
E-Beam Lithography Overlapping aware Stencil Planning for ...

RESEARCH ARTICLE
�EF1 and SIP1 Are Differentially Expressedand Have Overlapping Activities DuringXenopus EmbryogenesisLeo A. van Grunsven,1† Vincent Taelman,2† Christine Michiels,1 Karin Opdecamp,2
Danny Huylebroeck,1* and Eric J. Bellefroid2
The zinc finger/homeodomain transcription factor (zfhx1) family in vertebrates consists of two members,�EF1 and SIP1. They have been proposed to display antagonistic activities in the interpretation of Smad-dependent TGF� signaling during mesoderm formation. We cloned Xenopus �EF1 cDNA, analyzed theexpression profile of the gene, and compared the inducing and interacting properties of the protein to thatof XSIP1. Whereas XSIP1 RNA is selectively expressed in the early developing nervous system, we show thatX�EF1 gene transcription is only activated during neurulation and that its expression is restricted to theparaxial mesoderm. From early tail bud stage, X�EF1 and XSIP1 are coexpressed in migratory cranialneural crest, in the retina, and in the neural tube. Overproduction of X�EF1 in RNA-injected embryos, likethat of XSIP1, reduced the expression of BMP-dependent genes but only XSIP1 has the ability to induceneural markers. We find that X�EF1 and XSIP1 can both form complexes, although with different efficiency,with Smad3, with the coactivators p300 and pCAF, and with the corepressor CtBP1. Together, these resultsindicate that �EF1 and SIP1 do not function as antagonists during Xenopus early embryogenesis but dodisplay different repression efficiencies and interaction properties. Developmental Dynamics 235:1491–1500,2006. © 2006 Wiley-Liss, Inc.
Key words: �EF1; neural induction; SIP1; Sox2; Zfhx1; ZEB
Accepted 26 January 2006
INTRODUCTIONOne classification of transcription fac-tors with C2H2 type zinc fingers isbased on the number and organizationof the fingers (Iuchi, 2001). The zincfinger homeobox 1 (zfhx1) proteins arecharacterized by two clusters of mul-
tiple zinc fingers: one located in theN-terminal part of the protein and onein the C-terminal part, separated by alinker domain containing a homeodo-main-like sequence. This family hastwo members only, �EF1 (ZEB1,Zfhx1a) and SIP1 (ZEB2, Zfhx1b),
which exist in zebrafish, Xenopus,chick, mouse, and human (vanGrunsven et al., 2001).
�EF1 null mutant mice have multi-ple skeletal malformations, absence ofjoints, a severe T-cell deficiency, andthey die immediately after birth
The Supplementary Material referred to in this article can be found at http://www.interscience.wiley.com/jpages/1058-8388/suppmat1Department of Developmental Biology (VIB7), Flanders Interuniversity Institute for Biotechnology (VIB) and Laboratory of MolecularBiology (Celgen), University of Leuven, Campus Gasthuisberg, Leuven, Belgium2Laboratoire d�Embryologie Moleculaire, Universite Libre de Bruxelles, Institut de Biologie et de Medecine Moleculaires (IBMM), Gosselies,BelgiumGrant sponsor: Belgian program Inter-Universitary Attraction Poles from the Prime Minister’s Office, Science Policy Programming; Grantnumber: P5/35; Grant sponsor: the Communaute Francaise de Belgique; Grant number: ARC 00/05-250; Grant sponsor: the Fund forScientific Medical Research; Grant numbers: FRSM 3.4555.01; 3.4556.01; Grant sponsor: the International Brachet Stiftung; Grant number:01-4/l; Grant sponsor: the Fondation Reine Elisabeth; Grant number: 11256; Grant sponsor: VIB7; Grant sponsor: the University of Leuven;Grant number: OT/00/41; Grant sponsor: the Fund for Scientific Research-Flanders; Grant number: FWO-V G.0105.02.†Drs. van Grunsven and Taelman contributed equally to this work.*Correspondence to: Danny Huylebroeck, Department of Developmental Biology (VIB7), University of Leuven, CampusGasthuisberg (Bldg. Ond&Nav, box 812), Herestraat 49, B-3000 Leuven, Belgium.E-mail: [email protected]
DOI 10.1002/dvdy.20727Published online 3 March 2006 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 235:1491–1500, 2006
© 2006 Wiley-Liss, Inc.

(Takagi et al., 1998). SIP1 null mouseembryos show multiple defects fromembryonic day (E) 8.5 onward, do notundergo embryonic turning, are se-verely growth retarded by E9.5, anddie shortly thereafter. Analysis ofthese embryos showed that SIP1 isessential for the correct specificationof the neuroepithelium and delinea-tion of the neural plate, for the devel-opment of vagal neural crest cells, andfor the delamination and migration ofcranial neural crest cells (Van dePutte et al., 2003). Patients with het-erozygous deletions or truncations ofthe ZFHX1B gene (chromosome 2q22)suffer from Mowat–Wilson syndrome(Wakamatsu et al., 2001; Mowat et al.,2003). In Xenopus, XSIP1 is expressedin the prospective neurectoderm dur-ing gastrulation (Eisaki et al., 2000;van Grunsven et al., 2000). Overpro-duction of XSIP1 induces neural-spe-cific gene expression in explants of ec-toderm and represses in the embryothe transcription of the pan-mesoder-mal and nodal-induced brachyury(Xbra) gene, of BMP4, and of othergenes in the presumptive epidermis(Eisaki et al., 2000; Lerchner et al.,2000; Papin et al., 2002; Postigo et al.,2003). Loss-of-function studies in Xe-nopus showed that XSIP1 is essentialfor neural induction in amphibian em-bryos (Nitta et al., 2004).
Studies aiming at elucidating thefunction(s) and mechanism(s) of ac-tion of the zfhx1 proteins mainly havebeen carried out in cultured cells. TheDNA binding activity and specificityof the zinc finger clusters of �EF1 andSIP1 and a model in which both zincfinger clusters bind preferentially totwo spaced binding sites (primarilyCACCT(G), and alternatively CA-CANNT) have been proposed (Sekidoet al., 1997; Remacle et al., 1999; Ver-schueren et al., 1999). Spaced CAC-CT(G) sites are found in the upstreamregulatory region of many potentialtarget genes (van Grunsven et al.,2001). Recently, the E-cadherin genehas been identified as a target fortranscriptional repression by �EF1and SIP1 in vitro (Grooteclaes andFrisch, 2000; Comijn et al., 2001) andin vivo (Van de Putte et al., 2003).
