Curcumin targets the AKT–mTOR pathway for uterine leiomyosarcoma tumor growth suppression
10
ORIGINAL ARTICLE Curcumin targets the AKT–mTOR pathway for uterine leiomyosarcoma tumor growth suppression Tze Fang Wong • Takashi Takeda • Bin Li • Kenji Tsuiji • Akiko Kondo • Mari Tadakawa • Satoru Nagase • Nobuo Yaegashi Received: 1 October 2012 / Accepted: 10 April 2013 / Published online: 11 May 2013 Ó Japan Society of Clinical Oncology 2013 Abstract Background Uterine leiomyosarcomas generally do not respond well to standard chemotherapy. We previously demonstrated that curcumin, the active ingredient derived from the herb Curcuma longa, inhibits uterine leiomyo- sarcoma cells in vitro via the inhibition of the AKT– mammalian target of rapamycin (mTOR) pathway. As a preclinical investigation, we performed an in vivo study using female nude mice to confirm the therapeutic potential of curcumin against uterine leiomyosarcoma. Methods Human leiomyosarcoma cells, SK-UT-1, were inoculated in female nude mice to establish subcutaneous tumors. Either vehicle control or 250 mg/kg curcumin was administered intraperitoneally every day for 14 consecutive days, and the mice were then killed. The tumors were measured every 2–3 days. The tumors were processed for immunohis- tochemical analyses to detect total AKT, phosphorylated AKT, total mTOR, phosphorylated mTOR, and phosphorylated S6. To detect apoptosis, the tumors were stained for cleaved PARP and TUNEL. Ki-67 immunohistochemistry was performed to determine cell viability of the tumors. Results Compared with the control, curcumin reduced uterine leiomyosarcoma tumor volume and mass signifi- cantly with a concordant decrease in mTOR and S6 phosphorylation. However, AKT phosphorylation was not significantly altered. Cleaved PARP and TUNEL staining increased significantly with curcumin administration, indicating the induction of apoptosis. There was no dif- ference in Ki-67 staining between the two groups. Conclusion Curcumin inhibited uterine leiomyosarcoma tumor growth in vivo by targeting the AKT–mTOR path- way for inhibition. Keywords Curcumin Á Uterine leiomyosarcoma Á AKT–mTOR pathway Á Apoptosis Á In vivo Introduction Uterine leiomyosarcoma is a rare malignant tumor that com- prises 1 % of all malignancies arising from the uterus. Because of its generally low response rate to standard adjuvant chemotherapy, high propensity for hematological metastases, and high frequency of recurrence, uterine leiomyosarcoma is especially difficult to treat. Currently, adjuvant treatment with doxorubicin shows the best efficacy to toxicity ratio profile for uterine leiomyosarcomas and has been the first-line therapy for the past 20 years [1]. Various other cytotoxic chemicals such as gemcitabine, docetaxel, etc. have been tested either alone or in combination, but no promising alternative to doxorubicin monotherapy has been discovered so far. Clinical trials to test molecular targeted approaches are under way. Hormone-targeted therapy using antiprogestin [2] or aroma- tase inhibitors [3], mammalian target of rapamycin (mTOR) inhibition using rapamycin analogues [4], and antiangiogen- esis using either monoclonal antibodies or target-specific small molecules [5] are some of the approaches being evalu- ated at various stages of clinical trials. The constitutive activation of the mTOR pathway is one of the most investigated mechanisms of tumorigenesis of T. F. Wong Á T. Takeda Á B. Li Á K. Tsuiji Á A. Kondo Á M. Tadakawa Á S. Nagase Á N. Yaegashi Department of Obstetrics and Gynecology, Tohoku University Graduate School of Medicine, Sendai, Miyagi, Japan Present Address: T. Takeda (&) Division of Women’s Health, Research Institute of Traditional Asian Medicine, Kinki University School of Medicine, 377-2 Ohno-Higashi, Osaka-Sayama, Osaka 589-8511, Japan e-mail: [email protected] 123 Int J Clin Oncol (2014) 19:354–363 DOI 10.1007/s10147-013-0563-4
-
Upload
independent -
Category
Documents
-
view
11 -
download
0
Transcript of Curcumin targets the AKT–mTOR pathway for uterine leiomyosarcoma tumor growth suppression

ORIGINAL ARTICLE
Curcumin targets the AKT–mTOR pathway for uterineleiomyosarcoma tumor growth suppression
Tze Fang Wong • Takashi Takeda •
Bin Li • Kenji Tsuiji • Akiko Kondo •
Mari Tadakawa • Satoru Nagase • Nobuo Yaegashi
Received: 1 October 2012 / Accepted: 10 April 2013 / Published online: 11 May 2013
� Japan Society of Clinical Oncology 2013
Abstract
Background Uterine leiomyosarcomas generally do not
respond well to standard chemotherapy. We previously
demonstrated that curcumin, the active ingredient derived
from the herb Curcuma longa, inhibits uterine leiomyo-
sarcoma cells in vitro via the inhibition of the AKT–
mammalian target of rapamycin (mTOR) pathway. As a
preclinical investigation, we performed an in vivo study
using female nude mice to confirm the therapeutic potential
of curcumin against uterine leiomyosarcoma.
