
sello - Chrysler Hyundai H100 motor 2.4 Lts Especificaciones ...
Upload
independentCategory
view
2download
0
doi:10.1016/j.jmb.2007.12.012 J. Mol. Biol. (2008) 376, 1060–1075
Available online at www.sciencedirect.com
Crystal Structure of Lactoperoxidase at 2.4 Å Resolution
Amit Kumar Singh, Nagendra Singh, Sujata Sharma, S. Baskar Singh,Punit Kaur, A. Bhushan, A. Srinivasan and Tej P. Singh⁎
Department of Biophysics,All India Institute of MedicalSciences, Ansari Nagar,New Delhi - 110 029, India
Received 21 September 2007;received in revised form5 December 2007;accepted 6 December 2007Available online14 December 2007
*Corresponding author. E-mail [email protected] used: ABTS,
2,2′-azino-bis(3-ethylbenzthiazoline-eosinophil peroxidase; GLPO, capriHEPO, human eosinophil peroxidasmyeloperoxidase; HTPO, human thLPO, lactoperoxidase; MAN, mannomyeloperoxidase; NAG, N-acetyl glSDS-PAGE, sodium dodecyl sulfateelectrophoresis; TPO, thyroid perox
0022-2836/$ - see front matter © 2007 E
Lactoperoxidase (LPO) is a member of the mammalian peroxidase super-family. It catalyzes the oxidation of thiocyanate and halides. Freshly isolatedand purified samples of caprine LPO were saturated with ammoniumiodide and crystallized using 20% polyethylene glycol 3350 in a hangingdrop vapor diffusion setup. The structure has been determined using X-raycrystallographic method and refined to Rcryst and Rfree factors of 0.196 and0.203, respectively. The structure determination revealed an unexpectedphosphorylation of Ser198 in LPO, which is also confirmed by anti-phosphoserine antibody binding studies. The structure is also notable forobserving densities for glycan chains at all the four potential glycosylationsites. Caprine LPO consists of a single polypeptide chain of 595 amino acidresidues and folds into an oval-shaped structure. The structure contains 20well-defined α-helices of varying lengths including a helix, H2a, unique toLPO, and two short antiparallel β-strands. The structure confirms that theheme group is covalently linked to the protein through two ester linkagesinvolving carboxylic groups of Glu258 and Asp108 and modified methylgroups of pyrrole rings A and C, respectively. The heme moiety is slightlydistorted from planarity, but pyrrole ring B is distorted considerably.However, an iron atom is displaced only by 0.1 Å from the plane of the hemegroup toward the proximal site. The substrate diffusing channel in LPO iscylindrical in shape with a diameter of approximately 6 Å. Two histidineresidues and six buried water molecules are connected through a hydrogen-bonded chain from the distal heme cavity to the surface of protein moleculeand seemingly form the basis of proton relay for catalytic action. Ten iodideions have been observed in the structure. Out of these, only one iodide ion islocated in the distal heme cavity and is hydrogen bonded to the watermolecule W1. W1 is also hydrogen bonded to the heme iron as well as todistal His109. The structure contains a calcium ion that is coordinated toseven oxygen atoms and forms a typical pentagonal bipyramidalcoordination geometry.
© 2007 Elsevier Ltd. All rights reserved.
Keywords: antimicrobial activity; heme; iodide binding site; peroxidase;crystal structure
Edited by M. Gussess:
sulfonic acid); EPO,ne lactoperoxidase;e; HMPO, humanyroid peroxidase;se; MPO,ucosamine;–polyacrylamide gelidase.
lsevier Ltd. All rights reserve
Introduction
Lactoperoxidase (LPO; EC 1.11.1.7) is a heme-containing glycoprotein with a single chain that hasa molecular mass of ≈68 kDa. It catalyzes theinactivation of a wide range of microorganisms.1–3
The other members of the mammalian peroxidasefamily include eosinophil peroxidase (EPO), thyroidperoxidase (TPO), and myeloperoxidase (MPO).4,5
LPO, EPO, and MPO contribute to the nonimmunehost defense system by oxidizing halide andpseudohalide ions to produce potent antimicrobial
d.

1061Crystal Structure of Caprine Lactoperoxidase
agents. LPO carries out this function in exocrinesecretions including milk, tears, and saliva,2 whileEPO and MPO play similar roles in the phagosomesof eosinophils5 and neutrophils,6 respectively, dur-ing engulfment of microorganisms. The fourthmember of the mammalian peroxidase family, TPOis an intracellular membrane-bound protein, whichis involved in the catalysis of the iodination andcoupling of thyroglobulin moieties in the biosynth-esis of thyroid hormones thyroxine and triiodothyr-onine. The mammalian peroxidases also participatein the oxidative metabolism of xenobiotics respon-sible for hypersensitivity reactions and other toxicsequelae.6–8 As reported in the literature, LPO,9,10EPO,11,12 and TPO13 are monomeric proteins whileMPO14–17 is a covalently linked dimer of two iden-tical halves each consisting of two polypeptidechains of 108 and 466 amino acid residues as aresult of a posttranslational deletion of 6 amino acidresidues. In contrast, LPO is a single chain of 595amino acid residues with an extension of 12 and2 residues with respect to MPO at the N- andC-termini, respectively. There are four potential gly-cosylation sites in LPO and EPO, while MPO andTPO have five glycosylation sites in each. There are15 cysteine residues in LPO, 14 each in MPO andEPO, while TPO contains 17 cysteine residues. Allthe four members of mammalian peroxidases con-tain a covalently linked heme group, which is aderivative of protoporphyrin IX. In LPO, EPO, andTPO, the heme is involved in two ester linkagesthrough Glu258 and Asp108 (the numbering schemeused is that of LPO),13,18–22 whereas MPO containsan additional linkage involving the sulfonium ion ofMet243. The residues corresponding to Met243 ofMPO are Gln, Thr, and Val in LPO, EPO, and TPO,respectively. The amino acid sequence identities ofLPO with MPO, EPO, and TPO are in the range of50–56%. So far, structural information is availableonly on MPO.15–17 Here, we present the first crystalstructure of LPO at 2.4 Å resolution.
Results
Sequence analysis
The complete amino acid sequence determinationof caprine (goat) LPO (GLPO) (EF363153) showsthat it is a single-chain polypeptide of 595 aminoacid residues including 15 cysteines. The sequenceof 20 amino acid residues from the N-terminus ofGLPO determined using automated protein sequen-cer established the correct sequence of the startingN-terminal residue of the matured protein. Theamino acid sequence of GLPO has been comparedwith sequences of LPOs from other species (Fig. 1a)and with those of human MPO (HMPO), humanEPO (HEPO), and human TPO (HTPO) (Fig. 1b).The GLPO sequence has four potential glycosylationsites with Asn-X-Ser/Thr sequence motifs, whichare at Asn95, Asn205, Asn241, and Asn332. It con-
tains 15 cysteine residues to form seven disulfidebridges between Cys6 and Cys167, between Cys15and Cys28, between Cys129 and Cys139, betweenCys133 and Cys157, between Cys237 and Cys248,between Cys456 and Cys573, and between Cys554and Cys579. Cys441 is unpaired. Although the struc-ture of canine MPO is also known,15 for comparisonpurposes, the 1.8-Å-resolution HMPO structure willbe used. It is important to note that GLPO is amonomeric protein whereas HMPO is a covalentlylinked homodimer. It is also pertinent to note herethat the sequence of matured HMPO (Fig. 1b) doesnot contain the first 12 residues of GLPO, which alsoinclude an important cysteine at position 6. Appar-ently, Cys6 is responsible for the monomeric form ofGLPO because it forms an intramolecular disulfidebridge with Cys167. Since Cys6 is absent in HMPO,the residue corresponding to Cys167 (Cys153) inHMPO forms an intermolecular disulfide bridgewith the same residue of the second molecule,resulting in the formation of a covalently linkedhomodimer. HEPO is reported to be a monomericprotein11,12 (Fig. 1b), although the absence of Cys6from its N-terminal region seems to suggest that itcould be a dimeric structure similar to that ofHMPO.16,17 Unfortunately, the crystal structure ofHEPO is not yet known. There is an excision of ahexapeptide inHMPO as the first chain terminates atresidue number 106 and the second chain begins at113. Thus, its dimeric half contains two polypeptidechains of 108 (after deletions) and 466 amino acidresidues. All the four peroxidases contain onecovalently linked heme group per monomer. How-ever, the number of covalent linkages in HMPO isthree, while in other peroxidases, there are only twocovalent bonds. The additional covalent bond inHMPO is provided by Met243 through the interac-tion between the sulfonium ion and the terminal β-carbon of the vinyl group on pyrrole ring A. Theresidue corresponding to Met243 is not methioninein other peroxidases (Fig. 1b). There is yet anotherunexpected difference between the sequences ofGLPO and HMPO where Ser198 is found to bephosphorylated in GLPO, while the correspondingresidue in HMPO is proline and, hence, cannot bephosphorylated.
Quality of the final model
The final model of GLPO consists of an entirepolypeptide chain with residues from 1 to 595, onecovalently linked heme group, a calcium ion, fourglycan chains containing 8 N-acetyl glucosamine(NAG) and 3 mannose (MAN) residues, and 421water oxygen atoms. The overall mean B-factor is27.3 Å2. The entire structure is well defined. ARamachandran plot of the main-chain torsion angles(ϕ,ψ)23 shows that 86.5%of the residues are in the coreregions as defined in the program PROCHECK.24
The 11 sugar residues included in the model areremarkably well defined in the electron density map.Four hundred twenty-one water positions fulfill thecriteria of good electron density in the 2Fo−Fc map at

1062 Crystal Structure of Caprine Lactoperoxidase
2.5σ cutoff and interactions with protein atoms orwith other water molecules.
