
1201447.pdf - Oxford Academic
14
Brain (1997), 120, 1447–1460 Increased immunoreactivity to two overlapping peptides of myelin proteolipid protein in multiple sclerosis Judith M. Greer, 1 Peter A. Csurhes, 1 Kaye D. Cameron, 1 Pamela A. McCombe, 1,2 Michael F. Good 3 and Michael P. Pender 1,2 1 Department of Medicine, University of Queensland, Correspondence to: Dr Judith Greer, Neuroimmunology 2 Department of Neurology, Royal Brisbane Hospital and Research Unit, Department of Medicine, University of 3 Queensland Institute of Medical Research, Brisbane, Queensland, Clinical Sciences Building, Royal Brisbane Australia Hospital, Herston, QLD 4029, Australia Summary We tested the proliferative responses of peripheral blood Status Scale . 4.0); the blood from 15 of 19 patients in this group reacted to one or both of the peptides. Both peptides mononuclear cells from 61 patients with multiple sclerosis, 56 healthy control subjects and 52 patients with other could be recognized by short-term T-cell lines specific for whole PLP, and lines specific for one or other of the two neurological diseases to seven synthetic peptides of myelin proteolipid protein (PLP) and 19 synthetic peptides of myelin overlapping peptides were able to recognize whole PLP, indicating that these peptides can be processed naturally from basic protein (MBP). Increased proliferative responses to two overlapping PLP peptides, PLP 184–199 and PLP 190–209 , the intact molecule. This region of PLP is encephalitogenic in a number of strains of mice. Samples from multiple sclerosis were found significantly more frequently in blood from patients with relapsing–remitting or secondary progressive patients did not react more frequently to any of the MBP peptides than those from healthy control subjects. The multiple sclerosis (52.3%), but not from those with primary progressive multiple sclerosis (18.2%), than in that from proportions of patients with other neurological diseases whose blood responded to the MBP peptides that most healthy control subjects (8.9%) and patients with other neurological diseases (20.8%). Reactivity to these PLP frequently elicited responses in blood from multiple sclerosis patients were significantly lower than the proportions of peptides was most frequently seen in blood from patients with multiple sclerosis of 6–15 years duration and with multiple sclerosis patients and healthy control subjects whose blood responded to these peptides. moderate to severe disability (Kurtzke’s Expanded Disability Keywords: multiple sclerosis; proteolipid protein; immunoreactivity; myelin basic protein; autoimmunity Abbreviations: ANOVA 5 analysis of variance; EAE 5 experimental autoimmune encephalomyelitis; EDSS 5 Kurtzke’s Expanded Disability Status Scale; HLA 5 human leucocyte antigen; MBP 5 myelin basic protein; MHC 5 major histocompatibility complex; PBMC 5 peripheral blood mononuclear cells; PCR 5 polymerase chain reaction; PLP 5 myelin proteolipid protein; SI 5 stimulation index Introduction Multiple sclerosis is a human demyelinating disorder that is (PLP) and myelin basic protein (MBP). PLP comprises more than 50% of the protein in CNS myelin. It is expressed in likely to have an autoimmune basis (Pender, 1995). As the demyelinated lesions in multiple sclerosis are usually small amounts (, 0.5% of total protein) by Schwann cells in the PNS (peripheral nervous system), but it is not restricted to the CNS (Lumsden, 1970; Sobel, 1995), it is likely that the target antigens of autoaggressive cells in incorporated into PNS myelin (Puckett et al., 1987). In experimental animals, immunization with PLP and adjuvants multiple sclerosis are expressed primarily or exclusively in the CNS. CNS myelin proteins are possible targets. The most results in demyelinated lesions that are restricted to the CNS (Tuohy et al., 1988; Greer et al., 1992, 1996a; Chalk et al., abundant proteins of CNS myelin are proteolipid protein © Oxford University Press 1997
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of 1201447.pdf - Oxford Academic

Brain (1997),120,1447–1460
Increased immunoreactivity to two overlappingpeptides of myelin proteolipid protein in multiplesclerosisJudith M. Greer,1 Peter A. Csurhes,1 Kaye D. Cameron,1 Pamela A. McCombe,1,2 Michael F. Good3
and Michael P. Pender1,2
1Department of Medicine, University of Queensland, Correspondence to: Dr Judith Greer, Neuroimmunology2Department of Neurology, Royal Brisbane Hospital and Research Unit, Department of Medicine, University of3Queensland Institute of Medical Research, Brisbane, Queensland, Clinical Sciences Building, Royal BrisbaneAustralia Hospital, Herston, QLD 4029, Australia
SummaryWe tested the proliferative responses of peripheral blood Status Scale. 4.0); the blood from 15 of 19 patients in this
group reacted to one or both of the peptides. Both peptidesmononuclear cells from 61 patients with multiple sclerosis,56 healthy control subjects and 52 patients with other could be recognized by short-term T-cell lines specific for
whole PLP, and lines specific for one or other of the twoneurological diseases to seven synthetic peptides of myelinproteolipid protein (PLP) and 19 synthetic peptides of myelin overlapping peptides were able to recognize whole PLP,
indicating that these peptides can be processed naturally frombasic protein (MBP). Increased proliferative responses totwo overlapping PLP peptides, PLP184–199 and PLP190–209, the intact molecule. This region of PLP is encephalitogenic in
a number of strains of mice. Samples from multiple sclerosiswere found significantly more frequently in blood frompatients with relapsing–remitting or secondary progressive patients did not react more frequently to any of the MBP
peptides than those from healthy control subjects. Themultiple sclerosis (52.3%), but not from those with primaryprogressive multiple sclerosis (18.2%), than in that from proportions of patients with other neurological diseases
whose blood responded to the MBP peptides that mosthealthy control subjects (8.9%) and patients with otherneurological diseases (20.8%). Reactivity to these PLP frequently elicited responses in blood from multiple sclerosis
patients were significantly lower than the proportions ofpeptides was most frequently seen in blood from patientswith multiple sclerosis of 6–15 years duration and with multiple sclerosis patients and healthy control subjects whose
blood responded to these peptides.moderate to severe disability (Kurtzke’s Expanded Disability
Keywords: multiple sclerosis; proteolipid protein; immunoreactivity; myelin basic protein; autoimmunity
Abbreviations: ANOVA 5 analysis of variance; EAE5 experimental autoimmune encephalomyelitis; EDSS5 Kurtzke’sExpanded Disability Status Scale; HLA5 human leucocyte antigen; MBP5 myelin basic protein; MHC5 majorhistocompatibility complex; PBMC5 peripheral blood mononuclear cells; PCR5 polymerase chain reaction; PLP5 myelinproteolipid protein; SI5 stimulation index
IntroductionMultiple sclerosis is a human demyelinating disorder that is (PLP) and myelin basic protein (MBP). PLP comprises more
than 50% of the protein in CNS myelin. It is expressed inlikely to have an autoimmune basis (Pender, 1995). Asthe demyelinated lesions in multiple sclerosis are usually small amounts (, 0.5% of total protein) by Schwann cells
in the PNS (peripheral nervous system), but it is notrestricted to the CNS (Lumsden, 1970; Sobel, 1995), it islikely that the target antigens of autoaggressive cells in incorporated into PNS myelin (Puckettet al., 1987). In
experimental animals, immunization with PLP and adjuvantsmultiple sclerosis are expressed primarily or exclusively inthe CNS. CNS myelin proteins are possible targets. The most results in demyelinated lesions that are restricted to the CNS
(Tuohy et al., 1988; Greeret al., 1992, 1996a; Chalk et al.,abundant proteins of CNS myelin are proteolipid protein
© Oxford University Press 1997

1448 J. M. Greeret al.
1994a, b), as in multiple sclerosis. In contrast, MBP is found presence of salts. This makes the PLP protein a poor antigenin in vitro tissue culture studies, as the precipitated, flocculentin significant quantities (20–30% of total protein) in both
CNS and PNS myelin and, in experimental animals, PLP can inhibit cell–cell contact and subsequent cellactivation. However, Markovic-Pleseet al. (1995), usingimmunization with MBP (or whole CNS tissue) and adjuvants
or the passive transfer of MBP-specific lymphocytes results multiplein vitro stimulations with human PLP, have foundthat the protein can be processed by antigen-presenting cellsin demyelinated lesions in both the CNS and PNS (Pender
and Sears, 1982, 1984; Pender, 1988a, b; Penderet al., 1989, into immunodominant epitopes within the 30–60 and 180–210 regions of the molecule. Peptides covering these regions1995; Chalket al., 1994a, b; Abromson-Leemanet al., 1995).
