
Multiple roles of phosphoinositide-specific phospholipase C ...
Upload
independentCategory
view
5download
0
doi:10.1006/jmbi.2001.4550 available online at http://www.idealibrary.com on J. Mol. Biol. (2001) 307, 1049±1059
Sequence and Crystal Structure Determination of aBasic Phospholipase A2 from Common krait(Bungarus caeruleus) at 2.4 AÊ Resolution:Identification and Characterization of itsPharmacological Sites
Garima Singh, S. Gourinath, Sujata Sharma, M. ParamasivamA. Srinivasan and Tej P. Singh*
Department of Biophysics, AllIndia Institute of MedicalSciences, New Delhi110029, India
E-mail address of the [email protected]
Abbreviations used: PLA2, phospKrait phospholipase A2; BPLA2, boA2; PPLA2, porcine pancreatic PLAatra PLA2; EPLA2, notexin; NPLA2PLA2; GPLA2, b2-bungarotoxin; SPLscutatus PLA2.
0022-2836/01/041049±11 $35.00/0
This is the ®rst phospholipase A2 (PLA2) structure from the family ofkraits. The protein was isolated from Bungarus caeruleus (common krait)and the primary sequence was determined using cDNA approach. Three-dimensional structure of this presynaptic neurotoxic PLA2 from group Ihas been determined by molecular replacement method using the modelof PLA2 component of b2-bungarotoxin (Bungarus multicinctus) andre®ned using CNS package to a ®nal R-factor of 20.1 % for all the data inresolution range 20.0-2.4 AÊ . The ®nal re®ned model comprises 897 pro-tein atoms and 77 water molecules. The overall framework of krait phos-pholipase A2 with three long helices and two short antiparallel b-strandsis extremely similar to those observed for other group I PLA2s. However,the critical parts of PLA2 folding are concerned with its various func-tional loops. The conformations of these loops determine the ef®ciency ofenzyme action and presence/absence of various pharmacological func-tions. In the present structure calcium-binding loop is occupied by asodium ion with a 7-fold co-ordination. The conformation of loop 55-75in krait PLA2 corresponds to a very high activity of the enzyme. A com-parison of its sequence with multimeric PLA2s clearly shows the absenceof critical residues such as Tyr3, Trp61 and Phe64, which are involved inthe multimerization of PLA2 molecules. The protein shows anticoagulantand neurotoxic activities.
# 2001 Academic Press
Keywords: Bungarus caeruleus; phospholipase A2; presynaptic neurotoxin;neurotoxic site; X-ray structure
*Corresponding authorIntroduction
Phospholipase A2 s (PLA2s) are widely distribu-ted in the animal world. Besides the digestive tract,the snake venoms constitute a particularly richsource of these enzymes. The low molecular massextracellular PLA2s have been classi®ed into two
ing author:
holipase A2; KPLA2,vine phospholipase2; APLA2, Naja naja, Naja naja najaA2, Notechis scutatus
main groups I and II. Group I contains enzymesfrom mammalian pancreatic species and venoms ofElapidae (cobras) and Hydrophiidae (sea snakes)snakes, whereas PLA2 s from Crotalidae (rattle-snakes and pit vipers), Viperidae (old world vipers)and the human synovial ¯uid phospholipasesbelong to group II. The members of the ®rst grouppossess an S-S bridge between the side-chains ofCys11 and Cys80. The enzymes from the secondgroup have a C-terminal extension of ®ve to sevenresidues including a C-terminal Cys residue linkedto the SH group of residue 50. The other six disul-®de bridges are conserved in the proteins fromboth groups. Venom PLA2s vary considerably intheir primary and quaternary structures, in theirtoxicity and in enzyme activity. During evolution
# 2001 Academic Press

Table 1. Summary of crystallographic re®nement
RefinementResolution limits (AÊ ) 20.0-2.4No. of reflections 4629R-factor (%) 20.1
Free-R factor (%) 27.1
ModelProtein atoms 897Sodium ion 1Water molecules 77
RMS deviations from ideal valuesBond lengths (AÊ ) 0.007Bond angles (deg.) 1.1Dihedral angles (deg.) 22.2Over all G-factor 0.25
Average B-factors (AÊ 2)Main-chain atoms 28.2Side-chain atoms and water 31.2For all atoms 30.1
Ramachandran plot(non-Gly, non-Pro)Residues in most allowed regions (%) 90Residues in additionally allowed regions (%) 10
1050 Basic Phospholipase A2 from Bungarus caeruleus
snake venom PLA2s have acquired a number ofdifferent physiological activities including presyn-aptic neurotoxicity. The PLA2s share a commonqualitative catalytic property but differ greatly intheir pharmacological properties. The residues inthe catalytic site, Ca2�-binding loop and the inter-face recognition site have been identi®ed andunderstood well. However, the relationshipsbetween catalytic ef®ciency of the enzyme and itsvarious pharmacological properties, the identi®-cation of pharmacological sites and their character-istic conformations and relationships betweenconformations and lethalities of these actions arenot yet clearly de®ned. Several attempts have beenmade to predict the site of toxicity of theseenzymes by comparing protein sequences in corre-lation with their lethalities but it has not yet beenunderstood adequately due to lack of consistencyin the correlation. (Dufton & Hider, 1983; Kini &Iwanaga, 1986a,b; Kini & Evans, 1987; Tsai et al.,1987; Kondo et al., 1989; Takasaki et al., 1990; Tsaiet al., 1995). The most realistic correlationship canbe developed only through their three-dimensionalstructures. Therefore, it is important to determinethe crystal structures of PLA2s from the represen-tative species particularly from group I where onlya few structures are available.
