
Cotton Insect Management With Special Reference to the Boll ...
610
/ ~N d States rtment of ulture ' ultural neaearch Service Agriculture Handbool< Number 589 Cotton Insect Management With Special Reference to the Boll W^täl
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Cotton Insect Management With Special Reference to the Boll ...

/
~N
d States rtment of ulture
' ultural neaearch Service
Agriculture Handbool< Number 589
Cotton Insect Management With Special Reference to the Boll W^täl


oSpartÄ^ Cotton InsGCt Management With Special Reference to the Boll Weevil
Agricultural Research Service
Agriculture Handbook Number 589
Issued November 1983
Edited by R.L Ridgway, E.P. Lloyd, and W. H. Cross
For sale l)y the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402

ABSTRACT
Ridgway, R. L., E. P. Lloyd, and W. H. Cross, editors. 1983. Cotton insect management with special reference to the boll weevil. U.S. Department of Agriculture, Agriculture Handbook No. 589. 612 p., illus.
This monograph describes the state of the art of management of boll weevils in the United States. It is, therefore, concerned with the biological, economic, and environmental effects of this introduced pest, the problems associated with its control, and alternative approaches to dealing with this pest.
Keywords: cotton losses, cotton culture, Anthonomus grandis, cotton insects, Heliothis spp., simulation models. Boll Weevil Eradication Trial, Optimum Pest Management Trial, insect manage- ment, eradication, chemical insecticides, insect growth regula- tors, pheromones, sterile insect releases, beneficial arthropods, microbial agents
Mention of a proprietary product in this publication does not constitute a guarantee or warranty of the product by the U.S. Department of Agriculture and does not imply its approval by the Department to the exclusion of other products that may also be suitable. Views expressed by the authors are not necessarily those of the U.S. Department of Agriculture.

FOREWORD
Profitable cotton production in the U.S. and effective insect control and management have been particularly closely related since the boll weevil, Anthonomus grandis Boheman, entered the U.S. in 1892. Since that time, much research has been directed toward the boll weevil and other cotton insects. Recently, scientists have conducted expanded field and laboratory investigations to gain in- creased insight and understanding of the biological, economic, and environmental impacts of alternative options for beltwide cotton insect management. Alternative options or strategies are contro- versial. To a degree they are controversial because of deep-seated philosophy and/or convictions on the part of individuals and the economic interest groups. In broad terms, one of the most impor- tant things that we have learned over the years is that no cotton producer is an island to himself or herself in terms of effective insect management programs. Certainly, all segments of the cotton econoiay are entitled to have access to the knowledge available on the subject and to the privilege of making their own decisions.
A pilot field experiment was conducted in 1971-73 in south Mississippi and Louisiana to determine whether eliminating the boll weevil by properly integrating the use of various techniques was technically and operationally feasible. Although the Technical Guidance Committee for that experiment concluded that it was tech- nically and operationally feasible to eliminate the boll weevil as an economic pest in the United States, a significant number of en- tomologists opposed the conclusion. The interpretation, therefore, of the results of the pilot field experiment led to divergent views that may be rectified only through further research and joint dis- cussions. It is of interest to note that current authorization in the Agricultural and Consumer Protection Act of 1973 states that "...Secretary is authorized and directed to carry out programs to destroy and eliminate cotton boll weevils in infested areas of the United States...if the Secretary determines that method and sys- tems have been developed to the point that success in eradication of such insects is assured." This authorization stimulated the development of the present boll weevil program with the overall objective of developing sound scientific information which may be utilized in making a recommendation for a beltwide cotton insect management program for the boll weevil infested areas of the Cotton Belt.
During the development of a format for the Biological Evalua- tion Team Report, it became obvious that a number of the topics could be developed into chapters for the publication of a mono- graph. Enthusiasm for the proposed treatise gained momentum as the potential for such an undertaking was discussed with colleagues, authors, and administrators. There was general agreement that the monograph could add stature and visibility to the comprehensive study while emphasizing areas of interest to a broad base of read- ers. It was also envisaged that the monograph would be an integral part of the Biological Evaluation Team Report as well as a separate

IV
publication. This sequence of collective thoughts and delibera- tions brought forth "Cotton Insect Management with Special Refer- ence to the Boll Weevil."
The purpose of this monograph is to present the results of research studies related to the biological, economic, and environ- mental effects of the boll weevil and other cotton insects on the crop with special emphasis on the biological aspects. In view of the emphasis placed on the Optimum Pest Management and Boll Weevil Eradication Trials, it seems appropriate to pay special attention to the biological phases and to provide a summary of the pertinent data together with detailed references.
This monograph is divided into five parts. The introductory chapters provide a historical background, and describe current practices and impacts, as well as concepts for future considera- tion. The second part provides an indepth discussion of newly developed and existing technology available for practical use now or in the future. In the third part, the supporting components of insect rearing, sampling, and modeling are reviewed. Alternative programs and then impacts are discussed in section four. Finally, in section five, opportunities and constraints of current and future programs are presented.
The authors have paid attention to the more recent and perti- nent literature. As in any scientific endeavor, some may chal- lenge the data presented in the monograph. The material is based, however, on such extensive experimental evidence that is not like- ly to be found incorrect.
Some of the authors project certain management strategies for the boll weevil and other cotton insects. However, it is expected that the results of the research findings present here will not only promote cotton insect strategies but also have application to numerous other crop commodities.
Even though scientists, specialists, consultants, and others representing various segments of the cotton economy may hold di- verse views regarding insect management programs for cotton, they should not differ on the desirability of learning the truth. This monograph, authoritatively written by a number of authors and edited by three prominent scientists, presents the facts, assem- bles present knowledge, and indicates areas along which further knowledge must be obtained.
K. R. KELLER Executive Coordinator Boll Weevil Policy Group

ACKNOWLEDGMENTS
The editors of this monograph are very grateful for the assis- tance of many who contributed to its preparation and publication. Thanks are due to the USDA Boll Weevil Policy Group, Anson R. Bertrand, Harry C. Mussman, and Kenneth R. Farrell, for their sup- port and encouragement as this endeavor was conceived and prepara- tions were begun. Thanks are also due to the authors who prepared their material quickly and cooperated fully with an accelerated review process; to the many anonymous reviewers who provided their comments on a timely basis; to Mrs. Jane Wall, Mrs. Fay Eggers, and Ms. Judith Smith who provided substantial editorial assistance; to Mrs. Marguerite Benedict who prepared the index; and Mr. Edgar Skelton, Mr. Roy Nash, and Mrs. Winnie Daves who supervised the preparation of illustrations and of final copy. In addition, thanks are due to Dr. Kenneth R. Keller, executive coordinator for the Boll Weevil Policy Group, for his assistance in securing anony- mous peer reviews and providing strong encouragement throughout its preparation. Special thanks are due Mrs. Sara Smith, Mrs. Dorothy Turner, and Mrs. Marijane Burns for their tireless and persistent efforts as they worked for more than a year in the preparation and revision of this monograph on word processing equipment at Raleigh, NC.


vil
CONTENTS
Foreword iii
Acknowledgments v
Contributors ix
I. Introduction
1. Evolution of Cotton Insect Management in the United States 3
R. L. Ridgway and E. P. Lloyd
2. The Cost of Insecticides Used on Cotton in the United States 29
F. T. Cooke, Jr. and D. W. Parvin, Jr.
3. Ecology of Cotton Insects with Special Reference to the Boll Weevil 53
W. H. Cross
II. Suppression Components
4. Plant Resistance and Modified Cotton Culture 73 L. N. Namken, M. D. Heilman, J. N. Jenkins, and P. A. Miller
5. Entomophagous Arthropods 103 J. R. Abies, J. L. Goodenough, A. W. Hartstack, and R. L. Ridgway
6. Microbial Agents 129 M. R. Bell
7. Boll Weevil Sterility 153 J. E. Wright and E. J. Villavaso
8. Pheromones for Survey, Detection, and Control . . . .179 E. P. Lloyd, G. H. McKibben, J. E. Leggett, and A. W. Hartstack
9. Insect Growth Regulators with Emphasis on the Use of Benzoylphenyl Ureas 207
D. L. Bull, J. R. Abies, and E. P. Lloyd
10. Insecticides for Control of Cotton Insects 237 C. R. Parencia, Jr., T. R. Pfrimmer, and A. R. Hopkins

VIII
III. Support Components
11. Mass Rearing Boll Weevils 265 J. G. Griffin, P. P. Sikorowski, and 0. H. Lindig
12. Sampling Arthropods in Cotton 303 J. W. Smith, W. A. Dickerson, and W. P. Scott
13. Losses in Yield of Cotton Due to Insects 329 P. H. Schwartz
14. Models for Cotton Insect Pest Management 359 A. W. Hartstack and J. A. Witz
IV. Alternative Programs
15. Optimum Pest Management Trial in Mississippi .... 385 J. L. Hamer, G. L. Andrews, R. W. Seward, D. F. Young, Jr., and R. B. Head
16. Analysis of Technology Available for Eradication of the Boll Weevil 409
E. F. Knipling
17. Cotton and Insect Management Simulation Model .... 437 L. G. Brown, R. W. McClendon, and J. W. Jones
18. Impact of Alternative Cotton Insect Management Strategies on Producer Income in Mississippi. . . . 481
E. H. Simpson, III and D. W. Parvin, Jr.
19. Economic Evaluation of the Boll Weevil Eradica- tion Trial in North Carolina, 1978-80 497
G. A. Carlson and L. F. Suguiyama
V. Future Prospects
20. Opportunities for Improving Cotton Insect Manage- ment Programs and Some Constraints on Beltwide Implementation 521
R. E. Frisbie, J. R. Phillips, W. R. A. Lambert, and H. B. Jackson
Index 559

IX
CONTRIBUTORS
J. R. ABLES Cotton Insects Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77841
G. L. ANDREWS Mississippi Cooperative Extension Service Batesville, MS 38606
M. R. BELL Western Cotton Insects Laboratory Agricultural Research Service U.S. Department of Agriculture Phoenix, AZ 85040
L. G. BROWN Department of Industrial Engineering Mississippi State University Mississippi State, MS 39762
D. L. BULL Cotton Insects Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77841
G. A. CARLSON Department of Economics and Business North Carolina State University Raleigh, NC 27650
F. T. COOKE, JR. Economic Research Service U.S. Department of Agriculture Stoneville, MS 38776
W. H. CROSS Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
W. A. DICKERSON Boll Weevil Eradication Research Agricultural Research Service U.S. Department of Agriculture Raleigh, NC 27607

R. E. FRISBIE Integrated Pest Management Coordinator Texas A&M University College Station, TX 77843
J. L. GOODENOUGH Pest Control Equipment and Methods Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77843
J. G. GRIFFIN Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
J. L. HAMER Mississippi Cooperative Extension Service Mississippi State, MS 39762
A. W. HARTSTACK Pest Control Equipment and Methods Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77843
R. B. HEAD Mississippi Cooperative Extension Service Mississippi State, MS 39762
M. D. HEILMAN Soil and Water Conservation Research Laboratory
Agricultural Research Service U.S. Department of Agriculture Weslaco, TX 78596
A. R. HOPKINS Cotton Production Research Agricultural Research Service U.S. Department of Agriculture Florence, SC 29502
H. B. JACKSON Plant Pest Regulatory Services Clemson University Clemson, SC 29631

J. N. JENKINS Crop Science and Engineering
Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
J. W. JONES Department of Agricultural Engineering University of Florida Gainesville, FL 32611
K. R. KELLER Executive Coordinator, Boll Weevil
Policy Group U.S. Department of Agriculture Raleigh, NC 27650
E. F. KNIPLING Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705
W. R. A. LAMBERT Entomology Department University of Georgia Tifton, GA 31794
J. E. LEGGETT Cotton Production Research Laboratory Agricultural Research Service U.S. Department of Agriculture Florence, SC 29503
0. H. LINDIG Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
E. P. LLOYD Boll Weevil Eradication Research Agricultural Research Service U.S. Department of Agriculture Raleigh, NC 27607
R. W. McCLENDON Department of Agricultural and
Biological Engineering Mississippi State University Mississippi State, MS 39762

XII
G. H. McKIBBEN Boll Weevil Eradication Research Agricultural Research Service U.S. Department of Agriculture Raleigh, NC 27607
P. A. MILLER National Program Staff Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705
L. N. NAMKEN Soil and Water Conservation Research Laboratory
Agricultural Research Service U.S. Department of Agriculture Weslaco, TX 78596
C. R. PARENCIA, JR. Bioenvironmental Insect Control Laboratory Agricultural Research Service U.S. Department of Agriculture Stoneville, MS 38776
D. W. PARVIN, JR. Department of Agricultural Economics Mississippi State University Mississippi State, MS 39762
T. R. PFRIMMER Bioenvironmental Insect Control Laboratory Agricultural Research Service U.S. Department of Agriculture Stoneville, MS 38776
J. R. PHILLIPS Entomology Department University of Arkansas Fayetteville, AR 72701
R. L. RIDGWAY National Program Staff Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705
P. H. SCHWARTZ National Program Staff Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705

XIII
W. P. SCOTT Bioenvironmental Insect Control Laboratory Agricultural Research Service U.S. Department of Agriculture Stoneville, MS 38776
R. W. SEWARD Mississippi Cooperative Extension Service Batesville, MS 38606
P. P. SIKOROWSKI Department of Entomology Mississippi State University Mississippi State, MS 39762
E. H. SIMPSON, III Department of Agricultural Economics Mississippi State University Mississippi State, MS 39762
J. W. SMITH Bioenvironmental Insect Control Laboratory Agricultural Research Service U.S. Department of Agriculture Stoneville, MS 38776
L. F. SUGUIYÀMA Department of Economics and Business North Carolina State University Raleigh, NC 27650
E. J. VILLAVASO Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
J. A. WITZ Pest Control Equipment and Methods Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77843
J. E. WRIGHT Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
D. F. YOUNG, JR. Mississippi Cooperative Extension Service Mississippi State, MS 39762


SECTION I INTRODUCTION


Chapter 1
EVOLUTION OF COTTON INSECT MANAGEMENT IN THE UNITED STATES
R. L. Ridgway National Program Staff U.S. Department of Agriculture Agricultural Research Service Beltsville, MD 20705
E. P. Lloyd Boll Weevil Eradication Research Agricultural Research Service U.S. Department of Agriculture Raleigh, NC 27607
ABSTRACT The invasion of the United States by such exotic insects as the boll weevil (Anthonomus grandis Boheman) and the pink bollworm (Pectinophora gossypiella (Saunders)) substantially increased losses in cotton production due to insect pests. The use of inorganic and synthetic organic insecti- cides provided temporary solutions by controlling both exotic and native insect pests. However, undesirable side effects, such as destruction of natural enemies of insect pests, development of insecticide resistance, and adverse effects on the environment and human health, led to expanded efforts to develop improved methods of control. As a result, entomologists developed two divergent paradigms or strategies for designing technologies for insect con- trol. In one paradigm, emphasis was placed on con- trol actions to be used on a field-by-field basis depending on the time when a certain economic thresh- old was reached. In the other paradigm, emphasis was placed on suppression of a total insect population, with considerable attention to eradication. However, much of the research conducted to support the two paradigms has resulted in the development of tech- nologies that have application to both approaches.
Two cotton insect management trials conducted during 1978-1980, the Optimum Pest (insect) Manage- ment Trial in Mississippi and the Boll Weevil Eradi- cation Trial in North Carolina, together with accel- erated efforts in other States to improve cotton insect control, have now provided a basis for a pos- sible merger of the divergent paradigms. Such a merger could result in the design and implementation of substantially improved cotton insect management programs.

R. L RIDGWAY AND E. P. LLOYD
INTRODUCTION
Significant advances in control of the principal insects at- tacking cotton including the boll weevil (Anthonomus grandis Bohe- man), bollworm (Heliothis zea (Boddie)), tobacco budworm (H. vires- cens (F.)), and pink bollworm (Pectinophora gossypiella (Sounders)) have been made in recent years (Newsom and Brazzel 1968, Adkisson 1972, Ridgway 1972a, Newsom 1974, Reynolds et al. 1975, Bottrell and Adkisson 1977, Parencia 1978, Graham 1980, and Phillips et al. 1980). However, a potential for substantial additional advances, particularly in implementation, is now indicated by the results of the large area trials conducted during 1978-80 in Mississippi by the Mississippi Cooperative Extension Service (Andrews 1981); in North Carolina by the Animal and Plant Health Inspection Service of the U.S. Department of Agriculture (USDA) (Ganyard et al. 1981); and by the results of other insect management programs (Phillips et al. 1980). The Mississippi and North Carolina trials and the sev- eral alternative boll weevil/cotton insect management programs that were defined for evaluation purposes by the USDA-State Evaluation Teams were designed to provide the basis for selecting the pre- ferred cotton insect management program or programs for use throughout the boll weevil infested areas of the Cotton Belt (Eco- nomics and Statistics Service 1981).
The evolution of cotton insect control in the United States is examined in an attempt to provide a useful perspective from which to evaluate new information now available concerning the selection of improved cotton insect management programs.
Newsom (1974) divided the history of cotton insect management into four periods: (1) the period before the boll weevil entered the United States (before 1892); (2) the initial years of boll wee- vil infestation (1892-1917); (3) the period when calcium arsenate was used to control the boll weevil (1917-1945); and (4) the period when treatment with synthetic organic insecticide was the principal method used to control cotton insects (after 1945). Later, Perkins (1980a) divided into three parts the period when synthetic organic insecticides were used: (1) euphoria and the crisis of residues (1945-1955); (2) confusion and the crisis of the environment or be- ginning of new directions (1954-1972), and (3) changing paradigms (after 1968). With some changes in terminology, the periods iden- tified by Newsom (1974) and by Perkins (1980a) plus an additional period, the future, are used here as the framework for a discus- sion of the evolution of cotton insect management.
BEFORE THE BOLL WEEVIL
The first attempts to grow cotton in the United States were made about 1600, but significant commercial production began in the late 1700's (Handy 1896). At an early date, a number of arti- cles about insect pests of cotton, particularly the cotton leaf- worm (Alabama argillacea (Hubner)), appeared in southern newspapers

EVOLUTION OF COTTON INSECT CONTROL
^nd magazines, but the literature increased rapidly after a partic- ularly destructive infestation in 1847. The cotton leafworm then continued to be the primary insect pest for a number of years. However, after Paris green was introduced in 1872 for control of the leafworm and was used widely, losses diminished. Later, as cotton production expanded in Texas, the bollworm became recognized as the chief insect pest of cotton (Comstock 1879), and a wide range of possible control measures was suggested such as light traps, poisoned baits, sprays or dusts containing arsenicals, and trap crops. All were of limited effectiveness; however, in seasons of great bollworm abundance, use of arsenicals would "result in saving at least a part of the crop'* (Howard 1896). Therefore, sig- nificant losses to the cotton crop due to insect pests occurred in the United States prior to the occurrence of the boll weevil.
INITIAL YEARS OF BOLL WEEVIL INFESTATION
The first report of the occurrence of the boll weevil in the United States was received in the U.S. Department of Agriculture in the fall of 1894. However, a careful survey of the area indicated that the boll weevil had been causing serious damage there since 1892 (Townsend 1895). As time passed and the boll weevil spread north and eastward, it caused serious losses. In 1907, it crossed the Mississippi River, and by 1917 it had infested cotton in most of Georgia. By 1922, the insect had infested cotton in the remain- ing southeastern States (Hunter and Goad 1923).
Initial attempts to control the boll weevil with insecticides such as Paris green were unsuccessful, and emphasis was placed on cultural control methods such as destruction of volunteer plants, manual collection of fallen infested squares, early stalk destruc- tion, and trap crops (Howard 1896). However, during the early 1900*s some very important research was conducted on the boll wee- vil and other cotton insects and a fairly effective insect manage- ment system was devised (Hunter and Hinds 1905, Hunter and Pierce 1912, and Pierce 1922).
The insect management system articulated by Pierce (1922) in- cluded the recommendations for the application of arsenate of lead for control of *'worms,'* presumably the cotton leafworm and boll- worm, and numerous cultural methods for control of the boll weevil. Also, the potential of calcium arsenate was recognized even though it was not recommended.
GALGIUM ARSENATE PERIOD
Goad (1918) reported on field experiments in which he found that calcium arsenate gave more effective control of the boll wee- vil than the other arsenical compounds he tested. Goad and Gassidy

R. L RI DG WAY AND E. P. LLOYD
(1920) recommended the initiation of calcium arsenate treatments when 15-20% of the cotton squares were punctured and continuation of subsequent treatments at 4- or 5-day intervals until the bolls were large enough to be safe from attack. The demonstration of ef- fectiveness of calcium arsenate was a very significant event, but the successful application of calcium arsenate by aircraft in 1922 (Coad et al. 1924) perhaps had greater long-term impact. Several other scientists then confirmed the effectiveness of aerial appli- cation of calcium arsenate for control of the boll weevil (Post 1924, Hinds 1925, and Wilson 1926). Control of boll weevils with calcium arsenate made it possible to produce cotton economically in the south and southeastern United States despite the presence of the boll weevil. However, some undesirable side effects were not- ed. For example, destruction of natural enemies of such insect pests as the bollworm and cotton aphid (Aphis gossypii Glover) in- creased the incidence of these pests (Ewing and Ivy 1943).
Another significant event, though it was not fully recognized at the time, occurred during the same period. Isley and Baerg (1924) reported on the use of field surveys or "scouting" to assist in making decisions on whether action should be taken to control a cotton insect pest. Scouting became a key component of later pest management programs.
INTRODUCTION OF SYNTHETIC ORGANIC INSECTICIDES
Ivy (1944) evaluated DDT for control of the cotton leafworm and boll weevil but found that it did not provide effective control of either pest. Shortly thereafter. Ivy and Ewing (1946) reported excellent control of the boll weevil with benzene hexachloride (BHC) dust in laboratory and cage tests, and subsequent field tests confirmed the effectiveness of the gamma isomer (Ewing et al. 1947, Gaines and Dean 1947). Then Gaines and Wipprecht (1948) reported that a mixture of gamma BHC, DDT, and sulphur was more effective than calcium arsenate in controlling late-season cotton insects in- cluding the boll weevil, bollworm, and spider mites (Tetranychus spp.). Shortly thereafter several other organochlorine insecti- cides were developed, tested, and recommended for boll weevil con- trol (Gaines 1957). Use of synthetic insecticides and other pesti- cides on cotton and other commodities increased annually at a very rapid rate (Shepard 1951) until the quantities of insecticide used on cotton began to stabilize in the 1960*s (Ridgway et al. 1978).
During the decade following the introduction of synthetic or- ganic insecticides, the almost complete reliance upon insecticides for control of insects attacking cotton and other crops had an adverse impact on the amount of research directed to the develop- ment of alternative approaches to control. However, the impor- tance of selective insecticides and of biological control con- tinued to receive the attention of some scientists (Pickett 1949, Ripper et al. 1948, 1949). Also, during this period, Ewing and Parencia (1950) proposed a community-wide program for boll weevil

EVOLUTION OF COTTON INSECT CONTROL
suppression based on early-season use of insecticides to kill emerging overwintered weevils. They recognized clearly the value of preserving naturally occurring beneficial arthropods for control of bollworms rather than relying entirely upon the use of broad spectrum insecticides. The program they advocated gave early- season control of the boll weevil, and treatments were terminated early enough so parasites and predators of the bollworm could re- bound in time to provide natural control of the bollworm until late-season boll weevil infestations developed and had to be treated again.
BEGINNINGS OF NEW DIRECTIONS
While the introduction of the synthetic organic insecticides provided cotton producers with a highly effective method of con- trolling their insect pests, the widespread use of these insecti- cides resulted in the emergence of new problems. For example, the appearance of resistant strains of insects, the disruption of natural control systems that resulted in the elevation of secondary or occasional pests to major pest status, and environmental con- cerns caused many entomologists to begin considering alternative approaches to insect control that did not rely entirely upon the use of insecticides.
A high level of resistance to chlorinated hydrocarbon insecti- cides was detected in boll weevil populations in several areas of Louisiana in 1954 (Roussel and Glower 1955). Occurrence of similar resistance in Texas was reported the following year, however, sev- eral organophosphorus insecticides and calcium arsenate were re- ported effective against these populations (Walker et al. 1956). The threat created by the development of insecticide resistance and concerns about the damage caused by the boll weevil resulted in the U.S. Congress directing USDA to review the boll weevil problem and to identify new research approaches. A working group appointed to study the matter concluded that the future of conventional chemi- cals was seriously threatened and recommended the establishment of a new interdisciplinary laboratory to initiate a broad-based re- search program designed to find ways of reducing losses to a mini- mum or of eliminating the problem altogether (USDA 1958). Mean- while, studies underway on integrated biological and chemical con- trol of alfalfa pests were to have significant impact on the future (Stern et al. 1959).
The threat to the cotton industry posed by insecticide resis- tance was intensified when resistance to DDT was reported in the tobacco budworm (Brazzel 1963) and the bollworm (Graves et al. 1963), and resistance to methyl parathion was reported in the to- bacco budworm (Nemec and Adkisson 1969).
Also, the importance of the disruption of natural control systems, particularly through the destruction of natural enemies, was being more widely recognized. Some specific examples were

R. L RI DG WAY AND E. P. LLOYD
documented experimentally when both foliar and systemic insecti- cides were used to control the boll weevil and cotton fleahopper (Pseudatomoscelis seriatus (Reuter)) (Ridgway et al. 1967, Ridgway 1969) and when foliar applications were used to control Lygus spp. (van den Bosch 1971).
While insecticide resistance was developing within cotton in- sects and disruption of natural systems was being documented, con- cerns about the potential adverse environmental and health effects of insecticides were increasing. The publication of "Silent Spring" greatly stimulated expression of that concern (Carson 1962). Also, the adverse environmental effects that occurred when fish kills in the Mississippi River were documented in 1965 had im- pact on the evolution of cotton insect control. The Congressional hearings that followed this incident (U.S. Congress 1965) resulted in an increase in the appropriations that were to be used in im- proving methods of controlling insects.
The opposition to the use of DDT increased greatly during the 1960's, and its use on cotton was cancelled on December 31, 1972 (Dunlap 1981). Therefore, this effective and economical insecti- cide was no longer available for use by growers in the south and southeastern U.S. where it was still useful for the control of the bollworm and tobacco budworm (Anonymous 1975).
The pink bollworm problem was somewhat peripheral to the boll weevil problem, but the basic issues associated with insecticide use were very similar, particularly in the western U.S. Although the pink bollworm first became established in Texas in 1922, it did not become a serious pest until the 1950's (Noble 1969). A compre- hensive research program, together with the almost simultaneous de- velopment and use of harvest-aid chemicals and mechanical harvest- ing, resulted in effective cultural control of this insect in those parts of Texas where it had been a serious pest. However, in the mid-1960's the pink bollworm invaded California and Arizona from Mexico (Graham 1980). Shortly thereafter, regulatory actions began limiting the availability of the inexpensive and most effective in- secticides. However, the climate favored long-season cotton pro- duction and strongly discouraged short-season cultural control of the pink bollworm. Consequently, the spread of the pink bollworm in parts of Arizona and California and the use of large quantities of organophosphate insecticides resulted in that area attaining the distinction of having the highest per acre cost for cotton insect control in the United States during the late I960's (Starbird and French 1972).
Thus, as the I960's came to a close, the combination of insec- ticide resistance, pest resurgence, and regulatory actions had re- sulted in substantially increased costs and greatly increased risks for the cotton producer who continued to rely upon insecticides to prevent substantial losses due to insect pests.

EVOLUTION OF COTTON INSECT CONTROL
PARADIGMS AND EMERGING TECHNOLOGIES
The years after 1968 were marked by the emergence of the two distinct paradigms for insect control research that were to become so important to the evolution of cotton insect management in the United States. These paradigms were discussed in detail by Perkins (1980a) as total population management (TPM) and integrated pest management (IPM). However, they were probably first described by Rabb (1972) who used the terms eradication and population manage- ment (Figure 1). Perkins (1980a) notes that both new paradigms were designed to be an alternative to the chemical control para- digm, which had dominated entomological research and practice after 1950. The two paradigms are partially described by the following quotation:
"Carl B. Huffaker and Ray F. Smith, University of California) are not thinking integrated control in the sense that I am. I'm thinking integrated control in the sense that you're taking advantage of the char- acteristics of different systems and putting them to- gether for total management of a population. They're looking at integrated control . . . (as being) based on assessment of economic threshold levels and not to use control measures until they reach that goal ..." (E. F. Knipling, personal interview, 1976, as reported by Perkins 1980a.)
A somewhat more philosophical description of the contrasts be- tween the paradigms is provided by examining the naturalistic and humanistic presuppositions proposed by Perkins (1980a) for IPM and TPM, respectively, as follows:
"Naturalistic: A belief system that man is a part of the bio- sphere but that he cannot be the total master of it. He may manip- ulate it for his own benefit, but there are intrinsic limits to his manipulative powers that reside in the properties of the material world.
"Humanistic: A belief system that man is part of the bio- sphere and that he can be master of it. He may manipulate it for his benefit, and there are no intrinsic limits to his manipulative powers that reside in the properties of the material world. The limits, such as they are, are derived from his current ignorance of natural processes."
Although recognition of the IPM and TPM paradigms was useful in designing improved communications, there was also a danger in oversimplifying the situation. For instance, IPM as envisioned by C. B. Huffaker and R. F. Smith is considerably broader than econom- ic thresholds and action on a field-by-field basis (Huffaker and
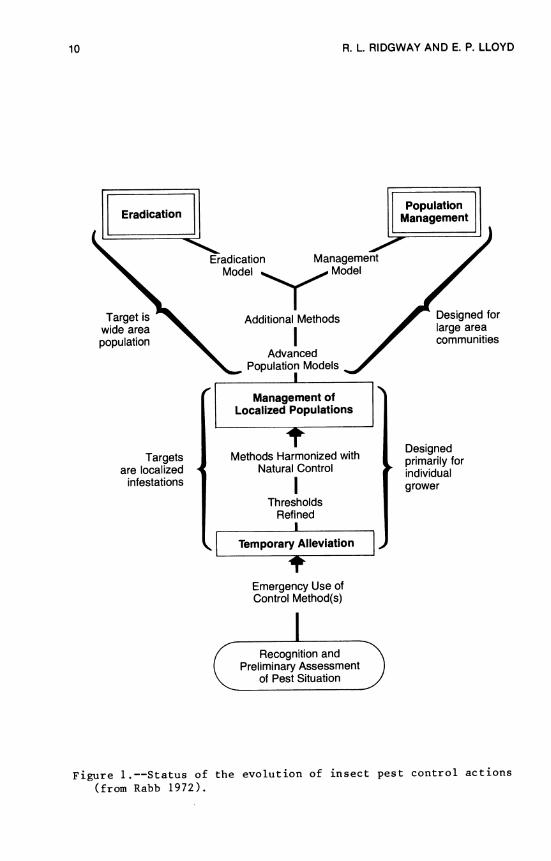
10 R. L RIDGWAY AND E. P. LLOYD
Eradication Model
Management Model
Target is wide area population
Additional Methods
I Advanced
Population Models
Targets are localized
infestations
Management of Localized Populations
Methods Harmonized with Natural Control
Thresholds Refined I
Temporary Alleviation
Designed for large area communities
Designed primarily for individual grower
Emergency Use of Control Method(s)
Recognition and Preliminary Assessment
of Pest Situation
Figure 1.—Status of the evolution of insect pest control actions (from Rabb 1972).

EVOLUTION OF COTTON INSECT CONTROL 11
Smith 1980), and insect management as envisioned by E. F. Knipling includes strategies other than TPM (Knipling 1978).
The TPM and IPM paradigms can readily be identified with re- cent activities designed to improve cotton insect control. For ex- ample, the TPM paradigm was pursued in the Pilot Boll Weevil Eradi- cation Experiment (PBWEE) in Mississippi in 1971-73 (Lloyd 1972), and the IPM paradigm was pursued in the "Huffaker Project" con- ducted from 1972 to 1977 (Huffaker and Smith 1980) and, to some ex- tent, in the cotton insect pest management pilot projects conducted in 14 States beginning in 1972 by Extension personnel with funds provided by USDA (Good and Bonell 1976). Disagreements over the interpretation of the results of the PBWEE increased the polariza- tion of viewpoints associated with the two paradigms (Perkins 1980b).
In 1975, the USDA proposed that funds be included in the fis- cal year 1977 budget to conduct the Boll Weevil Eradication Trial (BWET). When the Office of Management and Budget (OMB) reviewed the proposal, it advised USDA that the BWET should be accompanied by (1) a pest management trial; and (2) consideration of continuing current practices. OMB also advised USDA to conduct a comprehen- sive evaluation of the trials and of the biological, environmental, and economic impacts of alternative boll weevil/cotton insect man- agement programs. Thus, the USDA budget submitted to Congress in January 1976 requested funds to conduct eradication and management trials concurrently. The funds were appropriated; however, the House Agricultural Appropriations Committee instructed USDA not to use these funds in the eradication trial until additional technical advances were made and the affected State governments had author- ized the trials. In the fall of 1977, USDA and the House Committee agreed that the trials should proceed. In 1978, the two large areawide trials—BWET and Optimum Pest (insect) Management Trial (OPMT)—were initiated in North Carolina and Mississippi, respec- tively (USDA 1981).
The components of the BWET included: (1) mandatory partici- pation based on a grower referendum; (2) areawide diapause boll weevil control; (3) monitoring and suppression of boll weevils with pheromone traps; (4) selective use of an insect growth regulator; (5) release of sterile boll weevils; (6) increased field scouting; (7) quarantine procedures; and (8) increased education and techni- cal assistance to growers (Ganyard et al. 1981).
The components of the OPMT included: (1) voluntary participa- tion with full reimbursement to producers for diapause control and pinhead square treatments, i.e., incentive payments; (2) overwin- tered boll weevil control in the spring as needed; (3) monitoring of the density of boll weevils with pheromone traps; (4) increased field scouting and improved timing of insecticide treatments; and (5) increased education and technical assistance to growers (An- drews 1981).

12 R. L RIDGWAY AND E. P. LLOYD
Thus the conduct of the BWET and OPMT resulted, at least in part, from a policy directive ordering a comparison of the technol- ogies derived from the TPM and IPM paradigms. However, the inclu- sion of an incentive payment in the OPMT introduced a component that was not a characteristic of the IPM paradigm.
Although the BWET and OPMT were designed to test a number of new technologies for use in two specific areas of the Cotton Belt, additional technologies should be considered when one is designing improved programs applicable to different cotton growing areas. Many of these emerging technologies are becoming available because of the changes in direction of research initiated in the late 1950's by USDA (Hoffmann 1970), because of the expanded public sup- port for research that occurred in the 1960's and early 1970's, and because of increased efforts in the private sector to develop new technologies. Many of the accomplishments of these expanded thrusts are documented in two recent reviews (Knipling 1979, Huf- faker 1980). In addition, major advancements related to cotton in- sect control have been made in the development of modified cotton cultures (Walker and Niles 1971, Bird et al. 1977, and Heilman et al. 1977); in the improvement of microbial agents (ignoffo et al. 1965, Dulmage 1970, Coppedge et al. 1972, and Yearian and Young 1978); in the augmentation of predators and parasites (Ridgway and Vinson 1977); in the discovery and commercialization of insect pheromones (Mitchell 1981); in the development of insect growth regulators (Lloyd et al. 1977, Ganyard et al. 1978); in the discov- ery and improvement of autocidal methods (Laster et al. 1976, Earle et al. 1979, and Wright et al. 1980); and in plant growth regula- tion (Namken and Gausman 1978). Similar advances have also been made in the study of damage thresholds, survey techniques, fore- casting, and systems management (Anonymous 1972, DeMichele and Bottrell 1976, Hartstack et al. 1976, Hartstack et al. 1977, Anony- mous 1978, Sterling 1979, Brown et al. 1979, and Gutierrez et al. 1980).
During the 1970*s while the TPM and IPM paradigms were being scrutinized, the chemical insecticide paradigm that began develop- ing in the mid-1950*s was still very much in existence. The peti- tion for an emergency exemption from registration for DDT on cotton filed with the Environmental Protection Agency by the State of Louisiana (Anonymous 1975) and the petition for emergency exemption from registration for the synthetic pyrethroid insecticides filed by eleven States in 1977 (USDA 1977) constituted a continued ex- pression of the importance of insecticides for control of cotton insects. Also, the role of the private consultant became increas- ingly significant. At the beginning of the decade, in 1970, there were perhaps 500 to 1000 pest control and crop consultants in the U.S. By 1980, the number had increased to about 4000, and this number is likely to increase 4 or 5 fold during the next ten years (Gordon Berg, editor, Ag-Fieldman and Consultants, personal commun- ication). Also, several State consulting associations and a Na- tional Alliance of Independent Crop Consultants were formed during this period.

EVOLUTION OF COTTON INSECT CONTROL 13
The advances that were made during the 1970's in the develop- ment of improved insect control methods and in their application by both the public and private sectors provide a broad base for de- signing and implementing improved cotton insect management systems.
THE FUTURE
The conduct of the BWET and OPMT and the evaluation of alter- native cotton insect management programs for use in the boll weevil infested areas of the Cotton Belt may provide a unique opportunity to influence scientific change. Kuhn (1970), who utilized the con- cept of paradigms to analyze scientific and technical change, pre- sented a general model based upon the emergence, use, and eventual discard of paradigms in scientific communications. Perhaps the en- tomology profession, and particularly that portion of it that will be especially concerned with future directions for cotton insect management, has arrived at a point where the eradication (TPM) and population management (IPM) paradigms described by Rabb (1972) and Perkins (1980a) might be merged into a single paradigm, i.e., area- wide population management (Figure 2). Such a merger would appear to be logical since BWET and OPMT had a number of common elements: (1) high percentage of producer participation; (2) intense monitor- ing of insect populations; (3) application of suppression methods to boll weevil populations in an organized manner; and (4) improved management of local populations of the bollworm and tobacco bud- worm. However, the successful merger of the paradigms will require the establishment of criteria to be used to determine which strat- egy, i.e., local population management, areawide management, or eradication, is most appropriate for use against a particular in- sect pest. For instance, although Rabb (1972) considered eradica- tion an appropriate strategy, he noted that it should be used with considerable caution: "The decision to eradicate ... is a grave responsibility that should be attempted only after careful study involving diverse perspectives has produced convincing evidence that the benefits to be accrued more than balance the resulting ecological impoverishment caused by removal of the pest species." We agree with the need for considerable caution. But when exotic species such as the boll weevil are involved, ecolog- ical disruption that might be caused by the suppression methods used and the risk of reinfestation may be of more concern than any ecological disruption that might be caused by removing a single exotic insect species. Other discussions of the pros and cons provide a useful basis for establishing criteria for selecting the preferred insect management strategy (Eden 1978, Knipling 1978, Newsom 1978, and Rabb 1978).
A broad-based commitment to the merger of the TPM and 0PM par- adigms and the application of the merged paradigm to cotton insect management could provide a basis for areawide management of the boll weevil throughout much of its range and could also provide the framework for areawide management programs for the pink bollworm, bollworm, and tobacco budworm as applicable emerging technologies

14 R. L RI DG WAY AND E. P. LLOYD
Target is insects migrating or being transported
Target is total population over a large area
Containment
I Eradication
I
Designed to prevent reinfestation
Designed for geographically defined area that can be protected from reinfestation
Additional Methods
Target is a high percentage of a population over a large area
Areawide Population
Management
T Additional Methods
Designed for large area communities'
Targets are localized
infestations
Advanced Population Models I
IVIanagement of Localized Populations
♦ Methods Harmonized with
Natural Control
Thresholds Refined
Temporary Alleviation
Designed primarily for individual grower
Emergency Use of Control Method(s)
C Recognition and Preliminary Assessment
of Pest Situation
Figure 2.—Desired evolution of insect pest control actions (modi- fied from Rabb 1972).

EVOLUTION OF COTTON INSECT CONTROL 15
become available. Indeed, the areawide approach may be essential to effective implementation of such new technologies as mating dis- ruption or mass trapping via insect pheromones. In addition, cul- tural controls, including uniform late planting and use of plant growth regulators for host-plant-specific insects like the boll weevil and pink bollworm, would benefit from an areawide approach. The same would be true for such technologies as autocidal methods and augmentation of natural enemies. Also, implementation of area- wide management programs for such exotic insects as the boll weevil and pink bollworms, both of which have a limited host range, could be precursors to eradication of these species from a geographically defined area. At the same time, management of localized popula- tions would continue to be the desired approach for insect pests that only occasionally cause losses.
The design and implementation of desirable cotton insect management programs will require the effective integration of all the groups, organizations, and institutions that can make signifi- cant contributions. The importance of such an effort has been emphasized previously, particularly in the matter of biological control of insects (Figure 3, Ridgway 1972b). Some of the insti- tutional arrangements for deploying technologies in various pest management strategies have been identified (Starler and Ridgway 1977). These institutional arrangements include:
-Private producers of technologies selling directly to users. -Private producers of technologies selling directly to users
along with pest management consultant services. -Private consultants purchasing technologies and using them in
private pest management service programs. -Cooperatives producing the technologies, applying them, and
providing pest management services. -Pest management districts (with special taxation and enforce-
ment authorities) purchasing and applying technologies. -State and Federal agencies producing and/or purchasing the
technologies and implementing them.
The knowledge base that has been developed relative to cotton insect management programs provides a rather unique opportunity to develop more effective integration of the efforts of (1) suppliers of insect control materials; (2) private consultants; (3) commer- cial applicators; (4) education, information, and Extension agen- cies; and (5) action and regulatory agencies. All of these utilize the results of research to assist the cotton producer (Figure 4). Useful models are provided by the approaches used in the PBWEE in southern Mississippi (Lloyd 1972), in the BWET and OPMT (USDA 1978), in the community programs in Arkansas (Phillips et al. 1980), the High Plains boll weevil containment program, and in county programs facilitated by the Texas Pest Management Associa- tion (Haney and Frisbie 1981).
In view of the technologies that are available and of the suc- cessful experiences in implementing many of these technologies.
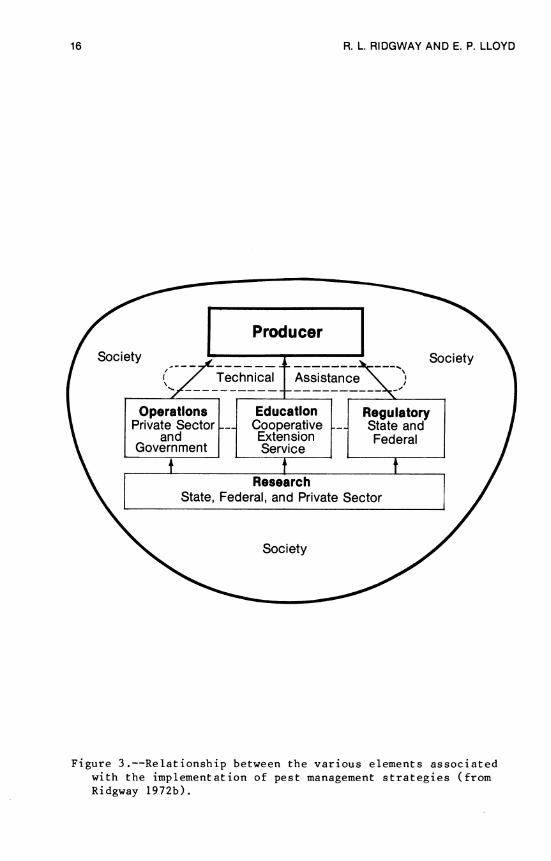
16 R. L RI DG WAY AND E. P. LLOYD
Figure 3.—Relationship between the various elements associated with the implementation of pest management strategies (from Ridgway 1972b).

EVOLUTION OF COTTON INSECT CONTROL 17
RESEARCH (State, Federal and Private Sector)
Figure 4.—Institutions that can contribute to development and im- plementation of improved cotton insect management practices.

18 R. L RIDGWAY AND E. P. LLOYD
increased emphasis should be placed on the design and implementa- tion of improved cotton insect management programs. Since cotton insect management programs will vary from area to area, no attempt has been made in this introductory chapter to describe specific programs; however, this review of the evolution of cotton insect control in the United States was written, and the monograph on "Cotton Insect Management with Special Reference to the Boll Wee- vil" was assembled to facilitate scientific communication and im- proved decision making on the selection of such cotton insect pro- grams. Hopefully, as professional entomologists, cotton producers, and policy makers proceed to influence the future direction of cot- ton insect management, they will critically examine the scientific record, objectively evaluate the most appropriate roles for various institutions, and carefully consider the long range impacts on both agriculture and consumers.