A number of studies have investigated whether multiple are included in the current study.sclerosis patients have an increased frequency of cells reactiveto myelin antigens, including PLP or MBP, in their blood orCSF. These studies have been performed using a variety ofMethodsmethodologies and antigen preparations, often on only smallPatients and control subjectsnumbers of subjects, and the results have been inconsistent.The subjects of this study consisted of 61 patients withMany of these studies have involved the production of long-clinically definite or laboratory-supported definite multipleterm T-cell lines and clones. Only a minority of the studiessclerosis (Poseret al., 1983), 56 healthy control subjects andhave looked at the immediate proliferative response of52 patients with other neurological diseases. Of the multipleperipheral blood mononuclear cells (PBMC); these studiessclerosis patients, 28 had relapsing–remitting multiplehave investigated responses to whole MBP (Lisak andsclerosis, 19 had secondary progressive multiple sclerosisZweiman, 1977; Brinkmanet al., 1982; Johnsonet al., 1986; and 14 had primary progressive multiple sclerosis. MultipleVandenbarket al., 1989; Trotteret al., 1991; Kerlero de sclerosis patients had not received corticosteroids for atRosbo et al., 1993; Zhanget al., 1993), MBP fragments least 2 months prior to being studied. Patients with other(Baxevaniset al., 1989), whole PLP and/or two synthetic neurological diseases had the following diagnoses: epilepsyPLP peptides (Johnsonet al., 1986; Trotteret al., 1991; (12), Parkinson’s disease (seven), Wilson’s disease (one),Kerlero de Rosboet al., 1993), and two quantitatively Huntington’s chorea (one), brain tumour (three), cerebralminor myelin components, MOG (myelin/oligodendrocyte aneurysm (two), intracranial arteriovenous malformationglycoprotein) (Kerlero de Rosboet al., 1993) and MAG (one), cerebrovascular disease (five), hydrocephalus (one),(myelin-associated glycoprotein) (Zhanget al., 1993). These neurofibromatosis type I (one), neurosarcoidosis (one),have produced diverse results with little concurrence. sleeping disorder (one), spinocerebellar degeneration (one),
Because there exists a correlation between the expressionmotor neuron disease (three), multifocal motor neuropathyof the major histocompatibility complex (MHC) class II (one), hereditary sensory neuropathy (one), Guillain–Barre´haplotypes, human leucocyte antigens (HLA) DR2, DW2syndrome (two), non-specified peripheral neuropathy (four),(HLA-DRB1*1501, -DQA1*0102 and -DQB1*0602), and C8 nerve root lesion (one), intercostal nerve entrapmentsusceptibility to multiple sclerosis (Stewartet al., 1981; (one), myasthenia gravis (one) and proximal myopathy (one).Hillert et al., 1994), it is thought that the pathogenesis ofmultiple sclerosis may involve T cells that recognize theautoantigen(s) in the context of these MHC class II molecules.
Tissue typingAntigens presented by MHC class II molecules are generallyGenomic DNA was prepared from Epstein–Barr-virus-12–25 amino acids in length (Rammenseeet al., 1995).transformed lymphoblastoid cell lines from each individualTherefore, the aim of the current study was to assessby chloroform/phenol extraction. HLA-DR typing was carriedthe proliferative responses of freshly isolated PBMC fromout at the Queensland Institute of Medical Research, Brisbane,multiple sclerosis patients, healthy control subjects andAustralia, and HLA-DQ typing at the Department ofpatients with other neurological diseases to multiple shortImmunology, Westmead Hospital, Sydney, Australia. For(12–20 amino acid residues) synthetic peptides of PLP andboth HLA-DR and -DQ typing, DNA was initially amplifiedMBP, in order to determine if there is a correlation betweenby the polymerase chain reaction (PCR) using generic primermultiple sclerosis and reactivity to these antigens.pairs. Specific HLA-DRB1 alleles were then determined bySeveral PLP epitopes that are immunodominant in a varietyhybridization of the PCR products with a panel of sequence-of genetically diverse mouse strains (Greeret al., 1996a)specific oligonucleotides. HLA-DQA1 and -DQB1 allelesand in humans (Markovic-Pleseet al., 1995) have notwere determined by restriction fragment length polymorphismpreviously been tested for their ability to stimulate freshlyanalysis of the PCR products.isolated human PBMC, and no previous studies have
assessed reactivity to peptides within all four major isoformsof human MBP. The use of synthetic peptides is particularlywarranted in the case of PLP which, because of its veryAntigens
Human PLP was prepared from human brain white matterhydrophobic nature, has a strong tendency to form highmolecular weight aggregates that readily precipitate in the as previously described (Greeret al., 1996a). PLP peptides

Immunoreactivity to PLP in multiple sclerosis 1449
Fig. 1 (A) The sequence of human PLP and its proposed orientation in the oligodendrocyte cellmembrane (Greeret al., 1996b) showing the peptides used in the current study in bold. (B) The fourisoforms of human MBP, showing the exon 2 insert (encoding 26 amino acids) in the 21.5 and20.2 kDa isoforms, and exon 5 (encoding 11 amino acids) which is deleted in the 20.2 and 17.3 kDaisoforms.
were prepared according to the human sequence (Hudson peptides (numbered according to the human sequence andlisted in Table 6) were synthesized (at the Queenslandet al., 1989) and were.90% pure by HPLC analysis. The
sequence of PLP showing the peptides used in the present Institute of Medical Research) to represent the sequences ofthe 4 major isoforms (21.5 kDa, 20.2 kDa, 18.5 kDa andstudy and the putative orientation of PLP in the myelin
membrane is shown in Fig. 1A. PLP41–58 (GTEKLIETY- 17.3 kDa; shown in Fig. 1B) of human MBP (Rothet al.,1987) and were.80% pure. Peptides 5b and 5c representFSKNYQDYE) and PLP95–114 (AVRQIFGDYKTTICGK-
GLSA) were synthesized at the Queensland Institute of the exon 2 insert (encoding 26 amino acids) in the 21.5 and20.2 kDa isoforms. Peptide 10b covers the junctional zoneMedical Research; PLP178–191 (NTWTTCQSIAFPSK),
PLP184–199(QSIAFPSKTSASIGSL) and PLP190–209(SKTSA- formed by deletion of exon 5 (encoding 11 amino acids) inthe 20.2 and 17.3 kDa isoforms. All MBP peptides areSIGSLCADARMYGVL) were synthesized by Auspep
(Melbourne, Australia); and PLP100–119 (FGDYKTTICGK- water-soluble.GLSATVTGG) and PLP139–151(HCLGKWLGHPDKF) werethe kind gift of Dr M. B. Lees, Shriver Center, Waltham,Mass., USA. PLP100–119 and PLP139–151 are water-soluble. Proliferation assays
Heparinized peripheral blood (~60 ml) was collected byThe other PLP peptides are moderately hydrophobic, andwere dissolved at a concentration of 5 mg/ml in 0.2 M venepuncture from each subject after informed written
consent had been obtained. PBMC were separated fromacetic acid prior to dilution in tissue-culture medium forproliferation assays. blood by centrifugation through Histopaque (Sigma Chemical
Co., St Louis, USA) and washed twice. Thein vitro responsesHuman MBP was extracted from human brain accordingto the method of Deibleret al. (1972). Overlapping MBP of PBMC were assayed in triplicate in 96-well round-

