arundinum Boddaert 1785) in the Kruger National Park. Mammalia
Depicta - Paul Parey, Hamburg and Berlin.
KEAST, A. 1968. The response of herbivores to vegetation. Pp. 258 - 264.
In: A practical guide to the study of the productivity of large
herbivores. Eds. FoB. Golley & H.K. Buechner. IBP Handbook No. 7.
Blackwell, Oxford.
KILLICK, D.JoB. 1963. An account of the plant ecology of the Cathedral
Peak Area of the Natal Drakensberg. Mem. Bot. Surv. S. Afr. 34: 1 - 178.
Government Printer, Pretoria.
KING, LoC. 1962. The morphology of the earth. Oliver and Boyd, Edinburgh.
KING, L.C. 1963. South African scenery. Third edition. Oliver and Boyd,
Edinburgh.
KITCHEN, D.W. 1974. Social behavior and ecology of the pronghorn.
Wildl. Monogr. Chestertown 38.
KNOBEL, R. 1958. Present day status of certain ungulates in the Union
of South Africa. Mammalia 22 : 498 - 503.
KOK, OoB. 1975. Gedrag en eko1ogie van die rooihartebees (Alcelaphus
buselaphus caama). Alg. publikasie nr. 4. Natuurbewaring, Prov.
Admin. O.V.So

261
KRUUK, H. 1972. The Spotted Hyaena. University of Chicago Press,
Chicago and London.
KUCHLER, A.W. 1967. Vegetation mapping. The Ronald Press Co.,
New York.
LAMPREY, H.F. 1963. Ecological seperation of the large mammal
species in the Tarangire Game Reserve, Tanganyika. ~~ Afr. Wildl. J.
1: 63 - 92.
LE RICHE, M. 1970. Birth of an oribi. Africana 4(4): 40- 42.
LEUTHOLD, W. 1977. African ungulates. A comparative review of their
ethology and behavioural ecology. Springer-Verlag, Berlin, Heidel
berg and New York.
LEYHAUSEN, P. 1965. The communal organization of solitary mammals.
Symp. zool. Soc. Lond. 14: 249 - 263.
LOXTON, R.F. 1966. A simplified soil survey procedure for farm
planning. Sci. Bull. Dept. Agr. Tech. Serv. 383. Government
Printer, Pretoria.
LYNCH, C.D. 1974. A behavioral study of the blesbok, Damaliscus
dorcas phillipsi, with special reference to territoriality.
Navors. nas. Mus., Bloemfontein 8: 1- 83.
MALOIY, G.M.O" 1973. The water metabolism of a small East African
antelope, the dik-dik. Proc. R. Soc. 184: 167 - 178.
MANNING, A. 1970. An introduction to animal behaviour. Edward Arnold
(Publishers) Ltd., London.
MARASCUILO, L.A. 1966.
65: 280 - 290.
Large-sample multiple comparisons. Psychol. Bull.
MASON, D.R. 1973. Range use and population structure of ungulates in
the Jack Scott Nature Reserve. MeSc. Thesis. University of Pretoria.
MENTIS, MoT. 1972. A review of some life history features of the large
herbivores of Africa. The Lammergeyer 76: 1 - 89.

262
MENTIS, M.To 1974 Distribution of some wild animals in Natal.
Lammergeyer 20: 1 - 68.
MILLAR, J.C.Go 1970. The past and present numerical status of the
oribi Ourebia ourebi (Zimmerman) in the Cape Province. Dept. of
Nature Conservation Investigational Report no. 17.
of Cape of Good Hope.
Prov. Admin.
MONFORT, A. & N. MONFORT. 1974. Notes sur L'Ecologie et le Comporte
ment des Oribis (Ourebia ourebi, Zimmerman, 1783). Terre Vie 28(2):
169 - 208.
+MORAVEC, J. 1969. Die Vorbedingungen der Pflazensoziologiegischen
Nomenklaturregeln. Acta bot. croat. 28: 259 - 265.
MORRISON, J.A. & E.W. MENZEL JR. 1972. Adaptation of a free-ranging
rhesus monkey group to division and transplantation. Wildl. Monogr.
Chestertown 31.
MULLER, P.Jo, M.JoAo WERGER, B.J. COETZEE, D. EDWARDS & NoG. JARMAN. 1972.
An apparatus for facilitating the manual tabulation of phytosociologi
cal data. Bothalia 10: 579 - 581.
+MYKYTOWYCZ, R. 1955. The behavioural role of the mammalian skin glands.
Naturwissenschaften 59: 133 - 139.
NORTON, P.M. 1980. The habitat and feeding ecology of the klipspringer
Oreotragus oreotragus (Zimmermann, 1783) in two areas of the Cape
Province. M.Sc. Thesis. University of Pretoria.
NORTON-GRIFFITHS, M. 1975. Counting animals. AWFL-handbook no. 1.
Serengeti Ecological Monitoring Programme, Nairobi, Kenya.
NOVELLI, P.A. 1975. Comparative social behaviour of springbok, Antidorcas
m. marsupialis (Zimmermann, 1780), and blesbok, Damaliscus dorcas
phillipsi Harper 1939, on the Jack Scott Nature Reserve, Transvaal.
M.Sc. Thesis. University of Pretoria.
OATES, LoGo 1972. A note on the sex ratio of steenbok Rhaphicerus campestris
in Transvaal Lowveld Mopane Colophospermum mopane Woodland. Koedoe 15:
141 - 142.

263
OLIVER, M.D.N., NoR.M. SHORT & J. HANKS. 1978., Population ecology of
oribi, grey rhebuck and mountain reedbuck in Highmoor State Forest
Land. s. Afr. J. Wildl. Res. 8: 95 - 106.
PALGRAVE, KoC. 1977. Trees of Southern Africa. C. Struik Publishers,
Cape Town and Johannesburg.
PENZHORN, B.L. 197la. A note on the sex ratio of steenbok Raphicerus
campestris in the Kalahari Gemsbok National Park. Koedoe 14: 61 - 64.
