
Cox-2 inhibition can lead to adverse effects in a rat model for temporal lobe epilepsy
8
Epilepsy Research (2010) 91, 49—56 journal homepage: www.elsevier.com/locate/epilepsyres Cox-2 inhibition can lead to adverse effects in a rat model for temporal lobe epilepsy Linda Holtman a , Erwin A. van Vliet a,b , Peter M. Edelbroek b , Eleonora Aronica c , Jan A. Gorter a,b,∗ a Swammerdam Institute for Life Sciences, Center for Neuroscience, University of Amsterdam, Science Park 904, 1098 XH Amsterdam, The Netherlands b Epilepsy Institute in The Netherlands Foundation (SEIN), Achterweg 5, 2103 SW Heemstede, The Netherlands c Academic Medical Center, Department of (Neuro)Pathology, University of Amsterdam, Meibergdreef 9, 1105 AZ Amsterdam, The Netherlands Received 23 March 2010; received in revised form 2 June 2010; accepted 23 June 2010 KEYWORDS Status epilepticus; Neuroinflammation; Temporal lobe epilepsy; P-glycoprotein; Pharmacoresistance Summary Purpose: Status epilepticus (SE) leads to upregulation of pro-inflammatory proteins including cyclooxygenase-2 (cox-2) which could be implicated in the epileptogenic process and epileptic seizures. Recent studies show that cox-2 can regulate expression of P-glycoprotein (P-gp) during epileptogenesis and epilepsy. P-gp could cause pharmacoresistance by reducing brain entry of anti-epileptic drugs such as phenytoin (PHT). Here we have investigated the effects of cox- 2 inhibition on epileptogenesis, spontaneous seizures and PHT treatment in a rat model for temporal lobe epilepsy (TLE). Methods: A 3-day treatment with the cox-2 inhibitor SC-58236 (SC) was started 1 day before electrically induced SE. Chronic epileptic rats were treated with SC for 14 days, which was followed by a 7-day period of SC/PHT combination treatment. Seizure activity was monitored continuously using electroencephalography. Results: SC treatment did not affect SE duration, but led to an increased number of rats that died during the first 2 weeks after SE. Cox-2 inhibition during the chronic period led to an increased number of seizures in the 2nd week of treatment in 50% of the rats. SC/PHT treatment reduced seizures significantly for only 2 days. Conclusions: Both SC treatment that started before SE and the 14-day treatment in chronic epileptic rats led to adverse effects in the TLE rat model. Despite a temporal reduction in seizure frequency with SC/PHT treatment, SC does not seem to be a suitable approach for anti-epileptogenic or anti-epileptic therapy. © 2010 Elsevier B.V. All rights reserved. ∗ Corresponding author at: Swammerdam Institute for Life Sciences, Center for Neuroscience, University of Amsterdam, Science Park 904, 1098 XH Amsterdam, The Netherlands. Tel.: +31 205257893; fax: +31 205257709. E-mail address: [email protected] (J.A. Gorter). 0920-1211/$ — see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.eplepsyres.2010.06.011
Transcript of Cox-2 inhibition can lead to adverse effects in a rat model for temporal lobe epilepsy

Epilepsy Research (2010) 91, 49—56
journa l homepage: www.e lsev ier .com/ locate /ep i lepsyres
Cox-2 inhibition can lead to adverse effects in a ratmodel for temporal lobe epilepsy
Linda Holtmana, Erwin A. van Vlieta,b, Peter M. Edelbroekb,Eleonora Aronicac, Jan A. Gortera,b,∗
a Swammerdam Institute for Life Sciences, Center for Neuroscience, University of Amsterdam, Science Park 904,1098 XH Amsterdam, The Netherlandsb Epilepsy Institute in The Netherlands Foundation (SEIN), Achterweg 5, 2103 SW Heemstede, The Netherlandsc Academic Medical Center, Department of (Neuro)Pathology, University of Amsterdam, Meibergdreef 9, 1105 AZ Amsterdam,The Netherlands
Received 23 March 2010; received in revised form 2 June 2010; accepted 23 June 2010
KEYWORDSStatus epilepticus;Neuroinflammation;Temporal lobeepilepsy;P-glycoprotein;Pharmacoresistance
SummaryPurpose: Status epilepticus (SE) leads to upregulation of pro-inflammatory proteins includingcyclooxygenase-2 (cox-2) which could be implicated in the epileptogenic process and epilepticseizures. Recent studies show that cox-2 can regulate expression of P-glycoprotein (P-gp) duringepileptogenesis and epilepsy. P-gp could cause pharmacoresistance by reducing brain entry ofanti-epileptic drugs such as phenytoin (PHT). Here we have investigated the effects of cox-2 inhibition on epileptogenesis, spontaneous seizures and PHT treatment in a rat model fortemporal lobe epilepsy (TLE).Methods: A 3-day treatment with the cox-2 inhibitor SC-58236 (SC) was started 1 day beforeelectrically induced SE. Chronic epileptic rats were treated with SC for 14 days, which wasfollowed by a 7-day period of SC/PHT combination treatment. Seizure activity was monitoredcontinuously using electroencephalography.Results: SC treatment did not affect SE duration, but led to an increased number of rats that diedduring the first 2 weeks after SE. Cox-2 inhibition during the chronic period led to an increasednumber of seizures in the 2nd week of treatment in 50% of the rats. SC/PHT treatment reduced
seizures significantly for only 2 days.Conclusions: Both SC treatment that started before SE and the 14-day treatment in chronic epileptic rats led to adverse efseizure frequency with SC/PHTanti-epileptogenic or anti-epilep© 2010 Elsevier B.V. All rights re∗ Corresponding author at: Swammerdam Institute for Life Sciences, C904, 1098 XH Amsterdam, The Netherlands. Tel.: +31 205257893; fax: +3
E-mail address: [email protected] (J.A. Gorter).
