
Denitrification in extreme alkaline saline soils of the former lake Texcoco
11
REGULAR ARTICLE Denitrification in extreme alkaline saline soils of the former lake Texcoco E. Ruiz-Romero & R. Alcántara-Hernández & C. Cruz-Mondragon & R. Marsch & M. L. Luna-Guido & L. Dendooven Received: 30 June 2008 / Accepted: 15 December 2008 / Published online: 6 February 2009 # Springer Science + Business Media B.V. 2009 Abstract Extreme alkalinity and salinity affects different processes involved in the N cycle, such as mineralization and nitrification, so it might affect the denitrification process and the N 2 O-to-N 2 ratio. An alkaline saline soil of the former lake Texcoco with electrolytic conductivity (EC) 12.6 dS m -1 and pH 10.0, and an ‘extreme’ alkaline saline soil with EC 56 dS m -1 and pH 10.4 were amended with 100 mg NO 3 - -N kg -1 , purged of all O 2 , shaken and incubated anaerobically for 72 h. A ‘normal’ agricul- tural soil of Acolman with EC 0.8 dS m -1 and pH 6.3 was used as control. Treatments were with or without acetylene (C 2 H 2 ) (found to inhibit reduction of N 2 O to N 2 ) and with or without chloramphenicol (found to inhibit de novo synthesis of reduction enzymes), while emission of CO 2 and N 2 O and concentrations of NO 3 - and NO 2 - were monitored. The NO 3 - reduc- tion rate was not affected by C 2 H 2 or chloramphen- icol in the Acolman soil, but decreased >3.5 times when chloramphenicol was added to the Texcoco soils. The concentration of NO 2 - increased 3 times when chloramphenicol was added to the Acolman soil after 72 h, was similar in the Texcoco soil with EC 12 dS m -1 , but chloramphenicol decreased it 14 times in the Texcoco soil with EC 56 dS m -1 . The emission of N 2 O in the unamended Acolman soil was 0.09 mg N kg -1 and N 2 6.5 mg N kg -1 soil after 72 h resulting in aN 2 O-to-N 2 ratio of 0.014, while N 2 O production was 0.05 mg N kg -1 in the Texcoco soil with EC 12.6 dS m -1 and N 2 1.6 mg N kg -1 resulting in a N 2 O-to- N 2 ratio of 0.031. The emission of N 2 O in the Texcoco soil with EC 56 dS m -1 was 1.3 mg N kg -1 soil after 72 h and that of N 2 was 2.0 mg N kg -1 resulting in a N 2 O-to-N 2 ratio of 0.65. It was found that the activity of NO 2 - and N 2 O reductase decreased with increased pH and EC, so that the concentrations of NO 2 - ,N 2 O and the N 2 O-to-N 2 ratio increased with increased EC and pH, although that the effect of other soil characteristics can not be excluded. Keywords Acetylene alkaline saline soil . Anaerobic incubation . Production of CO 2 and N 2 O . The denitrification process . N 2 O-to-N 2 ratio Introduction Irrigation and fertilization of crops has increased salinity of soils in large parts of the world (Rengasamy 2006). Increases in salinity will not only have a pro- found effect on crop yields, but also on soil micro- organisms and thus on soil processes (Liang et al. Plant Soil (2009) 319:247–257 DOI 10.1007/s11104-008-9867-y Responsibility Editor: Elizabeth (Liz) A. Stockdale. E. Ruiz-Romero : R. Alcántara-Hernández : C. Cruz-Mondragon : R. Marsch : M. L. Luna-Guido : L. Dendooven (*) Laboratory of Soil Ecology, Department of Biotechnology and Bioengineering, Cinvestav, Mexico City, Mexico e-mail: [email protected]
Transcript of Denitrification in extreme alkaline saline soils of the former lake Texcoco

REGULAR ARTICLE
Denitrification in extreme alkaline saline soilsof the former lake Texcoco
E. Ruiz-Romero & R. Alcántara-Hernández &
C. Cruz-Mondragon & R. Marsch &
M. L. Luna-Guido & L. Dendooven
Received: 30 June 2008 /Accepted: 15 December 2008 / Published online: 6 February 2009# Springer Science + Business Media B.V. 2009
Abstract Extreme alkalinity and salinity affectsdifferent processes involved in the N cycle, such asmineralization and nitrification, so it might affect thedenitrification process and the N2O-to-N2 ratio. Analkaline saline soil of the former lake Texcoco withelectrolytic conductivity (EC) 12.6 dS m−1 andpH 10.0, and an ‘extreme’ alkaline saline soil withEC 56 dS m−1 and pH 10.4 were amended with100 mg NO3
− -N kg−1, purged of all O2, shaken andincubated anaerobically for 72 h. A ‘normal’ agricul-tural soil of Acolman with EC 0.8 dS m−1 and pH 6.3was used as control. Treatments were with or withoutacetylene (C2H2) (found to inhibit reduction of N2Oto N2) and with or without chloramphenicol (found toinhibit de novo synthesis of reduction enzymes),while emission of CO2 and N2O and concentrationsof NO3
− and NO2− were monitored. The NO3
− reduc-tion rate was not affected by C2H2 or chloramphen-icol in the Acolman soil, but decreased >3.5 timeswhen chloramphenicol was added to the Texcocosoils. The concentration of NO2
− increased 3 times
when chloramphenicol was added to the Acolman soilafter 72 h, was similar in the Texcoco soil with EC 12dS m−1, but chloramphenicol decreased it 14 times inthe Texcoco soil with EC 56 dS m−1. The emission ofN2O in the unamended Acolman soil was 0.09 mg Nkg−1 and N2 6.5 mg N kg−1 soil after 72 h resulting ina N2O-to-N2 ratio of 0.014, while N2O productionwas 0.05 mg N kg−1 in the Texcoco soil with EC 12.6dS m−1 and N2 1.6 mg N kg−1 resulting in a N2O-to-N2 ratio of 0.031. The emission of N2O in theTexcoco soil with EC 56 dS m−1 was 1.3 mg Nkg−1 soil after 72 h and that of N2 was 2.0 mg N kg−1
resulting in a N2O-to-N2 ratio of 0.65. It was foundthat the activity of NO2
− and N2O reductase decreasedwith increased pH and EC, so that the concentrationsof NO2
−, N2O and the N2O-to-N2 ratio increased withincreased EC and pH, although that the effect of othersoil characteristics can not be excluded.
