
Deletion of flavoredoxin gene in Desulfovibrio gigas reveals its participation in thiosulfate...
15
Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin Manuela Broco a , Cla ´udio M. Soares a , Solange Oliveira a,b , Stephen G. Mayhew c , Claudina Rodrigues-Pousada a, * a Instituto de Tecnologia Quı ´ mica e Biolo ´ gica (ITQB), Universidade Nova de Lisboa, Av. Repu ´ blica (EAN), 2784-505 Oeiras, Portugal b Departamento de Biologia, Universidade de E ´ vora, Apartado 94, 7002-554 E ´ vora, Portugal c School of Biomolecular and Biomedical Science, Centre for Synthesis and Chemical Biology, Conway Institute, University College Dublin, Dublin 4, Ireland Received 3 July 2007; revised 20 July 2007; accepted 1 August 2007 Available online Edited by Miguel De la Rosa This work is dedicated to the memory of Professor Anto ´nio Xavier deceased on May 7th 2006. Abstract Flavoredoxin participates in Desulfovibrio gigas thio- sulfate reduction pathway. Its 3-dimensional model was gener- ated allowing the oxidized riboflavin-5 0 -phosphate (FMN) site to be predicted. Residues likely to be involved in FMN-binding were identified (N29, W35, T56, K92, H131 and F164) and mutated to alanine. Fluorescence titration with apoprotein showed that FMN is strongly bound in the wild-type protein. Comparison of K d values for mutants suggests that interactions with the phosphate group of FMN, contribute more to binding than the interactions with the isoalloxazine ring. The redox potential of bound FMN determined for wild-type and mutants revealed shifts to less negative values. These findings were corre- lated with the protein structure in order to contribute to a better understanding of the structure–function relationships in flavore- doxin. Ó 2007 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. Keywords: Desulfovibrio gigas; Flavoredoxin; FMN-binding site-directed mutagenesis; Molecular modeling 1. Introduction Flavoredoxin (Flr) is a 194 amino acid protein, isolated as a homodimer containing one oxidized riboflavin-5 0 -phosphate (FMN) per 25 kDa monomer [1]. Initially proposed to act as a redox carrier in bisulfite reduction by molecular hydrogen, recent studies have however shown its involvement in thiosul- fate reduction [2]. Proteins from the flavin reductase family catalyse the reduc- tion of free flavins by reduced nicotinamide nucleotides, the majority of which prefer FMN rather than FAD both as cofac- tor in the enzyme and as the reducible substrate. Yet, in spite of their involvement in a variety of biological processes [3], the function of the reduced free flavin produced in the reaction is poorly understood. In most cases, the flavin cofactor remains bound to the protein during enzyme isolation, indicating that the protein–flavin interaction is strong [4]. In other cases, the cofactor is weakly bound since the enzyme is isolated as an apoprotein [5]. Sequence analysis of Flr did not allow to identify motifs likely to form the FMN-binding site [6]. Three-dimensional structures have been determined for several proteins in the flavin reductase family [4,7,8], two of them sharing sequence homology, namely ferric reductase from Archaeoglobus fulgi- dus (PDB 1I0R) and an FMN-binding protein from Methano- bacterium thermoautotrophicum (PDB 1EJE). These structures and comparative modeling allowed us to identify a potential FMN-binding site and to predict amino acids likely to interact with the flavin. Experimental validation of these predictions showed that, by comparison with other flavin reductases, Flr has a high affinity for FMN and they provide evidence that residues N29, T56 and K92, located near the FMN phosphate group, are important for FMN-binding. 2. Materials and methods 2.1. Comparative modeling of Flr A model for the structure of Flr dimer was derived using compara- tive modeling methods as described in Supplementary data. 2.2. Cloning, expression, purification and site-directed mutagenesis The gene encoding Flr (flr) was cloned in the expression vector pET30a (+) (Novagen, Darmstadt, Germany), site-directed mutagene- sis was carried out according to the quikchange Ò site-directed muta- genesis kit instruction manual (Stratagene, Beverly, MA, USA) and molecular mass and protein concentration were estimated as described in Supplementary data. Abbreviations: FAD, oxidized and reduced flavin adenine dinucleotide; FMN, FMNH 2 , oxidized and reduced riboflavin-5 0 -phosphate; Flr, flavoredoxin; apoFlr, apoprotein form of flavoredoxin; flr, gene encoding flavoredoxin; H 6 Flr, 6His-tagged recombinant flavoredoxin; K d , dissociation constant; FMN-bp, FMN-binding protein; K d,ox , K d,red , dissociation constant in the oxidized and reduced forms; E f , E b , redox potential of free and bound FMN * Corresponding author. Fax: +351 214433644. E-mail address: [email protected] (C. Rodrigues-Pousada). He played a crucial role in the field of Bioinorganic Chemistry being the founder of ITQB and contributing to the understanding of the energy transduction of the anaerobe bacteria in particular the one of D. gigas. 0014-5793/$32.00 Ó 2007 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2007.08.009 FEBS Letters xxx (2007) xxx–xxx ARTICLE IN PRESS Please cite this article in press as: Broco, M. et al., Molecular determinants for FMN-binding in Desulfovibrio gigas ..., FEBS Lett. (2007), doi:10.1016/ j.febslet.2007.08.009
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Deletion of flavoredoxin gene in Desulfovibrio gigas reveals its participation in thiosulfate...

FEBS Letters xxx (2007) xxx–xxx
ARTICLE IN PRESS
Molecular determinants for FMN-binding in Desulfovibrio gigas
flavoredoxin
Manuela Brocoa, Claudio M. Soaresa, Solange Oliveiraa,b, Stephen G. Mayhewc,Claudina Rodrigues-Pousadaa,*
a Instituto de Tecnologia Quımica e Biologica (ITQB), Universidade Nova de Lisboa, Av. Republica (EAN), 2784-505 Oeiras, Portugalb Departamento de Biologia, Universidade de Evora, Apartado 94, 7002-554 Evora, Portugal
c School of Biomolecular and Biomedical Science, Centre for Synthesis and Chemical Biology, Conway Institute, University College Dublin,Dublin 4, Ireland
Received 3 July 2007; revised 20 July 2007; accepted 1 August 2007
Available online
Edited by Miguel De la Rosa
This work is dedicated to the memory of Professor Antonio Xavier deceased on May 7th 2006.�
Abstract Flavoredoxin participates in Desulfovibrio gigas thio-sulfate reduction pathway. Its 3-dimensional model was gener-ated allowing the oxidized riboflavin-5 0-phosphate (FMN) siteto be predicted. Residues likely to be involved in FMN-bindingwere identified (N29, W35, T56, K92, H131 and F164) andmutated to alanine. Fluorescence titration with apoproteinshowed that FMN is strongly bound in the wild-type protein.Comparison of Kd values for mutants suggests that interactionswith the phosphate group of FMN, contribute more to bindingthan the interactions with the isoalloxazine ring. The redoxpotential of bound FMN determined for wild-type and mutantsrevealed shifts to less negative values. These findings were corre-lated with the protein structure in order to contribute to a betterunderstanding of the structure–function relationships in flavore-doxin.� 2007 Federation of European Biochemical Societies. Publishedby Elsevier B.V. All rights reserved.
Keywords: Desulfovibrio gigas; Flavoredoxin; FMN-bindingsite-directed mutagenesis; Molecular modeling
1. Introduction
Flavoredoxin (Flr) is a 194 amino acid protein, isolated as a
homodimer containing one oxidized riboflavin-5 0-phosphate
(FMN) per 25 kDa monomer [1]. Initially proposed to act as
Abbreviations: FAD, oxidized and reduced flavin adenine dinucleotide;FMN, FMNH2, oxidized and reduced riboflavin-5 0-phosphate; Flr,flavoredoxin; apoFlr, apoprotein form of flavoredoxin; flr, geneencoding flavoredoxin; H6Flr, 6His-tagged recombinant flavoredoxin;Kd, dissociation constant; FMN-bp, FMN-binding protein; Kd,ox,Kd,red, dissociation constant in the oxidized and reduced forms; Ef, Eb,redox potential of free and bound FMN
*Corresponding author. Fax: +351 214433644.E-mail address: [email protected] (C. Rodrigues-Pousada).
�He played a crucial role in the field of Bioinorganic Chemistry beingthe founder of ITQB and contributing to the understanding of theenergy transduction of the anaerobe bacteria in particular the one ofD. gigas.
0014-5793/$32.00 � 2007 Federation of European Biochemical Societies. Pu
doi:10.1016/j.febslet.2007.08.009
Please cite this article in press as: Broco, M. et al., Molecular determinants foj.febslet.2007.08.009
a redox carrier in bisulfite reduction by molecular hydrogen,
recent studies have however shown its involvement in thiosul-
fate reduction [2].
Proteins from the flavin reductase family catalyse the reduc-
tion of free flavins by reduced nicotinamide nucleotides, the
majority of which prefer FMN rather than FAD both as cofac-
tor in the enzyme and as the reducible substrate. Yet, in spite
of their involvement in a variety of biological processes [3], the
function of the reduced free flavin produced in the reaction is
poorly understood. In most cases, the flavin cofactor remains
bound to the protein during enzyme isolation, indicating that
the protein–flavin interaction is strong [4]. In other cases, the
cofactor is weakly bound since the enzyme is isolated as an
apoprotein [5].
Sequence analysis of Flr did not allow to identify motifs
likely to form the FMN-binding site [6]. Three-dimensional
structures have been determined for several proteins in the
flavin reductase family [4,7,8], two of them sharing sequence
homology, namely ferric reductase from Archaeoglobus fulgi-
dus (PDB 1I0R) and an FMN-binding protein from Methano-
bacterium thermoautotrophicum (PDB 1EJE). These structures
and comparative modeling allowed us to identify a potential
FMN-binding site and to predict amino acids likely to interact
with the flavin. Experimental validation of these predictions
showed that, by comparison with other flavin reductases, Flr
has a high affinity for FMN and they provide evidence that
residues N29, T56 and K92, located near the FMN phosphate
group, are important for FMN-binding.
2. Materials and methods
2.1. Comparative modeling of FlrA model for the structure of Flr dimer was derived using compara-
tive modeling methods as described in Supplementary data.
