
Evolution of the central Catalan margin of the Valencia trough (western Mediterranean

Estuarine, Coastal and Shelf Science 57 (2003) 515–522
Denitrification in sediments of the LaurentianTrough, St. Lawrence Estuary, Quebec, Canada
Fenghai Wanga, S. Kim Junipera,*, Sılvia P. Pelegrıb, Stephen A. Mackoc
aDepartement des Sciences Biologiques, Geochronology and Geodynamics Research Centre (GEOTOP), Universite du Quebec a Montreal,
C.P. 8888, Succursale Centre-Ville, Montreal, Quebec, Canada H3C 3P8bLaboratoire d’Oceanographie et de Biogeochimie, UMR CNRS 6535, Campus de Luminy, Case 901, 163 Avenue de Luminy,
F-13288 Marseille, Cedex 9, FrancecDepartment of Environmental Sciences, University of Virginia, Clark Hall, Charlottesville, VA, 22903, USA
Received 19 July 2000; received in revised form 18 October 2002; accepted 22 October 2002
Abstract
The isotope pairing technique was used to obtain the first sedimentary denitrification rates for the St. Lawrence Estuary/Gulfsystem (Quebec, Canada). Denitrification rates were measured on intact sediment cores collected at three stations down the slope ofthe Laurentian Trough at 103, 200 and 320m depth. For samples collected in June 1997, total denitrification rates ranged from 1.8
to 3.3lmolNm�2 h�1 among the three stations. Only 3–5% of the NO3� consumed by the sediments was subsequently denitrified.
Nitrification was the major source of NO �3 for denitrification, representing 56–79% of total denitrification rates. Sediment con-
sumption of water column O2 (102–187lmolO2m�2 h�1) and NO �
3 (13.9–25.5NO �3 m�2 h�1) increased significantly downslope, as
did total denitrification rates. Denitrification made a negligible (<0.2%) contribution to carbon mineralization in our cores, but mayhave a greater impact on the nitrogen cycle. In addition, data suggest the presence of a major dissimilatory nitrate sink in thesesediments that far exceeds the requirements of denitrification. The latter observation is considered in terms of proposed interactions
between the nitrogen cycle and metal reduction/oxidation. We conclude by considering limitations to the extrapolation of these datato annual and regional scale nitrogen budgets for the deep waters of the St. Lawrence Estuary.� 2003 Elsevier Science B.V. All rights reserved.
Keywords: denitrification; nitrification; MnO2; isotope pairing technique; St. Lawrence Estuary
1. Introduction
In the marine environment, denitrification, the step-wise reduction of NO �
3 toN2, mainly occurs in sedimentsand is considered to be the major process removingnitrogen from the oceans (Seitzinger, 1988). Losses ofnitrogen through sedimentary denitrification may alterstoichiometric ratios of nutrient available to primaryproducers (Nixon, 1981) and contribute to the nitrogenlimitation commonly reported for marine systems(Ryther & Dunstan, 1971). Denitrification activity ismostly concentrated in estuaries, where there are largeterrestrial inputs of NO �
3 (Yeats, 1990) and organic
* Corresponding author.
E-mail address: [email protected] (S.K. Juniper).
0272-7714=03=$ - see front matter � 2003 Elsevier Science B.V. All rights
doi:10.1016/S0272-7714(02)00396-7
matter (Gearing & Pocklington, 1990). High rates oforganic matter decomposition in estuaries can depletesupplies of dissolved oxygen and cause anions such asNO �
3 to be used as terminal electron acceptors by hetero-trophic microbes.
The sub-arctic St. Lawrence Estuary is among theworld’s most intensively studied estuaries (e.g. El-Sabh& Silverberg, 1990). It is, in fact, a continuous systemthat includes several types of estuaries. The Lower orMaritime Estuary is more oceanic than most estuaries,because of its great dimensions and unimpeded con-nection with the Labrador Sea and slope waters of theAtlantic Ocean (El-Sabh & Silverberg, 1990). The sedi-mentation regime in this deep-water (max. depth 350m)estuary is characteristic of a coastal setting, while bottomwater conditions are more typical of the outer shelfand upper continental slope (El-Sabh & Silverberg,
reserved.

516 F. Wang et al. = Estuarine, Coastal and Shelf Science 57 (2003) 515–522
1990; Lavigne, Juniper, & Silverberg, 1997). A series ofstations along the floor of the Laurentian Trough havebeen the focus of many biological and biogeochemicalstudies as have coastal and intertidal sediments in thearea (Colombo, Silverberg, & Gearing, 1995a; Colombo,Silverberg, & Gearing, 1995b; Lavigne et al., 1997;Silverberg, Bakker, Edenborn, & Sundby, 1987). Thisstudy provides the first direct measurements of deni-trification in sediments of the Lower St. LawrenceEstuary. We used the isotope pairing technique (Nielsen,1992) to quantify denitrification at three differentstations situated over a ca. 220m depth range. Coor-dination of sample collection with a pore fluid geochem-istry study by Anschutz, Sundby, Lefrancois, Luther,and Mucci (2000) also permitted us to consider recentlyproposed interactions between the nitrogen and man-ganese cycles.