1450 J. M. Greeret al.
Table 1 Characteristics of multiple sclerosis patients and study (25 and 12.5µg/ml final concentration for PLP peptidescontrol subjects or 30 µg/ml final concentration for MBP peptides) were
those at which the most responses occurred in the initialParameter PBMC tested against*studies. Cultures were incubated for 6 days, with 0.5µCi
Group PLP peptides MBP peptides [3H]thymidine being added during the last 18 h. Cultureswere harvested and thymidine uptake was measured in
Number (female : male) c.p.m. (counts per minute) in aβ-plate counter (LKB). TheMS (whole group) 55 (40 : 15) 61 (45 : 16)stimulation index (SI) was determined by the formula: SI5PP-MS 11 (7 : 4) 14 (9 : 5)(mean c.p.m. of peptide-containing triplicate wells)/(meanRR-MS 26 (21 : 5) 28 (22 : 6)
SP-MS 18 (12 : 6) 19 (14 : 5) c.p.m. of control triplicate wells, without peptide). TheHealthy controls 54 (32 : 22) 56 (29 : 27) mean control ranged from 191 to 6071 c.p.m. A positiveOND 48 (19 : 29) 52 (23 : 29) proliferative response for each sample was considered to beMean age in years (range)
one in which the SIù 3.0 and the mean from the controlMS (whole group) 44.3 (15–70) 45.0 (15–70)wells plus 3SD of the mean did not exceed the mean fromPP-MS 49.5 (31–70) 50.1 (31–70)
RR-MS 37.9 (15–57) 38.1 (15–57) the peptide-containing triplicate wells.SP-MS 50.4 (30–65) 50.1 (30–65)Healthy controls 36.3 (15–61) 35.7 (15–61)OND 49.5 (19–75) 50.3 (16–75)
Mean years duration (range)Generation and testing of short-term T-cell linesMS (whole group) 11.0 (1–31) 10.6 (1–28)Short-term T-cell lines were prepared from three multiplePP-MS 5.6 (1–11) 6.2 (1–14)
RR-MS 8.8 (1–26) 8.7 (1–26) sclerosis patients by incubation of PBMC (43106 cells perSP-MS 17.6 (6–31) 16.7 (6–28) well) with 25 µg/ml of human PLP, PLP184–199or PLP190–209Healthy controls Not applicable Not applicable
in 24-well plates. Lines were restimulated 10 days later withOND Not applicable Not applicableantigen plus irradiated (3000 rad) autologous PBMC andMean EDSS score (range)then tested 12–15 days later in proliferative assays. For theseMS (whole group) 5.4 (0–9.5) 5.5 (0–9.5)
PP-MS 5.9 (3.5–7.5) 6.1 (3.5–7.5) assays, 53104 T cells were added to 105 irradiated autologousRR-MS 3.6 (0–8.0) 3.9 (0–8.0) PBMC and antigen in 96-well microtitre trays. Cultures wereSP-MS 7.3 (5.0–9.5) 7.4 (5.0–9.5)
incubated for 4 days, with 0.5µCi [3H]thymidine beingHealthy controls Not applicable Not applicableadded during the last 18 h. Cultures were harvested andOND Not applicable Not applicablethymidine uptake was measured as described above.HLA-Dw21
MS (whole group) 80% 80%PP-MS 90% 87%RR-MS 69% 68%SP-MS 89% 94% Statistical analysisHealthy controls 34% 34%
Percentages of individuals making a positive proliferativeOND 36% 37%response to peptides were compared using theχ2 test with
*The peripheral blood mononuclear cells (PBMC) of some Yates’ correction applied as required. Mean SI values werepatients were only tested for reactivity to myelin proteolipid compared using analysis of variance (ANOVA) to compareprotein (PLP) or myelin basic protein (MBP) peptides as
all the groups simultaneously, followed by Student’st test todescribed in text. MS5 multiple sclerosis; PP-MS5 primarycompare the pairs of groups.progressive multiple sclerosis; RR-MS5 relapsing–remitting
multiple sclerosis; SP-MS5 secondary progressive multiplesclerosis; OND5 other neurological diseases; EDSS5 Kurtzke’sExpanded Disability Status Scale; HLA-Dw21 5 percentage ofindividuals expressing one or more of the alleles DRB1*1501, ResultsDQA1*0102 or DQB1*0602.
Characteristics of multiple sclerosis patients andcontrol subjectsbottomed microtitre plates. PBMC (23105) and medium
(RPMI 1640 containing 10% heat-inactivated pooled human The characteristics of multiple sclerosis patients and controlsubjects are detailed in Table 1. PBMC from all patients withserum, 2 mML-glutamine, 10 mM HEPES buffer and 50
µM 2-mercaptoethanol) with or without peptides were added other neurological diseases and all except one multiplesclerosis patient that were tested for reactivity to PLP peptidesto wells in a total volume of 200µl. Initial studies were
done using blood from five laboratory staff, all of whom had were concurrently tested for reactivity to the MBP peptides.Seven additional multiple sclerosis patients and fourbeen exposed to both PLP and MBP for several years, and
four multiple sclerosis patients to determine the optimal additional patients with other neurological diseases weretested for reactivity to MBP peptides, but not PLP peptides.concentration of antigen to use in the main part of the study.
Peptides were tested at concentrations ranging from 1 to In the healthy control group, three females were tested onlyfor reactivity to PLP peptides, and five males were tested100 µg/ml. The concentrations used in the main part of the

Immunoreactivity to PLP in multiple sclerosis 1451
these clusters were not significantly different from those ofhealthy control subjects or of patients with other neurologicaldiseases (Fig. 2). However, there was a slight increase in thepercentage of multiple sclerosis patients with cells respondingto PLP41–58, and the mean SI of multiple sclerosis patientsfor PLP41–58 was significantly higher than the mean SI ofcells from patients with other neurological diseases, but notof those from healthy control subjects (Table 2). All threegroups showed a relatively high percentage of subjects withcells responding to PLP95–114(Fig. 2) and the mean SI valuesfor this peptide were high in all groups compared with valuesfor other peptides (Table 2). This reactivity correlated stronglywith the expression of the HLA-DRB1*1101 allele: 93% ofall individuals expressing this allele had cells which respondedto PLP95–114, and ~60% of ‘responders’ in each of thethree groups expressed this allele. More patients with otherneurological diseases responded to PLP100–119and PLP139–151
than did healthy control subjects or multiple sclerosis patients;however, the reactivity in patients with other neurologicaldiseases did not correlate strongly with any particular diseaseor with expression of a particular HLA-DR or -DQ allele.Few subjects from any group responded to PLP178–191(Fig.
Fig. 2 Percentages of healthy control subjects (HC), and patients2 and Table 2).with multiple sclerosis (MS) and other neurological diseases
In contrast, the percentage of multiple sclerosis patients(OND), responding with SIù 3.0 to PLP peptides. TheP valuesfor comparison of the responses of the three groups together withwith PBMC in their blood that responded to PLP190–209wasthose for the comparisons of the pairs of groups (χ2 analysis) are significantly higher than the corresponding percentages ofshown directly below the peptide to which they refer. healthy control subjects and of patients with other
neurological diseases (Fig. 2). The difference betweenmultiple sclerosis patients and patients with other neurologicalonly for reactivity to MBP peptides. As has previously been
found for other groups of multiple sclerosis patients, our diseases could not be explained by the predominance ofmales in the group of patients with other neurologicalmultiple sclerosis patients were predominantly female, the
mean age of onset of multiple sclerosis was higher for diseases, as the percentage of male patients with otherneurological diseases responding to this peptide was greaterprimary progressive multiple sclerosis than for relapsing–
remitting multiple sclerosis and secondary progressive than the percentage of females. The percentage of multiplesclerosis patients responding to the overlapping peptidemultiple sclerosis, and the percentage of patients expressing
the HLA-Dw2 haplotype was increased in multiple sclerosis PLP184–199 was also increased, but only the comparisonwith healthy control subjects reached statistical significance.patients compared with control subjects.Reactivity to these peptides was not restricted to individualsexpressing any particular HLA-DR or -DQ allele. Themajority of responders in the multiple sclerosis group (91%)Reactivity to PLP peptides
Epitopes of PLP that are immunogenic in a variety of expressed HLA-Dw2 alleles. However, only 53% ofresponders in the control groups expressed these alleles. Theexperimental animals cluster in three regions of the molecule
(Greer et al., 1996a). Peptides covering these regions expression of DRB1*1501 in combination with DRB1*0401,DRB1*0404 or DRB1*0701 did, however, appear to be(residues 40–60, 90–120 and 180–210), together with PLP139–
151, which is an encephalitogenic epitope in SJL mice related to increased responses to PLP184–199and PLP190–209.The mean SI value of multiple sclerosis patients for PLP190–(Tuohy et al., 1989), were tested for their ability to induce
proliferative responses with SIù 3.0 in freshly isolated 209 was significantly higher than that of healthy controlsubjects, but not of patients with other neurological diseasesPBMC collected from 55 multiple sclerosis patients, 54
healthy control subjects and 48 patients with other (Table 2). The mean SI of multiple sclerosis patients forPLP184–199 was increased compared with that of healthyneurological diseases. Peptides from two of the PLP epitope
clusters (residues 40–60 and 90–120) have previously been control subjects and that of patients with other neurologicaldiseases, but comparison of the three groups by ANOVA didused to derive T-cell lines from multiple sclerosis patients’
lymphocytes (Pelfreyet al., 1993, 1994; Correaleet al., not reach statistical significance.Within the multiple sclerosis group, reactivity to1995; Markovic-Pleseet al., 1995; Ohashiet al., 1995). In
the present study, however, the percentages of multiple PLP184–199 and PLP190–209 occurred most frequently inpatients with relapsing–remitting multiple sclerosis orsclerosis patients with cells responding to peptides within

1452 J. M. Greeret al.
Table 2 Mean stimulation index (SI6 SE) of PBMC from healthy control subjects, multiplesclerosis patients and patients with other neurological diseases to PLP peptides
Peptide Healthy Multiple Other neurological P-valuecontrols sclerosis diseases ANOVA*
PLP41–58 1.8 6 0.4 2.26 0.3** 1.6 6 0.2 0.019PLP95–114 2.9 6 0.4 3.06 0.9 2.56 0.6 0.764PLP100–119 1.6 6 0.1 1.56 0.1 2.56 0.4 0.066PLP139–151 1.2 6 0.1 1.56 0.1 2.06 0.3 0.113PLP178–191 1.5 6 0.1 1.56 0.2 1.96 0.5 0.799PLP184–199 1.7 6 0.2 3.06 0.6 2.26 0.5 0.068PLP190–209 1.4 6 0.1 2.16 0.2*** 1.7 6 0.2 0.012
*If P , 0.05 with ANOVA, then Student’st test was used to compare pairs of groups.** Mean SI ofmultiple sclerosis patients is significantly different from the mean SI of patients with other neurologicaldiseases (Student’st test;P 5 0.037), but not of healthy control subjects.*** Mean SI of multiplesclerosis patients is significantly different from the mean SI of healthy control subjects (P 5 0.003), butnot of patients with other neurological diseases.
