!["% TZgZ]ZR_d \Z]]VU Z_ ?RXR]R_U - Daily Pioneer](https://static.fdokumen.com/doc/165x107/6321f3d0aa6c954bc7077bd8/-tzgzzrd-zvu-z-rxrru-daily-pioneer.jpg)
Delivery of nucleic acids with a stearylated (RxR) 4 peptide using a non-covalent co-incubation...
10
Delivery of nucleic acids with a stearylated (RxR) 4 peptide using a non-covalent co-incubation strategy Taavi Lehto a,c, ⁎, Rachida Abes b , Nikita Oskolkov a , Julia Suhorutšenko a , Dana-Maria Copolovici a , Imre Mäger a,c , Joana R. Viola d , Oscar E. Simonson d , Kariem Ezzat c , Peter Guterstam c , Elo Eriste a , C.I. Edvard Smith d , Bernard Lebleu b , Samir EL Andaloussi a,c , Ülo Langel a,c a Laboratory of Molecular Biotechnology, Institute of Technology, University of Tartu, Nooruse 1, 50411 Tartu, Estonia b UMR 5235 CNRS, Université Montpellier 2, Place Eugene Bataillon, 34095 Montpellier cedex 5, France c Department of Neurochemistry, The Arrhenius Laboratories for Natural Sciences, Stockholm University, SE-10691 Stockholm, Sweden d Karolinska University Hospital Huddinge, SE-141 86 Huddinge, Sweden abstract article info Article history: Received 21 May 2009 Accepted 28 August 2009 Available online 8 September 2009 Keywords: Cell-penetrating peptide Gene delivery Oligonucleotide delivery Splice correction Stearylation In recent years, oligonucleotide-based molecules have been intensely used to modulate gene expression. All these molecules share the common feature of being essentially impermeable over cellular membranes and they therefore require efficient delivery vectors. Cell-penetrating peptides are a group of delivery peptides that has been readily used for nucleic acid delivery. In particular, polyarginine and derivates thereof, i.e. the (RxR) 4 peptide, have been applied with success both in vitro and in vivo. A major problem, however, with these arginine-rich peptides is that they frequently remain trapped in endosomal compartments following internalization. The activity of polyarginine has previously been improved by conjugation to a stearyl moiety. Therefore, we sought to investigate what impact such modification would have on the pre-clinically used (RxR) 4 peptide for non-covalent delivery of plasmids and splice-correcting oligonucleotides (SCOs) and compare it with stearylated Arg9 and Lipofectamine™ 2000. We show that stearyl-(RxR) 4 mediates efficient plasmid transfections in several cell lines and the expression levels are significantly higher than when using unmodified (RxR) 4 or stearylated Arg9. Although the trans- fection efficiency is lower than with Lipofectamine™ 2000, we show that stearyl-(RxR) 4 is substantially less toxic. Furthermore, using a functional splice-correction assay, we show that stearyl-(RxR) 4 complexed with 2′-OMe SCOs promotes significant splice correction whereas stearyl-Arg9 fails to do so. Moreover, stearyl- (RxR) 4 promotes dose-dependent splice correction in parity with (RxR) 4 -PMO covalent conjugates, but at least 10-times lower concentration. These features make this stearic acid modified analog of (RxR) 4 an intriguing vector for future in vivo experiments. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Scientific advances in last decades have laid the foundation for once utopian gene therapy to be probably accessible in the near future. The underlying principle of gene therapy is to deliver plasmid DNA carrying genes of interest into the cell nucleus and express otherwise deficient gene product to the normal physiological levels. In recent years, other techniques for gene expression modulation have emerged, which affect biological processes at a post-transcriptional level. These antisense- based techniques are mediated by short oligonucleotides (ONs), for instance, splice-correcting oligonucleotides (SCOs). As many diseases are associated with incorrect splicing patterns, utilization of SCOs is a plausible strategy to interfere with splicing pathways and correct dis- ease phenotypes [1]. Most of the molecules used in gene expression modulation have large molecular weights and are hydrophilic. Such characteristics pre- vent these molecules to cross cellular membranes, unless associated with a suitable delivery vehicle. To solve this limitation, plenty of de- livery vectors, both viral and non-viral, have been developed. Viral vectors are highly efficient but suffer from low cargo carrying capacity, immunogenicity and possible viral recombination in vivo [2]. Those major drawbacks in viral delivery have initiated intense search for efficient non-viral delivery vectors. Cell-penetrating peptides, CPPs, are one group of such delivery vectors, which has gained wide interest since their initial discovery Journal of Controlled Release 141 (2010) 42–51 Abbreviations: 2′-OMe ON, Phosphorothioate 2′-O-methyl RNA; Arg9, Nona- arginine; CPP, Cell-penetrating peptide; CR, Charge ratio; DLS, Dynamic light scattering; HKR, HEPES-buffered Krebs Ringer solution; MR, Molar ratio; ON, Oligonucleotide; Pen, Penetratin; SCO, Splice-correcting oligonucleotide; TFA, Trifluoroacetc acid; TP10, Transportan 10. ⁎ Corresponding author. Laboratory of Molecular Biotechnology, Institute of Tech- nology, University of Tartu, Nooruse 1, 50411 Tartu, Estonia. Tel.: +372 7 37 4866; fax: +372 7 37 4900. E-mail address: [email protected] (T. Lehto). 0168-3659/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.jconrel.2009.08.028 Contents lists available at ScienceDirect Journal of Controlled Release journal homepage: www.elsevier.com/locate/jconrel
Transcript of Delivery of nucleic acids with a stearylated (RxR) 4 peptide using a non-covalent co-incubation...

Journal of Controlled Release 141 (2010) 42–51
Contents lists available at ScienceDirect
Journal of Controlled Release
j ourna l homepage: www.e lsev ie r.com/ locate / jconre l
Delivery of nucleic acids with a stearylated (RxR)4 peptide using a non-covalentco-incubation strategy
Taavi Lehto a,c,⁎, Rachida Abes b, Nikita Oskolkov a, Julia Suhorutšenko a, Dana-Maria Copolovici a,Imre Mäger a,c, Joana R. Viola d, Oscar E. Simonson d, Kariem Ezzat c, Peter Guterstam c, Elo Eriste a,C.I. Edvard Smith d, Bernard Lebleu b, Samir EL Andaloussi a,c, Ülo Langel a,c
a Laboratory of Molecular Biotechnology, Institute of Technology, University of Tartu, Nooruse 1, 50411 Tartu, Estoniab UMR 5235 CNRS, Université Montpellier 2, Place Eugene Bataillon, 34095 Montpellier cedex 5, Francec Department of Neurochemistry, The Arrhenius Laboratories for Natural Sciences, Stockholm University, SE-10691 Stockholm, Swedend Karolinska University Hospital Huddinge, SE-141 86 Huddinge, Sweden
Abbreviations: 2′-OMe ON, Phosphorothioate 2′-arginine; CPP, Cell-penetrating peptide; CR, Charge ratioHKR, HEPES-buffered Krebs Ringer solution; MR, Molar rPenetratin; SCO, Splice-correcting oligonucleotide; TFTransportan 10.⁎ Corresponding author. Laboratory of Molecular Bio
nology, University of Tartu, Nooruse 1, 50411 Tartu, Efax: +372 7 37 4900.
E-mail address: [email protected] (T. Lehto).
0168-3659/$ – see front matter © 2009 Elsevier B.V. Adoi:10.1016/j.jconrel.2009.08.028
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 May 2009Accepted 28 August 2009Available online 8 September 2009
Keywords:Cell-penetrating peptideGene deliveryOligonucleotide deliverySplice correctionStearylation
In recent years, oligonucleotide-based molecules have been intensely used to modulate gene expression. Allthese molecules share the common feature of being essentially impermeable over cellular membranes andthey therefore require efficient delivery vectors. Cell-penetrating peptides are a group of delivery peptidesthat has been readily used for nucleic acid delivery. In particular, polyarginine and derivates thereof, i.e. the(RxR)4 peptide, have been applied with success both in vitro and in vivo. A major problem, however, withthese arginine-rich peptides is that they frequently remain trapped in endosomal compartments followinginternalization. The activity of polyarginine has previously been improved by conjugation to a stearyl moiety.Therefore, we sought to investigate what impact such modification would have on the pre-clinically used(RxR)4 peptide for non-covalent delivery of plasmids and splice-correcting oligonucleotides (SCOs) andcompare it with stearylated Arg9 and Lipofectamine™ 2000.We show that stearyl-(RxR)4 mediates efficient plasmid transfections in several cell lines and the expressionlevels are significantly higher than when using unmodified (RxR)4 or stearylated Arg9. Although the trans-fection efficiency is lower than with Lipofectamine™ 2000, we show that stearyl-(RxR)4 is substantially lesstoxic. Furthermore, using a functional splice-correction assay, we show that stearyl-(RxR)4 complexed with2′-OMe SCOs promotes significant splice correction whereas stearyl-Arg9 fails to do so. Moreover, stearyl-(RxR)4 promotes dose-dependent splice correction in parity with (RxR)4-PMO covalent conjugates, but atleast 10-times lower concentration. These features make this stearic acid modified analog of (RxR)4 anintriguing vector for future in vivo experiments.
