
jcb0581.pdf - Oxford Academic
14
ARE PLEOPODS JUST ‘‘MORE LEGS’’? THE FUNCTIONAL MORPHOLOGY OF SWIMMING LIMBS IN EURYTHENES GRYLLUS (AMPHIPODA) Michel A. Boudrias Marine & Environmental Studies, University of San Diego, 5998 Alcala Park, San Diego, California 92110-2492 U.S.A. (e-mail: [email protected]) ABSTRACT Amphipods, like most swimming crustaceans, employ a drag-based mechanism to produce thrust. The propulsors are paddle-shaped pleopods that move parallel to the direction of motion. These paired abdominal limbs generate both the propulsive thrust and the respiratory currents that bathe the thoracic gills. This study addresses the basic kinematics of motion and the pleopodal skeletomusculature of the deep-sea scavenger Eurythenes gryllus. The limb beat cycle consists of a power stroke where the three pleopod pairs, with their setal fan outstretched, swing sequentially through an arc parallel to the body axis, and then return anteriorly in a collapsed and bent configuration. The joint connecting the body to the muscular peduncle is complex, allowing promotion and remotion along the main body axis. Several hard plates for extrinsic muscle attachment are surrounded by arthrodial membrane. The extrinsic musculature is proportioned accordingly, with a large mass of muscles controlling the power stroke and a few long muscles generating the recovery stroke forces. The intrinsic musculature within the peduncle and annular rami serves two functions: (1) many long, thin muscle fibers within the peduncle function as support muscles responsible for shape changes; (2) large, strap-like muscles control flexion for the recovery stroke and abduction-adduction of the exopod for the power stroke. Several ancillary skeletal structures enhance swimming efficiency: reinforcement in the pleonal wall for muscle attachment, coupling hooks between the pleopod pair to effectively create a single paddle, the complementary shapes of the exopod and endopod, and an exopodal hook that facilitates the complete collapse of the pleopods on the recovery stroke. The functional design of the pleopod maximizes efficient recovery stroke motion while still providing strong remotion during the recovery stroke. From patches of dead seaweed on beaches, to kelp forests offshore, and down to the great depths of the ocean, amphipods are among the most abundant crustaceans and are significant members of benthic food webs. They are often encountered in large numbers as mating swarms (Lobel and Randall, 1986) or when at- tracted to bait (Hessler et al., 1972, 1978; Thurston, 1979; Baldwin and Smith, 1987; Sainte-Marie and Hargrave, 1987; Steele, 1988). Amphipods break down kelp racks on beaches, attack captured fish (Templeman, 1967; Bowman, 1974; Stepien and Brusca, 1985; Sainte-Marie and Hargrave, 1987), and are themselves important prey items for fish (Bray and Ebeling, 1975; Hobson and Chess, 1976, 1978; Marshall, 1979). The versatility of amphipod limbs allows for a wide range of be- haviors, from tube building to long jumps on sand beaches, to swimming over great dis- tances to find food (Schram, 1986). These numerous behaviors are reflected in the morphological complexity of limb structure and function. General descriptions of their mor- phological characteristics can be found in ma- jor taxonomic texts (for example, Sars, 1895; Bousfield, 1973; Barnard and Karaman, 1991). Amphipods have many different limbs, each with a set of well-defined functions. Antennae and antennules are used primarily as chemore- ceptors in both food and mate detection (Dahl, 1979; Lowry, 1986). Mouthparts, mandibles, maxillae, and maxillipeds function as a unit in tearing, chewing, and crushing food (Dahl, 1979; Sainte-Marie, 1984). Gnathopods, the first two pairs of thoracic limbs, are important in mating, feeding, grooming, and aggressive intra- and interspecific behavior (Holmquist, 1982; Dunham et al., 1986, personal observa- tion). The pereopods, the last five pairs of tho- racic limbs, are mainly for walking (Hessler, 1982) but are also used in amplexus, the pre- copular clasping of females by males, and as rudders when swimming (Vogel, 1985a, perso- nal observation). The uropods, the most poste- rior three pairs of abdominal limbs, operate 581 JOURNAL OF CRUSTACEAN BIOLOGY, 22(3): 581–594, 2002 Downloaded from https://academic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 May 2022
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of jcb0581.pdf - Oxford Academic

ARE PLEOPODS JUST ‘‘MORE LEGS’’? THE FUNCTIONAL
MORPHOLOGY OF SWIMMING LIMBS IN EURYTHENESGRYLLUS (AMPHIPODA)
Michel A. Boudrias
Marine & Environmental Studies, University of San Diego, 5998 Alcala Park, San Diego,
California 92110-2492 U.S.A. (e-mail: [email protected])
A B S T R A C T
Amphipods, like most swimming crustaceans, employ a drag-based mechanism to produce thrust. The
propulsors are paddle-shaped pleopods that move parallel to the direction of motion. These paired
abdominal limbs generate both the propulsive thrust and the respiratory currents that bathe the thoracic
gills. This study addresses the basic kinematics of motion and the pleopodal skeletomusculature of the
deep-sea scavenger Eurythenes gryllus. The limb beat cycle consists of a power stroke where the three
pleopod pairs, with their setal fan outstretched, swing sequentially through an arc parallel to the body axis,
and then return anteriorly in a collapsed and bent configuration. The joint connecting the body to the
muscular peduncle is complex, allowing promotion and remotion along the main body axis. Several hard
plates for extrinsic muscle attachment are surrounded by arthrodial membrane. The extrinsic musculature
is proportioned accordingly, with a large mass of muscles controlling the power stroke and a few long
muscles generating the recovery stroke forces. The intrinsic musculature within the peduncle and annular
rami serves two functions: (1) many long, thin muscle fibers within the peduncle function as support
muscles responsible for shape changes; (2) large, strap-like muscles control flexion for the recovery stroke
and abduction-adduction of the exopod for the power stroke. Several ancillary skeletal structures enhance
swimming efficiency: reinforcement in the pleonal wall for muscle attachment, coupling hooks between
the pleopod pair to effectively create a single paddle, the complementary shapes of the exopod and
endopod, and an exopodal hook that facilitates the complete collapse of the pleopods on the recovery
stroke. The functional design of the pleopod maximizes efficient recovery stroke motion while still
providing strong remotion during the recovery stroke.