2. Materials and methods
2.1. Sampling location and strategy
Samples were collected using a 20� 30 cm2 box-corerin June 1997 during a research cruise on the R/V AlcideC. Horth in the Lower St. Lawrence Estuary (Fig. 1).Three sampling stations were selected, along the slope of
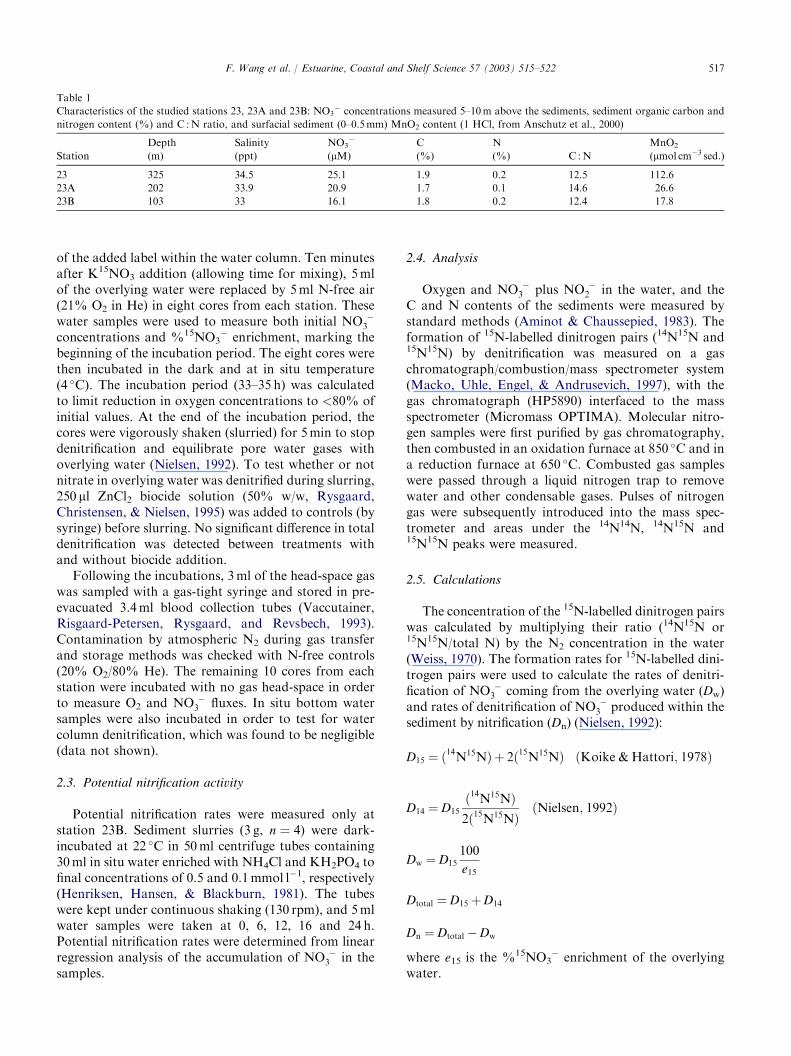
the Laurentian Trough. Station 23 was located on thefloor of the trough at a water depth of 325m, whilestations 23A and 23B were upslope on the southernshore at 202 and 103m depths, respectively (Table 1).The three stations showed very similar characteristicsexcept for surficial sediment MnO2 contents, which in-creased downslope from 17.8 to 112.6 lmolMnO2cm
�3
(Table 1). Once the box-corer was on deck, Plexiglassubcore tubes (12.8 cm long� 5 cm i.d.) were insertedthrough the overlying water into the sediment. Tubeswere then capped with rubber stoppers, removed fromthe box-core and refrigerated (Lamontagne & Valiela,1995). Cores from all three stations were collected thesame day and processed in a shore-based laboratoryduring the evening.
2.2. Main incubation
In the laboratory, the sediment column for 18 coresfrom each station was truncated at ca. 5 cm below thesediment–water interface, giving an overlying waterheight of ca. 8 cm. The overlying water was replacedby in situ bottom water, collected 5–10m above thesediments, and then enriched with K15NO3 (98 atom%15NO3
�, Aldrich) to final concentrations ranging from40 to 55 lMNO3
� (60.7–76.2% 15NO3�). A continuous,
gentle stirring system ensured homogeneous distribution
Fig. 1. Map of the study area—location of the stations 23, 23A and 23B.
517F. Wang et al. / Estuarine, Coastal and Shelf Science 57 (2003) 515–522
Table 1
Characteristics of the studied stations 23, 23A and 23B: NO3� concentrations measured 5–10m above the sediments, sediment organic carbon and
nitrogen content (%) and C :N ratio, and surfacial sediment (0–0.5mm) MnO2 content (1 HCl, from Anschutz et al., 2000)
Station
Depth
(m)
Salinity
(ppt)
NO3�
(lM)
C
(%)
N
(%) C :N
MnO2
(lmol cm�3 sed.)
23 325 34.5 25.1 1.9 0.2 12.5 112.6
23A 202 33.9 20.9 1.7 0.1 14.6 26.6
23B 103 33 16.1 1.8 0.2 12.4 17.8
of the added label within the water column. Ten minutesafter K15NO3 addition (allowing time for mixing), 5mlof the overlying water were replaced by 5ml N-free air(21% O2 in He) in eight cores from each station. Thesewater samples were used to measure both initial NO �
3
concentrations and %15NO3� enrichment, marking the
beginning of the incubation period. The eight cores werethen incubated in the dark and at in situ temperature(4 �C). The incubation period (33–35 h) was calculatedto limit reduction in oxygen concentrations to <80% ofinitial values. At the end of the incubation period, thecores were vigorously shaken (slurried) for 5min to stopdenitrification and equilibrate pore water gases withoverlying water (Nielsen, 1992). To test whether or notnitrate in overlying water was denitrified during slurring,250 ll ZnCl2 biocide solution (50% w/w, Rysgaard,Christensen, & Nielsen, 1995) was added to controls (bysyringe) before slurring. No significant difference in totaldenitrification was detected between treatments withand without biocide addition.
Following the incubations, 3ml of the head-space gaswas sampled with a gas-tight syringe and stored in pre-evacuated 3.4ml blood collection tubes (Vaccutainer,Risgaard-Petersen, Rysgaard, and Revsbech, 1993).Contamination by atmospheric N2 during gas transferand storage methods was checked with N-free controls(20% O2/80% He). The remaining 10 cores from eachstation were incubated with no gas head-space in orderto measure O2 and NO �
3 fluxes. In situ bottom watersamples were also incubated in order to test for watercolumn denitrification, which was found to be negligible(data not shown).