Keywords Acetylene alkaline saline soil .
Anaerobic incubation . Production of CO2 and N2O .
The denitrification process . N2O-to-N2 ratio
Introduction
Irrigation and fertilization of crops has increasedsalinity of soils in large parts of the world (Rengasamy2006). Increases in salinity will not only have a pro-found effect on crop yields, but also on soil micro-organisms and thus on soil processes (Liang et al.
Plant Soil (2009) 319:247–257DOI 10.1007/s11104-008-9867-y
Responsibility Editor: Elizabeth (Liz) A. Stockdale.
E. Ruiz-Romero : R. Alcántara-Hernández :C. Cruz-Mondragon :R. Marsch :M. L. Luna-Guido :L. Dendooven (*)Laboratory of Soil Ecology,Department of Biotechnology and Bioengineering,Cinvestav,Mexico City, Mexicoe-mail: [email protected]

2005; Tejada et al. 2006). Dynamics of nitrogen, animportant nutrient for plants, will be affected. It hasbeen reported that ammonification and nitrificationare inhibited when salt concentrations increased insoil (Oren 1999). Reduced ammonification will limitN availability for plants. The denitrification processmight also be affected and especially the N2O-to-N2
ratio. N2O is a trace gas, which plays a central role inatmospheric chemistry, and it has been reported that65% to 70% of this gas could be coming from soils(Sánchez et al. 2001; Kiese and Butterbach-Bahl2002). This gas has an approximately half-life of120 years and contributes to at least 5% of the globalwarming observed while it depletes the stratosphericozone layer (Sánchez et al. 2001; Wrage et al. 2001).The ratio of the nitrogen gases formed, i.e. N2O andN2, is largely controlled by pH, O2 and nitrate con-centrations and the status of the reduction enzymes(Firestone et al. 1980). N2O reductase is inhibited bya low pH and O2, so it might well be that large saltconcentrations will do the same. Little is known, howsalinity and alkalinity might affect the denitrificationprocess, the reduction enzymes involved and thus thedynamics of NO3
−, NO2−, N2O and N2 in soil under
anaerobic conditions.Most studies investigating the effects of salinity on
soil processes add salts to soil (e.g. Nelson et al.1996). Microorganisms in these soils, however, haveno time to adapt to the changed conditions. Theirresponse to salts might be different to those ofmicroorganisms living for an extended time in soilswith high amounts of salts. For instance, archaea areabundant in these soils, and might play an importantrole in these ‘extreme’ alkaline saline conditions(Valenzuela-Encinas et al. 2008). As such, studieswith ‘natural’ saline soil are more realistic. The soil ofTexcoco is alkaline saline with pH locally > 11 andelectrolytic conductivity (EC) > 80 dS m−1 (Beltrán-Hernández et al. 1999). Drainage with effluents sincethe 1970’s has created soils with different salt con-centrations and pH ideal to study the effect of thosecharacteristics on soil processes (Luna-Guido et al.2000). A disadvantage, however, of using ‘natural’saline alkaline soils of Texcoco is the variability inother soil characteristics, such as soil organic C, totalN and clay content, that might also affect soilprocesses (Castro-Silva et al. 2008). These variationsare a result of difference in lake deposition, vegetationand time of drainage (Luna-Guido et al. 2000).
In this study, we investigated how salinity andalkalinity affected the denitrification process, i.e.dynamics of NO3
−, NO2−, N2O, and the N2O-to-N2
ratio. Soil was sampled at two sites in the former lakeTexcoco with EC 56 and 12.6 dS m−1 and from anagriculture soil of Acolman with EC 0.8 dS m−1. Soilwas treated with or without acetylene (C2H2) (foundto inhibit reduction of N2O to N2) and with or withoutchloramphenicol (found to inhibit de novo synthesisof proteins) under strict anaerobic conditions so thatthe N2O-to-N2 ratio and the amount of reductionenzymes available could be investigated (Dendoovenet al. 1994a, b).
Materials and methods
Sampling sites
The first sampling site was located in the former lakeTexcoco in the valley of Mexico City (Mexico) at analtitude of 2,240 m above sea level with a meanannual temperature of 16°C and annual precipitationof 705 mm. The soil in the former lake Texcoco isformed from volcanic ash deposited in situ in alacustrine environment and covered recently bycolluvial materials. The aquifer is near to the surface(80–150 cm) and the groundwater is highly saline,with sodium (Na+), chloride (Cl−), bicarbonate(HCO3
−) and carbonate (CO32−) being dominant
(Gutiérrez-Castorena 1997). pH in undrained soilcan range from 9.8 to 11.7, and EC in saturationextracts ranges from 22 to 150 dS m−1 (Beltrán-Hernández 2001). Exchangeable sodium percentagesrange from 76 to 98% and sodium adsorption ratiofrom 103 to 1718 mmol l−1. Natural drainage is poor.Although the soils are deep, roots are restricted bya compact ash layer 5–20 cm thick at depths of16–40 cm.
The mineralogy is dominated by amorphous sili-ceous materials, smectites, kaolinite and cristobalite(Gutiérrez-Castorena 1997). The amorphous materialsgive the soil a very high water retention capacity (2.4to 5 kg kg−1), which decreases upon drying. Drainageof the area has created cracks 1 m deep and 10 cmwide. The amorphous material increases P fixation;retention is especially high in the moist subsoil(80%), while in the topsoil values range from 26 to31% (INEGI 1994).
