
Delayed-type hypersensitivity response in an isogenic murine model of paracoccidioidomycosis
10
Mycopathologia 126: 137-146, 1994. © 1994 KluwerAcademic Publishers. Printedin the Netherlands. Delayed-type hypersensitivity response in an isogenic murine model of paracoccidioidomycosis Raquel dos Anjos Fazioli, ~'2 Lticia Mary Singer-Vermes, 1 Suely Sanae Kashino, 1 Eva Burger, 1 Marcello Fabiano De Franco, 3 Maura Moscardi-Bacchi 3 & Vera Lticia Garcia Calich ~ t Departamento de Imunologia, Instituto de Ci6ncias Biomddicas, Universidade de S6o Paulo, S~o Paulo, Brasil; 2Sefdo de Imunologia, Instituto Adolfo Lutz, Sgo Paulo, Brasil; 3Departamento de Patologia, Faculdade de Medicina de Botucatu, UNESP, Botucatu, S~o Paulo, Brasil Received 13 October 1993; accepted in revisedform 25 January 1994 Abstract. The specific delayed-type hypersensitivity (DTH) response was evaluated in resistant (A/SN) and susceptible (B10.A) mice intraperitoneally infected with yeasts from a virulent (Pbl8) or from a non-virulent (Pb265) Paracoccidioides brasiliensis isolates. Both strains of mice were footpad challenged with homologous antigens. Pb18 infected A/SN mice developed an evident and persistent DTH response late in the course of the disease (90th day on) whereas B10.A animals mounted a discrete and ephemeral DTH response at the 14th day post-infection. A/SN mice infected with Pb265 developed cellular immune responses whereas B10.A mice were almost always anergic. Histological analysis of the footpads of infected mice at 48 hours after challenge showed a mixed infiltrate consisting of predominantly mononu- clear cells. Previous infection of resistant and susceptible mice with Pb18 did not alter their DTH responses against heterologous unrelated antigens (sheep red blood cells and dinitrofluorobenzene) indicating that the observed cellular anergy was antigen-specific. When fungal related antigens (candidin and histoplasmin) were tested in resistant mice, absence of cross-reactivity was noted. Thus, specific DTH responses against P. brasiliensis depend on both the host's genetically determined resistance and the virulence of the fungal isolate. Key words: Cellular immune response, Delayed-type hypersensitivity, Isogenic murine model, P. brasili- ensis Abbreviations: DTH, delayed-type hypersensitivity; DNFB, dinitrofluorobenzene; FN18, Fava Netto's antigen obtained from isolate Pb18; FN265, Fava Netto's antigen obtained from isolate Pb265; SRBC, sheep red blood cells Introduction Paracoccidioidomycosis (PCM) is a chronic gran- ulomatous disease caused by the dimorphic fungus Paracoccidioides brasiliensis [1]. As in other deep mycosis, cell-mediated immunity (CMI) has been ascribed as the most important host defense mechanism against this fungus [2- 4]. Usually progressive or disseminated forms of the disease are associated with depressed cellular immune responses and high levels of specific anti- bodies [5-7]. Our previous studies established an isogenic murine model of PCM. It was shown that A/SN and B10.A mice are respectively very resistant and susceptible animals to intraperitoneal (i.p.) infection with a virulent P. brasiliensis isolate (Pb18) [8] and that resistance was linked to a
-
Upload
andrelemos -
Category
Documents
-
view
0 -
download
0
Transcript of Delayed-type hypersensitivity response in an isogenic murine model of paracoccidioidomycosis

Mycopathologia 126: 137-146, 1994. © 1994 Kluwer Academic Publishers. Printed in the Netherlands.
Delayed-type hypersensitivity response in an isogenic murine model of paracoccidioidomycosis
Raquel dos Anjos Fazioli, ~'2 Lticia Mary Singer-Vermes, 1 Suely Sanae Kashino, 1 Eva Burger, 1 Marcello Fabiano De Franco, 3 Maura Moscardi-Bacchi 3 & Vera Lticia Garcia Calich ~ t Departamento de Imunologia, Instituto de Ci6ncias Biomddicas, Universidade de S6o Paulo, S~o Paulo, Brasil; 2Sefdo de Imunologia, Instituto Adolfo Lutz, Sgo Paulo, Brasil; 3 Departamento de Patologia, Faculdade de Medicina de Botucatu, UNESP, Botucatu, S~o Paulo, Brasil
Received 13 October 1993; accepted in revised form 25 January 1994
Abstract. The specific delayed-type hypersensitivity (DTH) response was evaluated in resistant (A/SN) and susceptible (B10.A) mice intraperitoneally infected with yeasts from a virulent (Pbl8) or from a non-virulent (Pb265) Paracoccidioides brasiliensis isolates. Both strains of mice were footpad challenged with homologous antigens. Pb18 infected A/SN mice developed an evident and persistent DTH response late in the course of the disease (90th day on) whereas B10.A animals mounted a discrete and ephemeral DTH response at the 14th day post-infection. A/SN mice infected with Pb265 developed cellular immune responses whereas B10.A mice were almost always anergic. Histological analysis of the footpads of infected mice at 48 hours after challenge showed a mixed infiltrate consisting of predominantly mononu- clear cells. Previous infection of resistant and susceptible mice with Pb18 did not alter their DTH responses against heterologous unrelated antigens (sheep red blood cells and dinitrofluorobenzene) indicating that the observed cellular anergy was antigen-specific. When fungal related antigens (candidin and histoplasmin) were tested in resistant mice, absence of cross-reactivity was noted. Thus, specific DTH responses against P. brasiliensis depend on both the host's genetically determined resistance and the virulence of the fungal isolate.