From patches of dead seaweed on beaches,to kelp forests offshore, and down to the greatdepths of the ocean, amphipods are among themost abundant crustaceans and are significantmembers of benthic food webs. They are oftenencountered in large numbers as matingswarms (Lobel and Randall, 1986) or when at-tracted to bait (Hessler et al., 1972, 1978;Thurston, 1979; Baldwin and Smith, 1987;Sainte-Marie and Hargrave, 1987; Steele,1988). Amphipods break down kelp racks onbeaches, attack captured fish (Templeman,1967; Bowman, 1974; Stepien and Brusca,1985; Sainte-Marie and Hargrave, 1987), andare themselves important prey items for fish(Bray and Ebeling, 1975; Hobson and Chess,1976, 1978; Marshall, 1979). The versatility ofamphipod limbs allows for a wide range of be-haviors, from tube building to long jumps onsand beaches, to swimming over great dis-tances to find food (Schram, 1986).
These numerous behaviors are reflected inthe morphological complexity of limb structure
and function. General descriptions of their mor-phological characteristics can be found in ma-jor taxonomic texts (for example, Sars, 1895;Bousfield, 1973; Barnard and Karaman, 1991).Amphipods have many different limbs, eachwith a set of well-defined functions. Antennaeand antennules are used primarily as chemore-ceptors in both food and mate detection (Dahl,1979; Lowry, 1986). Mouthparts, mandibles,maxillae, and maxillipeds function as a unit intearing, chewing, and crushing food (Dahl,1979; Sainte-Marie, 1984). Gnathopods, thefirst two pairs of thoracic limbs, are importantin mating, feeding, grooming, and aggressiveintra- and interspecific behavior (Holmquist,1982; Dunham et al., 1986, personal observa-tion). The pereopods, the last five pairs of tho-racic limbs, are mainly for walking (Hessler,1982) but are also used in amplexus, the pre-copular clasping of females by males, and asrudders when swimming (Vogel, 1985a, perso-nal observation). The uropods, the most poste-rior three pairs of abdominal limbs, operate
581
JOURNAL OF CRUSTACEAN BIOLOGY, 22(3): 581–594, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

either as rudders, as springs for jumping upfrom a substrate, or as anchoring appendages(Barnard, 1965; Conlan and Bousfield, 1982).All these appendages display significant taxo-nomic characters, so they have been drawn ex-tensively for most species, though often withoutfunctional interpretations.
In contrast, pleopods, the three anterior pairsof abdominal limbs, have been little studied.Pleopods generate respiratory currents thatbathe the thoracic gills and are the main, and inmost cases the only, swimming appendages.Some studies of pleopod beat frequency, as ameasure of respiratory rates (Waterman, 1961;Yayanos, 1978, 1981; George, 1979a) and afew morphological drawings in some major am-phipod textbooks (Sars, 1895; Bousfield, 1973)supply the bulk of the meager data on pleopods.There are no rigorous examinations of theirfunctional morphology. Only four papers focuson amphipod pleopod musculature and biome-chanics: (1) Stebbing (1888) illustrates the ba-sic intrinsic musculature, but gives no writtendescription or functional analysis; (2) Brusca(1981), studying hyperiids, includes a simpleaccount of extrinsic and intrinsic pleopodalmuscles; and (3) Vogel (1985a, b) presentssome data on the biomechanics and kinematicsof talitrids, with drawings of musculature andscanning electron microscopy of joints. Hisfunctional morphology paper (1985b) focusesmore on abdominal musculature than on themuscles that drive the pleopods.
Even for malacostracans as a whole, there isonly one publication on pleopod musculature(Kils, 1981, on euphausids), even though theseabdominal appendages are the main propulsivelimbs for a variety of pelagic and benthicmalacostracans (Hessler, 1981, 1985; Schram,1986). It is surprising that the biomechanicsand morphology of these appendages are notbetter understood.
My research addresses pleopod functionalmorphology by describing the skeletomuscula-ture and biomechanics. This paper will alsocharacterize the basic limb beat cycle to placethe morphology within the proper kinematiccontext. Although I have investigated pleopodalmusculature in several species of amphipods,I will limit my description of skeletomuscu-lature to Eurythenes gryllus (Lichtenstein,1887) (Fig. 1). Eurythenes gryllus is a deep-sealysianassid scavenger (Ingram and Hessler,1983; Smith and Baldwin, 1982), with a depthdistribution ranging from 100 m or less in polarenvironments to as deep as 5,800 m in the cen-tral oceans (Smith and Baldwin, 1984; Chris-tiansen et al., 1990). The vertical distribution(Smith and Baldwin, 1984), life history (Ingramand Hessler, 1987), respiration rates (George,1979a, b), and behavior (Ingram and Hessler,1983; Hargrave, 1985) of E. gryllus have beenstudied from shallow-water Arctic habitats toabyssal plains in the central Pacific. Descriptionof the morphology of the pleopods and the basickinematic description of the limb beat cycle is
Fig. 1. Lateral view of Eurythenes gryllus (Lichstenstein) female at the end of the pleopod power stroke. Scale bar ¼ 5 mm.
582 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 3, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

the first step in a more comprehensive analysisof amphipod swimming detailing the biome-chanics, kinematics, and fluid dynamics ofswimming based on the protocol described inBoudrias (1991).
MATERIALS AND METHODS
I chose Eurythenes gryllus as the focal species becauseits large size (1.5 cm for juveniles to more than 12 cm foradult females) facilitates dissection and provides clearerinsights into complex skeletal structures. My specimenswere collected at several sampling sites using free-vehicletrapping techniques (Ingram and Hessler, 1983; Smith andBaldwin, 1984; Baldwin and Smith, 1987). Furthermore, Iwas able to collect observational data on swimming be-havior, body positions and limb beat frequency from in situvideo recordings. Several hours of video data of E. gryllusgoing to bait at a study site at 4,100 m in the western Pacificwere analyzed frame by frame to gather appropriatekinematic parameters for an elementary description ofpleopod beat cycles. These kinematic data complementdescribed swimming rates for these deep-sea amphipods(Laver et al., 1985).
In order to better understand the fundamentals of pleopodbeat cycles and to collect more details on limb motionand limb position during a typical pleopod beat, I observedmany other species of swimming amphipods and studiedvideo recordings of Eurythenes obesus Chevreux, 1905, abathypelagic congener of E. gryllus. The data for E. obesuscame from video analysis of an animal collected in the SantaCatalina Basin and maintained in the laboratory. Detailedkinematic analysis will be discussed elsewhere (Boudrias, inpreparation), but the live and videotaped observations ofswimming motions are critical for explaining the morpho-logical patterns described here.
