
Spatial Memory Following Temporal Lobe Resection
41
Running head: SPATIAL MEMORY Spatial Memory Following Temporal Lobe Resection Catherine M. Diaz-Asper 1 , Stephen Dopkins 2 , Samuel J. Potolicchio, Jr. 3 , Anthony Caputy 4 George Washington University Washington, DC 1 Clinical Brain Disorders Branch, NIMH, NIH 2 Dept. of Psychology, The George Washington University 3 Dept. of Neurology, The George Washington University 4 Dept. of Neurosurgery, The George Washington University Please address correspondence to: Stephen Dopkins Department of Psychology 2125 G Street, NW Washington, DC 20052 Tel: (202) 994-6315 Fax: (202) 994-1602 E- mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Spatial Memory Following Temporal Lobe Resection

Running head: SPATIAL MEMORY
Spatial Memory Following Temporal Lobe Resection
Catherine M. Diaz-Asper1, Stephen Dopkins2, Samuel J. Potolicchio, Jr.3, Anthony Caputy4
George Washington University
Washington, DC
1 Clinical Brain Disorders Branch, NIMH, NIH 2 Dept. of Psychology, The George Washington University 3 Dept. of Neurology, The George Washington University 4 Dept. of Neurosurgery, The George Washington University Please address correspondence to: Stephen Dopkins Department of Psychology 2125 G Street, NW Washington, DC 20052 Tel: (202) 994-6315 Fax: (202) 994-1602 E-mail: [email protected]

2
Abstract
The present study sought a clearer understanding of spatial memory function consequent to
temporal lobe resection, and, in particular, of spatial memory function with respect to two- as well as
three-dimensional frames of reference. Relative to a group of 15 control participants, a group of 15
epilepsy patients with right temporal resections demonstrated deficits of memory for locations in a two-
dimensional display. A group of 13 epilepsy patients with left temporal resections did not demonstrate
such deficits. The right and the left resection groups both demonstrated deficits of memory for item-
location relationships in a two-dimensional display. The right but not the left resection group
demonstrated deficits of memory for item-location relationships in a three-dimensional display. The
differing results that were observed for item-location relationships in two- and three-dimensional displays
were attributed to differences in the way item information is bound with location information concerning
two- and three-dimensional domains.
KEYWORDS: spatial memory, visual memory, medial temporal lobe, temporal lobe epilepsy

3
Spatial memory following temporal lobe resection
There is widespread agreement that structures within the human temporal lobes play a critical role
in memory function. The initial evidence for this view emerged from studies of the patient H.M. After
undergoing bilateral resection of the medial temporal lobe, H.M. was found to have severe deficits of
long-term declarative memory (Miller, Corkin, & Tueber, 1968; Sagar, Cohen , Corkin, & Growdon,
1985; Scoville & Milner, 1957).
H.M.’s memory deficit has subsequently been modeled fairly successfully in non-human primates
(Gaffan, 1994; Murray & Mishkin, 1986; Ridley, Timothy, MacLean, & Baker, 1995; Suzuli, Zola-
Morgan, Squire, & Amaral 1993; Zola, Squire, Teng, Stefanacci, & Clark, 2000). Across delays ranging
from seconds to minutes, monkeys with similar medial temporal ablations have demonstrated deficits on
visual and tactual non-matching- to-sample tasks and on matching-to- location tasks (Squire, 1992; Zola-
Morgan & Squire, 1985). Further work with primates has attempted to localize memory function more
precisely. For example, deficits on simple non-matching-to-sample tasks have been demonstrated in
monkeys with damage to the perirhinal and entorhinal cortex (Murray & Mishkin, 1986; Suzuli et al.,
1993). In contrast, less severe impairments have been demonstrated on such tasks in monkeys with
damage restricted to the hippocampus [Zola et al., 2000).
At the same time, human neuropsychological research has sought to identify the substrates of
different sorts of long-term declarative memory within the medial temporal region. Early work explored
the lateralization of such memory. Dysfunction of the left medial temporal lobe was associated with
deficits of verbal memory (Blakemore & Falconer, 1967; Kimura, 1963; Pillon, Bazin, Deweer, Ehlre,
Baulac, & Dubois, 1999). Dysfunction of the right medial temporal lobe was associated with deficits of
nonverbal memory, including memory for visual and spatial information (Milner, 1968, Pigott & Milner,
1993; Pillon et al., 1999; Rains & Milner, 1994; Smith & Milner, 1981; Smith & Milner, 1989).

4
More recently, efforts have been made toward a fuller articulation of the effects of human medial
temporal dysfunction on long-term nonverbal memory. Few definitive conclusions have been reached,
however. One complicating factor is that many different facets of nonverbal memory can be
distinguished. Within spatial memory, for example, a distinction can be drawn on the basis of the
underlying reference frame, which can be either two- or three-dimensional (Stepankova, Fenton,
Pastalkova, Kalina, & Bohbot, 2004). An independent distinction can be drawn between location
memory, in which one simply remembers a particular location in a spatial domain, and item-location
memory, in which one remembers the location of a particular item in a domain (Rains & Milner, 1994;
Smith & Milner, 1981; Smith & Milner, 1989). Within three-dimensional spatial memory, a further
distinction can be drawn between egocentric memory, in which locations are remembered with reference
to one’s own location, and allocentric memory, in which locations are remembered without reference to
one’s own location (Feigenbaum & Morris, 2004; Goldstein, Canavan, & Polkey, 1989; Holdstock,
Mayes, Cezayirli, Aggleton, & Roberts, 1999; Incisa della Roccchetta, Samson, Ehrle, Denos, Hasboun,
& Baulac, 2004; O’Keefe & Nadel, 1978).
The present study attempted a relatively comprehensive and analytical examination of spatial
memory consequent to medial-temporal dysfunction. In particular, the object was to examine two- as well
as three-dimensional spatial memory. This was deemed to be important for two reasons. First, the bulk of
previous work in spatial memory has focused on memory either for three-dimensional displays or for
representations of three-dimensional scenes (Goldstein et al., 1989; Nunn, Graydon, Polkey, & Morris,
1999; Nunn, Polkey, & Morris, 1998; Pigott & Milner, 1993; Rains & Milner, 1994; Smith & Milner,
1981; Smith & Milner, 1989). Consequently, little is known about the effects of temporal lobe
dysfunction on two-dimensional spatial memory. As the results of the present study show, two- and three-
dimensional spatial memory do not always follow the same pattern. Second, imaging studies have

5
recently begun to explore two-dimensional spatial memory (Johnsrude, Owen, Crane, Milner, & Evans,
1999; Moscovitch, Kapur, Kohler, & Houle, 1995; Owen, Milner, Petrides, & Evans, 1996). It is
important to seek convergence between neuropsychological and imaging data.
Three groups of participants were tested: a group that had undergone resection of the anterior right
temporal lobe (RTLR), a group that had undergone resection of the anterior left temporal lobe (LTLR),
and a group of healthy adults. Portions of the hippocampus as well as the parahippocampal gyrus had
been resected in the RTLR and LTLR groups.
Experiment 1 examined two-dimensional spatial memory, making a distinction between location
and item-location memory. To establish a context for understanding item-location memory, the
experiment also tested two-dimensional visual memory for item information. Experiment 2 examined
three-dimensional item-location memory, making a distinction between egocentric and allocentric
memory.
The study differed in several methodological respects from many previous studies in this area.
First, the stimulus items for the tests of item and item-location memory were abstract visual figures rather
than objects. In this respect, the study followed Nunn et al. (1998) rather than several other previous
studies (Nunn et al., 1999; Owen et al., 1996; Smith & Milner, 1981; Smith & Milner, 1989). The
rationale here was that the use of abstract stimuli would limit the involvement of verbal processes in
performance of the tasks, and clarify the implications of the results with respect to spatial memory.
Second, the different tasks in each experiment were matched for difficulty on the basis of pilot
data. Thus, it was expected that the control participants would not differ in performance on the tasks for a
given experiment. The object was to assess the deficiency of the two TLR groups relative to the control
participants on each task. This is in contrast to past studies, which have either made no attempt to match
tasks for difficulty (Barr, 1997; Goldstein et al., 1989; Hermann, Seidenberg, Wyler, & Haltiner, 1993;