2.2. Cloning, expression, purification and site-directed mutagenesisThe gene encoding Flr (flr) was cloned in the expression vector
pET30a (+) (Novagen, Darmstadt, Germany), site-directed mutagene-sis was carried out according to the quikchange� site-directed muta-genesis kit instruction manual (Stratagene, Beverly, MA, USA) andmolecular mass and protein concentration were estimated as describedin Supplementary data.
blished by Elsevier B.V. All rights reserved.
r FMN-binding in Desulfovibrio gigas ..., FEBS Lett. (2007), doi:10.1016/

2 M. Broco et al. / FEBS Letters xxx (2007) xxx–xxx
ARTICLE IN PRESS
2.3. Cofactor identification and determination of the FMN:protein ratioThe flavin cofactor in the recombinant protein was extracted by boil-
ing the protein for 10 min. The nature of the cofactor was determinedby HPLC analysis (Nova Pak Waters Column, C18 60 A,3.9 · 150 mm), comparing its retention time with that of authenticFMN (Sigma, St Louis. MO, USA). The molar ratio of FMN to pro-tein was determined as previously described [9].
2.4. UV–visible absorption spectraUV–visible absorption spectra of holo- and apoproteins were re-
corded with Shimadzu Scientific Instruments UV-3100 or Cary 50spectrophotometers at room temperature in 20 mM Tris–HCl pH 7.5and 100 mM K2HPO4, 0.5 mM EDTA, pH 7.0, 25% (v/v) ethylene gly-col, respectively. Extinction coefficients for oxidized 6His-tagged re-combinant flavoredoxin (H6Flr) and its mutants were determined induplicate in 20 mM Tris–HCl pH 7.5 [9]. The absorption spectrumof 0.5 mg of protein in 0.5 ml was determined. The FMN was then re-moved and the supernatant containing the free FMN was measuredand its absorption spectrum recorded. The extinction coefficients usedfor free FMN were 12020 M�1 cm�1 at 445 nm and 10200 M�1 cm�1
at 380 nm [10].
2.5. Preparation of apoFlr and reconstitution of the holoproteinThe apoproteins of H6Flr and its mutants were obtained by chroma-
tography on phenyl-Sepharose (Sigma) in 1 M ammonium sulfate [11].FMN was removed from the bound protein by decreasing the pH to2.5–3. The apoflavoproteins were stored at 4 �C with 25% (v/v) ethyl-
Fig. 1. 3D model of Flr derived by comparative modeling: (a) FMN-bindinchains (labeled) rendered using thick sticks. A nickel ion is rendered using asticks. FMN carbon atoms are colored green to distinguish this group fromhydrogen bonds are represented by dashed lines. (b) Same as (a), but for ferrPymol (http://www.pymol.org).
Please cite this article in press as: Broco, M. et al., Molecular determinants foj.febslet.2007.08.009
ene glycol. The holoproteins were reconstituted by incubating apo-H6Flr with excess FMN [12]; and the excess removed with an AmiconUltra-30.
2.6. Determination of dissociation constantsDissociation constant values for the complex of FMN and apopro-
tein were determined by fluorescence titration of FMN with the apo-protein as described in Supplementary data.
2.7. Anaerobic redox titration and data analysisOxidation–reduction potentials were determined by potentiometry
with a platinum electrode to sense the potential and with a silver/silverchloride electrode as reference (Crison Instruments) and performed asin Supplementary data.
3. Results
3.1. Prediction of FMN-binding sites by comparative modeling
Most of the FMN interactions are with the backbone of the
protein, but some occur with amino acid side chains, as indi-
cated in Fig. 1c. The residues that make up the FMN-binding
pocket of Flr can be compared with: (i) those in ferric reduc-
tase and FMN-bp (FMN-binding protein), (Fig. 1a and b)
and (ii) those of highly homologous flavoredoxins from Desulf-
g pocket of the FMN-bp, with the FMN and directly contacting side-sphere and the main-chain of the protein around this zone using thinthe rest of the protein whose carbon atoms are in yellow. Potential
ic reductase. (c) Same as (a), but for Flr. Figures were produced using
r FMN-binding in Desulfovibrio gigas ..., FEBS Lett. (2007), doi:10.1016/
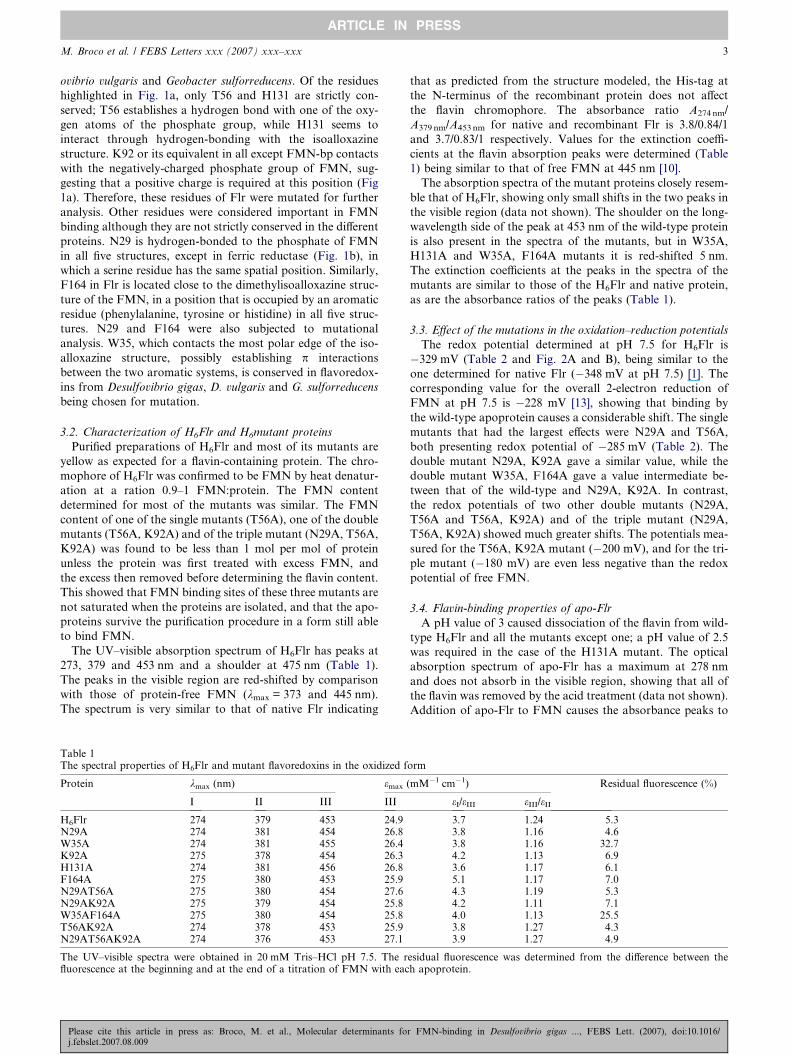
M. Broco et al. / FEBS Letters xxx (2007) xxx–xxx 3
ARTICLE IN PRESS
ovibrio vulgaris and Geobacter sulforreducens. Of the residues
highlighted in Fig. 1a, only T56 and H131 are strictly con-
served; T56 establishes a hydrogen bond with one of the oxy-
gen atoms of the phosphate group, while H131 seems to
interact through hydrogen-bonding with the isoalloxazine
structure. K92 or its equivalent in all except FMN-bp contacts
with the negatively-charged phosphate group of FMN, sug-
gesting that a positive charge is required at this position (Fig
1a). Therefore, these residues of Flr were mutated for further
analysis. Other residues were considered important in FMN
binding although they are not strictly conserved in the different
proteins. N29 is hydrogen-bonded to the phosphate of FMN
in all five structures, except in ferric reductase (Fig. 1b), in
which a serine residue has the same spatial position. Similarly,
F164 in Flr is located close to the dimethylisoalloxazine struc-
ture of the FMN, in a position that is occupied by an aromatic
residue (phenylalanine, tyrosine or histidine) in all five struc-
tures. N29 and F164 were also subjected to mutational
analysis. W35, which contacts the most polar edge of the iso-
alloxazine structure, possibly establishing p interactions
between the two aromatic systems, is conserved in flavoredox-
ins from Desulfovibrio gigas, D. vulgaris and G. sulforreducens
being chosen for mutation.
3.2. Characterization of H6Flr and H6mutant proteins
Purified preparations of H6Flr and most of its mutants are
yellow as expected for a flavin-containing protein. The chro-
mophore of H6Flr was confirmed to be FMN by heat denatur-
ation at a ration 0.9–1 FMN:protein. The FMN content
determined for most of the mutants was similar. The FMN
content of one of the single mutants (T56A), one of the double
mutants (T56A, K92A) and of the triple mutant (N29A, T56A,
K92A) was found to be less than 1 mol per mol of protein
unless the protein was first treated with excess FMN, and
the excess then removed before determining the flavin content.
This showed that FMN binding sites of these three mutants are
not saturated when the proteins are isolated, and that the apo-
proteins survive the purification procedure in a form still able
to bind FMN.
The UV–visible absorption spectrum of H6Flr has peaks at
273, 379 and 453 nm and a shoulder at 475 nm (Table 1).
The peaks in the visible region are red-shifted by comparison
with those of protein-free FMN (kmax = 373 and 445 nm).
The spectrum is very similar to that of native Flr indicating
Table 1The spectral properties of H6Flr and mutant flavoredoxins in the oxidized f
Protein kmax (nm) emax (
I II III III
H6Flr 274 379 453 24.9N29A 274 381 454 26.8W35A 274 381 455 26.4K92A 275 378 454 26.3H131A 274 381 456 26.8F164A 275 380 453 25.9N29AT56A 275 380 454 27.6N29AK92A 275 379 454 25.8W35AF164A 275 380 454 25.8T56AK92A 274 378 453 25.9N29AT56AK92A 274 376 453 27.1
The UV–visible spectra were obtained in 20 mM Tris–HCl pH 7.5. The rfluorescence at the beginning and at the end of a titration of FMN with eac
Please cite this article in press as: Broco, M. et al., Molecular determinants foj.febslet.2007.08.009
that as predicted from the structure modeled, the His-tag at
the N-terminus of the recombinant protein does not affect
the flavin chromophore. The absorbance ratio A274 nm/
A379 nm/A453 nm for native and recombinant Flr is 3.8/0.84/1
and 3.7/0.83/1 respectively. Values for the extinction coeffi-
cients at the flavin absorption peaks were determined (Table
1) being similar to that of free FMN at 445 nm [10].
The absorption spectra of the mutant proteins closely resem-
ble that of H6Flr, showing only small shifts in the two peaks in
the visible region (data not shown). The shoulder on the long-
wavelength side of the peak at 453 nm of the wild-type protein
is also present in the spectra of the mutants, but in W35A,
H131A and W35A, F164A mutants it is red-shifted 5 nm.
