
ddy219.pdf - Oxford Academic
12
ORIGINAL ARTICLE Ulk2 controls cortical excitatory–inhibitory balance via autophagic regulation of p62 and GABA A receptor trafficking in pyramidal neurons Akiko Sumitomo 1 , Hiroshi Yukitake 2 , Kazuko Hirai 1 , Kouta Horike 1 , Keisho Ueta 1 , Youjin Chung 2 , Eiji Warabi 3 , Toru Yanagawa 3 , Shiho Kitaoka 4,5 , Tomoyuki Furuyashiki 4,5 , Shuh Narumiya 4 , Tomoo Hirano 6 , Minae Niwa 2 , Etienne Sibille 7 , Takatoshi Hikida 1 , Takeshi Sakurai 1 , Koko Ishizuka 2 , Akira Sawa 2, * and Toshifumi Tomoda 1,7, * 1 Department of Research and Drug Discovery, Medical Innovation Center, Kyoto University Graduate School of Medicine, Kyoto 606-8507, Japan, 2 Department of Psychiatry and Behavioral Sciences, Johns Hopkins University School of Medicine, Baltimore, MD 21287, USA, 3 Faculty of Medicine, University of Tsukuba, Tsukuba 305-8575, Japan, 4 CREST Project, Medical Innovation Center, Kyoto University Graduate School of Medicine, Kyoto 606- 8507, Japan, 5 Department of Pharmacology, Kobe University Graduate School of Medicine, Hyogo 650-0017, Japan, 6 Department of Biophysics, Kyoto University Graduate School of Science, Kyoto 606-8502, Japan and 7 Campbell Family Mental Health Research Institute, Centre for Addiction and Mental Health, University of Toronto, Toronto, ON M5T 1R8, Canada *To whom correspondence should be addressed at: Department of Psychiatry and Behavioral Sciences, Johns Hopkins University School of Medicine, Baltimore, MD 21287, USA. Tel: þ1 4109554726; Email: [email protected] (A.S.); Campbell Family Mental Health Research Institute, Centre for Addiction and Mental Health, University of Toronto, Toronto, ON M5T 1R8, Canada. Tel: þ1 4165358501; Email: [email protected] (T.T.) Abstract Autophagy plays an essential role in intracellular degradation and maintenance of cellular homeostasis in all cells, including neurons. Although a recent study reported a copy number variation of Ulk2, a gene essential for initiating autophagy, associated with a case of schizophrenia (SZ), it remains to be studied whether Ulk2 dysfunction could underlie the pathophysiology of the disease. Here we show that Ulk2 heterozygous (Ulk2 þ/ ) mice have upregulated expression of sequestosome-1/p62, an autophagy-associated stress response protein, predominantly in pyramidal neurons of the prefron- tal cortex (PFC), and exhibit behavioral deficits associated with the PFC functions, including attenuated sensorimotor gating and impaired cognition. Ulk2 þ/ neurons showed imbalanced excitatory–inhibitory neurotransmission, due in part to selec- tive down-modulation of gamma-aminobutyric acid (GABA) A receptor surface expression in pyramidal neurons. Genetically reducing p62 gene dosage or suppressing p62 protein levels with an autophagy-inducing agent restored the GABA A receptor surface expression and rescued the behavioral deficits in Ulk2 þ/ mice. Moreover, expressing a short peptide that specifically interferes with the interaction of p62 and GABA A receptor-associated protein, a protein that regulates endocytic trafficking of GABA A receptors, also restored the GABA A receptor surface expression and rescued the behavioral deficits in Ulk2 þ/ mice. Received: January 26, 2018. Revised: May 22, 2018. Accepted: June 4, 2018 V C The Author(s) 2018. Published by Oxford University Press. All rights reserved. For permissions, please email: [email protected] 3165 doi: 10.1093/hmg/ddy219 Advance Access Publication Date: 8 June 2018 Original Article Human Molecular Genetics, 2018, Vol. 27, No. 18 3165–3176 Downloaded from https://academic.oup.com/hmg/article/27/18/3165/5034852 by guest on 06 September 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of ddy219.pdf - Oxford Academic

O R I G I N A L A R T I C L E
Ulk2 controls cortical excitatoryndashinhibitory balance via
autophagic regulation of p62 and GABAA receptor
trafficking in pyramidal neuronsAkiko Sumitomo1 Hiroshi Yukitake2 Kazuko Hirai1 Kouta Horike1Keisho Ueta1 Youjin Chung2 Eiji Warabi3 Toru Yanagawa3 Shiho Kitaoka45Tomoyuki Furuyashiki45 Shuh Narumiya4 Tomoo Hirano6 Minae Niwa2Etienne Sibille7 Takatoshi Hikida1 Takeshi Sakurai1 Koko Ishizuka2Akira Sawa2 and Toshifumi Tomoda171Department of Research and Drug Discovery Medical Innovation Center Kyoto University Graduate School ofMedicine Kyoto 606-8507 Japan 2Department of Psychiatry and Behavioral Sciences Johns Hopkins UniversitySchool of Medicine Baltimore MD 21287 USA 3Faculty of Medicine University of Tsukuba Tsukuba 305-8575Japan 4CREST Project Medical Innovation Center Kyoto University Graduate School of Medicine Kyoto 606-8507 Japan 5Department of Pharmacology Kobe University Graduate School of Medicine Hyogo 650-0017Japan 6Department of Biophysics Kyoto University Graduate School of Science Kyoto 606-8502 Japan and7Campbell Family Mental Health Research Institute Centre for Addiction and Mental Health University ofToronto Toronto ON M5T 1R8 Canada
To whom correspondence should be addressed at Department of Psychiatry and Behavioral Sciences Johns Hopkins University School of MedicineBaltimore MD 21287 USA Tel thorn1 4109554726 Email asawa1jhmiedu (AS) Campbell Family Mental Health Research Institute Centre for Addictionand Mental Health University of Toronto Toronto ON M5T 1R8 Canada Tel thorn1 4165358501 Email ttomoda1gmailcom (TT)
AbstractAutophagy plays an essential role in intracellular degradation and maintenance of cellular homeostasis in all cells includingneurons Although a recent study reported a copy number variation of Ulk2 a gene essential for initiating autophagyassociated with a case of schizophrenia (SZ) it remains to be studied whether Ulk2 dysfunction could underlie thepathophysiology of the disease Here we show that Ulk2 heterozygous (Ulk2thorn) mice have upregulated expression ofsequestosome-1p62 an autophagy-associated stress response protein predominantly in pyramidal neurons of the prefron-tal cortex (PFC) and exhibit behavioral deficits associated with the PFC functions including attenuated sensorimotor gatingand impaired cognition Ulk2thorn neurons showed imbalanced excitatoryndashinhibitory neurotransmission due in part to selec-tive down-modulation of gamma-aminobutyric acid (GABA)A receptor surface expression in pyramidal neurons Geneticallyreducing p62 gene dosage or suppressing p62 protein levels with an autophagy-inducing agent restored the GABAA receptorsurface expression and rescued the behavioral deficits in Ulk2thornmice Moreover expressing a short peptide that specificallyinterferes with the interaction of p62 and GABAA receptor-associated protein a protein that regulates endocytic trafficking ofGABAA receptors also restored the GABAA receptor surface expression and rescued the behavioral deficits in Ulk2thornmice
Received January 26 2018 Revised May 22 2018 Accepted June 4 2018
VC The Author(s) 2018 Published by Oxford University Press All rights reservedFor permissions please email journalspermissionsoupcom
3165
doi 101093hmgddy219Advance Access Publication Date 8 June 2018Original Article
Human Molecular Genetics 2018 Vol 27 No 18 3165ndash3176
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
Thus the current study reveals a novel mechanism linking deregulated autophagy to functional disturbances of the nervoussystem relevant to SZ through regulation of GABAA receptor surface presentation in pyramidal neurons
IntroductionMacroautophagy (hereafter referred to as autophagy) is a spe-cialized membrane trafficking machinery and a major cellularrecycling system responsible for degrading old proteins anddamaged organelles which ultimately serve as resources togenerate new proteins and energy and thus critical to the main-tenance of cellular homeostasis (1) In response to a range ofcellular stresses (eg depletion of nutrients and energy mis-folded protein accumulation and oxidative stress) autophagy isactivated to mitigate such stresses via activation of ULK1ULK2ATG1 serinethreonine protein kinases critical to the regulationof autophagy induction Subsequently microtubule-associatedprotein light chain 3 (LC3)-positive autophagosomes are formedto enwrap the proteins and organelles that are selectively la-beled with sequestosome-1p62 an autophagy-related stress re-sponse protein and then trafficked to fuse with lysosomes fordegradation
A role of autophagy in the nervous system was previouslyshown in nervous system-specific Atg5 and Atg7 conditionalknockout (cKO) mice which exhibit progressive neuronal deathaccompanied by accumulation of abnormal cytosolic proteinsand formation of p62-positive inclusions in neurons (2ndash4)These results suggested that continuous quality control of dif-fuse cytosolic proteins via basal autophagy is important for neu-ronal functions Subsequently we and others providedevidence that autophagy regulates synapse morphogenesis andfunction using a series of autophagy-deficient mutants inDrosophila melanogaster which demonstrate diminished size andnumber of synaptic boutons and a decrease in evoked neuro-transmitter release (5ndash8) Meanwhile ULK1ULK2ATG1 regulateendocytic trafficking of neuronal cargoes through associationwith a synaptic protein SynGAP (9) and also regulate traffickingof synaptic vesicles via association with additional neuronalproteins such as Unc-76FEZ1 and synaptotagmin-1 (5) In ac-cordance with these observations dopaminergic neuron-specific Atg7-cKO mice showed a role of autophagy in presynap-tic structure and neurotransmission exhibiting enlarged stria-tal axon terminal profiles and abnormally increased dopaminerelease (10) It is thus possible that autophagy-mediated regula-tion of pre- or post-synaptic components via modulation oftheir trafficking could contribute to neural plasticity Howeverquestions remain as to (i) whether neuronal autophagy is in-volved in higher brain functions such as cognition (ii) if sowhich brain regions and neuronal cell types are most vulnerableto impaired autophagy leading to specific behavioral outcomesand (iii) what are molecular mechanisms that account for suchoutcomes
A recent study showed that mice doubly deficient for Ulk1and Ulk2 in the nervous system have a slight increase in the lev-els of p62 protein a key target molecule in autophagic proteindegradation without showing a sign of p62-positive inclusions(11) Likewise mice deficient for Ulk1 had increased levels of p62protein expression in the prefrontal cortex (PFC) (12)Importantly unlike the knockout of non-redundant autophagygenes such as Atg5 and Atg7 that display early postnatal deathin mice and may possibly mask the underlying neural mecha-nisms Ulk1 and Ulk2 have redundant roles in autophagy andmice singly deficient for either of these genes are viable with no
overt deficits in overall physical conditions such as life spanbody weight and motor functions (12ndash15) Thus we regard thatperturbation of Ulk1 andor Ulk2 will provide the best model toaddress the current questions With this model it is expectedthat we could address whether this pathway plays a causal rolein neurobehavioral manifestation In this regard it is notablethat the copy number variations (CNVs) of ULK1ULK2ULK4family member genes were recently reported to be enriched inschizophrenia (SZ) patients recruited in a genome-wide associa-tion study (odds ratio frac14677) (16)
In the present study we set out to test whether Ulk2 heterozy-gous (Ulk2thorn) mice have signs of attenuated autophagy (ie ele-vated p62 expression) in certain regions or cell types within thebrain and addressed whether such changes can be linked to neu-robehavioral outcomes We hypothesized that elevated p62 lev-els have functional consequences on neurotransmission thatcould account for the observed behavioral deficits relevant to SZ
ResultsUlk2thornmice show elevated p62 protein levels in the PFC
As described above changes in the expression of p62 proteinhave been reported in both Ulk1 and Ulk2 knockout mice (1112)However a question remains as to which brain regions and cellsmost robustly represent such changes Thus we first examinedthe expression of p62 protein in various brain regions of Ulk2thorn
mice The mutants had significantly elevated expression of p62protein in the medial PFC (Fig 1A and B SupplementaryMaterial Fig S1A and B) which occurred in Ca2thorncalmodulin-dependent protein kinase II (CaMKII)-positive pyramidal neu-rons but not in the interneurons (Fig 1A SupplementaryMaterial Fig S1B) This elevated p62 expression was seen in thePFC but not in all other brain regions tested including the vi-sual cortex nucleus accumbens dorsal striatum hippocampusand ventral tegmental area (Supplementary Material Fig S1A)Taken together reduced expression of Ulk2 affects cellular pro-cesses involving p62 protein likely associated with autophagichomeostasis in a neuronal cell-type preferential and a brainregion-preferential manner
Ulk2thornmice show behavioral deficits associated withthe PFC function
We then questioned whether and how the reduction in Ulk2affects the PFC-relevant behaviors including sensorimotor gat-ing (17) and cognitive flexibility (18) Ulk2thorn mice had impairedsensorimotor gating showing lower levels of prepulse inhibi-tion (PPI) of acoustic startle stimuli (Fig 1C and D) To assess thePFC-dependent cognitive function Ulk2thornmice were tested in arule shifting paradigm (1920) in which mice were initiallytrained to associate food reward with a specific stimulus (ie ei-ther an odor or a digging medium) and subsequently evaluatedfor cognitive flexibility by changing the type of stimulus thatpredicts the reward (Fig 1E) Although both Ulk2thorn and WTmice learned the association rule in a similar number of trialsduring the initial association phase of trials Ulk2thorn mice werespecifically impaired during the rule shifting phase of trials(Fig 1F and G)
3166 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
WT WT
Saline CCI-779
Ulk2+-
Ulk2+-DAPI
WT
Ulk2+-
p62
LC3-I
α-Tubulin
LC3-II
p62 CaMKII
LC3-IITotal LC3
Nor
mal
ized
rat
iop62
Nor
mal
ized
leve
ls
1
0
2 (au)
CCI-779
WT
Ulk2+-
1
0
2 (au)
WT
Ulk2+-
50
(kDa)
17
72
WT + Veh Ulk2 + Veh Ulk2 + CCI-779Ulk2 + Clozapine
+-+-+-
0
500
(au)
Sta
rtle
res
pons
e
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
P
PI
0
20
40
60
80
A B
C D
E
F
WT
CCI-779
Ulk2+- Ulk2+-
+
O1TA
O2
TB
O1
TB
O2
TA
O1
TA
O2
TB
O1
TB
O2
TA
Initial association Rule shiftTrial 1 Trial n Trial 1 Trial n
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Ulk2 + CCI-779
WT + Veh Ulk2 + Veh
+-
+-
Num
ber
of p
erse
vera
tive
erro
rs
G
0
10
20
30
40
CCI-779
WT
Ulk2+-
WT
Ulk2+-
Veh+
Veh+
WT
CCI-779
Ulk2+- Ulk2+-
+Veh
+Veh
+
Figure 1 Upregulated p62 protein levels in pyramidal neurons of the PFC and PFC-associated behavioral deficits in Ulk2thorn mice (A) Selective upregulation of p62 pro-
tein in CaMKIIthorn neurons in the PFC of Ulk2thornmice Scale bar 10 lm (B) Western blot of p62 and LC3 in the frontal cortex of Ulk2thornmice chronically treated with saline
or a rapamycin analog CCI-779 The expression levels were measured by densitometric analysis using ImageJ Error bars indicate SEM P lt005 (C D) The amplitude of
startle response (C) and the percentage of PPI (D) were evaluated for WTthornsaline (nfrac1410) Ulk2thornthornsaline (nfrac1410) Ulk2thornthornCCI-779 (nfrac1410) and Ulk2thornthornclozapine (nfrac1410)
No significant difference in startle response [F(3 36)frac1409683 Pfrac1404183] (one-way ANOVA) Statistical significance for PPI F(3 36)frac141128 P lt00001 (two-way ANOVA with
repeated measures) P lt005 (Bonferroni post hoc test) (E) Schematic diagram of the rule shift assay Upon rule shifting the stimulus associated with food reward is
changed from odor cue 1 (underlined) to texture cue A (italicized) in this diagram (F) Numbers of trials to criterion during the initial association phase as well as the
rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thornthornCCI-779 (nfrac147) No significant difference during the initial association phase [F(2
20)frac14154 Pfrac140832] Statistical significance during the rule-shift phase F(2 20)frac14767 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (G) Numbers of per-
severative errors during the rule shift phase were scored F(2 20)frac14483 Pfrac140024 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3167|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
CaMKII-specific Ulk2thornmice show behavioral deficitsassociated with the PFC function
We next investigated specific roles of neuronal subtypes in theobserved phenotypes Mice heterozygous for conditional Ulk2knockout allele were crossed with mice expressing eitherpan-specific Cre (EIIa-Cre) pyramidal neuron-specific Cre(CaMKII-Cre) or interneuron-specific Cre (Viaat-Cre) and PPIwas measured When Ulk2 was ablated in an interneuron-specific manner PPI was not affected (Supplementary MaterialFig S1CndashE) In contrast pyramidal neuron-specific Ulk2 ablationresulted in attenuated PPI (Supplementary Material Fig S1C Dand F) These data are in line with the notion that homeostaticdeficits involving p62 accumulation in the CaMKII-positive py-ramidal neurons underlie the PPI deficits in Ulk2thornmice