248 Plant Soil (2009) 319:247–257

Parts of the former lake bed have been drained andirrigated with sewage effluent from a waste waterplant to remove excess of salt. Distichlis spicata, anindigenous grass with a high tolerance to salt and Na,and tamarix (Tamarix spp.) have been introducedsince the early 1970s to control erosion, and they nowcover much of the area. More details on thevegetation and the effluents used to drain the plotscan be found in Luna-Guido et al. (2000). At two siteswith different EC due to different periods of drainage,three approximately 400 m2 plots were defined. Eachplot was sampled by augering the 0–15 cm layer 30-times with an Edelman auger (combination type) withdiameter 10 cm as measured diagonally between theblades at the broadest part of the auger body(Eijkelkamp, Nl). The soil from each plot was pooledseparately, air-dried, 5 mm sieved and characterized(Table 1). The first soil with an EC 12.6 dS m−1 wassampled on 22nd June 2002 and the second with EC56 dS m−1 on 11th March 2002. The particle size distri-bution is different between the soils and that mightreflect variation of deposition in the former lake bed.
The third sampling site was located near the ex-convent of Acolman in the State of México (México)(Northern Latitude 19°38′ Western Longitude 98°55′)not far from the former lake Texcoco. Its averagealtitude is 2,250 m above sea level and characterizedby a sub-humid temperate climate with a mean annualtemperature of 14.9°C and average annual precipita-tion of 624 mm mainly from June through August(http://www.inegi.gob.mx). The soil in this area hasbeen cultivated with maize for > 20 years, receiving120 kg inorganic fertilizer ha−1 without being irrigated(http://www.inegi.gob.mx). The crop upon samplingwas maize.
The top 0–15 cm soil layer of three 400 m2 plotswas sampled by augering 30-times with an Edelman
auger (combination type) with diameter 10 cm as mea-sured diagonally between the blades at the broadestpart of the auger body (Eijkelkamp, Nl) on 14th April2003. The soil from each plot was pooled so that threesoil samples were obtained, air-dried, 5 mm sieved andcharacterized (Table 1). As such, a total of nine soilsamples was obtained; soil from three sampling sitesin triplicate.
Experimental procedure
Sixty sub-samples of 10 g air-dried soil from eachplot (three) of each sampling site (three) were addedto 120 ml flasks, adjusted to 40% water holdingcapacity (WHC) with distilled H2O and pre-incubatedat 25°C for 7 days in drums containing a flask with 1M NaOH to trap CO2 evolved and a jar with distilledwater to maintain the water content in the soil. After7 days, the water content in the flasks was tested andlosses were less than 2%.
Twenty ml of a degassed 3.57 mM KNO3 solutionwas added to half of the pre-incubated sub-samples,i.e. 30, from each plot of each sampling site while theother 30 pre-incubated sub-samples received 20 ml ofa solution containing 3.57 mM KNO3 plus 0.23 mMchloramphenicol (CHL treatment). The samples werewell mixed to create a soil slurry thereby minimizingthe influence of diffusion of nitrate through the soilmatrix. A concentration of 100 mg NO3
− -N kg−1 wasthus added. The concentration of chloramphenicol was150 mg kg−1, sufficient to inhibit protein synthesis,e.g. NO3
−, NO2− and N2O reductases, but not the
activity of existing enzymes (Dendooven et al. 1994a, b;Murray and Knowles 1999). After application of the3.57 mM KNO3 or 3.57 mM KNO3 plus 0.23 mMchloramphenicol solution, three sub-samples of eachtreatment were selected at random for assays of NO3
−
Table 1 Characteristics of soil of Texcoco and Acolman
Soil EC a pH WHC b Carbon Total nitrogen Particle size distribution
Organic Inorganic Clay Silt Sand Textural Classification(dS m−1) (g kg−1 soil)
Texcoco 56.0 10.4 740 53.0 11 6.7 220 180 600 Sandy clay loam12.6 10.0 600 38.8 8.0 2.9 50 90 860 Loamy sand
Acolman 0.8 6.3 896 19.0 0.6 1.4 440 300 260 Clay
a EC: Electrolytic conductivitybWHC: Water holding capacity
Plant Soil (2009) 319:247–257 249

and NO2−. Each sub-sample was shaken on an orbital
shaker at 180 rpm for 30 min and extracted with100 ml 0.5 M K2SO4. The extracts were filteredthrough Whatman No. 42 filter paper®.
The flasks were sealed air-tight with a rubberstopper, evacuated for 1 min and flushed with heliumfor 1 min. This procedure was repeated three times. Theproduction of N2O and N2 was determined using theC2H2 inhibition technique (Balderstone et al. 1976).After purging of all O2, 10 ml of C2H2 (10% v/v) wasadded to half of the sub-samples with KNO3 alone andto half of those with KNO3 + chloramphenicol. Allsub-samples were then kept at 25°C for 72 h. After 6,24, 48 and 72 h, three sub-samples were selected atrandom from each treatment. The headspace of eachflask was sampled and analysed for N2O and CO2. Theconcentrations of N2O and CO2 were corrected for gasdissolved in the water (Moraghan and Buresh 1977).After measurement of N2O and CO2, samples wereanalysed for NO2
− and NO3−. Data for NO2
− werecorrected for the formation of NO2
− through thedegradation of chloramphenicol (Dendooven et al.1994a, b) and standard curves for both NO3
− andNO2
− were determined for the three soils used.
Chemical analysis
Soil pH was measured in 1:2.5 soil-H2O suspensionusing a glass electrode (Thomas 1996). Large con-centrations of Cl− might affect the determination ofcarbon (C) by wet oxidation (Nelson and Sommers1996) so the total C in soil was determined byoxidation with potassium dichromate. The evolvedCO2 was trapped in NaOH and titrated with 0.1 MHCl (Amato 1983). Inorganic C in soil was deter-mined by adding 20 ml 1 M HCl solution to 1 g air-dried soil. The CO2 evolved was trapped in 20 ml 1 MNaOH and titrated with 0.1 M HCl to determine theCO2. The organic C was defined as the differencebetween total C and inorganic C. Total N was mea-sured by the Kjeldhal method using concentratedH2SO4, K2SO4 and HgO to digest the sample(Bremner 1996) and soil particle size distribution bythe hydrometer method as described by Gee andBauder (1996). Nitrate (NO3
−) and nitrite (NO2−) in
the K2SO4 extracts were determined colourimetrically(Mulvaney 1996).
