
Impact of Potassium Bromate and Potassium Iodate in a Pound Cake System

FEBS Letters xxx (2014) xxx–xxx
journal homepage: www.FEBSLetters .org
Cyclic mononucleotides modulate potassium and calcium flux responsesto H2O2 in Arabidopsis roots
http://dx.doi.org/10.1016/j.febslet.2014.01.0620014-5793/� 2014 The Authors. Published by Elsevier B.V.This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
⇑ Corresponding author.E-mail address: [email protected] (C. Gehring).
1 Both authors have contributed equally to the work.
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotides modulate potassium and calcium flux responses to H2O2 in Arabroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
Natalia Maria Ordoñez a,1, Claudius Marondedze a,1, Ludivine Thomas b, Stefania Pasqualini c,Lana Shabala d, Sergey Shabala d, Chris Gehring a,⇑a Biological and Environmental Sciences & Engineering Division, King Abdullah University of Science and Technology, Thuwal 23955-6900, Saudi Arabiab Bioscience Core Facility, King Abdullah University of Science and Technology, Thuwal 23955-6900, Saudi Arabiac Department of Applied Biology, University of Perugia, Borgo XX Giugno, 74, 06121 Perugia, Italyd School of Agricultural Science, University of Tasmania, Private Bag 54, Hobart, Tasmania 7001, Australia
a r t i c l e i n f o a b s t r a c t
Article history:Received 12 December 2013Revised 23 January 2014Accepted 24 January 2014Available online xxxx
Edited by Julian Schroeder
Keywords:Plant stressHydrogen peroxideCyclic mononucleotidecAMPcGMPIon fluxProteomicArabidopsis thaliana
Cyclic mononucleotides are messengers in plant stress responses. Here we show that hydrogen per-oxide (H2O2) induces rapid net K+-efflux and Ca2+-influx in Arabidopsis roots. Pre-treatment witheither 10 lM cAMP or cGMP for 1 or 24 h does significantly reduce net K+-leakage and Ca2+-influx,and in the case of the K+-fluxes, the cell permeant cyclic mononucleotides are more effective. Wealso examined the effect of 10 lM of the cell permeant 8-Br-cGMP on the Arabidopsis microsomalproteome and noted a specific increase in proteins with a role in stress responses and ion transport,suggesting that cGMP is sufficient to directly and/or indirectly induce complex adaptive changes tocellular stresses induced by H2O2.� 2014 The Authors. Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND
license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
1. Introduction
Environmental stresses may cause plants to generate reactiveoxygen species (ROS), such as hydrogen peroxide (H2O2), superox-ide ðO�2 Þ and hydroxyl radical (HO) [1]. ROS, which are highly reac-tive to membrane lipids, proteins, and DNA, are believed to be themajor contributing factors to stress injuries and to cause rapid cel-lular damage [2]. However, when ROS are generated at controlledlevels, cells recognise these reactive molecules as signals to acti-vate defence genes (for review, see [3]). The control of ROS level re-quires cells to rapidly produce and scavenge different forms of ROSin a coordinated manner. Enzymes, including superoxide dismu-tase (SOD), catalase (CAT), peroxidase (POX), ascorbate peroxidase(APX) and glutathione reductase (GR), and non-enzymatic antioxi-dants such as tocopherols, ascorbic acid (AsA), and glutathione(GSH) work in cooperation to the detoxification of ROS. Most ROSseem to exert signalling functions that lead to specific biological
processes. At the transcription level, H2O2-responsive genes wereinitially identified by microarray and AFLP-based technologies[4]. Later signalling properties and distinct transcriptional re-sponses were confirmed also for other ROS, including O�2 [5]. How-ever, H2O2 seems best suited to play a pivotal role as a signallingmolecule due to a comparatively high stability and long half-life.H2O2 generation occurs in plants following exposure to a widevariety of abiotic and biotic stresses. These include temperature,UV irradiation, excess excitation energy, ozone exposure, somephytohormones (e.g. ABA), drought, salt, flooding, heavy metals,wounding and pathogen challenge e.g. [6,7]. In shoots, generationof ROS during salt stress is mainly attributed to declining CO2 fix-ation, leading to higher leakage of electrons to O2. In salt-stressedroots, ROS production is attributed to the disruption of mitochon-drial function and salt-induced activation of the plasma membraneNADPH oxidases [8,9]. Furthermore, a number of studies show di-rect evidence of H2O2 accumulation in planta under salt stress [10]and it was reported that maize (Zea mays) plants pre-treated withH2O2 are capable of acquiring additional salt stress tolerance,suggesting that salt-induced ROS can induce antioxidant responsesat the cell level [11]. However, salt stress triggers not just an
idopsis

2 N.M. Ordoñez et al. / FEBS Letters xxx (2014) xxx–xxx
oxidative burst but also an osmotic stress and an ion imbalance[12]. Na+-influx into the cell results in a dramatic membrane depo-larization, resulting in substantial loss of cytosolic K+ [13]. BecauseK+ is critical for the activity of many enzymes as well as cellular os-motic adjustment, a lack of K+ can cause severe metabolic disorders[12] as well as trigger programmed cell death in salt-stressed tis-sues [14,15]. It has been shown that in plants K+ is transportedacross the plasma membrane in and out of the roots throughpotassium channels or transporters [13]. The inward-rectifyingK+-channels (KIRs) mediate K+-uptake, while the outward-rectify-ing K+-channels (KORs) mediate K+-release [16]. In addition, non-selective cation channels (NSCCs), the major system for Na+ entry,are also involved in K+-efflux and these channels have an impor-tant role in salt resistance in plants [13]. Constitutive NSCCs canbe subdivided according to their voltage dependence [17]. In addi-tion it has been demonstrated that ROS activated NSCCs are in-volved in HR response and in elongation/expansion of plant cells[18], and cyclic nucleotide-gated channels (CNGCs), which play arole in plant pathogen interaction in shoots [19] and in roots fornon-selective cation uptake [20]. Given that H2O2 [10] and cGMP[21,22] are components of stress responses in general and the re-sponse to NaCl and drought stress in particular, we have under-taken, firstly, to study H2O2-dependent ion fluxes in Arabidopsisthaliana roots and determine whether cyclic nucleotides, and espe-cially cGMP, may affect net K+- and Ca2+-fluxes. Secondly, weinvestigated if cGMP induces changes at the proteome level witha view to gain further insight into the possible physiological impli-cations for cyclic nucleotide-dependent changes in ion fluxes.