Reduced surface expression of GABAA receptors andimbalanced excitatoryndashinhibitory neurotransmission inUlk2thorn pyramidal neurons
To further investigate molecular and cellular basis that may un-derlie these behavioral changes electrophysiological propertiesof the cultured pyramidal neurons prepared from Ulk2thorn micewere examined Typically 80 of neurons in culture wereCaMKII-positive pyramidal neurons and 20 were glutamicacid decarboxylase 67 (GAD67)-positive interneurons under theculture condition as confirmed by immunostaining performedafter electrophysiological recording Patch-clamp recording ofUlk2thorn pyramidal neurons revealed altered excitatoryndashinhibitory neurotransmission the frequencies and amplitudesof miniature excitatory postsynaptic currents (mEPSC) were ele-vated whereas the miniature inhibitory postsynaptic currents(mIPSC) amplitudes were downregulated suggesting anexcitatory-dominant state at the resting condition (Fig 2A)Next depolarization-induced increases in calcium [Ca2thorn]i weremeasured in CaMKII-positive pyramidal neurons by transfectingGCaMP Ca2thorn sensor probe in culture (Supplementary MaterialFig S2) Ulk2thorn primary neurons showed higher levels of peakDFF0 and slower rates of fluorescence decay than WT neurons(Fig 2B) These data indicate higher excitability and lower re-sponsiveness to inhibitory signals in Ulk2thorn pyramidal neurons
To examine molecular underpinnings for the observed elec-trophysiological changes in Ulk2thorn neurons levels of surfaceexpression of the major excitatory and inhibitory neurotrans-mitter receptors [ie N-methyl-D-aspartate (NMDA) andgamma-aminobutyric acid (GABA)A receptors] were evaluatedin cultured cortical neurons via biotinylation experiments Bothtotal and surface expression levels of the NMDA receptors wereequivalent between WT and Ulk2thorn neurons (Fig 2C and D) Incontrast levels of surface expression of GABAA receptors (a2and a5 subunits) in Ulk2thorn neurons were lower than in WTwhile the total levels of these receptors remained unchangedbetween the two genotypes (Fig 2C and D)
Reducing the level of p62 expression restores GABAA
receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To address the role of p62 in the regulation of GABAA receptorsurface expression Ulk2thorn mice were bred with p62thorn miceand the levels of GABAA receptor surface expression in primarycortical neurons of Ulk2thorn p62thorn mice were evaluated whencompared with those in control or Ulk2thorn mice Reducing p62gene dosage in Ulk2thorn neurons led to significant up-regulation
of the GABAA receptor surface expression (Fig 3ASupplementary Material Fig S3A and B)
We next evaluated whether reducing p62 gene dosage canrescue behavioral deficits in Ulk2thorn mice PPI deficits were nor-malized to the levels equivalent to those in WT (Fig 3B) cognitiveinflexibility in the rule shifting assay was rescued (Fig 3C and D)Thus p62 not only serves as a marker for attenuated autophagyin neurons but also plays a functional role in the regulation ofGABAA receptor surface presentation as well as behaviors
Pharmacological intervention with rapamycin or its analogCCI-779 is known to enhance autophagic processes anddegrades p62 proteins (2122) Indeed we observed ameliorationof the pathologically elevated p62 levels in Ulk2thorn mice withchronic treatment with CCI-779 (Fig 1B) Importantly the sur-face expression of GABAA receptors in Ulk2thorn cultures was nor-malized by rapamycin treatment (Fig 2C and D) FurthermoreCCI-779 treatment normalized attenuated PPI (Fig 1C and D)and cognitive inflexibility (Fig 1F and G) These results supportthe notion that the observed phenotypes in Ulk2thorn mice arelikely caused by attenuation of autophagic processes involvingp62 Notably PPI deficits in Ulk2thorn mice were not normalizedby clozapine (Fig 1C and D)
Interfering with p62ndashGABARAP interaction restoresGABAA receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To further address the molecular mechanism by which the p62dosage affects GABAA receptor surface presentation we hypoth-esized that elevated p62 levels could titrate out proteins essen-tial for GABAA receptor trafficking p62 interacts with GABAA
receptor-associated protein (GABARAP) (23) a scaffolding pro-tein known to regulate GABAA receptor trafficking (24)Therefore we reasoned that interfering with p62ndashGABARAP in-teraction could liberate GABARAP leading to restoration ofGABAA receptor surface presentation Because 10-amino acidLIR (LC3-interacting region) motif located in the middle of thep62 protein was shown to bind GABARAP (23) we designed a26-amino acid peptide (26aa-peptide) that encompasses this re-gion and tested its potential to interfere with p62ndashGABARAP in-teraction (Fig 4A) When heterologously expressed in HEK293cells this 26aa-peptide inhibited p62ndashGABARAP interaction in adose-dependent manner (Fig 4A) Moreover when the 26aa-peptide was transfected in Ulk2thorn primary cortical neurons thelevel of cell surface GABAA receptor was upregulated whencompared with the neighboring non-transfected cells (Fig 4A)We next investigated its effects on behavioral deficits in Ulk2thorn
mice introducing the 26aa-peptide via adeno-associated virus(AAV)-mediated delivery into the medial PFC significantly im-proved PPI deficits (Fig 4B) and behavioral inflexibility (Fig 4Cand D) The results provide a proof of concept for the role of ele-vated p62 levels in sequestering GABARAP in Ulk2thornmice
p62 is elevated in olfactory neuronal cells in patientswith sporadic cases of SZ and bipolar disorder
Given that cortical excitatoryndashinhibitory imbalance PPI deficitsand behavioral inflexibility are generally seen in patients withpsychiatric conditions such as SZ and bipolar disorder (BP)(2526) we questioned how the molecular signatures associatedwith attenuated autophagic processes could generally be seenas a neuronal trait associated with such psychiatric conditionsAlthough induced pluripotent stem cells-derived neurons are
3168 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
useful for investigating specific disease variant-associated mo-lecular changes this labor-intensive and expensive experimen-tal system is not practically optimal to address questions tosporadic or general cases Instead the use of surrogate tissuesmay be considered (27) Thus we used olfactory neuronal cellsobtained via nasal biopsy (28ndash30) and addressed the questionwith these cells from patients with sporadic cases of SZ and BP
We examined levels of p62 and LC3 proteins in olfactoryneuronal cells biopsied from sporadic cases of SZ and BP whencompared with the age gender and race-matched healthy con-trols (Fig 5A) p62 protein levels were significantly upregulatedin cells derived from patients with SZ and BP (Fig 5BSupplementary Material Fig S4A and B) Furthermore the ratioof LC3-IILC3-I had a trend of upregulation a sign of LC3-II accu-mulation and hence attenuated autophagy (Fig 5CSupplementary Material Fig S4A and B) Note that p62 and Lc3mRNA levels showed no significant differences among groups
(Fig 5D) These results in particular the de-coupling of themRNA and protein levels indicate that attenuated autophagicprocess is a cellular signature in neuronal cells from patientswith sporadic SZ and BP
DiscussionThe present study demonstrates that the Ulk2-mediated processinvolving p62 regulation plays a critical role in higher brainfunctions at least in part through maintenance of theexcitatoryndashinhibitory balance in the PFC The finding also sug-gests that neuronal autophagy regulates not only cellular viabil-ity but also homeostasis of brain functions such as cognitionAlthough the CNV previously reported for Ulk2 likely representsan extremely rare case among SZ the underlying neurobiologi-cal deficit caused by Ulk2 dysfunction (ie excitatoryndashinhibitoryimbalance owing to elevated p62 expression) may provide a
Ulk2 WT
15
WT
mE
PS
C fr
eque
ncy
WT
Ulk2+-
Ulk2+- WT
mE
PS
C a
mpl
itude
Ulk2 +-
+-20pA
100msec
(sec)
WT
mIP
SC
freq
uenc
y
Ulk2+- WT
mIP
SC
am
plitu
de
Ulk2+-
(Hz)(pA)
Pea
k ΔF
F0
0
02
04
03
01
WT Ulk2
10
5
15
10
5
(Hz)
15
10
5
(pA)15
10
5
0000 +-
05
WT Ulk2+-
Nor
mal
ized
FD
R
0
1
15
05
0
02
04
03
01
05
05 1 15 20-05-01
06(au)
NR1GABAARα2
0
05
1
Nor
mal
ized
ratio
(Sur
face
Tot
al)
(au) (au)
0
05
1
A B
C D
GABAARα5
0
05
1
(au)
Rapa Veh Rapa Veh Rapa Veh
Ulk2 WT
+-
Sur
face
Tota
l
Ulk2+-Ulk2+-WT WT
Vehicle Rapa
NR1
NR1
α-Tubulin
p62
50
(kDa)
100
50
100
50
72
GABAARα2
50 GABAARα5
50
GABAARα2
GABAARα5ΔF
F0
Figure 2 Reduced surface expression of GABAA receptors and imbalanced excitatoryinhibitory neurotransmission in Ulk2thorn pyramidal neurons (A) Patch-clamp record-
ing of miniature synaptic currents from WT and Ulk2thorn primary neurons (nfrac1420 each) (16ndash25 DIV) Averages of mEPSC and mIPSC frequencies and amplitudes are plotted in
the graph Plt005 (MannndashWhitney test two-tailed) (B) WT and Ulk2thorn primary cortical neurons were transfected with pCAG-GCaMP6 and [Ca2thorn]i in each transfected neu-
ron was measured following depolarization Representative [Ca2thorn]i changes (DFF0) are plotted and the averaged peak value and the FDR are shown in the graph (nfrac1410 each)
Plt005 (MannndashWhitney test two-tailed) (C D) Surface-biotinylated cell lysates from primary neurons cultured with or without rapamycin (100 nM 30 min) were analyzed
by western blot using anti-GABAA receptor a2 and a5 subunit and NR1 antibodies Experiments were done in triplicate and densitometry analyses were done to evaluate the
surface levels of each receptor by normalizing them against the total levels of respective receptor and plotted in the graph Plt005 (KruskalndashWallis test)
3169|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
general pathophysiological mechanism pertinent to neuropsy-chiatric manifestation including the sporadic cases of SZ
Selective targeting of GABAA receptor by autophagy
Selective downmodulation of GABAA receptors on the surfaceof the pyramidal neuron likely leads to imbalancedexcitatoryndashinhibitory neurotransmission in Ulk2thorn mice Howcould neurons achieve such a selective regulation of a partic-ular neurotransmitter receptor through autophagy A previ-ous study reported selective regulation of GABAA receptor
surface presentation at the neuromuscular junction inCaenorhabditis elegans in which GABAA receptors expressedon the muscle cells are selectively sorted via endocytosisfrom the postsynaptic membrane surface to traffic to theautophagosome upon denervation whereas acetylcholinereceptors in the same cell did not traffic to the autophago-some (31) This study suggests a role of autophagy in selec-tive sorting of GABAA receptors responsible for attenuatedinhibitory neurotransmission At the mechanistic level weshowed that a specific interaction of p62 with GABARAPwhich positively regulates endocytic turnover of GABAA
Sur
face
GA
BA
AR
α1
Ulk2 +-
CaM
KII
Ulk2 p62+- +-
WT 0
1
05
(au)
Nor
mal
ized
exp
ress
ion
(GA
BA
AR
α1C
aMK
II)
WT
Ulk2+-
p62
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
PP
I
0
10
20
30
40
50
60
70WT Ulk2+-
Ulk2 p62+- +-
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
0
10
20
30
40
WT Ulk2+- Ulk2 p62+- +-
Ulk2 p62+- +-
WT Ulk2+-
Ulk2 p62+- +-
WT Ulk2+- Ulk2 p62+- +-
p
Figure 3 Reducing p62 gene dosage rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Primary cortical neurons were prepared
from mice with the indicated genotypes and the cell surface staining was done at 16 DIV with anti-GABAA receptor a1 subunit antibody followed by permeabilization
and further staining with anti-CaMKII and p62 antibodies Scale bar 20 lm Fluorescence intensities per soma were measured using ImageJ Plt 005 (KruskalndashWallis
test) Lower magnification images are shown in Supplementary Material Fig S3A (B) The amplitude of startle response (left) and PPI (right) were evaluated for WT
(nfrac149) Ulk2thorn (nfrac149) and Ulk2thorn p62thorn (nfrac1412) Statistical significance in startle response [F(2 28)frac145548 Pfrac1400093] (one-way ANOVA) and in PPI [F(2 28)frac148761
Pfrac1400011] (two-way ANOVA with repeated measures) P lt005 P lt001 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase
as well as the rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thorn p62thorn (nfrac147) No significant difference during the initial association
phase [F(2 20)frac14124 Pfrac140968] Statistical significance during the rule-shift phase F(2 20)frac14842 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (D)
Numbers of perseverative errors during the rule shift phase were scored F(2 20)frac14527 Pfrac140038 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3170 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
receptors (24) may be important elevated p62 expression inUlk2thorn pyramidal neurons appears to sequester a higher pro-portion of GABARAP and limit the amount of GABARAP avail-able for GABAA receptor surface presentation
Linking the data from the animal model and patientneuronal cells
Although both p62 and Ulk2 are ubiquitously expressed in allcell types in every tissue tested (1132) it is currently unknown
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p69-120 p79-120 p83-120
()
P
PI
0
10
20
30
40
50
60
70
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
WT Ulk2+- Ulk2 +-
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
+Cont +Cont +26aa
0
10
20
30
WT Ulk2+- Ulk2 +-
+Cont +Cont +26aa
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
GFP-p62 + + + + 26aa-peptide-FLAG
myc-GABARAP + + + +
50
100(kDa)
IP with myc
(IgG-HC)
WBanti-GFP
anti-GFP
IP
Input
50
anti-myc
26aa-FLAGanti-FLAG
Surface GABAARα1 GFP-26aa-peptide
Structure of Sequestosome-1p62
328-MESGNCSGGDDDWTHLSSKEVDPSTG-353
ZZ UBAPB1
LIR 4423911021
(26aa-peptide)
GABARAP
GFP-p62
GFP-p62
GABARAP
(dot blot)
Figure 4 Interfering with p62ndashGABARAP interaction rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Schematic illustration of
sequestosome-1p62 protein structure and designing strategy for the 26-amino acid peptide that interferes with p62ndashGABARAP interaction The LIR (LC3-interacting re-
gion) contains a DDDWxxL motif that was shown to interact with GABARAP (23) Primary cortical neurons prepared from Ulk2thorn mice were transfected at 16DIV with
an expression plasmid encoding GFP-26aa-peptide and the cell surface GABAA receptor (a1 subunit) was immuno-stained without cell permeabilization White arrow
indicates a cell transfected with GFP-26aa-peptide and open arrows indicate neighboring non-transfected cells Scale bar 20 lm (B) The amplitude of startle response
(left) and PPI (right) were evaluated for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (nfrac147) and Ulk2thorn with 26aa-peptide AAV (nfrac146) No significant
difference in startle response [F(2 17)frac141045 Pfrac140833] (one-way ANOVA) Statistical significance for PPI F(2 17)frac14853 P frac1400126 (two-way ANOVA with repeated meas-
ures) P lt005 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase as well as the rule shift phase of the assays were scored
for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (n frac147) and Ulk2thorn p62thorn with 26aa-peptide AAV (nfrac146) P lt005 P lt001 No significant difference
during the initial association phase [F(2 17)frac14102 P frac140827] Statistical significance during the rule-shift phase F(2 17)frac14774 Pfrac140018 (one-way ANOVA) P lt005
P lt001 (Bonferroni post hoc test) (D) Numbers of perseverative errors during the rule shift phase were scored F(2 17)frac14458 P 50046 (one-way ANOVA) P lt005Pfrac140057 (Bonferroni post hoc test)
3171|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

Thus the current study reveals a novel mechanism linking deregulated autophagy to functional disturbances of the nervoussystem relevant to SZ through regulation of GABAA receptor surface presentation in pyramidal neurons
IntroductionMacroautophagy (hereafter referred to as autophagy) is a spe-cialized membrane trafficking machinery and a major cellularrecycling system responsible for degrading old proteins anddamaged organelles which ultimately serve as resources togenerate new proteins and energy and thus critical to the main-tenance of cellular homeostasis (1) In response to a range ofcellular stresses (eg depletion of nutrients and energy mis-folded protein accumulation and oxidative stress) autophagy isactivated to mitigate such stresses via activation of ULK1ULK2ATG1 serinethreonine protein kinases critical to the regulationof autophagy induction Subsequently microtubule-associatedprotein light chain 3 (LC3)-positive autophagosomes are formedto enwrap the proteins and organelles that are selectively la-beled with sequestosome-1p62 an autophagy-related stress re-sponse protein and then trafficked to fuse with lysosomes fordegradation