So far, the structures of PLA2s analysed fromgroup I are bovine pancreatic PLA2 (BPLA2)(Dijkstra et al., 1981a,b), porcine pancreatic PLA2(PPLA2) (Dijkstra et al., 1983), Naja naja atra PLA2(APLA2) (Scott et al., 1990), Notexin (EPLA2)(Westerlund et al., 1992), Naja naja naja (NPLA2)(Fremont et al., 1993), b2-bungarotoxin (GPLA2)(Kwong et al., 1995) and Notechis scutatus scutatusPLA2 (SPLA2) (Carredano et al., 1998). Only two ofthem (SPLA2, EPLA2) show neurotoxicity but nosatisfactory explanation has yet been provided instructural terms. PLA2 from krait venom (Bungarus
caeruleus: KPLA2) belongs to group I and possessesseveral pharmacological properties. It shows asequence homology of 50-55 % with other group IPLA2s. It is strange that its sequence homologywith GPLA2, which is part of a complex withKunitz module in b2-bungarotoxin of banded krait(Bungarus multicictus) (Kwong et al., 1995) is of theorder of 46 %. However, it shows a very highsequence homology (85 %) with a recently reportedsequence of a free PLA2 from banded krait(Bungarus multicictus) (Danse, 1990). In order toelucidate the structural basis of toxic effects of kraitPLA2, we have determined its primary sequenceand crystal structure.
Sequence of KPLA2
The sequences of a number of group I PLA2shave been reported but none from the venom ofcommon krait. So far, the structures of sevenPLA2s from group I belonging to different specieshave been analysed. KPLA2 appears to be theshortest in length out of all the known sequencesof group I PLA2s. A comparison of the sequence ofKPLA2 with seven other PLA2s of group I revealssigni®cant differences in four regions: 10-23, 50-67,71-88 and 108-122 (Figure 1(a)). Most of theseregions correspond to important functional seg-ments of PLA2s and suggest differences in thestructures of these regions implying functionalvariations. It may be mentioned here that the pre-sent PLA2 shows a sequence homology of 85 %(Figure 1(b)) with a recently sequenced PLA2 frombanded krait (Danse, 1990).
Structure of KPLA2
The ®nal model consists of 897 protein atoms,one sodium ion and 77 water oxygen atoms. Theaccuracy of the model of KPLA2 was checkedduring re®nement by calculations of electron den-sity maps using coef®cients jFo ÿ Fcj, with selectedparts of the molecule deleted from the atomicmodel used for structure factor calculations. Theelectron density map j2Fo ÿ Fcj represents continu-ous and well-de®ned density (Figure 2) for thebackbone as well for the side-chains. The ®nal R-factor and Rfree in the resolution range 20.0-2.4 AÊ
were 0.201 and 0.271 respectively. The overall B-factor for the structure was 30.1 AÊ 2. The ®nalmodel of KPLA2 is of good stereochemical qualitywith r.m.s. deviations from ideal values being0.007 AÊ for bond lengths and 1.1o for bond angles(Table 1). A Ramachandran plot of the main-chaintorsion angles (f,c) (Ramachandran & Sasisekaran,1968) calculated using PROCHECK (Laskowskiet al., 1993) showed that 90 % of the residues werefound in the most allowed regions (Table 1), andrest of the residues were located in additionallyallowed regions with none in the generous and dis-allowed regions.
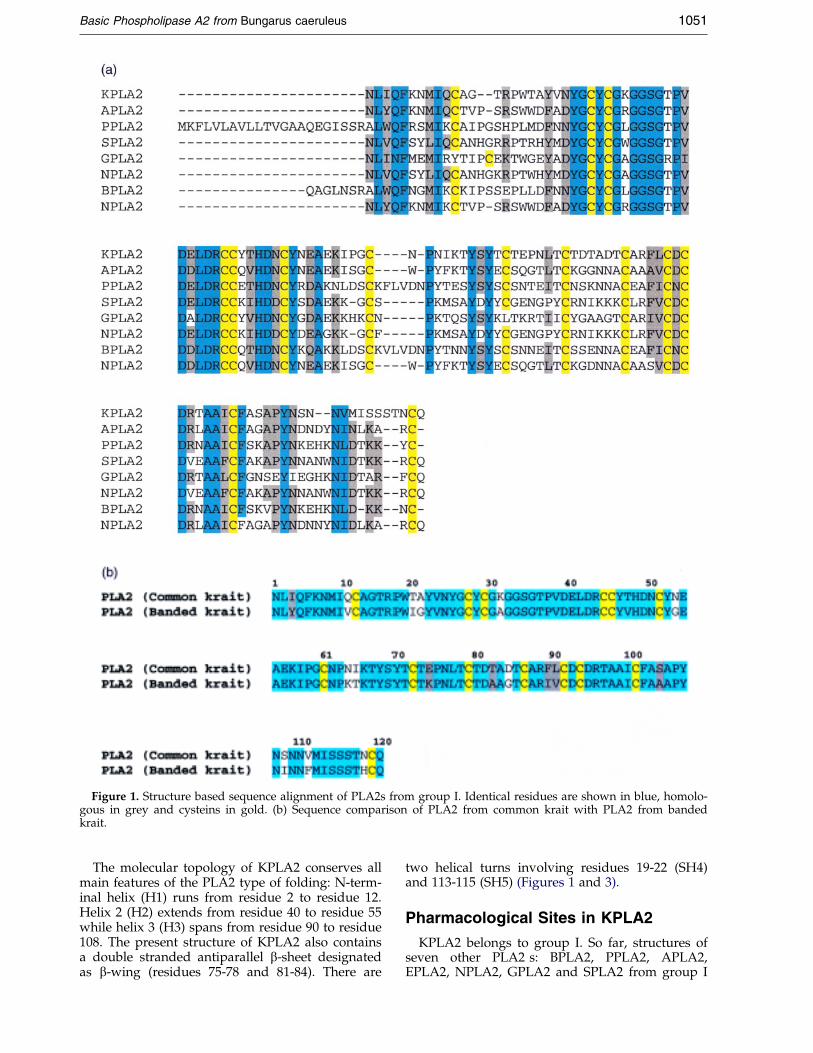
Figure 1. Structure based sequence alignment of PLA2s from group I. Identical residues are shown in blue, homolo-gous in grey and cysteins in gold. (b) Sequence comparison of PLA2 from common krait with PLA2 from bandedkrait.