2. Materials and methods
2.1. Plant material and growth conditions
A. thaliana, ecotype Col-0, seeds were sterilized and distributedover Petri dishes containing sterile Murashige and Skoog (MS)medium, pH 6.2. The plates were place at 4 �C for 2 days for seedstratification. Afterwards, they were placed into a growth roomand positioned vertically to allow roots to grow outside of themedium. Six day old seedlings were transferred into 55 � 15 mmPetri dishes so their roots were immersed in a shallow layer ofthe Basic Salt Medium (BSM) solution (0.1 mM KCl; 0.2 mM CaCl2;pH 6.0 adjusted with Tris base and 2-(N-morpholino) ethanesulfo-nic acid (MES) buffer). For pre-treatment with cyclic nucleotides,appropriate chemical (adenosine 3050- cyclic monophosphate so-dium salt monohydrate (cAMP; 10 lM); guanosine 3050- cyclicmonophosphate sodium salt (cGMP; 10 lM); or the cell permeantanalogues 8-Br-cAMP (10 lM) and 8-Br-cGMP (10 lM)) was addedto the Petri dish for either 1 or 24 h.
2.2. Non-invasive microelectrode ion flux measurements
Net fluxes of K+ and Ca2+ were measured using non-invasivemicroelectrode ion flux (the MIFE) measuring system (ROCU, Uni-versity of Tasmania) and are detailed elsewhere [23] as well as inthe Supplementary file 1.
2.3. A. thaliana cell suspension culture and microsomal proteinextraction
Cells derived from roots of A. thaliana (ecotype Columbia 0) andgrown in 100 mL of Gamborg0s B5 basal salt mixture (Sigma–Al-drich) with 2,4-dichlorophenoxyacetic acid (2,4-D; 1 mg mL�1)and kinetin (0.05 lg mL�1) in 250 mL sterile flask. Cells weregrown in a growth chamber (Innova� 43, New Brunswick ScientificCo., NJ) with shaking at 120 rpm, and subcultured every 10 days.
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotideroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
Light condition and cyclic nucleotide treatments and cell harvest-ing are further detailed in Supplementary file 1.
2.4. One-dimensional gel electrophoresis (1-DE) and in-gel trypsindigestion
Approximately 15 lg of the microsomal protein extract wasreduced in 2 � SDS reducing buffer and electrophoresed in 12%one-dimensional (1D) gel electrophoresis at 150 V for 30 min. Gelstaining, protein band section and in gel trypsin digestion are de-tailed in the Supplementary file 1.
2.5. Protein identification by LTQ-Orbitrap and functional analysis
Digested peptides were resuspended in 5% (v/v) ACN and 0.1%(v/v) formic acid (FA), and analysed with an LTQ Orbitrap massspectrometer (Thermo-Scientific, Bremen, Germany) coupled to ananoLC (Thermo-Scientific). Spectra were submitted to a localMASCOT (Matrix Science, London, UK) server and searched againstA. thaliana in the TAIR database (release 10), with a precursor masstolerance of 10 ppm, a fragment ion mass tolerance of ±0.5 Da,strict trypsin specificity allowing up to one missed cleavage, carb-amidomethyl modification on cysteine as a fixed modification andmethionine oxidation as a variable modification. Protein identifica-tion and data analysis methods are detailed in the Supplementaryfile 1.
2.6. Statistical analysis
All results are presented as mean ± S.E. (n = sample size). Datawere analysed with SPSS 16.0 for Windows (SPSS Inc., Chicago,IL, USA). Significant differences (at P < 0.05) between treatmentswere determined by one-way ANOVA based on Duncan’s multiplerange test and denoted by different lower case letters.
3. Results and discussion
Application of H2O2 to Arabidopsis roots induces a rapid andmassive K+-efflux. Just 1 h of treatment in either 10 lM of cellthe permeant cyclic nucleotide (CN) analogue 8-Br-cAMP or 8-Br-cGMP was effective in significantly (P < 0.05) reducing the extentof this H2O2-induced K+-leak (Fig. 1A). CN-treated roots lose >2-fold less K+ compared to non-treated controls (Fig. 1B). No statisti-cally significant (P < 0.05) differences between the Br-cAMP andBr-cGMP nucleotides were noted (Fig. 1). In the 1 h pre-treatment,membrane-permeable analogues were more efficient in preventingthe K+-efflux (significant at P < 0.05; Fig. 2A). Increasing the rootpre-treatment with cyclic nucleotides from 1 to 24 h has furtherdecreased the magnitude of H2O2-induced K+-efflux (Fig. 2B); ef-fect of timing was significant according to Duncan’s multiple rangetest (data not shown). No significant (P < 0.05) difference betweenpermeant and non-permeant analogues was found in this case(Fig. 2B); however, each CN has significantly (P < 0.05) reducedthe magnitude of H2O2-induced K+-loss compared with non-trea-ted controls (Fig. 2B). Consistent with the notion that salinity stressresults in a rapid elevation of H2O2 level in root tissues [8,9],root pre-treatment with CN reduced NaCl-dependent K+-efflux(Supplementary file 2).