2.3. Potential nitrification activity
Potential nitrification rates were measured only atstation 23B. Sediment slurries (3 g, n ¼ 4) were dark-incubated at 22 �C in 50ml centrifuge tubes containing30ml in situ water enriched with NH4Cl and KH2PO4 tofinal concentrations of 0.5 and 0.1mmol l�1, respectively(Henriksen, Hansen, & Blackburn, 1981). The tubeswere kept under continuous shaking (130 rpm), and 5mlwater samples were taken at 0, 6, 12, 16 and 24 h.Potential nitrification rates were determined from linearregression analysis of the accumulation of NO �
3 in thesamples.
2.4. Analysis
Oxygen and NO �3 plus NO �
2 in the water, and theC and N contents of the sediments were measured bystandard methods (Aminot & Chaussepied, 1983). Theformation of 15N-labelled dinitrogen pairs (14N15N and15N15N) by denitrification was measured on a gaschromatograph/combustion/mass spectrometer system(Macko, Uhle, Engel, & Andrusevich, 1997), with thegas chromatograph (HP5890) interfaced to the massspectrometer (Micromass OPTIMA). Molecular nitro-gen samples were first purified by gas chromatography,then combusted in an oxidation furnace at 850 �C and ina reduction furnace at 650 �C. Combusted gas sampleswere passed through a liquid nitrogen trap to removewater and other condensable gases. Pulses of nitrogengas were subsequently introduced into the mass spec-trometer and areas under the 14N14N, 14N15N and15N15N peaks were measured.
2.5. Calculations
The concentration of the 15N-labelled dinitrogen pairswas calculated by multiplying their ratio (14N15N or15N15N/total N) by the N2 concentration in the water(Weiss, 1970). The formation rates for 15N-labelled dini-trogen pairs were used to calculate the rates of denitri-fication of NO �
3 coming from the overlying water (Dw)and rates of denitrification of NO �
3 produced within thesediment by nitrification (Dn) (Nielsen, 1992):
D15 ¼ ð14N15NÞ þ 2ð15N15NÞ ðKoike & Hattori; 1978Þ
D14 ¼ D15
ð14N15NÞ2ð15N15NÞ
ðNielsen; 1992Þ
Dw ¼ D15
100
e15
Dtotal ¼ D15 þD14
Dn ¼ Dtotal �Dw
where e15 is the %15NO3� enrichment of the overlying
water.
518 F. Wang et al. = Estuarine, Coastal and Shelf Science 57 (2003) 515–522
Actual Dw rates at each station were calculated byextrapolating Dw values obtained in the laboratory to insitu NO �
3 concentrations.An analysis of variance (ANOVA) was carried out to
test the significance of between-treatment differences.
3. Results and discussion
Sediment O2 and NO �3 consumption rates, ranging
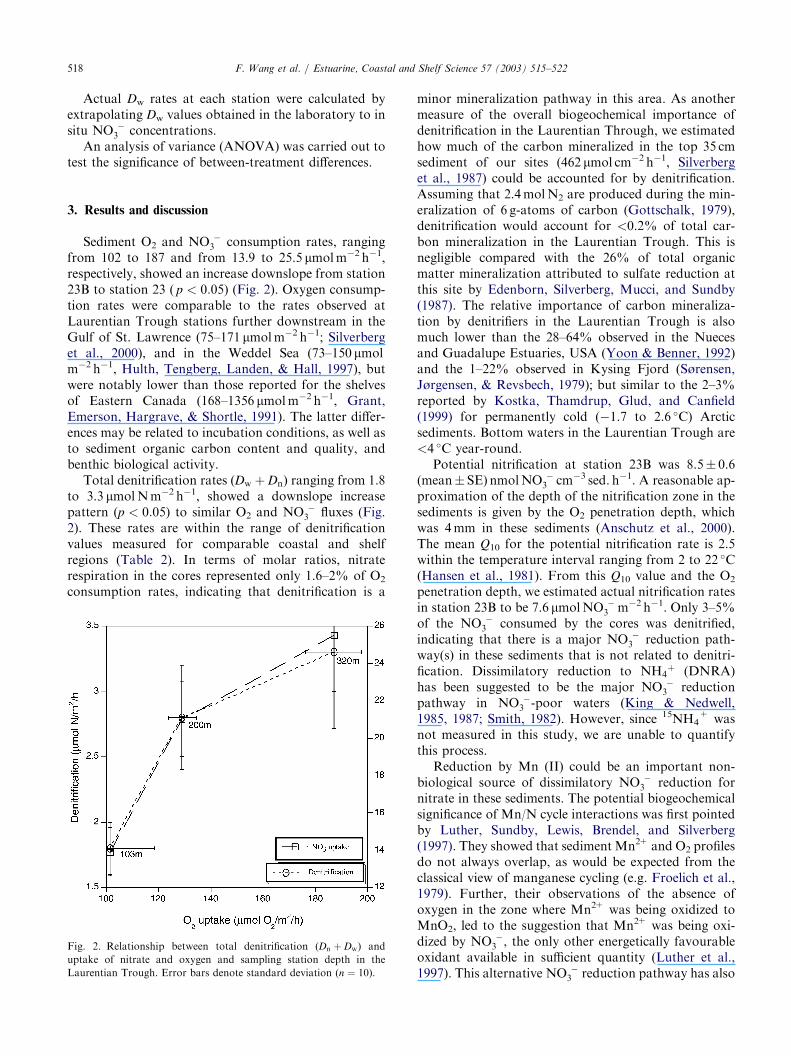
from 102 to 187 and from 13.9 to 25.5 lmolm�2 h�1,respectively, showed an increase downslope from station23B to station 23 ( p < 0:05) (Fig. 2). Oxygen consump-tion rates were comparable to the rates observed atLaurentian Trough stations further downstream in theGulf of St. Lawrence (75–171 lmolm�2 h�1; Silverberget al., 2000), and in the Weddel Sea (73–150 lmolm�2 h�1, Hulth, Tengberg, Landen, & Hall, 1997), butwere notably lower than those reported for the shelvesof Eastern Canada (168–1356 lmolm�2 h�1, Grant,Emerson, Hargrave, & Shortle, 1991). The latter differ-ences may be related to incubation conditions, as well asto sediment organic carbon content and quality, andbenthic biological activity.
