
Dna Barcoding a Collection of Ants (Hymenoptera: Formicidae) from Isla Del Coco, Costa Rica
Upload
independentCategory
view
0download
0
ARTICLE
Martin Pfeiffer · Karl Eduard Linsenmair
Territoriality in the Malaysian giant ant Camponotus gigas(Hymenoptera/Formicidae)
Received: October 17, 2000 / Accepted: May 14, 2001
Introduction
Most hitherto investigated ant species establish territoriesthat secure access to food resources. Depending on thespatiotemporal distribution of food availability, their strate-gies of territorial behavior vary widely (Hölldobler andLumsden 1980).
Territorial conflicts can be very costly for the ant colo-nies involved because they may lead to in wars causingmassive losses of workers on both sides (Mabelis 1979;Hölldobler 1983). In this context, ritual combat behaviormay have evolved as a means to solve territorial disputes ina less costly way. Especially well investigated are the ritualtournaments in Myrmecocystus mimicus (Hölldobler 1981).Such behavior is also known from Iridomyrmex purpureus(Ettershank and Ettershank 1982) and I. pruinosum(Hölldobler 1982), from harvester ants of the genus Messor(Yamaguchi 1995), and from interspecific interactionsbetween Polyrhachis laboriosa and Camponotus brutus(Mercier and Dejean 1996; Mercier et al. 1997). The ritualcombat of Formica lugubris and Formica rufa (Le Moli etal. 1982; Le Moli and Parmigiani 1982) includes a simulatedacid spraying, as is also found in Camponotus americanus(Carlin and Hölldobler 1983). Ritual fights are knownfrom Lasius niger (Czechowski 1984) and Leptothoraxacervorum (Dobrzanski 1966) as well.
The Malaysian giant forest ant Camponotus gigas exhib-its ritual and nonritual fighting behavior. In the lowlanddipterocarp rain forest of Sabah, on Borneo, we made a24-month study (spread over 5 years) of the behavioralecology of these mainly arboreal ants (Pfeiffer 1997). Herewe present our findings on the territorial habits of thisspecies. We especially wanted to know (1) whether majorsare distributed randomly or according to a certain patternamong the nests of a polydomous colony, (2) whichmeans are used to maintain territorial control, and (3) howterritorial conflicts are solved in intra- and interspecificconflicts.
M. Pfeiffer* (*) · K.E. LinsenmairTheodor-Boveri-Institute of the University of Wuerzburg,Department for Animal Ecology and Tropical Biology (Zoology III),Wuerzburg, Germany
Present address:* University of Ulm, Department of Experimental Ecology (Bio III),Albert-Einstein Allee 11, D-89069 Ulm, GermanyTel. �49-731-502-2679; Fax �49-731-502-2683e-mail: [email protected]
Abstract In a 5-ha area of primary lowland rain forestin Borneo, we observed four polydomous colonies of thenight-active giant ant Camponotus gigas. The nonover-lapping, three-dimensional territories in the canopy had aground size up to 0.8ha. C. gigas showed a distinct territo-rial behavior: (1) specific “barrack” nests, especially con-taining many majors, were situated at the borders and wereestablished during long-term territorial conflicts; (2) trunktrails were regularly patrolled by majors that attacked alienconspecifics and some other ant species violently; and (3)sentinels, often involved in long-enduring conflicts withneighboring ant colonies, defended the borders at bridge-heads. Interspecific conflicts with sympatric Camponotusspecies always led to violent, “bloody” fights of all castes.Intraspecific conflicts, however, were solved by ritual fights(“front leg boxing”) of majors. C. gigas performed a de-escalation strategy to end short periods of true intraspecific“ant war” that we provoked experimentally. Artificiallyinduced ritualized combats continued over weeks also inthe absence of baits, indicating that borders may becomeestablished by long-term conflicts of attrition. We discussthe differences between ritual fights in desert and rain-forest ants and apply Lanchester’s theory of battles to ourfindings.
Key words Camponotus gigas · Territory · Antagonistic be-havior · Ritualized fighting · Borneo · Rain forest
J Ethol (2001) 19:75–85 © Japan Ethological Society and Springer-Verlag Tokyo 2001

76
Material and methods
Observation area and time
Our observation plot was a 5-ha area of primary mixeddipterocarp lowland forest on Borneo in Kinabalu NationalPark, Malaysia (for details, see Pfeiffer and Linsenmair2000), where the ant community is the most species richso far described for a tropical rain forest, containing 524morphospecies from 73 genera (Brühl et al. 1998). BecauseC. gigas is predominantly night active, we usually startedour observations around 1700, 0.5 h before the ants’ mainforaging activity, and recorded data during the night-time using red-filtered headlamps. Observations tookplace during five periods totaling 2 years between July 1991and November 1995, with 1994 as the main observationtime.
Species studied
Camponotus (Dinomyrmex) gigas Latreille 1802 is one ofthe largest ant species in the world and a dominant memberof the ant communities of the Southeast Asian rain forests.Its aseasonal mating flight pattern combines phase-shiftedreproductive cycles that are not synchronized within thewhole population and an approximately semiannual rhythmresulting in a period of 188 � 5 days (Pfeiffer andLinsenmair 1997). The foraging strategy of Camponotusgigas functions according to the predictions of the centralplace foraging theory and is based on its polydomouscolony structure in combination with efficient communica-tion, polyethism, and ergonomic optimization (Pfeiffer andLinsenmair 1998).
Focus colony A had a territory of 0.8ha and about 7000foragers; roughly 13% of them were majors. Colony B hadan area of about 0.57ha, colony C about 0.35 ha, and colonyD about 0.66ha (Pfeiffer and Linsenmair 2000). Colonieswere polydomous, with 8 to 14 nests (Fig. 1).
Determination of the proportion of minors and majors atdifferent nests of the colony
To determine the number of foragers and the proportionsof majors at different nests of the polydomous colonies, wecounted the number of ants leaving the nests during theexodus period for 1.5h after the start of nocturnal foragingactivity (for details, see Pfeiffer and Linsenmair 2000).Within this time most foragers had left their nests and ma-jors had started patrolling.
According to their activity (high, medium, low), theirfood input (“sink” nest or “source” nest), and their positionwithin the colony (central, near foraging trees, at the bor-der) (see Pfeiffer and Linsenmair 1998, 2000), we distin-guished between “central” (n � 6), “minor” (n � 8), and“barrack” nests (n � 3). We tested for differences in majors’share of total activity among nests of these categories withan ANOVA.