PENZHORN, B.L. 1971b. A summary of the re-introduction of ungulates
into South African National Parks (to 31 December 1970) Koedoe 14:
145 - 159.
+PETRIDES, G.A.
in Nigeria.
tee tion.
1965. Advisory report on wildlife and national parks
American Committee for International Wildlife Pro-
PIENAAR, U. DE V. 1963. The large mammals of the Kruger National Park
their distribution and present-day status. Koedoe 6: 1 - 37.
PIENAAR, Uo DE V. 1974. Habitat-preference in South African antelope
species and its significance in natural and practical distribution
patterns. Koedoe 17: 185 - 195.
PIENAAR, U. DE V. & J.Wo VAN NIEKERK. 1963. The capture and transloca-
tion of three species of wild ungulates in the Eastern Transvaal with
special reference to RO 5-2807/B-SF (Roche) .as a tranquilizer in game
animals. Koedoe 6: 83 - 90.
POCOCK, R.I. 1910. On the specialized cutaneous glands of ruminants.
Proc. zool. Soc. Lond. : 840 - 986.
RALLS, Ko 1971.
443 - 449.
Mammalial scent marking. Science NoYo 171 (3970):
+REMANE, Ao 1960. Das soziale Leben der Tiere. Rowohlts deutsche
Enzyklopadie 97.
RINEY, T. & W.L. KETTLITZ. 1964. Management of large mammals in the
Transvaal. Mammalia 28: 189 - 248.

264
ROBERTS, A. 1954. The mammals of South Africa. Second Edition.
Central News Agency, Johannesburg.
+ROBERTSE, NoJo 1972.
Suidoos-Transvaal.
Pretoria.
Die doeltreffendheid van landbouvoorligting in
D.Sc. (Agric.)-tesis. Universiteit van
ROBINETTE, W.L. 1970. Game animal surveys. Joint WHO/FAO African
Trypanosomiasis Information Service, Segments FIJ.
ROLLINSON, DoH.I., K.Wo HARKER & J.I. TAYLOR. 1956. Studies on the
habits of zebu. cattle. IV. Errors associated with recording
technique.. J. agric. Sci., Camb. 47: 1 - 5.
ROOS, JoH., NoF.Go RETHMAN & GoDo KOTZE. 1973. Preliminary results on
species selection by animals on sour grassveld.
Proc. Grassld Soc. Sth. Afr. 8: 77 - 81.
ROOSEVELT, T. & E. HELLER. 1914. Life histories of African game mammals.
Vol. II. John Murray, London.
ROWE-ROWE, D.T. 1974. Flight behaviour and flight distances of blesbok.
z. Tierpsychol. 34 : 208 - 211.
SCHEEPERS, JoC. 1975. The plant ecology of the Kroonstad and Bethlehem
areas of the Highveld agricultural region. Vol. 1 and 2. D.Sc.
Thesis. University of Pretoria.
SCHENKEL, Ro 1966. On sociology and behaviour in impala (Aepyceros
melampus suara Matschie). z. Tierpsychol. 31: 177 - 205.
SCHMIDT-NIELSEN, Ko 1964. Desert animals, physiological problems of
heat and water. Clarendon Press, Oxford.
1930 .. Das Flehmen. I. Zool. Gart. 3: 183 - 198.
SCHOEN, A. 1972. Studies on the environmental physiology of a semi
desert antelope, the dik-dik· E. Afr. agric. For J. 40: 325 - 330.
SCHULZE, B.Ro
W.B. 28.
1965. Klirnaat van Suid-Afrika. Deel 8. Algernene oorsig.
Staatsdrukker, Pretoria.

265
SCLATER, W.L. 1900. The mammals of South Africa. Vol. 1. R.H. Porter, London.
SHORTRIDGE, GaCo 1934. The mammals of South West Africa. Vol. II.
Wm. Heinemann Ltd., London.
SKINNER, JoD., N. FAIRALL & Jo DU P. BOTHMAo 1977. South African Red
Data Book - Large Mammals. S.A. National Scientific Programmes
Report No. 18. CSIR, Pretoria.
SMITHERS, RaH.N. 1971. The mammals of Botswana. Mus. Memoir Nat.
Mus. Rhod. 4, Salisbury.
"" SMITHERS, RoH.N. & JoLoP. LOBAO TELLO. 1976. Check list and atlas of
the mammals of Mocambique.
SMITHERS, R.H.N. & VoJo WILSON.
mammals of Zimbabwe Rhodesia.
Salisbury.
Mus. Memoir Nat. Mus. Rhod. 8, Salisbury.
1979. Check list and atlas of the
Mus. Memoir Nat. Mus. Rhod. 9,
SMYTHE, N. 1970. On the existance of "persuit invitation" signals in
mammals. Am. Nat. 104: 491 - 494.
SOKAL, RoRo & FoJ. ROHLF. 1969. Biometry. W.H. Freeman and Co.,
San Francisco.
SPINAGE, CoAo 1968. A quantitative study of the daily activity of
the Uganda defassa waterbuck. E. Afr. Wildl. J. 6: 89 - 93.
+SPINAGE, CoAo 1969. Les Pares Nationaux du Rwanda. Rapport ecologique.
Ministere Britannique du Developpement d' Outre-mer, Londres.
+STEINHARDT, J. 1921. Von Wehrhaften Risen und Seinem Reiche. Alster
Verlag, Hamburg.
STEVENSON-HAMILTON, Jo 1929. The Lowveld - its wild life and people.
Cassel & Co., London.
STEWART, DoR.Mo & Jo STEWART. 1963. The distribution of some large
mammals in Kenya. Jl E. Africa nat. Hist. Soc. 24(3)107: 1 - 52.

266
+STEWART, G.A., R.A. PERRY, D.Mo TRAVES, S.J. PATTERSON, R.O.
SLAYTER, P.R. DUNN, P.J. JONES & J.R. SLEEMANo 1970. Lands
of the Crt-Victoria area, W.A. and N.T.