Total denitrification rates (Dw þ Dn) ranging from 1.8to 3.3 lmolNm�2 h�1, showed a downslope increasepattern (p < 0:05) to similar O2 and NO �
3 fluxes (Fig.2). These rates are within the range of denitrificationvalues measured for comparable coastal and shelfregions (Table 2). In terms of molar ratios, nitraterespiration in the cores represented only 1.6–2% of O2
consumption rates, indicating that denitrification is a
Fig. 2. Relationship between total denitrification (Dn þ Dw) and
uptake of nitrate and oxygen and sampling station depth in the
Laurentian Trough. Error bars denote standard deviation (n ¼ 10).
minor mineralization pathway in this area. As anothermeasure of the overall biogeochemical importance ofdenitrification in the Laurentian Through, we estimatedhow much of the carbon mineralized in the top 35 cmsediment of our sites (462 lmol cm�2 h�1, Silverberget al., 1987) could be accounted for by denitrification.Assuming that 2.4molN2 are produced during the min-eralization of 6 g-atoms of carbon (Gottschalk, 1979),denitrification would account for <0.2% of total car-bon mineralization in the Laurentian Trough. This isnegligible compared with the 26% of total organicmatter mineralization attributed to sulfate reduction atthis site by Edenborn, Silverberg, Mucci, and Sundby(1987). The relative importance of carbon mineraliza-tion by denitrifiers in the Laurentian Trough is alsomuch lower than the 28–64% observed in the Nuecesand Guadalupe Estuaries, USA (Yoon & Benner, 1992)and the 1–22% observed in Kysing Fjord (Sørensen,Jørgensen, & Revsbech, 1979); but similar to the 2–3%reported by Kostka, Thamdrup, Glud, and Canfield(1999) for permanently cold (�1.7 to 2.6 �C) Arcticsediments. Bottom waters in the Laurentian Trough are<4 �C year-round.
Potential nitrification at station 23B was 8.5� 0.6(mean� SE) nmolNO �
3 cm�3 sed. h�1. A reasonable ap-proximation of the depth of the nitrification zone in thesediments is given by the O2 penetration depth, whichwas 4mm in these sediments (Anschutz et al., 2000).The mean Q10 for the potential nitrification rate is 2.5within the temperature interval ranging from 2 to 22 �C(Hansen et al., 1981). From this Q10 value and the O2
penetration depth, we estimated actual nitrification ratesin station 23B to be 7.6 lmolNO �
3 m�2 h�1. Only 3–5%of the NO �
3 consumed by the cores was denitrified,indicating that there is a major NO �
3 reduction path-way(s) in these sediments that is not related to denitri-fication. Dissimilatory reduction to NH4
þ (DNRA)has been suggested to be the major NO �
3 reductionpathway in NO �
3 -poor waters (King & Nedwell,1985, 1987; Smith, 1982). However, since 15NH4
+ wasnot measured in this study, we are unable to quantifythis process.
Reduction by Mn (II) could be an important non-biological source of dissimilatory NO �
3 reduction fornitrate in these sediments. The potential biogeochemicalsignificance of Mn/N cycle interactions was first pointedby Luther, Sundby, Lewis, Brendel, and Silverberg(1997). They showed that sediment Mn2+ and O2 profilesdo not always overlap, as would be expected from theclassical view of manganese cycling (e.g. Froelich et al.,1979). Further, their observations of the absence ofoxygen in the zone where Mn2+ was being oxidized toMnO2, led to the suggestion that Mn2+ was being oxi-dized by NO �
3 , the only other energetically favourableoxidant available in sufficient quantity (Luther et al.,1997). This alternative NO �
3 reduction pathway has also
519F. Wang et al. / Estuarine, Coastal and Shelf Science 57 (2003) 515–522
Table 2
Denitrification rates of sediments from various continental shelf regions issued from the literature
Region
Denitrification
(lmolN2m�2 h�1)
Depth
(m)
Temperature
(�C)
Salinity
(ppt)
NO3�
(lM)
Organic
carbon (%) References
Washington Shelf 2.1 Christensen, Smethie, and Devol (1987)
Washington Shelf 13.3 98–630 29–48 0.7–1.6 Devol (1991)
Washington Shelf 3.6–19.4 42–630 19–23 1–2 Devol and Christensen (1993)
Pacific Ocean,
Mexican Shelf
5.4 42–388 10 30 Christensen, Murray, Devol,
and Codispoti (1987)
East China Sea 1.4 Aller et al. (1985)
North Atlantic
continental shelf
29.2 Seitzinger and Giblin (1996)
Bering Sea <1.8 19–144 �1.6 to 7.5 23–33 7–33 Haines, Atlas, Griffiths,
and Morita (1981)
Bering Sea 0.9 Koike and Hattori (1979)
Wadden Sea 0–16.7 Jensen, Jensen, and Kristensen (1996)
Arctic Sea, Svalbard 0–4.3 170–550 �2 to 3 34–36 9–12 Blackburn, Hall, Hulth,
and Landen (1996)
Gulf of Maine <3.3 32–290 6–10 6–20 Christensen, Murray, et al. (1987)
Northern Baltic
proper
6.3–27.1 30–446 0.8–4.9 10–10.5 5–10 Tuominen, Heinanen, Kuparinen,
and Nielsen (1998)
North Sea 9.8–13.2 9 34.5 0.6 0.12 Lohse, Kloosterhuis, Raaphorst,
and Helder (1996)
St.-Lawrence Estuary 1.7–3 102–320 4 34 16.1–25.1 1.7–1.9 This study
Water depth, temperature, salinity, NO3� concentration and sediment percent organic carbon content are given.
been proposed by other authors (Aller, 1990; Hulth,Aller, & Gilbert, 1999; Murray, Codispoti, & Friederich,1995; Schulz, Dahmke, Schinzel, Wallmann, & Zabel,1994; Sørensen et al., 1987; Tebo, 1991).