Analysis of major worker patrolling
We marked 90 major workers at three neighboring nests ofcolony A (at Q, 54 ants; at H, 30 ants; at E, 6 ants; see Fig.1) to analyze their patrolling behavior. For this purpose weelicited alarm by attacking the guards at the nest entrancewith tweezers and caught all majors that left the nests fordefense. These ants were marked individually with smallplastic tags and were observed during the following 10nights for 2 � 5 h and 2 � 3 h at three nests simultaneouslyby three observers, and for 6 � 1h by one observer walkingalong the trails between the nests.
Analysis of antagonistic behavior
We studied the fighting behavior of ants of two hostile C.gigas colonies at a natural tournament place at ground level
Fig. 1. Territorial distribution of four colonies of Camponotus gigasand colonies of other large, night-active, and competing ant species inour research area in June 1994. The territorial borders of the C. gigascolony A (bold solid lines) have been proven by experiments (seePfeiffer and Linsenmair 2000). The borders of other C. gigas territorieswere roughly determined by various observations (solid lines), all otherterritorial borders are only guessed at (dotted lines). The nests of C.gigas are symbolized by black dots (�). In the center is colony A, withthe black lines indicating the bamboo runway system between the nestsand letters marking some nests that we mention in this article. In colonyB, black lines and squares represent the bridges and towers of theCanopy Walk Way that served for ants as a trunk trail. This colony ofC. gigas consisted of two parts that had been divided by a colony ofOecophylla. White dots (�) represent nests of Camponotus c.f. festinus;( ), nests of two other Camponotus species (C. 64 and C. 75); , nestsof Oecophylla smaragdina (Oeco), , places of tournaments that hadbeen elicited by our experiments; , permanent sentries of C. gigas atthe base of trees; , permanent sentries of both C. gigas and C. c.f.festinus

77
during 1 week in April 1993. For a further experimentalanalysis of intraspecific aggression behavior, we chose colo-nies A and B in our field site. At their common border ineach of the territories, we built a table that was isolatedfrom the ground by use of “Tangle Foot” insect glue andconnected by laths of bamboo to the trail systems of theconcerned colonies. On these tables we fed ants at petridishes filled with sugar water for 1 week, until ants acceptedthese dishes as part of their territories. Then, both tableswere connected with bamboo laths (through a small stripeof territory of C. festinus; see Fig. 1), and ants of bothcolonies were fed jointly at one petri dish. Within 30min, an“ant war” broke out in the contact zone. Using this assay westarted long-term experiments on May 1, 1994, May 8, 1994,and October 20, 1995.
We observed the behavior of the fighting colonies overtime intervals of 2 weeks to 1.5 months. On each of 10 dayswe recorded about 30min of fights on a video recorder(Panasonic NV-MS95E, 12V red video lamp; Germany);these records were later analyzed to obtain the temporalparameters of the behavioral sequences.
The influence of colony-specific trail marking on thedominance behavior of ants was studied in 85 confrontationexperiments, by placing two ants of the same casteof antagonistic colonies on a 1-m-long section of trailand observing their interactions. Trail sections were re-newed after each confrontation and were taken fromthe colonies’ artificial trails that had been in use for atleast 1 week. We checked for two behaviors: dominancein the fight and jumping off the trail. In 20 similar experi-ments we tested with fresh bamboo laths (without chemicaltrails). We analyzed interspecific aggression behavior inlong-term experiments with C. gigas colonies A and B andone adjacent colony of Camponotus (Tanaemyrmex) festinusF.R. Smith (specimens are deposited with the collection ofthe University of Würzburg, Camponotus no. 11).
Colonies were connected by bamboo trails. To simulatean efficient trophobiont, we used infusion systems filled withsugar water (10% solution) that was continuously dripping(2–5 drops/min). These baits, which were refilled every day,were installed inside the territory of C. festinus, but veryclose to the borders of the two C. gigas territories. In eightsimilar assays we provided petri dishes with human urine(supplied as a nitrogen source; see Pfeiffer and Linsenmair2000) or sugar water at bamboo trails between coloniesof C. gigas and adjacent colonies of other Camponotusspecies. Interspecific competition behavior at ground levelwas examined at 300 petri dishes filled with urine that wereplaced in twenty 100-m transects throughout our plot. Re-cruitment behavior inside the territory and at the bordersseveral days after fighting was studied at petri dishes filledwith urine. We recorded the number of minors and majorsthat appeared at the baits and the time of arrival of the firstmajors.
Results
Majors activity pattern within the focus colony
The distribution of majors among the worker force of thedifferent nests of colony A was compared by analyzing thetotal numbers of majors and minors we had counted at eachnest during several observations of the evening exodus ofthe ants. Differences at the 17 nests of the colony werehighly significant (chi-square test; df � 33, �2 � 2723.8, P �0.001). When we tested differences between nest categories(see Methods), majors’ share of total activity differedsignificantly between the barrack nests [mean share of ma-jors (MSM), 24.9%] that lay near the colony’s borders andwere the starting points of majors for patrols to the frontier,and both other groups: central nests (MSM, 2.8%) andminor nests (MSM, 0.4%; ANOVA: F(2,14) � 32.3, P �0.001; Scheffé post hoc test: P � 0.001 for both groups; seeTable 1).
High numbers of majors were also found at the centralnests of the colony. Some of them were sentries at the nestentrances. When we elicited alarm by attacking sentrieswith tweezers, dozens of majors (up to 50 at central nestQ) rapidly left the nest and spread out to patrol the sur-rounding areas (about 1.50m around the nest) for about5–10min.
Patrolling of majors
The trail system and the areas around the nest entranceswere regularly patrolled by majors that attacked alien con-specifics or ants of some of the other species that wereoccasionally met on the trunk trails. These ants were eitherforced to leave the trail by jumping to the ground or theywere captured and carried to the nest. Patrolling startedshortly after exodus and stopped at dawn when the mainactivity period ceased. At ground, ants left the nest for shortround trips within an area of 1–3m2; on trails, they movedfor about 5–10m to the track’s next turnoff, then they cameback and scouted out the other direction before they re-turned to their nest. Extended patrols could reach the nextnests at the trails. Then, majors entered these nests in mostcases and stayed there for a few minutes before they re-turned to their starting point.
Of 90 majors that we had marked individually, we ob-served 50 again during the following 10 nights; 32 of thesethat had been tagged at the central nest Q were reobserved7.25 (mean) times; only 70.6% of these observations weremade at nest Q, and the rest at neighboring nests or on thetrunk roads between them. At nest H, 18 marked majorswere reobserved on average 5.94 times; 79.4% of theseobservations occurred at nest H. Patrolling behavior of the8 most active majors is shown in Table 2. Patrols of singleants were staggered. The trails between the nests were runfrom both sides, so patrols of neighboring nests led to com-plete guarding of the trails.