Methods Human leiomyosarcoma cells, SK-UT-1, were
inoculated in female nude mice to establish subcutaneous
tumors. Either vehicle control or 250 mg/kg curcumin was
administered intraperitoneally every day for 14 consecutive
days, and the mice were then killed. The tumors were measured
every 2–3 days. The tumors were processed for immunohis-
tochemical analyses to detect total AKT, phosphorylated AKT,
total mTOR, phosphorylated mTOR, and phosphorylated S6.
To detect apoptosis, the tumors were stained for cleaved PARP
and TUNEL. Ki-67 immunohistochemistry was performed to
determine cell viability of the tumors.
Results Compared with the control, curcumin reduced
uterine leiomyosarcoma tumor volume and mass signifi-
cantly with a concordant decrease in mTOR and S6
phosphorylation. However, AKT phosphorylation was not
significantly altered. Cleaved PARP and TUNEL staining
increased significantly with curcumin administration,
indicating the induction of apoptosis. There was no dif-
ference in Ki-67 staining between the two groups.
Conclusion Curcumin inhibited uterine leiomyosarcoma
tumor growth in vivo by targeting the AKT–mTOR path-
way for inhibition.
Keywords Curcumin � Uterine leiomyosarcoma �AKT–mTOR pathway � Apoptosis � In vivo
Introduction
Uterine leiomyosarcoma is a rare malignant tumor that com-
prises 1 % of all malignancies arising from the uterus.
Because of its generally low response rate to standard adjuvant
chemotherapy, high propensity for hematological metastases,
and high frequency of recurrence, uterine leiomyosarcoma is
especially difficult to treat. Currently, adjuvant treatment with
doxorubicin shows the best efficacy to toxicity ratio profile for
uterine leiomyosarcomas and has been the first-line therapy
for the past 20 years [1]. Various other cytotoxic chemicals
such as gemcitabine, docetaxel, etc. have been tested either
alone or in combination, but no promising alternative to
doxorubicin monotherapy has been discovered so far. Clinical
trials to test molecular targeted approaches are under way.
Hormone-targeted therapy using antiprogestin [2] or aroma-
tase inhibitors [3], mammalian target of rapamycin (mTOR)
inhibition using rapamycin analogues [4], and antiangiogen-
esis using either monoclonal antibodies or target-specific
small molecules [5] are some of the approaches being evalu-
ated at various stages of clinical trials.
The constitutive activation of the mTOR pathway is one
of the most investigated mechanisms of tumorigenesis of
T. F. Wong � T. Takeda � B. Li � K. Tsuiji � A. Kondo �M. Tadakawa � S. Nagase � N. Yaegashi
Department of Obstetrics and Gynecology, Tohoku University
Graduate School of Medicine, Sendai, Miyagi, Japan
Present Address:
T. Takeda (&)
Division of Women’s Health, Research Institute of Traditional
Asian Medicine, Kinki University School of Medicine,
377-2 Ohno-Higashi, Osaka-Sayama, Osaka 589-8511, Japan
e-mail: [email protected]
123
Int J Clin Oncol (2014) 19:354–363
DOI 10.1007/s10147-013-0563-4

leiomyosarcoma. Mice harboring smooth muscle-specific
knockout of the PTEN (phosphatase and tensin homologue
deleted from chromosome 10) gene died prematurely of
leiomyosarcoma; their survival was prolonged by treatment
with everolimus, a rapamycin analogue [6]. Incidentally,
59 % of uterine and extrauterine leiomyosarcoma cases
reported in one study had deletions of the chromosomal
region 10q that contains the PTEN gene [7]. Taken toge-
ther, these findings provide the evidence backing the tar-
geted approach against the mTOR pathway.
Our previous investigation explored the possibility of
using curcumin, an active ingredient derived from the
Indian spice, Curcuma longa, as a therapeutic candidate
against uterine leiomyosarcoma. Curcumin at a concen-
tration exceeding 25 lM in vitro was able to reduce uterine
leiomyosarcoma cell growth by more than 50 % and
induce apoptosis [8]. Building on the discovery that cur-
cumin inhibits mTOR in other tumor cells such as rhab-
domyosarcoma [9] and prostate cancer [10], we previously
demonstrated that curcumin was able to suppress mTOR
activity by downregulating the phosphorylation of mTOR
(Ser2448) and its downstream effectors, p70S6 (Thr389)
and S6 (Ser235/236), in uterine leiomyosarcoma cells.