EVOLUTION OF COTTON INSECT CONTROL 19
LITERATURE CITED
Adkisson, P. L. 1972. The integrated control of insect pests of cotton. Pro-
ceedings, Tall Timbers Conference on Ecological Animal Control by Habitat Management 4:175-188.
Andrews, G. L. 1981. Special insect projects: optimum insect management trial. Proceedings, Cotton Production and Mechanization Conference, National Cotton Council, Memphis, TN, p. 41-44.
Anonymous. 1972. Distribution, abundance and control of Heliothis species
in cotton and other host plants. Southern Cooperative Series Bulletin No. 169, Oklahoma Agricultural Experiment Station, 92 p.
1975. A petition for emergency use of DDT on cotton for con- trol of the tobacco budworm. State of Louisiana, unpublished report, 49 p.
1978. The boll weevil: management strategies. Southern Coop- erative Series Bulletin No. 228, Arkansas Agricultural Experi- ment Station, 130 p.
Bird, L. S., F. M. Bourland, R. G. Percy, J. E. Hood, and D. L. Bush.
1977. Additional progress in developing okra leaf, frego bract and glabrous multi-adversity resistant cottons. Proceedings, Beltwide Cotton Production Research Conference, National Cotton Council, Memphis, TN, p. 107-110.
Bottrell, D. G., and P. L. Adkisson. 1977. Cotton insect pest management. Annual Review of Ento- mology 22:451-481.
Brazzel, J. R. 1963. Resistance to DDT in Heliothis virescens. Journal of Economic Entomology 56:571-574.
Brown, L. G., R. W. McClendon, and J. W. Jones. 1979. Computer simulation of the interaction between the cotton
crop and insect pests. Transactions of the ASAE 22:771-774.
Carson, Rachel. 1962. Silent spring. Fawcett Books Group, Brooklyn, NY, 304 p.
Coad, B. R. 1918. Recent experimental work on poisoning cotton boll weevils. U.S. Department of Agriculture Bulletin 731, 15 p.

20 R. L RI DG WAY AND E. P. LLOYD
Coad, B. R., and T. P. Cassidy. 1920. Cotton boll weevil control by the use of poison. U. Department of Agriculture Bulletin 875, 31 p.
E. Johnson, and G. L. McNeil. 1924. Dusting cotton from aeroplanes. U.S. Department of Agriculture Bulletin 1204, 40 p.
Comstock, J. H. 1879. The boll worm, report on cotton insects. Government Printing Office, Washington, D.C., p. 287-315.
Coppedge, J. R., R. E. Kinzer, and R. L. Ridgway. 1972. Field evaluations of Bacillus thuringiensis and Heliothis nuclear polyhedrosis virus for control of Heliothis on cotton. Texas Agricultural Experiment Station PR-3086, p. 43-48.
DeMichele, D. W., and D. G. Bottrell. 1976. Systems approach to cotton insect pest management. Ln
Integrated Pest Management, J. L. Apple and R. F. Smith, edi- tors. Plenum Press, NY, 200 p.
Dulmage, H. T. 1970. Insecticide activity of HD-1, a new isolate of Bacillus
thuringiensis var. alesti. Journal of Invertebrate Pathology 15:232-239.
Dunlap, T. R. 1981. DDT: scientists, citizens and public policy. Princeton University Press, Princeton, NJ, 318 p.
Earle, N. W., L. A. Simmons, and S. S. Nilakhe. 1979. Laboratory studies of sterility and competitiveness of
boll weevils irradiated in an atmosphere of nitrogen, carbon dioxide or air. Journal of Economic Entomology 72:687-691.
Economics and Statistics Service. 1981. Overall evaluation of beltwide boll weevil/cotton insect management program. Economics and Statistics Service, Staff report. May.
Eden, W. G. 1978. Eradication of plant pests-pro. Bulletin of the Entomo-
logical Society of America 24:52-54
Ewing, K. P., and E. E. Ivy. 1943. Some factors influencing bollworm populations and damage.
Journal of Economic Entomology 36:602-606.

EVOLUTION OF COTTON INSECT CONTROL 21
Ewing, K. P., and C. R. Parencia, Jr. 1950. Early-season applications of insecticides on a community- wide basis for cotton-insect control in 1950. U.S. Department of Agriculture, Bureau of Entomology and Plant Quarantine E-810, 8 p.
C. R. Parencia, Jr., and E. E. Ivy. 1947. Cotton insect control with benzene hexachloride, alone or
in mixture with DDT. Journal of Economic Entomology 40:374- 381.
Gaines, J. C. 1957. Cotton insects and their control in the United States. Annual Review of Entomology 2:319-338.
and H. A. Dean. 1947. New insecticides for boll weevil, bollworm, and aphid con-
trol. Journal of Economic Entomology 40:363-370.
and R. Wipprecht. 1948. Effect of dusting schedules on yield of cotton during
1947. Journal of Economic Entomology 41:410-412.
Ganyard, M. C, J. R. Bradley, Jr., and J. R. Brazzel. 1978. Wide-area field test of diflubenzuron for control of an
indigenous boll weevil population. Journal of Economic Ento- mology 71:785-788.
J. R. Brazzel, J. H. Dillier, and A. E. Miller. 1981. Boll weevil eradication trial. Proceedings, Cotton Pro-
duction and Mechanization Conference, National Cotton Council, Memphis, TN, p. 38-40.
Good, J. M., and N. P. Bonell. 1976. Cotton insect pest management pilot projects, 1972-74.
U.S. Department of Agriculture, Exjiension Service 5-64 (4-76). Washington, D.C., 99 p.
Graham, H. M., editor. 1980. Pink bollworm control in the United States. U.S. Depart- ment of Agriculture, Science and Education Administration, Agricultural Research, ARM-W-16, 81 p.
Graves, J. B., J. S. Roussel, and J. R. Phillips. 1963. Resistance to some chlorinated hydrocarbon insecticides
in the bollworm, Heliothis zea. Journal of Economic Entomology 56:442-444.
Gutierrez, A. P., D. W. DeMichele, Y. Wang, G. L. Curry, R. Skeith, and L. G. Brown.
1980. The systems approach to research and decision making for cotton pest control. In New Technology of Pest Control. C. B. Huffaker, editor, John Wiley and Sons, NY, p. 155-186.

22 R. L RIDGWAY AND E. P. LLOYD
Handy, R. B. 1896. History and general statistics of cotton, Iri The Cotton
Plant. U.S. Department of Agriculture Bulletin 33, p. 17-66.
Haney, R. L., and R. E. Frisbie. 1981. Texas producers, consumers to benefit from statewide pest management plan. The Cotton Gin and Oil Mill Press. Volume 18:15, p. 18.
Hartstack, A. W., Jr., J. A. Witz, J. P. Hollingsworth, R. L. Ridgway, and J. D. Lopez, Jr.
1976. MOTHZV-2: A computer simulation of Heliothis zea and Heliothis virescens population dynamics. U.S. Department of Agriculture ARS-5-127, 55 p.
J. L. Henson, J. A. Witz, J. A. Jackman, J. P. Hollings- worth, and R. E. Frisbie.
1977. The Texas program for forecasting Heliothis spp. infesta- tion on cotton. Proceedings, Beltwide Cotton Producers Re- search Conference, National Cotton Council, Memphis, TN, p. 151-154.
Heilman, M. D., M. J. Lukefahr, L. N. Namken, and J. W. Norman. 1977. Field evaluation of a short season production system in
the lower Rio Grande Valley of Texas. Proceedings, Beltwide Cotton Production Research Conference, National Cotton Council, Memphis, TN, p. 80-83.
Hinds, W. E. 1925. Airplane dusting of cotton for boll weevil control. Journal of Economic Entomology 19:607.
Hoffmann, C. H. 1970. Alternatives to conventional insecticides for control of
insect pests. Agricultural Chemicals 25:19, 21-23, 35.
Howard, L. 0. 1896. The insects which affect the cotton plant in the United
States. ^ The Cotton Plant. U.S. Department of Agriculture Bulletin 33, p. 317-350.
Huffaker, C. B., editor. 1980. New technology of pest control. John Wiley, NY, 500 p.
and R. F. Smith. 1980. Rationale, organization and development of a national in-
tegrated pest management project. Tn New Technology in Pest Control, C. B. Huffaker, editor. John Wiley, NY, p. 2-24.
Hunter, W. D., and B. R. Coad. 1923. The boll weevil problem. U.S. Department of Agriculture,
Farmers* Bulletin 1329-1330.

EVOLUTION OF COTTON INSECT CONTROL 23
Hunter, W. D., and W. E. Hinds. 1905. The Mexican cotton boll weevil. U.S. Department of Agri-
culture, Bureau of Entomology Bulletin 51. 181 p.
and W. D. Pierce. 1912. The Mexican boll weevil: a summary of the investigations
of the insect up to December 31, 1911. Senate Document 305, 188 p.
Ignoffo, C. M., A. J. Chapman, and D. F. Martin. 1965. The nuclear polyhedrosis virus of Heliothis zea (Boddie)
and Heliothis virescens (F.) III. Effectiveness of the virus against field populations of Heliothis on cotton, corn and grain sorghum. Journal of Invertebrate Pathology 7:227-235.
Isley, D., and W. J. Baerg. 1924. The boll weevil problem in Arkansas. Arkansas Agricul-
tural Experiment Station Bulletin 190, 22 p.
Ivy, E. E. 1944. Tests with DDT on the more important cotton insects. Journal of Economic Entomology 37:142.
and K. P. Ewing. 1946. Benzene hexachloride to control cotton insects. Journal of Economic Entomology 39:38-41.
Knipling, E. F. 1978. Advances in technology for insect population eradication
and suppression. Bulletin of the Entomological Society of America 24:44-52.
1979. The basic principles of insect population suppression and management. U.S. Department of Agriculture, Agricultural Hand- book 512, p. 658-659.
Kuhn, T. S. 1970. The structure of scientific revolutions. 2nd edition. University of Chicago Press, Chicago, 210 p.
Laster, M. L., D. F. Martin, and D. W. Parvin, Jr. 1976. Potential for suppressing tobacco budworm (Lepidoptera: Noctuidae) by genetic sterilization. Mississippi Agricultural and Forestry Experiment Station, Technical Bulletin No. 82, 9 p.
Lloyd, E. P. 1972. Progress report on the pilot boll weevil eradication ex-
periment. Proceedings, Beltwide Cotton Production Mechaniza- tion Conference, National Cotton Council, Memphis, TN, p. 46- 49.

24 R. L RI DG WAY AND E. P. LLOYD
Lloyd, E. P., R. H. Wood, and E. B. Mitchell. 1977. Suppression with Th-6040 applied in cottonseed oil as a
foliar spray. Journal of Economic Entomology 70:442-444.
Mitchell, E. R., editor. 1981. Management of insect pests with semiochemicals. Plenum Publishing Corporation, NY, 514 p.
Namken, L. N., and H. N. Gausman. 1978. Practical aspects of chemical regulation of cotton plant
growth and fruiting. Proceedings, Cotton Production Mechan- ization Conference, National Cotton Council, Memphis, TN, p. 23-25.
Nemec, S., and P. L. Adkisson. 1969. Laboratory tests of insecticides for bollworm, tobacco budworm and boll weevil control. Texas Agricultural Experiment Station Progress Report 2674, 4 p.
Newsom, L. D. 1974. Pest management: history, current status and future pro-
gress. Ln Proceedings of the Summer Institute of Biological Control of Plant Insects and Diseases. F. G. Maxwell and F. A. Harris, editors. University of Mississippi Press, Jackson, MS, p. 1-18.
1978. Eradication of plant pests—con. Bulletin of the Entomo- logical Society of America 24:35-40.
and J. R. Brazzel. 1968. Pests and their control. Ln Advances in Production and Utilization of Cotton, F. C. Elliott, et al., editors, Iowa State University Press, Ames, lA, p. 367-405.
Noble, L. W. 1969. Fifty years of research on the pink bollworm in the United
States. U.S. Department of Agriculture, Agriculture Handbook 357, 62 p.
Parencia, C. R., Jr. 1978. One-hundred twenty years of research on cotton insects in
the United States. Agricultural Research Service, U.S. Depart- ment of Agriculture, Agriculture Handbook 515, 75 p.
Perkins, J. H. 1980a. The quest for innovation in agricultural entomology,
1945-1978. Ir^ Pest Control: Cultural and Environmental Aspects. D. Pimentel and J, H. Perkins, editors, Westview Press, Boulder, CO, p. 23-80.'
1980b. Boll weevil eradication. Science 207:1044-1050.

EVOLUTION OF COTTON INSECT CONTROL 25
Phillips, J. R., A. P. Gutierrez, and P. L. Adkisson. 1980. Cotton. Ln New Technology of Pest Control, C. B. Huf-
faker, editor, John Wiley and Sons, NY, p. 123-153.
Pickett, A. D. 1949. A critique of insect control methods. Canadian Ento- mologist 81:67-76.
Pierce, W. D. 1922. How insects affect the cotton plant and means of combat-
ing them. U.S. Department of Agriculture, Farmers' Bulletin 890.
Post, G. B. 1924. Boll weevil control by airplane. Georgia State College of Agriculture, Extension Division, Bulletin 301, 13:22.
Rabb, R. L. 1972. Principles and concepts of pest management. In Proceed-
ings of the National Insect Pest Management Workshop, Purdue University, West Lafayette, IN, p. 6-29.
1978. Eradication of plant pests—con. Bulletin of the Entomo- logical Society of America 24:40-44.
Reynolds, H. T., P. L. Adkisson, and R. F. Smith. 1975. Cotton insect pest management. Tn Introduction to Insect Pest Management. R. L. Metcalf and W. Luckmann, editors, John Wiley and Sons, NY, p. 379-443.
Ridgway, R. L. 1969. Control of the bollworm and tobacco budworm through con-
servation and augmentation of predaceous insects. Proceedings, Tall Timbers Conference on Ecological Animal Control of Habitat Management No. 1, p. 127-144.
1972a. Integrated control of cotton insects in the United States. Paper presented at the 31st Plenary Meeting of the International Cotton Advisory Committee, Managua, Nicaragua, October 22-28, 1972, 20 p.
1972b. Use of parasites, predators, and microbial agents in management of insect pests of crops. Tn Proceedings of the National Insect Pest Management Workshop, Purdue University, West Lafayette, IN, p. 51-62.
P. D. Lingren, C. B. Cowan, Jr., and J. W. Davis. 1967. Populations of arthropod predators and Heliothis spp.
after application of systemic insecticides. Journal of Eco- nomic Entomology 60:1012-1016.

26 R. L RIDGWAY AND E. P. LLOYD
Ridgway, R. L., J. C. Tinney, J. T. MacGregor, and N. E. Starler. 1978. Pesticide use in agriculture. Environmental Health Per-
spectives 27:103-112.
and S. B. Vinson. 1977. Biological control by augmentation of natural enemies. Plenum Press, NY, 480 p.
Ripper, W. E., R. M. Greenslade, J. Heath, and C. H. Barker. 1948. New formulations of DDT with selective properties. Nature 161:484-485.
R. M. Greenslade, and L. A. Lickerish. 1949. Combined chemical and biological control of insects by means of a systemic insecticide. Nature 163:787-789.
Roussel, J. S., and D. F. Glower. 1955. Resistance to the chlorinated hydrocarbon insecticides in
the boll weevil. Louisiana Agricultural Experiment Station, Circular 41, 9 p.
Shepard, H. A. 1951. The chemistry and action of insecticides. McGraw Hill,
NY.
Starbird, I. R., and B. L. French. 1972. Costs of producing upland cotton in the United States.
1969. U.S. Department of Agriculture, Economic Research Ser- vice, 48 p.
Starler, N. H., and R. L. Ridgway. 1977. Economic and social considerations for the utilization of
augmentation of natural enemies. Iji Biological Control by Augmentation of Natural Enemies. R. L. Ridgway and S. B. Vinson, editors. Plenum Press, NY.
Sterling, W. L., editor. 1979. Economic thresholds and sampling of Heliothis spp. on
cotton, corn, soybeans and other host plants. Southern Coop- erative Series Bulletin No. 231, Texas Agricultural Experiment Station, 159 p.
Stern, V., R. F. Smith, R. van den Bosch, and K. S. Hagen. 1959. The integrated control concept. Hilgardia 29:81-101.
Townsend, C. H. T. 1895. Report on the Mexican cotton boll weevil in Texas
(Anthonomus grandis Boh.). Insect Life 7:295-309.
U.S. Congress. 1965. Agricultural Appropriations for 1966, Hearings, Part 1,
Senate Committee on Appropriations. Government Printing Office, Washington, D.C.

EVOLUTION OF COTTON INSECT CONTROL 27
USDA. 1958. The boll weevil problem and research and facility needs
to meet the problem. U.S. Department of Agriculture. A report prepared at the request of the U.S. Congress. December 30, 50 p.
1977. The bollworm and tobacco budworm on cotton: status of in- secticides and other methods of control for 1977. U.S. Depart- ment of Agriculture, Washington, D.C. Unpublished report. 46 p.
1978. Plans for the boll weevil eradication trial and the optimum pest management trial and their evaluations. U.S. De- partment of Agriculture, Washington, D.C. Unpublished report.
1981. Executive overview of alternative boll weevil/cotton in- sect management programs. U.S. Department of Agriculture. Un- published report. May 19, 25 p.
van den Bosch, R., T. F. Leigh, L. A. Falcon, V. M. Stern, D. Gonzalez, and K. S. Hagen.
1971, The developing program of integrated pest management of cotton insects in California. Tn Biological Control, D. B. Huffaker, editor. Plenum Press, NY, p. 377-394.
Walker, J. K., Jr., B. G. Hightower, R. L. Hanna, and D. F. Martin. 1956. Control of boll weevils resistant to chlorinated hydro-
carbons. Texas Agricultural Experiment Station, Progress Report 1902.
and G. A. Niles. 1971. Population dynamics of the boll weevil and modified cotton
types. Texas Agricultural Experiment Station, Bulletin B-1109, 14 p.
Wilson, R. J. 1926. Boll weevil control by airplane. Agriculture—an attrac-
tive field for commercial aviation. Congressional Record 67: 2887-2888.
Wright, J. E., R. Moore, J. McCoy, G. Wiygul, and J. Haynes. 1980. Comparison of three sterilization procedures on the
quality of the male boll weevil. Journal of Economic Ento- mology 73:493-496.
Yearian, W. C, and S. Y. Young. 1978. Application of microbial pesticides on field crops. Miscellaneous Publication of the Entomological Society of America 10:21-26.


29
Chapter 2
THE COST OF INSECTICIDES USED ON COTTON IN THE UNITED STATES
F. T. Cooke, Jr. Economic Research Service U.S. Department of Agriculture Stoneville, MS 38776
D. W. Parvin, Jr. Department of Agricultural Economics Mississippi Agricultural and
Forestry Experiment Station Mississippi State University Mississippi State, MS 39762
ABSTRACT Expenditures for cotton insecticides in the United States increased from 96 million dollars in 1971 to 235 million dollars in 1978. However, during about the same period (1971-1977), quantities of insecticides applied to cotton in the United States declined from 73 million pounds of active ingredients to something over 39 million pounds.
This apparent conflict in the data results, at least in part, from the fact that the most common treatment used against the bollworm (Heli- othis zea (Boddie)) and the tobacco budworm (H. virescens (F.)) in the early seventies, toxaphene at 2.0 pounds active ingredients (a.i.) plus 1.0 pound a.i. methyl parathion, had been replaced in part by the synthetic pyrethroid materials at 0.1 pound a.i. by 1978.
Infestations of cotton insect pests are highly variable. However, at the national lev- el, the cost per acre of cotton insecticide ma- terial has remained remarkably constant. In four of the six surveys made in the years be- tween 1966 and 1977, average material cost per treated acre has ranged between 20 and 25 dol- lars. Nevertheless, the Southeast and the Mid- south have been experiencing a trend to in- creasing material cost per acre while most of the Southern Plains (Texas and Oklahoma) have had decreasing cost.
30 F. T. COOKE, JR., AND D. W. PARVIN, JR.
INTRODUCTION
The cost of controlling insect pests of cotton constitutes an important part of the total cost of producing cotton in many areas of the United States. Because the kinds of insect pests and the severity of infestations are highly variable and difficult to pre- dict, cotton producers and policy makers have difficulty forecast- ing the expected cost of insect control for a given year and over time. Therefore, there is a need to examine carefully the current and past cost of controlling cotton insects in order to have a ba- sis for estimating the impacts of proposed future programs on in- secticide use.
The primary purpose of this chapter is to provide information concerning the use of insecticides on cotton in different cotton producing regions and subregions of the United States. Such in- formation will be useful in the evaluation of alternative beltwide cotton insect control strategies for cotton insects (Keller 1980).
OVERVIEW OF PESTICIDE USE
A perspective on pesticide use and the importance of pesti- cides (includes but is not limited to insecticides) is provided by an examination of the information on sales of pesticides in the world and in the United States for the past decade. Such data were obtained from confidential surveys of leading international market- ers as reported in Farm Chemicals (Anonymous 1975, 1977, 1979). In 1971, the pesticide market in the United States represented 40% of the world market, but in 1974 and 1978 the proportion stabilized at 33% (Table 1). Insecticide sales represented about 35% of
Table 1.—The world and U.S. pesticide markets, user's level J^/, selected years (constant dollars, 1979 = 100)
World 2/ United Stat es 1971 1974 1978 1971 1974 1978
Pesticide ■(millions dollars ;)
Herbicides 1,672 2,687 4,218 946 1,298 1,966 Insecticides 1,245 2,236 3,437 325 603 919 Fungicides 507 1,179 1,747 95 142 215 Soil fumigants 31 85 191 6 25 74 Defoliants and
desiccants 18 23 110 12 22 43 Other 3/ 59 94 137 27 36 30
Total 3,532 6,304 9,840 1,411 2,126 3,247
\J End use. 2^/ Includes U.S. figures. 3^/ Includes growth regulators, pheromones, attractants, and
viruses.
Source: Anonymous 1975, 1977, and 1979.

COST OF INSECTICIDES USED ON COTTON 31
pesticide sales in the world in 1971, 1974, and 1978, but in the United States, insecticide sales as a percentage of pesticide sales increased from 23 in 1971 to 28 in 1974 and 1978. Total pesticide sales increased from $1.4 billion in 1971 to nearly $3.3 billion in 1978. Herbicide sales increased about $1 billion and insecticide sales rose nearly $600 million increasing their total market share. The increased sales are, in part, a result of the use of more expensive insecticides.
Corn and cotton production provide the two largest single crop pesticide markets in the world and the United States (Table 2). At the world level, pesticide sales for cotton production exceeded those for corn production in 1971 and 1974. That relationship was reversed in 1978. In the United States, pesticide sales for corn production have consistently exceeded those for cotton production, but the figures reflect a strong, consistent upward trend for both.
Specifically, the world markets for cotton insecticides in- creased 122% (Table 3), and the U.S. market increased 145% during this period. The five largest single crop insecticide markets in the United States for 1971, 1974, and 1978 are listed in Table 4. In 1971 and 1974, the sales of cotton insecticides in the United States exceeded the sales of insecticides used for corn production. In 1978, expenditures for corn insecticides in the United States were approximately equal to expenditures for cotton insecticides. The acreage of corn produced in the United States has increased steadily over this period while cotton acreage has remained fairly constant.
Table 2.—Two largest world and U.S. single crop pesticide markets, user's level 1/, selected years (constant dollars 1979 = 100)
World y United States
1971 1974 1978 nillions
1971 1974 1978 Crop — — _ ~" I
Corn
Cotton
668
720
1,071
1,146
1,460
1,388
430
265
628 989
301 407
\J End use. Ij Includes U.S. figures.
Source: Anonymous 1975, 1977, and 1979.

32 F. T. COOKE, JR., AND D. W. PARVIN, JR.
Table 3.—World and U.S. cotton insecticide markets, user's level J^/, selected years (constant dollars 1979 = 100)
World l_l United States Year - - - - millions dollars -------
1971 455 96 1974 816 168 1978 1,012 235
\_l End use. Ij Includes U.S. figures.
Source: Anonymous 1975, 1977, and 1979.
Table 4.—Five largest U.S. single crop insecticide markets, user's level J^/, selected years (constant dollars 1979 = 100)
Crop
Cotton Corn Soybeans Alfalfa Tobacco
971 1974 1978 , . - millions dollars -
96 168 235 66 125 236
18 34 7 22 25 9 21 23
1_/ End use.
Source: Anonymous 1975, 1977, and 1979.
COTTON INSECTICIDE USE
The U.S. Department of Agriculture has completed four studies of farmer uses of pesticides on cotton and other crops. These sur- veys are available for the 1964, 1966, 1971, and 1976 crop years (Eichers et al. 1968, 1970, 1978; Andrilenas 1974). Also, the U.S. Department of Agriculture routinely publishes estimates of the cost of producing selected crops in the United States. These cotton cost of production estimates were supported by three surveys .(Starbird 1974, Krenz et al. 1976, U.S. Department of Agriculture 1979b), and the figures were used to develop the cost of insect control on cotton for 1969, 1972, and 1974 (Cooke and Parvin 1981b). In addition, a special cotton pesticide survey was con- ducted for the 1977 crop year (Lin et al. [in review]). Also, a 1961 survey was conducted in only the portions of the Cotton Belt that were heavily infested with boll weevils (Langsford 1964).

COST OF INSECTICIDES USED ON COTTON 33
Finally, the so-called "normalized" study (Cooke and Parvin 1981a) was made only in the States infested with boll weevils (Cal- ifornia, Arizona, and New Mexico were excluded) and insect pressure, normal weather, and 1979 technology were assumed. Information was obtained by interviewing knowledgeable individuals in the boll weevil infested portion of the Cotton Belt. A few of these were agronomists and producers, but most respondents were Extension entomologists. Respondents tended to consider the last 5-10 years and conceptually averaged insect pressure and weather over that time. Basically, 1979 technology implied insecticides with 1979 labels.
Results of all of the specific studies are summarized in Table 5, Part I. Sample size, that is, number of farmers surveyed, ranged from 143 in 1976 to 2019 in 1977 to 4200 in 1969. The size of each of the study areas in terms of planted and harvested acres is reported in columns 3 and 4, and the percentage of acres treated is given in column 5. The 1961 study was conducted in areas where the boll weevil was considered to be a severe problem; thus, it would be inappropriate to expand these results to other areas of the Cotton Belt.
The beltwide estimates are reported in Table 5, Part II. Col- umns 12 and 13 are estimates of acres planted and harvested by year. Column 14 reports the number of applications per harvested acre. Column 15 lists acre treatments. From these last two columns, it can be seen that 1972 and 1974 were years of heavy insecticide use; 1969 was a light year.
POOLING THE SURVEYS TO APPROXIMATE EXPECTED VALUES
The usual method of estimating the extent of insecticide use and the cost is by conducting a survey. Over a period of years, as noted, ten surveys have been conducted (see column 1 of Table 5) that provided information on insecticide use and cost. However, several problems prohibit the simple averaging of the information to obtain estimates of the expected level of insecticide use and cost for the various regions and subregions of the Cotton Belt that are required to evaluate alternative beltwide cotton insect control strategies.
Generally, the purposes or objectives of the surveys were dif- ferent. When one then tries to extract information from a survey that was not designed to provide such information, a large sample size is preferred. The sample size associated with a few of the surveys was too small to disaggregate the information into the re- quired sub-State regions. Also, the regions and subregions select- ed for sampling in the various surveys were different.
In addition, costs over time must be compared in terms of con- stant dollars, but no valid index of prices paid by farmers for

34 . T. COOKE, JR., AND D. W. PARVIN, JR.
T3 X (U eu 4-t ■U
C c ^ •^ CO .. cu
M o M c
cu XJ V-i u Q; CO U-(
çu eu 1 VJ
r«. 4-t r^ c ON eu
•iH
1 c (U
v£> > ON c
o o
c iu o o
•H M-i •u tu ^O
g (U Cfl
o P U-l
p eu u CO
Q) W CO :3 eu
CO a; (U
TJ ^ J-l
o C •iH eu •U >^ o CO <i) a
I CO
m 0)
(U g '-* D
CO N-^
H
1 O 4J CO O ^ <d Pá Pá VvD tó 00 m pá <^ <!
(U CO ■u o u^ Z ¡z; s CO ¡z; O r^ :zi oo iz; ^ eu C « i-H <f —t eX) o o j-i eu ^ •\ »» < 4-i
". <JN C^
in <j-
1 1 Ë CO
0 o u c -r^ (U
,-H Pu eu o- (U y-^ r^ r-H ^ txc a. CO >-( <i- < Oá Da . p¿¡ .
CO CO c o rH ¡23 Z ^ CN iZi <t <J- M o CO >^^ "^«v.
eu }-« T- -•1 > eu 4J
<: J3
T3 '^ (U O
C CO 4-» O <t <t vO vO vO vO cn ej\ r-- in O <u CO O /^-N C^ o CM ro on vD r^ in r^ v£> u ^-1 eu - CO vX5 o in O r-. 00 in 00 i-H e^ 4-1 o > r-( r—4 n r. » n r.
O CO VJ ^ in <r CJN ,—1 r-H OJ CN O m <N u CO X ""* '~' '"^ '~~* '"^ *"* •"^
(U
¡3
T3 O c CO eu o 00 CNI <t vD r^ en 00 <f r—1
o eu 4-. o /"-s 00 00 vO in CN r^ in •vi- <f 4-» u P « (N m r-«. m 00 m C3> C3N vo r^ 4-1 o ce ^ f—f « »> •» •N « »» O CO i-H Nw' vO <f o ,—1 CM (^ en ,—t c^ U O. X .—1 1—t
1 O 4J CO o CN pei »S CNI P^ r—t in Pi ,—H o
eu CO 4-> O ^N 00 IS !z¡ 00 13 O 00 ¡s vO <1- u eu c « r-H O <t o r^ r^ 00 o s^ CU «—1 1—1 »\ n « n r>
<: 4-1 £2 >-• CNI vO r^ O 00 00 X m CN <r in m CO
C T3 O CU
5-. -^ 4J CU 4-> CO ^^ ^ CO (U eu o CNI PS P£¡ '—• Pí <—' in pá CM Ë o >-i u r-> . S !z: • ^ • • 53 • D «IH 4-» o ^-^ r^ <^ vD vo in
^: -H CO a VJ a Q; CO (X
y~^ 1—< <^ vo e3> r—t CN 0> vO vO vO vO r^ r^ w o> 0> ON ejN C7> ON
cnl
o ¡3
M C CO 1
á eu 1
v£J
CU r^ r^ i-H O C7N ^ o O u u 1 .n
0- 1 ON
X
eu vO 00 VJ CJN ON (D r—1 r—1
> CU .n C CO o '1-*
> CO . CTN U
CO cô TJ >. ÇL,
CU ÛÛ . 00 ^-< O 1—1 T3 ON CU rH CO C
'O O CO •PH c 4-t C
CO ^ eu eu •r-" C o ^ > O 4-1 eu CO O }-l u r—t 4-t u O CO
eu <u u PH CO PQ CJN x: 1 CO r-. u 1 T3 1^ c C^N •iH <h C
o H r>. CO 1—1 4-t 1 CJN • I-' 4-t n (U > O }-i vO ,i¿ eu U eu v>D .n O eu rC C3> rO o 1^ eu 4-t
CO •"^
00 1 «—t 'T~> eu • r\ CJN 1 f—1 4-t :5 00 1—1 T3 o C vO <U
rO eu i-H C3> P N CO •i-<
eu 0) > 1—(
x: ^ }-l . U CO 4-t 4-t O i-H CO B
c CO PH u eu CO o }-i CO #« 4-1 X3 13 <u CO <u eu C ^ a }-• CO .*> ^ Ë D CO r—1
o CO u eu 15 CO u CO eu .^ (U CO c eu rC o eu eu >^ CJ o > >-- P- • r-) ^ eu CO o
4-1 ■TD 4-t 1 1 1
}^
^3 eu O 1 CM C eu CO > eu <t r<- M 4-1 4-t • r-i CO vO <7N b-_i
O 1—4 i-i c ON (D 0 eu •H ,—1 <
1—1 CO T3 .n r-H eu eu (U . n >^ CO CO >-t (U CîC >d- r-
X CO vD CJN 4-1 C T3 >-• C3> eu
C 4-t eu CO o > CO p
'O a c CO TJ CO •r-'
eu X }-• P hJ 4-t eu 'O CO O eu 1 O «—H eu M-l r—1 1 D o 3 4-1 CO •iH r«»
TJ 4-i o CO ÖC }-l r^ C o o C •13 CJN O eu •H CO P CJ 4-t
CO 1 <
1 _ CO •rH CO • i-< 1 1 oo" CO }-l r-H r^ ^ CUT? - vO r-v ON
O eu XI CJN CJN >^ U 4-t <u 1—1 r—1
XI a )-l N • D a o •iH .#< 1—1
4-1 CO a I—1 XI CO CO c (U CO <D
•rH V-i g CJ 00 4-t CO ^ }-• C3N eu
•IH (D 4-» o D ^ XI O ^ O CO H
TJ 13 W P
eu "*>«. T—1 "•»v. *»««^ '> jp -^ 3 CM co|
CO CJ
•i-i W

COST OF INSECTICIDES USED ON COTTON 35
4-) Cd 4J
4J ^ X
tí tí tD -H
(O .. QJ M (J M tí
<ü 4J ^4
Cd M-<
I »-• I
r^ 4J f^ tí
tí u o o
•H M-l
Cd 'ü
(O
<U <ü T3 ^ •H 4J O tí
4J V^ O cd (1) a<
I i CO • »-I
LO <U
^ tí Cd ^»^ H
1 4J
o (0 o ^ <d p£¡ P£3 vD pía 00 tn pá <t
(1) Cd 4-» o in â g ;z¡ CO a o r^ 23 00 »-I Q) tí - rH <!■ -H 00 O o u (U .-H s.^ n •» #» «
<»1 u B ON CO r^ .-1 X es m m <t
I I B ^ tí -H
cd Cd u
u <u
(U CO Í-I tí o o Cd
á
pc5 r^ rH vo CO
. Pá • • Pá • <Ö CM g <1- <!• Ä CO 55
<í ,û
n3 /"^ <ü O
CO *J O <U CO o ^1 di ^ O > -H Cd >-i
Cd X
'O o CO (ü o 0) jj o V4 tí o Cd .-H cd i-H
o- X
<t <t vD vo vo vO CO CJN CO O CM CO CO vO r-- LO vO O m o r^ 00 LO 00
LO vO CO
lO <}■ <7^
00 CM <f /-s 00 00 vO CM LO r-- CO
vO r^ CO lO CM r- 00 CO <JN
00 <1- rH «^ LO -^t ^Ct «J^ <jN vo r^ CO
• vO ^t o .H CM CO
I o 4-» CO o
<u Cd 4J o tí - B
X
CO tí Ti O <U
u •1-4 4-i <D 4J Cd
X> Cd CÜ Q) O u U
j3 •H 4-» O !z: iH
ex }-l CU Qi Cd ÇU
Cd
CMPCJPCJ cM^r-H lOtór-H ^-NOOZ^; OOZO ooavo iH O <t O r^ í^
o 00
vO CM
o lO
00 CO
p^ ^ PCJ ^ lO Pí CM
<t vO vO LO
lO
LO
U Cd
^-N rH <t vO 0^ ^ CM <!■ vO r^ B CJ> vO vO vO vO r^ r^ r-- p^ r^ i^ v-^ <T^ a\ C^ CJ^ CJ^ a\ <J^ CJN <JN O
•-H rH •H rH iH 1—1 .H rH fH 5a
M T) tí Cd 1
g (ü vO
r-í O C3N O ^
O U M 1 •- CU 1 ^
0> rH (U VO 00 M C7^ <7> a> rH «-H
> <ü .^ tí (0 O «H
Cd , rH cd Cd
-TS >. ÇU -H <u W) 00 u o rH T3 C7N Q) rH Cd tí -H
»0 o Cd •H tí 4J tí
CO ^ (U <U «H tí • o ^ > O 4J (U CO o J-l O 1-1 4-> »-I o Cd
Oí <ü u Cl^ CO PQ a^ ^ 1 Cd r^ o 1 T3 ^ tí C3> •H <f tí
O rH W r-. Cd rH 4J 1 os •H 4J n 1 .H (U > O M vo ^ <u u 0) vO .« o <U Xi .<3^ J3 o > (U 4-» ^ .-H U
V4 Cd 00 1 rH »H 0) ."<J^ 1 1-1 *J ^ 00 —1 T) O tí vO 0)
Xi Q) rH 0^ tí N Cd .H «H «H
<U (U a > •-( ^ rtí . »-I Cd 4-> 4-» o r-t Cd S
Cd PM C tí <U CO O Í-I CO n ■u T3 53 Q) Cd <ü 0) tí ^ a V4 Cd .« ^ Ë 3 CD .—.
CO M d) 1^ CO o CO <U ^ <U Cd tí (U ^ O -H QJ (U }-l o o > u a •H u Oí Cd 0 t¿l 1 V4
4J TJ +J 1 i T3 (U u 1 CM tí Q) CO > (U ^ r^ M 4_) 4_) •H CO vO <J^ •—' O 1-1 Wl tí <y\ -H CU ;3 (U «H rH
1-4 CO T3 .« rH
Q) <U <U .«<r Cd CO }-l <U 00 «^ r^
^ cd vO 0^ 4-»
tí 'O V4 0^ rH 0) •H tí 4J <ü rH
Cd O > CO tí TJ CU tí Cd TJ Cd «H
<ü X }^ tí hJ ■U (U ^3 CO O Û> 1 o iH <D ^ 1-1 1 :3 o 3 -«J CO -H r>.
»o 4-» o Cd W) V4 r^
tí ü o tí T3 CTN o <u •H Cd tí ^ o 4-» -O TJ ►J <J
Cd tí tí 1 1 •" CO T-l Cd «rH 1 1 00 Cd u »H rH r^ ^ OH -o = vO P^ (JN
O <ü -O CJ\ C7^ «—1
>^ >-l 4-» <Ü tH rH
^ a ^1 ^3 • 3 P- o -i-I .« iH
4-) Cd pu 1-1 .• ^ Cd CO tí (U Cd <ü rH
•H ^ Ë o 00 4J CO »-I CJN (U
•H (U 4J o :3 -H rtí ^ á? o CO H œ tí M
T3 •H <u ^^.'-' -v^ -«>>. > rtí
-H :3 CM Ico 1 M Ü o Cd -H 1^ PL) (xq

^^ F. T. COOKE, JR, AND D. W. PARVIN, JR.
Table 6.—Index of prices paid by farmers
Production indexes Cost of production Cost of agricultural
Year (all commodities) chemicals
1961 271 1964 275 269 1965 277 271 1966 289 274 1967 290 277 1968 290 279 1969 302 275 1970 313 270 1971 328 274 1972 351 284 1973 424 292 1974 481 329 1975 528 443 1976 559 481 1977 579 434 1978 626 407 1979 711 495
Source: U.S. Department of Agriculture 1979a.
cotton insecticides is available. Of those that are available, the most helpful would seem to be the index of prices paid by farmers for agricultural chemicals (Table 6). However, this index declined for the periods 1968-71 and 1977-78, and the cost of cotton insec- ticides did not. Therefore, the index of prices paid by farmers for all production inputs, which reflects a consistent increase in the cost of all production costs to farmers for 1961-79 (Table 6), was used to adjust reported costs to 1979 constant dollars.
DISCUSSION OF SURVEYS
The 1961 Cotton Insecticide Survey
The 1961 cotton insecticide study (Langsford 1964) was con- ducted in areas where the boll weevil was known to be a severe problem. It would not be appropriate to expand the results of this survey to the rest of the Cotton Belt. However, the results are included because the sample size is relatively large and because the survey provides some information on approximately one-third of the cotton acreage in the United States.

COST OF INSECTICIDES USED ON COTTON 37
The 1964, 1966, 1971, and 1976 National Pesticide Use Surveys
The national surveys of pesticide use of 1964 (Eichers et al. 1968), 1966 (Eichers et al. 1970), 1971 (Andrilenas 1974), and 1976 (Eichers et al. 1978) were made to arrive at an estimate of the total use of pesticides in the United States. The production re- gions used in these surveys (Figure 1) follow State lines and were extremely large. However, the cost of controlling cotton insects differs considerably from north to south in such large areas, es- pecially in the Delta States and the Southern Plains Regions. Ad- ditionally, the number of cotton producers sampled was not adequate for providing regional estimates. Certainly the information is "too thin" to be disaggregated into State or sub-State regions.
The 1969, 1972, and 1974 Cotton Cost of Production Surveys
The cotton cost of production surveys of 1969 (Starbird 1974), 1972 (Krenz et al. 1976) and 1974 (U.S. Department of Agriculture 1979b) were designed to determine the annual cost of producing cot- ton. The production regions used in these surveys (Figure 2) were smaller than those used in the national pesticide use surveys and were not aligned with State lines, but Southeast Regions 1 and 2 and South Central Region 1 were still too large for our purposes, i.e., to obtain average costs of cotton insect control within re- gions, especially the Mississippi Delta Region, which includes the bootheel of Missouri; the Mississippi River Delta of Arkansas, Louisiana, and Mississippi; and the Red River Valley of Louisiana.
The 1977 Cotton Pesticide Use Survey
The 1977 cotton pesticide use survey was designed to assist with the evaluation of alternative cotton insect control strate- gies, commonly referred to as the boll weevil evaluation. The production regions used in this survey (Figure 3) involved still another group of geographical regions, whereby cotton producing regions were effectively separated according to pesticide use pat- terns, with the exception of Regions 1 and 3. (These regions are too large, and the insect control costs vary widely within these regions. Consequently, the estimated costs do not accurately re- flect differences that exist within the regions.) Also, in this survey, Region 2 was not sampled, and minor cotton production re- gions (especially in Texas) were not included.
Normalized Estimates of Insecticide Use
The survey on normalized estimates of insecticide use (Cooke and Parvin 1981a), like the 1977 cotton pesticide use survey, was designed to assist with the evaluation of alternative cotton insect control strategies. Thus, the Cotton Belt was divided into six major production regions (Figure 4), and each of the six major

38 F. T. COOKE, JR, AND D. W. PARVIN, JR.
0^
Í5
U to 0)
c
> u CO
<U CO
(U 'O •H
(U
CO 3
c o
•H tvO (U
c o
•H 4J O
x) o v^
Pui I I
0)
00
COST OF INSECTICIDES USED ON COTTON 39
lis- 9-= B O - S E 5 « c w © 9
*-' • E i5 iS 2 — Ü 0Û CÛ
Q
Q- \
(^ m CO.
T-^ cvi CO ^ lo CD r^ 0)
<û.
CO
CJ> vO ON
0) >
CO
o
O •r-(
a;
tí o
ü
O U
I
0) u a
•H

40 F. T. COOKE, JR., AND D. W. PARVIN, JR.
> u :3
CO
•H O
<U
o •H ÖO 0) v^ c o
•H
o
o
'^ I

COST OF INSECTICIDES USED ON COTTON 41
o
CO C O
•H to 0) U
c o
3
O U
I I

42 F. T. COOKE, JR., AND D. W. PARVIN, JR.
Table 7.—Estimated harvested acres, number of harvested acre, selected years, by Delphi
1969 L972
Materials Materials Cotton 11 Har- cost Har- cost per produc- vested per acre vested acre
Delphi tion acres Number (1979 acres Number (1979 regior L sub- (x appli- constant (x appli- constant
1/ region 1,000) cations dollars) 1,000) cations dollars)
A SE-1 150 4.0 16.54 SE-2 492 7.0 19.27 540 13.0 43.30 SE-3 199 10.0 31.89 200 15.0 57.49
Total or average 841 7.2 21.77 740 13.5 47.13
B SE-7 530 4.0 11.02 713 5.0 12.73 SC-2 379 3.0 9.17 513 4.0 9.36
Total or average 909 3.6 10.25 1,226 4.6 11.32
C SE-4 296 4.0 14.22 365 5.0 19.43 SE-5 226 8.0 20.19 259 7.0 16.69 SE-6 148 6.0 20.02 SC-1 1,787 4.0 18.14 2,659 7.0 22.49
Total or average 2,457 4.5 17.97 3,283 6.8 21.69
D SC-3 750 2.0 6.30 744 2.0 6.15 SC-4 329 2.0 6.42 286 5.0 21.96 SC-5 290 6.0 18.33 299 9.0 37.11
Total or average 1,369 2.8 8.88 1,329 4.2 16.52
E SC-6 1,309 5.0 18.45 1,450 3.0 11.49 SC-7 1,920 1.0 4.51 2,357 2.0 10.62 W-6
or average 100
3,329 3.0 2.6
17.16 10.37 2.4 Total 3,807 10.95
F W-1 599 2.0 11.07 764 2.0 20.06 W-2 93 7.0 33.46 75 9.0 77.95 W-3 204 4.0 24.96 201 9.0 63.05 W-4 36 2.0 12.20 W-5 61 7.0 27.99 78 3.0 17.50
Total or average 993 3.2 17.10 1,118 3.8 31.49
j_/ See Figure 4. Ij See Figure 2. _3/ 5-year average, 1975-79.
Source: 1969, 1972, and 1974—Cooke and Parvin 1981b; 1977— Cooke and Parvin 1981a.