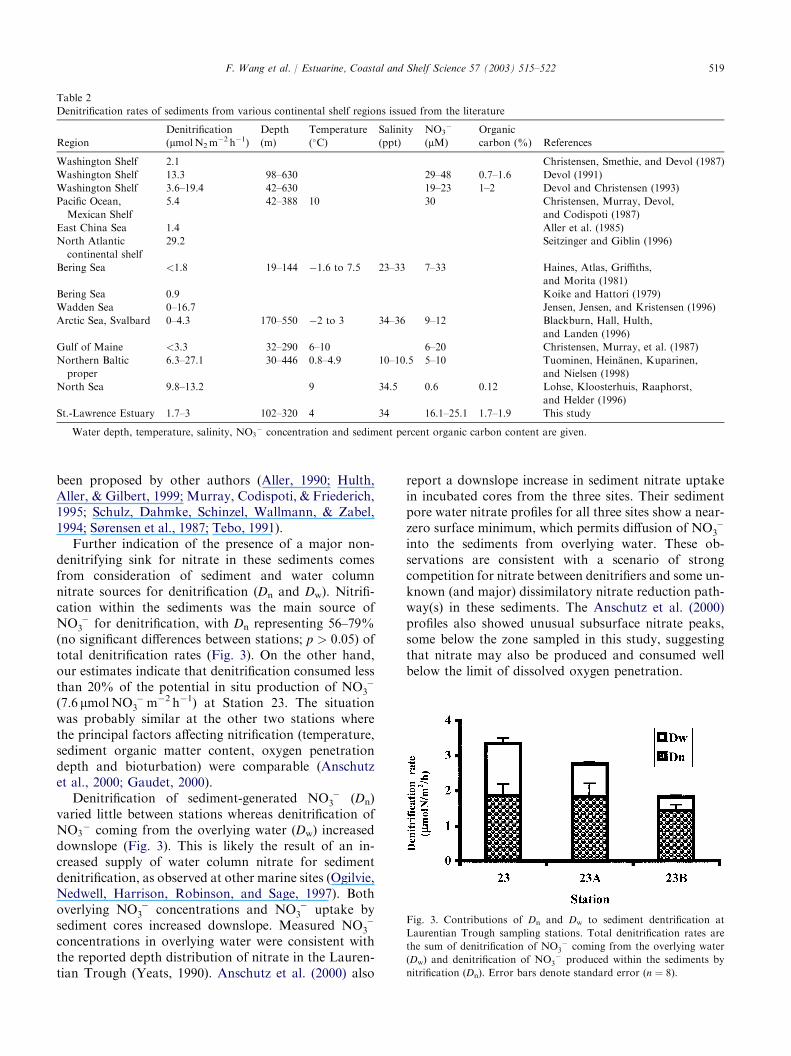
Further indication of the presence of a major non-denitrifying sink for nitrate in these sediments comesfrom consideration of sediment and water columnnitrate sources for denitrification (Dn and Dw). Nitrifi-cation within the sediments was the main source ofNO �
3 for denitrification, with Dn representing 56–79%(no significant differences between stations; p > 0:05) oftotal denitrification rates (Fig. 3). On the other hand,our estimates indicate that denitrification consumed lessthan 20% of the potential in situ production of NO �
3
(7.6 lmolNO �3 m�2 h�1) at Station 23. The situation
was probably similar at the other two stations wherethe principal factors affecting nitrification (temperature,sediment organic matter content, oxygen penetrationdepth and bioturbation) were comparable (Anschutzet al., 2000; Gaudet, 2000).
Denitrification of sediment-generated NO �3 (Dn)
varied little between stations whereas denitrification ofNO3
� coming from the overlying water (Dw) increaseddownslope (Fig. 3). This is likely the result of an in-creased supply of water column nitrate for sedimentdenitrification, as observed at other marine sites (Ogilvie,Nedwell, Harrison, Robinson, and Sage, 1997). Bothoverlying NO �
3 concentrations and NO �3 uptake by
sediment cores increased downslope. Measured NO �3
concentrations in overlying water were consistent withthe reported depth distribution of nitrate in the Lauren-tian Trough (Yeats, 1990). Anschutz et al. (2000) also
report a downslope increase in sediment nitrate uptakein incubated cores from the three sites. Their sedimentpore water nitrate profiles for all three sites show a near-zero surface minimum, which permits diffusion of NO �
3
into the sediments from overlying water. These ob-servations are consistent with a scenario of strongcompetition for nitrate between denitrifiers and some un-known (and major) dissimilatory nitrate reduction path-way(s) in these sediments. The Anschutz et al. (2000)profiles also showed unusual subsurface nitrate peaks,some below the zone sampled in this study, suggestingthat nitrate may also be produced and consumed wellbelow the limit of dissolved oxygen penetration.
Fig. 3. Contributions of Dn and Dw to sediment dentrification at
Laurentian Trough sampling stations. Total denitrification rates are
the sum of denitrification of NO �3 coming from the overlying water
(Dw) and denitrification of NO �3 produced within the sediments by
nitrification (Dn). Error bars denote standard error (n ¼ 8).