H2O2 application caused a rapid onset of transient Ca2+-influx(Fig. 3A–C). One hour pre-treatment with the cell permeant cyclicnucleotides, 8-Br-cAMP and 8-Br-cGMP, were similarly efficient inreducing both the magnitude of the peak H2O2-induced Ca2+-influx(�3-fold difference compared with non-treated controls; Fig. 3D;significant at P < 0.05). As a result, CN-treated roots accumulated2 to 3-fold less Ca2+ over the first 15 min after stress onset
s modulate potassium and calcium flux responses to H2O2 in Arabidopsis
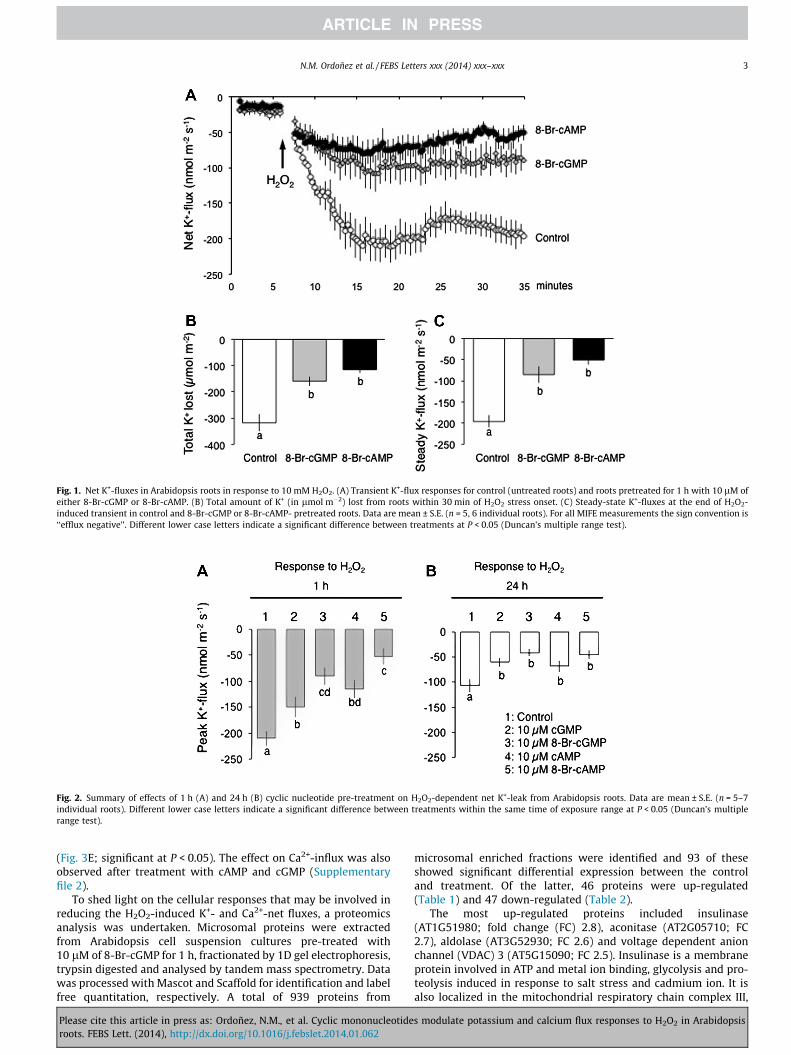
Fig. 1. Net K+-fluxes in Arabidopsis roots in response to 10 mM H2O2. (A) Transient K+-flux responses for control (untreated roots) and roots pretreated for 1 h with 10 lM ofeither 8-Br-cGMP or 8-Br-cAMP. (B) Total amount of K+ (in lmol m�2) lost from roots within 30 min of H2O2 stress onset. (C) Steady-state K+-fluxes at the end of H2O2-induced transient in control and 8-Br-cGMP or 8-Br-cAMP- pretreated roots. Data are mean ± S.E. (n = 5, 6 individual roots). For all MIFE measurements the sign convention is‘‘efflux negative’’. Different lower case letters indicate a significant difference between treatments at P < 0.05 (Duncan’s multiple range test).
Fig. 2. Summary of effects of 1 h (A) and 24 h (B) cyclic nucleotide pre-treatment on H2O2-dependent net K+-leak from Arabidopsis roots. Data are mean ± S.E. (n = 5–7individual roots). Different lower case letters indicate a significant difference between treatments within the same time of exposure range at P < 0.05 (Duncan’s multiplerange test).
N.M. Ordoñez et al. / FEBS Letters xxx (2014) xxx–xxx 3
(Fig. 3E; significant at P < 0.05). The effect on Ca2+-influx was alsoobserved after treatment with cAMP and cGMP (Supplementaryfile 2).
To shed light on the cellular responses that may be involved inreducing the H2O2-induced K+- and Ca2+-net fluxes, a proteomicsanalysis was undertaken. Microsomal proteins were extractedfrom Arabidopsis cell suspension cultures pre-treated with10 lM of 8-Br-cGMP for 1 h, fractionated by 1D gel electrophoresis,trypsin digested and analysed by tandem mass spectrometry. Datawas processed with Mascot and Scaffold for identification and labelfree quantitation, respectively. A total of 939 proteins from
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotideroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
microsomal enriched fractions were identified and 93 of theseshowed significant differential expression between the controland treatment. Of the latter, 46 proteins were up-regulated(Table 1) and 47 down-regulated (Table 2).
The most up-regulated proteins included insulinase(AT1G51980; fold change (FC) 2.8), aconitase (AT2G05710; FC2.7), aldolase (AT3G52930; FC 2.6) and voltage dependent anionchannel (VDAC) 3 (AT5G15090; FC 2.5). Insulinase is a membraneprotein involved in ATP and metal ion binding, glycolysis and pro-teolysis induced in response to salt stress and cadmium ion. It isalso localized in the mitochondrial respiratory chain complex III,
s modulate potassium and calcium flux responses to H2O2 in Arabidopsis
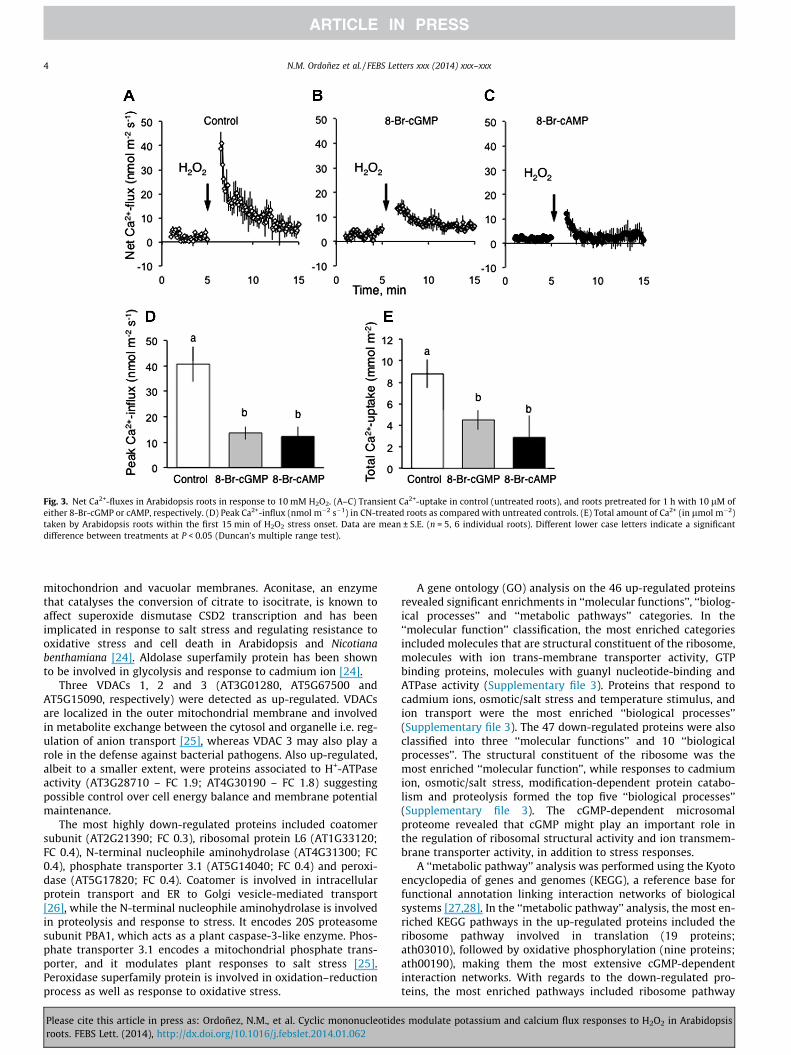
Fig. 3. Net Ca2+-fluxes in Arabidopsis roots in response to 10 mM H2O2. (A–C) Transient Ca2+-uptake in control (untreated roots), and roots pretreated for 1 h with 10 lM ofeither 8-Br-cGMP or cAMP, respectively. (D) Peak Ca2+-influx (nmol m�2 s�1) in CN-treated roots as compared with untreated controls. (E) Total amount of Ca2+ (in lmol m�2)taken by Arabidopsis roots within the first 15 min of H2O2 stress onset. Data are mean ± S.E. (n = 5, 6 individual roots). Different lower case letters indicate a significantdifference between treatments at P < 0.05 (Duncan’s multiple range test).