Basic Phospholipase A2 from Bungarus caeruleus 1051
The molecular topology of KPLA2 conserves allmain features of the PLA2 type of folding: N-term-inal helix (H1) runs from residue 2 to residue 12.Helix 2 (H2) extends from residue 40 to residue 55while helix 3 (H3) spans from residue 90 to residue108. The present structure of KPLA2 also containsa double stranded antiparallel b-sheet designatedas b-wing (residues 75-78 and 81-84). There are
two helical turns involving residues 19-22 (SH4)and 113-115 (SH5) (Figures 1 and 3).
Pharmacological Sites in KPLA2
KPLA2 belongs to group I. So far, structures ofseven other PLA2 s: BPLA2, PPLA2, APLA2,EPLA2, NPLA2, GPLA2 and SPLA2 from group I

Figure 2. Stereoview of a representative region of the electron density map (2Fo ÿ Fc) with a ®nal re®ned modelsuperimposed on it. The map was contoured at 1.6 s.
1052 Basic Phospholipase A2 from Bungarus caeruleus
have been determined. This is the ®rst PLA2 fromkrait venom. In addition to a characteristic catalyticsite KPLA2 possesses several other functional siteswhich are responsible for various pharmacologicalproperties. These sites are important since they dif-fer greatly in various PLA2s. There are such sixloop regions, which are held in different ways bycommon secondary structure motifs (Figure 4). Theconformational differences of these sites must bedirectly related to the differences in their pharma-cological functions. As seen from Figures 5(a)-(g),KPLA2 possesses a unique structure which is pre-sumably responsible for it having the largest num-ber of pharmacological actions.
Figure 3. Ribbon diagram showing the structure ofKPLA2. Three main helices are indicated as H1, H2 andH3, while two short helices are designated as SH4 andSH5. b-wing and disul®de linkages are also shown.Sodium ion can be seen in the calcium-binding loop.
Loop 1
As de®ned in Figure 4, this loop connects helixH1 and short helix SH4. In most cases, the size ofthis loop is larger than that observed in KPLA2.The short loop in KPLA2 adopts a type III b-turnconformation. The tightness of this loop seems todetermine the ¯exibility of calcium binding loopand de®nes the accessibility of catalytic residues(Figure 4).
Loop 2 (Calcium binding loop)
One of the highly conserved regions in theamino acid sequences of PLA2s is the segmentfrom Tyr25 to Gly35. The presence of two half
Figure 4. The backbone tracing of KPLA2. The loopregions are indicated in red with residue numbers at thebeginning as well as at the ends. Active site residuesHis48, Tyr52 and Asp99 are also indicated.

Basic Phospholipase A2 from Bungarus caeruleus 1053
cystein residues and four glycine residues suggesta well de®ned and speci®cally conformed back-bone. The cage-like structure with three oxygenatoms from carbonyl groups of Tyr28, Gly30 andGly32 residues protruding inwardly suggests apossible role in the sequestering of a cation. In thepresent structure a sodium ion is present with7-fold coordination. In the GPLA2 also an Na� waslocated in this loop but with 5-fold co-ordination.In this case, the two coordinating water moleculeswere absent. The structures of BPLA2, PPLA2 andNPLA2 were determined with Ca2� while those ofEPLA2 and SPLA2 did not have a metal ion. Astructural comparison of the metal binding loopsin various group I PLA2 structures shows a con-siderable similarity in the overall conformation ofthis loop. This implies that the presence of a cal-cium ion does not seem to alter the conformationof the loop appreciably but it seems to stabilize theloop and other parts of the protein structure byproviding strong co-ordinating interactions withGly28 O, Gly30 O, Gly32 O and Od1 and Od2 atomsof Asp49 together with two water oxygen atoms.Further evidence of the stability of calcium-bindingloop can be seen in the form of lower B factors forthe residues of the calcium-binding loop as com-pared to those where calcium site is occupiedby H2O or it is empty. (Westerlund et al., 1992;Carredano et al., 1998).