Key words: Cellular immune response, Delayed-type hypersensitivity, Isogenic murine model, P. brasili- ensis
Abbreviations: DTH, delayed-type hypersensitivity; DNFB, dinitrofluorobenzene; FN18, Fava Netto's antigen obtained from isolate Pb18; FN265, Fava Netto's antigen obtained from isolate Pb265; SRBC, sheep red blood cells
Introduction
Paracoccidioidomycosis (PCM) is a chronic gran- ulomatous disease caused by the dimorphic fungus Paracoccidioides brasiliensis [1]. As in other deep mycosis, cell-mediated immunity (CMI) has been ascribed as the most important host defense mechanism against this fungus [2- 4]. Usually progressive or disseminated forms of
the disease are associated with depressed cellular immune responses and high levels of specific anti- bodies [5-7].
Our previous studies established an isogenic murine model of PCM. It was shown that A/SN and B10.A mice are respectively very resistant and susceptible animals to intraperitoneal (i.p.) infection with a virulent P. brasiliensis isolate (Pb18) [8] and that resistance was linked to a

138
dominant autosomal gene [9]. Using susceptible mice we determined the virulence and immuno- genicity of several P. brasiliensis isolates and could classify isolate Pb18 as highly virulent and Pb265 isolate as non-virulent [10, 11].
Comparative studies using genetically defined hosts and microorganisms with known degrees of virulence are excellent tools to better understand the immunological basis for the resistance to vari- ous infectious agents. With this concept in mind we evaluated the pattern of DTH responses in A/SN and B10.A mice at various time intervals after i.p. infection with Pb18 or Pb265 isolates. The cellular immune responses to homologous and heterologous antigens were also investigated. Our results showed that P. brasiliensis infection determines an antigen-specific cellular immuno- depression and that both host and fungal factors affect the pattern of DTH responses.
Methods
Animals. Eleven week old female A/SN and B10.A mice, originally purchased from Jackson Laboratory (Bar Harbor, Maine), were used in all experiments. The animals were housed in the Department of Immunology, University of S~o Paulo and were given food and acidified-chlori- nated water ad libitum.
P. brasiliensis isolates and growth conditions. Two human isolates of P. brasiliensis were studied; Pbl8 (virulent) and Pb265 (non-virulent) [10, 11], obtained from our fungal culture collection. Both isolates were maintained at 37 °C in their yeast phase in semi-solid Fava Netto's medium [12] and used at the 7th day of growth culture. Virulence of Pb18 was controlled by means of LD 50% determination and at any evidence of virulence loss, the isolate was reisolated from susceptible mice.
Antigens. Two P. brasiliensis antigen prepara- tions obtained after chemical delipidation of the yeast cells, according to the Fava Netto's method
[12] were used. These preparations derived from isolates Pb18 and Pb265 were designated FN18 and FN265, respectively. FN18 and FN265 had protein concentrations of 310 and 332 ~zg m1-1, respectively; their carbohydrate content were respectively 153.4 and 49.5~gm1-1. Candida albicans or Histoplasma capsulatum antigens (kindly supplied by Dr Celeste Fava Netto and Celina Arruda) were also prepared according to Fava Netto's method [12]. The protein and the polysaccharide content of the C. albicans antigen (FNCa) were 322 txg m1-1 and 506.5 rag m1-1, respectively. The H. capsulatum antigen (FNHc) had a protein concentration of 315 ~g ml -~ and a carbohydrate content of 112.4 p~gm1-1. Protein and carbohydrate concentrations were always de- termined by the Lowry [13] and the a-naftol [14] methods, respectively.
Infection of mice. P. brasiliensis yeast cell suspen- sions, from 7 days-old cultures, were washed three times in phosphate-buffered saline (PBS), pH 7.2, and concentrations were adjusted to 10 × 106cells/ml based on hemocytometer co- unts. Viability was determined with the Janus Green vital stain [15] and was always higher than 85%. Animals were inoculated i.p. with 5.0 × 10 6 yeast cells contained in 0.5 ml of PBS. Cohorts of mice were evaluated regarding his- tology of the footpads, DTH response to homolo- gous and to heterologous antigens at various post infection times and were subsequently sacrificed.
Footpad tests to fungal antigens. DTH responses against P. brasiliensis antigens were evaluated during the evolution of the infectious process by the footpad test. The left hind footpad was in- jected subcutaneously with 25 ~1 of the antigen solution, homologous to the isolate used to infect the animals. The swelling was evaluated measur- ing the footpad thickness just before and 24 h after injection of antigen using a dial caliper (precision 0.01 ram, Mitutoyo, Tokyo, Japan) and the increase in thickness was calculated. The optimal concentrations and conditions for DTH assays determined after preliminary experiments

[16], were employed. The same procedure was used to evaluate the DTH response to C. albicans
and H. capsuIatum antigens in P. brasiliensis in- fected animals.