6
Owen et al., 1996; Smith & Milner, 1981; Smith & Milner, 1989) or, alternatively, have used a group-
matching procedure in which TLR and control participants are matched for performance on tasks of one
sort and then deficits are demonstrated in the TLR participants on tasks of another sort (Nunn et al.,
1998; Nunn et al., 1999). The virtues of the present procedure are twofold. First, it avoids problems
associated with the group-matching procedure, namely that TLR and control participants typically have to
be tested under different conditions (e.g. different retention intervals) in order to achieve equivalent levels
of performance on the task on which they are matched. It is conceivable that different sorts of memory
may be called into play under these different conditions (e.g. different sorts of memory may be involved
at different retention intervals). The second virtue of the present procedure is that it allows for more
efficient use of the experimental data. In particular contrast to the group matching procedure (Nunn et al.,
1998; Nunn et al., 1999), it is possible under the present procedure to assess the performance of the RTLR
and LTLR participants relative to the control participants on all tasks for a given experiment.
Experiment 1
This experiment tested the two-dimensional spatial memory of RTLR, LTLR, and control
participants, making a distinction between location and item-location memory. To aid in understanding the results
of the item-location memory test, the experiment also tested two-dimensional visual memory for item information.
The object was to discover whether the RTLR and/or LTLR participants differed from the control
participants on any of these forms of memory. Each of the experimental tasks consisted of an exposure
phase, in which a sequence of target stimuli was presented, and a test phase, in which the participant
attempted to distinguish the target stimuli from distractor stimuli.
The item task tested two-dimensional visual memory for the sorts of information used in
identifying items. Participants were required to remember the identities of abstract visual figures,
presented on a computer screen. The location task tested two-dimensional location memory. Participants
were required to remember locations on a computer screen. The item-location task tested two-dimensional

7
item-location memory. Participants were required to remember the locations of abstract visual figures on
a computer screen.
Method
Participants
Temporal lobe resection participants. These were drawn from a group of 43 patients who had
undergone a selective unilateral resection of the anterior temporal lobe as therapy for intractable epilepsy
(Weibe, Blume, Girvin, & Eliasziw, 2001). The seizure foci for the patients were idenitified with ictal
video-EEG monitoring. The extent of the resection for each patient was determined by intra-operative
EEG measurement. An initial portion of the anterior- lateral temporal lobe was first defined by
electrocortographic mapping. Following the resection of this portion, the electrocorticography array was
again placed on the temporal lobe. Recordings were obtained from the uncut surface of the temporal lobe,
from the resection surface, and from the hippocampus. Stimulation was again employed, including direct
stimulation of the hippocampus. If an electrically-active focus was defined, further cortical resection was
undertaken until the area was quiescent from an eleptogenic standpoint. The medial temporal structures,
including the amygdala and a portion of the hippocampus (3.0-cm) were then resected using
microsurgical techniques. The fornix was spared.
Because age at seizure onset, disease duration, seizure frequency (Dodrill & Matthews, 1992), and
anti-epileptic drug treatment (Devinsky, 1995), are known to influence memory abilities in temporal lobe
epileptics, participants in the two TLR groups were matched on these factors. The resulting sample
contained 31 patients (16 RTLR and 15 LTLR). One RTLR patient withdrew from the study after
completing one task, and two LTLR patients were excluded after being tested, due to the presence of
potentially confounding neurological conditions (cerebral palsy and spina bifida). Thus, twenty-eight
patients were included in the final analyses (15 RTLR and 13 LTLR). Two of the LTLR participants had

8
superior quadrant deficits. Because these participants achieved the required criterion on the
perceptual/attentional screening task (to be described later), they were retained in the study.
Surgeon’s notes indicated that the final group of RTLR participants had undergone resections of
the inferior, middle, and superior gyri, averaging, respectively, 3.29, 2.89, and 1.50 cm from the temporal
pole, with standard deviations of .34, 1.05, and 1.43 cm, respectively. The LTLR participants had
undergone resections averaging, respectively, 2.95, 2.55, and 1.05 cm, with standard deviations of .71,
1.16, and .99 cm, respectively. Notice that more cortex was resected from the inferior than the superior
gyrus; thus the resection involved an oblique cut extending from anterior/rostral to posterior/caudal.
Notice, also, that the amounts of cortex resected from the right and left TLR participants were more
nearly equivalent than would have been the case under some other resection procedures. In fact, the
amounts of tissue resected from the three cortical areas did not differ in the RTLR and LTLR groups
(inferior: F(1,22) = 1.43, MSe = .458; middle: F(1,22) = 1.93, MSe = .355; superior: F(1,22) < 1. These
and all other statistical tests were conducted against an alpha value of .05.). Analysis of post-surgery
imaging data for the final group of participants indicated that the procedure removed 25-40% of the
temporal lobe, including the amygdala and approximately 40% of the hippocampus and the surrounding
parahippocampal gyrus. Pathology reports indicated the presence of hippocampal atrophy or sclerosis for
9 of the RTLR and 9 of the LTLR participants. Tumors were resected from two of RTLR participants. All
TLR participants were tested at a minimum of 5 months following their surgery. All were left-hemisphere
dominant for speech.
Healthy control participants. These were drawn from a pool of 19 healthy volunteers. Criteria for
inclusion in the pool included the stated absence of any potentially confounding medical or neurological
conditions (e.g., history of head injury, serious cardiovascular disease, or stroke), and the stated absence
of drug and alcohol abuse. Six of the 19 participants in the pool were relatives or friends of TLR

9
participants. The remaining 13 participants were recruited from the surrounding community. From this
pool, a sample of 15 individuals was selected that were matched to the patient groups in terms of age, sex,
handedness, years of education, ethnicity, and estimated intelligence level.
Intelligence (IQ) was assessed for all participants using the Information and Picture Completion
subtests of the Wechsler Adult Intelligence Scale- Revised (WAIS-R) (Weschler, 1981). A composite
score was obtained from these subtests with an age-adjusted formula created by Kaufman, Ishikuma, and
Kaufman-Packer (1991) (see also Boone (1992)). Strength of handedness was ascertained using the
Edinburgh Handedness Inventory (Oldfield, 1971). No participant (TLR or control) had an estimated IQ
lower than 80, or an age less than 16 or greater than 60. All participants provided written informed
consent prior to undertaking the experiments.
Demographic information for the three participant groups is presented in Table 1. The groups did
not differ in terms of age (F(2,40)=1.01), sex (Χ2(2,N=43)=2.11), estimated IQ (F(2,40)=.48), handedness
(F(2,40)=.86), or education level (F(2,40)=.09). The two TLR groups did not differ in terms of age at
seizure onset (F(1,26)=.13), duration of epilepsy (F(1,26)=.29), current seizure frequency (F(1,26)=2.20),
or number of anti-epileptic medications taken (F(1,26)=.08).
Insert Table 1 here
Materials
Each target and distractor display for the item task consisted of an abstract visual figure in the
center of the computer screen. The figures were constructed, in the manner of Musen and Treisman
(1990) and Blaxton and Theodore (1997), by connecting points in a 3 X 3 array of points. To construct a
given figure, five pairs of points, chosen at random, were connected, with the constraints that (a) the
resulting line segments formed a connected figure, (b) no line segment was repeated in either direction,
and (c) a line did not simultaneously cross a column and a row in the point array. Any repetitions of