Overall molecular structure
The structural organization for the polypeptidechain of GLPO is shown as a ribbon diagram inFig. 2. The monomeric structure is largely α-helicalwith only two small antiparallel β-strands (residues357–359 and residues 373–375). The slightly elon-gated molecule of GLPO is packed with at least 20α-helices of varying lengths, while the structureof HMPO in its dimeric half contains only 19α-helices.16,17 An important α-helix, H2a (residues124–133) is absent in HMPO, whereas it is one of thevery important features of GLPO. Four out of sixresidues of the deleted hexapeptide in HMPO arepart of helix H2a. The central core of the moleculeconsists of five long α-helices, H2, H5, H6, H8, andH12, with a covalently attached heme group. TheN-terminus of GLPO does not form any repetitivesecondary structure until residue 75 of helix H1. H1is a short helix and is connected to H2 through a longchain. Helix H2 is a part of the core, and one of theresidues of this helix, Asp108, is covalently linked tothe heme group. Helix H2 is connected to H2a (124–133) through an extended chain. In HMPO, theresidues corresponding to 119 and 120 were notobserved in the electron density map while the nextsix residues are absent from the sequence. As aresult, helix H2a is absent in HMPO. Helix H2a isalmost perpendicularly aligned to the plane of theheme moiety and is located in the proximity of thesubstrate channel. This helix is followed by twoshort helices, H3 and H4. Helices H5 and H6 areconnected with each other by a V-shaped loop that isflanked by two extended chains. A core helix, H8,forms a triangle with helices H5 and H6. The hemegroup is sandwiched between helices H2 and H8.This is connected to the region consisting of helicesH13, H14, H15, and H16 and parts of helices H17, H18,and H19. This region represents the crown of theback face of the core region. The three helices H2, H5,and H6 also form a triangle, below which lies theheme group. The other two core helices, H8 and H12,which run parallel with each other, form the lowerwall on which the heme moiety rests.
Role of disulfide bonds in the structure of GLPO
The sequence of GLPO contains 15 cysteine resi-dues that form seven intrachain disulfide bonds,Cys6–Cys167, Cys15–Cys28, Cys129–Cys139,Cys133–Cys157, Cys237–Cys248, Cys456–Cys513,and Cys554–Cys579. In the structure of HMPO,there are six intramolecular disulfide bonds and oneintermolecular disulfide bond. It may be noted thatthe disulfide bond formed between Cys6 andCys167 in GLPO tethers the N-terminal segmentand a rigid loop, Phe165–Arg177. In HMPO, theN-terminal segment including Cys6 is absent andthe corresponding loop Pro151–Arg161 containingCys153 is shorter by two residues. As indicated by
B-factors, it is a relatively flexible loop also becauseit interacts poorly with other parts of the protein.Presumably, it helps it to protrude out of the mainbody of the molecule, facilitating the formation of anintermolecular Cys153–Cys153 disulfide bridge,resulting in the formation of a covalently linkedhomodimer.
Heme environment
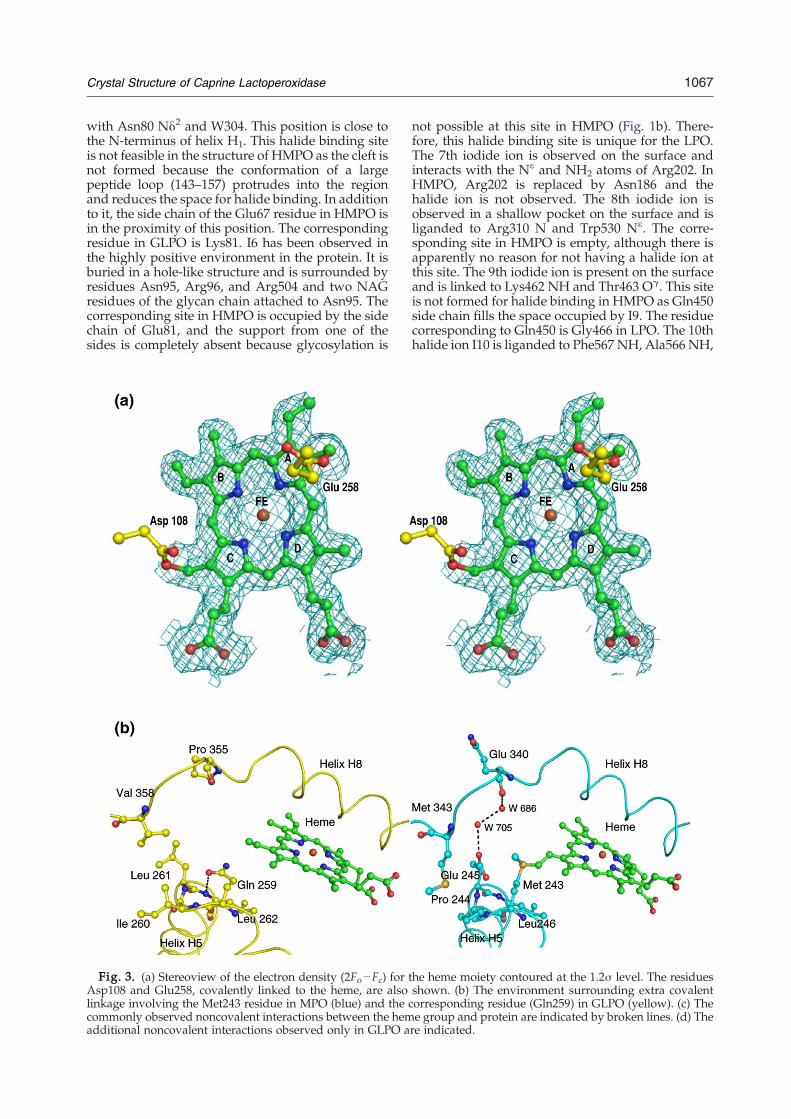
The heme group is a derivative of protoporphyrinIX26 in which the methyl groups on pyrrole rings Aand C are modified to allow formation of esterlinkages with the carboxyl groups of Glu258 andAsp108, respectively (Fig. 3a). Unlike in HMPOwhere Met243 also forms a covalent bond giving riseto a sulfonium ion linkage with the β-carbon of thevinyl group on pyrrole ring A, GLPO lacks thisinteraction because the corresponding residue isglutamine (Fig. 3b). As seen from Fig. 3b, theenvironment in the proximity of Met243 in HMPOfavors it to move closer to the heme group to form acovalent linkage as the adjacent residues Pro244 andGlu245 and the distant residue Met343 do not allowenough space for flexibility. On the other hand, inGLPO, Gln259 turns toward the distal heme cavitywhile the adjacent residues Ile260 and Leu261 movetoward the hydrophobic pocket containing Val358and other hydrophobic residues. The noncovalentinteractions between the protein and the hememoiety that are similar in both GLPO and HMPOare given in Fig. 3c. There are also some noncovalentinteractions between protein residues and the hemegroup in GLPO (Fig. 3d), which are not present inHMPO. This might compensate partly for the lack ofsulfonium ion linkage of HMPO. Gln259 in GLPO isa part of the type III β-turn formed by the segmentGln259-Ile260-Leu261-Leu262. The observed β-turnconformation is stabilized by a hydrogen bondbetween Gln259 O and Leu262 NH. The cornerresidues Ile260 and Leu261 protrude into theadjacent hydrophobic pocket formed with residuesLeu98, Val358, Leu376, Leu379, Phe384, Leu395, andLeu399. Apart