
Soil organism influence on ecosystem-level processes – bypassing the ecological hierarchy?

Palaeogeography, Palaeoclimatology, Palaeoecology, 23(1978): 263--278 © Elsevier Scientific Publishing Company, Amsterdam -- Printed in The Netherlands.
D E E P - S E A I C H N O F A C I E S : M O D E R N O R G A N I S M T R A C E S ON AND IN P E L A G I C C A R B O N A T E S OF T H E W E S T E R N E Q U A T O R I A L P A C I F I C
A. A. EKDALE and W. H. BERGER
Department of Geology and Geophysics, University of Utah, Salt Lake City, Utah (U.S.A.) Scripps Institution of Oceanography, La Jolla, Calif. (U.S.A.)
(Received November 29, 1976; accepted April 4, 1977)
ABSTRACT
Ekdale, A. A. and Berger, W. H., 1978. Deep-sea ichnofacies: modern organism traces on and in pelagic carbonates of the western equatorial Pacific. Palaeogeogr., Palaeoclimatol Palaeoecol., 23: 263--278.
Modern abyssal organism traces were investigated in bottom photographs and box cores collected on Ontong Java Plateau, western equatorial Pacific, in water depths of 1597--4441 m. All the sediments were highly burrowed, white to medium brown, calcareous nannofossil ooze. Trails of echinoids and holothuroids and also surface traces resembling Glockeria were recognized in the photographs. The box cores contained specimens of Chondrites, Planolites, Skolithos, Spiroraphe, Teichichnus and ?Zoophycos. Bioturbation at this locality is intense, and so surface trails and other features appear to be destroyed following burial by the activity of infaunal burrowers.
INTRODUCTION
Macroscopic m e m b e r s of the abyssal and hadal b o t t o m fauna are rare ly preserved in anc ien t deep-sea deposi ts , because of being w i t h o u t ha rd par ts (e.g., coe len te ra tes and var ious worm s ) or because the i r CaCO3 ske le tons are dissolved a f te r dea th (e.g., e c h i n o d e r m s and molluscs) . A l m o s t all o f the deep- sea m a c r o i n v e r t e b r a t e s can p roduce t races (i.e., t racks , trails, bu r rows) in the sea b o t t o m , a l though m o s t do n o t themse lves en te r the geologic r eco rd as b o d y fossils. Such t race fossils, in c o n j u n c t i o n with l i thologic and strati- graphic i n f o r m a t i o n , can he lp us recognize depos i t iona l cond i t ions in deep- sea depos i t s on land and unde r the sea. This pape r presen ts a b r ie f desc r ip t ion of the m o d e r n deep-sea ichnofacies a t a selected local i ty in the wes te rn equa tor ia l Pacific Ocean.
We s tud ied t races of m o d e r n abyssal organisms in pelagic c a r b o n a t e sedi- men t s co l lec ted dur ing April and May, 1975, on the Euryd ice E x p e d i t i o n o f Scripps Ins t i t u t ion o f O c e a n o g r a p h y to O n t o n g Java Plateau, n o r t h e a s t o f the S o l o m o n Is lands in the wes te rn equa to r i a l Pacific. Tracks and trails p r o d u c e d on the sea f loor b y ep i fauna l animals were scru t in ized in m o r e t h a n 200
264
TABLE I
Leg 9 Eurydice box core samples
Sample Depth (m) Latitude Longitude
77BX 3585 4°51.1 'N 156°37.0'E 79BX 2766 2°47.1~N 156°13.8'E 83 BX 2343 1°24.1'N 157°18.6'E 86BX 1929 0 ° 0.8'N 155°52.1'E 87BX 1930 0 ° 0.7'N 155°51.8'E 88BX 1923 0 ° 2.9'S 155°52.0'E 92BX 1597 2°13.5'S 156°59.9'E
102BX 2300 3°36.3'S 161°19.1'E 108BX 3383 1°44.7'S 160°48.4'E 112BX 2183 1°37.5'S 159°14.1'E 120BX 2247 0 ° 0.9'S 158°41.6'E 123BX 2949 0 ° 1.3'S 160°24.9'E 125BX 3368 0 ° 0.2'S 160°59.9'E 128BX 3732 0 ° 0.2'S 161°25.5fE 129BX 4169 0 ° 0.3'S 161°58.5'E 131BX 4441 0 ° 1.5'S 162°58.5'E 135BX 3509 0 ° 52.5'N 160 ° 59.6'E 136BX 3848 1 ° 6.0'N 161°36.3'E 139BX 4118 1 ° 21.8'N 162° 23.6'E 141 BX 4325 2 ° 21.7'N 163~42.3'E
