
Mapping hypersensitivity/allergic diseases in the International ...
Upload
khangminh22Category
view
1download
0
Kinetics of Delayed-Type Hypersensitivity to Tuberculin
Induced by Bacille Calmette-Guérin Vaccination in
Northern Malawi
Sian Floyd, Jorg M Pönnighaus, Lyn Bliss, Prince Nkhosa, Lifted Sichali, Glyn
Msiska, Paul E. M. Fine
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022

807
Kinetics of Delayed-Type Hypersensitivity to Tuberculin Induced by BacilleCalmette-Guerin Vaccination in Northern Malawi
Sian Floyd,1 Jorg M Ponnighaus,2,a Lyn Bliss,1
Prince Nkhosa,2 Lifted Sichali,2 Glyn Msiska,2
and Paul E. M. Fine1
1Department of Infectious and Tropical Diseases, Infectious DiseasesEpidemiology Unit, London School of Hygiene and Tropical Medicine,
London, United Kingdom; 2Karonga Prevention Study, Chilumba,Karonga District, Malawi
During 1986–1989, a bacille Calmette-Guerin (BCG) vaccine trial was carried out in north-ern Malawi. The effects of age, sex, and prevaccination delayed-type hypersensitivity (DTH)on the time course of the DTH response over 1–36 months after vaccination were studied in2418 persons. DTH response increased rapidly, to peak at 31–90 days after vaccination, whenmost persons had a measurable response. This was followed by a marked decline by 181–365days, particularly in those !15 years old at vaccination, followed by a more gradual decline.Prevaccination DTH was the single best predictor of postvaccination DTH. BCG-inducedDTH responsiveness appears to decline more rapidly in tropical than in temperate environ-ments. This may reflect high prevalence of exposure to other infections, which induce a Th2bias or compete for “space” within the T lymphocyte compartment. The inability of somepersons to mount a persistent DTH response probably reflects genetic background and/orenvironmental exposure history.
The discussion of the relationship between bacille Calmette-Guerin (BCG) vaccination and delayed-type hypersensitivity(DTH) to tuberculin has concentrated mainly on 3 issues. First,is the induction of tuberculin sensitivity an indication of inducedprotection against tuberculosis (TB)? It is now widely agreed thatthis is not so. Even though quality-control screening of BCGvaccines requires the induction of DTH in guinea pigs, there isno evidence that vaccine-induced DTH is a correlate of inducedprotection in humans [1]. The second issue concerns the impli-cations of BCG-associated tuberculin hypersensitivity for controlpolicies. The use and interpretation of tuberculin tests must takeinto account a person’s history of BCG vaccination [2]. The thirdissue concerns the implications of BCG-induced DTH when tu-berculin surveys are used to estimate the annual risk of Myco-bacterium tuberculosis infection. Such studies typically excludepersons with a history of BCG vaccination [3, 4].
Most discussion of these issues has neglected the fact that
Received 11 February 2002; revised 15 May 2002; electronically published12 August 2002.
Presented in part: 32nd World Conference on Lung Health, InternationalUnion Against Tuberculosis and Lung Disease, Paris, 1–4 November 2001(poster 5211).
Financial support: British Leprosy Relief Association; International Fed-eration of Anti-Leprosy Organizations; World Health Organization/UnitedNations Development Programme/World Bank Special Programme for Re-search and Training in Tropical Diseases; Wellcome Trust.
a Present affiliation: Lugala Lutheran Hospital, Lugala, Tanzania.Reprints or correspondence: Ms. Sian Floyd, Infectious Diseases Epi-
demiology Unit, Dept. of Infectious and Tropical Diseases, London Schoolof Hygiene and Tropical Medicine, Keppel St., London WC1E 7HT, UnitedKingdom ([email protected]).
The Journal of Infectious Diseases 2002;186:807–14� 2002 by the Infectious Diseases Society of America. All rights reserved.0022-1899/2002/18606-0011$15.00
the degree of BCG-induced DTH depends on time and geog-raphy, as first noted long ago. In 1952, it was noted that thedistribution of tuberculin sensitivity 1 year after BCG vacci-nation differed among populations in Denmark, Greece (Ath-ens), and Egypt (Cairo). The Danish vaccinees had the strongestresidual DTH, and the Egyptians had the least [5]. This ob-servation is consistent with several others in the literature. Forexample, BCG-vaccinated Danish children who were tested fortuberculin either 10 weeks or 5 years after vaccination appearedto maintain their tuberculin hypersensitivity [6–8], whereas per-sons in a large south India BCG vaccine trial who were testedat 2.5 months or 2.5 years after vaccination showed consid-erable loss of hypersensitivity over time [9]. In a study in Ma-lawi, only a small proportion (∼10%) of BCG-vaccinated per-sons maintained BCG-attributable hypersensitivity for manyyears after vaccination [3].
These observations raise 3 important questions. What arethe kinetics of the induction and persistence of DTH respon-siveness after BCG vaccination? What are the implications ofthe kinetics of BCG-induced DTH for immunity and T cellmemory? Finally, why do these kinetics appear to differ betweenpopulations? Published studies are limited in their ability toanswer these questions. Few have measured the DTH responseat several time points after BCG vaccination, few have includedboth adults and children, and only one has included personswho received a repeat BCG vaccination [10].