A role of autophagy in the nervous system was previouslyshown in nervous system-specific Atg5 and Atg7 conditionalknockout (cKO) mice which exhibit progressive neuronal deathaccompanied by accumulation of abnormal cytosolic proteinsand formation of p62-positive inclusions in neurons (2ndash4)These results suggested that continuous quality control of dif-fuse cytosolic proteins via basal autophagy is important for neu-ronal functions Subsequently we and others providedevidence that autophagy regulates synapse morphogenesis andfunction using a series of autophagy-deficient mutants inDrosophila melanogaster which demonstrate diminished size andnumber of synaptic boutons and a decrease in evoked neuro-transmitter release (5ndash8) Meanwhile ULK1ULK2ATG1 regulateendocytic trafficking of neuronal cargoes through associationwith a synaptic protein SynGAP (9) and also regulate traffickingof synaptic vesicles via association with additional neuronalproteins such as Unc-76FEZ1 and synaptotagmin-1 (5) In ac-cordance with these observations dopaminergic neuron-specific Atg7-cKO mice showed a role of autophagy in presynap-tic structure and neurotransmission exhibiting enlarged stria-tal axon terminal profiles and abnormally increased dopaminerelease (10) It is thus possible that autophagy-mediated regula-tion of pre- or post-synaptic components via modulation oftheir trafficking could contribute to neural plasticity Howeverquestions remain as to (i) whether neuronal autophagy is in-volved in higher brain functions such as cognition (ii) if sowhich brain regions and neuronal cell types are most vulnerableto impaired autophagy leading to specific behavioral outcomesand (iii) what are molecular mechanisms that account for suchoutcomes
A recent study showed that mice doubly deficient for Ulk1and Ulk2 in the nervous system have a slight increase in the lev-els of p62 protein a key target molecule in autophagic proteindegradation without showing a sign of p62-positive inclusions(11) Likewise mice deficient for Ulk1 had increased levels of p62protein expression in the prefrontal cortex (PFC) (12)Importantly unlike the knockout of non-redundant autophagygenes such as Atg5 and Atg7 that display early postnatal deathin mice and may possibly mask the underlying neural mecha-nisms Ulk1 and Ulk2 have redundant roles in autophagy andmice singly deficient for either of these genes are viable with no
overt deficits in overall physical conditions such as life spanbody weight and motor functions (12ndash15) Thus we regard thatperturbation of Ulk1 andor Ulk2 will provide the best model toaddress the current questions With this model it is expectedthat we could address whether this pathway plays a causal rolein neurobehavioral manifestation In this regard it is notablethat the copy number variations (CNVs) of ULK1ULK2ULK4family member genes were recently reported to be enriched inschizophrenia (SZ) patients recruited in a genome-wide associa-tion study (odds ratio frac14677) (16)
In the present study we set out to test whether Ulk2 heterozy-gous (Ulk2thorn) mice have signs of attenuated autophagy (ie ele-vated p62 expression) in certain regions or cell types within thebrain and addressed whether such changes can be linked to neu-robehavioral outcomes We hypothesized that elevated p62 lev-els have functional consequences on neurotransmission thatcould account for the observed behavioral deficits relevant to SZ
ResultsUlk2thornmice show elevated p62 protein levels in the PFC
As described above changes in the expression of p62 proteinhave been reported in both Ulk1 and Ulk2 knockout mice (1112)However a question remains as to which brain regions and cellsmost robustly represent such changes Thus we first examinedthe expression of p62 protein in various brain regions of Ulk2thorn
mice The mutants had significantly elevated expression of p62protein in the medial PFC (Fig 1A and B SupplementaryMaterial Fig S1A and B) which occurred in Ca2thorncalmodulin-dependent protein kinase II (CaMKII)-positive pyramidal neu-rons but not in the interneurons (Fig 1A SupplementaryMaterial Fig S1B) This elevated p62 expression was seen in thePFC but not in all other brain regions tested including the vi-sual cortex nucleus accumbens dorsal striatum hippocampusand ventral tegmental area (Supplementary Material Fig S1A)Taken together reduced expression of Ulk2 affects cellular pro-cesses involving p62 protein likely associated with autophagichomeostasis in a neuronal cell-type preferential and a brainregion-preferential manner
Ulk2thornmice show behavioral deficits associated withthe PFC function
We then questioned whether and how the reduction in Ulk2affects the PFC-relevant behaviors including sensorimotor gat-ing (17) and cognitive flexibility (18) Ulk2thorn mice had impairedsensorimotor gating showing lower levels of prepulse inhibi-tion (PPI) of acoustic startle stimuli (Fig 1C and D) To assess thePFC-dependent cognitive function Ulk2thornmice were tested in arule shifting paradigm (1920) in which mice were initiallytrained to associate food reward with a specific stimulus (ie ei-ther an odor or a digging medium) and subsequently evaluatedfor cognitive flexibility by changing the type of stimulus thatpredicts the reward (Fig 1E) Although both Ulk2thorn and WTmice learned the association rule in a similar number of trialsduring the initial association phase of trials Ulk2thorn mice werespecifically impaired during the rule shifting phase of trials(Fig 1F and G)
3166 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
WT WT
Saline CCI-779
Ulk2+-
Ulk2+-DAPI
WT
Ulk2+-
p62
LC3-I
α-Tubulin
LC3-II
p62 CaMKII
LC3-IITotal LC3
Nor
mal
ized
rat
iop62
Nor
mal
ized
leve
ls
1
0
2 (au)
CCI-779
WT
Ulk2+-
1
0
2 (au)
WT
Ulk2+-
50
(kDa)
17
72
WT + Veh Ulk2 + Veh Ulk2 + CCI-779Ulk2 + Clozapine
+-+-+-
0
500
(au)
Sta
rtle
res
pons
e
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
P
PI
0
20
40
60
80
A B
C D
E
F
WT
CCI-779
Ulk2+- Ulk2+-
+
O1TA
O2
TB
O1
TB
O2
TA
O1
TA
O2
TB
O1
TB
O2
TA
Initial association Rule shiftTrial 1 Trial n Trial 1 Trial n
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Ulk2 + CCI-779
WT + Veh Ulk2 + Veh
+-
+-
Num
ber
of p
erse
vera
tive
erro
rs
G
0
10
20
30
40
CCI-779
WT
Ulk2+-
WT
Ulk2+-
Veh+
Veh+
WT
CCI-779
Ulk2+- Ulk2+-
+Veh
+Veh
+
Figure 1 Upregulated p62 protein levels in pyramidal neurons of the PFC and PFC-associated behavioral deficits in Ulk2thorn mice (A) Selective upregulation of p62 pro-
tein in CaMKIIthorn neurons in the PFC of Ulk2thornmice Scale bar 10 lm (B) Western blot of p62 and LC3 in the frontal cortex of Ulk2thornmice chronically treated with saline
or a rapamycin analog CCI-779 The expression levels were measured by densitometric analysis using ImageJ Error bars indicate SEM P lt005 (C D) The amplitude of
startle response (C) and the percentage of PPI (D) were evaluated for WTthornsaline (nfrac1410) Ulk2thornthornsaline (nfrac1410) Ulk2thornthornCCI-779 (nfrac1410) and Ulk2thornthornclozapine (nfrac1410)
No significant difference in startle response [F(3 36)frac1409683 Pfrac1404183] (one-way ANOVA) Statistical significance for PPI F(3 36)frac141128 P lt00001 (two-way ANOVA with
repeated measures) P lt005 (Bonferroni post hoc test) (E) Schematic diagram of the rule shift assay Upon rule shifting the stimulus associated with food reward is
changed from odor cue 1 (underlined) to texture cue A (italicized) in this diagram (F) Numbers of trials to criterion during the initial association phase as well as the
rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thornthornCCI-779 (nfrac147) No significant difference during the initial association phase [F(2
20)frac14154 Pfrac140832] Statistical significance during the rule-shift phase F(2 20)frac14767 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (G) Numbers of per-
severative errors during the rule shift phase were scored F(2 20)frac14483 Pfrac140024 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3167|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
CaMKII-specific Ulk2thornmice show behavioral deficitsassociated with the PFC function
We next investigated specific roles of neuronal subtypes in theobserved phenotypes Mice heterozygous for conditional Ulk2knockout allele were crossed with mice expressing eitherpan-specific Cre (EIIa-Cre) pyramidal neuron-specific Cre(CaMKII-Cre) or interneuron-specific Cre (Viaat-Cre) and PPIwas measured When Ulk2 was ablated in an interneuron-specific manner PPI was not affected (Supplementary MaterialFig S1CndashE) In contrast pyramidal neuron-specific Ulk2 ablationresulted in attenuated PPI (Supplementary Material Fig S1C Dand F) These data are in line with the notion that homeostaticdeficits involving p62 accumulation in the CaMKII-positive py-ramidal neurons underlie the PPI deficits in Ulk2thornmice
Reduced surface expression of GABAA receptors andimbalanced excitatoryndashinhibitory neurotransmission inUlk2thorn pyramidal neurons
To further investigate molecular and cellular basis that may un-derlie these behavioral changes electrophysiological propertiesof the cultured pyramidal neurons prepared from Ulk2thorn micewere examined Typically 80 of neurons in culture wereCaMKII-positive pyramidal neurons and 20 were glutamicacid decarboxylase 67 (GAD67)-positive interneurons under theculture condition as confirmed by immunostaining performedafter electrophysiological recording Patch-clamp recording ofUlk2thorn pyramidal neurons revealed altered excitatoryndashinhibitory neurotransmission the frequencies and amplitudesof miniature excitatory postsynaptic currents (mEPSC) were ele-vated whereas the miniature inhibitory postsynaptic currents(mIPSC) amplitudes were downregulated suggesting anexcitatory-dominant state at the resting condition (Fig 2A)Next depolarization-induced increases in calcium [Ca2thorn]i weremeasured in CaMKII-positive pyramidal neurons by transfectingGCaMP Ca2thorn sensor probe in culture (Supplementary MaterialFig S2) Ulk2thorn primary neurons showed higher levels of peakDFF0 and slower rates of fluorescence decay than WT neurons(Fig 2B) These data indicate higher excitability and lower re-sponsiveness to inhibitory signals in Ulk2thorn pyramidal neurons
To examine molecular underpinnings for the observed elec-trophysiological changes in Ulk2thorn neurons levels of surfaceexpression of the major excitatory and inhibitory neurotrans-mitter receptors [ie N-methyl-D-aspartate (NMDA) andgamma-aminobutyric acid (GABA)A receptors] were evaluatedin cultured cortical neurons via biotinylation experiments Bothtotal and surface expression levels of the NMDA receptors wereequivalent between WT and Ulk2thorn neurons (Fig 2C and D) Incontrast levels of surface expression of GABAA receptors (a2and a5 subunits) in Ulk2thorn neurons were lower than in WTwhile the total levels of these receptors remained unchangedbetween the two genotypes (Fig 2C and D)
Reducing the level of p62 expression restores GABAA
receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To address the role of p62 in the regulation of GABAA receptorsurface expression Ulk2thorn mice were bred with p62thorn miceand the levels of GABAA receptor surface expression in primarycortical neurons of Ulk2thorn p62thorn mice were evaluated whencompared with those in control or Ulk2thorn mice Reducing p62gene dosage in Ulk2thorn neurons led to significant up-regulation
of the GABAA receptor surface expression (Fig 3ASupplementary Material Fig S3A and B)
We next evaluated whether reducing p62 gene dosage canrescue behavioral deficits in Ulk2thorn mice PPI deficits were nor-malized to the levels equivalent to those in WT (Fig 3B) cognitiveinflexibility in the rule shifting assay was rescued (Fig 3C and D)Thus p62 not only serves as a marker for attenuated autophagyin neurons but also plays a functional role in the regulation ofGABAA receptor surface presentation as well as behaviors
Pharmacological intervention with rapamycin or its analogCCI-779 is known to enhance autophagic processes anddegrades p62 proteins (2122) Indeed we observed ameliorationof the pathologically elevated p62 levels in Ulk2thorn mice withchronic treatment with CCI-779 (Fig 1B) Importantly the sur-face expression of GABAA receptors in Ulk2thorn cultures was nor-malized by rapamycin treatment (Fig 2C and D) FurthermoreCCI-779 treatment normalized attenuated PPI (Fig 1C and D)and cognitive inflexibility (Fig 1F and G) These results supportthe notion that the observed phenotypes in Ulk2thorn mice arelikely caused by attenuation of autophagic processes involvingp62 Notably PPI deficits in Ulk2thorn mice were not normalizedby clozapine (Fig 1C and D)
Interfering with p62ndashGABARAP interaction restoresGABAA receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To further address the molecular mechanism by which the p62dosage affects GABAA receptor surface presentation we hypoth-esized that elevated p62 levels could titrate out proteins essen-tial for GABAA receptor trafficking p62 interacts with GABAA
receptor-associated protein (GABARAP) (23) a scaffolding pro-tein known to regulate GABAA receptor trafficking (24)Therefore we reasoned that interfering with p62ndashGABARAP in-teraction could liberate GABARAP leading to restoration ofGABAA receptor surface presentation Because 10-amino acidLIR (LC3-interacting region) motif located in the middle of thep62 protein was shown to bind GABARAP (23) we designed a26-amino acid peptide (26aa-peptide) that encompasses this re-gion and tested its potential to interfere with p62ndashGABARAP in-teraction (Fig 4A) When heterologously expressed in HEK293cells this 26aa-peptide inhibited p62ndashGABARAP interaction in adose-dependent manner (Fig 4A) Moreover when the 26aa-peptide was transfected in Ulk2thorn primary cortical neurons thelevel of cell surface GABAA receptor was upregulated whencompared with the neighboring non-transfected cells (Fig 4A)We next investigated its effects on behavioral deficits in Ulk2thorn
mice introducing the 26aa-peptide via adeno-associated virus(AAV)-mediated delivery into the medial PFC significantly im-proved PPI deficits (Fig 4B) and behavioral inflexibility (Fig 4Cand D) The results provide a proof of concept for the role of ele-vated p62 levels in sequestering GABARAP in Ulk2thornmice
p62 is elevated in olfactory neuronal cells in patientswith sporadic cases of SZ and bipolar disorder
Given that cortical excitatoryndashinhibitory imbalance PPI deficitsand behavioral inflexibility are generally seen in patients withpsychiatric conditions such as SZ and bipolar disorder (BP)(2526) we questioned how the molecular signatures associatedwith attenuated autophagic processes could generally be seenas a neuronal trait associated with such psychiatric conditionsAlthough induced pluripotent stem cells-derived neurons are
3168 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
useful for investigating specific disease variant-associated mo-lecular changes this labor-intensive and expensive experimen-tal system is not practically optimal to address questions tosporadic or general cases Instead the use of surrogate tissuesmay be considered (27) Thus we used olfactory neuronal cellsobtained via nasal biopsy (28ndash30) and addressed the questionwith these cells from patients with sporadic cases of SZ and BP
We examined levels of p62 and LC3 proteins in olfactoryneuronal cells biopsied from sporadic cases of SZ and BP whencompared with the age gender and race-matched healthy con-trols (Fig 5A) p62 protein levels were significantly upregulatedin cells derived from patients with SZ and BP (Fig 5BSupplementary Material Fig S4A and B) Furthermore the ratioof LC3-IILC3-I had a trend of upregulation a sign of LC3-II accu-mulation and hence attenuated autophagy (Fig 5CSupplementary Material Fig S4A and B) Note that p62 and Lc3mRNA levels showed no significant differences among groups
(Fig 5D) These results in particular the de-coupling of themRNA and protein levels indicate that attenuated autophagicprocess is a cellular signature in neuronal cells from patientswith sporadic SZ and BP
DiscussionThe present study demonstrates that the Ulk2-mediated processinvolving p62 regulation plays a critical role in higher brainfunctions at least in part through maintenance of theexcitatoryndashinhibitory balance in the PFC The finding also sug-gests that neuronal autophagy regulates not only cellular viabil-ity but also homeostasis of brain functions such as cognitionAlthough the CNV previously reported for Ulk2 likely representsan extremely rare case among SZ the underlying neurobiologi-cal deficit caused by Ulk2 dysfunction (ie excitatoryndashinhibitoryimbalance owing to elevated p62 expression) may provide a
Ulk2 WT
15
WT
mE
PS
C fr
eque
ncy
WT
Ulk2+-
Ulk2+- WT
mE
PS
C a
mpl
itude
Ulk2 +-
+-20pA
100msec
(sec)
WT
mIP
SC
freq
uenc
y
Ulk2+- WT
mIP
SC
am
plitu
de
Ulk2+-
(Hz)(pA)
Pea
k ΔF
F0
0
02
04
03
01
WT Ulk2
10
5
15
10
5
(Hz)
15
10
5
(pA)15
10
5
0000 +-
05
WT Ulk2+-
Nor
mal
ized
FD
R
0
1
15
05
0
02
04
03
01
05
05 1 15 20-05-01
06(au)
NR1GABAARα2
0
05
1
Nor
mal
ized
ratio
(Sur
face
Tot
al)
(au) (au)
0
05
1
A B
C D
GABAARα5
0
05
1
(au)
Rapa Veh Rapa Veh Rapa Veh
Ulk2 WT
+-
Sur
face
Tota
l
Ulk2+-Ulk2+-WT WT
Vehicle Rapa
NR1
NR1
α-Tubulin
p62
50
(kDa)
100
50
100
50
72
GABAARα2
50 GABAARα5
50
GABAARα2
GABAARα5ΔF
F0