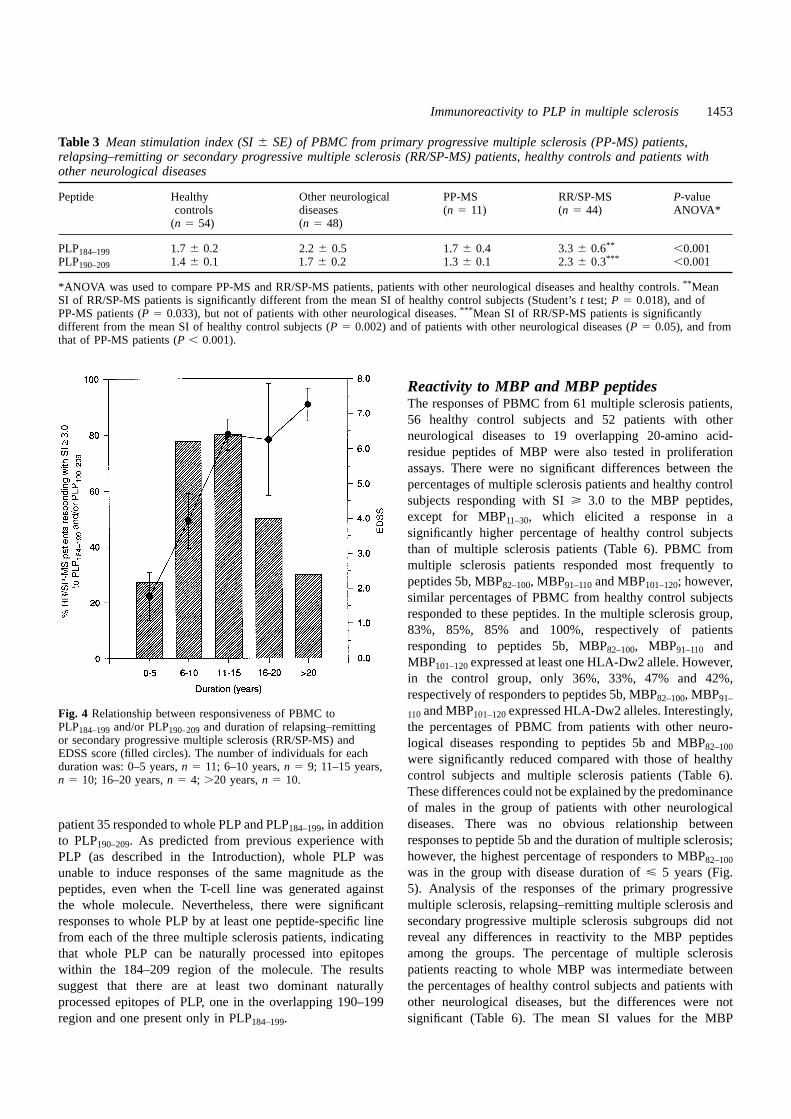
of primary progressive multiple sclerosis patients for thesepeptides were significantly lower than those of patients withrelapsing–remitting/secondary progressive multiple sclerosis(Table 3). Thehighest frequency of reactivity to PLP184–199and/or PLP190–209occurred in those relapsing–remitting/secondaryprogressive multiple sclerosis patients who had a diseaseduration of 6–15 years (Fig. 4). Fifteen of 19 patients with thisduration of disease responded to one or both peptides. Themean SI values of patients with relapsing–remitting/secondaryprogressive multiple sclerosis peaked in the 11–15 yearsduration group, at which time the mean SI values weresignificantly different from those of both healthy controlsubjects and patients with other neurological diseases (Table4). The differences between primary progressive multiplesclerosis patients and relapsing–remitting/secondaryprogressive multiple sclerosis patients are unlikely to be solelydue to the shorter disease duration in primary progressivemultiple sclerosis patients, as the mean SI values for primaryprogressive multiple sclerosis patients with a disease durationof 6–11 years were substantially lower than those for patientswith relapsing–remitting/secondary progressive multiplesclerosis with a duration of 6–10 years (Table 4).
To determine whether the PLP184–199 and PLP190–209
peptides can be naturally processed from the whole PLPFig. 3 Percentages of healthy control subjects (HC), andmolecule, short-term T-cell lines specific for whole humanpatients with primary progressive multiple sclerosis (PP-MS),PLP, PLP184–199 or PLP190–209 were prepared from threerelapsing–remitting or secondary progressive multiple sclerosis
(RR/SP-MS), or other neurological diseases (OND), responding tomultiple sclerosis patients who responded to these peptidesPLP184–199and/or PLP190–209. TheP values are shown directly in the initial assays. The T-cell lines were then tested forbelow the peptide(s) to which they refer (χ2 analysis). their ability to proliferate in response to the three antigens.
Short-term PLP-specific lines from multiple sclerosis patientsecondary progressive multiple sclerosis, who were grouped22 and multiple sclerosis patient 45 showed increasedtogether (relapsing–remitting/secondary-progressive multipleresponses to PLP184–199 and PLP190–209; however, the PLP-sclerosis) (Fig. 3) because the results were similar in the twospecific line from multiple sclerosis patient 35 did not respondgroups. Reactivity to these peptides was found significantlysignificantly to either peptide (Table 5). All of the linesmore frequently in patients with relapsing–remitting/ specific for PLP184–199 responded significantly to all threesecondaryprogressivemultiplesclerosis than inhealthycontrolantigens. In contrast, the response of PLP190–209-specific linessubjects or patients with other neurological diseases (Fig. 3).was variable, with lines from multiple sclerosis patient 22Few patients with primary progressive multiple sclerosisand multiple sclerosis patient 45 responding significantly
only to PLP190–209, whereas the line from multiple sclerosisreacted to these peptides (Fig. 3), and the mean SI values
Immunoreactivity to PLP in multiple sclerosis 1453
Table 3 Mean stimulation index (SI6 SE) of PBMC from primary progressive multiple sclerosis (PP-MS) patients,relapsing–remitting or secondary progressive multiple sclerosis (RR/SP-MS) patients, healthy controls and patients withother neurological diseases
Peptide Healthy Other neurological PP-MS RR/SP-MS P-valuecontrols diseases (n 5 11) (n 5 44) ANOVA*
(n 5 54) (n 5 48)
PLP184–199 1.7 6 0.2 2.26 0.5 1.76 0.4 3.36 0.6** ,0.001PLP190–209 1.4 6 0.1 1.76 0.2 1.36 0.1 2.36 0.3*** ,0.001
*ANOVA was used to compare PP-MS and RR/SP-MS patients, patients with other neurological diseases and healthy controls.** MeanSI of RR/SP-MS patients is significantly different from the mean SI of healthy control subjects (Student’st test;P 5 0.018), and ofPP-MS patients (P 5 0.033), but not of patients with other neurological diseases.*** Mean SI of RR/SP-MS patients is significantlydifferent from the mean SI of healthy control subjects (P 5 0.002) and of patients with other neurological diseases (P 5 0.05), and fromthat of PP-MS patients (P , 0.001).
Reactivity to MBP and MBP peptidesThe responses of PBMC from 61 multiple sclerosis patients,56 healthy control subjects and 52 patients with otherneurological diseases to 19 overlapping 20-amino acid-residue peptides of MBP were also tested in proliferationassays. There were no significant differences between thepercentages of multiple sclerosis patients and healthy controlsubjects responding with SIù 3.0 to the MBP peptides,except for MBP11–30, which elicited a response in asignificantly higher percentage of healthy control subjectsthan of multiple sclerosis patients (Table 6). PBMC frommultiple sclerosis patients responded most frequently topeptides 5b, MBP82–100, MBP91–110and MBP101–120; however,similar percentages of PBMC from healthy control subjectsresponded to these peptides. In the multiple sclerosis group,83%, 85%, 85% and 100%, respectively of patientsresponding to peptides 5b, MBP82–100, MBP91–110 andMBP101–120expressed at least one HLA-Dw2 allele. However,in the control group, only 36%, 33%, 47% and 42%,respectively of responders to peptides 5b, MBP82–100, MBP91–
110and MBP101–120expressed HLA-Dw2 alleles. Interestingly,Fig. 4 Relationship between responsiveness of PBMC toPLP184–199and/or PLP190–209and duration of relapsing–remitting the percentages of PBMC from patients with other neuro-or secondary progressive multiple sclerosis (RR/SP-MS) and logical diseases responding to peptides 5b and MBP82–100EDSS score (filled circles). The number of individuals for each were significantly reduced compared with those of healthyduration was: 0–5 years,n 5 11; 6–10 years,n 5 9; 11–15 years,
control subjects and multiple sclerosis patients (Table 6).n 5 10; 16–20 years,n 5 4; .20 years,n 5 10.These differences could not be explained by the predominanceof males in the group of patients with other neurologicaldiseases. There was no obvious relationship betweenpatient 35 responded to whole PLP and PLP184–199, in additionresponses to peptide 5b and the duration of multiple sclerosis;to PLP190–209. As predicted from previous experience withhowever, the highest percentage of responders to MBP82–100PLP (as described in the Introduction), whole PLP waswas in the group with disease duration ofø 5 years (Fig.unable to induce responses of the same magnitude as the5). Analysis of the responses of the primary progressivepeptides, even when the T-cell line was generated againstmultiple sclerosis, relapsing–remitting multiple sclerosis andthe whole molecule. Nevertheless, there were significantsecondary progressive multiple sclerosis subgroups did notresponses to whole PLP by at least one peptide-specific linereveal any differences in reactivity to the MBP peptidesfrom each of the three multiple sclerosis patients, indicatingamong the groups. The percentage of multiple sclerosisthat whole PLP can be naturally processed into epitopespatients reacting to whole MBP was intermediate betweenwithin the 184–209 region of the molecule. The resultsthe percentages of healthy control subjects and patients withsuggest that there are at least two dominant naturallyother neurological diseases, but the differences were notprocessed epitopes of PLP, one in the overlapping 190–199
region and one present only in PLP184–199. significant (Table 6). The mean SI values for the MBP