Loop 3 (anticoagulant site)
It was shown that the anticoagulant action wasindependent of enzymatic activity and wasmimicked by lysoderivative and fatty acid (Boffaet al., 1976). By complexing to phospholipids, theenzyme interferes with the activation of coagu-lation protein factors. It has been indicated that thestretch of residues 55 to 67, rich in positivelycharged residues, displays a strong anticoagulantactivity (Kini & Evans, 1987; Gowda et al., 1994;Carredano et al., 1998). It appears that a positivelycharged residue free for intermolecular interactionsat the corner of molecule corresponding to thestretch of residues 55 to 67 seems to be a commonfeature of most of the anticoagulant PLA2s regard-less of their group or the class (Carredano et al.,1998). In KPLA2 there are two positively chargedresidues in this segment, which are orientedfavourably to induce strong anticoagulant effect.As seen from Figures 5(a) and (b), the confor-mation of this loop is similar to those observed inNPLA2 and APLA2, which possess strong anticoa-gulant activity. The corresponding loop in otherPLA2s of group I adopts signi®cantly different con-formations (Figures 5(c)-(g)) resulting in arelatively lower or no anticoagulant activity.
Loop 4
It connects loop 3 and the b-wing of the protein.The length and conformation of loop 4 determinethe shape of loop 3 and the orientation of the
b-wing. The conformation of loop 4 in KPLA2 issimilar to those observed in NPLA2, APLA2,GPLA2 and BPLA2. The corresponding loops inPPLA2, SPLA2 and EPLA2 adopt different confor-mations. The lengths of these loops vary from 11to 18 residues. The length with 18 residues is thelargest for this loop in BPLA2. It has been shownin BPLA2 that the removal of a stretch of residues62 to 66 and with certain other replacements ofspeci®c residues such as Asp59 by Ser59, Ser60 byGly60 and Asn67 by Tyr67 enhanced the enzymeactivity by 16 times (Kuipers et al., 1989). Thelength of this loop in KPLA2 with just 11 residuesis among the smallest ones thus indicating thatthe enzyme possesses a highly ef®cient nativestructure.
Loop 5
Loop 5 is a short turn between two b-strands 75-78 and 81-85. The turn in KPLA2 overlaps verywell with that observed in NPLA2 (Figure 5(a))and differs greatly from those in APLA2(Figure 5(b)), BPLA2 (Figure 5(g)), EPLA2(Figure 5(f)), GPLA2 (Figure 5(e)), SPLA2(Figure 5(d)) and PPLA2 (Figure 5(c)). It is note-worthy that the sequence of the reverse turnGlu78-Pro79-Asn80-Leu81 in KPLA2 does not haveany sequence homology with corresponding frag-ments in other group I PLA2s from other species.
Loop 6 (neurotoxic site)
Several attempts have been made to predict theneurotoxic site of PLA2 enzymes by theoreticalmethods (Dufton & Hider, 1983). It is widelyaccepted that each presynaptic neurotoxin of PLA2type from the snake venom has an individual neu-rotoxic site separated from the PLA2 catalytic site(Yang, 1994 and Kini & Evans, 1989). Such a neu-rotoxic site enables the neurotoxin to speci®callybind to the receptor on the neuromembrane. How-ever, the location of neurotoxic site on the neuro-toxin molecule and details of its interactions withthe receptor have not yet been clearly understood.Presynaptic neurotoxic potential is predicted byhydropathy pro®les and phospholipases have dis-tinct hydrophobic regions around residues 80 to110 in group I PLA2s (Kini & Iwanaga, 1986a,b).
The structural comparisons of this loop inKPLA2 with presynaptic, EPLA2 (Westerlund et al.,1992) (Figure 5(f)) and SPLA2 (Carredano et al.,1998) (Figure 5(d)) show a striking similarity,whereas it displays clear differences with the corre-sponding loops in NPLA2 (Fremont et al., 1993)(Figure 5(a)), APLA2 (White et al., 1990)(Figure 5(b)) and BPLA2 (Dijkstra et al., 1981b)(Figure 5(g)), which are not neurotoxic. In case ofGPLA2, which is associated with a Kunitz proteaseinhibitor as a heterodimer, this loop has a differentconformation like those in the above PLA2s but itis suggested that upon binding of Kunitz inhibitorto the neuromembrane, the conformation of this

Figure 5 (legend opposite)
1054 Basic Phospholipase A2 from Bungarus caeruleus

Figure 5. Superimposition of Ca tracing of KPLA2 (continuous lines) on the Ca tracing of (a) NPLA2, (b) APLA2,(c) PPLA2, (d) SPLA2, (e) GPLA2, (f) EPLA2 and (g) BPLA2 (broken lines). L1-L6 indicate various loops, and N andC correspond to N and C termini.
Basic Phospholipase A2 from Bungarus caeruleus 1055
loop undergoes a change (Kwong et al., 1995). Fur-thermore, residues 58, 59, 60 and 61 of KPLA2form a unique b-turn II conformation with anH-bond between Ile58 O and Cys61 N(O � � �N � 3.00 AÊ ) while in non-neurotoxic PLA2s itadopts a random conformation. Also, the com-pactly organized local structure of the stretch 80-110 in KPLA2 is sharply different from the looselyextended conformations in non-neurotoxic PLA2s.In KPLA2, six main-chain/side-chain hydrogenbonds (Ser69 N-Asp81 Od1, Asp81 N-Asp81 Od1,Thr82 O-Arg88 NH2, Thr82 O-Arg88 Ne, Asp84 O-Arg88 N and Ala87 O-Cys91 N) bring residue 88closer to the main body of the molecule and con-tribute to the stability of the conformation of thestretch 80-110. Compared with the non-neurotoxicstructures, the peptidyl plane between residues 86and 88 ¯ips by about 180 �. This may be due tothe sequence changes in neurotoxic and non-
neurotoxic PLA2 s as the two consecutive residues86 and 88 of the non-neurotoxic PLA2s are Glyand Asp while the corresponding residues ofKPLA2 are Lys and Gly. Therefore, these structuraland electrostatic properties might be of functionalsigni®cance.