78
Sentries at the colony borders
At strategically most important points (e.g., at the base offoraging trees and at the end points of the bamboo trails), C.gigas established steady sentinels. Every night small groupsof major workers (mostly two to four ants) went out tostand sentry at these well-defined places; at dawn they re-treated to their nests. Inside our observation area we de-tected 12 of these sentries (see triangles in Fig. 1), all nearground level. When we carried out spot checks we foundthat ants guarded these posts over long periods of time(mean, 77 days; n � 6; range, 30–139; SD, 40.7). When wemarked majors of three colonies of C. gigas individually attheir posts, we recorded that single individuals were presentat one spot for a mean of 8.7 consecutive nights (n � 41;range, 2–20; SD, 5.04), thus performing a special form ofortstreue (site fidelity).
In many cases these sentinels were involved in long-termconflicts with rivaling C. gigas colonies or colonies of othersympatric Camponotus species. Small groups of their oppo-nents were found in their immediate neighborhood. From
time to time single majors of both sides stalked to the an-tagonistic sentries, and often these “scout patrols” resultedin combats (see following).
Results of the fighting experiments: starting an “ant war”
We provoked three long-term conflicts between adjacentcolonies to obtain more information about the conflict strat-egies of C. gigas. In our first experiment about 50 minors ofboth sides gathered at one bait, and fighting among thembroke out within 20 min. Ants threatened each other withopened mandibles and body jerking; some gripped eachother at the mandibles and pulled in different directions.Colony A recruited about 90 majors from central nest Qthat first gathered in a large column on their part of thebamboo trail and investigated their withdrawing foragers byintensive antennation. Suddenly, all majors of A attackedsimultaneously and occupied the tabletop, killing or badlyinjuring several minors of B that mostly fled and gathered attheir trail.
Table 1. The number of ants and the portion of majors leaving and entering at “central” (Q, D,T, F, Z, B), “minor” (E, H, N, R, L, W, M, V), and “barrack” (O, J, P) nests of colony A duringpeak activity time at sunset
Nest Observation- Mean number of Mean number of Percent of majorperiods each 15min ants/period majors/period workers
Q 155 149.3 3.7 2.5D 18 141.1 2.1 1.5T 30 195.7 6.9 3.5F 88 53.3 1.6 3.0Z 35 35.8 0.8 2.1B 12 45.5 2 5.0E 148 130.4 0.2 0.1H 107 103.1 1.0 1.0N 39 50.5 0.2 0.4R 27 43.1 0.3 0.7L 23 40.0 0.0 0.0W 21 20.1 0.0 0.0M 15 150.7 1.0 0.7V 9 39.2 0.1 0.3O 9 30.2 3.7 12.1J 7 32.4 11.7 36.2P 7 10.8 2.9 26.4
Activity was measured at the nest entrances in periods of 15 min; the share of majors at barracknests differed significantly from those of both other categories
Table 2. Activity pattern of eight major workers most active in patrolling during our examination
No. of Camponotus gigas Observation Number of patrolsindividuals nights
Nest Q Nest H Nest E n
S 139 5 16 1 0 17S 200 3 4 10 2 16S 208 5 2 9 3 14S 247 3 12 2 0 14S 249 7 20 3 1 24S 267 5 2 25 0 27S 277 3 2 11 0 13
Mean 3.88 7.25 7.62 0.75 15.6
Listed are ant numbers, the number of nights we noticed the ant, the number of patrols at thedifferent nests, and total n
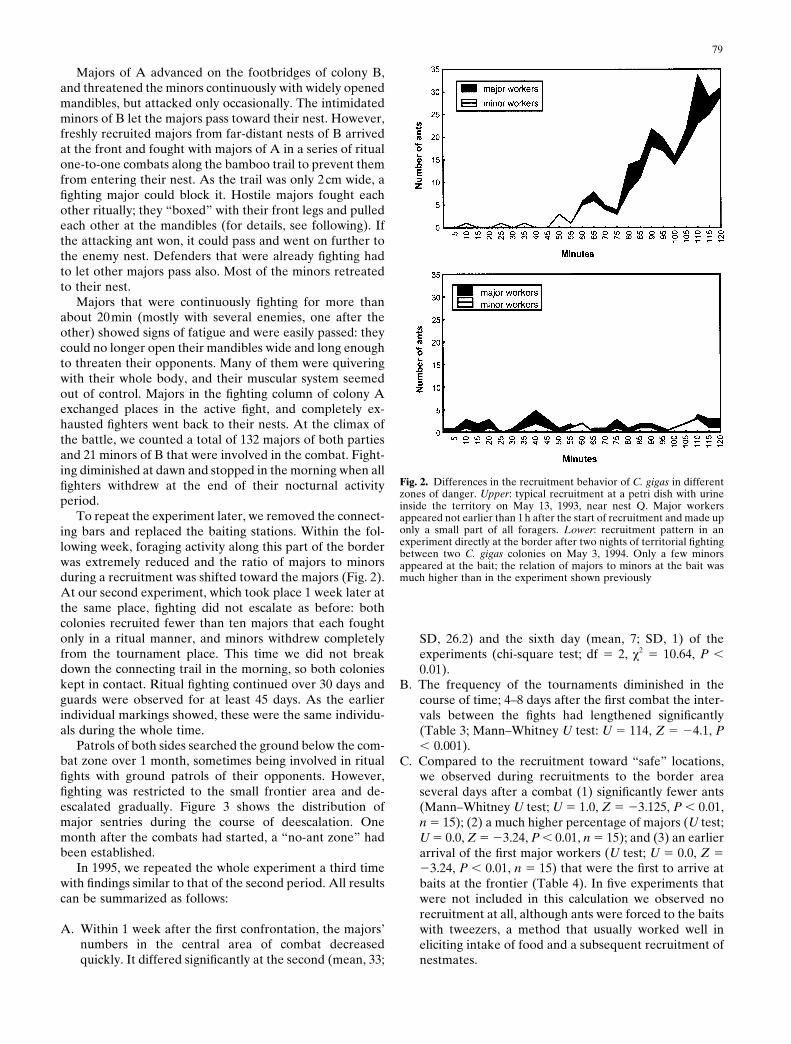
79
Majors of A advanced on the footbridges of colony B,and threatened the minors continuously with widely openedmandibles, but attacked only occasionally. The intimidatedminors of B let the majors pass toward their nest. However,freshly recruited majors from far-distant nests of B arrivedat the front and fought with majors of A in a series of ritualone-to-one combats along the bamboo trail to prevent themfrom entering their nest. As the trail was only 2cm wide, afighting major could block it. Hostile majors fought eachother ritually; they “boxed” with their front legs and pulledeach other at the mandibles (for details, see following). Ifthe attacking ant won, it could pass and went on further tothe enemy nest. Defenders that were already fighting hadto let other majors pass also. Most of the minors retreatedto their nest.