COST OF INSECTICIDES USED ON COTTON 43
applications per treated acre, and materials cost per region and cotton production subregion.
1974 1977 Normalij sed
Materia Is Materials Materials
cost pe r cost pe !r < :ost per
acre acre acre
Har- (1979 Har- (1979 Har- (1979
vested Number con- vested Number con- vested con-
acres appli- stant acres appli" - stant acres 3/ Number stant
(x ca- dol- (x ca- dol- (x appli- dol-
1,000) tions lars) 1,000) tions lars) 1,000) cations lars)
50 6.5 38.32
460 14.0 51.58 228 18.9 64.08 229 12.7 66.38
191 14.0 67.77 206 12.5 39.86 105 13.6 71.10
651 14.0 56.35 434 15.9 52.58 384 12.5 64.02
791 7.0 13.94 292 0.9 2.69 512 6.2 23.73
444 5.0 16.95 818 2.2 6.83 659 2.0 5.57
1,235 6.3 15.02 1,110 1.8 5.74 1,171 3.7 13.51
350 8.0 19.95 229 5.5 18.08 115 5.5 17.03
236 8.0 22.84 207 12.5 39.86 174 11.5 54.13
207 12.5 39.86 88 7.9 24.42
2,401 7.0 24.54 1,692 8.5 24.77 1,826 8.9 41.98
2,987 7.2 23.87 2,335 8.9 26.79 2,203 9.0 40.94
680 4.0 9.09 560 3.7 7.80 429 2.2 2.40
205 4.0 19.98 685 6.7 20.90 194 5.5 17.03
316 10.0 5.6
43.32 19.96 5.4 15.01
273 896
5.3 3.8
20.17
1,201 1,245 10.98
1,440 5.0 13.14 1,797 0.6 2.26 1,511 1.4 3.61
2,379 2.0 5.18 3,496 0.3 .87 2,764 1.0 6.65
3.1 8.18 0.4 1.34 54
4,329 2.3 1.1
9.89
3,819 5,293 5.81
1,082 2.0 12.21 1,212 1.8 7.05
131 7.0 41.11 641 9.4 33.70 286 7.0 37.74
80 1.1 2.55
3.4 19.61 81
2,014 1.1 4.1
2.55 15.17 1,499
Cooke and Parvin 1981a; and Lin et al. [in review]; normalized-

^^ F. T. COOKE, JR, AND D. W. PARVIN, JR.
production regions was divided into several subregions with similar insect control practices so as to facilitate collection of esti- mates of insecticide use and cost from entomological experts within the subregions. The entomological experts were interviewed, the estimates were summarized, and the summaries were submitted to two to four rounds of review by the experts. This approach to collect- ing expert opinion is a modification of the Delphi method (Linstone and Turoff 1975), and the six regions (A-F in Figure 4) are re- ferred to as Delphi regions. Region F was excluded from this study because boll weevils are not a threatening pest there. The experts in this inquiry consisted chiefly of one Extension entomologist per State in 11 weevil-infested States. A more comprehensive Delphi inquiry was conducted in 1980 and 1981 and reported elsewhere (Eco- nomic Research Service, U.S. Department of Agriculture, 1981).
INTERPRETATION OF SURVEY RESULTS
The number of insecticide applications used on cotton in any given year provides some insight as to the relative severity and duration of infestations of insect pests. It may also provide some insight into the effectiveness of insecticides against the target pests. A change in the number of treatments over time may reflect a change in insect management strategies or improved technology.
In Table 7, the normalized estimates are compared with the previously discussed estimates of insecticide use and costs for 1969, 1972, 1974, and 1977. Plainly, the number of insecticide ap- plications per acre in Delphi Region A ranged from 7.2 in 1969 to 15.9 in 1977, but the estimated cost per acre ranged from $21.77 in 1969 to $56.35 in 1974. The normalized estimate of the number of insecticide applications (12.5) is thus within the range observed. However, the normalized material cost of $64.02 is greater than any value produced in any of the surveys. (This probably reflects an increasing availability and acceptance of the more expensive syn- thetic pyrethroid insecticides.) Region A had the highest per acre cost of insecticide materials of any of the regions, but between 1969 and 1977, cotton acreage decreased from 841,000 to 434,000 acres.
In Delphi Region B, the average number of insecticide applica- tions was 1.8 per acre in 1977 and 6.3 in 1974. Average cost of insecticides ranged from $5.74 to $15.02 during these years. Thus the normalized estimates of 3.7 applications and $13.51 material cost are within the range obtained in the other surveys.
In Delphi Region C, the average number of insecticide treat- ments ranged from 4.5 in 1969 to 8.9 in 1977. The average cost for insecticides ranged from $17.97 in 1969 to $26.79 in 1977. The number of cotton acres declined from 3,282,000 in 1972 to 2,335,000 in 1977. The normalized estimates for the number of applications reflect this trend. However, the number of applications and the increased use of synthetic pyrethroid insecticides put the normal- ized estimate of material cost outside the range.

COST OF INSECTICIDES USED ON COTTON 45
In Delphi Region D, the average number of insecticide treat- ments ranged from 2.8 to 5.6 between 1969 and 1974. Insecticide costs ranged from $8.88 per acre (1969) to $19.96 per acre (1974). Cotton acreage declined from 1,369,000 acres in 1969 to 1,201,000 acres in 1974. The normalized estimates of the number of applica- tions and costs are consistent with these values.
In Delphi Region E, the number of insecticide applications ranged from 0.4 per acre (1977) to 3.1 per acre (1974). Per acre insecticide costs ranged from $1.34 to $10.95. Cotton acreage in- creased from 3,329,000 acres in 1969 to 5,293,000 in 1977. Region E had the lowest cost of insecticides of any of the regions.
Cost of Insecticides (Materials)
The cost of insecticides for the United States and for cotton producing States obtained from surveys made to determine the cost of producing cotton and from the Farm Enterprise Data System is reported in Tables 8 and 9, respectively. According to Table 8, insecticide costs have remained fairly constant from 1964 to 1980, increasing from $14.74 to $21.90 (1979 constant dollars). Accord- ing to Table 9, States with the highest insecticide cost per acre in the boll weevil infested part of the Cotton Belt were South Car- olina, Georgia, Alabama, and North Carolina. These States with the highest cost per acre are located in the southeastern United States. Louisiana, Mississippi, and Tennessee had the next highest insecticide cost. Texas and Oklahoma had the lowest insecticide cost of any of the boll weevil infested States.
Table 8.—U.S. cost of insecticide materials per harvested acre
1979 constant Year Dollars dollars
1964 5.69 14.74 1966 6.42 15.79 1969 6.79 15.96 1971 4.66 10.11 1972 7.35 14.92 1974 12.35 18.28 1976 15.83 20.10 1977 24.68 30.36 1978 21.49 24.50 1979 21.90 21.90 1980 25.31
Source: U.S. Department of Agriculture 1979b, Krenz et al. 1976, Starbird 1974.

46 F. T. COOKE, JR., AND D. W. PARVIN, JR.
Table 9.—Average insecticide material cost per harvested acre by States (1979 constant dollars)
State 1977 1978 1979
Alabama 82.82 72.00 Georgia 84.66 73.61 North Carolina 81.88 71.19 South Carolina 88.45 76.90 Arkansas 27.79 24.16 Louisiana 65.12 56.61 Mississippi 57.29 49.82 Missouri 19.56 17.01 Tennessee 40.53 35.24 New Mexico 4.59 3.99 Oklahoma 7.05 8.39 Texas 5.17 4.35 Arizona 139.06 120.91 California 44.96 39.09
64.42 65.86 63.70 68.81 21.61 50.65 44.57 15.22 31.53 3.57 7.37 3.80
108.18 34.98
Source: Krenz et al. 1976.
Cost of Applying Insecticides
The cost of applying insecticides estimated for Delphi regions 1-33 (Figure 4) is shown in Table 10. These costs ranged from $0.94 to $2.25 per acre.
Quantities of Insecticides Applied to Cotton
Relative quantities and per acre treatments of insecticides used on cotton are summarized in Tables 11, 12, and 13. It is clear from Table 11 that more insecticide is used in the Southeast and Delta States and that the decline in quantity used in the Southern Plains is substantial. Data on per acre treatments of specific insecticides are reported in Table 12. Methyl parathion, EPN, and toxaphene were the most extensively used materials. Data on pounds of specific insecticides used on cotton are reported in Table 13; for example, subsequent to cancellation of the registra- tion of DDT on December 31, 1972, the use of methyl parathion and toxaphene increased sharply through 1974. However, by 1976 and 1977, the use of methyl parathion and toxaphene had declined, and increased usage of EPN, monocrotophos, and methorny1 was observed in 1977. Also, in 1977, an experimental use permit allowed use of new synthetic pyrethroid insecticides (permethrin and fenvalerate) for control of Heliothis spp. Subsequently, long-term normalized esti- mates indicated a major shift to the synthetic pyrethroid insecti- cides (permethrin, fenvalerate) for control of bollworms (Heliothis zea (Boddie)) and tobacco budworms (li. virescens (F.)). They also

COST OF INSECTICIDES USED ON COTTON 47
Table 10.—Range in cost of one application per acre by Delphi subregion
Application cost per acre
Region 1/ (1979 constant dollars)
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18
19 20 21 22 23 24 25 26 27
28 29 30 31 32 33 34 35
1 .50 1 .50 1 .25 1 .71
1. 50 ■ - 1. 58 1 .71 1 .71 1 .80 1 .80
37 - 1. 45 25 - 1. 74 ,37 - 2. 25 .37 - 2. 25 ,75 - 2. 00 .25 - 1. 50 .25 - 1. 50 .10 ~ 1. ,75 .25 - 1. ,50
.27 - 1, .33
.25 - 1. .31
.81 - 2, .00
.81 - 2, .00
.50 - 2, .00
.57 - 2 .25 2 .00 - 2 .25 1 .70 - 2 .00 2 .00 - 2 .25
0 .94 - 1 .50 1 .75 - 2 .00
2 i.25 2 '.00 2 L25 ¿ L25 2/ 2/
\J See Figure 4. 1) Application costs in Regions 34 and 35 were not reported sep-
arately. There aerial applicators charge for materials and appli- cation.
Source: Cooke and Parvin 1981a.
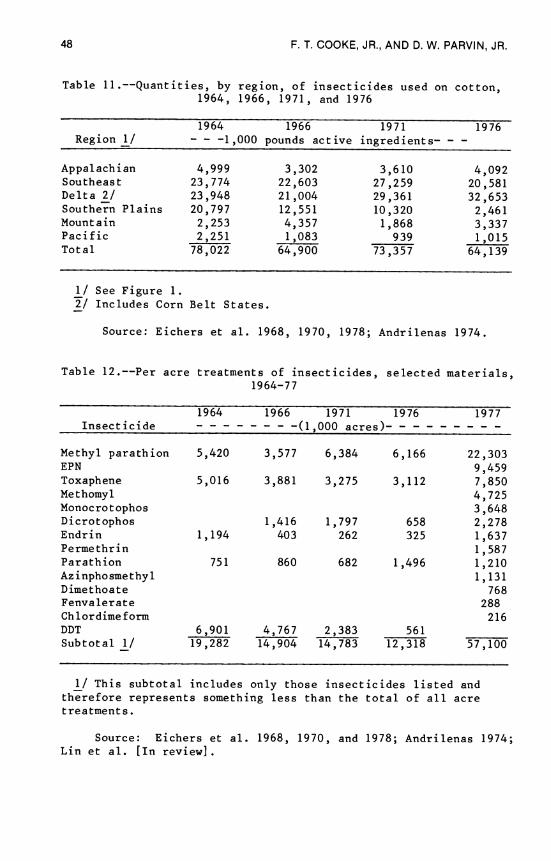
48 F. T. COOKE, JR., AND D. W. PARVIN, JR.
Table 11.—Quantities, by region, of insecticides used on cotton, 1964, 1966, 1971, and 1976
1964 1966 1971 1976 Region y 1,000 pounds active ingredients- - - -
Appalachian 4,999 3,302 3,610 4,092 Southeast l^.llk 22,603 27,259 20,581 Delta 2/ 23,948 21,004 29,361 32,653 Southern Plains 20,797 12,551 10,320 2,461 Mountain 2,253 4,357 1,868 3,337 Pacific 2,251 1,083 939 1,015 Total 78,022 64,900 73,357 64,139
XJ See Figure 1. y Includes Corn Belt States.
Source: Eichers et al. 1968, 1970, 1978; Andrilenas 1974.
Table 12.—Per acre treatments of insecticides, selected materials, 1964-77
1964 1966 1971 1976 1977 Insecticide _ - _ -^1 ,000 acres ) - - - -vi — — — —
Methyl parathion 5,420 3,577 6,384 6,166 22,303 EPN 9,459 Toxaphene 5,016 3,881 3,275 3,112 7,850 Methomyl 4,725 Monocrotophos 3,648 Dicrotophos 1,416 1,797 658 2,278 Endrin 1,194 403 lei 325 1,637 Permethrin 1,587 Parathion 751 860 682 1,496 1,210 Azinphosmethyl 1,131 Dimethoate 768 Fenvalerate 288 Chiordimeform 216 DDT 6,901
19,282 14,904 2,383 14,783
561 12,318 Subtotal \J 57,100
\J This subtotal includes only those insecticides listed and therefore represents something less than the total of all acre treatments.
Source: Eichers et al. 1968, 1970, and 1978; Andrilenas 1974; Lin et al. [in review].

COST OF INSECTICIDES USED ON COTTON 49
I
T3 0)
•p-i vO ■U vD C 0^ CO P
1 1
CO ,—1 -it
v£) <u ON
i—t
rû CO H
r>. CO ON 'd
D O a o o
r^ o ON «
00 vO vO <í vO CO ^ <D m en <ï CO CO •—t lO f^ lO ON
00 ^ CM ON ON CO CO CM r-l <j- ON CO ON <t CO vD
ooco«—»CO»—t-HONoooNcomoNvo CM—HON00<l-vDr^C0O00Cv|CMr-H li^CSI^<l-Or^vOCMONCOCM CM
in in .—I .-4 CM
•—I o ON 00 <i" 00 ON —I cs
f-H 1—( o m f-H 00 CM CO VO
f-H
o ON 00 ON in
in O ON
CM CM ON CJN CO vD
in CO
<t o CO ^ CO 00
in C7N in
o o 00
00 CM 00 »-• C3N ^
o m 00 vO CM r-H CM o 00 CJN
•» CM <i-
^ CO
00 o 00 VO VO 1^ o in
CO
r>. o in ^ 00 liO
o in in VO 00 <1- VD i—i vO CO 00 vO 1^ <JN 00 vO in rv.
oo" vO CM
i-H i-H CO CM
CM VO
c o
• H T-H
^ CO >. g 4J o J= }u
CO rC CO iJ (U o -v^
u Cu o c Q) Q) 4-í U-4 1—1
CO Q) O rC • p-i c B 4-t CO d) et c i-H 4-1 p- u o CO CO VJ B p-t
<U >^ O o X •H o á <u •H CO r-H ^ U JJ c >u JC ^ r-H TJ 4-»
>% Qu O o o •H (U 4-> O- 4-) CO »H O
X CO JO o »H ÍH s CO C (U > o 4-1 4J ¡S >< ■u c o T3 C ÍH •H g c i-H H ,0 <D P^ O (U o •r-l C (u CO N •H (U ^ Q 3 S H H S s Q w fi^ PL. <3 Q ptH u Q œ
O)
CO
0) CO <U )H P- <u »H
CO
O) CO OJ
42
na
CO
13
CO
T3
O)
c •H
00 CJN
> }H CO
O,
c CO
O)
o
<t
>H
<
CO 'O C c
CO
<u » CO • O o CO r^ xí <u C3N 4-» T3 1—(
•H >«. O *«
r-H .pJ 00 C 4J VO O O ON
<D CO CO 4J C • C 'H i-H
dJ CO CO (U (U jC 4-i >H 4-t QJ (X <]) rH CO »H rH U
CO <v i-H JO
CO MH o 4-» O •H O w 4-1 (U XI öß :3 CO >* • CO T3 o; r—1
C o > CO 3 u o;
•H O 0 • H x: a o > H CO <u

50 F. T. COOKE, JR., AND D. W. PARVIN, JR.
indicated that in the future, reduced amounts of methyl parathion, toxaphene, methomyl, and monocrotophos will be used for cotton in- sect control. For example, recommended dosages of the methyl para- thion plus toxaphene combination commonly used for bolIworm-budworm control are 1.0 and 2.0 pounds active ingredients (a.i.), respec- tively, per acre per application, but the recommended dosages of the synthetic pyrethroid insecticides range from 0.1 to 0.2 pound a.i. per acre per application. The shift to the synthetic pyre- throid insecticides will therefore result in a significantly smaller total number of pounds of insecticide being applied to cotton.

COST OF INSECTICIDES USED ON COTTON 51
LITERATURE CITED
Andrilenas, P. A. 1974. Farmers' use of pesticides in 1971—quantities. U.S. De-
partment of Agriculture, Economic Research Service, Agricul- tural Economic Report No. 252, 56 p.
Anonymous. 1975. World pesticide markets. Farm Chemicals 138:45-48.
1977. A look at world pesticide markets. Farm Chemicals 140: 38-43.
1979. A look at world pesticide markets. Farm Chemicals 142: 61-68.
Cooke, F. T., Jr., and D. W. Parvin, Jr. 1981a. The cost of controlling cotton insects in the boll weevil
infested States, Agricultural Economics Technical Report No. 28, Department of Agricultural Economics, Mississippi State University.
and D. W. Parvin, Jr. 1981b. Insecticides used to produce cotton in selected major production regions in the United States, 1969, 1972, and 1974. Economics and Statistics Service, U.S. Department of Agricul- ture, Economics and Statistics Staff Report No. AGESS810331, April 1981.
Eichers, T. R., P. A. Andrilenas, and T. W. Anderson. 1978. Farmers' use of pesticides in 1976. U.S. Department of Agriculture, Economics, Statistics, and Cooperative Service, Agricultural Economic Report No. 418, 58 p.
P. Andrilenas, H. Blake, R. Jenkins, and A. Fox. 1970. Quantities of pesticides used by farmers in 1966. U.S. Department of Agriculture, Economic Research Service, Agricul- tural Economic Report No. 179, 61 p.
P. Andrilenas, R. Jenkins, and A. Fox. 1968. Quantities of pesticides used by farmers in 1964. U.S. Department of Agriculture, Economic Research Servicie, Agricul- tural Economic Report No. 131, 37 p.
Keller, K. R. 1980. Status of the boll weevil eradication and optimum pest management trials. Tn Proceedings, 1980, Beltwide Cotton Production-Mechanization Conference (St. Louis, January 9-10, 1980), p. 29-34.

52 F. T. COOKE, JR., AND D. W. PARVIN, JR.
Krenz, R., C. Michael, G. Garst, D. Fawcett, and S. Rogers. 1976. Cost of producing food grains, feed grains, oil seeds, and
cotton, selected years. U.S. Department of Agriculture, Eco- nomic Research Service, Agricultural Economic Report No. 338,
40 p.
Langs ford, E. L. ^ 1964. Extent and cost of using chemicals in cotton production in
selected areas, 1961. U.S. Department of Agriculture, Economic Research Service, Farm Production Economics Division, ERS-155,
15 p.
Lin, Y. N., D. F. Caillavet, and D. W. Parvin, Jr. Estimation of insecticide, miticide, and nematicide applications in the Cotton Belt, 1977. Mississippi Agriculture and Forestry Experiment Station. [in review.]
Linstone, H. A., and M. Turoff, editors. 1975. The Delphi method: techniques and applications. Addison- Wesley Publishing Company, Reading, MA.
Starbird, I. R. 1974. Costs of producing upland cotton (selected years). U.S. Department of Agriculture, CS-265.
U.S. Department of Agriculture. 1979a. Agricultural Statistics 1979, p. 453-454.
1979b. Costs of producing selected crops in the United States selected years. Prepared for the Committee on Agriculture, Nutrition and Forestry, United States Senate.
1981. Economic Research Service. The Delphi: insecticide use and lint yields, beltwide boll weevil/cotton insect management programs, overall evaluation. Appendix E, ERS Staff Report No. AGESS810507.

53
Chapter 3
ECOLOGY OF COTTON INSECTS WITH SPECIAL REFERENCE TO THE BOLL WEEVIL
W. H. Cross Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
ABSTRACT Study of the biology, behavior, and ecology of the boll weevil (Anthonomus grandis Boheman) reveals that this insect can easily adapt to new environments. However, there are critical weak points in its life cycle, and these provide opportunities for effective management of this pest.
For example, alternate host plants act as reservoirs for native cotton insect pests, but the boll weevil, as a relatively recent intro- duced pest, largely lacks alternate host plants in the United States.
In the Boll Weevil Eradication and Optimum Pest Management Trials, more than 50 miles of cotton rows were sampled in 1978-79 for infest- ing arthropods. These (more than 1 million) were identified by group and converted to per acre estimates to obtain a better understanding of the trophic structure of the cotton field arthropod community.
A case history of a Mississippi cotton field demonstrates how poor cotton insect management can increase pest problems and increase produc- tion costs.
INTRODUCTION
The cotton field is a dynamic community with a physical envi- ronment, a crop phenology, and a structured animal population that is much influenced by the presence of infield "weeds" and even more by the array of surrounding crops and native vegetation. These an- imal populations are also influenced by sequences of weather (vary- ing annually) and by interplay among the species. This interplay of animal populations is often greatly simplified by using the categories "pests" and "beneficiáis."

54 W. H. CROSS
BOLL WEEVIL BIOLOGY, BEHAVIOR, AND ECOLOGY
The boll weevil (Anthonomus grandis Boheman) is a rather re- cent introduction from a tropical environment. (Only about 280 generations have developed in the United States.) Unlike native species, which tend to have many hosts, it survives in this country on cotton except for limited populations found on Cienfuegosia and Thespesia in southern Texas (Cross et al. 1975). Even in the Trop- ics, the boll weevil has a limited host range. For many years, cotton (Gossypium hirsutum L. and £. barbadense L.) was the only host known, but more recently, the weevil was found to reproduce on several other species of Gossypium and on several species in at least four other genera of the Malvaceae (Thespesia, Cienfuegosia, Hampea, and Hibiscus (one species)), especially those from the more southern parts of the range of the insect—Mexico, Central America, Colombia, and Venezuela (Cross et al. 1975). Thus, small popula- tions are maintained in areas remote from cultivated cotton.
In fact, relatively few of the overwintering generation sur- vive the winter in temperate climates. An average 10% is normal, and the rate is much lower after colder winters. Although many fe- male boll weevils overwinter with viable sperm in their spermathe- cae and can thus lay fertile eggs without having to find a male in the spring, much cotton in fringe areas becomes infested only after reimmigration of adults (Figure 1). Thus, this pest at times main- tains a tenuous existence in the United States.
Literature on the boll weevil has been summarized by Dunn (1964), Mitlin and Mitlin (1968), and Cross (1973). Basic studies of behavior of the boll weevil reported subsequently have been re- ported by Cross and Mitchell (1966), Lloyd et al. (1967), Cross et al. (1969), Hardee et al. (1969a, 1969b), Mitchell and Cross (1969, 1971), Walker and Bottrell (1970), and Roach et al. (1971).
The boll weevil is a typical snout beetle in the Curculioni- nae, the largest subfamily of the Curculionidae and one that in- cludes many pests of crops and trees. The genus Anthonomus con- tains 113 known species in America north of Mexico (H. R. Burke personal communication). This phytophagous group demonstrates a high degree of host specificity and has a general preference for feeding and reproducing in flower buds.
General Annual Cycle
Boll weevils overwinter as adults in forest and hedgerow lit- ter, usually near cotton fields. They return to cotton fields in the spring where they feed on the terminals of seedling plants and then feed on and reproduce in squares (flower buds). Later in the season, they attack and reproduce in bolls (fruit). Eggs are de- posited singly at the bottom of punctures made with the slender rostrum of the female. The larvae feed for 7-14 days in either the squares (which abscise and normally fall to the ground 3-5 days
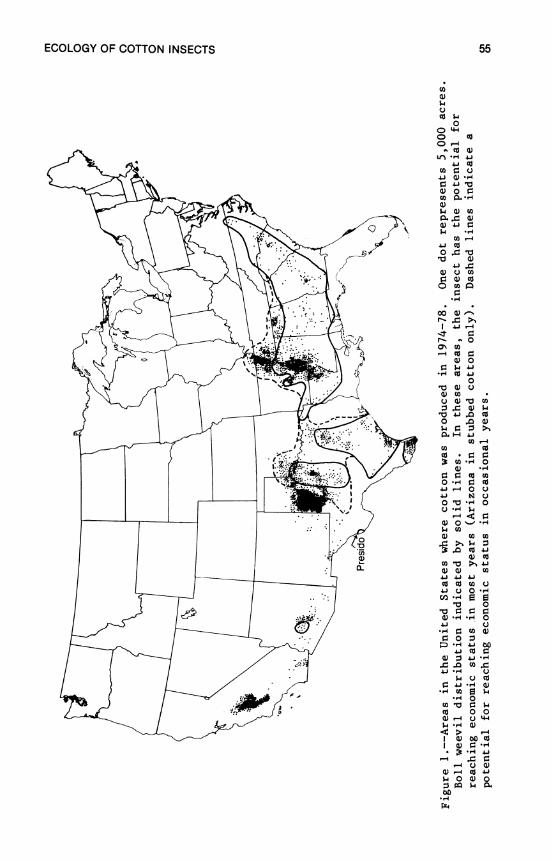
ECOLOGY OF COTTON INSECTS 55
CO d) u u CO t-i
o o M-i o CO o r-l
^ CO <U in -H 4J
•M CO CO a CJ JJ Q) «H
C -M XJ <U O C CO a«H Q) U Q) Oi ax <ü Q) U 0 U -H
CO 1-1 U CO O jC TJ
TJ 0) ^ X
Q) o CO a 0) CO o CO Q
Ö •H
00 0) -^ r^ X >^
1 4J ^ <f c r^ •» o ON CO ^ CO C
Q) o a M -p
•H CO ^ o
13 QJ o (U CO • o 0) TS CO 3 X <ü M
-V U Xi CO O XI 0) M tí 3 >. Oi M 4J
CO r-< CO CO CO . tí tí ¡^ CO «H O
(U •H tí tí CO CO o -H tí CO *J .—1 O O 4-1 N O o T> «H O O 'r^ U ^ < tí <U O ^ •H U CO (U CO CO
^ X CO tí
4-1 (U CO
CO 73 >% JJ <U Oí CO 4J 4-) 4J CO CO (0 o *-> O O •H CO «H e g
T) O Tí tí tí tí (ü «H «H o 4J o •H tí CO <ü tí O 3
|Z> «H 4J 60 4-í CO tí
0) 3 4J •H X J3 CO xi 4J .H O
V4 O CO tí 4-» .H <u
*H CO g Ul •H O
CO 73 tí u CO O o <U .-1 O **^ V^ «H <ü
<í > TH
1 0) M) CO 1 (U tí •H . > .H 4-1
-H 43 tí .-< O O)
0) rH CO 4J M O <U O tí PQ J-i o. bO

56 W. H. CROSS
after egg hatch) or the bolls and pupate; then after 3-5 days, the adults emerge through a hole cut to the outside. Variations in this cycle depend on average temperatures. Females of the new gen- eration begin laying eggs after 3-4 days. As many as five genera- tions may develop each season.
Emerging Overwintered Population
Although small numbers of overwintering boll weevils may move about during the warmer periods that occur throughout the winter, the maximum exodus from forest litter occurs from late April to early June. Thus, in areas where cotton is usually planted between May 1 and 15, weevils may be captured on sticky flight screens or on pheromone-baited traps several weeks before the cotton seedlings emerge. (Peak numbers may emerge following rains, especially after extended dry periods.) However, recent observations of native wee- vils marked with Zinc-65 and earlier observations of weevils caged above their hibernation sites with fruiting cotton plants indicate that these early emerging weevils lack interest in cotton for some period of time after they leave the litter. (Dissections of the early-season weevils disclose a considerable reserve of body fat and small gonads, indicating that they are still in diapause (W. H. Cross unpublished data).)
Although sampling of forest litter has revealed that at least small numbers of overwintering weevils are present every month of the year, most have emerged by early June. There is no evidence that these emerging adults feed on another plant before cotton is available. For example, when large numbers trapped May 1 as they entered a field that had just sprouted cotton seed were dissected, none contained any recognizable plant material in their alimentary tracts. Also, some of these weevils lived an average 11.0 days when they were placed in small cages in the field where nothing was available except small droplets of dew in the morning; some held similarly but with a wet cotton ball lived an average 13.6 days; and others caged with fruits and buds of a variety of native plants, including two species of wild Malvaceae, survived ca. 11 days (W. H. Cross unpublished data).
The total number of weevils emerging from hibernation varies greatly by location and year and depends on both the number enter- ing hibernation the previous fall and the rate of survival. On the basis of sampling in forest litter in fall and spring, an average 5-10% survive. Mortality is greatest during winters with sharp freezes when no insulating snow is present, and especially during winters when freezes and intermittent warm periods occur. (in fact, such a sequence can eradicate a population locally.) Survi- val is greatest during mild winters: after such a winter, as many as several hundred weevils per acre of cotton may emerge in areas with a preponderance of small fields and correspondingly more for- est edge. Males consistently represent 60-65% of an emerging

ECOLOGY OF COTTON INSECTS 57
population, and there is some evidence that the first few weevils that emerge are usually males.
The most interesting and valuable studies of behavioral pat- terns in the boll weevil relate to their response to olfactory and visual stimuli. Repeated tests over several years suggest that emerging overwintered weevils (and weevils of other generations as well) cannot recognize cotton at distances greater than perhaps a few inches. However, a chemical arrestant has been isolated from cotton plants (with increasing concentration gradients towards ter- minals and buds) that elicits feeding by the weevil. Also, a dis- tinct plant attractant has been isolated that is attractive to boll weevils when they are held in an olfactometer, but cages with plants are no more attractive to field weevils than cages without plants. Therefore, the first boll weevils seeking cotton in the early season apparently find it only through random movement.
There is likewise no attraction between the sexes or between individual weevils in the unfed emerging population. However, an isolated overwintered male weevil that finds and feeds on cotton squares for several days begins to emit a strong aggregating phero- mone that attracts both sexes from proved distances of 100 feet and perhaps farther when wind conditions are optimum. Indeed, if male weevils feed on cotton terminals or on seedling cotton instead of cotton squares, they produce lesser amounts of the pheromone. Also, if they are fed an apple, banana, or other fruits in the lab- oratory, they elicit a limited response in an olfactometer, but evoke no response in field trapping tests.
The females attracted to the male boll weevil by the aggrega- ting pheromone may or may not be stimulated to mate. However, in the laboratory, when females have been isolated and fed squares for several days, they usually respond to males and mate. In this in- stance (as also with weevils of other generations), the male does not recognize that a female is approaching until she is within less than 1 inch, but at this distance she seems to emit a short-range aphrodisiac that may excite him. If not, the female may stroke him with her antennae or walk over him one or more times until he re- sponds and mates with her (Cross and Mitchell 1966). In a number of observations, mating of overwintered females lasted from 13 to 37 minutes (mean 27 minutes), and multiple matings were observed on the same and subsequent days (Mitchell and Cross 1971). Also, as noted, some overwintered females (as many as 60% or more) mate in the fall before hibernating and so, by retaining viable sperm in their spermathecae, can deposit fertile eggs without a spring mat- ing though fecundity and fertility are reduced. Overwintered fe- males were observed to excavate the puncture they had made for 1-24 minutes (mean 5.2 minutes) before turning 180°and inserting the ovipositor; they then took 0.2-3.2 minutes (mean 1.0 minute) to complete oviposit ion. They laid an average of 0.74 egg per hour during daylight, and none at night (Mitchell and Cross 1969).

58 W. H. CROSS
Overwintered weevils have been reported to live about 30 days after they enter cotton fields, but the period is difficult to de- termine precisely. When large numbers of weevils in forest litter, or just emerging from litter, were marked for identification, con- siderable numbers survived through July and August, and the males had decidedly greater longevity than females. For example, the last recapture of an overwintered, marked female occurred September 14; the last recapture of a similar male occurred October 13. How- ever, when a few of the overwintered weevils collected in late sea- son were dissected, all were highly reproductive, and none showed evidence of diapause development in preparation for a second winter.
Emerging overwintered boll weevils may have to move considera- ble distances before finding cotton. For example, some marked wee- vils re-released in forest litter were recovered in pheromone- baited traps at distances of 1 and 1.7 miles from the release site, and 4.5% of 3380 marked weevils either missed or left nearby cotton and were recaptured in cotton that was 0.06 - 0.6 mile distant (W. H. Cross unpublished data.).
—1 and Subsequent Generations
As a general rule, F adult weevils begin emerging from squares when the flowers appear on the cotton plant (assuming that the overwintered weevils entered the field before or about the time the squares began to form). These adults walk from the fallen square to the cotton plant and immediately eat a hole in a leaf. (This is the only foliar feeding observed except when weevils are deprived of cotton for a number of hours.) A tenfold rate of pop- ulation increase per generation has been considered a reasonable approximation of the normal field situation, but this rate has not been confirmed. The ratio of sexes is about 1:1, and parthenogen- esis probably does not occur. Both sexes of the F, generation are receptive to mating after about 3 days.
Once a moderately large population of weevils is present, the emission of pheromone may not be important to mating success be- cause the newly emerged adults tend to congregate in flowers for early feeding. However, the pheromone emitted by males may produce limited aggregation or clumping since the number of males was higher than normal among 4250 weevils found together in cotton fruiting structures. With small populations, the pheromone is im- portant in assuring mating and also in increasing the extent of range of the female since she flies most often in response to the male pheromone.
A virgin female weevil begins to oviposit after the third to fourth day. She may make a typical puncture in a square, appar- ently in preparation for the egg, and may insert her ovipositor within the puncture, but she always deposits these infertile eggs

ECOLOGY OF COTTON INSECTS 59
on the outside of the square. Indeed, some of these eggs are de- posited at random on the plant.
In cotton fields, boll weevils make occasional short flights that are not a response to pheromone. However, flights are rarely initiated at wind velocities above 3 miles per hour; and at 5 miles per hour or above, flying weevils tend to settle back onto the plants. Weevils generally begin flights by facing into the wind, but the flights are random in direction so far as wind direction or position of the sun is concerned. In flight, the front legs are held high above the head, a position that probably aids the weevil in its habit of landing on the underside of leaves.
In field tests in which 6-day-old or older virgin females were released downwind of pheromone-producing males, the responses to the pheromone were very positive. Typically, a female moved to the top of the plant and often to the edge of the leaf nearest to the emission, faced into the wind with rostrum and antennae raised, and perhaps lifted the elytra a few times. Individual flights were us- ually not more than 3 or 4 feet and often shorter so a female might make as many as 12 flights before reaching the male. However, once she reached the plant containing the male, flights were rare, and her search for him normally involved rapid crawling over the plant. Mating of current-season weevils was observed to last from 12 to 51 minutes (mean 28 minutes). Multiple matings were observed the same and subsequent days, and first oviposit ion after mating often occurred on a square near the mating site and within 4.8-52.0 min- utes (mean 20.9 minutes) of the mating. However, the female seems not to oviposit in squares that are already infested unless most of the squares are infested. These current-season mated females ex- cavated the puncture for 0.8-17.2 minutes (mean 3.8 minutes) before turning 180** and inserting the ovipositor; they then took 0.2-3.2 minutes (mean 0.7 minute) to complete oviposit ion. They laid an average of 0.97 egg per hour. Thus, overall, the F. generation female (and subsequent females) were somewhat more efficient at oviposit ion than the overwintered females observed.
To find uninfested squares, newly mated females move from square to square, move onto an adjacent plant, or occasionally make short flights. Since multiple short flights of greater distances (10-30 feet) are almost always made to a male, the response to the male pheromone tends to increase the total area over which a female oviposits.
Late-Season Population
In late August, which is when the F. generation of boll wee- vils begins to appear, the behavioral patterns of the boll weevil change. By this time, fields in infested areas where insecticides have not been used probably have several thousand adults per acre. Also, at this time, the plants have ceased fruit production because

60 W. H. CROSS
of (1) a heavy feeding pressure by the large population of weevils, (2) the determinate nature of the cultivated varieties, and (3) possibly a deficiency of soil moisture and soil fertility. How- ever, many females are still actively producing eggs and are searching for uninfested squares or young bolls for oviposition sites. Therefore, large numbers of weevils migrate, apparently in search of uninfested cotton. As a result, in some cotton producing areas such as west Texas, isolated fields that do not have suitable adjacent litter or moisture for overwinter survival may become in- fested late in the season by weevils migrating as much as 50 miles, and migrating weevils have been captured in traps at considerable distances from cotton. Examination of 98, 309, and 1961 adult wee- vils collected on cotton in three fields newly infested by such migrating populations indicated that 63% were females (so more fe- males than males may migrate) and that the migrating weevils were mature and mostly reproductive adults (W. H. Cross unpublished data).
Also, beginning in late August (on the average), a segment of the population of boll weevils begins to develop the physiological and morphological conditions of diapause. The percentage increases in September, but it then decreases in October because the older weevils have moved from the field to litter and new adults are con- tinuing to emerge. (in the field, some percentage of older adults are highly reproductive as late as the first frost, probably a ge- netic mechanism for survival in the event of a change in climate.) This diapause condition in the boll weevil may be associated with boll feeding, scarcity of food, exposure of adults to cool night temperatures, or exposure of larvae and pupae to short days. The importance of these several factors seems to vary in different parts of the range of the weevil.
Weevils in firm diapause do not take part in long-range migra- tion. Rather, they move relatively short distances to suitable hi- bernation sites. Also, many weevils not in firm diapause seek pro- tection in litter with the onset of cool weather and may survive there through part of the winter though they all probably die, as do many in firm diapause, even when winter conditions are average. Both migratory and diapausing populations are extremely responsive to the male pheromone, even within forest environments.
Weevils involved in long-range dispersal, especially in the western parts of their range, are often transported passively by thermal currents (Fye 1968), but they do have some capability of sustained flight as indicated by flight mill studies of tethered weevils (W. H. Cross unpublished data). These males and females flew an average 1.32 and 1.43 miles, respectively, in the initial sustained flight and an average total of 2 and 2.2 miles to exhaus- tion. One female flew 11 miles before exhaustion. Average rate of flight was 180-200 feet per minute. Greater activity was observed at 29.5*^0 than at either 26.7°C or 32.3*^0. High-speed movies indi- cated the rate of wing beats at about 55 per second.

ECOLOGY OF COTTON INSECTS 61
Late-season boll weevils and early-season weevils respond ac- tively to pheromone-baited traps; midseason weevils respond much less. The greatest number of weevils respond when the traps are coated with daylight fluorescent paints that appear lemon yellow to our eyes but peak near the 525-nanometer (blue-green) range of the spectrum. Other regions of the spectrum are progressively less at- tractive; red is apparently not attractive.
THE COTTON FIELD ANIMAL COMMUNITY
Cotton fields are not, however, inhabited entirely, or even mainly, by boll weevils. A whole community of animals is present; thus, the ecology is dynamic. It changes because of or is changed by the interplay between species of animals searching for food as much as by the developing crop, the sequence of weather, and the presence of competing "weeds" and other vegetation.
Some idea of the makeup of the community can be obtained from Table 1. This table reflects some of the information obtained by two teams of entomologists \J during the cotton growing seasons of 1978 and 1979 on populations of arthropods in many of the cotton fields in North Carolina-Virginia and in Mississippi that were in- cluded in the Boll Weevil Eradication Trial and the Optimum Pest Management Trial. (The total collection is probably among the largest ever gathered from a single crop.) In these 2 years, 7066 D-Vac samples of 40-row feet per field, a total of 52.65 miles of cotton row (about 8.4 hectares), were taken at intervals designed to reflect all growth and development stages of cotton. All 1,037,067 arthropods collected in this sampling were segregated taxonomically by life stage and by trophic level by the two teams. See Chapter 12 of this handbook for a complete description of the sampling, for the technique employed in converting the counts of arthropods captured by D-Vac to a per area basis, and for the D-Vac-unit area ratios used to correct the numbers in Table 1.
The more than 1 million arthropods captured in the 2-year sampling are listed by major groups in Table 1 and show corrected number per acre and number per row-foot. These values represent the number of animals (arthropods) present and thus available to the diurnally operated D-Vac. Other animals known to inhabit cot- ton fields that were either poorly or not sampled by this method include those present on the cotton plants only nocturnally, those that occur below ground or on the soil surface, or those that are extremely active and so consistently escape the sampler. Among
\J Biological Evaluation Research Teams: one at the U.S. Department of Agriculture Boll Weevil Eradication Laboratory at Raleigh, NC—E. P. Lloyd, W. A. Dickerson, G. H. McKibben, T. J. Bradway, and D. C. Pierce; and one at the U.S. Department of Agri- culture Bioenvironmental Laboratory at Stoneville, MS—C. R. Parencia, J. W. Smith, W. P. Scott, and T. C. Lockley.