4 N.M. Ordoñez et al. / FEBS Letters xxx (2014) xxx–xxx
mitochondrion and vacuolar membranes. Aconitase, an enzymethat catalyses the conversion of citrate to isocitrate, is known toaffect superoxide dismutase CSD2 transcription and has beenimplicated in response to salt stress and regulating resistance tooxidative stress and cell death in Arabidopsis and Nicotianabenthamiana [24]. Aldolase superfamily protein has been shownto be involved in glycolysis and response to cadmium ion [24].
Three VDACs 1, 2 and 3 (AT3G01280, AT5G67500 andAT5G15090, respectively) were detected as up-regulated. VDACsare localized in the outer mitochondrial membrane and involvedin metabolite exchange between the cytosol and organelle i.e. reg-ulation of anion transport [25], whereas VDAC 3 may also play arole in the defense against bacterial pathogens. Also up-regulated,albeit to a smaller extent, were proteins associated to H+-ATPaseactivity (AT3G28710 – FC 1.9; AT4G30190 – FC 1.8) suggestingpossible control over cell energy balance and membrane potentialmaintenance.
The most highly down-regulated proteins included coatomersubunit (AT2G21390; FC 0.3), ribosomal protein L6 (AT1G33120;FC 0.4), N-terminal nucleophile aminohydrolase (AT4G31300; FC0.4), phosphate transporter 3.1 (AT5G14040; FC 0.4) and peroxi-dase (AT5G17820; FC 0.4). Coatomer is involved in intracellularprotein transport and ER to Golgi vesicle-mediated transport[26], while the N-terminal nucleophile aminohydrolase is involvedin proteolysis and response to stress. It encodes 20S proteasomesubunit PBA1, which acts as a plant caspase-3-like enzyme. Phos-phate transporter 3.1 encodes a mitochondrial phosphate trans-porter, and it modulates plant responses to salt stress [25].Peroxidase superfamily protein is involved in oxidation–reductionprocess as well as response to oxidative stress.
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotideroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
A gene ontology (GO) analysis on the 46 up-regulated proteinsrevealed significant enrichments in ‘‘molecular functions’’, ‘‘biolog-ical processes’’ and ‘‘metabolic pathways’’ categories. In the‘‘molecular function’’ classification, the most enriched categoriesincluded molecules that are structural constituent of the ribosome,molecules with ion trans-membrane transporter activity, GTPbinding proteins, molecules with guanyl nucleotide-binding andATPase activity (Supplementary file 3). Proteins that respond tocadmium ions, osmotic/salt stress and temperature stimulus, andion transport were the most enriched ‘‘biological processes’’(Supplementary file 3). The 47 down-regulated proteins were alsoclassified into three ‘‘molecular functions’’ and 10 ‘‘biologicalprocesses’’. The structural constituent of the ribosome was themost enriched ‘‘molecular function’’, while responses to cadmiumion, osmotic/salt stress, modification-dependent protein catabo-lism and proteolysis formed the top five ‘‘biological processes’’(Supplementary file 3). The cGMP-dependent microsomalproteome revealed that cGMP might play an important role inthe regulation of ribosomal structural activity and ion transmem-brane transporter activity, in addition to stress responses.
A ‘‘metabolic pathway’’ analysis was performed using the Kyotoencyclopedia of genes and genomes (KEGG), a reference base forfunctional annotation linking interaction networks of biologicalsystems [27,28]. In the ‘‘metabolic pathway’’ analysis, the most en-riched KEGG pathways in the up-regulated proteins included theribosome pathway involved in translation (19 proteins;ath03010), followed by oxidative phosphorylation (nine proteins;ath00190), making them the most extensive cGMP-dependentinteraction networks. With regards to the down-regulated pro-teins, the most enriched pathways included ribosome pathway
s modulate potassium and calcium flux responses to H2O2 in Arabidopsis

Table 1cGMP-responsive up-regulated microsomal proteins.