The potential of curcumin as an anticancer agent has been
explored in numerous studies. Despite its widely touted
efficacy against many cancer types regardless of their tissue
origins, curcumin has been limited by its poor bioavailability
and pharmacodynamics [11]. The most preferred mode of
administration for curcumin was the oral route, which has
been investigated in mouse models of ovarian cancer [12],
breast cancer [13], and prostate cancer [14]. One compre-
hensive study on the bioavailability of curcumin that com-
pared the oral route (p.o.) with intraperitoneal (i.p.)
administration suggests that i.p. administration is 100 times
(maximum plasma curcumin concentration, 2.25 lg/ml, at
100 mg/kg i.p.) more efficient than oral administration
(maximum plasma curcumin concentration, 0.22 lg/ml, at
1000 mg/kg p.o.) [15]. Thus, we have chosen to test curcu-
min’s action against a uterine leiomyosarcoma mouse model
in vivo using intraperitoneal curcumin administration. By
investigating curcumin’s tumor suppressive effect and
its effect on mTOR activity in vivo, we hope that our cur-
rent study will provide the necessary rationale for using
curcumin against uterine leiomyosarcoma in future clinical
investigations.
Materials and methods
Chemicals
Curcumin (from Curcuma longa, turmeric) and corn oil
were purchased from Wako Pure Chemical Industries
(Osaka, Japan). Curcumin was dissolved in dimethyl sulf-
oxide (DMSO) (Sigma–Aldrich, St. Louis, MO, USA) at
1 M concentration as a stock solution that was stored at
-20 �C. For intraperitoneal administration of curcumin,
curcumin was dissolved in 20 % DMSO and suspended in
corn oil at 250 mg/kg/50 ll per animal.
Cell culture
The uterine LMS cell line, SK-UT-1 was purchased from
American Type Culture Collection (ATCC, Manassas, VA,
USA). The cells were revived in Eagle’s minimum essen-
tial medium (Gibco, Invitrogen, Carlsbad, CA, USA)
supplemented with 10 % fetal bovine serum (FBS) and
maintained at 37 �C in a humidified 5 % CO2 atmosphere.
Recipient animal and tumor model
A uterine LMS tumor model was established in BALB/cAJC
female nude mice using SK-UT-1 tumor cells. Briefly, female
mice 5–6 weeks old were purchased from CLEA Japan and
housed in a specific-pathogen-free animal facility at the
Center for Laboratory Animal Research, Tohoku University
Graduate School of Medicine, with the approval from the
University’s committee on laboratory animal research. The
research was conducted in accordance with the regulations
laid out by the Ministry of Education, Culture, Sports, Science
and Technology, the Ministry of Health, Labour and Welfare,
and the Ministry of the Environment, Japan.
Before subcutaneous implantation of SK-UT-1 tumor
cells into nude mice, the cells were grown in monolayer
culture in medium supplemented with 20 % FBS: 107 SK-
UT-1 cells were suspended in 0.05 ml medium mixed with
0.05 ml Matrigel (Becton, Dickinson, Franklin Lakes, NJ,
USA), and then injected subcutaneously into both flanks of
mice. Subcutaneous tumor nodules appeared 7 days po-
stimplantation. Daily intraperitoneal curcumin administra-
tion (250 mg/kg/50 ll) was started on the 8th day
postimplantation. Control mice received intraperitoneal
injections of 20 % DMSO suspended in corn oil (total
volume, 50 ll per animal) as curcumin was dissolved in the
same ratio of DMSO to corn oil. The day of the start of
treatment was counted as day 1. The tumor volume was
measured every 1–3 days and was calculated as previously
described [16]. The length, width, and depth of the tumors
were measured and the volume calculated using the fol-
lowing formula: volume = R1 9 R2 9 R3 9 0.52. The
mice were killed after 14 days of treatment.
Immunohistochemistry
The subcutaneous tumors harvested were formalin fixed
and paraffin embedded. Immunohistochemical analyses
Int J Clin Oncol (2014) 19:354–363 355
123

were performed using a streptavidin–biotin amplification
method using the Histofine Kit (Nichirei, Kyoto, Japan).
Briefly, paraffin-embedded tumors were sliced, mounted on
slides, and deparaffinized for immunohistochemistry. The
slides were autoclaved at 121 �C for 5 min in citrate buffer
(pH 6.0) for antigen retrieval. For detection of the AKT–
mTOR pathway activity, antibodies detecting phosphory-
lated AKT (Thr308; P-AKT), total AKT, phosphorylated
mTOR (Ser2448; P-mTOR), total mTOR, and phosphory-
lated S6 (Ser235/236; P-S6) were purchased from Cell
Signaling Technology (Beverly, MA, USA). The staining
as defined by H scores was calculated as the product of
intensity (0–3) and percentage of area positively stained
(score range: 0–300). At least three independent micro-
scopic fields were evaluated from each sample. To check
the viability of tumor cells, antibody against Ki-67 was
purchased from Dako (Carpinteria, CA, USA). Ki-67
staining was calculated as the percentage of nuclei posi-
tively stained. For the detection of apoptosis, antibodies
detecting cleaved PARP (Cell Signaling Technology) and
TdT-mediated dUTP nick end labeling (TUNEL) staining
were used. TUNEL staining was performed using an
In Situ Cell Death Detection Kit, POD (Roche Diagnostics,
Basel, Switzerland) according to the manufacturer’s
instructions. Both cleaved PARP and TUNEL staining
were scored as the number of nuclei positively stained in
2,000 cells counted, referred to as apoptotic index (AI).
