
A shared cortical bottleneck underlying Attentional Blink and Psychological Refractory Period

Response Activation in Overlapping Tasks and the Response-SelectionBottleneck
Torsten Schubert, Rico Fischer, and Christine StelzelHumboldt-University Berlin
The authors investigated the impact of response activation on dual-task performance by presenting asubliminal prime before the stimulus in Task 2 (S2) of a psychological refractory period (PRP) task.Congruence between prime and S2 modulated the reaction times in Task 2 at short stimulus onsetasynchrony despite a PRP effect. This Task 2 congruence effect was paralleled by a Task 1 congruenceeffect and emerged exclusively under conditions of cross talk, whereas it did not occur under dual-taskconditions preventing cross talk between tasks. This suggests that response activation operates during thePRP in dual tasks and affects the response times in Task 2 via cross talk between common processingelements at prebottleneck stages but not by directly affecting the postbottleneck stages.
Keywords: overlapping tasks, central bottleneck, response activation, subliminal response priming,psychological refractory period
According to an important assumption in psychology, a centralcapacity limitation of the cognitive system is the reason tworesponse-selection processes cannot be performed simultaneouslybut only serially (Pashler, 1994; Welford, 1952). When the pro-cesses in one task are engaged in response selection, the responseselection in a second task cannot start until the response selectionin the first task has finished. This is the response-selection bottle-neck (RSB) assumption that was introduced in order to explain animportant finding in dual-task research, the so-called effect of thepsychological refractory period (PRP). The PRP effect is obtainedwhen two choice reaction time (RT) tasks are carried out insuccession with a short but variable interval between them, thestimulus onset asynchrony (SOA). The effect describes the findingthat the shorter the SOA, the longer the RT in the second task(RT2) whereas RTs on the first task (RT1) are often reported to beunaffected. The RSB assumption predicts the PRP because it isassumed that Task 2 processing will be interrupted completely atthe response-selection stage until response-selection processes inTask 1 are in progress.
The present article addresses the extent to which fast-actingresponse-activation processes may operate during the PRP of aninterrupted Task 2 and whether they may affect RT2 through a
bottleneck. This question is of considerable theoretical importancebecause a positive answer might indicate that the RSB is penetra-ble for certain types of response-selection processes, and this mayrepresent a serious challenge for the RSB theory. In fact, recentdual-task studies reported evidence suggesting that response-activation processes are not delayed by a bottleneck between tasks.Many of these studies used the so-called cross talk logic by whichresearchers show an influence of the compatibility between Task 2response characteristics and Task 1 processing characteristics onRT1 (Hommel, 1998; Logan & Schulkind, 2000).
However, as will be shown later, the theoretical implications ofthese findings for the RSB theory are not yet clear because ofmethodological caveats applying to the research logic used. Herewe report the findings of four experiments in which we used aresearch logic different than the cross-talk studies. This new re-search methodology allows testing the effects of response activa-tion on Task 2 performance independently of any compatibilityrelations between the component tasks of a dual-task situation.Based on this research methodology, we tested the generalizabilityof response-activation effects in dual tasks and, more importantly,tested two main models of how response activation may affect theprocessing time of Task 2 under conditions of a presumed RSB.According to the first model, the bypass model, the results ofresponse activation are completely passing through the bottleneckdelay in Task 2 and have direct input into the bottleneck stages ofTask 2. Evidence for this model would suggest that the RSB iscompletely penetrable by the results of response activation. Suchpenetrability had been reported for response-activation processesin very specific and selected dual-task situations (e.g., Pashler,Carrier, & Hoffman, 1993) and evidence demonstrating that this isa generalizable mechanism would strongly contradict the RSBassumption.
An alternative assumption, however, presumes that even ifresponse activation affects RT2 in Task 2, the results of theresponse activation have not really passed through the entirebottleneck between tasks. Due to a mechanism that we call an
Torsten Schubert, Rico Fischer, and Christine Stelzel, Department ofPsychology, Humboldt-University Berlin, Germany.
The research described in this article was supported by German Re-search Council Grant DFG Schu 1397/ 2-1, 2-3 to Torsten Schubert. Weare grateful to Peter A. Frensch for helpful discussions, to BernhardHommel, Iring Koch, Mei-Ching Lien, Dave Meyer, Harold Pashler, andMichael Ziessler for helpful comments on earlier versions of the manu-script, and to Marina Palazova and Franziska Plessow for help in dataprocessing.
Correspondence concerning this article should be addressed to TorstenSchubert, Department of Psychology, Humboldt-University Berlin, Wolf-gang Kohler-Haus, Rudower Chaussee 18, 12489 Berlin. E-mail:[email protected]
Journal of Experimental Psychology: Copyright 2008 by the American Psychological AssociationHuman Perception and Performance2008, Vol. 34, No. 2, 376–397
0096-1523/08/$12.00 DOI: 10.1037/0096-1523.34.2.376
376

indirect influence mechanism and that is mediated by cross talkbetween tasks, response activation may evoke effects on the per-formance of the interrupted Task 2. This mechanism merely mim-ics a bypass of the bottleneck by response activation and may bereconciled with a modified RSB assumption.
The relevance of distinguishing between these alternative viewsis stressed by the fact that the RSB theory has been challenged bya number of alternative ideas. These ideas have evoked severalcontroversies in experimental psychology concerning the architec-ture of the cognitive system and, specifically, the nature of pro-cessing limitations in dual-task situations. The controversies con-cern, for example, the precise location of a capacity limitation, thatis, centrally at the response-selection stage (McCann & Johnston,1992; Pashler, 1994) or peripherally at the motor stages (Keele,1973; Meyer & Kieras, 1997; see also De Jong, 1993), the natureof the RSB, that is, structurally (Pashler, 1994) or strategically(Meyer & Kieras, 1997), or the nature of the capacity allocation(Tombu & Jolicoeur, 2003). The study reported here was notdesigned to resolve these controversies. However, compared to itscompetitors, the RSB theory is unique regarding its prediction ofserial response-selection processes, and this makes the theory mostvulnerable to phenomena like response-activation effects. There-fore, we will first relate our findings to the RSB theory and later(particularly in the General Discussion) relate them to the predic-tions of other theories about bottleneck processing in overlappingtasks, for example, the theory of strategic response deferment(SRD; Meyer & Kieras, 1997) and of capacity sharing (Tombu &Jolicoeur, 2003).
Response Activation in Dual-Task Studies
The systematic observation of response-activation effects indual tasks began with the so-called cross-talk studies in whichresearchers investigate the impact of the compatibility relationbetween tasks on the RT1 of an overlapping dual task. In a pioneerstudy of this line of research, Hommel (1998; Experiment 1)presented subjects with red or green letters, that is, a red “H” or agreen “S.” In Task 1, participants pressed a left response key whenthey were shown a red letter and a right response key when theletter was green. In Task 2, participants gave a verbal response tothe identity of the letter. They were to say “left” when the pre-sented letter was an “H,” and “right” when it was an “S.” The mainfinding of this experiment was a specific cross-talk effect, theso-called backward compatibility effect; RT1 is affected by thecompatibility between the motor responses in Task 2 (R2) andTask 1 (R1). More specifically, participants performed faster left-key presses when the response to the letter required a “left” than a“right” response. In addition to RT1, compatibility between R2and R1 affected RT2 as well, which was of little importance in theHommel study but, as will be seen later, is of special importancefor the present article. In a further experiment, Hommel (1998;Experiment 2) extended these findings to a situation in which crosstalk emerged between the lexical information of the verbal motorresponse R2 (i.e., saying “red” on “H” and “green” on “S”) and thespecific color of the stimulus in Task 1 (S1; i.e., green or red).
Based on these findings, Hommel argued that if the RSB model(Pashler, 1994; Welford, 1952) were correct, then the responseselection in Task 2 should have begun only after response selectionin Task 1 had finished and parameters of R2 could not have
affected RT1. Because there was an effect, processes involved inthe translation of stimulus information into response informationcannot have been subjected to an RSB. As an alternative, Hommel(1998) proposed a model in which the response-selection mecha-nism is not a monolithically discrete process but derives fromdifferent subprocesses that are subject to capacity limitations todifferent extents (e.g., Kornblum, Hasbroucq, & Osman, 1990).More specifically, two main subprocesses are distinguished: re-sponse activation and response identification.1 Response-activation processes are assumed not to be affected by capacitylimitations and, thus, are not delayed by the response selection inTask 1. Therefore, parameters of the Task 2 motor response mayinfluence the computation of processing parameters in Task 1 at atime when the response selection in Task 1 has not yet finished. Ifthere is compatibility between R2 and Task 1 parameters, theselection of R1 will be facilitated and RT1 will be shorter com-pared to incompatible situations. In contrast to response activation,response identification (i.e., the response verification of Kornblumet al.) is a slow acting process required for a final test of theactivated response. Serial processing of response-identificationprocesses in dual-task situations is responsible for the emergenceof the PRP effect, even though it is not clear whether seriality iscaused by a capacity limitation or for other reasons (Hommel,1998).
The assumption of different subprocesses of the response-selection mechanism and of automatic, that is, nondelayed, re-sponse activation may explain cross-talk effects obtained in anumber of studies (e.g., Hommel & Eglau, 2002; Koch & Prinz,2002; Lien & Proctor, 2000; Logan & Gordon, 2001; Logan &Schulkind, 2000). However, two caveats limit the value of theevidence about response activation as observed in cross-talk stud-ies and obscure its theoretical implications for the RSB theory.
First, the experimental situations in cross-talk studies are spe-cific in a sense that may limit the generalizability of the theory(Logan & Gordon, 2001). That is, effects of cross talk are usuallydemonstrated when dual-task situations consist of component tasksthat involve processing elements common to both tasks. For ex-ample, in Task 2 of the Hommel (1988) Experiment 2, participantshad to respond to a letter by verbalizing the name of a color. Thiscolor-naming response (R2), however, was the natural response to
1 Hommel (1998) used the term response selection instead of responseidentification and the terminology stimulus–response translation for re-sponse activation. To avoid confusion, we use response selection to de-scribe the process by which stimulus characteristics are translated intoresponse characteristics. Response identification is only one subprocess ofthe more general concept of response selection. In his model, Hommel, forthe sake of simplicity, proposed that response identification drives aresponse above its selection threshold. Thereafter, the motor response willbe initiated immediately. We share his assumption and add an elaboratedconception for the checking mechanism of the response-identificationcomponent. This conception is based on assumptions of Kornblum, Has-broucq, and Osman (1990). According to these authors, response identifi-cation (they call this response verification) uses the accumulated responseactivation as input for the checking processes to assess whether the pre-activated response is the correct one. Different checking mechanisms arepossible, that is, search in a look-out table, rule-dependent search, and soforth. When the checking process finds an incorrectly activated response,it initiates a new response-selection process that uses the describedresponse-identification mechanisms.
377RESPONSE ACTIVATION IN OVERLAPPING TASKS

the stimulus for Task 1. In Task 1, participants performed anarbitrary finger response on the color of S1 (see other cross-talkstudies for overlap at other parts of the involved task sets, Koch &Prinz, 2002; Lien & Proctor, 2000; Logan & Schulkind, 2000). Inthe particular context of overlapping tasks, the very emergence ofresponse activation might be limited to situations with informa-tional overlap between tasks (Kornblum et al., 1990). Therefore, inthe study reported here, we asked whether response activationrepresents a generalizable phenomenon and may be generated insituations where the two component tasks are completely indepen-dent of each other (see also Miller & Alderton, 2006). To that end,we designed a research methodology that, unlike the cross-talkmethodology, allows that response activation in Task 2 can beevoked and manipulated independent of any informational overlapwith Task 1 components.
The second caveat directly concerns the question of whetherresponse activation may affect RT2 through the presumed bottle-neck between tasks. By concentrating primarily on the interpreta-tion of RT1 effects, cross-talk studies do not address this importantquestion, although two principal alternatives are conceivable onthe basis of the Hommel model (see also Lien & Proctor, 2002).According to the first alternative, the bypass model, Task 2 re-sponse activation genuinely passes through the entire bottleneckand has direct access to the bottleneck stages in Task 2. In moredetail, after finishing the Task 1 bottleneck stage, the persistingvalue of the Task 2 response activation is read out by the responseidentification in Task 2 and, therefore, may directly determine theduration of Task 2 bottleneck stages through the PRP. Possibledifferences between the accumulated response activation values indifferent Task 2 conditions should, therefore, directly determinedifferences in the duration of the Task 2 response identificationand directly cause RT2 differences. In addition to that, cross talkmay emerge between simultaneously ongoing processes in Task 1and Task 2 and may cause additional backward compatibilityeffects on RT1 (e.g., Hommel, 1998).
The bypass model capitalizes on findings of earlier studies thatdemonstrated for very specific situations the principal possibilityof a complete penetration of the RSB by response activation(Pashler et al., 1993; Vince & Welford, 1967). For example,Pashler et al. (1993) reported that the activation of saccadic eyemovements is not subjected to an RSB when the saccades areperformed on a peripheral instead of a central visual cue (thestimulus in Task 2, S2) in a PRP task. Although the authors foundan SOA-dependency of RT2 in the latter condition, there was noPRP effect in the first condition, suggesting that the visual cueevoked continuously operating response activation that has di-rectly determined the processing time in Task 2. The bypass modelassumes that response activation usually operates as in the situa-tion with peripheral stimuli. The response identification proceedsserially between tasks, although in specific cases (e.g., Pashler etal.) R2 may even be executed without waiting for the final re-sponse identification.
Figure 1, however, depicts a different alternative, which we callthe indirect influence model (Lien & Proctor, 2002). According tothis model, Task 2 response activation may operate during the PRPbut, unlike in the bypass model, its results have no direct access tothe bottleneck stages in Task 2. Instead the accumulated responseactivation is reset to an initial value before the start of theresponse-identification stage in Task 2 (see Footnote 2 for a
different mechanism preventing input from activation values to theidentification stage).2 Several authors (e.g., Gilbert & Shallice,2002; Logan & Delheimer, 2001; Logan & Gordon, 2001) pro-posed a reset assumption in order to explain how interference canbe prevented, which may result from activation values persistingfrom earlier task performance on subsequent task performance. Ina PRP situation, a complete reset of Task 2 values implies that theresults of response activation have no direct access to the response-identification stage in Task 2. In that case, the parameters of thefinally executed R2 are exclusively determined by the response-identification processes, similar to a process operating as the
2 Note that a reset is distinct from a premature decay of the responseactivation, which may also prevent input of the response-activation valuesto the response identification and which bears on the possibility of achanging value of the response activation during the bottleneck delay inTask 2. According to the decay assumption, response activation simplydecays to zero before the start of the response identification during a longbottleneck delay, that is, during the PRP, although, in principle, theactivation values may directly access the identification stage of Task 2. Inthe Experiment sections, we return to the possibility of premature decay.
AP1 RA1 RI1 M1
P2 RA2 RI2 M2
P1 RA1 RI1 M1
RI2 M2RA2
RT2
RT2
B
P2
RT1
RT1
Figure 1. Indirect influence model: Processing diagram of the two taskswith the following stages and processes. RT1, RT2 � response times inTasks 1 and 2, respectively; P1 and P2 � perception stages in Tasks 1 and2, respectively; RA1 and RA2 � response activation in Tasks 1 and 2,respectively; RI1 and RI2 � response-identification processes in Tasks 1and 2, respectively; M1 and M2 � motor stages on Task 1 and 2,respectively. The diagram illustrates the assumption that the responseactivation proceeds during the interruption of Task 2 but does not directlyaffect RI2. The vertical two-headed black-colored arrows denote cross talkbetween response-activation processes in the two tasks. Panel A representsa facilitative situation where cross talk leads to a shortening of the pre-bottleneck stages (P1 and RA1) in Task 1 compared to a nonfacilitativesituation in panel B, where cross talk leads to a prolongation of theprebottleneck stages in Task 1. The change of the processing time of theTask 1 prebottleneck stages will immediately propagate into Task 2 pro-cessing time.
378 SCHUBERT, FISCHER, AND STELZEL