Here, we report tuberculin-sensitivity data from northernMalawi for 12000 children and adults. Their DTH responseswere measured from 31 days to 36 months after either a firstor a second BCG vaccination. We explore the implications ofage, sex, initial BCG scar status, and prevaccination DTH forthe magnitude and time trend of the postvaccination DTH
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022

808 Floyd et al. JID 2002;186 (15 September)
response. We also assess detailed age trends in tuberculin sen-sitivity among young children as a function of BCG scar status,which reflects vaccination in infancy.
Methods
During 1986–1989, the Karonga Prevention Study implementeda randomized, double-blind vaccine trial of BCG-containing vac-cine versus repeat inoculation with the BCG vaccine versus BCGvaccine plus killed M. leprae in the Karonga District of northernMalawi. The background, design, recruitment methods, and trialresults are described in detail elsewhere [11, 12]. Trained vaccinatorsgave all vaccinations.
Subjects were eligible for the trial if they were 13 months old,were born after 1914, had no evidence of past or present TB orleprosy, and had no evidence of current malnutrition or severeillness. Those with scar evidence of previous BCG vaccination atrecruitment (“scar positive”) were given either a repeat BCG vac-cination, a repeat BCG vaccination plus killed M. leprae,86 � 10or placebo (dextran). Persons with no evidence of a scar at re-cruitment (“scar negative”) were given either BCG vaccination,BCG vaccination plus killed M. leprae, or BCG vaccination75 � 10plus killed M. leprae. BCG vaccine was provided by Glaxo86 � 10(strain 1077) and was given in a standard dose (0.1 mL) in thedeltoid region of the right arm.
Of 1120,000 subjects in the trial, 2418 had tuberculin skin testsbetween 31 days and 3 years after receipt of a BCG-containingvaccine (6 subjects were tested twice), and 452 were tested afterreceiving placebo. Of these 2870 persons, 1725 were also tested 1or 2 days before vaccination or on the day of vaccination.
All tuberculin skin tests used RT23 (2 tuberculin units; StatensSeruminstitut) and were done on the volar surface of a forearm byintradermal injection of 0.1 mL of reagent with a sterile 26-gaugeneedle. Tests were read at 48–72 h after vaccination; indurationdiameters were measured along and across the arm. Mean diam-eters are used in the analyses presented here. Data were coded,checked, and entered into computers at the project headquartersin Malawi and analyzed using STATA software (StataCorp) at theLondon School of Hygiene and Tropical Medicine (London).
The proportions of persons with an induration diameter of 10,15, and 10 mm before and after vaccination were analyzed bylogistic regression to explore the effects of age at vaccination, sex,initial BCG scar status, vaccine type, and prevaccination DTH onthe time course of the response. Subjects were divided into 2 agegroups (!15 and �15 years old) for analysis of whether the timecourse of the DTH response after vaccination depended on age atvaccination. Subjects were further subdivided into age groups (�4,5–9, 10–14, 15–24, 25–44, and �45 years), to control for the effectof age on the prevaccination response. The division by age !15 or�15 years distinguished children and adults, or “young” and“older” persons, and a finer division was not possible owing tosample size constraints. Prevaccination DTH was categorized as0, 1–5, 6–10, and 110 mm, a standard categorization. The effectsof age at vaccination, sex, initial BCG scar status, and time aftervaccination on the pre- and postvaccination DTH response wereall estimated, adjusting for each other. The effect of prevaccination
DTH on the postvaccination DTH response was estimated afteradjusting for age, sex, initial BCG scar status, and time since vac-cination. Median responses were also analyzed.
Analyses were also carried out, allowing for nonindependenceof observations made on the same person (because most individualswere measured twice, once before and once after vaccination).These analyses gave virtually identical results to those that assumedindependence. For simplicity, only those that assumed indepen-dence are presented here.
We also analyzed data for 14,666 BCG scar–positive childrenand 9220 BCG scar–negative children, who had tuberculin skintests at age �10 years, during a population survey of Karongadistrict in 1980–1985 [13]. The percentage of children with a skin-test response above the thresholds of 5 and 10 mm was calculatedby BCG scar status and age.
Results
Data are summarized in table 1 as the percentage of subjectswith an induration diameter of 15 mm before vaccination andat 5 time points after vaccination. The trends were similar whenwe used 0- or 10-mm thresholds (data not shown). As expected,the prevalence of DTH responsiveness prior to vaccination in-creased with age (table 1 and figure 1). The time course of theDTH response was similar for persons who received BCG withor without killed M. leprae (table 1) and, thus, data from thesegroups are combined in subsequent analyses.
Figure 1 shows the frequency distributions of DTH responsebefore and at 5 time points after vaccination for subjects !15and �15 years old at vaccination among persons who lackeda BCG scar at the time of vaccination. There was a sharpincrease in response 2–3 months after vaccination in both agegroups, at which time almost all persons had a DTH response(10 mm). This was followed by a marked decline in DTH re-sponse that was larger among younger than among older sub-jects. The evidence that the time course of the postvaccinationDTH response differed by age was strong ( andP p .012 P !
for the 5- and 10-mm thresholds, respectively) and was.001also evident in the time trend of the median response (data notshown).