1454 J. M. Greeret al.
Table 4 Mean stimulation index (SI6 SE) of PBMC from patients with relapsing–remitting or secondary progressivemultiple sclerosis (RR/SP-MS) and primary progressive multiple sclerosis (PP-MS) of different disease durations
Peptide HC OND RR/SP-MS duration (years) PP-MS duration (years)(n 554) (n 548)
0–5 6–10 11–15 16–20 .20 0–5 6–11(n 511) (n 5 9) (n 5 10) (n 5 4) (n 5 10) (n 5 5) (n 5 6)
PLP184–199 1.7 6 0.2 2.26 0.5 1.76 0.3 2.86 0.6* 7.5 6 2.7*** 2.1 6 0.7 1.86 0.4 1.96 0.8 1.66 0.5PLP190–209 1.4 6 0.1 1.76 0.2 1.56 0.3 2.26 0.5** 3.9 6 0.7*** 2.2 6 0.7* 1.5 6 0.5 1.16 0.2 1.46 0.1
When ANOVA was used to compare healthy control (HC) subjects, patients with other neurological diseases (OND), the five groups ofRR/SP-MS patients and the two groups of PP-MS patients, thenP , 0.000035 for each peptide.*P , 0.048 when compared with HCsubjects (in Student’st test), but not significant compared with OND patients.** P 5 0.003 when compared with HC subjects, but notsignificant compared with OND patients.*** P , 0.002 compared with both HC subjects and OND patients.
Table 5 Reactivity of short-term T-cell lines to whole myelin proteolipid protein (PLP), PLP184–199 or PLP190–209
Patient SI in initial proliferation Antigen Reactivity of the line (c.p.m.) accordingHLA haplotype assay on PBMC to antigen used to generate line
DRB1 DQA1 DQB1 PLP184–199 PLP190–209 PLP PLP184–199 PLP190–209
MS22 0401, 1501 0102, 0301 0301, 0602 4.9 1.7 – 15296 1121 14926 435 17616 630PLP 44936 1343 66266 2989* 21876 596PLP184–199 50716 426* 46,7426 4994* 31436 1054PLP190–209 50576 621* 18,9406 2578* 69406 540*
MS35 0701, 1501 0102, 0201 0201, 0602 3.1 8.2 – 3356 40 5556 487 3826 34PLP 5536 35 33136 774* 18846 390*PLP184–199 460 6 36 24986 461* 26906 786*PLP190–209 617 6 58 44306 612* 35,8316 958*
MS45 0401, 1101 0301, 0501 0301, 0301 3.5 5.8 – 18846 953 5216 134 6036 100PLP 38406 1121 16286 329* 8306 67PLP184–199 10,9116 2402* 11,3626 1568* 12856 143PLP190–209 63466 1453* 24376 170* 70146 396*
*Positive responses.
peptides and MBP (Table 7) showed the same pattern of peptides and multiple sclerosis. However, the differences inresponses of multiple sclerosis patients and patients withreactivity as that indicated by theχ2 analysis.other neurological diseases to MBP peptides do raise somequestions regarding regulation of responses to MBP, whichwill be discussed below.Discussion
This study indicates that there is increased reactivity againstPLP184–199 and PLP190–209 in relapsing–remitting multiplesclerosis and secondary-progressive multiple sclerosisAutoreactivity to PLP
Three previous studies have investigated proliferativepatients, but not primary-progressive multiple sclerosispatients, compared with healthy control subjects and patients responses of PLP-specific lymphocytes in the blood of
multiple sclerosis patients. Two of these studies reported nowith other neurological diseases (Fig. 3). Although stimula-tion assays with synthetic peptides can potentially introduce differences between multiple sclerosis patients and control
subjects (Johnsonet al., 1986; Kerlero de Rosboet al., 1993),a bias for cryptic epitopes, we have shown that PLP184–199-specific T cells and PLP190–209-specific T cells can respond whereas Trotteret al. (1991) showed a difference between
the responses of chronic progressive (primary progressive1to whole PLP and vice versa, indicating that these epitopesof PLP can be processed naturally from whole PLP by antigen- secondary progressive) multiple sclerosis patients and healthy
control subjects to whole PLP. None of these studiespresenting cells of multiple sclerosis patients. Autoimmuneresponses to epitopes within the same region of PLP (residues investigated the response to peptides within the 180–210
region of the molecule. In the two studies (Johnsonet al.,178–209) mediate experimental autoimmune encephalo-myelitis (EAE) in a variety of mice of different genetic 1986; Kerlero de Rosboet al., 1993) that found no differences
between multiple sclerosis patients and control subjects,backgrounds (Greeret al., 1992, 1996a). We did not find acorrelation between PBMC proliferative responses to MBP bovine PLP was used, whereas in the study of Trotteret al.

Immunoreactivity to PLP in multiple sclerosis 1455
Table 6 Percentage of subjects responding to myelin basic protein (MBP) and MBP peptides
MBP peptide Sequence Percentage with SIù 3.0
No. According to human 18.5-kDa isoform HC MS OND
1 1–20 ASQKRPSQRHGSKYLATAST 7.0 3.3 02 11–30 GSKYLATASTMDHARHGFLP 26.3* 3.3 3.93 21–40 MDHARHGFLPRHRDTGILDS 12.3 4.9 7.74 31–50 RHRDTGILDSIGRFFGGDRG 14.0 3.3 3.95 41–60 IGRFFGGDRGAPKRGSGKDS 19.3 13.1 9.65b (49–58)1 (exon 2, 1–10) RGAPKRGSGKVPWLKPGRSP 17.5 19.7 1.9**
5c (exon 2, 1–20) VPWLKPGRSPLPSHARSQPG 12.3 8.2 7.76 51–70 APKRGSGKDSHHPARTAHYG 12.3 9.8 7.77 61–80 HHPARTAHYGSLPQKSHGRT 21.1 11.5 11.58 71–90 SLPQKSHGRTQDENPVVHFF 8.8 3.3 5.89 82–100 DENPVVHFFKNIVTPRTPP 24.6 23.0 5.8**
10 91–110 KNIVTPRTPPPSQGKGRGLS 24.6 18.0 9.610b (96–105)1 (117–126) PRTPPPSQGKGAEGQRPGFG 5.3 8.2 1.911 101–120 PSQGKGRGLSLSRFSWGAEG 14.0 19.7 9.612 111–130 LSRFSWGAEGQRPGFGYGGR 15.8 16.4 13.513 122–140 RPGFGYGGRASDYKSAHKG 8.8 3.3 7.714 131–150 ASDYKSAHKGFKGVDAQGTL 15.8 6.6 9.615 141–160 FKGVDAQGTLSKIFKLGGRD 21.1 13.1 7.716 151–170 SKIFKLGGRDSRSGSPMARR 8.8 11.5 7.7MBP – 35.1 24.6 17.3
The 332 χ2 analysis was significant (P , 0.05) for peptides 2, 4, 5b, and 9, but not for the other peptides.*Percentage of healthycontrol subjects (HC) responding to peptide is significantly different from the percentages of both multiple sclerosis patients (MS)(P , 0.001,χ2) and patients with other neurological diseases (OND) (P 5 0.002) responding to the same peptide.** Percentage of ONDpatients responding to peptide is significantly different from the percentages of both MS patients and healthy control subjects respondingto the same peptide (P , 0.02)
however, differs from human PLP at residues 88, 188 and198 (Laursenet al., 1984). These differences could affectthe ability of human T cells to respond to bovine PLP,particularly since two of these differences occur in the regionof the molecule that we have identified in the present studyas being a target of autoreactive cells in multiple sclerosis.It is known that one of the murine encephalitogenic epitopeswithin the 180–209 region, PLP178–191, is encephalitogenicin SJL mice only when synthesized according to the murinesequence; it is not encephalitogenic when synthesizedaccording to the bovine sequence (Greeret al., 1997). Inaddition, in this murine model, mice immunized with murinePLP or the murine peptide respond only to murine PLP/peptide in proliferation assays; there is no cross-reactivitywith the bovine peptide. Thus, if a similar situation occursin humans, responses to epitopes in this region could bemissed if bovine PLP is used as the stimulating antigen.