so-called indirect route of the response selection in the Kornblumet al. (1990) model.
In the indirect influence model, Task 2 response activation mayaffect RT2 exclusively via an indirect path that is mediated bycross talk between tasks. This is illustrated in Figure 1, whichshows a situation in which response activation operates simulta-neously in Tasks 1 and 2. During the time of simultaneous re-sponse activation, there may be cross talk in situations with infor-mational overlap between the two tasks. Response activation willbe facilitated in compatible situations when parameters in the twotasks match each other and will be slowed in incompatible situa-tions when the parameters in both tasks require contradictorycomputations. Note that response activation in Task 2 may alsoreveal cross talk with other than response-activation processes inTask 1, for example, perceptual or lexical processes (Hommel,1998, Experiment 2; Koch & Prinz, 2002). Figure 1 illustrates thatcross talk will change the duration of the prebottleneck stages inTask 1 and, consequently, of RT1. Via the bottleneck mechanism,the effects of the changed duration of the prebottleneck stages inTask 1 will propagate completely into the Task 2 processing time.Importantly, the indirect influence mechanism predicts effects onRT2 that are of the same size as the effects on RT1.
Rationale of the Present Article and the ResearchMethodology
We used a new paradigm that allows manipulating response-activation processes in Task 2 of a PRP situation independently ofany characteristics of Task 1 and investigated response-activationeffects in dual tasks. In that paradigm, we present a prime stimulusbriefly before the stimulus in Task 2 (S2) of a PRP dual-tasksituation, as shown in Figure 2. The prime should modulateresponse-activation processes tied to S2. By manipulating thecongruence between the prime and S2, we are able to manipulateresponse activation in Task 2 independently of any informational
overlap between Tasks 1 and 2. Response-activation processesshould lead to faster RT2 when prime and S2 indicate the samemotor responses in Task 2 (congruent situation) compared to asituation where they indicate different motor responses (incongru-ent situation). It is a decisive question in this paradigm whethersuch RT2 congruence effects can be observed under conditions ofshort SOA between the two tasks. The idea is that, according to theRSB assumption, the PRP effect indicates a bottleneck interruptingTask 2 at short SOA (Pashler, 1994; Welford, 1952; but see Meyer& Kieras, 1997). Thus, if we find an RT2 congruence effect atshort SOA and a PRP effect, then not all response-selection pro-cesses in Task 2 could have been interrupted by the bottleneckbetween the tasks.
Importantly, we use subliminal primes to evoke response acti-vation in Task 2. It has been shown that sensory primes influenceRTs in choice RT tasks even when they are not detectable byparticipants (Eimer, 1999; Greenwald, Klinger, & Schuh, 1995;Leuthold & Kopp, 1998; Neumann & Klotz, 1994; Vorberg,Mattler, Heinecke, Schmidt, & Schwarzbach, 2003). Several re-searchers explain these findings by the existence of direct sensory-motor links (Goodale & Milner, 1992; Klotz & Neumann, 1999).Processes evoked by the presentation of the prime lead to a directactivation of the motor responses, thereby bypassing consciouscomponents of stimulus processing and response selection. Green-wald (1992) and others (e.g., Bargh & Chartrand, 1999) regard thisas an example of basic information processing that is not affectedby capacity limitations inherent to information processing requir-ing attentional control. Here, we exploit the mechanism of sub-liminal priming in order to test whether response activation canbypass the bottleneck or not.
The use of subliminal instead of supraliminal primes was addi-tionally motivated by methodological reasons such as the contro-versy about whether the RSB is a structural (Pashler, 1994) or astrategic (Meyer & Kieras, 1997) one. In detail, Meyer and Kieras(1997) assume that the PRP effect is due not to a capacity limita-tion but to a strategic decision and that participants may use astrategy avoiding an RSB under certain task conditions. In fact,Schumacher et al. (2001) showed that under conditions of exten-sive practice, equal priority of the two tasks, and different percep-tual and response modalities, there may be no increase of the RTsin two simultaneously performed choice RT tasks compared tosingle task RTs. This, in particular, has been interpreted as anindicator for the possible avoidance of the RSB (Schumacher et al.,2001; but see Anderson, Taatgen, & Byrne, 2005; Levy & Pashler,2001; Ruthruff, Johnston, Van Selst, Whitsell, & Remington,2003, for alternative interpretations). We accounted for the debateabout the origin of an RSB by using subliminal instead of supra-liminal primes because that should avoid any conscious influenceof the priming manipulation on participants’ processing strategies.Furthermore, we created a particular dual-task situation, wheremany authors would agree on the emergence of an RSB. In detail,we stressed serial order of Task 1 and Task 2 processing, includingthe motor responses, administered a low amount of practice (rel-ative to Schumacher et al., 2001), and used the same responsemodalities (Experiments 1, 2, 4). Such particular conditions shouldnot promote processing strategies avoiding an RSB in PRP tasks(see also Meyer & Kieras, 1999, p. 55).
S1 150 ms
S2 150 ms
R2
TASK 1 TASK 2
R1
prime-S2 interval85 ms for each SOASOA
prime 34 ms
SOA 85 ms or 800 ms
Figure 2. Combination of the subliminal priming paradigm with thedual-task paradigm of the psychological refractory period. S1, S2, R1,R2 � stimuli and motor responses in Task 1 and 2, respectively. Stimulusonset asynchrony (SOA) � time between S1 (an auditory stimulus) and S2(a visual stimulus, as shown). In Task 2, the time interval between primeonset and S2 was constant (85 ms) at each SOA. A congruent situation isillustrated where prime and S2 arrows point in the same (right) direction.For the present S2, an incongruent prime would point to the left direction;a neutral prime would not point to any direction because of a cutoff at eachend (adopted from Vorberg et al. (2003).
379RESPONSE ACTIVATION IN OVERLAPPING TASKS

Predictions of the Bypass and of the Indirect InfluenceModel
The bypass and the indirect influence models make severalcritical predictions about whether an RT2 congruence effectshould emerge at short SOA depending on the specific conditionsof informational overlap between tasks. According to the bypassmodel, the results of the response-activation processes in Task 2have direct access to the bottleneck stages in Task 2. Therefore, asa critical prediction, RT2 congruence effects should emerge inde-pendently of any commonalities between tasks, that is, underconditions without informational overlap and under conditionswith informational overlap between tasks. For illustration, con-sider the specific case of the condition without informationaloverlap, as shown in Figure 3. The figure represents a so-calledpure bypass model, where the RT2 congruence effect originatesexclusively from the influence of Task 2 response activation onTask 2 bottleneck stages because different activation values haveaccess to the response identification in congruent and incongruenttrials. As can be seen, the time for successful Task 2 responseidentification is shorter in congruent trials where prime and S2activate the same R2 parameters compared to incongruent trialswhere incorrect R2 parameters are activated by the prime. In thelatter case, the contradicting information of prime and S2 infor-mation should lead to longer processing time for the response-identification stage due to iterative processing (e.g., Kornblum et
al., 1990) and, consequently, to longer RT2 than in the congruentcase. Note that, in a situation with informational overlap, theillustrated mechanism may be supplemented by cross talk at theprebottleneck stages, which may cause an additional indirect in-fluence on RT2 (Figure 1).
In the indirect influence model, however, response activationcan influence RT2 only indirectly via cross talk between commonprocessing elements of Tasks 1 and 2 (Figure 4A). Therefore, as acritical prediction, RT2 congruence effects should occur only insituations with informational overlap mediating cross talk betweentasks. In that case, the RT2 congruence effect ought to be paral-leled by an RT1 congruence effect because of the proposed prop-agation of the RT1 prebottleneck effects into RT2. For a situationwithout informational overlap, the indirect influence model pre-dicts that response activation in Task 2 will not affect RT2 at shortSOA. As can be seen in Figure 4B, although response activationproceeds for some time in parallel to the response selection of Task1, it will affect neither the Task 1 prebottleneck nor the Task 2bottleneck time and, consequently, not RT2 at a short SOA.
We will follow these critical predictions as a red line throughoutthe following experiments. However, for a conclusive assessmentof the two models, we need to specify additional hypotheses thatbear on the temporal dynamics of response activation during thebottleneck delay and on its possible influence on the congruenceeffect. This is necessary because the hypotheses of the bypass
RT1
P1 RA1 RI1 M1
P2
RT2 incongruent
SP RT2 congruency effect
in a situation without
cross-talk
P1 RA1 RI1 M1
RA2P2
prime – S2congruent
RT2 congruent
RI2 M2
RI2 M2
RA2
prime – S2incongruent
SP
Figure 3. Bypass model: Predictions about the influence of subliminal priming on response times in a dual taskunder conditions of short stimulus onset asynchrony and without informational overlap. RT1, RT2 � responsetimes in Tasks 1 and 2, respectively; P1, P2, RI1, RI2, M1, and M2 � perception, response identification, andmotor stages in Tasks 1 and 2, respectively. RA1, RA2 � response activation in Tasks 1 and 2. SP �subliminally presented prime illustrated by the triangle, S2 � stimulus in Task 2. The arrow depicting responseactivation in Task 2 is drawn through the whole psychological refractory period illustrating the assumption thatresponse activation directly affects the RI2 processes. Top: In a congruent condition, prime and S2 are associatedwith identical motor responses. The prime activates the correct motor response in Task 2; this leads to ashortening of the processing time for RI2 compared to an incongruent condition. Bottom: In an incongruentcondition the prime activates the incorrect motor response in Task 2; this leads to a prolongation of the RI2 timecompared to the congruent condition.
380 SCHUBERT, FISCHER, AND STELZEL

model hold for the specific case that different values of responseactivation are transmitted to the response identification in congru-ent and incongruent trials and this, importantly, may depend on theduration of the bottleneck delay. Note that the values of response
activation do not represent a fixed entity but change over timeduring the accumulation process (Eimer, 1999; Vorberg et al.,2003). In extreme cases, Task 2 response activation may prema-turely decay, as discussed in Footnote 2, before the start of the
RT1 (Task 2 congruent)
AAP1 RA1 RI1 M1
P2
P1 RI1 M1
RI2 M2
RT2 incongruent
P2
RI2
RA1
M2RA2
P1 RA1 RI1 M1
P2
P1 RI1 M1
RI2 M2P2
RT1 congruency effectRT1
RI2 M2SP
SP
SP
RA1
incongruentRT1 (Task 2 )(Task 2
prime – S2congruent
– S2congruent
RA2
RA2
RT2 congruency effectRT2
propagation of RT1
effects into RT2 effects
when cross- talk-
SPprime – S2incongruentprime – S2incongruent
RT1 (Task 2 congruent)
BBP1 RA1 RI1 M1
P2
RI2 M2
RT2 incongruent
P2
SP
SP
P1 RA1 RI1 M1
RI2 M2RA2
P1 RA1 RI1 M1
P2
RI2 M2
RT2
P2
SP
no congruency
effects if no cross--
talkbetween tasks
-
SPprime – S2incongruentprime – S2incongruent
prime – S2congruentprime – S2congruent
P1 RA1 RI1 M1
RI2 M2RA2
RA2
incongruentRT1 (Task 2 )(Task 2
Figure 4. Indirect influence model: Situation with short interval between both tasks. RT1, RT2 � responsetimes in Tasks 1 and 2, respectively; P1, P2, RI1, RI2, M1, M2 � perception, response-identification, and motorstages in Tasks 1 and 2, respectively; RA1, RA2 � response activation in Tasks 1 and 2, respectively. SP �subliminally presented prime as illustrated by a triangle; S2 � stimulus in Task 2. The down-sided flashillustrates the assumption that the RA2 values have no direct access to RI2. Therefore, congruence relations inTask 2 may affect RT2 exclusively via cross talk with prebottleneck stages in Task 1 and via propagation of theseeffects into RT2. Panel A: Situation with informational overlap. Cross talk between Tasks 1 and 2 is illustratedby the vertical two-headed arrows. The congruence relation between prime and S2 facilitates or prolongs RA2depending on whether both activate identical (congruent condition) or contradictory (incongruent condition)motor responses. The difference between congruent and incongruent conditions leads to RT1 congruence effectsbecause of cross talk at prebottleneck stages in Task 1. The size of the RT1 congruence effect propagates backinto Task 2, which causes an RT2 congruence effect. Panel B: Situation without informational overlap betweenTasks 1 and 2. The congruence relation between prime and S2 facilitates or prolongs RA2 depending on thecongruence condition in Task 2. However, due to the lack of cross talk, RA2 will affect neither Task 1prebottleneck nor Task 2 bottleneck processing time and, consequently, not affect RT2.
381RESPONSE ACTIVATION IN OVERLAPPING TASKS