bottom photographs. Burrows produced within the sediment by infauna were studied in 20 box cores that were retrieved from the plateau and its flanks in water depths ranging from 1597 to 4441 m (Table I).
The box cores were collected with the corer of Hessler and Jumars (1974). This device recovers a relatively undisturbed sample of the sea floor by isolating a 50 by 50 cm area of the bottom and penetrating the sediment to a depth of 40- -50 cm.
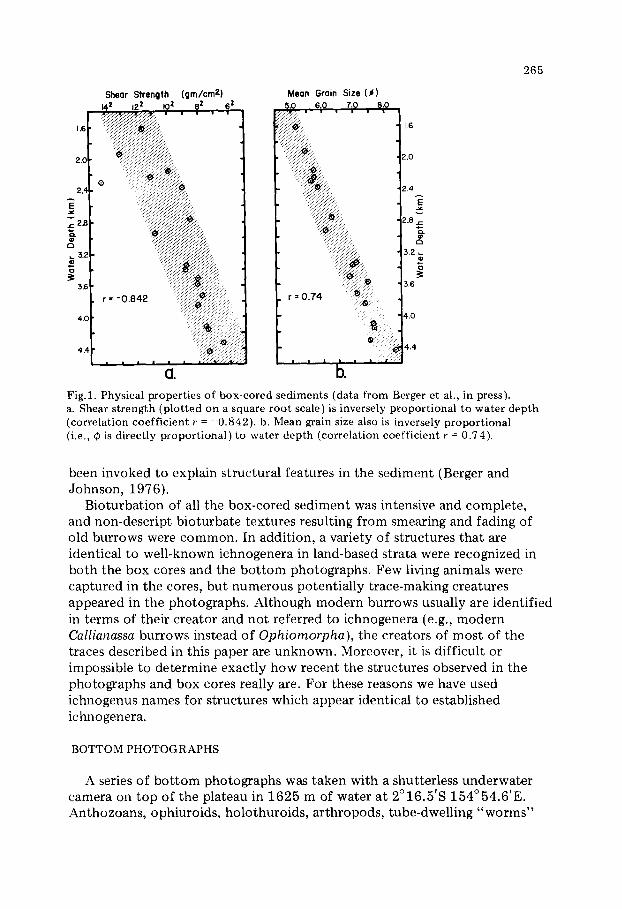
All the sediments recovered were highly burrowed, white to medium brown, calcareous nannofossil ooze. Physical properties (grain size and shear strength) show distinct depth gradients (Fig.l) . Average grain size ranges from 5.2 to 8.3 ¢, with the coarsest sediments on top of the plateau and the finest ones at depth. Both winnowing (especially above the lysocline) and dissolution (below the lysocline) contribute to this effect (Berger et al., in press). Shear strength also is clearly depth-dependent, the most firm sediment being on top of the plateau and the least firm at the foot of its flanks.
Sediment collected from the top of the plateau appeared to be undis- turbed and displayed an obvious vertical component of burrow orientation. On the other hand, sediments from below the lysocline exhibited sedimentary folding, shearing of layers and smearing of burrows. Some type of gravitational sediment movement ("ooze f low") resulting from carbonate dissolution has
265
Shear Strength (grn/cm 2) Mean Grain Size ( J ) i4 2 12 2 10 2 8 2 6 2
t 1 •
_ _ . ~ ! ~ .
40~ 4
CI. I~.' Fig.1. Physical properties of box-cored sediments (data from Berger et al., in press). a. Shear strength (plotted on a square root scale) is inversely proportional to water depth (correlation coefficient r = -0.842). b. Mean grain size also is inversely proportional (i.e., ~ is directly proportional) to water depth (correlation coefficient r = 0.74).
been invoked to explain s t ruc tura l fea tures in the s ed imen t (Berger and Johnson , 1976) .
B io tu rba t ion of all the b o x - c o r e d s ed i m en t was intensive and c o m p l e t e , and non-desc r ip t b i o t u r b a t e t ex tu res resul t ing f r o m smear ing and fading of old bur rows were c o m m o n . In addi t ion , a var ie ty o f s t ruc tures t ha t are ident ical to we l l -known i chnogenera in land-based s t ra ta were recognized in b o t h the b o x cores and the b o t t o m pho tog raphs . Few living animals were c a p t u r e d in the cores, bu t n u m e r o u s po ten t i a l ly t r ace -mak ing c rea tures a p p e a r e d in the pho tog raphs . Al though m o d e r n bu r rows usual ly are ident i f ied in t e rms of thei r c rea to r and no t re fe r red to i chnogenera (e.g., m o d e r n Callianassa bur rows ins tead o f Ophiomorpha), the c rea to rs of m o s t of the t races descr ibed in this p a p e r are u n k n o w n . Moreover , it is d i f f icul t or imposs ib le to de t e rmine exac t ly h o w recen t the s t ruc tures observed in the p h o t o g r a p h s and b o x cores real ly are. For these reasons we have used i chnogenus names for s t ruc tures which appea r ident ical to es tab l i shed ichnogenera .
BOTTOM PHOTOGRAPHS
A series o f b o t t o m p h o t o g r a p h s was t a k e n wi th a shut ter less u n d e r w a t e r c amera on t o p o f the p la teau in 1625 m of wa te r a t 2 °16 .5 'S 1 5 4 ° 5 4 . 6 ' E . An thozoans , ophiuro ids , ho lo thuro ids , a r t h ropods , tube-dwel l ing " w o r m s "