O-methyl RNA; Arg9, Nona-; DLS, Dynamic light scattering;atio; ON, Oligonucleotide; Pen,A, Trifluoroacetc acid; TP10,
technology, Institute of Tech-stonia. Tel.: +372 7 37 4866;
ll rights reserved.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Scientific advances in last decades have laid the foundation for onceutopian gene therapy to be probably accessible in the near future. Theunderlying principle of gene therapy is to deliver plasmid DNA carryinggenes of interest into the cell nucleus and express otherwise deficientgene product to the normal physiological levels. In recent years, othertechniques for gene expressionmodulation have emerged, which affect
biological processes at a post-transcriptional level. These antisense-based techniques are mediated by short oligonucleotides (ONs), forinstance, splice-correcting oligonucleotides (SCOs). As many diseasesare associated with incorrect splicing patterns, utilization of SCOs is aplausible strategy to interfere with splicing pathways and correct dis-ease phenotypes [1].
Most of the molecules used in gene expression modulation havelarge molecular weights and are hydrophilic. Such characteristics pre-vent these molecules to cross cellular membranes, unless associatedwith a suitable delivery vehicle. To solve this limitation, plenty of de-livery vectors, both viral and non-viral, have been developed. Viralvectors are highly efficient but suffer from low cargo carrying capacity,immunogenicity and possible viral recombination in vivo [2]. Thosemajor drawbacks in viral delivery have initiated intense search forefficient non-viral delivery vectors.
Cell-penetrating peptides, CPPs, are one group of such deliveryvectors, which has gained wide interest since their initial discovery

Table 1Sequences of cell-penetrating peptides and 2′-OMe and PMO oligonucleotides.
Name Sequence
(RxR)4 RXRRXRRXRRXRa
stearyl-(RxR)4 bRXRRXRRXRRXRa
stearyl-Arg9 bRRRRRRRRRa
Phosphorothioate2′-OMe RNA
c5′-CCU CUU ACC UCA GUU ACA-3′
PMO705 XB-CCT CTT ACC TCA GTT ACA-3′
X — 6-aminohexanoic acid.B — β-Alanine.
a C-terminal amide for co-incubation strategy.b N-terminally stearylated.c Cy5-labeled.
43T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
in 1994 [3]. CPPs are cationic and/or amphipathic peptides whichhave been shown to successfully deliver a wide range of differentmacromolecules across biological membranes, both in vivo and invitro [4–6].
Arginine-rich peptides are one of the most widely utilized class ofCPPs (reviewed in [7,8]). Recently one particular arginine-rich peptidehasbeenusedwith substantial success. (RxR)4 is anoligoarginine analog,which has an α-aminohexanoic acid linker (x) between arginineresidues [9]. (RxR)4 has been extensively studied after it was foundthat it is capable of mediating delivery of SCOs, consequently enablingredirection of pre-mRNA splicing and production of functional protein.It hasbeendescribed in in vitro systems— like theHeLapLuc 705 cell line[10,11], but it has also been shown to facilitate splicing redirectionin vivo. Conjugates of (RxR)4 to phosphorodiamidate morpholinooligomer (PMO) or peptide nucleic acid (PNA) ON analogs, were capableof promoting exon skipping and production of functional dystrophinin the mdx Duchenne muscular dystrophy murine model [12,13].
The internalization mechanism of CPPs remains debated but it iswidely accepted that most arginine-rich CPPs utilize some pathwaysof endocytosis to enter cells, at least at lower concentrations [14,15].Emanating from an endocytotic internalization pathway, CPPs remainmostly entrapped in endocytic vesicles, thus limiting the cargo avail-ability in the intracellular milieu, such as the cytoplasm or nucleus[16,17]. This limitation is now recognized as themajor barrier to over-come for the delivery of molecules that have to reach intracellularcompartments in order to exert their biological effect. Different strat-egies have been used to overcome endosomal entrapment, as recentlyreviewed for arginine-rich peptides [18]. One easily applicablemethodis to exploit lysosomotropic agents, such as chloroquine. However, itis only applicable in vitro, due to chloroquine pharmacological effectsin vivo, but even in vitro it has been shown to be substantially toxic[19]. As a result, intensified search for less toxic possibilities, whichwould be applicable in therapeutic settings, has been ongoing. For ex-ample, the Influenza virus-derived fusogenic HA2 peptide has beencovalently linked to CPPs, thereby facilitating endosomal escape [19,20]. Also, Lundberg et al. designed an endosomolytic analog of pene-tratin, EB1, which is thought to form an amphiphatic alpha helix in theslightly acidic environment of early–late endosomes through intro-duction of histidine moieties and to disrupt endosomal membranes[20].
Recently some groups have shown that stearylation of CPPs can be asuccessful strategy to improve delivery of ONs and plasmids [21–23].We have shown previously that stearylation of TP10 promotes efficientSCO delivery utilizing a non-covalent vectorization approach [23]. Al-though delivery of plasmidswith CPPs has been shownnumerous times[24–27], the relative efficacy of unmodified CPPs in promoting plasmiddelivery has been poor in general. Stearyl modification has been one ofthe most efficient strategies for plasmid delivery, as a lonemodificationon polyarginine [21,28] or if incorporated into more sophisticated de-livery devices, like multifunctional envelope-type nano device (MEND)[29,30].
In this study, we evaluate the effect of N-terminal stearylation of the(RxR)4 peptide on the delivery of plasmids and 2′-OMe SCOs utilizing anon-covalent co-incubation strategy. We also compare the effect of thesame modification on Arg9 peptide. Our results show that stearylationsignificantly increases plasmid deliverywith the (RxR)4 peptide and thedelivery efficacy is clearly superior to both unmodified (RxR)4 and ste-arylated Arg9. Plasmid delivery with stearyl-(RxR)4 is not as efficient aswith commercially available transfection agent Lipofectamine™ 2000,but at the same time stearyl-(RxR)4 is substantially less toxic. We alsoshow that stearyl-(RxR)4 significantly promotes the delivery of 2′-OMeSCOs, utilizing the same co-incubation approach, and the effects are inparitywith that of the pre-clinically used (RxR)4-PMO conjugate used at10-times higher concentrations. The versatility of the stearyl-(RxR)4 incombinationwith the non-toxic propertiesmakes it a highly interestingvector for future in vivo use.
2. Materials and methods
2.1. Synthesis of oligonucleotides and peptides
Phosphorothioate 2′-O-methyl RNA oligonucleotides were synthe-sized on an ÄKTA™ oligopilot™ plus 10 synthesizer with Oligosynt™15 (GE Healthcare, Sweden) pre-packed synthesis columns as pre-viously described [31]. The molarity of ONs was determined by OD-measurements.
The peptides (Table 1) were synthesized in a stepwise mannerat a 0.1mmol scale on an automated peptide synthesizer (AppliedBiosystems™model 433A) using Fmoc (fluorenylmethyloxycarbonyl)solid-phase peptide synthesis strategy. Rink-amide MBHA (methyl-benzylhydrylamine) resin (Fluka)was used as a solid support to obtainC-terminally amidated peptides. N-terminally stearylated peptideswere prepared by treatment of peptide resins with 4 Eq. stearic acid(Sigma, Germany) and 4 Eq. HOBt/HBTU (MultiSynTech, Germany)and 8 Eq. DIEA (Fluka) in DMF for 60 min. The final cleavage wasperformed using a standard protocol (95% TFA/2.5% TIS/2.5% water)for 2 h at room temperature. Peptides were purified by a reversed-phase HPLC using C18 column (SP 250/10 Nucleosil Macherey-Nagel,Germany) and a gradient of 5–80% acetonitrile/water containing 0.1%TFA. The molecular weight of peptides was analyzed by MALDI-TOFmass-spectroscopy and purity was N 95% as determined by analyticalHPLC. Themolarity of the peptides was determined based on dilutionsof accurately weighed substances. (RxR)4-PMO was synthesized aspreviously described [10].