response identification or, alternatively, it may be accumulated tothe same values in congruent and incongruent cases during bottle-neck delay. We will specify corresponding hypotheses that con-sider these possibilities for the bypass model in several parts of theExperiment sections and of the General Discussion section. Thiswill allow distinguishing modified versions of the bypass modelfrom the indirect influence model.
In all experiments, we assessed whether interindividual differ-ences in the perceptibility of the prime affected the size of the RTcongruence effects. For that purpose, we performed a prime signal-detection task after the dual-task RT part in all experiments andsubsequently correlated the corresponding d� values with the con-gruence effects. Because the results of these analyses allow ex-cluding a potentially confounding influence of the perceptibility onthe RT congruence effects, we report the methodological details ofthe prime-detection part and its findings only in the Appendix.
Experiment 1
Experiment 1 investigated subliminally evoked congruence ef-fects on RT2 in a PRP task that involved stimuli with differentmodalities (auditory S1 and visual S2) and required finger motorresponses with the left and right hand. The SOA between S1 andS2 was 85 ms (short SOA) and 800 ms (long SOA). The first aimwas to test whether the congruence relation between the subliminalprime and S2 modulates the PRP effect on RT2 at short SOAbecause this would suggest that response activation operated inTask 2 despite a bottleneck.
As a further aim, we conducted an initial test of the bypass andthe indirect influence models. For reasons of simplicity, we firstfocused on the simplest version of the bypass model, that is, thepure bypass model (Figure 3). According to that model, RT2congruence effects emerge exclusively from the influence of re-sponse activation on Task 2 bottleneck stages, and we shouldexpect that congruence affects exclusively the RTs in Task 2 (andnot in Task 1). On the contrary, an indirect influence modelpredicts that RT2 congruence effects at short SOA are necessarilyaccompanied by a similar congruence effect on RT1 because theRT2 effect results from cross talk at prebottleneck stages (seeFigure 4A and B).
In Experiment 1, cross talk was not prevented because the twocomponent tasks involved common processing elements concern-ing the spatial dimensions of the left- and right-sided motor effec-tors (R1 and R2). The spatial compatibility relations between R1and R2 may represent one source for cross talk at prebottleneckstages (Lien & Proctor, 2000, Experiments 2 and 3; 2002). Impor-tantly, because the accumulation of R2 information is itself mod-ulated by the Task 2 congruence manipulation, congruence-modulated cross talk between R1 and R2 information mayrepresent a mechanism that mediates effects of the Task 2 congru-ence manipulation on RT1.
Of course further mechanisms are conceivable that might causean effect of the Task 2 congruency manipulation on RT1 as well(i.e., note assumptions about strategic motor response grouping,Hommel, 1998; Pashler & Johnston, 1989, and others). Conse-quently, if we find a congruence effect on RT1 in addition to oneon RT2 at the short SOA, additional analyses become necessary inorder to examine its origin and to assess its consistence with thebypass or the indirect influence model.
Method
Participants. Twenty-eight students (18–35 years of age) par-ticipated in the experiment in return for payment (€8).
Experimental situation. Participants performed a dual-task RTpart and a subsequent prime signal-detection task. In the dual-taskRT part, participants performed two RT tasks together. In Task 1,they were presented with a tone of a frequency of either 350 or 900Hz. The tone was presented for 150 ms by a speaker connected toa standard personal computer. Half of the participants responded tothe low tone with a key press of the left middle finger and to thehigh tone with a key press of the left index finger on two responsebuttons located at the left side of an external keyboard. The otherhalf responded with the right index finger and the right middlefinger to the presentation of the low and high tones.
In Task 2, participants responded to a visually presented left- orright-pointing arrow presented for 150 ms (Figure 2). Half of theparticipants responded to the left-pointing arrow with a key pressof the right index finger and to the right-pointing arrow with a keypress of the right middle finger on two response buttons located atthe right side of an external keyboard. The remaining half re-sponded with the left middle and left index finger to the presen-tation of the left- and right-pointing arrows, respectively.
Procedure. At the beginning of a trial, a fixation cross waspresented at the center of the screen. After 1,000 ms, S1 waspresented for 150 ms followed, after an SOA of either 85 or 800ms, by the presentation of S2, a black colored left- or right-pointing arrow with a white inner cut-out (Figure 2). The arrowextended a visual angle of 1.09° � 3.47° and was randomlypresented 1.38° above or below the fixation cross. A prime (34 ms)was subliminally presented 85 ms before onset of S2 at the sameposition as S2. Prime stimuli were left- or right-pointing blackarrows extending a visual angle of 0.8° � 1.86°. Masking of theprime was achieved via metacontrast from the following target. Incongruent trials, prime and target arrows pointed in the samedirection, whereas in incongruent trials they pointed in oppositedirections. In neutral trials the prime did not point in any direction.The material and the time parameters for the prime and S2 stim-ulus were adopted from Vorberg et al. (2003).
Design. Variation of SOA and congruence was realized as a2 � 3 within-subjects design. The order of tones, SOAs, S2s, S2locations, and the congruence conditions varied randomly for eachparticipant in each block of trials. We presented 10 blocks of 48trials, which led to a total of 480 trials in the dual-task RT part.
Instruction. Participants were instructed to respond as quicklyand accurately as possible in both tasks and were told that the twotasks and, especially, the perception of both types of stimuli shouldbe emphasized equally. They were also informed of the order ofstimulus presentation and were asked to follow this order withtheir reactions. Participants were not informed about the existenceof subliminally presented prime stimuli.
Results
In all reported experiments, we first excluded trials with incor-rect answers in either task from the analysis of the RTs in thedual-task RT part. Then, RTs for Task 2 and Task 1 were sepa-rately analyzed with repeated measures analyses of variance(ANOVAs) including congruence (between prime and S2) andSOA as factors of central interest across all experiments. For
382 SCHUBERT, FISCHER, AND STELZEL

further clarification of the findings, we performed post hoc t testsand separate ANOVAs. For all repeated measures ANOVAs, wereport the corrected degrees of freedom with the corresponding εand corrected the p values if the sphericity assumption was notmet. Correction was performed according to the lower-boundprocedure. For error analysis, we analyzed the number of errorsseparately for each task with the same repeated measuresANOVAs as were used for the analysis of RTs.
Second reaction. As can be seen in Figure 5 (top), RT2 de-creased significantly with increasing SOA, F(1, 27) � 411.5, p �.01, suggesting the emergence of the expected PRP effect on RT2and pointing to the existence of a bottleneck in overlapping tasks.Most importantly for the present investigation, congruence af-fected RT2, F(2, 54) � 132.5, p � .01. The congruence effect wasmore pronounced at long compared to short SOA, as indicated bya significant interaction between SOA and congruence on RT2,F(2, 54) � 25.6, p � .01. A separate analysis showed that at shortSOA, RT2 was 33 ms shorter in the congruent (mean, M � 788ms) compared to the incongruent trials (M � 821 ms), t(27) �4.33, p � .01. Further multiple t tests showed that Task 2 re-sponses in neutral trials (M � 803 ms) were significantly slowercompared to congruent, t(27) � 2.4, p � .05, and significantlyfaster compared to incongruent trials, t(27) � 2.7, p � .05. Thelatter findings suggest that the subliminally presented primeevoked response-activation processes in Task 2 that were operat-ing and, more importantly, that influenced the RTs in Task 2despite the bottleneck.
First reaction. RT1 was slightly prolonged by 34 ms at shortcompared to long SOA, F(1, 27) � 9.8, p � .01. Congruencebetween prime and S2 in Task 2 affected the RT1 significantly,F(2, 54) � 4.99, p � .05. However, as can be seen in Figure 5(bottom), congruence affected RT1 at short but not at long SOA,
F(2, 54) � 3.22, p � .05. A separate analysis of the RT1 congru-ence effect, that is, the difference between RT1 incongruent andcongruent, at short SOA showed that RT1 was significantly shorterin the congruent (M � 564 ms) compared to the incongruent trials(M � 600 ms): 36 ms, t(27) � 5.9, p � .01. In addition, RT1 wasshorter in congruent compared to neutral (M � 572 ms) trials,although this effect did not reach significance, t(27) � 1.5, p �.16. In addition, RT1 was significantly shorter in the congruentcompared to the incongruent trials, t(27) � 4.1, p � .01. Thesefindings suggest that the congruence manipulation in Task 2 af-fected RT2 and RT1 almost to the same extent at short SOA.
Error analysis. The error rates for Tasks 2 and 1 are presentedin Table 1. In Task 2, congruence affected the error rate ofparticipants, F(2, 54) � 8.833, p � .01. That is, more errors werecommitted in incongruent (.07) compared to congruent (.05) andneutral (.05) trials averaged across the separate SOAs. The signif-icant interaction of SOA and congruence on error rates, F(2, 54) �17.093, p � .01, expresses the fact that the error rate was espe-cially increased in incongruent (.09) compared to congruent (.03)trials at long SOA. The specific pattern of error rates at long SOAparallels usual findings with the subliminal priming paradigm. It isconsistent with the assumption that at long SOA, participantshandle Task 2 as a single subliminal priming task. All in all, theresults of the error analysis exclude the possibility of a speed–accuracy trade off on dual-task RT performance.
In Task 1, participants committed more errors at short (.04)compared to long (.01), SOA, F(1, 27) � 89.1, p � .01, whichreflects the increased task difficulty under conditions of temporallyoverlapping task performance. In addition, the error rate wasslightly increased in the neutral (.03) compared to the incongruentcondition (.022), leading to a significant main effect of congru-ence, F(2, 54) � 3.2, p � .05. No other effects reached signifi-cance.
Further Analyses of the Congruence Effects on RT2 andRT1 at the Short SOA
In order to gain more insights into the origins of the similarcongruence effects in the two component tasks, we performedadditional analyses of the congruence effects at short SOA.
Temporal scheduling of the two motor responses. First, wetested for a possible motor grouping strategy of the participants atshort SOA. In motor grouping, participants postpone the executionof the motor response in Task 1 in order to tune it with the motorresponse in Task 2; and, therefore, manipulations changing thetime until the execution of R2 will similarly affect RT1 (Pashler &Johnston, 1989). Hommel (1998) proposed testing the possibleinfluence of response grouping on RT1 and RT2 effects by ana-lyzing the influence of the interresponse interval (IRI) between R1and R2 on the size of the experimental effect on RT1. If the RT1effect is caused by response grouping, then it should be especiallylarge at short IRIs and smaller or absent at long IRIs because agrouping tendency is expected to lead to small IRIs.
For the purpose of this analysis, we determined quartiles acrossthe distribution of IRIs (25, 50, 75, and 95%) at short SOA andcalculated the corresponding RT2 and RT1 (Figure 6). Subse-quently, we performed separate ANOVAs with the factors con-gruence and IRI quartiles on RT2 and RT1. In Task 2, the ANOVAshowed a significant effect of IRI quartiles on RT2, F(1, 27) �
SOA (in ms)
85 800
RT
(in
ms)
400
500
600
700
800RT1
400
500
600
700
800RT2 incongruent
neutralcongruent
Figure 5. Experiment 1: Mean reaction times (RT) for Task 2 (RT2) andTask 1 (RT1) depending on stimulus onset asynchrony (SOA) and con-gruence conditions in Task 2.
383RESPONSE ACTIVATION IN OVERLAPPING TASKS

59.06, p � .01, ε � 0.333, expressing the fact that the longer theIRI, the larger the RT2. As expected on the basis of the previousanalysis of mean RT2, congruence affected RT2, F(2, 54) � 19.2,p � .01. Most importantly, the effect of congruence on RT2 wasindependent of the IRI quartiles, as suggested by the nonsignificantinteraction of the two factors on RT2, F(6, 162) � 1.45, p � .19.In Task 1, there was no significant effect of IRI quartiles on RT1,F(1, 27) � 1.43, p � .20, ε � 0.333, suggesting that participantsscheduled their R1 motor responses independently of the R2 motorresponses. Congruence affected RT1, F(2, 54) � 19.06, p � .01.The interaction between congruence and IRI quartiles on RT1 wasfar from significant, F(6, 162) � 1.39, p � .20. This rules outgrouping strategy as a cause of the observed RT1 congruenceeffect at short SOA.
R1-Task 2 spatial compatibility. Next, we tested whether theobserved congruence effects at short SOA were modulated by thespatial compatibility relations between R1 and Task 2 parameters.In order to do this, we performed separate ANOVAs on RT2 andRT1 specifically at short SOA, including the factors congruenceand spatial compatibility between tasks.
R1–R2 compatibility. Spatial R1–R2 compatibility wasgiven when participants performed motor responses withspatially compatible fingers, that is, Task 1 left middle finger/
Task 2 right index finger and Task 1 left index finger/Task 2right middle finger. In Task 2, R1–R2 compatibility signifi-cantly affected RT2 at short SOA, F(1, 27) � 4.3, p � .05,replicating the findings of Hommel (1998); Lien and Proc-tor (2000), and others. RT2 was faster in compatible (M � 797ms) compared to incompatible conditions (M � 812 ms).Most importantly, however, the effects of congruence andR1–R2 compatibility on RT2 at short SOA were completelyadditive, as suggested by the lack of an interaction betweenthe two factors, F(2, 54) � 0.36, p � .65. Thus, the observedRT2 congruence effect at short SOA was just as large un-der conditions of spatially compatible and incompatible re-sponses (mean for both differences RT2 incongruent–congruent � 33 ms).
As in Task 2, R1–R2 compatibility in Task 1 significantlyaffected RT1 at short SOA, F(1, 27) � 14.5, p � .01, replicat-ing the findings of Hommel (1998); Lien and Proctor (2000)and others on backward compatibility affecting RT1. However,as before, the effects of congruence and R1–R2 compatibilitywere additive, as indicated by the lack of an interaction betweenthe two factors, F(2, 54) � 0.29, p � .75. Thus, the observedRT1 congruence effect at short SOA was as large under con-ditions of compatible motor responses (M � 35 ms) as underconditions of incompatible motor responses (M � 39 ms). Thus,the RT1 congruence effect was not modulated by R1–R2 com-patibility.
R1–Prime compatibility. Importantly, RT1 was not affectedby a direct influence of the prime in Task 2 onto the R1. Thisis so because the spatial relation between prime and R1 had nosignificant main effect on RT1 when tested independently of theR1–R2 compatibility and independently of the Task 2 congru-ence relation, F(1, 27) � 0.15, p � .90. In sum, these analysesrevealed that the RT1 congruence effect at short SOA wasindependent of any R1–Task 2 compatibility relations be-tween tasks.
Discussion
Participants responded more slowly in Task 2 at short comparedto long SOA, indicating the PRP effect and the involvement of abottleneck in Task 2. In addition, the congruence between thesubliminal prime and S2 modulated RT2 at short SOA, withshorter RT2 in congruent compared to neutral and incongruentconditions. Together, these results suggest that the subliminalprime evoked response-activation processes in Task 2 operating
Table 1Error Rates in Task 2 and 1 Depending on SOA and Congruence in Experiment 1
SOA in ms
Congruence condition
Task 2 Task 1
Congruent Neutral Incongruent Congruent Neutral Incongruent
85 0.06 0.05 0.05 0.05 0.05 0.03800 0.03 0.04 0.09 0.01 0.01 0.01
IRI Quartile
1 2 3 4
RT
(in
ms)
500
600
700
800
900
1000
1100 incongruentneutralcongruent
RT2
RT1
Figure 6. Experiment 1: Mean reaction times (RT) by quartile for Task2 (RT2) and Task 1 (RT1) depending on the interresponse intervalsbetween the motor reactions in the two tasks (IRI) and congruence condi-tions in Task 2 at a short SOA.
384 SCHUBERT, FISCHER, AND STELZEL