Majors that were continuously fighting for more thanabout 20min (mostly with several enemies, one after theother) showed signs of fatigue and were easily passed: theycould no longer open their mandibles wide and long enoughto threaten their opponents. Many of them were quiveringwith their whole body, and their muscular system seemedout of control. Majors in the fighting column of colony Aexchanged places in the active fight, and completely ex-hausted fighters went back to their nests. At the climax ofthe battle, we counted a total of 132 majors of both partiesand 21 minors of B that were involved in the combat. Fight-ing diminished at dawn and stopped in the morning when allfighters withdrew at the end of their nocturnal activityperiod.
To repeat the experiment later, we removed the connect-ing bars and replaced the baiting stations. Within the fol-lowing week, foraging activity along this part of the borderwas extremely reduced and the ratio of majors to minorsduring a recruitment was shifted toward the majors (Fig. 2).At our second experiment, which took place 1 week later atthe same place, fighting did not escalate as before: bothcolonies recruited fewer than ten majors that each foughtonly in a ritual manner, and minors withdrew completelyfrom the tournament place. This time we did not breakdown the connecting trail in the morning, so both colonieskept in contact. Ritual fighting continued over 30 days andguards were observed for at least 45 days. As the earlierindividual markings showed, these were the same individu-als during the whole time.
Patrols of both sides searched the ground below the com-bat zone over 1 month, sometimes being involved in ritualfights with ground patrols of their opponents. However,fighting was restricted to the small frontier area and de-escalated gradually. Figure 3 shows the distribution ofmajor sentries during the course of deescalation. Onemonth after the combats had started, a “no-ant zone” hadbeen established.
In 1995, we repeated the whole experiment a third timewith findings similar to that of the second period. All resultscan be summarized as follows:
A. Within 1 week after the first confrontation, the majors’numbers in the central area of combat decreasedquickly. It differed significantly at the second (mean, 33;
SD, 26.2) and the sixth day (mean, 7; SD, 1) of theexperiments (chi-square test; df � 2, �2 � 10.64, P �0.01).
B. The frequency of the tournaments diminished in thecourse of time; 4–8 days after the first combat the inter-vals between the fights had lengthened significantly(Table 3; Mann–Whitney U test: U � 114, Z � �4.1, P� 0.001).
C. Compared to the recruitment toward “safe” locations,we observed during recruitments to the border areaseveral days after a combat (1) significantly fewer ants(Mann–Whitney U test; U � 1.0, Z � �3.125, P � 0.01,n � 15); (2) a much higher percentage of majors (U test;U � 0.0, Z � �3.24, P � 0.01, n � 15); and (3) an earlierarrival of the first major workers (U test; U � 0.0, Z ��3.24, P � 0.01, n � 15) that were the first to arrive atbaits at the frontier (Table 4). In five experiments thatwere not included in this calculation we observed norecruitment at all, although ants were forced to the baitswith tweezers, a method that usually worked well ineliciting intake of food and a subsequent recruitment ofnestmates.
Fig. 2. Differences in the recruitment behavior of C. gigas in differentzones of danger. Upper: typical recruitment at a petri dish with urineinside the territory on May 13, 1993, near nest Q. Major workersappeared not earlier than 1h after the start of recruitment and made uponly a small part of all foragers. Lower: recruitment pattern in anexperiment directly at the border after two nights of territorial fightingbetween two C. gigas colonies on May 3, 1994. Only a few minorsappeared at the bait; the relation of majors to minors at the bait wasmuch higher than in the experiment shown previously
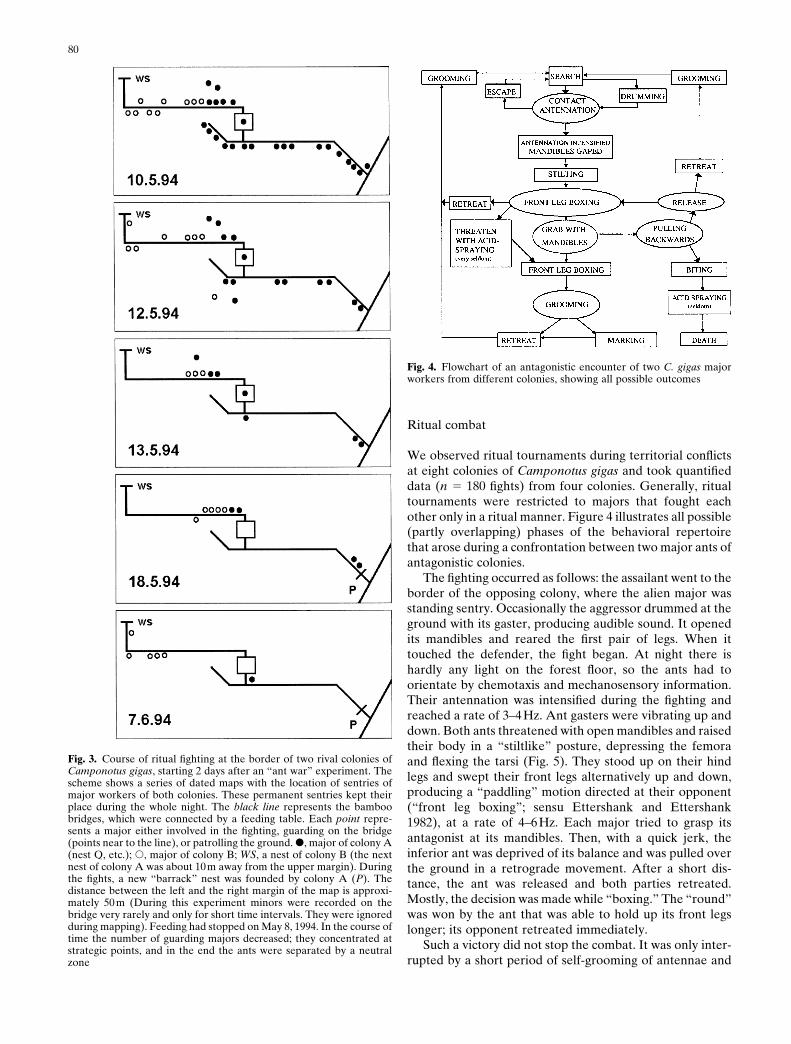
80
Ritual combat
We observed ritual tournaments during territorial conflictsat eight colonies of Camponotus gigas and took quantifieddata (n � 180 fights) from four colonies. Generally, ritualtournaments were restricted to majors that fought eachother only in a ritual manner. Figure 4 illustrates all possible(partly overlapping) phases of the behavioral repertoirethat arose during a confrontation between two major ants ofantagonistic colonies.