The sex difference in DTH response among those !15 yearsold was small and not statistically significant, but, at older ages,the responses were higher among men (figure 2A) both beforeand 4–36 months after vaccination (test for age-sex interaction4–36 months after vaccination, and for useP p .006 P ! .001of 5- and 10-mm thresholds, respectively). The exception wasat the peak period 2–3 months after vaccination when men andwomen had similar DTH responses. Of importance, the maleexcess after vaccination could be explained entirely by the DTHdifference at the time of vaccination, because there was noevidence of an age-sex interaction in DTH response after vac-cination after adjustment for the prevaccination response (anal-ysis based on 1467 skin tests) or when analysis was restricted
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022

JID 2002;186 (15 September) Kinetics of BCG-Induced DTH 809
Table 1. Subjects with a delayed-type hypersensitivity response to tuberculin of 15 mm, by time sincevaccination, initial bacille Calmette-Guerin (BCG) scar status, age at vaccination, sex, and vaccine type.
BCG scar status at vaccination, variableBefore
vaccination
Test time after vaccination
31–90days
91–180days
181–365days
13–24months
25–36months
NegativeAge, years
�4 9 (248) 82 (45) 57 (77) 50 (24) 48 (130) 40 (156)5–9 15 (126) 95 (21) 65 (40) 69 (16) 43 (63) 62 (76)10–14 33 (144) 86 (28) 79 (42) 58 (12) 68 (82) 70 (63)15–24 47 (96) 88 (17) 76 (25) 73 (11) 78 (58) 62 (39)25–44 51 (202) 90 (40) 83 (84) 74 (27) 71 (76) 73 (89)�45 64 (171) 100 (34) 96 (54) 73 (22) 83 (92) 73 (74)
SexMale 37 (434) 90 (83) 76 (140) 67 (46) 61 (224) 57 (220)Female 33 (553) 89 (102) 76 (182) 65 (66) 66 (277) 61 (277)
VaccineBCG 36 (456) 87 (78) 74 (160) 67 (57) 64 (235) 59 (188)BCG � 5 � 107 killed M. leprae 33 (135) 93 (29) 72 (47) 38 (13) 62 (80) 53 (91)BCG � 6 � 108 killed M. leprae 34 (396) 91 (78) 79 (115) 74 (42) 65 (186) 63 (218)
PositiveAge, years
�4 16 (128) 100 (20) 61 (38) 75 (16) 59 (27) 55 (49)5–9 28 (243) 97 (32) 74 (38) 76 (17) 68 (93) 59 (75)10–14 34 (134) 94 (17) 86 (22) 100 (10) 77 (44) 77 (47)15–24 57 (161) 97 (30) 94 (32) 74 (19) 75 (44) 82 (57)25–44 54 (69) 100 (13) 80 (10) 50 (8) 69 (26) 67 (21)�45 100 (3) 100 (1) — — — 100 (1)
SexMale 38 (370) 9 (61) 78 (63) 76 (37) 71 (115) 70 (136)Female 34 (368) 100 (52) 77 (77) 76 (33) 69 (119) 65 (114)
VaccineBCG 31 (292) 98 (46) 78 (120) 77(66) 69 (120) 69 (143)BCG � 6 � 108 killed M. leprae 40 (184) 97 (67) 75 (20) 50 (4) 71 (114) 66 (107)Placebo 38 (262) 56 (50) 41 (97) 42 (45) 45 (141) 45 (119)
NOTE. Data are percentage (no.) of subjects. M. leprae, Mycobacterium leprae.
to persons who were BCG scar negative and had no DTHresponse before vaccination (analysis based on 349 skin tests;figure 2B).
Initial BCG scar status influenced the postvaccination re-sponse, with higher responses in those with a prior BCG scar( , 5- and 10-mm thresholds; figure 3). This differenceP ! .001was also evident in the median responses (data not shown).The association remained strong even after stratification of theanalysis according to prevaccination DTH response and alsoafter restriction of the analysis to persons with a prevaccinationresponse of 0 mm (data not shown). The effects of initial BCGscar status and sex on the postvaccination DTH response weresmall relative to the effects of age at vaccination and time sincevaccination (figures 1, 2, and 3).
There was strong evidence that DTH prior to vaccinationwas an important determinant of postvaccination DTH re-sponsiveness ( ) for children (!15 years old) and adultsP ! .001(�15 years old), regardless of BCG scar status at vaccination.Prevaccination DTH was a much better predictor of the post-vaccination DTH response than age, sex, or initial BCG scarstatus. Figure 4 provides illustrative data for adults (�15 yearsold) without a BCG scar at vaccination. Most subjects, re-