Interestingly, Trotteret al.(1991) found that the percentageof patients with early (disease duration ofø 2 years) andlow-disability (EDSS , 3.0) relapsing–remitting multiple
Fig. 5 Relationship between responsiveness to MBP peptide 5bsclerosis reacting to PLP was lower than that of patientsand MBP82–100and duration of multiple sclerosis. The number ofwith more severe progressive disease. This is in agreementindividuals for each duration was: 0–5 years,n 5 16; 6–10 years,
n 5 18; 11–15 years,n 5 14; .16 years,n 5 13. with the present study, in which we found that the greatestreactivity to PLP184–199 and/or PLP190–209 occurred inrelapsing–remitting multiple sclerosis and secondary(1991) human PLP was used. PLP is a highly conservedprogressive multiple sclerosis patients with a disease durationmolecule; there is, 1% difference in PLP among mammalianof 6–15 years. The low frequency of reactivity to PLPspecies, and murine and human PLP are completely identical
(Macklin et al., 1987; Hudsonet al., 1989). Bovine PLP, peptides in patients with a short disease duration may indicate

1456 J. M. Greeret al.
Table 7 Mean SI6 SE of PBMC of healthy controls, multiple sclerosis patients and patients with other neurologicaldiseases for MBP and MBP peptides
MBP peptide Mean SI6 SE P-value(ANOVA)
No. According to human 18.5 kDa isoform HC MS OND
1 1–20 1.56 0.2* 1.2 6 0.1 1.16 0.1 0.0412 11–30 2.16 0.3* 1.3 6 0.2 1.36 0.2 0.0083 21–40 1.66 0.2 1.96 0.7 1.36 0.1 0.5634 31–50 1.66 0.2 1.26 0.1 1.36 0.1 0.0985 41–60 2.26 0.3* 1.5 6 0.2 1.46 0.2 0.0195b (49–58)1 (exon 2, 1–10) 1.86 0.2 1.86 0.2 1.16 0.1** 0.0055c (exon 2, 1–20) 1.66 0.2 1.96 0.4 1.86 0.6 0.9266 51–70 1.76 0.3 1.36 0.2 1.36 0.1 0.1507 61–80 2.16 0.3 1.66 0.3 1.66 0.2 0.2578 71–90 1.36 0.2 1.36 0.1 1.36 0.1 0.9089 82–100 2.76 0.4 2.16 0.2 1.56 0.2** 0.029
10 91–110 2.66 0.4 2.56 0.7 2.16 0.7 0.83110b (96–105)1 (117–126) 1.26 0.2 1.46 0.3 0.96 0.1 0.23611 101–120 2.36 0.7 2.26 0.4 1.66 0.3 0.57512 111–130 2.06 0.4 1.86 0.2 1.66 0.2 0.54413 122–140 1.26 0.1 1.46 0.5 1.36 0.3 0.82614 131–150 1.66 0.2 1.46 0.3 1.36 0.1 0.44415 141–160 2.76 0.5* 1.8 6 0.2 1.66 0.3 0.04816 151–170 1.66 0.2 1.56 0.2 1.36 0.1 0.368
MBP 3.0 6 0.4 2.26 0.2 2.36 0.3 0.118
*Mean SI of healthy control (HC) subjects is significantly different from mean SI of multiple sclerosis (MS) patients (Student’st test;P , 0.049) and patients with other neurological diseases (OND) (P , 0.021). **Mean SI of OND patients is significantly different frommean SI of MS patients (P , 0.04) and HC subjects (P , 0.002).
that the 184–209 region of PLP is not the target of the initial DRB4 (DR53), but not DR3 (Markovic-Pleseet al., 1995).Interestingly, Ito et al. (1996) have recently shown thatautoimmune attack in most cases of multiple sclerosis.
Reactivity to this region of PLP might develop only after transgenic mice expressing the human HLA-DR4 moleculeare susceptible to induction of EAE with PLP175–192, ansubstantial myelin damage has occurred. Alternatively, the
frequency of specific lymphocytes in the periphery may have epitope overlapping PLP184–199. Markovic-Plese et al.(1995) also showed that PLP180–199 is a dominant regionbeen low if the disease was quiescent when PBMC were
collected. recognized by human T cells (control and multiple sclerosis)that have been stimulated several timesin vitro with PLPThe association between multiple sclerosis and the expres-
sion of HLA-DRB1*1501, -DQA1*0102 and -DQB1*0602 (Markovic-Pleseet al., 1995). We have also found that whole-PLP-stimulated T-cell lines can recognize both PLP184–199(Dw2) alleles (Hillertet al., 1994) suggests that the target
antigen(s) in multiple sclerosis may have a high affinity for and PLP190–209and, in addition, that T-cell lines specific forthese peptides can recognize the whole PLP molecule (Tablethese molecules, in the same way that certain self peptides
preferentially bind to HLA molecules associated with the 5). The specific epitopes recognized will depend largely onthe particular HLA genotype of the individual. These resultsautoimmune disease pemphigus vulgaris (Wucherpfennig
et al., 1995). The majority of relapsing–remitting multiple clearly demonstrate that the intact PLP molecule can benaturally processed into epitopes contained within the PLP184–sclerosis and secondary progressive multiple sclerosis patients
reacting to PLP184–199or PLP190–209expressed these multiple 199 or PLP190–209 peptides. These features of the immuneresponse to PLP may allow susceptible individuals to developsclerosis-associated alleles. However, our results suggest that
the development of reactivity to PLP184–199 and/or PLP190– increased reactivity to PLP, once myelin breakdown hasoccurred.209 may be related to the presence of other alleles in
the genotype, in particular DRB1*0401, DRB1*0404, or As already mentioned, PBMC from most of the primaryprogressive multiple sclerosis patients did not respond toDRB1*0701 (or other alleles in linkage disequilibrium with
them). The most common genotype encountered in the group PLP184–199or PLP190–209. Several studies have suggested thatprimary progressive multiple sclerosis differs fromof multiple sclerosis patients tested in this study, HLA-
DRB1*0301,1501, did not correlate with reactivity to the relapsing–remitting and secondary progressive multiplesclerosis (Confavreuxet al., 1980; Verjanset al., 1983;two peptides. Previous binding assays have shown that
PLP180–199 and PLP190–209 bind with relatively high affinity Larsenet al., 1985; Poseret al., 1986; Olerupet al., 1989;Thompsonet al., 1991; Reveszet al., 1994). Both histologicalto a variety of murine (Greeret al., 1996a) and human class
II MHC molecules, including HLA-DRB1*1501, DR4 and (Reveszet al., 1994) and MRI (Thompsonet al., 1991)

Immunoreactivity to PLP in multiple sclerosis 1457
findings indicate that there is less inflammation in primary that multiple sclerosis might result from an inability to deleteautoreactive T cells in the CNS (Tabiet al., 1995). Oneprogressive multiple sclerosis than in the other forms of
multiple sclerosis. It has also been suggested that expression possible explanation for our findings is that, in healthy controlsubjects, these MBP-reactive cells make a primary responseof certain HLA genes, in addition to the Dw2 susceptibility
alleles, may predispose towards primary progressive multiple to the peptidesin vitro; in patients with other neurologicaldiseases, in whom some damage to the CNS or PNS hassclerosis (Olerupet al., 1989), although this evidence is
inconclusive. Furthermore, primary-progressive multiple occurred, the T cells have already encountered the MBPpeptidesin vivoand have been deleted in the nervous system,sclerosis and relapsing–remitting/secondary progressive
multiple sclerosis also differ in the age of onset (Confavreux as occurs during spontaneous recovery from EAE (Tabiet al.,1994, 1995); whereas, in multiple sclerosis patients, these Tet al., 1980; Verjans et al., 1983), initial symptoms
(Confavreuxet al., 1980; Larsenet al., 1985) and prognosis cells have not been deleted, even after repeated exposure toantigen in vivo. The same pattern of responses was notwith regard to disability (Confavreuxet al., 1980; Verjans
et al., 1983) and mortality (Poseret al., 1986). Our present observed with all PLP peptides. However, this may be dueto the absence of PLP in PNS myelin, and a consequent lackresults support the notion that primary progressive multiple
sclerosis differs from relapsing–remitting multiple sclerosis of deletion of PLP-reactive cells in patients with otherneurological diseases of the PNS. Further study of theand secondary progressive multiple sclerosis.activation status of the responding cells from healthy controlsubjects, multiple sclerosis patients, and patients with otherneurological diseases to certain MBP and PLP peptides willAutoreactivity to MBP
Some previous studies of the PBMC proliferative responses be required to resolve this issue. The lack of increasedreactivity to MBP peptides in patients with multiple sclerosisto MBP or MBP fragments have shown significantly higher
reactivity to MBP in multiple sclerosis patients than in normal might have been anticipated from the fact that EAE inducedby immunization with MBP (or whole CNS tissue), orcontrol subjects or patients with other neurological diseases
(Lisak and Zweiman, 1977; Brinkmanet al., 1982; Baxevanis by the passive transfer of MBP-sensitized lymphocytes, ischaracterized by major involvement of the proximal PNSet al., 1989; Vandenbarket al., 1989), but other studies have
not (Johnsonet al., 1986; Kerlero de Rosboet al., 1993). (Pender and Sears, 1982, 1984; Pender, 1988a, b; Penderet al., 1989, 1995; Chalket al., 1994a, b; Abromson-LeemanZhanget al. (1994) have reported that there is an increased