0920-1211/$ — see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.eplepsyres.2010.06.011
fects in the TLE rat model. Despite a temporal reduction intreatment, SC does not seem to be a suitable approach fortic therapy.served.
enter for Neuroscience, University of Amsterdam, Science Park1 205257709.

5
I
Ttaaadcp(2nMmtsbacsarneewtPpd
SorTpearaioIivtPlwwpw
M
E
ANa
wha
Ei
IsaewDP(nhmaCcuSrta0tmdca(
s
2tca4teid
D
FSri
0
ntroduction
here is ample evidence that temporal lobe epilepsy andhe development of epilepsy (epileptogenesis) are associ-ted with inflammatory processes in the brain (Gorter etl., 2006; Vezzani and Granata, 2005). Inflammatory medi-tors are abundantly present in brain tissue from animalsuring the epileptogenic (latent) period and during thehronic epileptic phase as well as in resected human hip-ocampal tissue from medical intractable epileptic patientsGorter et al., 2006; Jamali et al., 2006; Ravizza et al.,008). Inhibition of inflammation may therefore provide aew therapeutic strategy for (pharmacoresistant) epilepsy.oreover, it may even be possible to prevent the develop-ent of epilepsy. A potential target for anti-inflammatory
reatment in epilepsy is cyclooxygenase-2 (cox-2). Severaltudies have shown neuroprotective effects of cox-2 inhi-ition (Hewett et al., 2006; Iadecola et al., 2001; Kunznd Oliw, 2001). However, in a recent study we could notonfirm a neuroprotective effect when we used the highlypecific cox-2 inhibitor SC-58236 (SC) that was administeredfter the status epilepticus (SE) in a rat model for tempo-al lobe epilepsy. More importantly, the SC treatment wasot anti-epileptogenic and did not produce an anticonvulsiveffect when administered to chronic epileptic rats (Holtmant al., 2009). Interestingly, in a subsequent study in whiche started to apply the cox-2 inhibitor before the induc-
ion of SE, we were able to prevent the upregulation of-glycoprotein (P-gp), a protein that is involved in the trans-ort of many compounds including several anti-epilepticrugs (AEDs) (van Vliet et al., 2010).
Since this SC treatment protocol prevents the normallyE-induced P-gp upregulation, we reasoned that timingf the cox-2 inhibition could be crucial for an effectiveeduction of inflammation and an anti-epileptogenic effect.herefore we tested the effects of this cox-2 inhibitionrotocol on early epileptogenesis in rats in which SE waslectrically induced. Moreover, since P-gp expression after2-week SC treatment in chronic epileptic rats could be
educed to levels similar to control expression (van Vliet etl., 2010), we also tested the effects of SC on seizure activ-ty in chronic epileptic rats in the absence and presencef phenytoin (PHT), an AED that is a substrate for P-gp.n a previous study we detected that PHT brain levels arencreased in SC-treated rats after PHT administration vs.ehicle-treated rats (van Vliet et al., 2010), a phenomenonhat can be expected if this AED is a substrate for P-gp. SinceHT treatment in untreated chronic epileptic rats does notead to complete seizure reduction (van Vliet et al., 2006),e expected that PHT administration in SC-treated rats, inhich P-gp expression has been shown to be reduced com-ared to untreated epileptic rats (van Vliet et al., 2010),ould lead to improved seizure control.
aterials and methods
xperimental animals
dult male Sprague—Dawley rats (Harlan Netherlands, Horst, Theetherlands) weighing 400—600 g were used in this study which waspproved by the University Animal Welfare committee. The rats
acSt1
L. Holtman et al.
ere housed individually in a controlled environment (21 ± 1 ◦C;umidity 60%; lights on 08:00 a.m.—8:00 p.m.; food and water avail-ble ad libitum).
lectrode implantation and status epilepticus (SE)nduction
n order to record hippocampal EEG and to induce SE, insulatedtainless steel electrodes were implanted in the left dentate gyrusnd angular bundle respectively, as described previously (Gortert al., 2001). Two weeks after recovery from the operation, ratsere connected to a stimulation and recording system (NeuroDataigital Stimulator, Cygnus Technology Inc., Delaware Water Gap,A, USA), with a shielded multistrand cable and electrical swivelAir Precision, Le Plessis Robinson, France). Hippocampal EEG sig-als were amplified (10×) via a FET transistor that connected theeadset to a differential amplifier (20×; CyberAmp, Axon Instru-ents, Burlingame, CA, USA), filtered (1—60 Hz) and digitized bycomputer. EEG software (Harmonie, Stellate Systems, Montreal,
anada) sampled the incoming signal at a frequency of 200 Hz perhannel. A week after after habituation to the new condition, ratsnderwent tetanic stimulation (50 Hz) of the angular bundle until aE was evoked, as described previously (Gorter et al., 2001). Shortly,ats underwent tetanic stimulation (50 Hz) of the hippocampus inhe form of a succession of trains of pulses every 13 s. Each train hadduration of 10 s and consisted of biphasic pulses (pulse duration
.5 ms, maximal intensity 500 �A). Stimulation was stopped whenhe rats displayed sustained forelimb clonus and salivation for a fewinutes, which usually occurred within 1 h. Behavior was observeduring electrical stimulation and several hours thereafter. SE washaracterized by periodic epileptiform discharges (PEDs) of 1—2 Hznd was accompanied by behavioral and electroencephalographicEEG) seizures, which lasted for several hours.
Behavior during SE was scored according to the Racine scale foreizure severity (Racine, 1972):
A stage 1 seizure was characterized by behavioral arrest, eye clo-sure, facial clonus and ‘wet dog shakes’.Stage 2: head nodding associated with more severe facial clonus.Stage 3: clonus of one forelimb.Stage 4: bilateral clonus accompanied by rearing.Stage 5: rearing and falling accompanied by generalized clonicseizures.