The extinction coefficients at the peaks in the spectra of the
mutants are similar to those of the H6Flr and native protein,
as are the absorbance ratios of the peaks (Table 1).
3.3. Effect of the mutations in the oxidation–reduction potentials
The redox potential determined at pH 7.5 for H6Flr is
�329 mV (Table 2 and Fig. 2A and B), being similar to the
one determined for native Flr (�348 mV at pH 7.5) [1]. The
corresponding value for the overall 2-electron reduction of
FMN at pH 7.5 is �228 mV [13], showing that binding by
the wild-type apoprotein causes a considerable shift. The single
mutants that had the largest effects were N29A and T56A,
both presenting redox potential of �285 mV (Table 2). The
double mutant N29A, K92A gave a similar value, while the
double mutant W35A, F164A gave a value intermediate be-
tween that of the wild-type and N29A, K92A. In contrast,
the redox potentials of two other double mutants (N29A,
T56A and T56A, K92A) and of the triple mutant (N29A,
T56A, K92A) showed much greater shifts. The potentials mea-
sured for the T56A, K92A mutant (�200 mV), and for the tri-
ple mutant (�180 mV) are even less negative than the redox
potential of free FMN.
3.4. Flavin-binding properties of apo-Flr
A pH value of 3 caused dissociation of the flavin from wild-
type H6Flr and all the mutants except one; a pH value of 2.5
was required in the case of the H131A mutant. The optical
absorption spectrum of apo-Flr has a maximum at 278 nm
and does not absorb in the visible region, showing that all of
the flavin was removed by the acid treatment (data not shown).
Addition of apo-Flr to FMN causes the absorbance peaks to
orm
mM�1 cm�1) Residual fluorescence (%)
eI/eIII eIII/eII
3.7 1.24 5.33.8 1.16 4.63.8 1.16 32.74.2 1.13 6.93.6 1.17 6.15.1 1.17 7.04.3 1.19 5.34.2 1.11 7.14.0 1.13 25.53.8 1.27 4.33.9 1.27 4.9
esidual fluorescence was determined from the difference between theh apoprotein.
r FMN-binding in Desulfovibrio gigas ..., FEBS Lett. (2007), doi:10.1016/

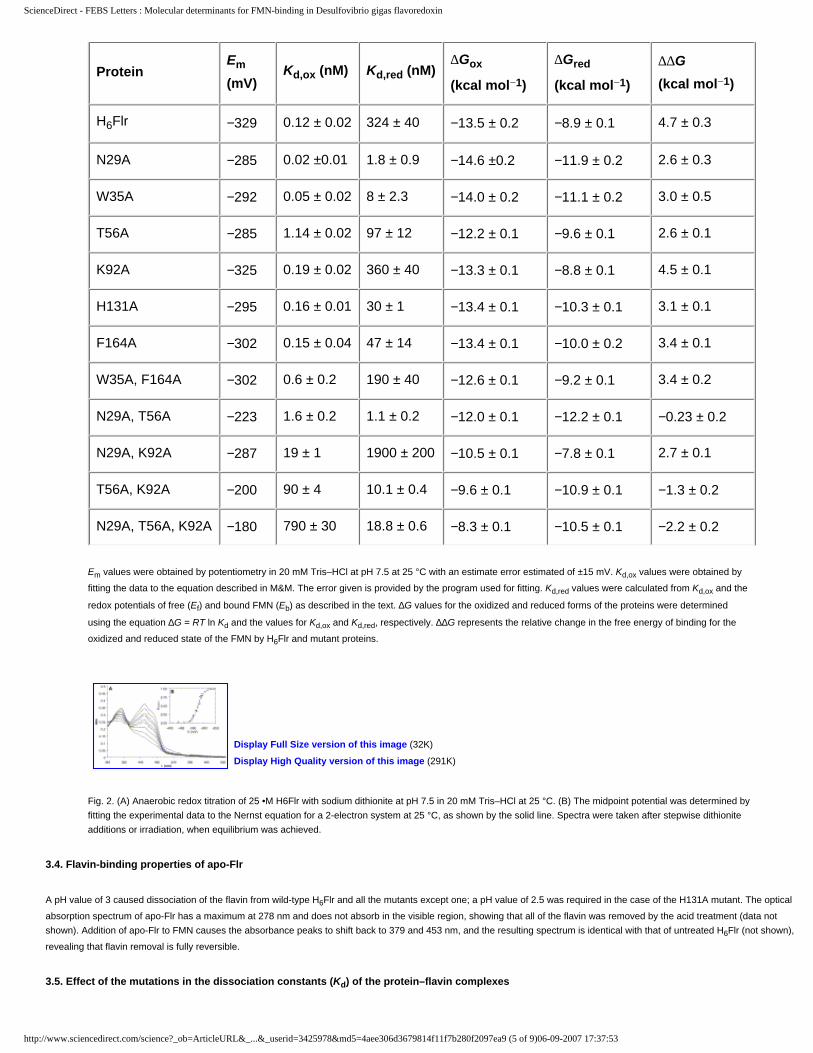
Table 2FMN dissociation constants, midpoint oxidation–reduction potentials and Gibbs free energy of FMN-binding for H6Flr and mutant flavoredoxins
Protein Em (mV) Kd,ox (nM) Kd,red (nM) DGox (kcal mol�1) DGred (kcal mol�1) DDG (kcal mol�1)
H6Flr �329 0.12 ± 0.02 324 ± 40 �13.5 ± 0.2 �8.9 ± 0.1 4.7 ± 0.3N29A �285 0.02 ±0.01 1.8 ± 0.9 �14.6 ±0.2 �11.9 ± 0.2 2.6 ± 0.3W35A �292 0.05 ± 0.02 8 ± 2.3 �14.0 ± 0.2 �11.1 ± 0.2 3.0 ± 0.5T56A �285 1.14 ± 0.02 97 ± 12 �12.2 ± 0.1 �9.6 ± 0.1 2.6 ± 0.1K92A �325 0.19 ± 0.02 360 ± 40 �13.3 ± 0.1 �8.8 ± 0.1 4.5 ± 0.1H131A �295 0.16 ± 0.01 30 ± 1 �13.4 ± 0.1 �10.3 ± 0.1 3.1 ± 0.1F164A �302 0.15 ± 0.04 47 ± 14 �13.4 ± 0.1 �10.0 ± 0.2 3.4 ± 0.1W35A, F164A �302 0.6 ± 0.2 190 ± 40 �12.6 ± 0.1 �9.2 ± 0.1 3.4 ± 0.2N29A, T56A �223 1.6 ± 0.2 1.1 ± 0.2 �12.0 ± 0.1 �12.2 ± 0.1 �0.23 ± 0.2N29A, K92A �287 19 ± 1 1900 ± 200 �10.5 ± 0.1 �7.8 ± 0.1 2.7 ± 0.1T56A, K92A �200 90 ± 4 10.1 ± 0.4 �9.6 ± 0.1 �10.9 ± 0.1 �1.3 ± 0.2N29A, T56A, K92A �180 790 ± 30 18.8 ± 0.6 �8.3 ± 0.1 �10.5 ± 0.1 �2.2 ± 0.2
Em values were obtained by potentiometry in 20 mM Tris–HCl at pH 7.5 at 25 �C with an estimate error estimated of ±15 mV. Kd,ox values wereobtained by fitting the data to the equation described in M&M. The error given is provided by the program used for fitting. Kd,red values werecalculated from Kd,ox and the redox potentials of free (Ef) and bound FMN (Eb) as described in the text. DG values for the oxidized and reducedforms of the proteins were determined using the equation DG = RT lnKd and the values for Kd,ox and Kd,red, respectively. DDG represents the relativechange in the free energy of binding for the oxidized and reduced state of the FMN by H6Flr and mutant proteins.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
340 390 440 490 540 590 640 690λ (nm)
Abs 0.00
0.25
0.50
0.75
1.00
-450 -400 -350 -300 -250E (mV)
A45
3nm
A B
Fig. 2. (A) Anaerobic redox titration of 25 lM H6Flr with sodium dithionite at pH 7.5 in 20 mM Tris–HCl at 25 �C. (B) The midpoint potential wasdetermined by fitting the experimental data to the Nernst equation for a 2-electron system at 25 �C, as shown by the solid line. Spectra were takenafter stepwise dithionite additions or irradiation, when equilibrium was achieved.
4 M. Broco et al. / FEBS Letters xxx (2007) xxx–xxx
ARTICLE IN PRESS
shift back to 379 and 453 nm, and the resulting spectrum is
identical with that of untreated H6Flr (not shown), revealing
that flavin removal is fully reversible.
3.5. Effect of the mutations in the dissociation constants (Kd) of
the protein–flavin complexes
The value determined for dissociation constant (Kd) for the
wild-type protein, 0.12 ± 0.01 nM, indicates that the non-cova-
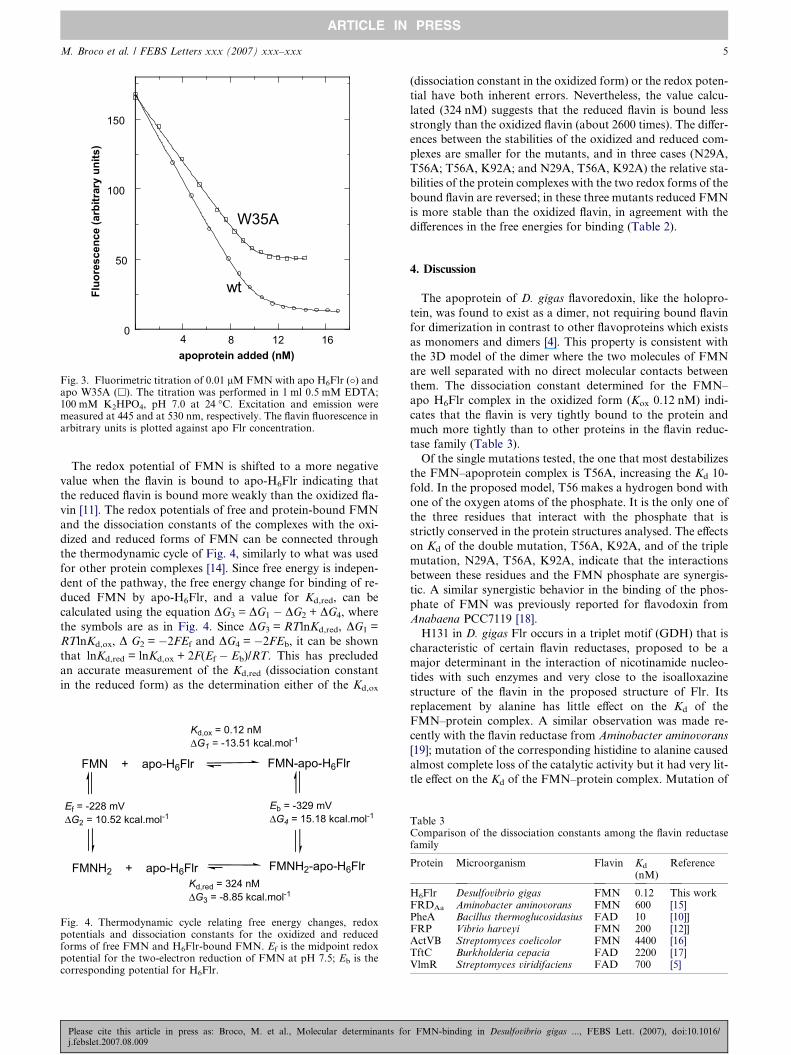
lent interactions between FMN and apo-H6Flr are very strong
(Table 2, Fig. 3), confirming the earlier qualitative observa-
tions that FMN is difficult to remove from the protein (see
Supplementary data).