Figure 2 Reduced surface expression of GABAA receptors and imbalanced excitatoryinhibitory neurotransmission in Ulk2thorn pyramidal neurons (A) Patch-clamp record-
ing of miniature synaptic currents from WT and Ulk2thorn primary neurons (nfrac1420 each) (16ndash25 DIV) Averages of mEPSC and mIPSC frequencies and amplitudes are plotted in
the graph Plt005 (MannndashWhitney test two-tailed) (B) WT and Ulk2thorn primary cortical neurons were transfected with pCAG-GCaMP6 and [Ca2thorn]i in each transfected neu-
ron was measured following depolarization Representative [Ca2thorn]i changes (DFF0) are plotted and the averaged peak value and the FDR are shown in the graph (nfrac1410 each)
Plt005 (MannndashWhitney test two-tailed) (C D) Surface-biotinylated cell lysates from primary neurons cultured with or without rapamycin (100 nM 30 min) were analyzed
by western blot using anti-GABAA receptor a2 and a5 subunit and NR1 antibodies Experiments were done in triplicate and densitometry analyses were done to evaluate the
surface levels of each receptor by normalizing them against the total levels of respective receptor and plotted in the graph Plt005 (KruskalndashWallis test)
3169|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
general pathophysiological mechanism pertinent to neuropsy-chiatric manifestation including the sporadic cases of SZ
Selective targeting of GABAA receptor by autophagy
Selective downmodulation of GABAA receptors on the surfaceof the pyramidal neuron likely leads to imbalancedexcitatoryndashinhibitory neurotransmission in Ulk2thorn mice Howcould neurons achieve such a selective regulation of a partic-ular neurotransmitter receptor through autophagy A previ-ous study reported selective regulation of GABAA receptor
surface presentation at the neuromuscular junction inCaenorhabditis elegans in which GABAA receptors expressedon the muscle cells are selectively sorted via endocytosisfrom the postsynaptic membrane surface to traffic to theautophagosome upon denervation whereas acetylcholinereceptors in the same cell did not traffic to the autophago-some (31) This study suggests a role of autophagy in selec-tive sorting of GABAA receptors responsible for attenuatedinhibitory neurotransmission At the mechanistic level weshowed that a specific interaction of p62 with GABARAPwhich positively regulates endocytic turnover of GABAA
Sur
face
GA
BA
AR
α1
Ulk2 +-
CaM
KII
Ulk2 p62+- +-
WT 0
1
05
(au)
Nor
mal
ized
exp
ress
ion
(GA
BA
AR
α1C
aMK
II)
WT
Ulk2+-
p62
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
PP
I
0
10
20
30
40
50
60
70WT Ulk2+-
Ulk2 p62+- +-
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
0
10
20
30
40
WT Ulk2+- Ulk2 p62+- +-
Ulk2 p62+- +-
WT Ulk2+-
Ulk2 p62+- +-
WT Ulk2+- Ulk2 p62+- +-
p
Figure 3 Reducing p62 gene dosage rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Primary cortical neurons were prepared
from mice with the indicated genotypes and the cell surface staining was done at 16 DIV with anti-GABAA receptor a1 subunit antibody followed by permeabilization
and further staining with anti-CaMKII and p62 antibodies Scale bar 20 lm Fluorescence intensities per soma were measured using ImageJ Plt 005 (KruskalndashWallis
test) Lower magnification images are shown in Supplementary Material Fig S3A (B) The amplitude of startle response (left) and PPI (right) were evaluated for WT
(nfrac149) Ulk2thorn (nfrac149) and Ulk2thorn p62thorn (nfrac1412) Statistical significance in startle response [F(2 28)frac145548 Pfrac1400093] (one-way ANOVA) and in PPI [F(2 28)frac148761
Pfrac1400011] (two-way ANOVA with repeated measures) P lt005 P lt001 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase
as well as the rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thorn p62thorn (nfrac147) No significant difference during the initial association
phase [F(2 20)frac14124 Pfrac140968] Statistical significance during the rule-shift phase F(2 20)frac14842 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (D)
Numbers of perseverative errors during the rule shift phase were scored F(2 20)frac14527 Pfrac140038 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3170 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
receptors (24) may be important elevated p62 expression inUlk2thorn pyramidal neurons appears to sequester a higher pro-portion of GABARAP and limit the amount of GABARAP avail-able for GABAA receptor surface presentation
Linking the data from the animal model and patientneuronal cells
Although both p62 and Ulk2 are ubiquitously expressed in allcell types in every tissue tested (1132) it is currently unknown
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p69-120 p79-120 p83-120
()
P
PI
0
10
20
30
40
50
60
70
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
WT Ulk2+- Ulk2 +-
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
+Cont +Cont +26aa
0
10
20
30
WT Ulk2+- Ulk2 +-
+Cont +Cont +26aa
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
GFP-p62 + + + + 26aa-peptide-FLAG
myc-GABARAP + + + +
50
100(kDa)
IP with myc
(IgG-HC)
WBanti-GFP
anti-GFP
IP
Input
50
anti-myc
26aa-FLAGanti-FLAG
Surface GABAARα1 GFP-26aa-peptide
Structure of Sequestosome-1p62
328-MESGNCSGGDDDWTHLSSKEVDPSTG-353
ZZ UBAPB1
LIR 4423911021
(26aa-peptide)
GABARAP
GFP-p62
GFP-p62
GABARAP
(dot blot)
Figure 4 Interfering with p62ndashGABARAP interaction rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Schematic illustration of
sequestosome-1p62 protein structure and designing strategy for the 26-amino acid peptide that interferes with p62ndashGABARAP interaction The LIR (LC3-interacting re-
gion) contains a DDDWxxL motif that was shown to interact with GABARAP (23) Primary cortical neurons prepared from Ulk2thorn mice were transfected at 16DIV with
an expression plasmid encoding GFP-26aa-peptide and the cell surface GABAA receptor (a1 subunit) was immuno-stained without cell permeabilization White arrow
indicates a cell transfected with GFP-26aa-peptide and open arrows indicate neighboring non-transfected cells Scale bar 20 lm (B) The amplitude of startle response
(left) and PPI (right) were evaluated for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (nfrac147) and Ulk2thorn with 26aa-peptide AAV (nfrac146) No significant
difference in startle response [F(2 17)frac141045 Pfrac140833] (one-way ANOVA) Statistical significance for PPI F(2 17)frac14853 P frac1400126 (two-way ANOVA with repeated meas-
ures) P lt005 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase as well as the rule shift phase of the assays were scored
for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (n frac147) and Ulk2thorn p62thorn with 26aa-peptide AAV (nfrac146) P lt005 P lt001 No significant difference
during the initial association phase [F(2 17)frac14102 P frac140827] Statistical significance during the rule-shift phase F(2 17)frac14774 Pfrac140018 (one-way ANOVA) P lt005
P lt001 (Bonferroni post hoc test) (D) Numbers of perseverative errors during the rule shift phase were scored F(2 17)frac14458 P 50046 (one-way ANOVA) P lt005Pfrac140057 (Bonferroni post hoc test)
3171|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
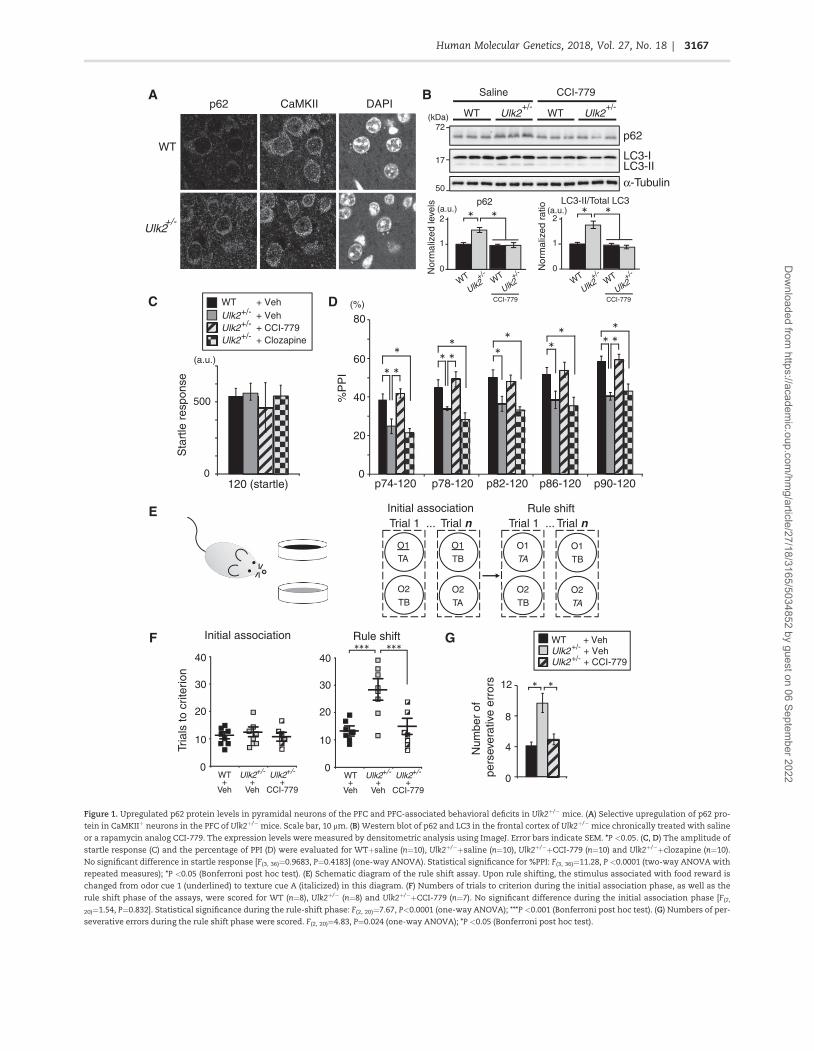
WT WT
Saline CCI-779
Ulk2+-
Ulk2+-DAPI
WT
Ulk2+-
p62
LC3-I
α-Tubulin
LC3-II
p62 CaMKII
LC3-IITotal LC3
Nor
mal
ized
rat
iop62
Nor
mal
ized
leve
ls
1
0
2 (au)
CCI-779
WT
Ulk2+-
1
0
2 (au)
WT
Ulk2+-
50
(kDa)
17
72
WT + Veh Ulk2 + Veh Ulk2 + CCI-779Ulk2 + Clozapine
+-+-+-
0
500
(au)
Sta
rtle
res
pons
e
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
P
PI
0
20
40
60
80
A B
C D
E
F
WT
CCI-779
Ulk2+- Ulk2+-
+
O1TA
O2
TB
O1
TB
O2
TA
O1
TA
O2
TB
O1
TB
O2
TA
Initial association Rule shiftTrial 1 Trial n Trial 1 Trial n
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Ulk2 + CCI-779
WT + Veh Ulk2 + Veh
+-
+-
Num
ber
of p
erse
vera
tive
erro
rs
G
0
10
20
30
40
CCI-779
WT
Ulk2+-
WT
Ulk2+-
Veh+
Veh+
WT
CCI-779
Ulk2+- Ulk2+-
+Veh
+Veh
+
Figure 1 Upregulated p62 protein levels in pyramidal neurons of the PFC and PFC-associated behavioral deficits in Ulk2thorn mice (A) Selective upregulation of p62 pro-
tein in CaMKIIthorn neurons in the PFC of Ulk2thornmice Scale bar 10 lm (B) Western blot of p62 and LC3 in the frontal cortex of Ulk2thornmice chronically treated with saline
or a rapamycin analog CCI-779 The expression levels were measured by densitometric analysis using ImageJ Error bars indicate SEM P lt005 (C D) The amplitude of
startle response (C) and the percentage of PPI (D) were evaluated for WTthornsaline (nfrac1410) Ulk2thornthornsaline (nfrac1410) Ulk2thornthornCCI-779 (nfrac1410) and Ulk2thornthornclozapine (nfrac1410)
No significant difference in startle response [F(3 36)frac1409683 Pfrac1404183] (one-way ANOVA) Statistical significance for PPI F(3 36)frac141128 P lt00001 (two-way ANOVA with
repeated measures) P lt005 (Bonferroni post hoc test) (E) Schematic diagram of the rule shift assay Upon rule shifting the stimulus associated with food reward is
changed from odor cue 1 (underlined) to texture cue A (italicized) in this diagram (F) Numbers of trials to criterion during the initial association phase as well as the
rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thornthornCCI-779 (nfrac147) No significant difference during the initial association phase [F(2
20)frac14154 Pfrac140832] Statistical significance during the rule-shift phase F(2 20)frac14767 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (G) Numbers of per-
severative errors during the rule shift phase were scored F(2 20)frac14483 Pfrac140024 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3167|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
CaMKII-specific Ulk2thornmice show behavioral deficitsassociated with the PFC function
We next investigated specific roles of neuronal subtypes in theobserved phenotypes Mice heterozygous for conditional Ulk2knockout allele were crossed with mice expressing eitherpan-specific Cre (EIIa-Cre) pyramidal neuron-specific Cre(CaMKII-Cre) or interneuron-specific Cre (Viaat-Cre) and PPIwas measured When Ulk2 was ablated in an interneuron-specific manner PPI was not affected (Supplementary MaterialFig S1CndashE) In contrast pyramidal neuron-specific Ulk2 ablationresulted in attenuated PPI (Supplementary Material Fig S1C Dand F) These data are in line with the notion that homeostaticdeficits involving p62 accumulation in the CaMKII-positive py-ramidal neurons underlie the PPI deficits in Ulk2thornmice
Reduced surface expression of GABAA receptors andimbalanced excitatoryndashinhibitory neurotransmission inUlk2thorn pyramidal neurons
To further investigate molecular and cellular basis that may un-derlie these behavioral changes electrophysiological propertiesof the cultured pyramidal neurons prepared from Ulk2thorn micewere examined Typically 80 of neurons in culture wereCaMKII-positive pyramidal neurons and 20 were glutamicacid decarboxylase 67 (GAD67)-positive interneurons under theculture condition as confirmed by immunostaining performedafter electrophysiological recording Patch-clamp recording ofUlk2thorn pyramidal neurons revealed altered excitatoryndashinhibitory neurotransmission the frequencies and amplitudesof miniature excitatory postsynaptic currents (mEPSC) were ele-vated whereas the miniature inhibitory postsynaptic currents(mIPSC) amplitudes were downregulated suggesting anexcitatory-dominant state at the resting condition (Fig 2A)Next depolarization-induced increases in calcium [Ca2thorn]i weremeasured in CaMKII-positive pyramidal neurons by transfectingGCaMP Ca2thorn sensor probe in culture (Supplementary MaterialFig S2) Ulk2thorn primary neurons showed higher levels of peakDFF0 and slower rates of fluorescence decay than WT neurons(Fig 2B) These data indicate higher excitability and lower re-sponsiveness to inhibitory signals in Ulk2thorn pyramidal neurons
To examine molecular underpinnings for the observed elec-trophysiological changes in Ulk2thorn neurons levels of surfaceexpression of the major excitatory and inhibitory neurotrans-mitter receptors [ie N-methyl-D-aspartate (NMDA) andgamma-aminobutyric acid (GABA)A receptors] were evaluatedin cultured cortical neurons via biotinylation experiments Bothtotal and surface expression levels of the NMDA receptors wereequivalent between WT and Ulk2thorn neurons (Fig 2C and D) Incontrast levels of surface expression of GABAA receptors (a2and a5 subunits) in Ulk2thorn neurons were lower than in WTwhile the total levels of these receptors remained unchangedbetween the two genotypes (Fig 2C and D)
Reducing the level of p62 expression restores GABAA
receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To address the role of p62 in the regulation of GABAA receptorsurface expression Ulk2thorn mice were bred with p62thorn miceand the levels of GABAA receptor surface expression in primarycortical neurons of Ulk2thorn p62thorn mice were evaluated whencompared with those in control or Ulk2thorn mice Reducing p62gene dosage in Ulk2thorn neurons led to significant up-regulation
of the GABAA receptor surface expression (Fig 3ASupplementary Material Fig S3A and B)
We next evaluated whether reducing p62 gene dosage canrescue behavioral deficits in Ulk2thorn mice PPI deficits were nor-malized to the levels equivalent to those in WT (Fig 3B) cognitiveinflexibility in the rule shifting assay was rescued (Fig 3C and D)Thus p62 not only serves as a marker for attenuated autophagyin neurons but also plays a functional role in the regulation ofGABAA receptor surface presentation as well as behaviors
Pharmacological intervention with rapamycin or its analogCCI-779 is known to enhance autophagic processes anddegrades p62 proteins (2122) Indeed we observed ameliorationof the pathologically elevated p62 levels in Ulk2thorn mice withchronic treatment with CCI-779 (Fig 1B) Importantly the sur-face expression of GABAA receptors in Ulk2thorn cultures was nor-malized by rapamycin treatment (Fig 2C and D) FurthermoreCCI-779 treatment normalized attenuated PPI (Fig 1C and D)and cognitive inflexibility (Fig 1F and G) These results supportthe notion that the observed phenotypes in Ulk2thorn mice arelikely caused by attenuation of autophagic processes involvingp62 Notably PPI deficits in Ulk2thorn mice were not normalizedby clozapine (Fig 1C and D)
Interfering with p62ndashGABARAP interaction restoresGABAA receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To further address the molecular mechanism by which the p62dosage affects GABAA receptor surface presentation we hypoth-esized that elevated p62 levels could titrate out proteins essen-tial for GABAA receptor trafficking p62 interacts with GABAA
receptor-associated protein (GABARAP) (23) a scaffolding pro-tein known to regulate GABAA receptor trafficking (24)Therefore we reasoned that interfering with p62ndashGABARAP in-teraction could liberate GABARAP leading to restoration ofGABAA receptor surface presentation Because 10-amino acidLIR (LC3-interacting region) motif located in the middle of thep62 protein was shown to bind GABARAP (23) we designed a26-amino acid peptide (26aa-peptide) that encompasses this re-gion and tested its potential to interfere with p62ndashGABARAP in-teraction (Fig 4A) When heterologously expressed in HEK293cells this 26aa-peptide inhibited p62ndashGABARAP interaction in adose-dependent manner (Fig 4A) Moreover when the 26aa-peptide was transfected in Ulk2thorn primary cortical neurons thelevel of cell surface GABAA receptor was upregulated whencompared with the neighboring non-transfected cells (Fig 4A)We next investigated its effects on behavioral deficits in Ulk2thorn
mice introducing the 26aa-peptide via adeno-associated virus(AAV)-mediated delivery into the medial PFC significantly im-proved PPI deficits (Fig 4B) and behavioral inflexibility (Fig 4Cand D) The results provide a proof of concept for the role of ele-vated p62 levels in sequestering GABARAP in Ulk2thornmice
p62 is elevated in olfactory neuronal cells in patientswith sporadic cases of SZ and bipolar disorder
Given that cortical excitatoryndashinhibitory imbalance PPI deficitsand behavioral inflexibility are generally seen in patients withpsychiatric conditions such as SZ and bipolar disorder (BP)(2526) we questioned how the molecular signatures associatedwith attenuated autophagic processes could generally be seenas a neuronal trait associated with such psychiatric conditionsAlthough induced pluripotent stem cells-derived neurons are
3168 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
useful for investigating specific disease variant-associated mo-lecular changes this labor-intensive and expensive experimen-tal system is not practically optimal to address questions tosporadic or general cases Instead the use of surrogate tissuesmay be considered (27) Thus we used olfactory neuronal cellsobtained via nasal biopsy (28ndash30) and addressed the questionwith these cells from patients with sporadic cases of SZ and BP