Histology. Histological analysis of the footpad of normal or Pb18 infected mice challenged with the homologous antigen (FN18) was done at 24 and 48 hours after injection to ascertain the presence and the intensity of the cellular infiltrate. The left hind footpads of B10.A and A/SN mice at different time intervals were removed, fixed in 10% formalin, embedded in paraffin, sectioned and stained with hematoxylin-eosin (HE). The cellular influx was characterized according to the cell type predominance and the intensity of the reaction was registered.
D TH reactions to unrelated antigens. DTH reac- tions to 2,4-dinitro-l-fluorobenzene (DNFB) were evaluated in A/SN and B10.A mice at 7, 30, 60, 90 and 120 days post infection. The contact hypersensitivity test was assayed in mice sensit- ized with DNFB (Merck, Germany) according to a previously described method [17-19] with minor modifications. Briefly, seven days before challenge, 100 ~1 of a 0.5% DNFB solution in acetone/olive oil was applied on the abdomen of the animals and the challenge was performed applying 20 ~1 of the same solution on the back side of the left ear. The right ear received only acetone/olive oil as a control for irritation caused by the solvents. Ear thickness of each animal was measured just before DNFB application and 24 h later with a dial caliper (precision 0.01 mm, Mitu- toyo, Tokyo, Japan), and the difference in ear thickness was calculated. Both, infected and sa- line control groups were assayed under the same conditions. As additional controls, infected and non infected mice were challenged, but not pre- viously sensitized with DNFB. DTH responses to sheep red blood cells (SRBC) were also evaluated in normal and Pb18 infected mice which were i.v. sensitized with 5 x 105 SRBC in 0.5 ml of PBS. Four days later, mice were challenged in the left hind footpad with 5 x 108 SRBC suspended in
139
50 ~1 of PBS [20]. Footpad swelling was measured immediately before and 24 hours later. Normal non-sensitized but only footpad challenged mice were included as another control group. The dif- ferences in the footpad thicknesses, before and 24 hours after antigen injection, were expressed in ram.
Statistical analysis. Means, standard errors of the means (SEM) and analysis of variance followed by the Tukey or the Dunnett test were used to analyze the data [21]. Significance was deter- mined at the p < 0.05 level.
Results
D T H responses in susceptible and resistant mice infected with a highly virulent or an avirulent iso- late o f P. brasiliensis. Preliminary investigations demonstrated that, among other antigen prepara- tions, Fava Netto antigen was the most reliable since it did not induce inflammatory responses in normal animals and evoked the highest DTH reactions in infected mice. It was also observed that the most adequate protein concentrations of Pb18 and Pb265 antigens were 310 and 332 ~zg m1-1, respectively and that the best time to determine the footpad swelling was twenty four hours after antigen challenge [16].
A/SN and B10.A mice i.p. infected with 5 X 10 6 Pb18 or Pb265 cells were footpad tested with homologous Fava Netto's antigen at selected intervals over a period of 150 days. Control non infected A/SN and B10.A mice were also assayed by the footpad reaction using the same antigen preparations. The results of DTH responses of Pb18 and Pb265 infected mice, during the evol- ution of the disease are shown in Fig. 1. Data obtained in non infected control mice are also depicted in this figure. Comparing the mean foot- pad swelling of infected mice, at the different post infection periods, with their control counter- parts, we observed that: (a) the non-specific in- flammatory reaction induced by FN18 and FN265 in resistant and susceptible normal animals is