520 F. Wang et al. = Estuarine, Coastal and Shelf Science 57 (2003) 515–522
At the time that cores were collected for this study,reduced metal species such as Mn (II) may have beencompeting with heterotrophic microorganisms for oxi-dants such as nitrate, as has been proposed by Lutheret al. (1997). We have no direct evidence for this but thepotential for such interference is high given the reactivityof the species involved and the presence of abundantreduced manganese and iron near the sediment–waterinterface (Anschutz et al., 2000). Further investigationof competitive interactions between microbial andstrictly chemical oxidations is warranted, particularlyin deep-water coastal sediments that are sinks forcontinentally derived metals but have relatively loworganic content. Ultimately, microbial oxidation of or-ganic matter is responsible for the reduction of metals inthese sediments, but metal reduction need not be insteady state equilibrium with metal oxidation. It isbecoming increasingly apparent that episodes of intensemicrobial oxidative activity linked to organic mattersedimentation events are common in temperate marineenvironments such as the St. Lawrence system (Lavigneet al., 1997). During such events, dissolved oxidants canbe quickly consumed, leading to the reduction ofparticulate metal oxides. Anschutz et al. (2000) pointout that particulate metal oxides in these sedimentsprovide a potential electron sink of far greater oxidativecapacity than that of typical dissolved oxygen concen-trations. The role of particulate iron and manganeseoxides as terminal electron acceptors for organic matteroxidation is of increasing interest in microbial ecology(Lovley, 1993; Nealson, 1997). Faunal bioturbation willalso likely increase during sedimentation events (Des-rosiers et al., 2000), augmenting the downward advec-tion of particulate oxides into anoxic layers where theywill react. Following periods of intensified benthic bio-logical activity, upwardly diffusing reduced iron andmanganese species may then have a predominatinginfluence on chemistry and microbiology in surfacesediments.
Compared to the situation in shallower estuaries andpossibly on continental slopes, denitrification, at levelsmeasured here and in other continental shelf areas, isnot an important electron sink for oxidative processessuch as organic carbon mineralization. Its primarybiogeochemical significance appears to be limited toits impact on the nitrogen cycle within the sediments,although our data show that <20% of potential NO �
3
production is subsequently denitrified. Since we detectedonly minor changes in denitrification rates in ourupslope transect, we conclude that our data are repre-sentative of the slope and floor of the Laurentian Troughin this area off Rimouski, at the time of sampling in June1997. Although we also observed similar denitrificationrates at the same three stations in October 1997 (datanot shown), it would be premature to consider annualdenitrification budgets. Since primary productivity and
organic matter flux to the seafloor in the region arestrongly seasonal (e.g. Lavigne et al., 1997), estimatingannual denitrification loss of N should await a moreextensive seasonal coverage. We would advise evengreater caution in extrapolating denitrification ratesfrom this single transect to the deep waters of the entireSt. Lawrence Estuary. The approximate 200 km estuar-ine portion of the Laurentian Trough is characterizedby a downstream transition from terrigenous to marinesedimentation regimes (Silverberg et al., 1987). Thistransition affects the quality and quantity of sedimentorganic matter, as well as the sedimentation of redoxactive metals such as Mn and Fe. Such a gradient inthe supply of organic matter and potentially interferingmetals may have considerable impact on the magnitudeof nitrogen loss via sediment denitrification.
Acknowledgements
We thank the Captain and crew of the R/V AlcideC. Horth for their enthusiastic collaboration duringsampling operations at sea. Bjorn Sundby kindly pro-vided laboratory space at the Universite du Quebeca Rimouski for the core incubations. This work wasfunded by the Natural Sciences and Engineering Re-search Council of Canada, through grants to the Cana-dian JGOFS program. Discussions with Bjorn Sundbyand Pierre Anschutz, and input from three anonymousreviewers aided in the development and improvement ofthis paper.
References
Aller, R. C. (1990). Bioturbation and manganese cycling in hemi-
pelagic sediments. Philosophical Transactions of the Royal Society of
London 331, 51–68.
Aller, R. C., Mackin, J. E., Ullman, W. J., Chen-Hou, W., Shing-Min,
T., Jian-Cai, J., Yong-Nian, S., & Jia-Zhen, H. (1985). Early
chemical diagenesis, sediment–water solute exchange, and storage
of reactive organic matter near the mouth of the Changjiang, East
China Sea. Continental Shelf Research 4, 227–251.
Aminot, A., & Chaussepied, M. (1983). Manuel des analyses chimiques
en milieu marin (pp. 395). Paris: Centre National pour l’Exploita-
tion des Oceans (CNEXO).
Anschutz, P., Sundby, B., Lefrancois, L., Luther, G. W., & Mucci, A.
(2000). Interactions between metal oxides and species of nitrogen
and iodine in bioturbated marine sediments. Geochimica et
Cosmochimica Acta 64, 2751–2763.
Blackburn, T. H., Hall, P. O. J., Hulth, S., & Landen, A. (1996).
Organic-N loss by efflux and burial associated with a low efflux of
inorganic N and with nitrate assimilation in Arctic sediments
(Svalbard, Norway). Marine Ecology Progress Series 141, 283–293.
Christensen, J. P., Murray, J. W., Devol, A. H., & Codispoti, L. A.
(1987). Denitrification in continental shelf sediments has major
impact on the oceanic nitrogen budget. Global Biogeochemical
Cycles 1, 97–116.
Christensen, J. P., Smethie, W. H., Jr., & Devol, A. H. (1987). Benthic
nutrient regeneration and denitrification on the Washington
continental shelf. Deep-Sea Research 34, 1027–1049.

521F. Wang et al. / Estuarine, Coastal and Shelf Science 57 (2003) 515–522
Colombo, J. C., Silverberg, N., & Gearing, J. N. (1995). Biogeochem-
istry of organic matter in the Laurentian Trough; I, Composition
and vertical fluxes of rapidly settling particles. Marine Chemistry
51, 277–293.