SE severity was classified as follows: Mild SE: Racine’s scale 1 andseizures and exploratory behavior (>4 h), which is different from
heir natural exploratory behavior since the rats do not walk in aontinuous pace. Moderate SE: head bobbing, periods of immobilitynd occasional generalized seizures (stage 4 or 5) lasting more thanh. Severe SE: continuous stage 4—5 seizure activity (>4 h). Con-
rol rats (n = 5) were implanted with electrodes and handled likexperimental rats but they were not electrically stimulated. Dur-ng the experiments, EEG was recorded continuously (24 h/day), asescribed previously (Gorter et al., 2001).
rug treatment during epileptogenesis
or this study we have used the highly selective cox-2 inhibitorC-58236 (SC) (Pfizer-Kalamazoo, MI, USA), as for this inhibitor, neu-oprotective effects have been found in rats after a focally inducedschemic insult (Govoni et al., 2001).
To assess the effects of the cox-2 inhibitor on epileptogenesis,
group of rats (n = 9) was treated with SC and SE was induced. Foromparison, a group of vehicle-treated animals (n = 9) underwentE induction in an identical manner. For both the SC-treated andhe vehicle-treated group, EEG was recorded for 15 days, startingday prior to SE induction.

or te
H
Ttwt3iopbusfdTebt1
a(s8nw
S
AAtfupSf
R
Ea
Snt(ii4owd(aggd
Cox-2 inhibition can lead to adverse effects in a rat model f
SC treatment was started 1 day before SE-induction and lastedfor 3 days. A daily dosage of 10 mg/kg SC in water was given orallyvia a gastric tube under short isoflurane anesthesia. This dosageeffectively reduces prostaglandin E2 levels (Holtman et al., 2009).Rats were sacrificed 2 weeks after SE-induction and SC-treated ratswere compared to rats that were treated with vehicle.
EEG monitoring in chronic epileptic rats
To determine seizure frequency and duration in chronic epilepticrats, continuous EEG recordings were started 4 months after SEinduction. As previously described (Gorter et al., 2001; van Vlietet al., 2004b, 2008, 2006), a stable baseline of seizure frequencywas reached in chronic epileptic rats at this time point, and seizureclusters did not occur. Seizure parameters were recorded continu-ously, but were evaluated for 8 h/day, since this was the therapeuticwindow of PHT. Only rats that had frequent daily seizures (at least3 seizures from 09:00 a.m. to 05:00 p.m.) with a stable frequencyfor at least 5 days, were included in the study (n = 6).
Drug treatment in chronic epileptic rats
Rats that undergo SE develop a stable seizure frequency in thechronic epileptic phase approximately 3—4 months after SE induc-tion (Gorter et al., 2001). For this experiment, chronic epilepticrate were used that had a stable seizure frequency. These ratswere treated with SC (once daily 10 mg/kg, orally via gastric tubeunder short isoflurane anesthesia) for 14 days (n = 6). We previouslyshowed that this protocol reduced P-gp expression in chronic epilep-tic rats to control levels (van Vliet et al., 2010). Anesthesia neverlasted longer than 1—2 min and was necessary to handle epilepticrats. This did not affect daily spontaneous seizure frequency (vanVliet et al., 2006).
On the 14th day of SC treatment, phenytoin (PHT) treatmentwas started, while continuing SC treatment: the chronic epilep-tic rats received SC (10 mg/kg) and in addition once daily PHTsodium (5,5-diphenylhydantoin sodium salt dissolved in distilledwater alkalinized with NaOH, Katwijk Chemie, Katwijk, The Nether-lands; 75 mg/kg on the first day i.p., 50 mg/kg on the following days,i.p.). We previously showed that this protocol provided therapeu-tic blood levels of PHT (10—20 �g/ml) in rats for at least 8 h (vanVliet et al., 2006). PHT was given in combination with SC for 7 days.Hereafter PHT and SC treatment was stopped.
Phenytoin analysis in blood
Whole blood was taken from the tail vein under isoflurane anaesthe-sia and directly spotted on filter paper (Whatman 3). The dried bloodspots (8 mm) were punched into a test tube and shaken for 30 minwith 300 �l of an acetonitrile/methanol (1:3; v:v) mixture contain-ing 2.5 �g/ml ETB (internal standard). The extract was mixed with700 �l 10 mM KH2PO4 and 200 �l of the solvent was directly injectedinto the chromatograph. The HPLC system consisted of a P1000 sol-vent delivery system (Thermo Electron), a MIDAS automatic sampleinjector (Spark) and an UV 6000LP diode-array detector (ThermoElectron) at a wavelength of 205 nm. Data processing was per-formed with Spectrasystem PC1000 software (Thermo SeparationProducts). Separation was performed on a home-made reversedphase analytical column (15 cm × 0.46 cm) packed with Alltima 3C18(Alltech/Applied Science, Breda, The Netherlands) kept at a con-stant temperature of 35 ◦C. The mobile phase consisted of a mixtureof acetonitrile (14 vol%), methanol (29 vol%) and 40 mM phosphate
buffer pH 7.0 (57%) and was delivered isocratically at a flow rate of1.0 ml/min. ETB and PHT retention times were 11.2 and 13.8 min,respectively. The extraction recoveries were >95%. Within-day pre-cision for PHT was 3.1% for 10.8 �g/ml blood spot samples (n = 20)and the assay was linear in the range to at least 25 �g/ml.aalta
mporal lobe epilepsy 51
istology and immunocytochemistry
o determine the effects of SC on epileptogenesis, SC and vehicle-reated rats were sacrificed 14 days after SE-induction. Ratsere deeply anesthetized with pentobarbital (Nembutal, intraperi-
oneally, 60 mg/kg) and perfused through the ascending aorta with00 ml 0.37% Na2S solution followed by 300 ml 4% paraformaldehyden 0.1 M phosphate buffer, pH 7.4. The brains were post-fixed in situvernight at 4 ◦C. Brains were dissected and cryoprotected in 30%hosphate-buffered sucrose solution (pH 7.4). After overnight incu-ation at 4 ◦C, brains were frozen in isopentane (−32 ◦C) and storedntil sectioning (−80 ◦C). Brains were cut into 40 �m, horizontalections. Thereafter, sections of the hippocampus (5.32—5.82 mmrom Bregma, (Paxinos and Watson, 1998)) were processed asescribed previously for NeuN and OX-42 immunocytochemistry andimm’s stainings of the mossy fibers (Holtman et al., 2009). Thextent of synaptic reorganization of the mossy fibers was evaluatedy two observers according to a standardized 0—5 scale accordingo the extent and density of zinc stained granules (Cavazos et al.,992).