62 W. H. CROSS
I u
I—I
(U
CO o> •H F-1
s na
a (0 CO
00 CO r^
•H ON
00 M
•H > I CO c
o >
M r-l o CO
¡z; ö u
•H «H
CO
O Jd a, o TJ
6
Pu
O
ÖO
O •»-i
CO
f
CO H
-o (U
i o o < i
o CO
è< O
u 0)
d) 3
O
U u o
o 00 u CO
. u u
CO
u
O) o
O
>
I tí Cu
O U O
00 o> vO CO
lo o m vo o in CO <^
r^ vo rH <í- o 00 c>i -^ in m
coococovo mON^fcM cvimr^oooo vocsooco
CO vx> 00 LO VO t-H in ON vo r>»
<t r^ ON •—I en r«. r>» r«. 00 00
m>ßr>.oooN o«-Hf-icNi 0000000000 o\ o^ ^ <J\
00 »-» v£) r^
VO rH o r-» VO CO CO rH
rH ON m CO VO N^ CO •—< rH ON
CO r^ CO O CO r^ CNj CM r-4 o
ON -^ m 00 r>» NO m «^t
CO <N CS| CM
rH o 00 m CO CM es es •—I o o O o
ON I—I o o
CO CO CO m o o r-. ^
o »-H ON ON r^ CO CM —1 r-H i-H
m t—• r-l O ON t-H »—I I—4 t—4 o
f^ vo in <i- o o o o
^ o o o o o o o o o o o o o o o o o
o \0 CM r-l
1^ rH CM o CM NO VO ON sj- CO ir> 00
00 NO m CO <í- cN in r^ m m ON r*» <f <f CM
ON 00 r-i r^ vo 00 m rH vo 00 ON «^ ^ CM .-H
m in ON CM .-H CO CO in ON r^ vO m
r^ vo CM CM
CO <N CM CM CM
<3 PQ < < CO r-l rH CO »-H r-4
3 ,0 CO
CO
CO -O T3 o •H u a s o <u o CO rH »H
o H
< < < < CM CM »-I CM CO
pq <î <î PQ CM CM CM rH
PQ <: rH CO CO CM
(U (U CO CO
•H «H (U 0) i-i ^ CO CO 1-1 O^ »O X)
<U O 'H «H »0 rH ex. TJ
CO >^«H «H O V4 M 42
•H O J3 a u o H <:
CO (U
T3 •H o
T3
CO -H
ü O
>^ ̂ -s CO CO W 'H ^^ »J /—\
. G >> (0 •u r-i
^ CO 4J
4J
•H rH O O) 4-t CO B
(U €0 O CO >-• CO 'V tí ^w' <u
T) «H 0) Í3 CO CO • H no CO a CO 6 <u T> (U -• O (U 73 M S CO •H CO -^ a CO •H CO to <ü tí 'O •H T)
CO 0) o T3 C T3 •H 4J o •H -CJ •H C c jû •H O •H M a o ^ •H tí
•H •H o »^ •H (U o o 4J o U O }-l o «H CO rH C o xi (U •H JC o CO <J 1-4 P (U CO o 4J CO rH 4J CO O O o >>t O M O) Kl Xí CO CO u < o Q S W < o O u s hJ pq

ECOLOGY OF COTTON INSECTS 63
ON m 00 r-. CM vx>
O^ O^ \D o <t 00
<Tx ^o 00 o r-4 r- ON CM
CM CO <1- CO <3N 00 00 ON O
CM en CO o> ON cr>
<t -^ -^ cr> o> CTN
in m in vO ON ON ON ON ON ON ON
v£> vO CJN ON
vO O <JN o
r^ v£5 CO <f -d- <!•
I—I O r^ <t <i- CO
CO vO Cvl Cvl CO C>J CM C>4
CM CM
O O O O C^J r-l
O d
CJN vO O o o r-l
O CO
o o o o
<»■ :í :^ o o o -^ <!• CO o o o
CO CM C^l CM o o o o
CM o
CM f-^ o o o o
o r^ o CM
o o o o
ON in CM CO CM ON in in <!■
•—• —• 00 r- \0 CM <f <t -^t
CO 00 r^ in 00 ON in m CO CM CM CM
ON
CM
CM P- ^ Cvl CM ^
O m o ^
PQ < fCl w < PQ PQ « ÇQ < <! <í < CO CM 1—1 r—1 <^ T—4 CM CM -H CM r-i CO r-H 1—í r-l r-l
/-N Cd . • ■u CQ ^-N
fa cd Í) ^ ^-\ «w' ■U 4-) Cd CO o Cd en
•H CO C •H ^-^ o 3 ;3 VJ > c Ö- <u CO . 3 • H 0 CO Î3 CO
. cd 4J •H • r-H •H CO QJ J-l O P eJ CO •H 'O
•i-í PQ (U <ü (U -H ^ c u p. Tí ^ Cd r-* • 3 Cd Cd (U 03 {3 u u <ü O. c v^
T-t 'd (U • 13 Cd O) O cu 00 O Cd 0) > Çh .CQ M-l CO CO CO .
Cd <u (U 4J (U cd Cd a 0) Cd .•H O ^-N . 3 (0 CO r-^ CO c o Tí -ö p CO CO cd o •H r-« B í-i CO Ou 3 ÍJ O T) • H CO •H • H 0) 3 .T3 • H la i-t o (ü •I-I I—t 0 <ü 1—1 ^ «H r-4 •ft O U ■u <u v^ Cd • l-l 4J i-t 9¿ 4J 4J ,c: o o 4= Cd
B o .«-< VJ O PM cd 0) ö o O Cd a Cd 3 4-1 CJ e: •y XJ Q) «H CO Cd o Cd o •X3 n3 (U •H U 1^ 1*^
Q) o o o o o :^e :3 Ä ÖOr-t o •H 0 4-» JC ^ o CO íí Píl •l-l U rC H
bO r-< Cd o U3 ts CO 4-t Cd Xí •H O) N>X r-t 0 4J r-4
o o >N 3 X! CO Cd o ?^ C2 •H OJ CO <U <v C r-l
o fa a fa PA çu s o CO <: Q < ÇU 50 z < <:
>% U cd CO e «-t
•H Cd »-I 4J a c
(U }-4 .-Ö o -H e o
•H C
fï pQ CO
^ r—1 o« .»« o .« CO Wl
/-^ n ü
Í s 0) CO Í) ^ 0) CO 4-t ou C3
w o o O 4J
CO VJ >^'0
g ^ XJ :i ^ Cd CO C r-l C O 0) o Ü u o Q)
CO CO
>^ Cd >-l V4 Cd o CO
e a V4
V4 6 i cu 1 0
1 CO 4J PQ c: C CM o Cd Ü 4J ..»
í-l ^^ >% O CO M CU r-l Cd a Cd 'X3
.3 .r^ C 1 O O 1 -r^ o < CM 0) ^ (U CO
a 0) 4-> .. ^ g
p-^-^ CÎ o U CO V4 Ü VJ o
a g cu O 3 o 4J CO u J-» a o o o o o CU
xi o í^ 4-1 4-» U
cd o 13 Tí 4J Q) C *J O r-l cd ü cd
r-4 Q) u O) CO c¡ »-I
4-> 1 CO C •H Cd cd u
u 4J r-l M <u
CU o 13
> Q* d) 0 4J r-l .3 O
1 c =o¿! V4 • rl Cvl 0) ^ x:
CU »-^ 4-1 o CO •rl u u Q)
t s ^ :3 •
■^1 w O) ^ CÎ *
o •H o ^-^

64 W. H. CROSS
these are butterflies, larger grasshoppers (Melanoplus, Schisto- cerca, Dissosteira, Orchelimum, Conocephalus), Gryllidae, Carabi- dae, Collembola, Lycosidae, Odonata (Anax, lÊrythemis, Pantala), Annelida, amphibians (Bufo, Rana), reptiles, birds, and mammals (mice, shrews). In addition, the cotton growing seasons of both 1978 and 1979 followed colder than average winters, and the cotton fields sampled were usually included in the trial programs (about two-thirds of the fields were in the actual evaluation areas); con- sequently, pest populations were generally low, and beneficial pop- ulations were generally protected.
Other biases of the values are more specific. The family Chloropidae at more than two per row-foot includes several species of small gnats that were probably in the cotton foliage seeking a favorable physical environment and perhaps feeding on nectar. Whiteflies (Trialeurodes abutilonea), though present at almost the same density, did not cause significant damage. Leafhoppers (Cica- dellidae) tend to be more abundant on grasses than on cotton, so the large numbers (excluding a few genera such as Graphocephala) must indicate the presence of grass weeds in the cotton. The family Thripidae includes some banded-winged thrips (Aeolothrips) that are predaceous, but these are not identified separately. Similarly, the order Acariña includes a few predaceous Oribatidae, and the order Orthoptera includes a few Conocephalus and Oecanthus that are sometimes predators. Finally, Table 1 is oversimplified in that it does not take into account the effects of hyperpara- sit ism or prédation on predators. For example, a numerical summa- tion (see footnote 1, Table 1) shows that lA + IB = 53.67% of the total, 2A + 2B = 15.01% of the total, and 3 = 28.22% of the total, so one might roughly assume that 15% of the secondary consumers (2A + 2B) are in fact tertiary consumers. Clearly, many direct field observations are required to measure the impact of secondary and tertiary consumers. Thus, many feeding relationships are in play within the community of the cotton field that are not readily apparent when a few pests and a few beneficial insects are sampled.
Nevertheless, those relationships that are important to the producer are readily apparent in Table 1. For example, the pests that a trained scout records in the eastern parts of the U.S. Cot- ton Belt are whiteflies, thrips, aphids, Lygus, fleahoppers (Pseudatomoscelis seriatus), Heliothis spp., clouded plant bugs (Neurocolpus nubilus), and boll weevils (Anthonomus grandis). Others may be listed but they are only occasionally economic pests (Little and Martin 1942, Young 1969). Principal beneficial insects that would be noted are indicated in Table 1—some Chalcidoidea and Braconidae (parasites) and a number of predators including Orius, ladybird beetles (Coccinellidae), spiders (Araneida), big-eyed bugs (Geocoris), and Nabidae. Others may play a role at times (Whitcomb and Bell 1964, Young 1969) and certainly have impact in composite.
Also, some pests are seasonal in importance. Thrips and aphids may appear on cotton plants throughout the season, but

ECOLOGY OF COTTON INSECTS 65
thrips usually damage only young plants, and this is often true of aphids. Then there is the cotton leafworm (Alabama argillacea (Hübner)), which is not listed in Table 1. Formerly this immigrant from the Tropics was occasionally a devastating pest over wide areas, but in other years populations increased so late in the season that the insect simply helped producers defoliate the cotton in preparation for harvest. On the other hand, there are major pests such as Heliothis spp. and the boll weevil that develop sev- eral generations on cotton through the growing season. Plant bugs of several species, including Lygus spp. and, recently, Neurocolpus nubilus, also tend to have this characteristic.
A CASE HISTORY OF MISMANAGEMENT OF COTTON ECOLOGY
Experts have used the information on the biology, behavior, and ecology of the boll weevil and the composite picture of the cotton field community (summarized in Table 1) to devise techniques for managing pest populations in cotton. However, the justifica- tion for these techniques and their effects on the ecology of the cotton field can be understood most easily by presenting a case history of mismanagement. 2_/
The 23-acre cotton field in question is in northern Pontotoc County, MS, and was one of several small fields in the area farmed by a local grower. It was one of the several current practice fields for the Optimum Pest Management Trial (OPMT) in Panola Coun- ty, MS. Therefore, the arthropod population was sampled each sea- son from 1977 to 1980. In addition, a team of entomologists V intensively sampled the field each 2 weeks in 1980. However, the results of the samplings were not conveyed to the grower in order to avoid influencing normal management in any way, thereby intro- ducing bias into the check.
The peripheral area around the field had several good over- wintering sites for boll weevils, and the winter of 1979 was mild enough to ensure good survival of overwintering weevils. In fact, four infield boll weevil traps placed on stakes around the field captured 31 weevils (7-8 per trap) in the 9-week period (April 23 to June 30) before the first puncture-sized squares were found in 1980. With optimum pest management, this number of captured wee- vils would have been the signal for close scouting to determine
2/ History largely prepared by Gordon L. Snodgrass. V W. H. Cross, G. L. Snodgrass, P. R. Miller, J. T. Robbins,
III, "et al.. Agricultural Research Service, U.S. Department of Agriculture, Boll Weevil Research Laboratory, Mississippi State, MS; and J. W. Smith and T. C. Lockley, U.S. Department of Agricul- ture, Bioenvironmental Laboratory, Stoneville, MS.

66 W. H. CROSS
,r o 00 CO 'Cl- C\l û. CO CsJ CM eg CNJ
O 4J O CO
o U U
O a
•H Q)
2 ^ (U O
•H M
O
Cd o I
en CO CNl (U
f-4 (U cu
42 e u td
CQ c
Cd CO >
Tí I o o o u u o
Xí ^
Cd Q) *J
r-l o Cd a>
•H o M-l o
C 3 Cd c
■u u •« o CO
e o •H CO
Cd ^-1 (U o CO
CO 00 c c o »H
•H l^ 4-» O Cd u
.-» ÖO
Cu CU o <u a u
T3 4J <u
•H tí
-5 *^ 0 3
<ü «
1 •»
es u tí
V4 o

ECOLOGY OF COTTON INSECTS 67
whether there was need for a pinhead square application of insecti- cide for boll weevils. (Capture of 4-5 per trap was used in the OPMT as the threshold for this application.) However, the grower did not use a cotton scout and made no application.
The overwintered weevils that entered the field infested 1.5- 3% of the squares; the F, generation emerged around the third week of July and infested 7-9% of the squares, and the F^ generation emerged around the first week of August and infested 51.5% of the squares. (Control measures are recommended when the rate exceeds 10%.) Meanwhile, a sizable population of beneficial arthropods was helping control the bollworm populations in the field (bollworms were first detected there July 17). On July 30, the grower made his first application of insecticide (1/2 pound methyl parathion plus 1/8 pound methomyl per acre); he then made three more appli- cations (each 3 pound methyl parathion plus 3 pound toxaphene per acre) on August 8, 15, and 23. Plainly, at the time of the first application, the highest rate of square infestation had been reached. However, the grower probably sprayed because his neighbor sprayed, not because he realized the extent of the problem (the neighbor used a cotton scout). Also, the addition of methomyl to the material applied July 30 probably indicated that the grower did not know what his insect problems were (bollworms were not a prob- lem in the field at that time). Finally, the timing of the last three applications was incorrect. They should have been 4 or 5 days apart instead of a week or more apart if they were to provide optimal boll weevil and bollworm control.
One obvious result of the series of events was a continuation of the boll weevil problem: boll weevils remained at an econom- ically damaging level for the rest of the growing season. Another was the creation of a bollworm problem. One week after the first insecticide application, the bollworm population in the field reached an economically damaging level and remained there for the rest of the growing season.
The reason for the bollworm problem is clear from Figure 2, which reflects the combined populations of important beneficial ar- thropods (nabids, spiders, lacewings, ladybird beetles, Orius sp., and Geocoris sp.) present in the field during the 1978, 1979, and 1980 growing seasons. These levels were determined from D-Vac sam- ples taken as part of the Current Insect Control practice compari- son with the OPMT. In 1978 and 1979, when no insecticides were used in the field through August, the populations of beneficial arthropods peaked in July and August and began to decline during the last week of August. Thus, before the natural decline in the populations of beneficial arthropods that would probably have oc- curred around the last week of August 1980, the grower could have received the benefit of their presence to contain his bollworm problem. Instead, the four insecticide applications reduced these populations from 22,200 per acre on July 24 to 0 per acre on Auguát 26.

68 W. H. CROSS
Additional information on the effect of insecticides on the populations of beneficial arthropods in the field was obtained July 30-31 in two special surveys. On July 30, D-Vac samples were taken before the first application of insecticide, which was made that afternoon. A second set of D-Vac samples was taken during the morning of July 31. In addition, on July 31, the dead arthro- pods in 375 feet of row were picked up by the team and taken back to the laboratory for identification. From the D-Vac samples, the population which consisted of the same six groups of beneficial arthropods sampled earlier, was 20,234 per acre in the field before the first application and 8794 per acre after the application, a reduction of 56.5% in less than 24 hours. From the collection of dead arthropods on the ground in the field, the reduction was only to 5161 per acre, which was expected since it was difficult to find all the dead arthropods. However, the hand collection also showed that the insecticide had killed at least 586 boll weevils and 37 adult Heliothis spp. per acre, and both of these insects are poorly sampled by use of the D-Vac.
In sum, the producer's failure to use optimum pest management practices including a cotton scout led to additional insect prob- lems in his field. Moreover, the use of insecticides to control the boll weevil was the key. If the boll weevil had been con- trolled by insecticide during the early part of the growing season, the need for insecticide treatments for boll weevil control would have been delayed until August. Better management of insect pests may or may not have increased yield of cotton lint, which was 479 pounds per acre in 1980, about average for the county. However, proper timing of one boll weevil treatment in the early fruiting period would almost certainly have resulted in less total expense for insecticide."
Once again it has been demonstrated that the boll weevil is the key pest because of inadequate natural biological controls. It therefore requires the early use of insecticide, which, in turn, kills predators of bollworm eggs and young larvae and so allows the pest population to explode. There may be exceptions. For example, in areas where large corn crops mature before the cotton crop, Heliothis spp. may shift so heavily to cotton that the predators cannot control them. However, the sequence described here has oc- curred over and over since the boll weevil first appeared in the United States. It is still occurring.

ECOLOGY OF COTTON INSECTS 69
LITERATURE CITED
Cross, W. H. 1973. Biology, control, and eradication of the boll weevil. An- nual Review of Entomology 18:17-46.
D. D. Hardee, F. Nichols, H. C. Mitchell, E. B. Mitchell, P. M. Huddleston, and J. H. Tumlinson.
1969. Attraction of female boll weevils to traps baited with males or extracts of males. Journal of Economic Entomology 62: 154-161.
M. J. Lukefahr, P. A. Fryxell, and H. R. Burke. 1975. Host plants of the boll weevil. Environmental Entomology 4:19-26.
and H. C. Mitchell. 1966. Mating behavior of the female boll weevil. Journal of Ec-
onomic Entomology 59:1503-1507.
Dunn, H. A. 1964. Cotton boll weevil (Anthonomus grandis Boheman), abstracts
of research publications, 1943-60. U.S. Department of Agricul- ture, Miscellaneous Publication No. 985, 194 p.
Fye, R. E. 1968. Spread of the boll weevil by drainage water and air cur-
rents. Journal of Economic Entomology 61:18-24.
Hardee, D. D., W. H. Cross, and E. B. Mitchell. 1969a. Male boll weevils are more attractive than cotton plants
to boll weevils. Journal of Economic Entomology 62:165-169.
W. H. Cross, E. B. Mitchell, P. M. Huddleston, H. C. Mitchell, M. E. Merkl, and T. B. Davich.
1969b. Biological factors influencing responses of the female boll weevil to the male sex pheromone in field and large-cage tests. Journal of Economic Entomology 62:161-165.
Little, V. A., and D. F. Martin. 1942. Cotton insects of the United States. Minnesota Burgess Publishing Company, Minneapolis, 130 p.
Lloyd, E. P., F. C. Tingle, and R. T. Gast. 1967. Environmental stimuli inducing diapause in the boll wee-
vil. Journal of Economic Entomology 60:99-102.
Mitchell, H. C, and W. H. Cross. 1969. Oviposition by the boll weevil in the field. Journal of Economic Entomology 62:604-605.

^^ W. H. CROSS
Mitchell, H. C, and W. H. Cross. 1971. Mating of boll weevils in the field. Journal of Economic Entomology 64:773-774.
Mitlin, L. L., and N. Mitlin. 1968. Boll weevil, Anthonomus grandis Boh., abstracts of re-
search publications, 1961-65. U.S. Department of Agriculture, Miscellaneous Publication No. 1092, 32 p.
Roach, S. H., H. M. Taft, L. Ray, and A. R. Hopkins. 1971. Population dynamics of the boll weevil in an isolated cot-
ton field in South Carolina. Annals of the Entomological Sou- cie ty of America 64:394-399.
Walker, J. K., Jr., and D. G. Bottrell. 1970. Infestations of boll weevils in isolated plots of cotton
in Texas, 1960-69. Journal of Economic Entomology 63:1646- 1650.
Whitcomb, W. H., and K. Bell. 1964. Predaceous insects, spiders, and mites in Arkansas cotton
fields. Arkansas Agricultural Experiment Station, University of Arkansas, Bulletin No. 690, 84 p.
Young, D. F., Jr. 1969. Cotton insect control. Oxmoor House, Birmingham, AL,
185 p.

SECTION il SUPPRESSION COMPONENTS


73
Chapter 4
PLANT RESISTANCE AND MODIFIED COTTON CULTURE
L. N. Namken and M. D. Heilman Soil and Water Conservation Research Laboratory
Agricultural Research Service U.S. Department of Agriculture Weslaco, TX 78596
J. N. Jenkins Crop Science and Engineering Research Laboratory
Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
P. A. Miller National Program Staff Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705
ABSTRACT The potential of genetic alterations plus improved pest management and cultural prac- tices in cotton growing systems in the regions of the Cotton Belt where boll weevils (Anthonomus grandis Boheman) occur are evaluated. Studies involving use of frego bract, red plant color, and male sterility resistance traits are reviewed relative to overall cotton pest management prac- tices and production efficiency. The advantages of rapid-fruiting cotton genotypes in short-season production systems are discussed, and data are presented to show that simultaneous improvement in rapid fruiting and fiber quality is possible. A short-season cotton production system developed for the south Texas environment is described.
INTRODUCTION
Research to develop cotton cultivars that have improved re- sistance to pest insects and modification of cotton production sys- tems so as to manage these pests more effectively and efficiently are being emphasized across the Cotton Belt. In the portions in- fested with boll weevils (Anthonomus grandis Boheman), particular attention is being given to cultivars and production systems di- rected against boll weevils, tobacco budworms (Heliothis virescens

'^^ L. N. NAMKEN ET AL
(F.)), bollworms (Heliothis zea (Boddie)), and tarnished plant bugs (Lygus lineolaris (Palisot de Beauvois)). The range of environmen- tal conditions in these areas and the diversity of insect problems these cotton growers face make it doubtful that any one cultivar or production system will suffice. Nevertheless, the probability that we can develop efficient systems capable of dealing with local or area problems has recently been greatly enhanced by (1) the availa- bility and selection of more efficient genetic material, (2) the development of optimum pest management practices based on a better understanding of ways of minimizing boll weevil damage while sup- pressing other pests, (3) the development of more efficient cultur- al practices, and (4) the realization that it is through the integrated efforts of agronomists, plant breeders, entomologists, engineers, and economists that significant gains will be made.
Our primary objective here is to evaluate current advances in genetic alteration of the cotton plant, or those soon to be made available, and to consider their potential impact when they are coupled with improved pest management and cultural practices in cotton production systems in the regions of the Cotton Belt that are infested with boll weevils.
CURRENT COTTON CULTURE IN INFESTED AREAS
The average harvested acreage of cotton, the yield per acre, and the production of 480-pound bales for 1974-78 are summarized in Table 1. Similar data for 1970 and 1979 are shown in Table 2. These tables report data for four regions.
However, for the purposes of this discussion, which has to do with areas of the United States infested with boll weevils, the West Region has been dropped, and the other three regions have been rearranged as follows:
Southeast: Georgia, North Carolina, South Carolina Midsouth: Alabama, Arkansas, Louisiana, Mississippi,
Missouri, Tennessee Southwest: Oklahoma, Texas
Thus, Alabama is considered a part of the Midsouth Region (term re- placing Delta), not a part of the Southeast Region. The West Re- gion is not involved since boll weevils do not occur there.
Southeast Region
Major cotton cultivars planted in the Southeast Region are listed in Table 3. The Coker, McNair, and Stoneville cultivars used in the Southeast Region are full-season or midseason culti- vars with open-type bolls adapted to spindle-type harvesting. These cottons have relatively long (1 1/16-1 1/8 inch) fibers of high fiber tensile strength and relatively high micronaire (mature fiber) readings.
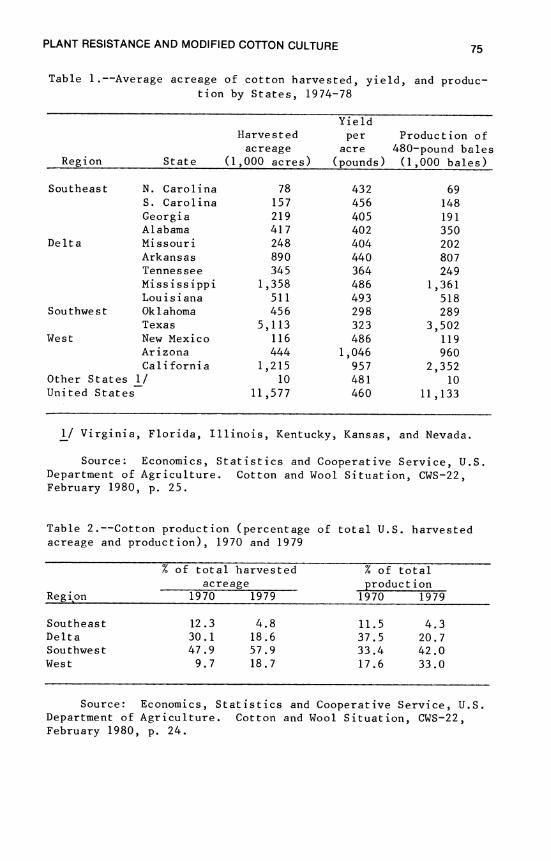
PLANT RESISTANCE AND MODIFIED COTTON CULTURE 75
Table 1.—Average acreage of cotton harvested, yield, and produc- tion by States, 1974-78
Yield Harvested per Production of acreage acre 480- -pound bales
Region State (1,000 acres) (pounds) (1 ,000 bales)
Southeast N. Carolina 78 432 69 S. Carolina 157 456 148 Georgia 219 405 191 Alabama 417 402 350
Delta Missouri 248 404 202 Arkansas 890 440 807 Tennessee 345 364 249 Mississippi 1,358 486 1,361 Louisiana 511 493 518
Southwest Oklahoma 456 298 289 Texas 5,113 323 3,502
West New Mexico 116 486 119 Arizona 444 1,046 960 California 1,215 957 2,352
Other States 1/ 10 481 10 United States 11,577 460 11,133
JL_/ Virginia, Florida, Illinois, Kentucky, Kansas, and Nevada.
Source: Economics, Statistics and Cooperative Service, U.S. Department of Agriculture. Cotton and Wool Situation, CWS-22, February 1980, p. 25.
Table 2.—Cotton production (percentage of total U.S. harvested acreage and production), 1970 and 1979
% of total harvested % of total acreage prodi
1970 iction
Region 1970 1979 1979
Southeast 12.3 4.8 11.5 4.3 Delta 30.1 18.6 37.5 20.7 Southwest 47.9 57.9 33.4 42.0 West 9.7 18.7 17.6 33.0
Source: Economics, Statistics and Cooperative Service, U.S. Department of Agriculture. Cotton and Wool Situation, CWS-22, February 1980, p. 24.

^^ L N. NAMKEN ETAL
Table 3.—Major cotton cultivars planted in the Southeast Region, 1979
Estimated cotton acreage (%) planted 1/
Cultivar North Carolina South Carolina Georgia
Coker 304 10 41 21 Coker 310 61 37 38 McNair 220 24 8 Stoneville 825 5
y Total acres planted: North Carolina, 50,000; South Carolina, 115,000; Georgia, 120,000.
Source: U.S. Department of Agriculture, Agricultural Market- ing Service, Cotton Division, Memphis, TN, September 1979.
Most cotton in this region is rain grown, though supplemental irrigation is applied in some areas. Chemical defoliants are gen- erally used as a harvest aid, and a field can be harvested two or more times with a spindle-type, selective harvester. Both the boll weevil and the bollworm/budworm complex inflict substantial damage. Fusarium wilt (Fusarium oxysporum Schlecht. f_. vasinfectum [Atk.] Snyd. & Hans.) and root-knot nematodes (Meloidogyne incognita (Ko- foid & White) Chitwood) are prevalent, particularly on the more sandy, acid soils.
Midsouth Region
The major cultivars planted in this region are listed in Table 4. Stoneville and Deltapine types, which are full season or midseason in maturity, predominate. These cottons have open-type bolls and are adapted to spindle harvesting. Lint characteristics are similar to those of the southeast cultivars in that the fiber is relatively long, strong, and mature.
Some supplemental irrigation is used in the region, though most cotton in the region is rain grown. For the most part, the crop is spindle picked, and chemical defoliants are used to facili- tate harvest. Plant bugs, the bollworm/budworm complex, and the boll weevil are important insect pests. Fusarium wilt, root-knot nematodes, and Verticillium wilt (Verticillium dahliae Kleb.) are prevalent. Boll rots (various organisms), particularly in the southern part of the region, often cause significant losses in yield.

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 77
Table 4.—Major cotton cultivars planted in the Midsouth Region, 1979
Estimated cotton acreage (%) pL anted 1/ Cultivar MO TN AR AL MS LA
Coker 310 15 Deltapine 16 8 10 8 11 Deltapine 55 15 9 10 7 18 21 Deltapine 61 11 10 23 15 20 29 Stoneville 213 58 48 52 27 40 27 Stoneville 825 7 10 6 6
1/ Total acres planted: Missouri, 170,000; Tennessee, 250,000; Arkansas, 700,000; Alabama, 335,000; Mississippi, 1,050,000; Louisiana, 480,000.
Source: U.S. Department of Agriculture, Agricultural Market- ing Service, Cotton Division, Memphis, TN, September 1979.
Southwest Region
The Southwest Region includes both the semiarid High Plains of Oklahoma and Texas (10-20 inches of annual precipitation) and the more humid areas (40-50 inches) of eastern Oklahoma and Texas. Cultivars and cultural practices, therefore, vary so greatly that the major cultivars planted in the region are listed in Table 5 by marketing service area (Figure 1). The major characteristics of these cultivars are compared with those of all cultivars grown in the three regions in Table 6.
The more arid western areas of Oklahoma and Texas, including the High Plains, Rolling Plains, and Blacklands, produce cotton for once-over stripper harvest so cultivars adapted to this type of harvest predominate. Seedling diseases (Rhizoctonia solani Kühn, Pythium spp., and Fusarium spp.), Verticillium wilt, and bacterial blight (Xanthomonas malvacearum [E.F. Sm.] Dows.) can cause major losses in these areas. The boll weevil is a problem, and the boll- worm/budworm complex and the plant bug complex occasionally cause local problems.
Northeastern Texas, the river bottoms of central Texas, the Lower Rio Grande Valley, and the Coastal Bend areas have substan- tially more rainfall than the western areas, and growers spindle harvest a major portion of their crop. Stoneville, Deltapine, and McNair (open-boll cultivars) are therefore grown as are intermedi- ate boll-type cottons adapted to both stripper- or spindle-type harvest. However, growers in the Coastal Bend area of Texas are shifting rapidly to short-season cultivars and are using once-over stripper harvest; and producers in the Lower Rio Grande Valley are also making some transition to short-season cottons that tend to

78 L N. NAMKEN ET AL
O •ri > U Q) œ
txû tí
M U CO IS <D U 3
<
tí (U 6
■u u CO Ou <ü Q
I I

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 79
o •H
t>Û (U
Oí
4J
O OS
<D
00 tí
u tu 6
CO (U u CO
Ü CO (U
(U
O O
U O
to f
0)
CO H
Q) tí <U
1—1 •H "*»>^ U3
-^1 <3 T3 (U •U tí CO
1—1
tí a <>0 tí Q) •H bû t—1 CO u Oí CO M P3 o CO
tí o 4-> •H u to 4J o 3 CO o a •H
V4 }^
r>. rH in es
86
rol
CO CO
CNJ CO tí
i-H
O ^ m .-H
O 0\ \D
•<t CO <J- CN CM -H
I—I vO .—I í^ -H X
0) 0) in vo H-3 tí tí •H J_»
Pu VJ CO CO
^ '^ »-I f-H tí <U 0) CO Q Q HJ
CO
en o CO
u
cvi CM -H r«H CM en
<U (U I I
CO CO
u u CO CO
tí í CO CO
M CO >> 4-1 4-1 •H to (Ü 0) o o to e tí tí o Ü ÏS >^ O O ß Ë U to 4-J 4-> to to S f^ CO CO H H
^ H
^ en
•H CO JH tí Pu cu B v^ Q) o S o .«« tí o O o •H o CO •» •H CX) > c^ •H en Q
» tí tí o •H 4J 4-) 4-) to O tí ü <í « .-O <ü o o O o o •H o - >
«en V4 o -H Q) o o CO en •» -^ 00 tí œ « to . •H CO ^ CO CO ■U
r-t o X g 0) rH o (U O ^
CO ^ H JC V4 Q 43 CO CO
tí tí rH s .». HJ .H 4í5 O O r-i o »»^ CO CO o o 0) tí U
•»o M «H tí CM O U •U vO ^ to CO rH en C>J <ü tí
"CX) O V4 O rH —1 O O •H
« o CO ^1 « rH •» 00
to ON O <3 tí «m O *j 0) r^ o M
rH tí <ü <3 <u Tí o U ^ tí -< tí
•H tO 4J .. Xí -o rH
-o < 2 tí tí (u 6 to O 4J .« o •H tí o X CO »-I CO o CO CO 00
.-• o rH X <í a -^ 0)
<!■ o H MH <1> <JN O öOen tí tí CO «H «H ■P <ü « tí M tí CO CO o <ü (U <u g CO 00 M V4 4J
Coo U rH «H CO CO CO
CO r-H a 4J ^1 o o (U o CO o o Q 4J EC o o
M-i .« en o CO o o o ON
o v£> CM ¿ . CO O c^ 0) •« CO CO r-^ 4J en (U <u .. a^ CO CO na -o Q) rH e en tí tí o
•H ,H rH u u 4-» •» u o tí <u CO «H tí tí o Xi
|jq 4J M M CO g CO (ü
4J rnl Í-I CM jen 1 Cu
\C ' ' Q) U CO

80 L N. NAMKEN ET AL
o
Xi
73
CO U CO
T3
C D
öocnl C ' 4J O CO ^
> ai CO
o >>
ON ^ CO
CO
so <u
r-l
•§ H
00 JC CO 4J bO Ö0 tí X (U o »^ tí
CO I
jc: CO
tí o (U tí
Q Q) Q) Q) Q) 0) (U <U (U
Tí "TlJ TlJ "TJ tJ C tí C tí tí 42 JC JC ^ tí X tí tí tí „ _ „ «H 4J «H . . pua4a.Puao o o pL» o OiCucxo o COWCncOCOPQPQPQCOPQWWCOPQPQ
OOOOOPOP£5tóOPE!OOO0cS0cí CO CO en CO CO CO
TJi-trH'Oi-l'O O'd'O OrHrH'O O O
SfefaSfaSÖSgiScoptHfeScoco
CO<l-COCM<t'-HCM.-(-<î-CMCOCSmC>ICS CMCMCMCSCMCNCNCSCMCMCSC^JCMCSCN
cncMmtnr^cna>cni-HLnr^moor^c30
som<t->d-m-H^Osocs^>í-*cncs mcoc^cocnmcococncnmcocncom
m so m rH m —1 in CM
so lO 1-H r^ o CM CM 00 ^ rs. FH m so
r>- ^ X m so iJ
en CM O <U <ü <U 1
en
•4- O <D (U (U CM l-l rH f-l i-l pLi P4 M o —• tí tí tí CM QÍ f-i f-l i-l CO CO CO en m •H «H «H u u XJ U «H «H «H > a a Pu M M M H CO > > > 4-» 4J
•H u u CO CO CO CO CO CO *H CO 0) 0) 0) O O 4J 0) Q) u u u ^ ^ *M CO s tí tí tí o
S23 ^ o o o i <J
r-l M ^ r-« rH 1—1 tí tí tí B S O o (U (U 0) CO CO CO U CO -M 4J 4J CO CO Ü ü u Q Ö Q HJ HJ HJ s P^ CO CO CO H H
CO
CO 73
<f I—I
I m m
tí o CO CO (U (O
73 •H a
T3 •H
73 tí CO
0)
O
73 tí
CO
o
o u
H
a O)
> •H Q tí o
O
CO
00 tí
0)
•H o 73 CO »-I ^ tí S 3 •H 4J (1) a f—1 CO PL, CO CO g >. M •
u u tí CM o 1 o 4J 4J tí ^ 1—1 •
Q) tí Ok Ö0 a tí o tí o o •H •«
•H •H U as iJ il «U 00 1^ tí CO <: <3N CO O .u •—1
f-i a •« a 73 CO (U M
tí 73 M 0) a CO CO :3 n o ^ a M ^-^ u r^ Û)
M-l 0) o tí JJ *J M-l u a
CO CO •H <U >% • •H TS M CO CO ^^ 73 <ü 00
73 CO 0) 4J <í >> 1 c; O CO M-l 0>* 00 73 0) r-l • O r^ f-l +J <ü u 1
1 m tí CO <ü u m ITS m •H •u tí r>. <!■ rH v^ CO CO Q) ON r-l 1 CO <ü B »-H N-^ O 4J ^ > 4J
CM tí u M -d tí ^ CO »-I CO CO <ü O w 4J CO 43 (X u CO CO > Q) tí CO tí •H «H U o CO (U o CO 4J Q) I—1 CO CO <ü f-l a . O«
CO u a eu CO l-l Q) u •H • CO l-l CO
s CO U CD <ü
3 u •H M-l 4J o <u CO 4.)
M 4J 4J .. <u II O CO CO <D <ü .H
X O a U U l-l CO II «H >> u CO f—1 . 'Ö 4J tí > tí II (^ tí 1 O
PC4 CO M M CO tí 4-1 0) o
"^1 ^ "•»^ "**>, Ö0 ■u
-H O CM jen tí 4J X •H o CO M-4 ü

L N. NAMKEN ETAL 81
have shorter lint, less tensile strength, and somewhat lower micro- naire readings than the midseason or full-season cultivars. Most of the river bottoms and about half of the Lower Valley are irri- gated. Growers may have serious problems with the bollworm/budworm complex as well as with the boll weevil. Root rot (Phyinatotrichum omnivorum [Shear] Duggar) may cause severe losses in local areas.
POTENTIAL OF RESISTANT COTTONS
Plants differ in their ability to produce under attack by pest insects, an ability that is commonly called resistance. Therefore, resistance of an experimental strain is usually measured by compar- ing the experimental strain with a cultivar known to be "suscepti- ble." However, total resistance would be immunity, which would mean that a cultivar or strain is not attacked by a given pest un- der any condition, an exceedingly rare occurrence. Resistance is thus relative and encompasses any degree of favorable plant reac- tion less than immunity. Most often the level of resistance en- countered in plant cultivars is moderate, that is, partial or intermediate. Practically, a resistant cultivar is resistant to a given pest when it produces a larger crop of satisfactory quality than other cultivars when all are challenged equally by that pest.
Usually, in cotton fields, there are several insects of impor- tance. Therefore, strains that are resistant to one species of insect can be only one of the components of an integrated pest man- agement program. One must consider the population of the pest to which the strain or cultivar is resistant and its response to other insects and to diseases in formulating an integrated program.
Frego Bract Resistance to Boll Weevils
The boll weevil is considered a key pest in areas where it oc- curs, so much effort has been expended in a search for resistance to this pest. One morphological trait of the cotton plant that has been investigated thoroughly is frego bract, a name applied to a rolled bract mutant found by a Mr. Frego in a field of Stoneville 2-B cotton near Manila, AR, in 1943 or 1944 (Waddle 1972). Frego is imparted by a single recessive gene designated _fg (Green 1955). Jones et al. (1964) recorded a 56% reduction in oviposition by boll weevils in field plots of frego cotton. Hunter et al. (1965) veri- fied that the trait inhibited weevil feeding and oviposition when weevil populations were moderate to low. Resistance was also noted by Lincoln and Waddle (1965).
Investigating further, Jenkins et al. (1969) reported reduced oviposition by boll weevils in field plots planted to five frego strains compared with plots planted to a nonfrego control. How- ever, when weevils were confined in the laboratory on bracts of frego cottons, there was no reduction in oviposition. Mitchell et al. (1973) then found that overwintered and current season female

82 L. N. NAMKEN ET AL
weevils oviposited about 50% fewer eggs on frego cotton in field plots than on control cotton. Moreover, the marked weevils de- parted the frego cotton about twice as rapidly as they departed nonfrego cotton plots: 64% were recovered from squares in control plants and only 20% were recovered from frego buds. Also, weevils were frequently found on leaves and stems of frego cotton and more often on terminals than on squares, which were preferred locations in control plots. Thus, there was more plant-to-plant movement of females on frego cotton than on the control, which suggested sev- eral ways of using the frego character in an integrated control program. For example, pheromone traps might be more effective in plots of frego cotton.
Meanwhile Jenkins and Parrott (1971) planted 10-20 acres of frego cotton on four grower farms in an effort to estimate the cum- ulative effects of frego cotton and appropriate controls on popula- tions of boll weevils exposed for three or four generations. They then compared oviposit ion (an indirect measure of population sup- pression) on the two types of cotton. Oviposition was 66, 71, 75, and 94% less in the four fields of frego cotton than in the control fields. Moreover, the lesser reductions each succeeding generation reflected smaller numbers of overwintering weevils present in the fields and different control measures used during the season. On two farms, no insecticides were necessary in the frego fields, but the cotton in the control fields had to have normal amounts.
Subsequently, the effects of frego cotton and reproduction- diapause control were studied by Jenkins et al. (1973) at 22 sites in Yalobusha County, MS. Populations of boll weevils in 4-acre fields of frego cotton that received or did not receive diapause control were suppressed 69 and 79%, respectively, compared with ad- jacent, similarly treated 4-acre control fields. Still more im- portant, the beginning of weekly applications of insecticides for boll weevil control could be delayed 4 weeks in the frego cotton. Thus, growers who plant cotton cultivars that are resistant to boll weevils should be able to withhold applications of insecticides against boll weevils until populations of Heliothis spp. have peaked, thereby protecting the populations of beneficial insects that can contribute to control of the bo11worm/budworm complex. As other types of resistance to the boll weevil become available, they should be as useful as, or more useful than, the frego character in integrated pest management programs.
The interaction between insecticide and cotton cultivar was investigated in 12 fields planted to frego cotton (90 acres) and in 12 fields planted to nonfrego cotton (128 acres) in Copiah County, MS, in 1972 (Jenkins 1976). The populations of weevils in all fields averaged 42 per acre at time of pinhead square, the time when one application of azinphosmethyl (0.25 pound per acre) was made. Thereafter, fields were treated individually if more than 10% of the squares had damage due to weevil oviposition. In the fields of frego cotton, there were 50% fewer damaged squares than in the control fields and 46% fewer applications of insecticide.

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 83
Yields of cottons were essentially identical in all fields, 1.7 bales per acre.
More recently, reasons other than simple nonpreference have been advanced to explain the "resistance" of frego cotton to boll weevils. Parrott et al. (1973) recovered approximately seven times more insecticide from frego squares than from nonfrego squares, which should mean that insecticides applied to frego squares would give better control of boll weevils and other square feeding in- sects. Also, insecticides applied to frego cotton for control of Heliothis spp. were more effective (Schuster and Anderson 1976). McGovern and Cross (1976) found a higher rate of parasitism of boll weevil larvae by Bracon mellitor Say in frego cotton than in non- frego cotton. They attributed the difference to the fact that frego has an exposed flower bud, which increases the efficiency of the parasite in its search for squares containing developing boll weevil larvae.
In view of the changes in behavior of boll weevils exposed to frego cotton, the effectiveness of the trait might well be enhanced by interplanting strips of nonfrego cotton with frego cotton. Wea- ver and Baker (1972), therefore, planted mixed strips of 40 rows of frego cotton and 8 rows of nonfrego cotton in a 10-acre field. Only 6 applications of insecticides had to be made to the frego cotton to control boll weevils; 14 were needed to protect the non- frego cotton. Such an approach would increase the opportunities of boll weevils to express nonpreference, and so it has definite pos- sibilities for pest management programs.
Other Potential Sources of Resistance
Red leaf (red stems and leaves) plant color has long been known as a nonpreferred cotton phenotype when boll weevils have a choice. For example, Isley (1928) reported that a red leaf strain had fewer damaged squares than a green leaf strain. Jones et al. (1978) discussed the potential of this and other plant resistance traits as management strategies and reported results obtained by several researchers in Georgia, Louisiana, and North Carolina. For example, when insecticides were applied only to strips of green, normal bract cotton planted in fields of red leaf cotton in Louisi- ana, in-season (July and August) control of boll weevils was ob- tained with 80% less insecticide. They also reported unpublished data of D. F. Clower, J. E. Jones, L. Williams, and J. R. Phillips who interplanted 88 rows of red leaf cotton with 8-row strips of a green, prolific-fruiting cotton. Again, good control of boll wee- vils was obtained until "cut out" by applying insecticides to only the 8-row strips of green cotton.
Weaver and Reddy (1977) studied the effects of frego, red stem, and male sterile cottons on activity of boll weevils. As a result, they suggested a system whereby 80-90% of a field is plant- ed to frego bract, red leaf cotton and the remaining 10-20% to

84 L N. NAMKEN ETAL
green, male sterile, super okra leaf cotton, which the weevils should prefer. The male sterile cotton would act as a biological trap (reduced adult emergence); in addition, insecticides could be applied to the green strips. Also, in another experiment, they harvested significantly more frego cotton, red stem strain, than frego cotton, green strain (515 v. 370 pounds per acre), and found no significant difference in yields when no insecticides were used (979 V. 960 pounds per acre). These data show that even though frego bract does have boll weevil resistance, large populations of weevils can reduce yields of frego bract cotton significantly.
There has been a systematic search for other sources of boll weevil resistance in cotton. Buford et al. (1968) observed slight reductions in oviposition of weevils exposed to 26 of 252 cotton strains taken mostly from the regional collection. A strain of Gossypium barbadense L., Sea Island (Sl) Seaberry, was the most resistant. Jenkins et al. (1969) confirmed these reductions in small field-plot tests and also found that five frego, four red leaf, and several SI strains showed resistance. Day neutral selec- tions from race stocks of Gossypium hirsutum L. T-25, T-80, and T-209 were reported by McCarty et al"! (1977) as carrying low levels of boll weevil resistance. Also, lines with genomes from Upland and cytoplasms from G^. arboreum L. , G_. herbaceum L. , and G^. anomalum Wawra ex Wawra & Peyr. reduced boll weevil oviposition about 15% (McCarty 1976), probably because these three lines have fewer anthers.
However, primitive races of G^. hirsutum are especially promis- ing as a source of resistance that could be bred into cultivated cottons. In a study of boll weevil oviposition on 191 of such ac- cessions, 69 showed significantly less boll weevil oviposition than commerical cultivars (Jenkins et al. 1978). Most of these lines are photoperiodic in their flowering response, so Jenkins is pres- ently converting them to day neutrality. Then the lines can be evaluated in field plots and other tests for resistance to boll weevil and other cotton insects.
Resistance to the Heliothis Complex
Breeding for resistance to boll weevil cannot be approached apart from proper consideration of other cotton insects. In areas infested by the boll weevil, this usually will involve Heliothis spp. and species of plant bugs.
Glabrous cotton plants are less preferred for oviposition, and nectariless plants that may be a poorer source of food (Lukefahr and Rhyne 1960, Lukefahr et al. 1965, 1971) have shown resistance ^^ Heliothis spp. Also, various types of larval antibiosis due to naturally occurring compounds have been reported (Lukefahr and Martin 1966, Oliver et al. 1967, Lukefahr and Houghtaling 1969). High terpenoid (commonly called high gossypol) is the most re- searched of these compounds, but tannins and related compounds have

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 85
recently been suggested as sources of resistance (Stipanovic et al. 1977, Waiss et al. 1977). Fourteen lines, mostly from the £. hir- sutum race collection, reduced growth of larvae relative to growth on Deltapine 16 though no causal factor was identified (Parrott et al. 1978). These cottons are available as breeding materials.
On the other hand, frego bract, which is resistant to the boll weevil, is more susceptible to plant bugs, and cotton with a high content of gossypol has increased susceptibility to thrips (Thripi- dae) of various species.
Even glandless cottons show increased susceptibility to Helio- this spp. However, Meredith et al. (1979) identified nine gland- less strains (apparently related) that were no more susceptible than Stoneville 7A glanded; and Oliver et al. (1970) found a wide range of oviposition and damage in glandless lines when 12 isogenic glandless and glanded lines were grown in field plots. Thus, sig- nificant resistance to Heliothis spp. is present in glandless cot- tons., but these lines are usually more susceptible to tarnished plant bugs. In contrast, Meredith et al. (1979) showed that when glandless was combined with nectariless, hirsutenèss, or rapid fruiting ability, susceptibility to plant bugs was reduced.
These problems may now be possible of solution. Methods of producing uniform infestations of Heliothis spp. on thousands of progeny rows (Hall et al. 1980, Jenkins et al. 1982) have recently been made available. These procedures should permit rapid evalua- tion of germplasm and should expedite the development of cottons that are resistant to Heliothis spp.
Resistance to Tarnished Plant Bugs
The nectariless cotton cultivars that are now available to growers have a relatively high degree of resistance to plant bugs. For example, Meredith et al. (1973) found that nectariless strains reduced populations of these pests about 60%; and their results were confirmed by Schuster et al. (1976), who also found reduced populations of fleahoppers (Pseudatomoscelis seriatus (Reuter)) on nectariless. This nonpreference may reflect the fact that almost 45% of the feeding time of tarnished plant bugs on cotton is spent on squares and that an additional 25% is spent at leaf nectaries (Latson et al. 1977).
However, additional sources of resistance to plant bugs are needed because the nectariless trait alone does not give adequate protection when populations of this insect are high, so research is continuing. Timok 811 (Jenkins et al. 1977) crosses involving 9.' hirsutum race stocks (Jenkins and Parrott 1976) and cottons from Bulgaria (Jenkins and Parrott 1976, Lambert et al. 1980) have been reported to show resistance, and these germplasm sources have been released for use by breeders. Pubescence too has been

86 L. N. NAMKEN ET AL
implicated in resistance (Meredith and Schuster 1979). The inheri- tance of plant bug resistance is simple enough so that progress can be expected to proceed at a fairly rapid rate (Meredith and Laster 1975, Jenkins et al. 1977).
Jones (1972), Waddle (1972), and Jenkins et al. (1973) report- ed increased susceptibility of frego cottons to tarnished plant bugs and other plant bugs. These workers thus concluded that even if a high degree of resistance to boll weevils was present in frego cottons, the increased sensitivity to fleahoppers and to tarnished plant bugs must be overcome before the trait could be potentially useful.
Also, glabrous cottons are more susceptible to tarnished plant bugs than are normal cultivars. Adding the nectariless trait to these cottons will increase their resistance to plant bugs, but they are still less resistant than normal cultivars. Bailey et al. (1980) concluded that frego or glabrous cottons, even after the ad- dition of the nectariless character, will probably have formidable problems with plant bugs in the Mississippi Delta. Although breed- ers are attempting to find other sources of plant bug resistance that can be combined with nectariless, the sensitivity of frego and glabrous cottons to plant bugs remains a consideration.
RAPID FRUITING-EARLY MATURITY POTENTIAL
Rapid fruiting and early maturity have long been perceived by cotton breeders as traits that would help growers cope with pest insects, particularly the boll weevil (Parencia 1978). The impor- tance of these characteristics was appreciated first when the boll weevil invaded the United States in the early 1900's. They became important again in the mid to late I960's and 1970*s when insect resistance to organophosphate insecticides was seen as a serious problem. Low input, short-season cotton management systems coupled with appropriate insect management practices are now recognized by plant breeders, agronomists, and entomologists as a promising method of noninsecticidal control. Rapid fruiting cotton genotypes could increase the probability that a crop would mature early enough to escape much of the heavy midseason to late-season insect pressures experienced in many cotton producing areas.
Cultivar Fruiting and Fiber Characteristics
One of the striking differences between the more recently re- leased cotton cultivars and the older varieties selected in the Delta is the much faster flowering rate of the early maturing types, particularly during the early part of the bloom period (Walker and Niles 1971, Namken and Heilman 1973, Gerard and Reeves 1975, Namken et al. 1975, Wolfenbarger et al. 1979).