The amount of CO2 and N2O was determined withan Agilent 4890D gas chromatograph fitted with an
electron capture detector. A J&W Scientific GS-Qcolumn was used to separate CO2 and N2O from theother gases; the flow rate of the carrier gas, N2, was5 ml min−1. Injection, detection and column-oventemperatures were set at 100°C, 225°C, and 35°C,respectively. An aliquot of 1 cm3 was injected into thechromatograph using a Teflon sealed glass syringe(Hamilton®, USA).
The emission of N2 was calculated as the differ-ence in N2O produced between the untreated soil andsoil treated with C2H2 multiplied with 0.8, i.e. fiveelectrons (e−) are used when NO3
− is reduced to N2,but only four when reduced to N2O (Dendooven et al.1994a, b).
Statistical analyses
Emission of N2O and CO2 was regressed on elapsedtime using a linear regression model which wasforced to pass through the origin but allowed differentslopes (production rates) for each treatment. Thisapproach is supported by theoretical considerationsthat no N2O and CO2 was produced at time zero andthe atmosphere in the flask contained no N2O andCO2 as the headspace was flushed with He. Produc-tion of NO2
− and NO3− was regressed on elapsed time
using a linear model not forced to pass through theorigin and allowed different slopes (production rates)for each treatment.
Soil characteristics were subjected to a one-wayanalysis of variance (ANOVA) using PROC GLM(SAS 1988) to test for significant differences betweentreatments with the Tukey’s Studentized Range test.Significant differences between treatments for N2O,NO2
−, NO3− and CO2 production rates were deter-
mined using PROC MIXED (SAS 1988). All analyseswere performed using SAS statistical package (1988).Each point in the figures is the mean of nine mea-surements, i.e. data from three plots in triplicate.
Results
Concentration of NO3−
The concentration of NO3− decreased linearly over
time in the Acolman soil (Fig. 1a). The NO3− reduction
rate was 0.78 mg N kg−1 h−1 in the unamended soil.C2H2 had no effect on the amount of NO3
− reduced
250 Plant Soil (2009) 319:247–257

compared to the unamended soil, but chloramphenicoldecreased the amount reduced (Table 2).
The concentration of NO3− decreased linearly over
time in the Texcoco soil with EC 12.6 dS m−1 (Fig. 1b).C2H2 had no significant effect on the concentrations ofNO3
− compared to the unamended soil, but chloram-phenicol inhibited the reduction of NO3
− significantly(P<0.05). The NO3
− reduction rate was 0.76 mg Nkg−1 h−1 in the unamended soil and 0.22 mg N kg−1 h−1
in the chloramphenicol amended soil.The concentration of NO3
− decreased linearlyover time in the unamended Texcoco soil with EC
56 dS m−1 and when treated with C2H2, but not inthe chloramphenicol-amended soil (Fig. 1c). TheNO3
− reduction rate was 1.16 mg N kg−1 h−1 in theunamended soil and no significant reduction tookplace in the chlorampehicol-amended soil.
Concentration of NO2−
The concentration of NO2− increased in the unamend-
ed Acolman soil to a maximum of 12 mg NO2− -N
kg−1 soil after 72 h (Fig. 2a). C2H2 had no significanteffect on the concentration of NO2
−, compared to the
Fig. 1 Concentration of NO3− (mg N kg−1 soil) from a Acolman
soil, b Texcoco soil with EC 12.6 dS m−1 and c Texcoco soil withEC 56 dS m−1 untreated (CON, □), treated with acetylene (ACE,
■), chloramphenicol (CHL, ○) or chloramphenicol plus acetylene(CHA, ●) incubated under strict anaerobic conditions at 25°C for72 h. Bars are ± 1 standard error of the estimates (P<0.05 and n=9)
Plant Soil (2009) 319:247–257 251

unamended soil, but chloramphenicol increased itsignificantly. The concentration of NO2
− increasednearly linearly in the chloramphenicol-amended soilto 36 mg NO2
− -N kg−1 soil after 72 h.The concentration of NO2
− increased in theunamended Texcoco soil with EC 12 dS m−1 to amaximum of 30 mg NO2
− -N kg−1 soil after 48 h(Fig. 2b). Chloramphenicol had no significant effecton the concentration of NO2
−, compared to theunamended soil, but C2H2 increased it. The NO2
−
production rate was 0.44 mg N kg−1 h−1 in theunamended soil and 0.55 mg N kg−1 h−1 in the C2H2-treated soil.
The concentration of NO2− increased in the
unamended Texcoco soil with EC 56 dS m−1 to amaximum of 52 mg NO2
− -N kg−1 soil after 48 h(Fig. 2c). C2H2 had no significant effect on theconcentration of NO2
−, compared to the unamendedsoil, but chloramphenicol decreased it significantly.The NO2
− production rate was 0.78 mg N kg−1 h−1
in the unamended soil, but no significant increaseoccurred in the chloramphenicol-amended soil.
Emission of N2O and N2
The emission of N2O in the unamended Acolman soilremained < 0.2 mg N kg−1 soil (Fig. 3a). Addition ofC2H2 increased the emission of N2O significantly and8.24 mg N kg−1 soil was produced after 72 h. Theemission of N2 was thus 6.52 mg N kg−1 soil after
72 h and calculated as the difference in N2O producedbetween the untreated soil and soil treated with C2H2
multiplied with 0.8. Chloramphenicol increased theproduction of N2O to 0.53 mg N kg−1 compared tothe untreated soil and the combined application ofchloramphenicol and C2H2 increased it to 1.06 mg Nkg−1. As such, the emission of N2 was 0.42 mg Nkg−1 in the chloramphenicol-amended soil after 72 h.
The emission of N2O in the unamended Texcocosoil with EC 12.6 dS m−1 was 0.01 mg N kg−1
(Fig. 3b). Addition of C2H2 increased the emission ofN2O significantly and 1.98 mg N kg−1 soil wasproduced after 72 h. The emission of N2 was thus1.58 mg N kg−1 soil after 72 h. Chloramphenicolincreased the production of N2O to 0.05 mg N kg−1
soil and the combined application of chloramphenicoland C2H2 to 0.20 mg N kg−1 soil. The emission of N2
was thus 0.09 mg N kg−1 soil in the chloramphenicol-amended soil after 72 h.