266
Fig.2. Eehinoid trails photographed on the sea bottom in 1625 m of water at 2~16.5'S O F • 154 54.6 E (Ontong Java Plateau).
and rat.tail fish were recognized in the photographs. Moreover, the sediment appeared to be thoroughly re-worked by organisms, and a variety of distinctive organism traces on the sediment surface were evident.
Echinoid trails
The most abundant and easily recognized trails in the bo t tom photographs are those of echinoids plowing across the sediment surface. Such trails typically consist o f a ridge of sediment (often bi-lobed with a central furrow), and usually they occur in loose, irregular meander patterns (Fig.2). Although

267
we saw no creatures in the act of making the trails, Heezen and Hollister (1971, figs. 4.22 and 4.23) show that identical trails are indeed produced by wandering echinoids elsewhere in the deep sea. One small spatangoid echinoid was recovered in one of the box cores, but no trail was preserved on the surface of that core.
Echinoid trails resembling those in our photographs have been seen at abyssal depths all over the world (e.g., see Fig.3). Similar trace fossils in land- based sections have been described as Scolicia by Kern and Warme (1974, figs. 7e and f) in the Cretaceous Point Loma Formation near San Diego, although deep-sea echinoid trails apparently lack the characteristic central ridge of Scolicia. In gross morphology the echinoid trails in our bot tom photographs also resemble the Taphrhelminthopsis described by Ksi~2kiewicz (1970, pls. 2a to g and 3f to h) in Eocene Carpathian Flysch of Poland.
Holothuroid trails
Several holothuroids were captured on film at the end of their trails in one photograph (Fig.4). The trails appear to be broader and higher than those of the echinoids, and their width and shape are very irregular. Moreover, they lack the central furrow that characterizes many of the echinoid trails. The coiled fecal string of a small holothuroid, probably produced in the manner described by Heezen and Hollister (1971, fig. 5.14), appeared in one photograph.
Clumps of holes
Tiny pits or holes in the sea floor, clumped together in groups ranging from six to thirteen holes of uniform diameter, were noticed in several photographs. Although ophiuroids buried just beneath the sediment surface may leave a regular ring of holes with one hole per arm (Heezen and Hollister, 1971, figs. 6.12 and 6.21), the clumps of holes in our photographs never are so regular, and the number of holes varies markedly from clump to clump. We have no evidence to suggest the type of organism (or organisms) producing the holes, although obviously it must be an infaunal creature. Clumps of holes in the deep-sea floor are distributed worldwide (Heezen and Hollister, 1971, figs. 6.23, 6.24 and 6.26).
Glockeria
Traces resembling the ichnogenus Glockeria were photographed on the ocean bottom. These consist of large numbers of thin, straight, shallow grooves in the sediment radiating outwards from a central hole (Fig.5). The overall aspect of the trace is that of a spoked wheel or of a daisy-like flower with very thin petals. Lack of a scale in the photographs precludes our estimating its diameter, although specimens of different sizes were photo-