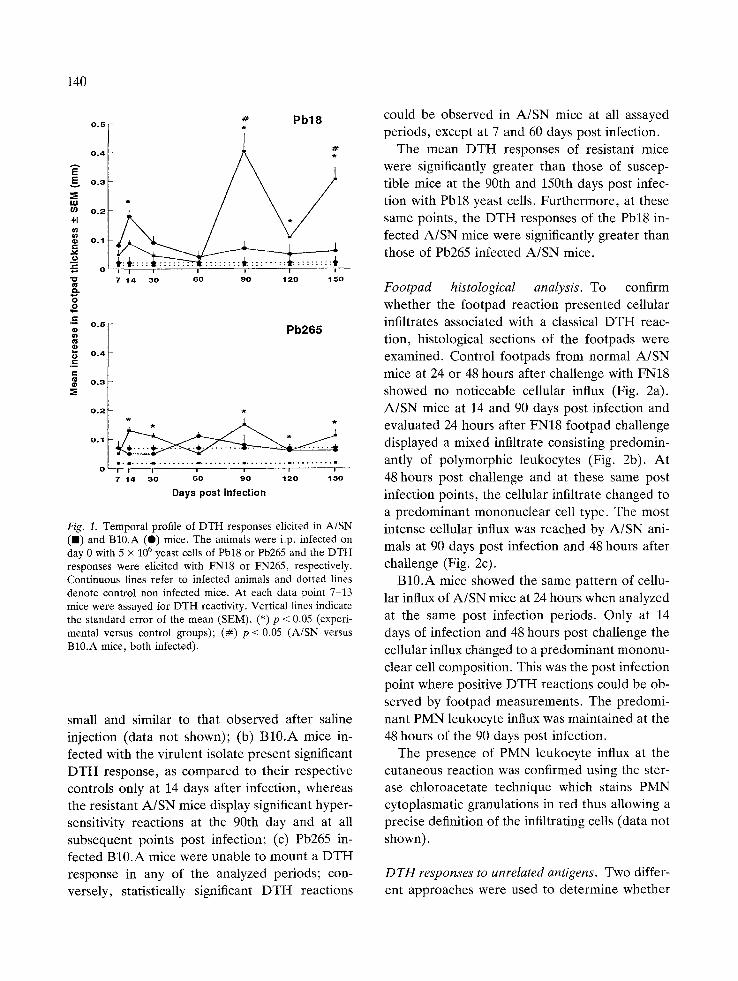
140
E
U J m
+1 uJ
e -
.2
e~
O
2 .=
I1)
o .E
I11
O . S
0 . 4
0 . 3
0 . 2
0 . 1
o i ! i
7 1 4 3 0
# P b 1 8 w
i i t i
6 0 9 0 1 2 0 1 5 0
0 . 5
0 . 4
0 . 3
0 . 2
0 . 1
0
P b 2 6 5
~e
~ ! i z z i 1 - -
1 4 3 0 6 0 9 0 1 2 0 1 5 0
Days post infection
Fig. 1. Temporal profile of DTH responses elicited in A/SN (B) and B10.A (e) mice. The animals were i.p. infected on day 0 with 5 x 106 yeast cells of Pbl8 or Pb265 and the DTH responses were elicited with FN18 or FN265, respectively. Continuous lines refer to infected animals and dotted lines denote control non infected mice. At each data point 7-13 mice were assayed for DTH reactivity. Vertical lines indicate the standard error of the mean (SEM). (*) p < 0.05 (experi- mental versus control groups); (#) p < 0.05 (A/SN versus B10.A mice, both infected).
small and similar to that observed after saline injection (data not shown); (b) B10.A mice in- fected with the virulent isolate present significant DTH response, as compared to their respective controls only at 14 days after infection, whereas the resistant A/SN mice display significant hyper- sensitivity reactions at the 90th day and at all subsequent points post infection; (c) Pb265 in- fected B10.A mice were unable to mount a DTH response in any of the analyzed periods; con- versely, statistically significant DTH reactions
could be observed in A/SN mice at all assayed periods, except at 7 and 60 days post infection.
The mean DTH responses of resistant mice were significantly greater than those of suscep- tible mice at the 90th and 150th days post infec- tion with Pb18 yeast cells. Furthermore, at these same points, the DTH responses of the Pb18 in- fected A/SN mice were significantly greater than those of Pb265 infected A/SN mice.
Footpad histological analysis. To confirm whether the footpad reaction presented cellular infiltrates associated with a classical DTH reac- tion, histological sections of the footpads were examined. Control footpads from normal A/SN mice at 24 or 48 hours after challenge with FN18 showed no noticeable cellular influx (Fig. 2a). A/SN mice at 14 and 90 days post infection and evaluated 24 hours after FN18 footpad challenge displayed a mixed infiltrate consisting predomin- antly of polymorphic leukocytes (Fig. 2b). At 48 hours post challenge and at these same post infection points, the cellular infiltrate changed to a predominant mononuclear cell type. The most intense cellular influx was reached by A/SN ani- mals at 90 days post infection and 48 hours after challenge (Fig. 2c).
B10.A mice showed the same pattern of cellu- lar influx of A/SN mice at 24 hours when analyzed at the same post infection periods. Only at 14 days of infection and 48 hours post challenge the cellular influx changed to a predominant mononu- clear cell composition. This was the post infection point where positive DTH reactions could be ob- served by footpad measurements. The predomi- nant PMN leukocyte influx was maintained at the 48 hours of the 90 days post infection.
The presence of PMN leukocyte influx at the cutaneous reaction was confirmed using the ster- ase chloroacetate technique which stains PMN cytoplasmatic granulations in red thus allowing a precise definition of the infiltrating cells (data not shown).
DTH responses to unrelated antigens. Two differ- ent approaches were used to determine whether

141
Fig. 2. Histological sections of footpads from: (a) normal A/SN mice, obtained 24 hours after footpad challenge with FN18 (HE x 160); (b) infected A/SN mice, obtained at 90 days post infection and 24 h after footpad challenge with FN18 (HE x 400); and (c) infected A/SN mice taken at 90 days post infection and 48 h after footpad challenge with FN18 (HE x 400).