Res. Ser. no. 28. CSIRO Aust. Land
STOKER, D.Jo
stad •
.. STRYBIS, CoBo
1977.
1974.
Statistiese tabelle. Academica, Pretoria en Kaap-
More oribi for Golden Gate. Custos 3(11): 10.
TAINTON, NoM., Dolo BRANSBY & P. DE Vo BOOYSEN. 1976. Common veld and
pasture grasses of Natal. Shuter & Shooter, Pietermaritzburg.
TAIT, W.J.Eo 1969.
154 - 160.
Elusive and rare oribi. Afr. Wild Life 23(2):
TALBOT, L.Mo & M.H. TALBOTo 1963. The wildebeest in western Masailand,
East Africa.Wildl. Monogr. Chestertown. 12.
TAYLOR, CoR. 1968. Hygroscopic food: a source of water for desert
antelopes? Nature, Land. 219: 181 - 182.
THERON, G.Ko 1973. n Ekologiese studie van die plantegroei van die
Loskopdam-natuurreservaat. DoSc.-proefskrif. Universiteit van
Pretoriao
THOMSON, P.J. 1973. Notes on the oribi (Mammalia, Bovidae) in
Rhodesia. Arnoldia Rhod. 6(21): 1 - 5.
THOMSON, WoR. 1975.
cover density.
A photographic technique to quantify lateral
J. South Afr. Wildl. Manage. Assoc. 5: 75 - 78.
+THUNBERG, C~P. 1795. Travels in Europe, Africa and Asia made between
the years 1770 and 1779. Second edition. Fo and C. Rivington,
London.
TINBERGEN, N. 1940. Die Ubersprungsbewegung. z. Tierpsychol. 4: 1 - 40.

267
TINLEY, K.L. 1969. Dik-dik, Madogua kirki in South West Africa:
notes on distribution, ecology and behaviour. Madogua 1: 7 - 33.
TINLEY, K.L. 1977. Framework of the Gorongosa ecosystem.
(Wildl. Mgmt.) Thesis. University of Pretoria.
D.Sc.
TREWARTHA, G.T. 1954. An introduction to clirnat~. McGraw-Hill Book
Co., Inc., New York.
+VALENTINE, K.W.G. 1970. Use of land classification concepts as a basis
for a reconnaissance soil survey in northern British Columbia.
Can. J. Soil Sci. 50: 71 - 77.
VAN LAVIEREN, LoP. & J.D. ESSER. 1979. Numbers, distribution and habitat
preference of large mammals in Bouba Ndjida National Park, Cameroon.
Afr. J. Ecol. ~1: 141 - 153. If
VAN ZYL, J.HoMo 1965. The vegetation of the SoAo Lombard Nature Reserve
and its utilization by certain antelope. Zool. Afr. 1: 55 - 72.
VINCENT, J. 1962. The distribution of ungulates in Natal. Ann. Cape
Provo Mus. 2: 110 - 117.
VON RICHTER, W. 1971. Survey of rare and endagered mammal species in
South Africa.
Group.
Project Report CT (2): Fauna Survey IBP/LT Working
VON RICHTER, Wo 1972.
Connochaetes gnou.
VON RICHTER, Wo 1974.
Territorial behaviour of the black wildebeest,
Zool. Afr. 7: 207 - 231.
Survey of the adequacy of existing conserved
areas in relation to wild animal species. Koedoe 17: 39 - 69.
VON RICHTER, W.CoDo LYNCH & T. WESSELSo 1972. Status and distribution
of the larger mammal species on farmland in the Orange Free State.
Nature Conserv. Research Report No. 1. O.F.S. Provincial Administra-
tion.
VON WILHELM, W.H. 1933 Das wild des Okawangogebietes und des Caprivizipfels.
JaS.Wo Africa Scient. Soc. 6: 51 - 74.

268
WALTER, H. 1971. Ecology of tropical and subtropical vegetation.
Oliver & Boyd, Edinburgh.
WALTHER, Fo 1958.
z. Tierpsychol.
Zum Kampf- und Paarungsverhalten einiger Antilopen.
15: 340 - 380.
WALTHER, F. 1964a. Verhaltensstudien an der Gattung Tragelaphus de
Blainville (1816) in Gefangenschaft, unter besonderer Berlicksichtigung
des Sozialverhaltens. z. Tierpsychol. 21: 393 - 467.
WALTHER, Fo 1964b. Einige Verhaltungsbeobachtungen an Thomsonsgazellen
(Gazella thomsoni GUnther, 1884) im Ngorongoro-Krater. Z. Tierpsychol.
21: 871 - 890.
WALTHER, F. 1965. Verhaltensstudien an der Grantgazelle (Gazella granti
Brooke, 1872) im Ngorongoro-Krater. z. Tierpsychol. 22: 167 - 208.
WALTHER, Fo 1969. Flight behaviour and avoidance of predators in Thomson's
gazelle Gazella thomsoni Gunther 1884. Behaviour 34: 184 - 221.
WALTHER, F. 1974. Some reflections on expressive behaviour in combats
and courtship of certain horned ungulates. Pp. 56 - 106. In: The
behaviour of ungulates and its relation to management. Eds. V. Geist
and F. Walther. IUCN Publ. New Series 24, Merges Switzerland.
WASER, Pa 1975. Diurnal and nocturnal strategies of the bushbuck
Tragelaphus scriptus (Pallas). E. Afr. Wildl. J. 13: 49 - 63.
WEERBURO. 1954. Klimaat van Suid-Afrika. Deel 1. Klimaatstatistieke.
WoB• 19. Staatsdrukker, Pretoria.
WEERBURO. 1965. Klimaat van Suid-Afrika. Deel 9. Gemiddelde
maandelikse reenval tot einde 1960. W.B. 29. Staatsdrukker, Pretoria.
WEERBURO. 1967a. Report on meteorological data of the year 1965.
Government Printer, Pretoria.