268
Fig.3. Echinoid trails p h o t o g r a p n e a on the sea b o t t o m in 2320 m water at 2 1 ~ 2 6 . 0 t N 85 ° 23.5 ~ W (Car ibbean Sea).

269
Fig.4. Holothuroid trails photographed on the sea bottom in 1625 m of water at 2°16.5'S 154°54.6'E (Ontong Java Plateau); h = holothuroid.
graphed. Very similar t races have been discovered in Cre taceous f lysch- type deposi ts in Poland (Ksig~,kiewicz, 1970, fig. 7) and also on the m o d e r n deep- sea f loor (Heezen and Hollister, 1971, fig. 6.13).
BOX CORES
The box core samples revealed tha t abyssal deposi ts in the vicini ty of On tong Java Plateau are indeed heavily b io tu rba ted . With one side o f the box

270
Fig.5. Glockeria photographed on the sea bottom in 1625 m of water at 2°16.5tS 154 ° 54.6 t E (Ontong Java Plateau ).
removed , the bu r rowed sed iment could be s tudied in vert ical cross-section. Most o f the sediments exh ib i t ed b io tu rba t e t ex tu res in which the general o r i en ta t ion o f bur rows was apparent , bu t individual ly dis t inct bur rows could no t easily be fo l lowed for any length in mos t cases.
A grid pa t te rn o f b io tu rba t ion , consist ing o f vert ical and hor izon ta l burrows, seemed to charac ter ize sediments col lec ted at shal lower dep ths (e.g., 1 5 0 0 - - 2 5 0 0 m) on top o f the plateau (Fig.6}. Vert ical burrows, how- ever, were rare to absent in sediments col lec ted at greater dep ths (e.g., more than 3300 m) on the pla teau 's f lank (Fig.7). Incl ined and (or} smeared

271
Fig.6. Grid pattern of bioturbation in a vertical face of Box Core No. 120 recovered from 2247 m of water (Ontong Java Plateau).
Fig.7. Predominantly horizontal burrows in a vertical face of Box Core No. 129 recovered from 4169 m of water (Ontong Java Plateau).
burrows (Fig.8), p robab ly resulting f rom mass flow down the slope, are c o m m o n in these deeper-water cores (Berger and Johnson , 1976).
Some box cores con ta ined distinctive traces on the surface and within the sediment tha t resemble wel l -known ichnogenera in land-based strata. The originators o f these traces appeared to be rarely present, and no e f for t was made to systemat ical ly find and capture them.
Chondri tes
This is a dendri t ic bur row sys tem c o m p o s e d of numerous branching tunnels o f un i fo rm diameter. Branching occurs as various side tunnels

272
Fig.8. Deformed burrows in a vertical face of Box Core No. 139 recovered from 4118 m of water (Ontong Java Plateau).
angling of f a previous or main tunnel at 30 ° or 40 ° angles rather than as bifurcations at "Y-shaped" junctions.
Burrows resembling Chondrites were found on the surfaces of three box cores f rom depths of 1597--2949 m near Ontong Java Plateau (Fig.9). The tunnel diameters ranged from 2 to 4 mm. All the specimens appeared to be surface features; some were preserved in convex epirelief, and others were preserved in full relief as coherent agglutinated burrow systems lying loose on the sediment surface. Although no Chondrites were seen inside the sediment in vertical cross-section when the box cores were dissected, it is possible that specimens were overlooked due to poor color contrast. Ancient Chondrites specimens preserved in pelagic carbonates in Deep Sea Drilling Project cores are typically oblique to bedding and exhibit branching at all angles from the vertical (Ekdale, 1974; Chamberlain, 1975).
Spiroraphe
This distinctive spiralling trail actually consists of a double spiral in which the course winds inward, doubles back on itself at the center, and then winds outward again alongside the previous, inward spiralling trail. Traces resembling Spiroraphe occurred in convex epirelief on the surfaces of three box cores collected in water depths ranging from 3368 to 4169 m (Fig.10).
?Zoophycos
A "roos ter tail" trace resembling Zoophycos was preserved some 35 cm below the sediment surface in one box core taken from 2183 m of water