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 87
Namken et al. (1975) found that Tamcot SP-37 produced 1.7 times more blooms during the first 20 days of the bloom period than did Stoneville 213, a difference they attributed to the sig- nificantly shorter vertical and horizontal flowering intervals for Tamcot SP-37.
Additional data (L. N. Namken unpublished data) supporting earlier findings are shown in Figure 2A. By June 1, 1977, Tamcot SP-37 had produced nearly twice as many cumulative blooms per acre as Stoneville 213; and GH-11-9, an experimental strain selected from Tamcot SP-37, had produced 2.5 times more blooms per acre than Stoneville 213 and 1.3 times more blooms per acre than Tamcot SP- 37. Some of this accelerated production by GH-11-9 and Tamcot SP- 37 is explained by earlier first flowering (5-7 days earlier). However, regression of rate of fruiting against time showed in- creased rates of bloom. For example, when one eliminates the ef- fect of earliness of first bloom, Tamcot SP-37 produced 1.4 times more blooms than Stoneville 213 during the first 20 days of bloom, and GH-11-9 produced 1.4 and 1.9 times more blooms than Tamcot SP- 37 and Stoneville 213, respectively. Yields of lint cotton fol- lowed the same trends. At 123 days from planting (July 8), yields were 513, 329, and 89 pounds of lint per acre for GH-11-9, Tamcot SP-37, and Stoneville 213, respectively. These yields represented 58, 53, and 14%, respectively, of the total yields for the three cottons.
Strain GH-11-9 was also compared with Tamcot SP-37, Stoneville 213, and Stoneville 825 in 1978 and 1979 at Weslaco, TX (L. N.
Namken unpublished data). The results from the four-row trials are presented in Table 7. Cotton yields were generally high in south Texas in 1978; yields in 1979 were relatively low, but some part of the low yield from the 1979 test field could be attributed to a moderate infestation of nematodes in the test field. Nevertheless,
Table 7.—Yields of lint cotton for three cotton genotypes in 1978 and 1979 variety tests at Weslaco, TX
Variety Lint cotton yields (pounds per acre) J^/
or 1978 2/ 1979 3/
strain 130 days Final 130 days Final
GH-11-9 1211 a 1398 a 458 a 761 a Tamcot SP-37 1090 b 1225 ab 204 b 398 b Stoneville 213 673 c 984 c 61 b 337 b
_!/ Means followed by the same letters are not significantly dif- ferent at the 0.05% level according to Duncan's multiple range test.
2J Means of 4 replications. 3/ Means of 3 replications.
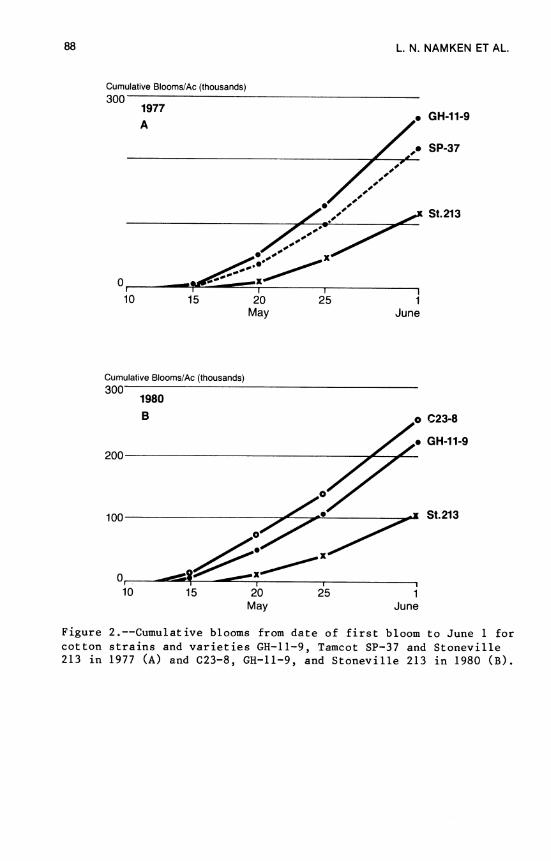
88 L N. NAMKEN ET AL
Cumulative Blooms/Ac (thousands)
300"
Cumulative Blooms/Ac (thousands)
300"
200
100
Figure 2.—Cumulative blooms from date of first bloom to June 1 for cotton strains and varieties GH-11-9, Tamcot SP-37 and Stoneville 213 in 1977 (A) and C23-8, GH-11-9, and Stoneville 213 in 1980 (B).

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 89
an important aspect of these findings was that the yields followed the same trends in both years; that is, each held the same rank in terms of portion of the total crop harvested at 130 days. Thus, there is potential for further increases in earliness and rapid fruiting.
Poor fiber quality, that is, short staple length, low micro- naire readings, and low tensile strength, has been a problem with many short-season cotton cultivars, particularly in the Southwest Region where short-season production is more practiced. (in many areas, grower acceptance of the newer short-season cottons has been slow because buyers are reluctant to offer forward contracts for the lint of these cottons.) Thus, researchers have been encouraged to investigate the relationship between rapid fruiting and fiber quality.
At Weslaco, TX, (L. N. Namken unpublished data) in 1977, crosses were made between rapid fruiting selections and cottons with high lint quality. Seed from F progeny rows of selections that combined rapid fruiting with high lint quality were then com- posited and put into a replicated, two-row test in 1980. The ex- perimental strain, GH-11-9, McNair 220 (a midseason cultivar), and Stoneville 213 (a full-season cultivar) were the checks. Earliness of the new strains was then checked by comparative testing. Re- sults (cumulative blooms from first bloom to June 1) for the exper- imental strain C23-8 and for GH-11-9 and Stoneville 213 are presented in Figure 2B. The trends are similar to those obtained in the 1977 comparisons (Figure 2A): strain C23-8 had an increased bloom rate compared with GH-11-9, which had the highest bloom rate in the 1977 test.
The next step was a comparison of the yields of lint and the fruiting characteristics of five rapid fruiting experimental strains and two checks, McNair 220 and Stoneville 213. The re- sults are presented in Table 8. (One of the experimental strains, GH-11-9, has already been described.) The other four strains were F, selections from CAMD-E x Paymaster 4298 and Tamcot SP-37 x Pay- master 4298 crosses. The four F, strains and GH-11-9 had signifi- cantly earlier bloom, significantly more cumulative blooms by June 1, and significantly shorter vertical flowering intervals than either McNair 220 or Stoneville 213. However, the C23-8 strain also had significantly higher yields at 129 days from planting than any other entry in the test. Since this strain did not differ sig- nificantly from the other experimental strains in any of the earli- ness indexes (Table 8), the significantly increased yield at this time must be attributed to some other factor. (in this regard, we note that earliness indexes reflect potential, and yield data re- flect how much of the potential a particular cotton reaches in a given environment.) Also, the C23-8 strain had significantly higher final yield than any entry except the C27-5 strain.

90 L N. NAMKEN ET AL
Table 8.—Fruiting characteristics and yields of lint cotton for five rapid fruiting strains and for McNair 220 and Stoneville 213, Weslaco, TX, 1980 1/, 2/
Lint cotton Cumulative yie lids
Variety Mean days Mean blooms (pounds per acre) or to first VFI 3/ per acre 129
strain bloom (days) by June 1 days Final
C23-8 68.5 a 2.01 a 254,700 a 753 a 1171 a
C27-5 69.5 a 1.98 a 248,200 a 558 b 1009 ab C27-1 68.5 a 1.98 a 264,000 a 596 b 984 b C26-6 69.5 a 1.81 a 256,300 a 564 b 988 b
GH-11-9 70.5 ab 1.99 a 224,300 a 557 b 921 b
McNair 220 72.0 b 2.52 b 141,300 b 470 b 946 b
Stoneville 213 72.5 b 2.51 b 109,700 b 288 c 888 b
Ij Means of 4 replications. 2/ Means followed by the same letters are not significantly dif-
ferent at the 0.05 probability level according to Duncan's multiple
range test. 3/ Vertical flowering interval.
The fiber properties of the same seven entries are summarized in Table 9. The GH-11-9 selection, which generally has a rela- tively low micronaire reading and has fiber strength similar to its
Table 9.—Properties of lint of five rapid fruiting strains and of McNair 220 and Stoneville 213, Weslaco, TX, 1980 1/, 2/
Micro- Strength Unifor-
Variety Lint Length naire (1/8-inch mity
or strain (%) (inches) reading gage) (m/uhm)
C23-8 33.9 be 1.09 e 4.8 ab 20.6 cd 82.9 be
C27-5 35.8 a 1.17 a 4.2 cd 20.8 bed 81.9 c C27-1 33.0 c 1.11 de 4.3 c 23.4 ab 82.0 c C26-6 34.6 ab 1.13 be 4.0 de 23.3 abc 84.4 ab
GH-11-9 34.7 ab 1.11 cde 3.9 e 19.2 d 83.1 be
McNair 220 34.8 ab 1.13 bed 4.6 b 25.0 a 85.7 a
Stoneville 213 34.3 ab 1.14 b 5.0 a 25.0 a 84.8 ab
Ij Means of 4 replications. 2j Means followed by the same letters are not significantly
different at the 0.05 probability level according to Duncan's multiple range test.

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 91
parent, Tamcot SP-37, was nevertheless within the acceptable range (3.9); however, it had the lowest micronaire reading of any entry in the test. Three of the four F, strains had significantly higher micronaire readings and two (C27-1 and C26-6) had signifi- cantly higher fiber tensile strength than GH-11-9. There were significant differences in fiber length among the entries, but all had fiber length in the acceptable range.
Thus, the results of the Weslaco tests indicate that simultan- eous improvement of rapid fruiting and fiber quality of cotton is possible.
Impact of Rapid Fruiting on Boll Weevil Populations
Walker and Niles (1971) described the value of rapid fruiting cotton genotypes in the management of the boll weevil and to in- creased production efficiency. First, rapid fruiting increases the probability that an acceptable crop will be set before weevil in- festations reach damaging levels. (This probability is increased if management practices are used to reduce populations of overwin- tered weevils that enter the fields at pinhead to one-third-grown square stages of plant development.) Second, earlier maturity al- lows for earlier stalk destruction which, in itself, reduces the number of overwintering weevils.
They therefore concluded that damaging infestations of weevils will not occur until after the second generation emerges if over- wintered populations of female weevils are no higher than 20 per acre. (As few as 60 female weevils per acre would be a damaging first-generation infestation.) They also established that the time of emergence of the first and second generations is predictable; that is, the second generation will emerge approximately 45 days after the crop first reaches the one-third-grown square stage or about 30 days into the bloom period. Then they used the data for the rapid fruiting genotypes available at the time to develop a model that showed that rapid fruiting genotypes produce a majority of their cumulative blooms and set bolls in the 30-day period be- fore the second generation of weevils emerge. Furthermore, the average age of bolls of the rapid fruiting genotypes is greater (lower percentage of bolls younger than 12 days) at this time than the average age of bolls of the long-season genotypes. (These had fewer bolls after 30 days of blooming, and a greater percentage of the total fruit was less than 12 days old.)
Studies by Walker et al. (1977) and Parker et al. (1980) have since indicated that bolls 12 days or older are not as susceptible to weevil damage as younger bolls. Thus, it is important to obtain a high percentage of 12-day-old or older bolls before the second generation of weevils emerge. Otherwise, acceptable yields cannot be obtained without intensive late-season control.

92 L N. NAMKEN ETAL
Further development of rapid fruiting cultivars may thus have a significant impact on the efficiency of future short-season cot- ton production systems. The following are some of the goals: (1) a higher percentage of total bolls set in the first 20-25 days of the bloom period, (2) reduction in the total number of insecticide applications needed, and (3) earlier harvest and stalk destruction to further suppress populations of overwintering boll weevils.
SHORT-SEASON COTTON MANAGEMENT SYSTEM
An integrated short-season cotton production system for south Texas was field tested in 1976, 1977, and 1978 (Heilman et al. 1977, 1979). In each year, 10- to 15-acre fields (seven in 1976, five in 1977, and four in 1978) were selected where a short-season production system could be compared with a conventional production system. Test sites had either medium- or fine-textured soils so the impact of soil, climate, and insect populations could be con- trasted. Short-season cultivars (Tamcot SP-37 and Tamcot CAMD-E), a midseason cultivar (McNair 220), and a full-season cultivar (Stoneville 213) were planted in late February or early March each year. Fields were listerbedded, preirrigated, or irrigated for seed germination and treated with a preplant herbicide. Nitrogen applications were approximately one-half (30 pounds per acre) the rate commonly used by south Texas cotton growers. A maximum of one postplant irrigation was used on the medium-textured soil and no more than two on the fine-textured soils; the timing and the number depended on visual evaluation of crop stress during the fruiting period.
In 1976 and 1977, grower fields of Stoneville 213 adjacent to each test site were used as checks for yield and production inputs (pesticides, irrigation, fertilizer). In 1978, yields and produc- tion inputs at the test sites were compared with means for growers of Stoneville 213 who were participating in the Integrated Pest Management Program of the Texas A&M University Extension Service.
With short-season cotton production, pest management is impor- tant because bolls on the bottom and in the middle of the plant must develop unharmed to achieve maximum production. Therefore, in these tests, 0.25 pound azinphosmethyl per acre was applied when approximately 25-50% of the plant population had first pinhead-size squares. A second application of azinphosmethyl (0.33 pound per acre) was made 7-10 days later, at or just before the first one- third-grown square was observed. The primary purpose of these early-season applications was to reduce the overwintering boll wee- vil population so damaging infestations would not be present until after the second generation emerged, which is about 30 days after first bloom in south Texas. The early applications also gave at least partial protection from plant bugs (particularly fleahoppers in south Texas) during this critical stage of plant development. (No additional insecticide was needed for fleahopper control at any of the test sites during the 3-year period.)

1976 4/18 1977 kll^ 1978 5/1 Mean
PLANT RESISTANCE AND MODIFIED COTTON CULTURE 93
Table 10.—Average dates and time intervals between second applica- tion to control overwintering boll weevils and any subsequent application to control boll weevil or bollworm-budworm later in season (Heilman et al. 1979)
Date of— Days between 2nd overwintering Next required 2nd and 3rd
Year weevil application \J application Ij applications
6/15 58 6/15 50 6/10 40
49
1/ Second application of 0.33 pound of azinphosmethyl per acre applied at one-third-grown square stage of plant growth.
2/ Next required application applied when economic threshold reached for boll weevil or Heliothis spp. complex.
The average dates of applications of insecticides are shown in Table 10. By June 15, the latest date for the third application, second-generation boll weevils were just about to emerge so this first postbloom application was often applied for control of the Heliothis spp. complex rather than for control of boll weevils. However, at times, control of both insect pests was needed. The applications therefore controlled insect problems during the first 30-35 days of the bloom period. In addition, populations of bene- ficial insects monitored at the test sites following the two early- season applications were generally greater than those in the check fields where overwintered boll weevil control measures were not applied.
The yields of lint cotton from the rapid fruiting (short- season cottons) and the longer season cottons are compared in Table 11. Generally, the faster fruiting cottons had acceptable or
Table 11.—Average yields of lint cotton 1976-78 (Heilman et al. 1979)
Yield (pounds per acre) \J Variety 1976 1977 1978 Mean
Tamcot SP-37 698 b 570 a 673 be 647 ab McNair 220 812 a 663 a 919 a 798 a CAMD-E 652 a 671 a 803 ab 709 ab Stoneville 213 619 b 490 b 685 be 598 b Grower check 646 b 525 b 554 c 575 b
1/ Mean yields followed by the same letter within each column are noT significantly different at the 0.05 level according to Duncan's multiple range test.

94 L N. NAMKEN ET AL
Table 12.—Comparison of average production inputs for short-season cotton and grower check, 1976-78 (Heilman et al. 1979)
Short- season Grower
Production inputs cotton check
Seasonal insecticide applications 6 10 Irrigations (postplant) Medium-textured soil 1 2 Fine-textured soils 2 3
Fertilizer (pounds nitrogen per acre) 30 60
better than average yield levels. Then, since a large portion of the fruits were set during the 30-day period between first bloom and emergence of second-generation boll weevils, the grower had only to protect bolls that were already set and were more than 12 days old, which is much easier than protecting squares until they reach bloom and then protecting the young bolls.
The major production inputs for the short-season test fields and the grower check fields are summarized in Table 12.
CURRENT STATUS OF THE TECHNOLOGY
Plant Resistance
No glabrous, male sterile, red plant color, frego bract, or high gossypol cultivars are presently available for grower use; one nectariless variety, Stoneville 825, is. Although breeders have germplasm sources for each of these traits and for various combi- nations of these traits, cultivars that have the traits and that give acceptable agronomic performance over wide geographic areas have not been developed. For example, frego bract strains perform as well as normal bract strains in the absence of plant bugs; how- ever, when plant bugs are present and uncontrolled, frego strains yield poorly. Breeders are thus attempting to combine frego with nectariless and other resistance to plant bugs in order to utilize the frego bract resistance to boll weevils.
Rapid Fruiting-Early Maturity
A number of rapid fruiting (short-season) cultivars are available to growers, but performance varies in different areas so individual State variety yield trials are the best guide to per- formance. The Tamcot, CAMD, Cascot, Lankart, Paymaster, and G&P short-season types were developed in Texas and are primarily adapt- ed to the Southwest Region. Their performance in the Southeast and

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 95
Midsouth Regions has been highly erratic, an indication of a lack of adaptation to environmental conditions in these regions.
The primary advantage of the rapid fruiting genotypes is that they have proved to be very productive in high management level production systems (both cultural and pest management). The major problems include marginal lint quality and some evidence of a lack of drought tolerance in a number of the low rainfall dryland pro- duction areas of the Southwest.
Modified Cotton Production System
From data presented in this chapter and from the studies of Walker and Niles (1971), Walker et al. (1976), and Parker et al. (1980), an integrated cotton production system is an efficient way to grow cotton in many of the boll weevil infested areas of the Southwest Region. This system should combine: (1) rapid fruiting genotypes with a pest management program designed to minimize the damaging effects of the boll weevil, and (2) agronomic practices designed to hasten maturity so as to escape much of the late-season Heliothis spp. infestation. This type of production system would not necessarily be as effective in the other regions of the Cotton Belt where boll weevils occur. However, similar systems may be de- veloped in the future. We need more efficient rapid fruiting geno- types adapted to these regions; and we need to incorporate more pest resistance into the rapid fruiting genotypes. We also need to develop improved agronomic practices including the use of bioregu- lators to control plant fruiting and vegetative growth. Finally, we need more information on insect dynamics so pest management practices can be improved.

96 L N. NAM KEN ETAL
LITERATURE CITED
Bailey, J. C, B. W. Hanny, and W. R. Meredith, Jr. 1980. Combinations of resistant traits and insecticides: effect
of cotton yield and insect populations. Journal of Economic Entomology 73:58-60.
Buford, W. T., J. N. Jenkins, and F. G. Maxwell. 1968. A boll weevil oviposition suppression factor in cotton.
Crop Science 8:647-649.
Gerard, C. J., and S. A. Reeves. 1975. Influence of climatic conditions and moisture levels on
earliness and yields of different cotton cultivars on irrigated vertisols. Ln Proceedings, Beltwide Cotton Production Research Conference, p. 76-78.
Green, J.M. 1955. Frego bract: a genetic marker in upland cotton. Journal
of Heredity 46:232.
Hall, P. K., W. L. Parrott, J. N. Jenkins, and J. C. McCarty, Jr. 1980. Use of tobacco budworm, eggs and larvae for establishing
field infestations on cotton. Journal of Economic Entomology 73:393-395.
Heilman, M. D., M. J. Lukefahr, L. N. Namken, and J. W. Norman. 1977. Field evaluation of a short season production system in Lower Rio Grande Valley of Texas. Iji Proceedings, Beltwide Cotton Production Research Conference, p. 80-83.
L. N. Namken, J. W. Norman, and M. J. Lukefahr. 1979. Evaluation of an integrated short-season management pro-
duction system for cotton. Journal of Economic Entomology 72:896-900.
Hunter, R. C., T. F. Leigh, C. Lincoln, B. A. Waddle, and L. A. Bariola.
1965. Evaluation of a selected cross section of cottons for re- sistance to the boll weevil. Arkansas Experiment Station, Bul- letin No. 700.
Isley, D. 1928. The relation of leaf color and leaf size to boll weevil
infestation. Journal of Economic Entomology 21: 553-559.
Jenkins, J. N. 1976. Boll weevil resistant cotton varieties. In Boll Weevil
Suppression, Management, and Elimination Technology. U.S. De- partment of Agriculture, ARS-S-71, p. 45-49.

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 97
Jenkins, J. N., F. G. Maxwell, W. L. Parrott, and W. T. Buford. 1969. Resistance to boll weevil Anthonomus grandis oviposition
in cotton. Crop Science 9:369-372.
J. C. McCarty, Jr., and W. L. Parrott. 1977. Inheritance of resistance to tarnished plant bugs in a
cross of Stoneville 213 by Timok 811. [Abstract] Ln Proceed- ings, Beltwide Cotton Production Research Conference, p. 97.
and W. L. Parrott. 1971. Effectiveness of frego bract as a boll weevil resistance
character in cotton. Crop Science 11:739-743.
and W. L. Parrott. 1976. Plant bug resistance in upland cotton. [Abstract] In. Proceedings, Beltwide Cotton Production Research Conference, p. 87.
V7. L. Parrott, and J. C. McCarty, Jr. 1973. The role of a boll weevil resistant cotton in pest manage- ment research. Journal of Environmental Quality 2:337-340.
W. L. Parrott, J. C. McCarty, Jr., and A. T. Earnheart. 1978. Evaluation of primitive races of Gossypium hirsutum L. for
resistance to boll weevil. Mississippi Agricultural and For- estry Experiment Station, Technical Bulletin No. 91, 13 p.
W. L. Parrott, J. C. McCarty, Jr,. and W. H. White. 1982. Breeding cotton for resistance to tobacco budworm; tech- niques for uniform field infestations. Journal of Crop Science 22:400-404.
Jones, J. E. 1972. Effect of morphological characters of cotton on insects
and pathogens. In. Proceedings, Beltwide Cotton Conference, p. 88-92.
J. B. Weaver, and M. F. Schuster. 1978. Host plant resistance to the boll weevil. In The Boll Weevil: Management Strategies. Southern Cooperative Series Bulletin No. 228, p. 50-73.
Jones, J. S., L. D. Newsom, and K. W. Tifton. 1964. Differences in boll weevil infestations among several bio-
types of upland cotton. Tn Proceedings, Beltwide Cotton Pro- duction Research Conference, p. 48-55.
Lambert, L., J. N. Jenkins, W. L. Parrott, and J. C. McCarty, Jr. 1980. Evaluation of 38 foreign and domestic cotton cultivars
for tarnished plant bug resistance. Mississippi Agricultural and Forestry Experiment Station, Research Report, Volume 5, No. 1, 4 p.

98 L N. NAMKEN ET AL
Latson, L. N., J. N. Jenkins, W. L. Parrott, and F. G. Maxwell. 1977. Behavior of the tarnished plant bug Lygus lineolaris on
cotton Gossypium hirsutum and horse weed Erigeron canadensis. Mississippi Agricultural and Forestry Experiment Station, Technical Bulletin No. 85, 6 p.
Lincoln, C, and B. A. Waddle. 1965. Insect-resistance of frego bract cotton. Arkansas Farm Research 15:5.
Lukefahr, M. J., and J. E. Houghtaling. 1969. Resistance of cotton strains with high gossypol content
to Heliothis spp. Journal of Economic Entomology 62:588-591.
J. E. Houghtaling, and H. M. Graham. 1971. Suppression of Heliothis populations with glabrous cotton
strains. Journal of Economic Entomology 64:486-488.
and D. F. Martin. 1966. Cotton plant pigments as a source of resistance to the bollworm and tobacco budworm. Journal of Economic Entomology 59:176-179.
D. F. Martin, and J. R. Meyer. 1965. Plant resistance to five Lepidoptera attacking cotton. Journal of Economic Entomology 58:516-518.
and C. Rhyne. 1960. Effects of nectariless cottons on populations of three
lepidopterous insects. Journal of Economic Entomology 53:242- 244.
McCarty, J. C., Jr. 1976. Evaluation of species and primitive races of cotton for
boll weevil resistance and agronomic qualities. Ph.D. disser- tation. Agronomy Department, Mississippi State university, 66 p.
J. N. Jenkins, and W. L. Parrott. 1977. Boll weevil resistance, agronomic characteristics, and
fiber quality in progenies of a cotton cultivar crossed with 20 primitive stocks. Crop Science 17:5-7.
McGovern, W. L., and W. H. Cross. 1976. Effect of two cotton varieties on level of boll weevil parasitism (Col: Curculionidae). Entomophaga 21:123-125.
Meredith, W. R., Jr., B. W. Hanny, and J. C. Bailey. 1979. Genetic variability among glandless cottons for resistance
to two insects. Crop Science 19:651-653.

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 99
Meredith, W. R. , Jr., and M. L. Laster. 1975. Agronomic and genetic analyses of tarnished plant bug tol-
erance in cotton. Crop Science 15:535-538.
C. D. Ranney, M. L. Laster, and R. R. Bridge. 1973. Agronomic potential of nectariless cotton. Journal of Environmental Quality 2:141-144.
and M. F. Schuster. 1979. Tolerance of glabrous and pubescent cotton to tarnished
plant bug. Crop Science 19:484-488.
Mitchell, H. C, W. H. Cross, W. L. McGovern, and E. M. Dawson. 1973. Behavior of the boll weevil on frego bract cotton. Jour-
nal of Economic Entomology 66:677-680.
Namken, L. N., and M. D. Heilman. 1973. Determinate cotton cultivars for more efficient cotton
production on medium-textured soils in the Lower Rio Grande Valley of Texas. Agronomy Journal 65:953-956.
M. D. Heilman, and R. G. Brown. 1975. Flowering intervals, days to initial flower, and seedling
uniformity as factors for development of short-season cotton cultivars. ^ Proceedings, Beltwide Cotton Production Research Conference, p. 80-85.
Oliver, B. F., F. G. Maxwell, and J. N. Jenkins. 1967. Measuring aspects of antibiosis in cotton lines to the
bollworm. Journal of Economic Entomology 60:1459-1460.
F. G. Maxwell, and J. N. Jenkins. 1970. A comparison of the damage done by the bollworm to glanded
and glandless cottons. Journal of Economic Entomology 63:1328- 1329.
Parencia, C. R., Jr. 1978. One hundred twenty years of research on cotton insects in
the United State«. U.S. Department of Agriculture, Agriculture Handbook No. 515.
Parker, R. D., J. K. Walker, G. A. Niles, and J. R. Mulkey. 1980. The short-season effect and escape from the boll weevil.
Texas Agricultural Experiment Station, Bulletin No. B-1315, 45 p.
Parrott, W. L., J. N. Jenkins, J. C. McCarty, Jr., and L. Lambert. 1978. A procedure to evaluate for antibiosis in cotton to the
tobacco budworm. Journal of Economic Entomology 71:310-311.

100 L N. NAMKEN ETAL
Parrott, W. L., J. N. Jenkins, and D. B. Smith. 1973. Frego bract cotton and normal bract cotton: how morphol-
ogy affects control of boll weevils by insecticides. Journal of Economic Entomology 66:222-225.
Schuster, M. F., and R. E. Anderson. 1976. Insecticidal efficacy on insect resistant cottons. Jour-
nal of Economic Entomology 69:691-692.
M. J. Lukefahr, and F. G. Maxwell. 1976. Impact of nectariless cotton on plant bugs and natural
enemies. Journal of Economic Entomology 69:400-402.
Stipanovic, R. D., A. A. Bell, and M. J. Lukefahr. 1977. Natural insecticides from cotton (Gossypium). In Host
Plant Resistance to Pests. American Chemical Society Symposium Series 62:197-214.
Waddle, B. A. 1972. The breeding behavior of the frego bract character in
cotton. In Proceedings, Beltwide Cotton Production Research Conference, p. 62.
Waiss, A. C, Jr., B. G. Chan, and C. A. Elliger. 1977. Host plant resistance to insects. ^ Host Plant Resis-
tance to Pests. American Chemical Society Symposium Series 62:115-128.
Walker, J. K., J. R. Gannaway, and G. A. Niles. 1977. Age distribution of cotton bolls and damage from the boll
weevil. Journal of Economic Entomology 70:5-8.
and G. A. Niles. 1971. Population dynamics of the boll weevil and modified
cotton types. Texas Agricultural Experiment Station Bulletin, No. 1109, 14 p.
G. A. Niles, J. R. Gannaway, R. D. Bradshaw, and R. E. Glodt.
1976. Narrow-row planting of cotton genotypes and boll weevil damage. Journal of Economic Entomology 69:249-253.
Weaver, J. B., Jr., and S. Baker 1972. Studies on boll weevil non-preference, boll rot and agro-
nomic characteristics of frego bract cotton. Iri Proceedings, Beltwide Cotton Production Research Conference, p. 60-61.
and M. S. Reddy. 1977. Boll weevil nonpreference, antibiosis, and hatchability
studies utilizing cotton lines with multiple nonpreferred characters. Journal of Economic Entomology 70:283-285.

PLANT RESISTANCE AND MODIFIED COTTON CULTURE 101
Wolfenbarger, D. A., R. H. Dilday, and J. W. Davis. 1979. Fruiting requirements of short-season cottons to avoid damage by the boll weevil in the Lower Rio Grande Valley. In Proceedings, Beltwide Cotton Production Research Conference p. 88-90.


103
Chapter 5
ENTOMOPHAGOUS ARTHROPODS
J. R. Abies Cotton Insect Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77841
J. L. Goodenough and A. W. Hart stack Pest Control Equipment and Methods Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77843
R. L. Ridgway National Program Staff Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705
ABSTRACT Entomophagous arthropods can be of great value in suppressing populations of cot- ton insects, especially Heliothis zea (Boddie) and jl. virescens (F.). Because of the particu- lar importance of naturally occurring popula- tions of predators in suppressing populations of these two important pests, this chapter fo- cuses on the efficacy of the complex of ca. 15 species of predators that regularly attack Heliothis spp. in cotton. Efficacy of these predators may be affected by: (1) density of predators and prey, (2) distribution of prey and area of the habitat searched, and (3) prey preferences and available alternative foods plus the disruptive influence of broad-spectrum insecticides and other factors as well.
MOTHZV-2, the Heliothis spp. population dynamics model, is useful in predicting the effect of various predator-prey ratios. These ratios can perhaps be used to make decisions re- garding integrated pest management on a field- by-field basis. The model may also be useful in estimating the potential value of beneficial in- sects, particularly those that may be destroyied by insecticides, and in identifying species that could be used in augmentation programs.

104 J. R. ABLES ET AL
Insecticides applied to cotton to con-
trol codistributed pests such as the boll weevil (Anthonomus grandis Boheman), plant bugs (Lygus spp.), and the pink bollworm (Pec- tinophora gossypiella (Saunders)) release Heliothis spp. from natural control by killing the entomophagous arthropods. Pest management efforts should primarily be concentrated on selective methods of controlling these codis- tributed pests.
INTRODUCTION
Although ecologists often remark on the reduced diversity of plant and animal life in modern agroecosystems, such crop units as cotton production areas are, in fact, complex systems composed of numerous interacting entities. Thousands of species of arthropods thus may be present in a cotton field at one time or another; still only 60-70 species, ca. 2%, are potential phytophagous pests. Fur- thermore, only a few key species normally cause at least 80% of all cotton losses due to pest insects (Newsom and Brazzel 1968). The other phytophagous species rarely cause significant damage because their populations are suppressed by natural limiting factors in- cluding entomophagous arthropods, i.e., predaceous and parasitic insects, mites, and spiders.
The importance of entomophagous arthropods in regulating the species of phytophagous insects that damage cotton has been recog- nized for a long time (Gorham 1847, Riley 1873). However, a few of these pests, most notably the boll weevil (Anthonomus grandis Boheman), apparently are much less affected. For example, an in- tensive search revealed that over 50 species of parasites and pred- ators, all except Ectatomma tuberculatum (Olivier) native to the
United States, attack the boll weevil (Pierce et al. 1912), but none has ever managed to give adequate control. This situation is consistent with the generally accepted theory of biological con- trol whereby an introduced pest (such as the boll weevil), which is adapted to survive in a location to which it is not indigenous, will escape natural controls in the new location. Thus, efforts are now focused on the identification of exotic parasites and predators that might be introduced for control of the boll weevil, and studies in southern Mexico and other subtropical areas have re- vealed several potential parasites (Cross and Chesnut 1971, J. R. Cate unpublished data). Introduction and possible augmentation of hymenopteran parasites for this purpose have been considered for Bracon kirkpatricki (Wilkinson) (Cross et al. 1969) and for Cato- laccus grandis (Burks) (Johnson et al. 1973). Also, the red im- ported fire ant (Solenopsis invicta Buren), an introduced insect that is generally considered a pest, has recently proved capable of

ENTOMOPHAGOUS ARTHROPODS 105
providing some biological control of local populations of boll wee- vils (sterling 1978). However, there are many potential problems involved in establishment of introduced natural enemies in an annual crop such as cotton (Price 1981), so additional approaches should be considered. For example, Bracon mellitor Say, a native parasite, has at times been an effective suppressant (Bottrell 1976, J. R. Cate personal communication). Because relatively lit- tle is known of the biology or ecology of this parasite, further work based on those studies summarized and continued by Adams et al. (1969) could provide a way of using B^. mellitor in augmentative biological control of the boll weevil.
Likewise, naturally occurring populations of entomophagous arthropods seem to have less effect on cotton pests such as the in- troduced pink bollworm (Pectinophora gossypiella (Saunders)) and plant bugs. Thus, importation of natural enemies of these pests has recently been emphasized. However, in these cases, native nat- ural enemies have not been studied in detail, and further work ap- pears warranted to determine their true value.
On the other hand, several lepidopteran pests of cotton, army- worms (Spodoptera spp.), the cabbage looper (Trichoplusia ni (Hüb- ner)), the bollworm (Heliothis zea (Boddie)), and the tobacco bud- worm (li. virescens (F.)), are attacked by complexes of entomophagous arthropods that, if undisturbed by chemical insecticides, are fre- quently capable of suppressing populations of these pests to densi- ties that cause no significant damage to cotton (van den Bosch et al. 1971, Ehler et al. 1973, van Steenwyk et al. 1976, McDaniel and Sterling 1979). These entomophagous complexes may be very diverse and extensive. For example, Whitcomb and Bell (1964) reported that over 600 predaceous species representing a total of 68 families of insects, spiders, and mites were found in cotton fields in Arkan- sas; and van den Bosch and Hagen (1966) provide a detailed list of ca. 60 species of parasites and predators of lepidopteran and other pests of cotton.
Natural populations of entomophagous arthropods are therefore of great value in suppressing certain phytophagous species in cot- ton. Furthermore, the use of entomophages through conservation, augmentation, and importation is increasingly seen as a viable ap- proach to insect control; and the use of parasites is of consider- able promise and importance. However, in the case of cotton insect management in multipest areas, the most immediate opportunities for substantial use of entomophages in the near term are most likely to be related to increased use of natural populations of predators for control of Heliothis spp. Therefore, in this chapter, we emphasize this particular interaction. Such a focus is further justified be- cause Heliothis spp. may often account for most of the cotton yield lost because of damage by insects and mites. For example, in 1976 alone, 26 million pounds of insecticides were applied to cotton for control of Heliothis spp. (Liapis 1980).

106 J. R. ABLES ET AL
POPULATIONS OF KEY PREDATORS OF HELIOTHIS SPP.
Among the numerous naturally occurring entomophagous arthro- pods, 15-20 species of the predaceous arthropods that occur in cotton are usually predominant, and evidence indicates that these are the most important predators of Heliothis spp. Some of these more common predators are listed in Table 1. The relative abun- dance and phenological occurrence of these and other predators in cotton are apparently governed by several interacting factors in- cluding geographical location and attendant climate and cultural practices, crop phenology, application of insecticides, and avail- ability and type of prey. These factors are discussed in the fol- lowing section.
Table 1.—Some common predators or groups of predators known to attack Heliothis spp. (Adapted from Ridgway and Lingren 1972)
Stage of Heliothis spp. primarily attacked
Egg Larvae (by instar) Predator 1-2 3 4-5
Hemiptera Lygaeidae (big-eyed bugs)
Geocoris punctipes (Say) x x Nabidae (damsel bugs)
Reduviolus americoferus (Carayon) x x x _R. alternatus (Parshley) x x x
Anthocoridae (minute pirate bugs) Orius insidiosus (Say) x x Orius tristicolor (White) x x
Neuroptera Chrysopidae (green lacewings)
Chrysopa carnea Stephens x x x Chrysopa oculata Say x x x Chrysopa rufilabris Burmeister x x x
Coleóptera Coccinellidae (lady beetles) Coleomegilla maculata (De Geer) x x Hippodamia convergens x x
Guerin-Meneville Scymnus spp. x x
Araneida (spiders) Araneidae (orb weavers) x x Oxyopidae (lynx spiders)
Oxyopes salticus Hentz x x Salticidae (jumping spiders) x x x x Thomisidae (crab spiders) x x

ENTOMOPHAGOUS ARTHROPODS 107
Geographical Location and Cultural Practices
The differences in predator populations among geographical areas certainly reflect differences in climate and probably differ- ences in local cultural practices and phenotypic differences among varieties of cotton. Table 2 is presented to show the percentage relative abundance of four major predators in three diverse cotton growing areas of the United States. In California, the predaceous Hemiptera, Orius and Geocoris, are generally more abundant and spi- ders are less abundant than in the other locations. Also, Collops and Notoxus beetles tend to be more abundant in the arid Southwest and in eastern areas when rainfall is below normal (J. R. Abies un- published data). Indeed, Gonzalez et al. (1977) reported that pop- ulations of all predators were much greater in California than in other locations.
Likewise, the abundance of predaceous arthropods appears to be affected by differences among cotton varieties. For example, Shepard et al. (1972) demonstrated that field populations of preda- tors were more abundant on early maturing glabrous and Stoneville cottons than on a pilose variety, and Schuster et al. (1976) found that populations were 17-35% lower on nectariless cottons than on cottons with extrafloral nectaries. However, despite the numerous commercial varieties of cotton available to growers, ca. 80% of the total U.S. cotton acreage in 1979 was planted to ca. seven major varieties. Thus, regionwide acceptance of a limited number of spe- cific varieties, though they do have different agronomic character- istics, may confound the factors responsible for regional differ- ences in the species composition and abundance of predaceous arthropods.
Table 2.—Relative abundance of selected predators in cotton fields in different cotton growing areas of the United States
Cotton growing % re lat ive abundance area Orius Geocor is Coccinellidae Spiders
Texas Rio Grande Valley 1/ 13 11 20 30 Brazos River Bottoms 2/ 8 22 18 20
California Indio 3/ 35 43 6 Rosedale 3/ 21 47 3
Mississippi Delta 4/ 74 11 22 30 Blackbelt 4/ 22 35 7 28
\J Schuster and Boling 1974. 2/ J. R. Abies et al. unpubl'ished data. J/ Gonzalez et al. 1977. 4/ Pitre et al. 1978.

108 J.R. ABLESETAL
Cultural practices such as irrigation, cultivation, and appli- cation of insecticides can also be expected to influence the abun- dance of entomophagous arthropods in agricultural crops such as cotton (van den Bosch and Telford 1964).
Crop Phenology, Insecticides, and Prey Availability
The pest species that are the primary source of prey for pre- daceous arthropods change in relative abundance during the cotton growing season as the cotton crop itself progresses phenologically. Generally, thrips, aphids, boll weevils, and plant bugs are early- season pests and tend to be present from the plant seedling stage to initiation of plant fruiting; boll weevils, Heliothis spp., and spider mites are predominant midseason and late-season pests. Be- cause of this seasonal shift in pest species, types and rates of insecticides may tend to change during the season though most gen- erally reduce populations of entomophagous arthropods directly by toxic effects or indirectly by depletion of the number of preferred prey. For example, the systemic insecticides that are normally ap- plied at planting or shortly thereafter to control thrips and aphids were observed to severely reduce populations of entomopha- ges (Ridgway et al. 1967). Also, a single application of an organ- ophosphate insecticide against plant bugs and other early-season pests significantly reduced the number of entomophagous arthropods in two studies (Dinkins et al. 1970, Shepard and Sterling 1972). However, populations usually returned to near their original levels in ca. 14 days if no further applications were made. (Continued applications resulted in consistently low populations.) In sum, many conventional insecticides applied to early-season cotton are deleterious to populations of entomophagous arthropods, but the ad- verse effects vary with the rate, time, frequency, and method of application.
Several studies have suggested that populations of entomoph- agous arthropods usually decline during midseason and late season. Schuster and Boling (1974) attributed the decline to the end of a generation cycle, to a dispersal phase, or to prey reduction. Dinkins et al. (1970) suggested that late-season applications of insecticides were responsible. Smith and Stadelbacher (1978), after 3 consecutive years of study in untreated fields, concluded that peak populations of predators coincided with peak squaring of cotton and considered the seasonal decline of predators as natural. Yokayama (1978) provided further evidence that plant phenology and attendant changes in the nutritional value of the cotton plant may result in changes in populations of entomophages. Nevertheless, drift of insecticide from nearby treated fields, insecticidal de- pletion of entomophages in reservoir crops, and entrapment of im- migrating predators in treated cotton fields, especially in areas of extensive acreage that undergo treatments at 3- to 5-day inter- vals, likely contribute to the decline.