Sections were photographed using bright-field illumination onZeiss Axiophot microscope, equipped with a digital camera
MicroFire, Optronics, Golate, CA, USA), using StereoInvestigatoroftware (version 6) and imported into Adobe Photoshop (version.0). This program was used to optimize contrast and bright-ess, but not to enhance or change the image content in anyay.
tatistical comparisons
ll data are expressed as mean ± standard error of the mean (sem).two-tailed Fisher’s exact test was performed to test whether SC
reatment affected mortality or SE severity. Statistical comparisonsor SC and PHT treatment in chronic epileptic rats were performedsing the Wilcoxon Matched-pairs Signed-rank test. All the otherarameters were tested for statistical significance by the two-tailedtudent’s t-test. p < 0.05 was assumed to indicate a significant dif-erence.
esults
ffects of SC-58236 pretreatment on SE durationnd epileptogenesis
E-duration in the SC-treated group did not differ sig-ificantly from vehicle-treated animals: 11.3 ± 1.3 h (SCreatment) compared to 9.7 ± 0.2 h (vehicle treatment)Fig. 1(A)). An example of PED activity during SE is shownn Fig. 5(A). SE severity was scored according to the behav-or the animals showed during SE. In the SC-treated group,
out of 9 animals experienced severe SE, whereas 2 outf 9 from the vehicle-treated group experienced severe SE,hich was not significantly different. SE may lead to deathuring SE (acute death) or within days after SE inductionsubacute death). There was a significant higher mortalityfter SE in the SC-treated group than in the vehicle-treatedroup (p < 0.05, Fisher’s exact test): in the vehicle-treatedroup, 1 out of 9 animals died during SE. None of the ratsied later. However, in the SC-treated group, SE led to sub-
cute death in 6 out of 9 animals. None of the SC-treatednimals died during SE. Two SC-treated rats died after theatent period. All of the animals in the SC-treated grouphat experienced severe SE died subacutely (n = 4), as wells two animals that experienced mild SE. In the vehicle-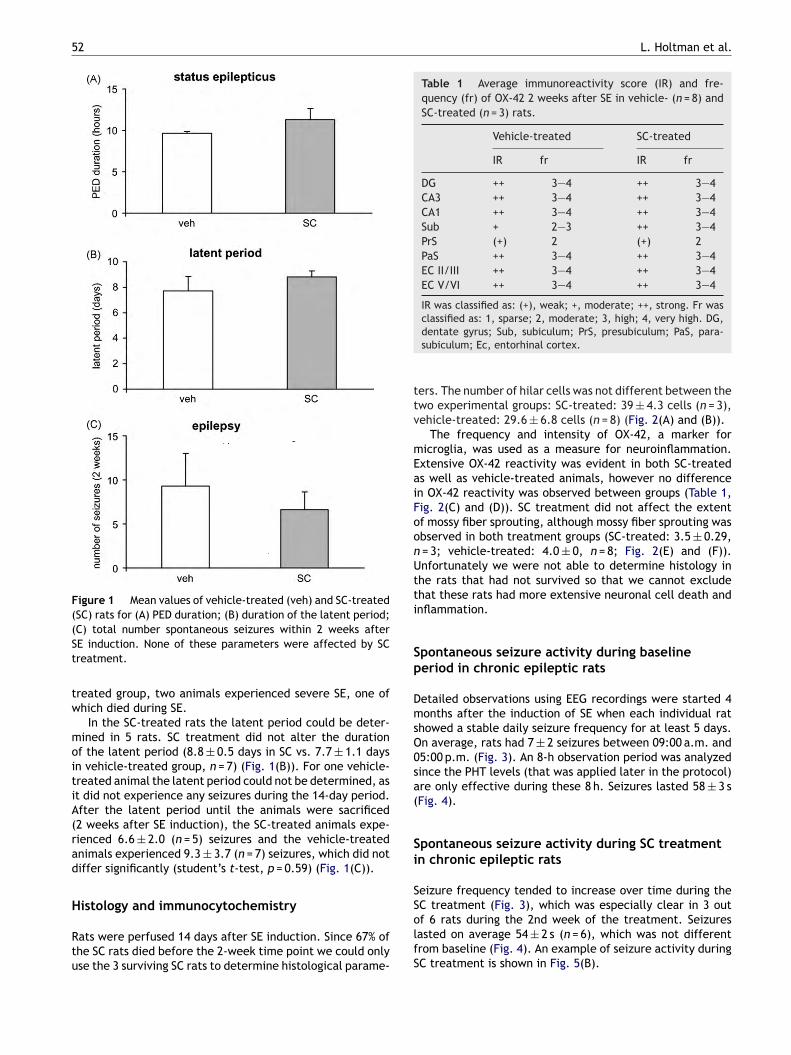
52 L. Holtman et al.
Figure 1 Mean values of vehicle-treated (veh) and SC-treated(SC) rats for (A) PED duration; (B) duration of the latent period;(St
tw
moitiA(rad
H
Rtu
Table 1 Average immunoreactivity score (IR) and fre-quency (fr) of OX-42 2 weeks after SE in vehicle- (n = 8) andSC-treated (n = 3) rats.