Catalytic Activity of KPLA2
The residues His48, Tyr52, Asp99 and Asp49,which are responsible for catalytic activity have anideal stereochemistry with the presence of the socalled ``catalytic network'', a system of hydrogenbonds which involves the catalytic triad. Inaddition to catalytic triad, the active site includes anucleophile attacking water molecule, which has awell de®ned electron density and forms twohydrogen bonds with His48 Nd1 and Asp49 Od1
(3.00 and 2.58 AÊ , respectively) (Figure 6). In a
Figure 6. Stereoview of the activesite residues in KPLA2. The resi-dues are labeled.
1056 Basic Phospholipase A2 from Bungarus caeruleus
similar way, a complete overlap with other class IPLA2s is observed for the invariant hydrophobicside-chains of Leu2, Leu/Ile3, Ile9, Tyr22, Cys29,Cys45 and Ala107 which form the walls of thehydrophobic channel for the binding of the sub-strate in PLA2. The side-chains of these residuesare directed towards, and thus might interact withthe phospholipid layer.
Materials and Methods
Purification PLA2
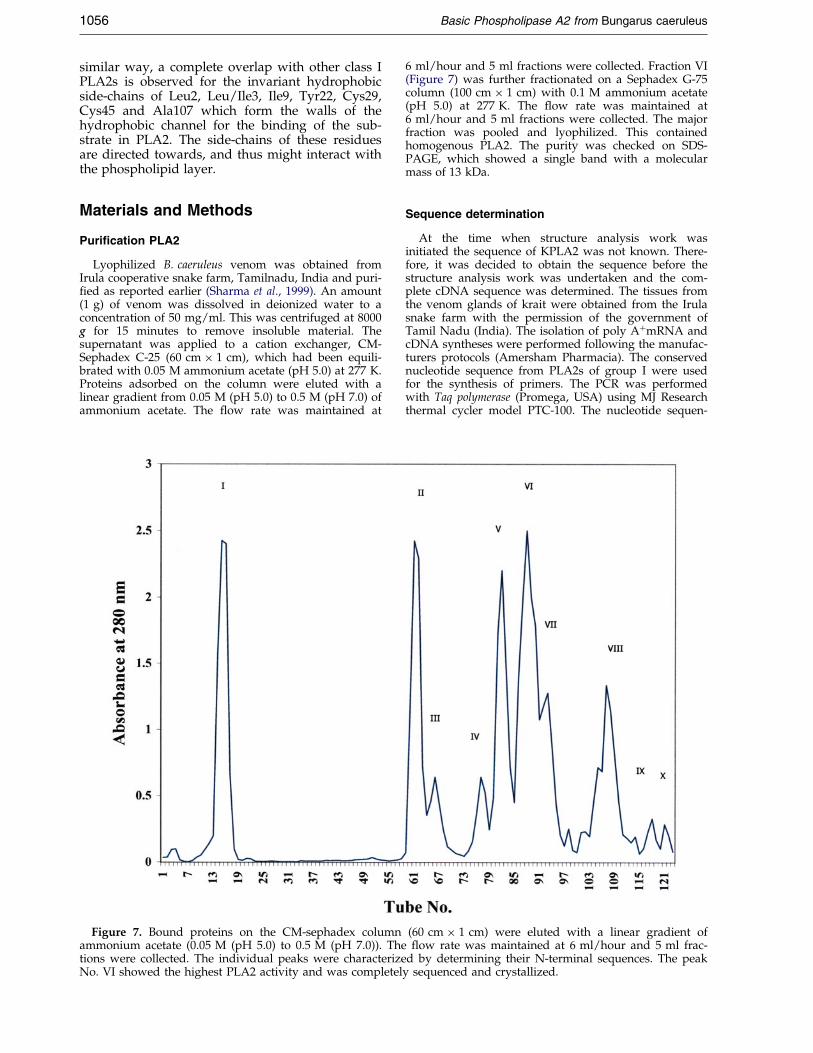
Lyophilized B. caeruleus venom was obtained fromIrula cooperative snake farm, Tamilnadu, India and puri-®ed as reported earlier (Sharma et al., 1999). An amount(1 g) of venom was dissolved in deionized water to aconcentration of 50 mg/ml. This was centrifuged at 8000g for 15 minutes to remove insoluble material. Thesupernatant was applied to a cation exchanger, CM-Sephadex C-25 (60 cm � 1 cm), which had been equili-brated with 0.05 M ammonium acetate (pH 5.0) at 277 K.Proteins adsorbed on the column were eluted with alinear gradient from 0.05 M (pH 5.0) to 0.5 M (pH 7.0) ofammonium acetate. The ¯ow rate was maintained at
Figure 7. Bound proteins on the CM-sephadex columnammonium acetate (0.05 M (pH 5.0) to 0.5 M (pH 7.0)). Thetions were collected. The individual peaks were characterizeNo. VI showed the highest PLA2 activity and was completel
6 ml/hour and 5 ml fractions were collected. Fraction VI(Figure 7) was further fractionated on a Sephadex G-75column (100 cm � 1 cm) with 0.1 M ammonium acetate(pH 5.0) at 277 K. The ¯ow rate was maintained at6 ml/hour and 5 ml fractions were collected. The majorfraction was pooled and lyophilized. This containedhomogenous PLA2. The purity was checked on SDS-PAGE, which showed a single band with a molecularmass of 13 kDa.