frequency of IL-2-responsive T cells specific for MBP in et al., 1995), which does not occur in typical multiplesclerosis.peripheral blood and CSF of multiple sclerosis patients. In
the present study, the proportion of multiple sclerosis patientsreacting to whole MBP was intermediate between the propor-tions of healthy control subjects and patients with other
Conclusionsneurological diseases, but the differences were not significant.In the present study we have demonstrated increased T-cellIn the present study, the peptides that were recognizedreactivity to two overlapping PLP peptides in patients withmost often by PBMC from multiple sclerosis patients wererelapsing–remitting or secondary progressive multiple scler-MBP82–100, MBP91–110, MBP101–120 and peptide 5b whichosis, but not primary progressive multiple sclerosis. Both ofcovers a region expressed in the 21.5 and 20.2 kDa isoformsthe peptides can be processed naturally from the intact PLPof MBP. Previously, we (Penderet al., 1996) and othersmolecule. The region of PLP that is the target of this(Martin et al., 1990; Otaet al., 1990; Zhanget al., 1992)autoreactivity is encephalitogenic in a wide variety of micehave found that MBP82–100 (which is the same sequence asof different genetic backgrounds. It is also a region in whichthe peptide numbered MBP84–102by Ota et al., 1990) is anspecies differences in the PLP sequence occur, and this mayimmunodominant region, as T cells stimulated with wholeaccount for the failure of two previous studies, which usedMBP frequently react with peptides in this region. Valliet al.bovine rather than human PLP, to identify PLP as a target(1993) showed that MBP82–100 is promiscuous in its abilityof autoreactive cells in the blood of multiple sclerosis patients.to bind with high affinity to many different HLA-DRThe strongest and most frequent reactivity to these PLPmolecules. In contrast, they found that MBP101–120binds withpeptides occurred in those relapsing–remitting multiple scler-only low affinity to several HLA-DR molecules, includingosis and secondary progressive multiple sclerosis patientsHLA-DRB1*1501. Not all individuals expressing HLA-Dw2with multiple sclerosis of 6–15 years duration and withalleles reacted to MBP82–100, MBP91–110, MBP101–120 andmoderate to severe disability. In contrast, multiple sclerosispeptide 5b but, of those multiple sclerosis patients who didpatients did not react more frequently to any of the MBPreact, nearly all expressed Dw2.peptides than did healthy control subjects, suggesting that inSimilar proportions of healthy control subjects and multiplethe majority of cases of multiple sclerosis, autoreactivity tosclerosis patients had PBMC that recognized MBP82–100,MBP may not play a significant role in the disease process.MBP91–110, MBP101–120 and peptide 5b, but the proportionsIt remains to be determined whether T-cell responses to theseof patients with other neurological diseases responding to
these peptides were considerably lower. We have suggested PLP peptides play a pathogenic role in multiple sclerosis.

1458 J. M. Greeret al.
The HLA-Dw2 haplotype segregates closely with multiple sclerosisAcknowledgementsin multiplex families. J Neuroimmunol 1994; 50: 95–100.We would like to thank Dr B. Bennetts and Dr E. Jazwinska
for the HLA-typing.This work was supported by the National Hudson LD, Puckett C, Berndt J, Chan J, Gencic S. Mutation ofMultiple Sclerosis Society of Australia (Elizabeth Albiez the proteolipid protein gene PLP in a human X chromosome-linked
myelin disorder. Proc Natl Acad Sci USA 1989; 86: 8128–31.Fellowship to J.M.G.) and by the National Health and MedicalResearch Council of Australia. Ito K, Bian H-J, Molina M, Han J, Magram J, Saar E, et al. HLA-
DR4-IE chimeric class II transgenic, murine class II-deficient miceare susceptible to experimental allergic encephalomyelitis. J Exp
References Med 1996; 183: 2635–44.Abromson-Leeman S, Bronson R, Dorf ME. Experimental
Johnson D, Hafler DA, Fallis RJ, Lees MB, Brady RO, Quarles RH,autoimmune peripheral neuritis induced in BALB/c mice by myelinet al. Cell-mediated immunity to myelin-associated glycoprotein,basic protein-specific T cell clones. J Exp Med 1995; 182: 587–92.proteolipid protein, and myelin basic protein in multiple sclerosis.
Baxevanis CN, Reclos GJ, Servis C, Anastasopoulos E, Arsenis P,J Neuroimmunol 1986; 13: 99–108.Katsiyiannis A, et al. Peptides of myelin basic protein stimulate T
Kerlero de Rosbo N, Milo R, Lees MB, Burger D, Bernard CC,lymphocytes from patients with multiple sclerosis. J NeuroimmunolBen Nun A. Reactivity to myelin antigens in multiple sclerosis.1989; 22: 23–30.Peripheral blood lymphocytes respond predominantly to myelin
Brinkman CJ, Nillesen WM, Hommes OR, Lamers KJ, de Pauwoligodendrocyte glycoprotein. J Clin Invest 1993; 92: 2602–8.BE, Delmotte P. Cell-mediated immunity in multiple sclerosis as
Larsen JP, Kvaale G, Riise T, Nyland H, Aarli JA. Multipledetermined by sensitivity of different lymphocyte populations tosclerosis – more than one disease? Acta Neurol Scand 1985; 72:various brain tissue antigens. Ann Neurol 1982; 11: 450–5.145–50.
Chalk JB, McCombe PA, Pender MP. Conduction abnormalities areLaursen RA, Samiullah M, Lees MB. The structure of bovine brainrestricted to the central nervous system in experimental autoimmunemyelin proteolipid and its organization in myelin. Proc Natl Acadencephalomyelitis induced by inoculation with proteolipid proteinSci USA 1984; 81: 2912–6.but not with myelin basic protein. Brain 1994a; 117: 975–86.
Chalk JB, McCombe PA, Smith R, Pender MP. Clinical and Lisak RP, Zweiman B. In vitro cell-mediated immunity ofhistological findings in proteolipid protein-induced experimental cerebrospinal-fluid lymphocytes to myelin basic protein in primaryautoimmune encephalomyelitis (EAE) in the Lewis rat. Distribution demyelinating diseases. N Engl J Med 1977; 297: 850–3.of demyelination differs from that in EAE induced by other antigens.
Lumsden CE. The neuropathology of multiple sclerosis. In: VinkenJ Neurol Sci 1994b; 123: 154–61.PJ, Bruyn GW, editors. Handbook of clinical neurology, Vol. 9.
Confavreux C, Aimard G, Devic M. Course and prognosis of Amsterdam: North-Holland, 1970: 217–309.multiple sclerosis assessed by the computerized data processing of
Macklin WB, Campagnoni CW, Deininger PL, Gardinier MV.349 patients. Brain 1980; 103: 281–300.Structure and expression of the mouse myelin proteolipid protein
Correale J, Gilmore W, McMillan M, Li S, McCarthy K, Le T, gene. J Neurosci Res 1987; 18: 383–94.et al. Patterns of cytokine secretion by autoreactive proteolipid
Markovic-Plese S, Fukaura H, Zhang J, al-Sabbagh A, Southwoodprotein-specific T cell clones during the course of multiple sclerosis.S, Sette A, et al. T cell recognition of immunodominant and crypticJ Immunol 1995; 154: 2959–68.proteolipid protein epitopes in humans. J Immunol 1995; 155:
Deibler GE, Martenson RE, Kies MW. Large scale preparation of982–92.myelin basic protein from central nervous tissue of several
Martin R, Jaraquemada D, Flerlage M, Richert J, Whitaker J, Longmammalian species. Prep Biochem 1972; 2: 139–65.EO, et al. Fine specificity and HLA restriction of myelin basic
Greer JM, Kuchroo VK, Sobel RA, Lees MB. Identification and protein-specific cytotoxic T cell lines from multiple sclerosis patientscharacterization of a second encephalitogenic determinant of myelinand healthy individuals. J Immunol 1990; 145: 540–8.proteolipid protein (residues 178–191) for SJL mice. J Immunol
Meinl E, Weber F, Drexler K, Morelle C, Ott M, Saruhan-Direskeneli1992; 149: 783–8.G, et al. Myelin basic protein-specific T lymphocyte repertoire in
Greer JM, Sobel RA, Sette A, Southwood S, Lees MB, Kuchroomultiple sclerosis: complexity of the response and dominance ofVK. Immunogenic and encephalitogenic epitope clusters of myelinnested epitopes due to recruitment of multiple T cell clones. J Clinproteolipid protein. J Immunol 1996a; 156: 371–79. Invest 1993; 92: 2633–43.Greer JM, Dyer CA, Pakaski M, Symonowicz C, Lees MB. Ohashi T, Yamamura T, Inobe J-I, Kondo T, Kunishita T, Tabira T.Orientation of myelin proteolipid protein in the oligodendrocyte Analysis of proteolipid protein (PLP)-specific T cells in multiplecell membrane. Neurochem Res 1996b; 21: 431–40. sclerosis: identification of PLP 95–116 as an HLA-DR2,w15-
associated determinant. Int Immunol 1995; 7: 1771–8.Greer JM, Klinguer C, Trifilieff E, Sobel RA, Lees MB.Encephalitogenicity of murine, but not bovine, DM20 in SJL mice
Olerup O, Hillert J, Fredrikson S, Olsson T, Kam-Hansen S, Molleris due to a single amino acid difference in the immunodominant
E, et al. Primarily chronic progressive and relapsing/remittingencephalitogenic epitope. Neurochem Res 1997; 22: 541–7.
multiple sclerosis: two immunogenetically distinct disease entities.Proc Natl Acad Sci USA 1989; 86: 7113–7.Hillert J, Kall T, Vrethem M, Fredrikson S, Ohlson M, Olerup O.