We examined levels of p62 and LC3 proteins in olfactoryneuronal cells biopsied from sporadic cases of SZ and BP whencompared with the age gender and race-matched healthy con-trols (Fig 5A) p62 protein levels were significantly upregulatedin cells derived from patients with SZ and BP (Fig 5BSupplementary Material Fig S4A and B) Furthermore the ratioof LC3-IILC3-I had a trend of upregulation a sign of LC3-II accu-mulation and hence attenuated autophagy (Fig 5CSupplementary Material Fig S4A and B) Note that p62 and Lc3mRNA levels showed no significant differences among groups
(Fig 5D) These results in particular the de-coupling of themRNA and protein levels indicate that attenuated autophagicprocess is a cellular signature in neuronal cells from patientswith sporadic SZ and BP
DiscussionThe present study demonstrates that the Ulk2-mediated processinvolving p62 regulation plays a critical role in higher brainfunctions at least in part through maintenance of theexcitatoryndashinhibitory balance in the PFC The finding also sug-gests that neuronal autophagy regulates not only cellular viabil-ity but also homeostasis of brain functions such as cognitionAlthough the CNV previously reported for Ulk2 likely representsan extremely rare case among SZ the underlying neurobiologi-cal deficit caused by Ulk2 dysfunction (ie excitatoryndashinhibitoryimbalance owing to elevated p62 expression) may provide a
Ulk2 WT
15
WT
mE
PS
C fr
eque
ncy
WT
Ulk2+-
Ulk2+- WT
mE
PS
C a
mpl
itude
Ulk2 +-
+-20pA
100msec
(sec)
WT
mIP
SC
freq
uenc
y
Ulk2+- WT
mIP
SC
am
plitu
de
Ulk2+-
(Hz)(pA)
Pea
k ΔF
F0
0
02
04
03
01
WT Ulk2
10
5
15
10
5
(Hz)
15
10
5
(pA)15
10
5
0000 +-
05
WT Ulk2+-
Nor
mal
ized
FD
R
0
1
15
05
0
02
04
03
01
05
05 1 15 20-05-01
06(au)
NR1GABAARα2
0
05
1
Nor
mal
ized
ratio
(Sur
face
Tot
al)
(au) (au)
0
05
1
A B
C D
GABAARα5
0
05
1
(au)
Rapa Veh Rapa Veh Rapa Veh
Ulk2 WT
+-
Sur
face
Tota
l
Ulk2+-Ulk2+-WT WT
Vehicle Rapa
NR1
NR1
α-Tubulin
p62
50
(kDa)
100
50
100
50
72
GABAARα2
50 GABAARα5
50
GABAARα2
GABAARα5ΔF
F0
Figure 2 Reduced surface expression of GABAA receptors and imbalanced excitatoryinhibitory neurotransmission in Ulk2thorn pyramidal neurons (A) Patch-clamp record-
ing of miniature synaptic currents from WT and Ulk2thorn primary neurons (nfrac1420 each) (16ndash25 DIV) Averages of mEPSC and mIPSC frequencies and amplitudes are plotted in
the graph Plt005 (MannndashWhitney test two-tailed) (B) WT and Ulk2thorn primary cortical neurons were transfected with pCAG-GCaMP6 and [Ca2thorn]i in each transfected neu-
ron was measured following depolarization Representative [Ca2thorn]i changes (DFF0) are plotted and the averaged peak value and the FDR are shown in the graph (nfrac1410 each)
Plt005 (MannndashWhitney test two-tailed) (C D) Surface-biotinylated cell lysates from primary neurons cultured with or without rapamycin (100 nM 30 min) were analyzed
by western blot using anti-GABAA receptor a2 and a5 subunit and NR1 antibodies Experiments were done in triplicate and densitometry analyses were done to evaluate the
surface levels of each receptor by normalizing them against the total levels of respective receptor and plotted in the graph Plt005 (KruskalndashWallis test)
3169|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
general pathophysiological mechanism pertinent to neuropsy-chiatric manifestation including the sporadic cases of SZ
Selective targeting of GABAA receptor by autophagy
Selective downmodulation of GABAA receptors on the surfaceof the pyramidal neuron likely leads to imbalancedexcitatoryndashinhibitory neurotransmission in Ulk2thorn mice Howcould neurons achieve such a selective regulation of a partic-ular neurotransmitter receptor through autophagy A previ-ous study reported selective regulation of GABAA receptor
surface presentation at the neuromuscular junction inCaenorhabditis elegans in which GABAA receptors expressedon the muscle cells are selectively sorted via endocytosisfrom the postsynaptic membrane surface to traffic to theautophagosome upon denervation whereas acetylcholinereceptors in the same cell did not traffic to the autophago-some (31) This study suggests a role of autophagy in selec-tive sorting of GABAA receptors responsible for attenuatedinhibitory neurotransmission At the mechanistic level weshowed that a specific interaction of p62 with GABARAPwhich positively regulates endocytic turnover of GABAA
Sur
face
GA
BA
AR
α1
Ulk2 +-
CaM
KII
Ulk2 p62+- +-
WT 0
1
05
(au)
Nor
mal
ized
exp
ress
ion
(GA
BA
AR
α1C
aMK
II)
WT
Ulk2+-
p62
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
PP
I
0
10
20
30
40
50
60
70WT Ulk2+-
Ulk2 p62+- +-
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
0
10
20
30
40
WT Ulk2+- Ulk2 p62+- +-
Ulk2 p62+- +-
WT Ulk2+-
Ulk2 p62+- +-
WT Ulk2+- Ulk2 p62+- +-
p
Figure 3 Reducing p62 gene dosage rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Primary cortical neurons were prepared
from mice with the indicated genotypes and the cell surface staining was done at 16 DIV with anti-GABAA receptor a1 subunit antibody followed by permeabilization
and further staining with anti-CaMKII and p62 antibodies Scale bar 20 lm Fluorescence intensities per soma were measured using ImageJ Plt 005 (KruskalndashWallis
test) Lower magnification images are shown in Supplementary Material Fig S3A (B) The amplitude of startle response (left) and PPI (right) were evaluated for WT
(nfrac149) Ulk2thorn (nfrac149) and Ulk2thorn p62thorn (nfrac1412) Statistical significance in startle response [F(2 28)frac145548 Pfrac1400093] (one-way ANOVA) and in PPI [F(2 28)frac148761
Pfrac1400011] (two-way ANOVA with repeated measures) P lt005 P lt001 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase
as well as the rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thorn p62thorn (nfrac147) No significant difference during the initial association
phase [F(2 20)frac14124 Pfrac140968] Statistical significance during the rule-shift phase F(2 20)frac14842 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (D)
Numbers of perseverative errors during the rule shift phase were scored F(2 20)frac14527 Pfrac140038 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3170 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
receptors (24) may be important elevated p62 expression inUlk2thorn pyramidal neurons appears to sequester a higher pro-portion of GABARAP and limit the amount of GABARAP avail-able for GABAA receptor surface presentation
Linking the data from the animal model and patientneuronal cells
Although both p62 and Ulk2 are ubiquitously expressed in allcell types in every tissue tested (1132) it is currently unknown
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p69-120 p79-120 p83-120
()
P
PI
0
10
20
30
40
50
60
70
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
WT Ulk2+- Ulk2 +-
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
+Cont +Cont +26aa
0
10
20
30
WT Ulk2+- Ulk2 +-
+Cont +Cont +26aa
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
GFP-p62 + + + + 26aa-peptide-FLAG
myc-GABARAP + + + +
50
100(kDa)
IP with myc
(IgG-HC)
WBanti-GFP
anti-GFP
IP
Input
50
anti-myc
26aa-FLAGanti-FLAG
Surface GABAARα1 GFP-26aa-peptide
Structure of Sequestosome-1p62
328-MESGNCSGGDDDWTHLSSKEVDPSTG-353
ZZ UBAPB1
LIR 4423911021
(26aa-peptide)
GABARAP
GFP-p62
GFP-p62
GABARAP
(dot blot)
Figure 4 Interfering with p62ndashGABARAP interaction rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Schematic illustration of
sequestosome-1p62 protein structure and designing strategy for the 26-amino acid peptide that interferes with p62ndashGABARAP interaction The LIR (LC3-interacting re-
gion) contains a DDDWxxL motif that was shown to interact with GABARAP (23) Primary cortical neurons prepared from Ulk2thorn mice were transfected at 16DIV with
an expression plasmid encoding GFP-26aa-peptide and the cell surface GABAA receptor (a1 subunit) was immuno-stained without cell permeabilization White arrow
indicates a cell transfected with GFP-26aa-peptide and open arrows indicate neighboring non-transfected cells Scale bar 20 lm (B) The amplitude of startle response
(left) and PPI (right) were evaluated for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (nfrac147) and Ulk2thorn with 26aa-peptide AAV (nfrac146) No significant
difference in startle response [F(2 17)frac141045 Pfrac140833] (one-way ANOVA) Statistical significance for PPI F(2 17)frac14853 P frac1400126 (two-way ANOVA with repeated meas-
ures) P lt005 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase as well as the rule shift phase of the assays were scored
for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (n frac147) and Ulk2thorn p62thorn with 26aa-peptide AAV (nfrac146) P lt005 P lt001 No significant difference
during the initial association phase [F(2 17)frac14102 P frac140827] Statistical significance during the rule-shift phase F(2 17)frac14774 Pfrac140018 (one-way ANOVA) P lt005
P lt001 (Bonferroni post hoc test) (D) Numbers of perseverative errors during the rule shift phase were scored F(2 17)frac14458 P 50046 (one-way ANOVA) P lt005Pfrac140057 (Bonferroni post hoc test)
3171|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

CaMKII-specific Ulk2thornmice show behavioral deficitsassociated with the PFC function
We next investigated specific roles of neuronal subtypes in theobserved phenotypes Mice heterozygous for conditional Ulk2knockout allele were crossed with mice expressing eitherpan-specific Cre (EIIa-Cre) pyramidal neuron-specific Cre(CaMKII-Cre) or interneuron-specific Cre (Viaat-Cre) and PPIwas measured When Ulk2 was ablated in an interneuron-specific manner PPI was not affected (Supplementary MaterialFig S1CndashE) In contrast pyramidal neuron-specific Ulk2 ablationresulted in attenuated PPI (Supplementary Material Fig S1C Dand F) These data are in line with the notion that homeostaticdeficits involving p62 accumulation in the CaMKII-positive py-ramidal neurons underlie the PPI deficits in Ulk2thornmice
Reduced surface expression of GABAA receptors andimbalanced excitatoryndashinhibitory neurotransmission inUlk2thorn pyramidal neurons
To further investigate molecular and cellular basis that may un-derlie these behavioral changes electrophysiological propertiesof the cultured pyramidal neurons prepared from Ulk2thorn micewere examined Typically 80 of neurons in culture wereCaMKII-positive pyramidal neurons and 20 were glutamicacid decarboxylase 67 (GAD67)-positive interneurons under theculture condition as confirmed by immunostaining performedafter electrophysiological recording Patch-clamp recording ofUlk2thorn pyramidal neurons revealed altered excitatoryndashinhibitory neurotransmission the frequencies and amplitudesof miniature excitatory postsynaptic currents (mEPSC) were ele-vated whereas the miniature inhibitory postsynaptic currents(mIPSC) amplitudes were downregulated suggesting anexcitatory-dominant state at the resting condition (Fig 2A)Next depolarization-induced increases in calcium [Ca2thorn]i weremeasured in CaMKII-positive pyramidal neurons by transfectingGCaMP Ca2thorn sensor probe in culture (Supplementary MaterialFig S2) Ulk2thorn primary neurons showed higher levels of peakDFF0 and slower rates of fluorescence decay than WT neurons(Fig 2B) These data indicate higher excitability and lower re-sponsiveness to inhibitory signals in Ulk2thorn pyramidal neurons
To examine molecular underpinnings for the observed elec-trophysiological changes in Ulk2thorn neurons levels of surfaceexpression of the major excitatory and inhibitory neurotrans-mitter receptors [ie N-methyl-D-aspartate (NMDA) andgamma-aminobutyric acid (GABA)A receptors] were evaluatedin cultured cortical neurons via biotinylation experiments Bothtotal and surface expression levels of the NMDA receptors wereequivalent between WT and Ulk2thorn neurons (Fig 2C and D) Incontrast levels of surface expression of GABAA receptors (a2and a5 subunits) in Ulk2thorn neurons were lower than in WTwhile the total levels of these receptors remained unchangedbetween the two genotypes (Fig 2C and D)
Reducing the level of p62 expression restores GABAA
receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To address the role of p62 in the regulation of GABAA receptorsurface expression Ulk2thorn mice were bred with p62thorn miceand the levels of GABAA receptor surface expression in primarycortical neurons of Ulk2thorn p62thorn mice were evaluated whencompared with those in control or Ulk2thorn mice Reducing p62gene dosage in Ulk2thorn neurons led to significant up-regulation
of the GABAA receptor surface expression (Fig 3ASupplementary Material Fig S3A and B)
We next evaluated whether reducing p62 gene dosage canrescue behavioral deficits in Ulk2thorn mice PPI deficits were nor-malized to the levels equivalent to those in WT (Fig 3B) cognitiveinflexibility in the rule shifting assay was rescued (Fig 3C and D)Thus p62 not only serves as a marker for attenuated autophagyin neurons but also plays a functional role in the regulation ofGABAA receptor surface presentation as well as behaviors
Pharmacological intervention with rapamycin or its analogCCI-779 is known to enhance autophagic processes anddegrades p62 proteins (2122) Indeed we observed ameliorationof the pathologically elevated p62 levels in Ulk2thorn mice withchronic treatment with CCI-779 (Fig 1B) Importantly the sur-face expression of GABAA receptors in Ulk2thorn cultures was nor-malized by rapamycin treatment (Fig 2C and D) FurthermoreCCI-779 treatment normalized attenuated PPI (Fig 1C and D)and cognitive inflexibility (Fig 1F and G) These results supportthe notion that the observed phenotypes in Ulk2thorn mice arelikely caused by attenuation of autophagic processes involvingp62 Notably PPI deficits in Ulk2thorn mice were not normalizedby clozapine (Fig 1C and D)
Interfering with p62ndashGABARAP interaction restoresGABAA receptor surface presentation and rescues thebehavioral deficits in Ulk2thornmice
To further address the molecular mechanism by which the p62dosage affects GABAA receptor surface presentation we hypoth-esized that elevated p62 levels could titrate out proteins essen-tial for GABAA receptor trafficking p62 interacts with GABAA
receptor-associated protein (GABARAP) (23) a scaffolding pro-tein known to regulate GABAA receptor trafficking (24)Therefore we reasoned that interfering with p62ndashGABARAP in-teraction could liberate GABARAP leading to restoration ofGABAA receptor surface presentation Because 10-amino acidLIR (LC3-interacting region) motif located in the middle of thep62 protein was shown to bind GABARAP (23) we designed a26-amino acid peptide (26aa-peptide) that encompasses this re-gion and tested its potential to interfere with p62ndashGABARAP in-teraction (Fig 4A) When heterologously expressed in HEK293cells this 26aa-peptide inhibited p62ndashGABARAP interaction in adose-dependent manner (Fig 4A) Moreover when the 26aa-peptide was transfected in Ulk2thorn primary cortical neurons thelevel of cell surface GABAA receptor was upregulated whencompared with the neighboring non-transfected cells (Fig 4A)We next investigated its effects on behavioral deficits in Ulk2thorn
mice introducing the 26aa-peptide via adeno-associated virus(AAV)-mediated delivery into the medial PFC significantly im-proved PPI deficits (Fig 4B) and behavioral inflexibility (Fig 4Cand D) The results provide a proof of concept for the role of ele-vated p62 levels in sequestering GABARAP in Ulk2thornmice
p62 is elevated in olfactory neuronal cells in patientswith sporadic cases of SZ and bipolar disorder
Given that cortical excitatoryndashinhibitory imbalance PPI deficitsand behavioral inflexibility are generally seen in patients withpsychiatric conditions such as SZ and bipolar disorder (BP)(2526) we questioned how the molecular signatures associatedwith attenuated autophagic processes could generally be seenas a neuronal trait associated with such psychiatric conditionsAlthough induced pluripotent stem cells-derived neurons are
3168 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
useful for investigating specific disease variant-associated mo-lecular changes this labor-intensive and expensive experimen-tal system is not practically optimal to address questions tosporadic or general cases Instead the use of surrogate tissuesmay be considered (27) Thus we used olfactory neuronal cellsobtained via nasal biopsy (28ndash30) and addressed the questionwith these cells from patients with sporadic cases of SZ and BP
We examined levels of p62 and LC3 proteins in olfactoryneuronal cells biopsied from sporadic cases of SZ and BP whencompared with the age gender and race-matched healthy con-trols (Fig 5A) p62 protein levels were significantly upregulatedin cells derived from patients with SZ and BP (Fig 5BSupplementary Material Fig S4A and B) Furthermore the ratioof LC3-IILC3-I had a trend of upregulation a sign of LC3-II accu-mulation and hence attenuated autophagy (Fig 5CSupplementary Material Fig S4A and B) Note that p62 and Lc3mRNA levels showed no significant differences among groups
(Fig 5D) These results in particular the de-coupling of themRNA and protein levels indicate that attenuated autophagicprocess is a cellular signature in neuronal cells from patientswith sporadic SZ and BP