The skeletomusculature was elucidated through a combi-nation of techniques. Working with Formalin-fixed, alcohol-preserved animals, most of the research was accomplishedthrough visual inspection of gross and fine dissections andcamera lucida drawings. The large size of the musclesallowed simple motion tests in which the muscle was movedby pulling it with forceps and the resulting motion of thelimb was noted. Some specimens were fixed in mercuric-chloride (HgCl2) for better muscle fixation, while otherswere macerated in potassium hydroxide (KOH) to removesoft tissue for better viewing of skeletal elements. Clearedspecimens were stained with aniline blue to discriminatehard cuticular structures from softer, more pliable arthrodialmembrane.
RESULTS
Basic Kinematics
The three pairs of pleopods exhibit the sameset of movements. The main motion parallelsthe sagittal plane of the body. Transverse move-ments occur only for the exopod of each pleo-pod and follow the spread and collapse cycleof the power and recovery strokes. At the onsetof the power stroke, all pairs are extended for-ward into the cavity under the body with thelateral walls formed by the thoracic coxalplates and the abdominal pleura. The cycle
begins with the posteriorly directed powerstroke of pleopod III. The force of water flowflares the setae to form a fan normal to theflow (Fig. 2A). Muscular abduction of the exo-pod widens the setal fan by spreading the tworami apart. This power stroke is followedsequentially by the posteriorly directed sweepof pleopod II, then pleopod I. At the end of thepower stroke, the exopods of pleopod III areadducted, the setal fan collapses, and pleopodsIII contact uropod I. This is followed by subse-quent contact of pleopod II on pleopod III andpleopod I on pleopod II. In most instances, allthree pleopod pairs begin their recovery strokesimultaneously. The setal fan is completelyfolded, the pleopods are bent at the peduncle-ramal joint, and the promotion closely followsthe narrow confines of the sagittal plane (Fig.2B). At the end of the recovery stroke, the ramiextend forward again. The cycle then repeatsitself at different beat rates dependent on theswimming speed required.
Skeletomusculature
An amphipod pleopod consists of a heavilymuscularized peduncle and two tapered flagel-lar rami fringed by plumose setae (Fig. 2A).The main skeletal structures will be describedfirst, followed by a detailed description of theextrinsic and intrinsic musculature.
The body-peduncle joint is composed ofsclerotized plates embedded in convoluted andpliable arthrodial membrane (Figs. 3, 4). Theanterior-posterior sweep of the pleopods thatproduces the propulsive thrust is controlled bythree condyles attaching laterally. These threecondyles define two hinge lines. The uppercondyles connect the body wall to the proximaledge of the joint (Figs. 3, 4). The lower lateralcondyle joins the distal end of the complexjoint structure with the proximal edge of thepeduncle (Figs. 3, 4). There is no lower medialcondyle but two sclerotized plates, separatedby arthrodial membrane, might provide someadditional motion (Fig. 3).
The medial hinge is formed by attachment ofthe peduncle directly to a bulge on the bodywall (Figs. 3A, C, 4). The upper lateral condyleconnects the body wall with a separate sclero-tized ring of cuticle (Figs. 3A, B, D, 4). A longcuticular crook leaving the peduncle and hing-ing to the distal end of the separate cuticularring (Fig. 3B, D) forms the lower lateral con-dyle. The upper hinges are responsible for themain sweep of the pleopods; some extension
583BOUDRIAS: PLEOPOD FUNCTIONAL MORPHOLOGY
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

may occur on the medial side due to the con-certina-like cuticle on the body bulge (Fig. 3A).The convoluted arthrodial membrane permitsenormous freedom of motion and provides amatrix for the many sclerotized plates.
In addition to the complex array of stiffenedstructures forming the intricate body-pedunclejoint, a hardened plate is invaginated from theposterior edge of the proximal cuticle into thecenter of the joint (Fig. 4A). This large, thickplate functions as a major location for extrinsicmuscle insertion. Smaller sclerotized structuresattached to the top of the peduncle serve as in-sertion sites for other muscles (Fig. 4B).
Similar planes of motion occur at the pedun-cle-ramal joint, but the extent of movement isreduced and consequently joint construction issimplified. The main axis of motion is definedby a single hinge line on the anterior side andinfolded arthrodial membrane around much ofthe remaining portion of the joint (Fig. 5). Thehinge line for the endopod confers anterolat-eral-posteromedial motion to this ramus (Fig.5A, B). The hinge for the exopod is formed byan evagination of hardened cuticle at the ante-
rior edge of the exopod combined with an in-vagination of soft cuticle at the anterodistaledge of the peduncle (Fig. 5C). The only defi-nite condyle at this joint is lateroposterior with-in the exopod (Fig. 5). It permits theabduction-adduction of the exopod. Thesehinges control the anterior-posterior flexion ofthe rami. With the arthrodial membrane on theposterior side of the joint, flexion is posteriad,diametrical to the body-peduncle joint. This ar-rangement permits an almost complete foldingof the pleopods on the recovery stroke.
The exopod and endopod are both designedas a series of tapering, tubular annuli bilaterallyfringed with setae (Fig. 6A, B). There is mini-mal arthrodial membrane anteriorly between theannuli. The rami can thus curl posteriad duringthe flexed recovery stroke and remain straightand rigid during the power stroke. Each annulushas a pair of setae angled outward and slightlyposteriorly, attached to the distal edge. The se-tae are themselves bilaterally fringed withfine setules. The interaction of adjacent plu-mose setae creates an intricate mesh network(Fig. 6C).
Fig. 2. Pleopods in power and recovery strokes. (A) anterior view showing overlap of setae on power stroke and extent ofpropulsive setal fan; (B) lateral view of pleopod on recovery stroke showing collapsed setal fan, flexed rami, and contractedpeduncle. Scale bar ¼ 5 mm.
584 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 3, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

These skeletal structures define the limits ofmotion and insertions for the musculature. Themuscles are functionally divided in two majorgroups: (1) extrinsic ones that generate thepower needed for promotion and remotion, and(2) intrinsic ones that provide structural supportand flexural control.
Extrinsic Muscles
There are two functional groups of musclesoriginating on the pleonal wall and inserting atthe complex body-peduncle joint. These mus-cles control the major movements of the limb.Five muscle bundles (M1–5), arranged in twolayers, originate on the anterior edge of the
Fig. 3. Body-peduncle joint with pleopod in power stroke position. (A) anterior view; (B) posterior view; (C) medial viewshowing the upper medial condyle; (D) lateral view showing both upper and lower condyles. Shaded structures ¼ arthrodialmembrane. * ¼ condyles. Scale bar ¼ 5 mm.