The emission of N2O in the unamended Texcocosoil with EC 56 dS m−1 was 1.32 mg N kg−1 soil after72 h (Fig. 3c). Addition of C2H2 increased theemission of N2O significantly and 3.87 mg N kg−1
soil was produced after 72 h. The emission of N2 wasthus 2.04 mg N kg−1 soil after 72 h. Chloramphenicoldecreased the emission of N2O to 0.01 mg N kg−1 soiland a similar amount was produced when chloram-phenicol and C2H2 were added to soil. No detectableN2 was produced in the chloramphenicol-amendedsoil after 72 h.
Table 2 Production of CO2 (mg C kg−1), NO3−, NO2
−, N2Oand N2 (mg N kg−1) from the Acolman, Texcoco soil withelectrolytic conductivity (EC) 12.6 dS m−1 (Texcoco-A) and EC
56 dS m−1 (Texcoco-B) treated with acetylene (C2H2),chloramphenicol and chloramphenicol+C2H2 under anaerobicconditions after 72 h
Layer Treatment Production of e− to reduce NOx
CO2-C e− NO3− -N NO2
− -N N2O-N N2-N
Acolman Unamended 13.9 56 −62.2 9.6 0.09 6.52 52Acetylene (C2H2) 16.8 67 −73.6 9.4 8.24 ND 52Chloramphenicol 18.4 74 −38.0 34.5 0.53 0.42 73Chloramphenicol+C2H2 19.3 77 −40.6 29.4 1.06 ND 72
Texcoco-A Unamended 3.4 14 −54.2 28.1 0.01 1.58 64Acetylene 3.8 15 −55.9 37.5 1.98 ND 83Chloramphenicol 7.2 29 −31.7 32.6 0.05 0.09 66Chloramphenicol+C2H2 10.6 42 −26.1 32.9 0.20 ND 66
Texcoco-B Unamended 13.1 52 −76.9 50.7 1.32 2.04 117Acetylene 10.8 43 −74.6 48.3 3.87 ND 112Chloramphenicol 13.8 55 −6.7 2.8 0.01 ND 6Chloramphenicol+C2H2 17.8 71 −13.1 2.8 0.01 ND 6
252 Plant Soil (2009) 319:247–257

Emission of CO2
The emission of CO2 resembled a zero-order kineticin the Acolman soil (Fig. 4a). Addition of C2H2 and/or chloramphenicol had no significant effect on theCO2 production rate. The emission of CO2 in theunamended Texcoco soil with EC 12.6 dS m−1 showeda flush at the onset of the incubation (Fig. 4b).Addition of C2H2 had no significant effect on theCO2 production rate, but addition of chloramphenicolto soil increased the emission of CO2 significantlycompare to the untreated soil (P<0.05). The emissionof CO2 in the unamended Texcoco soil with EC 56dS m−1 showed a lag at the onset of the incubation(Fig. 4c). Addition of C2H2 and/or chloramphenicolhad no significant effect on the CO2 production rate.
Discussion
In the unamended Acolman soil, the amount of NO3−
-N reduced was 4.4 times larger than the CO2-Cemitted after 72 h (Table 2). The amount of NO3
− -Nreduced in the Acolman soil amended with chloram-phenicol was 2.1 times larger than the CO2-Cproduced after 72 h. The ratio between CO2-Cproduced and NO3
− -N reduced is normally 1 whenall NO3
− -N is reduced to N2O-N or 0.80 whenreduced to N2-N. In the Acolman soil, however, largeconcentrations of NO2
− were detected, especially inthe chloramphenicol amended soil. Considering thatall e− generated by the oxidation of organic material,i.e. the CO2 produced is directly related to the e−
generated, are used to reduce NO3− to NOx, i.e. 2e
−
NO3− to NO2
−, 4 e− NO3− to N2O-N and 5 e− for
NO3− to N2-N, then the ratio between CO2-C and
Time (days)
Prod
uctio
n of
NO
2- (m
g N
kg-1
dry
soi
l)
0
12
24
36
48
60ACE CHACHL CON
0
12
24
36
48
60
0
12
24
36
48
60
0 10 20 30 40 50 60 70
a)
b)
c)
Fig. 2 Concentration of NO2− (mg N kg−1 soil) from a
Acolman soil, b Texcoco soil with EC 12.6 dS m−1 and cTexcoco soil with EC 56 dS m−1 untreated (CON, □), treatedwith acetylene (ACE, ■), chloramphenicol (CHL, ○) orchloramphenicol plus acetylene (CHA, ●) incubated under strictanaerobic conditions at 25°C for 72 h. Bars are ± 1 standard error ofthe estimates (P<0.05 and n=9)
Time (days)
Em
issi
on o
f N
2O (
mg
N k
g-1 d
ry s
oil)
0
2
4
6
8
10
12
ACE CHACHL CON
0.0
0.6
1.2
1.8
2.4
3.0
0
1
2
3
4
5
0 10 20 30 40 50 60 70
a)
b)
c)
Fig. 3 Emission of N2O (mg N kg−1 soil) from a Acolman soil,b Texcoco soil with EC 12.6 dS m−1 and c Texcoco soil withEC 56 dS m−1 untreated (CON, □), treated with acetylene(ACE, ■), chloramphenicol (CHL, ○) or chloramphenicol plusacetylene (CHA, ●) incubated under strict anaerobic conditions at25°C for 72 h. Bars are ± 1 standard error of the estimates (P<0.05and n=9)
Plant Soil (2009) 319:247–257 253
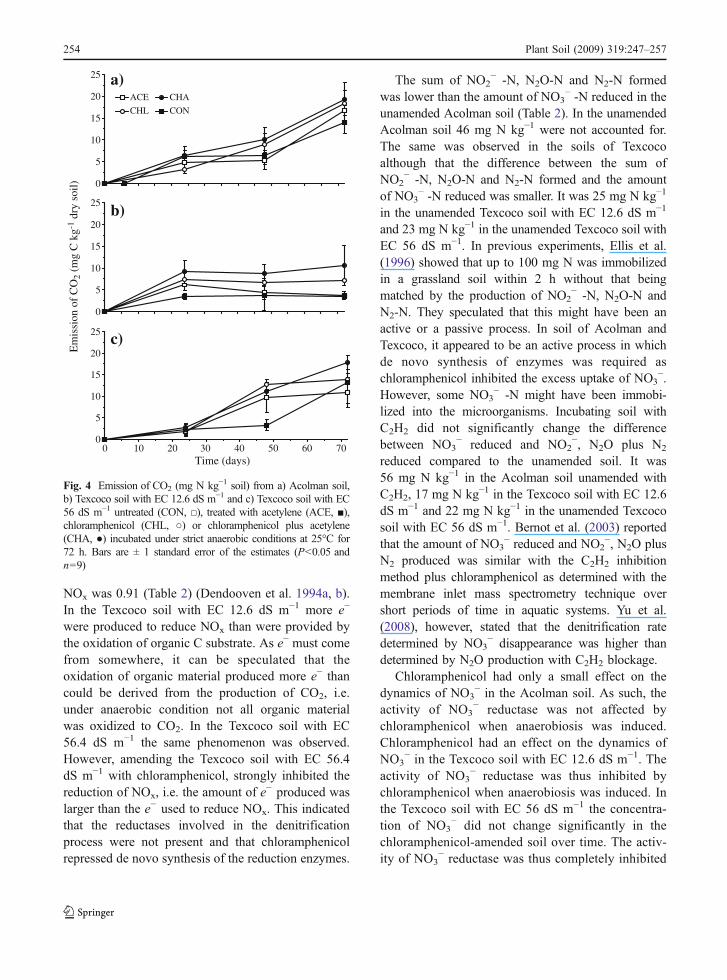
NOx was 0.91 (Table 2) (Dendooven et al. 1994a, b).In the Texcoco soil with EC 12.6 dS m−1 more e−