142
g
1.1,1 u}
+1 14
O
P
P
|
e -
IN
0.5
0.4
0.3
0.2
0.1
A / $ N
I- - - i , - - - - 1 - - - 4 - - - 4
I I I I I
0.5
0.4
0.3
7 3O
B 1 0 . A
60 90 120
0,2 /
0.1 t ~- _ _ ~ - _ _ _ , _ _ _ _ ~ _ _ _ .~
/ 0 I I t i
7 30 60 90 120
Days after Infection
Fig..3. Contact hypersensitivity reactions to DNFB in normal ( I ) and infected (0 ) A/SN and B10.A mice. The animals were infected with 5 × 106 yeast cells of Pbl8. The dotted lines refer to challenged, but not previously DNFB sensitized mice. At each point 6-7 mice were assayed for DNFB re- sponse. Vertical lines indicate the standard error of the mean (SEM).
A E g IS I I I
I t l
4 t
= @
O
e
@
e ,
@
~ . . A / S N
1"
11,8.
o.(1-
0.2"
0 14
1.2-
1
0.(1
0.6-
(1,4-
0
.,-T- :i~: : / ! i '/ / / ',//, _~ :::i:i i l / / ,
/ ! / , ' / / i / / / i//
,7/ i ,} .......... I ::~'!!! / / / , , , I ,-;,/
B 1 0 . A
.............. , ' / / ii::i!ii;i'~i ii:iii~:i~ i / / ~i!i!i~!:i:!~i
%i~i / ! / ii:i]i:iii
)iiiiii!iiiiii? . , , ,~':'!~i:i:~i~ !~i~i~:~!~i!~i! ':':' !~!i:!~!~i~!~i~ ......
1,t
Days after Infection
/ i , l
'//,! ://j ,'//.~ t
/ / / , L/ ,~/'/ ~ /7 / '!/ i ,'//J / l'/i
Fig. 4. DTH responses against SRBC in mice infected with 5 × 106 yeast cells of Pb18 and in normal animals. Three different groups were tested at 14th and 90th days post infec- tion: normal mice only footpad challenged with SRBC ([]); sensitized and footpad challenged normal mice (D); sensitized and footpad challenged infected mice (N). Each bar repre- sents the mean increase in footpad thickness of 10-12 mice. Vertical lines indicate standard error of the mean (SEM). (*) p < 0.05 (experimental versus control normal groups, only footpad challenged).
previous infection with P. brasiliens& influences the ability of mice to develop D T H responses to heterologous antigens: contact hypersensitivity to DNFB and D T H response to SRBC.
Contact hypersensitivity to DNFB was assayed in groups of A/SN and B10.A mice infected with 5 × 106 yeast cells of Pbl8 at various periods after infection. Non infected control animals were equivalently assayed. All data obtained in the non-sensitized and challenged controls were corn-
bined since these animals presented similar in- crease in ear thickness.
Normal as well as infected A/SN and B10.A mice displayed significant contact sensitivity to DNFB, at all assayed data points, as compared to their challenged but not sensitized control counterparts. The intensity of the reactions to DNFB found in infected and non infected A/SN and B10.A mice, both sensitized and challenged with DNFB, were similar at all assayed points, as depicted in Fig. 3. Thus, previous infection did

not alter their ability to elicit response to this unrelated antigen.
The DTH response to SRBC was assayed in A/SN and B10.A mice at the 14th and 90th days post infection. These points were chosen because they represent the highest cutaneous reactivity of B10.A (14th day) and A/SN (90th day) mice to P. brasiliensis antigen (FN18). Both mouse strains were able to mount DTH responses to SRBC, as verified by the significantly higher foot- pad swelling observed in control non infected mice when compared to challenged, but not sensi- tized mice. Infected A/SN and B10.A mice, at both assayed times after infection, displayed DTH responses similar to their control non in- fected counterparts (Fig. 4), showing that pre- vious infection did not alter their ability to de- velop DTH response to SRBC.
Specificity of DTH responses to P. brasiliensis. Pb18 infected A/SN mice were tested with related polysaccharide fungal antigens (FNCa or FNHc) at the 90th day post infection, when they show the most prominent DTH response to the hom- ologous antigen preparation. These animals did not show a positive cutaneous reactivity to either FNCa or FNHc antigens, since their DTH re- sponses did not differ significantly from the nor- mal non infected counterparts (Fig. 5). These re- sults indicate that cross reactivity to related fungal antigens is absent in this mouse model.
D i s c u s s i o n
The association between cellular immune depres- sion and the severity of the disease in patients with PCM was noted by several investigators [3, 4, 6, 7]. These observations suggested that cellular immune response is one of the most im- portant resistance mechanisms operating in P. brasiliensis infected hosts [22, 23]. To gain further insights on the phenomenon, an inbred mouse model has been used in our laboratory to study the correlation between DTH reactivity and resis- tance to P. brasiliensis infection.
143
A
E o.s- E
O.4-
O i-
"0 0 . 3 -
ft.
- - 0 . 2 - O
.o o _g 0 . 1 -
C
Q
' O .C
f J f J
f J
I j f l
C. albicans H. capsulatum
ANTIGENS
#
R brasiliensJs
Fig. 5. Footpad hypersensitivity responses in A/SN mice, 90 days post infection with 5 × 106 Pb18 yeast cells (hatched bars) and in control noninfected animals (empty bars). DTH was assayed using C. albicans, H. capsulatum or P. brasiliensis (Pbl8) derived antigens and footpad swelling were measured 24 hours after challenge. Vertical lines indicate the standard error of the mean (SEM) of groups with 13-22 mice. (*) p < 0.05 (experimental versus control groups); (#) p < 0.05 (FN18 versus heterologous antigens in infected mice).