Despite the vast number of studieson potential target genes for zfhx1family proteins, the mechanism(s) bywhich these proteins regulate target
genes remains unclear. They do bindto the MH2 domain of activated Smadproteins (Verschueren et al., 1999;Postigo, 2003), the intracellular effec-tors of transforming growth factor-beta (TGF�) family signalling (Hill,2001; Shi and Massague, 2003). �EF1and SIP1 interact with the corepres-sor CtBP (carboxyl terminal bindingprotein; Chinnadurai, 2003), whichcontributes to their repressive activityin vitro (Furusawa et al., 1999;Postigo and Dean, 1999; Grooteclaesand Frisch, 2000; Shi et al., 2003; vanGrunsven et al., 2003). However,when full-length proteins, which aredefective for CtBP binding, are used inthese in vitro assays, no clear correla-tion between the repressive activity ofthe zfhx1 proteins and CtBP bindingcan be established for the target genesstudied thus far (van Grunsven et al.,2003). Different repressive potentialshave been attributed to the differentdomains of �EF1 and SIP1, but evi-dence showing a correlation betweenthese domains and the function of�EF1 and SIP1 in vivo has been lack-ing. Postigo and coworkers (Postigo etal., 2003) have shown that �EF1 andSIP1 can regulate TGF�/bone mor-phogenetic protein (BMP) signallingin opposite ways, with �EF1 synergiz-ing with Smad-mediating transcrip-tional activation and SIP1 repressingit. It was suggested that this differen-tial effect is because SIP1 functionsonly as a transcriptional repressor byrecruiting CtBP to the Smads,whereas �EF1 also binds to the coac-tivators p300 and pCAF, which dis-place CtBP and allow it to function asan activator (Postigo, 2003; Postigo etal., 2003). In Xenopus, XSIP1 has beenshown to repress directly Xbra expres-sion during gastrulation, therebyplaying a role in the differentiationbetween mesodermal and ectodermallayers (Lerchner et al., 2000). In con-trast, overproduction of human �EF1induces Xbra expression ectopically,suggesting that both proteins competefor the same binding sites on the Xbrapromoter and act as antagonists(Postigo et al., 2003).
Here, we cloned a Xenopus �EF1cDNA and show that the gene is acti-vated later than XSIP1 and that it ispredominantly expressed in the em-bryonic paraxial mesoderm. By usingan identical RNA injection approach
used by others, we show that X�EF1,like XSIP1, represses the expressionof BMP-dependent genes, the inhibi-tory effect observed being less dra-matic, however, than that of XSIP1,which may explain the inability ofX�EF1 to neuralize ectodermal cells.We show that both X�EF1 and XSIP1can interact with the coactivatorspCAF and p300 and the corepressorCtBP1. In contrast to XSIP1, X�EF1needs p300 binding to be able to bindto Smads. Together, these data indi-cate that X�EF1 and XSIP1 do notfunction antagonistically but haveoverlapping, yet slightly different ac-tivities during early Xenopus develop-ment.
RESULTS
�EF1 and SIP1 AreDifferentially ExpressedDuring XenopusEmbryogenesis
We and others have found that Xeno-pus SIP1 (XSIP1) is expressed in theearly embryo in the neurectoderm andplays a role in neural differentiation(Eisaki et al., 2000; van Grunsven etal., 2000; Nitta et al., 2004). In thismanuscript, we wanted to knowwhether SIP1 and �EF1 have overlap-ping, counteracting, or distinct func-tions in the regulation of early embry-ogenesis. Therefore, we cloned theXenopus �EF1 (X�EF1) cDNA andcompared its expression and activitiesin the embryo to that of XSIP1. Figure1A shows an amino acid alignment ofhuman, mouse, and Xenopus laevis�EF1. The domains that were identi-fied in earlier studies in mouse andhuman �EF1 are conserved in Xeno-pus �EF1. An exception is the C3Hzinc finger that is present in m�EF1(from amino acid 499 to 519), and inmouse and Xenopus SIP1 but is ab-sent from human and Xenopus �EF1(Fig. 1B). This zinc finger has beenshown to be necessary for the bindingof the POU-domain of Oct1 to rat�EF1 (Smith and Darling, 2003). Inaddition, the first CtBP binding sitethat is present in mouse and human�EF1 and in SIP1 proteins is not con-served in Xenopus. During the courseof this study, the NIH Xenopus initia-tive reported a full-length Xenopuslaevis clone very similar to our Xeno-
1492 VAN GRUNSVEN ET AL.

pus �EF1 gene (GenBank accessionno. BC073606, further referred to asX�EF1b; 92% homology) that has thefirst CtBP site intact (Klein et al.,
2002). A schematic representation ofXenopus laevis and mouse �EF1 andSIP1 is given in Figure 1B, showingthat the overall similarity is low be-
tween SIP1 and �EF1, except for thewell-known domains. In particular,the amino-terminal (NZF) and car-boxy-terminal (CZF) zinc fingers are
Fig. 1. �EF1 amino acid and XZfhx1 family comparison. A: Amino acid sequence comparison between human (hs), mouse (mm), and Xenopus laevis(xla, this study; xlb, �EF1 submitted to GenBank by an NIH Xenopus initiative; GenBank accession nos. U12170, D76432, DQ319867, and BC073606,respectively). The boxed sequences are previously determined functional domains in mouse or human �EF1. C2H2 and C3H indicate the type of zincfinger; SBD, Smad binding domain; CtBP, CtBP binding site; HD, homeo-like domain. B: Schematic representation of X�EF1 and its similarities withm�EF1 and XSIP1. As a reference the homology between XSIP1 and mSIP1 is also represented. Dark gray boxes represent C2H2-type zinc fingers;light gray boxes C3H-type zinc-fingers; stripes represent individual CtBP binding sites (CBS); NZF, N-terminal zinc finger domain; CZF, C-terminal zincfinger domain. Comparisons at the amino acid level were carried out using ClustalW analysis.
�EF1 AND SIP1 DURING XENOPUS EMBRYOGENESIS 1493

highly conserved as well as the home-odomain-like domain (HD; Genettaand Kadesch, 1996). Striking is thelow homology between the respectiveSmad-binding domain (SBD) regions,i.e., 15% between X�EF1 and XSIP1,and 28% between m�EF1 and mSIP1.
Expression profiling showed thatX�EF1 transcripts can be detectedfrom the mid-neurula stage, while
Fig. 2. Expression of X�EF1 and XSIP1. A:Reverse transcriptase-polymerase chain reac-tion (RT-PCR) analysis of X�EF1 expression atthe indicated embryonic stages, comparedwith that of XSIP1. Control RT-PCR of histoneH4 was used as a reaction and loading control.B–O: Whole-mount in situ hybridization ofX�EF1 and XSIP1 expression at the indicatedstages. Sections through the spinal cord of theembryos in D,E,J, and K are shown in F,G,L,and K, respectively. ba, branchial arches; ev,eye vesicle; n, notochord; nc, neural crest; nt,neural tube; s, somites; tt, tail tip. B–E: Dorsalview, anterior toward the right. J,K: Dorsalview, anterior toward the left. (H,I,L,M) Lateralview, anterior toward the right.
Fig. 3.