Vehicle-treated SC-treated
IR fr IR fr
DG ++ 3—4 ++ 3—4CA3 ++ 3—4 ++ 3—4CA1 ++ 3—4 ++ 3—4Sub + 2—3 ++ 3—4PrS (+) 2 (+) 2PaS ++ 3—4 ++ 3—4EC II/III ++ 3—4 ++ 3—4EC V/VI ++ 3—4 ++ 3—4
IR was classified as: (+), weak; +, moderate; ++, strong. Fr was
ttv
mEaiFoonUtti
Sp
DmsO0sa(
Si
SS
C) total number spontaneous seizures within 2 weeks afterE induction. None of these parameters were affected by SCreatment.
reated group, two animals experienced severe SE, one ofhich died during SE.
In the SC-treated rats the latent period could be deter-ined in 5 rats. SC treatment did not alter the duration
f the latent period (8.8 ± 0.5 days in SC vs. 7.7 ± 1.1 daysn vehicle-treated group, n = 7) (Fig. 1(B)). For one vehicle-reated animal the latent period could not be determined, ast did not experience any seizures during the 14-day period.fter the latent period until the animals were sacrificed2 weeks after SE induction), the SC-treated animals expe-ienced 6.6 ± 2.0 (n = 5) seizures and the vehicle-treatednimals experienced 9.3 ± 3.7 (n = 7) seizures, which did notiffer significantly (student’s t-test, p = 0.59) (Fig. 1(C)).
istology and immunocytochemistry
ats were perfused 14 days after SE induction. Since 67% ofhe SC rats died before the 2-week time point we could onlyse the 3 surviving SC rats to determine histological parame-
olfS
classified as: 1, sparse; 2, moderate; 3, high; 4, very high. DG,dentate gyrus; Sub, subiculum; PrS, presubiculum; PaS, para-subiculum; Ec, entorhinal cortex.
ers. The number of hilar cells was not different between thewo experimental groups: SC-treated: 39 ± 4.3 cells (n = 3),ehicle-treated: 29.6 ± 6.8 cells (n = 8) (Fig. 2(A) and (B)).
The frequency and intensity of OX-42, a marker foricroglia, was used as a measure for neuroinflammation.
xtensive OX-42 reactivity was evident in both SC-treateds well as vehicle-treated animals, however no differencen OX-42 reactivity was observed between groups (Table 1,ig. 2(C) and (D)). SC treatment did not affect the extentf mossy fiber sprouting, although mossy fiber sprouting wasbserved in both treatment groups (SC-treated: 3.5 ± 0.29,= 3; vehicle-treated: 4.0 ± 0, n = 8; Fig. 2(E) and (F)).nfortunately we were not able to determine histology inhe rats that had not survived so that we cannot excludehat these rats had more extensive neuronal cell death andnflammation.
pontaneous seizure activity during baselineeriod in chronic epileptic rats
etailed observations using EEG recordings were started 4onths after the induction of SE when each individual rat
howed a stable daily seizure frequency for at least 5 days.n average, rats had 7 ± 2 seizures between 09:00 a.m. and5:00 p.m. (Fig. 3). An 8-h observation period was analyzedince the PHT levels (that was applied later in the protocol)re only effective during these 8 h. Seizures lasted 58 ± 3 sFig. 4).
pontaneous seizure activity during SC treatmentn chronic epileptic rats
eizure frequency tended to increase over time during theC treatment (Fig. 3), which was especially clear in 3 out
f 6 rats during the 2nd week of the treatment. Seizuresasted on average 54 ± 2 s (n = 6), which was not differentrom baseline (Fig. 4). An example of seizure activity duringC treatment is shown in Fig. 5(B).
Cox-2 inhibition can lead to adverse effects in a rat model for temporal lobe epilepsy 53
Figure 2 Cell death (NeuN), microglia activation (OX-42) and mossy fiber sprouting (Timm) of horizontal sections (5.32—5.82 mmfrom Bregma) in vehicle-treated (A, C, E) and SC-treated (B, D, F) rats 2 weeks after SE. In vehicle-treated rats, only a few NeuN-positive hilar cells were present (A). In the surviving SC-treated rats, a similar pattern was observed (B). Extensive OX-42 reactivitywas evident in both vehicle-treated (C) as SC-treated (D) animals. No difference in OX-42 reactivity was observed between groups.
lecuuting
nSP
nlso
D
Tce
Timm’s staining showed mossy fiber sprouting into the inner morats. SC treatment did not affect the extent of mossy fiber spro
Spontaneous seizure activity during combinedSC/PHT treatment in chronic epileptic rats
One hour after the administration of PHT (75 mg/kg on thefirst day, 50 mg/kg on the consecutive 6 days), average PHTplasma levels were 18.4 ± 1.5 �g/ml. Eight hours after PHTadministration this level was reduced to 9.5 ± 1.4 �g/mlwhich is still within the therapeutic range of 10—20 �g/ml.No changes in PHT plasma values were observed duringtreatment.
Seizure frequency was significantly decreased (WilcoxonMatched-pairs Signed-rank test, p < 0.05) on the first 2days of PHT/SC treatment compared to the last dayof SC-treatment period (Fig. 3). Hereafter, seizure fre-quency increased and was not significantly different fromthe last day of SC treatment. No significant differences
were observed when the seizure frequency during PHT/SCtreatment was compared to the seizures that occurredduring the last 7 days of SC treatment. An exampleof seizure activity during SC/PHT treatment is shown inFig. 5(C).Sw3cc
lar layer of the dentate gyrus of vehicle (E) and SC-treated (F). Scale bar = 250 �m.