2.2. Cell cultures
CHO cells were grown at 37 °C, 5% CO2 in Dulbecco's ModifiedEagle's Medium F12 (DMEM-F12) with glutamax supplement with0.1 mMnon-essential amino acids, 1.0 mM sodiumpyruvate, 10% fetalbovine serum, 100U/ml penicillin and 100 mg/ml streptomycin (PAALaboratories GmbH, Germany).
BHK21 cells were grown at 37 °C, 5% CO2 in Glasgow's ModifiedEagle's Medium (GMEM) with glutamax supplement with 0.1 mMnon-essential amino acids, 1.0 mM sodium pyruvate, with tryptosephosphate (AppliChem, Germany), 10% fetal bovine serum, 100U/mlpenicillin and 100 mg/ml streptomycin (PAA Laboratories GmbH,Germany).
HEK293 cells were grown at 37 °C, 5% CO2 in Dulbecco's ModifiedEagle's Medium (DMEM) with glutamax supplemented with 0.1 mMnon-essential amino acids, 1.0 mM sodium pyruvate, 10% fetal bovineserum, 100 U/ml penicillin and 100 mg/ml streptomycin (PAA Labora-tories GmbH, Germany).
HeLa pLuc 705 cells, kindly provided by Ryszard Kole, were grownat 37 °C, 5% CO2 in Dulbecco's Modified Eagle's Medium (DMEM)withglutamax supplemented with 0.1 mM non-essential amino acids,1.0 mM sodium pyruvate, 10% fetal bovine serum, 100U/ml penicillinand 100 mg/ml streptomycin (PAA Laboratories GmbH, Germany).

44 T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
2.3. Complex formation
0.5 µg of pGL3 luciferase expressing plasmid was mixed with CPPsat different charge ratios (CRs) (1:1–1:5) in MQwater in 1/10th of thefinal treatment volume (i.e. 50 µl). CRs were calculated theoretically,taking into account the positive charges of the peptide and negativecharges of the plasmid. For instance, final concentration of stearyl-(RxR)4 was 0.38 μM at the 1:1 CR. Complexes were formed for 1 h atroom temperature. Meanwhile, cell medium was replaced in 24-wellplates for fresh serum-free media (450 µl). In case of Lipofectamine™2000 (Invitrogen, Sweden), the complexes were formed according tothe manufacturer's protocol.
DNA condensationwas analyzed using an ethidiumbromide (EtBr)(Sigma, Sweden) exclusion assay. Briefly, complexes were formed asdescribed above. After 1 h incubation, 135μl MQ water was addedto each sample and transferred into a black 96-well plate (NUNC,Sweden). Thereafter, 15 µl of EtBr solution was added to give a finalEtBr concentration of 400nM. After 10 min, fluorescence was mea-sured on a Spectra Max Gemini XS fluorometer (Molecular Devices,Palo Alto, CA, USA) at λex=518 nm and λem=605 nm. Results aregiven as relative fluorescence and a value of 100% is attributed to thefluorescence of naked DNA with EtBr.
Stability of plasmid:stearyl-(RxR)4 complexes was evaluated inthe presence of serum. Briefly, complexes were formed as describedabove. Thereafter, serum was added to the complexes at differentconcentrations (5, 10 and 50%) and incubated over different periods oftime. At 0, 1, 4 and 24 h samples were loaded on an agarose gel (2%)and imaged by staining the gel with EtBr (0.5 µg/ml).
2′-OMe ON was mixed with CPPs at different molar ratios in MQwater in 1/10th of the final treatment volume (i.e. 50 µl). Complexeswere formed for 1 h at room temperature and meanwhile the cellmedium was replaced in 24-well plates to fresh serum-free DMEM(450 µl). Thereafter, complexes were added to each well. When usingLipofectamine™ 2000 (Invitrogen, Sweden), the complexes wereprepared according to the manufacturer's protocol. Complexes for-mation between 2′-OMe ON and peptides was analyzed in the EtBrexclusion assay in the same way as with plasmids (see above).Stearyl-(RxR)4:2′-OMe SCO complexes were analyzed towards serumstability as described above for the plasmid:peptide complexes.
2.4. Plasmid delivery assays
4×104 CHO cells, 3.5×104 BHK 21 cells, or 5×104 HEK cells wereseeded 24 h before the experiment into 24-well plates. Cells weretreated with plasmid:CPP complexes at different charge ratios (1:1,1:2, 1:3, 1:5) for 4 h in serum-freemedia followedby addition of 500 µlof full growth media and incubated for another 20 h. Thereafter, thecells were washed twice with HKR and lysed using 100 µl 0.2% TritonX-100 in HKR buffer for 30 min at room temperature. Luciferaseactivity was measured using Promega's luciferase assay system onGLOMAX™ 96 microplate luminometer (Promega, Sweden) and nor-malized to protein content (Lowry, BioRad, USA). Lipofectamine™2000 (Invitrogen, Sweden) was used according to the manufacturer'sprotocol.
7×104 CHO cells were seeded 24 h before experiment onto 13 mmtissue culture coverslips that were placed into a 24-well plate. Cellswere treated with pEGFP-C1 plasmid (plasmid expressing enhancedgreen fluorescent protein) and CPP complexes at different charge ra-tios (1:3, 1:5) for 4 h in serum-free media followed by addition of500 µl of full growth media and incubated for another 20 h at 37 °C.Lipofectamine™ 2000 (Invitrogen, Sweden)was used according to themanufacturer's protocol. Thereafter, cells were washed with PBS andfixed by using 4% formaldehyde solution at room temperature for10 min. Imageswere capturedusing 60-fold objective onNikonEclipseTE2000-U inverted microscope and a Digital Camera DXM1200C, andprocessed with EZ-C1 software V.2.30 (Nikon, Japan).
2.5. Splice-correction assay
HeLa pLuc 705 cells (5×104) were seeded 24 h prior to experimentsinto 24-well plates. Cells were treated with peptide:2′-OMe ON com-plexes at differentmolar ratios (2:1, 3:1, 5:1, 6:1, 10:1, and 20:1) for 4 hin serum-free media (or in serum-containing media) followed by ad-dition of 500 µl serum-containing medium and incubated for another20 h. For (RxR)4-PMO conjugates, cells were treated for 4 h in Opti-MEM®, followed by replacement to serum-containing medium and in-cubated additionally for 20 h. Thereafter, the cells were washed andlysed using 100 µl 0.2% Triton X-100 in HKR buffer for 30 min at roomtemperature. Luciferase activity was measured using Promega's lucif-erase assay system on GLOMAX™ 96 microplate luminometer (Pro-mega, Sweden) and normalized to protein content (Lowry, BioRad,USA).
2.6. Flow cytometry
HeLa pLuc 705 cells (2×105) were seeded 24 h prior to experi-ments into 24-well plates. Cells were treated with peptide and Cy5-labeled 2′-OMe ON complexes at molar ratio 1:6 and incubated for 2 hin serum-free Opti-MEM® media. Thereafter, cells were washed twicewith PBS and detached from the plate by trypsinization for 5 min at37 °C. Cells were suspended in ice-cold PBS containing 5% FBS andcentrifuged (900×g for 5 min). Supernatant was removed and theresulting cell pellet was resuspended in ice-cold PBS containing 0.5%FBS and 0.05 mg/ml propidium iodide (PI) (Molecular Probes, Eugene,OR, USA) for staining cells with non-intact plasma membrane. Flowcytometry analysis was carried out with a BD FacsCanto flow cyto-meter (BD Biosciences, San Jose, CA, USA).
Population of viable cells was determined from a scatter plot (for-ward scattered light (FSC) vs. side scattered light (SSC) plot). PI stainingwas used as a control to verify that plasma membranes of cells in theviable population were intact. A minimum of 10,000 events from theviable cell population per sample were analyzed. The percentage oftransfected cells was determined by counting the cells displaying Cy5fluorescent signal above a threshold level. The uptakewas characterizedby calculating mean Cy5 fluorescence value for transfected cells andcomparing it with the same parameter of untreated cells. Aforemen-tioned analysiswas carried out using FacsDiva software (BDBiosciences,San Jose, CA, USA).
2.7. LDH leakage assay
Membrane integrity was evaluated using LDH leakage assay(SIGMA-ALDRICH™ St. Louise, MO, USA), which measures lactatedehydrogenase release. Briefly, 1×104 cells were seeded 1 day beforetreatment into 96-well plate. Cells were treated with plasmid:CPPcomplexes at different charge ratios in serum-free media for 30 min.Thereafter, LDH leakage assay was used according to the manu-facturer's protocol. Untreated cells were defined as no leakage and100% leakage was defined as total LDH release by lysing cells in 0.2%Triton X-100 in HKR buffer.