Fig. 3. Overexpression of X�EF1 in embryosrepresses less efficiently than XSIP1 bone mor-phogenetic protein (BMP) -dependent genes.A: Four-cell stage embryos were injected withX�EF1 or XSIP1 encoding RNA into one blas-tomere (500 pg) in the equatorial region andanalyzed for Xbra expression. Both X�EF1 andXSIP1 repress Xbra expression. B–E: Eight- tosixteen-cell embryos injected into one animalblastomere with low (50 pg) or high (500 pg)doses of the indicated mRNA and analyzed forGata2, Msx1, and EK expression. F: Westernblot analysis of X�EF1- and XSIP-injected em-bryos showing that the proteins are equally ex-pressed. �-Tubulin antibody is used as a load-ing control. A–C: Respective repression 100%,n � 54 for XSIP1 and 100%, n � 50 for X�EF1(A); (B) 100%, n � 24 for XSIP1 and 100%, n �34 for X�EF1; (C) 100%, n � 16 for XSIP1 and100%, n � 14 for X�EF1. (D) 100%, n � 44 forXSIP1 and 100%, n � 36 for X�EF1; (E) all noneor only partially inhibited, n � 30 for X�EF1injected and all repressed, n � 32 for XSIP1. A:Embryos at gastrula stage are viewed from thevegetal pole. Embryos in D and E are viewedlaterally, anterior to the right; B is a ventral view;C a dorsal view, anterior to the right. The in-jected area (indicated by a black arrow) ismarked by blue staining for nuclear LacZ ex-pression obtained after coinjection of LacZRNA. All control embryos are viewed dorsolat-erally except Xbra, which is a vegetal view.
1494 VAN GRUNSVEN ET AL.

XSIP1 transcripts are already de-tected at early gastrula stage (Fig.2A). By in situ hybridization, X�EF1expression is observed first at stage13–14 in the paraxial mesoderm,while XSIP1 expression is restrictedto the neurectoderm (Fig. 2B,C). Bylate neurula to early tail bud stage,X�EF1 expression is also detected inmigratory cranial neural crest, in theretina and in the neural tube. Withinthe spinal cord, X�EF1 mRNA is con-fined to the dorsal lateral edges (Fig.2D,F,H,J,L). By late tadpole, X�EF1RNA levels remain high in the headand the tail (Fig. 2N,O). The expres-sion profile of X�EF1b is identical tothat of X�EF1a (see SupplementaryFigure S1, which can be viewed athttp://www.interscience.wiley.com/jpages/1058-8388/suppmat). From lateneurula stage, expression of X�EF1overlaps that of XSIP1 in the eye andcranial neural crest (Fig. 2D,E,H–M)but not in the dorsal midline whereX�EF1 expression is excluded frompresumptive premigratory trunk neu-ral crest cells, which strongly expressXSIP1 (Fig. 2F,G). Thus, �EF1 andSIP1 show distinct temporal and spa-tial expression patterns during Xeno-pus embryogenesis, although substan-tial overlap of expression also exists.Besides the expression of X�EF1 inthe eye, these results corroborate theexpression domain of �EF1 observedin mouse and chick embryos (Fu-nahashi et al., 1993; Takagi et al.,1998).
X�EF1 Represses, LessEfficiently Than XSIP1,BMP-Dependent Genes andDoes Not Cause Induction ofNeural Markers inEctodermal Cells
Injection of RNA encoding XSIP1 orhuman SIP1 in blastomeres of one- tofour-cell stage Xenopus embryos hasbeen shown to cause repression ofXbra in the mesoderm and of the ec-todermal marker epidermal keratin(EK), and induction of the neural platemarker Sox2, a gene critical for earlyneurogenesis (Papin et al., 2002; Nittaet al., 2004). To determine whetherX�EF1 has transcription regulatoryeffects similar to XSIP1 in this type ofexperiments, we injected RNA encod-
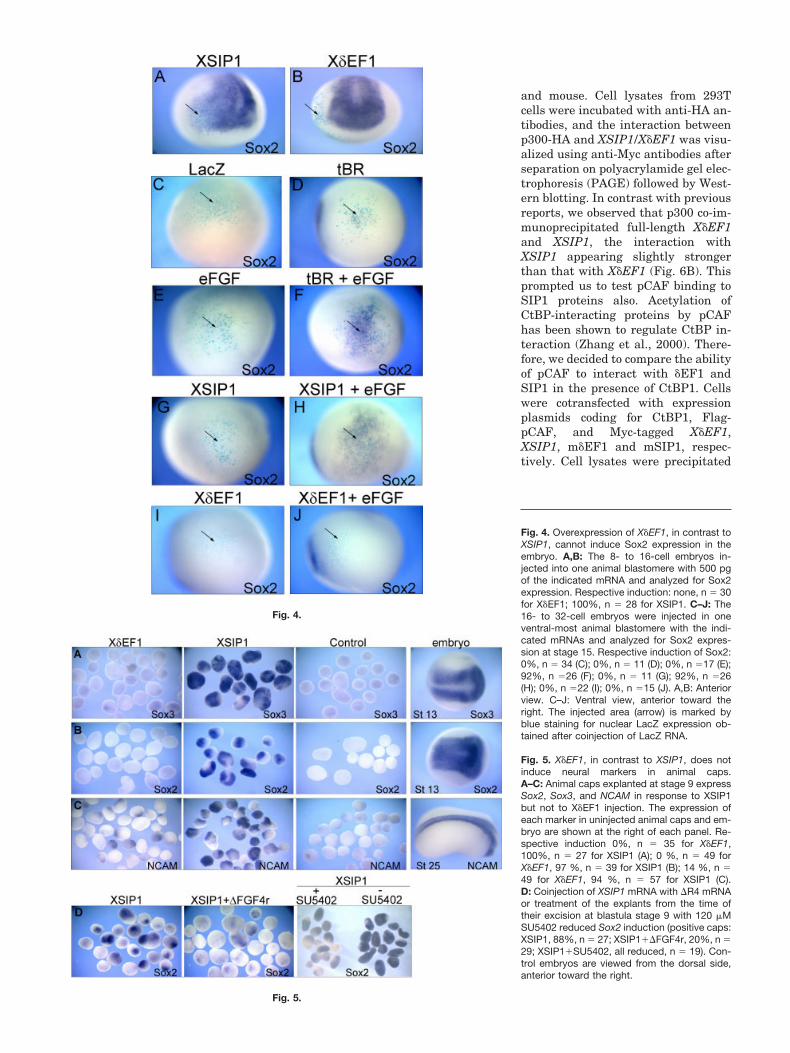
ing a myc-tagged X�EF1 or XSIP1 intoXenopus embryos (250–500 pg RNA).Western blot detection of protein ex-tracts from XSIP1 and X�EF1 injectedembryos shows that the proteins areequally well expressed (Fig. 3F). Over-production of X�EF1, like that ofXSIP1, was found to repress Xbra(Fig. 3A). At these doses, the expres-sion of the BMP-targets Msx1 (Suzukiet al., 1997), Gata2 (Walmsley et al.,1994), and E.K. was also repressed byboth proteins (Fig. 3B–D), the repres-sion observed in the X�EF1-injectedembryos appearing often less dra-matic than that observed in XSIP-in-jected embryos. At lower doses ofX�EF1 (25–50 pg RNA), the expres-sion of these marker genes was unal-tered, whereas their expression wasstill totally abolished by XSIP1 (Fig.3E, and data not shown). In X�EF1-injected embryos, no ectopic Sox2 orSox3 expression was observed,whereas it was found expanded in em-bryos injected with XSIP1 RNA. Iden-tical results were obtained using non–myc-tagged versions of the constructs(Fig. 4A,B, and data not shown).