273
Fig.9. Chondrites on the surface of Box Core No. 123 recovered from 2949 m of water (Ontong Java Plateau).
(Fig . l l ) . The trace is a highly lobate, horizontal spreite structure with radiating arcuate grooves similar to Zoophycos specimens described by Simpson (1970, fig. lc, d, e) and others. However, a marginal tube on the spreite and "U-shaped" aspect of the burrow as a whole, which are typical characteristics of the ichnogenus, are not evident in our specimen.
Most workers believe that the typical Zoophycos preserved in ancient rocks on land is a burrow system produced within the sediment by an infaunal animal (Taylor, 1967; Simpson, 1970; H~intzschel, 1975; and many others) rather than a surface trail. The specimen from Ontong Java Plateau supports that hypothesis.
Planolites
This trace is simply an unornamented, unbranched, horizontal or sub- horizontal trail that may be straight or slightly sinuous. In vertical cross- sections tl~rough box cores Planolites appears as a solid circle or ellipse (Figs. 6--8). This was the most common burrow type in all the samples, and specimens ranged in diameter from 2 or 3 mm to 2 or 3 cm. Two box cores had small (2 mm wide) sinusoidal trails on the surface, and possibly these also should be referred to Planolites.
Skolithos
Vertical cylindrical burrows, ranging from approximately 1 to 3 cm in diameter and 5--10 cm long, were common in most of the shallower-water (i.e., less than 2500 m) cores (Fig.6). Except for their upright orientation, they closely resemble the dominantly horizontal Planolites in size, shape and

274
Fig.10. Spiroraphe o n the surface o f Box Core No. 108 recovered f r o m 3383 m of wa te r ( O n t o n g Java Plateau) .
color. Burrows of this type in bathyal sediments in New Zealand have been referred to Tigillites by Hayward (1976). Alpert (1974) and H~intzschel (1975), however, suggest that Tigillites is synonymous with Skolithos and possibly Monocraterion, which are known to be especially characteristic of shallow-water environments. Frey (1970} refers to similar deep-water vertical burrows in the Niobrara Chalk of Kansas as "cylindrical shafts". Although the ichnogenus Skolithos generally is applied to vertical burrows in shallow- marine sandstones of Cambrian through Devonian age, the morphologic descriptions of Alpert and H~intzschel also fit the structures found in our box cores. Skolithos was rare to absent in the deeper-water cores, possibly because
Fig.11. ?Zoophycos on a ho r i zon ta l p lane in Box Core No. 112 recovered f rom 2183 m of wate r ( O n t o n g Java Plateau).