Colombo, J. C., Silverberg, N., & Gearing, J. N. (1995). Biogeochem-
istry of organic matter in the Laurentian Trough; II, Bulk
composition of the sediments and relative reactivity of major
components during early diagenesis. Marine Chemistry 51,
295–314.
Desrosiers, G., Savenkoff, C., Olivier, M., Stora, G., Juniper, S. K.,
Caron, A., Gagne, J.-P., Legendre, L., Mulsow, S., Roy, S.,
Grehan, A., Scaps, P., Silverberg, N., Klein, B., Tremblay, J.-E., &
Therriault, J.-C. (2000). Trophic structure of benthos in the Gulf of
St. Lawrence and on the Scotian shelf. Deep-Sea Research Part II
47, 663–697.
Devol, A. H. (1991). Direct measurement of nitrogen gas fluxes from
continental shelf sediments. Nature (London) 349, 319–321.
Devol, A. H., & Christensen, J. P. (1993). Benthic fluxes and nitrogen
cycling in sediments of the continental margin of the eastern North
Pacific. Journal of Marine Research 51, 345–372.
Edenborn, H. M., Silverberg, N., Mucci, A., & Sundby, B. (1987).
Sulfate reduction in deep coastal marine sediments. Marine
Chemistry 21, 329–345.
El-Sabh, M. I., & Silverberg, N. (1990). The St. Lawrence Estuary:
Introduction. In M. I. El-Sabh, & N. Silverberg (Eds.), Ocean-
ography of a large-scale estuarine system, the St.-Lawrence. Coastal
and estuarine studies Vol. 39 (pp. 1–6). New York: Springer-Verlag.
Froelich, P. N., Klinkhammer, G. P., Bender, M. L., Luedtke, N. A.,
Heath, G. R., Cullen, D., Dauphin, P., Hammond, D., Hartman,
B., & Maynard, V. (1979). Early oxidation of organic matter in
pelagic sediments of the eastern equatorial Atlantic: suboxic
diagenesis. Geochimica et Cosmochimica Acta 43, 1075–1090.
Gaudet, A. M. (2000). Caracterisation et evaluation du potentiel
bioturbateur des populations benthiques, le long d’un gradient
bathymetrique, de l’estuaire maritime du St-Laurent. MSc. thesis in
Environmental Science, Universite du Quebec a Montreal.
Gearing, J. N., & Pocklington, R. (1990). Organic geochemical studies
in the St. Lawrence Estuary. In M. I. El-Sabh, & N. Silverberg
(Eds.), Oceanography of a large-scale estuarine system, the
St.-Lawrence. Coastal and estuarine studies Vol 39 (pp. 170–195).
New York: Springer-Verlag.
Gottschalk, G. (1979). Bacterial metabolism (pp. 101–103). New York:
Sringer-Verlag.
Grant, J., Emerson, C. W., Hargrave, B. T., & Shortle, J. L. (1991).
Benthic oxygen consumption on continental shelves of Eastern
Canada. Continental Shelf Research 11, 1083–1097.
Haines, J. R., Atlas, R. M., Griffiths, R. P., & Morita, R. Y. (1981).
Denitrification and nitrogen fixation in Alaskan continental shelf
sediments. Applied and Environmental Microbiology 41, 412–421.
Hansen, J. I., Henriksen, K., & Blackburn, T. H. (1981). Seasonal
distribution of nitrifying bacteria and rates of nitrification in
coastal marine sediments. Marine Ecology 7, 297–304.
Henriksen, K., Hansen, J. I., & Blackburn, T. H. (1981). Rates of
nitrification, distribution of nitrifying bacteria, and nitrate fluxes in
different types of sediment from danish waters. Marine Biology 61,
299–304.
Hulth, S., Aller, R. C., & Gilbert, F. (1999). Coupled anoxic
nitrification/manganese reduction in marine sediments. Geochimica
et Cosmochimica Acta 63, 49–66.
Hulth, S., Tengberg, A., Landen, A., & Hall, P. O. J. (1997).
Mineralization and burial of organic carbon in sediments of the
southern Weddell Sea (Antarctica). Deep-Sea Research 44, 955–
981.
Jensen, K. M., Jensen, M. H., & Kristensen, E. (1996). Nitrification
and denitrification in Wadden Sea sediments (Konigshafen, Island
of Sylt, Germany) as measured by nitrogen isotope pairing and
isotope dilution. Aquatic Microbial Ecology 11, 181–191.
King, D., & Nedwell, D. B. (1985). The influence of nitrate
concentration upon the endproducts of nitrate dissimilation by
bacteria in anaerobic salt marsh sediments. FEMS Microbiology
Letters 31, 23–28.
King, D., & Nedwell, D. B. (1987). The adaptation of nitrate reducing
bacterial communities in estuarine sediments in response to
overlying nitrate load. FEMS Microbiology Letters 35, 278–285.
Koike, I., & Hattori, A. (1978). Simultaneous determination of
nitrification and nitrate reduction in coastal sediments by a 15N
dilution technique. Applied and Environmental Microbiology 35,
853–857.
Koike, I., & Hattori, A. (1979). Estimates of denitrification in sedi-
ments of the Bering Sea shelf. Deep-Sea Research 26, 409–415.
Kostka, J. E., Thamdrup, B., Glud, R. N., & Canfield, D. E. (1999).
Rates and pathways of carbon oxidation in permanently cold
Arctic sediments. Marine Ecology Progress Series 187, 7–21.
Lamontagne, M. G., & Valiela, I. (1995). Denitrification measured by
a direct N2 flux method in sediments of Waquoit Bay, MA.
Biogeochemistry 31, 63–83.
Lavigne, C., Juniper, S. K., & Silverberg, N. (1997). Spatio-temporal
variability in benthic microbial activity and particle flux in the
Laurentian Trough. Deep-Sea Research 44, 1793–1813.
Lohse, L., Kloosterhuis, H. T., Raaphorst, W.W., &Helder,W. (1996).