Our first set of experiments (Fig. 1) demon- strated that susceptible and resistant mice de- veloped totally distinct patterns of DTH re- sponse. B10.A mice showed a single and low intensity peak of cutaneous positivity 2 weeks after Pbl8 infection, behaving as anergic animals at all subsequent post infection assayed periods. Conversely, A/SN mice were able to mount an intense DTH reaction three months after P. brasi- liensis (Pb18) inoculation and a weak cutaneous activity at 2 weeks post infection. The suscepti- bility of B10.A mice to the i.p. route of infection cannot be attributed to their intrinsic inability to mount a DTH response since these animals are able to develop DTH reactivity in the beginning of the Pb18 infection; most probably suscepti- bility might be linked to the development of anergy to fungal antigens during the course of the

144
active disease. In contrast, resistance of A/SN mice is associated to the development of an evi- dent and persistent DTH response in the course of the disease. Comparing the DTH results ob- tained in A/SN mice (Fig. 1) it can be observed that the virulent Pbl8 isolate evokes greater DTH responses than does the non-virulent Pb265 iso- late. B10.A mice however develop anergy in cu- taneous reactivity independently from the in- fecting fungal isolate. These negative responses after cutaneous challenge observed in B10.A mice can be linked to immunosuppressive me- chanisms induced by the virulent isolate and to the weak immunostimulation induced by the non- virulent isolate. Another attractive interpretation is that susceptible hosts mount an immune re- sponse which do not favor cellular immune me- chanisms.
Whatever are the immunoreguiatory mechan- isms involved in the different patterns of DTH responses of A/SN and B10.A mice infected with virulent or non-virulent P. brasiliensis isolates, cellular immunity in our model depends on both fungal and host's intrinsinc factors.
The results herein obtained using susceptible mice parallel the chronic progressive form of the human PCM~ where the progression of the dis- ease is associated to cellular immunodepression. Conversely, the behavior of resistant animals is similar to those of the localized or benign forms of the human disease in which a correlation be- tween a good prognosis and an overt DTH reac- tivity is commonly found. Furthermore, our re- sults are similar to those observed in the pulmonary infection of mice with Coccidioides immitis [24] as well as in outbred animals infected by the i.v. route with Pb18 and Pb265 yeast cells [25].
Fluctuation of cell mediated immune response as observed in the present study (Fig. 1) was also noted by other investigators working either with P. brasiliensis [26] or with H. capsulatum [27] infected animals. This phenomenon has been as- cribed to immunoregulatory mechanisms associ- ated to T dependent specific immunodepression, antibody production or by the presence of circul- ating immune complexes [26, 27].
The footpad swelling observed in P. brasiliensis infected mice challenged with the homologous antigen seems to be a classical DTH reaction, as evidenced by the predominant mononuclear leukocytes influx observed 48 hours after antigen challenge [28].
To further investigate the cell mediated im- munity in murine PCM, two heterologous anti- gens were selected: SRBC and DNFB. Normal as well as P. brasiliensis infected B10.A and A/SN animals were able to mount a cutaneous reactivity to these antigen preparations. Thus, the capacity of B10.A animals to maintain their respon- siveness to SRBC and DNFB, but not to P. brasil- iensis antigens, allows us to state that the immunodepression presented by these animals is antigen-specific. Similar results were obtained in other murine models of paracoccidioidomycosis [29] or cryptococcosis [30]. Injected mice pre- sented depressed DTH responses to homologous antigens but their ability to develop DTH reac- tivity to the heterologous antigen assayed (DNFB or SRBC) was preserved. Conversely, Castafieda et al. [31] observed a non-specific impairment of cell mediated immunity in P. brasiliensis infected BALB/c mice but this phenomenon was observed only late in the disease. Further studies per- formed by Castafieda et al. (32) demonstrated that impairment of lymphocyte proliferation was mediated by T suppressor cells and specific DTH responses were reduced when the animals were pretreated with immune serum. Despite the dis- crepancies between our results and those found by the above cited authors, both models show that severe paracoecidioidomycosis is associated with impairment in cellular immunity.
Cross-reactivity with related fungal antigens is frequently observed in human PCM [33, 34]. In our experimental model, however, the obtained DTH responses were specific to P. brasiliensis since C. albicans and H. capsulatum antigens were not able to induce footpad swelling in in- fected animals. Our results are in agreement with those reported by Jimenez-Finkel and Murphy [29] who observed that CBA/J mice immunized with P. brasiliensis antigens and footpad chal- lenged with H. capsulatum or C. immitis antigen

preparations presented only specific DTH re- sponses. The discrepancies detected in the cu- taneous reactivity of human and murine PCM could be ascribed to a more frequent infection with other fungal species by humans and/or to a more specific repertoire of T cells used by P. brasiliensis infected mice.