Sequence determination
At the time when structure analysis work wasinitiated the sequence of KPLA2 was not known. There-fore, it was decided to obtain the sequence before thestructure analysis work was undertaken and the com-plete cDNA sequence was determined. The tissues fromthe venom glands of krait were obtained from the Irulasnake farm with the permission of the government ofTamil Nadu (India). The isolation of poly A�mRNA andcDNA syntheses were performed following the manufac-turers protocols (Amersham Pharmacia). The conservednucleotide sequence from PLA2s of group I were usedfor the synthesis of primers. The PCR was performedwith Taq polymerase (Promega, USA) using MJ Researchthermal cycler model PTC-100. The nucleotide sequen-
(60 cm � 1 cm) were eluted with a linear gradient of¯ow rate was maintained at 6 ml/hour and 5 ml frac-
d by determining their N-terminal sequences. The peaky sequenced and crystallized.

Table 2. Crystallographic data
Space group R32Cell dimensions a � b � c � 57.9 AÊ
a � b � g � 92.0 �Z 6Matthews coefficient (Vm) (AÊ 3/Da) 2.5Solvent content (%) 50.0Resolution range (AÊ ) 20.0-2.4Number of crystals used 4Number of unique reflections 4629Overall completeness (20.0-2.4 AÊ )% 93Redundancy 17.2Completeness in the highest resolution
shell (2.9-2.4 AÊ ) (%)92
Rmerge (%) 9.2Rsym in the highest resolution
shell (2.9-2.4) (%)17.4
Mean I/s(I) 7.2
Basic Phospholipase A2 from Bungarus caeruleus 1057
cing was performed on the cloned double standard DNA(pGEM-T) using automatic sequencer model ABI-377.Both strands were used for sequencing. The cDNA ofKPLA2 reported here is 464 base-pairs (bp) in length. Itcomprises the open reading frame encoding 147 aminoacid residues and part of the 30-UTR (18 bp). The signalpeptide is 27 amino acid long. The nucleotide andderived amino acid sequences are given in Figure 8(Gene bank accession number: AF297663).
Crystallization of PLA2
Crystals of KPLA2 were obtained using sitting-dropvapour diffusion method. The protein was dissolved at10 mg/ml in 50 mM Tris. HCl buffer (pH 8.5) containing1.4 M NaCl, 1 mM NaN3. It was equilibrated with thesame buffer containing 3.5 M NaCl. After twenty days ofincubation at 8 �C, crystals had grown to about0.3 mm � 0.2 mm � 0.2 mm in size.
Data collection
The crystals were mounted in the glass capillaries.X-ray intensities were measured at 285 K using a MARResearch imaging plate scanner with a diameter of300 mm. The crystal to detector distance was kept at200 mm. Monochromatic CuKa radiation was obtainedwith the help of a graphite crystal monochromatormounted on a Rigaku RU-200 rotating anode X-ray gen-erator operating at 40 kV and 100 mA with a focal pointof 0.3 mm � 3 mm. A total of 135 images were collectedeach with 1 � rotation. The total exposure time for indi-vidual image was 1800 seconds. The crystals were small,but diffracted very well and were stable in the X-raybeam. The complete data set to 2.4 AÊ resolution wasobtained using four crystals.
Data processing
The HKL package (Otwinowski, 1993; Minor, 1993)and MARXDS and MARSCALE (Kabsch, 1988) wereused for the determination of unit cell parameters, data
processing and scaling of the data. The crystals belong torhombohedral space group R32 with one molecule in theasymmetric unit (Table 2).
Structure determination
The structure was solved by molecular replacementusing AMoRe program (Navaza, 1994) from the CCP4software suite (CCP4, 1994; Dodson et al., 1997). Therotation function was calculated using diffraction datafrom 10 to 4.5 AÊ resolution with a sphere radius of 18 AÊ .The ®rst peak appeared as the distinct solution in therotation function. This peak grew more distinct in thetranslation calculations with correlation coef®cient of34.3 % and R-factor of 49.9 %. The rigid body re®nementfurther improved the correlation coef®cient to 42.0 %with an R-factor of 46.2 %.
Structure refinement
The solution was applied to model coordinates usingLSQKAB (CCP4, 1994) and these coordinates were used
Figure 8. Nucleotide anddeduced amino acid sequence ofKPLA2. The amino acid residuesare shown in three letter code. Thearrow marks the N-terminal aminoacid of the mature protein. Thestop codon is indicated by ***.