WEERBUROo 1967b. Report on meteorological data of the year 1966.
Government Printer, Pretoria.
WEINMANN, H. 1955. The chemistry and physiology of grasses. Pp. 5 71 -
600. In: The grasses and pastures of South Africa.
Central News Agency, Johannesburg.
Ed. D. Meredith.

269
WELLINGTON, JoH. 1955. Southern Africa. Vol. 1. Cambridge University
Press, Cambridge.
WERGER, M.J.A. 1973. Phytosociology of the upper Orange River Valley,
South Africa: a syntaxonomical and synecological study. Doctors
proefschrift. De Katholieke Universiteit te Nijmegen, Nederland.
WERGER, MoJ.A. 1974a.
vegetation science.
109.
The place of the Zurich-Montpellier method in
Folia Geobot. & Phytotax Bohemoslov. 9: 99 -
WERGER, M.J.A. 1974b. On concepts and techniques applied in the
Zurich-Montpellier method of vegetation survey. Bothalia 11:
309 - 323.
+WIGHT, H.Mo 1938. Field and laboratory technics in wildlife management.
University of Michigan. Ann Arbor.
WILSON, D.Eo 1975. Factors affecting roan and sable antelope popula-
tions on nature reserves in the Transvaal with particular reference
to ecophysiological aspects. D.Sc. Thesis. University of Pretoria.
WYATT, JoR. & SoK. ELTRINGHAM. 1974. The daily activity of the elephant
in the Rwenzori National Park, Uganda. E. Afr. Wildl. J. 12: 273 - 289.
·WYNNE-EDWARDS, VoCo 1962. Animal dispersion in relation to social be
haviour. Oliver and Boyd, London.
YOUNG, E. 1970. Water as n faktor in die ekologie van wild in die
Nasionale Krugerwildtuin. M.Sc./D.Sc. (Natuurbeheer)-verhandeling.
Universiteit van Pretoria.
ZAGATHA, M.D. & A.Oo HAUGEN. 1973. Winter movement and home range of
white-tailed deer at Pilot Knob State Park, Iowa. Proc. Iowa Acad.
~· 79: 74 - 78.
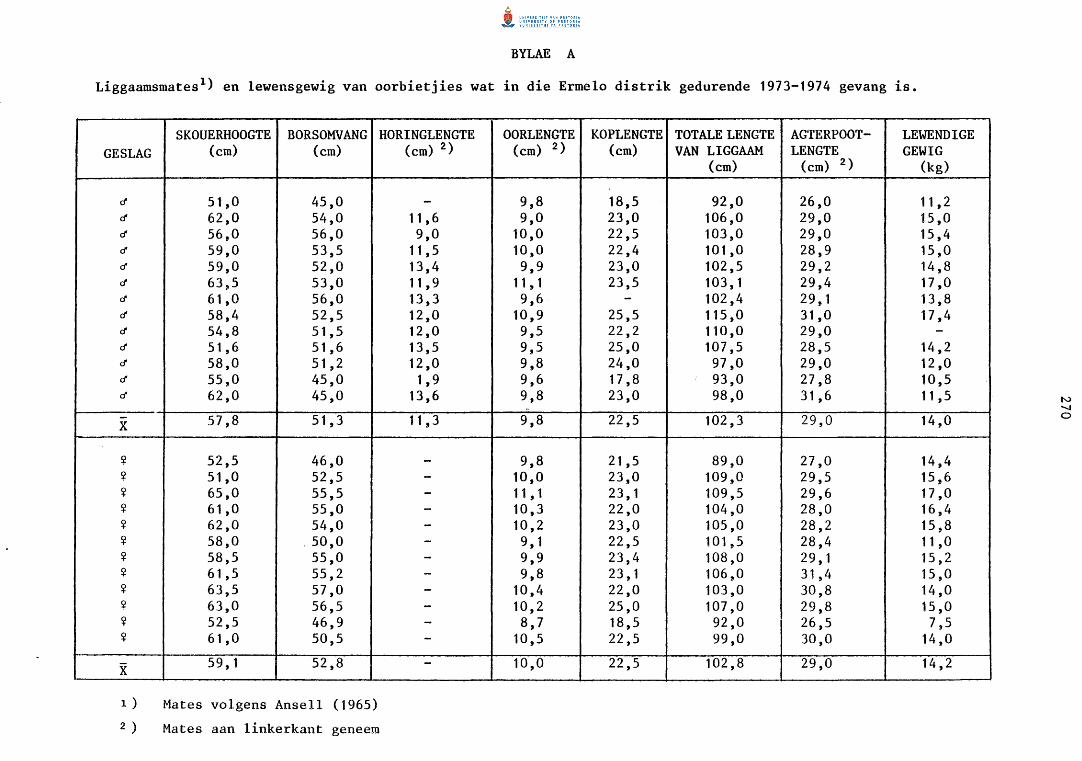
BYLAE A
Liggaamsmates 1 ) en lewensgewig van oorbietjies wat in die Ermelo distrik gedurende 1973-1974 gevang is.