275
the m u c h lower shear s t rengths of sub-lysocl inal sed iments do n o t p e r m i t vert ical bu r rows to r ema in open w i t h o u t collapsing.
Teichichnus
This b u r r o w is a vert ical spreite p r o d u c e d by an infaunal o rgan ism moving back and fo r th in a ho r i zon ta l d i rec t ion while con t inua l ly shif t ing its bu r row u p w a r d (or, less c o m m o n l y , d o w n w a r d ) in the sediment . The ne t resul t is a wall-like s t ruc tu re tha t , in cross-sect ion in a b o x core, appea r s as a s tack of concave-up dish s t ruc tu res wi th the last tunne l o f the organism at the t op of the s tack (Fig.12). Teichichnus was d iscovered in a b o x core col lec ted at a
Fig.12. Teichichnus in a vertical face of Box Core No. 92 recovered from 1597 m of water (Ontong Java Plateau).
d e p t h o f 1597 m. Excava t ion o f the s ed imen t in the core revealed t ha t the b u r r o w e x t e n d e d in to the core for a ho r i zon ta l d is tance o f m o r e t h a n 15 cm. The spreite, including the r o u n d tunne l at the top , was 31 m m high. The tunne l a t the t o p was 8 m m in d iamete r .
DISCUSSI ON
The b o x cores and b o t t o m p h o t o g r a p h s f r o m O n t o n g Java Pla teau provide a gl impse of t he m o d e r n abyssal ichnofac ies a t one local i ty. Traces p r o d u c e d b y organisms b o t h on and in the s ed imen t have been observed, and there is no doub t ing the abyssal s e d i m e n t a r y e n v i r o n m e n t in which t hey were f o r m e d

276
Thus, conclusions drawn from this study may aid in the recognition of ancient deep-sea deposits on land.
The abyssal sediments on and around Ontong Java Plateau are moderately productive; they reflect relatively high fertility in the overlying waters, and they support a rich benthic fauna. The sea bottom here is thoroughly re- worked by organisms digging in, plowing through or crawling across the sediment. All the box cores and all the bottom photographs demonstrate that bioturbation is intense and universal. In fact, the myriad traces produced on top of the sea floor (i.e., those that we discovered in the bottom photographs and on the surfaces of box cores) appear to be destroyed following burial due to the extensive burrowing activities of infaunal organisms.
Wherever continuous sedimentation precludes the formation of bedding planes, surface trails rarely will enter the fossil record. Surface (or bedding plane) traces will be preserved only if there is (1) a break in sedimentation, causing a bedding plane to form, or (2) a sudden change in living conditions at the bottom, temporarily halting the bioturbation activities of the infauna. Neither possibility is likely to occur frequently in the stable deep-sea environment. Therefore, we can assume that only those traces produced last and deepest in the sediment by burrowers will be preserved. This has been shown to be true for trace fossils in massive, pelagic deposits in Deep Sea Drilling Project cores (Ekdale, 1974; Chamberlain, 1975), and the box cores studied for this report confirm it for modern pelagic deposits.
Seilacher (1964, 1967} has suggested that abyssal deposits are characterized by the "Nereites ichnofacies", which includes a wide variety of highly patterned, dominantly horizontal, feeding trails. Presumably these are produced by deposit feeders attempting to mine the water-sediment interface (or perhaps a lithologic interface within a turbidite sediment) for food as efficiently as possible. Seilacher's interpretation, however, is based primarily on observations of trace fossils in turbidite sequences (see also Seilacher, 1962) rather than in abyssal pelagic deposits far-removed from land sources, such as the sediments investigated for this paper.
Certainly the Spiroraphe, Glockeria and meandering echinoid trails discovered in our samples fall into the category of horizontal, patterned, feeding trails. These are bedding plane traces that are best known in land- based sections in turbidites and other sequences representing discontinuous, deep-water sedimentation. They are unknown, however, in massive deep-sea deposits representing continuous pelagic sedimentation, such as those preserved in Deep Sea Drilling Project cores, due to lack of preservation (Ekdale, 1974; Chamberlain, 1975). Their absence in pelagic deposits seems to be due to the absence of bedding planes, because they are produced on the sea bottom and then apparently destroyed by extensive infaunal bioturbation following burial.
Kennedy (1975} and Ekdale (in press) have suggested that the typical ichnofacies in deep-sea carbonates consists of Chondrites, Planolites, Teichichnus and Zoophycos. These are the most characteristic traces in