The fighting occurred as follows: the assailant went to theborder of the opposing colony, where the alien major wasstanding sentry. Occasionally the aggressor drummed at theground with its gaster, producing audible sound. It openedits mandibles and reared the first pair of legs. When ittouched the defender, the fight began. At night there ishardly any light on the forest floor, so the ants had toorientate by chemotaxis and mechanosensory information.Their antennation was intensified during the fighting andreached a rate of 3–4Hz. Ant gasters were vibrating up anddown. Both ants threatened with open mandibles and raisedtheir body in a “stiltlike” posture, depressing the femoraand flexing the tarsi (Fig. 5). They stood up on their hindlegs and swept their front legs alternatively up and down,producing a “paddling” motion directed at their opponent(“front leg boxing”; sensu Ettershank and Ettershank1982), at a rate of 4–6Hz. Each major tried to grasp itsantagonist at its mandibles. Then, with a quick jerk, theinferior ant was deprived of its balance and was pulled overthe ground in a retrograde movement. After a short dis-tance, the ant was released and both parties retreated.Mostly, the decision was made while “boxing.” The “round”was won by the ant that was able to hold up its front legslonger; its opponent retreated immediately.
Such a victory did not stop the combat. It was only inter-rupted by a short period of self-grooming of antennae and
Fig. 3. Course of ritual fighting at the border of two rival colonies ofCamponotus gigas, starting 2 days after an “ant war” experiment. Thescheme shows a series of dated maps with the location of sentries ofmajor workers of both colonies. These permanent sentries kept theirplace during the whole night. The black line represents the bamboobridges, which were connected by a feeding table. Each point repre-sents a major either involved in the fighting, guarding on the bridge(points near to the line), or patrolling the ground. �, major of colony A(nest Q, etc.); �, major of colony B; WS, a nest of colony B (the nextnest of colony A was about 10m away from the upper margin). Duringthe fights, a new “barrack” nest was founded by colony A (P). Thedistance between the left and the right margin of the map is approxi-mately 50 m (During this experiment minors were recorded on thebridge very rarely and only for short time intervals. They were ignoredduring mapping). Feeding had stopped on May 8, 1994. In the course oftime the number of guarding majors decreased; they concentrated atstrategic points, and in the end the ants were separated by a neutralzone
Fig. 4. Flowchart of an antagonistic encounter of two C. gigas majorworkers from different colonies, showing all possible outcomes

81
front legs. In these breaks, ants often retreated to theirterritories, where two or three of their major nestmateswere still on guard. Sometimes (n � 18 of 180) the fightersstarted boxing again when they met their nestmates, butthey did it in a weakened form. The sequence of the move-ments was much slower in comparison to real fights. The
mandibles were opened only half way, and the tarsi werenot raised as high and never touched the nestmate.
When an assailant retreated, its opponent marked itstrack, pulling its gaster close over the ground. We observeddrops coming out from the acidoporus, so a marking sub-stance may have come from the hindgut or poison gland.However, as the gaster was dragged along the ground withits full length, additional glands may be involved in thismarking. After a few minutes’ break, ants returned to thetournament site: if they were tired, they were relieved bytheir nestmates. Usually fighting lasted the whole night.Only very seldom, and just for a short time, did one groupretreat as a whole.
On average, a single bout including antennation, stilting,front leg boxing, and grasping with the mandibles lasted86.43s (n � 62; SD � 79.86), and the interval between thefights 335s (n � 63; SD � 600.89). However, length of thedifferent parts was quite flexible. Mostly, just one or twopairs of majors were fighting, while the other majors wereinvolved in the combat only rarely. We rarely observed twoor three majors attacking one opponent on the ground, but,if so, this never happened other than in a ritual way. Yet, wenoted some majors carrying on their bodies the cutoff headsof dead majors that had taken firm hold with their man-dibles; these were clear signs of nonritual fights among C.
Table 3. Duration of ritual fights and the intervals between them, measured in different periodsafter three “ant wars”
Period Fight [s] SD n Interval [s] SD n
After 1–2 days 66.0 44.4 32 64.9*** 97.2 22After 4–8 days 108.2 101.8 30 527.5*** 801.8 31
Mean/total 86.4 79.9 62 335.5 653.9 53
In the first 2 days after the war, ritual tournaments between single majors of each colony werecarried out in short intervals; 4–8 days later, the intervals between the fights had lengthenedsignificantly (marked as ***); the length of the fights, however, did not differ significantly
Table 4. Recruitments of majors versus minors to baits inside the territory and at the borders, several days after fighting Group T (upper):recruitments of C. gigas to baits inside its territory; group B (lower): recruitments at the borders, several days after fighting
Date Duration of n Group Arrival of Mean SD Mean SD Mean Percent ofexperiment first major number number of ants majors(min) (min) of majors of minors
13.05.93 120 24 T 55 1.88 2.64 7.83 9.18 9.71 1917.05.93 160 83 T 54 2.87 2.50 15.93 11.27 18.80 1520.05.93 110 85 T 42 3.84 2.95 13.89 9.74 17.73 2217.02.94 180 17 T 135 0.29 0.59 4.53 6.23 4.82 618.02.94 90 16 T 73 0.56 0.81 9.75 10.01 10.31 531.03.94 180 123 T 15 0.13 0.34 6.69 3.62 6.82 201.07.94 60 59 T 50 0.14 0.43 16.31 7.17 16.44 103.05.94 120 24 B 0 1.04 1.23 0.83 0.82 1.88 5604.05.94 90 11 B 0 3.64 1.57 2.27 0.90 5.91 6206.05.94 60 14 B 0 1.07 0.83 1.50 1.22 2.57 4209.05.94 120 12 B 0 0.86 0.38 0 0.00 0.86 10009.05.94 110 7 B 0 0.67 0.52 0 0.00 0.67 10030.10.95 390 13 B 0 0.25 0.46 0 0.00 0.25 10030.10.95 390 13 B 0 0.75 1.39 0 0.00 0.75 10031.10.95 60 6 B 0 1.00 1.00 1 1.00 2.00 50
Given are date, the length of the experiment, the number of observations, the arrival of the first major after recruitment had started (0min meansthat a major was the first at the bait), the mean numbers of majors and minors and their SDs, the total number of ants, and the percentage of themajors. The small number of observations in group B is due to the low activity at the border. During these observations there were hardly anychanges in the number of ants visiting the baits
Fig. 5. Two major workers of Camponotus gigas in a ritual fight

82
gigas. In only 2 of 180 fights that we observed did aggressionbecome so intense that neither fighter released their man-dible grip; instead, they sprayed acid on each other and diedwithin minutes.