Accession No. Protein description Fold change GO enrichment
AT1G51980 Insulinase (Peptidase family M16) protein 2.8 OiM, RtOSAT2G05710 Aconitase 3 2.7 RCi, RtOS, CWAT3G52930 Aldolase superfamily protein 2.6 RCi, RtOS, CWAT5G15090 Voltage dependent anion channel 3 2.5 IT, OM, RBSAT3G27240 Cytochrome C1 family 2.4 OP, OiMAT4G02520 Glutathione S-transferase PHI 2 2.4 RCi, RtOS, RTS, RBS, MAT1G04820 Tubulin alpha-4 chain 2.3 RCi, RtOS, CWAT1G77120 Alcohol dehydrogenase 1 2.3 RCi, RtOS, PMAT2G36530 Enolase 2.3 RCi, RtOS, RTS, MAT1G64520 Regulatory particle non-ATPase 12A 2.2 RTS, RBS, M⁄AT2G33040 ATP3 c subunit of Mt ATP synthase 2.2 IT, OP, OiMAT4G13010 Oxidoreductase, zn-binding dehydrogenase 2.1 OiMAT4G20890 Tubulin beta-9 chain 2.1 RCi, Vac M, PMAT4G37910 Mitochondrial heat shock protein 70–1 2.1 RCi, RtOS, CWAT5G40810 Cytochrome C1 family 2.1 OiMAT3G14990 Glutamine amidotransferase-like protein 2 RCi, PM, VacAT1G53240 Lactate/malate dehydrogenase family protein 1.9 RCi, RtOS, RTS, RBS, CWAT1G55490 Chaperonin 60 beta 1.9 RTS, RBS, MAT2G42210 Mitochondrial import translocase 1.9 OiMAT3G02360 6-Phosphogluconate dehydrogenase protein 1.9 RtOS, MAT3G08580 ADP/ATP carrier 1 1.9 OiMAT3G28710 ATPase, V0/A0 complex, subunit C/D 1.9 IT, OP, PMAT3G46430 Unknown protein 1.9 OiMAT4G32470 Cytochrome b ubiquinol oxidase 1.9 mATPet, OP, OiMAT5G16050 General regulatory factor 5 1.9 RCi, CWAT5G62690 Tubulin beta chain 2 1.9 RCi, RtOS, CWAT5G66760 Succinate dehydrogenase 1–1 1.9 mATPet, OP, OiMAT5G67500 Voltage dependent anion channel 2 1.9 IT, OM, RBSAT1G20620 Catalase 3 1.8 RCi, RTS, CWAT2G39460 Ribosomal protein L23AA 1.8 RTS, CWAT4G24190 Chaperone protein htpG family protein 1.8 RCi, RtOS, RTS, M, ERAT4G30190 H+-ATPase 2 1.8 IT, OP, OiMAT5G40770 Prohibitin 3 1.8 RtOS, MAT1G07920 GTP-binding Elongation factor Tu protein 1.7 RCi, M, VacAT3G42050 Vacuolar ATP synthase subunit H protein 1.7 IT, OP, RBS, Vac M, PMAT4G34200 D-3-phosphoglycerate dehydrogenase 1.7 IT, MAT5G15650 Reversibly glycosylated polypeptide 2 1.7 RCi, RtOS, CWAT3G16450 Mannose-binding lectin superfamily protein 1.6 RTS, OMAT3G23990 Heat shock protein 60 1.6 RCi, RTS, Vac MAT4G11150 Vacuolar ATP synthase subunit E1 1.6 IT, OP, OiM, RtOS, RTS, RBSAT5G08670 ATP synthase alpha/beta family protein 1.6 IT, OiMAT5G35530 Ribosomal protein S3 family protein 1.6 RtOS, MAT3G01280 Voltage-dependent anion channel 1 1.5 IT, OM, RBSAT3G15950 DNA topoisomerase-related 1.5 RtOS, MAT3G52990 Pyruvate kinase family protein 1.5 RCi, MAT5G13450 Delta subunit of Mt ATP synthase 1.5 IT, OiM
CW-cell wall, ER-endoplasmic reticulum, IT-ion transport, mATPet-mitochondrion ATP synthase coupled electron transport, M-membrane (based on TAIR annotation), M⁄-proteosome and chloroplast envelope, OiM-organelle inner membrane, OM-outer membrane, OP-oxidative phosphorylation, PM- plasma membrane, RTS-response totemperature stimulus, RBS-response to biotic stimulus, RCi-response to cadmium ion, RtOS-response to osmotic/salt stress, Vac-vacuole.
N.M. Ordoñez et al. / FEBS Letters xxx (2014) xxx–xxx 5
(nine proteins) and biosynthesis of phenylpropanoids (seven pro-teins; ath01061). The KEGG analysis indicated that many of the dif-ferentially expressed microsomal proteins play a role inmodulating the level of translation and oxidative phosphorylationnetworks. The cGMP-dependent increase in oxidative phosphory-lation associated proteins as well as proteins with a role in abioticstress responses is consistent with findings in animal systems thatshowed that cGMP-dependent stress responses are linked to in-creases in energy demand [29,30].
Cyclic nucleotides and notably, cGMP and cAMP are increas-ingly becoming recognized as important second messengers inhigher plants. Cyclic nucleotide-dependent signalling is oftenlinked to transient increases in cytosolic Ca2+, and it is likely thatthe cooperation of both messengers is important in modulatingabiotic stress [22] and biotic stress responses e.g. [31,32].Cooperation between these second messengers is further sup-ported by the fact that Cyclic Nucleotide Gated Channels (CNGCs)harbour a calmodulin-binding site in their C-terminal cytosolicdomain [33]. Furthermore, cGMP has also been shown to cause a
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotideroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
time-dependent increase in the content of oxidised methionineresidues in a group of proteins functionally enriched for stress re-sponses, suggesting that this cGMP-dependent post-translationalmodification may act as a cellular signal [34] that induce a cellulardefence response.
Activation of Ca2+-permeable channels in the plasma membranehave been reported in several plant systems including guard cells[35], and it was shown recently that AtCNGC5 and AtCNGC6 areunique cGMP-activated non-selective Ca2+-permeable cation chan-nels in the plasma membrane of Arabidopsis guard cells [36]. Fur-thermore, Ca2+-currents mediated by CNGC6 channels inArabidopsis root epidermal protoplasts were activated by cytosoliccAMP [37] under the heat treatment. Such a cAMP-dependent acti-vation is not seen in our in planta studies (Fig. 3). There are twopossible explanations for this. Firstly, it could be that in ourexperiments, channels other than CNGC5 or CNGC6 may mediateH2O2-induced Ca2+-uptake. Indeed, it was shown recently that aROS-regulated Ca2+-transport protein, Annexin 1 (AtANN1), canmediate H2O2-induced Ca2+-uptake in Arabidopsis roots and is
s modulate potassium and calcium flux responses to H2O2 in Arabidopsis

Table 2cGMP-responsive down regulated microsomal proteins.