despite a bottleneck.3 We also obtained a congruence effect at longSOA which was larger than that at the short SOA. The resultingunderadditive SOA � Congruence interaction on RT2 is consistentwith the assumption that response activation is operating duringthe PRP and that large parts of the difference in the processingtime between congruent and incongruent trials (Vorberg et al.,2003) are absorbed into slack at short SOA (McCann & Johnston,1992; Schubert, 1999; see also the General Discussion).
A further important result is that the congruence between primeand S2 affected RT1 in addition to RT2 at short SOA. The size ofthe congruence effect on RT1 paralleled that on RT2, which isimportant for the predictions of the pure bypass and the indirectinfluence model. The indirect influence model predicts that RT2congruence effects are accompanied by similar RT1 effects be-cause of cross talk at prebottleneck stages. Therefore the presentfindings are entirely consistent with the predictions of the indirectinfluence model, although the specific origin of such a cross talkin the context of the present paradigm remains to be specified later.
On the other hand, the observation of an RT1 congruence effectcontrasts strongly with a pure bypass model predicting that re-sponse activation exclusively affects the duration of the bottleneckstages in Task 2. Several additional assumptions that might ac-count for the emergence of an RT1 congruence effect within theframework of a pure bypass model do not appear likely. Forexample, an RT1 congruence effect might emerge if a bypass ofthe bottleneck were accompanied by a simple motor groupingstrategy of the participants (Pashler & Johnston, 1989), which,however, was ruled out by the findings of a corresponding post hocanalysis. Another possibility for the emergence of an RT1 congru-ence effect accompanying a bypass of the bottleneck would be thatthe subliminal prime provided a direct influence on Task 1 re-sponse selection and/or motor execution processes. However, thefindings of the prime–R1 compatibility analysis (independent ofthe prime–S2 relation) showed no specific impact of the primestimulus in Task 2 on the processes in Task 1, which is indepen-dent of, and separable from, the congruence relation betweenprime and S2. This, conflicts with the idea of nonconscious primeinformation being automatically tied to the motor processes in theunrelated Task 1 and affecting R1 execution times directly, that is,independently of its interaction with S2 (Azuma, Prinz, & Koch,2004).
In our view, the RT1 congruence effect emerges because ofspatial cross talk between R1 selection processes and R2 response-activation processes at prebottleneck stages of the two tasks (seealso Lien & Proctor, 2000, 2002). However, we will postpone thespecification of that particular cross-talk mechanism to the Dis-cussion of Experiment 2 (and will discuss further possible expla-nations about the origin of the RT1 congruence effect in theGeneral Discussion). It is important to note that although cross talkis an essential mechanism in the indirect influence model, crosstalk is not generally precluded by the bypass model in situationswith informational overlap between tasks (except the pure bypassversion). Therefore, in Experiments 2–4, we continued testing thetwo models by focusing on predictions that allow for a moreconclusive decision between them, for example, predictions aboutthe impact of the temporal and of the informational overlap be-tween tasks on the Task 2 congruence effects.
Experiment 2
In Experiment 2, we varied the degree of the temporal overlapbetween the two tasks by manipulating the SOA in three stepsinstead of two. In addition to the short (85 ms) and long (800 ms)SOA, we included a medium SOA of 160 ms. This specificmanipulation allows distinguishing between the indirect influenceand the bypass models in the following way: According to anindirect influence model, the RT2 congruence effect results fromcross talk between response-activation processes at prebottleneckstages in the two tasks. According to Hommel (1998), the amountof cross talk diminishes with decreasing temporal overlap betweenthe tasks. This is plausible because with decreasing temporaloverlap between the two tasks, the temporal overlap of theresponse-activation processes is reduced as well. Therefore, therewill be less time available for ongoing cross talk in a situation witha small compared to a large temporal overlap. In a PRP dual-tasksituation, the temporal overlap between the two tasks and, conse-quently, between the response-activation processes is larger at ashort compared to a medium SOA. Therefore, the time for crosstalk between response-activation processes and, consequently, theamount of the resulting congruence effect should be larger in thecondition of short compared to medium SOA.
The bypass model predicts the opposite effect: Assuming thatthe size of the Task 2 congruence effect depends on the directimpact of response-activation values on the Task 2 identificationstage, the congruence effect should be smaller at a short comparedto a medium SOA. This prediction derives from findings about thetemporal dynamics of the size of the congruence effects in sub-liminal priming tasks. A number of authors have shown thatincreasing the delay between prime processing and final responseexecution leads to a severe decrease of the resulting congruenceeffect in task situations in which the prime presentation is sepa-rated from further processing (Eimer & Schlaghecken, 1998; Ei-mer, 1999 and others). In detail, with increasing delay, the size ofthe congruence effect turns from a well-pronounced positive to areversed-congruence effect (Eimer, 1999; Experiments 1 and 3).According to Eimer (1999), this decrease of the congruence effectresults from ongoing decay of the accumulated response activation
3 It might still be conceivable that the bottleneck generally interrupts anyresponse-activation processes and that Task 2 response activation is notoperating during the bottleneck delay at all. In that case, the observed RT2congruence effect may originate solely from a small portion of trials inwhich very fast Task 2 response-activation processes entered the bottle-neck before the Task 1 processes. This assumption can be tested in adistribution analysis of the RT2 congruence effects depending on responsespeed. Assuming that only the fastest Task 2 response activation may occurbefore Task 1 enters the bottleneck, Task 2 congruence effects should beespecially large the faster the responses in Task 2 and should decline withslower responses. In order to assess this possibility, we analyzed thecongruence effects separately at the .05 (fastest), .1, .25, .5, .75, .9, and .95bins of the Vincentized cumulative probabilities of the RT2 and obtainedcongruence effects of �2, 10, 22, 40, 47, 46, and 34 ms, respectively.Multiple t tests revealed significant congruence effects in all bins ( ps � .05not corrected for multiple comparisons) except that in the .05 and the .95bin, ps � .70 and .11, respectively. Certainly, the observed results do notsupport the predictions of this alternative assumption. Because this con-clusion holds true for the findings of a similar analysis in Experiment 2, wewill not focus on that argument further.
385RESPONSE ACTIVATION IN OVERLAPPING TASKS

if controlled-motor processes are delayed after subliminal primepresentation.
In the present dual-task paradigm, the bottleneck delays the timeinterval between prime representation and response execution inTask 2 and, importantly, this delay is larger at a short SOA than ata medium SOA. A potential decay of the response activationshould progress more the larger the delay between prime presen-tation and R2 execution; that is, it should progress more at a shortcompared to a medium SOA. Therefore, smaller congruence ef-fects are predicted at the short SOA than at the medium SOA in abypass model that assumes the RT2 congruence effect to resultfrom a direct impact of the persisting activation values on theidentification stage.
In sum, an indirect influence model predicts a larger congruenceeffect at short compared to medium SOA, whereas a bypass modelpredicts the opposite finding. In addition to these assumptions, it isworth mentioning that the indirect influence model necessarilypredicts that RT2 congruence effects are accompanied by similarRT1 effects.
Method
Thirty-three students (18–35 years of age) participated in thisexperiment in return for payment (€8). Participants performed adual-task RT part and a subsequent prime signal-detection task(see Appendix for details of the latter task). Design, procedure,stimulus material, initial data processing, and instruction for thedual-task RT part were identical to the corresponding part of theExperiment 1 with the following exceptions. All participants re-sponded with the left hand in Task 1 and with the right hand inTask 2. In Task 1, they responded to the low and high tones withkey presses of the left middle and left index fingers, respectively.In Task 2, participants pressed the right index finger on thepresentation of a left-pointing and the right middle finger on thepresentation of a right-pointing arrow. We presented S2 randomlyat SOAs of 85, 160, or 800 ms after the presentation of S1. Eachcombination of tones, SOAs, S2s, S2 locations, and congruenceconditions was presented in random order during blocks of 72trials. Blocks were repeated four times during the dual-task part.After each 36 trials we allowed for a break of self-paced duration.
Results
Second reaction. As can be seen in Figure 7 (top), RT2 de-creased significantly with increasing SOA, F(1, 32) � 629.0, p �.01, ε � 0.5, indicating a PRP effect. As in Experiment 1, con-gruence affected RT2, F(2, 64) � 66.38, p � .01. Participantsresponded faster on congruent compared to neutral and to incon-gruent trials. The RT2 congruence effect was significant at eachSOA, as indicated by the results of separate t tests, ts (32) � 4.5,3.02, 11.11, all ps � 0.01, for the short, medium, and large SOA,respectively. The size of the congruence effect differed at differentSOAs, as indicated by the significant interaction of SOA andcongruence on RT2, F(1, 32) � 7.14, p � .05, ε � 0.25. The mainquestion of this experiment, however, was whether the size of theRT2 congruence effect is larger at short than at medium SOA. Ascan be seen in Figure 7 (top), consistent with the prediction of theindirect influence model, the RT2 congruence effect was larger atshort (M � 43 ms) than at medium SOA (M � 24 ms), t(32) �
1.75, p � .05, one-tailed. Further separate analyses with multiplet tests showed that at short SOA, RT2 was significantly smaller inthe congruent trials (M � 815 ms) compared to the neutral trials(M � 837 ms), t(32) � 2.49, p � .05. In addition, RT2 in neutraltrials was significantly smaller than in incongruent trials (M � 858ms), t(32) � 2.64, p � .05. At medium SOA, RT2 was smaller incongruent trials (M � 763) and neutral trials (M � 762 ms) thanin the incongruent trials (M � 787 ms), ts (32) � 3.02, and 3.2,ps � 0.01, respectively.
First reaction. In Task 1, SOA did not affect RT1, F(1, 32) �0.503, p � .45, ε � 0.5. As in the previous experiment, congruenceaffected RT1, F(2, 64 � 11.31, p � .01. However, as can be seenin Figure 7 (bottom), congruence affected RT1 only at short andmedium SOAs but not at the long SOA. This is supported by thefinding of a significant interaction between congruence and SOAon RT1, F(1, 32) � 4.34, p � .05, ε � 0.25. In order to assesswhether the pattern of the congruence effect on RT1 paralleled thaton RT2, we assessed the congruence effect on RT1 at short andmiddle SOAs with separate analyses. As with RT2, the size of theRT1 congruence effect was larger at short (M � 41 ms) than atmedium SOA (M � 22 ms), t(32) � 1.78, p � .05, one-tailed.Further separate analyses with multiple t tests showed that at theshort SOA, RT1 was smaller for congruent trials (M � 590 ms)than for neutral trials (M � 614 ms), t(32) � 3.41, p � .01. RT1in neutral trials was smaller than in incongruent trials (M � 630ms), t(32) � 2.22, p � .05. At medium SOA, RT1 was smaller incongruent trials (M � 613 ms) and neutral trials (M � 612 ms)compared to incongruent trials (M � 635 ms), ts (32) � 3.02, 3.03,both ps � 0.01. Taken together, like the findings of Experiment 1,the present results suggest similar effects of the congruence ma-nipulation in Task 2 on RT2 and RT1 at short SOA and, inaddition, at the medium SOA.
SOA (in ms)
85 160 800
RT
(in
ms)
400
500
600
700
800RT1
400
500
600
700
800incongruentneutralcongruent
RT2
Figure 7. Experiment 2: Mean reaction times (RT) for Task 2 (RT2) andTask 1 (RT1) depending on stimulus onset asynchrony (SOA) and con-gruence conditions in Task 2.
386 SCHUBERT, FISCHER, AND STELZEL

Error analysis. In Task 2, congruence affected the error rate,F(2, 64) � 9.15, p � .01. Participants made more errors in incon-gruent (.07) compared to congruent (.05) and neutral (.055) trials (seeTable 2). The significant interaction of SOA and congruence on errorrates, F(1, 32) � 18.3, p � .01, ε � 0.25, expresses the fact thatparticipants had an especially large error rate in incongruent trials(.11) compared to congruent (.03) and neutral (.05) trials at the longSOA. In Task 1, participants committed more errors at short (.05) andmedium (.04) compared to long (.02) SOAs, F(1, 32) � 29.1, p � .01,ε � 0.5. No other effects reached significance.
Further Analyses of the Congruence Effects on RT2 andRT1 at Short and Medium SOAs
We again tested for a possible influence of the temporal sched-uling of the motor responses and of the spatial compatibilitybetween R1 and Task 2 characteristics on the size of the RT2 andRT1 congruence effects at short and medium SOA.
Temporal scheduling of motor responses. We determined quar-tiles across the distribution of the IRIs and calculated the correspond-ing RT2 and RT1 at short and medium SOA (see Figure 8). Subse-quently, we performed ANOVAs for RT2 and RT1 with the factorsIRI, congruence, and SOA (85 vs. 160 ms). For Task 2, the ANOVAshowed a significant effect of IRI quartiles on RT2, F(1, 32) � 62.53,p � .01, ε � 0.333, expressing the fact that the larger the IRI betweenR1 and R2, the larger RT2. IRI affected RT2 to a similar extent ateach SOA, as indicated by the nonsignificant interaction between bothfactors on RT2, F(3, 96) � 1.62, p � .15. Congruence affected RT2,F(2, 64) � 14.1, p � .01. Although, Task 2 responses were unex-pectedly fast in the neutral compared to the congruent condition atlong IRI, the size of the congruence effect was not affected by the IRIquartiles. This is shown by a nonsignificant Congruence � IRIinteraction on RT2, F(1, 32) � 2.52, p � .10, ε � 0.167. There wasalso no significant SOA � Congruence � IRI interaction, F(6,192) � 0.96, p � .45. In Task 1, there was no significant effect of IRIquartiles on RT1, F(1, 32) � 2.06, p � .15, ε � 0.333, suggesting thatparticipants scheduled their R1 motor responses independently of theR2 motor responses. As before with RT2, this pattern was similar forthe short as well as for the medium SOA, as revealed by the nonsig-nificant interaction of IRI and SOA on RT1, F(3, 96) � 1.36, p � .25.Congruence affected RT1, F(2, 64) � 13.96, p � .01. The interactionof congruence and IRI quartiles on RT1, however, was not significant,F(1, 32) � 2.14, p � .10, ε � 0.167, as was the SOA � Congru-ence � IRI interaction, F(6, 192) � 0.881, p � .35. Taken together,these results rule out the possibility that the observed congruenceeffects on RT1 at short and medium SOAs were due to a groupingstrategy.
R1–R2 spatial compatibility. We performed separateANOVAs including the factors congruence, R1–R2 compatibil-ity, and SOA (85 vs. 160 ms) on RT2 and RT1. In Task 2, themain effect of R1–R2 compatibility on RT2 approached signif-icance, F(1, 32) � 3.3, p � .07. However, the interaction ofR1–R2 compatibility and SOA significantly affected RT2, F(1,32) � 5.12, p � .05. This interaction indicates that the size ofthe compatibility effect (RT2 incompatible – compatible) de-creased from the short SOA (compatibility effect � 16 ms) tothe medium SOA (compatibility effect � 2 ms), as suggested byprevious findings of Hommel (1998). The interaction of con-gruence and compatibility did not significantly affect RT2, F(2,64) � 1.39, p � .25. The SOA � Congruence � Compatibilityinteraction was not significant, F(2, 64) � 0.15, p � .85. Thus,like the findings in Experiment 1, congruence and R1–R2compatibility additively affected RT2 at short and mediumSOAs. In Task 1, the ANOVA revealed an effect of R1–R2compatibility on RT1, F(1, 32) � 23.14, p � .01. However, theeffect of R1–R2 compatibility on RT1 was larger at the shortSOA (compatibility effect � 23 ms) than at the medium SOA(compatibility effect � 10 ms), as expressed by the significantinteraction between R1–R2 compatibility and SOA on RT1,F(1, 32) � 6.34, p � .05, (Hommel, 1998). Most importantly,the interaction of congruence and compatibility was not signif-
IRI Quartile
1 2 3 4
RT
(in
ms)
500
600
700
800
900
1000
1100 incongruentneutralcongruent RT2
RT1
Figure 8. Experiment 2: Mean reaction times (RT) by quartile for Task2 (RT2) and Task 1 (RT1) depending on the interresponse intervalsbetween the motor reactions in the two tasks (IRI) and congruence condi-tions in Task 2 averaged for short and medium SOAs.
Table 2Error Rates in Task 2 and 1 Depending on SOA and Congruence in Experiment 2
SOA in ms
Congruence condition
Task 2 Task 1
Congruent Neutral Incongruent Congruent Neutral Incongruent
85 0.06 0.06 0.05 0.05 0.06 0.05160 0.05 0.05 0.05 0.05 0.04 0.04800 0.03 0.05 0.11 0.02 0.01 0.02
387RESPONSE ACTIVATION IN OVERLAPPING TASKS