10
figures, or highly verbalizable figures (e.g., common shapes or alphanumeric characters) were eliminated
through visual inspection. The array points were erased for the final presentation of the figures. Each
figure was approximately 2 cm X 2 cm in size.
Each target and distractor display for the location task consisted of a 2 cm X 2 cm white square at
a certain location within a region that was defined on the computer screen by a continuous irregular
border. The border remained constant across all displays. The rationale for including this border depended
on the distinction that has been proposed between the categorical and the coordinate modes of spatial
processing. According to this proposal, relationships such as “above,” “beside,” etc. are assessed with
categorical processing, whereas quantitative measures of distance are assessed with coordinate
processing. Categorical processing is based in the left hemisphere, whereas coordinate processing is based
in the right hemisphere (Kosslyn, 1987; Kosslyn, 1994). The border was included to encourage nonverbal
coordinate involvement in the location task. It was reasoned that participants would be encouraged to
locate the white square in a given display in terms of a set of distances from points on the border.
Each target and distractor display for the item-location task consisted of an abstract visual figure,
of the same sort used for the item task, at a certain location within the same irregular-bordered region that
was used for the location task (See Figure 1).
Insert Figure 1 here
Because the testing phase for each task required participants to emit positive responses to target
displays and negative responses to distractors, the stimulus set for each task was composed of pairs of
displays, with one member of each pair serving as the target, and the other as the distractor. For the item
task, the members of each target/distractor (T/D) pair were visually similar (with at least 3 out of 5 line
segments in common). For the location task, the members of each T/D pair resembled one another in the
placement of the white square. The position of the square differed by no less than 1 cm but no more than 3

11
cm. For the item-location task, a distractor was formed for each target by placing the visual figure of the
target in a position previously occupied by the visual figure of a different target during the exposure
phase. The displays for the exposure and test phases of the three tasks were presented in the same quasi-
random order to all participants.
Task matching. The tasks for the experiment were matched for difficulty using the procedure of
Chapman and Chapman (1978). The procedure involved pilot testing two parallel sets of 15 T/D pairs for
each task. Hence, 90 T/D pairs were tested overall. Participants comprised a convenient sample of 136
undergraduate students, who received class credit for their participation. Sixty-three participants piloted
one set of T/D pairs for all three tasks, and 67 piloted a second set. The testing procedure was the same as
is later described for the experiment proper. When pilot testing was completed, the corrected recognition
(hits – false alarms) score for each T/D pair was calculated. Fifteen T/D pairs were then selected for the
final version of each task, with the corrected recognition score for each selected pair coming from the
middle of the distribution for its set of T/D pairs. Different visual figures were used in the pairs for the
three tasks. The 15 T/D pairs that were selected for the item, location, and item-location tasks had mean
corrected recognition scores of .29, .31, and .32, respectively. The Kuder-Richardson formula 20 (a
coefficient of internal consistency that is used for items scored dichotomously) was then computed for the
T/D pairs that had been selected for each task. The Kuder-Richardson scores for the item, location, and
item-location tasks were .75, .84, and .93, respectively. Finally, an additional 6 participants were tested on
the T/D pairs that had been selected for the three tasks. In this sample, the mean corrected recognition
scores for the three tasks were .31, .34, and .35. The Kuder-Richardson scores were .76, .86, and .93.
Procedure
Perceptual/attentional screening. Although recent work suggests that temporal lobectomy patients
do not suffer from simple deficits of perception and attention, all participants were screened to ensure

12
adequate perceptual and attentional capacities prior to undertaking the experimental tasks (Mendola,
Rizzo, Cosgrove, Cole, Black, & Corkin, 1999). The screening task was computer-based and included
components that tested perception and attention with respect to item and location information. All
participants completed the item component first. This consisted of 30 trials (excluding two practice trials
that served to familiarize participants with the task). At the beginning of each trial, a visual figure (of the
same sort as was used in the experiment) was presented for 150 ms (Hellige & Michimata, 1989; Rybash
& Hoyer, 1992). Following a 1-second delay, a second figure was presented for 150 ms. Participants were
required to indicate, as quickly as possible, whether the second figure matched the first. The location
component was similar, except that the visual figure was presented at a certain location within the region
that was defined in the center of the computer screen by the irregular border. Participants were required
to determine whether the second presentation of the figure occurred in the same location as the first. The
delay interval between the two presentations in each trial was kept deliberately brief (1 sec) to ensure that
perceptual and attentional, rather than mnemonic, processes were engaged (Hellige & Michimata, 1989).
On the basis of findings with a pilot sample, participants were required to achieve at least 25 out of a
possible 30 correct responses on each component of the screening task in order to be included in the
study. Both of the LTLR participants with superior quadrant deficits attained this level of performance.
One TLR participant was excluded for failure to attain this level of performance.
Overview of experiment proper. Each of the three tasks consisted of an exposure phase and a test
phase. The test phase for each task followed its associated exposure phase (e.g., item exposure phase,
followed by item test phase; location exposure phase, followed by location test phase, etc.). Previous
studies assessing long-term memory function have used varying retention intervals. The delay between
the exposure and the test phase has ranged from minutes to weeks (Abrahams, Pickering, Polkey, &
Morris, 1997; Smith & Milner, 1981). Previous studies have also varied in whether or not the participant

13
was actively engaged in a “filler” task between exposure and test. The present experiment used a three-
minute, filled, retention interval. The filler task consisted of questions pertaining to the participant’s
medical or social background. The presentation order of the three tasks was counterbalanced across
participants.
Exposure phase. The exposure phase for the item task consisted of 15 trials, on each of which a
different visual figure was presented in the center of the computer screen (without the irregular border).
Each trial lasted for 5 seconds, and was followed by a 3-second inter-trial interva l. Participants were
instructed to remember the figures for later testing. The exposure phase for the location task consisted of
15 trials, on each of which a white square was presented in a different location within the region that an
irregular border defined on the computer screen. The white square and the irregular border were the same
for all trials. Each trial lasted for 5 seconds and was followed by a 3-second inter-trial interval.
Participants were instructed to remember the indicated locations for later testing. The exposure phase for
the item-location task consisted of 15 trials, on each of which a different visual figure was presented in a
different location within the irregular-bordered region. Each trial lasted for 5 seconds, and was followed
by a 3-second inter-trial interval. Participants were instructed to remember the figures and their locations
for later testing.
Test phase. The test phase for each of the three tasks commenced immediately following the three-
minute filled delay. Participants received both verbal and on-screen instructions immediately prior to each
testing phase. For the item task, 30 displays (15 targets and 15 distractors) were presented, in sequence.
Each display consisted of a visual figure in the center of the computer screen (without the irregular
border). Participants indicated whether the figure had been presented during the exposure phase. For the
location task, 30 displays (15 targets and 15 distractors) were presented. Each display consisted of the
white square in a certain location within the irregular-bordered region. Participants indicated whether or