585BOUDRIAS: PLEOPOD FUNCTIONAL MORPHOLOGY
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

pleonal wall and insert medially on the anteriorface at the top of the peduncle (Fig. 7A, B).These muscles are the main recovery strokepromotors. One large fiber (M6) (Fig. 7A) orig-inates on the fascia of the medial layer ofmuscles and inserts on a small cuticular evagi-nation on the medial side of the body-pedunclejoint close to the promotor bundles (see Fig. 3).Its function is more difficult to clarify, but itsinsertion suggests a part in the promotion ofthe pleopods.
The major muscle mass in the pleopods ismultilayered but still retains a single function.The bulk of the musculature is a thick mass ofmultilayered muscle fibers (M7) that originateslaterally on the pleonal wall (Fig. 7) and insertson the large cuticular plate (see Fig. 4). Themost medial muscle layer, with its wide originhigh on the pleonal wall, inserts medially onthe plate while the deeper layers insert progres-sively more laterally. This large muscle is the
power stroke remotor. Because the powerstroke produces the thrust necessary for swim-ming and must pull a large resistive surface oneach stroke, it is not surprising that this muscleforms the bulk of the extrinsic pleopodal mus-cles. The most lateral layer of extrinsic musclefibers (M8) (Fig. 7C) originates low on thepleonal wall and inserts on the lateral edge ofthe large cuticular plate. It also acts as a remo-tor during the power stroke. Small muscles(M9) connecting the pleonal wall to portions of
Fig. 4. Plan view of body-peduncle joint, looking in frombody into peduncle. (A) large cuticular plate covering mostof the joint opening with clear view of upper condyles; (B)large cuticular plate removed showing underlying cuticularrings and lower lateral condyle. Muscle insertion siteshighlighted. * ¼ condyles. Shaded structures ¼ arthrodialmembrane. Scale bar ¼ 5 mm.
Fig. 5. Plan view of peduncle-ramal joint with rami ex-tended. (A) view from peduncle into rami showing lateralcondyle, apodemes for muscle insertion, and hinge line forflexion; (B) view from the rami into the peduncle to showthe lateral condyle and adductor apodeme more clearly; (C)lateral view showing main fulcrum for exopodal abduction-adduction; note the complementary evagination-invagina-tion at the anterior edge which forms the main part of the hingeline. * ¼ condyle. Shaded structures ¼ arthrodial mem-brane. Scale bar ¼ 5 mm.
586 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 3, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

the complex body-peduncle joint probably of-fer additional structural support (Fig. 7).
Intrinsic Muscles
Peduncle Structural Support.—The peduncleis an elongated rectangular structure with angledproximal and distal ends. It is a cuticular sackfilled by muscle fibers (Fig. 8A–D). Themajority of these fibers are small and whollycontained within the peduncle. They originateon small apodemes at the proximal posterior
corners of the peduncle (Fig. 8A–C) and fan outalong its length. Some fibers insert at inter-mediate positions along the anterior face of thepeduncle while others insert at the distal edge ofthe peduncle along both sides (Fig. 8A–C).These muscle fibers do not cross the peduncle-ramal joint (Fig. 8D). They most likely functionas support muscles controlling the shape of thepeduncle on the power and recovery strokes.The proximal end of the peduncle containshardened cuticular plates for muscle origins andsofter cuticle that is designed to flex when thesestructural muscles contract. This is an unusualconstruction, which permits changes in shape ofthe peduncle, by buckling the walls, withoutaffecting the flexion and rocking at either thebody-peduncle joint or the peduncle-ramal joint.
Ramal Motion Control.—The flagellar ramican flex posteriad but most of the flexion andextension seems to be passive. Muscular flexionhappens only at the endopod. A wedge-shapedmass of muscle fibers (M10) originates midwayalong the anterior face of the peduncle andinserts in the endopod on a small, hardenedcuticular pad on the posterior surface (Fig. 9).There is no definite extensor within thepeduncle. The flexion of the endopod, inconjunction with the exopodal clasping hook,seems to hold both rami together on therecovery stroke. The second major axis ofmotion at the peduncle-ramal joint is theabduction-adduction of the exopod. Threemuscles are responsible for the flaring andclosing of the exopod (Fig. 9). A large muscle(M11) originates on the anterior face of thepeduncle, away from the body-peduncle joint
Fig. 7. Medial view of extrinsic musculature. Multiple views show layering of muscle fibers. Insertion of promotors notvisible. Scale bar ¼ 5 mm.
Fig. 6. Ramal structure of endopod. (A) medial viewshowing lateroposterior attachment of setae; (B) anteriorview; (C) close-up of plumose setae showing overlap ofsetules. Scale bar ¼ 5 mm.
587BOUDRIAS: PLEOPOD FUNCTIONAL MORPHOLOGY
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

Fig. 8. Intrinsic peduncle support muscles with peduncle extended in power stroke position. (A) lateral view; (B) medialview; (C) anterior view showing both surficial and deep muscle fibers; (D) dorsal view. Shaded structures ¼ arthrodialmembrane. Scale bar ¼ 5 mm.
588 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 3, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

and inserts on the anterolateral edge of theexopod (Fig. 9 and Fig. 5C). A small, wedge-shaped muscle (M12) extends diagonally acrossthe distal portion of the peduncle from themedial wall to a small apodeme on the lateralside of the exopod (Fig. 9 and Fig. 5C). TheM11 and M12 muscles act in concert asabductors for the power stroke. This active,muscular component of exopod abduction isamplified by the external force of the fluid. Theadduction of the exopod is controlled by a set ofthree long muscle fibers (M13), which originateon a pad of reinforced cuticle on the anteriorside of the peduncle and insert along a largeapodeme centrally located in the exopod (Fig. 9and Fig. 5). The three large adductors areattached to the apodeme along its entire length.The apodeme has a very wide base within theexopod, spanning almost the whole width ofthe limb, which provides a strong fulcrum foradduction (Fig. 5C).
Within both the exopod and endopod, twomuscle fibers run the whole length of the fla-gellar rami, originating proximally within each
ramus on small cuticular pads and insertingalong its length to the tip (Fig. 9B). One origi-nates close to the insertion of the flexor in theendopod; the other originates near the insertionof the adductor in the exopod. They probablyact in concert with the endopodal flexor duringthe promotion of the limb. They may also stiff-en the rami during the power stroke.