Accession No. Protein description Fold change GO enrichment
AT2G21390 Coatomer subunit 0.3 VMAT1G33120 Ribosomal protein L6 family 0.4 PM, Vac MAT4G31300 N-terminal nucleophile aminohydrolases 0.4 RtOS, Vac MAT5G14040 Phosphate transporter 3,1 0.4 CWAT5G17820 Peroxidase superfamily protein 0.4 CWAT5G19990 Regulatory particle triple-A ATPase 6A 0.4 MAT1G47260 Gamma carbonic anhydrase 2 0.5 RtOS, MAT1G53750 Regulatory particle triple-A 1A 0.5 PMAT1G70600 Ribosomal protein L18e/L15 protein 0.5 MAT2G21660 Cold, circadian rhythm & RNA binding 2 0.5 RCi, RtOS, M⁄AT2G22420 Peroxidase superfamily protein 0.5 ExRAT2G44350 Citrate synthase family protein 0.5 RCi, CWAT3G02530 TCP-1/cpn60 chaperonin family protein 0.5 RCi, PMAT3G09440 Heat shock protein 70 0.5 RCi, CWAT3G09630 Ribosomal protein L4/L1 family 0.5 CWAT3G09840 Cell division cycle 48 0.5 RCi, CWAT3G13790 Glycosyl hydrolases family 32 protein 0.5 CWAT4G13930 Serine hydroxymethyltransferase 4 0.5 RCi, PMAT4G16720 Ribosomal protein L23/L15e protein 0.5 PMAT5G02870 Ribosomal protein L4/L1 family 0.5 M, Vac MAT5G20010 RAS-related nuclear protein-1 0.5 RCi, RtOS, CWAT5G64100 Peroxidase superfamily protein 0.5 CWAT1G08360 Ribosomal protein L1p/L10e family 0.6 PMAT1G20200 PAM domain protein 0.6 PMAT1G66280 Glycosyl hydrolase superfamily protein 0.6 RtOS, MAT1G71695 Peroxidase superfamily protein 0.6 CWAT1G78900 Vacuolar ATP synthase subunit A 0.6 RtOS, CWAT1G79530 Glyceraldehyde-3-phosphate dehydr. 0.6 MAT2G04390 Ribosomal S17 family protein 0.6 PMAT2G41810 Protein of unknown function, DUF642 0.6 ExRAT3G03960 TCP-1/cpn60 chaperonin family protein 0.6 MAT3G04720 Pathogenesis-related 4 0.6 RtOS, ExRAT3G07110 Ribosomal protein L13 family protein 0.6 MAT3G07390 Auxin-responsive family protein 0.6 PMAT3G08530 Clathrin, heavy chain 0.6 VMAT3G11130 Clathrin, heavy chain 0.6 VMAT3G11400 Eukaryotic translation IF factor 3G1 0.6 PMAT3G14600 Ribosomal protein L18ae/LX protein 0.6 MAT3G43190 Sucrose synthase 4 0.6 PMAT3G51800 Metallopeptidase M24 family protein 0.6 PM, NucAT3G54400 Eukaryotic aspartyl protease protein 0.6 CWAT4G24820 26S proteasome, regulatory subunit Rpn7 0.6 MAT4G39260 Cold, circadian rhythm & RNA binding 1 0.6 RtOS, CWAT5G02500 Heat shock cognate protein 70–1 0.6 RCi,M, CWAT5G20290 Ribosomal protein S8e family protein 0.6 CWAT5G26260 TRAF-like family protein 0.6 ExRAT5G52470 Fibrillarin 1 0.6 M, Nuc
CW-cell wall, ExR-extracellular region, M-membrane (based on TAIR annotation), M⁄-peroxisome, Nuc-Nucleus, PM- plasma membrane RCi-response to cadmium ion, RtOS-response to osmotic/salt stress, Vac-vacuole, VM-vesicle membrane.
6 N.M. Ordoñez et al. / FEBS Letters xxx (2014) xxx–xxx
responsible for the stress-induced elevation of cytosolic free Ca2+
[38]. Secondly, it is conceivable that effects of CNs on CNGC activityin planta may be indirect and mediated by messengers absent inpatch-clamp experiments.
Since the treatment of roots with H2O2 has induced massive K+-efflux that is reduced by cyclic nucleotides, we propose that theCNGC are at least in parts responsible for the flux. This hypothesisis supported by several findings. Firstly, CNGCs are gated by cyclicnucleotides on the cytosolic side that harbours the cyclic nucleo-tide-binding site. This would also explain why the cell permeantanalogue is more effective at preventing K+-loss. Secondly, CNGCsbelong to the group of non-selective cation channels (NSCC) and,as such, are permeable to both K+ and Ca2+ therefore accountingfor the modulation of the Ca2+-flux by the cyclic nucleotide pre-treatment. Importantly, the role of CNGC in plant adaptive re-sponses to salt stress was clearly demonstrated (e.g. CNGC10;[39,40]). Thirdly, patch-clamp experiments have suggested thatsome NSCC channels may indeed be activated by either H2O2
[41] or hydroxyl radicals [42]. However, the molecular identity of
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotideroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
these channels remains to be discovered, and it remains to be an-swered of whether the observed H2O2-induced changes in K+- andCa2+-fluxes are mediated by the same specific CNGC, or by differentchannels from this group. A further possibility is that observed ef-fects of CN on net K+-fluxes in CN-pre-treated roots may be indirectand mediated by the voltage-gated K+-selective (e.g. GORK; [13])channels. Indeed, the observed 1.8–1.9 increase in amount of H+-ATPase or its subunits may be indicative of CN activation of thiselectrogenic transport system. This hypothesis should be testedin direct experiments including net H+-flux and membrane poten-tial measurements, as well as biochemical assays of H+-ATPasetransport activity.
Given that cyclic nucleotide pre-treatment does markedlychange the proteome, we were interested to learn if the changesin proteomic signature can shed some light on the systems re-sponse that in turn can explain the protective effect of the treat-ment. The two most noteworthy characteristics of the proteomicsresponse induced within 1 h post-cyclic nucleotide treatmentwere, firstly, the up-regulation of proteins involved in abiotic stress
s modulate potassium and calcium flux responses to H2O2 in Arabidopsis

N.M. Ordoñez et al. / FEBS Letters xxx (2014) xxx–xxx 7
responses and secondly proteins with a direct or indirect role in iontransport. These would suggest that cyclic nucleotides act as mes-sengers that are sufficient to trigger an abiotic defense responsethat in turn has a marked effect on H2O2-dependent ion fluxes.The cyclic nucleotide-dependent accumulation of proteins in-volved in ion transport are consistent with changes in H2O2-depen-dent ion flux signatures and also with specific cGMP-dependenttranscript accumulation of genes that encode monovalent cationtransporters such as non-selective ion channels and cation:protonantiporters [43]. However, it also appears that cGMP is sufficient todirectly and/or indirectly induce a highly selective change in theproteome showing that it does induce many proteins, some thatare directly involved in ion transport and others annotated as hav-ing a role in metabolism and abiotic stress responses. It is conceiv-able that cGMP does so indirectly by activating via CNGCs that inturn increase cytosolic Ca2+ that then acts as a second messengerin downstream signaling. The changes in the cGMP-dependentproteome that result in changed H2O2-dependent ion fluxes areindication of the complex mechanisms that govern ion transportand ion homeostasis at the systems level.