from stabilizing the turn conforma-tion, it also enriches the core structure of LPO withhydrophobic interactions. Gln259 is also involved ina hydrogen-bonded network with Gln102 andGln105 in the distal heme region. Gln105 forms ahydrogen bond with the nitrogen atom of pyrrolering B of the heme moiety. Such a hydrogen-bondednetwork and the interaction with the heme groupare absent in HMPO. It may be noted that thesequence of the corresponding segment in HMPO isMet243-Pro244-Glu245-Leu246, which is remark-ably different from that in GLPO. Although it alsoadopts a β-turn conformation, the side chains ofcorner residues Pro244 and Glu245 are in differentorientations. Met243 is covalently linked to the hemegroup. Pro244 is not involved in any specificinteraction, while Glu245 forms a hydrogen bondwith its own NH group. The corresponding Leu262in GLPO is a part of one of the sides of the distalheme cavity. The heme moiety is deeply buried

1063Crystal Structure of Caprine Lactoperoxidase
inside the protein molecule, while the heme cavity issurrounded by a number of helices from three sides.It is open only from one side. The two strands, S1
Fig. 1 (legend o
and S2 (Fig. 2), are situated on the upper portion ofthe opening to the heme cavity. Apart from the twostrands, H4 and H9 are the two short helices that are
n next page)
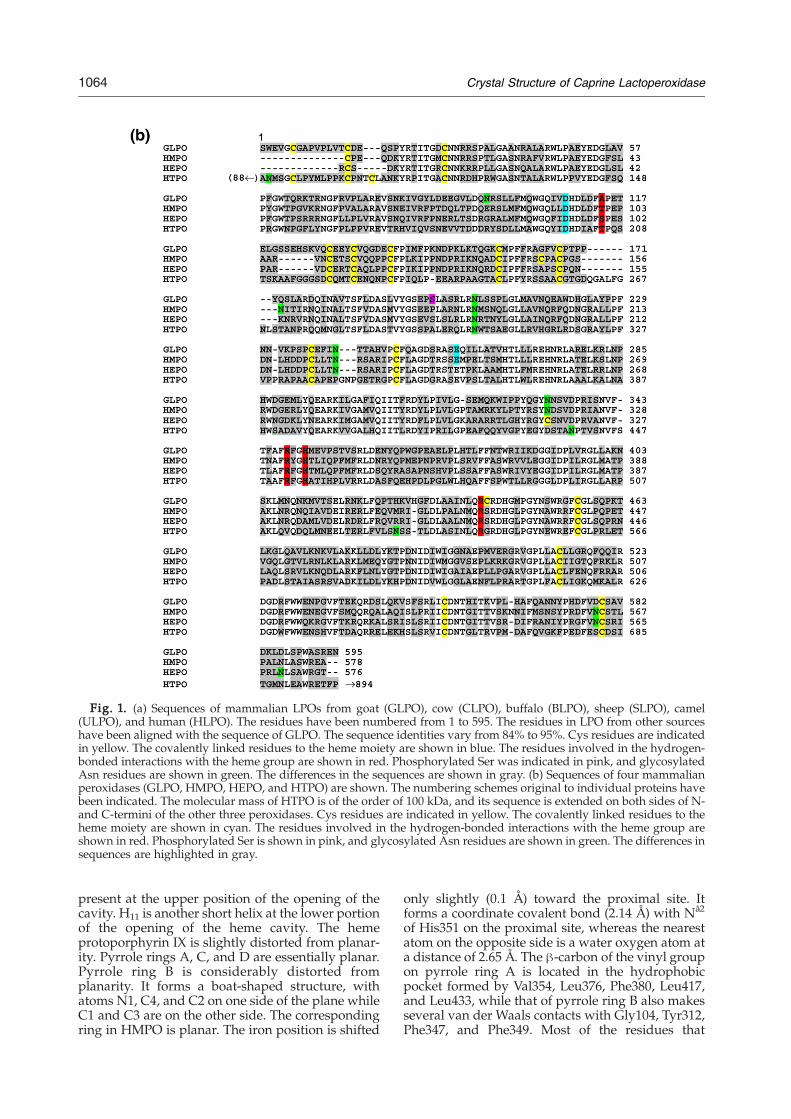
Fig. 1. (a) Sequences of mammalian LPOs from goat (GLPO), cow (CLPO), buffalo (BLPO), sheep (SLPO), camel(ULPO), and human (HLPO). The residues have been numbered from 1 to 595. The residues in LPO from other sourceshave been aligned with the sequence of GLPO. The sequence identities vary from 84% to 95%. Cys residues are indicatedin yellow. The covalently linked residues to the heme moiety are shown in blue. The residues involved in the hydrogen-bonded interactions with the heme group are shown in red. Phosphorylated Ser was indicated in pink, and glycosylatedAsn residues are shown in green. The differences in the sequences are shown in gray. (b) Sequences of four mammalianperoxidases (GLPO, HMPO, HEPO, and HTPO) are shown. The numbering schemes original to individual proteins havebeen indicated. The molecular mass of HTPO is of the order of 100 kDa, and its sequence is extended on both sides of N-and C-termini of the other three peroxidases. Cys residues are indicated in yellow. The covalently linked residues to theheme moiety are shown in cyan. The residues involved in the hydrogen-bonded interactions with the heme group areshown in red. Phosphorylated Ser is shown in pink, and glycosylated Asn residues are shown in green. The differences insequences are highlighted in gray.
1064 Crystal Structure of Caprine Lactoperoxidase
present at the upper position of the opening of thecavity. H11 is another short helix at the lower portionof the opening of the heme cavity. The hemeprotoporphyrin IX is slightly distorted from planar-ity. Pyrrole rings A, C, and D are essentially planar.Pyrrole ring B is considerably distorted fromplanarity. It forms a boat-shaped structure, withatoms N1, C4, and C2 on one side of the plane whileC1 and C3 are on the other side. The correspondingring in HMPO is planar. The iron position is shifted
only slightly (0.1 Å) toward the proximal site. Itforms a coordinate covalent bond (2.14 Å) with Nå2
of His351 on the proximal site, whereas the nearestatom on the opposite side is a water oxygen atom ata distance of 2.65 Å. The β-carbon of the vinyl groupon pyrrole ring A is located in the hydrophobicpocket formed by Val354, Leu376, Phe380, Leu417,and Leu433, while that of pyrrole ring B also makesseveral van der Waals contacts with Gly104, Tyr312,Phe347, and Phe349. Most of the residues that
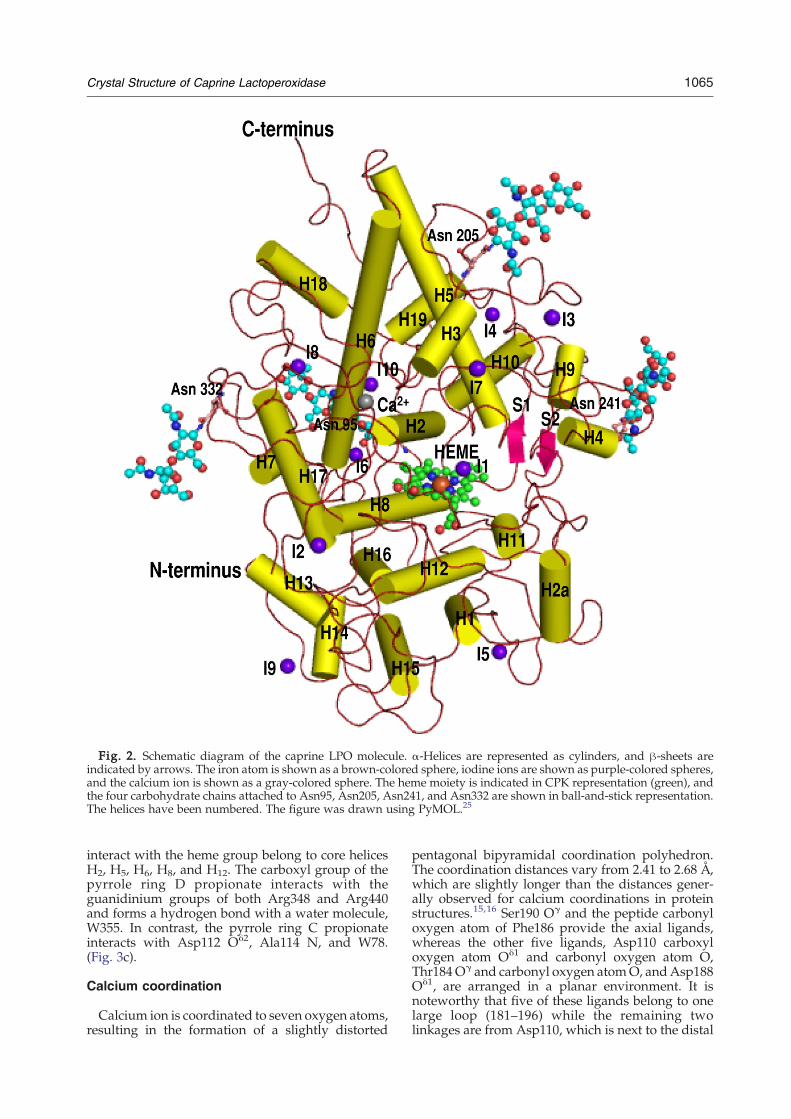
Fig. 2. Schematic diagram of the caprine LPO molecule. α-Helices are represented as cylinders, and β-sheets areindicated by arrows. The iron atom is shown as a brown-colored sphere, iodine ions are shown as purple-colored spheres,and the calcium ion is shown as a gray-colored sphere. The heme moiety is indicated in CPK representation (green), andthe four carbohydrate chains attached to Asn95, Asn205, Asn241, and Asn332 are shown in ball-and-stick representation.The helices have been numbered. The figure was drawn using PyMOL.25
1065Crystal Structure of Caprine Lactoperoxidase
interact with the heme group belong to core helicesH2, H5, H6, H8, and H12. The carboxyl group of thepyrrole ring D propionate interacts with theguanidinium groups of both Arg348 and Arg440and forms a hydrogen bond with a water molecule,W355. In contrast, the pyrrole ring C propionateinteracts with Asp112 Oδ2, Ala114 N, and W78.(Fig. 3c).