The Kd value decreases the affinity for FMN (10-fold) in
T56A mutant, while the affinity increased in N29A for about
6-fold. The other four single mutants (W35, K92, H131 and
F164) did not significantly change the affinity of the apopro-
Please cite this article in press as: Broco, M. et al., Molecular determinants foj.febslet.2007.08.009
tein for FMN. In contrast, replacement of a pair of the three
amino acids predicted to interact with the FMN phosphate
(N29, T56 and K92) increased Kd between 13-fold (N29A,
T56A) and 750-fold (T56A, K92A), while replacement of all
three residues caused the greatest effect, increasing Kd by
6600-fold to 790 lM.
The flavin fluorescence is largely quenched by interaction
with the apoproteins except in the mutants W35A and
W35A, F164A, where the residual fluorescence at the end of
the titration was about 33% and 25%, respectively (Table 1
and Fig. 3). Similar levels of flavin fluorescence were observed
from the isolated forms of the holoproteins of these two
mutant proteins, suggesting that the flavin in the W35 mutants
is more exposed to the solvent, and that in the native protein
the side chain of W35 is responsible for much of the fluores-
cence quenching.
r FMN-binding in Desulfovibrio gigas ..., FEBS Lett. (2007), doi:10.1016/
0
50
100
150
Fluo
resc
ence
(arb
itrar
y un
its)
apoprotein added (nM)
wt
W35A
4 8 12 16
Fig. 3. Fluorimetric titration of 0.01 lM FMN with apo H6Flr (�) andapo W35A (h). The titration was performed in 1 ml 0.5 mM EDTA;100 mM K2HPO4, pH 7.0 at 24 �C. Excitation and emission weremeasured at 445 and at 530 nm, respectively. The flavin fluorescence inarbitrary units is plotted against apo Flr concentration.
M. Broco et al. / FEBS Letters xxx (2007) xxx–xxx 5
ARTICLE IN PRESS
The redox potential of FMN is shifted to a more negative
value when the flavin is bound to apo-H6Flr indicating that
the reduced flavin is bound more weakly than the oxidized fla-
vin [11]. The redox potentials of free and protein-bound FMN
and the dissociation constants of the complexes with the oxi-
dized and reduced forms of FMN can be connected through
the thermodynamic cycle of Fig. 4, similarly to what was used
for other protein complexes [14]. Since free energy is indepen-
dent of the pathway, the free energy change for binding of re-
duced FMN by apo-H6Flr, and a value for Kd,red, can be
calculated using the equation DG3 = DG1 � DG2 + DG4, where
the symbols are as in Fig. 4. Since DG3 = RTlnKd,red, DG1 =
RTlnKd,ox, D G2 = �2FEf and DG4 = �2FEb, it can be shown
that lnKd,red = lnKd,ox + 2F(Ef � Eb)/RT. This has precluded
an accurate measurement of the Kd,red (dissociation constant
in the reduced form) as the determination either of the Kd,ox
FMN + apo-H6Flr FMN-apo-H6Flr
FMNH2 + apo-H6Flr FMNH2-apo-H6Flr
Ef = -228 mVΔG2 = 10.52 kcal.mol-1
Eb = -329 mVΔG4 = 15.18 kcal.mol-1
Kd,ox = 0.12 nMΔG1 = -13.51 kcal.mol-1
Kd,red = 324 nMΔG3 = -8.85 kcal.mol-1
Fig. 4. Thermodynamic cycle relating free energy changes, redoxpotentials and dissociation constants for the oxidized and reducedforms of free FMN and H6Flr-bound FMN. Ef is the midpoint redoxpotential for the two-electron reduction of FMN at pH 7.5; Eb is thecorresponding potential for H6Flr.
Please cite this article in press as: Broco, M. et al., Molecular determinants foj.febslet.2007.08.009
(dissociation constant in the oxidized form) or the redox poten-
tial have both inherent errors. Nevertheless, the value calcu-
lated (324 nM) suggests that the reduced flavin is bound less
strongly than the oxidized flavin (about 2600 times). The differ-
ences between the stabilities of the oxidized and reduced com-
plexes are smaller for the mutants, and in three cases (N29A,
T56A; T56A, K92A; and N29A, T56A, K92A) the relative sta-
bilities of the protein complexes with the two redox forms of the
bound flavin are reversed; in these three mutants reduced FMN
is more stable than the oxidized flavin, in agreement with the
differences in the free energies for binding (Table 2).
4. Discussion
The apoprotein of D. gigas flavoredoxin, like the holopro-
tein, was found to exist as a dimer, not requiring bound flavin
for dimerization in contrast to other flavoproteins which exists
as monomers and dimers [4]. This property is consistent with
the 3D model of the dimer where the two molecules of FMN
are well separated with no direct molecular contacts between
them. The dissociation constant determined for the FMN–
apo H6Flr complex in the oxidized form (Kox 0.12 nM) indi-
cates that the flavin is very tightly bound to the protein and
much more tightly than to other proteins in the flavin reduc-
tase family (Table 3).
Of the single mutations tested, the one that most destabilizes
the FMN–apoprotein complex is T56A, increasing the Kd 10-
fold. In the proposed model, T56 makes a hydrogen bond with
one of the oxygen atoms of the phosphate. It is the only one of
the three residues that interact with the phosphate that is
strictly conserved in the protein structures analysed. The effects
on Kd of the double mutation, T56A, K92A, and of the triple
mutation, N29A, T56A, K92A, indicate that the interactions
between these residues and the FMN phosphate are synergis-
tic. A similar synergistic behavior in the binding of the phos-
phate of FMN was previously reported for flavodoxin from
Anabaena PCC7119 [18].
H131 in D. gigas Flr occurs in a triplet motif (GDH) that is
characteristic of certain flavin reductases, proposed to be a
major determinant in the interaction of nicotinamide nucleo-
tides with such enzymes and very close to the isoalloxazine
structure of the flavin in the proposed structure of Flr. Its
replacement by alanine has little effect on the Kd of the
FMN–protein complex. A similar observation was made re-
cently with the flavin reductase from Aminobacter aminovorans
[19]; mutation of the corresponding histidine to alanine caused
almost complete loss of the catalytic activity but it had very lit-
tle effect on the Kd of the FMN–protein complex. Mutation of
Table 3Comparison of the dissociation constants among the flavin reductasefamily
Protein Microorganism Flavin Kd
(nM)Reference
H6Flr Desulfovibrio gigas FMN 0.12 This workFRDAa Aminobacter aminovorans FMN 600 [15]PheA Bacillus thermoglucosidasius FAD 10 [10]]FRP Vibrio harveyi FMN 200 [12]]ActVB Streptomyces coelicolor FMN 4400 [16]TftC Burkholderia cepacia FAD 2200 [17]VlmR Streptomyces viridifaciens FAD 700 [5]
r FMN-binding in Desulfovibrio gigas ..., FEBS Lett. (2007), doi:10.1016/

6 M. Broco et al. / FEBS Letters xxx (2007) xxx–xxx
ARTICLE IN PRESS
both aromatic residues (mutant W35A, F164A) caused a slight
increase in Kd, suggesting that the hydrophobic region pro-
vided by these residues on the edge of the isoalloxazine struc-
ture has some importance stabilizing the FMN–protein
interaction, however, very little effect on the affinity for
FMN was observed for single mutations (Table 2).
The dissociation constants estimated for the complexes of
FMNH2 (reduced riboflavin-5 0-phosphate) and apoprotein re-
veal that wild-type H6Flr provides less stability to the hydro-
quinone than to the oxidized FMN. The hydroquinone is
similarly destabilized in most of the mutants (Table 2). In
the model developed for Flr, the NH from the main chain of
alanine 34 is near the N(5) of the isoalloxazine ring, destabiliz-
ing the complex FMNH2-Flr. Like other flavin reductases [15],
reduction of the FMN in Flr and its mutants occurs smoothly
with increasing dithionite with no accumulation of flavin sem-
iquinone or other intermediates.
The changes in the redox potential of H6Flr in the mutants
further support the assignment of the flavin-binding site in the
model. Surprisingly, the mutations close to the isoalloxazine
structure of the flavin (W35, F164 and H131) cause rather
modest changes. This probably indicates that the important
interactions with this region of the flavin involve the main
chain of the protein, rather than the side chains of individual
residues. The overall structure of the b-barrel domain to which
FMN is attached, not only in D. gigas Flr, but also to ferric
reductase from A. fulgidus and the FMN-bp from M. thermo-
autotrophicum, is more important for flavin stabilization than
the side chains of the residues [6]. In the case of flavoredoxin,
W35 and H131 contribute to the b-barrel, and the small effects
that result from their replacement by alanine are not therefore
too surprising. Similarly to what was observed for the dissoci-
ation constants, replacement of residues forming hydrogen
bonds with phosphate causes the largest effects on the redox
potential. Such effect, presumably result from changes in the
relative affinities of the protein for the oxidized and reduced
flavin and associated modifications in the flavin environment.
In conclusion, we show here that the residues interacting
with the phosphate of FMN in Flr contribute most to the bind-
ing of the flavin and to the redox properties of the flavin–pro-
tein complex.