SKOUERHOOGTE BORSOMVANG HORINGLENGTE GESLAG (em) (em)
cJ 51,0 45,0 cJ 62,0 54,0 cJ 56,0 56,0 cJ 59,0 53,5 cJ 59,0 52,0 cJ 63,5 53,0 cJ 61,0 56,0 cJ 58,4 52,5 cJ 54,8 51 ,5 cJ 51 ,6 51 ,6 cJ 58,0 51 ,2 cJ 55,0 45,0 cf 62,0 45,0
X 57,8 51,3
~ 52,5 46,0 ~ 51 ,0 52,5 ~ 65,0 55,5 ~ 61 ,0 55,0 ~ 62,0 54,0 ~ 58,0 . 50,0 ~ 58,5 55,0 ~ 61,5 55,2 ~ 63,5 57,0 ~ 63,0 56,5 ~ 52,5 46,9 ~ 61 ,o 50,5
x 59,1 52,8
1) Mates volgens Ansell (1965)
2 ) Mates aan linkerkant geneem
(em) 2 )
-11,6 9,0
11 '5 13,4 11 , 9 13,3 12,0 12,0 13,5 12,0 1,9
13,6
11 '3
-------------
OORLENGTE (em) 2 )
9,8 9,0
10,0 10,0 9,9
11 ' 1 9,6
10,9 9,5 9,5 9,8 9,6 9,8
,,
9,8
9,8 10,0 11 ' 1 10,3 10,2 9, 1 9,9 9,8
10,4 10,2 8,7
10,5
10,0
KOPLENGTE TOTALE LENGTE AGTERPOOT- LEWENDIGE (em) VAN LIGGAAM LEN GTE GEWIG
(em) (em) 2 ) (kg)
18,5 92,0 26,0 11 '2 23,0 106,0 29,0 15,0 22,5 103,0 29,0 15,4 22,4 101 ,0 28,9 15,0 23,0 102,5 29,2 14,8 23,5 103' 1 29,4 17,0
- 102,4 29' 1 13,8 25,5 115,0 31 ,0 17,4 22,2 110 ,o 29,0 -25,0 107,5 28,5 14,2 24,0 97,0 29,0 12,0 17,8 93,0 27,8 10,5 23,0 98,0 31,6 11 , 5
22,5 102,3 29,0 14,0
21 ,5 89,0 27,0 14,4 23,0 109,0 29,5 15,6 23, 1 109,5 29,6 17,0 22,0 104,0 28,0 16,4 23,0 105,0 28,2 15,8 22,5 101,5 28,4 11 '0 23,4 108,0 29, 1 15,2 23' 1 106,0 31,4 15,0 22,0 103,0 30,8 14,0 25,0 107,0 29,8 15,0 18,5 92,0 26,5 7,5 22,5 99,0 30,0 14,0
22,5 102,8 29,0 14,2
I
N ........ 0
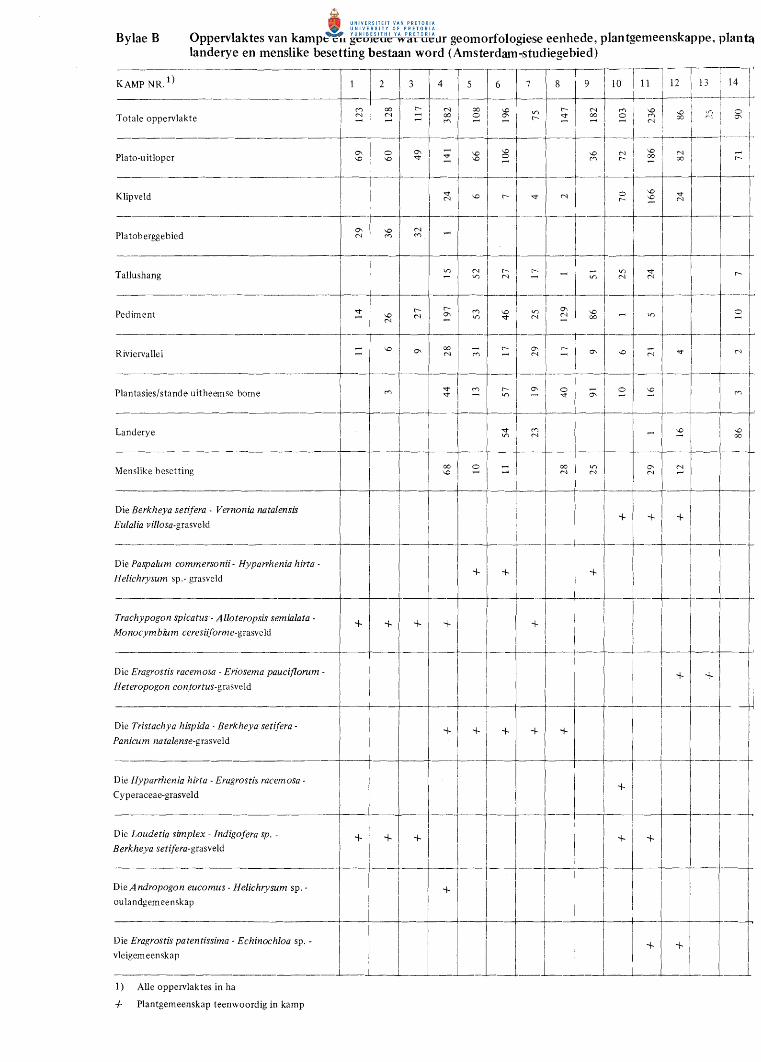
Bylae B Oppervlaktes van kampe en gebiede wat deur geomorfologiese eenhede, plantgemeenskappe, plant~ landerye en menslike besetting bestaan word (Amsterdam-studiegebied)
KAMPNR.l) "-12 ~ 5 16 vrl 8 -19l~~T~ ~I --------------+-1 M-~ 00-+1-~T--;;-I~i---::t-V>- r- N ~--:-t- \0 : "~1 0 i
Tot ale oppervlakte i ~ ~ 1 :::: I ~ 1 S I ;:: I r- 1 :::: I ::S \ S I ~ ( 00 i ~ 1 1 a- :
-------------+-:-~ ~-+--1 -:,t-=~1 \0 r-=-r-r-r--:-+1 --=-t~-,-:~-r~-t Plato-uitloper -o -o 71 ;::: -o I S L I I M r- I ::S \ 00 1 1 r- ;
-+-t- ~-r--r-~ -+---1----~-~+ --+--t-t K!ipveld I I I ;:!; I -o r- I 7 I N I ;2 ~ I ;:!; I I I
------------- -=-~-~ ~ 8--r I --+ -t-+--t- -=-t-t--~-t Platoberggebied N M I M I -1 1 I I I 1 I 1 I / I
T------------- ~- t1V> I ~i--=-t-:t
1
-+-:-t--~1-:t--i-i--t allushang L I l V> t- N I - -J V> 1 N 1 N L 1 r- 1