Calcium coordination
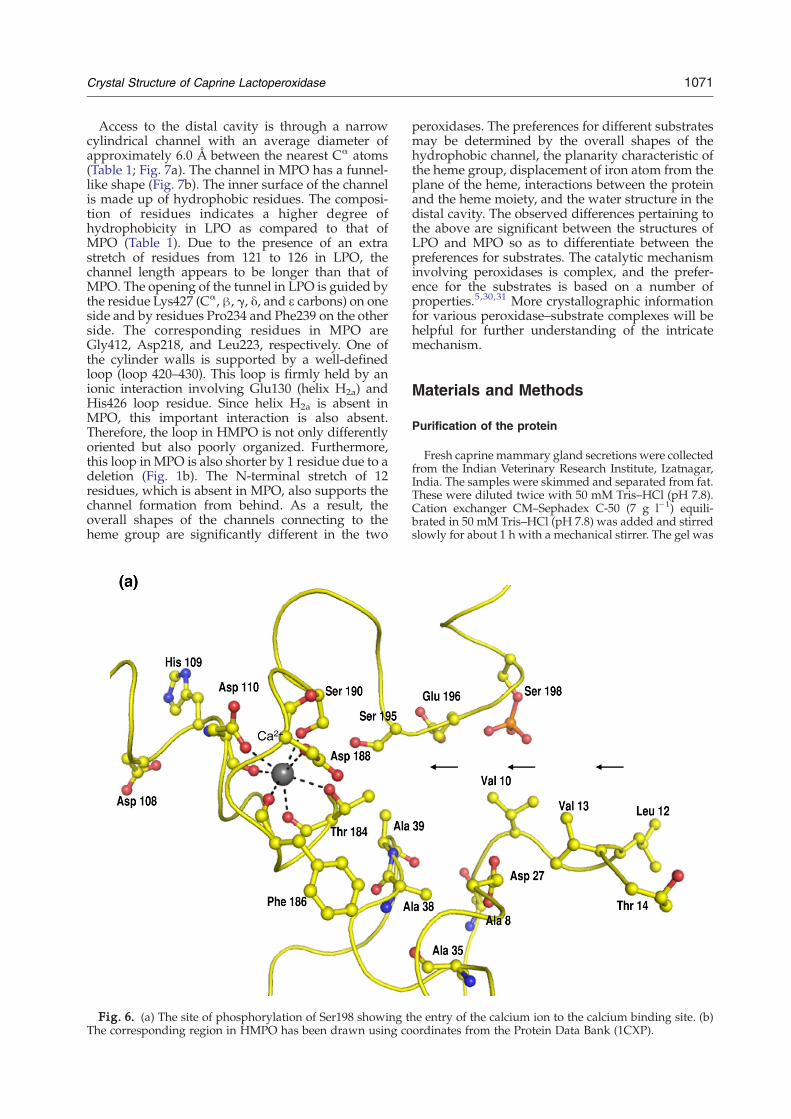
Calcium ion is coordinated to seven oxygen atoms,resulting in the formation of a slightly distorted
pentagonal bipyramidal coordination polyhedron.The coordination distances vary from 2.41 to 2.68 Å,which are slightly longer than the distances gener-ally observed for calcium coordinations in proteinstructures.15,16 Ser190 Oγ and the peptide carbonyloxygen atom of Phe186 provide the axial ligands,whereas the other five ligands, Asp110 carboxyloxygen atom Oδ1 and carbonyl oxygen atom O,Thr184Oγ and carbonyl oxygen atomO, andAsp188Oδ1, are arranged in a planar environment. It isnoteworthy that five of these ligands belong to onelarge loop (181–196) while the remaining twolinkages are from Asp110, which is next to the distal

1066 Crystal Structure of Caprine Lactoperoxidase
His109 in the sequence, indicating the significance ofcalcium ion coordination in the structure andfunction of LPO.
Proximal interacting residues
The proximal side of the heme moiety shows thatthe amino acid side chains from two of the corehelices, H8 and H12 (Fig. 2), are involved ininteractions. His351 Nε2 of helix H8 is coordinatedto the iron atom (2.14 Å), while Nδ1 forms ahydrogen bond with Asn437 Oδ1 (2.78 Å) fromhelix H12. Asn437 O forms a hydrogen bond withpeptide NH of Cys441. The side chain of Cys441 isfree in the structure of LPO and is buried in thehydrophobic pocket built by Phe345, Ala434,Leu438, and Trp493. In contrast, the correspondingsite is significantly less hydrophobic in HMPO as ithas not only Asn345 and Ser434 in place of Phe345and Ala434 but also serine at position 441. Thisshows that Cys441 is very tightly packed in thehydrophobic pocket and is protected well fromgetting oxidized. However, the significance of freeCys441 is not yet clearly understood. The carboxylicgroup of the ring D propionate not only interactswith the guanidinium groups of both Arg348 andArg440 but also forms a hydrogen bond with awater molecule. The ring C propionate interactswith Asp112 Oδ2, Ala114 N, and W38. Gln423interacts with the carboxyl oxygen of the propionatethrough water molecule W355. This interaction isabsent in HMPO as the corresponding residue isGlu423 and its side chain is oriented in the oppositedirection. It is interesting to note that Cys441 with afree side chain is located in the region of proximalcavity.
Distal heme cavity
The distal heme cavity is located on the oppositeside of the proximal cavity and is known to be thesite of catalysis in heme-containing peroxidases. Itextends from the heme pyrrole rings to the surface ofthe protein in a well-formed channel. Its sides aremarked with the side chains of Phe113, Leu199,Asn230, Pro234, Pro236, Phe381, and Pro424. It isremarkable to observe that helix H2a (124–133) isinvolved in stabilizing the shape of the distal hemecavity through interactions with loops that are partof the cavity walls. Helix H2a is a very stablestructure with two disulfide bridges through Cys129and Cys133. The helix corresponding to H2a is notpresent in HMPO.16 Instead, it forms a loop with avery different orientation. There is another loop,420–430, which is of considerable interest because ithas a remarkably different conformation from thatof the corresponding segment in HMPO. This loopsupports a part of one of the sides of the distalcavity, resulting in the difference in the shape ofdistal cavities of GLPO and HMPO. The residue at430 is Gly in both structures, which can impartflexibility to this loop. The residue next to it isHis429 in GLPO, which forms a hydrogen bond
(His429Nε2⋯His377 O=2.81 Å) with a well-orderedloop structure containing His377. The residuecorresponding to His429 is absent in HMPO. Yetanother important interaction is observed in GLPObetween His426 and Glu130 Oε1 (His426 Nδ1⋯Glu130 Oδ1=2.51 Å). Glu130 is part of a very stablehelix, H2a. On the other hand, the residue corre-sponding to His426 is Met411 in HMPO. Loop 420–430 is also characterized by a tight β-turn for theresidues Gln423-Pro424-Thr425-His426. In thisarrangement, Gln423 turns toward the heme groupand interacts with the propionate carboxylate ofpyrrole ring A through a water molecule. It alsoimparts structural uniqueness to the conformationof loop 420–430 as well as to the heme moiety. It isclearly shows that the interactions involving theheme group (Fig. 3c) and the residues that con-tribute to the size and shape of the distal cavity arenot identical in GLPO and HMPO. The channel onthe side of the distal heme cavity is filled with watermolecules as well as with the side chains of Gln105,His109, and Arg255. In the present structure, thedistal heme cavity contains one iodide ion I1, whichis hydrogen bonded to W1, which, in turn, is hy-drogen bonded to the heme iron atom and distalHis109 Nε2. His109 Nδ1 forms a very strong hy-drogen bond (2.45 Å) with W2. W2 forms two otherhydrogen bonds, one with His266 Nε2 and the otherwith Asp253 Oδ2. His266 Nδ1 forms another hydro-gen bond with W3, which is involved in two otherhydrogen bonds with W4 and His266 O. W4 ishydrogen bonded to W5, which is further hydrogenbonded to W6 and Gln250 O. W6 forms two hy-drogen bonds with surface residues Ala214 O andLeu203 O. The linking of Asn437–His351 (on theproximal site)–heme iron–W1–His109–W2–His266–W3–W4–W5–W6–Ala214/Leu203 (Fig. 4) is animportant structural feature of peroxidase enzymes,which facilitates proton relay from distal histidineaway to the surface of the protein.
Characterization of halide binding
The solution of native protein was incubated inthe protein buffer solution containing 2 mM NH4Ifor 24 h. The iodide-saturated GLPO was crystal-lized. The structure revealed 10 positions of iodideions (I1–I10) (Fig. 2). Out of 10 iodide ions, I1 isfound in the distal heme cavity. It is liganded toHis109 and three water molecules including W1. Asimilar halide position has also been observed inHMPO.16 A second iodide has been located in theproximal site, which is hydrogen bonded to thepeptide NH groups of Trp46 and Val342. It may benoted that this site is located in the proximity of theamino-terminus of the core helix H8. The 3rd and 4thhalide ions are found in shallow clefts on the surfaceof the protein but are liganded well to the proteinNH/NH2 groups and water molecules. All thesehalide positions were also reported in the HMPOstructure.16 Additional halide ions (I5–I10) havebeen observed in the LPO structure. I5 is observed ina cleft on the surface. It forms two hydrogen bonds
1067Crystal Structure of Caprine Lactoperoxidase
with Asn80 Nδ2 and W304. This position is close tothe N-terminus of helix H1. This halide binding siteis not feasible in the structure of HMPO as the cleft isnot formed because the conformation of a largepeptide loop (143–157) protrudes into the regionand reduces the space for halide binding. In additionto it, the side chain of the Glu67 residue in HMPO isin the proximity of this position. The correspondingresidue in GLPO is Lys81. I6 has been observed inthe highly positive environment in the protein. It isburied in a hole-like structure and is surrounded byresidues Asn95, Arg96, and Arg504 and two NAGresidues of the glycan chain attached to Asn95. Thecorresponding site in HMPO is occupied by the sidechain of Glu81, and the support from one of thesides is completely absent because glycosylation is
Fig. 3. (a) Stereoview of the electron density (2Fo−Fc) for tAsp108 and Glu258, covalently linked to the heme, are alsolinkage involving the Met243 residue in MPO (blue) and the ccommonly observed noncovalent interactions between the hemadditional noncovalent interactions observed only in GLPO a
not possible at this site in HMPO (Fig. 1b). There-fore, this halide binding site is unique for the LPO.The 7th iodide ion is observed on the surface andinteracts with the Nε and NH2 atoms of Arg202. InHMPO, Arg202 is replaced by Asn186 and thehalide ion is not observed. The 8th iodide ion isobserved in a shallow pocket on the surface and isliganded to Arg310 N and Trp530 Nε. The corre-sponding site in HMPO is empty, although there isapparently no reason for not having a halide ion atthis site. The 9th iodide ion is present on the surfaceand is linked to Lys462 NH and Thr463 Oγ. This siteis not formed for halide binding in HMPO as Gln450side chain fills the space occupied by I9. The residuecorresponding to Gln450 is Gly466 in LPO. The 10thhalide ion I10 is liganded to Phe567 NH, Ala566 NH,
he heme moiety contoured at the 1.2σ level. The residuesshown. (b) The environment surrounding extra covalentorresponding residue (Gln259) in GLPO (yellow). (c) Thee group and protein are indicated by broken lines. (d) There indicated.