Trail marking and dominance
We tried to establish by confrontation experiments whetherterritorial marking of trails affected the dominance of mi-nors from different colonies toward each other or thechance of majors to win in ritual tournaments. We com-pared behavior on own and alien trails (Table 5). Minorswere more dominant on their own trails than on alien trails[a; G test (df � 1), G � 5.01, n � 98; Bonferroni corrected,P � 0.05] and jumped more often from alien trails [b; G test(df � 1), G � 5.39, n � 98; Bonferroni corrected, P � 0.05].Majors showed a similar tendency; however, we found nosignificant influence of trail marking on their behavior.(This finding was probably because the experimental settingwas too artificial and put both parties under stress.)
When tested on a neutral trail both castes showed signifi-cant behavioral differences: 8 of 18 minors (44.4%) with-drew and tried to hide below the trail, whereas only 1 of 22majors (4.5%) did so (Fisher test; n � 18, Bonferroni cor-rected, P � 0.05). Taken together, all workers jumped lessoften from their own trails (14 of 84) than from neutral trails[16 of 40; G test (df � 1), G � 7.52, n � 124; Bonferronicorrected, P � 0.05].
Interspecific competition
Some ant species establish absolute territories by excludingnearly all other ants (Hölldobler and Wilson 1990). How-ever, this was not the case in the large territories of C. gigasthat interacted in various ways with sympatric ant species1
(for a complete list of species, see Pfeiffer 1997).
A series of smaller, night-active, nondominant species(including two species of Camponotus and one ofCrematogaster) were more or less tolerated on Camponotusgigas trails and even at (arboreal) baits. Another group(mostly Dolichoderinae and several species of Polyrhachis,especially P. ypsilon) used these trails only during daytimeand replaced C. gigas inside its arboreal territory. A thirdgroup of ants, including large Ponerinae, interacted with C.gigas on the ground mostly by scramble competition; onlysome species of tiny Myrmicinae monopolized resources. Alast group of (night-active) sympatric Formicinae (mostlyCamponotus species), however, had arboreal territoriesthat were defended against C. gigas, and vice versa.
One of these Camponotini, Camponotus festinus, wasvery abundant in our field site, established several coloniesthere (see Fig. 1), and even settled at deserted nest sites ofC. gigas (n � 2). Interactions with C. gigas differed in dis-tinct strata of the forest. At 10 of 300 ground-level baits weobserved both species feeding simultaneously, thus display-ing scramble competition. However, when we presentedsimilar baits (n � 9) on bamboo trails 1.50m in heightbetween neighboring territories, both species changed to“interference competition.” Following mass recruitments ofboth sides, the ants fought violently at the baits, causingseveral deaths. During these interspecific combats we neverobserved ritual fighting behavior; in most cases the ants (ofall castes) attacked each other with their mandibles but alsoused their poison gland and sprayed acid.
In several long-term experiments, we provoked fightsof C. gigas with neighboring colonies of C. festinus. Bybaiting with infusion systems that dispensed only smalldrops of sugar water, we avoided mass recruitments andtherefore large battles between the ants. Usually several C.gigas majors occupied the bait, but workers of C. festinuswere sitting below the bamboo trail or in the vegetationabove it and attacked the larger C. gigas from there. Most ofthe few major workers of C. gigas that guarded the baits lostparts of their antennae and legs within a few nights. Thissituation continued over days: C. gigas dominated at thebait; however, C. festinus controlled the trail to it, thuspreventing C. gigas from larger exploitation. The “pin-prick” strategy of C. festinus succeeded in the end, andC. gigas had to retreat from all baits. However, even afterthe baits were removed, sentries of both sides were found atthe borders for a mean of 31.4 days (n � 12, SD � 36.1); thisresult showed that not only the baits, but also the territoryborder, were guarded by them. Similar results were alsoobtained with two other sympatric Camponotus species (seeFig. 1).
Discussion
Lumsden and Hölldobler (1983) claimed, for a detailedanalysis of the territorial strategy of ants, the examinationof (1) the distribution of resources within the territory, (2)the territorial “architecture,” (3) the social organization ofits defense, and (4) its spatiotemporal structure.
1 It must be mentioned that majors of all these species are smaller thanmost minors of C. gigas
Table 5. Influence of trail marking on the dominance of C. gigas inintraspecific confrontations of similar castes
Minors (n � 98) Majors (n � 68)
Own trail Alien trail Own trail Alien trail
Number of ants 50 48 34 34
Dominant 34.0%a 14.6%a 20.6% 17.6%Not dominant 66.0%a 85.4%a 79.4% 82.4%
Jump off the trail 12.0%b 31.3%b 23.5% 44.1%No jump 88.0%b 68.8%b 76.5% 55.9%
We noted whether the ants were dominant in the fights and whetherthey jumped off the trail during combatFrequencies of behavior shown on own and alien trails were comparedin G tests with Bonferroni correction; significant differences aremarked with lettersa G test (df � 1), G � 5.01, n � 98; Bonferroni corrected, P � 0.05b G test (df � 1), G � 5.39, n � 98; Bonferroni corrected, P � 0.05

83
Resources used by C. gigas are randomly distributed:invertebrates, bird droppings, and cadavers, on which C.gigas feeds occasionally, are widely dispersed within therain forest, as well as groups of trophobiotic Homoptera(Pfeiffer 1997). Territories of C. gigas colonies had clear-cutborders that were defended even in absence of a baitagainst intra- and interspecific enemies. However, this de-fense was restricted to the arboreal parts of the territory andtook place at “bottlenecks” that gave the opportunity toguard them with a few majors, e.g., the base of the trees. Onthe ground, C. gigas guarded only the immediate surround-ing of their nest entrances, and resources were used inscramble competition. Similar findings were reported byJackson (1984) from Cameroon, who found exclusive terri-tories only in tree-dwelling ant species; ground-dwellingants had overlapping territories because of the lack of“bridgeheads” that could be easily protected. Partly, thisfinding may also reflect problems with territorial marking,which is much easier on arboreal trails than on large groundareas. Our data show that in C. gigas territorial markingsupported the dominance of territory owners.
In Kinabalu Park, several abundant species defendedtheir mutually exclusive, arboreal territories during thenight: Camponotus gigas, C. festinus, Camponotus 64,Camponotus 75, and Oecophylla smaragdina. However, atleast C. gigas tolerated several nondominant species evenon its trunk trails. The three-dimensional territories ofthe giant ants extended over wide areas and are among thelargest we know (Hölldobler and Wilson 1990). Thepolydomic nesting structure of C. gigas intensified territo-rial control. Majors were concentrated in specialized “bar-rack nests” at the endangered territorial borders and in thecentral nests of the colony to protect the queen, brood, andfood supplies. Majors in central nests may have a secondfunction as food storage as is shown, e.g., from majors ofColobopsis nipponicus (Hasegawa 1993). In “minor” neststhat lay within the territory near foraging trees (Pfeiffer andLinsenmair 2000) the percentage of majors was low. Duringterritorial conflicts, high numbers of majors could easily bemobilized by long-range recruitment from central nests tothe borders (as we have already shown by computer countsfor minors during foraging; see Pfeiffer and Linsenmair1998). Compared with a random arrangement of majors atall nests, this strategy allowed colonies to keep a lesserinvestment for producing major workers. Border defense asa primary task of the majors is also known from other antspecies, e.g., Pheidole dentata, Pheidole titanis, or Aztecatrigona (Wilson 1976; Feener 1988; Adams 1994).