1058 Basic Phospholipase A2 from Bungarus caeruleus
as the starting model for the re®nement using CNS pack-age (BruÈ nger et al., 1998). The re¯ections were treated astwo data sets, one data set consisting of 10 % of re¯ec-tions (500 re¯ections) which were randomly selected forthe Rfree calculations (BruÈ nger, 1992) and the other setconsisting of the rest of the data (4535 re¯ections).During the re®nement, the bond lengths and bondangles were restrained close to the standard values, andrestraints were also placed on the planarity of groupsand non-bonded contacts as de®ned by Engh & Huber(1991). Several cycles of re®nement using rigid-body con-jugate-gradient minimization, simulated-annealing andmodel building with the program ``O'' (Jones et al.,1991), brought the R-factor down to 0.30 and the Rfree to0.415 for all the data in the resolution range of 20.0-2.4 AÊ . At this stage, the individual isotropic B-factorre®nement was carried out which reduced the R-factorto 0.260 and Rfree to 0.334. Manual model building usingj2Fo ÿ Fcj and jFo ÿ Fcj electron density maps and furtherre®nement by simulated-annealing with moleculardynamics using a slow-cooling protocol from 3000 K to300 K (BruÈ nger et al., 1990), brought the R-factor downto 0.234 and Rfree to 0.321 for the re¯ections in the resol-ution range 20.0-2.4 AÊ . Water molecules were includedin the model, provided the peaks were greater than 2.5sin [Fo ÿ Fc] maps, hydrogen bond partners with appro-priate distance and angle geometry and B-values lessthan 50 AÊ 2 after re®nement. The re®nement improvedthe map and enabled the addition of more water mol-ecules. After each set of cycles of re®nement, j2Fo ÿ Fcjand jFo ÿ Fcj maps were calculated using MAPMAN(Kleywegt & Jones, 1996) and corrections to the side-chain orientations were done using program ``O'' (Joneset al., 1991) on silicon graphics Indigo-2 workstation. Themodel was checked by calculating a series of omit mapsat each stage of re®nement. The ®nal R-factor was 0.201and Rfree was 0.271 for all the data in the resolutionrange of 20.0 to 2.4 AÊ .
Protein Data Bank and gene bankaccession numbers
The atomic coordinates of the re®ned structure ofKPLA2 have been deposited with RCSB Protein DataBank with the entry code of 1FE5. The nucleotidesequence has been deposited in gene bank with the entrycode of AF297663.
Acknowledgments
The ®nancial support from the Department of Scienceand Technology, New Delhi is gratefully acknowledged.G.S. and M.P. thank the Council of Scienti®c and Indus-trial Research, New Delhi for the award of fellowships.
References
Boffa, G. A., Boffa, M. C. & Winchenne, J. J. (1976).A Phospholipase A2 with anticoagulant activity. I.Isolation from Vipera berus venom and properties.Biochim. Biophys. Acta, 429, 828-836.
BruÈ nger, A. T. (1992). The free R-value: a novel statisti-cal quantity for assessing the accuracy of crystalstructures. Nature, 355, 472-474.
BruÈ nger, A. T., Krukowski, A. & Erickson, J. (1990).Slow-cooling protocols for crystallographic re®ne-
ment by simulated annealing. Acta Crystallog. sect.A, 46, 585-593.
BruÈ nger, A. T., Adams, P. D., Clore, G. M., Delano,W. L., Gros, P. & Gorsse-Kunstleve, R. W. et al.(1998). Crystallography and NMR system (CNS): anew software system for macromolecular structuredetermination. Acta Crystallog. sect. D, 54, 905-921.
Carredano, E., Westerlund, B., Persson, B., Saarinen, M.,Ramaswamy, S., Eaker, D. & Eklund, H. (1998). Thethree-dimensional structures of two toxins fromsnake venom throw light on the anticoagulant andneurotoxic sites of phospholipase A2. Toxicon, 36,75-92.
Collaborative Computational Project Number 4 (1994).The CCP4 suite: programs for protein crystallogra-phy. Acta Crystallog. sect. D, 50, 760-763.
Danse, J. M. (1990). Nucleotide sequence encoding fornon-toxic phospholipase: A2 from Bungarus multi-cinctus. Nucl. Acids Res. 18, 4608.
Dijkstra, B. W., Drenth, J. & Kalk, K. H. (1981a). Activesite and catalytic mechanism of phospholipase A2.Nature, 289, 604-606.
Dijkstra, B. W., Kalk, K. H., Hol, W. G. & Drenth, J.(1981b). Structure of bovine pancreatic phospho-lipase A2 at 1.7 AÊ resolution. J. Mol. Biol. 147, 97-123.
Dijkstra, B. W., Renetseder, R., Kalk, K. H., Hol, W. G.& Drenth, J. (1983). Structure of porcine pancreaticphospholipase A2 at 2.6 AÊ resolution and compari-son with bovine phospholipase A2. J. Mol. Biol. 168,163-179.
Dodson, E. J., Winn, M. & Ralph, A. (1997). Collabora-tive Computational Project 4: providing programsfor protein crystallography. Methods Enzymol. 277,620-633.
Dufton, M. J. & Hider, R. C. (1983). Classi®cation ofphospholipase A2 according to sequence. Evolution-ary and pharmacological implications. Eur. J.Biochem. 137, 545-551.
Engh, R. A. & Huber, R. (1991). Accurate bond andangle parameters for X-ray protein structure re®ne-ment. Acta Crystallog. sect. A, 47, 392-400.
Fremont, D. H., Anderson, D. H., Wilson, I. A., Dennis,E. A. & Xuong, N. H. (1993). Crystal structure ofphospholipase A2 from Indian cobra reveals atrimeric association. Proc. Natl Acad. Sci. USA, 90,342-346.
Godwa, V. T., Schmidt, J. & Middlebrook, J. L. (1994).Primary sequence determination of the most basicmyonecrotic phospholipase A2 from venom ofVipera russelli. Toxicon, 32, 665-673.
Jones, T. A., Zou, J., Cowan, S. W. & Kjeldgaad, M.(1991). Improved methods for building models inelectron density maps and the location of errors inthese models. Acta Crystallog. sect. A, 47, 110-119.