Acknowledgements: This work was financially supported by POCTI/BME/37480/2001 to C.R-P., and POCTI/BME/32789/2000 to C.S., thefellowship PRAXIS XXI/BD/21527/99 to M.B., from FCT, and byEnterprise Ireland SC/2002/0052 and IRCSET SC/2003/018 to S.G.M.
Appendix A. Supplementary data
Supplementary data associated with this article can be
found, in the online version, at doi:10.1016/j.febslet.2007.
08.009.
References
[1] Chen, L., Liu, M.Y. and LeGall, J. (1993) Isolation andcharacterization of flavoredoxin, a new flavoprotein that permitsin vitro reconstitution of an electron transfer chain frommolecular hydrogen to sulfite reduction in the bacterium Desulf-ovibrio gigas. Arch. Biochem. Biophys. 303, 44–50.
Please cite this article in press as: Broco, M. et al., Molecular determinants foj.febslet.2007.08.009
[2] Broco, M., Rousset, M., Oliveira, S. and Rodrigues-Pousada, C.(2005) Deletion of flavoredoxin gene in Desulfovibrio gigas revealsits participation in thiosulfate reduction. FEBS Lett. 579, 4803–4807.
[3] Galan, B., Diaz, E., Prieto, M.A. and Garcia, J.L. (2000)Functional analysis of the small component of the 4-hydroxy-phenylacetate 3-monooxygenase of Escherichia coli W: a proto-type of a new flavin:AD(P)H reductase subfamily. J. Bacteriol.182, 627–636.
[4] van den Heuvel, R.H., Westphal, A.H., Heck, A.J., Walsh, M.A.,Rovida, S., van Berkel, W.J. and Mattevi, A. (2004) Structuralstudies on flavin reductase PheA2 reveal binding of NAD in anunusual folded conformation and support novel mechanism ofaction. J. Biol. Chem. 279, 12860–12867.
[5] Xun, L. (1996) Purification and characterization of chlorophenol4-monooxygenase from Burkholderia cepacia AC1100. J. Bacte-riol. 178, 2645–2649.
[6] Schroder, I., Johnson, E. and de Vries, S. (2003) Microbial ferriciron reductases. FEMS Microbiol. Rev. 27, 427–447.
[7] Chiu, H.J., Johnson, E., Schroder, I. and Rees, D.C. (2001)Crystal structures of a novel ferric reductase from the hyper-thermophilic archaeon Archaeoglobus fulgidus and its complexwith NADP+. Structure 9, 311–319.
[8] Koike, H., Sasaki, H., Kobori, T., Zenno, S., Saigo, K., Murphy,M.E., Adman, E.T. and Tanokura, M. (1998) 1.8 A crystalstructure of the major NAD(P)H:FMN oxidoreductase of abioluminescent bacterium, Vibrio fischeri: overall structure, cofac-tor and substrate-analog binding, and comparison with relatedflavoproteins. J. Mol. Biol. 280, 259–273.
[9] Macheroux, P. (1999) UV–visible spectroscopy as a tool to studyflavoproteins in: Flavoprotein Protocols (Chapman, S.K. and dnsReid, G.A.E., Eds.), pp. 1–7, Humana Press, Edinburgh, UK.
[10] Lostao, A., Gomez-Moreno, C., Mayhew, S.G. and Sancho, J.(1997) Differential stabilization of the three FMN redox forms bytyrosine 94 and tryptophan 57 in flavodoxin from Anabaena andits influence on the redox potentials. Biochemistry 36, 14334–14344.
[11] Van Berkel, W.J., Van den Berg, W.A. and Muller, F. (1988)Large-scale preparation and reconstitution of apo-flavoproteinswith special reference to butyryl-CoA dehydrogenase fromMegasphaera elsdenii. Hydrophobic-interaction chromatography.Eur. J. Biochem. 178, 197–207.
[12] Watanabe, M., Nishino, T., Takio, K., Sofuni, T. and Nohmi, T.(1998) Purification and characterization of wild-type and mutant‘‘classical’’ nitroreductases of Salmonella typhimurium. L33Rmutation greatly diminishes binding of FMN to the nitroreduc-tase of S. typhimurium. J. Biol. Chem. 273, 23922–23928.
[13] Draper, R.D. and Ingraham, L.L. (1968) A potentiometric studyof the flavin semiquinone equilibrium. Arch. Biochem. Biophys.125, 802–808.
[14] Schejter, A., Ryan, M.D., Blizzard, E.R., Zhang, C., Margoliash,E. and Feinberg, B.A. (2006) The redox couple of the cytochromec cyanide complex: the contribution of heme iron ligation to thestructural stability, chemical reactivity, and physiological behav-ior of horse cytochrome c. Protein Sci. 15, 234–241.
[15] Filisetti, L., Fontecave, M. and Niviere, V. (2003) Mechanism andsubstrate specificity of the flavin reductase ActVB from Strepto-myces coelicolor. J. Biol. Chem. 278, 296–303.
[16] Valton, J., Filisetti, L., Fontecave, M. and Niviere, V. (2004) Atwo-component flavin-dependent monooxygenase involved inactinorhodin biosynthesis in Streptomyces coelicolor. J. Biol.Chem. 279, 44362–44369.
[17] Gisi, M.R. and Xun, L. (2003) Characterization of chlorophenol4-monooxygenase (TftD) and NADH:flavin adenine dinucleotideoxidoreductase (TftC) of Burkholderia cepacia AC1100. J. Bacte-riol. 185, 2786–2792.
[18] Lostao, A., El Harrous, M., Daoudi, F., Romero, A., Parody-Morreale, A. and Sancho, J. (2000) Dissecting the energetics ofthe apoflavodoxin–FMN complex. J. Biol. Chem. 275, 9518–9526.
[19] Russell, T.R. and Tu, S.C. (2004) Aminobacter aminovoransNADH:flavin oxidoreductase His140: a highly conserved residuecritical for NADH binding and utilization. Biochemistry 43,12887–12893.
r FMN-binding in Desulfovibrio gigas ..., FEBS Lett. (2007), doi:10.1016/

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
Login: Register
Home
Browse Search My Settings Alerts Help
Quick Search Title, abstract, keywords Author e.g. j s smith
Journal/book title Volume Issue Page
Article in Press, Corrected Proof - Note to users
SummaryPlusFull Text + LinksPDF (688 K) View thumbnail images | View full size images
doi:10.1016/j.febslet.2007.08.009 Copyright © 2007 Federation of European Biochemical Societies Published by Elsevier B.V.
Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
Edited by Miguel De la Rosa
This work is dedicated to the memory of Professor António Xavier deceased on May 7th 2006†
Manuela Brocoa, Cláudio M. Soaresa, Solange Oliveiraa, b, Stephen G. Mayhewc and Claudina Rodrigues-Pousadaa, , aInstituto de Tecnologia Qu•´mica e Biológica (ITQB), Universidade Nova de Lisboa, Av. República (EAN), 2784-505 Oeiras, PortugalbDepartamento de Biologia, Universidade de Évora, Apartado 94, 7002-554 Évora, PortugalcSchool of Biomolecular and Biomedical Science, Centre for Synthesis and Chemical Biology, Conway Institute, University College Dublin, Dublin 4, Ireland Received 3 July 2007; revised 20 July 2007; accepted 1 August 2007. Available online 17 August 2007.
Abstract
Flavoredoxin participates in Desulfovibrio gigas thiosulfate reduction pathway. Its 3-dimensional model was generated allowing the oxidized riboflavin-5′-phosphate (FMN) site to be predicted. Residues likely to be involved in FMN-binding were identified (N29, W35, T56, K92, H131 and F164) and mutated to alanine. Fluorescence titration with apoprotein showed that FMN is strongly bound in the wild-type protein. Comparison of Kd values for mutants suggests that interactions with the phosphate group of FMN, contribute more to
binding than the interactions with the isoalloxazine ring. The redox potential of bound FMN determined for wild-type and mutants revealed shifts to less negative values. These findings were correlated with the protein structure in order to contribute to a better understanding of the structure–function relationships in flavoredoxin.
Keywords: Desulfovibrio gigas; Flavoredoxin; FMN-binding site-directed mutagenesis; Molecular modeling
Abbreviations: FAD, oxidized and reduced flavin adenine dinucleotide; FMN, FMNH2, oxidized and reduced riboflavin-5′-phosphate; Flr, flavoredoxin; apoFlr, apoprotein form of
flavoredoxin; flr, gene encoding flavoredoxin; H6Flr, 6His-tagged recombinant flavoredoxin; Kd, dissociation constant; FMN-bp, FMN-binding protein; Kd,ox, Kd,red, dissociation
constant in the oxidized and reduced forms; Ef, Eb, redox potential of free and bound FMN
Article Outline
1. Introduction
2. Materials and methods
2.1. Comparative modeling of Flr
2.2. Cloning, expression, purification and site-directed mutagenesis
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (1 of 9)06-09-2007 17:37:53

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
2.3. Cofactor identification and determination of the FMN:protein ratio
2.4. UV–visible absorption spectra
2.5. Preparation of apoFlr and reconstitution of the holoprotein
2.6. Determination of dissociation constants
2.7. Anaerobic redox titration and data analysis
3. Results
3.1. Prediction of FMN-binding sites by comparative modeling
3.2. Characterization of H6Flr and H6mutant proteins
3.3. Effect of the mutations in the oxidation–reduction potentials
3.4. Flavin-binding properties of apo-Flr
3.5. Effect of the mutations in the dissociation constants (Kd) of the protein–flavin complexes
4. Discussion
Acknowledgements
Appendix A. Supplementary data
References
1. Introduction
Flavoredoxin (Flr) is a 194 amino acid protein, isolated as a homodimer containing one oxidized riboflavin-5′-phosphate (FMN) per 25 kDa monomer [1]. Initially proposed to act
as a redox carrier in bisulfite reduction by molecular hydrogen, recent studies have however shown its involvement in thiosulfate reduction [2].
Proteins from the flavin reductase family catalyse the reduction of free flavins by reduced nicotinamide nucleotides, the majority of which prefer FMN rather than FAD both as cofactor in the enzyme and as the reducible substrate. Yet, in spite of their involvement in a variety of biological processes [3], the function of the reduced free flavin produced in
the reaction is poorly understood. In most cases, the flavin cofactor remains bound to the protein during enzyme isolation, indicating that the protein–flavin interaction is strong [4].