gardless of prevaccination DTH, mounted a DTH response31–90 days after vaccination, but most tended to revert to theirprevaccination status by 2 years after vaccination. Of interest,of 33 subjects with an initial response of 0 mm, 20 (61%) hadno DTH response when tested �1 year later, whereas only 8%( ) with an initial response of 110 mm had no responsen p 51�1 year after vaccination ( ).P ! .001
The raw data, with a threshold of 15 mm (table 1), showedan increase in DTH response in the placebo group 31–90 daysafter vaccination. This increase was also evident when a 110-mm threshold was used (data not shown). Further analysis ofthe placebo group suggested that the magnitude of this boostingeffect might be larger in persons !15 years old than in those�15 years old, whereas there was little evidence that the mag-nitude of the boosting effect changed with time since vacci-nation. However, the analysis was limited by small sample size,and confidence intervals were very wide. Nevertheless, in com-parison with the dramatic changes in DTH response observedin vaccinated subjects (table 1), the boosting effect was small.Additional evidence that the boosting effect was minor wasthat, among persons in the placebo group who were testedbefore vaccination, the percentage whose DTH response had
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022
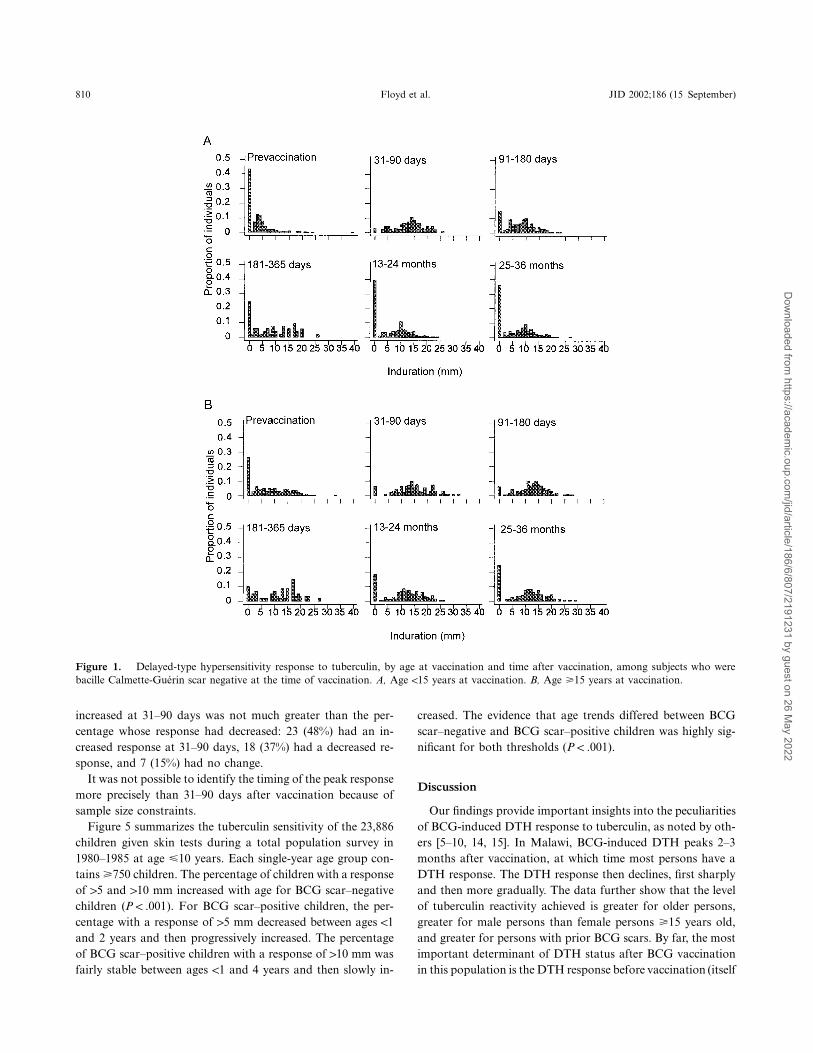
810 Floyd et al. JID 2002;186 (15 September)
Figure 1. Delayed-type hypersensitivity response to tuberculin, by age at vaccination and time after vaccination, among subjects who werebacille Calmette-Guerin scar negative at the time of vaccination. A, Age !15 years at vaccination. B, Age �15 years at vaccination.
increased at 31–90 days was not much greater than the per-centage whose response had decreased: 23 (48%) had an in-creased response at 31–90 days, 18 (37%) had a decreased re-sponse, and 7 (15%) had no change.
It was not possible to identify the timing of the peak responsemore precisely than 31–90 days after vaccination because ofsample size constraints.
Figure 5 summarizes the tuberculin sensitivity of the 23,886children given skin tests during a total population survey in1980–1985 at age �10 years. Each single-year age group con-tains �750 children. The percentage of children with a responseof 15 and 110 mm increased with age for BCG scar–negativechildren ( ). For BCG scar–positive children, the per-P ! .001centage with a response of 15 mm decreased between ages !1and 2 years and then progressively increased. The percentageof BCG scar–positive children with a response of 110 mm wasfairly stable between ages !1 and 4 years and then slowly in-
creased. The evidence that age trends differed between BCGscar–negative and BCG scar–positive children was highly sig-nificant for both thresholds ( ).P ! .001
Discussion
Our findings provide important insights into the peculiaritiesof BCG-induced DTH response to tuberculin, as noted by oth-ers [5–10, 14, 15]. In Malawi, BCG-induced DTH peaks 2–3months after vaccination, at which time most persons have aDTH response. The DTH response then declines, first sharplyand then more gradually. The data further show that the levelof tuberculin reactivity achieved is greater for older persons,greater for male persons than female persons �15 years old,and greater for persons with prior BCG scars. By far, the mostimportant determinant of DTH status after BCG vaccinationin this population is the DTH response before vaccination (itself
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022
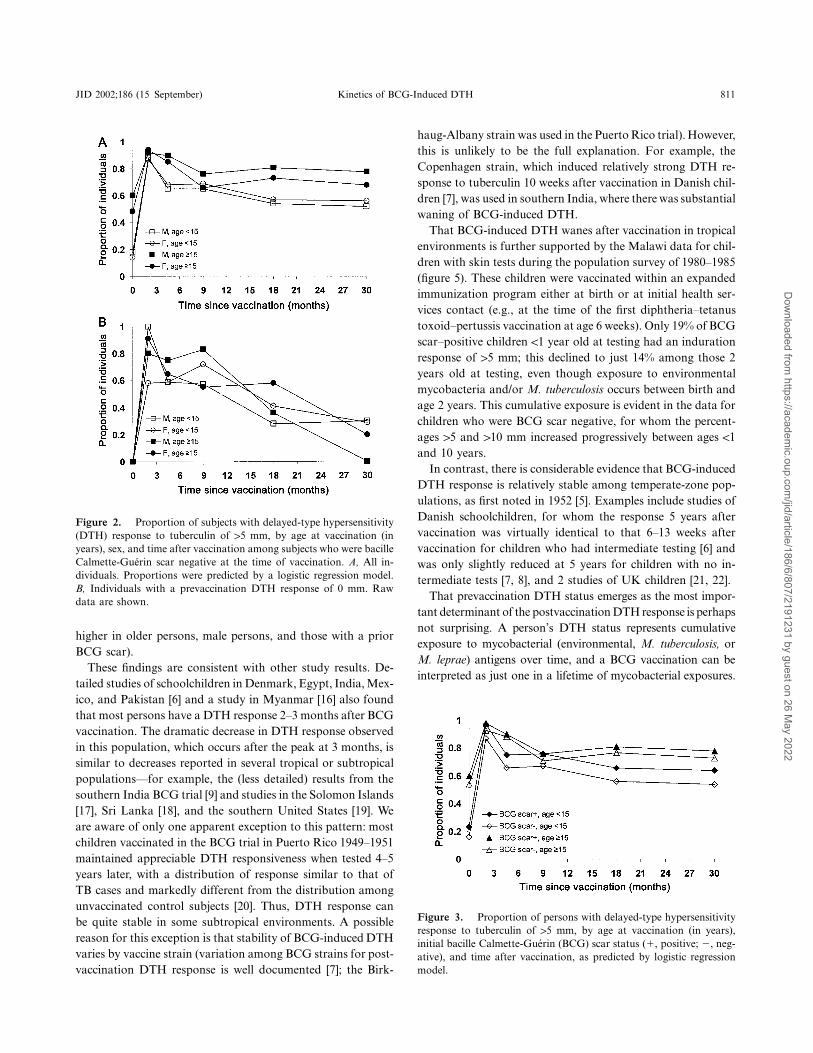