2.8. MTS proliferation assay
Long-term toxicity was evaluated using an MTS proliferationassay (Promega, Madison, WI, USA). The MTS assay measures theactivity of mitochondrial dehydrogenases to convert tetrazoliumsalts into formazan. Briefly, 1×104 cells were seeded 1 day beforetreatment into 96-well plate. Cells were treated with plasmid:CPPcomplexes at different charge ratios in serum-free media for 4 h,after which serum-containing medium was added and incubated foranother 20 h. Thereafter, MTS proliferation assay was used accordingto the manufacturer's protocol. Untreated cells were defined as 100%viable.

Fig. 1. Complex formation, effect of stearylation and the impact of an endosomolyticagent on transfection of plasmids with CPPs. (A) Ability of CPPs to form complexes witha pGL3 plasmid at different plasmid:peptide charge ratios (1:0.5–1:5), was analyzedusing an ethidium bromide (EtBr) exclusion assay. (B) For comparison of transfectionefficiencies of stearyl-(RxR)4 and (RxR)4 peptide, 4×104 CHO cells/well were seeded24 h prior experiment in 24-well plates. Cells were treated with complexes at 4different charge ratios (1:1, 1:2, 1:3 and 1:5), using 0.5 μg of plasmid per well, for 4 h inserum-free media followed by replacement to 10% serum-containing medium andincubated additionally for 20 h. Cells were washed with HKR buffer and lysed in 0.2%Triton X-100, luciferase activity was measured and normalized against the proteincontent in each well. (C) Experiments were carried in the same manner at charge ratio1:5, but with addition of 100 μM chloroquine (CQ) to the complexes before celltreatments. The values represent the mean of at least three independent experimentsperformed in duplicate (mean±SEM). (B) ***pb0.001, ANOVA Dunnett's multiplecomparison test.
45T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
2.9. Dynamic light scattering (DLS) studies
Hydrodynamic mean diameter of the DNA nanoparticles was deter-mined by dynamic light scattering studies using a Zetasizer Nano ZSapparatus (Malvern Instruments, United Kingdom). Plasmid DNA(pDNA) complexes resulting from the addition of stearyl-(RXR)4 wereformulated according to the protocol for in vitro transfection and as-sessed indisposable lowvolume cuvettes. Briefly, pDNAwas formulatedin deionizedwater, in 20 µl volume, at afinal concentration of 0.01 µg/µlof pDNA. After 30 min incubation at room temperature, the DNA com-plexeswere diluted in Opti-MEM® into a final volume of 200 µl. All datawas converted to “relative intensity” plots fromwhere themean hydro-dynamic diameter was derived.
2.10. Statistics
Values in all experiments are represented asmean±SEMof at least 3independent experiments done in duplicate. CPP- and Lipofectaminemediated delivery efficiency of plasmid or SCO was consideredsignificant at ***pb0.001 using ANOVA Dunnett's multiple comparisontest or ANOVA Bonferroni's multiple comparison test. In toxicitymeasurements, a decrease in viability was considered significant at***pb0.001 using ANOVA Dunnett's multiple comparison test.
3. Results
3.1. Complex formation efficacy of CPPs
The (RxR)4 peptide has been successfully used in recent years forthe delivery of covalently linked SCOs [10,11,32]. To our knowledge,there are no reports showing that the (RxR)4 peptide would be able tofacilitate delivery of plasmids. Therefore, for the formation of plasmid:CPP complexes a simple non-covalent co-incubation strategy was used.When utilizing that approach, positively charged CPPs and negativelycharged cargo are simply mixed together to form steady complexes[33,34].
The non-covalent complexes formed between CPPs and plasmidDNA were characterized by an EtBr exclusion assay, essentially as de-scribed previously [20,35]. A clear increase in quenching was observedwith all CPPs (Fig. 1A), thus indicating the inability of EtBr to bind toDNA and indirectly showing the ability of the peptides to interact withplasmid DNA. At a charge ratio (CR) of 1:5, all used CPPs quenched EtBralmost completely. Stearyl-(RxR)4 had a more pronounced condensingeffect in this assay at higher CRs while (RxR)4 was more efficient atlower CRs. These results were confirmed by a gel retardation assay,where similar effects were seen in the retardation of complexmigration(data not shown).
Particle size of plasmid:stearyl-(RxR)4 complexes was characterizedusing the dynamic light scattering (DLS) analysis. At a CR of 1:3,diameter of the complexes was around 450 nm. Plasmid DNAwas evenfurther condensed at CR of 1:5, resulting in the formation of nanopart-icles with a size of around 370 nm (Table 2).
Stability of plasmid:stearyl-(RxR)4 complexes towards the degrad-ing capacity of serum was evaluated by co-incubating the complexeswith various concentrations of serum and analyzed on agarose gel. AtCRs of 1:3 and 1:5, the peptidewas able to shield the plasmid from deg-radation at least for 24 h, whereas naked plasmid underwent completedegradation after 24 h (Supplementary Fig. 1).
3.2. Impact of N-terminal stearylation on cell transfections
After the characterization of plasmid:stearyl-(RxR)4 complexes, wesought to investigate if they were also able to promote the internali-zation of plasmid DNA expressing luciferase. The delivery of plasmidswith CPPs, for instance with Tat, has been described previously butthe efficacy of plasmid transfection was poor [25]. In line with these
observations, luciferase expression in CHO cells transfected withplasmid:(RxR)4 complexes at all CRs was negligible (Fig. 1B). Derivingfrom the success of stearylation of TP10 for SCO delivery, we sought toadd the hydrophobic stearic acidmoiety to theN-terminus of the (RxR)4

Fig. 2. Delivery of luciferase expressing plasmid using stearyl-(RxR)4, (RxR)4 or stearyl-Arg9 peptide in different cell lines. 3.5×104 BHK21 cells (A), 4×104 HEK293 (B) and4×104 CHO (C) cells/well were seeded 24 h prior experiment into 24-well plates. Cellswere treated and analyzed as in Fig. 1B.
Table 2Particle size of plasmid:stearyl-(RxR)4 complexes, determined by dynamic lightscattering (DLS) measurements.
Charge ratio (plasmid:stearyl-(RXR)4) Average diameter (nm) ± SD
1:3 450.8±84.01:5 372.9±68.3
46 T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
peptide. Stearylation had a substantial effect on luciferase activity at allCRs (Fig. 1B). Significantly high transfection levels were seen at CR 1:3and the effect was even more pronounced at CR 1:5, generating up to170-fold increase in luciferase expression compared to untreated cells(Fig. 1B).
3.3. Endocytic uptake of plasmid:CPP complexes
Chloroquine is an agent known to facilitate destabilization ofendosomes and subsequent release of entrapped biomolecules. Toconfirm that plasmid:CPP complexes were taken up by an endocyticpathway, plasmid transfections were carried out in CHO cells in thepresence of chloroquine at a final concentration of 100 μM (Fig. 1C).In case of both stearyl-(RxR)4 and stearyl-Arg9, a significant increasein luciferase expression levels was observed when simultaneouslytreated with chloroquine. Unmodified (RxR)4 efficiently condensedDNA, but there was only hardly detectable effect on plasmid trans-fections, even in the presence of chloroquine, suggesting that onlysmall amounts of (RxR)4 complexes were taken up by cells. The effectof chloroquine was more pronounced with stearyl-Arg9, showinghigher increase and absolute values of luciferase expression thanstearyl-(RxR)4. Such substantial increase with chloroquine treat-ment shows that despite promoting endosomal escape much of theentrapped material still remains in endosomal compartments, whichis in parity with our earlier results seen with stearyl-TP10 on SCOdelivery [23]. These results also demonstrate that stearyl-(RxR)4is far more potent than the previously described stearyl-Arg9 pep-tide (in the absence of chloroquine), emphasizing that even smallstructural differences (i.e insertion of an 6-aminohexanoic acidspacer between the Arg residues) can have a significant effect on thedelivery efficacy.
To confirm that stearyl-(RxR)4 is capable of promoting plasmiddelivery in additional cell lines, similar experiments were carried outin BHK21 and HEK293 cells. Transfection efficiencies with stearyl-(RxR)4 in BHK21 (Fig. 2A) andHEK293 (Fig. 2B) cells were even higherthan in CHO cells (Fig. 2C). The same tendency was seen when cellswere transfected with a commercially available transfection agent —Lipofectamine™ 2000 (data not shown). At themost efficient CR of 1:5of all tested peptides, stearyl-(RxR)4 was significantly more effectivethan (RxR)4 or stearyl-Arg9.