Acknowledgments
We wish to thank Madhava Yada from the Bioscience Core Lab-oratory, King Abdullah University of Science and Technology, forproviding technical support. Sergey Shabala acknowledges thefinancial support from the Australian Research Council. Mr. Hong-hong Wu is acknowledged for his assistance with data statisticalanalysis.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.febslet.2014.01.062.
References
[1] Bolwell, G.P. (1999) Role of active oxygen species and NO in plant defenceresponses. Curr. Opin. Plant Biol. 2, 287–294.
[2] Prasad, T.K., Anderson, M.D., Martin, B.A. and Stewart, C.R. (1994) Evidence forchilling-induced oxidative stress in maize seedlings and a regulatory role forhydrogen peroxide. Plant Cell 6, 65–74.
[3] Mittler, R. et al. (2011) ROS signaling: the new wave? Trends Plant Sci. 16,300–309.
[4] Sarry, J.E. et al. (2006) The early responses of Arabidopsis thaliana cells tocadmium exposure explored by protein and metabolite profiling analyses.Proteomics 6, 2180–2198.
[5] Vranova, E., Atichartpongkul, S., Villarroel, R., Van Montagu, M., Inze, D. andVan Camp, W. (2002) Comprehensive analysis of gene expression in Nicotianatabacum leaves acclimated to oxidative stress. Proc. Natl. Acad. Sci. USA 99,10870–10875.
[6] Karpinski, S., Reynolds, H., Karpinska, B., Wingsle, G., Creissen, G. andMullineaux, P. (1999) Systemic signaling and acclimation in response toexcess excitation energy in Arabidopsis. Science 284, 654–657.
[7] Rodrigo-Moreno, A., Poschenrieder, C. and Shabala, S. (2013) Transition metals:a double edge sward in ROS generation and signaling. Plant Signal. Behav. 8.
[8] Bose, J., Rodrigo-Moreno, A., Shabala, S. (2013) ROS homeostasis in halophytesin the context of salinity stress tolerance, J. Exp. Bot. http://dx.doi.org/10.1093/jxb/ert430.
[9] Pottosin, I., Velarde-Buendía, A.M., Bose, J., Zepeda-Jazo, I., Shabala, S.,Dobrovinskaya, O. (2014) Cross-talk between ROS and polyamines inregulation of ion transport across plasma membrane: implications for plantadaptive responses. J. Exp. Bot. http://dx.doi.org/10.1093/jxb/ert423.
[10] Borsani, O., Valpuesta, V. and Botella, M.A. (2001) Evidence for a role ofsalicylic acid in the oxidative damage generated by NaCl and osmotic stress inArabidopsis seedlings. Plant Physiol. 126, 1024–1030.
[11] de Azevedo Neto, A.D., Prisco, J.T., Eneas-Filho, J., Medeiros, J.V. and Gomes-Filho, E. (2005) Hydrogen peroxide pre-treatment induces salt-stressacclimation in maize plants. J. Plant Physiol. 162, 1114–1122.
[12] Zhu, J.K. (2003) Regulation of ion homeostasis under salt stress. Curr. Opin.Plant Biol. 6, 441–445.
[13] Shabala, S. and Cuin, T.A. (2008) Potassium transport and plant salt tolerance.Physiol. Plant 133, 651–669.
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotideroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
[14] Shabala, S., Cuin, T.A., Prismall, L. and Nemchinov, L.G. (2007) Expression ofanimal CED-9 anti-apoptotic gene in tobacco modifies plasma membrane ionfluxes in response to salinity and oxidative stress. Planta 227, 189–197.
[15] Demidchik, V., Cuin, T.A., Svistunenko, D., Smith, S.J., Miller, A.J., Shabala, S.,Sokolik, A. and Yurin, V. (2010) Arabidopsis root K+-efflux conductanceactivated by hydroxyl radicals: single-channel properties, genetic basis andinvolvement in stress-induced cell death. J. Cell Sci. 123, 1468–1479.
[16] Véry, A.A. and Sentenac, H. (2002) Cation channels in the Arabidopsis plasmamembrane. Trends Plant Sci. 7, 168–175.
[17] Demidchik, V. and Maathuis, F.J. (2007) Physiological roles of nonselectivecation channels in plants: from salt stress to signalling and development. NewPhytol. 175, 387–404.
[18] Foreman, J. et al. (2003) Reactive oxygen species produced by NADPH oxidaseregulate plant cell growth. Nature 422, 442–446.
[19] Yoshioka, K., Moeder, W., Kang, H.G., Kachroo, P., Masmoudi, K., Berkowitz, G.and Klessig, D.F. (2006) The chimeric Arabidopsis CYCLIC NUCLEOTIDE-GATEDION CHANNEL11/12 activates multiple pathogen resistance responses. PlantCell 18, 747–763.
[20] Gobert, A., Park, G., Amtmann, A., Sanders, D. and Maathuis, F.J. (2006) Arabidopsisthaliana cyclic nucleotide gated channel 3 forms a non-selective ion transporterinvolved in germination and cation transport. J. Exp. Bot. 57, 791–800.
[21] Pasqualini, S., Meier, S., Gehring, C., Madeo, L., Fornaciari, M., Romano, B. andEderli, L. (2009) Ozone and nitric oxide induce cGMP-dependent and -independent transcription of defence genes in tobacco. New Phytol. 181, 860–870.
[22] Donaldson, L., Ludidi, N., Knight, M.R., Gehring, C. and Denby, K. (2004) Saltand osmotic stress cause rapid increases in Arabidopsis thaliana cGMP levels.FEBS Lett. 569, 317–320.