14
not the square had been presented in the indicated location during the exposure phase. For the item-
location task, 30 displays (15 targets and 15 distractors) were presented. Each display consisted of a visual
figure in a certain location within the irregular-bordered region. Participants indicated whether the figure
had been presented in the indicated location. For all three tasks, participants pressed the “Y” and the “N”
key to make their responses.
Participants also completed a de-briefing questionnaire regarding their performance in the three
tasks. The questionnaire asked participants to describe the mnemonic strategies that they had used in
performing each task. For the item task, participants were asked whether they had been reminded of
familiar things when studying the figures, whether they had formulated names or verbal descriptions for
the figures, whether they had used some other strategy, or whether they had used no particular strategy.
For the location task, participants were asked whether they had remembered the location of the white
square relative to the contour of the irregular border, whether they had remembered the location relative
to the shape of the computer screen, whether they had remembered the location in some other way, or
whether they had used no particular strategy. For the item-location task, participants were asked whether
they had used a combination of the strategies indicated earlier, whether they had used some other strategy,
or whether they had used no particular strategy.
Results
Memory Tests
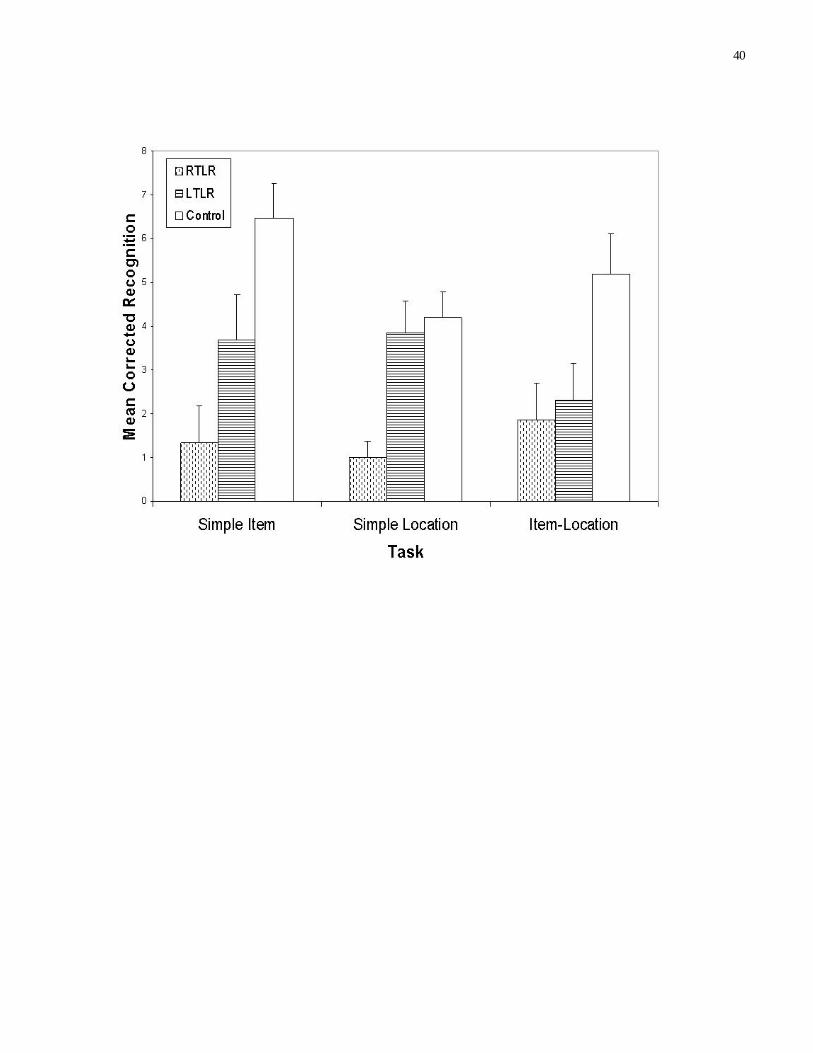
Figure 2 illustrates the mean corrected recognition score for each of the three tasks as a function of
group membership. A series of one-way ANOVA’s revealed significant group differences on the Item
task (F(2,40)=8.96, MSe=.049), the Location task (F(2,40)=10.21, MSe=.020), and the Item-Location task
(F(2,40)=4.52, MSe=.048).
Insert Figure 2 here

15
Planned comparisons revealed significant differences between the RTLR participants and the
control participants on the item task (F(1,28) = 19.97, MSe = .043, Partial Eta Squared = .42), the location
task (F(1,28) = 21.42, MSe = .015, Partial Eta Squared = .43), and the item-location task (F(1,28) = 7.38,
MSe = .05, Partial Eta Squared = .21). In addition, significant differences were revealed between the
LTLR participants and the control participants on the item (F(1,26) = 4.76, MSe = .05, Partial Eta
Squared = .16) and the item-location tasks (F(1,26) = 5.31, MSe = .049, Partial Eta Squared = .17), and
between the RTLR and the LTLR participants on the location task (F(1,26) = 13.88, MSe = .018, Partial
Eta Squared = .35).
Ninety-five percent confidence intervals were examined for the corrected recognition data from
each group to ascertain the likelihood that memory capacity was completely absent, with the result that
the data reflected random performance. For the LTLR and the control participants, confidence intervals
ranged from 0.47 to 8.20 across the three tasks. As a result, it is reasonably certain that the observed data
do not reflect random performance. The same can be said for the performance of the RTLR participants
on the item-location task (CI = .16 to 3.58). However, the performance of the RTLR participants on the
item (CI = -.40 to 3.07) and the location (CI = -.10 to 2.10) tasks suggests that performance may have
been random here.
Although the differences were not significant, the three participant groups did nonetheless differ in
estimated IQ and distribution of males and females. Therefore, one-way ANCOVAs were calculated for
the three groups on the three tasks, with sex and estimated IQ as covariates, to ascertain whether these
variables affected performance. The main effect of the covariates was not significant for the location and
the item-location tasks, but there was a significant effect of estimated IQ (F(1,38)=4.68, MSe=.041) and
sex (F(1,38)=4.83, MSe=.041) on item task performance. After controlling for the effects of estimated IQ
and sex, the main effect of group on the item task remained highly significant (F(2,38)=9.66, MSe=.041).

16
Pairwise comparisons revealed the same pattern as reported above: The RTLR (F(1,26) = 19.63, MSe =
.041, Partial Eta Squared = .34) and the LTLR (F(1,24) = 8.86, MSe = .036, Partial Eta Squared = .27)
participants both performed significantly more poorly than the control participants on this task.
A multivariate repeated-measures ANOVA confirmed that the control group’s performance was
matched across the three experimental tasks (F(2,13)=1.63).
Mnemonic Strategies
In order to determine whether the groups differed in the strategies used for memorizing the target
stimuli, chi-square analyses were performed on the results of the de-briefing questionnaire. On the item
task, a majority of the participants in each group (11/13 LTLR, 13/15 RTLR, and 11/15 control
participants) stated that they were reminded of familiar things when studying the figures. The groups did
not differ in their use of this strategy (Χ2 (2,N=43)=1.01). On the location task, a majority of the
participants in each group (12/13 LTLR, 14/15 RTLR, and 14/15 control) stated that they remembered the
location of the white square relative to the contour of the irregular border. Again, the groups did not differ
in their use of this strategy (Χ2 (2,N=43)=.02). On the item-location task, a majority of the participants in
each group (8/13 LTLR, 11/15 RTLR, and 11/15 control) used the former two strategies in combination.
Again, the three groups did not differ in their choice of strategy for this task (Χ2 (2,N=43)=4.72).
Memory Test Performance and Size of Resection
The surgeon’s notes for the TLR participants listed measurements across three regions of the
temporal lobe: the inferior gyrus, the middle gyrus, and the superior gyrus. Analyses were conduced for
14 of the 15 RTLR participants to assess the correlation between measures of performance on the three
tasks and amounts of tissue resected from the three regions. (The surgeon’s note for one RTLR participant
provided non-specific resection information). A constant amount of hippocampal tissue was removed in
each resection, so this structure was not considered in the analysis. None of the correlations between the