This calculation was based on a method previously
described [17]. The slides were observed and the images
taken using a Nikon Coolscope Digital Microscope (Nikon,
Tokyo, Japan).
Statistical analysis
Each experimental group experiment consisted of five mice
(n = 5). Student’s t test was used to evaluate statistical
significance. The statistical software JMP (SAS, Cary, NC,
USA) was used for analyses and graphical representations.
Differences were considered statistically significant at
p \ 0.05.
Results
Curcumin inhibits uterine leiomyosarcoma tumor
growth in vivo
Intraperitoneal administration of 250 mg/kg curcumin
dissolved in DMSO and corn oil significantly (p \ 0.05)
inhibited uterine leiomyosarcoma tumor growth compared
to tumors in control animals from day 9 (Fig. 1a). Tumors
in animals treated with curcumin continued to grow, albeit
at a slower pace, until the animals were killed at day 14,
and the size of tumors remained significantly smaller than
in the control group (curcumin, 138.0 ± 91.4 mm3, vs.
control, 291.1 ± 133.8 mm3; p \ 0.01) (Fig. 1b). Tumors
excised from the animals were weighed (results shown
in Fig. 1c). Tumors treated with curcumin had significantly
smaller mass compared with the control group (curcu-
min, 314.7 ± 163.0 mg, vs. control, 489.3 ± 170.1 mg;
p \ 0.05).
Curcumin downregulates mTOR (Ser2448) and S6
(Ser235/236) phosphorylation, but not AKT
phosphorylation (Thr308) in vivo
Our previous in vitro data suggest that curcumin inhibits
uterine leiomyosarcoma cell growth by inhibiting the
activity of mTOR. We detected the phosphorylation of
AKT, mTOR, and S6 in the excised tumors using immu-
nohistochemistry of paraffin-embedded tissue slides. Fig-
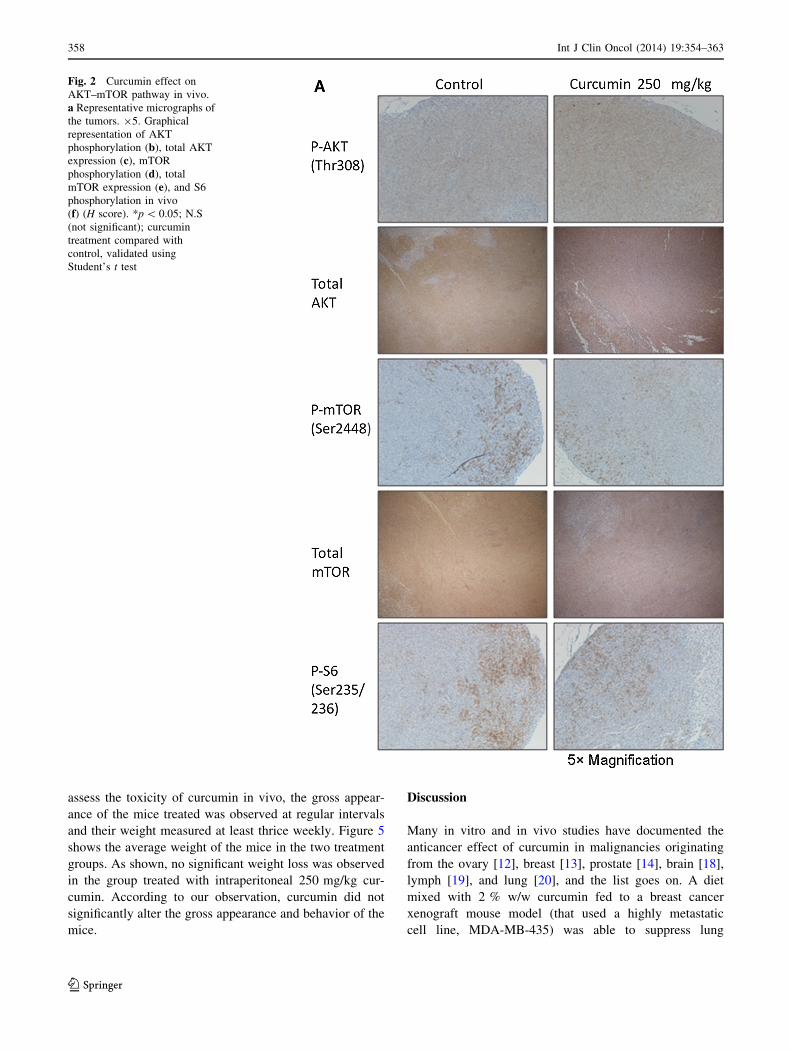
ure 2a shows representative images of tumors from both
control and curcumin-treated groups. From gross observa-
tion, curcumin seemed to lower the intensity and the area of
positive staining for mTOR and S6, whereas AKT phos-
phorylation seemed to increase slightly. Figure 2b shows
box graphs representing the results of quantification of
P-AKT, P-mTOR, and P-S6 staining. As shown, curcumin
treatment significantly downregulated mTOR (H score:
curcumin, 10.2 ± 6.4, vs. control, 18.2 ± 8.9; p \ 0.05)
and S6 phosphorylation (H score: curcumin, 12.5 ± 16.3,
vs. control, 48.5 ± 39.0; p \ 0.05) in the tumors.