[23] Ordonez, N.M., Shabala, L., Gehring, C. and Shabala, S. (2013) Noninvasivemicroelectrode ion flux estimation technique (MIFE) for the study of theregulation of root membrane transport by cyclic nucleotides. Methods Mol.Biol. 1016, 95–106.
[24] Moeder, W., Del Pozo, O., Navarre, D.A., Martin, G.B. and Klessig, D.F. (2007)Aconitase plays a role in regulating resistance to oxidative stress and celldeath in Arabidopsis and Nicotiana benthamiana. Plant Mol. Biol. 63, 273–287.
[25] Heazlewood, J.L., Tonti-Filippini, J.S., Gout, A.M., Day, D.A., Whelan, J. andMillar, A.H. (2004) Experimental analysis of the Arabidopsis mitochondrialproteome highlights signaling and regulatory components, providesassessment of targeting prediction programs, and indicates plant-specificmitochondrial proteins. Plant Cell 16, 241–256.
[26] Zhang, Y., Feng, S., Chen, F., Chen, H., Wang, J., McCall, C., Xiong, Y. and Deng,X.W. (2008) Arabidopsis DDB1-CUL4 ASSOCIATED FACTOR1 forms a nuclearE3 ubiquitin ligase with DDB1 and CUL4 that is involved in multiple plantdevelopmental processes. Plant Cell 20, 1437–1455.
[27] Kanehisa, M. et al. (2006) From genomics to chemical genomics: newdevelopments in KEGG. Nucleic Acids Res. 34, D354–D357.
[28] Moriya, Y., Itoh, M., Okuda, S., Yoshizawa, A.C. and Kanehisa, M. (2007) KAAS:an automatic genome annotation and pathway reconstruction server. NucleicAcids Res. 35, W182–W185.
[29] Laustiola, K., Vuorinen, P. and Metsa-Ketela, T. (1984) 8-bromo-cGMPimproves energy state in hypoxic rat atria. Acta Pharm. Toxicol. 55, 21–24.
[30] Young, M.E. and Leighton, B. (1998) Fuel oxidation in skeletal muscle isincreased by nitric oxide cGMP – evidence for involvement of cGMP-dependent protein kinase. FEBS Lett. 424, 79–83.
[31] Meier, S., Madeo, L., Ederli, L., Donaldson, L., Pasqualini, S. and Gehring, C.(2009) Deciphering cGMP signatures and cGMP-dependent pathways in plantdefence. Plant Signal. Behav. 4, 307–309.
[32] Qi, Z., Verma, R., Gehring, C., Yamaguchi, Y., Zhao, Y., Ryan, C.A. and Berkowitz,G.A. (2010) Ca2+ signaling by plant Arabidopsis thaliana Pep peptides dependson AtPepR1, a receptor with guanylyl cyclase activity, and cGMP-activatedCa2+ channels. Proc. Natl. Acad. Sci. USA 107, 21193–21198.
[33] Zelman, A.K., Dawe, A., Gehring, C. and Berkowitz, G.A. (2012) Evolutionaryand structural perspectives of plant cyclic nucleotide-gated cation channels.Front. Plant Sci. 3, 95.
[34] Marondedze, C., Turek, I., Parrott, B., Thomas, L., Jankovic, B., Lilley, K.S. andGehring, C. (2013) Structural and functional characteristics of cGMP-dependent methionine oxidation in Arabidopsis thaliana proteins. CellCommun. Signal. 11, 1.
[35] Pei, Z.M., Murata, Y., Benning, G., Thomine, S., Klusener, B., Allen, G.J., Grill, E.and Schroeder, J.I. (2000) Calcium channels activated by hydrogen peroxidemediate abscisic acid signalling in guard cells. Nature 406, 731–734.
[36] Wang, Y.F., Munemasa, S., Nishimura, N., Ren, H.M., Robert, N., Han, M.,Puzorjova, I., Kollist, H., Lee, S., Mori, I. and Schroeder, J.I. (2013) Identificationof cyclic GMP-activated nonselective Ca2+-permeable cation channels andassociated CNGC5 and CNGC6 genes in Arabidopsis guard cells. Plant Physiol.163, 578–590.
[37] Gao, F., Han, X.W., Wu, J.H., Zheng, S.Z., Shang, Z.L., Sun, D.Y., Zhou, R.G. and Li,B. (2012) A heat-activated calcium-permeable channel – Arabidopsis cyclicnucleotide-gated ion channel 6 – is involved in heat shock responses. Plant J.70, 1056–1069.
[38] Richards, S.L., Laohavisit, A., Mortimer, J.C., Shabala, L., Swarbreck, S.M.,Shabala, S. and Davies, J.M. (2014) Annexin 1 regulates the H2O2-inducedcalcium signature in Arabidopsis thaliana roots. Plant J. 77, 136–145.
[39] Guo, K.M., Babourina, O., Christopher, D.A., Borsic, T. and Rengel, Z. (2010) Thecyclic nucleotide-gated channel AtCNGC10 transports Ca2+ and Mg2+ inArabidopsis. Physiol. Plant. 139, 303–312.
s modulate potassium and calcium flux responses to H2O2 in Arabidopsis

8 N.M. Ordoñez et al. / FEBS Letters xxx (2014) xxx–xxx
[40] Guo, K.M., Babourina, O., Christopher, D.A., Borsics, T. and Rengel, Z. (2008)The cyclic nucleotide-gated channel, AtCNGC10, influences salt tolerance inArabidopsis. Physiol. Plant. 134, 499–507.
[41] Demidchik, V., Shabala, S.N. and Davies, J.M. (2007) Spatial variation in H2O2response of Arabidopsis thaliana root epidermal Ca2+ flux and plasmamembrane Ca2+ channels. Plant J. 49, 377–386.
Please cite this article in press as: Ordoñez, N.M., et al. Cyclic mononucleotideroots. FEBS Lett. (2014), http://dx.doi.org/10.1016/j.febslet.2014.01.062
[42] Demidchik, V., Shabala, S.N., Coutts, K.B., Tester, M.A. and Davies, J.M. (2003)Free oxygen radicals regulate plasma membrane Ca2+- and K+-permeablechannels in plant root cells. J. Cell Sci. 116, 81–88.
[43] Maathuis, F.J. (2006) CGMP modulates gene transcription and cation transportin Arabidopsis roots. Plant J. 45, 700–711.
s modulate potassium and calcium flux responses to H2O2 in Arabidopsis
Copyright © 2022 FDOKUMEN



