All factors considered, our results demon- strated that susceptibility to P. brasiliensis is as- sociated with impaired cellular immunity and re- sistance with the presence of adequate specific DTH reactions. Previous studies showed that sus- ceptible animals present increased counts of vi- able yeasts in various organs, high levels of speci- fic antibodies and impaired macrophage activation as determined by its spreading ability on glass slides and H202 production [35-37]. This kind of immune response is associated with a progressive disease characterized by the presence of numerous and non-organized granulomatous lesions [8, 11]. The same studies demonstrated that resistant mice show a self-healing infection with a limited number of well organized granulo- matous lesions containing few viable yeasts, low specific antibody levels besides an early and sus- tained macrophage activation [8, 11, 35-37]. Taken together these data suggest that resistant mice handle the immune activation to a preferen- tial cellular response while susceptible animals develop a predominant humoral response.
Studies concerning T cell proliferation and major interleukins production by susceptible and resistant animals in the course of murine PCM would allow us to better define the resistance mechanisms operating in murine paracoccidioido- mycosis.
Acknowledgments
We thank Roberto Nilo Pinheiro for taking care of the animals. This work was supported by Fun- dag~o de Amparo h Pesquisa do Estado de Silo Paulo (FAPESP), Conselho Nacional de Pes- quisas (CNPq) e Financiadora de Estudos e Pro- jetos (FINEP).
145
References
1. Restrepo AM. Immune response to Paracoccidioides bra- siliensis in human and animal hosts. Cur Top Med Mycol 1988; 2: 239-77.
2. Brummer E, Restrepo A, Stevens DA, Azzi R, Gomez AM, Hoyos GL, McEwen JG, Cano LE, Bedout C. Mu- rine model of paracoccidioidomycosis. Production of fatal" acute pulmonary or chronic pulmonary and disseminated disease: immunological and pathological observations. J Exp Pathol 1984; 1: 241-55.
3. Mota NGS, Rezkallah-Iwasso MT, Peragoli MTS, Audi RC, Mendes RP, Marconde J, Marques SA, Dillon NL, Franco MF. Correlation between cell-mediated immunity and clinical forms of paracoccidioidomycosis. Trans R Soc Trop Med Hyg 1985; 79: 765-72.
4. Restrepo A, Restrepo M, Restrepo F, Ariztiz~ibal LH, Moncada LH, V61ez H. Immune responses in paracocci- dioidomycosis: A controlled study of 16 patients before and after treatment. Sabouraudia 1978; 16: 151-63.
5. Defaveri J, Rezkallah-Iwasso MT, Franco MF. Experi- mental pulmonary paracoccidioidomycosis in mice: morphology and correlations of lesions with humoral and cellular immune response. Mycopathologia 1982; 77: 3- 11.
6. Mok PWY, Greer DL. Cell-mediated immune responses in patients with paracoccidioidomycosis. Clin Exp Immu- nol 1977; 28: 89-98.
7. Musatti CC, Rezkallah-Iwasso MT, Mendes E, Mendes, NF. In vivo and in vitro evaluation of cell mediated im- munity in patients with paracoccidioidomycosis. Cell Im- munol. 1976; 24: 365-78.
8. Calich VLG, Singer-Vermes LM, Siqueira AM, Burger E. Susceptibility and resistance of inbred mice to Paracoc- cidioides brasiliensis. Br J Exp Pathol 1985; 66: 585-94.
9. Calich VLG, Burger E, Kashino SS, Fazioli RA, Singer- Vermes LM. Resistance to Paracoccidoides brasiliensis in mice is controlled by a single dominant autosomal gene. Infect Immun 1987; 55: 1919-23.
10. Kashino SS, Calich VLG, Burger E, Singer-Vermes LM. In vivo and in vitro characteristics of six Paracoccidioides brasiliensis strains. Mycopathologia 1987; 92: 173-78.
11. Singer-Vermes LM, Burger E, Franco MF, Moscardi- Bacchi M, Mendes-Giannini MJS, Calich VLG. Evalu- ation of the pathogenicity and immunogenicity of seven Paracoccidioides brasiliensis isolates in susceptible inbred mice. J Med Vet Mycol 1989; 27: 71-82.
12. Fava Netto C. Estudos quantitativos sobre a fixa~ao do complemento na blastomicose sul-americana, corn antfgeno polissacarfdico. Arq Cir Clin Exp S Paulo 1955; 18: 197-254.
13. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Pro- tein measurement with the Folin phenol reagent. J Biol Chem 1951; 193: 265-75.
14. Kabat EA, Manfred MM. Imunoqufmica experimental, 2nd ed. M6xico: Prensa Medica Mexicana, 1968: 31-32.
15. Berliner MD, Reca ME. Vital staining of Histoplasma capsulatum with Janus Green B. Sabouraudia 1966; 5: 26-29.
16. Fazioli RA, Singer-Vermes LM, Kashino SS, Burger E,

146
Calich VLG. Standardization of the intradermal test in experimental murine paracoccidioidomycosis. Braz J Med Biol Res 1986; 19: 641A.
17. Corsini AC, Oliveira OLP, Costa MG. Unimpaired de- layed type hypersensitivity reactions in mice infected with Trypanosoma cruzi strain Y. Z Parasitenkunde 1980; 61: 179-85.