Ancillary Skeletal Structures
In addition to the skeletal elements responsi-ble for pleopod kinematics, Eurythenes grylluspleopods exhibit many structural refinementsthat improve limb function. The most obviousof these ancillary structures is the reinforce-ment of the pleonal wall where the main remo-tor muscle originates. Smaller ridges within thepleon wall also reinforce the sites of origin forthe promotors. The pattern of cuticular but-tressing reflects the multiple muscle layers (seeFig. 7). It is found only in the three pleonalsegments. The reinforcement has an outercuticle component where the muscles originate;it is a crystal-like structure that creates a
Fig. 9. Intrinsic ramal control muscles. (A) anterior view of muscle origins and insertions, with surficial support musclesremoved; (B) details of insertion of main intrinsic peduncle muscles and origin of ramal flexors. Shaded structures ¼arthrodial membrane. Scale bar ¼ 5 mm.
589BOUDRIAS: PLEOPOD FUNCTIONAL MORPHOLOGY
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022
fingerprint pattern on the outside of the cuticle.Beneath this layer of crystal-like elements andwithin the cuticle of the pleonal wall, deep fur-rows correspond to the origin of the musclelayers.
Two other ancillary structures related to theproficiency of the power stroke are: (1) the bi-laterally fringed rami with plumose setae, and(2) the small hooks on the distomedial surfaceof each peduncle (Fig. 10). The fringed ramioverlap to such an extent they behave function-ally as a full paddle (see Fig. 2A). The hooksclasp with those of the opposite limb, enablingthe pleopod pair to function as a single unitwith maximal resistive area (Fig. 10). The jux-taposition of the two peduncles and the addi-tional overlap of the two endopods affect thedrag of the whole limb (Boudrias, 1992).
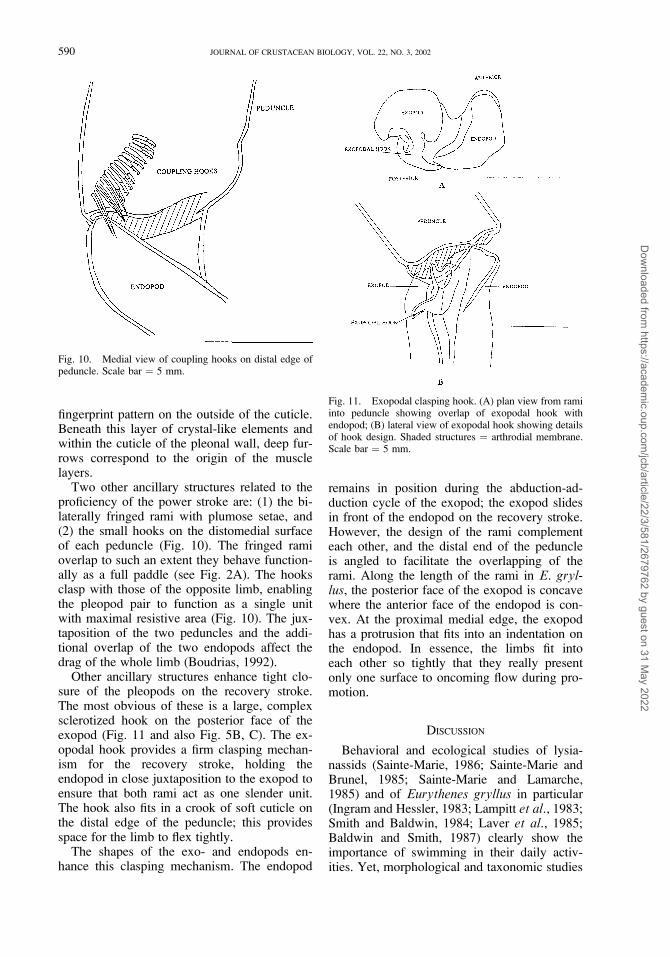
Other ancillary structures enhance tight clo-sure of the pleopods on the recovery stroke.The most obvious of these is a large, complexsclerotized hook on the posterior face of theexopod (Fig. 11 and also Fig. 5B, C). The ex-opodal hook provides a firm clasping mechan-ism for the recovery stroke, holding theendopod in close juxtaposition to the exopod toensure that both rami act as one slender unit.The hook also fits in a crook of soft cuticle onthe distal edge of the peduncle; this providesspace for the limb to flex tightly.
The shapes of the exo- and endopods en-hance this clasping mechanism. The endopod
remains in position during the abduction-ad-duction cycle of the exopod; the exopod slidesin front of the endopod on the recovery stroke.However, the design of the rami complementeach other, and the distal end of the peduncleis angled to facilitate the overlapping of therami. Along the length of the rami in E. gryl-lus, the posterior face of the exopod is concavewhere the anterior face of the endopod is con-vex. At the proximal medial edge, the exopodhas a protrusion that fits into an indentation onthe endopod. In essence, the limbs fit intoeach other so tightly that they really presentonly one surface to oncoming flow during pro-motion.
DISCUSSION
Behavioral and ecological studies of lysia-nassids (Sainte-Marie, 1986; Sainte-Marie andBrunel, 1985; Sainte-Marie and Lamarche,1985) and of Eurythenes gryllus in particular(Ingram and Hessler, 1983; Lampitt et al., 1983;Smith and Baldwin, 1984; Laver et al., 1985;Baldwin and Smith, 1987) clearly show theimportance of swimming in their daily activ-ities. Yet, morphological and taxonomic studies
Fig. 10. Medial view of coupling hooks on distal edge ofpeduncle. Scale bar ¼ 5 mm.
Fig. 11. Exopodal clasping hook. (A) plan view from ramiinto peduncle showing overlap of exopodal hook withendopod; (B) lateral view of exopodal hook showing detailsof hook design. Shaded structures ¼ arthrodial membrane.Scale bar ¼ 5 mm.
590 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 3, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

have emphasized the anterior half of the body.The role of pleopods in swimming and respira-tion should warrant more than passing referenceand simple descriptions.
Previously published papers on pleopodmorphology only describe basic kinematics orpresent simple musculature drawings. Stebbing(1888) presents a few drawings of intrinsicmuscles in the peduncle of Orchomene abys-sorum Stebbing, 1888. He draws the internallongitudinal muscles (M10, M11), the wedge-shaped diagonal spreader (M12), and mentionsthe muscular peduncles of a few other lysianas-sids. Sars (1895) shows a few drawings ofpleopods with sketchy details of pleopodalmusculature that resemble the basic arrange-ment described here. Bousfield (1973) depictsmany pleopods and adequately describes theessential swing motion of these appendages. Inthe few papers that particularly emphasize themusculature of pleopods (Brusca, 1981; Vogel,1985a, b), simplified drawings and succinct de-scriptions of muscle layout explain the remo-tion-promotion cycle reasonably well. Yet noneof these works define the functional arrange-ment of joints and muscles, and few examinethe ancillary skeletal structures or the interac-tion of fluid dynamics and limb construction.