The effect of curcumin on total AKT and total mTOR
expression in vivo was also investigated. Total mTOR
expression was also reduced with curcumin treatment
although the effect was not statistically significant
(H score: curcumin, 125 ± 38.1, vs. control, 97.5 ± 29.0;
p = 0.0859). Both AKT phosphorylation (H score: curcu-
min, 45.5 ± 22.0, vs. control, 27.0 ± 17.5; p = 0.0523)
and total AKT expression (H score: curcumin,
112.0 ± 49.4, vs. control, 91.5 ± 26.5; p = 0.2625)
increased slightly as a result of treatment with curcumin
but the results were not statistically significant.
Curcumin induces apoptosis in uterine leiomyosarcoma
subcutaneous xenograft
Our previous in vitro data demonstrated that curcumin
induces apoptosis in uterine leiomyosarcoma cells. We
detected cleaved PARP and fragmented DNA (TUNEL
staining) in the excised tumors using immunohistochem-
istry of paraffin-embedded tissue slides. Figure 3a shows
representative images of tumors from both control and
curcumin-treated groups. From gross observation, curcu-
min seemed to increase the number of apoptotic cells in the
tumors. Figure 3b,c shows box graphs representing the
356 Int J Clin Oncol (2014) 19:354–363
123

quantification of positive cleaved PARP and TUNEL
staining. As shown, curcumin treatment significantly
increased PARP cleavage (AI: curcumin, 9.2 ± 3.3, vs.
control, 5.0 ± 2.8; p \ 0.01) and DNA fragmentation (AI:
curcumin, 8.3 ± 4.0, vs. control, 4.6 ± 2.6; p \ 0.05) in
the tumors.
Curcumin does not affect Ki-67 expression in viable
cells
Ki-67 is frequently used to evaluate the proliferative abil-
ities of cells within a tissue. Figure 4a shows representative
images of tumors treated with either curcumin or control.
No obvious difference was observed between the two
groups. Figure 4b is a graph showing the quantified stain-
ing percentage using anti-Ki-67 antibody. Curcumin did
not significantly alter Ki-67 expression in the tumors,
although the average staining was slightly reduced, to
39.7 ± 8.4 % from 44.5 ± 7.6 % of the control group
(p = 0.202).
Treatment with curcumin is well tolerated in vivo
Standard chemotherapy often causes side effects such as
anorexia, nausea, vomiting, diarrhea, and alopecia that may
affect the overall well-being of the individual treated. To
Fig. 1 Uterine leiomyosarcoma SK-UT-1 subcutaneous tumor
growth in female nude mice, n = 5 per group. a Tumor growth in
control (squares) versus 250 mg/kg curcumin (triangles). The tumors
were measured with a caliper. Graphical representation of tumor
volume on the 14th day of treatment (b) and tumor weight on the 14th
day of treatment (c). *p \ 0.05; **p\ 0.01; curcumin treatment
compared with control, validated using Student’s t test
Int J Clin Oncol (2014) 19:354–363 357
123
assess the toxicity of curcumin in vivo, the gross appear-
ance of the mice treated was observed at regular intervals
and their weight measured at least thrice weekly. Figure 5
shows the average weight of the mice in the two treatment
groups. As shown, no significant weight loss was observed
in the group treated with intraperitoneal 250 mg/kg cur-
cumin. According to our observation, curcumin did not
significantly alter the gross appearance and behavior of the
mice.