Pedime-nt -------r ~I :t;r E I ;1-;r- ~ t ~ 1--;r--=-t-=-1 ~-~ s1 ----------+-::+-:-r-l: +-:.- j--:-r-:-+--t-t-:+---+-t----t Riviervallei I -±~ I ~ I N I ~ - N I - I ~ I ~ I N I " I I N I -------------+- -t-+-r--i-- --r----t--r--+--t---t-+--t Pbntosies/sbndeuitheemse borne I ~ I I ~ I ::: I :;; I ;': I ;;' I ;; I S I :0: I I I ~
Lan:=------------r-1 _l+--~-r ~- ~ rrt-~~rj-~~-r ~ r ______________ L_t I _L_+-+-t-i--+- +--LL_j--_l Menslike besotting I I I I ~ I S I = I I ~ I ~ I I ~ I ~ I I I
T-f---r-+-t-T-i-t--r-·-i--r--t--t---~ ~~;.~::~~::,:.:::~:;/ emonW nata/ens~ I L I I I I I I I ~ l ~ I ~ f t ; ~: Paspalu: commnsonU. Hypan"henia hir:+ I --j-1 J-+-r~t~r-~1 -r:-Lt--I -+II --~---~--r Helzchrysum sp.- grasveld I 1 1 I 1 I )
1
I I
-----------1+-1 _i_-t _ _j_~-i---~ -+---t-i--+---t Trachypog~n spicatu~.- A lloteropsis semialata - +-+- I -+- .,._ I .,._ I I ~~ .,._I I \ ! . \ [ \i I
Mo:cymbTUm cer:ufo~e-grasveld_____ -lf-J--i-t-- -~--~--+J-+--+-~ Die Eragrostis racemosa - Eriosema pauciflorum - I I 1 I I t tl ~~ I I I -+-II -+- I I Heteropogon con.tortus-grasveld i 1 I I I I \ I ( I: ------------ ---+--i-+--+--+-+-r--t-+--t--t---t--i- -f Die .Tristachya hispida- Berkheya setifera- Dl II I -+- I -+- I -+- I -+-
1
1 -+- I I \ I I I I Panzcum natalense-grasveld _j i I J I I I 1 1 1 1 I -------------- -+ ~-~--t-- -t--+-+-r--r-t--t--i Die Hyparrhenia hirta- Eragrostis racemosa- I I I I i 1
1 I I l I I I Cyperaceae-grasveld 1 I t I I I I I I -+- I \ t I I --------------1---r- --t-~ t-+-t-- I +-+-t-- -+-t Die Loudetia simplex - !ndigofera sp. -\I -+- -+- I -+- I r I I I \i I -+- I -+- I I I I
Berkheya setifera-grasveld I I 1
\ I 1 I 1
J I I I I
----------1- ~--t-1--r---t---t---t--+--- -t-T-+--r DieAndropogon eucomus- Helichrysum sp.- Jl I 1 -+- I I I I
11 I I 11 I II
oulandgemeenskap I I I I I 1 I I
-------------+- --t--+-__j I -+-i--r1--t-t-i---t--+ ~:~g~::~:~::•tentissima · Echinochloa sp · I I I I iJ I I I ~ I ~1 I I ______________ j__L_l_j __ L_ _l_ ___ _J_ -k- _1 _ _1 1) Aile oppervlaktes in ha
+ Plantgemeenskap teenwoordig in kamp
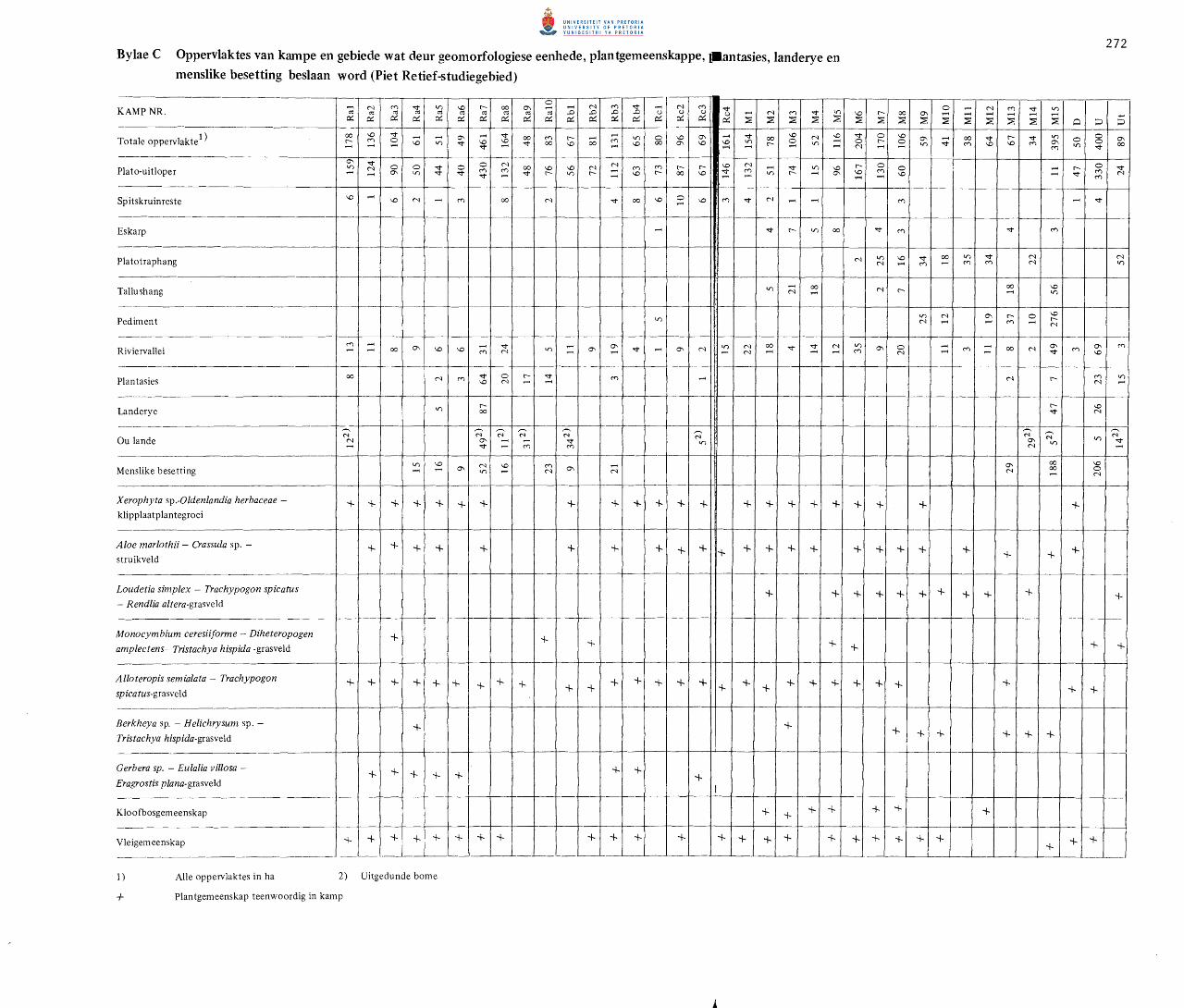
Bylae C Oppervlaktes van kampe en gebiede wat deur geomorfologiese eenhede, plantgemeenskappe, •antasies, landerye en
menslike besetting beslaan word (Piet Retief-studiegebied)
KAMP NR.