were produced to reduce NOx than were provided bythe oxidation of organic C substrate. As e− must comefrom somewhere, it can be speculated that theoxidation of organic material produced more e− thancould be derived from the production of CO2, i.e.under anaerobic condition not all organic materialwas oxidized to CO2. In the Texcoco soil with EC56.4 dS m−1 the same phenomenon was observed.However, amending the Texcoco soil with EC 56.4dS m−1 with chloramphenicol, strongly inhibited thereduction of NOx, i.e. the amount of e− produced waslarger than the e− used to reduce NOx. This indicatedthat the reductases involved in the denitrificationprocess were not present and that chloramphenicolrepressed de novo synthesis of the reduction enzymes.
The sum of NO2− -N, N2O-N and N2-N formed
was lower than the amount of NO3− -N reduced in the
unamended Acolman soil (Table 2). In the unamendedAcolman soil 46 mg N kg−1 were not accounted for.The same was observed in the soils of Texcocoalthough that the difference between the sum ofNO2
− -N, N2O-N and N2-N formed and the amountof NO3
− -N reduced was smaller. It was 25 mg N kg−1
in the unamended Texcoco soil with EC 12.6 dS m−1
and 23 mg N kg−1 in the unamended Texcoco soil withEC 56 dS m−1. In previous experiments, Ellis et al.(1996) showed that up to 100 mg N was immobilizedin a grassland soil within 2 h without that beingmatched by the production of NO2
− -N, N2O-N andN2-N. They speculated that this might have been anactive or a passive process. In soil of Acolman andTexcoco, it appeared to be an active process in whichde novo synthesis of enzymes was required aschloramphenicol inhibited the excess uptake of NO3
−.However, some NO3
− -N might have been immobi-lized into the microorganisms. Incubating soil withC2H2 did not significantly change the differencebetween NO3
− reduced and NO2−, N2O plus N2
reduced compared to the unamended soil. It was56 mg N kg−1 in the Acolman soil unamended withC2H2, 17 mg N kg−1 in the Texcoco soil with EC 12.6dS m−1 and 22 mg N kg−1 in the unamended Texcocosoil with EC 56 dS m−1. Bernot et al. (2003) reportedthat the amount of NO3
− reduced and NO2−, N2O plus
N2 produced was similar with the C2H2 inhibitionmethod plus chloramphenicol as determined with themembrane inlet mass spectrometry technique overshort periods of time in aquatic systems. Yu et al.(2008), however, stated that the denitrification ratedetermined by NO3
− disappearance was higher thandetermined by N2O production with C2H2 blockage.
Chloramphenicol had only a small effect on thedynamics of NO3
− in the Acolman soil. As such, theactivity of NO3
− reductase was not affected bychloramphenicol when anaerobiosis was induced.Chloramphenicol had an effect on the dynamics ofNO3
− in the Texcoco soil with EC 12.6 dS m−1. Theactivity of NO3
− reductase was thus inhibited bychloramphenicol when anaerobiosis was induced. Inthe Texcoco soil with EC 56 dS m−1 the concentra-tion of NO3
− did not change significantly in thechloramphenicol-amended soil over time. The activ-ity of NO3
− reductase was thus completely inhibited
Time (days)
Em
issi
on o
f C
O2
(mg
C k
g-1 d
ry s
oil) 0
5
10
15
20
25
ACE CHA
CHL CON
0
5
10
15
20
25
0
5
10
15
20
25
0 10 20 30 40 50 60 70
a)
b)
c)
Fig. 4 Emission of CO2 (mg N kg−1 soil) from a) Acolman soil,b) Texcoco soil with EC 12.6 dS m−1 and c) Texcoco soil with EC56 dS m−1 untreated (CON, □), treated with acetylene (ACE, ■),chloramphenicol (CHL, ○) or chloramphenicol plus acetylene(CHA, ●) incubated under strict anaerobic conditions at 25°C for72 h. Bars are ± 1 standard error of the estimates (P<0.05 andn=9)
254 Plant Soil (2009) 319:247–257

by chloramphenicol when anaerobiosis was induced. Itappears that the activity of NO3
− reductase in soil de-creased with increased pH and EC. It has to be remem-bered, however, that not only salt concentration and pHwere different between the soils used in this study butalso other characteristics, such as particle size distri-bution and organic matter content (Table 1). Little in-formation exists about how specific soil characteristicsmight affect reduction enzymes involved in the denitri-fication process, but possible effects cannot be exclud-ed although they are presumably small. For instance,the clay content is known to affect decomposition of14C labelled glucose in soil (van Veen et al. 1985) so itmight have an effect on the reductases studied.