ENTOMOPHAGOUS ARTHROPODS 109
EFFICACY OF PREDATORS IN SUPPRESSION OF HELIOTHIS SPP.
Methods of Evaluating Efficacy
Determination of the effects of predators on a prey population is among the most challenging problems of population ecology stud- ies. For example, destruction of a large part of the prey popula- tion may or may not affect net survival of prey because other factors may cause mortality in the population of the prey. How- ever, there is some degree of irreplaceable mortality due to pred- ators, and its relationship to net prey mortality must be deter- mined (Kiritani and Dempster 1973).
Two general methods, correlative (indirect) and experimental (direct), are widely used to assess the efficacy of predators. The correlative method mainly involves statistical methods of relating life table and density data of predators and prey to depict a cause-and-effeet relationship for the changes in density of the two populations (Stehr 1975). Proponents of this technique contend that predator efficiency can be evaluated without disturbing the ecosystem under study. If one uses both deterministic and stochas- tic processes, correlative methods allow rapid assessment. Fur- thermore, correlative methods appear more amenable to the eventual development and use of decision-making index charts and provide a convenient method of field testing or model validation. However, the methods are inferential, and the results may be misleading (Huffaker and Kennett 1969) because extrapolation beyond the bounds of existing data is usually required.
The experimental method often involves the use of paired com- parison analysis of plots that have natural enemies with plots that lack natural enemies. Usually, the natural enemies are removed through exclusion techniques (chemicals, cages, hand removal), a process that can disrupt the system being studied. A useful, but onerous alternative method is direct observation of the frequency of prédation by a particular predator upon a specific prey (Whit- comb and Bell 1964).
Perhaps neither method of evaluation is adequate in itself for assessing the role of predators in the suppression of Heliothis spp. on cotton, though both have advantages. A complex of preda- tors such as that associated with Heliothis spp. is easier to study by the correlative method because it can often be used to infer the impact of each predator species on the prey population. On the other hand, experimental methods can provide basic biological in- formation that serves as a basis for developing functional models and can provide a means of obtaining data that may have been masked in a correlative method.

110 J. R. ABLESETAL
Experimental Data
Since the work of Quaintance and Brues (1905), there has been little question about the importance of predators of the Heliothis spp. complex in cotton, and researchers have repeatedly demon- strated that the more efficient predator species have a significant effect on populations of this pest. For example, Fletcher and Thomas (1943), in a 7-year study, demonstrated that arthropod pred- ators consumed 15.3-32.9% of the eggs produced by the bollworm and in addition 12.6-59.6% of the first-instar larvae present. Bell and Whitcomb (1962) likewise reported 6-38% prédation of Heliothis spp. eggs in cotton, and Whitcomb (1967a, 1967b) reported that predators in cotton consumed 12-26% of Heliothis spp. eggs and 25-56% of their larvae. Lingren et al. (1968) found that the more effective predator species consumed 27-130 Heliothis spp. eggs per predator per day when confined with prey in petri dishes. They also reported a 76% reduction in tobacco budworms in field cages and a 96% reduction in Heliothis spp. larvae when Chrysopa carnea larvae were introduced at a rate of 420,000 per acre. Likewise, when Lopez et al. (1976) released C. carnea into field cages at a rate of 100,000 per acre, the population of tobacco budworms was reduced to 133 larvae per acre compared with 3491 per acre in the control cages. In California, naturally occurring predators de- stroyed 66-90% of the bollworm larvae present in field cages (van den Bosch et al. 1969) and 96% of bollworm eggs when predators were confined with prey on cotton terminals (Tejada 1971). Recently, McDaniel and Sterling (1979) used an improved radioactive labeling technique to determine that ca. 15 species of predators collec- tively preyed on 87-93% (seasonal average) of Heliothis spp. eggs, thereby preventing significant damage to cotton by the pest.
Despite the knowledge thus gained concerning the biology, ecology, and behavior of the species of predaceous arthropods asso- ciated with cotton, there remains a tendency to define their bio- logical characteristics vaguely and to group them as beneficial in- sects. We are therefore concerned in this chapter to recognize and to define the differences among the more important species. Eight major predators or predator groups that attack Heliothis spp. have been selected as most important on the basis of several years of sampling of natural populations in central and south Texas. In ad- dition, there is general agreement about their effectiveness as key predators, and our selections are quite similar to those selected for study in the Boll Weevil Eradication Trial in North Carolina and the Optimum Pest Management Trial in Mississippi. By using data on prey consumed per predator per day found in available literature or, when data were lacking, by using intuitive judgment, we de- veloped a set of "normalized relative efficiency indices" for some of these groups (Table 3). The larger numbers indicate relatively higher efficiency (their use in computer modeling will be discussed in relation to decision making). The groups are by no means com- plete; that is, they do not include all predators, and other in- sects such as ants may be very important in certain areas of the

ENTOMOPHAGOUS ARTHROPODS 111
Cotton Belt. Furthermore, eventual recognition and quantitative evaluation at the species level are needed.
Plainly, the results of the various studies of predator effi- ciency that were used were quite variable because of differences in sampling techniques, methodology, and the temporal aspects of each study. Furthermore, conclusions were sometimes made without regard to densities of predators and their prey, variables that are basic to the dynamic interaction between predator and prey populations. Nevertheless, an understanding of these and other factors that de- termine the efficacy of predators in suppressing Heliothis spp. is essential to successful use of these natural controls in integrated pest management (IPM).
Table 3.—Relative efficiency indices for prédation on Heliothis spp. eggs and larvae by eight predator groups i/
Index for Index for egg prédation larval prédation
Predator factor factor
Scymnus 0.21 0.22 Other Coccinellidae 2/ 2.70 0.69 Other Coleóptera 3/ 0.23 0.086 Orius 0.31 0.40 Geocoris 0.92 2.30 Other Hemiptera 4/ 0.69 1.00 Chrysopa 2.50 2.90 Spiders 0.49 0.35
\J Original data (consumption of eggs or larvae per predator per day) were adjusted to reflect factors for each predator group rela- tive to each other (i.e., a higher value represents a higher effi- ciency); due to normalization, the total for all factors is eight, which is the total number of predator groups.
2j Primarily the genera Hippodamia, Coleomegilla, and Cycloneda. V Primarily Collops spp., Notoxus spp., and small Carabidae. k_l Primarily Nabidae, Reduviidae, Pentatomidae (Asopinae), and
Miridae.
FACTORS AFFECTING PREDATOR EFFICACY
The extent to which predator populations can exploit popula- tions of prey is determined mainly by the density of the predators and their ability to locate and consume prey. Thus, one must dis- tinguish between factors affecting predator abundance and those af- fecting predator efficiency (Hassell 1978). Earlier, we reviewed some of the factors that determine predator abundance. Some of them, for example, climate and actual predator abundance, also

^^^ J. R. ABLES ETAL
affect predator efficiency. In addition, predator efficiency is affected by (1) the interaction between prey density and predator density and their rates of increase, (2) the interaction between prey distribution and habitat area searched, and (3) the interac- tion between prey preferences and alternate foods.
Predator and Prey Density
One intuitively expects that a predator population will find and consume more prey as the population of the prey or the popula- tion of the predator increases. Thus, Solomon (1949) proposed the term "functional response" to describe the change in rate of préda- tion as prey density increases and the term "numerical response" to describe the changes in the density of predators as the density of prey changes. He further stated that a natural enemy must be den- sity dependent and must take a greater percentage of the prey popu- lation as prey density increases if it is to control the prey popu- lation effectively.
The results of both empirical and theoretical studies suggest that most predaceous arthropods do not exhibit a really strong density-dependent functional response to prey populations. They generally show a decelerating increase in rate of prédation (to a limited maximum) as prey density increases. Such a response re- flects the behavioral and physiological activities associated with prédation such as killing, eating, and digestion of prey; as these activities increase, the time spent in search of prey decreases. Also, the numerical response depends upon predator developmental response, rate of survival, and adult fecundity. Additionally, there is a lower threshold of prey consumption below which the predator cannot reproduce (Hassell 1978).
According to theory, the characteristic functional response of predaceous arthropods will lead to increasing or widely fluctuating densities of prey (Solomon 1949). Despite these apparently un- stable interactions, several factors may produce stable prey popu- lations (Price 1975, Hassell 1978). For example, the predators that attack Heliothis spp. in cotton are generally polyphagous and feed on several types of prey that are distributed nonrandomly. Predators therefore might conceivably switch to the more abundant species of prey that may be disproportionately attacked by individ- ual predators and the predator population as a whole. Such behav- ioral switching could have a short-term lag effect on pest popula- tions to the point of producing stability.
After analyzing several years of data on populations of preda- ceous arthropods and Heliothis spp. eggs and larvae in cotton (j. R. Abies and A. W. Hartstack unpublished data), we are unable to discern any regular patterns of increase or decrease of preda- ceous species relative to the number of Heliothis spp. present. This failure to achieve close correlations is not surprising be- cause the predator populations were generally sampled by the D-Vac

ENTOMOPHAGOUS ARTHROPODS 113
method, which has inherently great variability, and because of the predators' general lack of prey specificity. However, J. R. Abies (unpublished data) determined that increased numbers of predators result in lower numbers of Heliothis spp. larvae (Figure 1); and similar relationships have been demonstrated by comparing popula- tions of Heliothis spp. in untreated cotton fields with populations in fields where predators were depleted by treatment with chemical insecticides (Ridgway 1969, van den Bosch et al. 1971, van Steenwyk et al. 1976).
No. Heliothis larvae (x 1000) 25
1 \ r 5 10 15 20 25 30 35 40
No. predators (x 1000)'
Figure 1.—The relationship between numbers of predators per acre and number of Heliothis spp. larvae per acre. Data points represent means for July (1979) in six cotton fields near College Station, TX (J. R, Abies unpublished data).
Prey Distribution and Habitat Searched
Most theoretical appraisals of the interaction between preda- tors and prey assume a uniform distribution of prey in a uniform environment. However, Heliothis spp. eggs and larvae are not uni- formly distributed among cotton plants nor does the environment remain uniform; for example, size of individual plants varies, and the plants grow. Usually, Heliothis spp. eggs are found on the upper one-third of the cotton plant, and larvae, except those re- cently hatched, usually feed within the relative shelter of fruit- ing parts (squares and bolls) of the plant. Thus, to locate Helio- this spp., predators must not only search a growing plant habitat but must also restrict their search to specific plant locations. Knipling (1979) accounted for the expanding search area in his sim- ulation codeveloping models by assuming that the cotton plant in- creases by a factor of 1.0 every 10 days. Donahoe and Pitre (1977)

114 J. R. ABLES ET AL
documented this inverse relationship when they demonstrated that Reduviolus roseipennis (Reuter) captured fewer first-instar boll- worm larvae as plant age and size increased. They also identified the underside of leaves of the upper one-third of the plant as the major area of search by this predator.
Prey Preferences and Alternate Foods
Although the predators found in cotton may show distinct pref- erence for particular types of prey, they are generally polyphagous in their habits. For example, Abies (1978) determined that an as- sassin bug (Zelus renardii Kolenati) was most adapted to the cap- ture of flying insects but fed on various forms of at least 15 spe- cies of arthropods, 32% of which were potential cotton pests; 12% of the total diet consisted of Heliothis spp. larvae.
If a predator can feed on several prey types, then preferences or selection of prey may shift in accordance with the relative abundance of the various available types of prey (Price 1975). In fact, Abies et al. (1978) noted that prédation on tobacco budworm eggs by five species of predators was reduced in the presence of aphids. Also, many predators feed on extrafloral nectaries of the cotton plant, and the time spent feeding on such plant materials may limit time spent feeding on prey (J. R. Abies unpublished data).
When Heliothis spp. is the selected prey, different predator species and different ages of the same predator species may often show an apparent preference for prey at different stages of devel- opment. For example, Lawrence and Watson (1979) showed that first- instar Geocoris punctipes nymphs fed almost exclusively on tobacco budworm eggs; older nymphs and adults fed on eggs and larvae (pri- marily first-instar). Likewise, many predators in cotton such as Orius and Geocoris are relatively small and are able to feed only on eggs and small larvae; asopine pentatomids and vespid wasps feed primarily on larger larvae.
MODELING POPULATIONS OF ENTOMOPHAGOUS ARTHROPODS AND HELIOTHIS SPP. FOR IPM DECISION MAKING
Two general types of modeling of predator-prey systems, com- plex (component) and simple, are presently used. Published models of the complex type include those of Watt (1959) and of Holling (1966), both of which model the searching, attacking, digesting, and resting time of each species of predator. Development of com- plex models for use in cotton would sometimes require that each of the pest species, each stage of the predator, and each stage of the prey be modeled. Also, accurate estimates of host crop phenology (leaf area, height of plant, distribution of fruit as to location as well as age) would be needed. We feel that these models are far more complex than is probably necessary; however, resulting

ENTOMOPHAGOUS ARTHROPODS 115
information could be a tremendous help in developing or testing simpler models.
Simple predator-prey models have been reported by Knipling and McGuire (1968), Knipling (1979), and Hartstack et al. (1976). In these cases, the hypotheses are such that many factors considered by the complex models are simplified; also, no attempt is made to measure minute details on each predator or prey. This more sim- plistic approach is taken, at least in part, because of the enor- mous task of measuring the host population accurately and the even more difficult task of monitoring the predator populations.
The simple model proposed by Knipling and McGuire (1968) for Trichogramma, an egg parasite of Heliothis spp., and later adapted and suggested for general use in cotton models by Hartstack et al. (1975) in the dynamic Heliothis spp. model MOTHZV-2 is,
M = 1 - EXP (-0.693)(P^)
(N) (S)"" where,
M = Probability of mortality of Heliothis spp. eggs or larvae P = Number of effective predators N = Number of effective predators required to cause a 50% mor-
tality of prey S = Relative search area (1 = first square to 6 = mature
plant)
The daily rate of prédation of larvae is reduced as the larvae increase in size with age as follows:
M = (M) EXP -0.42 (A) 0.657
where A = age of larvae in days.
To improve the effectiveness and versatility of this basic model, the life histories for Heliothis spp. and each predator group were divided into different age distribution classes, and rel- ative prédation efficiency indices were assigned to each predator species or group. The end result was a flexible predator-prey sub- routine to the Heliothis spp. population dynamics model MOTHZV-2. We revised the relative efficiency indices according to latest available literature of eggs removed per predator per day (Table 3). When we then applied the MOTHZV-2 model—with modifications de- scribed by Hartstack and Witz (Chapter 14 this handbook)—to data from 19 cotton fields, we were able to predict the survival of large Heliothis spp. larvae with reasonable accuracy (Figure 2). We have also used MOTHZV-2 to determine the conceptual effects of different densities of total predators on the survival of Heliothis spp. and on subsequent plant damage and potential trends in yield. The predicted means for these parameters when we multiplied actual data on populations of Heliothis spp. from eight fields in the presence of actual populations of predators by factors of 0, 1, 2,

116 J. R. ABLES ET AL
Actual no. large Heliothis larvae
15,000 "
1 1 1 2500 7500 12,500 Predicted no. large Heliothis larvae
Figure 2.—Comparison of MOTHZV-2 computer-simulated population and actual field populations of large Heliothis spp. larvae in 19 south Texas cotton fields. Data are maximum number per acre of either predicted (x-axis) or actual (y-axis) populations before July 12, 1973.
and 4 are shown in Table 4. As the predator population was in- creased, the number of Heliothis spp. larvae and plant damage de- creased, and the total production of bolls increased significantly enough to suggest the potential of considerably higher crop yield. Note that while we use the general phrase "number of effective predators" (P in previous equation and in Figure 3), our inputs in- clude specific data on abundance and relative efficiency for each of the predators listed, in Table 3.
Hartstack et al. (1975) suggested that MOTHZV-2 might be used to include entomophages in deciding whether an insecticide should be applied. Practitioners of integrated pest management in cotton recognize the value of conserving entomophagous arthropods but presently do not have well-defined guidelines for including the presence and potential impact of entomophages in their decision- making process. A trichotomous decision chart was therefore developed. A recently modified version is presented in Figure 3. If the data on effective natural predators (the sum of field estimates times the relative efficiency values shown in Table 3, x-axis) and the number of Heliothis spp. eggs (y-axis) plot to the right of the decision line, the predator population is pre- dicted as large enough to prevent the Heliothis spp. population

ENTOMOPHAGOUS ARTHROPODS 117
Table 4,—Predicted mean maximum (X) number of large Heliothis spp. larvae and total bolls and percentage of square and boll damage in the presence of four levels of naturally occurring predators in cotton 1/
X maximum X maximum
X number number number X maximum X maximum predators large larvae bolls % damaged % damaged per acre per acre 2/ per acre squares 3/ bolls
0 14,381 41,706 27.9 21.9 9,566 4/ 3,132 84,842 5.4 5.1 19,133 1,384 107,032 2.2 1.6 38,264 556 115,257 1.2 0.4
\J Means were predicted by MOTHZV-2 model based on one to three samples per week taken in eight fields in Frio County, TX, during mid-May to mid-July 1973.
Ij Assumed threshold: 2500 large larvae per acre. _3/ Peak square production occurred in early July. 4/ Actual mean.
from exceeding an assumed threshold (action level) of 2500 large larvae per acre (Adkisson et al. 1964, Ridgway 1969, Hartstack et al. 1978). If the populations plot to the left side of the deci- sion line, predator populations are predicted as too small to ade- quately suppress the Heliothis spp. populations, and the need for corrective action is assumed. If populations plot at the approxi- mate center of the chart, the impact of predators is unpredictable, and populations should be monitored daily until a decision can be made.
A close look at the quantitative relationships apparent in Figure 3 suggests that a ratio of at least two effective predators per one Heliothis spp. egg will usually eliminate the need for cor- rective action; as the ratio decreases below this level, some type of action may be required. However, time of initiation of action should also depend on the choice of action selected. For example, action directed against the larval population resulting from eggs that have escaped prédation should not be taken until small larvae are actually found and until the recommended plant damage thresh- olds are reached. (Small larvae are also susceptible to prédation so the predators that attack them may still prevent economic losses to the crop.) Actions directed against larvae may be of several kinds (chemical or microbial insecticide or the augmentative re- lease of a larval parasite or predator), and the effective time of use may vary. On the other hand, if action is to be applied against the egg stage (chemical ovicides or the augmentative re- lease of an egg parasite or predator), the action must be made on the basis of the natural ratios of predators to eggs.

118 J. R. ABLES ET AL
Action No No. Of Heiiothis Required Undecided Action eggs (x 1000)
100
75
50 «s I \ 1 1 \ 1 1
0 10 20 30 40 50 60 70 No. of effective natural predators (x 1000)
Figure 3.—A decision-making chart showing the number of naturally occurring predators and Heiiothis spp. eggs per acre of cotton. A required corrective action is expected when the number of large Heiiothis spp. larvae reaches or exceeds 2500 per acre.
Although results of our field studies and of those of McDaniel and Sterling (1979) indicate that our predicted results for spe- cific ratios of predators to Heiiothis spp. are reasonably accu- rate, there is need for continued improvement of the predator-prey subroutine to provide greater precision and flexibility in the ap- plication of MOTHZV-2 to IPM assessments in highly variable field situations. Hartstack and Witz (Chapter 14 in this handbook) de- tail the improvements that are needed.
APPROACHES TO THE USE OF ENTOMOPHAGOUS ARTHROPODS IN IPM IN COTTON
Most of this chapter has been concerned with the impact of predators on populations of Heiiothis spp. in cotton. The role or value of these entomophages in IPM programs has been best demon- strated by those studies in which predators were eliminated by insecticides. This elimination has frequently been followed by a resurgence of key pests and by severe outbreaks of secondary pests. Furthermore, the increased use of insecticides that is then neces- sary to deal with the resulting catastrophic populations of pests has probably been a major reason for the development of insecticide resistance in such pests as the tobacco budworm, boll weevil,

ENTOMOPHAGOUS ARTHROPODS 119
spider mites, and lygus bugs (Newsom and Brazzel 1968, van den Bosch et al. 1971, Reynolds et al. 1975).
For example, in California, van den Bosch et al. (1971) demon- strated that three applications of azodrin reduced the populations of predators so severely that populations of bollworms and boll damage were greater in treated fields than in untreated fields. Also, van Steenwyk et al. (1976) reported that predator numbers were higher and bollworm populations lower in California fields treated with insecticides one to three times for pink bollworms than in fields treated six to seven times (eight to nine treatments apparently suppressed both pests). Although Heliothis spp. have been controlled by entomophagous arthropods in the absence of broad-spectrum insecticides, management programs for Heliothis are complicated by the fact that Heliothis spp. in cotton are often co- distributed with such other pests as the boll weevil, pink boll- worm, and plant bugs. When insecticides must be applied against these codistributed pests, the end result is often an outbreak of Heliothis spp. and/or other lepidopteran pests. Therefore, IPM recommendations and supporting research should be focused simultan- eously on control of the codistributed pests and conservation of the entomophages that control Heliothis spp. The advantage of any such cotton IPM strategy was demonstrated by Pimentel et al. (1980) who estimated that 40% of all insecticide costs in cotton result from the loss of natural enemies; the total added cost was esti- mated to be more than $119 million. An additional added cost due to increased resistance of pests to insecticides was estimated to be ca. $36 million.
Conservation
Conservation of entomophagous arthropods in cotton is simply the avoidance or reduction of activities that are deleterious to these natural enemies of pest insects. Perhaps the most important method of conservation is the use of selective control strategies such as host plant resistance, selective insecticides, or judicious use of certain broad-spectrum insecticides. A successful IPM pro- gram for cotton would likely include all of these methods as well as others that are ecologically sound. For example, Ridgway et al. (1977) suggest that a cotton insect management program designed for optimum use of entomophages would consist of "(1) the production of an early crop through selection of varieties and other management practices, (2) the use of resistant or tolerant varieties where available, and (3) a method of controlling injurious plant bugs and boll weevils that will have the minimum effects on natural ene- mies." These strategies should also be supported by precise deci- sion-making procedures.

120 J. R. ABLESETAL
Augmentation
Although naturally occurring entomophagous arthropods often suppress pest populations adequately, ecological restrictions may prevent numbers from increasing to an effective level. In such cases, one solution is augmentation, the deliberate increase of the populations or their beneficial effects by the release of labora- tory-reared species, the provision of supplemental hosts or other foods, the use of behavioral chemicals, or any combination of these methods (Abies and Ridgway 1981). In an appraisal of augmentative programs in the United States, Abies and Ridgway (1981) noted that the technical knowledge about the use of most entomophagous arthro- pods in ecological pest control is not adequate to allow the deter- mination of when and where augmentation (or alternatives) should be used. However, the technology is most advanced for the egg para- sites Trichogramma spp., the predator Chrysopa carnea, and other species that have been studied for use in cotton. Indeed, the benefits to be derived from augmentation are likely to be high- est when the approach is used in the type of situation common in cotton fields where insecticides must be used to control pests such as pink bollworm, Heliothis spp., and boll weevils.
Importation
Importation, the classical approach to biological control, in- volves foreign exploration for and introduction of natural enemies of insect pests that are not native to the United States and that are therefore not suppressed adequately by the native entomophagous arthropods. Because this is true for the boll weevil, the pink bollworm, and perhaps other pests such as lygus bugs, foreign ex- ploration for natural enemies of these pests is in process. Scien- tists at Texas A&M University are exploring areas of southern Mex- ico in attempts to screen potential natural enemies of the boll weevil. Also, scientists at the University of California, River- side, have for several years explored a variety of geographical re- gions and have introduced several promising species of parasites of the pink bollworm. In addition, a Federal and State cooperative program has recently been developed for the exploration and intro- duction of natural enemies of plant bugs that attack cotton. Con- tinued research and education in these different approaches will be necessary to implement the various forms of biological control in cotton.
ACKNOWLEDGMENTS
Outputs from the MOTHZV model were based on data collected by R. E. Kinzer, S. L. Jones, and D. Kizer to whom we are grateful. We thank D. W. McCommas, Jr., for preparation of the figures.

ENTOMOPHAGOUS ARTHROPODS 121
LITERATURE CITED
Abies, J. R. 1978. Feeding behavior of an assassin bug, Zelus renardii. An-
nals of the Entomological Society of America 71:476-478.
S. L. Jones, and D. W. McCommas, Jr. 1978. Response of selected predator species to different densi-
ties of Aphis gossypii and Heliothis virescens eggs. Environ- mental Entomology 7:402-404.
and R. L. Ridgway. 1981. Augmentation of entomophagous arthropods to control pest
insects and mites. Ln Biological Control in Crop Production, p. 273-303. G. C. Papavizas, editor. Allenheld, Osmun, and Company.
Adams, C. H., W. H. Cross, and H. C. Mitchell. 1969. Biology of Bracon mellitor, a parasite of the boll weevil. Journal of Economic Entomology 62:889-896.
Adkisson, P. L., R. L. Hanna, and C. F. Bailey. 1964. Estimates of the number of Heliothis larvae per acre of
cotton and their relation to the fruiting cycle and yield of the host. Journal of Economic Entomology 57:657-663.
Bell, K. 0., Jr., and W. H. Whitcomb. 1962. Efficiency of egg predators of the bollworm. Arkansas Farm Research 11:9.
Bottrell, D. G. 1976. Biological control agents of the boll weevil. In Boll Weevil Suppression, Management, and Elimination Technology. U.S. Department of Agriculture, ARS-S-71, p. 22-25.
Cross, W. H., W. L. McGovern, and H. C. Mitchell. 1969. Biology of Bracon kirkpatricki and field releases of the
parasite for control of the boll weevil. Journal of Economic Entomology 62:448-454.
and T. L. Chesnut. 1971. Arthropod parasites of the boll weevil, Anthonomus grandis: 1. An annotated list. Annals of the Entomological Society of America 64:516-527.
Dinkins, R. L., J. R. Brazzel,. and C. A. Wilson. 1970. Seasonal incidence of major predaceous arthropods in Mis-
sissippi cotton fields. Journal of Economic Entomology 63:814- 817.

122 j. R. ABLESETAL
Donahoe, M. C, and H. N. Pitre. 1977. Reduviolus roseipennis behavior and effectiveness in re-
ducing numbers of Heliothis zea on cotton. Environmental Ento- mology 6:872-876.
Ehler, L. E., K. G. Eveleens, and R. van den Bosch. 1973. An evaluation of some natural enemies of cabbage looper on
cotton in California. Environmental Entomology 2:1009-1015.
Fletcher, R. K., and F. L. Thomas. 1943. Natural control of eggs and first instar larvae of Helio-
this armigera. Journal of Economic Entomology 36:557-560.
Gonzalez, D., D. A. Ramsey, T. F. Leigh, B. S. Ekbom, and R. van den Bosch.
1977. A comparison of vacuum and whole-plant methods for samp- ling predaceous arthropods on cotton. Environmental Entomology 6:750-760.
Gorham, D. P. 1847. The cotton worm. DeBow's Commercial Review 3:535.
Hartstack, A. W., Jr., R. L. Ridgway, and S. L. Jones. 1978. Damage to cotton by the bollworm and tobacco budworm. Journal of Economic Entomology 71:239-243.
J. A. Witz, J. P. Hollingsworth, R. L. Ridgway, and J. D. Lopez.
1976. MOTHZV-2: a computer simulation of the Heliothis zea and Heliothis virescens population dynamics. Users manual. U.S. Department of Agriculture, ARS-S-127.
J. A. Witz, and R. L. Ridgway. 1975. Suggested applications of a dynamic Heliothis model
(MOTHZV-1) in pest management decision making. Proceedings, 1973 Beltwide Cotton Production Research Conference, Phoenix, AZ, January 9-10, 1973, p. 118-122.
Hassell, M. P. 1978. The dynamics of arthropod predator-prey systems. In Monograph in Population Research Biology No. 13. PrinceTon University Press, Princeton, NJ, 237 p.
Holling, C. S. 1966. The functional response of invertebrate predators to prey
density. Memoirs Entomological Society of Canada. No. 48, 86 p.
Huffaker, C. B., and CE. Kennett. 1969. Some aspects of assessing efficiency of natural enemies. Canadian Entomologist 101:425-447.

ENTOMOPHAGOUS ARTHROPODS 123
Johnson, W. L., W. H. Cross, W. L. McGovern, and H. C. Mitchell. 1973. Biology of Heterolaccus grandis in a laboratory culture
and its potential as an introduced parasite of the boll weevil in the United States. Environmental Entomology 2:112-118.
Kiritani, K., and J. P. Dempster. 1973. Quantitative evaluations of natural enemy effectiveness. Journal of Applied Ecology 10:323-330.
Knipling, E. F. 1979. The basic principles of insect population suppression and management. U.S. Department of Agriculture, Handbook No. 512, 659 p.
and J. U. McGuire, Jr. 1968. Population models to appraise the limitations and potenti-
alities of Trichogramma in managing host insect populations. U.S. Department of Agriculture, Technical Bulletin No. 1386, 44 p.
Lawrence, R. K., and T. F. Watson. 1979. Predator-prey relationship of Geocoris punctipes and Heli-
othis virescens. Environmental Entomology 8:245-248.
Liapis, P. S. 1980. Controlling Heliothis spp. on cotton through the release
of Trichogramma pretiosum and applications of Bacillus thurin- giensis: An economic assessment. U.S. Department of Agricul- ture Economics, Statistics, and Cooperative Services, Natural Resources Branch, Staff Report, 36 p.
Lingren, P. D., R. L. Ridgway, and S. L. Jones. 1968. Consumption by several common arthropod predators of eggs
and larvae of two species of Heliothis that attack cotton. An- nals of the Entomological Society of America 61:613-618.
Lopez, J. D., Jr., R. L. Ridgway, and R. E. Pinnell. 1976. Comparative efficacy of four insect predators of the boll- worm and tobacco budworm. Environmental Entomology 5:1160- 1164.
McDaniel, S. G., and W. L. Sterling. 1979. Predator determination and efficiency on Heliothis vires-
cens eggs in cotton using P. Environmental Entomology 8: TÖ83-1087.
Newsom, L. D., and J. R. Brazzel. 1968. Pests and their control. In. Advances in Production and Utilization of Quality Cotton: Principles and Practices, p. 367-405. F. C. Elliot, M. Hoover, and W. K. Porter, editors. Iowa State Universit,y Press, Ames.

124 J. R. ABLESETAL
Pierce, W. D., R. A. Cushman, and C. E. Hood. 1912. The insect enemies of the cotton boll weevil. U.S. De-
partment of Agriculture, Bureau of Entomology Bulletin No. 100, 99 p.
Pimentel, D. , D. Andow, D. Gallahan, I. Schreiner, T. E. Thompson, R. Dyson-Hudson, S. N. Jacobson, M. A. Irish, S. F. Kroop, A. M. Moss, M. Shepard, and B. G. Vinzant.
1980. Pesticides; environmental and social costs. In Pest Control: Cultural and Environmental Aspects, p. 99-158. D. Pimentel and J. H. Perkins, editors. American Association for Advancement of Science Selected Symposium No. 43. Westview Press, Boulder, CO.
Pitre, H. N., T. L. Hillhouse, M. C. Donahoe, and H. C. Kinard. 1978. Beneficial arthropods on soybeans and cotton in different
ecosystems in Mississippi. Mississippi Agricultural and Fores- try Experiment Station, Technical Bulletin No. 90, 9 p.
Price, P. W. 1975. Insect ecology. ♦ Wiley and Sons, New York, 514 p.
1981. Relevance of ecological concepts to practical biological control, Tn Biological Control in Crop Production, p. 3-19. G. C. Papavizas, editor. Allenheld, Osmun, and Company.
Quaintance, A. L., and C. T. Brues. 1905. The cotton bollworm. U.S. Department of Agriculture, Bu-
reau of Entomology Bulletin No. 50, 155 p.
Reynolds, H. T., P. L. Adkisson, and R. F. Smith. 1975. Cotton insect pest management. I_n Introduction to Insect
Pest Management, p. 379-443. R. L. Metcalf and W. Luckman, editors. Wiley and Sons, New York.
Ridgway, R. L. 1969. Control of the bollworm and tobacco budwoirm through con-
servation and augmentation of predaceous insects. In Proceed- ings, Tall Timbers Conference of Ecological Animal Control by Habitat Management, p. 127-144.
E. G. King, and J. L. Carrillo. 1977. Augmentation of natural enemies for control of plant pests
in the Western Hemisphere. In Biological Control by Augmenta- tion of Natural Enemies, p. 379-416. R. L. Ridgway and S. B. Vinson, editors. Plenum Press, New York.
and P. D. Lingren. 1972. Predaceous and parasitic arthropods as regulators of Heli- othis populations. In. Bollworm and Tobacco Budworm in Cotton. Southern Cooperative Series Bulletin No. 169, p. 48-56.

ENTOMOPHAGOUS ARTHROPODS 125
Ridgway, R. L., P. D. Lingren, C. B. Cowan, Jr., and J. W. Davis. 1967. Populations of arthropod predators and Heliothis spp. af-
ter applications of systemic insecticides to cotton. Journal of Economic Entomology 60:1012-1016.
Riley, C. V. 1873. Fifth annual report of the noxious, beneficial and other
insects of the State of Missouri, made to the State Board of Agriculture, pursuant to an appropriation for this purpose from the legislature of the State. Eighth Annual Report, State Board of Agriculture, 1872, p. 160-168.
Schuster, M. F., and J. C. Boling. 1974. Phenology of early and mid-season predatory and phytopha-
gous insects in cotton in the Lower Rio Grande Valley of Texas. Texas Agricultural Experiment Station, Miscellaneous Publica- tion No. 1133, 31 p.
M. J. Lukefahr, and F. G. Maxwell, 1976. Impact of nectariless cotton on plant bugs and natural
enemies. Journal of Economic Entomology 69:400-402.
Shepard, M., and W. L. Sterling. 1972. Effects of early season applications of insecticides on beneficial insects and spiders in cotton. Texas Agricultural Experiment Station, Miscellaneous Publication No. 1045, 14 p.
W. L. Sterling, and J. K. Walker, Jr. 1972. Abundance of beneficial arthropods on cotton genotypes. Environmental Entomology 1:117-121.
Smith, J. W., and E. A. Stadelbacher. 1978. Predatory arthropods: seasonal rise and decline of popu-
lations in cotton fields in the Mississippi Delta. Environmen- tal Entomology 7:367-371.
Solomon, M. E. 1949. The natural control of animal populations. Journal of Animal Ecology 18:1-35.
Stehr, F. W. 1975. Parasitoids and predators in pest management. In, Intro- duction to Insect Pest Management, p. 147-188. R. L. Metcalf and W. Luckman, editors. Wiley and Sons, New York.
Sterling, W. L. 1978. Fortuitous biological suppression of the boll weevil by
the red imported fire ant. Environmental Entomology 7:564-568.

126 j. R. ABLESETAL
Tejada, L. O. 1971. Evaluation of the effectiveness of some general predators
at different predator-prey densities, with special reference to their attack on Heliothis zea (Boddie). Ph.D. dissertation. University of California, Riverside.
van den Bosch, R., and K. S. Hagen. 1966. Predaceous and parasitic arthropods in California cotton
fields. California Agricultural Experiment Station, Bulletin No. 820, 32 p.
T. F. Leigh, L. A. Falcon, V. M. Stern, D. Gonzalez, and K. S. Hagen.
1971. The developing program of integrated control of cotton pests in California. In Biological Control, p. 377-394. C. B. Huffaker, editor. Plenum Press, New York.
T. F. Leigh, D. Gonzalez, and R. E. Stinner. 1969. Cage studies on predators of the bollworm in cotton. Journal of Economic Entomology 62:1486-1489.
and A. D. Telford. 1964. Environmental modification and biological control. In Biological Control of Insect Pests and Weeds, p. 459-488. P. DeBach, editor. Chapman and Hall, London.
van Steenwyk, R. A., N. C. Toscano, G. R. Ballmer, K. Kido, and W. T. Reynolds.
1976. Increased insecticide use in cotton may cause secondary pest outbreaks. California Agriculture 30:14-15.
Watt, K. E. F. 1959. A mathematical model for the effect of densities of at-
tacked and attacking species on the number attacked. Canadian Entomologist 111:129-144.
Whitcomb, W. H. 1967a. Field studies on predators of the second instar bollworm, Heliothis zea (Boddie) (Lepidoptera: Noctuidae). Journal of the Georgia Entomological Society 2:113-118.
1967b. Bollworm predators in northeast Arkansas. Arkansas Farm Research 16:2.
and K. 0. Bell, Jr. 1964. Predaceous insects, spiders, and mites of Arkansas cotton
fields. Arkansas Agricultural Experiment Station, Bulletin No. 690, 84 p.

ENTOMOPHAGOUS ARTHROPODS 127
Yokayama, V. Y. 1978. Relation of seasonal changes in extrafloral nectar and fo-
liar protein and arthropod populations in cotton. Environmen- tal Entomology 7:799-802.


129
Chapter 6
MICROBIAL AGENTS
M. R. Bell Western Cotton Insects Laboratory Agricultural Research Service U.S. Department of Agriculture Phoenix, AZ 85040
ABSTRACT Although applications of protozoan and fungal pathogens will produce infection in boll weevils (Anthonomous grandis Boheman) in the field, the pathogens act too slowly to pro- tect the plants. At present, the only micro- bial agents used on cotton are the Heliothis nuclear polyhedrosis virus and Bacillus thurin- giensis Berliner, both of which can suppress moderate to low populations or even control some large populations of the bollworm (Helio- this zea (Boddie)) and the tobacco budworm (Heliothis virescens (F.)). However, proper timing, application techniques, and special formulations are necessary. Because these mi- crobial agents are highly selective (compared with chemical insecticides), they are particu- larly useful for control of Heliothis spp. when broad-spectrum insecticides are not needed to control other insects such as the boll weevil.
INTRODUCTION
The early studies of diseases of insects were concerned al- most entirely with diseases of such useful species as the silkworm (Bombyx mori (L.)) and the honey bee (Apis mellifera L.). Then as these studies slowly spread to include diseases of other insects, the discipline of insect pathology evolved, and scientists began to think of using pathogenic micro-organisms to control pest insects. Insect pathology is therefore relatively young as a separate disci- pline of entomology. Indeed, the first international conference was held in Prague in 1958. Since that time, the growing number of scientists working specifically in this area of research has pro- duced a rapid increase in information about insect diseases, but diseases of pest species continue ta be the most studied.
Some degree of success has been achieved in the use of insect pathogens (now termed microbial insecticides) in management of pest

""^^ M. R. BELL
insects in crops, but they are presently used much less for this purpose than are chemical insecticides. The simple fact is that the potential for microbial control envisioned in the 1950*s and I960's has not been realized, not because that potential was un- real but because new methods necessary for effective use have not been developed as rapidly as anticipated. For example, at present, only two insect pathogens are registered for use on cotton in the United States, the nuclear polyhedrosis virus (NPV) from Heli- this zea (Boddie) and the bacterium Bacillus thuringiensis Ber- liner, variety HD-1 (B ._t. ). Both have been exempted by the Envi- ronmental Protection Agency from the requirement of a tolerance for residues, and treated fields may be reentered immediately after application.
Another difficulty with microbial insecticides has been lack of consistent economic control, which sometimes stems from lack of knowledge of the user. For example, a user such as a cotton pro- ducer is probably skilled in the application of chemical insecti- cides and knows what to expect from them. However, entirely new problems can arise when pathogens are used in pest management sys- tems, and both the limitations of the pathogens and their potential must be understood. Some of these problems may be specific to one insect pest on one crop; others may be inherent in the pathogen it- self. Nevertheless, laboratory and field studies conducted with a variety of microbial agents against several major cotton insect pests indicate that such problems can be solved.
The purpose of this chapter is to review the major character- istics of pathogens that are being used or are being studied for use in controlling cotton insects in the United States and to re- port the results of field trials. Hopefully, it will give those persons already familiar with cotton insect pest management a bet- ter understanding of the potential of microbial control.
GENERAL CHARACTERISTICS OF PATHOGENS IN COTTON INSECT MANAGEMENT
A pest control advisor who recommends the use of pathogens for control of insects, like those who recommend the use of chemi- cal insecticides, must have an understanding of the terms used to describe pathogens and disease processes and must be familiar with the specific characteristics of the microbial. Steinhaus and Mar- tignoni (1970) present short definitions of terms used in inverte- brate pathology (e.g., pathogen, disease, infection, incubation period, epizootic). Following are brief descriptions of the micro- bial agents most often considered for use against cotton insects.
Viruses
More than 700 species of insects and mites have been reported to harbor viruses (David 1975), but viruses in a genus termed

MICROBIAL AGENTS 131
"Baculovirus" are considered to have the greatest potential for con- trol of insects in agriculture. Baculoviruses (identified in more than 300 species of insects) are of two types, the nuclear polyhe- drosis viruses (NPV*s) and the granulosis viruses (GV's). Both have infectious units ("virions") that are composed of deoxyribonu- cleic acid and are enclosed in a capsid and surrounded by an enve- lope. In addition, NPV*s form polyhedral inclusion bodies (PIB's) in the nuclei of infected cells (Figure 1), and these polyhedra are composed of several virions embedded in a proteinaceous matrix. The only virus so far registered for agricultural use on row crops in the United States is the NPV from Heliothis zea. It is produced by Sandoz, Inc., under the trade name Elcar.
All presently known insect viruses must be ingested by the in- sect pest if they are to be effective. In the case of an NPV, which generally infects only larvae, the protein matrix dissolves in the gut and releases the virions, which then move into other tissues of the host. Within 24 hours, the nuclei of cells in cer- tain tissues of the larvae may contain polyhedra, but external appearance and behavior may not be noticeably changed. Indeed, during the succeeding incubation period, the larvae may continue to feed on the plant and to grow, though this may be affected by such factors as the quantity of active virus consumed, the size of the larvae, the temperature, and the food supply. Among the first symptoms of nucleopolyhedrosis (the disease caused by NPV) are sluggishness and decreased feeding. Then the larvae soon cease feeding and die within about 3 days, depending on the suscepti- bility of the host. The time from ingestion of a lethal dose of NPV to death for lepidopteran larvae on field cotton ranges from about 3 to 15 days. Other pathological characteristics of insect viruses are detailed by Steinhaus (1963), Smith (1967, 1977), and David (1975).
Plainly, the incubation period of an NPV and the fact that the material must be ingested should be recognized when an NPV is considered for control of cotton insects. In addition, NPV*s may be destroyed by ultraviolet radiation, pH, and plant defenses (Bullock 1967, Young and Yearian 1974, Jacques 1977), problems that can be partly overcome by using protective materials to increase the field persistence of the virus (ignoffo and Batzer 1971, Bull et al. 1976). Also, the stored virus may be destroyed or its ac- tivity reduced by exposure to high temperatures (McLeod et al. 1977). Finally, timing of an application of NPV is critical if larvae are to be infected as soon as possible after hatch and be- fore they can do serious plant damage.
On the other hand, NPV*s are usually selective and host spe- cific (i.e., the Heliothis NPV infects only Heliothis spp.), which can be a disadvantage or an asset; for example, an application against a pest insect would not affect beneficial insects, but it also would not affect any other pest species present. Finally, none of the baculoviruses has been found to be hazardous to man or other mammals, and they can be produced in the quantities needed

132 M. R. BELL
■5.1) -^ • '' *'i-^. ^w^
f4 ^^
« ^
v; :• §àt; (A) Cell nucleus contain- ing immature polyhedra of a nuclear polyhe- drosis virus
•7v \
V-..:
4
(B) Mature polyhedra of a nuclear polyhedrosis virus
(C) Virions of a nuclear polyhedrosis virus
Figure 1.—Tissue infected with a nuclear polyhedrosis virus. Original electron microphotography by F. D. Stewart, U.S. Department of Agriculture, Animal and Plant Health Inspection Service, Phoenix, AZ.