Denitrification rates as measured by the isotope pairing method and
by the acetylene inhibition technique in continental shelf sediments
of the North Sea. Marine Ecology Progress Series 132, 169–179.
Lovley, D. R. (1993). Dissimilatory Fe and Mn reduction. Microbial
Reviews 55, 259–287.
Luther, G. W., Sundby, B., Lewis, B. L., Brendel, P. J., & Silverberg,
N. (1997). Interactions of manganese with the nitrogen cycle:
alternative pathways to dinitrogen. Geochimica et Cosmochimica
Acta 61, 4043–4045.
Macko, S. A., Uhle, M. E., Engel, M. H., & Andrusevich, V. (1997).
Stable nitrogen isotope analysis of amino acid enantiomers by gas
chromatography/combustion/isotope ratio mass spectrometry.
Analytical Chemistry 69, 926–929.
Murray, J. W., Codispoti, L. A., & Friederich, G. E. (1995).
Oxidation–reduction environments, the suboxic zone of the Black
Sea. In C. P. Huang, C. R. O’Melia, & J. J. Morgan (Eds.), Aquatic
chemistry: Interfacial and interspecies processes Vol. 244 (pp. 157–
176). Washington D.C.: American Chemical Society.
Nealson, K. H. (1997). Sediment bacteria: who’s there, what are they
doing and what’s new. Annual Reviews of Earth and Planetary
Sciences 25, 403–434.
Nielsen, L. P. (1992). Denitrification in sediment determined from
nitrogen isotope pairing. FEMS Microbiology Ecology 86, 357–362.
Nixon, S. W. (1981). Remineralization and nutrient cycling in coastal
marine ecosystems. In B. J. Nielson, & L. E. Cronin (Eds.),
Estuaries and nutrients (pp. 111–138). Clifton: Humana Press.
Ogilvie, B., Nedwell, D. B., Harrison, R. M., Robinson, A., & Sage, A.
(1997). High nitrate, muddy estuaries as nitrogen sinks: the
nitrogen budget of the River Colne Estuary (United Kingdom).
Marine Ecology Progress Series 150, 217–228.
Risgaard-Petersen, N., Rysgaard, S., & Revsbech, N. P. (1993). A
sensitive assay for determination of 14N/15N isotope distribution in
nitrate. Journal of Microbiological Methods 17, 155–164.
Rysgaard, S., Christensen, P. B., & Nielsen, L. P. (1995). Seasonal
variation in nitrification and denitrification in estuarine sediment
colonized by benthic microalgae and biooturbating infauna. Marine
Ecology Progress Series 126, 111–121.
Ryther, J. H., & Dunstan, W. M. (1971). Nitrogen, phosphorus, and
eutrophication in the coastal marine environment. Science 171,
1008–1013.
Schulz, H. D., Dahmke, A., Schinzel, U., Wallmann, K., & Zabel, M.
(1994). Early diagenetic processes, fluxes, and reaction rates in
sediments of the South Atlantic. Geochimica et Cosmochimica Acta
58, 2041–2060.

522 F. Wang et al. = Estuarine, Coastal and Shelf Science 57 (2003) 515–522
Seitzinger, S. P. (1988). Denitrification in freshwater and coastal
marine ecosystems: ecological and geochemical significance.
Limnology and Oceanography 33, 702–724.
Seitzinger, S. P., & Giblin, A. E. (1996). Estimating denitrification in
North Atlantic continental shelf sediments. Biogeochemistry 35,
235–260.
Silverberg, N., Bakker, J., Edenborn, H. M., & Sundby, B. (1987).
Oxygen profiles and organic carbon fluxes in Laurentian Trough
sediments. Netherlands Journal of Sea Research 21, 95–105.
Silverberg, N., Sundby, B., Mucci, A., Zhong, S., Arakaki, T., Hall, P.,
Landen, A., & Tengberg, A. (2000). Remineralization of organic
carbon in eastern Canadian continental margin sediments. Deep-
Sea Research Part II 47, 699–731.
Smith, M. S. (1982). Dissimilatory reduction of NO2� to NH4
+ and
N2O by a soil Citrobacter sp. Applied and Environmental Micro-
biology 43, 854–860.
Sørensen, J., Jørgensen, K. S., Colley, S., Hydes, D. J., Thomson, J., &
Wilson, T. R. S. (1987). Depth localization of denitrification in a
deep-sea sediment from the Madeira abyssal plain. Limnology and
Oceanography 32, 758–762.
Sørensen, J., Jørgensen, B. B., & Revsbech, N. P. (1979). A
comparison of oxygen, nitrate, and sulphate respiration in coastal
marine sediments. Microbial Ecology 5, 105–115.
Tebo, B. M. (1991). Manganese(II) oxidation in the suboxic zone of
the Black Sea. Deep-Sea Research 38, 883–905.
Tuominen, L., Heinanen, A., Kuparinen, J., & Nielsen, L. P. (1998).
Spatial and temporal variability of denitrification in the sediments
of the northern Baltic Proper. Marine Ecology Progress Series 172,
13–24.
Weiss, R. F. (1970). The solubility of nitrogen, oxygen and
argon in water and seawater. Deep-Sea Research 17, 721–
735.
Yeats, P. A. (1990). Reactivity and transport of nutrients and metals in
the St. Lawrence Estuary. In M. I. El-Sabh, & N. Silverberg (Eds.),
Oceanography of a large-scale estuarine system, the St.-Lawrence.
Coastal and estuarine studies Vol. 39 (pp. 155–165). New York:
Springer-Verlag.
Yoon, W. B., & Benner, R. (1992). Denitrification and oxygen
consumption in sediments of two south Texas estuaries. Marine
Ecology Progress Series 90, 157–167.
Copyright © 2022 FDOKUMEN




