Discussion
Many in vitro and in vivo studies have documented the
anticancer effect of curcumin in malignancies originating
from the ovary [12], breast [13], prostate [14], brain [18],
lymph [19], and lung [20], and the list goes on. A diet
mixed with 2 % w/w curcumin fed to a breast cancer
xenograft mouse model (that used a highly metastatic
cell line, MDA-MB-435) was able to suppress lung
Fig. 2 Curcumin effect on
AKT–mTOR pathway in vivo.
a Representative micrographs of
the tumors. 95. Graphical
representation of AKT
phosphorylation (b), total AKT
expression (c), mTOR
phosphorylation (d), total
mTOR expression (e), and S6
phosphorylation in vivo
(f) (H score). *p \ 0.05; N.S
(not significant); curcumin
treatment compared with
control, validated using
Student’s t test
358 Int J Clin Oncol (2014) 19:354–363
123

metastasis. The study did not compare the tumor size
between treatment groups, but was able to demonstrate
that curcumin significantly downregulated Ki-67 expres-
sion in the tumors [13]. An in vivo ovarian cancer model
showed that daily oral curcumin at 500 mg/kg dosage
could reduce tumor growth by as much as 50 %,
although only marginal statistical significance was
achieved [12]. In the prostate cancer mouse model, 5 mg/
day curcumin was fed to mice harboring prostate cancer
cells PC-3 xenografts and tumor growth was monitored
for at least 30 days [14]. In this study, curcumin was
able to suppress tumor growth by almost 50 % compared
with the control, although the statistical significance of
this finding was not mentioned.
Daily intratumoral injection of 100 mg/kg curcumin in an
in vivo glioma mouse model significantly reduced the size of
the subcutaneous tumor xenografts compared with the con-
trol group [18]. This study demonstrated that curcumin
inhibits the AKT–mTOR pathway in glioma cells in vitro,
but the tumors excised were only analyzed for LC-3 (a
marker for autophagy) expression. In a melanoma mouse
model, curcumin was administered intraperitoneally, similar
Fig. 2 continued
Int J Clin Oncol (2014) 19:354–363 359
123

to our study. The dosage used was 25 mg/kg daily, and no
therapeutic effect was observed when curcumin was
administered alone. However, in mice inoculated with anti-
bodies directed against the melanoma cells, curcumin was
able to prolong their survival compared with the untreated
controls [21].
In vivo studies investigating curcumin’s effect in mouse
xenograft models so far could not consistently demonstrate
curcumin’s efficacy. Our study showed that 250 mg/kg
curcumin administered intraperitoneally was able to slow
uterine leiomyosarcoma tumor growth, and the effect was
statistically significant. Although other studies have dem-
onstrated curcumin’s inhibitory effect on various other
pathways, we are the first to show that intraperitoneal
curcumin could inhibit mTOR, S6 phosphorylation, indi-
cating that curcumin is indeed effective against the AKT–
mTOR pathway in vivo. More notably, we have quantified
the effect of curcumin on the tumor tissues and analyzed
the results for statistical significance.
We attribute our favorable results to two factors: first,
the intraperitoneal route of administration, which has
shown higher bioavailability for curcumin in vivo [15]; and
second, a sufficiently high dose, 250 versus 25 mg/kg in
the melanoma mouse model [21]. Assuming that curcumin
bioavailability in nude mice is similar with the study pre-
viously described [15], 250 mg/kg curcumin will yield a
maximum plasma concentration of 5.63 lg/ml or 15.3 lM.
This concentration should be sufficiently high to cause
cytotoxicity to SK-UT-1 cells, as predicted by our previous
in vitro study [8].
We noticed that intraperitoneal administration of
250 mg/kg curcumin could not reduce AKT phosphory-
lation. In contrast, P-AKT staining increased slightly in
the tumor cells in the curcumin-treated group. Our
Fig. 3 Curcumin effect on apoptosis in vivo. a Representative
micrographs of the tumors. 920. Graphical representation of PARP
cleavage (b) and TUNEL staining in vivo (c). AI apoptotic index,
number of darkly stained nuclei in 2,000 cells calculated. *p \ 0.05;
**p \ 0.01; curcumin treatment compared with control, validated
using Student’s t test
360 Int J Clin Oncol (2014) 19:354–363
123

previous investigation on uterine leiomyosarcoma cells
in vitro suggested that low curcumin concentration
could not inhibit AKT phosphorylation [22]. A higher
concentration or enhanced bioavailability using an adju-
vant such as epigallocatechin-3-gallate (EGCG) was nee-
ded to effectively inhibit AKT activity. Perhaps this was
the reason why intraperitoneal 250 mg/kg curcumin daily
could not entirely eliminate uterine leiomyosarcoma in our
mouse model. Sustained AKT phosphorylation as a result
of low plasma curcumin concentration probably enabled
the survival of the tumor cells, as AKT not only reacti-
vates mTOR, but also suppresses apoptosis [23]. It has
been shown that specific inhibition of mTOR by rapa-
mycin causes a negative feedback reactivation of AKT [8,
9]. Although low curcumin concentrations can inhibit
mTOR, a higher concentration is required to inhibit AKT
as well.