------
Totale oppervlakte1)
Plato-uitloper
------------
Spitskruinreste
--------
Eskarp
--
Platotraphang
Tallushang
Pediment
Riviervallei
Plantasies
l.anderye --
Ou lande
--
Mcnslike besetting
Xerophyta sp.-Oldenlandia herbac eae-
klipplaatplantegroei
Aloe marlothii- Crassula sp. -
struikveld
---------------
Loudetia simplex - Trachypogon
- Rendlia altera-grasveld
spicatus
-------------
Monocymbium ceresiiforme - Dih
amplectens- Tristachya hispida -gr
eteropogen
as veld
--
--
-----
Alloteropis semialata - Trachypog
spica tus-grasvcld
Berkheya sp. - Helichrysum sp.
Tristachya hispida-grasveld
Gerbera sp. - Eulalia villosa -
Eragrostis plana-grasveld --------------
Kloofbosgemeenskap
Vleigemeenskap
on
Alle oppervlaktes in ha
-----
r-· ....... N M 1 o.n ell ell ell ell
0:: 0:: 0:: 0:: 0:: -
00 \0 ~ ....... ....... r- M 0 \C) o.n ...... ....... .......
0'1 ~ o.n N 0 0
""" ....... ....... 0'1 o.n """
\C) ....... \0 N .......
- t----
- r-- - ,---
M -....... ....... 00 0\ \C)
1---- - ----00 N
r- - r--- --o.n
,.-._ t---- - t----('I
N ...... - ,---
o.n \C)
....... .......
.,._ .,._ .,._ .,._ .,._
.,._ .,._ .,._ .,._
.,._
.,._ .,._ .,._ .,._ .,._
.,._
t----
.,._ .,._ .,._ .,._
1---·
.,._ .,._ .,._ .,._ .,._
--'-----
2) Uitgedunde borne 1)
+ Plantgemeenskap teenwoordig in kamp
\C) r- 00 ell ell ell
0:: 0:: 0::
0\ ....... ~ \0 \C)
~ ~ .......
0 N 0 M M ~ ~ .......
M 00
- """ \0 M N
1--
M ~ 0 \C) N
r-00
- -('I ('I
0\ ....... ~ .......
0\ N \C)
o.n .......
.,._ .,._
.,._
.,._ .,._ .,._
.,._
.,._ .,._ .,._
L-_1_
0 ,.----~-~ 0'1 .......
~ N M -.:t ...-\ N M ~ o.n ell ell .D .D .D () () () () ....... N M ~
0:: 0:: 0:: 0:: 0:: 0:: 0:: 0:: 0:: 0:: :::E :::E :::E :::E :::E 1--- · ...
00 M r- ....... ....... o.n 0 \C) 0'1 ....... ~ 00 \C) N ~
~ 00 00 M \C) 00 0'1 \C) \0 o.n r- s o.n \C) ....... ....... ....... .......
t----1--N \C) N ....... 00 \C) \C) N ....... M M r- r-
::!; M ~ o.n \C)
""" r- o.n r- ....... \0 r- 00 \C) ....... o.n r- ....... 0'1
N ~ 00 \C) 0 ....... \C) M
""" N ....... .......
....... """
r- o.n 00
o.n ....... 00 N .......
t---
o.n
t---· ....... 0\ o.n N 00 ~ ~ N o.n ....... 0\ ....... """
....... 0'1 N ....... N ....... ....... .......
r- ~ M ....... ....... .......
- - ,.-._ ('I ('I ('I .......
""" o.n M M
....... M 0\ N N
r---r--
.,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._
1---f-·
.,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._
1----1--·
.,._ .,._
.,._ .,._ .,._
1--t---1----t--1---
.,._ .,._ .,._ .,._ .,._ .... .,._ .,._ .,._ .... .,._ .,._ .,._ .,._
.,._
1---f-·
.,._ .,._ .,._
I r---f-- 1---f-
.,._ .,._ .,.. .,._
1---1--.,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._
'---'- '--· --
272
0 ....... N M ~ o.n \0 r- 00 0'1 ....... ....... ....... ....... .......
~ 5 :::E :::E :::E :::E :::E :::E :::E :::E :::E 0 ;:::::>
~ 0 \0 0'1 ~ 00
""" r- ~ o.n 0 0 0'1 0 r- 0 o.n M \C) \0 M 0'1 o.n 0 00 N ....... ....... M
""" r- 0 'I ....... r- 0 ~ \C) M 0 M N ....... ....... \C) .......