DiscussionThe present study demonstrates that the Ulk2-mediated processinvolving p62 regulation plays a critical role in higher brainfunctions at least in part through maintenance of theexcitatoryndashinhibitory balance in the PFC The finding also sug-gests that neuronal autophagy regulates not only cellular viabil-ity but also homeostasis of brain functions such as cognitionAlthough the CNV previously reported for Ulk2 likely representsan extremely rare case among SZ the underlying neurobiologi-cal deficit caused by Ulk2 dysfunction (ie excitatoryndashinhibitoryimbalance owing to elevated p62 expression) may provide a
Ulk2 WT
15
WT
mE
PS
C fr
eque
ncy
WT
Ulk2+-
Ulk2+- WT
mE
PS
C a
mpl
itude
Ulk2 +-
+-20pA
100msec
(sec)
WT
mIP
SC
freq
uenc
y
Ulk2+- WT
mIP
SC
am
plitu
de
Ulk2+-
(Hz)(pA)
Pea
k ΔF
F0
0
02
04
03
01
WT Ulk2
10
5
15
10
5
(Hz)
15
10
5
(pA)15
10
5
0000 +-
05
WT Ulk2+-
Nor
mal
ized
FD
R
0
1
15
05
0
02
04
03
01
05
05 1 15 20-05-01
06(au)
NR1GABAARα2
0
05
1
Nor
mal
ized
ratio
(Sur
face
Tot
al)
(au) (au)
0
05
1
A B
C D
GABAARα5
0
05
1
(au)
Rapa Veh Rapa Veh Rapa Veh
Ulk2 WT
+-
Sur
face
Tota
l
Ulk2+-Ulk2+-WT WT
Vehicle Rapa
NR1
NR1
α-Tubulin
p62
50
(kDa)
100
50
100
50
72
GABAARα2
50 GABAARα5
50
GABAARα2
GABAARα5ΔF
F0
Figure 2 Reduced surface expression of GABAA receptors and imbalanced excitatoryinhibitory neurotransmission in Ulk2thorn pyramidal neurons (A) Patch-clamp record-
ing of miniature synaptic currents from WT and Ulk2thorn primary neurons (nfrac1420 each) (16ndash25 DIV) Averages of mEPSC and mIPSC frequencies and amplitudes are plotted in
the graph Plt005 (MannndashWhitney test two-tailed) (B) WT and Ulk2thorn primary cortical neurons were transfected with pCAG-GCaMP6 and [Ca2thorn]i in each transfected neu-
ron was measured following depolarization Representative [Ca2thorn]i changes (DFF0) are plotted and the averaged peak value and the FDR are shown in the graph (nfrac1410 each)
Plt005 (MannndashWhitney test two-tailed) (C D) Surface-biotinylated cell lysates from primary neurons cultured with or without rapamycin (100 nM 30 min) were analyzed
by western blot using anti-GABAA receptor a2 and a5 subunit and NR1 antibodies Experiments were done in triplicate and densitometry analyses were done to evaluate the
surface levels of each receptor by normalizing them against the total levels of respective receptor and plotted in the graph Plt005 (KruskalndashWallis test)
3169|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
general pathophysiological mechanism pertinent to neuropsy-chiatric manifestation including the sporadic cases of SZ
Selective targeting of GABAA receptor by autophagy
Selective downmodulation of GABAA receptors on the surfaceof the pyramidal neuron likely leads to imbalancedexcitatoryndashinhibitory neurotransmission in Ulk2thorn mice Howcould neurons achieve such a selective regulation of a partic-ular neurotransmitter receptor through autophagy A previ-ous study reported selective regulation of GABAA receptor
surface presentation at the neuromuscular junction inCaenorhabditis elegans in which GABAA receptors expressedon the muscle cells are selectively sorted via endocytosisfrom the postsynaptic membrane surface to traffic to theautophagosome upon denervation whereas acetylcholinereceptors in the same cell did not traffic to the autophago-some (31) This study suggests a role of autophagy in selec-tive sorting of GABAA receptors responsible for attenuatedinhibitory neurotransmission At the mechanistic level weshowed that a specific interaction of p62 with GABARAPwhich positively regulates endocytic turnover of GABAA
Sur
face
GA
BA
AR
α1
Ulk2 +-
CaM
KII
Ulk2 p62+- +-
WT 0
1
05
(au)
Nor
mal
ized
exp
ress
ion
(GA
BA
AR
α1C
aMK
II)
WT
Ulk2+-
p62
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
PP
I
0
10
20
30
40
50
60
70WT Ulk2+-
Ulk2 p62+- +-
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
0
10
20
30
40
WT Ulk2+- Ulk2 p62+- +-
Ulk2 p62+- +-
WT Ulk2+-
Ulk2 p62+- +-
WT Ulk2+- Ulk2 p62+- +-
p
Figure 3 Reducing p62 gene dosage rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Primary cortical neurons were prepared
from mice with the indicated genotypes and the cell surface staining was done at 16 DIV with anti-GABAA receptor a1 subunit antibody followed by permeabilization
and further staining with anti-CaMKII and p62 antibodies Scale bar 20 lm Fluorescence intensities per soma were measured using ImageJ Plt 005 (KruskalndashWallis
test) Lower magnification images are shown in Supplementary Material Fig S3A (B) The amplitude of startle response (left) and PPI (right) were evaluated for WT
(nfrac149) Ulk2thorn (nfrac149) and Ulk2thorn p62thorn (nfrac1412) Statistical significance in startle response [F(2 28)frac145548 Pfrac1400093] (one-way ANOVA) and in PPI [F(2 28)frac148761
Pfrac1400011] (two-way ANOVA with repeated measures) P lt005 P lt001 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase
as well as the rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thorn p62thorn (nfrac147) No significant difference during the initial association
phase [F(2 20)frac14124 Pfrac140968] Statistical significance during the rule-shift phase F(2 20)frac14842 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (D)
Numbers of perseverative errors during the rule shift phase were scored F(2 20)frac14527 Pfrac140038 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3170 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
receptors (24) may be important elevated p62 expression inUlk2thorn pyramidal neurons appears to sequester a higher pro-portion of GABARAP and limit the amount of GABARAP avail-able for GABAA receptor surface presentation
Linking the data from the animal model and patientneuronal cells
Although both p62 and Ulk2 are ubiquitously expressed in allcell types in every tissue tested (1132) it is currently unknown
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p69-120 p79-120 p83-120
()
P
PI
0
10
20
30
40
50
60
70
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
WT Ulk2+- Ulk2 +-
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
+Cont +Cont +26aa
0
10
20
30
WT Ulk2+- Ulk2 +-
+Cont +Cont +26aa
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
GFP-p62 + + + + 26aa-peptide-FLAG
myc-GABARAP + + + +
50
100(kDa)
IP with myc
(IgG-HC)
WBanti-GFP
anti-GFP
IP
Input
50
anti-myc
26aa-FLAGanti-FLAG
Surface GABAARα1 GFP-26aa-peptide
Structure of Sequestosome-1p62
328-MESGNCSGGDDDWTHLSSKEVDPSTG-353
ZZ UBAPB1
LIR 4423911021
(26aa-peptide)
GABARAP
GFP-p62
GFP-p62
GABARAP
(dot blot)
Figure 4 Interfering with p62ndashGABARAP interaction rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Schematic illustration of
sequestosome-1p62 protein structure and designing strategy for the 26-amino acid peptide that interferes with p62ndashGABARAP interaction The LIR (LC3-interacting re-
gion) contains a DDDWxxL motif that was shown to interact with GABARAP (23) Primary cortical neurons prepared from Ulk2thorn mice were transfected at 16DIV with
an expression plasmid encoding GFP-26aa-peptide and the cell surface GABAA receptor (a1 subunit) was immuno-stained without cell permeabilization White arrow
indicates a cell transfected with GFP-26aa-peptide and open arrows indicate neighboring non-transfected cells Scale bar 20 lm (B) The amplitude of startle response
(left) and PPI (right) were evaluated for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (nfrac147) and Ulk2thorn with 26aa-peptide AAV (nfrac146) No significant
difference in startle response [F(2 17)frac141045 Pfrac140833] (one-way ANOVA) Statistical significance for PPI F(2 17)frac14853 P frac1400126 (two-way ANOVA with repeated meas-
ures) P lt005 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase as well as the rule shift phase of the assays were scored
for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (n frac147) and Ulk2thorn p62thorn with 26aa-peptide AAV (nfrac146) P lt005 P lt001 No significant difference
during the initial association phase [F(2 17)frac14102 P frac140827] Statistical significance during the rule-shift phase F(2 17)frac14774 Pfrac140018 (one-way ANOVA) P lt005
P lt001 (Bonferroni post hoc test) (D) Numbers of perseverative errors during the rule shift phase were scored F(2 17)frac14458 P 50046 (one-way ANOVA) P lt005Pfrac140057 (Bonferroni post hoc test)
3171|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

useful for investigating specific disease variant-associated mo-lecular changes this labor-intensive and expensive experimen-tal system is not practically optimal to address questions tosporadic or general cases Instead the use of surrogate tissuesmay be considered (27) Thus we used olfactory neuronal cellsobtained via nasal biopsy (28ndash30) and addressed the questionwith these cells from patients with sporadic cases of SZ and BP
We examined levels of p62 and LC3 proteins in olfactoryneuronal cells biopsied from sporadic cases of SZ and BP whencompared with the age gender and race-matched healthy con-trols (Fig 5A) p62 protein levels were significantly upregulatedin cells derived from patients with SZ and BP (Fig 5BSupplementary Material Fig S4A and B) Furthermore the ratioof LC3-IILC3-I had a trend of upregulation a sign of LC3-II accu-mulation and hence attenuated autophagy (Fig 5CSupplementary Material Fig S4A and B) Note that p62 and Lc3mRNA levels showed no significant differences among groups
(Fig 5D) These results in particular the de-coupling of themRNA and protein levels indicate that attenuated autophagicprocess is a cellular signature in neuronal cells from patientswith sporadic SZ and BP
DiscussionThe present study demonstrates that the Ulk2-mediated processinvolving p62 regulation plays a critical role in higher brainfunctions at least in part through maintenance of theexcitatoryndashinhibitory balance in the PFC The finding also sug-gests that neuronal autophagy regulates not only cellular viabil-ity but also homeostasis of brain functions such as cognitionAlthough the CNV previously reported for Ulk2 likely representsan extremely rare case among SZ the underlying neurobiologi-cal deficit caused by Ulk2 dysfunction (ie excitatoryndashinhibitoryimbalance owing to elevated p62 expression) may provide a
Ulk2 WT
15
WT
mE
PS
C fr
eque
ncy
WT
Ulk2+-
Ulk2+- WT
mE
PS
C a
mpl
itude
Ulk2 +-
+-20pA
100msec
(sec)
WT
mIP
SC
freq
uenc
y
Ulk2+- WT
mIP
SC
am
plitu
de
Ulk2+-
(Hz)(pA)
Pea
k ΔF
F0
0
02
04
03
01
WT Ulk2
10
5
15
10
5
(Hz)
15
10
5
(pA)15
10
5
0000 +-
05
WT Ulk2+-
Nor
mal
ized
FD
R
0
1
15
05
0
02
04
03
01
05
05 1 15 20-05-01
06(au)
NR1GABAARα2
0
05
1
Nor
mal
ized
ratio
(Sur
face
Tot
al)
(au) (au)
0
05
1
A B
C D
GABAARα5
0
05
1
(au)
Rapa Veh Rapa Veh Rapa Veh
Ulk2 WT
+-
Sur
face
Tota
l
Ulk2+-Ulk2+-WT WT
Vehicle Rapa
NR1
NR1
α-Tubulin
p62
50
(kDa)
100
50
100
50
72
GABAARα2
50 GABAARα5
50
GABAARα2
GABAARα5ΔF
F0
Figure 2 Reduced surface expression of GABAA receptors and imbalanced excitatoryinhibitory neurotransmission in Ulk2thorn pyramidal neurons (A) Patch-clamp record-
ing of miniature synaptic currents from WT and Ulk2thorn primary neurons (nfrac1420 each) (16ndash25 DIV) Averages of mEPSC and mIPSC frequencies and amplitudes are plotted in
the graph Plt005 (MannndashWhitney test two-tailed) (B) WT and Ulk2thorn primary cortical neurons were transfected with pCAG-GCaMP6 and [Ca2thorn]i in each transfected neu-
ron was measured following depolarization Representative [Ca2thorn]i changes (DFF0) are plotted and the averaged peak value and the FDR are shown in the graph (nfrac1410 each)
Plt005 (MannndashWhitney test two-tailed) (C D) Surface-biotinylated cell lysates from primary neurons cultured with or without rapamycin (100 nM 30 min) were analyzed
by western blot using anti-GABAA receptor a2 and a5 subunit and NR1 antibodies Experiments were done in triplicate and densitometry analyses were done to evaluate the
surface levels of each receptor by normalizing them against the total levels of respective receptor and plotted in the graph Plt005 (KruskalndashWallis test)
3169|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
general pathophysiological mechanism pertinent to neuropsy-chiatric manifestation including the sporadic cases of SZ
Selective targeting of GABAA receptor by autophagy
Selective downmodulation of GABAA receptors on the surfaceof the pyramidal neuron likely leads to imbalancedexcitatoryndashinhibitory neurotransmission in Ulk2thorn mice Howcould neurons achieve such a selective regulation of a partic-ular neurotransmitter receptor through autophagy A previ-ous study reported selective regulation of GABAA receptor
surface presentation at the neuromuscular junction inCaenorhabditis elegans in which GABAA receptors expressedon the muscle cells are selectively sorted via endocytosisfrom the postsynaptic membrane surface to traffic to theautophagosome upon denervation whereas acetylcholinereceptors in the same cell did not traffic to the autophago-some (31) This study suggests a role of autophagy in selec-tive sorting of GABAA receptors responsible for attenuatedinhibitory neurotransmission At the mechanistic level weshowed that a specific interaction of p62 with GABARAPwhich positively regulates endocytic turnover of GABAA
Sur
face
GA
BA
AR
α1
Ulk2 +-
CaM
KII
Ulk2 p62+- +-
WT 0
1
05
(au)
Nor
mal
ized
exp
ress
ion
(GA
BA
AR
α1C
aMK
II)
WT
Ulk2+-
p62
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
PP
I
0
10
20
30
40
50
60
70WT Ulk2+-
Ulk2 p62+- +-
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
0
10
20
30
40
WT Ulk2+- Ulk2 p62+- +-
Ulk2 p62+- +-
WT Ulk2+-
Ulk2 p62+- +-
WT Ulk2+- Ulk2 p62+- +-
p
Figure 3 Reducing p62 gene dosage rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Primary cortical neurons were prepared
from mice with the indicated genotypes and the cell surface staining was done at 16 DIV with anti-GABAA receptor a1 subunit antibody followed by permeabilization
and further staining with anti-CaMKII and p62 antibodies Scale bar 20 lm Fluorescence intensities per soma were measured using ImageJ Plt 005 (KruskalndashWallis
test) Lower magnification images are shown in Supplementary Material Fig S3A (B) The amplitude of startle response (left) and PPI (right) were evaluated for WT
(nfrac149) Ulk2thorn (nfrac149) and Ulk2thorn p62thorn (nfrac1412) Statistical significance in startle response [F(2 28)frac145548 Pfrac1400093] (one-way ANOVA) and in PPI [F(2 28)frac148761
Pfrac1400011] (two-way ANOVA with repeated measures) P lt005 P lt001 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase
as well as the rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thorn p62thorn (nfrac147) No significant difference during the initial association
phase [F(2 20)frac14124 Pfrac140968] Statistical significance during the rule-shift phase F(2 20)frac14842 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (D)
Numbers of perseverative errors during the rule shift phase were scored F(2 20)frac14527 Pfrac140038 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3170 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
receptors (24) may be important elevated p62 expression inUlk2thorn pyramidal neurons appears to sequester a higher pro-portion of GABARAP and limit the amount of GABARAP avail-able for GABAA receptor surface presentation
Linking the data from the animal model and patientneuronal cells
Although both p62 and Ulk2 are ubiquitously expressed in allcell types in every tissue tested (1132) it is currently unknown
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p69-120 p79-120 p83-120
()
P
PI
0
10
20
30
40
50
60
70
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
WT Ulk2+- Ulk2 +-
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
+Cont +Cont +26aa
0
10
20
30
WT Ulk2+- Ulk2 +-
+Cont +Cont +26aa
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
GFP-p62 + + + + 26aa-peptide-FLAG
myc-GABARAP + + + +
50
100(kDa)
IP with myc
(IgG-HC)
WBanti-GFP
anti-GFP
IP
Input
50
anti-myc
26aa-FLAGanti-FLAG
Surface GABAARα1 GFP-26aa-peptide
Structure of Sequestosome-1p62
328-MESGNCSGGDDDWTHLSSKEVDPSTG-353
ZZ UBAPB1
LIR 4423911021
(26aa-peptide)
GABARAP
GFP-p62
GFP-p62
GABARAP
(dot blot)
Figure 4 Interfering with p62ndashGABARAP interaction rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Schematic illustration of
sequestosome-1p62 protein structure and designing strategy for the 26-amino acid peptide that interferes with p62ndashGABARAP interaction The LIR (LC3-interacting re-
gion) contains a DDDWxxL motif that was shown to interact with GABARAP (23) Primary cortical neurons prepared from Ulk2thorn mice were transfected at 16DIV with
an expression plasmid encoding GFP-26aa-peptide and the cell surface GABAA receptor (a1 subunit) was immuno-stained without cell permeabilization White arrow
indicates a cell transfected with GFP-26aa-peptide and open arrows indicate neighboring non-transfected cells Scale bar 20 lm (B) The amplitude of startle response
(left) and PPI (right) were evaluated for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (nfrac147) and Ulk2thorn with 26aa-peptide AAV (nfrac146) No significant