The acquisition of neural fate in Xe-nopus, like in chick embryos, does notoccur solely by inhibition of BMP li-gand activity (the “default” model) butinvolves BMP-independent fibroblastgrowth factor (FGF) signaling (Linkerand Stern, 2004; Delaune et al., 2005).We asked whether increasing FGFsignaling in the embryo would helpX�EF1 and XSIP1 to induce neuraltissue. As reported, injection of RNAencoding a dominant-negative form ofthe BMP receptor (tBR, 200 pg/blas-tomere) together with 0.16 pg of eFGFRNA into ventral animal cells in 16-cell stage embryos induces Sox2 inventral ectodermal cells. At thesedoses, neither tBR nor eFGF alone ac-tivates mesoderm formation or in-duces Sox2 (Fig. 4D–F; Delaune et al.,2005). X�EF1 and XSIP1 RNA wereinjected alone or together with 0.16 pgof eFGF RNA. We found that, similarto the coinjection of tBR�eFGF RNA, acombination of XSIP1�eFGF RNA in-duced Sox2 in ventral ectodermalcells, whereas neither XSIP1 noreFGF could do this on their own (re-spectively, Fig. 4E,G,H). In identicalconditions, no activation of Sox2 wasobserved in embryos injected withX�EF1�eFGF RNAs (Fig. 4I,J).
We next used animal cap assays fordocumenting the effects of X�EF1. An-imal caps, cut from embryos injectedat the two-cell stage in each blas-tomere with 250 pg of myc-taggedXSIP1 RNA, revealed a dramatic in-crease in the levels of Sox3, Sox2, andNCAM transcripts. We did not ob-serve this increase in myc-taggedX�EF1 RNA injected caps (Fig. 5A–C)or X�EF1b injected caps (see Supple-mentary Figure S2). Identical resultswere obtained with non–myc-taggedconstructs (data not shown). Thesedramatic increases of the expressionof neural markers (not seen in intactembryos) is most probably due to anincreased level of FGF signaling incaps compared with the correspondingdomain in intact embryos (Delaune etal., 2005). Indeed, inhibition of FGFsignaling either by �R4 RNA, whichencodes a dominant-negative mem-brane-bound form of the FGF receptorFGFR4 (Hongo et al., 1999), or treat-ment of the explants with SU5402, apharmacological inhibitor of FGFRs(Mohammadi et al., 1998), reducedXSIP1-mediated induction of Sox2(Fig. 5D). Thus, X�EF1 and XSIP1 ap-pear to share similar regulatory prop-erties, XSIP1 being, however, a morepotent inhibitor than X�EF1 of theknown anti-neuralizing effects ofBMPs.
Both X�EF1 and XSIP1Interact With theCoactivators p300 and pCAFand the Corepressor CtBP1
As our results show that X�EF1 is lesseffective than XSIP1 to inhibit the ex-pression of BMP-dependent genes, wetested the possibility whether this dif-ferent activity is due to differentialbinding to coactivators or corepres-sors. First, we tested whether the Xe-nopus proteins, like their mammalianorthologues, can interact with thecorepressor CtBP1. Flag-tagged CtBP1and Myc-tagged XSIP1 or X�EF1 werecoexpressed in 293T cells and immu-noprecipitations (IP) on the cell ly-sates were done using anti-Flag anti-bodies. CtBP1 immunoprecipitatedXSIP1 and X�EF1 very efficiently(Fig. 6A).
Next, we tested the interaction be-tween p300 (HA-tagged) and the full-length zfhx1 proteins of both Xenopus
�EF1 AND SIP1 DURING XENOPUS EMBRYOGENESIS 1495
and mouse. Cell lysates from 293Tcells were incubated with anti-HA an-tibodies, and the interaction betweenp300-HA and XSIP1/X�EF1 was visu-alized using anti-Myc antibodies afterseparation on polyacrylamide gel elec-trophoresis (PAGE) followed by West-ern blotting. In contrast with previousreports, we observed that p300 co-im-munoprecipitated full-length X�EF1and XSIP1, the interaction withXSIP1 appearing slightly strongerthan that with X�EF1 (Fig. 6B). Thisprompted us to test pCAF binding toSIP1 proteins also. Acetylation ofCtBP-interacting proteins by pCAFhas been shown to regulate CtBP in-teraction (Zhang et al., 2000). There-fore, we decided to compare the abilityof pCAF to interact with �EF1 andSIP1 in the presence of CtBP1. Cellswere cotransfected with expressionplasmids coding for CtBP1, Flag-pCAF, and Myc-tagged X�EF1,XSIP1, m�EF1 and mSIP1, respec-tively. Cell lysates were precipitated
Fig. 4.
Fig. 5.
Fig. 4. Overexpression of X�EF1, in contrast toXSIP1, cannot induce Sox2 expression in theembryo. A,B: The 8- to 16-cell embryos in-jected into one animal blastomere with 500 pgof the indicated mRNA and analyzed for Sox2expression. Respective induction: none, n � 30for X�EF1; 100%, n � 28 for XSIP1. C–J: The16- to 32-cell embryos were injected in oneventral-most animal blastomere with the indi-cated mRNAs and analyzed for Sox2 expres-sion at stage 15. Respective induction of Sox2:0%, n � 34 (C); 0%, n � 11 (D); 0%, n �17 (E);92%, n �26 (F); 0%, n � 11 (G); 92%, n �26(H); 0%, n �22 (I); 0%, n �15 (J). A,B: Anteriorview. C–J: Ventral view, anterior toward theright. The injected area (arrow) is marked byblue staining for nuclear LacZ expression ob-tained after coinjection of LacZ RNA.
Fig. 5. X�EF1, in contrast to XSIP1, does notinduce neural markers in animal caps.A–C: Animal caps explanted at stage 9 expressSox2, Sox3, and NCAM in response to XSIP1but not to X�EF1 injection. The expression ofeach marker in uninjected animal caps and em-bryo are shown at the right of each panel. Re-spective induction 0%, n � 35 for X�EF1,100%, n � 27 for XSIP1 (A); 0 %, n � 49 forX�EF1, 97 %, n � 39 for XSIP1 (B); 14 %, n �49 for X�EF1, 94 %, n � 57 for XSIP1 (C).D: Coinjection of XSIP1 mRNA with �R4 mRNAor treatment of the explants from the time oftheir excision at blastula stage 9 with 120 �MSU5402 reduced Sox2 induction (positive caps:XSIP1, 88%, n � 27; XSIP1��FGF4r, 20%, n �29; XSIP1�SU5402, all reduced, n � 19). Con-trol embryos are viewed from the dorsal side,anterior toward the right.