C. gigas showed two different strategies to defend itsterritory: (1) ritual combat during intraspecific competitionand (2) “true,” violent fights in interaction with other antspecies. Ritualization seems to have evolved in many antspecies because the lack of a fixed territory can lead tofrequent confrontations of workers from neighboring colo-nies. These species have overlapping territories, foragers ofdifferent colonies that meet in the field perform short dis-plays of ritual aggression behavior; this is the case in M.mimicus (Hölldobler 1976), in the Australian Iridomyrmexpurpureus (Ettershank and Ettershank 1982), in Messor
aciculatus (Yamaguchi 1995), or in the interspecific compe-tition of Polyrhachis laboriosa and Camponotus brutus(Mercier and Dejean 1996). In I. purpureus and M.aciculatus, minors perform front leg boxing; the behavioralpatterns resembled those of C. gigas, but periods of fightingare only 12s or up to 1min, respectively.
In contrast to all other species, in C. gigas ritual fightswere carried out merely by major workers that met regu-larly at fixed bridge points to defend the borders ofnonoverlapping territories. These majors performed an ex-treme form of site fidelity. Up to now, ortstreue has only bereported in connection with foraging areas (Hölldobler andWilson 1990); for C. gigas, however, we use this term todescribe the fidelity of majors to their posts and tournamentplaces.
In the tournaments of M. mimicus, Lumsden andHölldobler (1983) distinguished between two models thatwould allow the opposing colonies to assess each other’sstrength: head counting and cast-polling (Hölldobler 1984).Head counting could be measured most easily in the arbo-real fights of C. gigas, as fighting majors formed pairs alongthe trails during combat. Additional majors could pass thesefighters and reach the nest of their opponents, thus takingadvantage of their local majority. Cast-polling was alsoeasy: smaller majors had less strength and were tired withina shorter time; they had to be replaced more frequently. Asmost of the combats were carried out by only a few majors(especially when the “hot phase” was over), it seems thatcast-polling was enough to determine the strength of acolony. This hypothesis is supported by the fact that minorshad a strong tendency to withdraw from prolonged agonis-tic encounters. One strong ant could stop a number of oppo-nents when it blocked the narrow arboreal trails. It is likelythat, for energetic reasons, the strongest majors could beproduced by large colonies only.
All combats that we observed were solved by a strategyof de-escalation. Although in Myrmecocystus mimicus(Hölldobler 1976) or I. purpureus (Ettershank andEttershank 1982) hundreds of ants are involved in gradu-ally growing ritual tournaments, in C. gigas the numberof fighters was reduced to a handful of majors within acouple of days, while breaks between the single fights wereprolonged. Not only was the method of the combat ritual-ized, in the end the fight itself involved representatives,possibly because its arboreal territory could be protectedmost effectively by a few strong majors. In the rain forest,it seems to be more appropriate to guard certain bridge-heads of a fixed territory over a long time, whereas in adesert environment a ritual fighting strategy needs moreparticipants.
Davidson (1997) stressed the importance ofcarbohydrate-rich diets (e.g., honeydew) for ant protectionof (absolute) territories. From our point of view, high di-etary ratios of CHO:protein seem to be especially impor-tant for the development of a ritual competition patternthat results in purely energetic costs. The ritualization ofaggressive behavior in C. gigas may be based on a cost–benefit calculation. When energy is at low cost, nitrogen (asan essential factor in the production of a large worker force)

84
is a minimum factor. Considering the enormous territoriesof C. gigas and its relatively small colony size, the costs fordefending the territory in nonritual fights may be too high.The very large majors are especially valuable and are, ac-cording to our observations, in many situations timid ratherthan aggressive.
Our study points to a series of long-term ritual conflictsof attrition between small groups of majors with which acolony of C. gigas is faced at many border sections of itsterritory. Only if the ownership of the territory was unclear,as at the start of our first experiment, did territorial conflictsresult in major warfare. In contrast to our observations, Tho(1981), D. Jones, and C. Brühl (personal communication),reported intraspecific warfare in which majors killed eachother in higher numbers. Why fighting was nonritual inthese cases we can only guess: perhaps the strength of thecolonies differed greatly and the territories were not only tobe maintained but enlarged. In our study area, at least onone occasion, a founding colony of C. gigas seemed to havebeen eliminated by the old owners of the territory, but wedid not observe this directly.
Interspecific conflicts, however, resulted in nonritualcombats, which shows that ritual fights are only possible ifboth sides have same risks. The huge majors of C. gigaswere tough fighters, but not very quick; even much smallerspecies were able to hurt them seriously by attacking theirlegs or antennae. Although at first dominant in most of theconflicts, in the course of time they could be overcomeby pinprick strategies. For smaller ants (e.g., otherCamponotus species, or minors of C. gigas), the risk ofbeing injured in a ritual fight was high, and the chance towin it was negligible. Thus, minors withdrew from mostintraspecific fights. On the other hand, the potential loss ofbiomass in a violent fight is lower in smaller ant species,compared to the risk of C. gigas, especially when smallerants apply a guerilla tactic. This relationship may explainwhy interspecies combats were nonritual.
Another access to this problem is given by Lanchester’s(1916) theory of combat (see Franks and Partridge 1993, fordetails) that concisely presented two models relevant also incombats of ants: the linear law and the square law. Initiallyused to calculate losses in nonritual combats, they are alsosuited to discuss the differences of intra- and interspecificconflicts in C. gigas. The linear law will apply if the battle isa series of one-to-one conflicts with excess individuals wait-ing for a free opponent, and it predicts that a few goodfighters are better than many poor ones. This is the case inthe intraspecific ritual duels of C. gigas in which some indi-viduals with massive individual fighting value can decide thebattle. If, however, all individuals are equally vulnerable toattacks from every individual on the opposing side, lossesare proportional to the number of the opposing side andmany poor fighters should succeed over a small number ofgood ones. This scenario happened in interspecific fightswhen several majors of smaller Camponotus species at-tacked one C. gigas in a violent “freestyle” manner, thusenlarging their chance to win. The same was true whennumerous minors of C. gigas attacked alien ants on theirtrails.