difference in startle response [F(2 17)frac141045 Pfrac140833] (one-way ANOVA) Statistical significance for PPI F(2 17)frac14853 P frac1400126 (two-way ANOVA with repeated meas-
ures) P lt005 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase as well as the rule shift phase of the assays were scored
for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (n frac147) and Ulk2thorn p62thorn with 26aa-peptide AAV (nfrac146) P lt005 P lt001 No significant difference
during the initial association phase [F(2 17)frac14102 P frac140827] Statistical significance during the rule-shift phase F(2 17)frac14774 Pfrac140018 (one-way ANOVA) P lt005
P lt001 (Bonferroni post hoc test) (D) Numbers of perseverative errors during the rule shift phase were scored F(2 17)frac14458 P 50046 (one-way ANOVA) P lt005Pfrac140057 (Bonferroni post hoc test)
3171|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

general pathophysiological mechanism pertinent to neuropsy-chiatric manifestation including the sporadic cases of SZ
Selective targeting of GABAA receptor by autophagy
Selective downmodulation of GABAA receptors on the surfaceof the pyramidal neuron likely leads to imbalancedexcitatoryndashinhibitory neurotransmission in Ulk2thorn mice Howcould neurons achieve such a selective regulation of a partic-ular neurotransmitter receptor through autophagy A previ-ous study reported selective regulation of GABAA receptor
surface presentation at the neuromuscular junction inCaenorhabditis elegans in which GABAA receptors expressedon the muscle cells are selectively sorted via endocytosisfrom the postsynaptic membrane surface to traffic to theautophagosome upon denervation whereas acetylcholinereceptors in the same cell did not traffic to the autophago-some (31) This study suggests a role of autophagy in selec-tive sorting of GABAA receptors responsible for attenuatedinhibitory neurotransmission At the mechanistic level weshowed that a specific interaction of p62 with GABARAPwhich positively regulates endocytic turnover of GABAA
Sur
face
GA
BA
AR
α1
Ulk2 +-
CaM
KII
Ulk2 p62+- +-
WT 0
1
05
(au)
Nor
mal
ized
exp
ress
ion
(GA
BA
AR
α1C
aMK
II)
WT
Ulk2+-
p62
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p74-120 p78-120 p82-120 p86-120 p90-120
()
PP
I
0
10
20
30
40
50
60
70WT Ulk2+-
Ulk2 p62+- +-
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
40
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
0
10
20
30
40
WT Ulk2+- Ulk2 p62+- +-
Ulk2 p62+- +-
WT Ulk2+-
Ulk2 p62+- +-
WT Ulk2+- Ulk2 p62+- +-
p
Figure 3 Reducing p62 gene dosage rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Primary cortical neurons were prepared
from mice with the indicated genotypes and the cell surface staining was done at 16 DIV with anti-GABAA receptor a1 subunit antibody followed by permeabilization
and further staining with anti-CaMKII and p62 antibodies Scale bar 20 lm Fluorescence intensities per soma were measured using ImageJ Plt 005 (KruskalndashWallis
test) Lower magnification images are shown in Supplementary Material Fig S3A (B) The amplitude of startle response (left) and PPI (right) were evaluated for WT
(nfrac149) Ulk2thorn (nfrac149) and Ulk2thorn p62thorn (nfrac1412) Statistical significance in startle response [F(2 28)frac145548 Pfrac1400093] (one-way ANOVA) and in PPI [F(2 28)frac148761
Pfrac1400011] (two-way ANOVA with repeated measures) P lt005 P lt001 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase
as well as the rule shift phase of the assays were scored for WT (nfrac148) Ulk2thorn (nfrac148) and Ulk2thorn p62thorn (nfrac147) No significant difference during the initial association
phase [F(2 20)frac14124 Pfrac140968] Statistical significance during the rule-shift phase F(2 20)frac14842 Plt00001 (one-way ANOVA) P lt0001 (Bonferroni post hoc test) (D)
Numbers of perseverative errors during the rule shift phase were scored F(2 20)frac14527 Pfrac140038 (one-way ANOVA) P lt005 (Bonferroni post hoc test)
3170 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
receptors (24) may be important elevated p62 expression inUlk2thorn pyramidal neurons appears to sequester a higher pro-portion of GABARAP and limit the amount of GABARAP avail-able for GABAA receptor surface presentation
Linking the data from the animal model and patientneuronal cells
Although both p62 and Ulk2 are ubiquitously expressed in allcell types in every tissue tested (1132) it is currently unknown
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p69-120 p79-120 p83-120
()
P
PI
0
10
20
30
40
50
60
70
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
WT Ulk2+- Ulk2 +-
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
+Cont +Cont +26aa
0
10
20
30
WT Ulk2+- Ulk2 +-
+Cont +Cont +26aa
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
GFP-p62 + + + + 26aa-peptide-FLAG
myc-GABARAP + + + +
50
100(kDa)
IP with myc
(IgG-HC)
WBanti-GFP
anti-GFP
IP
Input
50
anti-myc
26aa-FLAGanti-FLAG
Surface GABAARα1 GFP-26aa-peptide
Structure of Sequestosome-1p62
328-MESGNCSGGDDDWTHLSSKEVDPSTG-353
ZZ UBAPB1
LIR 4423911021
(26aa-peptide)
GABARAP
GFP-p62
GFP-p62
GABARAP
(dot blot)
Figure 4 Interfering with p62ndashGABARAP interaction rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Schematic illustration of
sequestosome-1p62 protein structure and designing strategy for the 26-amino acid peptide that interferes with p62ndashGABARAP interaction The LIR (LC3-interacting re-
gion) contains a DDDWxxL motif that was shown to interact with GABARAP (23) Primary cortical neurons prepared from Ulk2thorn mice were transfected at 16DIV with
an expression plasmid encoding GFP-26aa-peptide and the cell surface GABAA receptor (a1 subunit) was immuno-stained without cell permeabilization White arrow
indicates a cell transfected with GFP-26aa-peptide and open arrows indicate neighboring non-transfected cells Scale bar 20 lm (B) The amplitude of startle response
(left) and PPI (right) were evaluated for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (nfrac147) and Ulk2thorn with 26aa-peptide AAV (nfrac146) No significant
difference in startle response [F(2 17)frac141045 Pfrac140833] (one-way ANOVA) Statistical significance for PPI F(2 17)frac14853 P frac1400126 (two-way ANOVA with repeated meas-
ures) P lt005 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase as well as the rule shift phase of the assays were scored
for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (n frac147) and Ulk2thorn p62thorn with 26aa-peptide AAV (nfrac146) P lt005 P lt001 No significant difference
during the initial association phase [F(2 17)frac14102 P frac140827] Statistical significance during the rule-shift phase F(2 17)frac14774 Pfrac140018 (one-way ANOVA) P lt005
P lt001 (Bonferroni post hoc test) (D) Numbers of perseverative errors during the rule shift phase were scored F(2 17)frac14458 P 50046 (one-way ANOVA) P lt005Pfrac140057 (Bonferroni post hoc test)
3171|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

receptors (24) may be important elevated p62 expression inUlk2thorn pyramidal neurons appears to sequester a higher pro-portion of GABARAP and limit the amount of GABARAP avail-able for GABAA receptor surface presentation
Linking the data from the animal model and patientneuronal cells
Although both p62 and Ulk2 are ubiquitously expressed in allcell types in every tissue tested (1132) it is currently unknown
0
500
(au)
Sta
rtle
resp
onse
120 (startle) p69-120 p79-120 p83-120
()
P
PI
0
10
20
30
40
50
60
70
A
B
C
Tria
ls to
crit
erio
n
0
10
20
30
Initial association Rule shift
0
4
8
12
Num
ber o
f p
erse
vera
tive
erro
rs
D
WT Ulk2+- Ulk2 +-
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
+Cont +Cont +26aa
0
10
20
30
WT Ulk2+- Ulk2 +-
+Cont +Cont +26aa
WT + Cont Ulk2 + Cont+-
Ulk2 + 26aa+-
GFP-p62 + + + + 26aa-peptide-FLAG
myc-GABARAP + + + +
50
100(kDa)
IP with myc
(IgG-HC)
WBanti-GFP
anti-GFP
IP
Input
50
anti-myc
26aa-FLAGanti-FLAG
Surface GABAARα1 GFP-26aa-peptide
Structure of Sequestosome-1p62
328-MESGNCSGGDDDWTHLSSKEVDPSTG-353
ZZ UBAPB1
LIR 4423911021
(26aa-peptide)
GABARAP
GFP-p62
GFP-p62
GABARAP
(dot blot)
Figure 4 Interfering with p62ndashGABARAP interaction rescues GABAA receptor surface presentation and behavioral deficits in Ulk2thorn mice (A) Schematic illustration of
sequestosome-1p62 protein structure and designing strategy for the 26-amino acid peptide that interferes with p62ndashGABARAP interaction The LIR (LC3-interacting re-
gion) contains a DDDWxxL motif that was shown to interact with GABARAP (23) Primary cortical neurons prepared from Ulk2thorn mice were transfected at 16DIV with
an expression plasmid encoding GFP-26aa-peptide and the cell surface GABAA receptor (a1 subunit) was immuno-stained without cell permeabilization White arrow
indicates a cell transfected with GFP-26aa-peptide and open arrows indicate neighboring non-transfected cells Scale bar 20 lm (B) The amplitude of startle response
(left) and PPI (right) were evaluated for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (nfrac147) and Ulk2thorn with 26aa-peptide AAV (nfrac146) No significant
difference in startle response [F(2 17)frac141045 Pfrac140833] (one-way ANOVA) Statistical significance for PPI F(2 17)frac14853 P frac1400126 (two-way ANOVA with repeated meas-
ures) P lt005 (Bonferroni post hoc test) (C) Numbers of trials to criterion during the initial association phase as well as the rule shift phase of the assays were scored
for WT injected with control AAV (nfrac147) Ulk2thorn with control AAV (n frac147) and Ulk2thorn p62thorn with 26aa-peptide AAV (nfrac146) P lt005 P lt001 No significant difference
during the initial association phase [F(2 17)frac14102 P frac140827] Statistical significance during the rule-shift phase F(2 17)frac14774 Pfrac140018 (one-way ANOVA) P lt005
P lt001 (Bonferroni post hoc test) (D) Numbers of perseverative errors during the rule shift phase were scored F(2 17)frac14458 P 50046 (one-way ANOVA) P lt005Pfrac140057 (Bonferroni post hoc test)
3171|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

why p62 protein accumulation is predominantly observed inthe pyramidal neurons of the PFC in Ulk2thorn mice It is possiblethat the levels of autophagic activity necessary to maintain cel-lular homeostasis are differentially regulated via specific physi-ological demands determined by each cell type in a given tissueIndeed Ulk1Ulk2 double knockout mice show severer neuronalloss in the hippocampus than in the cerebellum (11)Nevertheless in a pathological context we observed attenuatedautophagic activity even in olfactory neuronal cells frompatients with sporadic SZ and BP Thus pathological deregula-tion of autophagic processes may occur in a cell type-preferential manner through combination of both intrinsic cel-lular vulnerability and circuit-wide physiological demandsthereby underlying sporadic psychiatric conditionsFurthermore our data from an animal model suggests that
deregulated autophagy may play a causal role in neuropsychiat-ric manifestations rather than representing mere incidental orcompensatory changes
Translational potential of the present study
Although there are means of intervening with a type of clinicalmanifestations (eg positive symptoms) observed in patientswith SZ via the use of antipsychotics the medications may notbe effective to other critical symptoms such as negative symp-toms and cognitive dysfunction (33) Clozapine is known as themedication with superior efficacy to all other antipsychoticsbut some patients show treatment resistance to clozapine evenin positive symptoms Thus new therapeutic strategies differ-ent from clozapine and other antipsychotics are awaited In the
A B
C
Normal Control Schizophrenia Bipolar disorder (NC) (SZ) (BP)
N 22 23 19
Age 3441 plusmn 253 3617 plusmn 257 4216 plusmn 247
Gender (MF) 1804 1707 712
Race 13900 14621 11413 (AACauAsianmix)
P lt 005 Fisherrsquos exact test
NC SZ BP
(au)
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3
LC3-II
Nor
mal
ized
pro
tein
leve
ls
0
1
2
3 (au)
NC SZ BP
LC3-I
Nor
mal
ized
ratio
NC SZ BP0
1
2
3
LC3-IILC3-I
p62(au)
0
1
2
3
4
5
NC SZ BP Nor
mal
ized
pro
tein
leve
ls
(au)
D
Nor
mal
ized
mR
NA
leve
ls
p62
0
05
1
15
NC SZ BP
(au)Lc3
NC SZ BP 0
05
1
15
Nor
mal
ized
mR
NA
leve
ls
(au)
para
Figure 5 Attenuated autophagy in olfactory neuronal cells in sporadic cases of psychiatric disorders (A) Demographics of human subjects in this study P lt005
(Fisherrsquos exact test) (B) The relative protein expression levels of p62 for each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot Plt005
Plt001 (one-way ANOVA) See also Supplementary Material Fig S4A and B (C) The relative protein expression levels of LC3-I LC3-II and their ratio (LC3-IILC3-I) for
each subject of control (circle) SZ (square) and BP (triangle) evaluated by western blot paraPfrac140067175 Pfrac140054516 (D) qPCR analysis of p62 and Lc3 mRNA expression in
control or patients-derived olfactory neuronal cells No statistical significance (one-way ANOVA)
3172 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

present study we observed that sensorimotor gating deficitsand cognitive inflexibility in Ulk2thorn mice could be amelioratedwhen p62 gene dosage was reduced or by reducing the levels ofp62 protein using a rapamycin analogue CCI-779 but not withclozapine We cautiously but optimistically interpret thatp62-associated autophagic signaling cascade may be a potentialtarget for drug development at least for SZ In summary thepresent study provides a novel concept that p62-associatedautophagic signaling cascade may be an important pathophysi-ological mediator for sporadic cases of SZ and BP by affectingthe cortical excitatoryndashinhibitory balance and higher brain func-tions such as sensorimotor gating and cognitive flexibility
Materials and MethodsAnimals
Conditional Ulk2 knockout mice (Ulk2tm1ThsnJ) and transgenicmice that express EIIa-Cre CaMKIIa-CreERT2 or Viaat-Cre wereobtained from The Jackson Laboratory (Bar Harbor USA) Ulk2heterozygous mice were generated as previously described (14)p62 knockout mice were generated as described (32) Mice weremaintained on the C57BL6J genetic background for at least 12generations Eight- to 12-week old male mice were used for be-havioral analysis All behavioral experiments and data collec-tion were performed by the experimenters who were blind toanimal genotypes and identity of treatment conditionsMaintenance of mouse colonies and experiments using micewere in accordance with the NIH Guide for the Care and Use ofLaboratory Animals and approved by the Committee on AnimalResearch at Kyoto University Graduate School of Medicine
Prepulse inhibition
The startle response and PPI were measured using a startle re-flex measurement system (SR-LAB) as described (34) with mi-nor modifications The test session began by placing a malemouse in a plastic cylinder and leaving it undisturbed for30 min The background white noise level in the chamber was70 dB A prepulsendashpulse trial started with a 50-ms null periodfollowed by a 20-ms prepulse white noise (74 78 82 86 or90 dB) After a 100-ms delay the startle stimulus (a 40-ms120 dB white noise) was presented followed by a 290-ms record-ing time The total duration of each trial was 500 ms A test ses-sion consisted of six trial types (pulse-only trial and five typesof prepulsendashpulse trial) Six blocks of the six trial types werepresented in a pseudorandomized order such that each trialtype was presented once within a block The formula100 [(response on acoustic prepulsepulse stimulus trialsstartle response on pulse-only trials) 100)] was used to calcu-late PPI
Rule shift assay
Cognitive flexibility was evaluated in the rule-shifting assay es-sentially as described (1920) In brief mice were habituated tofood odor cues (ie coriander and garlic powder) texture cues(ie fine and coarse digging media) and feeding apparatus priorto testing and then food-deprived a day before the assays Micewere initially trained in a sequence of trials to associate a foodreward with a specific stimulus (ie either an odor or a diggingmedium) A varying combination of stimulus and food rewardwas presented to mice per trial Eight consecutive correctresponses to the food reward were considered reaching
criterion (ie successful establishment of association betweenthe stimulus and the food reward) and the number of trials toreach criterion were scored for each mouse tested Upon ruleshifting (eg from an odor cue to a different texture cue to pre-dict reward) numbers of errors owing to perseveration to an oldrule were scored before reaching new criterion
Drug delivery
CCI-779 was dissolved in phosphate-buffered saline (PBS) (pH76) and injected daily (ip 10 mgkg body weight) for 8ndash14 daysClozapine was dissolved in PBS (pH 52) and injected daily (ip15 mgkg body weight) for 2 weeks Tamoxifen (Sigma) was dis-solved in corn oil (10 mgml) and injected daily (ip 1 mgmouse) for five consecutive days
Quantitative reverse transcription-polymerase chainreaction
Total RNA was extracted from PFC using RNeasy Mini kit(Qiagen) and reverse-transcribed with a ReverTra Ace cDNAsynthesis kit (Toyobo) TaqMan probes were purchased fromApplied Biosystems Inc (ABI) All data were normalized withGapdh as reference