with anti-Flag antibody, and the pre-cipitates were analyzed for the pres-ence of pCAF, �EF1 or SIP1, andCtBP. pCAF, like p300, was capable offorming complexes with all full-lengthSIP1 and �EF1 proteins, the interac-tion with SIP1 being much more effi-cient than that with �EF1 proteins(the X�EF1 band becoming visible af-ter longer exposure of the gel shownhere; Figure 6C, lanes 2–5). We ob-
served that CtBP is not present in thepCAF-�EF1 complexes but found thisto be true for pCAF-SIP1 complexes aswell. In the same experiment, Flag-SIP1 was used to immunoprecipitateCtBP, showing that a CtBP-SIP1 in-teraction occurs under these condi-tions (Fig. 6C, lane 6). Thus, both Xe-nopus �EF1 and SIP1 are found tointeract, although with different effi-ciency, to the corepressor CtBP and
the coactivators p300 and pCAF, theinteraction with pCAF prohibiting therecruitment of CtBP to �EF1/SIP1.
X�EF1 Binding to Smad3Requires p300
Smads are proteins that may have aninfluence on the transcriptional activ-ity of �EF1 and SIP1 as well. To fur-ther investigate on a molecular inter-action level, the differences inrepressive capacity of X�EF1 andXSIP1, we also addressed whetherthere was a difference in Smad-bind-ing capacity. SIP1 has been found tointeract with Smads 1, 2, 3, and 5, anda Smad-binding domain (SBD) suffi-cient for interaction has been mappedin SIP1 (Verschueren et al., 1999). Us-ing a yeast two-hybrid approach, wewere unable to detect interactions be-tween mouse �EF1 polypeptides andSmads 1 through 8 (van Grunsven etal., 2003), but a Smad-�EF1 complexhas been observed by other means(Postigo et al., 2003). To explain thediscrepancy between these and our re-sults, we hypothesized that �EF1 andSIP1 both can bind to Smads but notin a similar manner. In particular, wetook into account the possibility thatSIP1 would bind to Smads directly,whereas �EF1 would bind Smadsthrough p300. This would explain whyin yeast the �EF1-Smad interactioncould not be confirmed. First, we in-vestigated whether the Smad3-p300interaction was influenced by thepresence of XSIP1 or X�EF1. Expres-sion plasmids encoding Flag-Smad3,Myc-X�EF1 or Myc-XSIP1, and HA-tagged p300 were transfected in 293Tcells in the presence or absence of aconstitutively active TGF� type I re-ceptor (CA Alk4), and precipitationswere done using anti-Flag antibodies.A representative Western blot analy-sis of the proteins that coprecipitatedwith Flag-Smad3 is shown in Figure7. As shown by Pouponnot and co-workers (Pouponnot et al., 1998),p300-Smad3 interaction can be evi-denced when TGF� signaling is acti-vated but it is absent without signal-ing (Fig. 7, lanes 1 and 2). Theinteraction was enhanced whenX�EF1 or XSIP1 was coexpressed(lanes 3–4). Together with our findingthat X�EF1 and XSIP1 can both inter-act with p300 (Fig. 6B), this suggests
Fig. 6. X�EF1, like XSIP1, interacts with the coactivators p300 and pCAF and the corepressorCtBP. Plasmids encoding the indicated proteins were transfected in 293T cells and cell lysatesprocessed for immunoprecipitation with the appropriate antibodies and protein complexes (IP)separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and analyzed by Westernblotting. A portion of each lysate was analyzed for expression of transfected proteins by Westernblot (Lysate). A: CtBP can coimmunoprecipitate XSIP1 and X�EF1, immunoprecipitations weredone with anti-Flag antibodies. B: p300 can immunoprecipitate X�EF1 and XSIP1. Immunoprecipi-tations were carried out with anti-HA antibodies. C: pCAF interacts with XSIP1 and X�EF1 and thepCAF -SIP1 and pCAF-�EF1 complexes do not contain CtBP. Here, anti-Flag antibodies wereused. Note that the synthesis levels of the different Zfhx1 proteins are equal but that SIP1 forms astronger complex with pCAF than �EF1 does (compare lanes 2–5).
Fig. 7. In contrast to SIP1, �EF1-Smad3 binding requires p300. Lanes 1–2: Smad3 forms acomplex with p300 in the presence of a constitutively active TGF� receptor (CA-ALK4; lane 1).Lanes 3–4: In the presence of X�EF1 or XSIP1, this Smad3-p300 complex-formation is enhanced.Lane 6: Smad3 can coimmunoprecipitate XSIP1 in the presence of CA-ALK4. Lanes 7–8: p300cotransfection results in a gain of Smad3-X�EF1 complex, while the XSIP1-Smad3 complexformation is enhanced. We used anti-Flag antibodies in these experiments to perform the immu-noprecipitations.
�EF1 AND SIP1 DURING XENOPUS EMBRYOGENESIS 1497

that both X�EF1 and XSIP1 can sta-bilize the Smad3-p300 interaction.
The same Flag-Smad3 precipitateswere analyzed for the presence ofX�EF1 or XSIP1 in the presence orabsence of p300, and under TGF�–Smad pathway activation (Fig. 7, lane5–8, the precipitated complexes visu-alized in lanes 3–4 were loaded againin 7–8). In the absence of p300, but inthe presence of TGF� signaling, onlyXSIP1 interacted with Smad3 (com-pare lane 5 to 6). However, when p300was coexpressed, a clear interactionbetween Smad3 and full-lengthX�EF1 was also evidenced, as well asan increase in Smad3-XSIP1 interac-tion (lanes 7–8). These data indicatethat full-length X�EF1 and XSIP1 canboth interact with Smad proteins, theX�EF1–Smad interaction however be-ing one that is strictly dependent onthe presence of p300 in the complex.
DISCUSSION
The existence of families of transcrip-tion factor, like the zfhx1 family stud-ied here, does not only point out thepossibility for a different role of eachmember during development but alsodoes not exclude that their mecha-nism of action may vary. We used Xe-nopus embryos to study the expres-sion, the function, and the mechanismof action of SIP1 and �EF1 in verte-brates. We find that X�EF1 is onlyexpressed during postgastrulationstages. We demonstrate that both�EF1 and SIP1 have, to different ex-tends, the capacity to transcription-ally repress BMP-dependent genesbut only XSIP1 has strong neuralizingactivity. This activity of XSIP1 re-quires intact FGF signaling. LikeX�EF1, XSIP1 is found to bind to thecoactivators p300 and pCAF and, inaddition, and unlike XSIP1, X�EF1binding to Smad3 is strictly depen-dent on the presence of p300 in thecomplex.