EEG seizures lasted on average 36 ± 2 s, which was sig-ificantly shorter compared to the baseline (58 ± 3 s) andC-period (54 ± 2 s; Fig. 4). One rat died 6 days after startingHT/SC treatment.
After the combined PHT/SC treatment was stopped, theumber of seizures initially increased and returned to base-ine values within a few days (Fig. 3). One rat died on theecond day of the wash-out period and two more rats diedn the third day of the wash-out period.
iscussion
he findings in this study indicate that treatment with theox-2 inhibitor SC-58236 starting before SE led to adverseffects with more rats dying within the first 2 weeks after
E. Moreover, 2 week SC treatment in chronic epileptic ratsith daily frequent seizures led to an increase of seizures inout of 6 rats during the 2nd week of treatment. Subsequentombined SC/PHT treatment transiently improved seizureontrol. Below we discuss the significance of these findings.
54 L. Holtman et al.
Figure 3 Effects of SC and SC/PHT on seizure frequency.During SC treatment seizure frequency tended to increaseover time, which was especially evident 13—14 days after SCtreatment. During SC/PHT treatment, the seizure frequency sig-nificantly decreased (Wilcoxon Matched-pairs Signed-rank test,p < 0.05) on the first 2 days of the treatment compared to thelast day of SC-treatment period. Hereafter, seizure frequencyincreased and was not significantly different from the last dayoda
P
TeaeeecIb
Figure 4 Effects of SC and SC/PHT on seizure duration. Onaverage seizures lasted 58 ± 3 s during baseline (n = 6). SC treat-mEs
pttma(tdrmavSr
FS
f SC treatment. After the end of the PHT/SC treatment anduring the recovery period, 3 animals died (indicated by therrows).
retreatment with SC and epileptogenesis
here are conflicting results concerning cox-2 inhibition inpileptic rats. Several studies indicate a neuroprotectivend anti-epileptogenic effect (Jung et al., 2006). Oth-rs have indicated no protective and no anti-epileptogenic
ffect (Holtman et al., 2009) or even a seizure aggravatingffect (Baik et al., 1999; Kim et al., 2008). These discrepan-ies could be in part due to the timing of the cox-2 inhibition.n the present study the cox-2 inhibition with SC was startedefore SE and was continued for 2 more days after SE. In atdviT
igure 5 Electrographic seizure activity. Electrographic recordingsC treatment in the chronic epileptic phase (B) and a seizure during
ent did not alter seizure duration. During SC/PHT treatment,EG seizures lasted on average 36 ± 2 s, which was significantlyhorter compared to the baseline and SC-period.
revious study we showed that we could effectively controlhe expression levels of P-gp (van Vliet et al., 2010) usinghis protocol. The hypothesis was therefore, that pretreat-ent with this cox-2 inhibitor would reduce inflammation
nd epileptogenesis. In agreement with a previous studyvan Vliet et al., 2010) SE severity was not different fromhe vehicle-treated rats and none of the SC-treated rats dieduring SE. However, we observed that considerably moreats died during the next 2 weeks after the SE/SC treat-ent. In the SC-treated rats that survived we did not detectchange in the duration of the latent period compared to
ehicle-treated rats. Although we cannot exclude that theC pretreatment also led to an increase or decrease of neu-onal cell death (we have no information on histology in
he rats that did not survive the treatment), neuronal celleath in the surviving SC-treated rats was not different fromehicle-treated rats. We observed an increase in mortalityn animals that were treated with SC before SE induction.his does not occur when SC treatment was started after SEshowing the 1—2 Hz PED activity during SE (A); a seizure duringSC/PHT treatment in the chronic epileptic phase (C).

or te
2IcospadV(fwaeptorwTis2
C
Citttcai2t
A
WDlo
FSa(
R
B
B
Cox-2 inhibition can lead to adverse effects in a rat model f
induction (Holtman et al., 2009), which might implicate thatsevere seizure activity, such as during SE, in combinationwith cox-2 inhibition can be lethal. An increased mortalityrate has also been observed after treatment with a cox-2inhibitor that starts before kainate-induced SE (Baik et al.,1999). Similarly, it has been shown that the combination ofsevere seizure activity and the presence of celecoxib can belethal (Kim et al., 2008). However, since these pretreatmenteffects are not found in the pilocarpine-induced SE model,such an effect might be dependent on the animal modelor on the pharmacokinetic properties of the cox-2 inhibitor(Bauer et al., 2008; Zibell et al., 2009).
Two-week cox-2 inhibition treatment in chronicepileptic rats
Previously, we studied the effects of a 5-day SC treatmentin chronic epileptic rats. This did not affect seizure activity(Holtman et al., 2009) or P-gp expression (van Vliet, 2007).However, in a later study we observed that a 2-week SCtreatment reduced P-gp expression to normal control levels(van Vliet et al., 2010). Since the extent of P-gp expres-sion has been related to seizure activity (van Vliet et al.,2004a) we reasoned that the reduced P-gp expression couldbe a reflection of a reduction in chronic seizure activity in 2-week SC-treated rats. However, contrary to our expectationthis was not the case and an increase in seizure frequencywas observed during the 2nd week of SC treatment in 3out of 6 rats. After the 2nd week of SC treatment 4 outof 6 rats died unexpectedly without known cause. Since thisonly rarely occurs in untreated chronic epileptic rats, it islikely that the chronic SC and/or SC/PHT treatment wasresponsible for this. Slow clearance and drug accumulationby repeated administration could eventually lead to too highdrug levels in the body and lead to renal, cardiovascular andcerebrovascular dysfunction (e.g. stroke), a side-effect thathas been reported for several cox-2 inhibitors after chronictreatment (Simmons et al., 2004). It is also possible thatthe normal upregulation of P-gp is a compensatory responseto protect the brain from xenobiotics that can more easilycross the leaky blood—brain barrier in an epileptic brain. Areduction of P-gp could then cause entrance of unwantedsubstances that are normally extruded by P-gp and that nowcan lead to detrimental effects in the brain. More studiesusing other cox-2 inhibitors should be performed to con-clude more definitively whether the combination of frequentseizures and chronic cox-2 inhibition will have debilitatingeffects on the course of epilepsy. This finding might warrantcaution for chronic use in epileptic patients, since manycox-2 inhibitors are commonly used as non-steroidal anti-inflammatory drugs (Simmons et al., 2004).