MICROBIAL AGENTS 133
at an economically competitive price (Shieh and Bohmfalk 1980). For further information on problems and benefits, the reader is referred to in-depth reviews of the subject (Cameron 1963, Stairs 1971, Ignoffo 1974, Falcon 1976).
Bacteria
Entomogenous bacteria may be either sporeformers (bacteria that produce persistent endospores) or nonsporeformers. At pres- ent, only the sporeformers are being seriously investigated for control of cotton insect pests. However, nonspore-forming patho- gens are found in cotton insects. For example, the bacterium Serrâtia marcescens Bizi o sometimes causes mortality of a wide range of insect hosts including the boll weevil (Anthonomus grandis Boheman) (McLaughlin and Keller 1964). It is particularly associ- ated with insects when they are being reared in the laboratory or are otherwise subjected to stress (Angus 1974).
By far the most promising bacterium available for microbial control of cotton insects is B^._^. The strain used at present (the HD-1 isolate) produces a proteinaceous parasporal crystal inclusion that is toxic to lepidopteran larvae but has little or no effect on other animals. Soon after the parasporal crystals are ingested, the gut cells of larvae of susceptible species are affected in such a way as to inhibit feeding by the larvae. In fact, treatment with B._t^. normally results in less feeding by the larval pest during the incubation period than treatment with an NPV or a GV, a matter of considerable importance when minimum damage to foliage is of concern.
Also, sublethal doses of B^._^. provide some control of larvae of at least some cotton insect pests. They may recover from the dose, but they feed less than untreated larvae and grow slowly for some time after exposure (Dulmage et al. 1978; M. R. Bell unpub- lished data). In addition, with this extended larval development period, they may be exposed to adverse environmental factors that may produce a control effect.
Bacteria, like viruses, must be ingested by an insect pest be- fore they can have any effect. Therefore, oviposition site, feed- ing habits of the larvae, and methods of application are important. Also, bacteria are usually adversely affected by the environment, and repeated applications may be necessary to maintain plant pro- tection. Finally, bacteria, especially those presently registered for use, are (like the viruses) considered environmentally safe. However, B^._t. has two major advantages over NPV ' s as a microbial agent. It has a wider host range, and large volumes can be readily produced by using fermentation equipment. This wide host range of the HD-1 isolate (it affects most Lepidoptera but is selective enough to be safe) gives it a large market. For this reason, the HD-1 isolate is marketed by several companies under various trade names (e.g., Dipel, Thuricide, Bactur). Many

134 M. R. BELL
publications are available that deal with various aspects of B.t. as a microbial insecticide (Heimpel and Angus 1963, Heimpel iTóT, Dulmage 1970, Burgerjon and Martouret 1971, Cooksey 1971, Falcon 1971b).
Fungi
Fungi differ from the bacteria, viruses, and protozoa that are being considered for control of agricultural pests in that inges- tion is not usually necessary for infection: the organisms can penetrate through the surface of the integument. Success of this mode of invasion, however, seems to depend on certain environmental conditions and on the physiological condition of the host cuticle. These conditions vary with different hosts and fungi. Temperature, humidity, and light are among the factors known to affect disease incidence.
Once cotton insects have been infected by a fungus, the gen- eral pathology of the disease (mycosis) is as follows: the infec- tive unit, usually the protected resting stage or spore, germinates on the cuticle, and the germ tube penetrates to the hemocoel. With most entomogenous fungi, fragments of the mycelium, so-called hy- phal bodies, then multiply in the body cavity. Thus, by the time of the death of the host, the host body is filled with mycelium and may become firm. At this point, the resting stages of the fungus may then be formed, usually as spores on conidiophores that have erupted from the insect body, which causes the body to take on a fuzzy appearance.
People working in cotton insect control have seen epizootics of fungal diseases that dramatically reduced the insect population. As a result, there is considerable interest in fungi as control agents. However, no fungi are presently registered for use on cot- ton in the United States. In general, the fungi have the advantage of a relatively wide host range and external invasion, and they can be produced by using culture methods. On the other hand, this wide host range may encompass beneficial insects, the necessary environ- mental conditions are sometimes limiting, and the incubation period may be long.
Nevertheless, the fungi have potential as microbial agents on cotton, especially in areas of normally high humidity such as the southeastern part of the Cotton Belt. More research is needed to obtain the knowledge required for decision making. In-depth de- scriptions of mycoses and reports on the use of fungi in microbial control are available (Madelin 1966, Roberts and Yendol 1971, Bell 1974, McCoy 1974, Ferron 1978).
Protozoa
Few protozoan pathogens of insects have been field tested as microbial control agents of crop pests, perhaps because the

MICROBIAL AGENTS I35
incubation period is so long that crop damage usually is not con- trolled (McLaughlin 1971). This is because few protozoa are highly virulent. Instead they tend to cause slow, debilitative symptoms that do not lead to the rapid control needed for most crops. This is a disadvantage when the concern is protection of a crop but could be an advantage when spread and persistence of the disease are more important factors.
In general, the protozoan pathogens that are being studied as microbial agents are applied in the resistant stage (spores or cysts). Once this stage has been ingested by the host, the invasive stage emerges in the gut, penetrates the gut wall, and begins to multiply in the host tissue. Most protozoan pathogens can only be produced in insect hosts, so it may sometimes be difficult to pro- duce large quantities. However, some are transmitted from genera- tion to generation within the egg, which enhances persistence of the disease in the population. Because the life cycles and types of protozoan pathogens are diverse, the reader is referred to more detailed reviews and descriptions for further information (Tañada 1959, Weiser 1963, McLaughlin 1971).
General Considerations
From the previous brief discussions, certain characteristics are obviously common to most pathogens that are used or are under study as microbial agents for control of cotton insects. Since control usually cannot occur until the pathogen is consumed, the techniques used to apply the pathogen are often very important. For example, even with fungi, which can penetrate the integument, the pathogen must contact the host. Therefore, a technique is needed that places the pathogen close to the target pest. However, there are also considerations such as the persistence of the mi- crobe, the formulation used (e.g., baits, stickers, protectants), and the possibility that rapid plant growth may dilute the micro- bial. The user must consider the biology of the host insect and the symptomatology of the disease, particularly as it affects the activity of the insect pest during the incubation period (e.g., where and how much feeding will occur). Finally, the methods used to evaluate the effects of microbial treatments should take into account the expected impact of an epizootic of the disease under study.
REVIEW OF MICROBIAL CONTROL OF COTTON INSECT PESTS
Boll Weevil
Few pathogens have been found in field populations of boll weevils, and none is registered for use at this time. Even though McLaughlin et al. (1966) isolated a variety of bacteria and fungi from dead, field-collected weevils, none of those tested proved to

136 M. R. BELL
be pathogenic. The researchers did, however, note that the micro- bial flora might have caused mortality of otherwise stressed wee- vils. Also, Serratia marcescens caused an epizootic in caged weevils that could only be controlled by feeding the weevils anti- biotics (McLaughlin and Keller 1964). (The mechanism of pathogen- icity of this bacterium was described (Slatten and Carson 1967).) Further, boll weevils were found to be susceptible to infection by the nonoccluded Chilo irridescent virus (McLaughlin et al. 1972), the fungus Beauveria bassiana (Balsamo) Vuillemin (McLaughlin 1962), and the fungi Metarrhizium anisopliae (Metschnikoff) Sorokin and Nomuraea rileyi (Farlow) Samson (R. E. McLaughlin and M. R. Bell unpublished data). Although all these infect boll weevils in the field, results of tests have been inconclusive as to their pos- sible use in control systems.
More extensive trials were conducted in 1965-67 with two pro- tozoan pathogens, both isolated from colonies where boll weevils were being mass reared (Gast 1966). One protozoan was Mattesia grandis McLaughlin (McLaughlin 1965), and the other was Glugea gast i McLaughlin (McLaughlin 1969). Both protozoa also infect the bollworm, the tobacco budworm (H_. virescens (F.)), and the pink bollworm (Pectinophora gossypiella (Saunders)) (ignoffo and Garcia 1965). In all cage tests and field trials, the protozoa were ap- plied to cotton in a bait formulation based on cottonseed oil that enhanced feeding activity of weevils (Daum et al. 1967). An oil- soluble dye added to mark weevils that fed on the bait formulation aided in evaluating the effectiveness of treatments. The percent- age of weevils marked by the dye and the incidence of the proto- zoan diseases were high early and late in the growing season but relatively low throughout the period of rapid development of the cotton during the middle of the season (McLaughlin 1967, McLaughlin et al. 1968, 1969). For example, about 80% of the marked weevils were infected by G_. gasti in late June to mid-July, but only 11-25% were infected from mid-July through August. Later in the season, the weevils again responded, and as many as 70% were diseased. Also, over 60% of the weevils collected from winter trash around the field were diseased, and winter mortality of diapausing weevils was higher in the test area.
Thus the research showed that epizootics could be induced in a weevil population with a bait formulation though the protozoan pathogens acted too slowly to protect the crop.
Pink Bollworm
At present, no pathogens are recommended for use in control- ling the pink bollworm. However, results of some field tests have been promising. Ignoffo (1962a, 1962b) found that pink bollworm larvae were susceptible to injections of _B.^. spores, determined the lethal dose, and showed that field conditions would probably be such that the bacterium could be used for control of pink bollworms. Subsequently, in field-cage studies, about 40% of

MICROBIAL AGENTS 137
mature larvae were killed by incorporating ^»t_, into soil (ignoffo and Graham 1967); however, the rates necessary to achieve this re- duction were not economically feasible. Also, in other field-cage studies, Bullock and Dulmage (1969) obtained significant control (only 10% of the bolls had larval mines) when jB.t^. was sprayed on cotton subsequently infested with pink bollworms; in the untreated control, 80% of the bolls had larval mines. However, the results were not reproducible in subsequent experiments. Graves and Watson (1970) also reported poor control with a commercial preparation of
Bacillus thuringiensis thus has promise for control of the pink bollworm because isolates are highly virulent. However, con- sistent results have not been obtained because no effective method of usage has been developed. As with other pathogens and other hosts, the major problem is getting the target larvae to ingest the pathogen, but the difficulty is especially great with the pink bollworm. Green bolls are a favored site for oviposition for pink bollworm females, and a high percentage of eggs on bolls are placed under the calyx (Brazzel and Martin 1957). However, larvae from eggs hatched on the bolls enter the bolls within minutes after hatching, and even larvae from eggs oviposited on other parts of the plant enter fruit within about 40 minutes (McLaughlin 1972). This short exposure to the external environment means that larvae have little time to consume an effective dose of pathogen.
The matter of time for ingestion is also a problem with viral pathogens. The NPV isolated from the alfalfa looper (Autographa californica (Speyer)) the so-called ACNPV, also infects pink boll- worm larvae (Vail et al. 1972) as well as several other lepidop- teran pests of cotton (Vail et al. 1970, Vail and Jay 1973). How- ever, in field tests involving weekly applications of ACNPV, only 1% of 3500 larvae examined (Vail et al. 1978) were infected, pre- sumably because the larvae did not ingest the pathogen.
An attempt was therefore made to increase the ingestion of virus by first-instar pink bollworm larvae by incorporating the pathogen in a feeding stimulant (Bell and Kanavel 1975). When this formulation was applied to greenhouse plants, 60.7% of larvae from eggs placed randomly on the plants were infected; when plants were treated with virus in water alone, only 28.7% of the larvae were infected. Also, in a subsequent field test, application of ACNPV plus bait (the feeding stimulant) reduced the number of pink boll- worm larvae in bolls 69% compared with the controls, and the per- centage of damaged bolls decreased (Table 1) (Bell and Kanavel 1977). In fact, the feeding adjuvant alone reduced the numbers of pink bollworm larvae in bolls 43%, apparently because the feeding behavior of newly hatched larvae was altered. The virus alone had no effect. Although this degree of control was not deemed practi- cal because of the quantity and cost of materials, the results in- dicated than an early-season application of such a formulation might be useful as a suppression method.

138 M. R. BELL
Table 1.—Effect of nuclear polyhedrosis virus from Autographa californica in water or in bait on pink bollworm damage to cot- ton, 1975 1,2/
No. pink bollworms per %
Treatment 3/ 50 bolls damaged bolls
Untreated control 37.3 a 37.6 a 2.97X10^^ PIB*s per ha w/o bait 34.6 a 33.0 ab 935 liters bait per ha w/o virus 21.8 ab 24.0 be 2.97X10 PIB's per ha in 935 liters
bait 11.6 b 14.1 c
\J Average of four replications per treatment during the 1975 growing season.
1_/ Values within column followed by the same letter are not sig- nificantly different at the 5% level as determined by LSD.
_3/ FIB = polyhedral inclusion body.
Other pathogens infect the pink bollworm and can be considered potential control agents. For example, the protozoa earlier identified as a boll weevil pathogen also infected pink bollworms (no field tests were conducted) (ignoffo and Garcia 1965). Also, a cytoplasmic polyhedrosis virus found in a laboratory culture of pink bollworms (ignoffo and Adams 1966) produced chronic, débili- tât ive effects on the insect. At present, this pathogen is not considered a promising candidate as a microbial agent for control of pink bollworms because of the quantity of virus required to pro- duce mortality and problems associated with production (Bell and Henneberry 1980).
Cotton Bollworm and Tobacco Budworm
Most of the research effort involving microbial control of cotton insects in the United States has had to do with the boll- worm and tobacco budworm. Both B^.^. and the NPV from H. zea (so- called B£CU]JOVÍ£US h£]^^^ are registered for use against these pests, and the ACNPV, though not yet registered, has shown promise against tobacco budworms.
However, the level of control of Heliothis spp. on cotton pro- duced by multiple applications of the Heliothis NPV has been errat- ic. In some tests, the control obtained has been comparable to that obtained with chemical insecticides. For example, Ignoffo et al. (1965) reported yields of 560 kilograms and 715 kilograms of seed-cotton per 0.4 hectare from plots treated with 6x10 and 6x10 PIB's per 0.4 hectare, respectively, in 25.4 liters of spray per 0.4 hectare; yields from untreated plots were 294 kilograms per 0.4 hectare. Also, Allen et al. (1967) found that in small plots.

MICROBIAL AGENTS 139
applications of 1.2x10 PIB's per 0.4 hectare were as effective as the insecticide standard (toxaphene-DDT). In large plots, ap- plications of the virus plus the naturally occurring predator-para- site complex gave satisfactory control (Allen et al. 1966). Shieh and Bohmfalk (1980) also found that yields of seed cotton increased 10-40% over yields from check plots when multiple treatments of Elcar were applied to cotton against relatively low populations of Heliothis spp. However, other researchers have obtained marginal to no control with Heliothis NPV used alone in field tests (McGarr and Ignoffo 1966, McGarr 1968, Pfrimmer 1979).
Generally, the control of Heliothis spp. obtained with B^.^. variety HD-1 has been more consistent than that obtained with the Heliothis NPV. Control comparable to that obtained with a chemical insecticide was reported by McGarr et al. (1970), but the quantity of formulation necessary was too great for applications to be eco- nomically feasible. Others report that application of 3.6-7.3x10 International Units (lU) per 0.4 hectare will suppress a larval population and result in increased yield (Pfrimmer et al. 1971, Bull et al. 1979, Pfrimmer 1979). However, the level of control obtained was usually not comparable to that obtained with chemical insecticides. On the other hand. Bull et al. (1979) found that B._t.and the Heliothis NPV gave comparable control of moderate popu- Tations of larvae when applied at their respective recommended rates.
Several attempts have been made to increase the effectiveness of the Heliothis NPV and B.t_. against Heliothis spp. One possi- bility that was explored because of the known rapid inactivation of NPV on cotton (Yearian and Young 1974, Young and Yearian 1974, Ig- noffo et al. 1976b) was spray formulations that would improve the persistence of the pathogens on field cotton. In fact, formula- tions that protected the polyhedra from sunlight did increase the field persistence of NPV (Bull et al. 1976) but did not signifi- cantly increase efficacy. Apparently what was needed was an ad- juvant that included a gustatory stimulant, a sunlight protectant, and an evaporation retardant (ignoffo et al. 1976a), but the most important of these additions would be a gustatory stimulant capable of increasing foliar feeding of the larvae since that is of major importance for the consistent effectiveness of microbial treatments. A number of the materials that elicit feeding responses in Heliothis spp. have been identified and tested for use (Montoya et al. 1966, McLaughlin et al. 1971, Patti and Garner 1974, Bell and Kanavel 1978). In most instances, the addition of these materials to NPV or B.t. sprays has increased the effectiveness of the microbials.
Two gustatory-type spray adjuvants are presently marketed for commercial use with NPV or B^._t. on cotton. One is Gustol, devel- oped and manufactured by Sandoz, Inc. The other is COAX, manufac- tured by Traders Oil Mill Co. Both increase foliar feeding of Heliothis spp. larvae and thus increase effectiveness of the micro- bials. Both also increase the persistence of NPV on cotton. In ten field tests, applications of Heliothis NPV and Gustol to

140 M. R. BELL
early-season cotton resulted in 3.4% of the squares being damaged and 2.8% of the bolls; 6.2% of the squares and 5.2% of the bolls were damaged when the NPV was applied without the adjuvant (L, Russo, Sandoz, Inc., personal communication). Likewise, in full- season trials (average, three tests), yields of seed cotton were 505, 622, and 723 kilograms per 0.4 hectare for untreated fields, fields treated with virus alone, and fields treated with virus plus Gustol, respectively.
Much the same thing has happened when adjuvants have been added to ACNPV. Field tests conducted on late-planted cotton in Arizona indicated that applications of a mixture of ACNPV plus B^._t. plus COAX gave acceptable control of a moderate to heavy infesta- tion of Heliothis spp. (93-98% H. virescens) (Bell and Romine 1980). Only <10% of the squares treated with the mixture were dam- aged; 30-60+% were damaged in untreated cotton. Moreover, yield was greatly increased (Table 2).
Thus the mixture of B^.£. and virus appeared to give more effi- cacious control than the other treatments even though these two pathogens were antagonistic in laboratory tests (M. R. Bell un- published data). Possibly the sublethal effect of JB._t. (reducing growth rate) described by Dulmage et al. (1978) was involved. The larvae, though alive, remained small for some time and may have been killed by the virus before they recovered from the JB._t.
Microbials have also been mixed with chemical insecticides to improve the level of control of Heliothis spp. in cotton (Pieters et al.1978, Luttrell et al. 1979, Yearian et al. 1980). However, results reported to date have indicated that little was gained by mixing NPV or ^.t_. with these materials.
In summary, procedures that tend to increase the probability that Heliothis spp. larvae will ingest an active microbial at the proper time have been relatively effective in increasing the level of control. These procedures include the use of a feeding-stimu- lant adjuvant, optimum dosages, spray methods that result in maxi- mum deposits on the target area, and proper timing of the applica- tion so it affects larvae of the right age (Falcon 1971a, Chapman and Ignoffo 1972, Stacey et al. 1977a, 1977b). Another possibility that has not been much investigated is the effect of growth and fruiting pattern of the cotton on damage. Falcon (1974) discussed the possible effect of the presence of squares and small bolls on cotton during the latter part of the season. Such fruit would not normally mature and be harvested, but could provide feeding sites for young Heliothis spp. larvae during the incubation period of the viral pathogen. This feeding damage apparently would not reduce yield, and the preference of the larvae for the small bolls during that period would presumably protect the more mature, harvestable bolls.

MICROBIAL AGENTS 141
Table 2.—Yield of late-planted (June 26) cotton treated with mi- crobials for control of Heliothis spp. , Phoenix, AZ, 1978 l_l
_____
seed cotton Treatment Rate per ha 2/ (kg/ha)
Test 1 Bacillus thuringiensis (B^._t. ) 560 g 1427 a
+ACNPV V 7.41x10 FIB 4/ +adjuvant 3.36 kg
ACNPV 7.41x10^^ PIB 1066 b +adjuvant 3.36 kg
ACNPV alone 7.41x10^^ PIB 744 c
Untreated control 322 d
Test 2 B.£. 560 g 1108 a +adjuvant * 3.36 kg
B.£. alone 560 g 837 b
Untreated control 328 c
1/ Average of four replicates; 15 row-meters handpicked per rep- licate. Means within columns not followed by the same letter are significantly different at the 5% level of confidence (Duncan's multiple range test—analysis of variance of Latin square).
2^/ All treatments were applied in a total volume of 93.5 liters water per hectare.
_3/ ACNPV = nuclear polyhedrosis virus of the alfalfa looper. 4/ PIB = polyhedral inclusion body.
Other Cotton Insects
The NPV's of the cabbage looper (Trichoplusia ni (Hübner)) and the beet armyworm (Spodoptera exigua (Hübner)) occur naturally in populations of larvae in cotton and are important in the regulation of their respective hosts. When densities of larvae are high,the natural epizootics that usually follow greatly reduce the popula- tions. The feeding behavior of these two species favors ingestion and rapid spread of the viral diseases; thus when augmentation is necessary, these viruses can be applied with standard equipment and without elaborate preparations (Falcon 1971a). However, control of

142 M. R. BELL
these species is seldom needed because cotton can tolerate consid- erable damage to the foliage at the time populations are normally high. The viruses could nevertheless be important in an area pest management program when reduction of a population on cotton might prevent excessive damage to fall crops.
The cotton leafperforator (Bucculatrix thurberiella Busck) is considered a sporadic pest of cotton in the western United States. Although this insect is a leaf miner during most of its larval stage and thus is not exposed to microbial applications dur- ing that time, Vail et al. (1977) obtained partial control with multiple applications of the ACNPV. Further, multiple applications of B^._t^. at normal recommended rates (0.227-0.454 kg per ha) re- sulted in an acceptable level of control (M. R. Bell unpublished data). Such methods probably would not be used to control this pest, but treatments directed against other pest insects could re- duce populations of cotton leafperforators.
CONCLUSION
Although often ignored, pathogens act continuously to limit populations of some cotton pests such as the cabbage looper. Their usefulness in regulating populations of Heliothis spp. in cotton has also been demonstrated. In addition, insect pathogens may interfere with development or reproduction of pest insects and can increase their susceptibility to predators, parasites, or chemical insecticides.
Many efforts are being made to increase the effectiveness and use of microbial agents in management of cotton insects. These in- clude but are not limited to: (1) identifying new, more virulent pathogens or new strains of known pathogens; (2) improving spray formulations; (3) developing new methods of applying or dissemina- ting pathogens; and (4) determining plant-insect-pathogen interac- tions. The development of these more effective methods or materi- als, plus the fact that users are more knowledgeable about the factors involved in the use of microbials, should result in more consistent control of cotton insect pests.

MICROBIAL AGENTS 143
LITERATURE CITED
Allen, G. E., B. G. Gregory, and J. R. Brazzel. 1966. Integration of the Heliothis nuclear polyhedrosis virus
into a biological control program for cotton. Journal of Eco- nomic Entomology 59:1333-1336.
B. G. Gregory, and T. L. Pate. 1967. Field evaluation of a nuclear-polyhedrosis virus in the
control of Heliothis zea and Heliothis virescens on cotton. Journal of Invertebrate Pathology 9:40-42.
Angus, T. A. 1974. Microbiological control of insects: bacterial pathogens.
In Proceedings, Summer Institute on Biological Control of Plant Insects and Diseases, p. 532-540. Mississippi State Univer- sity, University Press of Mississippi.
Bell, J. V. 1974. Mycoses. In Insect Diseases. Volume 1, p. 185-236.
G. E. Cantwell, editor. Marcell Dekker, Inc., New York.
Bell, M. R., and T. J. Henneberry. 1980. Entomopathogens for pink bollworm control. In Pink Boll-
worm Control in the Western United States. H. M. Graham, edi- tor. U.S. Department of Agriculture, ARM-W-16, p. 76-81.
and R. F. Kanavel. 1975. Potential of bait formulations to increase effectiveness
of nuclear polyhedrosis virus against the pink bollworm. Jour- nal of Economic Entomology 68:389-391.
and R. F. Kanavel. 1977. Field tests of a nuclear polyhedrosis virus in a bait for- mulation for control of pink bollworms and Heliothis spp. on cotton in Arizona. Journal of Economic Entomology 70:625-629.
and R. F. Kanavel. 1978. Tobacco budworm: development of a spray and adjuvant to
increase effectiveness of a nuclear polyhedrosis virus. Jour- nal of Economic Entomology 71:350-352.
and C. L. Romine. 1980. Tobacco budworm: field evaluation of microbial control in
cotton using Bacillus thuringiensis and a nuclear polyhedrosis virus with a feeding adjuvant. Journal of Economic Entomology 73:427-430.
Brazzel, J. R., and D. F. Martin. 1957. Oviposit ion sites of the pink bollworm on the cotton
plant. Journal of Economic Entomology 50:122-124.

144 M. R. BELL
Bull, D. L., V. S. House, J. R. Ables, and R. K. Morrison. 1979. Selective methods for managing insect pests of cotton.
Journal of Economic Entomology 72:841-846.
R. L. Ridgway, V. S. House, and N. W. Pryor. 1976. Improved formulation of the Heliothis nuclear polyhedrosis
virus. Journal of Economic Entomology 69:731-736.
Bullock, H. R. 1967. Persistence of Heliothis nuclear polyhedrosis virus on
cotton foliage. Journal of Invertebrate Pathology 9:434-436.
and H. T. Dulmage. 1969. Bacillus thuringiensis against pink bollworms on cotton in
field cages. Journal of Economic Entomology 62:994-995.
Burgerjon, A., and D. Martouret. 1971. Determination and significance of the host spectrum of
Bacillus thuringiensis. In Microbial Control of Insects and Mites, p. 305-325. H. D. Burges and N. W. Hussey, editors. Academic Press, New York.
Cameron, J. W. M. 1963. Factors affecting the use of microbial pathogens in insect
control. Annual Review of Entomology 8:265-286.
Chapman, A. J., and C. M. Ignoffo. 1972. Influence of rate and spray volume of nucleopolyhedrosis virus on control of Heliothis in cotton. Journal of Inverte- brate Pathology 20:183-186.
Cooksey, K. E. 1971. The protein crystal toxin of Bacillus thuringiensis: biochemistry and mode of action. In Microbial Control of In- sects and Mites, p. 247-274. H. D. Burges and N. W. Hussey, editors. Academic Press, New York.
Daum, R. J., R. E. McLaughlin, and D. D. Hardee. 1967. Development of the bait principle for boll weevil control:
cottonseed oil, a source of attractants and feeding stimulants for the boll weevil bait. Journal of Economic Entomology 60: 321-325.
David, W. A. L. 1975. The status of viruses pathogenic for insects and mites. Annual Review of Entomology 20:97-117.
Dulmage, H. T. 1970. Insecticidal activity of HD-1, a new isolate of Bacillus
thuringiensis var. alesti. Journal of Invertebrate Pathology 15:232-239.

MICROBIAL AGENTS ""^S
Dulmage, H. T., H. M. Graham, and E. Martinez. 1978. Interactions between the tobacco budworm, Heliothis vires-
cens, and the ô-endotoxin produced by the HD-l isolate of Bacillus thuringiensis var. kurstaki: relationship between length of exposure to the toxin and survival. Journal of Invertebrate Pathology 32:40-50.
Falcon, L. A. 1971a. Microbial control as a tool in integrated control pro-
grams. I_n Biological Control, p. 346-364. C. B. Huffaker, editor. Plenum Press, New York.
1971b. Use of bacteria for microbial control. In Microbial Con- trol of Insects and Mites, p. 67-69. H. D. Burges and N. W. Hussey, editors. Academic Press, New York.
1974. Insect pathogens: Integration into a pest management system. Ln Proceedings, Summer Institute on Biological Control of Plant Insects and Diseases. Mississippi State University, p. 618-627. University Press of Mississippi.
1976. Problems associated with the use of arthropod viruses in pest control. Annual Review of Entomology 21:305-324.
Ferron, P. 1978. Biological control of insect pests by entomogenous fungi. Annual Review of Entomology 23:409-442.
Gast, R. T. 1966. Control of four diseases of laboratory-reared boll weevils. Journal of Economic Entomology 59:793-797.
Graves, G. N., and T. F. Watson. 1970. Effect of Bacillus thuringiensis on the pink bollworm. Journal of Economic Entomology 63:1828-1830.
Heimpel, A. M. 1967. A critical review of Bacillus thuringiensis var. thurin- giensis Berliner and other crystalliferous bacteria. Annuaf Review of Entomology 12:287-322.
and T. A. Angus. 1963. Diseases caused by certain spore forming bacteria. In_ In-
sect Pathology, an Advanced Treatise, p. 21-73. Volume 2. E. A. Steinhaus, editor. Academic Press, New York.
Ignoffo, C. M. 1962a. Susceptibility of Pectinophora gossypiella (Saunders) to
intrahemocoelic injections of Bacillus thuringiensis Berliner. Journal of Insect Pathology 4:34-40,

146 M. R. BELL
Ignoffo, C. M. 1962b. The effects of temperature and humidity on mortality of
larvae of Pectinophora gossypiella (Saunders) injected with Bacillus thuringiensis Berliner. Journal of Insect Pathology 4:63-71.
1974. Microbiological control of insects: viral pathogens. ^E Proceedings, Summer Institute of Biological Control of Plant Insects and Diseases. Mississippi State University, p. 541- 557. University Press of Mississippi.
and J. R. Adams. 1966. A cytoplasmic-polyhedrosis virus. Smithiavirus pecti- nophorae sp. n. of the pink bollworm, Pectinophora gossypiella (Saunders). Journal of Invertebrate Pathology 8:59-66.
and 0. F. Batzer. 1971. Microencapsulation and ultraviolet protectants to increase
sunlight stability of an insect virus. Journal of Economic En- tomology 64:850-853.
A. J. Chapman, and D. F. Martin. 1965. The nuclear-polyhedrosis virus of Heliothis zea (Boddie)
and Heliothis virescens (Fabricius). III. The effectiveness of the virus against field populations of Heliothis on cotton, corn and grain sorghum. Journal of Invertebrate Pathology 7: 227-235.
and C. Garcia. 1965. Infection of the cabbage looper, bollworm, tobacco bud-
worm, and pink bollworm with spores of Mattesia grandis McLaughlin collected from boll weevils. Journal of Inverte- brate Pathology 7:260-262.
and H. M. Graham. 1967, Laboratory and field cage tests with Bacillus thuringien-
sis against pink bollworm larvae. Journal of Invertebrate Pathology 9:390-394.
D. L. Hostetter, and D. B. Smith. 1976a. Gustatory stimulant, sunlight protectant, evaporation re-
tardant: three characteristics of a microbial insecticidal ad- juvant. Journal of Economic Entomology 69:207-210.
W. C. Yearian, S. Y. Young, D. L. Hostetter, and D. L. Bull.
1976b. Laboratory and field persistence of new commercial for- mulations of the Heliothis nucleopolyhedrosis virus, Baculo- virus heliothis. Journal of Economic Entomology 69:233-236.

MICROBIAL AGENTS 147
Jacques, R. P. 1977, Stability of entomopathogenic viruses. In Environmental
Stability of Microbial Insecticides. Entomology Society of America, Miscellaneous Publication No. 10, p. 99-116.
Luttrell, R. G., W. C. Yearian, and S. Y. Young. 1979. Laboratory and field studies on the efficacy of selected
chemical insecticide-Elcar® (Baculovirus heliothis) combina- tions against Heliothis spp. Journal of Economic Entomology 72:57-60.
McCoy, C. W. 1974. Fungal pathogens and their use in the microbial control
of insects and mites. ^ Proceedings, Summer Institute on Biological Control of Plant Insects and Diseases. Mississippi State University, p. 564-575. University Press of Mississippi.
McGarr, R. L. 1968. Field tests with a nuclear polyhedral virus against the bollworm and tobacco budworm, 1964-1966. Journal of Economic Entomology 61:342.
H. T. Dulmage, and D. A. Wolfenbarger. 1970. The a-endotoxin of Bacillus thuringiensis, HD-1, and
chemical insecticides for control of the tobacco budworm and the bollworm. Journal of Economic Entomology 63:1357-1358.
and C. M. Ignoffo. 1966. Control of Heliothis spp. with a nuclear polyhedrosis virus, EPN, and two newer insecticides. Journal of Economic Entomology 59:1284-1285.
McLaughlin, J. R. 1972. Bionomics of the pink bollworm, Pectinophora gossypiella
(Saunders), in the southern desert cotton region of California. Ph.D. dissertation. University of California, Riverside. 98 p.
McLaughlin, R. E. 1962. Infectivity tests with Beauveria bassiana (Balsamo) Vuillemin on Anthonomus grandis Boheman. Journal of Insect Pathology 4:386-388.
1965. Mattesia grandis n. sp., a sporozoan pathogen of the boll weevil, Anthonomus grandis Boheman. Journal of Protozoology 12:405-4TT:
1967. Development of the bait principle for boll weevil control. II. Field-cage tests with a feeding stimulant and the proto- zoan Mattesia ^randi£. Journal of Invertebrate Pathology 9: 70-77.

148 M. R. BELL
McLaughlin, R. E. 1969. Glugea gasti sp. n., a microsporidian pathogen of the boll weevil, Anthonomus grandis. Journal of Protozoology 16:84-92.
1971. Use of protozoans for microbial control of insects. In Microbial Control of Insects and Mites, p. 151-172. H. DT~ Burges and N. W. Hussey, editors. Academic Press, New York.
G. L. Andrews, and M. R. Bell. 1971. Field tests for control of Heliothis spp. with a nuclear
polyhedrosis virus included in a boll weevil bait. Journal of Invertebrate Pathology 18:304-305.
M. R. Bell, and S. D. Veal, 1966. Bacteria and fungi associated with dead boll weevils (An-
thonomus grandis) in a natural population. Journal of Inverte- brate Pathology 8:401-408.
T. C. Cleveland, R. J. Daum, and M. R. Bell. 1969. Development of the bait principle for boll weevil control.
IV. Field tests with a bait containing a feeding stimulant and the sporozoans Glugea gasti and Mattesia grandis. Journal of Invertebrate Pathology 13:429-441.
^___^_ R. J. Daum, and M. R. Bell. 1968. Development of the bait principle for boll weevil control.
III. Field-cage tests with a feeding stimulant and the proto- zoans Mattesia grandis (Neogregarinida) and a microsporidian. Journal of Invertebrate Pathology 12:168-174.
and J. C. Keller. 1964. Antibiotic control of an epizootic caused by Serratia marcescens Bizio in the boll weevil, Anthonomus grandis Bohe- man. Journal of Insect Pathology 6:481-485.
H. A. Scott, and M. R. Bell, 1972. Infection of the boll weevil by Chilo irridescent virus. Journal of Invertebrate Pathology 19:285-290.
McLeod, P. J., W. C. Yearian, and S. Y. Young, III. 1977. Inactivation of Baculovirus heliothis by ultraviolet ra-
diation, dew, and temperature. Journal of Invertebrate Pathol- ogy 30:237-241.
Madelin, M. F. 1966. Fungal parasites of insects. Annual Review of Entomology
11:423-448.
Montoya, E. L., C. M. Ignoffo, and R. L. McGarr. 1966. A feeding stimulant to increase effectiveness of, and a
field test with a nuclear polyhedrosis virus of Heliothis. Journal of Invertebrate Pathology 8:320-324.

MICROBIAL AGENTS 149
Patti, J. H., and G. R. Garner. 1974. Bacillus thuringiensis investigations for control of Heliothis spp. on cotton. Journal of Economic Entomology 67: 415-418.
Pfrimmer, T. R. 1979. Heliothis spp. control on cotton with pyrethroids, car-
bamates, organophosphates, and biological insecticides. Jour- nal of Economic Entomology 72:593-598.
R. E. Furr, and E. A. Stadelbacher. 1971. Materials for control of boll weevils, bollworm, and to- bacco budworms on cotton at Stoneville, Mississippi. Journal of Economic Entomology 64:475-478.
Pieters, E. P., S. Y. Young, W. G. Yearian, W. L. Sterling, D. R. Melville, and F. R. Gilliland.
1978. Efficacy of Elcar^-chlordimeform and Dipel®-chlordimeform mixtures for control of Heliothis spp. on cotton. Southwestern Entomologist 3:237-240.
Roberts, D. W., and W. G. Yendol. 1971. Use of fungi for microbial control of insects. In Micro- bial Gontrol of Insects and Mites, p. 125-150. H. D. Burges and N. W. Hussey, editors. Academic Press, New York.
Shieh, T. R., and G. T. Bohmfalk. 1980. Production and efficacy of baculoviruses. Biotechnology
and Bioengineering XXII:1357-1375.
Slatten, B. H., and A. D. Carson. 1967. Mechanism of pathogenicity of Serratia marcescens. I. Virulence for the adult boll weevil. Journal of Invertebrate Pathology 9:78-81.
Smith, K. M. 1967. Insect virology. Academic Press, London, 256 p.
1977. Virus-insect relationships. Longman, New York, 287 p.
Stacey, A. L., W. C. Yearian, and S. Y. Young, III. 1977a. Evaluation of Baculovirus heliothis with feeding stimu-
lants for control of Heliothis larvae on cotton. Journal of Economic Entomology 70:779-784.
S. Y. Young, III, and W. G. Yearian. 1977b. ^Effect of larval age and mortality level on damage to
cotton by Heliothis zea infected with Baculovirus heliothis. Journal of Economic Entomology 70:383-386.

^^^ M. R. BELL
Stairs, G. R. 1971. Use of viruses for microbial control of insects. In Mi-
crobial Control of Insects and Mites, p. 97-124. H. D."Yurges and N. W. Hussey, editors. Academic Press, New York.
Steinhaus, E. A., editor. 1963. Insect pathology, an advanced treatise. Volume 1. Aca-
demic Press, New York.
and M. E. Martignoni. 1970. An abridged glossary of terms used in invertebrate pathol-
ogy. 2nd edition. Miscellaneous Publication.
Tañada, Y. 1959. Microbial control of insect pests. Annual Review of Ento- mology 4:277-302.
Vail, P. v., T. J. Henneberry, L. A. Bariola, R. L. Wilson, F. D. Wilson, D. L. Kittock, and H. F. Arle.
1978. Evaluation of several control techniques as potential com- ponents of an integrated system for pink bollworm suppression in the Southwest. U.S. Department of Agriculture, Production Research Report No. 172, 18 p.
T. J. Henneberry, and M. R. Bell. 1977. Cotton leafperforator: effect of a nuclear polyhedrosis
virus on field populations. Journal of Economic Entomology 70: 727-728.
D. L. Jay. 1973. Pathology of a nuclear polyhedrosis virus of the alfalfa
looper in alternate hosts. Journal of Invertebrate Pathology 21:198-204.
D. L. Jay, and D. K. Hunter, 1970. Cross infectivity of a nuclear polyhedrosis virus isola-
ted from the alfalfa looper, Autographa californica. In Pro- ceedings, Fourth International Colloquium On Insect Pathology, p. 297-304.
D. L. Jay, D. K. Hunter, and R. T. Staten. 1972. A nuclear polyhedrosis virus infective to the pink boll-
worm, Pectinophora gossypiella. Journal of Invertebrate Pa- thology 20:124-128.
Weiser, J. 1963. Sporozoan infections. In Insect Pathology, an Advanced
Treatise, p. 291-334. Volume 2. E. A. Steinhaus, editor. Academic Press, New York.

MICROBIAL AGENTS 151
Yearian, W. C, R. G. Luttrell, A. L. Stacy, and S. Y. Young. 1980. Efficacy of Bacillus thuringiensis and Baculovirus helio- this-chlorodimeform spray mixtures against Heliothis spp on cotton. Journal of the Georgia Entomological Society 15:258- 260.
and S. Y. Young. 1974. Persistence of Heliothis nuclear polyhedrosis virus on
cotton plant parts. Environmental Entomology 3:1035-1036.
Young, S. Y., and W. C. Yearian. 1974. Persistence of Heliothis NPV on foliage of cotton. En- vironmental Entomology 3:253-255.


153
Chapter 7
BOLL WEEVIL STERILITY
J. E. Wright and E. J. Villavaso Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
ABSTRACT Early sterilization of boll weevils (Anthonomus grandis Boheman) with gamma irradi- ation was not satisfactory because the treatment caused high mortality and did not completely sterilize the males and females. Chemosteri- lants were therefore evaluated, and the weevils used in the Pilot Boll Weevil Eradication Exper- iment in southern Mississippi in the early 1970's were treated with these materials. How- ever, chemosterilization was effective only for male boll weevils, so errors in separating the sexes resulted in the release of some fertile females. Also the effects of the chemosterilants on all males were not permanent; some recovered fertility before death. Research produced other candidate sterilization procedures such as irra- diation of pupae with fractionated doses of irradiation, fumigation of adults, and gamma ir- radiation plus immersion in solutions of chemo- sterilants. Finally, a new method was devised that sterilized both sexes. Newly emerged adults are fed for 5 days on a thin layer (slab) of adult diet containing 100 ppm of diflubenzuron; and on the 6th day both sexes are exposed to 10,000 rad of gamma irradiation in a nitrogen atm.osphere from a cesium-137 source. Male and female boll weevils treated by this method are 99.99% sterile and are dead within 14 days after irradiation. Thus, no regeneration of reproductive tissues occurs.
Tests of the field competitiveness of the sterilized weevils show that they are about 25% as competitive as untreated weevils from days 0 through 4 after release; then competitiveness rapidly decreases.