JID 2002;186 (15 September) Kinetics of BCG-Induced DTH 811
Figure 2. Proportion of subjects with delayed-type hypersensitivity(DTH) response to tuberculin of 15 mm, by age at vaccination (inyears), sex, and time after vaccination among subjects who were bacilleCalmette-Guerin scar negative at the time of vaccination. A, All in-dividuals. Proportions were predicted by a logistic regression model.B, Individuals with a prevaccination DTH response of 0 mm. Rawdata are shown.
Figure 3. Proportion of persons with delayed-type hypersensitivityresponse to tuberculin of 15 mm, by age at vaccination (in years),initial bacille Calmette-Guerin (BCG) scar status (�, positive; �, neg-ative), and time after vaccination, as predicted by logistic regressionmodel.
higher in older persons, male persons, and those with a priorBCG scar).
These findings are consistent with other study results. De-tailed studies of schoolchildren in Denmark, Egypt, India, Mex-ico, and Pakistan [6] and a study in Myanmar [16] also foundthat most persons have a DTH response 2–3 months after BCGvaccination. The dramatic decrease in DTH response observedin this population, which occurs after the peak at 3 months, issimilar to decreases reported in several tropical or subtropicalpopulations—for example, the (less detailed) results from thesouthern India BCG trial [9] and studies in the Solomon Islands[17], Sri Lanka [18], and the southern United States [19]. Weare aware of only one apparent exception to this pattern: mostchildren vaccinated in the BCG trial in Puerto Rico 1949–1951maintained appreciable DTH responsiveness when tested 4–5years later, with a distribution of response similar to that ofTB cases and markedly different from the distribution amongunvaccinated control subjects [20]. Thus, DTH response canbe quite stable in some subtropical environments. A possiblereason for this exception is that stability of BCG-induced DTHvaries by vaccine strain (variation among BCG strains for post-vaccination DTH response is well documented [7]; the Birk-
haug-Albany strain was used in the Puerto Rico trial). However,this is unlikely to be the full explanation. For example, theCopenhagen strain, which induced relatively strong DTH re-sponse to tuberculin 10 weeks after vaccination in Danish chil-dren [7], was used in southern India, where there was substantialwaning of BCG-induced DTH.
That BCG-induced DTH wanes after vaccination in tropicalenvironments is further supported by the Malawi data for chil-dren with skin tests during the population survey of 1980–1985(figure 5). These children were vaccinated within an expandedimmunization program either at birth or at initial health ser-vices contact (e.g., at the time of the first diphtheria–tetanustoxoid–pertussis vaccination at age 6 weeks). Only 19% of BCGscar–positive children !1 year old at testing had an indurationresponse of 15 mm; this declined to just 14% among those 2years old at testing, even though exposure to environmentalmycobacteria and/or M. tuberculosis occurs between birth andage 2 years. This cumulative exposure is evident in the data forchildren who were BCG scar negative, for whom the percent-ages 15 and 110 mm increased progressively between ages !1and 10 years.
In contrast, there is considerable evidence that BCG-inducedDTH response is relatively stable among temperate-zone pop-ulations, as first noted in 1952 [5]. Examples include studies ofDanish schoolchildren, for whom the response 5 years aftervaccination was virtually identical to that 6–13 weeks aftervaccination for children who had intermediate testing [6] andwas only slightly reduced at 5 years for children with no in-termediate tests [7, 8], and 2 studies of UK children [21, 22].