My descriptions of the pleopods in Eur-ythenes gryllus add many details to the basicarrangement of the skeletomusculature of am-phipod pleopods. The standard plan for E. gry-llus consists of: (1) two sets of extrinsicmuscles for promotion and remotion; (2) an in-trinsic flexor important for the recovery stroke;(3) intrinsic support muscles; (4) intrinsic mus-cles responsible for abduction and adduction ofthe exopod; and (5) a long flexor running thelength of each ramus. Some intrinsic musclesaffect the shape of the peduncle, while others,like the long M11, the small fan-shaped M12,and the large M13, function by changing thewidth of the setal fan. The rami only need aflexor to curl the ramus on the recovery strokebecause the passive force of water unfurls therami during the power stroke.
Although the design of the pleopdal skeleto-musculature for Eurythenes gryllus generallyfollows the amphipod plan, these wide-rangingscavengers appear to have a more complicatedfunctional design than other amphipods. Forexample, the multiple layers of muscles form-ing a large proportion of pleonal tissue areespecially remarkable. The multiple layers eachhave their own originating line, though they
combine to function as one muscle mass. Thiscontrasts markedly with other amphipods thattypically have a single layer of remotor-promo-tor muscle bundles (Brusca, 1981; Vogel,1985b). The cuticular reinforcement of thepleonal wall in E. gryllus at the position ofmuscle origin may have become necessary be-cause of the large forces applied there by themassive muscles.
The basic arrangement of extrinsic musclesfunctions within an intricate body-pedunclejoint. This joint in E. gryllus is designed withextensive convolutions of arthrodial membrane,an unusual arrangement of condyles, and theexistence of a large cuticular plate. The divi-sion of this joint into separate sclerotized partsadds complexity to a joint that seems to have arather simple function, i.e., determining the ex-tent of swing of the pleopods. The design ofthis elaborate joint may refine the movementof the pleopods by allowing more stretching ofthe joint. This may facilitate the long sweep an-gles that extend from the thoracic cavity to thetail cone produced by the uropods and the tel-son. The double hinge line only provides ante-rior-posterior motion. The convolutions ofarthrodial membrane and the many plates inter-act to produce an effective power and recoverystroke cycle. The large cuticular plate whichserves as a strong anchor for muscle insertionadds more complexity to this joint. It is unclearwhy this complex design is needed, becausethe kinematics are rather simple. The designcomplexity of E. gryllus may reflect its activeswimming and scavenging lifestyle.
Furthermore, the intrinsic musculature ofE. gryllus is unusual because the bulk of thefibers are contained within the peduncle with-out crossing the proximal or distal joints. Thisis the first evidence of a series of muscleswholly contained in a limb segment. This pat-tern of muscle arrangement may be more prev-alent in swimming limbs of other crustaceansbut has yet to be explored. The unusual patternof muscle fibers seems to function by not bend-ing the peduncle but rather changing its shape.The location and layout of the fibers suggestthe potential for altering the elongated, flatanterior face to a shorter, more convex, andslightly pointed facade. The fibers along theposterior face can shorten the rectangular pe-duncle in conjunction with the buckling of theanterior face and the shortening of the sides.This would reduce the surface area of the pe-duncle on the recovery stroke, an important
591BOUDRIAS: PLEOPOD FUNCTIONAL MORPHOLOGY
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

gain in the optimization of net thrust (Boudrias,1992). This unique design may be crucial tocrustaceans that can not rotate the peduncle oftheir swimming appendages. Changing theshape to reduce the surface area may be a novelway to reduce drag on the recovery stroke.
The other intrinsic muscles have two majorfunctions: (1) abduction-adduction to controlthe width of the setal fan, and (2) flexion of theexopod and rami to further increase the effi-ciency of the recovery stroke. Muscular flexionoccurs only in the endopod, controlled by ashort, triangular-shaped muscle (M9). The skel-etal structure of the peduncle-ramal joint is suchthat the endopod proximal edge is always ante-rior in position to the exopodal hook (Figs. 5,11). By flexing the endopod within its coupledarrangement, the exopod will also be bent pos-teriad. This muscular flexion combines with animportant passive component determined byfluid forces produced by the rapid promotion ofthe recovery stroke. The extension of the ramiis almost entirely passive during the strong re-motion of the limbs. The remotion controlledextrinsically and the fluid forces straighten therami and spread the setae and setules.
The abductors and adductors of the exopodact antagonistically. The two intrinsic abduc-tors, the long, strap-like M11 and the smallwedge-shaped M12, provide the muscular com-ponent to the spreading of the setal fan. Theflaring of the setae and setules and the mainte-nance of the wide setal fan is a passive effectof the fluid dynamic forces acting on the pleo-pods. The unusual construction of the exopodaladductor (M13) clearly indicates a strong mus-cular component to the closing of the setal fan.The three muscle bundles surround a long apo-deme with a wide insertion at the peduncle-ramal joint. This design seems to imply that themuscular abduction-adduction of the exopod iscritical for an efficient pleopod beat. This is re-inforced by the skeletal construction of the pe-duncle-ramal joint that only has a definitelateral condyle. The transverse motion of theexopod appears to govern the design of thepeduncle-ramal joint.
The fundamental design of the pleopods inE. gryllus permits remotion of a wide setal fanfor the power stroke and promotion of a tightlycollapsed pleopod pair on the recovery stroke.Net thrust develops from the positive drag pro-vided by a fully expanded setal fan reduced bythe negative drag engendered on the recoverystroke (Boudrias, in preparation). Ancillary
skeletal structures, particularly those affectingthe rami, can further influence the efficiency ofthe power and recovery strokes. The plumosesetae form a tight meshwork and the pedunclesare hooked together at their distal edge. Thiswide fan leads to an effective power strokewhere the pleopods functionally behave as asolid paddle. This should increase the thrustproduced on the power stroke.
However, it appears that E. gryllus mainlyoptimizes net propulsive thrust by reducing theresistive area of the recovery stroke. This maybe partly accomplished by the intrinsic supportmuscles that change the shape of the peduncle.In addition, the design of the rami appears tocontribute to an efficient recovery stroke. Theyare built to complement each other with theconcave surface of the endopod and the convexsurface of the exopod fitting tightly together onthe recovery stroke. Thus, they effectively pre-sent a single limb to the flow. Moreover, theexopodal hook enhances the paired flexion ofthe rami and clasps them tightly to further re-duce their surface area. The combination offundamental skeletomuscualture and ancillarystructures in E. gryllus leads to an efficientpleopod beat cycle with efficient production ofnet thrust (Boudrias, 1992).