In the unamended Acolman soil, the NO2− con-
centration was 9.6 mg N kg−1 soil after 72 h, 28.1 mgN kg−1 soil in the unamended Texcoco soil with EC12.6 dS m−1 and 56.7 mg N kg−1 in the unamendedTexcoco soil with EC 56 dS m−1. This would indicatethat the activity of NO2
− reductases or its de novosynthesis was inhibited with increased pH and saltconcentration. However, difference in the amount ofdenitrifying microorganisms in the different soilsmight have contributed to the differences in concen-trations of NO2
− observed.Production of N2O was low in the Acolman soil
and the Texcoco soil with EC 12.6 dS m−1 comparedto the production of N2. Nearly all N2O produced wasreduced to N2 so the activity of N2O reductase and/orits de novo synthesis was not inhibited. Simek et al.(2002) concluded in a study on the optimum pH fordenitrification in soil that at pH above 7, N2 is a moreimportant denitrification product than N2O. In theTexcoco soil with EC 56 dS m−1 the N2O-to-N2 ratiowas 0.65 so it appears that N2O reductases and/or itsde novo synthesis was inhibited. De novo synthesis ofN2O reductases occurred in the three soils aschloramphenicol reduced the production of N2. Initialconcentrations of N2O reductases, however, appearsto decrease with increased pH and EC as the produc-tion of N2 decreased with increased pH and EC inchloramphenicol-amended soil, but as mentionedbefore other soil characteristics might have affectedthe results obtained.
It has been reported that chloramphenicol notonly inhibits de novo synthesis of reduction enzymes,but might also reduce the activity of existingreduction enzymes (Brooks et al. 1992; Pell et al.
1996; Murray and Knowles 1999). Murray andKnowles (1999) stated that chloramphenicol at con-centrations of > 0.1 g l−1 or 0.3 mM, inhibited theactivity of the existing nitrate reductase, but not atlower concentrations. The amount of chloramphenicoladded in the experiment reported here was 0.23 mMand lower than the amount recommended by Murrayand Knowles (1999), but small negative effects on theactivity of the reduction enzymes can not be excluded(Pell et al. 1996). In the Acolman soil amended withchloramphenicol, the amount of e− generated byoxidation of organic material was matched by theamount of e− used to reduce NOx (Table 2). Thiswould indicate that chloramphenicol might changethe concentrations of NO3
−, NO2−, N2O and N2 in the
denitrification process through inhibition of theexisting enzymes, but not the reduction process itself.
The amount of CO2 produced was larger in thechloramphenicol amended soils compared to theunamended soils (Table 2). Similar results have beenreported for grassland soils (Dendooven et al. 1994a, b).The increase in C mineralization was presumably dueto a diversion of C supply from protein synthesis torespiratory processes (Smith and Tiedje 1979) and abiological degradation of chloramphenicol (Smith andWorrell 1950).
Conclusions
Under anaerobic conditions, NO3− was taken up in
excess of what was reduced to NO2− -N, N2O-N plus
N2. The process appeared to be active as chloram-phenicol negated that effect. The production of N2
was larger than that of N2O resulting in a low N2O-to-N2 ratio in soil with moderate amount of salts, butmore N2O was produced than N2 in the ‘extreme’alkaline saline soil under strict anaerobic conditions.The activity of NO2
− reductase appeared to decreasewith increased pH and EC, although other soilcharacteristics might have contributed to the process.The activity of the N2O reductase was only inhibitedin soil with EC 56 dS m−1.
Acknowledgements We thank A. Meza for technical assistanceand ‘Comisión Nacional de Agua’ (CNA) for access to theformer lake Texcoco. The research was funded by ‘ConsejoNacional de Ciencia y Tecnología’ (CONACyT) Méxicoresearch grant 39801-Z.
Plant Soil (2009) 319:247–257 255

References
Amato M (1983) Determinations of carbon 12C and 14C in plantand soil. Soil Biol Biochem 15:107–114 doi:10.1016/0038-0717(83)90059-7
Balderstone WL, Scherr B, Payne WJ (1976) Blockage byacetylene of nitrous oxide reduction in Pseudomonasperfectomarinus. Appl Environ Microbiol 31:504–508
Beltrán-Hernández RI (2001) Descomposición de lodos residualesen suelos del ex-Lago de Texcoco. PhD thesis Cinvestav,México DF, México
Beltrán-Hernández RI, Coss-Muñoz E, Luna-Guido ML,Mercado-García F, Siebe C, Dendooven L (1999) C andN dynamics in alkaline saline soil of the former LakeTexcoco (Mexico) as affected by application of sewagesludge. Eur J Soil Sci 50:601–608 doi:10.1046/j.1365-2389.1999.00270.x
Bernot MJ, Dodds WK, Gardner WS, McCarthy MJ, Sobolev D,Tank JL (2003) Comparing denitrification rates estimatesfor a Texas estuary by using acetylene inhibition andmembrane inlet mass spectrometry. Appl Environ Micro-biol 69:5950–5956 doi:10.1128/AEM.69.10.5950-5956.2003
Bremner JM (1996) Total nitrogen. In: Sparks DL (ed) Methodsof soil analysis: Chemical methods. Soil Science Societyof America, Madison, pp 1085–1121 part 3
Brooks MH, Smith RL, Macalady DL (1992) Inhibition ofexisting denitrification enzyme-acyivity by chloramphenicol.Appl Environ Microbiol 58:1746–1753
Castro-Silva C, Luna-Guido M, Marsch R, Ceballos JM,Dendooven L (2008) Production of CO2 and N2O inalkaline saline soil of Texcoco at different water contentsamended with urea. Soil Biol Biochem 40:1813–1822doi:10.1016/j.soilbio.2008.03.004