icant for RT1, F(2, 64) � 2.4, p � .09, just as the SOA �Congruence � Compatibility interaction was not, F(2, 64) �0.77, p � .45. The latter findings suggest that the congruencebetween prime and S2 and the R1–R2 compatibility additivelyaffected RT1 at short and medium SOAs, again replicating thefindings of Experiment 1.
Prime–R1 compatibility. A further ANOVA revealed that thespatial relation between the prime and R1 did not significantlyaffect RT1, F(1, 32) � 2.22, p � .14, when tested independentlyof the R1–R2 compatibility and of the congruence relation in Task2. There was also no interaction between SOA and prime–R1compatibility on RT1, F(1, 32) � 0.51, p � .47. Consequently,RT1 was not affected by a direct influence of the prime in Task 2.
Discussion
In Experiment 2, we replicated subliminally evoked congruenceeffects at a short SOA and found congruence effects at a mediumSOA. Thus, the triggering of response-activation processes bysubliminal primes despite a bottleneck is a replicable phenomenon.In addition, we replicated the finding of Experiment 1 that thecongruence manipulation in Task 2 evokes similar congruenceeffects on RT2 and RT1.
The main issue of Experiment 2 was the analysis of the size ofthe RT2 congruence effect at different SOAs. As predicted by theindirect influence model and contrary to the bypass model, the sizeof the RT2 and RT1 congruence effects decreased from the shortto the medium SOA. This is consistent with the assumption that thecongruence effect emerges from cross talk between common pro-cesses located at prebottleneck stages. Such cross talk leads to aneffect of congruence on Task 1 prebottleneck processing time thatpropagates into Task 2 processing time. With an increase of theSOA from short to medium, there is less temporal overlap betweenthe two tasks and, consequently, less time available for cross talk.This diminishes the size of the resulting congruence effect, con-trary to the predictions of the bypass model.
An important question requiring consideration concerns thepossible nature of cross talk that may have caused congruenceeffects on RT1. How can the congruence relation between theprime and S2 affect the prebottleneck processing time in Task 1and, consequently, RT1? Findings of cross-talk studies suggestthat a precondition for an influence of Task 2 parameters on Task1 processing is the existence of informational overlap betweentasks (Logan & Gordon, 2001). In the present article, informationaloverlap between tasks had been present by the left–right spatialdimensions of the manual responses R1 and R2, which may haveled to spatial cross-talk effects between the tasks (i.e., the back-ward compatibility effect; see Hommel, 1998; Koch & Prinz,2002; Lien & Proctor, 2000). Indeed, the findings of the additionalanalyses of the RT1 and RT2 effects support this assumptionbecause they show spatial R1–R2 compatibility effects for theshort SOA (and for the medium SOA in Experiment 2). Responseswere faster in spatially compatible compared to incompatibleconditions suggesting that spatial R1–R2 cross talk was operatingin the present paradigm.
However, a pure spatial R1–R2 cross-talk assumption alone is notsufficient to explain how Task 2 congruence affected RT1. As ananswer on that question, we propose the assumption of a congruence-modulated R1–R2 cross talk where the completion time for the final
R1 selection depends on the particular congruence condition in Task2. In detail, we assume that the priming manipulation in Task 2influences the accumulation time for the spatial R2 parameter that byitself interacts with the R1 information during cross talk at prebottle-neck stages. In other words, the priming manipulation in Task 2affects the time for activating the critical R2 parameter that is spatiallycompatible or incompatible to the R1 parameters and, importantly,that interacts with these R1 parameters during cross talk. Because, thecritical compatible/incompatible R2 parameter will be provided ear-lier in congruent compared to incongruent Task 2 conditions, crosstalk may be completed earlier in the first compared to the latterconditions and so will the Task 1 response-identification stage. As aconsequence, this should lead to a decrease of the RT1 under condi-tions of congruent Task 2 compared to incongruent Task 2 trials,which is consistent with the present observation of RT1 congruenceeffects at short and medium SOAs.
Note that the earlier availability of the critical R2 information in thecongruent compared to the incongruent Task 2 conditions does notcorrespond with an idea of more time for cross talk between tasks inthe present paradigm.4 If congruent conditions in Task 2 wouldprovide more time for cross talk, then one would expect an increasedsize of cross talk effects in this condition compared to that in incon-gruent ones (Hommel, 1998). This, in particular, should have led to aninteraction between congruence and R1–R2 compatibility on RT1,which, however, was not the case in Experiments 1 and 2. On thecontrary, we found spatial R1–R2 compatibility effects that were ofthe same size irrespective of the underlying Task 2 congruenceconditions. This, in particular, suggests that in congruent Task 2conditions, cross talk was not only begun earlier but was also com-pleted earlier compared to incongruent Task 2 conditions. In that case,the time for cross talk may remain constant in the two congruenceconditions, and R1–R2 compatibility effects can emerge, which are ofthe same size in congruent and incongruent Task 2 conditions.
In sum, the proposed assumption can explain the present obser-vations of an RT1 congruence effect and of a spatial R1–R2compatibility effect on RT1 with the size of the latter beingindependent of the Task 2 congruence condition. Therefore, andbecause alternative assumptions like motor grouping or a potentialdirect influence of the prime on R1 (as discussed earlier) seem notto be adequate, we assume that cross talk between R1 and R2parameters mediates the influence of the congruence relation inTask 2 on RT1. It is important to note that each part of theresponse-selection subprocesses in Task 1, that is, the responseactivation5 as well as the response-identification component, could
4 This idea was proposed by two reviewers of this article.5 It could be argued, by referring to the logic of the Additive Factor
Method (Sternberg, 1969), that the finding of additive effects betweencongruence and R1–R2 compatibility on RTs suggests separable effects ofthe two factors on separate processing stages. This may object to theassumption that cross talk was located at the response-activation stage ofboth tasks because the congruence manipulation is assumed to affect thatstage. However, the validity of this objection is not guaranteed because theAdditive Factor Method logic is reversed; that is, it predicts that if twofactor manipulations affect independent stages, then their effects on RTshould be additive. This does not allow the unequivocal deduction of thereversed conclusion, that is, if two factor manipulations have additiveeffects then they affect necessarily independent processing stages (seePrinz, 1972, and also Sternberg, 1969 for that implication).
388 SCHUBERT, FISCHER, AND STELZEL
be affected by cross talk from Task 2. Changes of the processingduration in each of these processes would lead to identical effectson RT1 and RT2 because of the propagation of the Task 1 effectsonto RT2 via the bottleneck.
Experiment 3
In Experiment 3, we tested response-activation effects in a PRPtask without informational overlap between the component tasks.For that purpose, we presented participants with the same type ofS1, S2, and prime stimuli as in Experiments 1 and 2. However,now participants performed verbal responses to the tones in Task1 by saying the German words for “low” and “high” on thepresentation of a low or high tone, respectively. In Task 2, theyperformed a manual response with the left and right index fingerson the presentation of the left- and right-pointing S2 arrows.
Although a few studies have shown that vertical and horizontalresponse dimensions may interfere with each other in single tasks (foran overview, see Proctor & Cho, 2001; Vu & Proctor, 2001), there isno evidence for such cross talk in dual-task studies combining verticalauditory–verbal and horizontal visual–manual response dimensions.Therefore, the selected combination of input and output modalitiesand response dimensions in Experiment 3 ensured minimizing infor-mational overlap between processing elements of the two tasks com-pared to the experimental conditions in Experiments 1 and 2.
The specific conditions of Experiment 3 lead to opposite pre-dictions for the indirect influence model and the bypass model.The indirect influence model predicts no congruence effects at theshort SOA on RT2 and RT1 because there are no common pro-cessing elements between tasks (see Figure 4B). In contrast, thebypass model predicts that response activation may bypass thebottleneck mechanism directly and independently of any Task 1characteristics (see Figure 3). Therefore, given the present exper-imental conditions, response activation should evoke congruenceeffects on RT2 at both short and long SOAs.
Method
Twenty-eight students (18–35 years of age) participated in returnfor payment (€8). Participants performed a dual-task RT part and asubsequent prime signal-detection task (see Appendix for details).Design, procedure, stimulus material, initial data processing, andinstruction for the dual-task RT part were identical to the correspond-ing part in Experiment 1 with the following exceptions. In Task 1,participants said “tief” and “hoch,” the German words for “low” and“high,” on the presentation of a low and high tone, respectively. InTask 2, participants pressed the left index finger on the presentation ofa left-pointing arrow and the right index finger on the presentation ofa right-pointing arrow. Each block, including 48 trials, was repeatedeight times, leading to a total of 384 trials in the dual-task RT part. Forthe subsequent prime signal-detection part, see Appendix.
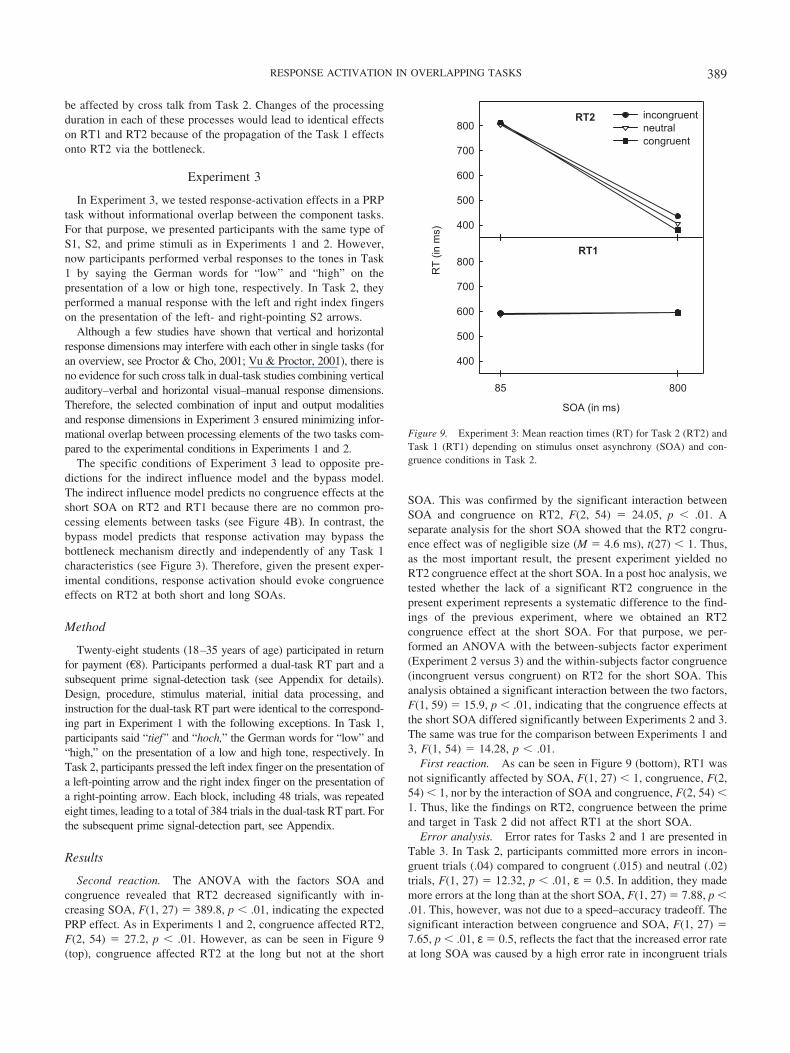
Results
Second reaction. The ANOVA with the factors SOA andcongruence revealed that RT2 decreased significantly with in-creasing SOA, F(1, 27) � 389.8, p � .01, indicating the expectedPRP effect. As in Experiments 1 and 2, congruence affected RT2,F(2, 54) � 27.2, p � .01. However, as can be seen in Figure 9(top), congruence affected RT2 at the long but not at the short
SOA. This was confirmed by the significant interaction betweenSOA and congruence on RT2, F(2, 54) � 24.05, p � .01. Aseparate analysis for the short SOA showed that the RT2 congru-ence effect was of negligible size (M � 4.6 ms), t(27) � 1. Thus,as the most important result, the present experiment yielded noRT2 congruence effect at the short SOA. In a post hoc analysis, wetested whether the lack of a significant RT2 congruence in thepresent experiment represents a systematic difference to the find-ings of the previous experiment, where we obtained an RT2congruence effect at the short SOA. For that purpose, we per-formed an ANOVA with the between-subjects factor experiment(Experiment 2 versus 3) and the within-subjects factor congruence(incongruent versus congruent) on RT2 for the short SOA. Thisanalysis obtained a significant interaction between the two factors,F(1, 59) � 15.9, p � .01, indicating that the congruence effects atthe short SOA differed significantly between Experiments 2 and 3.The same was true for the comparison between Experiments 1 and3, F(1, 54) � 14.28, p � .01.
First reaction. As can be seen in Figure 9 (bottom), RT1 wasnot significantly affected by SOA, F(1, 27) � 1, congruence, F(2,54) � 1, nor by the interaction of SOA and congruence, F(2, 54) �1. Thus, like the findings on RT2, congruence between the primeand target in Task 2 did not affect RT1 at the short SOA.
Error analysis. Error rates for Tasks 2 and 1 are presented inTable 3. In Task 2, participants committed more errors in incon-gruent trials (.04) compared to congruent (.015) and neutral (.02)trials, F(1, 27) � 12.32, p � .01, ε � 0.5. In addition, they mademore errors at the long than at the short SOA, F(1, 27) � 7.88, p �.01. This, however, was not due to a speed–accuracy tradeoff. Thesignificant interaction between congruence and SOA, F(1, 27) �7.65, p � .01, ε � 0.5, reflects the fact that the increased error rateat long SOA was caused by a high error rate in incongruent trials
SOA (in ms)
85 800
RT
(in
ms)
400
500
600
700
800RT1
400
500
600
700
800incongruentneutralcongruent
RT2
Figure 9. Experiment 3: Mean reaction times (RT) for Task 2 (RT2) andTask 1 (RT1) depending on stimulus onset asynchrony (SOA) and con-gruence conditions in Task 2.
389RESPONSE ACTIVATION IN OVERLAPPING TASKS