17
task variables and the resection measurements was significant at the .05 level. The same analyses were
also conducted for 10 of the 13 LTLR participants, revealing no significant relationships. The results of
the correlational analyses are presented in Table 2.
Insert Table 2 here
Discussion
None of the tasks was more difficult than any other for the control participants. By implication,
the preliminary task-matching process was successful.
The RTLR and LTLR participants both performed more poorly than the control participants on the
Item task. These results are somewhat unexpected, given that the majority of previous reports have linked
visual memory deficits with right temporal dysfunction (Barr, 1997; Jones-Gotman, Zatorre, Olivier,
Andermann, Cendes, Staunton, McMackin, Siegel, & Wieser, 1997; Milner, 1968). The results are not
unprecedented, however. Several previous reports have linked visual memory deficits with left temporal
dysfunction or with left and right dysfunction (Hornak, Oxbury, Oxbury, Iverson, & Gaffan, 1997;
Maguire & Cipolotti, 1998). Future work will be necessary to disentangle the factors that control visual
memory deficits in temporal lobe patients.
The results for the item task imply that the information needed to identify the stimulus items was
bilaterally represented. Several conceptions of this bilateral representation are possible. On one hand, the
items might have been represented differently in the two hemispheres – in the verbal modality in the left
hemisphere and in the visual modality in the right hemisphere. On the other hand, the stimuli might have
been represented in the same way in the two hemispheres - in terms that were neither verbal nor visual but
that instead might be called amodal. Several points can be made against the joint verbal-visual
conception. First, it goes against the principle of parsimony. Second, the visual figures used in the present
study were designed to be resistant to verbal coding. Third, a majority of participants in all of the groups

18
indicated on the de-briefing questionnaire that they performed the item task by noting resemblances
between the figures and familiar things but not by formulating names or verbal descriptions for the
figures. For example participants may have called up a conceptual representation of a thing that a given
figure resembled. Because the representation existed at the conceptual level it would have reflected visual,
auditory, olfactory, and tactile aspects of the object. In general, then, the results are consistent with the
view that the item memory process represented the figures in amodal terms.
The RTLR participants performed more poorly than the LTLR and the control participants on the
location task. These results reinforce previous reports of a location memory deficit in RTLR patients. The
results constitute useful new information, however. Whereas most previous studies have shown deficits of
three-dimensional location memory in RTLR patients (O’Reilly & Rudy, 2001), the present results show
deficits of two-dimensional location memory. As subsequent results will show, two- and three-
dimensional spatial memory do not always show the same pattern.
The RTLR and LTLR participants performed more poorly than the control participants on the
item-location task. These results again reinforce previous reports of an item-location memory deficit in
RTLR patients. Again, however, the results constitute new information. Whereas most previous studies
have shown deficits of three-dimensional item-location memory (Goldstein et al., 1989; Nunn et al., 1998;
Nunn et al., 1999; Smith & Milner, 1981; Smith & Milner, 1989), the present results show deficits of two-
dimensional item-location memory. Furthermore, whereas previous RTLR deficits of three-dimensional
item-location memory have occurred in the context of intact LTLR performance, the present RTLR
deficits of two-dimensional item-location memory occur in the context of similar LTLR deficits.
In interpreting the results for the item-location task, we must bear in mind that the item and
location tasks, on one hand, and the item-location task, on the other hand, made independent demands
upon participants. Whereas the item and location tasks required participants to remember the figures and

19
locations that had been presented, the item-location task provided participants with the figures and
locations that had been presented and required them to remember the way the figures and locations had
been linked. Thus, the item-location task tapped the binding of items and locations (Chalfonte & Johnson,
1996; O’Reilly & Rudy, 2001; Shimamura, 2002). What, then, are the implications of these results?
Notably, the information that bound items and locations was bilaterally recorded. Thus, the pattern was
the same as for item information and different from the pattern for location information. This issue will be
discussed in greater detail later.
Experiment 2
This experiment tested the three-dimensional item-location memory of the RTLR, LTLR, and
control participants, making a distinction between egocentric and allocentric item-location memory. The
object was to discover whether the RTLR and/or LTLR participants differed from the control participants
in either egocentric or allocentric three-dimensional item-location memory and to compare the degree to
which the RTLR and/or LTLR participants differed from the controls in the two sorts of memory.
During the exposure phase of the egocentric item-location task, a series of displays was presented,
with each display consisting of an abstract two-dimensional visual figure in a certain location on a
tabletop surface. During the test phase, participants distinguished displays that had been presented during
the exposure phase from new displays in which old figures were presented in new locations. The
procedure for the allocentric item-location task was the same as for the egocentric item-location task,
except that the displays were rotated either 90, 180, or 270 degrees between the exposure and the test
phase. Goldstein et al. (1989) used a similar procedure to test egocentric and allocentric item-location
memory.
Method

20
Materials
Each target and distractor display was constructed in the same way as for the item-location task of
Experiment 1. The displays were printed in black ink on sheets of 8.5”X11” paper, approximately the
same size as in Experiment 1. The sheets on which the displays were printed were contained within a 3”
ring-binder. Separate sheets of green paper were placed between the display sheets for masking purposes.
The 15 T/D pairs for both the egocentric and allocentric item-location tasks were formed in the
same way as the T/D pairs for the item-location task of Experiment 1. For the test phase of the allocentric
item-location task, the sheets on which the target displays were printed were rotated clockwise in
increments of 90°. (Notice that the target figures remained in the same orientation relative to the irregular
border, because the whole sheet was rotated). Nine sheets were rotated 90° at test, 11 were rotated 180°,
and 10 were rotated 270°. The displays for the exposure and test phases of the two tasks were presented
in the same quasi-random order to all participants.
Task matching. This was carried out as for Experiment 1. Two parallel sets of 15 T/D pairs were
pilot tested for each task. Hence, 60 T/D pairs were tested overall. The participants comprised a
convenient sample of 139 undergraduate students, who received class credit for their participation. Fifty-
two participants piloted the egocentric item-location task with one set of stimuli and the allocentric item-
location task with another set of stimuli. A further 52 participants piloted the two tasks in the opposite
manner. The testing procedure was the same as is later described for the experiment proper. The 15 T/D
pairs that were initially selected for the egocentric and allocentric item-location tasks had mean corrected
recognition scores of .18 and .19, respectively. Because accuracy was rather low, alterations were made to
the targets and distractors to make them more discriminable (the visual figures were moved closer to the
border). The altered T/D pairs for the two tasks were subsequently tested again with a different sample of
35 participants. The altered T/D pairs for the egocentric and allocentric item-location tasks achieved mean

21
corrected recognition scores of .29 and .30, respectively. The Kuder-Richardson scores for the egocentric
and allocentric item-location pairs were .88 and .92, respectively. Different visual figures were used in the
displays for the two tasks.
Procedure
Overview. The testing phase for each task followed the exposure phase for that task, with a three-
minute interval inserted between exposure and test. This interval was usually filled with unrelated
conversation. The presentation order of the tasks was counterbalanced across participants.
Exposure phase. The exposure phase for both the egocentric and the allocentric item-location tasks
consisted of 15 trials, on each of which the loose- leaf binder was laid on the table, directly in front of the
participant, open to the display sheet for the current trial. The participant saw the sheet for 5 seconds and
was instructed to remember the location of the figure relative to the irregular border. Each trial was
followed by a 3-second inter-trial interval.
Test phase. The test phase for both of the tasks commenced immediately following the three-
minute filled delay. Thirty displays (15 previously-seen targets and 15 distractors), were presented on the
table, in sequence, directly in front of the participant. For the egocentric item-location task, participants
were required to indicate whether or not each figure had appeared previously in the indicated location.
For the allocentric item-location task, participants were told that the page had been rotated from its
orientation at exposure and required to indicate whether or not each figure had appeared in the indicated
location relative to the irregular border. For both tasks, participants indicated their responses by writing
“Y” or “N” on pre-printed answer sheets.
Results
Memory Tests
Figure 3 shows the mean corrected recognition scores for the two tasks as a function of group