Primary cortical neuron culture
Primary cortical neurons were prepared from E135 frontal cor-tex through papain treatment (05 lgml in Earlersquos balanced saltsolution supplemented with 5 mM ethylenediaminetetraaceticacid (EDTA) and 200 lM L-cysteine) followed by mechanical trit-uration using fire-bore glass pipettes and plated on poly-D-ly-sine-coated glass cover slips or glass-bottom dishes (MatTek)The cultures were recovered in serum-containing media[Neurobasal media supplemented with 10 horse serum 5 fe-tal bovine serum and 2 mM glutamine (Gibco)] for 4 h and main-tained in serum-free media [Neurobasal media supplementedwith B-27 (150 diluted) 2 mM glutamine 50 IUml penicillinand 50 lgml streptomycin] with half of media being replacedwith fresh media every 2ndash3 days The cultures were used for im-munocytochemical analysis surface biotinylation followed bywestern blot analysis patch-clamp recording and Ca2thorn imaginganalysis as described below
Immunohistochemistry and immunocytochemistry
For immunohistochemical analysis brains of perfused mice(nfrac14 4 per group) were serially cut into 50 lm-thick coronal sec-tions using vibratome (VT1200S Leica) and the sections fromone cohort of mice (a littermate pair of wild-type and Ulk2thorn
mice) were permeabilized in PBS containing 005 Triton X-100for 1 h and incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS and simultaneously immunos-tained for 16 h at 4C with primary antibodies followed by AlexaFluorVR 488- or 546-conjugated secondary antibodies (MolecularProbes) for 1 h at room temperature The stained samples wereobserved using a confocal microscope (SP8 Leica 40 objectivelens NA frac1413) images of one optical section (1 lm thick) wereacquired from three to six non-overlapping areas per sectionrandomly chosen in each brain region (PFC visual cortex nu-cleus accumbens dorsolateral striatum hippocampus ventraltegmental area) and three to four serial sections were analyzedThe fluorescence intensities of p62 immunostaining were
3173|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

measured from each neuronal soma (30ndash50 somas per section)using ImageJ (NIH) with the background levels of staining in ad-jacent regions being subtracted and the average immunofluo-rescence intensity was calculated across all serial sections fromevery mouse used
For immunocytochemical analysis cultured primary neu-rons were fixed with 4 PFA in PBS for 15 min and permeabi-lized in PBS containing 005 Triton X-100 for 1 h When thesurface epitope was stained the primary antibody was includedin PBS containing 10 goat serum before permeabilization Thesamples were incubated for 30 min at room temperature in 10goat serum (Chemicon) in PBS followed by primary antibodiesfor 16 h at 4C and by secondary antibodies (Molecular Probes)for 1 h at room temperature
The primary antibodies used were anti-p62 (guinea pig1400 MBL) anti-CaMKIIa [mouse monoclonal (6G9) 1500Stressmarq] anti-GAD67 [mouse monoclonal (1G102) 11000Chemicon] anti-GABAA receptor a1 subunit [rabbit 1500Alomone labs or mouse monoclonal (S95ndash35) 1500 LSBio] andanti-ULK2 (rabbit 150 Thermo)
Surface biotinylation and western blots
Biotinylation of cell surface proteins was performed in primaryneuron cultures using the cell surface protein isolation kit(Pierce) according to the manufacturerrsquos protocol Briefly cellswere incubated with ice-cold PBS containing Sulfo-NHS-SS-Biotin (Pierce) for 30 min with gentle rocking at 4C Cells werethen lysed and precipitated with NeutrAvidin beads Precipitatedproteins were eluted from the NeutrAvidin beads with loadingbuffer containing dithiothreitol and heated for 5 min at 95C andthen analyzed by western blot The primary antibodies usedwere anti-GABAA receptor a2 subunit (rabbit 11000 Abgent)anti-GABAA receptor a5 subunit (rabbit 11000 RampD Systems)anti-NR1 [rabbit monoclonal (11726) 11000 Millipore] anti-p62(guinea pig 11000 MBL) anti-a-tubulin [mouse monoclonal (B-5-1-2) 18000 Sigma] and anti-LC3B (rabbit 11000 Novus)
Patch-clamp recording
Primary cortical neurons prepared as above were plated onpoly-D-lysine-coated glass cover slips Whole-cell clamp record-ings were performed on the cultured neurons at 16ndash25 daysin vitro (DIV) with EPC10 (HEKA) amplifier at room temperatureFire-polished glass patch pipettes of 3ndash6 MX were used and themembrane current was recorded at a holding potential of50 mV mEPSCs or mIPSCs larger than 4 pA were analyzedThe composition of the external saline was (in mM) 145 sodiumchloride (NaCl) 5 potassium hydroxide (KOH) 10 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (HEPES) 2 calciumchloride (CaCl2) 1 magnesium chloride (MgCl2) and 10 glucosepH 73 The composition of the intracellular solution was 140 D-glucuronate 155 KOH 7 potassium chloride (KCl) 5 ethyleneglycol-bis(b-aminoethyl ether)-NNN0N0-tetraacetic acid and 10HEPES pH 73 The junction potential of 14 mV was corrected asin a previous study (35) Recorded neurons were fixed andstained with anti-CaMKIIa (1500 Stressmarq) to confirm theiridentity as pyramidal neurons
Ca2thorn imaging in dissociated culture
Primary cortical neurons prepared from E135 frontal cortex asabove were plated on poly-D-lysine-coated glass-bottom dishes
(MatTek) Cells were transfected at 14ndash18 DIV with pCAG-GCaMP6m (36) using lipofectamine 2000 (Invitrogen) accordingto the standard protocol Two days post-transfection cellswere depolarized by final concentration of KCl (25 mM) plusCaCl2 (30 mM) and imaged with an IX81 fluorescence micro-scope (Olympus) equipped with an objective lens (UPlanSApoNA 135 60 for oil Olympus) a heated stage (37C) anauto-injector for drug delivery and a CoolSNAP CCD camera(HQ2 Photometrics) A series of images were acquired fromsoma and dendrites at 30 framess for 5 s prior to and25 s following depolarization using MetaMorph (MolecularDevices) Relative changes in fluorescence (DFF0) were calcu-lated by normalizing the fluorescence intensity at a given timewith the level of baseline fluorescence measured before theonset of depolarization Peak values for DFF0 were scored andthe fluorescence decay rate (FDR) within 02 s from the peakfluorescence time point was calculated using the followingformula
Normalized FDR frac14frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor Ulk2thorn=
frac12DF=F0ethtfrac14peakTHORN DF=F0ethtfrac14peakthorn 02THORN ethfor WT
Cultured neurons were fixed and stained with anti-CaMKIIa(1500 Stressmarq) to confirm their identity
AAV-mediated gene delivery
Injection of AAV9 encoding a cytomegalovirus promoter-driven green fluorescence protein (GFP)-26aa-peptide (5 1012 gcml) into the medial PFC was performed in 8-week-oldmice Mice were anesthetized by isoflurane inhalation andstereotaxic surgery was performed according to the standardprocedures The coordinate of the injection site was 20 mmanterior and 05 mm lateral to bregma at a depth of 15 mmfor the PFC A total of 05 ll of purified virus was deliveredon each hemisphere over a 3 min period The infected micewere used for behavioral assessment 2ndash3 weeks after surgicaloperations
Clinical subjects and nasal biopsy
Patients were recruited from the outpatient psychiatric clinicsof the Johns Hopkins Medical Institutions Diagnosis was per-formed according to criteria of the Diagnostic and StatisticalManual of Mental Disorders-5th Edition (DSM-5) (AmericanPsychiatric Association) Subjects were excluded from thestudy if they had a history of traumatic brain injury with lossof consciousness for gt1 h a history of drug abuse within 6months of the study drug dependence within 12 months ofthe study or a history of untreated major medical illnessesOlfactory epithelial neuron cultures were prepared as de-scribed (37) The study was approved by the Johns HopkinsInstitutional Review Board and all subjects gave written con-sent for their participation
Statistical analysis
All data were represented as mean 6standard error of the mean(SEM) and were analyzed by KruskalndashWallis test followed byDunnrsquos multiple comparison test unless otherwise notedBehavioral assay data were analyzed by one-way or two-wayanalysis of variance (ANOVA) followed by Bonferroni post hoc
3174 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

test using Prism statistics software (GraphPad) Statistical sig-nificance in figures Plt 005 Plt 001 Plt 0001
Supplementary MaterialSupplementary Material is available at HMG online
Acknowledgements
We thank Shuhei Ueda Makiko Morita Yuichiro Hayashi TomMacpherson Hiroyuki Okuno Yoshiaki Tagawa and YojiNakamura for advice and Ayumi Saka and Sayaka Mauchi fortechnical assistance
Conflict of Interest statement None declared
FundingThis work was supported by grants from the NationalIinstitutes of Health [MH-084018 MH-094268 Silvio O ConteCenter MH-069853 MH-085226 MH-088753 and MH-092443 toASa MH105660 to KI ASa] Stanley S-R and RUSKFoundations (to ASa) NARSAD and Maryland Stem CellResearch Fund (to KI ASa) Department of DefenseCongressionally Directed Medical Research Program [W81XWH-11-1-0269 to TT] Childrends Tumor Foundation-DrugDiscovery Initiative (to TT) Japan Society of Promotion ofScience [49170001 to TY 24689015 to TF 25110717 and15H04259 to THir 15H01285 and 16K01948 to TS 15H0427516H06568 and 16K14579 to THik] Takeda Science Foundationthe Naito Foundation the Kato Memorial Trust for NambyoResearch and the International Cooperative Research Programof Institute for Protein Research at Osaka University [ICRa-17-13] (to THik) and partly supported by Takeda PharmaceuticalCo Ltd
References1 Mizushima N and Komatsu M (2011) Autophagy renova-
tion of cells and tissues Cell 147 728ndash7412 Hara T Nakamura K Matsui M Yamamoto A
Nakahara Y Suzuki-Migishima R Yokoyama MMishima K Saito I Okano H and Mizushima N (2006)Suppression of basal autophagy in neural cells causes neu-rodegenerative disease in mice Nature 441 885ndash889
3 Komatsu M Waguri S Chiba T Murata S Iwata JTanida I Ueno T Koike M Uchiyama Y Kominami Eand Tanaka K (2006) Loss of autophagy in the central ner-vous system causes neurodegeneration in mice Nature 441880ndash884
4 Komatsu M Wang QJ Holstein GR Friedrich VL JrIwata J Kominami E Chait BT Tanaka K and Yue Z(2007) Essential role for autophagy protein Atg7 in themaintenance of axonal homeostasis and the prevention ofaxonal degeneration Proc Natl Acad Sci USA 10414489ndash14494
5 Toda H Mochizuki H Flores III R Josowitz R KrasievaTB LaMorte VJ Suzuki E Gindhart JG Furukubo-Tokunaga K and Tomoda T (2008) UNC-51ATG1 kinaseregulates axonal transport by mediating motor-cargo as-sembly Genes Dev 22 3292ndash3307
6 Wairkar Y Toda H Mochizuki H Furukubo-Tokunaga KTomoda T and DiAntonio A (2009) Unc-51 controls active
zone density and protein composition by downregulatingERK signaling J Neurosci 29 517ndash528
7 Shen W and Ganetzky B (2009) Autophagy promotes syn-apse development in Drosophila J Cell Biol 187 71ndash79
8 Mochizuki H Toda H Ando M Kurusu M Tomoda Tand Furukubo-Tokunaga K (2011) UNC-51ATG1 controlsaxonal and dendritic development via kinesin-mediatedvesicle transport in the Drosophila brain PLoS One 6 e19632
9 Tomoda T Kim J-H Zhan C and Hatten ME (2004) Roleof Unc511 and its binding partners in CNS axon outgrowthGenes Dev 18 541ndash558
10 Hernandez D Torres CA Setlik W Cebrian CMosharov EV Tang G Cheng HC Kholodilov NYarygina O Burke RE Gershon M and Sulzer D (2012)Regulation of presynaptic neurotransmission by macroau-tophagy Neuron 74 277ndash284
11 Joo JH Wang B Frankel E Ge L Xu L Iyengar R Li-Harms X Wright C Shaw TI Lindsten T et al (2016) Thenoncanonical role of ULKATG1 in ER-to-Golgi trafficking isessential for cellular homeostasis Mol Cell 62 491ndash506
12 Sumitomo A Ueta K Mauchi S Hirai K Horike KHikida T Sakurai T Sawa A and Tomoda T (2017) Ulk1protects against ethanol-induced neuronal stress andcognition-related behavioral deficits Neurosci Res 11754ndash61
13 Kundu M Lindsten T Yang CY Wu J Zhao F Zhang JSelak MA Ney PA and Thompson CB (2008) Ulk1 plays acritical role in the autophagic clearance of mitochondria andribosomes during reticulocyte maturation Blood 1121493ndash1502
14 Cheong H Lindsten T Wu J Lu C and Thompson CB(2011) Ammonia-induced autophagy is independent ofULK1ULK2 kinases Proc Natl Acad Sci USA 10811121ndash11126
15 Cheong H Wu J Gonzales LK Guttentag SHThompson CB and Lindsten T (2014) Analysis of a lung de-fect in autophagy-deficient mouse strains Autophagy 1045ndash56
16 Lang B Pu J Hunter I Liu M Martin-Granados C ReillyTJ Gao GD Guan ZL Li WD Shi YY et al (2014)Recurrent deletions of ULK4 in schizophrenia a gene crucialfor neuritogenesis and neuronal motility J Cell Sci 127630ndash640
17 Swerdlow NR and Geyer MA (1998) Using an animalmodel of deficient sensorimotor gating to study the patho-physiology and new treatments of schizophrenia SchizophrBull 24 285ndash301
18 Kellendonk C Simpson EH and Kandel ER (2009)Modeling cognitive endophenotypes of schizophrenia inmice Trends Neurosci 32 347ndash358
19 Bissonette GB Martins GJ Franz TM Harper ESSchoenbaum G and Powell EM (2008) Double dissociationof the effects of medial and orbital prefrontal cortical lesionson attentional and affective shifts in mice J Neurosci 2811124ndash11130
20 Cho KK Hoch R Lee AT Patel T Rubenstein JL andSohal VS (2015) Gamma rhythms link prefrontal interneu-ron dysfunction with cognitive inflexibility in Dlx56thorn
mice Neuron 85 1332ndash134321 Schmelzle T and Hall MN (2000) TOR a central controller
of cell growth Cell 103 253ndash26222 Ravikumar B Vacher C Berger Z Davies JE Luo S
Oroz LG Scaravilli F Easton DF Duden R OrsquoKane CJand Rubinsztein DC (2004) Inhibition of mTOR induces
3175|Human Molecular Genetics 2018 Vol 27 No 1 8
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022
autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022

autophagy and reduces toxicity of polyglutamine expan-sions in fly and mouse models of Huntington disease NatGenet 36 585ndash595
23 Pankiv S Clausen TH Lamark T Brech A Bruun JAOutzen H Oslashvervatn A Bjoslashrkoslashy G and Johansen T (2007)p62SQSTM1 binds directly to Atg8LC3 to facilitate degrada-tion of ubiquitinated protein aggregates by autophagy J BiolChem 282 24131ndash24145
24 Wang H Bedford FK Brandon NJ Moss SJ and OlsenRW (1999) GABA(A)-receptor-associated protein linksGABA(A) receptors and the cytoskeleton Nature 397 69ndash72
25 Perry W Minassian A Feifel D and Braff DL (2001)Sensorimotor gating deficits in bipolar disorder patientswith acute psychotic mania Biol Psychiatry 50 418ndash424
26 Green MF (2006) Cognitive impairment and functional out-come in schizophrenia and bipolar disorder J ClinPsychiatry 67(Suppl) e12ndashe18 discussion 36ndash42
27 Gamo NJ and Sawa A (2014) Human stem cells and surro-gate tissues for basic and translational study of mental dis-orders Biol Psychiatry 75 918ndash919
28 English JA Fan Y Focking M Lopez LM HryniewieckaM Wynne K Dicker P Matigian N Cagney G Mackay-Sim A and Cotter DR (2015) Reduced protein synthesis inschizophrenia patient-derived olfactory cells TranslPsychiatry 5 e663
29 Lavoie J Gasso Astorga P Segal-Gavish H Wu YCChung Y Cascella NG Sawa A and Ishizuka K (2017)The olfactory neural epithelium as a tool in neuroscienceTrends Mol Med 23 100ndash103
30 Lavoie J Sawa A and Ishizuka K (2017) Application ofolfactory tissue and its neural progenitors to schizophreniaand psychiatric research Curr Opin Psychiatry 30 176ndash183
31 Rowland AM Richmond JE Olsen JG Hall DH andBamber BA (2006) Presynaptic terminals independentlyregulate synaptic clustering and autophagy of GABAA recep-tors in Caenorhabditis elegans J Neurosci 26 1711ndash1720
32 Komatsu M Waguri S Koike M Sou YS Ueno T HaraT Mizushima N Iwata J Ezaki J Murata S et al (2007)Homeostatic levels of p62 control cytoplasmic inclusionbody formation in autophagy-deficient mice Cell 1311149ndash1163
33 Nucifora FC Jr Mihaljevic M Lee BJ and Sawa A (2017)Clozapine as a model for antipsychotic developmentNeurotherapeutics 14 750ndash761
34 Takahashi N Sakurai T Bozdagi-Gunal O Dorr NPMoy J Krug L Gama-Sosa M Elder GA Koch RJWalker RH et al (2011) Increased expression of receptorphosphotyrosine phosphatase-bf is associated with molec-ular cellular behavioral and cognitive schizophrenia phe-notypes Transl Psychiatry 1 e8
35 Hirano T and Kasono K (1993) Spatial distribution of excit-atory and inhibitory synapses on a Purkinje cell in a rat cere-bellar culture J Neurophysiol 70 1316ndash1325
36 Chen TW Wardill TJ Sun Y Pulver SR Renninger SLBaohan A Schreiter ER Kerr RA Orger MB JayaramanV et al (2013) Ultrasensitive fluorescent proteins for imagingneuronal activity Nature 499 295ndash300
37 Kano S Colantuoni C Han F Zhou Z Yuan Q WilsonA Takayanagi Y Lee Y Rapoport J Eaton W et al (2013)Genome-wide profiling of multiple histone methylations inolfactory cells further implications for cellular susceptibilityto oxidative stress in schizophrenia Mol Psychiatry 18740ndash742
3176 | Human Molecular Genetics 2018 Vol 27 No 18
Dow
nloaded from httpsacadem
icoupcomhm
garticle271831655034852 by guest on 06 September 2022




