at this SOA (see Table 3). In Task 1, participants committed moreerrors at the short (.035) than at the long (.02) SOA, F(1, 27) �11.09, p � .01. No other effects reached significance.
Discussion
In Experiment 3, we did not obtain any congruence effects at theshort SOA on RT2 and RT1 under conditions that avoided infor-mational overlap between the component tasks of a dual-tasksituation. This result is opposed to the predictions of the bypassmodel. However, it is entirely consistent with the assumptions ofthe indirect influence model predicting that response-activationprocesses affect RT2 indirectly via cross talk at prebottleneckstages. Taken together, the findings of all three experiments areconsistent with the assumption that response activation indirectlyinfluences the processing time in Task 2 via cross talk and not bydirectly affecting the duration of the bottleneck stages in Task 2.We give further explanations for the present data in the Discussionsection for Experiment 4 and the General Discussion.
Experiment 4
Experiment 4 tests an important alternative explanation for thelack of evidence for the bypass model in the previous experiments.This alternative explanation focuses on a potential influence of thebottleneck delay in the Task 2 processing chain on the dynamics ofthe response-activation processes. In overlapping tasks, the pro-cessing chain of Task 2 will be delayed for the amount of the PRP,that is, the bottleneck delay. One could argue that the amount ofthe PRP in Experiments 1–3 was too large to reveal a direct effectof persisting response activation in Task 2 onto RT2.
This argument is plausible because the amount of responseactivation accumulated on the subliminal prime stimulus changesacross time (Eimer, 1999; Vorberg et al., 2003). After responseactivation has reached peak level, decay processes may have led toa rapid decrease of the accumulated response activation (Eimer,1999). Consequently, in case of a large PRP, there might simplynot have been enough response activation left to directly affect thebottleneck stages in Task 2. This might have obscured a directbypass of the bottleneck by response activation and would haveimplications for the interpretation of some of our previous find-ings. For example, one could assume that in Experiments 1 and 2,the response activation has indeed passed through the bottleneckand has affected the Task 2 bottleneck stages, but the residualactivation values have prematurely decayed because of a longbottleneck delay. Therefore, the effects we observed in Experi-ments 1 and 2 at short SOA may stem from the indirect influencemechanism, although a bypass had also occurred (with the effects
of the bypass being lost due to decay).6 Consequently, if theassumption of premature decay were true, then we should acceptthe validity of both models, that is, of the bypass model and of theindirect influence model, which must be integrated in a hybridmodel combining elements of both.
In order to test the premature decay assumption, we askedparticipants to perform a dual task similar to that in Experiment 1but with a much smaller PRP than in the experiments before. Theamount of the PRP can be estimated by calculating RT2 in acondition with temporal overlap minus RT2 in a condition withoutany temporal overlap (Pashler, 1994). In the previous experiments,the smallest PRP was 331 ms for the medium SOA compared tothe long SOA in Experiment 2.7 According to locus-of-slack logic(McCann & Johnston, 1992) and latent network theory (Schwe-ickert, 1983), the amount of the PRP depends on the prebottleneckand bottleneck processing time in Task 1. If this time decreases,the time for the Task 2 processes waiting for the completion ofbottleneck processes in Task 1 diminishes as well.
In Experiment 4, participants performed a simple instead of achoice reaction in Task 1. In simple RT tasks, perception anddecision stages are typically shortened (Frith & Done, 1986; Schu-bert, 1999), which leads to a dramatic decrease of the PRP whensimple RT tasks as in Task 1 are combined with choice RT tasksas in Task 2 (e.g., Karlin & Kestenbaum, 1968).
If a large PRP was indeed responsible for the lack of bypasseffects in Experiments 1–3, we should obtain an RT2 congruenceeffect at short SOA because of the expected short PRP under thepresent experimental condition. In contrast, if the indirect influ-ence model provides the more accurate explanation of the presentdata, then we do not expect congruence effects on either RT1 orRT2 at the short SOA. According to the indirect influence model,congruence effects emerge at short SOA because of cross talkbetween activation processes involving a spatial left–right re-sponse dimension. However, the possibility for such crosstalk isnot provided when performing a simple RT task as Task 1.
Method
Twenty-four students (18–35 years of age) participated in thisexperiment in return for payment (€8). Participants performed adual-task RT part and a subsequent prime signal-detection task
6 We are grateful to Mei-Ching Lien and Dave Meyer for suggesting thispossibility.
7 The PRP effects as measured by RT2 at a short SOA minus RT2 at along SOA amounted to 408, 427, and 402 ms in Experiments 1, 2, and 3,respectively.
Table 3Error Rates in Task 2 and 1 Depending on SOA and Congruence in Experiment 3
SOA in ms
Congruence condition
Task 2 Task 1
Congruent Neutral Incongruent Congruent Neutral Incongruent
85 0.01 0.02 0.02 0.03 0.04 0.04800 0.02 0.02 0.07 0.02 0.02 0.02
390 SCHUBERT, FISCHER, AND STELZEL

(see Appendix for details). Design, procedure, stimulus material,and instruction for the dual-task RT part were identical to that ofExperiment 1 with the following exceptions. In Task 1, partici-pants responded to the presence of a tone irrespective of itsfrequency. Half of the participants using their left index finger forthe tone task responded with their right index and middle finger toa left- or right-pointing arrow in Task 2. The other half of theparticipants, performing the tone task with their right hand, re-sponded with their left index and middle finger to the arrows.Participants performed 12 warm-up trials for each component taskbefore beginning the main part of the experiment. They wereinstructed not to respond prematurely to the presentation of thetone in Task 1.
Results
Second reaction. As in the previous experiments, the ANOVArevealed a decrease of RT2 with increasing SOA, F(1, 23) �173.22, p � .01, indicating a PRP effect. The size of the PRP effectas measured by RT2 at the short (491 ms) minus RT2 at the longSOA (351 ms) was 140 ms (see Figure 10, top). Congruenceaffected RT2, F(1, 23) � 154.77, p � .01, ε � 0.5. This effect waslocated at the long SOA, as confirmed by the significant interac-tion between SOA and congruence, F(1, 23) � 266.96, p � .01, ε� 0.5. A separate analysis showed a nonsignificant RT2 congru-ence effect (M � 5 ms), t(23) � 1.7, p � .08, at the short SOA. Inaddition, the size of the RT2 congruence effect at the short SOAwas far smaller than the size of the RT2 congruence effects inExperiments 1 and 2 (i.e., 33 and 42 ms, respectively). A subse-quent cross-experiment ANOVA with the between-subjects factorexperiment (Experiment 2 versus 4) and the within-subjects factorRT2 congruence effect (incongruent versus congruent) showed asignificant interaction between experiment and congruence, F(1,55) � 10.74, p � .01. Thus, in Experiment 4, the RT2 congruenceeffect at the short SOA was significantly smaller than that obtained
in Experiment 2. The same was true for the comparison betweenRT2 congruence effects in Experiments 1 and 4, F(1, 50) � 10.12,p � .01. These findings are inconsistent with the assumption thata bypass of response activation was prevented in Experiment 2because of a large PRP effect.
First reaction. RT1 was increased by 68 ms at the long com-pared to the short SOA, F(1, 23) � 90.68, p � .01, expressing aslight tendency of participants to group motor responses in Tasks1 and 2 (Pashler & Johnston, 1989). Despite this slight groupingtendency, there was no significant effect of congruence, F(1, 23) �2.14, p � .15, ε � 0.5, and no significant interaction of congruenceand SOA, F(2, 46) � 0.22, p � .64, on RT1 (Figure 10 bottom).
Error analysis. Error rates for Tasks 2 and 1 are presented inTable 4. We performed repeated measures ANOVAs including thefactors SOA and congruence for Tasks 2 and 1. In Task 2,participants committed more errors in incongruent trials (.06)compared to congruent (.02) and neutral (.02) trials, F(1, 23) �17.53, p � .01, ε � 0.5. They made more errors at the longcompared to the short SOA, F(1, 23) � 29.14, p � .01. Thesignificant interaction between congruence and SOA, F(1, 23) �19.92, p � .01, ε � 0.5, reflects the fact that the increased errorrate at long SOA was not due to a general speed–accuracy tradeoff, but was due to the particularly high error rate in incongruenttrials at the long SOA. In Task 1, participants committed slightlymore errors at the short SOA (.003) than at the long SOA (.002),F(1, 23) � 3.2, p � .08. No other effects reached significance.
Discussion
The findings of Experiment 4 allow the exclusion of an alter-native assumption for the obtained lack of bypass effects in Ex-periments 1–3. According to that assumption, the size of the PRPwas too large to allow response activation affecting the bottleneckstages in Task 2 directly through the PRP. If this were the case,then a strong decrease of the PRP in Task 2 should have led to theemergence of RT2 congruence effects in the present dual task.However, although the PRP effect in Experiment 4 was reduced tohalf of the smallest PRP effect in the previous experiments, wecould not find significant RT2 congruence effects. Therefore, thealternative assumption does not appear to be correct.
On the other hand, the present findings are consistent with theassumption of an indirect influence of the response activation onthe processing time in Task 2. In the present task situation, Task 1consisted of a simple RT involving a prespecified motor responsewith one finger. This task did not require a distinction between leftand right responses as was required in Task 2. Because the spatialresponse dimensions in the two tasks did not overlap, cross talk didnot emerge and the congruence effect could not arise.
In addition to the above mentioned decay assumption, there is afurther alternative account that should be mentioned. It is conceiv-able that a full completion of response activation during thePRP-caused delay of Task 2 may have obscured bypass effects ofthe response activation in our experiments. In more detail, re-sponse activation may have approached the same final values inincongruent trials as in congruent trials because of a prolongedtime for the accumulation of response activation for incongruenttrials during the PRP. However, if this were the case, the bypassmodel would not predict an RT2 congruence effect at short SOA.This is because the same values of response activation would have
SOA (in ms)
85 800
RT
(in
ms)
300
400
500
600RT1
300
400
500
600incongruentneutralcongruent
RT2
Figure 10. Experiment 4: Mean reaction times (RT) for Task 2 (RT2) andTask 1 (RT1), depending on stimulus onset asynchrony (SOA) and con-gruence conditions in Task 2.
391RESPONSE ACTIVATION IN OVERLAPPING TASKS

had input into the response-identification processes in incongruentand in congruent trials and this should prevent the emergence of acongruence effect. Note that this assumption, just as the decayassumption, predicts that a smaller PRP than that in Experiments1–3 should have led to the emergence of RT2 congruence effectsat short SOA. Because this was not the case in Experiment 4, weconclude that no matter which of the two mechanisms, that is,decay or full completion, a large PRP was not the reason why wedid not find evidence for a direct effect of the response activationon the bottleneck stages in Experiments 1–3.
General Discussion
Research Objectives
Findings of cross-talk studies (e.g., Hommel, 1998; Lien &Proctor, 2002) have shown that fast-acting response-activationprocesses may not be delayed by a bottleneck in overlapping dualtasks. Given the high importance of this finding for the RSBtheory, we investigated the generalizability of this phenomenonand asked how response activation may affect the processing timein Task 2 of a PRP task. For that purpose, we used a new paradigmin which a subliminal prime presented briefly before S2 modulatesresponse-activation processes related to that stimulus. As an ad-vantage, this paradigm allows testing the effects of response acti-vation on Task 2 performance independently of any compatibilityrelations between the component tasks. In this paradigm, thefinding of a congruence effect between prime and S2 on RT2 at ashort SOA is indicative for response-activation processes operat-ing in Task 2 despite a bottleneck.
Response Activation and the RSB: the Bypass and theIndirect Influence Model
Importantly, we observed RT2 congruence effects in Experi-ments 1 and 2 in different SOA conditions, including short andmedium SOAs. This specific finding suggests that not all processesassociated with the selection of motor reactions in Task 2 stopwhen response-selection processes in Task 1 are operating, whichextends previous results obtained with a different research meth-odology (Hommel, 1998; Koch & Prinz, 2002; Lien & Proctor,2002). However, as argued at the beginning of this article, differentviews are possible as to how response activation may affectprocessing times in overlapping tasks given an RSB: the bypassmodel and the indirect influence model.
Four major findings of the present experiments favor the indi-rect influence model over the bypass model. First, we observed
congruence effects on RT2 that paralleled those on RT1 in Exper-iments 1 and 2. This is consistent with the prediction of the indirectinfluence model that Task 2 response activation affects RT2 onlyvia an indirect influence mechanism and not by directly affectingthe bottleneck stages in Task 2. According to the indirect influencemodel, Task 2 response activation affects RT1 via cross talkbetween common processing elements of the component tasks.Because cross talk affects the duration of prebottleneck stages inTask 1, the effects on RT1 are propagated back into RT2 (McCann& Johnston, 1992; Schweickert, 1983), which explains the findingof similar RT2 and RT1 effects in Experiments 1 and 2. Second, aspredicted by the indirect influence model, we found RT2 and RT1congruence effects at short SOA only under conditions of infor-mational overlap between the tasks (Experiments 1 and 2) but notwhen informational overlap was precluded (Experiments 3 and 4).Importantly, this finding is in opposition to the predictions of thebypass model predicting the emergence of congruence effects evenunder conditions of no informational overlap between tasks. Third,again as predicted by the indirect influence model and contrary tothe predictions of the bypass model, we found that the size of thecongruence effects decreased with increasing SOA—short versusmedium (Hommel, 1998). And finally, a small size of the bottle-neck delay does not represent a sufficient condition for the occur-rence of bypass effects as shown by the lack of an RT2 congruenceeffect at short SOA in Experiment 4. In contrast, robust congru-ence effects were demonstrated despite a rather large bottleneckdelay under conditions of informational overlap between tasks inExperiments 1 and 2. This specific result pattern makes modifiedversions of the bypass model unlikely, according to which re-sponse activation has virtually bypassed the bottleneck but itspossible effects on RT2 have been blurred either due to prematuredecay or due to full completion of the response activation. Notethat even hybrid models combining elements of the indirect influ-ence model and of the bypass model appear to be unlikely asexplanations for the findings in Experiments 1 and 2 as long asadditional assumptions rest on premature decay conjectures.
In sum, all these findings are consistent with the indirect influ-ence model and are either not predicted by, or are even contradic-tory to, assumptions of the bypass model (see Figure 3). In addi-tion, our results do not support modified versions of the bypassmodel, which incorporate additional hypotheses about the possibleinfluence of the temporal dynamics of response activation duringthe bottleneck delay.
One could argue that some results of previous cross-talk studiesare at odds with some of the evidence reported in the presentarticle and may, therefore, object to the generalizability of the
Table 4Error Rates in Task 2 and 1 Depending on SOA and Congruence in Experiment 4
SOA in ms
Congruence condition
Task 2 Task 1
Congruent Neutral Incongruent Congruent Neutral Incongruent
85 0.02 0.01 0.01 0.003 0.005 0.0033800 0.02 0.02 0.12 0.001 0.001 0.002
Note. The errors in Task 1 are anticipatory reactions (response time on the first task � 0 ms).
392 SCHUBERT, FISCHER, AND STELZEL