Immunoreactivity to PLP in multiple sclerosis 1459
Ota K, Matsui M, Milford EL, Mackin GA, Weiner HL, Hafler DA. comparison of the pathology of primary and secondary progressivemultiple sclerosis. Brain 1994;117: 759–65.T-cell recognition of an immunodominant myelin basic protein
epitope in multiple sclerosis. Nature 1990; 346: 183–7.Roth HJ, Kronquist KE, Kerlero de Rosbo N, Crandall BF,Campagnoni AT. Evidence for the expression of four myelin basicPelfrey CM, Trotter JL, Tranquill LR, McFarland HF. Identificationprotein variants in the developing human spinal cord through cDNAof a novel T cell epitope of human proteolipid protein (residuescloning. J Neurosci Res 1987; 17: 321–8.40–60) recognized by proliferative and cytolytic CD41 T cells
from multiple sclerosis patients. J Neuroimmunol 1993; 46: 33–42.Sobel RA. The pathology of multiple sclerosis. [Review]. NeurolClin 1995; 13: 1–21.Pelfrey CM, Trotter JL, Tranquill LR, McFarland HF. Identification
of a second T cell epitope of human proteolipid protein (residuesStewart GJ, McLeod JG, Basten A, Bashir HV. HLA family studies89–106) recognized by proliferative and cytolytic CD41 T cells and multiple sclerosis: a common gene, dominantly expressed. Humfrom multiple sclerosis patients. J Neuroimmunol 1994; 53: 153–61.Immunol 1981; 3: 13–29.
Pender MP. The pathophysiology of myelin basic protein-inducedTabi Z, McCombe PA, Pender MP. Apoptotic elimination of Vβ8.21
acute experimental allergic encephalomyelitis in the Lewis rat. Jcells from the central nervous system during recovery fromNeurol Sci 1988a; 86: 277–89. experimental autoimmune encephalomyelitis induced by the passive
transfer of Vβ8.21 encephalitogenic T cells. Eur J Immunol 1994;Pender MP. Demyelination of the peripheral nervous system causes24: 2609–17.neurologic signs in myelin basic protein-induced experimental
allergic encephalomyelitis. Implications for the etiology of multiple Tabi Z, McCombe PA, Pender MP. Antigen-specific down-regulationsclerosis. Ann NY Acad Sci 1988b; 540: 732–4. of myelin basic protein-reactive T cells during spontaneous recovery
from experimental autoimmune encephalomyelitis: further evidencePender MP. Multiple sclerosis. In: Pender MP, McCombe PA,
of apoptotic deletion of autoreactive T cells in the central nervouseditors. Autoimmune neurological disease. Cambridge: Cambridge
system. Int Immunol 1995; 7: 967–73.University Press, 1995: 89–154.
Thompson AJ, Kermode AG, Wicks D, MacManus DG, KendallPender MP, Sears TA. Conduction block in the peripheral nervousBE, Kingsley DP, et al. Major differences in the dynamics ofsystem in experimental allergic encephalomyelitis. Nature 1982;primary and secondary progressive multiple sclerosis. Ann Neurol296: 860–2. 1991; 29: 53–62.
Pender MP, Sears TA. The pathophysiology of acute experimentalTrotter JL, Hickey WF, van der Veen RC, Sulze L. Peripheral bloodallergic encephalomyelitis in the rabbit. Brain 1984; 107: 699–726.mononuclear cells from multiple sclerosis patients recognize myelin
proteolipid protein and selected peptides. J Neuroimmunol 1991;Pender MP, Nguyen KB, Willenborg DO. Demyelination and early33: 55–62.remyelination in experimental allergic encephalomyelitis passively
transferred with myelin basic protein-sensitized lymphocytes in theTuohy VK, Sobel RA, Lees MB. Myelin proteolipid protein-inducedLewis rat. J Neuroimmunol 1989; 25: 125–42. experimental allergic encephalomyelitis: variations of disease
expression in different strains of mice. J Immunol 1988; 140:Pender MP, Tabi Z, Nguyen KB, McCombe PA. The proximal1868–73.peripheral nervous system is a major site of demyelination in
experimental autoimmune encephalomyelitis induced in the LewisTuohy VK, Lu Z, Sobel RA, Laursen RA, Lees MB. Identificationrat by a myelin basic protein-specific T cell clone. Acta Neuropatholof an encephalitogenic determinant of myelin proteolipid protein(Berl) 1995; 89: 527–31. for SJL mice. J Immunol 1989; 142: 1523–7.
Pender MP, Csurhes PA, Houghten RA, McCombe PA, Good MF.Valli A, Sette A, Kappos L, Oseroff C, Sidney J, Miescher G, et al.A study of human T-cell lines generated from multiple sclerosisBinding of myelin basic protein peptides to human histocompatibilitypatients and controls by stimulation with peptides of myelin basicleukocyte antigen class II molecules and their recognition by Tprotein. J Neuroimmunol 1996; 70: 65–74. cells from multiple sclerosis patients. J Clin Invest 1993; 91: 616–28.
Vandenbark AA, Chou YK, Bourdette D, Whitham R, Chilgren J,Poser CM, Paty DW, Scheinberg L, McDonald WI, Davis FA, EbersChou CH, et al. Human T lymphocyte response to myelin basicGC, et al. New diagnostic criteria for multiple sclerosis: guidelinesprotein: selection of T lymphocyte lines from MBP-responsivefor research protocols. Ann Neurol 1983; 13: 227–31.donors. J Neurosci Res 1989; 23: 21–30.
Poser S, Poser W, Schlaf G, Firnhaber W, Lauer K, Wolter M, et al.Verjans E, Theys SP, Delmotte P, Carton H. Clinical parametersPrognostic indicators in multiple sclerosis. Acta Neurol Scand 1986;and intrathecal IgG synthesis as prognostic features in multiple74: 387–92.sclerosis. Part 1. J Neurol 1983; 229: 155–65.
Puckett C, Hudson L, Ono K, Friedrich V, Benecke J, Dubois-Wucherpfennig KW, Yu B, Bhol K, Monos DS, Argyris E, KarrDalcq M, et al. Myelin-specific proteolipid protein is expressed inRW, et al. Structural basis for major histocompatibility complexmyelinating Schwann cells but is not incorporated into myelin(MHC)-linked susceptibility to autoimmunity: charged residues ofsheaths. J Neurosci Res 1987; 18: 511–8.a single MHC binding pocket confer selective presentation of self-
Rammensee H-G, Friede T, Stevanovic S. MHC ligands and peptidepeptides in pemphigus vulgaris. Proc Natl Acad Sci USA 1995; 92:motifs: first listing. [Review]. Immunogenetics 1995; 41: 178–228. 11935–9.
Zhang J, Medaer R, Hashim GA, Chin Y, van den Berg-Loonen E,Revesz T, Kidd D, Thompson AJ, Barnard RO, McDonald WI. A

1460 J. M. Greeret al.
Raus JC. Myelin basic protein-specific T lymphocytes in multiple Zhang J, Markovic-Plese S, Lacet B, Raus J, Weiner HL, HaflerDA. Increased frequency of interleukin-2-responsive T cells specificsclerosis and controls: precursor frequency, fine specificity, andfor myelin basic protein and proteolipid protein in peripheral bloodcytotoxicity. Ann Neurol 1992; 32: 330–8.and CSF of patients with multiple sclerosis. J Exp Med 1994; 179:
Zhang Y, Burger D, Saruhan G, Jeannet M, Steck AJ. The T-973–84.lymphocyte response against myelin-associated glycoprotein andmyelin basic protein in patients with multiple sclerosis. Neurology1993; 43: 403–7. Received February 13, 1997. Accepted March 13, 1997




