That prevaccination DTH status emerges as the most impor-tant determinant of the postvaccination DTH response is perhapsnot surprising. A person’s DTH status represents cumulativeexposure to mycobacterial (environmental, M. tuberculosis, orM. leprae) antigens over time, and a BCG vaccination can beinterpreted as just one in a lifetime of mycobacterial exposures.
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022
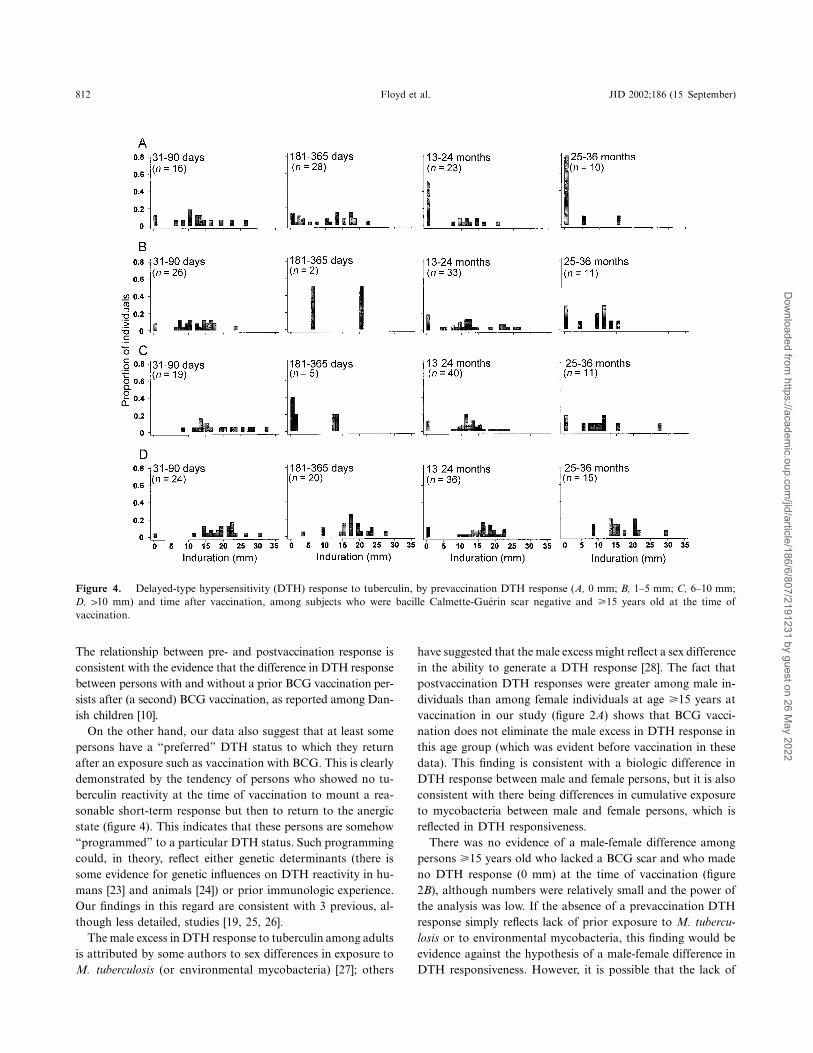
812 Floyd et al. JID 2002;186 (15 September)
Figure 4. Delayed-type hypersensitivity (DTH) response to tuberculin, by prevaccination DTH response (A, 0 mm; B, 1–5 mm; C, 6–10 mm;D, 110 mm) and time after vaccination, among subjects who were bacille Calmette-Guerin scar negative and �15 years old at the time ofvaccination.
The relationship between pre- and postvaccination response isconsistent with the evidence that the difference in DTH responsebetween persons with and without a prior BCG vaccination per-sists after (a second) BCG vaccination, as reported among Dan-ish children [10].
On the other hand, our data also suggest that at least somepersons have a “preferred” DTH status to which they returnafter an exposure such as vaccination with BCG. This is clearlydemonstrated by the tendency of persons who showed no tu-berculin reactivity at the time of vaccination to mount a rea-sonable short-term response but then to return to the anergicstate (figure 4). This indicates that these persons are somehow“programmed” to a particular DTH status. Such programmingcould, in theory, reflect either genetic determinants (there issome evidence for genetic influences on DTH reactivity in hu-mans [23] and animals [24]) or prior immunologic experience.Our findings in this regard are consistent with 3 previous, al-though less detailed, studies [19, 25, 26].
The male excess in DTH response to tuberculin among adultsis attributed by some authors to sex differences in exposure toM. tuberculosis (or environmental mycobacteria) [27]; others
have suggested that the male excess might reflect a sex differencein the ability to generate a DTH response [28]. The fact thatpostvaccination DTH responses were greater among male in-dividuals than among female individuals at age �15 years atvaccination in our study (figure 2A) shows that BCG vacci-nation does not eliminate the male excess in DTH response inthis age group (which was evident before vaccination in thesedata). This finding is consistent with a biologic difference inDTH response between male and female persons, but it is alsoconsistent with there being differences in cumulative exposureto mycobacteria between male and female persons, which isreflected in DTH responsiveness.