22
membership. One-way ANOVA’s failed to reveal significant group differences on the egocentric item-
location task (F(2,40)=1.78, MSe=.024, p = .182). There was a trend towards group differences on the
allocentric item-location task (F(2,40)=3.02, MSe=.041, p=.06). A post-hoc comparison revealed that the
performance of the RTLR group on the egocentric and allocentric item-location tasks was significantly
poorer than the performance of the control group on these tasks (F(1,28) = 8.64, MSe = .032, Partial Eta
Squared = .24), and that the difference between the LTLR and the control participants did not differ on
the two tasks (F(1,28) < 1).
Insert Figure 3 here
Ninety-five percent confidence intervals were examined for the corrected recognition data for each
group, to ascertain the likelihood that the data reflect random performance. Across all three participant
groups, CIs were > 0 for the egocentric item-location task, suggesting that the observed data do not reflect
random performance. The same can be said for the LTLR and control participants on the allocentric item-
location task (CIs ranged from .67 to 5.46). However, the RTLR group’s performance on the allocentric
item-location task suggests that performance may have been random here (CI = -.46 to 2.73).
One-way ANCOVAs were also conducted, with estimated IQ and gender as covariates. No
significant main effects of the covariates were observed for either the egocentric or the allocentric item-
location tasks.
A multivariate repeated-measures ANOVA confirmed that the control group’s performance was
matched across the two experimental tasks, (F(1,14)=3.16).
Task Performance and Size of Resection
Measures of performance on both tasks were correlated with the resection measurements for 14 of
the 15 RTLR participants. None of the correlations between the task variables and the resection variables
was significant at the .05 level. The same correlations were also conducted for 10 of the 13 LTLR

23
participants, revealing no significant relationships. The results of the correlational analyses are presented
in Table 3.
Insert Table 3 here
Discussion
None of the tasks was more difficult than any other for the control participants. The preliminary
task-matching process was evidently successful.
In comparison to the control participants, the RTLR participants performed significantly more
poorly on the egocentric and allocentric item-location tasks. These results reinforce previous reports of a
three-dimensional item-location deficit in RTLR patients (Nunn et al., 1998; Nunn et al., 1999; Smith &
Milner, 1981; Smith & Milner, 1989). Furthermore, as in previous work, the RTLR deficits occurred in
the context of intact LTLR performance.
The RTLR participants were equally deficient on the tests of egocentric and allocentric item-
location memory. These results contrast with previous reports that RTLR participants are relatively more
deficient on tests of allocentric than egocentric item-location memory (Gaffan, 1994).
The results of the present experiment offer an interesting contrast to the results of Experiment 1. In
Experiment 1, both RTLR and LTLR participants showed deficits of two-dimensional item-location
memory. In the present experiment, RTLR but not LTLR participants showed deficits of three-
dimensional item-location memory. In both experiments, the item-location tasks tapped information on
the binding of item and location information. The stimulus items and the procedures for testing item-
location memory were the same in the two experiments. The primary difference between the two
experiments was that the stimulus items were located in a two-dimensional domain in Experiment 1 and a
three-dimensional domain in Experiment 2. The contrast between the results of the two experiments will
be addressed below.

24
General Discussion
The present results contribute to an emerging picture of spatial memory consequent to temporal
lobe dysfunction. These results reinforce earlier reports regarding three-dimensional spatial memory.
More importantly, the results augment the extant picture with new information regarding two-dimensional
spatial memory. In Experiment 1, RTLR but not LRLR patients showed deficits of two-dimensional
location memory. At the same time, both RTLR and LTLR patients showed deficits of two-dimensional
item-location memory. The exploration of two-dimensional spatial memory consequent to temporal lobe
resection is particularly useful in light of the importance of the two-dimensional reference frame in
imaging studies of spatial memory (Johnsrude et al., 1999; Moscovitch et al., 1995; Owen et al, 1996).
In fleshing out the picture of two-dimensional spatial memory consequent to temporal lobe
dysfunction, the present results suggest complexities that have been heretofore unnoted. Of greatest
interest, in this respect, are the results for the two- and three-dimensional item-location tasks. Whereas the
RTLR and LTLR participants both showed deficits in the two-dimensional task, only the RTLR
participants showed deficits in the three-dimensional task. Although the two- and three- dimensional
item-location tasks were not explicitly matched for difficulty, the two tasks were quite similar except for
the difference in dimensionality. Thus, the differences that were observed across tasks may require an
explanation.
In attempting to explain these differences, it is useful to consider the item-location tasks in the
context of the item task. The RTLR and LTLR participants both showed deficits in the latter task. Thus,
across participant groups, performance in the two- but not the three-dimensional item-location task
followed the same pattern as performance in the item task. Why was this the case? It is worth
remembering that the two-dimensional item-location task involved a simpler form of space than the three-
dimensional item-location task. The space of the two-dimensional task paralleled the space of the retina.

25
In contrast, the space of the three-dimensional task had to be inferred from the two-dimensional retinal
image on the basis of depth cues. In light of this difference between the two item-location tasks, one
possible explanation of the observed pattern runs as follows: Item information was dominant in the
memory representation that was created to bind item and two-dimensional location information. At the
core of a given representation was the item in question. The item’s location was recorded as an added
piece of information associated with the item. In contrast, location information was dominant in the
memory representation that was created to bind item and three-dimensional location information. At the
core of a given representation was the location in question. The presence of the item at that location was
recorded as an added piece of information associated with the location. The difference between the two
cases arose because three-dimensional location is more complex than two-dimensional location. An
item’s three-dimensional location was too complex to be recorded as an added piece of information.
This proposed explanation can accommodate the present results as follows: Consider, first, the fact
that RTLR and LTLR participants both showed deficits of two-dimensional item-location memory. These
deficits derived from deficits of item memory; the binding of item and two-dimensional location
information depended on the representation of item information. Consider, second, the fact that RTLR but
not LTLR participants showed deficits of three-dimensional item-location memory. We assume that
RTLR but not LTLR participants had deficits of three-dimensional location memory. Although this was
not shown in the present study, it has been shown in previous work (Rybash & Hoyer, 1992). The
assumption derives further support from the fact that RTLR but not LTLR participants showed deficits of
two-dimensional location memory in the present study. Given this assumption, the deficits that RTLR but
not LTLR participants showed in three-dimensional item-location memory derived from deficits of three-
dimensional location memory; the binding of item and three-dimensional location information depended
on the representation of location information.

26
The proposed explanation is consistent with results from imaging studies. Imaging results
comport, in particular, with the idea that the binding of item and two-dimensional location information
depends on the representation of item information. Imaging studies have shown the entorhinal cortex to be
activated during item memory tasks (Fried, Cameron, Yashar, Fong, & Morrow, 2002). (Results from
non-human primates provide a similar picture; if non-matching and matching- to-sample are taken as
analogs of recognition, deficits in item recognition have been associated with ablation of the entorhinal
cortex (Murray, 2000). Imaging studies have also shown the entorhinal cortex to be activated during two-
dimensional item-location memory tasks (Owen et al., 1996). The present results are consistent with the
implication that the same brain area is involved in memory for item and two-dimensional item-location
memory. In addition, the present results are consistent with the view that the entorhinal cortex is the
particular brain area involved, inasmuch as this area was included in the resected portion of the temporal
lobe for both the RTLR and LTLR groups.
Of course, the proposed explanation remains purely speculative at this point and will need to be
borne out by further results. Regardless of the ultimate fate of this particular explanation, the results that
are demonstrated here for two-dimensional item-location memory will require a future accounting.
The present study also produced results that are inconsistent in several respects with previous
work in nonverbal memory. The results bear, first, on the question of the relative deficiency of visual and
spatial memory consequent to right medial temporal dysfunction. Conflicting positions have been taken
with respect to this question, with Barr (1997) and Nunn et al. (1998) arguing that right medial temporal
dysfunction produces larger deficits of visual and spatial memory, respectively. The present results
support neither of these previous positions; in Experiment 1, the RLTR participants were equally deficient
relative to the control participants on the tests of visual memory (the item task) and spatial memory (the
location and item-location tasks). The present results also bear on the question of the relative deficiency