Moreover, curcumin has varying affinity for different
kinases [24]. The inhibitory effect of curcumin was
compared between protein kinase A (PkA), protein kinase
C (PkC), protamine kinase (cPK), phosphorylase kinase
(PhK), autophosphorylation-activated protein kinase
(AK), and pp60c-src tyrosine kinase. At about 0.1 mM
(100 lM) curcumin, PhK, pp60c-src, PkC, PkA, AK, and
cPK were inhibited by 98 , 40 , 15 , 10 , l , and 0.5 %,
respectively. Therefore, it is considered reasonable that
AKT requires a higher concentration of curcumin to be
inhibited because different proteins have different affini-
ties for curcumin.
Fig. 4 Curcumin effect on
tumor cell viability in vivo.
a Representative micrographs of
the tumors. 920. Graphical
representation of Ki-67 staining
in vivo (b). N.S, statistically not
significant; curcumin treatment
compared with control,
validated using Student’s t test
Fig. 5 Posttreatment body weight of nude mice (g). Black bars,
control; white bars, curcumin 250 mg/kg
Int J Clin Oncol (2014) 19:354–363 361
123

Excluding clinical trials investigating chemoprevention
using curcumin, phase I or II clinical trials using curcumin
so far were only conducted on pancreatic cancer cases [25,
26]. Curcumin was administered orally in these studies.
Although curcumin was generally well tolerated clinically
and produced very few adverse effects, the plasma con-
centration of curcumin ranged from 29 to 412 ng/ml [27],
which translates to a maximum concentration of
7.87–111.84 nM. This concentration is well below the
cytotoxic dose against cancer cells as predicted by in vitro
studies [28]. As a result, oral curcumin in those clinical
trials failed to significantly improve the survival of the
patients, and the average response rates were about 3 %.
Experimenting with different modes of administration
has certainly reached its limitation in improving curcumin
bioavailability. As demonstrated by our previous study [22]
and another group that combined piperine with curcumin
[29], adding an adjuvant may improve the cellular uptake
and absorption of curcumin in the digestive tract. Another
approach is to prepare curcumin as encapsulated nanopar-
ticles [30]. Such an approach has proven to improve oral
bioavailability of curcumin by as much as ninefold.
However, more research is needed to make curcumin fully
ready to be utilized effectively in a clinical setting.
In conclusion, our study was able to prove the efficacy
of intraperitoneal curcumin in vivo. Intraperitoneal curcu-
min was able to reduce uterine leiomyosarcoma cell growth
in vivo via the inhibition of mTOR activity that resulted in
the induction of apoptosis. Our study and analyses suggest
that, although promising, the bioavailability of curcumin
needs to be significantly improved for curcumin to be
effectively utilized clinically.
Acknowledgments We thank Ms. Emi Endo, Ms. Etsuko Oba, and
Ms. Etsuko Tomita for assistance in tissue processing and immuno-
histochemical analysis. This study was approved by the Ethics
Committee for animal research and was supported by a Grant-in-aid
from the Japanese Ministry of Education, Science, Sports, and Cul-
ture, Tokyo, Japan (23592430).
Conflict of interest The authors report no conflict of interest.
References
1. Amant F, Coosemans A, Debiec-Rychter M et al (2009) Clinical
management of uterine sarcomas. Lancet Oncol 10:1188–1198
2. Koivisto-Korander R, Leminen A, Heikinheimo O (2007) Mife-
pristone as treatment of recurrent progesterone receptor-positive
uterine leiomyosarcoma. Obstet Gynecol 109:512–514
3. O’Cearbhaill R, Zhou Q, Iasonos A et al (2010) Treatment of
advanced uterine leiomyosarcoma with aromatase inhibitors.
Gynecol Oncol 116:424–429
4. Quek R, Wang Q, Morgan JA et al (2011) Combination mTOR
and IGF-1R inhibition: phase I trial of everolimus and
figitumumab in patients with advanced sarcomas and other solid
tumors. Clin Cancer Res 17:871–879
5. Ganjoo K, Jacobs C (2010) Antiangiogenesis agents in the
treatment of soft tissue sarcomas. Cancer (Phila) 116:1177–1183
6. Hernando E, Charytonowicz E, Dudas ME et al (2007) The
AKT–mTOR pathway plays a critical role in the development of
leiomyosarcomas. Nat Med 13:748–753
7. Hu J, Rao UNM, Jasani S et al (2005) Loss of DNA copy number
of 10q is associated with aggressive behavior of leiomyosarco-
mas: a comparative genomic hybridization study. Cancer Genet
Cytogenet 161:20–27
8. Wong TF, Takeda T, Li B et al (2011) Curcumin disrupts uterine
leiomyosarcoma cells through AKT–mTOR pathway inhibition.