This clasping mechanism on the exopod ofE. gryllus is unusual and is rarely depicted inother amphipods. It is difficult to determinehow prevalent the exopodal hook is, but DeBroyer and Klages (1991) show it in theirdrawing of Epimeria rubrieques De Broyerand Klages, 1991. Similarly, the shape of theexopods and endopods are rarely mentioned,and the design of the joints is rarely described.It would not be surprising if other amphipodshad crucial ancillary structures that improve theproduction of net thrust. A comprehensive sur-vey of pleopod design, with particular attentionto joint construction and special features of therami could provide the necessary data for a sig-nificant interspecific comparison.
A close inspection of pleopod design mighteven reveal new character traits that could beused in distinguishing familial lines within theamphipods and possibly higher taxonomic af-finities. External characters, such as couplinghooks or the exopodal clasping mechanism,could be described and quantified. Internal de-sign constraints, like the presence of cuticularreinforcement in the pleonal wall, the numberand layout of muscles, and the shape and de-sign of the peduncle and the rami, could com-
592 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 3, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

plement existing phylogenetic analyses of thisgroup. My preliminary research on five otherspecies, including Jassus falcata Montagu,1808, Perampithoe humeralis Stimpson, 1864,and Orchomene abyssorum Stebbing, 1888,shows differences in pleopodal design andbody shape among them.
In summary, the design of the pleopods inEurythenes gryllus illustrates complex skeleto-musculature construction. The ancillary struc-tures of these propulsive limbs interact stronglywith the fluid dynamic forces affecting their lo-comotion. The insights gained using both func-tional morphology and behavioral analyses ofswimming have provided the basis for moredetailed kinematic and dynamic analysis of theforces acting on the body and limbs of theseactive swimmers. Because pleopods providepropulsion during swimming and also generatethe water currents needed for respiration, acomplete analysis of their biomechanics, kine-matics, and fluid dynamics is crucial (Boudrias,1991). With more information on pleopods andtheir interaction with body drag, we can beginto understand energy budgets of individual spe-cies, interspecific behavioral and distributionaldifferences, and effects of body and limb de-sign on the hydrodynamics of swimming.Thus, the analysis of the functional morphol-ogy of pleopods may provide more than simpledescriptions of muscles and joints.
ACKNOWLEDGEMENTS
I express my thanks to Drs. Robert R. Hessler, William
A. Newman, and Ken L. Smith, Jr., from the Scripps
Institution of Oceanography, for their insightful comments
and their knowledge of amphipod functional morphology
and ecology. I am particularly indebted to Robert Hessler
and Ken Smith for the use of their collection of deep-sea
amphipods and deep-sea video footage of swimming
amphipods. I was supported by a Natural Science and
Engineering Research Council Scholarship from Canada.
Comments from two anonymous reviewers greatly im-
proved the focus of this manuscript.
LITERATURE CITED
Baldwin, R. J., and K. L. Smith, Jr. 1987. Temporalvariation in catch rate, length, color and sex of thenecrophagous amphipod, Eurythenes gryllus, from thecentral and eastern North Pacific.—Deep-Sea Research34: 425–439.
Barnard, J. L. 1965. Marine Amphipoda of the familyAmpithoidae from southern California.—Proceedings ofthe U.S. National Museum 118: 1–46.
———, and G. S. Karaman. 1991. The families and generaof marine gammaridean Amphipoda (except marinegammaroids).—Records of the Australian Museum,Supplement 13. 866 pp.
Boudrias, M. A. 1991. Methods for the study of amphipodswimming: behavior, morphology, and fluid dynamics.—-Hydrobiologia 223: 11–25.
———. 1992. The biomechanics, kinematics, and fluid
dynamics of swimming in the deep-sea lysianassid
amphipod Eurythenes gryllus (Lichtenstein).—Ph.D.
thesis. Scripps Institution of Oceanography, University
of California, San Diego. 240 pp.Bousfield, E. L. 1973. Shallow-water Gammaridean Am-
phipoda of New England. Cornell University Press,London. 312 pp.
Bowman, T. E. 1974. The ‘‘sea flea’’ Dolobrotus mardeni n.gen., n. sp., a deep-water American lobster bait scavenger(Amphipoda: Eusiridae).—Proceedings of the BiologicalSociety of Washington 87: 129–138.
Bray, R. N., and A. W. Ebeling. 1975. Food, activity, andhabitat of three ‘‘picker-type’’ microcarnivorous fishes inthe kelp forests off Santa Barbara, California.—FisheriesBulletin (U.S.) 73: 815–829.
Brusca, G. J. 1981. On the anatomy of Cystisoma (Am-phipoda: Hyperiidea).—Journal of Crustacean Biology 1:358–375.
Christiansen, B. O., O. Pfannkuche, and H. Thiel. 1990.Vertical distribution and population structure of thenecrophagous amphipod Eurythenes gryllus in the westEuropean basin.—Marine Ecology Progress Series 66:35–45.
Conlan, K. E., and E. L. Bousfield. 1982. The amphipodsuperfamily Corophioidea in the northeastern Pacificregion. Family Ampithoidae: systematics and distribu-tional ecology.—Publications in Biological Oceanogra-phy of National Museums of Canada 10: 41–75.
Dahl, E. 1979. Deep-sea carrion feeding amphipods: evolu-tionary patterns in niche adaptation.—Oikos 33: 167–175.
De Broyer, C., and M. Klages. 1991. A new Epimeria(Crustacea, Amphipoda, Paramphithoidae) from the Wed-dell Sea.—Antarctic Science 3: 159–166.
Dunham, P., T. Alexander, and A. Hursham. 1986.Precopulatory mate guarding in an amphipod, Gammaruslawrencianus Bousfield.—Animal Behaviour 34: 1680–1686.
George, R. Y. 1979a. What adaptive strategies promoteimmigration and speciation in deep-sea environments?—-Sarsia 64: 61–65.
———. 1979b. Behavioral and metabolic adaptations of
polar and deep-sea crustaceans: a hypothesis concerning
physiological basis for evolution of cold adapted
crustaceans.—Bulletin, Biological Society of Washington
3: 283–296.Hargrave, B. T. 1985. Feeding rates of abyssal scavenging
amphipods (Eurythenes gryllus) determined in situ bytime-lapse photography.—Deep-Sea Research 32: 443–450.