In accordance with previous obser-vations made in chick and mouse (Fu-nahashi et al., 1993; Takagi et al.,1998), we find that X�EF1 is activatedin the postgastrulation embryo only,predominantly in mesodermal tissues,and later it is also expressed in thenervous system. In contrast, XSIP1 isexpressed from early gastrula onwardand is highest in the dorsal epithelial
cells (Fig. 2; Eisaki et al., 2000; vanGrunsven et al., 2000). Consequently,in contrast to the model proposed byPostigo et al. (2003), SIP1 is the onlymember of the zfhx1 family to func-tion during early Xenopus embryogen-esis in the differentiation of mesoder-mal and neural tissues.
When we analyzed the activities offull-length X�EF1 and XSIP1 pro-teins, we observed that both proteinsinhibit the expression of BMP-depen-dent genes. A difference in transcrip-tional repression capacity was ob-served when lower amounts of RNAwere injected, which could explain theinefficiency of X�EF1 to induce neuraltissue (Figs. 4, 5). Thus, these resultsdiffer from those obtained by Postigoand collaborators using human �EF1and showing that overexpression ofhuman �EF1 from injected RNA (us-ing 250–500 pg RNA) has no effect onEK expression but induces ectopicXbra expression. Therefore, these au-thors proposed a model in which �EF1and XSIP1 function as antagonists inthe control of the activation of Xbra(Postigo et al., 2003). The basis of thecontradiction between their resultsand ours is unclear. It may reflect dis-tinct specificity of action of the humanand Xenopus �EF1 proteins. It shouldbe pointed out however that the modelof an activating function of �EF1 inthe Xenopus embryo, put forth byPostigo et al. (2003), relies mainly onthe effect of a hypothetical dominant-negative form of �EF1 and on the hy-pothesis that �EF1 is expressed ear-lier during Xenopus development thanSIP1. However, the latter hypothesisis proven wrong (Fig. 2). Our observa-tions rather suggest that the two pro-teins have overlapping functions inthe embryo in those sites where theirexpression overlaps.
We observed that intact FGF signal-ing is required for XSIP1 to neuralizeectodermal cells (Fig. 4). This findingagrees with recent observations in Xe-nopus, like in amniote and ascidianembryos, that early FGF signaling be-sides BMP inhibition is required forneural induction (Linker and Stern,2004; Delaune et al., 2005). It is un-known what the FGF signaling meansfor XSIP1’s activity. FGF has beenshown to inhibit BMP activity and,hence, to promote neural developmentby phosphorylating the linker region
of Smad1 (Pera et al., 2003). Alterna-tively, FGF could influence XSIP1 ac-tivity through modification of SIP1 orthrough cooperation with other FGF-induced proteins, such as Gata andEts factors (Bertrand et al., 2003) orother factors that remain to be identi-fied.
Human �EF1 has been reported toact as an activator and repressor oftranscription, but human SIP1 wouldonly be a repressor, due to the inca-pacity of SIP1 to bind to coactivators(Postigo et al., 2003). We have not ob-served this in our experiments. Ourstudies show that both XSIP1 andX�EF1 bind to the coactivators p300and pCAF and that their ectopic ex-pression decreases BMP-dependentgene transcription. Again the basis ofthis contradiction is unclear. It shouldbe pointed out that most of the pro-moter studies performed to demon-strate the functional interaction be-tween the coactivators and �EF1 wererather artificial and that SIP1 wasnever tested in these assays (Postigoet al., 2003).
The function of pCAF or p300 bind-ing to XSIP1 may be to acetylate it,thereby leading to the release of CtBP,as has been postulated for �EF1(Postigo et al., 2003) and shown forother proteins like E1A (Zhang et al.,2000). Our results also show that bothX�EF1 and XSIP1 stabilize Smad3-p300 interaction and that the Smad3-X�EF1 interaction, but not Smad3-XSIP1 interaction, requires thepresence of p300 (Fig. 7). The molecu-lar basis of this difference is not clear.SIP1 and �EF1 proteins have quitedivergent SBD regions (Fig. 1B), andit would be interesting to determinewhether their SBD domain is respon-sible for this difference by construct-ing, for example, chimera proteins be-tween XSIP1 and X�EF1.
In conclusion, we suggest that theproposed model by Postigo and collab-orators (Postigo et al., 2003) in whichSIP1 and �EF1 function particularlyas antagonists in the regulation ofTGF� family signaling during earlyXenopus development needs to be re-defined. We rather suggest that thetwo proteins have overlapping, yetslightly different, activities in the em-bryo in those sites where their expres-sion overlaps. A way to achieve a bet-ter understanding of the action of
1498 VAN GRUNSVEN ET AL.

X�EF1 and XSIP1 on BMP/TGF� sig-naling would be to determine whethertheir differential interaction withSmads and with the p300/pCAF coac-tivators explain their difference intranscriptional repressive activity.
EXPERIMENTALPROCEDURES
Expression Plasmids
All plasmids used were cloned usingstandard cloning techniques. X�EF1(GenBank accession no. DQ319867)was cloned using degenerate primersand standard PCR techniques fromneural stage Xenopus poly(A) RNAand subsequently subcloned in pCS2in frame with 6xMyc (pCS3). TheX�EF1b clone was obtained fromOpen Biosystems (Biocat, Heidelberg,Germany) and recloned in pCS3 usingthe gateway system (Invitrogen,Merelbeke, Belgium). pCS2 with in-serts of Myc-tagged wild-type mouseSIP1 (Verschueren et al., 1999), Xeno-pus SIP1 (van Grunsven et al., 2000),and mouse �EF1 (Funahashi et al.,1993) cDNA and pCDNA3 with in-serts of Flag-tagged wild-type mouseSIP1 have been described previously(Remacle et al., 1999; Papin et al.,2002). Flag-CtBP1, Flag-Smad3,p300HA, and Flag-pCAF expressionplasmids have been described else-where (Eckner et al., 1994; Vers-chueren et al., 1999; van Grunsven etal., 2003).
Xenopus EmbryoManipulations and Injections
Xenopus embryos were obtained fromadult frogs by hormone-induced egg-laying and in vitro fertilization usingstandard methods (Sive et al., 2000)and staged according to Nieuwkoop andFaber (1997). Capped mRNAs weresynthesized in vitro by using the mMes-sage mMachine kit (Ambion, Austin,TX). mRNA was injected as described atthe indicated stage. Synthetic nuc-LacZRNA (from 100 to 500 pg) was used as alineage tracer. Initial references for thepreviously described expression con-struct used are �FGFR4 (Hongo et al.,1999) and eFGF (Delaune et al., 2005).Animal cap explants were prepared atlate blastula (stage 9) and cultured in1� Steinberg medium supplemented
with 0.1% bovine serum albumin untilearly (Sox2, Sox3) or late (NCAM) neu-rula stage. SU5402 (Calbiochem) wasdissolved in dimethyl sulfoxide (120mM), and diluted 1,000-fold in 0.1�MBS for whole-embryo treatments, andin 1� MBS for animal cap explants. Allinjections were performed at leasttwice, and the result of one representa-tive experiment is shown.