Lipofection using cationic lipids is the most common and efficientstrategy to introduce ONs, including plasmid DNA, into the cells. Lipo-fectamine™ 2000, the most widely used transfection agent, was usedas an internal positive control to compare the delivery efficacy withCPPs. Lipofectamine™ 2000was superior to stearyl-(RxR)4 in terms oftransfection efficiency, but at the same time substantial differencesbetween toxicity profiles of stearyl-(RxR)4 and Lipofectamine™ 2000were observed (Fig. 3). Both long-term (MTS assay) and short-term(LDH leakage) toxicities were assessed. Although there were no toxiceffects with stearyl-(RxR)4 or Lipofectamine™ 2000 in the membraneintegrity assay (data not shown), Lipofectamine™ 2000 had a signif-icantly higher toxic effect on cell viability in the cell proliferation assay(Fig. 3B). Viability of the cells was reduced more than 30% as com-pared to untreated cells. At the same time no toxicity was observedwith any of the CPP:plasmid complexes at any of the tested CRs.Similar toxicity profiles were also observed in the protein determi-nation assay when normalizing the transfection efficiencies to proteinamount in each well (data not shown).
To address the fraction of cells transfected with plasmid:stearyl-(RxR)4 complexes, we transfected CHO cells with EGFP expressingplasmid and utilized confocal microscopy for the analysis. Plasmid:stearyl-(RxR)4 complexes transfected uniformly most of the cellpopulation (Fig. 3C2), in line with the result we obtained with SCOs.However, Lipofectamine™ 2000:plasmid complexes did not seem toreach thewhole cell population (which is also in parity with what weobserved in the SCO transfection experiments), although lipofectionenabled higher per-cell transfection efficiency (Fig. 3C3).
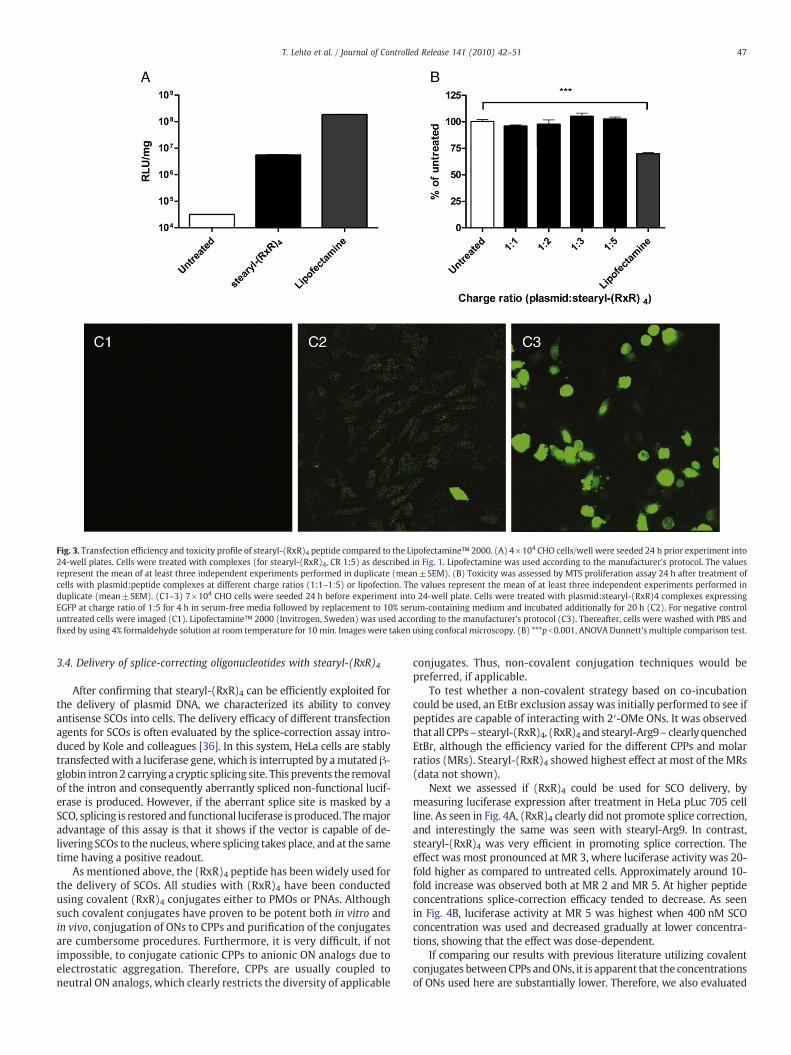
Fig. 3. Transfection efficiency and toxicity profile of stearyl-(RxR)4 peptide compared to the Lipofectamine™ 2000. (A) 4×104 CHO cells/well were seeded 24 h prior experiment into24-well plates. Cells were treated with complexes (for stearyl-(RxR)4, CR 1:5) as described in Fig. 1. Lipofectamine was used according to the manufacturer's protocol. The valuesrepresent the mean of at least three independent experiments performed in duplicate (mean±SEM). (B) Toxicity was assessed by MTS proliferation assay 24 h after treatment ofcells with plasmid:peptide complexes at different charge ratios (1:1–1:5) or lipofection. The values represent the mean of at least three independent experiments performed induplicate (mean±SEM). (C1–3) 7×104 CHO cells were seeded 24 h before experiment into 24-well plate. Cells were treated with plasmid:stearyl-(RxR)4 complexes expressingEGFP at charge ratio of 1:5 for 4 h in serum-free media followed by replacement to 10% serum-containing medium and incubated additionally for 20 h (C2). For negative controluntreated cells were imaged (C1). Lipofectamine™ 2000 (Invitrogen, Sweden) was used according to the manufacturer's protocol (C3). Thereafter, cells were washed with PBS andfixed by using 4% formaldehyde solution at room temperature for 10 min. Images were taken using confocal microscopy. (B) ***pb0.001, ANOVA Dunnett's multiple comparison test.
47T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
3.4. Delivery of splice-correcting oligonucleotides with stearyl-(RxR)4
After confirming that stearyl-(RxR)4 can be efficiently exploited forthe delivery of plasmid DNA, we characterized its ability to conveyantisense SCOs into cells. The delivery efficacy of different transfectionagents for SCOs is often evaluated by the splice-correction assay intro-duced by Kole and colleagues [36]. In this system, HeLa cells are stablytransfectedwith a luciferase gene, which is interrupted by amutated β-globin intron 2 carrying a cryptic splicing site. This prevents the removalof the intron and consequently aberrantly spliced non-functional lucif-erase is produced. However, if the aberrant splice site is masked by aSCO, splicing is restored and functional luciferase is produced. Themajoradvantage of this assay is that it shows if the vector is capable of de-livering SCOs to the nucleus, where splicing takes place, and at the sametime having a positive readout.
As mentioned above, the (RxR)4 peptide has been widely used forthe delivery of SCOs. All studies with (RxR)4 have been conductedusing covalent (RxR)4 conjugates either to PMOs or PNAs. Althoughsuch covalent conjugates have proven to be potent both in vitro andin vivo, conjugation of ONs to CPPs and purification of the conjugatesare cumbersome procedures. Furthermore, it is very difficult, if notimpossible, to conjugate cationic CPPs to anionic ON analogs due toelectrostatic aggregation. Therefore, CPPs are usually coupled toneutral ON analogs, which clearly restricts the diversity of applicable
conjugates. Thus, non-covalent conjugation techniques would bepreferred, if applicable.
To test whether a non-covalent strategy based on co-incubationcould be used, an EtBr exclusion assay was initially performed to see ifpeptides are capable of interacting with 2′-OMe ONs. It was observedthat all CPPs– stearyl-(RxR)4, (RxR)4 and stearyl-Arg9– clearly quenchedEtBr, although the efficiency varied for the different CPPs and molarratios (MRs). Stearyl-(RxR)4 showed highest effect at most of the MRs(data not shown).
Next we assessed if (RxR)4 could be used for SCO delivery, bymeasuring luciferase expression after treatment in HeLa pLuc 705 cellline. As seen in Fig. 4A, (RxR)4 clearly did not promote splice correction,and interestingly the same was seen with stearyl-Arg9. In contrast,stearyl-(RxR)4 was very efficient in promoting splice correction. Theeffect was most pronounced at MR 3, where luciferase activity was 20-fold higher as compared to untreated cells. Approximately around 10-fold increase was observed both at MR 2 and MR 5. At higher peptideconcentrations splice-correction efficacy tended to decrease. As seenin Fig. 4B, luciferase activity at MR 5 was highest when 400 nM SCOconcentration was used and decreased gradually at lower concentra-tions, showing that the effect was dose-dependent.