277
ancient carbonate oozes in Deep Sea Drilling Project cores; they are preserved within the sediment rather than on bedding planes. All four ichnogenera, including our tentative designation of ?Zoophycos, were found in the box cores collected for this study. With the possible exception of Planolites, these traces also fit Seilacher's general model of deposit-feeding organisms systematically mining the sediment for food in highly organized and efficient burrow patterns.
ACKNOWLE DGEM ENTS
This research was funded by grants from the Office of Naval Research (USN-N-00014-69-A-0200-6049 to Berger), the National Science Foundation (OCE-75-04335-A01 to Berger and OCE-75-21601 to Ekdale) and the University of Utah Research Committee (No. 1732 to Ekdale). We thank the many individuals who made this work possible: T. C. Johnson assisted in all the phases of the expedition; G. Wilson was in charge of the box-coring operation; T. Walsh took the deep-sea photographs; D. Ripley assisted in the laboratory; T. C. Johnson and E. L. Hamilton made measurements on physical properties. J. Philip Kern, Peter H. Roth and Susan F. Ekdale offered suggestions for improvement of the manuscript.
REFERENCES
Alpert, S. P., 1974. Systematic review of the genus Skolithos. J. Paleontol., 48: 661--669. Berger, W. H. and Johnson, T. C., 1976. Deep-sea carbonates: Dissolution and mass
wasting on Ontong-Java Plateau. Science, 192: 785--787. Berger, W. H., Johnson, T. C. and Hamilton, E. L., in press. Sedimentation on Ontong-
Java Plateau: Observations on a classic "carbonate monitor". In: Fate of Fossil Fuel CO2. Off. Naval Res. Conf., Hawaii, 1976.
Chamberlain, C. K., 1975. Trace fossils in DSDP cores of the Pacific. J. Paleontol., 49: 1074--1096.
Ekdale, A. A., 1974. Geologic History of the Abyssal Benthos: Evidence from Trace Fossils in Deep Sea Drilling Project Cores. Dissertation, Rice University, Houston, Texas.
Ekdale, A. A., in press. Trace fossils in Leg 42A cores. Init. Rep. Deep Sea Drill. Proj., 42. Frey, R. W., 1970. Trace-fossils of the Fort Hayes Limestone Member of the Niobrara
Chalk (Upper Cretaceous) of West-central Kansas. Univ. Kansas Paleontol. Contrib., Art. 53 :41 pp.
H/intzschel, W., 1975. Trace fossils and problematica, second edition. In: C. Teichert (Editor), Treatise on Invertebrate Paleontology. Part W, Supp. 1, Miscellanea, Univ. of Kansas and Geol. Soc. Am., Lawrence, Kansas.
Hayward, B. W., 1976. Lower Miocene bathyal and submarine canyon ichnocoenoses from Northland, New Zealand. Lethaia, 9: 149--162.
Heezen, B. C. and Hollister, C. D., 1971. The Face of the Deep. Oxford University Press, New York, 659 pp.
Hessler, R. R. and Jumars, P. A., 1974. Abyssal community analysis from replicate box cores in the central North Pacific. Deep-sea Res., 21: 185--209.
Kennedy, W. J., 1975. Trace fossils in carbonate rocks. In: R. W. Frey (Editor), The Study of Trace Fossils. Springer-Verlag, New York, N.Y.
Kern, J. P. and Warme, J. E., 1974. Trace fossils and bathymetry of the Upper Cretaceous

278
Point Loma Formation, San Diego, California. Bull. Geol. Soc. Am., 85: 893--900. Ksi}~kiewicz, M., 1970. Observations on the ichnofauna of the Polish Carpathians. In:
T. P. Crimes and J. C. Harper (Editors), Trace Fossils. Geol. J. Spec. Iss., 3 :283- -322 Seilacher, A., 1962. Paleontological studies on turbidite sedimentation and erosion. J.
Geol., 70: 227--234. Seilacher, A., 1964. Biogenic sedimentary structures. In: J. Imbrie and N. D. Newell
(Editors), Approaches to Paleoecology. Wiley, New York, N~Y., pp. 296--316. Seilacher, A., 1967. Bathymetry of trace fossils. Mar. Geol., 5: 413--428. Simpson, S., 1970. Notes on Zoophycos and Spirophyton. In: T. P. Crimes and J. C.
Harper (Editors), Trace Fossils. Geol. J. Spec. Iss., 3: 505--514. Taylor, B. J., 1967. Trace fossils from the Fossil Bluff Series of Alexander Island. Brit.
Antarctic Surv. Bull., 13: 1--30.
Copyright © 2022 FDOKUMEN




