There was no evidence of a male-female difference amongpersons �15 years old who lacked a BCG scar and who madeno DTH response (0 mm) at the time of vaccination (figure2B), although numbers were relatively small and the power ofthe analysis was low. If the absence of a prevaccination DTHresponse simply reflects lack of prior exposure to M. tubercu-losis or to environmental mycobacteria, this finding would beevidence against the hypothesis of a male-female difference inDTH responsiveness. However, it is possible that the lack of
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022

JID 2002;186 (15 September) Kinetics of BCG-Induced DTH 813
Figure 5. Proportions of children �10 years old tested in a popu-lation survey who had a DTH response to tuberculin of 15 mm or 110mm, by bacille Calmette-Guerin (BCG) scar status (�, positive; �,negative) and age.
an initial response in this group reflects not just lack of previousexposure but also a relative inability to respond (or program-ming not to respond). If a relative inability to respond werethe explanation, then the finding could still be consistent witha general sex difference in immune responsiveness among those�15 years old.
It is not easy to explain the finding that, after restriction ofthe analysis to those who made no DTH response prior tovaccination, the postvaccination DTH response was greateramong persons with an initial BCG scar than among thosewithout such a scar. A plausible explanation is an excess offalse-negative responses among those who were BCG scar pos-itive at vaccination, compared with those who were BCG scarnegative.
This is a large data set, and the trends are clear and con-vincing. However, we could not study trends in individuals overtime. Ideally, one would like to test subjects repeatedly tomap individual trajectories. This would enable assessment ofwhether, for example, individual DTH responses fluctuate ap-preciably. If so, data such as these would reflect only the pro-portion of persons who happened to be in a certain state atsuccessive times after vaccination. However, such an approachis not possible because of boosting attributable to repeatedtuberculin tests [29].
The prevalence of human immunodeficiency virus (HIV) inKaronga District was !3% among adults 15–49 years old atthe time of the surveys [30]. Thus, our findings refer effectivelyto HIV-negative subjects. There were no differences in resultswhen our data were stratified by whether the tests were readat 48 or 72 h [31].
Our central finding was that, in Malawi, BCG-induced DTHto tuberculin increases rapidly to a peak at 2–3 months aftervaccination and then decreases markedly. This is consistent withfindings in several other studies indicating that DTH respon-siveness decreases more rapidly in tropical environments thanin some temperate environments [3, 5, 7, 9]. Returning to the
second question posed in the introduction about the implica-tions of the kinetics of BCG-induced DTH for immunity andT cell memory, it is tempting to relate the difference in kineticsbetween temperate and tropical environments to the fact thatBCG is known to provide better protection against TB in tem-perate than in tropical environments [32]. In this context, wenote that the Puerto Rican data, which, in contrast to severalother studies in tropical or subtropical environments, indicatedrelative stability of postvaccination DTH, also showed thatBCG imparted a moderate level of protection (∼30%) againstTB in that population [20]. The relationship is not a simpleone, however, because there is no evidence that tuberculin sen-sitivity (i.e., as measured 1 year after vaccination) is a correlateof BCG-induced protection [1, 33].
In regard to the third question posed in the introduction,which asked why do the kinetics of BCG-induced DTH appearto differ between populations, the reasons for the geographicdifferences are not clear. A possible explanation is that theyreflect the high prevalence of exposure to other infections intropical (but not in nontropical) environments, including par-asites, which bias the immune system away from Th1 and DTHresponsiveness [34] or which may compete for attention or spacewithin the T lymphocyte compartment [35, 36]. A better un-derstanding of the kinetics of the immune responses underlyingthese observations will provide an important clue to the naturalhistory of mycobacterial infections.
References
1. Hart P D’A, Sutherland I, Thomas J. The immunity conferred by effectiveBCG and vole bacillus vaccines, in relation to individual variations intuberculin sensitivity and to technical variations in the vaccines. Tubercle1967;48:201–10.
2. Chin J. Control of communicable diseases. 17th ed. Washington, DC: Amer-ican Public Health Association, 2000.
3. Fine PEM, Bruce J, Ponnighaus JM, Nkhosa P, Harawa A, Vynnycky E.Tuberculin sensitivity: conversions and reversions in a rural African pop-ulation. Int J Tuberc Lung Dis 1999;3:962–75.
4. Rieder HL. Methodological issues in the estimation of the tuberculosis prob-lem from tuberculin surveys. Tuber Lung Dis 1995;76:114–21.
5. Palmer CE. BCG vaccination and tuberculin allergy. Lancet 1952;1:935–40.6. World Health Organization Tuberculosis Research Office. Certain charac-
teristics of BCG-induced tuberculin sensitivity. Bull World Health Organ1955;12:123–41.
7. Horwitz O, Bunch-Christensen K. Correlation between tuberculin sensitivityafter 2 months and 5 years among BCG vaccinated subjects. Bull WorldHealth Organ 1972;47:49–58.
8. Guld J, Wallgreen A, Sundaresan T, Kaufmann SHE, ten Dam G. The du-ration of BCG-induced tuberculin sensitivity in children, and its irrele-vance for revaccination. Bull World Health Organ 1968;39:829–36.
9. Trial of BCG vaccines in South India for tuberculosis prevention. Tuber-culosis Prevention Trial. Indian J Med Res 1980;72(Suppl):1–74.
10. Nyboe J. The immediate effects of BCG revaccination. Bull World HealthOrgan 1969;41:63–73.
11. Ponnighaus JM, Fine PEM, Gruer PJK, et al. The Karonga Prevention Trial:a leprosy and tuberculosis vaccine trial in northern Malawi. I. Methodsof the vaccination phase. Lepr Rev 1993;64:338–56.