Despite the differences in tactics between ritual andnonritual fighting, the result was the same in both cases. Theterritorial interactions were so intensive that they resultedin a no-ants zone between the rivaling colonies. Thus, bothmechanisms seem to be equally effective within their eco-logical context by providing clear territorial borders.
Acknowledgments We are grateful to Sabah Parks for assistance andhelp that allowed this study to be conducted. We especially thankDatuk Dr. Lamri Ali, Mr. Eric Wong, Dr. Jamili Nais, and Mr. KasitahKarim for various kinds of support. We thank Antonia Uecker,Sabine Hussmann, and Matthias Dolek for help in the field work, Dr.Carsten Brühl for the determination of C. festinus, Dr. AndreasFloren, Prof. Dr. A. Buschinger, and three anonymous referees forhelpful comments on the manuscript, and Prof. Dr. Herbert Vogt forhelp with the statistics. This study was funded by the DAAD(Deutscher Akademischer Austauschdienst). We also thank the“Deutsche Forschungsgemeinschaft” (DFG) for granting the infra-structure for our activities within the framework of the “DFGSchwerpunktprogramm Mechanismen der Aufrechterhaltungtropischer Diversität.”
References
Adams ES (1994) Territory defense by the ant Azteca trigona:maintainance of an arboreal ant mosaic. Oecologia (Berl) 97:202–208
Brühl CA, Gunsalam G, Linsenmair KE (1998) Stratification of ants(Hymenoptera, Formicidae) in a primary rain forest in Sabah,Borneo. J Trop Ecol 14:285–297
Carlin NF, Hölldobler B (1983) Nestmate and kin recognition ininterspecific mixed colonies of ants. Science 222:1027–1029
Czechowski W (1984) Tournaments and raids in Lasius niger (L.)(Hymenoptera, Formicidae). Ann Zool (Wars) 38:81–91
Davidson DW (1997) The role of resource imbalances in the evolu-tionary ecology of tropical arboreal ants. Biol J Linn Soc 61:153–181
Dobrzanski J (1966) Contribution to the ethology of Leptothoraxacervorum (Hymenoptera: Formicidae). Acta Biol Exp (Wars)26:71–78
Ettershank G, Ettershank JA (1982) Ritualised fighting in the meat antIridomyrmex purpureus (Smith) (Hymenoptera: Formicidae). J AustEntomol Soc 21:97–102
Feener DH Jr (1988) Effects of parasites on foraging and defensebehavior of a termitophagous ant, Pheidole titanis Wheeler (Hy-menoptera: Formicidae). Behav Ecol Soc 22:421–427
Franks NR, Partridge LW (1993) Lanchester battles and the evolutionof combat in ants. Anim Behav 45:197–199
Hasegawa E (1993) Caste specialisation in food storage in the dimor-phic ant Colobopsis nipponicus (Wheeler). Insectes Soc 40:261–271
Hölldobler B (1976) Tournaments and slavery in a desert ant. Science192:912–914
Hölldobler B (1981) Foraging and spatiotemporal territories in thehoney ant Myrmecocystus mimicus Wheeler (Hymenoptera:Formicidae). Behav Ecol Soc 9:301–314
Hölldobler B (1982) Interference strategy of Iridomyrmex pruinosum(Hymenoptera: Formicidae) during foraging. Oecologia (Berl)52:208–213
Hölldobler B (1983) Territorial behavior in the green tree ant(Oecophylla smaragdina). Biotropica 15:241–250
Hölldobler B (1984) Konkurrenzverhalten und Territorialität inAmeisenpopulationen. In: Eisner T, Hölldobler B, Lindauer M (eds)Chemische Ökologie Territorialität Gegenseitige Verständigung.Fischer, New York, pp 25–70
Hölldobler B, Lumsden CJ (1980) Territorial strategies in antsOecophylla longinoda, Pogonomyrmex, Mymecocystus mimicus. Sci-ence 210:732–739
Hölldobler B, Wilson EO (1990) The ants. Belknap Press, HarvardUniversity Press, Cambridge

85
Jackson DA (1984) Ant distribution patterns in a Cameroonian cocoaplantation: investigation of the ant mosaic hypothesis. Oecologia(Berl) 62:318–324
Lanchester FW (1916) Aircraft in warfare: the dawn of the fourth arm.Constable, London
Le Moli F, Parmigiani S (1982) Intraspecific combat in the red woodant (Formica lugubris, Zett.). Aggress Behav 8:145–148
Le Moli F, Mori A, Parmigiani S (1982) Agonistic behaviour ofFormica rufa L. (Hymenoptera: Formicidae). Monit Zool ital (NS)16:325–331
Lumsden CJ, Hölldobler B (1983) Ritualized combat and intercolonycommunication in ants. J Theor Biol 100:81–98
Mabelis AA (1979) Wood ant wars: the relationship between aggres-sion and predation in the red wood ant (Formica polyctena Forst.).Neth J Zool 29:451–620
Mercier JL, Dejean A (1996) Ritualized behavior during competitionfor food between two Formicinae. Insectes Soc 43:17–29
Mercier JL, Lenoir A, Dejean A (1997) Ritualized versus aggressivebehaviours displayed by Polyrhachis laboriosa (F. Smith) duringinterspecific competition. Behav Proc 41:39–50
Pfeiffer M (1997) Sozialstruktur und Verhaltensökologie vonRiesenameisen Camponotus gigas Latreille 1802 im RegenwaldMalaysias auf Borneo. Wissenschaft und Technik Verlag, Berlin
Pfeiffer M, Linsenmair KE (1997) Reproductive synchronization in thetropics: the circa-semiannual rhythm in the nuptial flight of the giantant Camponotus gigas Latreille (Hym./Form.). Ecotropica 3:21–32
Pfeiffer M, Linsenmair KE (1998) Polydomy and the organization offoraging in a colony of the Malaysian giant ant Camponotus gigas(Hym./Form.). Oecologia (Berl) 117:579–590
Pfeiffer M, Linsenmair KE (2000) Contributions to the life historyof the Malaysian giant ant Camponotus gigas (Hymenoptera/Formicidae). Insectes Soc 47:123–132
Tho YP (1981) The giant forest ant Camponotus gigas, Malaysia. NatMalays 6:32–35
Wilson EO (1976) The organization of colony defense in the antPheidole dentata Mayr (Hymenoptera: Formicidae). Behav Ecol Soc1:63–81
Yamaguchi T (1995) Intraspecific competition through food robbing inthe harvester ant, Messor aciculatus (Fr. Smith) and its consequenceson colony survival. Insectes Soc 42:89–101
Copyright © 2022 FDOKUMEN




