""" M
M ....... ~
""" M ~ M
N o.n \C) ~ 00 o.n ~ N N N ....... M ....... M M N o.n
N r- 00 \C) ...... o.n
o.n N 0'1 r- 0 \C) r-N ....... ....... M ....... N
o.n 0'1 0 ....... M
....... 00 N 0'1 M 0'1 M M N ...... ....... ~ \0
N r- M o.n N .......
r- \C)
""" N
- - ,.-._ ('I ('I
('I o.n 0'1 o.n ~ N .......
00 \0 0\ N 00 0
....... N
r--
.,._ .,._ .,._ .,._
.,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._
.,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._ .,._
.,._ .,._ .,._
.,._ .... .,._ .,._ .,._ .,._
.,._ .,._ .,._ .,._ .,._ .,._
.,._ .,._ .,._
t--r--.,._ .,._ .,._ .,._ .,._ .,._ .,._
.,._

273
BYLAE D
Alfabetiese lys van die plantspesies wat in die Amsterdam- en Piet
Retief-studiegebiede geidentifiseer is.
Acacia mearnsii De Wild
Acalypha caperonioides
Acalypha schinzii Pax
Aloe marlothii Berg.
Baill.
Alloteropsis semialata (R. Br.) Hitch.
Andropogon appendiculatus Nees
Andropogon eucomus Nees
Aristida junciformis Trin. & Rupr.
Asparagus africanus Lam.
Athrixia phylicoides DC.
Berkheza echinacea Burtt Davy
Berkheza insisnis (Hans.) Thell.
Berkheia set if era DC.
Brachiaria serrata (Spreng) Stapf.
Calpurnia ~ (Ait.) Benth.
Cassia biensis (Steyaert) Medonca & Torre
Conyza bonariensis Cronquist
·crassula mollus Thunb.
Ctenium concinnum Nees
Cussonia spicata Thunbe
Cymbopogon excavatus (Hochst.) Stapf
Dalbergia obovata E. Meyer
Dicoma zeyheri Sand.
Digitaria tricholaenoides Stapf
Diheteropogon amplectens (Nees) W.D. Clayton

Diose.lros whlteana (Hi ern) F. White
El,lonurus muticus (Spring.) Kunt
Era~rostis caeensis (Thunb.)
Era~rostis curvula (Schrad.)
Eragrostis gummiflua Nees
Eragrostis patentissima Hack
Eragrostis £lana Nees
Trin.
Nees
Eragrostis racemosa (Thunb.) Stend.
Eriosema cordatum E. Mey
Eriosema pauciflorum Klotzsch
Eriosema salignum E. Mey.
Eulalia villosa (Thunb.) Nees
Euphorbia pulvinata Marl.
Faurea speciosa (Welw.) Welw.
~ capensis Thunb.
Gnaphalium luteo-album L.
Gnidia capitata L.f.
Haplocarpha scaposa Harv.
274
Harpechloa falx (L.) Kuntze
Helichrysum latifolium (Thunb.) Less.
Helichrysum nudiflorum (L.) Less.
Helichrysum polycladum Klatt
Helichrysum swynnertonii S. Moore
Heteropogon contortus (L.) Beauv.
Hyparrhenia ~ (Stapf) Stent
Hyparrhenia hirta (L.) Stapf
Hypoxis rigidula Bak.
Hypoxis rooperi S. Moore

Kalanchoe thyrsiflora Harv.
Koderia cristata (L.) Pers.
Leonotis dysophylla Benth.
275
Loudetia flavida (Stapf) C.E. Hubb.
Loudetia simplex (Nees) C.E. Hubb.
Microchloa caffra Nees
Monocymbium ceresiiforme (Nees) Stapf
Nidorella resedifolia DC.
Oldenlandia herbacea (L.) Roxb.
Panicum na~alense Hochst.
Panicum ecklonii Nees
Paspalum commersonii Lam.
Pearsonia sessiflora Duemmer
Pellaea calomelanos (Sw.) Link
Pellaea viridis (Forsk.) Prantl
Pelargonium luridum (Andr.) Sweet
Pentanisia angustifolia Hochst.
Phyllanthus glaucophyllus Sond.
Phyllanthus parvulus Sond.
Pteridium aquilinum (L.) Kuhn
Raphionacme hirsuta (E. Mey.) R.A. Dyer
Rendlia altera (Rendle) Chiov.
Rhamnus prinoides L'Herit
!Yll!!.. dentata Thunb.
~ eckloniana Sond.
!Yll!!.. disco lor E. Mey.
Rhus tranEvaalensis Engl.
Rhus pyroides Burch.
Rhynchelytrum repens (Willd.) C.E. Hubb.

276
Rhynchelytrum setifolium (Stapf) Chiov.
Schkuhria pinnata (Lam.) Kuntze
Schistostephium crataegifolium (DC • .) Fenzl
Senecio affinis DC.
Senecio hieracioides DC.
Setaria splendida Stapf
Setaria neglecta de Wit
Setaria nigrirostris (Nees) Dur. & Schinz
Sporobolus pectinatus Hark.
Stiburus conrathii Hack.
Syzygium cordatum Hochst.
Themeda triandra Forsk.
Trachypogon spicatus (L.f.) Kuntze
Tristachya hispida (L.f.) K. Schum.
Tristachya rehmannii Hack.
Vernonia natalensis Sch. Bip.
Vernonia poskeana Vatke & Hildebr.











![Die rol van Nederland in die transnasionale beweging van enkele Afrikaanse skrywers [Andre Brink, Breyten Breytenbach, Etienne van Heerden, Antjie Krog, Marlene van Niekerk]](https://static.fdokumen.com/doc/165x107/63193579e9c87e0c091003c7/die-rol-van-nederland-in-die-transnasionale-beweging-van-enkele-afrikaanse-skrywers.jpg)