12. Randomized controlled trial of single BCG, repeated BCG, or combined
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022

814 Floyd et al. JID 2002;186 (15 September)
BCG and killed Mycobacterium leprae vaccine for prevention of leprosyand tuberculosis in Malawi. Karonga Prevention Trial Group. Lancet1996;348:17–24.
13. Ponnighaus JM, Fine PEM, Bliss L, Sliney IJ, Bradley DJ, Rees RJW. TheLepra Evaluation Project (LEP), an epidemiological study of leprosy innorthern Malawi. I. Methods. Lepr Rev 1987;58:359–75.
14. Menzies D, Vissandjee B. Effect of Bacille Calmette-Guerin vaccination ontuberculin reactivity. Am Rev Respir Dis 1992;145:621–5.
15. Menzies D. What does tuberculin reactivity after bacille Calmette-Guerinvaccination tell us? Clin Infect Dis 2000;31(Suppl 3):S71–4.
16. Tun KM, Win H, Oo AK, Myint SS, Hla SK, Naing H. Tuberculin conversionafter BCG vaccination: comparison by gestation and by age at immu-nization in Myanmar. Trans R Soc Trop Med Hyg 2000;94:219–20.
17. Eason RJ. Tuberculin sensitivity. Ann Trop Paediatr 1987;7:87–90.18. Karalliede S, Katugaha LP, Uragoda CG. Tuberculin response of Sri Lankan
children after BCG vaccination at birth. Tubercle 1987;68:33–8.19. Shaw LW. Field studies on immunization against tuberculosis. I. Tuberculin
allergy following BCG vaccination of schoolchildren in Muscogee County,Georgia. Public Health Rep 1951;66:1415–26.
20. Comstock GW, Livesay VT, Woolpert SF. Evaluation of BCG vaccinationamong Puerto Rican children. Am J Public Health 1974;64:283–91.
21. Crawshaw PAG, Thomson AH. Heaf test results after neonatal BCG. ArchDis Child 1988;63:1490–1.
22. Ormerod LP, Garnett JM. Tuberculin skin reactivity four years after neonatalBCG vaccination. Arch Dis Child 1992;67:530–1.
23. Jepson AP, Fowler A, Banya WAS, et al. Genetic regulation of acquiredimmune responses to antigens of Mycobacterium tuberculosis: a study oftwins in West Africa. Infect Immun 2001;69:3989–94.
24. Magnusson M, Bentzon MW, Fennestad KL, Maxild J, Moller S. The roleof genetics in the response to tuberculin and to other mycobacterial sen-sitins. Bull Int Union Tuberc Lung Dis 1988;63:13–5.
25. Wijsmuller G. Relation between pre-vaccination and post-vaccination tu-berculin sensitivity. Bull World Health Organ 1966;35:459–78.
26. Narain R, Vallishayee RS, Mayurnath S. Post-vaccination tuberculin sensi-
tivity for assessing BCG vaccination in areas with high prevalence of non-
specific sensitivity. Tubercle 1981;62:231–9.
27. Roelsgaard E, Iversen E, Bløcher C. Tuberculosis in tropical Africa: an epi-
demiological study. Bull World Health Organ 1964;30:459–518.
28. Fine PEM, Floyd S, Stanford JL, et al. Environmental mycobacteria in north-
ern Malawi: implications for the epidemiology of tuberculosis and leprosy.
Epidemiol Infect 2001;126:379–87.
29. Thompson NJ, Glassroth JL, Snider DE Jr, Farer LS. The booster phenom-
enon in serial tuberculin testing. Am Rev Respir Dis 1979;119:587–97.
30. Glynn JR, Warndorff DK, Fine PEM, Msiska GK, Munthali MM, Ponnig-
haus JM. The impact of HIV on morbidity and mortality from tuber-
culosis in sub-Saharan Africa: a study in rural Malawi and review of the
literature. Health Transit Rev 1997;7(Suppl 2):75–87.
31. Cobelens F, Borgdorff MW. Boosting, BCG, and time of reading in tuberculin
skin testing. Lancet 2001;358:2014.
32. Fine PEM. Variation in protection by BCG: implications of and for heter-
ologous immunity. Lancet 1995;346:1339–45.
33. Fine PEM, Sterne JAC, Ponnighaus JM, Rees RJW. Delayed-type hyper-
sensitivity, mycobacterial vaccines and protective immunity. Lancet 1994;
344:1245–9.
34. Elias D, Wolday D, Akuffo H, Petros B, Bronner U, Britton S. Effect of
deworming on human T cell responses to mycobacterial antigens in hel-
minth-exposed individuals before and after Bacille Calmette-Guerin
(BCG) vaccination. Clin Exp Immun 2001;123:219–25.
35. Messele T, Abdulkadir M, Fontanet AL, et al. Reduced naive and increased
activated CD4 and CD8 cells in healthy adult Ethiopians compared with
their Dutch counterparts. Clin Exp Immunol 1999;115:443–50.
36. Selin LK, Lin MY, Kraemer KA, et al. Attrition of T-cell memory: selective
loss of LCMV epitope-specific memory CD8 T-cells following infections
with heterologous viruses. Immunity 1999;11:733–42.
Dow
nloaded from https://academ
ic.oup.com/jid/article/186/6/807/2191231 by guest on 26 M
ay 2022
Copyright © 2022 FDOKUMEN




