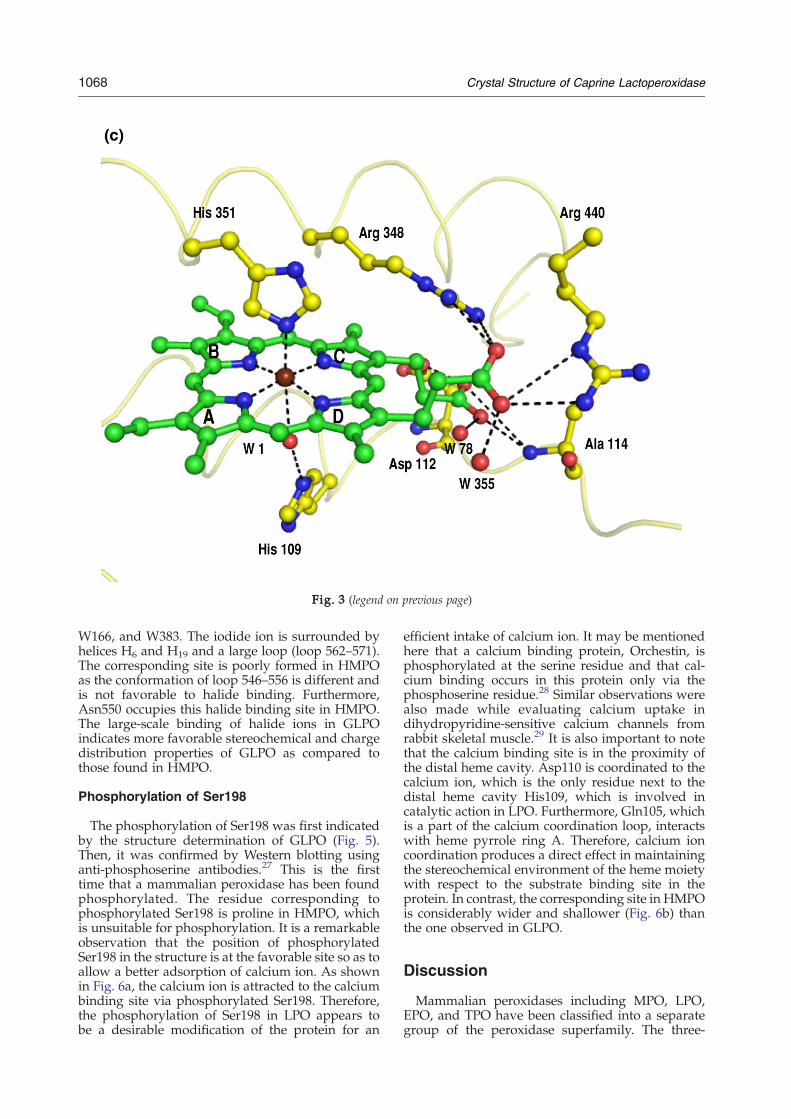
Fig. 3 (legend on previous page)
1068 Crystal Structure of Caprine Lactoperoxidase
W166, and W383. The iodide ion is surrounded byhelices H6 and H19 and a large loop (loop 562–571).The corresponding site is poorly formed in HMPOas the conformation of loop 546–556 is different andis not favorable to halide binding. Furthermore,Asn550 occupies this halide binding site in HMPO.The large-scale binding of halide ions in GLPOindicates more favorable stereochemical and chargedistribution properties of GLPO as compared tothose found in HMPO.
Phosphorylation of Ser198
The phosphorylation of Ser198 was first indicatedby the structure determination of GLPO (Fig. 5).Then, it was confirmed by Western blotting usinganti-phosphoserine antibodies.27 This is the firsttime that a mammalian peroxidase has been foundphosphorylated. The residue corresponding tophosphorylated Ser198 is proline in HMPO, whichis unsuitable for phosphorylation. It is a remarkableobservation that the position of phosphorylatedSer198 in the structure is at the favorable site so as toallow a better adsorption of calcium ion. As shownin Fig. 6a, the calcium ion is attracted to the calciumbinding site via phosphorylated Ser198. Therefore,the phosphorylation of Ser198 in LPO appears tobe a desirable modification of the protein for an
efficient intake of calcium ion. It may be mentionedhere that a calcium binding protein, Orchestin, isphosphorylated at the serine residue and that cal-cium binding occurs in this protein only via thephosphoserine residue.28 Similar observations werealso made while evaluating calcium uptake indihydropyridine-sensitive calcium channels fromrabbit skeletal muscle.29 It is also important to notethat the calcium binding site is in the proximity ofthe distal heme cavity. Asp110 is coordinated to thecalcium ion, which is the only residue next to thedistal heme cavity His109, which is involved incatalytic action in LPO. Furthermore, Gln105, whichis a part of the calcium coordination loop, interactswith heme pyrrole ring A. Therefore, calcium ioncoordination produces a direct effect in maintainingthe stereochemical environment of the heme moietywith respect to the substrate binding site in theprotein. In contrast, the corresponding site in HMPOis considerably wider and shallower (Fig. 6b) thanthe one observed in GLPO.
Discussion
Mammalian peroxidases including MPO, LPO,EPO, and TPO have been classified into a separategroup of the peroxidase superfamily. The three-
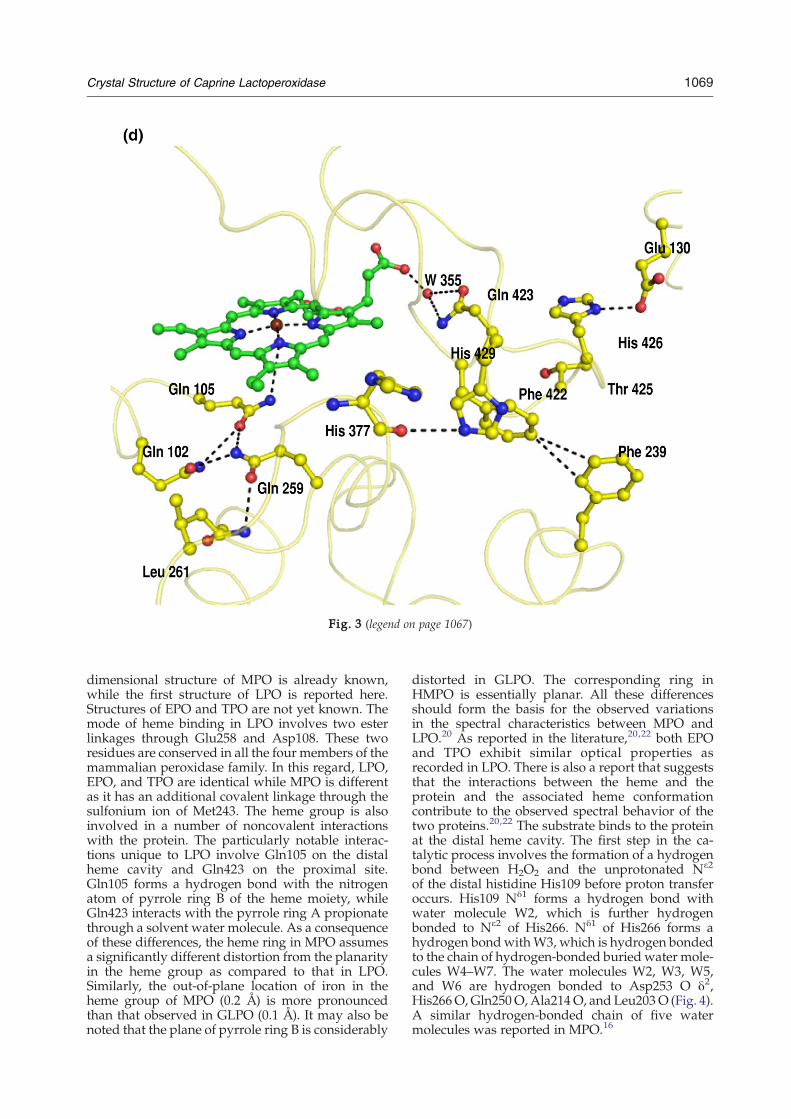
Fig. 3 (legend on page 1067)
1069Crystal Structure of Caprine Lactoperoxidase
dimensional structure of MPO is already known,while the first structure of LPO is reported here.Structures of EPO and TPO are not yet known. Themode of heme binding in LPO involves two esterlinkages through Glu258 and Asp108. These tworesidues are conserved in all the four members of themammalian peroxidase family. In this regard, LPO,EPO, and TPO are identical while MPO is differentas it has an additional covalent linkage through thesulfonium ion of Met243. The heme group is alsoinvolved in a number of noncovalent interactionswith the protein. The particularly notable interac-tions unique to LPO involve Gln105 on the distalheme cavity and Gln423 on the proximal site.Gln105 forms a hydrogen bond with the nitrogenatom of pyrrole ring B of the heme moiety, whileGln423 interacts with the pyrrole ring A propionatethrough a solvent water molecule. As a consequenceof these differences, the heme ring in MPO assumesa significantly different distortion from the planarityin the heme group as compared to that in LPO.Similarly, the out-of-plane location of iron in theheme group of MPO (0.2 Å) is more pronouncedthan that observed in GLPO (0.1 Å). It may also benoted that the plane of pyrrole ring B is considerably
distorted in GLPO. The corresponding ring inHMPO is essentially planar. All these differencesshould form the basis for the observed variationsin the spectral characteristics between MPO andLPO.20 As reported in the literature,20,22 both EPOand TPO exhibit similar optical properties asrecorded in LPO. There is also a report that suggeststhat the interactions between the heme and theprotein and the associated heme conformationcontribute to the observed spectral behavior of thetwo proteins.20,22 The substrate binds to the proteinat the distal heme cavity. The first step in the ca-talytic process involves the formation of a hydrogenbond between H2O2 and the unprotonated Nε2