indirect influence model. For example, Hommel (1998) and Loganand Schulkind (2000) reported effects of task compatibility onRT2, which, although not specifically discussed, seem numericallylarger than those on RT1. In the case of larger RT2 effects, onlypart of the RT2 compatibility effect can be explained by an impactof the indirect influence mechanism via effect propagation fromRT1 to RT2 because the latter predicts similar sizes of the RT1 andRT2 effects. Theoretically, the remaining part of the RT2 effectmay reflect some residual Task 2 activation that might havesurvived the influence of the bottleneck mechanism between tasks(e.g., see Logan & Gordon, 2001). However, the specific cross-talkmethodology remains inconclusive to such an interpretation be-cause it does not allow locating the RT2 effect unequivocally toresidual Task 1 or to Task 2 activation. It is conceivable thatforward interference between postbottleneck motor processes ofthe two tasks may be responsible for such overadditive RT2compatibility effects. For example, such interference effects mayexplain the findings of the Hommel study (Experiment 2), assum-ing that verbal-motor coding of S1 interferes in forward directionwith the verbal motor responses in Task 2. A similar mechanismmay also explain the overadditive compatibility effect on RT2 inLogan and Schulkind (2000). Therefore, the finding of a largercompatibility effect on RT2 than on RT1, as observed in somecross-talk studies, does not necessarily contradict the present con-clusion of an indirect influence mechanism.
What mechanism may hinder a bypass of the bottleneck by theresponse activation in Task 2? A mechanism preventing a genuinebypass appears when all response activation is completely reset atthe time of the start of the bottleneck stages in Task 2 (see Pashler,1993, as cited in Lien & Proctor, 2002, for a different proposal).Although based on a post hoc interpretation, the present datapattern suggests that such a potential reset of response activationseems unlikely to be of passive nature. As has been previouslydiscussed, a decay assumption cannot explain the actual pattern ofthe RT2 congruence effects at short SOA under conditions of long(Experiments 1 and 2) and of short bottleneck delays (Experiment4). Furthermore, the present findings also suggest that the resetshould not be tied to the conscious awareness of the processedstimulus information because response activation in Task 2 wasmodulated by primes that were not consciously processed by theparticipants (see Appendix).
Several models have incorporated a reset assumption in order toexplain the serial processing of actions in task contexts such asPRP dual-task (Logan & Gordon, 2001) and task-switching situ-ations (Gilbert & Shallice, 2002). In these models, the need toassume a reset of former activation values results from the theo-retical presumptions of the underlying simulation models. Forexample, in the Logan and Gordon model, a reset of the R1activation after the completed motor response (R1) is necessary inorder to avoid perseveration of R1 during Task 2 response execu-tion. In contrast, the indirect influence model suggests that Task 2response-activation values are generally reset before R2 responseidentification and that this is not specifically related to the mereavoidance of response perseveration. For example, consider Ex-periments 3 and 4, in which the risk of response perseveration wasminimized because R1 and R2 involved different response dimen-sions. Here, we did not find RT2 congruence effects at the shortSOA, which is consistent with the idea of a reset preventing directeffects of the response activation on the computational processes
in Task 2 bottleneck stages. Note that alternative assumptionsabout premature decay or a complete accumulation of activationdo not provide sufficient explanations for both the data pattern inExperiment 3 and the data pattern in Experiment 4.
Implications for the RSB Assumption
The present findings have different implications for severalsubaspects of the RSB assumption, including the aspects of thediscreteness and the seriality of the response selection and of acapacity limitation.
First of all, the finding of Task 2 response activation persistingduring response-selection processes in Task 1 is difficult to rec-oncile with the assumption of a discrete response-selection mech-anism that, as a whole, is subject to a capacity limitation (Hommel,1998; Lien & Proctor, 2000). Similar to the findings from otherresearch areas (e.g., Kornblum et al., 1990), the present resultssupport an elaborate conception of the response-selection mecha-nism incorporating the distinction between response activation andresponse-identification processes (see also Hommel, 1998). This,in particular, provides an integrated account for a wide range ofinterference phenomena obtained in single- and dual-task studies(Lien & Proctor, 2002). A challenge for future research might bethe integration of assumptions about the possible continuous char-acter of response activation and the presumed discrete type of theresponse-identification stage (but see Kornblum, Stevens,Whipple, & Requin, 1999). Miller (1988) formulated several con-straints concerning the character of the information-transformation-within and the information-transmission-betweenprocessing stages, that is, continuous or discrete. For example,assuming discrete information transformation for response identi-fication would require discrete transmission of information fromresponse activation to response identification. Cross-talk effects indual tasks, however, require that information had been exchangedbetween processing stages of different tasks. Continuous responseactivation may represent a location for that exchange, for examplein terms of coactivation between the response activation in the twotasks (Miller, 1998). However, other mechanisms are conceivable.Lien, McCann, Ruthruff, and Proctor (2005) assumed that cross-talk related information exchange occurs in an amodal response-selection stage operating on the abstract response codes, whereasresponse activation operates independently between the two com-ponent tasks.
Further, the important seriality assumption of the RSB theory isnot violated by the present findings. Although the data indicate thatresponse activation may proceed in parallel to early parts of theresponse selection in another task, they are consistent with theassumption that the response identification proceeds serially afterthe response selection in Task 1. This is so because the present dataare in accord with the assumption that the accumulated responseactivation is completely reset after the end of the response selec-tion in Task 1. A reset, however, requires that the responseidentification should operate in Task 2 after the completion ofTask 1 bottleneck processes. Note that correct R2 informationwould not systematically be available if the Task 2 responseidentification did not proceed after a reset of the accumulatedmotor information (Miller & Reynolds, 2003). In sum, the assump-tion that response identification proceeds serially between tasks isnot necessarily violated in task situations allowing response acti-
393RESPONSE ACTIVATION IN OVERLAPPING TASKS

vation to persist during the response selection of another task. Thisallows reconciling response-activation effects in dual tasks asobserved with the present paradigm and in cross-talk studies witha framework assuming a bottleneck at the final stage of theresponse selection, that is, the response identification.
Finally, the present data are not conclusive about whether theseriality of response-selection stages is caused by a capacity lim-itation (McCann & Johnston, 1992; Pashler, 1994) or by otherreasons, for example a strategic decision of the participants (Meyer& Kieras, 1997; Schumacher et al., 2001). Although this remainsa controversial issue, it is fair to say that response-activationeffects do not contradict the assumption of a capacity limitation atthe response-selection stage even though other reasons for serialscheduling are discussed (Hommel, 1998; Logan & Gordon, 2001;Meyer & Kieras, 1997).
A further issue concerns the generalizability of response-activation effects in overlapping dual tasks. In our study, we foundevidence for response activation affecting the dual-task perfor-mance despite a PRP in situations with informational overlapbetween the component tasks. This corroborates the findings ofother cross-talk studies (Logan & Gordon, 2001; Logan & Schul-kind, 2000) and raises the question of whether automatic responseactivation may generally operate in any dual-task situation orwhether the phenomenon might be specific only for situations withinformational overlap between tasks. According to some authors(Hommel, 1998; Hommel, Musseler, Aschersleben, & Prinz,2001), a motor response becomes automatically activated when-ever a stimulus is processed regarding its behavioral consequences.Other authors (Kornblum et al., 1990), however, suggested thatresponse activation occurs in selected situations only, for example,situations with overlap between the stimulus and response dimen-sions of a task. Such dimensional overlap is particularly pro-nounced in dual-task situations with informational overlap be-tween the component tasks. The present findings show thatresponse activation can be evoked in dual-task situations withoutany informational overlap between tasks. However, the effects ofresponse activation on RT2 at short SOA were only measurable insituations with informational overlap between tasks. Future re-search, therefore, will be necessary to further uncover the role ofthe task content for the occurrence and the action of responseactivation in dual tasks (Koch & Prinz, 2002).
Relation to Theories Competing With the RSB Assumption
The strategic response deferment (SRD) model (Meyer &Kieras, 1997) explains the PRP effect by the assumption of astrategic interruption of the Task 2 processing chain, which may belocated at various processing stages, that is, at the perception, theresponse-selection, or the motor stages. Although the present find-ings do not contradict these assumptions, our findings have inter-esting implications for that model. First of all, in our view, there isno reason to assume that response activation would affect thebottleneck stages in Task 2 in different ways in the case of astructural or a strategic RSB. Thus, we believe that in the case ofa strategic instead of a structural RSB, response activation wouldnot bypass the bottleneck either.
Importantly, the SRD model is able to accommodate some of thepresent findings from different theoretical perspectives than can amodel assuming a structural RSB. For example, the lack of con-gruence effects at the short SOA in Experiments 3 and 4 couldtheoretically be interpreted with the assumption of a strategicbottleneck that is located not between the response selection butbetween the motor stages of the tasks, that is, a strategic motorbottleneck. In this case, the entire response-selection processwould be performed in parallel between the two tasks without anypostponement of the response-activation and the response-identification processes in Task 2. For this case, the locus-of-slacklogic (Meyer & Kieras, 1997; McCann & Johnston, 1992; Schu-bert, 1999) predicts that any change in the duration of the responseselection in Task 2, which was caused by the congruence manip-ulation, should be absorbed into the cognitive slack at short SOA.This should result in an underadditive interaction of the congru-ence manipulation and the SOA on RT2, as would be consistentwith the findings of Experiments 3 and 4.
However, the assumption of a strategic motor bottleneck and ofentirely parallel response-selection processes is not sufficient toexplain all the data of the present article as a sole account. Forexample, it remains an open question how Task 2 congruence mayaffect RT1 and RT2 in Experiments 1 and 2 if the congruenceeffects were completely absorbed by the PRP. There is nothing inthe SRD model that allows for directly predicting these effects.Like the RSB model, the SRD model, therefore, needs additionalassumptions to explain the observation of RT1 and RT2 congru-ence effects. One way, for example, to align the present findingswith the assumption of a motor bottleneck and parallel response-selection processes would be to assume cross talk at prebottleneckstages8 and a propagation mechanism as proposed in the presentarticle. The incorporation of related assumptions seems to be aconsiderable challenge for the SRD model because its underlyingproduction-based assumptions do not, a priori, allow for informa-tion exchange between the discrete processing stages of the twotasks (Logan & Gordon, 2001).
Additionally, there are further reasons why we refrain frominterpreting the observed underadditive Congruence � SOA inter-action in terms of an absorption of the complete response-selectionstage into cognitive slack. First of all, previous studies highlightedthe role of sufficient dual-task practice for the occurrence ofstrategies avoiding an RSB. In detail, Schumacher et al. (1999)reported underadditivity between a response-selection manipula-tion and the SOA for the practice sessions 3 and 2 in a dual-tasksituation with either two manual or a manual and a verbal motor
8 Another way to reconcile the present findings with a motor bottleneckmodel is to assume that motor interference at postbottleneck stagesemerges during the execution of the manual motor responses in Experi-ments 1 and 2. However, although this assumption may explain why weobserved faster RT1 and RT2 under conditions of spatially compatiblecompared to incompatible R1 and R2, it does not explain why we foundadditional congruence effects on RT1 and RT2. Note that one cannotexplain the congruence effects by assuming that the scheduling of R1execution was tuned to the time of R2 selection. The latter assumptionwould mean that motor grouping had caused the congruence effects, andthis assumption was excluded by the IRI analysis in Experiments 1 and 2.
394 SCHUBERT, FISCHER, AND STELZEL

response (Experiments 3 and 4), respectively.9 Certainly, this ismore practice than applied in any of the present experiments.However, even more importantly, whereas Schumacher et al. ma-nipulated the duration of the response selection by manipulatingthe spatial compatibility of the stimulus–response mapping (Ex-periments 3 and 4), we used a priming manipulation to affect anearly subcomponent of the response selection. Probably, this prim-ing manipulation affects mechanisms associated with the fastactivation of a response, whereas the compatibility manipulation ofSchumacher et al. affects mainly the response selection proper(Sanders, 1980), that is, the response identification (according tothe present terminology). Therefore, we assume that, in the presentstudy, the response activation had been processed during cognitiveslack.
Yet another theory recently formulated as an alternative to theinitial RSB model assumes that limited central resources areshared between the response-selection stages instead of an all-or-none allocation between Task 1 and 2 (Tombu & Jolicoeur, 2003).Although this theory has no problems with the assumption ofparallel response-selection mechanisms, it additionally preservesthe assumption of a capacity limitation. As an important charac-teristic, the Tombu and Jolicoeur model allows for the explanationof various findings that concern an impact of Task 2 processingdifficulty on Task 1 processing time. Such an impact is explainedby assuming that, at short SOA, resources are shared between thetemporally overlapping response-selection processes of two tasks.Under conditions of a resource allocation differing from a 100%(Task 1) to 0% (Task 2) ratio, this, in particular, leads to a changedresource supply to Task 1 processes depending on the difficulty ofTask 2 processes. This in turn leads to changes of Task 1 process-ing time, which will be propagated into the processing times ofTask 2 (Tombu & Jolicoeur, 2003).
The assumption of resource sharing between response-selectionstages may explain the observed RT1 and RT2 congruence effectsas seen in Experiments 1 and 2. Assuming that incongruent trialsare more time consuming and, probably, more resource consumingthan congruent trials (Vorberg et al., 2003) allows the prediction oflarger RT1 and RT2 in incongruent compared to congruent trials.However, this assumption does not explain why we did not findcongruence effects in Experiments 3 and 4. In order to explain thedifference in findings between Experiments 1 and 2 and Experi-ments 3 and 4 with a resource-sharing account, one has to assumea different ratio for the resource allocation between experiments.However, in the study reported here, we held constant the task-emphasize instructions. Thus, a resource sharing account for theresponse-selection mechanism seems to be inadequate to solelyaccount for the present data.
Conclusion
Nonconsciously evoked response-activation processes can op-erate even when response-selection processes in Task 1 have notyet finished (but see Ansorge, 2004). This result is consistent withfindings of earlier cross-talk studies. However, whereas the impli-cations of response-activation effects for the RSB assumptioncould not conclusively be analyzed with cross-talk studies, themethodology used in the present article does allow for such ananalysis. Our findings suggest that response-activation effects inoverlapping tasks do not necessarily contradict the notion of a
capacity limitation at the response-selection stage, as proposed bythe RSB assumption. Response-time effects evoked by persistingresponse activation rather challenge the assumption of a unique,discrete response-selection mechanism that, as a whole, is subjectto a capacity limitation. The assumption of different response-selection subprocesses with response activation proceeding in par-allel between tasks while not bypassing the final response identi-fication allows a proper account of many response-activationeffects in overlapping tasks.
9 Note that Schubert (1999) could not find an underadditive interactionbetween the response-selection difficulty manipulation and SOA in thethird practice session when applying a numerosity manipulation and com-paring conditions with more than two alternatives.
References
Anderson, J. R., Taatgen, N. A., & Byrne, M. D. (2005). Learning toachieve perfect timesharing: Architectural implications of Hazeltine,Teague, and Ivry (2002). Journal of Experimental Psychology: HumanPerception and Performance, 31, 749–761.
Ansorge, U. (2004). Top-down contingencies of nonconscious primingrevealed by dual-task interference. Quarterly Journal of ExperimentalPsychology A, 57, 1123–1148.
Azuma, R., Prinz, W., & Koch, I. (2004). Dual-task slowing and the effectsof cross-task compatibility. Quarterly Journal of Experimental Psychol-ogy A, 57, 693–713.
Bargh, J., & Chartrand, T. L. (1999). The unbearable automaticity of being.American Psychologist, 54, 462–479.
De Jong, R. (1993). Multiple bottlenecks in overlapping task performance.Journal of Experimental Psychology: Human Perception and Perfor-mance, 19, 965–980.
Eimer, M. (1999). Facilitatory and inhibitory effects of masked primestimuli on motor activation and behavioral performance. Acta Psycho-logica, 101, 293–313.
Eimer, M., & Schlaghecken, F. (1998). Effects of masked stimuli on motoractivation: Behavioral and electrophysiological evidence. Journal ofExperimental Psychology: Human Perception and Performance, 24,1737–1747.
Frith, D. C., & Done, D. J. (1986). Routes to action in reaction tasks.Psychological Research, 48, 169–177.
Gilbert, S. J., & Shallice, T. (2002). Task switching: A PDP model.Cognitive Psychology, 44, 297–337.
Goodale, M. A., & Milner, D. A. (1992). Separate visual pathways forperception and action. Trends in Neuroscience, 15, 20–25.
Greenwald, A. G. (1992). Unconscious cognition claimed. American Psy-chologist, 47, 766–779.
Greenwald, A. G., Klinger, M. R., & Schuh, E. S. (1995). Activation bymarginally perceptible (“subliminal”) stimuli: Dissociation of uncon-scious from conscious cognition. Journal of Experimental Psychology:General, 124, 22–42.
Hommel, B. (1998). Automatic stimulus–response translation in dual-taskperformance. Journal of Experimental Psychology: Human Perceptionand Performance, 24, 1368–1384.
Hommel, B., & Eglau, (2002). Control of stimulus-response translation indual-task performance. Psychological Research, 66, 260–273.
Hommel, B., Musseler, J., Aschersleben, G., & Prinz, W. (2001). Thetheory of event coding (TEC): A framework for perception and actionplanning. Behavioural and Brain Sciences, 24, 849–937.
Karlin, L., & Kestenbaum, R. (1968). Effects of number of alternatives onthe psychological refractory period. Quarterly Journal of ExperimentalPsychology, 20, 167–178.
395RESPONSE ACTIVATION IN OVERLAPPING TASKS