18. Curotto MA, Abrahamsohn IA. Contact hypersensitivity to DNFB in Trypanosoma cruzi infected mice. Rev Inst Med Trop S Paulo 1982; 24: 69-74.
19. Sy MS, Moorhead JW, Claman HN. Regulation of cell mediated immunity by antibodies: possible role of anti- receptor antibodies in the regulation of contact sensitivity to DNFB in mice. J Immunol 1979; 123: 2593-98.
20. Marchal G, Seman M, Millon G, Truffa-Bachi P, Zilber- farb V. Local adoptive transfer of skin delayed-type hyp- ersensitivity initiated by a single T lymphocytes. J Immu- nol 1982; 129: 954-58.
21. Zar JH. Biostatistical analysis, 2nd ed. New Jersey: Pren- tice Hall, 1984.
22. Miyaji M, Nishimura K. Granuloma formation and killing functions of granuloma in congenitally athymic nude mice infected with Blastomyces dermatitidis and Paracocci- dioides brasiliensis. Mycopathologia 1983; 82: 129-41.
23. Robledo MA, Graybill JR, Ahrens J, Restrepo A, Drutz DJ, Robledo M. Host defense against experimental para- coccidioidomycosis. Am Rev Respir Dis. 1982; 125: 563- 67.
24. Cox RA, Kennell W, Boneyk L, Murphy JW. Induction ' and expression of cell-mediated immune responses in in- bred mice infected with Coccidioides immitis. Infect Immun 1988; 56: 13-17.
25. Zacharias D, Ueda A, Moscardi-Bacchi M, Franco M, San-Blas G. A comparative histopathological, immunol- ogical and biochemical study of experimental intravenous paracoccidioidomycosis induced in mice by three Paracoc- cidioides brasiliensis isolates. J Med Vet Mycol 1986; 24: 445-54.
26. Moscardi M, Franco MF. Paracoccidioidomicose experi- mental do camundongo, I: Aspectos imunopatol6gicos da infec~fto intraperitoneal. Rev Inst Med Trop S Paulo 1980; 22: 286-93.
27. Taylor ML, Dfaz S, Gonz~ilez PA, Sosa AC, Toriello C. Relationship between pathogenesis and immune regu- lation mechanisms in histoplasmosis: a hypothetical ap- proach. Rev Infect Dis 1984; 6: 775-82.
28. Martins AB, Raffel V. Cellular activities in hypersensitiv- ity reactions, I: Comparative cytology of delayed, Jones- Mote and Arthus reactions. J Immunol 1964; 93: 937-47.
29. Jimenez-Finkel BE, Murphy JW. Induction of antigen- specific T-suppressor cells by soluble Paracoccidioides brasiliensis antigen. Infect Immun 1988; 56: 734-43.
30. Murphy JW, Cox RA. Induction of antigen-specific suppr- ession by circulating Cryptococcus neoformans antigen. Clin Exp Immunol 1988; 73: 174-80.
31. Castaneda E, Brummer E, Pappagianis D, Stevens DA. Impairment of cellular but not humoral immune re- sponses in chronic pulmonary and disseminated paracoc- cidioidomycosis in mice. Infect Immun 1988; 56: 1771- 77.
32. Castafieda E, Brummer E, Pappagianis D, Stevens DA. Regulation of immune responses by T-suppressor cells and by serum in chronic paracoccidioidomycosis. Cell Im- munol 1988; 117: 1-11.
33. Restrepo AM, Schneidau JD JR. Nature of skin-reactive principle in culture filtrates prepared from Paracocci- dioides brasiliensis. J Bacteriol 1967; 93: 1741-48.
34. Fava Netto C, Guerra MAG, Costa EO. Contribuigfio ao estudo imunoldgico da paracoccidioidomicose. Rea96es intrgtdermicas em pacientes com dois antfgenos homd- logos e dois heteroldgos. Rev Inst Med Trop S Paulo 1976; 18: 186-90.
35. Singer-Vermes LM, Caldeira CB, Burger E, Calich VLG. Experimental murine paracoccidioidomycosis: relation- ship among the dissemination of the infection, humoral and cellular immune responses. Clin Exp. Immunol 1993; 94: 75-79.
36. Calich VLG, Fazioli RA., Teixeira HC, Russo M, Singer- Vermes LM, Burger E, Vaz CA.C. Mechanisms of host resistance to Paracoccidioides brasiliensis. In: Torres-Ro- driguez JM, ed. Proceedings of the 10th Congress of the International Society for Human and Animal Mycology. Barcelona J.R. Prous Science Ed, 1988: 154-159.
37. Vaz CA_C, MacKenzie DWR, Hearn V, Camargo ZP, Singer-Vermes LM, Burger E, Calich VLG. Specific re- cognition pattern of IgM and IgG antibodies produced in the course of experimental paracoccidioidomycosis. Clin Exp Immunol 1992: 88: 119-23.
Address for correspondence: Vera Lficia Garcia Calich, De- partamento de Imunologia, Instituto de Ci~ncias Biom6dicas, Universidade de Sao Paulo, S~o Paulo, 05508-900, SP, Brasil Phone: 55-011-818-7397; Fax: 55-011-813-0845




