If comparing our results with previous literature utilizing covalentconjugates betweenCPPs andONs, it is apparent that the concentrationsof ONs used here are substantially lower. Therefore, we also evaluated

48 T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
the relative efficacy of the stearyl-(RxR)4:2′-OMe ON non-covalentcomplexes compared to the previously described (RxR)4-PMO conju-gate. In accordancewith previous studies, a dose-dependent increase insplice correctionwasobserved from1 μMto4 μMof (RxR)4-PMO,whichis in parity with the effects seen with non-covalent stearyl-(RxR)4:2′-OMe SCO complexes at 10-times lower SCO concentration (Fig. 4B).
To assess the delivery efficacy in terms of fraction of cells transfectedwith stearyl-(RxR)4:2′-OMe SCO complexes,we utilized Cy5-labeled 2′-OMeSCOs anddetermined theuptakebyFACS analysis. Both stearylated
and unmodified peptide transfected almost 100% of the cell population,whereas (RxR)4-PMO conjugate and also Lipofectamine:SCO complexesfailed to do so (Fig. 4C). These results are in keeping with the obser-vation made with plasmid:stearyl-(RxR)4 complexes, where also mostof the cells were uniformly transfected with plasmid.
FACS analysis also enabled us to compare the uptake of unmodifiedand stearylated (RxR)4 peptide and Cy5-labeled SCO complexes. Inter-estingly, complexes with both peptides were taken up almost equally(Fig. 4D), with stearyl-(RxR)4 being a fraction better at lower

49T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
concentrations, therefore, we assume that these differences cannotaccount for large differences in biological activity and that the stearicacid modification has to be responsible for this effect.
To have insight to the potential of utilizing stearyl-(RxR)4 as andelivery vector in vivo, we conducted transfections in the presence ofserum (Fig. 4F). Unfortunately, considerable decrease in transfectionefficiency was observed, however splice-correction efficiency was notcompletely abrogated and 4-fold increase in splice correction wasdetected (Fig. 4F). Similar tendency was shown with (RxR)4-PMOconjugates,whichwas substantially less active in the presence of serum.Nevertheless, sensitivity to serum does not prevent (RxR)4-PMO conju-gates to be very potent in vivo.
We have previously reported that stearyl-TP10 vector is an efficientvector for the delivery of SCOs [23]. Stearylated TP10 is slightly morepotent for the delivery of 2′-OMe SCOs than stearyl-(RxR)4, the differ-ence in efficiency being roughly 2 times. However, stearyl-(RxR)4exerted the same efficiency of splice correction than (RxR)4-PMO, but atup to 20-fold lower concentrations, 200nM and 4 μM respectively.Nevertheless, these (RxR)4 peptides did not reach the transfectionefficiency levels of Lipofectamine™ 2000 (Fig. 4E).
4. Discussion
Nucleic acids used in genemodulation share the common feature ofinadequate bioavailability due to their physicochemical properties.Finding suitable delivery vehicles for these molecules is a prerequisitefor successful gene therapy in the future. Viral vectors are highly eff-icient, but several concerns related to immunogenicity and limitedpack-aging capacity hamper their use. Failings of viral delivery have openedavenue for non-viral delivery vectors. CPPs are one such group of non-viral delivery vectors that has been shown to mediate delivery of avariety of molecules used in gene modulation settings, both in vivo andin vitro [4,6]. Different ONs and their analogs are the foundation of mostof the compounds used in gene modulating settings, such as splice-correcting ONs, antimiRs, siRNAs, plasmids etc. In most cases ONs areattached to CPPs by a covalent bond, for instance via a disulfide bridge,but as aforementioned this strategy makes the production and puri-fication of these compounds complicated and time consuming. Al-though this conjugation approach will remain the main strategy foruncharged ONs analogues, like PNAs and PMOs, the possibility to com-plex negatively charged ONs by co-incubation strategy with CPPs hasgained wide interest recently.
The main drawback of CPPs is the intracellular fate of both CPP andrespective cargo, mainly because CPPs are prone to utilize endosomalpathways for internalization into cells with a concurrent trapping ofCPP-bound substances into endosomal compartments. Many conceptshave been proposed to overcome this limitation. Both design of newCPPs and modification of known CPPs to increase their endosomolyticproperties have been introduced and several applications have beensuccessfully implemented.Modifying CPPswith hydrophobicmoietieshas been shown to increase their cellular uptake [28,37–39]. More-over, stearylation has been shown to be successful in promoting ON
Fig. 4. Stearyl-(RxR)4 mediated non-covalent SCO delivery, dose-dependent effect on splicepLuc 705 cells were seeded 24 h prior experiment in 24-well plates. Cells were treated with dSCO. (B) Dose-dependent effect was evaluated at molar ratio 5, and the concentration of SCO10% serum-containing medium and incubated additionally 20 h. In case of (RxR)4-PMOreplacement to 10% serum-containing medium and incubated additionally 20 h. Cells were wand normalized against the protein content. Values obtained in RLU/mg were then convertemean of at least three independent experiments performed in duplicate (mean±SEM). (C) HCells were treated with peptide and Cy5-labeled 2′-OMe ON complexes at molar ratio 1:6 antrypsinization and suspended in ice-cold PBS containing 5% FBS and centrifuged. Supernatan0.5% FBS, after what flow cytometry analysis was carried out. The percentage of transfectedthreshold level. Aforementioned analysis was carried out using FacsDiva software. (D) Cecharacterized by calculating mean Cy5 fluorescence value for transfected cells and comparinFig. 4B. Stearyl-(RxR)4:SCO complexes were used at 200nM SCO (at molar ratio 3), wherea(at molar ratio 10). Lipofectamine™ 2000 (Invitrogen, Sweden) was used according to the mdescribed on (A) and (B), however, transfection was carried out in serum-containing media. Cwas used at 1 µM.
delivery [21,23,28]. The mechanism by which stearylation increasescytoplasmic and nuclear delivery is not yet clear. However, this modi-fication clearly renders the peptide more hydrophobic and enablesenhanced condensation of ON, presumably facilitating more stablenanoparticle formation. Probably it also facilitates the peptide inter-play with endosomal membranes, thus enabling some of the CPP andrespective cargo to escape from endosomal compartments.
As mentioned before, plasmid delivery with CPPs not carrying anymodifications seems to be rather ineffective [25]. For this reason, syner-gistic effect of known transfection agents has been utilized to achievereasonable transfection efficiency [40–42] or rather more complicatedsystems have been developed to facilitate plasmid transfections, likeMEND [43], and tetraarginine-PEG lipid-coated protamine/DNA [44].
We show here that an addition of a simple hydrophobic modifi-cation, stearic acid, to (RxR)4 peptide has a significant effect on the(RxR)4 peptide's ability to transfect plasmids into different cell lines.Whereas unmodified (RxR)4 had negligible effects on the transfectionsof plasmids, stearyl-(RxR)4 increased transfection by several logs insome cell lines. However, the levels of transfections were still morethan a log lower thanwith Lipofectamine™ 2000, showing that stearyl-(RxR)4 is a less potent vector for plasmid delivery than Lipofectamine™2000. However, it is known that Lipofectamine™ 2000 efficiency islargely associated with toxicity. Comparison of the toxicity profiles ofboth stearyl-(RxR)4 and Lipofectamine™ 2000 (complexed with plas-mid) showed that Lipofectamine™ 2000 decreased cell viability by atleast 30% in 24 h, while stearyl-(RxR)4 did not display any toxicity atused concentrations. Furthermore, stearyl-(RxR)4 enabled most of thecells in the population to be uniformly transfected, whereas it isquestionable in case of Lipofectamine™ 2000.
Our results demonstrate that the usefulness of stearylation as amethod to increase plasmid transfections differs between CPPs, as itclearly enhancesdeliverywith (RxR)4 buthas a significantly lower effecton Arg9. Interestingly, upon co-treatment with chloroquine, higher lu-ciferase expression levels were achieved with stearyl-Arg9 than withstearyl-(RxR)4, indicating that the stearic acid modification on (RxR)4enables more efficient endosomal escape compared to Arg9, which ismainly entrapped in the endosomes. The differences in delivery efficacypoint out that small structural differences, in this case aminohexanoicacid linker, can have significant effects. Despite the growing use ofstearylated oligoarginines for plasmid delivery, our comparative resultsclearly illustrate that (RxR)4 is superior to Arg9.
DLS measurements of plasmid:stearyl-(RxR)4 complexes clearlysupported the assumption of nanoparticle formation upon ethidiumbromide exclusion assay, as the uniform nanoparticles with a diameterof around 400 nmwere registered. In complex stability assay, formationof these nanoparticles enabled the shielding of the plasmid from theserum degradation over 24 h.