In other cases, the cofactor is weakly bound since the enzyme is isolated as an apoprotein [5].
Sequence analysis of Flr did not allow to identify motifs likely to form the FMN-binding site [6]. Three-dimensional structures have been determined for several proteins in the
flavin reductase family [4], [7] and [8], two of them sharing sequence homology, namely ferric reductase from Archaeoglobus fulgidus (PDB 1I0R) and an FMN-binding protein
from Methanobacterium thermoautotrophicum (PDB 1EJE). These structures and comparative modeling allowed us to identify a potential FMN-binding site and to predict amino acids likely to interact with the flavin. Experimental validation of these predictions showed that, by comparison with other flavin reductases, Flr has a high affinity for FMN and they provide evidence that residues N29, T56 and K92, located near the FMN phosphate group, are important for FMN-binding.
2. Materials and methods
2.1. Comparative modeling of Flr
A model for the structure of Flr dimer was derived using comparative modeling methods as described in Supplementary data.
2.2. Cloning, expression, purification and site-directed mutagenesis
The gene encoding Flr (flr) was cloned in the expression vector pET30a (+) (Novagen, Darmstadt, Germany), site-directed mutagenesis was carried out according to the quikchange® site-directed mutagenesis kit instruction manual (Stratagene, Beverly, MA, USA) and molecular mass and protein concentration were estimated as described in Supplementary data.
2.3. Cofactor identification and determination of the FMN:protein ratio
The flavin cofactor in the recombinant protein was extracted by boiling the protein for 10 min. The nature of the cofactor was determined by HPLC analysis (Nova Pak Waters Column, C18 60 Å, 3.9 × 150 mm), comparing its retention time with that of authentic FMN (Sigma, St Louis. MO, USA). The molar ratio of FMN to protein was determined as
previously described [9].
2.4. UV–visible absorption spectra
UV–visible absorption spectra of holo- and apoproteins were recorded with Shimadzu Scientific Instruments UV-3100 or Cary 50 spectrophotometers at room temperature in 20 mM Tris–HCl pH 7.5 and 100 mM K2HPO4, 0.5 mM EDTA, pH 7.0, 25% (v/v) ethylene glycol, respectively. Extinction coefficients for oxidized 6His-tagged recombinant
flavoredoxin (H6Flr) and its mutants were determined in duplicate in 20 mM Tris–HCl pH 7.5 [9]. The absorption spectrum of 0.5 mg of protein in 0.5 ml was determined. The FMN
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (2 of 9)06-09-2007 17:37:53

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
was then removed and the supernatant containing the free FMN was measured and its absorption spectrum recorded. The extinction coefficients used for free FMN were
12 020 M−1 cm−1 at 445 nm and 10 200 M−1 cm−1 at 380 nm [10].
2.5. Preparation of apoFlr and reconstitution of the holoprotein
The apoproteins of H6Flr and its mutants were obtained by chromatography on phenyl-Sepharose (Sigma) in 1 M ammonium sulfate [11]. FMN was removed from the bound
protein by decreasing the pH to 2.5–3. The apoflavoproteins were stored at 4 °C with 25% (v/v) ethylene glycol. The holoproteins were reconstituted by incubating apo-H6Flr with
excess FMN [12]; and the excess removed with an Amicon Ultra-30.
2.6. Determination of dissociation constants
Dissociation constant values for the complex of FMN and apoprotein were determined by fluorescence titration of FMN with the apoprotein as described in Supplementary data.
2.7. Anaerobic redox titration and data analysis
Oxidation–reduction potentials were determined by potentiometry with a platinum electrode to sense the potential and with a silver/silver chloride electrode as reference (Crison Instruments) and performed as in Supplementary data.
3. Results
3.1. Prediction of FMN-binding sites by comparative modeling
Most of the FMN interactions are with the backbone of the protein, but some occur with amino acid side chains, as indicated in Fig. 1c. The residues that make up the FMN-
binding pocket of Flr can be compared with: (i) those in ferric reductase and FMN-bp (FMN-binding protein), (Fig. 1a and b) and (ii) those of highly homologous flavoredoxins from
Desulfovibrio vulgaris and Geobacter sulforreducens. Of the residues highlighted in Fig. 1a, only T56 and H131 are strictly conserved; T56 establishes a hydrogen bond with one
of the oxygen atoms of the phosphate group, while H131 seems to interact through hydrogen-bonding with the isoalloxazine structure. K92 or its equivalent in all except FMN-bp contacts with the negatively-charged phosphate group of FMN, suggesting that a positive charge is required at this position (Fig 1a). Therefore, these residues of Flr were
mutated for further analysis. Other residues were considered important in FMN binding although they are not strictly conserved in the different proteins. N29 is hydrogen-bonded to the phosphate of FMN in all five structures, except in ferric reductase (Fig. 1b), in which a serine residue has the same spatial position. Similarly, F164 in Flr is located close to
the dimethylisoalloxazine structure of the FMN, in a position that is occupied by an aromatic residue (phenylalanine, tyrosine or histidine) in all five structures. N29 and F164 were also subjected to mutational analysis. W35, which contacts the most polar edge of the isoalloxazine structure, possibly establishing π interactions between the two aromatic systems, is conserved in flavoredoxins from Desulfovibrio gigas, D. vulgaris and G. sulforreducens being chosen for mutation.
Display Full Size version of this image (110K)
Display High Quality version of this image (480K)
Fig. 1. 3D model of Flr derived by comparative modeling: (a) FMN-binding pocket of the FMN-bp, with the FMN and directly contacting side-chains (labeled) rendered using thick sticks. A nickel ion is rendered using a sphere and the main-chain of the protein around this zone using thin sticks. FMN carbon atoms are colored green to distinguish this group from the rest of the protein whose carbon atoms are in yellow. Potential hydrogen bonds are represented by dashed lines. (b) Same as (a), but for ferric reductase. (c) Same as (a), but for Flr. Figures were produced using Pymol (http://www.pymol.org).
3.2. Characterization of H6Flr and H6mutant proteins
Purified preparations of H6Flr and most of its mutants are yellow as expected for a flavin-containing protein. The chromophore of H6Flr was confirmed to be FMN by heat
denaturation at a ration 0.9–1 FMN:protein. The FMN content determined for most of the mutants was similar. The FMN content of one of the single mutants (T56A), one of the double mutants (T56A, K92A) and of the triple mutant (N29A, T56A, K92A) was found to be less than 1 mol per mol of protein unless the protein was first treated with excess FMN, and the excess then removed before determining the flavin content. This showed that FMN binding sites of these three mutants are not saturated when the proteins are isolated, and that the apoproteins survive the purification procedure in a form still able to bind FMN.
The UV–visible absorption spectrum of H6Flr has peaks at 273, 379 and 453 nm and a shoulder at 475 nm (Table 1). The peaks in the visible region are red-shifted by
comparison with those of protein-free FMN (λmax = 373 and 445 nm). The spectrum is very similar to that of native Flr indicating that as predicted from the structure modeled, the
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (3 of 9)06-09-2007 17:37:53

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
His-tag at the N-terminus of the recombinant protein does not affect the flavin chromophore. The absorbance ratio A274 nm/A379 nm/A453 nm for native and recombinant Flr is
3.8/0.84/1 and 3.7/0.83/1 respectively. Values for the extinction coefficients at the flavin absorption peaks were determined (Table 1) being similar to that of free FMN at 445 nm
[10].
Table 1.
The spectral properties of H6Flr and mutant flavoredoxins in the oxidized form
Protein λmax (nm) εmax (mM−1 cm−1) Residual fluorescence (%)
I II III III εI/εIII εIII/εII
H6Flr 274 379 453 24.9 3.7 1.24 5.3
N29A 274 381 454 26.8 3.8 1.16 4.6
W35A 274 381 455 26.4 3.8 1.16 32.7
K92A 275 378 454 26.3 4.2 1.13 6.9
H131A 274 381 456 26.8 3.6 1.17 6.1
F164A 275 380 453 25.9 5.1 1.17 7.0
N29AT56A 275 380 454 27.6 4.3 1.19 5.3
N29AK92A 275 379 454 25.8 4.2 1.11 7.1
W35AF164A 275 380 454 25.8 4.0 1.13 25.5
T56AK92A 274 378 453 25.9 3.8 1.27 4.3
N29AT56AK92A 274 376 453 27.1 3.9 1.27 4.9
The UV–visible spectra were obtained in 20 mM Tris–HCl pH 7.5. The residual fluorescence was determined from the difference between the fluorescence at the beginning and at the end of a titration of FMN with each apoprotein.
The absorption spectra of the mutant proteins closely resemble that of H6Flr, showing only small shifts in the two peaks in the visible region (data not shown). The shoulder on the
long-wavelength side of the peak at 453 nm of the wild-type protein is also present in the spectra of the mutants, but in W35A, H131A and W35A, F164A mutants it is red-shifted 5 nm. The extinction coefficients at the peaks in the spectra of the mutants are similar to those of the H6Flr and native protein, as are the absorbance ratios of the peaks (Table 1).
3.3. Effect of the mutations in the oxidation–reduction potentials
The redox potential determined at pH 7.5 for H6Flr is −329 mV (Table 2 and Fig. 2A and B), being similar to the one determined for native Flr (−348 mV at pH 7.5) [1]. The
corresponding value for the overall 2-electron reduction of FMN at pH 7.5 is −228 mV [13], showing that binding by the wild-type apoprotein causes a considerable shift. The
single mutants that had the largest effects were N29A and T56A, both presenting redox potential of −285 mV (Table 2). The double mutant N29A, K92A gave a similar value,
while the double mutant W35A, F164A gave a value intermediate between that of the wild-type and N29A, K92A. In contrast, the redox potentials of two other double mutants (N29A, T56A and T56A, K92A) and of the triple mutant (N29A, T56A, K92A) showed much greater shifts. The potentials measured for the T56A, K92A mutant (−200 mV), and for the triple mutant (−180 mV) are even less negative than the redox potential of free FMN.
Table 2.