Seizure control after SC treatment
Previous research in our laboratory has indicated that pheny-toin (PHT) could not effectively control seizures in chronic
epileptic rats (van Vliet et al., 2006). Inhibition of P-gpwith tariquidar (TQD) however, effectively produced seizurecontrol during the first few days of PHT treatment. (vanVliet et al., 2006). The present strategy in which we targetcox-2 that leads to reduced P-gp expression (Bauer et al.,C
mporal lobe epilepsy 55
008) will have potential advantages over P-gp inhibition.nhibition of cox-2 will restore P-gp levels comparable toontrol levels and thereby maintain the protective functionf P-gp at the level of the blood—brain barrier. In previoustudies we could demonstrate that targeting cox-2 or therostaglandin EP1 receptor in the chronic epileptic brain ispromising strategy to control the expression levels of P-gpespite recurrent seizure activity (Pekcec et al., 2009; vanliet et al., 2010) and improves PHT delivery into the brainvan Vliet et al., 2010). The observed increase in seizurerequency at the end of the 2-week treatment with SC,as stopped with SC/PHT treatment. Although there wasreduction in seizure frequency for 2 days, this temporal
ffect on seizure frequency was not what was aimed for. Aossible explanation is that tolerance has developed. Dueo the decreased levels of P-gp, the resulting higher levelsf PHT in the brain might induce alterations in its targets,esulting in a reduced efficacy of PHT. This is comparable tohat we observed when P-gp was completely blocked withQD (van Vliet et al., 2006). TQD improved PHT delivery
nto the brain and led to an almost complete reduction ofeizures, but this effect was also temporarily and lasted fordays.
onclusion
ox-2 inhibition with SC-58236 had severe side effects withncreased mortality after SE as well as after 2-week chronicreatment. The results indicate that extreme care must beaken in case of chronic cox-2 inhibition in relation withhe disease process itself. Whether the effects of chronicox-2 inhibition on seizure activity is unique for SC-58236 orlso occur when other cox-2 inhibitors are used, needs to benvestigated. In any case, these data do no support a cox-
inhibition strategy as anti-epileptogenic or anti-epilepticherapy.
cknowledgements
e thank Pfizer for providing SC-58236. We also thank M.C.e Rijke and W.E. Dieters (Epilepsy Institute in The Nether-
ands Foundation, Heemstede, The Netherlands) for analysisf phenytoin in plasma.
This work was supported by the ‘‘Nationaal Epilepsieonds’’ grant 07-19 (JAG), The Netherlands Organisation forcientific Research (NWO) grant 863.08.017 (Veni) (EAvV)nd EU FP7 project NeuroGlia, grant agreement no. 202167EA).
eferences
aik, E.J., Kim, E.J., Lee, S.H., Moon, C., 1999. Cyclooxygenase-2 selective inhibitors aggravate kainic acid induced seizureand neuronal cell death in the hippocampus. Brain Res. 843,118—129.
auer, B., Hartz, A.M., Pekcec, A., Toellner, K., Miller, D.S.,
Potschka, H., 2008. Seizure-induced up-regulation of P-glycoprotein at the blood-brain barrier through glutamate andcyclooxygenase-2 signaling. Mol. Pharmacol. 73, 1444—1453.avazos, J.E., Golarai, G., Sutula, T.P., 1992. Septotemporal varia-tion of the supragranular projection of the mossy fiber pathway

5
G
G
G
H
H
I
J
J
K
K
P
P
R
R
S
v
v
v
v
v
v
V
6
in the dentate gyrus of normal and kindled rats. Hippocampus2, 363—372.
orter, J.A., van Vliet, E.A., Aronica, E., Lopes da Silva, F.H., 2001.Progression of spontaneous seizures after status epilepticus isassociated with mossy fibre sprouting and extensive bilateral lossof hilar parvalbumin and somatostatin-immunoreactive neurons.Eur. J. Neurosci. 13, 657—669.
orter, J.A., van Vliet, E.A., Aronica, E., Breit, T., Rauwerda, H.,Lopes da Silva, F.H., Wadman, W.J., 2006. Potential new anti-epileptogenic targets indicated by microarray analysis in a ratmodel for temporal lobe epilepsy. J. Neurosci. 26, 11083—11110.
ovoni, S., Masoero, E., Favalli, L., Rozza, A., Scelsi, R., Viappiani,S., Buccellati, C., Sala, A., Folco, G., 2001. The cycloxygenase-2inhibitor SC58236 is neuroprotective in an in vivo model of focalischemia in the rat. Neurosci. Lett. 303, 91—94.
ewett, S.J., Silakova, J.M., Hewett, J.A., 2006. Oral treatmentwith rofecoxib reduces hippocampal excitotoxic neurodegener-ation. J. Pharmacol. Exp. Ther. 319, 1219—1224.