of the distal histidine His109 before proton transferoccurs. His109 Nδ1 forms a hydrogen bond withwater molecule W2, which is further hydrogenbonded to Nε2 of His266. Nδ1 of His266 forms ahydrogen bondwithW3, which is hydrogen bondedto the chain of hydrogen-bonded buried water mole-cules W4–W7. The water molecules W2, W3, W5,and W6 are hydrogen bonded to Asp253 O δ2,His266O, Gln250O,Ala214O, and Leu203O (Fig. 4).A similar hydrogen-bonded chain of five watermolecules was reported in MPO.16
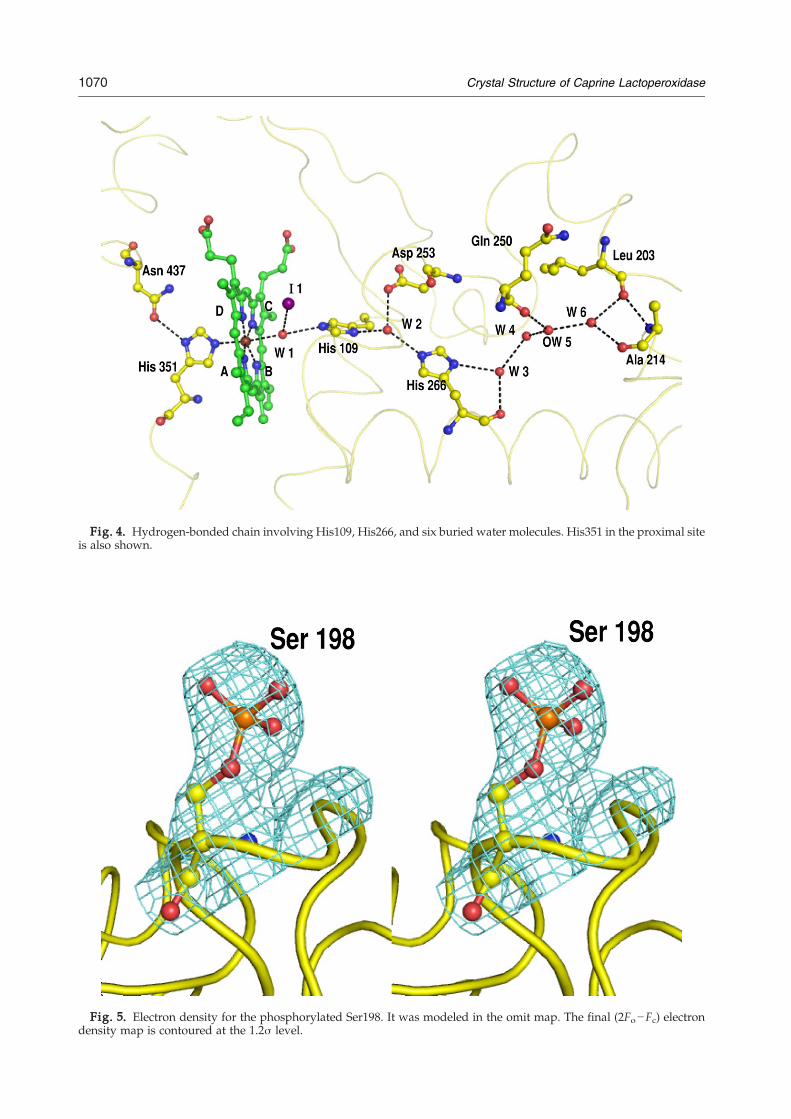
Fig. 4. Hydrogen-bonded chain involving His109, His266, and six buried water molecules. His351 in the proximal siteis also shown.
Fig. 5. Electron density for the phosphorylated Ser198. It was modeled in the omit map. The final (2Fo−Fc) electrondensity map is contoured at the 1.2σ level.
1070 Crystal Structure of Caprine Lactoperoxidase
1071Crystal Structure of Caprine Lactoperoxidase
Access to the distal cavity is through a narrowcylindrical channel with an average diameter ofapproximately 6.0 Å between the nearest Cα atoms(Table 1; Fig. 7a). The channel in MPO has a funnel-like shape (Fig. 7b). The inner surface of the channelis made up of hydrophobic residues. The composi-tion of residues indicates a higher degree ofhydrophobicity in LPO as compared to that ofMPO (Table 1). Due to the presence of an extrastretch of residues from 121 to 126 in LPO, thechannel length appears to be longer than that ofMPO. The opening of the tunnel in LPO is guided bythe residue Lys427 (Cα, β, γ, δ, and ε carbons) on oneside and by residues Pro234 and Phe239 on the otherside. The corresponding residues in MPO areGly412, Asp218, and Leu223, respectively. One ofthe cylinder walls is supported by a well-definedloop (loop 420–430). This loop is firmly held by anionic interaction involving Glu130 (helix H2a) andHis426 loop residue. Since helix H2a is absent inMPO, this important interaction is also absent.Therefore, the loop in HMPO is not only differentlyoriented but also poorly organized. Furthermore,this loop in MPO is also shorter by 1 residue due to adeletion (Fig. 1b). The N-terminal stretch of 12residues, which is absent in MPO, also supports thechannel formation from behind. As a result, theoverall shapes of the channels connecting to theheme group are significantly different in the two
Fig. 6. (a) The site of phosphorylation of Ser198 showing tThe corresponding region in HMPO has been drawn using co
peroxidases. The preferences for different substratesmay be determined by the overall shapes of thehydrophobic channel, the planarity characteristic ofthe heme group, displacement of iron atom from theplane of the heme, interactions between the proteinand the heme moiety, and the water structure in thedistal cavity. The observed differences pertaining tothe above are significant between the structures ofLPO and MPO so as to differentiate between thepreferences for substrates. The catalytic mechanisminvolving peroxidases is complex, and the prefer-ence for the substrates is based on a number ofproperties.5,30,31 More crystallographic informationfor various peroxidase–substrate complexes will behelpful for further understanding of the intricatemechanism.
Materials and Methods
Purification of the protein
Fresh caprine mammary gland secretions were collectedfrom the Indian Veterinary Research Institute, Izatnagar,India. The samples were skimmed and separated from fat.These were diluted twice with 50 mM Tris–HCl (pH 7.8).Cation exchanger CM–Sephadex C-50 (7 g l−1) equili-brated in 50 mM Tris–HCl (pH 7.8) was added and stirredslowly for about 1 h with a mechanical stirrer. The gel was
he entry of the calcium ion to the calcium binding site. (b)ordinates from the Protein Data Bank (1CXP).
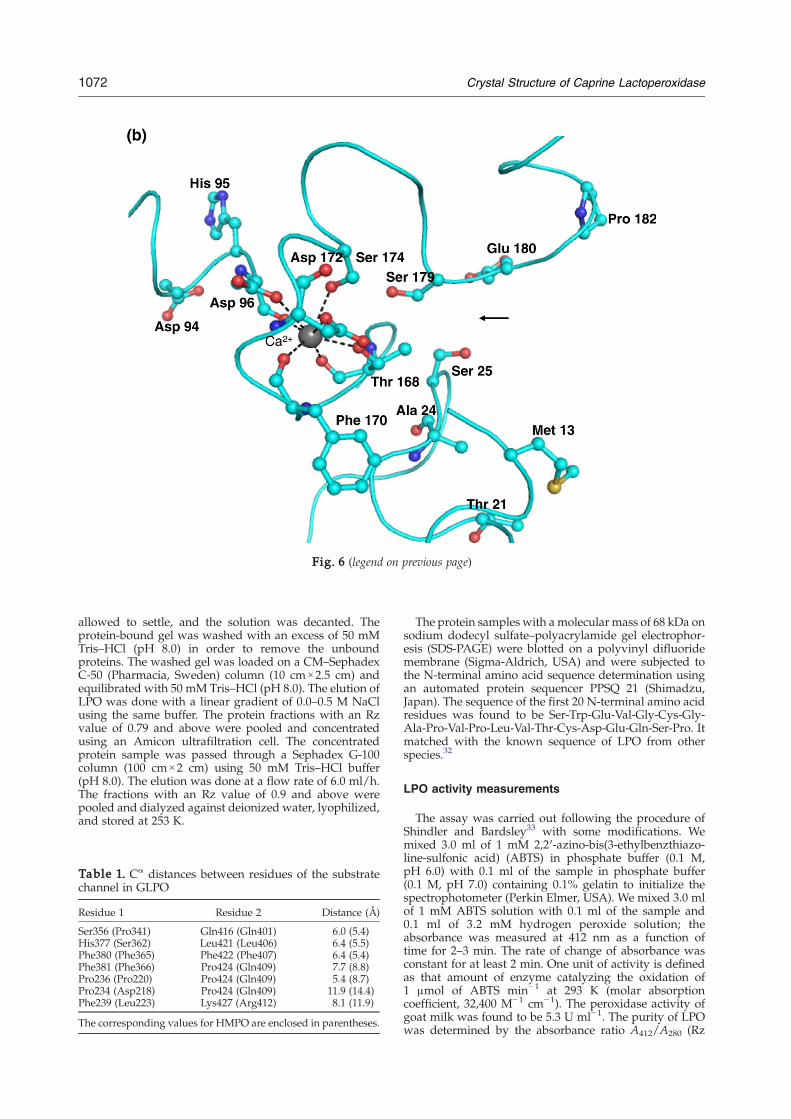
Fig. 6 (legend on previous page)
1072 Crystal Structure of Caprine Lactoperoxidase
allowed to settle, and the solution was decanted. Theprotein-bound gel was washed with an excess of 50 mMTris–HCl (pH 8.0) in order to remove the unboundproteins. The washed gel was loaded on a CM–SephadexC-50 (Pharmacia, Sweden) column (10 cm×2.5 cm) andequilibrated with 50 mM Tris–HCl (pH 8.0). The elution ofLPO was done with a linear gradient of 0.0–0.5 M NaClusing the same buffer. The protein fractions with an Rzvalue of 0.79 and above were pooled and concentratedusing an Amicon ultrafiltration cell. The concentratedprotein sample was passed through a Sephadex G-100column (100 cm×2 cm) using 50 mM Tris–HCl buffer(pH 8.0). The elution was done at a flow rate of 6.0 ml/h.The fractions with an Rz value of 0.9 and above werepooled and dialyzed against deionized water, lyophilized,and stored at 253 K.