FMN dissociation constants, midpoint oxidation–reduction potentials and Gibbs free energy of FMN-binding for H6Flr and mutant flavoredoxins
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (4 of 9)06-09-2007 17:37:53
ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
ProteinEm (mV)
Kd,ox (nM) Kd,red (nM)∆Gox
(kcal mol−1)
∆Gred
(kcal mol−1)
∆∆G (kcal mol−1)
H6Flr −329 0.12 ± 0.02 324 ± 40 −13.5 ± 0.2 −8.9 ± 0.1 4.7 ± 0.3
N29A −285 0.02 ±0.01 1.8 ± 0.9 −14.6 ±0.2 −11.9 ± 0.2 2.6 ± 0.3
W35A −292 0.05 ± 0.02 8 ± 2.3 −14.0 ± 0.2 −11.1 ± 0.2 3.0 ± 0.5
T56A −285 1.14 ± 0.02 97 ± 12 −12.2 ± 0.1 −9.6 ± 0.1 2.6 ± 0.1
K92A −325 0.19 ± 0.02 360 ± 40 −13.3 ± 0.1 −8.8 ± 0.1 4.5 ± 0.1
H131A −295 0.16 ± 0.01 30 ± 1 −13.4 ± 0.1 −10.3 ± 0.1 3.1 ± 0.1
F164A −302 0.15 ± 0.04 47 ± 14 −13.4 ± 0.1 −10.0 ± 0.2 3.4 ± 0.1
W35A, F164A −302 0.6 ± 0.2 190 ± 40 −12.6 ± 0.1 −9.2 ± 0.1 3.4 ± 0.2
N29A, T56A −223 1.6 ± 0.2 1.1 ± 0.2 −12.0 ± 0.1 −12.2 ± 0.1 −0.23 ± 0.2
N29A, K92A −287 19 ± 1 1900 ± 200 −10.5 ± 0.1 −7.8 ± 0.1 2.7 ± 0.1
T56A, K92A −200 90 ± 4 10.1 ± 0.4 −9.6 ± 0.1 −10.9 ± 0.1 −1.3 ± 0.2
N29A, T56A, K92A −180 790 ± 30 18.8 ± 0.6 −8.3 ± 0.1 −10.5 ± 0.1 −2.2 ± 0.2
Em values were obtained by potentiometry in 20 mM Tris–HCl at pH 7.5 at 25 °C with an estimate error estimated of ±15 mV. Kd,ox values were obtained by
fitting the data to the equation described in M&M. The error given is provided by the program used for fitting. Kd,red values were calculated from Kd,ox and the
redox potentials of free (Ef) and bound FMN (Eb) as described in the text. ∆G values for the oxidized and reduced forms of the proteins were determined
using the equation ∆G = RT ln Kd and the values for Kd,ox and Kd,red, respectively. ∆∆G represents the relative change in the free energy of binding for the
oxidized and reduced state of the FMN by H6Flr and mutant proteins.
Display Full Size version of this image (32K)
Display High Quality version of this image (291K)
Fig. 2. (A) Anaerobic redox titration of 25 •M H6Flr with sodium dithionite at pH 7.5 in 20 mM Tris–HCl at 25 °C. (B) The midpoint potential was determined by fitting the experimental data to the Nernst equation for a 2-electron system at 25 °C, as shown by the solid line. Spectra were taken after stepwise dithionite additions or irradiation, when equilibrium was achieved.
3.4. Flavin-binding properties of apo-Flr
A pH value of 3 caused dissociation of the flavin from wild-type H6Flr and all the mutants except one; a pH value of 2.5 was required in the case of the H131A mutant. The optical
absorption spectrum of apo-Flr has a maximum at 278 nm and does not absorb in the visible region, showing that all of the flavin was removed by the acid treatment (data not shown). Addition of apo-Flr to FMN causes the absorbance peaks to shift back to 379 and 453 nm, and the resulting spectrum is identical with that of untreated H6Flr (not shown),
revealing that flavin removal is fully reversible.
3.5. Effect of the mutations in the dissociation constants (Kd) of the protein–flavin complexes
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (5 of 9)06-09-2007 17:37:53

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
The value determined for dissociation constant (Kd) for the wild-type protein, 0.12 ± 0.01 nM, indicates that the non-covalent interactions between FMN and apo-H6Flr are very
strong (Table 2, Fig. 3), confirming the earlier qualitative observations that FMN is difficult to remove from the protein (see Supplementary data).
Display Full Size version of this image (15K)
Display High Quality version of this image (119K)
Fig. 3. Fluorimetric titration of 0.01 •M FMN with apo H6Flr ( ) and apo W35A (•). The titration was performed in 1 ml 0.5 mM EDTA; 100 mM K2HPO4, pH 7.0
at 24 °C. Excitation and emission were measured at 445 and at 530 nm, respectively. The flavin fluorescence in arbitrary units is plotted against apo Flr concentration.
The Kd value decreases the affinity for FMN (10-fold) in T56A mutant, while the affinity increased in N29A for about 6-fold. The other four single mutants (W35, K92, H131 and
F164) did not significantly change the affinity of the apoprotein for FMN. In contrast, replacement of a pair of the three amino acids predicted to interact with the FMN phosphate (N29, T56 and K92) increased Kd between 13-fold (N29A, T56A) and 750-fold (T56A, K92A), while replacement of all three residues caused the greatest effect, increasing Kd by
6600-fold to 790 •M.
The flavin fluorescence is largely quenched by interaction with the apoproteins except in the mutants W35A and W35A, F164A, where the residual fluorescence at the end of the titration was about 33% and 25%, respectively (Table 1 and Fig. 3). Similar levels of flavin fluorescence were observed from the isolated forms of the holoproteins of these two
mutant proteins, suggesting that the flavin in the W35 mutants is more exposed to the solvent, and that in the native protein the side chain of W35 is responsible for much of the fluorescence quenching.
The redox potential of FMN is shifted to a more negative value when the flavin is bound to apo-H6Flr indicating that the reduced flavin is bound more weakly than the oxidized
flavin [11]. The redox potentials of free and protein-bound FMN and the dissociation constants of the complexes with the oxidized and reduced forms of FMN can be connected
through the thermodynamic cycle of Fig. 4, similarly to what was used for other protein complexes [14]. Since free energy is independent of the pathway, the free energy change
for binding of reduced FMN by apo-H6Flr, and a value for Kd,red, can be calculated using the equation ∆G3 = ∆G1 − ∆G2 + ∆G4, where the symbols are as in Fig. 4. Since
∆G3 = RTlnKd,red, ∆G1 = RTlnKd,ox, ∆ G2 = −2FEf and ∆G4 = −2FEb, it can be shown that lnKd,red = lnKd,ox + 2F(Ef − Eb)/RT. This has precluded an accurate measurement of the
Kd,red (dissociation constant in the reduced form) as the determination either of the Kd,ox (dissociation constant in the oxidized form) or the redox potential have both inherent
errors. Nevertheless, the value calculated (324 nM) suggests that the reduced flavin is bound less strongly than the oxidized flavin (about 2600 times). The differences between the stabilities of the oxidized and reduced complexes are smaller for the mutants, and in three cases (N29A, T56A; T56A, K92A; and N29A, T56A, K92A) the relative stabilities of the protein complexes with the two redox forms of the bound flavin are reversed; in these three mutants reduced FMN is more stable than the oxidized flavin, in agreement with the differences in the free energies for binding (Table 2).
Display Full Size version of this image (16K)
Display High Quality version of this image (121K)
Fig. 4. Thermodynamic cycle relating free energy changes, redox potentials and dissociation constants for the oxidized and reduced forms of free FMN and H6Flr-bound FMN. Ef is the midpoint redox potential for the two-electron reduction of FMN at pH 7.5; Eb is the corresponding potential for H6Flr.
4. Discussion
The apoprotein of D. gigas flavoredoxin, like the holoprotein, was found to exist as a dimer, not requiring bound flavin for dimerization in contrast to other flavoproteins which exists as monomers and dimers [4]. This property is consistent with the 3D model of the dimer where the two molecules of FMN are well separated with no direct molecular
contacts between them. The dissociation constant determined for the FMN–apo H6Flr complex in the oxidized form (Kox 0.12 nM) indicates that the flavin is very tightly bound to
the protein and much more tightly than to other proteins in the flavin reductase family (Table 3).
Table 3.
Comparison of the dissociation constants among the flavin reductase family
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (6 of 9)06-09-2007 17:37:53

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
Protein Microorganism Flavin Kd (nM) Reference
H6Flr Desulfovibrio gigas FMN 0.12 This work
FRDAa Aminobacter aminovorans FMN 600 [15]
PheA Bacillus thermoglucosidasius FAD 10 [10]]
FRP Vibrio harveyi FMN 200 [12]]
ActVB Streptomyces coelicolor FMN 4400 [16]
TftC Burkholderia cepacia FAD 2200 [17]
VlmR Streptomyces viridifaciens FAD 700 [5]
Of the single mutations tested, the one that most destabilizes the FMN–apoprotein complex is T56A, increasing the Kd 10-fold. In the proposed model, T56 makes a hydrogen
bond with one of the oxygen atoms of the phosphate. It is the only one of the three residues that interact with the phosphate that is strictly conserved in the protein structures analysed. The effects on Kd of the double mutation, T56A, K92A, and of the triple mutation, N29A, T56A, K92A, indicate that the interactions between these residues and the
FMN phosphate are synergistic. A similar synergistic behavior in the binding of the phosphate of FMN was previously reported for flavodoxin from Anabaena PCC7119 [18].
H131 in D. gigas Flr occurs in a triplet motif (GDH) that is characteristic of certain flavin reductases, proposed to be a major determinant in the interaction of nicotinamide nucleotides with such enzymes and very close to the isoalloxazine structure of the flavin in the proposed structure of Flr. Its replacement by alanine has little effect on the Kd of
the FMN–protein complex. A similar observation was made recently with the flavin reductase from Aminobacter aminovorans [19]; mutation of the corresponding histidine to
alanine caused almost complete loss of the catalytic activity but it had very little effect on the Kd of the FMN–protein complex. Mutation of both aromatic residues (mutant W35A,
F164A) caused a slight increase in Kd, suggesting that the hydrophobic region provided by these residues on the edge of the isoalloxazine structure has some importance
stabilizing the FMN–protein interaction, however, very little effect on the affinity for FMN was observed for single mutations (Table 2).
The dissociation constants estimated for the complexes of FMNH2 (reduced riboflavin-5′-phosphate) and apoprotein reveal that wild-type H6Flr provides less stability to the
hydroquinone than to the oxidized FMN. The hydroquinone is similarly destabilized in most of the mutants (Table 2). In the model developed for Flr, the NH from the main chain of
alanine 34 is near the N(5) of the isoalloxazine ring, destabilizing the complex FMNH2-Flr. Like other flavin reductases [15], reduction of the FMN in Flr and its mutants occurs
smoothly with increasing dithionite with no accumulation of flavin semiquinone or other intermediates.