Keele, S. (1973). Attention and human performance. Pacific Palisades, CA:Goodyear.
Klotz, W., & Neumann, O. (1999). Motor activation without consciousdiscrimination in metacontrast masking. Journal of Experimental Psy-chology: Human Perception and Performance, 25, 976–992.
Koch, I., & Prinz, W. (2002). Process interference and code overlap indual-task performance. Journal of Experimental Psychology: HumanPerception and Performance, 28, 192–201.
Kornblum, S., Hasbroucq, T., & Osman, A. (1990). Dimensional overlap:Cognitive basis for stimulus–response compatibility—a model and tax-onomy. Psychological Review, 97, 253–270.
Kornblum, S., Stevens, G. T., Whipple, A., & Requin, J. (1999). The effectof irrelevant stimuli: 1. The time course of stimulus–stimulus andstimulus–response consistency effects with Stroop-like stimuli, Simon-like tasks, and their factorial combination. Journal of ExperimentalPsychology: Human Perception and Performance, 25, 688–715.
Leuthold, H., & Kopp, B. (1998). Mechanisms of priming by maskedstimuli: Inferences from event-related brain potentials. PsychologicalScience, 9, 263–269.
Levy, J., & Pashler, H. (2001). Is dual-task slowing instruction dependent?Journal of Experimental Psychology: Human Perception and Perfor-mance, 27, 862–869.
Lien, M-C., McCann, R. S., Ruthruff, E., & Proctor, R. W. (2005).Dual-task performance with ideomotor-compatible tasks: Is the centralprocessing bottleneck intact, bypassed, or shifted in locus? Journal ofExperimental Psychology: Human Perception & Performance, 31, 122–144.
Lien, M-C., & Proctor, R. W. (2000). Multiple spatial correspondenceeffects on dual-task performance. Journal of Experimental Psychology:Human Perception and Performance, 26, 1260–1280.
Lien, M-C., & Proctor, R. W. (2002). Stimulus-response compatibility andpsychological refractory period effects: Implications for response selec-tion. Psychonomic Bulletin & Review, 9, 212–238.
Logan, G. D., & Delheimer, J. A. (2001). Parallel memory retrieval indual-task situations: II. Episodic memory. Journal of ExperimentalPsychology: Human Perception and Performance, 27, 668–685.
Logan, G. D., & Gordon, R. D. (2001). Executive control of visualattention in dual-task situations. Psychological Review, 108, 393–434.
Logan, G. D., & Schulkind, M. D. (2000). Parallel memory retrieval indual-task situations: I. Semantic memory. Journal of Experimental Psy-chology: Human Perception and Performance, 26, 1072–1090.
McCann, R. S., & Johnston, J. C. (1992). Locus of the single-channelbottleneck in dual-task interference. Journal of Experimental Psychol-ogy: Human Perception and Performance, 18, 471–484.
Meyer, D. E., & Kieras, D. E. (1997). A computational theory of executivecognitive processes and multiple-task performance: Part 1. Basic mech-anisms. Psychological Review, 104, 3–65.
Meyer, D. E., & Kieras, D. E. (1999). Precis to a practical unified theoryof cognition and action: Some lessons from EPIC computational modelsof human multiple-task performance. In D. Gopher & A. Koriat (Eds.),Attention and performance XVII. Cognitive regulation of performance:Interaction of theory and application (pp. 17–88). Cambridge, MA: MITPress.
Miller, J. (1988). Discrete and continuous models of human informationprocessing. Theoretical distinctions and empirical results. Acta Psycho-logica, 67, 191–257.
Miller, J., & Alderton, M. (2006). Backward response-level crosstalk in thepsychological refractory period paradigm. Journal of Experimental Psy-chology: Human Perception and Performance, 32, 149–165.
Miller, J., & Reynolds, A. (2003). The locus of redundant-targets andnontarget effects: Evidence from the psychological refractory periodparadigm. Journal of Experimental Psychology: Human Perception andPerformance, 29, 1126–1142.
Neumann, O., & Klotz, W. (1994). Motor responses to nonreportablemasked stimuli: Where is the limit of direct parameter specification? InC. Umilta & M. Moscovitch (Eds.), Attention and performance 15:Conscious and nonconscious information processing (pp. 123–150).Cambridge, MA: The MIT Press.
Pashler, H. (1994). Dual-task interference in simple tasks: Data and theory.Psychological Bulletin, 116, 220–244.
Pashler, H., Carrier, M., & Hoffman, J. (1993). Saccadic eye movementsand dual-task interference. Quarterly Journal of Experimental Psychol-ogy A, 46, 51–82.
Pashler, H., & Johnston, J. C. (1989). Chronometric evidence for centralpostponement in temporally overlapping tasks. Quarterly Journal ofExperimental Psychology, 41A, 19–45.
Prinz, W. (1972). Reaktionszeit-Fraktionierung durch Varianzanalyse?[Fractionation of response times by analysis of variance?] Archiv furPsychologie, 124, 240–252.
Proctor, R. W., & Cho, Y. S. (2001). The up-right/down-left advantageoccurs for both participant- and computer-paced conditions: An empir-ical observation on Adam, Boon, Baas, & Umilta (1998). Journal ofExperimental Psychology: Human Perception and Performance, 27,466–471.
Ruthruff, E., Johnston, J. C., Van Selst, M., Whitsell, S., & Remington, R.(2003). Vanishing dual-task interference after practice: Has the bottle-neck been eliminated or is it merely latent? Journal of ExperimentalPsychology: Human Perception and Performance, 29, 280–289.
Sanders, A. F. (1980). Stage analysis of reaction processes. In G. E.Stelmach & J. Requin (Eds.), Tutorials in motor behaviour (pp. 331–353). North-Holland.
Schubert, T. (1999). Processing differences between simple and choicereactions affect bottleneck localization in overlapping tasks. Journal ofExperimental Psychology: Human Perception and Performance, 25,408–425.
Schumacher, E. H., Lauber, E. J., Glass, J. M., Zurbriggen, E. L., Gmeindl,L., Kieras, D. E., & Meyer, D. E. (1999). Concurrent response-selectionprocesses in dual-task performance: Evidence for adaptive executivecontrol of task scheduling. Journal of Experimental Psychology: HumanPerception and Performance, 25, 791–814.
Schumacher, E. H., Seymour, T. L., Glass, J. M., Lauber, E. J., Kieras,D. E., & Meyer, D. E. (2001). Virtually perfect time sharing in dual-taskperformance: Uncorking the central cognitive bottleneck. PsychologicalScience, 12, 101–108.
Schweickert, R. (1983). Latent Network Theory: Scheduling of processesin sentence verification and the Stroop effect. Journal of ExperimentalPsychology: Learning, Memory, and Cognition, 9, 353–379.
Sternberg, S. (1969). The discovery of processing stages: Extensions ofDonders’ method. In W. G. Koster (Ed.), Attention and performance II(pp. 276–315). Amsterdam: North-Holland.
Tombu, M., & Jolicoeur, P. (2003). A central capacity sharing model ofdual-task performance. Journal of Experimental Psychology, HumanPerception and Performance, 29, 3–18.
Vince, M. A., & Welford, A. T. (1967). Time taken to change speed of aresponse. Nature, 213, 532–533.
Vorberg, D., Mattler, U., Heinecke, A., Schmidt, T., & Schwarzbach, J.(2003). Different time courses for visual perception and action priming.Proceedings of the National Academy of Sciences, USA, 100, 6275–6280.
Vu, K-P. L., & Proctor, R. W. (2001). Determinants of right-left andtop-bottom prevalence for two-dimensional spatial compatibility. Jour-nal of Experimental Psychology: Human Perception and Performance,27, 813–828.
Welford, A. T. (1952). The “psychological refractory period” and the timing ofhigh speed performance. British Journal of Psychology, 43, 2–19.
396 SCHUBERT, FISCHER, AND STELZEL

Appendix
Prime Signal-Detection Task
In the prime signal-detection part, participants were informed thatprimes had been subliminally presented in Task 2. Examples ofdifferent prime stimuli were shown in slow motion. Then, participantswere presented with the same trials in the same block structure as inthe dual-task RT part and participants indicated with finger responsesif the prime and S2 pointed to the same direction or not. The latteralternative included cases where prime and S2 pointed in oppositedirections and cases where S2 was preceded by a neutral prime. Incase of uncertainty, they should respond intuitively. Participants wereasked to ignore the presentation of the auditory stimulus. They usedthe same hand for the responses in the prime signal-detection part thatthey had used for Task 2 in the preceding dual-task RT part. Thisprocedure was common to all experiments with some exceptionsspecified in the local parts below. In Experiments 1 and 4, half of theparticipants used the index finger for indicating identical and themiddle finger for indicating nonidentical directions of prime and S2.The other half used the reversed mapping.
We calculated d� values and conducted regression analyses sepa-rately for each experiment according to the procedure below. For thecalculation of d� values, we regarded the yes responses on congruenttrials as hits and the yes responses on neutral trials as false alarms. Inthe regression analysis (Greenwald et al., 1995), we regressed the RT2congruence effects as the dependent variable on the d� values (inde-pendent variable) separately for each SOA. The regression analysesfocused on the resulting slope coefficients (�) and on the resultingintercepts. The analysis of � allows testing whether individual differ-ences in the perceptibility of the prime affect the amount of the RT2congruence effect. If �, that is, the slope coefficient, does not differfrom zero, the RT2 congruence effect does not depend on the per-ceptibility of the prime. In addition, the intercept in the regressionanalysis represents the value of the dependent variable when theindependent variable is equal to zero. Therefore, in the present anal-ysis, the intercept predicts the amount of the RT2 congruence effectwhen d� is equal to zero, that is, under conditions of no perceptibility.An intercept that is significantly different from zero indicates signif-icant priming even under conditions of zero prime perceptibility(Greenwald et al., 1995).
Experiment 1
Results. The mean d� value was 0.32. The slope coefficient �did not differ from zero for the short SOA, � � �6.7, t(27) ��0.29, p � .70, and for the long SOA, � � 14, t(27) � 1.6, p �.10. This suggests that the size of the RT2 congruence effect wasnot affected by prime perceptibility at either SOA. The interceptsof the regression analyses at the short and long SOAs were 32 and89, respectively. Both intercepts differed significantly from zero, ts(27) � 3.1, and 18.1, both ps � 0.01, indicating that a significantRT2 congruence effect was associated with zero perceptibility ofthe primes at both SOAs.
Prime Signal-Detection Task in Experiment 2
Method. Sixteen participants used the index finger for indicat-ing identical directions and the left middle finger for indicating
nonidentical directions of prime and S2. The remaining partici-pants used the reversed mapping. Unlike the dual-task RT part,blocks were repeated three times during the prime signal-detectionpart.
Results. The mean d� value was 0.37. The subsequent regres-sion analyses of the RT2 congruence effects on d� values revealed� values of �22, �22, and 4 at short, medium, and long SOAs,respectively. None of these � values was significantly differentfrom zero, t(32) � �1.00, p � .30, t(32) � �1.6, p � .11, t(32) �0.28, p � .75, for the short, medium, and long SOAs, respectively.Thus, the size of the RT2 congruence effect was not affected by theperceptibility of the primes at either SOA. The calculated inter-cepts were 51, 31, and 78 for short, middle, and long SOAs,respectively. They differed significantly from zero at each SOA, ts(32) � 4.58, 3.22, and 8.08, all ps � 0.01, for the short, medium,and long SOAs, respectively. This indicates that the RT2 congru-ence effect was significant at each SOA even with zero percepti-bility of the primes.
Prime Signal-Detection Task in Experiment 3
Method. Half of the participants used the left index finger andthe remaining half used the right index finger for indicating iden-tity of prime and S2.
Results. The mean d� value was 0.21. The regression coeffi-cient � did not differ from zero for the short, � � 13, t(27) � 0.5,p � .6, and for the long SOA, � � �5, t(27) � �0.2, p � .8.Again, the size of the RT2 congruence effect did not depend onprime perceptibility at either SOA. The intercepts of the regressionanalyses at the short and the long SOAs were �6 and 58, respec-tively. At short SOA, the intercept was not significantly differentfrom zero, t(27) � - 0.85, p � .4, confirming the finding of nocongruence effects at this SOA in the dual-task RT part. At thelong SOA, the intercept was significantly larger than zero, t(27) �6.41, p � .01, indicating that a significant RT2 congruence effectoccurred even when prime perceptibility equals zero.
Prime Signal-Detection Task in Experiment 4
Results. The mean d� value was 0.29. The regression coeffi-cient � did not differ from zero for the short, � � 6.9, t(23) � 1.2,p � .25, and for the long SOA, � � 4, t(23) � 0.45, p � .65. Thisshows that the size of the RT2 congruence effect did not changewith changing prime perceptibility at either SOA. The intercepts ofthe regression analyses at short and long SOA were 3 and 88,respectively. At the short SOA, the intercept was not significantlydifferent from zero, t(23) � 0.88, p � .39, confirming the findingof no congruence effect at this SOA in the dual-task RT part. Atthe long SOA, the intercept was significantly larger than zero,t(23) � 15.5, p � .01, indicating a significant RT2 congruenceeffect even under conditions of zero prime perceptibility.
Received June 28, 2005Revision received May 10, 2007
Accepted June 11, 2007 �
397RESPONSE ACTIVATION IN OVERLAPPING TASKS
Copyright © 2022 FDOKUMEN




