27
of egocentric and allocentric memory consequent to right medial temporal dysfunction. In previous work,
it has been argued that allocentric memory is more deficient than egocentric memory (Goldstein et al.,
1989). The present results do not support this view; in Experiment 2, the RLTR participants were equally
deficient relative to the control participants on the tests of egocentric and allocentric item-location
memory. In considering the differences between the present and previous results, it is important remember
the measures that were taken here to match the tasks of the two experiments for difficulty. In light of these
measures, the present results may deserve more credence than previous findings.
The present results are also of some interest with respect to visual memory. In contrast to many
previous studies, in which visual memory deficits have been reported only for RTLR patients, both RTLR
and LTLR patients showed deficits of visual memory, as indexed by the item memory task. Further work
will be necessary to sort out the factors that determine visual memory deficits in temporal lobe patients.
Finally, the present results have implications for the pre- and post-surgical assessment of patients
undergoing temporal lobe resection. These results suggest that the pattern of visual-spatial deficits
following temporal lobe resection is complex. Ideally, pre- and post-assessments should tap item,
location, and item-location memory. Location and item-location memory should ideally be assessed in
two as well as three dimensions. In this context, it should be noted that an informal survey of memory
problems among the patient participants revealed fewer complaints in the RTLR than the LTLR group.
Several possible limitations of the study should be noted. First, the sample may have been too
small. Assuming the effect size of Nunn et al. (1998), a sample of 15 RTLR and 15 LTLR patients, and a
critical value of .05, pre-experimental calculations placed the power of the experimental tasks at .8. This
estimate may have been overly optimistic, however. A larger sample would have increased the power of
the tasks to detect differences among the groups. Second, the level of performance on the tests of the

28
study may have been low enough among the control participants as to cause floor effects among the
patient participants.
In conclusion, the present study demonstrated deficits for RTLR and LTLR patients on tests of
item memory and two-dimensional item-location memory. The study demonstrated deficits for RTLR but
not LTLR patients on tests of two-dimensional location memory and three-dimensional item-location
memory.

29
References
Abrahams, S., Pickering, A., Polkey, C., & Morris, R. (1997). Spatial memory deficits in patients
with unilateral damage to the right hippocampal formation. Neuropsychologia, 35, 11-24.
Barr, W. (1997). Examining the right temporal lobe’s role in nonverbal memory. Brain and
Cognition, 35, 26-41.
Blakemore, C. B., & Falconer, M. A. (1967). Long-term effects of anterior temporal lobectomy on
certain cognitive functions. Journal of Neurology, Neurosurgery, and Psychiatry, 30, 364-367.
Blaxton, T., & Theodore, W. (1997). The role of the temporal lobes in recognizing visuospatial
material: Remembering versus knowing. Brain and Cognition, 35, 5-25.
Boone, D. E. (1992). Evaluation of Kaufman’s short forms of the WAIS-R with psychiatric
inpatients. Journal of Clinical Psychology, 48, 239-245.
Chalfonte, B., & Johnson, M. K. (1996). Feature memory and binding in young and older adults.
Memory & Cognition, 24, 403-416.
Chapman, L. J., & Chapman, J. P. (1978). The measurement of differential deficit. Journal of
Psychiatric Research, 14, 303-311.
Devinsky, O. (1995). Cognitive and behavioral effects of antiepileptic drugs. Epilepsia, 36, 46-65.
Dodrill, C. B., & Matthews, C. G. (1992). The role of neuropsychology in the assessment and
treatment of persons with epilepsy. American Psychologist, 47, 1139-1142.
Feigenbaum, J. D., & Morris, R. G. (2004). Allocentric versus egocentric spatial memory after
unilateral temporal lobectomy in humans. Neuropsychology, 18, 462-472.
Fried, I., Cameron, K. A., Yashar, S., Fong, R., & Morrow, J. W. (2002). Inhibitory and excitatory
responses of single neurons in the human medial temporal lobe during recognition of faces and objects.
Cerebral Cortex, 12, 575-84.

30
Gaffan, D. (1994). Scene-specific memory for objects: A model of episodic memory impairment
in monkeys with fornix transection. Journal of Cognitive Neuroscience, 6, 305-320.
Goldstein, L. H., Canavan, A. G. M., & Polkey, C. E. (1989). Cognitive mapping after unilateral
temporal lobectomy. Neuropsychologia, 27, 167-177.
Haist, F., Bowden, G. J., & Mao, H. (2001). Consolidation of human memory over decades
revealed by functional magnetic imaging. Nature Neuroscience, 4, 1139-45.
Hellige, J. B., & Michimata, C. (1989). Categorization versus distance: Hemispheric differences
for processing spatial information. Memory and Cognition, 17, 770-776.
Hermann, B., Seidenberg, M., Wyler, A., & Haltiner, A. (1993). Dissociation of object
recognition and spatial localization abilities following temporal lobe lesions in humans.
Neuropsychology, 7, 343-350.
Holdstock, J. S., Mayes, A. R., Cezayirli, E., Aggleton, J. P., & Roberts, N. (1999). A comparison
of egocentric and allocentric spatial memory in medial temporal lobe and Korsakoff amnesics. Cortex, 35,
479-501.
Hornak, J., Oxbury, S., Oxbury, J., Iversen, S. D., & Gaffan, D. (1997). Hemifield-specific visual
recognition memory impairments in patients with unilateral temporal lobe removals. Neuropsychologia,
35, 1311-1315.
Incisa della Rocchetta, A., Samson, S, Ehrle, N., Denos, M., Hasboun, D., & Baulac, M. (2004).
Memory for visuospatial location following selective hippocampal sclerosis. Neuropsychology, 18, 15-28.
Jones-Gotman, M., Zatorre, R. J., Olivier, A., Andermann, F., Cendes, F., Staunton, H.,
McMackin, D., Siegel, A. M., & Wieser, H.-G. (1997). Learning and retention of words and designs
following excision from medial or lateral temporal- lobe structures. Neuropsychologia, 35, 963-973.

31
Johnsrude, I. S., Owen, A. M., Crane, J., Milner, B. & Evans, A. C. (1999). A cognitive activation
study of memory for spatial relationships. Neuropsychologia, 37, 829-841.
Kaufman, A. S., Ishikuma, T., & Kaufman-Packer, J. L. (1991). Amazingly short forms
of the WAIS-R. Journal of Psychoeducational Assessment, 9, 4-15.
Kimura, D. (1963). Right temporal lobe damage. Archives of Neurology, 8, 264-71.
Kosslyn, S. M. (1987). Seeing and imagining in the cerebral hemispheres: A computational
approach. Psychological Review, 94, 148-175.
Kosslyn, S. M. (1994). Image and brain: The resolution of the imagery debate. Cambridge, MA:
MIT Press.
Maguire, E. A., & Cipolotti, L. (1998). Selective sparing of topographic memory. Journal of
Neurology, Neurosurgery & Psychiatry, 65, 903-909.
Mendola, J. D., Rizzo, J. F., Cosgrove, G. R., Cole, A. J., Black, P, & Corkin, S. (1999). Visual
discrimination after anterior temporal lobectomy in humans. Neurology, 52, 1028-1037.
Milner, B. (1968). Visual recognition and recall after right temporal- lobe excision.
Neuropsychologia, 6, 191-209.
Milner, B., Corkin, S., & Teuber, H. L. (1968). Further analysis of the hippocampal amnesic
syndrome: 14-year followup study of H.M. Neuropsychologia, 6, 215-234.
Moscovitch, M., Kapur, S., Kohler, S., & Houle, S. (1995). Distinct neural correlates of visual
long-term memory for spatial location and object identity: A PET study in humans. Proceedings of the
National Academy of Science, 92, 3721-3725.
Murray, E. A. (2000). Memory for objects in nonhuman primates. In M. S. Gazzaniga (Ed.), The
New Cognitive Neurosciences, pp. 753-763. Cambridge, MA: MIT Press.