The changes in the redox potential of H6Flr in the mutants further support the assignment of the flavin-binding site in the model. Surprisingly, the mutations close to the
isoalloxazine structure of the flavin (W35, F164 and H131) cause rather modest changes. This probably indicates that the important interactions with this region of the flavin involve the main chain of the protein, rather than the side chains of individual residues. The overall structure of the β-barrel domain to which FMN is attached, not only in D. gigas Flr, but also to ferric reductase from A. fulgidus and the FMN-bp from M. thermoautotrophicum, is more important for flavin stabilization than the side chains of the residues [6]. In
the case of flavoredoxin, W35 and H131 contribute to the β-barrel, and the small effects that result from their replacement by alanine are not therefore too surprising. Similarly to what was observed for the dissociation constants, replacement of residues forming hydrogen bonds with phosphate causes the largest effects on the redox potential. Such effect, presumably result from changes in the relative affinities of the protein for the oxidized and reduced flavin and associated modifications in the flavin environment.
In conclusion, we show here that the residues interacting with the phosphate of FMN in Flr contribute most to the binding of the flavin and to the redox properties of the flavin–protein complex.
Acknowledgements
This work was financially supported by POCTI/BME/37480/2001 to C.R-P., and POCTI/BME/32789/2000 to C.S., the fellowship PRAXIS XXI/BD/21527/99 to M.B., from FCT, and by Enterprise Ireland SC/2002/0052 and IRCSET SC/2003/018 to S.G.M.
References
[1] L. Chen, M.Y. Liu and J. LeGall, Isolation and characterization of flavoredoxin, a new flavoprotein that permits in vitro reconstitution of an electron transfer chain from
molecular hydrogen to sulfite reduction in the bacterium Desulfovibrio gigas, Arch. Biochem. Biophys. 303 (1993), pp. 44–50. Abstract | PDF (491 K) | View Record in Scopus |
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (7 of 9)06-09-2007 17:37:53

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
Cited By in Scopus (6)
[2] M. Broco, M. Rousset, S. Oliveira and C. Rodrigues-Pousada, Deletion of flavoredoxin gene in Desulfovibrio gigas reveals its participation in thiosulfate reduction, FEBS Lett.
579 (2005), pp. 4803–4807. SummaryPlus | Full Text + Links | PDF (196 K) | View Record in Scopus | Cited By in Scopus (2)
[3] B. Galan, E. Diaz, M.A. Prieto and J.L. Garcia, Functional analysis of the small component of the 4-hydroxyphenylacetate 3-monooxygenase of Escherichia coli W: a prototype
of a new flavin:AD(P)H reductase subfamily, J. Bacteriol. 182 (2000), pp. 627–636. Full Text via CrossRef | View Record in Scopus | Cited By in Scopus (63)
[4] R.H. van den Heuvel, A.H. Westphal, A.J. Heck, M.A. Walsh, S. Rovida, W.J. van Berkel and A. Mattevi, Structural studies on flavin reductase PheA2 reveal binding of NAD in
an unusual folded conformation and support novel mechanism of action, J. Biol. Chem. 279 (2004), pp. 12860–12867. View Record in Scopus | Cited By in Scopus (14)
[5] L. Xun, Purification and characterization of chlorophenol 4-monooxygenase from Burkholderia cepacia AC1100, J. Bacteriol. 178 (1996), pp. 2645–2649. View Record in
Scopus | Cited By in Scopus (38)
[6] I. Schröder, E. Johnson and S. de Vries, Microbial ferric iron reductases, FEMS Microbiol. Rev. 27 (2003), pp. 427–447. Abstract | View Record in Scopus | Cited By in Scopus
(35)
[7] H.J. Chiu, E. Johnson, I. Schröder and D.C. Rees, Crystal structures of a novel ferric reductase from the hyperthermophilic archaeon Archaeoglobus fulgidus and its complex
with NADP+, Structure 9 (2001), pp. 311–319. SummaryPlus | Full Text + Links | PDF (572 K) | View Record in Scopus | Cited By in Scopus (27)
[8] H. Koike, H. Sasaki, T. Kobori, S. Zenno, K. Saigo, M.E. Murphy, E.T. Adman and M. Tanokura, 1.8 A crystal structure of the major NAD(P)H:FMN oxidoreductase of a
bioluminescent bacterium, Vibrio fischeri: overall structure, cofactor and substrate-analog binding, and comparison with related flavoproteins, J. Mol. Biol. 280 (1998), pp. 259–273. SummaryPlus | Full Text + Links | PDF (2891 K) | View Record in Scopus | Cited By in Scopus (44)
[9] P. Macheroux, UV–visible spectroscopy as a tool to study flavoproteins. In: S.K. Chapman and G.A.E. dns Reid, Editors, Flavoprotein Protocols, Humana Press, Edinburgh,
UK (1999), pp. 1–7. View Record in Scopus | Cited By in Scopus (4)
[10] A. Lostao, C. Gomez-Moreno, S.G. Mayhew and J. Sancho, Differential stabilization of the three FMN redox forms by tyrosine 94 and tryptophan 57 in flavodoxin
fromAnabaena and its influence on the redox potentials, Biochemistry 36 (1997), pp. 14334–14344. Full Text via CrossRef | View Record in Scopus | Cited By in Scopus (44)
[11] W.J. Van Berkel, W.A. Van den Berg and F. Müller, Large-scale preparation and reconstitution of apo-flavoproteins with special reference to butyryl-CoA dehydrogenase from
Megasphaera elsdenii. Hydrophobic-interaction chromatography, Eur. J. Biochem. 178 (1988), pp. 197–207. View Record in Scopus | Cited By in Scopus (15)
[12] M. Watanabe, T. Nishino, K. Takio, T. Sofuni and T. Nohmi, Purification and characterization of wild-type and mutant “classical” nitroreductases of Salmonella typhimurium.
L33R mutation greatly diminishes binding of FMN to the nitroreductase of S. typhimurium, J. Biol. Chem. 273 (1998), pp. 23922–23928. Full Text via CrossRef | View Record in
Scopus | Cited By in Scopus (26)
[13] R.D. Draper and L.L. Ingraham, A potentiometric study of the flavin semiquinone equilibrium, Arch. Biochem. Biophys. 125 (1968), pp. 802–808. Abstract | Full Text + Links |
PDF (549 K) | View Record in Scopus | Cited By in Scopus (57)
[14] A. Schejter, M.D. Ryan, E.R. Blizzard, C. Zhang, E. Margoliash and B.A. Feinberg, The redox couple of the cytochrome c cyanide complex: the contribution of heme iron
ligation to the structural stability, chemical reactivity, and physiological behavior of horse cytochrome c, Protein Sci. 15 (2006), pp. 234–241. Full Text via CrossRef | View Record
in Scopus | Cited By in Scopus (3)
[15] L. Filisetti, M. Fontecave and V. Nivière, Mechanism and substrate specificity of the flavin reductase ActVB from Streptomyces coelicolor, J. Biol. Chem. 278 (2003), pp. 296–
303. View Record in Scopus | Cited By in Scopus (12)
[16] J. Valton, L. Filisetti, M. Fontecave and V. Niviere, A two-component flavin-dependent monooxygenase involved in actinorhodin biosynthesis in Streptomyces coelicolor, J.
Biol. Chem. 279 (2004), pp. 44362–44369. Full Text via CrossRef | View Record in Scopus | Cited By in Scopus (12)
[17] M.R. Gisi and L. Xun, Characterization of chlorophenol 4-monooxygenase (TftD) and NADH:flavin adenine dinucleotide oxidoreductase (TftC) of Burkholderia cepacia
AC1100, J. Bacteriol. 185 (2003), pp. 2786–2792. Full Text via CrossRef | View Record in Scopus | Cited By in Scopus (22)
[18] A. Lostao, M. El Harrous, F. Daoudi, A. Romero, A. Parody-Morreale and J. Sancho, Dissecting the energetics of the apoflavodoxin–FMN complex, J. Biol. Chem. 275
(2000), pp. 9518–9526. Full Text via CrossRef | View Record in Scopus | Cited By in Scopus (37)
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (8 of 9)06-09-2007 17:37:53

ScienceDirect - FEBS Letters : Molecular determinants for FMN-binding in Desulfovibrio gigas flavoredoxin
[19] T.R. Russell and S.C. Tu, Aminobacter aminovorans NADH:flavin oxidoreductase His140: a highly conserved residue critical for NADH binding and utilization, Biochemistry
43 (2004), pp. 12887–12893. Full Text via CrossRef | View Record in Scopus | Cited By in Scopus (5)
Appendix A. Supplementary data
(1101 K)
mmc1.doc
Microsoft Word file 1.
Supplementary information. Supplementary material.
Corresponding author. Fax: +351 214433644.† He played a crucial role in the field of Bioinorganic Chemistry being the founder of ITQB and contributing to the understanding of the energy transduction of the anaerobe bacteria in particular the one of D. gigas.
Note to users: The section "Articles in Press" contains peer reviewed accepted articles to be published in this journal. When the final article is assigned to an issue of the journal, the "Article in Press" version will be removed from this section and will appear in the associated published journal issue. The date it was first made available online will be carried over. Please be aware that although "Articles in Press" do not have all bibliographic details available yet, they can already be cited using the year of online publication and the DOI as follows: Author(s), Article Title, Journal (Year), DOI. Please consult the journal's reference style for the exact appearance of these elements, abbreviation of journal names and the use of punctuation.
There are three types of "Articles in Press":
● Accepted manuscripts: these are articles that have been peer reviewed and accepted for publication by the Editorial Board. The articles have not yet been copy edited and/or formatted in the journal house style.
● Uncorrected proofs: these are copy edited and formatted articles that are not yet finalized and that will be corrected by the authors. Therefore the text could change before final publication.
● Corrected proofs: these are articles containing the authors' corrections and may, or may not yet have specific issue and page numbers assigned.
Article in Press, Corrected Proof
Home
Browse Search My Settings Alerts Help
About ScienceDirect | Contact Us | Terms & Conditions | Privacy Policy
Copyright © 2007 Elsevier B.V. All rights reserved. ScienceDirect® is a registered trademark of Elsevier B.V.
http://www.sciencedirect.com/science?_ob=ArticleURL&_...&_userid=3425978&md5=4aee306d3679814f11f7b280f2097ea9 (9 of 9)06-09-2007 17:37:53




