Kabsch, W. (1988). Evaluation of a single-crystal X-raydiffraction data from a position-sensitive detector.J. Appl. Crystallog. 21, 916-924.
Kini, R. M. & Evans, H. J. (1987). Structure-functionrelationship of phospholipases. The anticoagulantregion of phospholipase A2. J. Biol. Chem. 262,14402-14407.
Kini, R. M. & Evans, H. J. (1989). A common cytolyticregion in myotoxins, hemolysins, cardiotoxins andantibacterial peptides. Int. J. Peptide Protein Res. 34,277-286.
Kini, R. M. & Iwanaga, S. (1986a). Structure-functionrelationships of phospholipases. I: prediction of pre-synaptic neurotoxicity. Toxicon, 24, 527-41.

Basic Phospholipase A2 from Bungarus caeruleus 1059
Kini, R. M. & Iwanaga, S. (1986b). Structure-functionrelationships of phospholipases. II: charge densitydistribution and the myotoxicity of presynapticallyneurotoxic phospholipases. Toxicon, 24, 895-905.
Kleywegt, G. J. & Jones, T. A. (1996). xdlMAPMAN andxdlDATAMAN programs for reformatting, analysisand manipulating of biomacromolecular electrondensity maps and re¯ection data sets. Acta Crystal-log. sect. D, 52, 826-828.
Kondo, K., Zhang, J., Xu, K. & Kagamiyama, ?. (1989).Amino acid sequence of a presynaptic neurotoxin,agkistrodotoxin, from the venom of Agkistrodonhalys Pallas. J. Biochem. 105, 196-203.
Kuipers, O. P., Thunnissen, M. M., de Geus, P., Dijkstra,B. W., Drenth, J., Verheij, H. M. & de Haas, G. H.(1989). Enhanced activity and altered speci®city ofphospholipase A2 by deletion of a surface loop.Science, 244, 82-85.
Kwong, P. D., McDonald, N. Q., Sigler, P. B. &Hendrickson, W. A. (1995). Structure of beta2-bungarotoxin: potassium channel binding byKunitz modules and targeted phospholipase action.Structures, 3, 1109-1119.
Laskowski, R., Macarthur, M., Moss, D. & Thornton, J.(1993). Procheck: a program to check stereochemicalquality of protein structures. J. Appl. Crystallog. 26,283-290.
Minor, W. (1993). XDISPLAYF Program, Purdue Univer-sity.
Navaza, J. (1994). AmoRe: an Automated package forMolecular Replacement. Acta Crystallog. sect. A, 50,157-163.
Otwinowski, Z. (1993). Oscillation data reduction pro-gram. In Proceedings of the CCP4 Study Weekened(Sawyer, L., Issacs, N. & Bailey, S., eds), pp. 56-62,SERC Daresbury Laboratory, UK.
Ramachandran, G. N. & Sasisekharan, V. (1968). Confor-mation of polypeptides and proteins. Advan. ProteinChem. 23, 283-438.
Scott, D. L., White, S. P., Otwinowski, Z., Yuan, W.,Gelb, M. H. & Sigler, P. B. (1990). Interfacial cataly-sis: the mechanism of phospholipase A2. Science,250, 1541-1546.
Sharma, S., Karthikeyan, S., Betzel, C. & Singh, T. P.(1999). Isolation, puri®cation, crystallization andpreliminary X-ray analysis of beta 1-bungarotoxinfrom Bungarus caeruleus (Indian common krait).Acta Crystallog. sect. D, 55, 1093-1094.
Takasaki, C., Yutani, F. & Kajiyashiki, T. (1990). Aminoacid sequences of eight phospholipases A2 from thevenom of Australian king brown snake, Pseudechisaustralis. Toxicon, 28, 329-339.
Tsai, I. H., Liu, H. C. & Chang, T. (1987). Toxicitydomain in presynaptically toxic phospholipase A2of snake venom. Biochem. Biophys. Acta, 916, 94-99.
Tsai, I. H., Lu, P. J., Wang, Y. M., Ho, C. L. & Liaw,L. L. (1995). Molecular cloning and characterizationof a neurotoxic phospholipase A2 from the venomof Taiwan habu (Trimeresurus mucrosquamatus).Biochem. J. 311, 895-900.
Westerlund, B., Nordlund, P., Uhlin, U., Eaker, D. &Eklund, H. (1992). The three-dimensional structureof notexin, a presynaptic neurotoxic phospholipaseA2 at 2.0 AÊ resolution. FEBS Letters, 301, 159-164.
White, S. P., Scott, D. L., Otwinowski, Z., Gelb, M. H. &Sigler, P. B. (1990). Crystal structure of cobra-venom phospholipase A2 in a complex with atransition-state analogue. Science, 250, 1560-1563.
Yang, C. C. (1994). Structure-function relationship ofphospholipase A2 from snake venoms. J. Toxicol.Toxin Rev. 13, 125-177.
Edited by R. Huber
(Received 19 September 2000; received in revised form 7 February 2001; accepted 7 February 2001)
Copyright © 2022 FDOKUMEN



![â…^Ûnj‰];æ;êŠËßÖ];å^Ÿ÷];l]ˆÓi†Ú - saida.dz](https://static.fdokumen.com/doc/165x107/6313fe3ec72bc2f2dd0437e8/aunjaeesessoeayloiu-saidadz.jpg)
