As mentioned before, the (RxR)4 peptide has been successfully ex-ploited when conjugated to SCOs, both in vitro and in vivo [8,12,13,45].However, when utilizing a non-covalent co-incubation protocol, thepeptide was unable to mediate any splice correction in the HeLa pLuc705 cell assay. We have recently shown, that N-terminally stearylated
correction and comparison with the (RxR)4-PMO covalent conjugate. (A) 5×104 HeLaifferent peptides: 2′-OMe ON complexes at different molar ratios (2–20), using 200nMwas varied from 100 to 400nM for 4 h in serum-free media followed by replacement tocovalent conjugates, treatment was carried out for 4 h in Opti-MEM®, followed byashed with HKR buffer and lysed in 0.2% Triton X-100, luciferase activity was measuredd to fold-increase of treatments compared to untreated cells. The values represent theeLa pLuc 705 cells (2×105) were seeded 24 h prior to experiments into 24-well plates.
d incubated for 2 h in serum-free Opti-MEM®media. Thereafter, cells were detached byt was removed and the resulting cell pellet was resuspended in ice-cold PBS containingcells was determined by counting the cells displaying Cy5 fluorescent signal above a
lls were seeded, treated and analyzed as described in (C). However, the uptake wasg it with the same parameter of untreated cells. (E) Cells were treated as described ins (RxR)4-PMO conjugate was used at 4 µM and stearyl-TP10 was used at 200nM SCOanufacturer's protocol using 200 nM SCO. (F) Cells were seeded, treated and analyzed asoncentration of SCO complexed with stearyl-(RxR)4 was 400nM, whereas (RxR)4-PMO

50 T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
TP10 facilitates efficient splice correction if non-covalently complexedwith the 2′-OMe SCOs [23]. Introduction of the same modification,stearic acid, to the (RxR)4 peptidepromoted efficient splice correction inthe same co-incubation setting. Splice-correction efficacy as high as 20-fold was seen at an SCO concentration of 200nM at MR 3. Remarkably,concentrations needed to obtain efficient splice correctionwith stearyl-(RxR)4:SCO complexes are very low compared to the concentrationsneeded when utilizing the covalent strategy of (RxR)4-PMO, which arerather in μM range [11].
Interestingly, stearylation of Arg9 did not have any effect on thedelivery of 2′-OMe SCO, indicating that the successful implementa-tion of stearylation is dependent on the properties of the used CPP.Moreover it is highly dependent on the cargo, as stearyl-Arg9 wasrelatively effective for plasmid delivery but failed to deliver SCOs.Generally it seems that hydrophobicity plays a key role in the generalsuccess of CPPs to deliver plasmids and SCOs. Introduction of a hydro-phobic moiety to the (RxR)4 significantly increased both plasmid andSCOdelivery. Samemodification only partlyworkedwithArg9, but Arg9is hydrophilic in nature. As we have shown before [23], stearylation oftransportan 10 had an even greater effect on SCO delivery, than stearyl-(RxR)4 used in this study, which is in line with the amphipathic/hydrophobic character of transportans. Also, as it was seen that (RxR)4and stearyl-(RxR)4 complexes with Cy5-labeled 2′-OMe SCOs weretaken up in comparable quantities, it is reasonable to assume that smalldifferences in uptake could not account for the large differences inbiological activity, indicating the central role of the stearic acid moiety.Furthermore, if these complexes are expectedly taken up by endosomalpathway, it is another indication that stearic acid enables endosomalescape, at least in case of (RxR)4 peptide.
As aforementioned, delivery efficacy of stearyl-(RxR)4 was con-siderably decreased in the presence of serum, however splice correc-tion was still detectable. This is in accordance with the resultobtained with (RxR)4-PMO and this reduction in efficiency does notaffect (RxR)4-PMO of being very effective in vivo [12,45].
Considering, that many groups are extensively using (RxR)4 in pre-clinical settings for the delivery of SCOs, this new information might beof great importance. The simplicity of the experimental set-up in com-bination with the properties of stearyl-(RxR)4 makes it a highly in-teresting vector for future in vivo use.
Acknowledgements
The work presented in this article was supported by the SwedishResearch Council (VR-NT); by the Center for Biomembrane Research,Stockholm; by the Knut and Alice Wallenberg's Foundation; by the EUthrough the European Regional Development Fund through the Centerof Excellence in Chemical Biology, Estonia; by the targeted financingSF0180027s08 from the Estonian Government; by the DoRa Program ofThe European Social Fund; and by the Archimedes Foundation.Work inthe group of B. Lebleu was supported by the Association Françaisecontre les Myopathies and the Ligue Régionale Française contre lecancer. Work in the group of CIE Smith was supported by the SwedishResearch Council, KI Faculty funds for funding of postgraduate students,and the European Union FP6 grant EuroPharmacoGene (FP6-2005-037283).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.jconrel.2009.08.028.
References
[1] P. Sazani, R. Kole, Therapeutic potential of antisense oligonucleotides as modu-lators of alternative splicing, J. Clin. Invest. 112 (4) (2003) 481–486.
[2] A.D. Miller, Retrovirus packaging cells, Hum. Gene Ther. 1 (1) (1990) 5–14.
[3] D. Derossi, A.H. Joliot, G. Chassaing, A. Prochiantz, The third helix of the Anten-napedia homeodomain translocates through biological membranes, J. Biol. Chem.269 (14) (1994) 10444–10450.
[4] M. Mäe, Ü. Langel, Cell-penetrating peptides as vectors for peptide, protein andoligonucleotide delivery, Curr. Opin. Pharmacol. 6 (5) (2006) 509–514.
[5] K.M. Stewart, K.L. Horton, S.O. Kelley, Cell-penetrating peptides as delivery vehiclesfor biology and medicine, Org. Biomol. Chem. 6 (13) (2008) 2242–2255.
[6] E. Vives, J. Schmidt, A. Pelegrin, Cell-penetrating and cell-targeting peptides indrug delivery, Biochim. Biophys. Acta 1786 (2) (2008) 126–138.
[7] I. Nakase, T. Takeuchi, G. Tanaka, S. Futaki, Methodological and cellular aspects thatgovern the internalization mechanisms of arginine-rich cell-penetrating peptides,Adv. Drug Deliv. Rev. 60 (4–5) (2008) 598–607.
[8] B. Lebleu, H.M. Moulton, R. Abes, G.D. Ivanova, S. Abes, D.A. Stein, P.L. Iversen, A.A.Arzumanov, M.J. Gait, Cell penetrating peptide conjugates of steric block oligonu-cleotides, Adv. Drug Deliv. Rev. 60 (4–5) (2008) 517–529.
[9] J.B. Rothbard, E. Kreider, C.L. VanDeusen, L.Wright, B.L.Wylie, P.A.Wender, Arginine-rich molecular transporters for drug delivery: role of backbone spacing in cellularuptake, J. Med. Chem. 45 (17) (2002) 3612–3618.
[10] S. Abes, H.M. Moulton, P. Clair, P. Prevot, D.S. Youngblood, R.P. Wu, P.L. Iversen, B.Lebleu, Vectorization of morpholino oligomers by the (R-Ahx-R)4 peptide allowsefficient splicing correction in the absence of endosomolytic agents, J. ControlRelease 116 (3) (2006) 304–313.
[11] R. Abes, H.M. Moulton, P. Clair, S.T. Yang, S. Abes, K. Melikov, P. Prevot, D.S.Youngblood, P.L. Iversen, L.V. Chernomordik, B. Lebleu, Delivery of steric blockmorpholino oligomers by (R-X-R)4 peptides: structure–activity studies, NucleicAcids Res. 36 (20) (2008) 6343–6354.
[12] S. Fletcher, K. Honeyman, A.M. Fall, P.L. Harding, R.D. Johnsen, J.P. Steinhaus, H.M.Moulton, P.L. Iversen, S.D. Wilton, Morpholino oligomer-mediated exon skippingaverts the onset of dystrophic pathology in themdxmouse,Mol. Ther. 15 (9) (2007)1587–1592.
[13] G.D. Ivanova, A. Arzumanov, R. Abes, H. Yin,M.J.Wood, B. Lebleu,M.J. Gait, Improvedcell-penetrating peptide–PNA conjugates for splicing redirection in HeLa cells andexon skipping inmdxmousemuscle, Nucleic Acids Res. 36 (20) (2008) 6418–6428.
[14] F. Duchardt, M. Fotin-Mleczek, H. Schwarz, R. Fischer, R. Brock, A comprehensivemodel for the cellular uptake of cationic cell-penetrating peptides, Traffic 8 (7)(2007) 848–866.
[15] S. Futaki, I. Nakase, A. Tadokoro, T. Takeuchi, A.T. Jones, Arginine-rich peptides andtheir internalization mechanisms, Biochem. Soc. Trans. 35 (Pt 4) (2007) 784–787.