In Situ Hybridization
Embryos were fixed in MEMFA,stained for �-galactosidase activitywith 5-bromo 4-chloro-3-indolyl-�-ga-lactopyranoside (X-Gal), and pro-cessed for in situ hybridization usingdigoxigenin-labeled antisense RNAprobes according to (Sive et al., 2000).To produce the X�EF1 probe, X�EF1was cloned in PCRscript, linearizedwith NotI and transcribed with T7.Plasmids used for generating theother in situ probes were as describedpreviously: Xbra (Smith et al., 1991),EK (Jonas et al., 1985), Msx1 (Suzukiet al., 1997), Gata2 (Walmsley et al.,1994), Sox2 (Kishi et al., 2000), Sox3(Penzel et al., 1997), and NCAM(Krieg et al., 1989). For sections, em-bryos after completion of the whole-mount procedure were gelatin-embed-ded and vibratome-sectioned at 30 �mthickness.
Cell Culture, TransientTransfections, andImmunoprecipitations
All immunoprecipitations of SIP1 and�EF1 complexes, and PAGE andWestern analysis, were carried out asdescribed (van Grunsven et al., 1996)using the following buffer for lysesand at least 3 � 1 ml 10 min washingafter incubation of the cell extractswith antibody and protein A/G Sepha-rose beads: 50 mM Tris/HCl pH 7.6;170 mM NaCl; 0.1% (v/v) NP40; 50mM NaF; 2 mM Na3VO4; 0.2 mM di-thiothreitol; complete protease inhibi-tor cocktail; 10% (v/v) glycerol. Mouseanti–Myc-tag antibody and rabbit an-ti-CtBP antiserum were from SantaCruz. Anti-HA (clone 12CA5) antibod-ies were donated by Innogenetics S.A.(Zwijnaarde, Belgium). Anti-Flag an-tibodies were from SIGMA.
ACKNOWLEDGMENTSWe thank Kristin Verschueren andother members of her team for discus-sion of our results. We also thankHorst Grunz, Richard Harland, Lau-rent Kodjabachian, Roger Patient,and Thomas Sargent for providing uswith various constructs; and we aregrateful to Sadia Kricha for technicalassistance. E.B. and D.H. was fundedby the Belgian program Inter-Univer-sitary Attraction Poles from the PrimeMinister’s Office, Science Policy Pro-gramming. E.B. was also supported bythe Communaute Francaise de Bel-gique and the Fund for Scientific Med-ical Research. The work in the D.H.lab was supported by VIB, the Univer-sity of Leuven (OT/00/41), and theFund for Scientific Research-Flanders.L.A.v.G was a postdoctoral fellow fromFWO-V and V.T. a fellow of the Fondspour la Formation a la Recherchedans l�Industrie et l�Agriculture.
REFERENCES
Bertrand V, Hudson C, Caillol D, PopoviciC, Lemaire P. 2003. Neural tissue in as-cidian embryos is induced by FGF9/16/20, acting via a combination of maternalGATA and Ets transcription factors. Cell115:615–627.
Chinnadurai G. 2003. CtBP family pro-teins: more than transcriptional core-pressors. Bioessays 25:9–12.
Comijn J, Berx G, Vermassen P, Vers-chueren K, van Grunsven L, Bruyneel E,Mareel M, Huylebroeck D, van Roy F.2001. The two-handed E box binding zincfinger protein SIP1 downregulates E-cadherin and induces invasion. Mol Cell7:1267–1278.
Delaune E, Lemaire P, Kodjabachian L.2005. Neural induction in Xenopus re-quires early FGF signalling in additiontoBMPinhibition.Development132:299–310.
Eckner R, Ewen ME, Newsome D, GerdesM, DeCaprio JA, Lawrence JB, Living-ston DM. 1994. Molecular cloning andfunctional analysis of the adenovirusE1A-associated 300-kD protein (p300)reveals a protein with properties of atranscriptional adaptor. Genes Dev 8:869–884.
Eisaki A, Kuroda H, Fukui A, Asashima M.2000. XSIP1, a member of two-handedzinc finger proteins, induced anteriorneural markers in Xenopus laevis ani-mal cap. Biochem Biophys Res Commun271:151–157.
Funahashi J, Sekido R, Murai K, KamachiY, Kondoh H. 1993. Delta-crystallin en-hancer binding protein delta EF1 is azinc finger-homeodomain protein impli-cated in postgastrulation embryogenesis.Development 119:433–446.
�EF1 AND SIP1 DURING XENOPUS EMBRYOGENESIS 1499

Furusawa T, Moribe H, Kondoh H, HigashiY. 1999. Identification of CtBP1 andCtBP2 as corepressors of zinc finger-ho-meodomain factor deltaEF1. Mol CellBiol 19:8581–8590.
Genetta T, Kadesch T. 1996. Cloning of acDNA encoding a mouse transcriptionalrepressor displaying striking sequenceconservationacrossvertebrates.Gene169:289–290.
Grooteclaes ML, Frisch SM. 2000. Evi-dence for a function of CtBP in epithelialgene regulation and anoikis. Oncogene19:3823–3828.
Hill CS. 2001. TGF-beta signalling path-ways in early Xenopus development.Curr Opin Genet Dev 11:533–540.
Hongo I, Kengaku M, Okamoto H. 1999.FGF signaling and the anterior neuralinduction in Xenopus. Dev Biol 216:561–581.
Iuchi S. 2001. Three classes of C2H2 zincfinger proteins. Cell Mol Life Sci 58:625–635.
Jonas E, Sargent TD, Dawid IB. 1985. Epi-dermal keratin gene expressed in em-bryos of Xenopus laevis. Proc Natl AcadSci U S A 82:5413–5417.
Kishi M, Mizuseki K, Sasai N, YamazakiH, Shiota K, Nakanishi S, Sasai Y. 2000.Requirement of Sox2-mediated signalingfor differentiation of early Xenopus neu-roectoderm. Development 127:791–800.
Klein SL, Strausberg RL, Wagner L, Pon-tius J, Clifton SW, Richardson P. 2002.Genetic and genomic tools for Xenopusresearch: the NIH Xenopus initiative.Dev Dyn 225:384–391.
Krieg PA, Sakaguchi DS, Kintner CR.1989. Primary structure and develop-mental expression of a large cytoplasmicdomain form of Xenopus laevis neuralcell adhesion molecule (NCAM). NucleicAcids Res 17:10321–10335.
Lerchner W, Latinkic BV, Remacle JE,Huylebroeck D, Smith JC. 2000. Region-specific activation of the Xenopusbrachyury promoter involves active re-pression in ectoderm and endoderm: astudy using transgenic frog embryos. De-velopment 127:2729–2739.
Linker C, Stern CD. 2004. Neural induc-tion requires BMP inhibition only as alate step, and involves signals other thanFGF and Wnt antagonists. Development131:5671–5681.