32
Murray, E. A., & Mishkin, M. (1986). Visual recognition in monkeys following rhinal cortical
ablations combined with either amydalectomy or hippocampectomy. The Journal of Neuroscience, 6,
1991-2003.
Musen, G., & Treisman, A. (1990). Implicit and explicit memory for visual patterns. Journal of
Experimental Psychology: Learning, Memory, and Cognition, 16, 127-137.
Nunn, J. A., Graydon, F. J. X., Polkey, C. E., & Morris, R. (1999). Differential spatial memory
impairment after right temporal lobectomy demonstrated using temporal titration. Brain, 122, 47-59.
Nunn, J., Polkey, C., & Morris, R. (1998). Selective spatial memory impairment after right
unilateral temporal lobectomy. Neuropsychologia, 36, 837-848.
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford: Clarendon Press.
Oldfield, R. C. (1971). The assessment and analysis of handedness: The Edinburgh Inventory.
Neuropsychologia, 9, 97-113.
O’Reilly, R. C., & Rudy, J. W. (2001). Conjunctive representations in learning and memory:
Principles of cortical and hippocampal function. Psychological Review, 108, 311-345.
Owen, A. M., Milner, B., Petrides, M., & Evans, A. C. (1996). A specific role for the right
parahippocampal gyrus in the retrieval of object-location: A positron emission
tomography study. Journal of Cognitive Neuroscience, 8, 588-602.
Pigott, S., & Milner, B. (1993). Memory for different aspects of complex visual scenes after
unilateral temporal or frontal- lobe resection. Neuropsychologia, 31, 1-15.
Pillon, B., Bazin, B., Deweer, B., Ehrle, N., Baulac, M., & Dubois, B. (1999). Specificity of
memory deficits after right or left temporal lobectomy. Cortex, 35, 561-571.
Rains, G. D., & Milner, B. (1994). Right-hippocampal contralateral-hand effect in the recall of
spatial location in the tactual modality. Neuropsychologia, 32, 1233-1242.

33
Ridley, R.M., Timothy, C.J., Maclean, C. J., & Baker, H. F. (1995). Conditional learning and
memory impairments following neurotoxic lesion of the CA1 field of the hippocampus. Neuroscience, 67,
263-275.
Rybash, J. M., & Hoyer, W. J. (1992). Hemispheric specialization for categorical and coordinate
Spatial representations: A reappraisal. Memory and Cognition, 20, 271-276.
Sagar, H. H., Cohen, N. J.,Corkin, S. & Growdon, J. M. (1985). Dissociations among processes in
remote memory. In D. S. Olton, E. Gamzu, & S. Corkin (Eds.), Memory Dysfunctions, pp. 533-535. New
York: NY Academy of Science.
Scoville, W., & Milner, B. (1957). Loss of recent memory after bilateral hippocampal lesions.
Journal of Neurology, Neurosurgery, and Psychiatry, 20, 11-21.
Shimamura, A. P. (2002). Relational binding theory and the role of consolidation in memory
retrieval. In L. R. Squire, & D. L. Schacter (Eds.), Neuropsychology of memory (3rd ed.), pp. 61-72.
New York: Guilford Press.
Smith, M., & Milner, B. (1981). The role of the right hippocampus in the recall of spatial
location. Neuropsychologia, 19, 781-793.
Smith, M. L., & Milner, B. (1989). Right hippocampal impairment in the recall of spatial location:
Encoding deficit or rapid forgetting. Neuropsychologia, 27, 71-81.
Squire, L. (1992). Memory and the hippocampus: A synthesis from findings with rats, monkeys,
and humans. Psychological Review, 99, 195-231.
Stepankova, K., Fenton, A, Pastalkova, E., Kalina, M., & Bohbot, V. (2004). Object- location
memory impairment in patients with thermal lesions to the right or left hippocampus. Neuropsychologia,
42, 1017-1028.

34
Suzuki, W. A., Zola-Morgan, S., Squire, L. R., & Amaral, D. G. (1993). Lesions of the perirhinal
and parahippocampal cortices in the monkey produce long- lasting memory impairment in the visual and
tactual modalities. The Journal of Neuroscience, 13, 2430-2451.
Wechsler, D. (1981). WAIS-R manual: Wechsler Adult Intelligence Scale-Revised. San Antonio,
TX: Psychological Corporation.
Wiebe, S., Blume, W. T., Girvin, J. P., & Eliasziw, M. (2001). A randomized, controlled trial of
surgery for temporal- lobe epilepsy. New England Journal of Medicine, 345, 311-318.
Zola, S. M., Squire, L. R., Teng, E., Stefanacci, L., & Clark, R. E. (2000). Impaired recognition
memory in monkeys after damage to limited to the hippocampal region. The Journal of Neuroscience, 20,
451-463.
Zola-Morgan, S. M., & Squire, L. R. (1985). Medial temporal lesions in monkeys impair memory
on a variety of tasks sensitive to human amnesia. Behavioral Neuroscience, 99, 22-34.

35
Figure Captions
Figure 1. Example of the type of stimuli used in the Item-Location task of Experiment 1 and the
Egocentric and Allocentric Item-Location tasks of Experiment 2.
Figure 2. Mean corrected recognition score as a function of participant group on the Item, Location, and
Item-Location tasks of Experiment 1.
Figure 3. Mean corrected recognition score as a function of participant group on the Egocentric and
Allocentric Item-Location tasks of Experiment 2.

36
Table 1
Means and standard deviations of participant characteristics
RTLR LTLR Control
N 15 13 15
Male / female 7/8 3/10 7/8
Edinburgh Handedness Inventory [36] 77.90 (24.70) 74.12 (24.03) 62.81 (44.10)
Age 41.53 (11.89) 43.08 (11.28) 37.60 (8.55)
Years of Education 14.67 (2.50) 14.77 (3.09) 14.40 (1.53)
Estimated IQ 94.89 (13.98) 99.28 (16.91) 98.69 (7.23)
Age at seizure onset 16.83 (9.76) 17.63 (15.02) -
Duration of epilepsy 21.40 (9.59) 20.54 (11.57) -
No. anti-epileptic drugs taken 1.00 (0.78) 1.08 (0.64) -
Current seizure frequency (Engel∇) 1.20 (0.41) 1.46 (0.52) -
Age and time variables reported in years
∇ Engel level 1 = seizure free; 2 = rare seizures; 3 = worthwhile improvement; 4 = no worthwhile
improvement.

37
Table 2
Experiment 1: Pearson Correlations between Amounts of Tissue Resected and Task Variables
Identity Location Identity-Location
RTLR Group
Inferior gyrus -.06 -.02 .05
Middle gyrus .21 .02 -.09
Superior gyrus -.16 -.23 -.19
LTLR Group
Inferior gyrus .21 -.16 -.29
Middle gyrus -.02 -.29 -.46
Superior gyrus -.53 .08 -.38

38
Table 3
Experiment 2: Pearson Correlations between Amounts of Tissue Resected and Task Variables
Egocentric Allocentric
RTLR Group
Inferior gyrus .31 -.46
Middle gyrus .38 -.47
Superior gyrus .35 -.28
LTLR Group
Inferior gyrus .12 .17
Middle gyrus -.02 .25
Superior gyrus -.42 -.38

39
40

41




