[16] P. Lundin, H. Johansson, P. Guterstam, T.Holm,M.Hansen, Ü. Langel, S. El Andaloussi,Distinct uptake routes of cell-penetrating peptide conjugates, Bioconjug. Chem. 19(12) (2008) 2535–2542.
[17] S. El-Andaloussi, H. Johansson, P. Lundberg, U. Langel, Induction of splice correctionby cell-penetrating peptide nucleic acids, J. Gene Med. 8 (10) (2006) 1262–1273.
[18] A. El-Sayed, S. Futaki, H. Harashima, Delivery of macromolecules using arginine-richcell-penetrating peptides: ways to overcome endosomal entrapment, AAPS J. 11 (1)(2009) 13–22.
[19] J.S. Wadia, R.V. Stan, S.F. Dowdy, Transducible TAT-HA fusogenic peptide enhancesescape of TAT-fusion proteins after lipid raft macropinocytosis, Nat. Med. 10 (3)(2004) 310–315.
[20] P. Lundberg, S. El-Andaloussi, T. Sütlü, H. Johansson, Ü. Langel, Delivery of shortinterfering RNA using endosomolytic cell-penetrating peptides, FASEB J. 21 (11)(2007) 2664–2671.
[21] S. Futaki, W. Ohashi, T. Suzuki, M. Niwa, S. Tanaka, K. Ueda, H. Harashima, Y.Sugiura, Stearylated arginine-rich peptides: a new class of transfection systems,Bioconjug. Chem. 12 (6) (2001) 1005–1011.
[22] Y. Nakamura, K. Kogure, S. Futaki, H. Harashima, Octaarginine-modified multifunc-tional envelope-type nano device for siRNA, J. Control Release 119 (3) (2007)360–367.
[23] M. Mäe, S. El Andaloussi, P. Lundin, N. Oskolkov, H.J. Johansson, P. Guterstam, Ü.Langel, A stearylated CPP for delivery of splice correcting oligonucleotides usinga non-covalent co-incubation strategy, J. Control Release 134 (3) (2009)221–227.
[24] H. Hashida, M. Miyamoto, Y. Cho, Y. Hida, K. Kato, T. Kurokawa, S. Okushiba, S.Kondo, H. Dosaka-Akita, H. Katoh, Fusion of HIV-1 Tat protein transductiondomain to poly-lysine as a new DNA delivery tool, Br. J. Cancer 90 (6) (2004)1252–1258.
[25] I.A. Ignatovich, E.B. Dizhe, A.V. Pavlotskaya, B.N. Akifiev, S.V. Burov, S.V. Orlov, A.P.Perevozchikov, Complexes of plasmid DNA with basic domain 47–57 of the HIV-1Tat protein are transferred to mammalian cells by endocytosis-mediated path-ways, J. Biol. Chem. 278 (43) (2003) 42625–42636.
[26] Z. Liu, M. Li, D. Cui, J. Fei, Macro-branched cell-penetrating peptide design for genedelivery, J. Control Release 102 (3) (2005) 699–710.
[27] S.L. Lo, S. Wang, An endosomolytic Tat peptide produced by incorporation ofhistidine and cysteine residues as a nonviral vector for DNA transfection, Bio-materials 29 (15) (2008) 2408–2414.
[28] I.A. Khalil, S. Futaki, M. Niwa, Y. Baba, N. Kaji, H. Kamiya, H. Harashima, Mech-anism of improved gene transfer by the N-terminal stearylation of octaarginine:enhanced cellular association by hydrophobic core formation, Gene Ther. 11 (7)(2004) 636–644.
[29] R. Moriguchi, K. Kogure, H. Akita, S. Futaki, M. Miyagishi, K. Taira, H. Harashima,A multifunctional envelope-type nano device for novel gene delivery of siRNAplasmids, Int. J. Pharm. 301 (1–2) (2005) 277–285.
[30] I.A. Khalil, K. Kogure, S. Futaki, S. Hama, H. Akita,M. Ueno, H. Kishida,M. Kudoh, Y.Mishina, K. Kataoka, M. Yamada, H. Harashima, Octaarginine-modified multi-functional envelope-type nanoparticles for gene delivery, Gene Ther. 14 (8)(2007) 682–689.

51T. Lehto et al. / Journal of Controlled Release 141 (2010) 42–51
[31] P. Guterstam, M. Lindgren, H. Johansson, U. Tedebark, J. Wengel, S. El Andaloussi,Ü. Langel, Splice-switching efficiency and specificity for oligonucleotides withlocked nucleic acid monomers, Biochem. J. 412 (2) (2008) 307–313.
[32] H. Yin, H.M. Moulton, Y. Seow, C. Boyd, J. Boutilier, P. Iverson, M.J. Wood, Cell-penetrating peptide-conjugatedantisense oligonucleotides restore systemicmuscleand cardiac dystrophin expression and function, Hum. Mol. Genet. 17 (24) (2008)3909–3918.
[33] S. Deshayes,M.Morris, F. Heitz, G. Divita, Delivery of proteins and nucleic acids usinga non-covalent peptide-based strategy, Adv. Drug Deliv. Rev. 60 (4–5) (2008)537–547.
[34] B.R. Meade, S.F. Dowdy, Enhancing the cellular uptake of siRNA duplexes followingnoncovalent packaging with protein transduction domain peptides, Adv. DrugDeliv. Rev. 60 (4–5) (2008) 530–536.
[35] H. Petersen, K. Kunath, A.L. Martin, S. Stolnik, C.J. Roberts, M.C. Davies, T. Kissel,Star-shaped poly(ethylene glycol)-block-polyethylenimine copolymers enhanceDNA condensation of low molecular weight polyethylenimines, Biomacromole-cules 3 (5) (2002) 926–936.
[36] S.H. Kang, M.J. Cho, R. Kole, Up-regulation of luciferase gene expression with anti-sense oligonucleotides: implications and applications in functional assay develop-ment, Biochemistry 37 (18) (1998) 6235–6239.
[37] A.R. Nelson, L. Borland, N.L. Allbritton, C.E. Sims, Myristoyl-based transport of pep-tides into living cells, Biochemistry 46 (51) (2007) 14771–14781.
[38] J. Fernandez-Carneado, M.J. Kogan, N. VanMau, S. Pujals, C. Lopez-Iglesias, F. Heitz,E. Giralt, Fatty acyl moieties: improving Pro-rich peptide uptake inside HeLa cells,J. Pept. Res. 65 (6) (2005) 580–590.
[39] X.D. Guo, F. Tandiono, N. Wiradharma, D. Khor, C.G. Tan, M. Khan, Y. Qian, Y.Y.Yang, Cationic micelles self-assembled from cholesterol-conjugated oligopeptidesas an efficient gene delivery vector, Biomaterials 29 (36) (2008) 4838–4846.
[40] K. Kilk, S. El-Andaloussi, P. Järver, A. Meikas, A. Valkna, T. Bartfai, P. Kogerman, M.Metsis, Ü. Langel, Evaluation of transportan 10 in PEI mediated plasmid deliveryassay, J. Control Release 103 (2) (2005) 511–523.
[41] C. Rudolph, C. Plank, J. Lausier, U. Schillinger, R.H. Muller, J. Rosenecker, Oligomersof the arginine-rich motif of the HIV-1 TAT protein are capable of transferringplasmid DNA into cells, J. Biol. Chem. 278 (13) (2003) 11411–11418.
[42] J. Gadi, K. Ruthala, K.A. Kong, H.W. Park, M.H. Kim, The third helix of the Hoxc8homeodomain peptide enhances the efficiency of gene transfer in combinationwith Lipofectamine, Mol. Biotechnol. 42 (1) (2008) 41–48.
[43] K. Kogure, H. Akita, Y. Yamada, H. Harashima, Multifunctional envelope-type nanodevice (MEND) as a non-viral gene delivery system, Adv. Drug Deliv. Rev. 60 (4–5)(2008) 559–571.
[44] T. Fujita, M. Furuhata, Y. Hattori, H. Kawakami, K. Toma, Y. Maitani, High genedelivery in tumor by intratumoral injection of tetraarginine-PEG lipid-coatedprotamine/DNA, J. Control Release 129 (2) (2008) 124–127.
[45] H.M.Moulton, S. Fletcher, B.W.Neuman, G.McClorey, D.A. Stein, S. Abes, S.D.Wilton,M.J. Buchmeier, B. Lebleu, P.L. Iversen, Cell-penetrating peptide-morpholino conju-gates alter pre-mRNA splicing of DMD (Duchenne muscular dystrophy) and inhibitmurine coronavirus replication in vivo, Biochem. Soc. Trans. 35 (Pt 4) (2007) 826–828.




