Mohammadi M, Froum S, Hamby JM,Schroeder MC, Panek RL, Lu GH, Eli-seenkova AV, Green D, Schlessinger J,Hubbard SR. 1998. Crystal structure ofan angiogenesis inhibitor bound to theFGF receptor tyrosine kinase domain.EMBO J 17:5896–5904.
Mowat DR, Wilson MJ, Goossens M. 2003.Mowat-Wilson syndrome. J Med Genet40:305–310.
Nieuwkoop PD, Faber J. 1997. A normaltable of Xenopus laevis (Daudin). Am-sterdam: North Holland Publishing Co.
Nitta KR, Tanegashima K, Takahashi S,Asashima M. 2004. XSIP1 is essential for
early neural gene expression and neuraldifferentiation by suppression of BMPsignaling. Dev Biol 275:258–267.
Papin C, van Grunsven LA, VerschuerenK, Huylebroeck D, Smith JC. 2002. Dy-namic regulation of Brachyury expres-sion in the amphibian embryo by XSIP1.Mech Dev 111:37–46.
Penzel R, Oschwald R, Chen Y, Tacke L,Grunz H. 1997. Characterization andearly embryonic expression of a neuralspecific transcription factor xSOX3 in Xe-nopus laevis. Int J Dev Biol 41:667–677.
Pera EM, Ikeda A, Eivers E, De RobertisEM. 2003. Integration of IGF, FGF, andanti-BMP signals via Smad1 phosphory-lation in neural induction. Genes Dev17:3023–3028.
Postigo AA. 2003. Opposing functions ofZEB proteins in the regulation of theTGF{beta}/BMP signaling pathway.EMBO J 22:2443–2452.
Postigo AA, Dean DC. 1999. ZEB repressestranscription through interaction withthe corepressor CtBP. Proc Natl Acad SciU S A 96:6683–6688.
Postigo AA, Depp JL, Taylor JJ, Kroll KL.2003. Regulation of Smad signalingthrough a differential recruitment of co-activators and corepressors by ZEB pro-teins. EMBO J 22:2453–2462.
Pouponnot C, Jayaraman L, Massague J.1998. Physical and Functional Interac-tion of SMADs and p300/CBP. J BiolChem 273:22865–22868.
Remacle JE, Kraft H, Lerchner W,Wuytens G, Collart C, Verschueren K,Smith JC, Huylebroeck D. 1999. Newmode of DNA binding of multi-zinc fingertranscription factors: deltaEF1 familymembers bind with two hands to twotarget sites. EMBO J 18:5073–5084.
Sekido R, Murai K, Kamachi Y, Kondoh H.1997. Two mechanisms in the action ofrepressor deltaEF1: binding site compe-tition with an activator and active re-pression. Genes Cells 2:771–783.
Shi Y, Massague J. 2003. Mechanisms ofTGF-beta signaling from cell membraneto the nucleus. Cell 113:685–700.
Shi Y, Sawada J, Sui G, el Affar B, Whet-stine JR, Lan F, Ogawa H, Luke MP,Nakatani Y. 2003. Coordinated histonemodifications mediated by a CtBP co-re-pressor complex. Nature 422:735–738.
Sive H, Grainger RM, Harland RM. 2000.Early development of Xenopus Laevis. Alaboratory manual. Cold Spring Harbor,NY: Cold Spring Harbor, LaboratoryPress.
Smith GE, Darling DS. 2003. Combinationof a zinc finger and homeodomain re-quired for protein-interaction. Mol BiolRep 30:199–206.
Smith JC, Price BM, Green JB, Weigel D,Herrmann BG. 1991. Expression of a Xe-nopus homolog of Brachyury (T) is animmediate-early response to mesoderminduction. Cell 67:79–87.
Suzuki A, Ueno N, Hemmati-Brivanlou A.1997. Xenopus msx1 mediates epidermal
induction and neural inhibition byBMP4. Development 124:3037–3044.
Takagi T, Moribe H, Kondoh H, Higashi Y.1998. DeltaEF1, a zinc finger and home-odomain transcription factor, is requiredfor skeleton patterning in multiple lin-eages. Development 125:21–31.
Van de Putte T, Maruhashi M, Francis A,Nelles L, Kondoh H, Huylebroeck D, Hi-gashi Y. 2003. Mice lacking ZFHX1B, thegene that codes for Smad-interacting pro-tein-1, reveal a role for multiple neuralcrest cell defects in the etiology of Hirsch-sprung disease-mental retardation syn-drome. Am J Hum Genet 72:465–470.
van Grunsven LA, Billon N, Savatier P,Thomas A, Urdiales JL, Rudkin BB.1996. Effect of nerve growth factor on theexpression of cell cycle regulatory pro-teins in PC12 cells: dissection of the neu-rotrophic response from the anti-mito-genic response. Oncogene 12:1347–1356.
van Grunsven LA, Papin C, Avalosse B,Opdecamp K, Huylebroeck D, Smith JC,Bellefroid EJ. 2000. XSIP1, a Xenopuszinc finger/homeodomain encoding genehighly expressed during early neural de-velopment. Mech Dev 94:189–193.
van Grunsven LA, Schellens A, Huylebro-eck D, Verschueren K. 2001. SIP1 (Smadinteracting protein 1) and deltaEF1 (del-ta-crystallin enhancer binding factor)are structurally similar transcriptionalrepressors. J Bone Joint Surg Am 83:S40–S47.
van Grunsven LA, Michiels C, Van dePutte T, Nelles L, Wuytens G, Vers-chueren K, Huylebroeck D. 2003. Inter-action between Smad-interacting pro-tein-1 and the corepressor C-terminalbinding protein is dispensable for tran-scriptional repression of E-cadherin.J Biol Chem 278:26135–26145.
Verschueren K, Remacle JE, Collart C, KraftH, Baker BS, Tylzanowski P, Nelles L,Wuytens G, Su MT, Bodmer R, Smith JC,Huylebroeck D. 1999. SIP1, a novel zincfinger/homeodomain repressor, interactswith Smad proteins and binds to 5�-CACCT sequences in candidate targetgenes. J Biol Chem 274:20489–20498.
Wakamatsu N, Yamada Y, Yamada K, OnoT, Nomura N, Taniguchi H, Kitoh H,Mutoh N, Yamanaka T, Mushiake K,Kato K, Sonta S, Nagaya M. 2001. Mu-tations in SIP1, encoding Smad interact-ing protein-1, cause a form of Hirsch-sprung disease. Nat Genet 27:369–370.
Walmsley ME, Guille MJ, Bertwistle D,Smith JC, Pizzey JA, Patient RK. 1994.Negative control of Xenopus GATA-2 byactivin and noggin with eventual expres-sion in precursors of the ventral bloodislands. Development 120:2519–2529.
Zhang Q, Yao H, Vo N, Goodman RH. 2000.Acetylation of adenovirus E1A regulatesbinding of the transcriptional corepres-sor CtBP. Proc Natl Acad Sci U S A 97:14323–14328.
1500 VAN GRUNSVEN ET AL.
Copyright © 2022 FDOKUMEN




