Dendooven L, Splatt P, Anderson JM (1994a) The use ofchloramphenicol in the study of the denitrification process:some side-effects. Soil Biol Biochem 26:925–927doi:10.1016/0038-0717(94)90309-3
Dendooven L, Splatt P, Anderson JM, Scholefield D (1994b)Kinetics of the denitrification process in a soil underpermanent pasture. Soil Biol Biochem 26:361–370doi:10.1016/0038-0717(94)90285-2
Ellis S, Dendooven L, Goulding KWT (1996) Quantitativeassessment of soil nitrate disappearance and N2O evolutionduring denitrification: Nitrate disappearance during denitri-fication. Soil Biol Biochem 28:589–595 doi:10.1016/0038-0717(95)00181-6
Firestone MK, Firestone RB, Tiedje JM (1980) Nitrous oxidefrom soil denitrification: factors controlling its biologicalproduction. Science 208:749–751 doi:10.1126/science.208.4445.749
Gee GW, Bauder JW (1996) Particle-size analysis. In: Klute A(ed) Methods of soil analysis: Physical and mineralogicalmethods. Soil Science Society of America, Madison,pp 383–411 part 1
Gutiérrez-Castorena MC (1997) Los suelos de la rivera orientaldel ex Lago de Texcoco (macro- y micro-morfología).PhD thesis, Colegio de Postgraduados de Chapingo,México, p 158
INEGI (1994) Guide for technical tours ‘3’ and ‘12’ MexicoCity, Acapulco. Fifteenth World Congress of Soil Science,Acapulco, México, Instituto Nacional de Estadística,Geografía e Informática. Aguascalientes, México
Kiese R, Butterbach-Bahl K (2002) N2O and CO2 emissionsfrom three different tropical forest sites in the wet tropicsof Queensland, Australia. Soil Biol Biochem 34:975–987doi:10.1016/S0038-0717(02)00031-7
Liang Y, Nikolic M, Peng Y, Chen W, Jiang Y (2005) Organicmanure stimulates biological activity and barley growth insoil subject to secondary salinization. Soil Biol Biochem37:1185–1195 doi:10.1016/j.soilbio.2004.11.017
Luna-Guido ML, Beltrán-Hernández RI, Solis-Ceballos NA,Hernández-Chavez N, Mercado-García F, Olalde-PortugalV, Catt JA, Dendooven L (2000) Chemical and biologicalcharacteristics of alkaline saline soils from the formerLake Texcoco as affected by artificial drainage. Biol FertilSoils 32:102–108 doi:10.1007/s003740000223
Moraghan JT, Buresh RJ (1977) Correction for dissolvednitrous oxide in nitrogen studies. Soil Sci Soc Am J41:1201–1202
Mulvaney RL (1996) Nitrogen—Inorganic forms. In: Sparks DL(ed) Methods of soil analysis: Chemical methods Part 3.Soil Science Society of America Inc, American Society ofAgronomy, Inc Madison, Wisconsin, USA, pp 1123–1184
Murray RE, Knowles R (1999) Chloramphenicol inhibition ofdenitrifying enzyme activity in two agricultural soils. ApplEnviron Microbiol 65:3487–3492
Nelson PN, Ladd JN, Oades JM (1996) Decomposition of 14C-labelled plant material in salt-affected soil. Soil BiolBiochem 28:433–441 doi:10.1016/0038-0717(96)00002-8
Nelson DW, Sommers LE (1996) Total carbon, organic carbonand organic matter. In: Sparks DL (ed) Methods of soilanalysis: Chemical methods. Soil Science Society ofAmerica, Madison, pp 961–1010 part 3
Oren A (1999) Bioenergetic aspects of halophilism. MicrobiolMol Biol Rev 63:334–348
Pell M, Stenberg B, Stenström J, Thorstensson L (1996)Potential denitrification activity assay in soil- with orwithout chloramphenicol. Soil Biol Biochem 28:393–398doi:10.1016/0038-0717(95)00149-2
Rengasamy P (2006) World salinization with emphasis onAustralia. J Exp Bot 57:1017–1023 doi:10.1093/jxb/erj108
Sánchez L, Díez JA, Vallejo A, Cartagena MC (2001)Denitrification losses from irrigated crops in central Spain.Soil Biol Biochem 33:1201–1209 doi:10.1016/S0038-0717(01)00024-4
SAS Institute (1988) Statistic Guide for Personal Computers.Version 6.03, Edn SAS Institute, Cary
Simek M, Jisova L, Hopkins DW (2002) What is the so-calledoptimum pH for denirtrification in soil. Soil Biol Biochem34:1227–1234 doi:10.1016/S0038-0717(02)00059-7
Smith SM, TiedjeJM (1979) Phases of denitrification follow-ing oxygen depletion in soil. Soil Biol Biochem 11:261–267
Smith GN, Worrell CS (1950) Decomposition of chloromycetin(chloramphenicol) by micro-organisms. Arch Biochem28:232–242
Tejada M, Garcia C, Gonzalez JL, Hernandez MT (2006) Useof organic amendment as a strategy for saline soil
256 Plant Soil (2009) 319:247–257

remediation: influence on the physical, chemical andbiological properties of soil. Soil Biol Biochem 38:1413–1421
Thomas GW (1996) Soil pH and soil acidity. In: SparksDL (ed) Methods of soil analysis: chemical methods.Soil Science Society of America, Madison, pp 475–490part 3
van Veen JA, Ladd JN, Amato M (1985) Turnover of carbonand nitrogen through the microbial biomass in a sandyloam and a clay soil incubated with [14C(U)]glucose and[15N](NH4)2SO4 under different moisture regimes. SoilBiol Biochem 17:747–756
Valenzuela-Encinas C, Neria-González I, Alcántara-HernándezRJ, Enriques-Aragón JA, Hernandez-Rodriguez CH,Dendooven L, Marsch R (2008) Phylogenetic analysis ofarchaeal community in an alkaline-saline soil of the formerlake Texcoco (Mexico). Extremophiles 12:247–254
Wrage N, Velthof GL, van Beusichem ML, Oenema O (2001)Role of nitrifier denitrification in the production of nitrousoxide. Soil Biol Biochem 33:1723–1732
Yu K, Struwe S, Kjøller A, Chen G (2008) Denitrification ratedetermined by nitrate disappearance is higher thandetermined by nitrous oxide production with acetyleneblockage. Ecol Engin 32:90–96
Plant Soil (2009) 319:247–257 257




