oltman, L., van Vliet, E.A., van Schaik, R., Queiroz, C.M., Aronica,E., Gorter, J.A., 2009. Effects of SC58236, a selective COX-2inhibitor, on epileptogenesis and spontaneous seizures in a ratmodel for temporal lobe epilepsy. Epilepsy Res. 84, 56—66.
adecola, C., Niwa, K., Nogawa, S., Zhao, X., Nagayama, M., Araki,E., Morham, S., Ross, M.E., 2001. Reduced susceptibility toischemic brain injury and N-methyl-D-aspartate-mediated neu-rotoxicity in cyclooxygenase-2-deficient mice. Proc. Natl. Acad.Sci. U.S.A. 98, 1294—1299.
amali, S., Bartolomei, F., Robaglia-Schlupp, A., Massacrier, A., Per-agut, J.C., Regis, J., Dufour, H., Ravid, R., Roll, P., Pereira,S., Royer, B., Roeckel-Trevisiol, N., Fontaine, M., Guye, M.,Boucraut, J., Chauvel, P., Cau, P., Szepetowski, P., 2006. Large-scale expression study of human mesial temporal lobe epilepsy:evidence for dysregulation of the neurotransmission and com-plement systems in the entorhinal cortex. Brain 129, 625—641.
ung, K.H., Chu, K., Lee, S.T., Kim, J., Sinn, D.I., Kim, J.M.,Park, D.K., Lee, J.J., Kim, S.U., Kim, M., Lee, S.K., Roh,J.K., 2006. Cyclooxygenase-2 inhibitor, celecoxib, inhibits thealtered hippocampal neurogenesis with attenuation of sponta-neous recurrent seizures following pilocarpine-induced statusepilepticus. Neurobiol. Dis. 23, 237—246.
im, H.J., Chung, J.I., Lee, S.H., Jung, Y.S., Moon, C.H., Baik,E.J., 2008. Involvement of endogenous prostaglandin F2alphaon kainic acid-induced seizure activity through FP receptor: themechanism of proconvulsant effects of COX-2 inhibitors. BrainRes. 1193, 153—161.
unz, T., Oliw, E.H., 2001. The selective cyclooxygenase-2 inhibitorrofecoxib reduces kainate-induced cell death in the rat hip-pocampus. Eur. J. Neurosci. 13, 569—575.
axinos, G., Watson, C., 1998. The Rat Brain in Stereotaxic Coordi-nates, San Diego.
Z
L. Holtman et al.
ekcec, A., Unkruer, B., Schlichtiger, J., Soerensen, J., Hartz,A.M., Bauer, B., van Vliet, E.A., Gorter, J.A., Potschka, H.,2009. Targeting prostaglandin E2 EP1 receptors prevents seizure-associated P-glycoprotein up-regulation. J. Pharmacol. Exp.Ther. 330, 939—947.
acine, R.J., 1972. Modification of seizure activity by electricalstimulation. II. Motor seizure. Electroencephalogr. Clin. Neuro-physiol. 32, 281—294.
avizza, T., Gagliardi, B., Noe, F., Boer, K., Aronica, E., Vezzani,A., 2008. Innate and adaptive immunity during epileptoge-nesis and spontaneous seizures: evidence from experimentalmodels and human temporal lobe epilepsy. Neurobiol. Dis. 29,142—160.
immons, D.L., Botting, R.M., Hla, T., 2004. Cyclooxygenaseisozymes: the biology of prostaglandin synthesis and inhibition.Pharmacol. Rev. 56, 387—437.
an Vliet, E.A., 2007. The Role of the Blood—Brain Barrier and Mul-tidrug Transporters in Pharmacoresistant Epilepsy. Center forNeuroscience, University of Amsterdam, Amsterdam.
an Vliet, E.A., Aronica, E., Redeker, S., Marchi, N., Rizzi, M.,Vezzani, A., Gorter, J.A., 2004a. Selective and persistentupregulation of mdr1b mRNA and P-glycoprotein in the parahip-pocampal cortex of chronic epileptic rats. Epilepsy Res. 60,203—213.
an Vliet, E.A., Aronica, E., Tolner, E.A., Lopes da Silva, F.H., Gorter,J.A., 2004b. Progression of temporal lobe epilepsy in the ratis associated with immunocytochemical changes in inhibitoryinterneurons in specific regions of the hippocampal formation.Exp. Neurol. 187, 367—379.
an Vliet, E.A., Van Schaik, R., Edelbroek, P.M., Redeker, S., Aron-ica, E., Wadman, W.J., Marchi, N., Vezzani, A., Gorter, J.A.,2006. Inhibition of the multidrug transporter P-glycoproteinimproves seizure control in phenytoin-treated chronic epilepticrats. Epilepsia 47, 672—680.
an Vliet, E.A., van Schaik, R., Edelbroek, P.M., Lopes da Silva,F.H., Wadman, W.J., Gorter, J.A., 2008. Development of toler-ance to levetiracetam in rats with chronic epilepsy. Epilepsia 49,1151—1159.
an Vliet, E.A., Zibell, G., Pekcec, A., Schlichtiger, J., Edelbroek,P.M., Holtman, L., Aronica, E., Gorter, J.A., Potschka, H., 2010.COX-2 inhibition controls P-glycoprotein expression and pro-motes brain delivery of phenytoin in chronic epileptic rats.Neuropharmacology 58, 404—412.
ezzani, A., Granata, T., 2005. Brain inflammation in epilepsy:experimental and clinical evidence. Epilepsia 46, 1724—
1743.ibell, G., Unkruer, B., Pekcec, A., Hartz, A.M., Bauer, B., Miller,D.S., Potschka, H., 2009. Prevention of seizure-induced up-regulation of endothelial P-glycoprotein by COX-2 inhibition.Neuropharmacology 56, 849—855.




