Hessler, R. R. 1981. Evolution of arthropod locomotion: acrustacean model. Pp. 9–30 in C. F. Herreid and C. R.Fourtner, eds. Locomotion and Exercise of Arthropods.Plenum Press, New York.
———. 1982. The structural morphology of walking
mechanisms in eumalacostracan crustaceans.—Philo-
sophical Transanctions of the Royal Society of London
296B: 245–298.
———. 1985. Swimming in Crustacea.—Transactions of
the Royal Society of Edinburgh 76: 115–122.
———, C. L. Ingram, A. A. Yayanos, and B. R. Burnett.1978. Scavenging amphipods from the floor of thePhilippine Trench.—Deep-Sea Research 25: 1029–1047.
593BOUDRIAS: PLEOPOD FUNCTIONAL MORPHOLOGY
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022

———, J. D. Isaacs, and E. L. Mills. 1972. Giant amphipodfrom the abyssal Pacific Ocean.—Science 175: 636–637.
Hobson, E. S., and J. R. Chess. 1976. Trophic interactionsamong fishes and zooplankters nearshore at SantaCatalina Island, California, USA.—Fisheries Bulletin(U.S.) 74: 567–598.
———. 1978. Trophic relationships among fishes andplankton in the lagoon at Enewetak Atoll, MarshallIslands.—Fisheries Bulletin (U.S.) 76: 133–153.
Holmquist, J. G. 1982. The functional morphology ofgnathopods: importance in grooming and variation withregard to habitat in talitroidean amphipods.—Journal ofCrustacean Biology 2: 159–179.
Ingram, C. L., and R. R. Hessler. 1983. Distribution andbehavior of scavenging amphipods from the central NorthPacific.—Deep-Sea Research 30: 683–706.
———. 1987. Population biology of the deep-sea amphipodEurythenes gryllus: inferences from instar analyses.—Deep-Sea Research 34: 1889–1910.
Kils, U. 1981. The swimming behavior, swimmingperformance, and energy balance of Antarctic krill,Euphausia superba.—SCAR Biomass Report Series 30:122 pp.
Lampitt, R. S., N. R. Merrett, and M. H. Thurston. 1983.Inter-relations of necrophagous amphipods, a fish pred-ator, and tidal currents in the deep sea.—Marine Biology74: 73–78.
Laver, M. B., M. S. Olsson, J. L. Edelman, and K. L. Smith,Jr. 1985. Swimming rates of scavenging deep-seaamphipods recorded with a free-vehicle video camera.—-Deep-Sea Research 32: 1135–1142.
Lobel, P. S., and J. E. Randall. 1986. Swarming behavior ofthe hyperiid amphipod Anchylomera.—Journal of Plank-ton Research 8: 253–262.
Lowry, J. K. 1986. The callynophore, a eucaridean/pera-caridean sensory organ prevalent among the Amphipoda(Crustacea).—Zoologica Scripta 15: 333–349.
Marshall, N. B. 1979. Developments in Deep-Sea Biology.Blandford Press, Poole, Dorset. 566 pp.
Sainte-Marie, B. 1984. Morphological adaptations forcarrion feeding in four species of littoral or circalittorallysianassid amphipods.—Canadian Journal Zoology 62:1668–1674.
———. 1986. Feeding and swimming of lysianassidamphipods in a shallow cold-water bay.—Marine Biology91: 219–229.
———, and P. Brunel. 1985. Suprabenthic gradients ofswimming activity by cold-water gammaridean amphipodCrustacea over a muddy shelf in the Gulf of SaintLawrence.—Marine Ecology Progress Series 23: 57–69.
———, and B. T. Hargrave. 1987. Estimation of scavengerabundance and distance of attraction to bait.—MarineBiology 94: 431–443.
———, G. Lamarche. 1985. The diets of six species of thecarrion-feeding lysianassid amphipod genus Anonyx andtheir relation with morphology and swimming behav-ior.—Sarsia 70: 119–126.
Sars, G. O. 1895. An Account of the Crustacea of Norway:Amphipoda. Christiana and Copenhagen. Vol. 1, 711 pp.,240 pl.
Schram, F. 1986. Crustacea. Oxford University Press. 606 pp.Smith, K. L., Jr., and R. J. Baldwin. 1982. Scavenging deep-
sea amphipods: effects of food odor on oxygen consump-tion and a proposed metabolic strategy.—Marine Biology68: 287–298.
———, and ———. 1984. Vertical distribution of thenecrophagous amphipod, Eurythenes gryllus in the NorthPacific: spatial and temporal variation.—Deep-Sea Re-search 31: 1179–1196.
Stebbing, T. R. R. 1888. Report on the Amphipoda collectedby H.M.S. Challenger during the years 1873–1876.—Reports Voyages Challenger 1873–1876 (Zoology) 29: 1–1737, pl.
Steele, D. H. 1988. What is the amphipod life style?—Crustaceana Supplement 13: 134–142.
Stepien, C. A., and R. C. Brusca. 1985. Nocturnal attacks onnearshore fishes in southern California by crustaceanzooplankton.—Marine Ecology Progress Series 25: 91–105.
Templeman, W. 1967. Predation on living fishes on longlinein Baffin Bay by the amphipod Eurythenes gryllus(Lichtenstein), and a new distribution record.—Journal ofthe Fisheries Research Board of Canada 24: 215–217.
Thurston, M. H. 1979. Scavenging abyssal amphipods fromthe North-East Atlantic ocean.—Marine Biology 51:55–68.
Vogel, F. 1985a. Das Schwimmen der Talitridae (Crustacea:Amphipoda): Functionmorphologie, Phanomenologie undEnergetik.—Helgolander Meeresuntersuchungen 39:303–339.
———. 1985b. Die abdominale muskulatur von Orchestiacavimana Heller, 1865 (Amphipoda, Talitridae).—Crus-taceana 50: 11–26.
Waterman, T. H. (ed.). 1961. The Physiology of Crustacea.Vol. 1 and 2. Academic Press, New York
Yayanos, A. A. 1978. Recovery and maintenance of liveamphipods at a pressure of 580 bars from an ocean depthof 5700 m.—Science 200: 1056–1059.
———. 1981. Reversible inactivation of deep-sea amphi-pods (Paralicella capresca) by a decompression from 601bars to atmospheric pressure.—Comparative Biochemis-try Physiology 69A: 563–565.
RECEIVED: 15 April 1999.ACCEPTED: 26 November 2002.
594 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 22, NO. 3, 2002
Dow
nloaded from https://academ
ic.oup.com/jcb/article/22/3/581/2679762 by guest on 31 M
ay 2022




