Table 1. Cα distances between residues of the substratechannel in GLPO
Residue 1 Residue 2 Distance (Å)
Ser356 (Pro341) Gln416 (Gln401) 6.0 (5.4)His377 (Ser362) Leu421 (Leu406) 6.4 (5.5)Phe380 (Phe365) Phe422 (Phe407) 6.4 (5.4)Phe381 (Phe366) Pro424 (Gln409) 7.7 (8.8)Pro236 (Pro220) Pro424 (Gln409) 5.4 (8.7)Pro234 (Asp218) Pro424 (Gln409) 11.9 (14.4)Phe239 (Leu223) Lys427 (Arg412) 8.1 (11.9)
The corresponding values for HMPO are enclosed in parentheses.
The protein samples with a molecular mass of 68 kDa onsodium dodecyl sulfate–polyacrylamide gel electrophor-esis (SDS-PAGE) were blotted on a polyvinyl difluoridemembrane (Sigma-Aldrich, USA) and were subjected tothe N-terminal amino acid sequence determination usingan automated protein sequencer PPSQ 21 (Shimadzu,Japan). The sequence of the first 20 N-terminal amino acidresidues was found to be Ser-Trp-Glu-Val-Gly-Cys-Gly-Ala-Pro-Val-Pro-Leu-Val-Thr-Cys-Asp-Glu-Gln-Ser-Pro. Itmatched with the known sequence of LPO from otherspecies.32
LPO activity measurements
The assay was carried out following the procedure ofShindler and Bardsley33 with some modifications. Wemixed 3.0 ml of 1 mM 2,2′-azino-bis(3-ethylbenzthiazo-line-sulfonic acid) (ABTS) in phosphate buffer (0.1 M,pH 6.0) with 0.1 ml of the sample in phosphate buffer(0.1 M, pH 7.0) containing 0.1% gelatin to initialize thespectrophotometer (Perkin Elmer, USA). We mixed 3.0 mlof 1 mM ABTS solution with 0.1 ml of the sample and0.1 ml of 3.2 mM hydrogen peroxide solution; theabsorbance was measured at 412 nm as a function oftime for 2–3 min. The rate of change of absorbance wasconstant for at least 2 min. One unit of activity is definedas that amount of enzyme catalyzing the oxidation of1 μmol of ABTS min−1 at 293 K (molar absorptioncoefficient, 32,400 M−1 cm−1). The peroxidase activity ofgoat milk was found to be 5.3 U ml−1. The purity of LPOwas determined by the absorbance ratio A412/A280 (Rz

Fig. 7. The substrate channel and the hydrogen-bonded His109 and His266 and buried water molecules involved inproton relay. Loops 118–132, 230–240, 246–248, 375–383, and 420–423 in GLPO are indicated in red (a). The correspondingloops in HMPO (loops 101–118, 214–224, 230–232, 360–368, and 405–415) are shown in red (b). The N-terminal segment1–12 in GLPO is indicated in purple. The dotted line in (b) indicates a break in the chain of HMPO. Helices H2a and H4 inGLPO and H4 in HMPO are also indicated.
1073Crystal Structure of Caprine Lactoperoxidase
value). The Rz value for the purified LPO was found to be0.932.
Complete amino acid sequence determination
The mammary gland tissue of lactating goat wasobtained from the Indian Veterinary Research Institute.The total RNA was extracted by the phenol/chloroformmethod.34 The reaction with Moloney murine leukemiavirus–reverse transcriptase polymerase chain reaction wasused for polymerase chain reaction amplification of thegene. The conserved nucleotide sequences from otherproteins of the peroxidase family32 and the N-terminalsequence of GLPO as obtained using Edman degradationwere used for the design of primers. The nucleotidesequence was carried out on the cloned double-strandedDNA (pGEM-T) using automatic sequencer model ABI-377. The complete nucleotide and deduced amino acidsequences have been deposited in the gene bank withaccession number EF363153.
Phosphorylation detection
Purified LPO was run on SDS-PAGE using 10%polyacrylamide gel as described by Laemmli. The proteinwas transferred from SDS-PAGE to a polyvinyl difluoridemembrane (Sigma-Aldrich, USA) byWestern blotting, andnonspecific binding sites were blocked for 2–4 h at roomtemperature using 5% dried milk, Tris-buffered saline(pH 7.4), and 0.1% Tween 20. The membrane was probedovernight at 4 °C with primary anti-phosphoserine anti-body (1:5000) for the serine phosphorylation site in GLPO.
A secondary horseradish-peroxidase-labeled antibodywas raised in rabbit (goat anti-rabbit immunoglobulin G;Jackson Immunochemicals) in combination with enhancedchemiluminescence detection system (SuperSignal WestPico Chemiluminescent Substrate) to visualize the pri-mary antibodies.
Crystallization
The purified samples of protein were dissolved in0.01 M phosphate buffer (pH 7.0) to a concentration of25 mg/ml. A reservoir solution containing 0.2 M ammo-nium iodide and 20% (w/v) polyethylene glycol 3350 wasprepared. Protein solution (5 μl) was mixed with 5 μl ofreservoir solution to prepare 10 μl of drops for hangingdrop vapor diffusion method. The rectangular, brownishcrystals measuring up to 0.3 mm×0.2 mm×0.2 mm wereobtained after 1 week.
X-ray intensity data collection
The X-ray intensity data were collected at 287 K using a345-mm-diameter MAR Research dtb Imaging platescanner mounted on a Rigaku RU-300 rotating anodeX-ray generator operating at 50 kV and 100 mA. TheOsmic Blue confocal optics was used for focusing CuKαradiation. The data were indexed and scaled using theprograms DENZO and SCALEPACK.35 The crystalsbelong to monoclinic space group P21 with the followingcell parameters: a=54.2 Å, b=80.8 Å, c=77.0 Å, andβ=102.9°. The unit cell contains one molecule in theasymmetric unit. The final data show an overall complete-

Table 2.Data collection and refinement statistics of GLPO
Space group P21Unit cell dimensions
a (Å) 54.2b (Å) 80.8c (Å) 77.0β (°) 102.9
Number of molecules in the unit cell 2Resolution range (Å) 20.0–2.4Total number of measured reflections 141,640Number of unique reflections 24,962Overall completeness of data (%) 97.9Completeness in the highest-resolution shell(2.44–2.40 Å) (%)
90.1
Overall Rsym (%) 12.1Rsym in the highest-resolution shell (%) 32.3Overall I/σ(I) 5.9I/σ(I) in the highest-resolution shell (%) 2.3Rcryst (%) 19.6Rfree (%) 20.3Protein atoms 4757Heme group (1) atoms 43Iodide ions 10Calcium ion 1NAG residues (n=8) (N-linked) 112MAN residues (n=3) (N-linked) 33Water oxygen atoms 421r.m.s.d. in bond lengths (Å) 0.01r.m.s.d. in bond angles (°) 1.8r.m.s.d. in torsion angles (°) 24.1Mean B-factor for main-chain atoms 25.2Mean B-factor for side-chain atoms and waters 28.3Mean B-factor for all atoms 26.9Residues in the most allowed regions (%) 89.4Residues in the additionally allowed regions (%) 9.0Residues in the generously allowed regions (%) 1.6
1074 Crystal Structure of Caprine Lactoperoxidase
ness of 98% with an Rsym of 12.1% for 2.4 Å resolution.Data collection summary is shown in Table 2.
Structure determination and refinement
The structure was determined with molecular replace-ment method based on maximum likelihood in PHASER36
using the coordinates of one molecule of HMPO16 as thesearch model. The rotation and translation search func-tions were calculated, with data between 12.0 and 4.0 Åyielding a clear solution with a distinct peak. The stackingarrangement of molecules in the unit cell for this solutionyielded no unfavorable intermolecular contacts. Thetransformed coordinates using PHASER were subjectedto 20 cycles of rigid-body refinement with REFMAC537
from the CCP4i v4.2 software suit.38 This reduced theRcryst and Rfree factors to 33.4% and 42.6%, respectively(2% of the reflections were used for the calculation of Rfreeand were not included in the refinement). The manualmodel building of the protein using |2Fo−Fc| Fourier and|Fo−Fc| difference Fourier maps was carried out withgraphics program O39 and COOT40 on a Silicon GraphicsO2 Workstation. Further refinement cycles reduced Rcrystand Rfree factors to 0.251 and 0.284, respectively. Thedifference electron density |Fo−Fc| map computed at thisstage indicated the presence of four carbohydrate chains atAsn95 (2 NAG residues), Asn205 (2 NAG residues),Asn241 (2 NAG residues+1 MAN residue), and Asn332(2 NAG residues). Eleven strong peaks corresponding to10 iodide ions and 1 calcium ion were also observed. Theywere included in the subsequent refinement cycles.Further, difference Fourier |Fo−Fc| maps indicated an
extra electron density for the side chain of Ser198 at 3σcutoff into which a phosphorylated Ser side chain wasfitted well and refined (Fig. 6). The subsequent Fourier(2Fo−Fc) and difference Fourier (Fo−Fc) maps revealed thepositions of 421 water oxygen atoms. The refinementfinally converged to Rcryst and Rfree factors with values of0.196 and 0.203, respectively. The final data collection andrefinement statistics are given in Table 2.