Gynecol Oncol 122:141–148
9. Beevers CS, Chen L, Liu L et al (2009) Curcumin disrupts the
mammalian target of rapamycin–raptor complex. Cancer Res
69:1000–1008
10. Yu S, Shen G, Khor TO et al (2008) Curcumin inhibits Akt/
mammalian target of rapamycin signaling through protein phos-
phatase-dependent mechanism. Mol Cancer Ther 7:2609–2620
11. Anand P, Sundaram C, Jhurani S et al (2008) Curcumin and
cancer: an ‘‘old-age’’ disease with an ‘‘age-old’’ solution. Cancer
Lett 267:133–164
12. Lin YG, Kunnumakkara AB, Nair A et al (2007) Curcumin
inhibits tumor growth and angiogenesis in ovarian carcinoma by
targeting the nuclear factor-jB pathway. Clin Cancer Res
13:3423–3430
13. Aggarwal BB, Shishodia S, Takada Y et al (2005) Curcumin
suppresses the paclitaxel-induced nuclear factor-jB pathway in
breast cancer cells and inhibits lung metastasis of human breast
cancer in nude mice. Clin Cancer Res 11:7490–7498
14. Li M, Zhang Z, Hill DL et al (2007) Curcumin, a dietary com-
ponent, has anticancer, chemosensitization, and radiosensitization
effects by down-regulating the MDM2 oncogene through the
PI3K/mTOR/ETS2 pathway. Cancer Res 67:1988–1996
15. Pan M-H, Huang T-M, Lin J-K (1999) Biotransformation of
curcumin through reduction and glucuronidation in mice. Drug
Metab Dispos 27:486–494
16. Tsuiji K, Takeda T, Li B et al (2010) Establishment of a novel
xenograft model for human uterine leiomyoma in immunodefi-
cient mice. Tohoku J Exp Med 222:55–61
17. Bressenot A, Marchal S, Bezdetnaya L et al (2009) Assessment of
apoptosis by immunohistochemistry to active caspase-3, active
caspase-7, or cleaved PARP in monolayer cells and spheroid and
subcutaneous xenografts of human carcinoma. J Histochem Cy-
tochem 57:289–300
18. Aoki H, Takada Y, Kondo S et al (2007) Evidence that curcumin
suppresses the growth of malignant gliomas in vitro and in vivo
through induction of autophagy: role of Akt and extracellular
signal-regulated kinase signaling pathways. Mol Pharmacol
72:29–39
19. Uddin S, Hussain AR, Manogaran PS et al (2005) Curcumin
suppresses growth and induces apoptosis in primary effusion
lymphoma. Oncogene 24:7022–7030
20. Radhakrishna Pillai G, Srivastava AS, Hassanein TI et al (2004)
Induction of apoptosis in human lung cancer cells by curcumin.
Cancer Lett 208:163–170
21. Odot J, Albert P, Carlier A et al (2004) In vitro and in vivo anti-
tumoral effect of curcumin against melanoma cells. Int J Cancer
111:381–387
22. Kondo A, Takeda T, Li B et al (2012) Epigallocatechin-3-gallate
potentiates curcumin’s ability to suppress uterine leiomyosar-
coma cell growth and induce apoptosis. Int J Clin Oncol doi:
10.1007/s10147-012-0387-7
23. Scheid MP, Duronio V (1998) Dissociation of cytokine-induced
phosphorylation of Bad and activation of PKB/akt: involvement
362 Int J Clin Oncol (2014) 19:354–363
123

of MEK upstream of Bad phosphorylation. Proc Natl Acad Sci
USA 95:7439–7444
24. Reddy S, Aggarwal BB (1994) Curcumin is a non-competitive
and selective inhibitor of phosphorylase kinase. FEBS Lett
341:19–22
25. Dhillon N, Aggarwal BB, Newman RA et al (2008) Phase II trial
of curcumin in patients with advanced pancreatic cancer. Clin
Cancer Res 14:4491–4499
26. Vaklavas C, Tsimberidou A-M, Wen S et al (2011) Phase 1
clinical trials in 83 patients with pancreatic cancer. Cancer (Phila)
117:77–85
27. Kanai M, Yoshimura K, Asada M et al (2011) A phase I/II study
of gemcitabine-based chemotherapy plus curcumin for patients
with gemcitabine-resistant pancreatic cancer. Cancer Chemother
Pharmacol 68:157–164
28. Li L, Aggarwal BB, Shishodia S et al (2004) Nuclear factor-jB
and IjB kinase are constitutively active in human pancreatic
cells, and their down-regulation by curcumin (diferuloylmethane)
is associated with the suppression of proliferation and the
induction of apoptosis. Cancer (Phila) 101:2351–2362
29. Shoba G, Joy D, Joseph T et al (1998) Influence of piperine on
the pharmacokinetics of curcumin in animals and human volun-
teers. Planta Med 64:353–356
30. Shaikh J, Ankola DD, Beniwal V et al (2009) Nanoparticle
encapsulation improves oral bioavailability of curcumin by at
least 9-fold when compared to curcumin administered with pip-
erine as absorption enhancer. Eur J Pharm Sci 37:223–230
Int J Clin Oncol (2014) 19:354–363 363
123