Protein Data Bank accession number
The refined atomic coordinates of GLPO have beendeposited in the Protein Data Bank with accession code2R5L.
Acknowledgements
The authors acknowledge financial support fromthe Department of Science and Technology, NewDelhi. T.P.S. thanks the Department of Biotechnol-ogy, New Delhi, for the Distinguished Biotechnolo-gist award. A.K.S. thanks the Council of ScientificIndustrial Research, New Delhi, for having beenawarded a fellowship.
References
1. Soukka, T., Lumikari, M. & Tenovuo, J. (1991).Combined inhibitory effect of lactoferrin and lacto-peroxidase system on the viability of Streptococcusmutans, serotype c. Scand. J. Dent. Res. 99, 390–396.
2. Dionysius, D. A., Grieve, P. A. & Vos, A. C. (1992).Studies on the lactoperoxidase system: reactionkinetics and antibacterial activity using two methodsfor hydrogen peroxide generation. J. Appl. Bacteriol. 72,146–153.
3. Benoy, M. J., Essy, A. K., Sreekumar, B. & Haridas, M.(2000). Thiocyanate mediated antifungal and antibac-terial property of goat milk lactoperoxidase. Life Sci.66, 2433–2439.
4. Ueda, T., Sakamaki, K., Kuroki, T., Yano, I. & Nagata,S. (1997). Molecular cloning and characterization ofthe chromosomal gene for human lactoperoxidase.Eur. J. Biochem. 243, 32–41.
5. Furtmuller, P. G., Zederbauer, M., Jantschko, W.,Helm, J., Bogner, M., Jakopitsch, C. & Obinger, C.(2006). Active site structure and catalytic mechanismsof human peroxidases. Arch. Biochem. Biophys. 445,199–213.
6. Jiang, X., Khursigara, G. & Rubin, R. L. (1994). Trans-formation of lupus-inducing drugs to cytotoxic pro-ducts by activated neutrophils. Science, 266, 810–813.
7. Uetrecht, J. P. (1988). Mechanism of drug-inducedlupus. Chem. Res. Toxicol. 1, 133–143.
8. Chignell, C. F. (1985). Structure–activity relationshipsin the free-radical metabolism of xenobiotics. Environ.Health Perspect. 61, 133–137.
9. Sievers, G. (1981). Structure of milk lactoperoxidase.Evidence for a single polypeptide chain. FEBS Lett.127, 253–256.
10. Langbakk, B. & Flatmark, T. (1989). Lactoperoxidasefrom human colostrum. Biochem. J. 259, 627–631.
11. Bolscher, B. G., Plat, H. & Wever, R. (1984). Some

1075Crystal Structure of Caprine Lactoperoxidase
properties of human eosinophil peroxidase, a compar-ison with other peroxidases. Biochim. Biophys. Acta,784, 177–186.
12. Olsson, I., Persson, A. M., Stromberg, K., Winqvist, I.,Tai, P. C. & Spry, C. J. (1985). Purification of eosinophilperoxidase and studies of biosynthesis and processingin human marrow cells. Blood, 66, 1143–1148.
13. Kimura, S., Kotani, T., McBride, O. W., Umeki, K.,Hirai, K., Nakayama, T. & Ohtaki, S. (1987). Humanthyroid peroxidase: complete cDNA and proteinsequence, chromosome mapping, and identificationof two alternately spliced mRNAs. Proc. Natl Acad. Sci.USA, 84, 5555–5559.
14. Yamada, M. (1982). Myeloperoxidase precursors inhuman myeloid leukemia HL-60 cells. J. Biol. Chem.257, 5980–5982.
15. Zeng, J. & Fenna, R. E. (1992). X-ray crystal structureof canine myeloperoxidase at 3.0 Å resolution. J. Mol.Biol. 226, 185–207.
16. Fiedler, T. J., Davey, C. A. & Fenna, R. E. (2000).X-ray crystal structure and characterization of halide-binding sites of human myeloperoxidase at 1.8 Åresolution. J. Biol. Chem. 275, 11964–11971.
17. Blair-Johnson, M., Fiedler, T. & Fenna, R. (2001).Human myeloperoxidase: structure of a cyanidecomplex and its interaction with bromide and thio-cyanate substrates at 1.9 Å resolution. Biochemistry, 40,13990–13997.
18. DePillis, G. D., Ozaki, S., Kuo, J. M., Maltby, D. A. &Ortiz de Montellano, P. R. (1997). Autocatalyticprocessing of heme by lactoperoxidase produces thenative protein-bound prosthetic group. J. Biol. Chem.272, 8857–8860.
19. Colas, C., Kuo, J. M. & Ortiz de Montellano, P. R.(2002). Asp-225 and Glu-375 in autocatalytic attach-ment of the prosthetic heme group of lactoperoxidase.J. Biol. Chem. 277, 7191–7200.
20. Andersson, L. A., Bylkas, S. A. & Wilson, A. E. (1996).Spectral analysis of lactoperoxidase. J. Biol. Chem. 271,3406–3412.
21. Rae, T. D. & Goff, H. M. (1998). The heme prostheticgroup of lactoperoxidase. Structural characteristicsof heme l and heme l-peptides. J. Biol. Chem. 273,27968–27977.
22. Oxvig, C., Thomsen, A. R., Overgaard,M. T., Sorensen,E. S., Hojrup, P., Bjerrum,M. J. et al. (1999). Biochemicalevidence for heme linkage through esters with Asp-93and Glu-241 in human eosinophil peroxidase. Theester with Asp93 is only partially formed in vivo.J. Biol. Chem. 274, 16953–16958.
23. Ramachandran, G. N. & Sasisekharan, V. (1968). Con-formation of polypeptides and proteins. Adv. ProteinChem. 23, 283–438.
24. Laskowski, R. A., Macarthur, M., Moss, D. &Thornton, J. (1993). PROCHECK: a program to checkstereo chemical quality of protein structures. J. Appl.Crystallogr. 26, 283–290.
25. DeLano, W. L. (2002). The PyMOL Molecular Graphics
System. DeLano Scientific, San Carlos, CA; http://www.pymol.org.
26. Thanabal, V. & La Mar, G. N. (1989). A nuclearOverhauser effect investigation of the molecular andelectronic structure of the heme crevice in lactoper-oxidase. Biochemistry, 28, 7038–7044.
27. Colyer, J. & Wang, J. H. (1991). Dependence of cardiacsarcoplasmic reticulum calcium pump activity on thephosphorylation status of phospholamban. J. Biol.Chem. 266, 17486–17493.
28. Hecker, A., Testeniere, O., Marin, F. & Luquet, G.(2003). Phosphorylation of serine residues is funda-mental for the calcium-binding ability of Orchestin, asoluble matrix protein from crustacean calciumstorage structures. FEBS Lett. 535, 49–54.
29. Nunoki, K., Florio, V. & Catterall, W. A. (1989).Activation of purified calcium channels by stoichio-metric protein phosphorylation. Proc. Natl Acad. Sci.USA, 86, 6816–6820.
30. Ferrari, R. P., Ghibaudi, E.M., Traversa, S., Laurenti, E.,De Gioia, L. & Salmona, M. (1997). Spectroscopic andbinding studies on the interaction of inorganic anionswith lactoperoxidase. J. Inorg. Biochem. 68, 17–26.
31. Wever, R., Kast, W. M., Kasinoedin, J. H. & Boelens, R.(1982). The peroxidation of thiocyanate catalysed bymyeloperoxidase and lactoperoxidase. Biochim. Bio-phys. Acta, 709, 212–219.
32. Cals, M. M., Mailliart, P., Brignon, G., Anglade, P. &Dumas, B. R. (1991). Primary structure of bovinelactoperoxidase, a fourth member of a mammalianheme peroxidase family. Eur. J. Biochem. 198, 733–739.
33. Shindler, J. S. & Bardsley, W. G. (1975). Steady-statekinetics of lactoperoxidase with ABTS as chromogen.Biochem. Biophys. Res. Commun. 67, 1307–1312.
34. Chomczynski, P. & Sacchi, N. (1987). Single-stepmethod of RNA isolation by acid guanidinium thio-cyanate phenol chloroform extraction. Anal. Biochem.162, 156–159.
35. Otwinowski, Z. & Minor, W. (1997). Processing ofX-ray diffraction data collected in oscillation mode.Methods Enzymol. 276, 307–326.
36. McCoy, A. J., Grosse-Kunstleve, R. W., Storoni, L. C. &Read, R. J. (2005). Likelihood-enhanced fast transla-tion functions. Acta Crystallogr. D61, 458–464.
37. Murshudov, G. N., Vagin, A. A. & Dodson, E. J. (1997).Refinement of macromolecular structures by themaximum-likelihood method. Acta Crystallogr. D53,240–253.
38. Collaborative Computational Project, Number 4.(1994). The CCP4 suite: programs for protein crystal-lography. Acta Crystallogr. D50, 760–763.
39. Jones, T. A., Zou, J., Cowan, S. W. & Kjeldgaard, M.(1991). Improved methods for building models inelectron density maps and the location of errors inthese models. Acta Crystallogr. A47, 110–118.
40. Emsley, P. & Cowtan, K. (2004). Coot: model-buildingtools for molecular graphics. Acta Crystallogr. D60,2126–2132.
Copyright © 2022 FDOKUMEN




