154 j. E. WRIGHT AND E. VILLA VASO
Boll weevils sterilized by the new method were released in the Boll Weevil Eradication Trial in North Carolina and Virginia in 1979.
There was no evidence of reproduction by native or released weevils in the evaluation area following the release of more than 11 million sterile insects.
The development and successful use of sterile boll weevils is described, and the ramifications for further implementation are discussed.
INTRODUCTION
Since Knipling (1955) proposed that insect control or eradica- tion could be achieved by the introduction of sterile male insects into a natural population, scientists have sought a satisfactory method of sterilizing the boll weevil (Anthonomus grandis Boheman). Davich and Lindquist (1962) reported that gamma irradiation was not satisfactory because sterilization was not permanent and the dose required to produce the temporary sterility was the lethal dose. Davich et al. (1965) also calculated that the sterilized males lost about 80% vigor and were only about 20% as competitive for native females as the native (unirradiated) male weevils.
Meanwhile certain chemicals were reported to be effective sterilizing agents (Knipling 1960, Lindquist 1961), and LaBrecque (1961) reported that three alkylating agents (aphoxide, apholate, and aphomide) caused a high degree of sterility in house flies (Musca domestica L.). Haynes (1963) subsequently found that one of these, apholate, reduced subsequent hatch of eggs when male boll weevils were dipped in a 2% aqueous solution. Hedin et al. (1964) then reported that male boll weevils were sterilized when apholate was incorporated into the diet or applied as a foliar spray to cot- ton plants to which they were exposed. On the other hand, Lind- quist et al. (1964) reported failure of a small-scale eradication test against the boll weevil when apholate was used as the sterilant.
Davich et al. (1965) speculated that the reason for their failure to achieve eradication in a small-scale test was that sex- ual competitiveness was reduced in the males. Also, Borkovec (1962) reported that apholate and other chemosterilants had an effect on insects similar to that of gamma irradiation; i.e., in- duction of a dominant lethal effect (Knipling 1964).
Next, fractionated doses of gamma irradiation were tried as a method of sterilizing the boll weevil (Flint et al. 1966), but the lethal effects were such that this procedure was not considered

BOLL WEEVIL STERILITY 155
further at that time. Thermal neutrons, fast neutrons, and micro- waves were investigated by several scientists. Some sterility was induced but mortality was high (Klassen et al. 1969). Bartlett et al. (1968) reported that in laboratory studies, male boll weevils sterilized by gamma irradiation were relatively competitive for 4-5 days and might be able to suppress a native population during that time. Davich (1976) tested this idea in Texas in 1968 on 65 acres of cotton and achieved a reduction in the size of the native population. However, when Davich et al. (1967) attempted to eradi- cate a natural field population of weevils in a partly isolated area in Baldwin County, AL, by releasing males sterilized by dipping them twice in a 2% apholate solution (24-hour intervals), eradication was not achieved because of (1) lack of isolation, (2) initially high populations, (3) sterility of males ranging from 88 to 100% and averaging ca. 95%, (4) males that survived and regained fertility, and (5) necessity of sexing males by hand to remove fe- males that were not sterilized by the treatment. (The 2% error in separating the sexes resulted in the release of some fertile females.)
A chemosterilant, busulfan, was reported to sterilize male boll weevils without causing appreciable damage to vigor or mating ability (Klassen et al. 1968, Klassen and Earle 1970). The three major disadvantages of busulfan identified at the time were: (1) it was not an adequate female sterilant; (2) there was only a small margin of safety in the dosage requirements (again the sterilizing dose and the lethal dose were approximately the same); and (3) to obtain sterilized males, the males had to consume a sterilizing dose of the treated adult diet. Haynes et al. (1973) showed that a combination of hempa and busulfan was superior to busulfan alone. Lloyd et al. (1976) reported that feeding an adult diet containing busulfan or busulfan and hempa produced an average 93% sterility (from laboratory and field assays, the range was from 70.6 to 99.6%) in male boll weevils released during the Pilot Boll Weevil Eradication Experiment (PBWEE) though 30.3 ± 2.3 yg of busulfan had to be ingested to obtain this level of sterility. Even so, weevils were obviously not completely sterile, and those that survived re- gained fertility. In fact, in 1973, the final year of the PBWEE (Lloyd et al. 1976), males were sterilized by feeding a 0.09% con- centration of busulfan in the adult diet. During the PBWEE, fer- tile eggs and larvae were detected in two fields in the most iso- lated part of the eradication zone. Since these eggs were believed to be the progeny of released females, procedures were changed. A combination of 0.09% busulfan and 0.4% hempa was fed in the adult diet to increase the sterility induced in both males and females. The resulting level of sterility initially appeared to range from 94 to 100%. Knipling (1976) in the report of the Technical Guid- ance Committee for the Pilot Boll Weevil Eradication Experiment therefore indicated a need for (1) improved mass-rearing pro- cedures to assure the capability of producing adequate numbers of high quality boll weevils for sterilization and release, (2) an improved technique to assure maximum and consistently high levels of sterilization with a minimum detrimental effect on the vigor

156 j. E. WRIGHT AND E. VILLAVASO
and mating competitiveness of the males, and (3) new methods for sterilizing both sexes of the boll weevil. Subsequently, major research efforts were made to improve rearing procedures and to develop a new sterilization procedure.
DEVELOPMENT OF.NEW METHODS OF STERILIZATION
Since the results of the PBWEE had indicated that a new method of sterilizing both sexes of the boll weevil was needed, several were developed and tested.
In 1975, D. Birkenmeyer and D. Childress (unpublished data) investigated the effect of fractionated doses on adult boll wee- vils. Haynes and Mitchell (1977) and Haynes et al. (1977) found that fractionated doses of irradiation applied to pupae or newly emerged adult boll weevils (total dose of 6-8 krads) resulted in 98-100% sterility.
Gassner et al. (1974), Terry et al. (1974, 1977), and Earle and Leopold (1975) described procedures for application of chemo- sterilants to boll weevils based on fumigation in vacuum. McHaffey and Borkovec (1976) tested a reduced-pressure dipping method for the materials. Also, Borkovec and McHaffey (1977) chemosterilized adult boll weevils by fumigation (both sexes) at atmospheric pres- sure with alkylating aziridinyl phosphine sulfides and oxides. The chemosterilant, bisazir, was investigated by Borkovec et al. (1978) and McCoy and Wright (1979) and was found to sterilize about 99% of treated males. However, mortality was similar to that produced by sterilizing doses of radiation.
Either fumigation or irradiation of aged adults was therefore satisfactory for sterilizing male boll weevils, but the females would oviposit fertile eggs until the lethal effects of the treat- ments were expressed. The insect growth regulator, diflubenzuron, and its analog penfluron had been shown to prevent hatch of eggs from exposed female insects (Moore and Taft 1975, Wright and Spates 1976, Borkovec et al. 1978). Wright et al. (1979) dipped adult weevils in an acetone solution of diflubenzuron, which rendered the females infertile after the adults were irradiated (mode of action described in detail subsequently). However, Earle et al. (1979a) reported that the flight ability of irradiated male boll weevils treated with diflubenzuron in acetone was apparently impaired by the acetone. Also, Haynes et al. (1981) repo>-ted that the acetone decreased flight activity.
Fractionated irradiation treatment of young adult weevils was compared with acute irradiation of aged boll weevils by Haynes et al. (1979), who reported that mortality was less when weevils were treated 6 days after emergence. Earle et al. (1978) showed that acute irradiation of adults was at least as effective a procedure as fractionated irradiation of pupae. Subsequently, Haynes et al. (1978) evaluated the possibility of sterilizing large numbers of

BOLL WEEVIL STERILITY 157
weevils by exposing the pupal and early adult stages to fraction- ated doses of irradiation while they were still in the larval medi- um. This procedure was eventually eliminated from consideration because of the complexity of the problems involved in handling, irradiation, and transportation.
Wright et al. (1979) then examined the effects of aging the adults before exposing them to sterilizing procedures and compared the mating ability, sterility, and survival of weevils treated by irradiation or fumigation. They determined that more males were available to mate after the irradiation treatment because fumigated males had significantly higher mortality at 7 days posttreatment. Next, Wright et al. (1980a) compared the three most promising tech- niques for sterilization of the boll weevil: (1) irradiation in a nitrogen atmosphere plus dipping 6-day-old adults in a 0.1% di- flubenzuron solution; (2) irradiation of pupae with fractionated doses; and (3) fumigation of adults with bisazir plus dipping 6- day-old adults in penfluron. The results indicated that technique (1) was the best experimentally. Both sexes of weevils were steri- lized when diflubenzuron was administered in the adult diet in combination with irradiation or fumigation, though a dose of 500 ppm of diflubenzuron appeared to reduce sperm transfer and the antibiotics caused reduced production of pheromone by the males. In contrast, males fumigated for 90 minutes in bisazir and then dipped in an acetone solution containing 1.35% penfluron (Borkovec et al. 1978) were 23% as competitive as normal males.
When adult boll weevils were held for 5 days after emergence on diet containing 100 ppm diflubenzuron and then exposed on the 6th day to 10 krad of gamma irradiation from a cesium-137 source in a nitrogen atmosphere, both males and females were sterile (Wright et al. 1980b). There was no indication that this lower level of diflubenzuron affected sperm transfer or mating behavior. Moore et al. (1978) fed males 500 ppm of diflubenzuron in adult diet for 3 days or dipped males in a 0.6% acetone solution and observed no ad- verse effects on sperm transfer. Earle et al. (1978) reported that diflubenzuron reduced matings as much as 40-50% when adults were dipped in 0.01-0.1% acetone solutions, though such dipping did not affect the mating performance (Earle et al. 1979a).
Villavaso et al. (1977) stated that an acetone solution of di- flubenzuron applied to 4-day-old males following irradiation did not affect insemination of females by males.
In summation, the holding of both sexes of weevils on a slab of adult diet for 6 days so they could age before irradiation al- lowed mating to occur, but the addition of the diflubenzuron to the diet meant that this material was incorporated within the eggs of the females so the eggs did not hatch (Ivie and Wright 1978, Bull and Ivie 1980). The 10 krad of gamma irradiation that was then ad- ministered caused all the insects to die within 12-15 days post- treatment. The mortality occurred early enough to prevent recovery of fertility by the released adults.

"•58 j. E. WRIGHT AND E. VILLA VASO
This latest method of sterilizing boll weevils was now tested against 1,350,000 mass-reared weevils. When the eggs (1,701,140) produced by these treated adults were collected and evaluated for fertility (treated x treated crosses), the sterility factor was 99.999% (Wright and Roberson, 1981). Moreover, the 0.001% fertil- ity was believed to result from contamination by colony adults since the experiments were done in the mass-rearing facility where isolation was incomplete. The same procedure for sterilizing wee- vils was field tested in an isolated cotton planting in Florida (Mitchell et al. 1980). The more than 600,000 sterile boll weevils released there did not establish a population in the field.
COMPETITIVENESS OF STERILIZED WEEVILS
The detrimental effects of irradiation on the vigor of male boll weevils has been well-documented (Davich and Lindquist 1962, Flint et al. 1966). The impact of this reduced vigor on the com- petitiveness of treated males was equally evident. For example, Davich et al. (1965) reported that the mating competitiveness of irradiated males in a field-cage test was 20% that of normal males.
Chemosterilization too results in reduced vigor. Villavaso and Earle (1976) showed that males sterilized by feeding on a diet containing 0.08% busulfan for 6 days mated with only one-half as many females as did normal males in field tests. Also, Haynes et al. (1975), in laboratory tests, found that the mating competitive- ness of males sterilized by feeding on diet containing 0.08% busul- fan plus 0.4% hempa for 6 days was 25-33% that of normal males.
In 1977, Villavaso et al. (1979) devised field tests to deter- mine estimates of the competitiveness of sterile male boll weevils. Sterile and fertile males and fertile virgin females were released into isolated, weevil-free plots of cotton. After 7 days, squares with oviposition punctures were collected and incubated in the lab- oratory, and percentage egg hatch was determined. This experimen- tally determined value for percentage egg hatch in the field was then used along with the values for percentage egg hatch obtained from crosses between sterile males and normal females and normal males and normal females in estimating competitiveness by the for- mula of Fried (1971) in which no assumptions were made concerning the factors involved in the degree of competitiveness achieved as:
where
(——TT-) (-77) X 100 = % competitiveness Ee-Hs S ^
Ha = % egg hatch of N^î x N9 Hs = % egg hatch of S^ x N9 Ee = experimentally determined egg hatch from the
field N = the number of normal males used S = the number of sterile males used

BOLL WEEVIL STERILITY 159
Males sterilized by the pupal fractionation treatment of Haynes et al. (1977) were 23% as competitive as normal males. How- ever, males allowed to remain on the surface of the larval diet for 3-4 days after emergence were 36% as competitive as normal males when they were then sterilized by 7 krad of gamma irradiation fol- lowed by a 5-second dip in a 0.02% solution of diflubenzuron in acetone (R. A. Leopold, D. T. North, personal communication, Earle et al. 1978).
Males given 10 krad of gamma irradiation in a nitrogen atmos- phere and then dipped in an acetone solution containing 0.1% diflu- benzuron were 17% as competitive as normal males; the competitive- ness of males treated by the pupal fractionation treatment was 12% (Villavaso et al. 1980). The nitrogen was used because it anes- thetized the weevils so they were easier to handle. Furthermore, Earle et al. (1979b) showed that male mating capability was approx- imately the same whether the males were given 10 krad in nitrogen or 8 krad in air.
RELEASE OF STERILE MALES IN BOLL WEEVIL ERADICATION TRIAL
The procedure adopted in preparing the sterile boll weevils for release during 1979 in the Boll Weevil Eradication Trial (BWET) was as follows: 13-14 days after the eggs were implanted in the artificial medium, the newly emerged adults were collected into 30- X 40-centimeter, nylon marquisette bags (about 6000 per bag) that were placed on top of diet slabs (32 x 42 x 1 centimeters). These slabs had been treated by mixing antibiotics (streptomycin (0.01%), neomycin sulfate (0.01%), and polpet (0.03%) with sand and grits (Grit-o-Cobs 0-60) and spreading the mixture thinly over the surface of the slabs to prevent bacterial contamination. The second day, the bags were lifted, and slabs were turned over. New slabs were provided the third and the fifth days. The sixth day, the adults were chilled in a walk-in refrigerator (ca. 4°C) and placed in a seed separator for removal of dead adults. Then they were exposed to the 10 krad of irradiation, packaged, and shipped to North Carolina by commercial air carrier for release.
In all, 11 million adults were mass reared, sterilized, and released in the BWET in 1979. No evidence of reproduction attrib- utable to the released sterile boll weevils was found. Adults were exposed to a single dose of 10 krad of gamma irradiation in a cesium-137 source (3107 ± 5% curie). The doses were monitored by Fricke dosimetry and thermoluminescent dosimeters (Fricke and Hart 1966). The nitrogen atmosphere was used during irradiation to lower oxygen tension and according to Earle et al. (1979b) should have enhanced the competitiveness of the males.

^^^ J. E. WRIGHT AND E. VILLA VASO
SELECTION OF ARTIFICIAL DIET FOR MASS REARING
Vanderzant and Davich (1958) developed the first artificial diets for the boll weevil. Other diets were developed by several researchers including Earle et al. (1966), Sterling et al. (1965), Gast and Davich (1966), and Lindig and Malone (1973). However, most of the diets were designed for use with small laboratory colo- nies, and there were some questions concerning their suitability for mass production of boll weevils, including (1) cost, (2) the limited supply of specialized ingredients, and (3) possible incom- patibility with mechanized equipment.
Three experimental diets appeared to fit all parameters neces- sary for mass rearing boll weevils. One was a modification of a diet devised by Gast and Davich (1966) (Gast diet), one was a diet devised by Lindig and Malone (1973) (HG), and one was the cotton- seed flour diet (CSF) of R. F. Moore (unpublished information). The composition of these three diets is indicated in Table 1.
Because of the large numbers of adults required for sterile insect releases, these three diets were evaluated by feeding each to 3 million adult weevils per week. Eggs were mechanically ex- tracted from the diet pellets (Griffin and Lindig 1977), surface sterilized with 10% formalin solution (Sikorowski et al. 1977), and implanted in congealed larval test diets by spraying. (Concur- rently, small-scale tests were also conducted.) Data for the large-scale test (Table 2) showed that significantly more adults were produced from each tray of Gast diet (382.9) than from trays of the HG diet (326.2) or the CSF diet (198). No differences were detected in the small-scale tests. In the large-scale test, the average numbers of days necessary to obtain 80% of the total adult emergence from the Gast, CSF, and HG diets were 4.7, 4.6, and 4.2, respectively. The averages for the small-scale test were 4.3, 4.3, and 3.8 days, respectively. Adults emerging from the HG diet were slightly larger (12.8 mg) than adults emerging from the CSF (12.6) or Gast (12.1) diets. (Also, the weights of adults fed on these three diets for 15 days were 15.8, 16.9, and 17.1 mg, respec- tively.) In the concurrent small-scale tests, females exposed to the HG diet laid a significantly higher average number of eggs per female per day for 12 days (13.2) than females exposed to the CSF diet (11.7) or the Gast diet (12.3), but the average dropped to ca. 5.5 eggs per female per day when the females were handled according to mass-production procedures. Thus mechanical extraction and the handling of eggs in mass greatly reduced the number of eggs recovered.
The Gast diet produced more adults at the least cost (Table 2), but the HG diet consistently had a higher percentage of eggs hatching. Otherwise, there were few differences among the diets in ease of handling in the large-volume mechanized procedures (flash-sterilizing units, tray forming and filling, or diet prepa- rations) except that the dry ingredients used in the CSF diet were extremely dusty. This constituted a potential hazard, and more

BOLL WEEVIL STERILITY 161
Table 1.—Composition of 3 artificial diets for mass rearing of the boll weevil
Gast CSF 1/ HG \_l
Adult and Adult Larval Adult Larval larval
Ingredients diet diet diet diet diets
Agar (grams) 80 78.6 80.4 88.5 80 Sugar (grams 65 74 178.9 179 60
Wesson salt (grams) 10.5 12 13.2 13.1 10.5
Cholesterol (grams) 2.75 3.4 2.24 2.21 1.0 Vitamin mix (ml) 10.8 12 2.0 1.0 10.0
Ascorbic acid (grams ) 2.5 2.75 7.16 7.08 5.0
Potassium 5.4 6.0 2.2 8.7 5.4
sórbate (grams) Methyl paraben
(grams) 3.5 3.8 3.35 6.64 3.5 Cottonseed 265.0 310.75 meal (grams)
Cottonseed flour (grams) 223.6 221.3
Underflow (grams) 200 Cottonseed 100 101.75 meats (grams)
Promine D (grams) 100 116.5 100
Casein (grams) 150
Wheat germ (grams) 100
Corn cob grits (grams) 56 Choline chloride
(grams) 4.47 4.43
Tocopherol (grams) 0.72 0.71
Zinc acetate (grams) 0.044 0.044
Cobalt chloride (grams) 0.022 0.022
Sodium molybdate (grams) 0.022 0.022
Wheat germ oil (ml) 13.42 13.12
Methenamine (grams) 0.75 2/0.75
Biotin solution 0.6 0.6 0.6
(0.1%)(ml) B,^ solution 0.6 0.6 ^t0.01%)(ml)
HCL 6 N (ml) 4 4 Formaldehyde (ml) Water (ml) 3800 3800
1.12 1.15 3800 3800
0.6
4
3800
1/ CSF = cottonseed flour; HG = diet devised by Lindig and Malone (l'973).
2/ Adults only.

162 j. E. WRIGHT AND E. VILLA VASO
Table 2.—Production of adult boll weevils from test diets, mass-production procedures
Mean Mean cost Mean num~ weight of diet ber of of Mean % Mean % needed to adults emerging of 100 of traps produce
emerging weevils eggs yield- contami- 1000 adults 3/ Diet 1/ per tray 2/ (mg) ing adults nated (dollars)
Gast 382.9 a 12.1 15.4 2.1 0.21 CSF 198.0 c 12.6 8.1 1.4 0.58 HG 326.2 b 12.8 12.9 1.2 0.31
j_/ CSF = cottonseed flour; HG = diet devised by Lindig and Malone (1973).
Ij Means followed by different letters are significantly differ- ent at the 5% level of probability as determined by Duncan's new multiple range test.
_3/ Includes cost of tray and cover.
labor and equipment were needed for the dry-mixing stage of the CSF diet. The Gast diet was selected as the mass-rearing diet, and all insects used in the BWET were reared on this diet.
GENETICS OF STRAIN
Bartlett (1967) selected a strain of boll weevils with an ebony characteristic; i.e., it is identifiable by ebony color of the entire body of the adult. The trait is semidominant and re- quires selection and backcrossing to maintain. The genotypes and progeny phenotypes resulting from possible crosses are listed in Table 3. Jenkins et al. (1972) introduced a pearl-eyed ebony characteristic into the maintenance colony in 1969, but the pearl- eyed characteristic was removed from the colony in 1970-71 because behavioral characteristics indicated that the pearl-eyed mutant was not adapted for use in the field.
An ebony colony that was to provide identifiable weevils for the sterile releases in the BWET was set up in 1971 by collecting native specimens from North Carolina and crossing them with the laboratory ebony colony. By 1977, the ebony characteristic in the colony was only about 50%; thus, in October 1978, ebony adults were selected from pupal cells and a selected, isolated colony was begun for the ebony body characteristic. In 1979, when the adult boll weevils were sterilized and released in the BWET, 99% had the ebony trait.

BOLL WEEVIL STERILITY 163
Table 3.—Genotypes and progeny phenotypes resulting from crosses of ebony (e) and wild-type body (+)
Genotype Phenotype
e/e e/+ +/+ e/e X e/e e/e X e/+ e/+ X e/+ e/e X + /+ e/+ X +/+ + /+ X + /+
Ebony - black Bronze - (lOR 3/4 Munsell Notation) Wild-type (lOR 4/8 Munsell Notation) All ebony 1 ebony: 1 bronze 1 ebony: 2 bronze: 1 wild-type All bronze 1 bronze: 1 wild-type All wild-type
ADULT MARKING WITH CALCO OIL RED
Calco oil red N1700 dye was first used to mark the boll weevil by Gast and Landin (1966) who incorporated 1000 ppm within the larval diet. The result was that the oil was found within the fat bodies of the emerging adults. Although the dye had no adverse effects when small numbers of weevils were reared, Wilkinson et al. (1972) reported that it did produce significant differences in the pupal weight, adult longevity, and egg production of three lepidop- teran species. Since the dye might therefore affect the quality of mass-reared adult weevils, we analyzed its effects on the steril- ity, mortality, and pheromone production of mass-reared weevils. For this test, the dye, 550 ppm, was dissolved in ca. 100 ml of corn oil or wheat germ oil and added to standard larval diet (Lin- dig 1979) that had been flash-sterilized according to the proce-
dures of Griffin et al. (1974). Normal mass-rearing procedures (Roberson and Wright 1982) were used in forming trays, dispensing diet, and implanting eggs. Trays of control diet (no dye) were prepared and planted the same day for comparison. Weight of emerg- ing adults, oviposit ion, and hatch data were determined according to the procedure described by Lindig (1979); that is, the usual wax-coated adult diet pellets were fed to both marked and unmarked adults for the oviposition and hatch tests. The adults were then held on diet slabs for 6 days and exposed to 10 krad. The proce- dures used to determine the sterility, egg hatch, and mating abil- ity of both types of weevils were those described by Wright et al. (1979). Pheromone production was quantitated by the method of McKibben et al. (1976). Frass collected daily for days 1-4 and 5-8 was combined for extraction and analysis. Also, acetylcholines- terase and B^carboxylesterase enzyme activities in the 5-day-old dyed and control insects were determined according to the methods of Ellman et al. (1961) and Chambers (1973), respectively.
164 J. E. WRIGHT AND E. VILLA VASO
Table 4.—Effects of incorporating red dye in larval diet of boll weevils
No. % mor- Adults eggs Weight (mg) tality
per per % of emerg- at 14 tray day hatch ing adults days
Diet (±X SD) (±X SD) (±X SP) (±X SP) (±X SP)
Standard 646.1±148.1 8.6±1.4 72.6±4.2 13.2±0.4 8.5±4.9 Standard
+ dye 597.1±110.3 9.5±1.1 74.7±3.6 13.0±0.2 8.3
The results showed no significant differences between the dyed weevils and the control weevils (Lindig et al. 1980) (Table 4).
The Calco oil red dye provided a positive marking of all adults. They could easily be identified in the field or elsewhere by visual examination of the thin dorsal coverings of the abdomen under the elytra. Also, moribund adults could be identified by grinding them in a small amount of acetone and observing the red- dish color. Still more importantly, the eggs produced by the ster- ilized released females were dyed and therefore distinguishable from the eggs oviposited by native females in cotton squares. All sterile adults released in the BWET had been fed the calco oil red dye.
IRRAPIATION SOURCE
The cobalt-60 irradiation source at the Boll Weevil Research Laboratory, which had an output of 356 krad per minute, was used in the first efforts to sterilize the boll weevil by irradiation. However, this source had limited chamber space in which to place insects. A cesium-137 source was purchased subsequently so frac- tionated doses of irradiation could be applied to boll weevil pupae (Flint et al. 1966, Haynes et al. 1977). This cesium-137 source has a relatively low rad output—120 rad per minute—but its cham- ber capacity is large enough to accommodate ca. 125,000-150,000 adults. On the other hand, it took ca. 90 minutes in the cesium source to administer 10 krad and only ca. 15 minutes in the cobalt source. Thus the difference in the energy levels of radiation to which the adult boll weevils were exposed and the time required to reach the necessary dose of 10 krad could have an effect on the longevity and sterility of the treated weevils. The sensitivity of male and female boll weevils to lethal effects of gamma irradia- tion in the two chambers was evaluated by exposing 2-, 3-, and 6-day-old adults (sexes held separately after treatment) to 0, 1250, 1570, 2480, 3140, 4970, and 6800 rads. There were no signif- icant differences in the biological effects of the cesium and cobalt sources at the levels tested (Abdul Matin et al. 1980).

BOLL WEEVIL STERILITY 165
EVALUATION OF THE STERILE BOLL WEEVILS RELEASED IN THE BOLL WEEVIL ERADICATION TRIAL IN 1979
Mortality of sterile weevils held in the rearing laboratory was 50% at 4.6 days, 70% at 5.4 days, and 99% at 8.4 days. Mortal- ity of sterile weevils caged in the field was 50% at 5.8 days, 70% at 7.2 days, and 99% at 11.7 days. However, in the Florida test, mortality of sterile weevils held in the laboratory (50, 70, and 87% after 4.1, 5.2, and 9.1 days, respectively) was similar to that of weevils held in the field (50, 70, and 99% at 7.4, 8.7, and 13.2 days, respectively). Mortality was always directly correlated with the effects of the irradiation on the destruction of the midgut epithelial cells. Irradiation treatment also produced a decline in vigor the 7th day (Nilakhe and Earle 1976, Earle et al. 1978).
Sperm transfer by treated males taken at random from each of the 29 shipments to North Carolina averaged 46.6% compared with 58% for the controls. This was satisfactory since irradiated boll wee- vils produce no sperm 5 days after treatment (Nilakhe and Earle 1976), and released sterile males had the ability to transfer sperm the day they were released in the cotton field. Normal male boll weevils usually attain peak mating activity after they are 4 days old (Cross 1973).
According to McKibben et al. (1976) and Gueldner and Wiygul (1978), male boll weevils produce little pheromone (<20 nanograms per male) until they are more than 5 days old. McGovern et al. (1975) reported that exposure to 10 krad of gamma irradiation did not affect pheromone production by the males until about the sixth day after treatment; then production was only about one-third that of the controls. Earle et al. (1978) reported similar results. This decrease in pheromone was one of the reasons adult weevils were aged before release; the released steriles should be capable of pheromone production the day of release. McGovern et al. (1975) and Villavaso et al. (1979) showed that males produced pheromone on days 0-3 after emergence if they were fed fresh cotton squares.) The sterile male boll weevils released during the BWET averaged 290 nanograms pheromone per day when they were held in the laboratory; similar unsterilized males averaged only 221 nanograms per male per day (Wright and Thomas 1981).
The ebony marker and the presence of calco oil red dye within the adult boll weevils made it easy to identify the sterile weevils released in the BWET. Only 1.2% of the released weevils were bronze and only one wild-type weevil was observed among those sam- pled. Projection of these figures indicates that ca. 450 wild-type and 158,818 bronze weevils were present among the 13,050,000 ster- ile weevils shipped to North Carolina.
The flight behavior of adult weevils released in the BWET was tested to determine their locomotor activity because earlier, when diflubenzuron was applied in an acetone dip, movement was severely curtailed (19% flight v. 39% for controls) (Haynes et al. 1981).

166 J. E. WRIGHT AND E. VILLAVASO
The data for the sampled weevils showed that an average 28% of the sterile insects and 49% of the controls (untreated) were capable of flight. M. C. Ganyard (personal communication) of Animal and Plant Health Inspection Service reported that 76,000 of the released sterile insects were captured in traps at distances of >400 meters from their release points.
The High Pressure Liquid Chromâtography analysis of difluben- zuron in the treated boll weevils at day 6 showed that an average 182 and 221 nanograms were present in the released males and fe- males, respectively. Additional analysis of eggs laid by these weevils indicated that ca. 8.5 ppm diflubenzuron was present in the eggs. Both findings are important. In the first place, the incor- poration of diflubenzuron in the adult diet makes it possible to sterilize both sexes of boll weevils (Wright et al. 1980b, Wright and Roberson 1981) since diflubenzuron inhibits the synthesis of chitin and interferes with larval development in eggs of treated females as long as the material is present within the females. Ir- radiation then prevents further oogénesis after the initial eggs containing diflubenzuron are oviposited, which occurs within 4 days after treatment in the laboratory (Wright and Roberson 1981). In the second place, as Bull and Ivie (1980) noted in a study on the fate and activity of diflubenzuron in the boll weevil, 1.0 ppm diflubenzuron must be present in the egg to inhibit hatch.
The sterile boll weevils released in the BWET in the North Carolina-Virginia area in 1979 had been sterilized by acute irradi- ation plus diflubenzuron treatment. The method was selected be- cause of its simplicity and because of the potential health hazard associated with the fumigation treatment. Also, the diflubenzuron was fed (100 ppm for 5 days before irradiation) to newly emerged weevils (Wright et al. 1980b) because of the adverse effects of acetone on flight ability discovered in 1978 (Earle and Simmons 1979). Samples of weevils released in the BWET were observed to be 12% as competitive as normal males (Villavaso 1981).
In 1980 and 1981 other weevils treated with the same method were evaluated for competitiveness in Louisiana: treated weevils averaged 7 and 12%, respectively, over the 7-day posttreatment period (Villavaso unpublished data). The sterile weevils were 23% as competitive as untreated weevils for days 0-4 and 14% as compet- itive for days 0-5 after release. In addition, mortality and mat- ing studies of laboratory-held and caged males showed that the sterile males were most effective for the first 4 days (Villa- vaso 1981) (Table 5). The combined competitiveness data would sup- port the release of steriles for better effectiveness at intervals of 5 days rather than every 7 days.
The relative effectiveness of released sterile boll weevils with different ratings for competitiveness is difficult to demon- strate in the field, especially when native populations are very low and eradication is the goal. However, theoretical
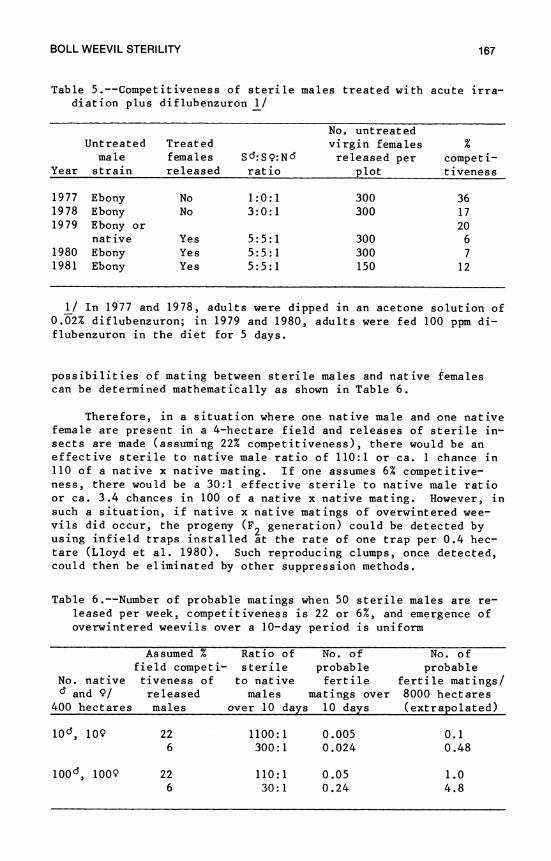
BOLL WEEVIL STERILITY 167
Table 5.—Competitiveness of sterile males treated with acute irra- diation plus diflubenzuron 1/
Untreated Treated male females ScJ:S9:N<?
Year strain released ratio
1977 Ebony No 1:0:1 1978 Ebony No 3:0:1 1979 Ebony or
native Yes 5:5:1 1980 Ebony Yes 5:5:1 1981 Ebony Yes 5:5:1
No, untreated virgin females % released per competi- plot tiveness
300 300
300 300 150
36 17 20 6 7
12
\J In 1977 and 1978, adults were dipped in an acetone solution of 0.02% diflubenzuron; in 1979 and 1980, adults were fed 100 ppm di- flubenzuron in the diet for 5 days.
possibilities of mating between sterile males and native females can be determined mathematically as shown in Table 6.
Therefore, in a situation where one native male and one native female are present in a 4-hectare field and releases of sterile in- sects are made (assuming 22% competitiveness), there would be an effective sterile to native male ratio of 110:1 or ca. 1 chance in 110 of a native x native mating. If one assumes 6% competitive- ness, there would be a 30:1 effective sterile to native male ratio or ca. 3.4 chances in 100 of a native x native mating. However, in such a situation, if native x native matings of overwintered wee- vils did occur, the progeny (F^ generation) could be detected by using infield traps installed at the rate of one trap per 0.4 hec- tare (Lloyd et al. 1980). Such reproducing clumps, once detected, could then be eliminated by other suppression methods.
Table 6.—Number of probable matings when 50 sterile males are re- leased per week, competitiveness is 22 or 6%, and emergence of overwintered weevils over a 10-day period is uniform
Assumed % Ratio of No. of No. of field competi- - sterile probable probable
No. native tiveness of to native fertile fertile matings/ ^ and 9/ released males matings over 8000 hectares
400 hectares males over 10 days ; 10 days (extrapolated)
10^, 109 22 1100:1 0.005 0.1 6 300:1 0.024 0.48
100^, 1009 22 110:1 0.05 1.0 6 30:1 0.24 4.8

168 J. E. WRIGHT AND E. VILLA VASO
When sterile boll weevils are to be used in efforts to eradi- cate, they should be used against an emerged overwintered popula- tion that has been reduced to a low level by other suppression technology. Once most of the overwintered weevils have emerged, releases of sterile weevils could be terminated, and a pheromone trapping system (Chapter 8 in this handbook) could be installed to detect reproducing clumps.
Releases of sterile boll weevils could also be considered for pest management since we now have reliable insect rearing and im- proved sterilizing procedures. Additional research is needed to determ.ine how such releases can be used as a component of existing pest management program-s for cotton.
REQUIREMENT FOR USING STERILE INSECTS
The analyses of the PBWEE by Whitten and Foster (1975), Eden et al. (1973), the National Academy of Sciences (1975), and Perkins (1980) all included remarks to the effect that technical shortcom- ings in the degree of sterility and behavioral competitiveness of the released sterilized weevils had seriously limited that eradica- tion experiment. Knipling (1979) listed the conditions he felt had to be met before the sterile male technique could be used in an eradication attempt. These were:
(1) Mass rearing of quality insects. (2) Method of sterilizing that produces males that are suf-
ficiently competitive. (3) Quantitation of natural populations of insects to deter-
mine numbers of sterile insects that should be released. (4) Distribution of released insects at the most opportune
time so that they will be spatially competitive with the natural populations for mating.
(5) Information available concerning the normal population dynamics at the time of release.
(6) Information concerning the degree of infiltration of wee- vils from areas outside the test area and its impact up- on the technology being used.
(7) Prior analysis of costs, effectiveness, and ecological effects of this autocidal method as opposed to other available methods.
(8) Information concerning the effects of released insects on crops, health, and man.
These requirements are formidable and indicate that sterile releases or other autocidal techniques cannot provide a practical way to deal with most pest problems. At the same time, the tech- nique does have potential in managing some important pests, espe- cially when the method is integrated with other suppression methods.

BOLL WEEVIL STERILITY 169
Knipling (1979) has pointed out that sterile insects may be useful in four situations:
(1) For suppression or elimination of low-level natural popu- lations.
(2) For elimination of incipient populations in a new area. (3) For prevention of the establishment of a new population
in a given area. (4) For pest management programs.
The sterile boll weevil should now be considered a candidate for these situations.
The effort to induce sterility in male and female boll weevils has extended over more than two decades. The search was for a treatment that (1) would not destroy the regenerative cells of the midgut so digestive enzjrmes would be produced; (2) would destroy all spermatogonia and oogonia so sterility would be permanent; and (3) would induce dominant lethal mutations in all gametes. The sterilization method developed and utilized sterilizes both sexes and yields a male that is about 23% competitive during the first 5 days or so after release. The effects of the irradiation are then manifested, competitiveness decreases, and the boll weevils die within 14 days.

170 J. E. WRIGHT AND E. VILLAVASO
LITERATURE CITED
Abdul Matin, A. S. M., J. E. Wright, and T. B. Davich. 1980. Effect of low levels of gamma irradiation on longevity and
fertility of the boll weevil. Southwestern Entomology 5:112- 117.
Bartlett, A. C. 1967. Genetic markers in boll weevil. Journal of Heredity 58:
159-163.
P. A. Hooker, and D. D. Hardee. 1968. Behavior of irradiated boll weevils. 1. Feeding, attrac-
tion, mating, and mortality. Journal of Economic Entomology 61:1677-1680.
Borkovec, A. B. 1962. Sexual sterilization of insects by chemicals. Science
137:1034-1037.
and D. G. McHaffey. 1977. Chemosterilization of the boll weevil by fumigation. Journal of Economic Entomology 70:424-426.
C. W. Woods, and P. H. Terry. 1978. Boll weevil: Chemosterilization by fumigation and dip-
ping. Journal of Economic Entomology 71:862-866.
Bull, D. L., and G. W. Ivie. 1980. Activity and fate of diflubenzuron and certain derivatives
in the boll weevil. Pesticide Biochemistry and Physiology 13: 41-52.
Chambers, H. 1973. Thiol esters as substrates for insect carboxylesterases. Annals of the Entomological Society of America 66:663-665.
Cross, W. H. 1973. Biology, control, and eradication of the boll weevil. An-
nual Review of Entomology 18:17-46.
Davich, T. B. 1976. Boll weevil sterility. In Boll Weevil Suppression, Man-
agement, and Elimination Technology. U.S. Department of Agri- culture, ARS-S-71, p. 53-58.
J. C. Keller, E. B. Mitchell, P. Huddleston, R. Hill, D. A. Lindquist, G. McKibben, and W. H. Cross.
1965. Preliminary field experiments with sterile males for erad- ication of the boll weevil. Journal of Economic Entomology 58: 127-131.

BOLL WEEVIL STERILITY 171
Davich, T. B. and D. A. Lindquist. 1962. Exploratory studies on gamma radiation for the steriliza-
tion of the boll weevil. Journal of Economic Entomology 55: 164-167.
M. E. Merkl, E. B. Mitchell, D. D. Hardee, R. T. Gast, G. H. McKibben, and P. M. Huddleston.
1967. Field experiments with sterile males for eradication of the boll weevil. Journal of Economic Entomology 60:1533-1538.
Earle, N. W., and R. Leopold. 1975. Sterilization of the boll weevil: Vacuum fumigation with hempa combined with feeding busulfan-treated diet. Journal of Economic Entomology 68:283-286.
S. S. Nilakhe, and L. A. Simmons. 1979a. Mating ability of irradiated male boll weevils treated with diflubenzuron or penfluron. Journal of Economic Ento- mology 72:334-336.
and L. A. Simmons. 1979. Boll weevil: Ability to fly affected by acetone, irradia-
tion, and diflubenzuron. Journal of Economic Entomology 72: 573-575.
L. A. Simmons, and S. S. Nilakhe. 1979b. Laboratory studies of sterility and competitiveness of
boll weevils irradiated in an atmosphere of nitrogen, carbon dioxide, or air. Journal of Economic Entomology 72:687-691.
L. A. Simmons, S. S. Nilakhe, E. J. Villavaso, G. H. McKibben, and P. Sikorowski.
1978. Pheromone production and sterility in boll weevils: effect of acute and fractionated gamma irradiation. Journal of Eco- nomic Entomology 71:591-595.
A. B. Walker, and M. L. Burks. 1966. Development of an artificial diet based on the analysis of
amino acids in cotton squares. Annals of the Entomological Society of America 59:664-669
Eden, W. G., H. C. Chiang, E. H. Glass, D. L. Haynes, P. Oman, and H. T. Reynolds.
1973. The pilot boll weevil eradication experiment. Bulletin of the Entomological Society of America 19:218-221.
Ellman, G. L., K. D. Courtney, V. Andres, Jr., and R. M. Featherstone.
1961. A new and rapid colorimetric determination of acetylcho- linesterase activity. Pharmacology (Basel) 7:88-95.

172 J. E. WRIGHT AND E. VILLAVASO
Flint, H. M., W. R. Bibow, and 0. K. Labren. 1966. Radiation studies witb tbe boll weevil: Letbal effects
on larvae, pupae, and adults: male sterility and dose frac- tionation. Journal of Economic Entomology 59:1249-1255.
Fricke, H., and E. J. Hart. 1966. Chemical dosimetry. In Radiation Dosimetry, p. 167-239. Volume 2: Instrumentation, 2nd edition. F. H. Attix and W. C. Roesch, editors. Academic Press, New York.
Fried, M. 1971. Determination of sterile insect competitiveness. Journal
of Economic Entomology 64:869-872.
Gassner, G., D. Childress, G. Pomonis, and J. Eaton. 1974. Boll weevil chemosterilization by hypobarometric distilla-
tion. Journal of Economic Entomology 67:278-280.
Gast, R. T., and T. B. Davich. 1966. Boll weevils. Ln Insect Colonization and Mass Production,
p. 405-419. C. N. Smith, editor. Academic Press, New York.
and M. Landin. 1966. Adult boll weevils and eggs marked with dye fed in the
larval diet. Journal of Economic Entomology 59: 474-475.
Griffin, J. G., and 0. H. Lindig. 1977. System for mechanical harvesting of boll weevil eggs from
diet pellets. Transactions of the ASAE 20:254-256.
0. H. Lindig, and R. E. McLaughlin. 1974. Flash sterilizers: Sterilizing diets for insects. Jour-
nal of Economic Entomology 67:689.
Gueldner, R. C, and G. Wiygul. 1978. Rhythms in pheromone production of the male boll weevil.
Science 199:984-986.
Haynes, J. W. 1963. Chemical sterility agents as they affect the boll weevil, Anthonomus grandis Boheman. M.S. thesis, Mississippi State University, State College, MS, 38 p.
Haynes, J. W., and E. B. Mitchell. 1977. Fractionated irradiation of boll weevils during pupal de- velopment: effect of sperm depletion and transfer as measured by female responsiveness. Journal of Economic Entomology 70:411-412.

BOLL WEEVIL STERILITY 173
Haynes, J. W., N. Mitlin, T. B. Davich, J. R. Dawson, W. L. McGovern, and G. H. McKibben.
1977. Sterilization of boll weevil pupae with fractionated doses of gamma irradiation. Entomologia Experimentalis et Applicata 21:57-62.
N. Mitlin, T. B. Davich, B. J. Nail, and J. R. Dawson. 1975. Mating and sterility of male boll weevils treated with busulfan plus hempa. Environmental Entomology 4:315-318.
N. Mitlin, C. E. Sloan, and J. R. Dawson. 1972. Busulfan: Development of improved methods of sterilizing boll weevils. Journal of Economic Entomology 66:619-622.
J. E. Wright, T. B. Davich, J. Roberson, J. G. Griffin, and E. Darden.
1978. Boll weevil: Experimental sterilization of large numbers by fractionated irradiation. Journal of Economic Entomology 71:943-946.
W. L. McGovern, and J. E. Wright. 1981. Diflubenzuron (solvent-water suspension) dip for boll weevils: effects measured by flight, sterility, and sperm transfer. Environmental Entomology 10:492-495.
J. E. Wright, and E. Mattix. 1979, Fractionated vs acute irradiation: The effects of treat-
ing adult boll weevils (Coleóptera: Curculionidae) at different ages. Entomologia Experimentalis et Applicata 26:61-66.
Hedin, P. A., C. P. Cody, and A. C. Thompson, Jr. 1964. Antifertility effect of the chemosterilant apholate on the male boll weevil. Journal of Economic Entomology 57:270-272.
Ivie, G. W., and J. E. Wright. 1978. Fate of diflubenzuron in the stable fly and house fly. Journal of Agricultural and Food Chemistry 26:90-94.
Jenkins, J. N., J. C. McCarty, Jr., W. L. Parrott, 0. H. Lindig, and R. E. McLaughlin.
1972. Genetic characteristics of an ebony pearl strain of boll weevils. Journal of Economic Entomology 65:1621-1624.
Klassen, W., and N. W. Earle. 1970. Permanent sterility produced in boll weevils with busulfan without reducing production of pheromone. Journal of Economic Entomology 63:1195-1198.
J. F. Norland, and A. B. Borkovec. 1968. Potential chemosterilants for boll weevils. Journal of Economic Entomology 61:401-407.

174 J. E. WRIGHT AND E. VILLAVASO
Klassen, W., J. F. Norland, and R. W. Briggs. 1969. Sterilization of boll weevils with combinations of chemo-
sterilants and X-rays, gamma rays, thermal neutrons, or fast neutrons. Journal of Economic Entomology 62:1204-1216.
Knipling, E. F. 1955. Possibilities of insect control or eradication through the
use of sexually sterile males. Journal of Economic Entomology 48:459-462.
1960. The eradication of the screwworm fly. Scientific American 203:54-61.
1964. The potential role of the sterility method for insect population control with special reference to combining this method with conventional methods. U.S. Department of Agricul- ture, ARS-33-98, 54 p.
1976, Report of Technical Guidance Committee for Pilot Boll Weevil Eradication Experiment. In Proceedings of a Conference on Boll Weevil Suppression, Management, and Elimination Tech- nology. U.S. Department of Agriculture, ARS-S-71, p. 122-126.
1979. The basic principles of insect population suppression and management. U.S. Department of Agriculture, Agriculture Hand- book No. 512, 623 p.
Labrecque, G. C. 1961. Studies with three alkylating agents as house fly steril-
ants. Journal of Economic Entomology 54:684-689.
Lindig, 0. H. 1979. A replacement for cottonseed meal and meats in boll weevil
diets. Journal of Economic Entomology 72:291-292.
and 0. L. Malone. 1973. Oviposition of boll weevils fed diets containing germi- nated cottonseed puree or cottonseed meats puree. Journal of Economic Entomology 66:566-567.
G. Wiygul, J. E. Wright, J. Roberson, and J. R. Dawson. 1980. Rapid method for mass-marking boll weevils. Journal of Economic Entomology 73:385-386.
Lindquist, A. W. 1961. Chemicals to sterilize insects. Journal of the Washington Academy of Sciences 51:109-114.

BOLL WEEVIL STERILITY 175
Lindquist, A. W., L. J. Gorzycki, M. S. Mayer, A. L. Scales, and T. B. Davich.
1964. Laboratory studies on sterilization of the boll weevil with apholate. Journal of Economic Entomology 57:746-750.
Lloyd, E. P., J. R. McCoy, and J. W. Haynes. 1976. Release of sterile boll weevils in the pilot boll weevil eradication experiment in 1972-73. In Boll Weevil Suppression, Management, and Elimination Technology, U.S. Department of Ag- riculture, ARS-S-71, p. 95-102.
G. H. McKibben, E. F. Knipling, J. A. Witz, A. W. Hart- stack, J. E. Leggett, and D. F. Lockwood.
1980. Mass trapping for detection, suppression, and integration with other suppression measures against the boll weevil. In Proceedings, International Colloquium on Management of Ins'ect Pests with Semio-Chemicals. Gainesville, FL. March 24-28.
McCoy, J. R., and J. E. Wright. 1979. Evaluation of bisazir and penfluron as sterilants for the boll weevil. Southwestern Entomologist 4:209-215.
McGovern, W. L., G. H. McKibben, R. C. Gueldner, and W. H. Cross. 1975. Irradiated boll weevils: Pheromone production determined by GLC analysis. Journal of Economic Entomology 68:521-523.
McHaffey, D. G., and A. B. Borkovec. 1976. Vacuum dipping: A new method of administering chemosteri-
lants to the boll weevil. Journal of Economic Entomology 69:139-143.
McKibben, G. H., W. L. McGovern, W. H. Cross, and 0. H. Lindig. 1976. Search for a super laboratory strain of boll weevils: A
rapid method for pheromone analysis of frass. Environmental Entomology 5:81-82.
Mitchell, E. B., M. E. Merkl, J. E. Wright, T. B. Davich, and R. F. Heiser. 1980. Sterility of boll weevils in the field following treatment
with diflubenzuron and gamma irradiation. Journal of Economic Entomology 73:824-826.
Moore, R. F., Jr., R. A. Leopold, and H. M. Taft. 1978. Boll weevils: Mechanism of transfer of diflubenzuron from male to female. Journal of Economic Entomology 71:587-590.
and H. M. Taft. 1975. Boll weevils: Chemosterilization of both sexes with busulfan plus Thompson-Hayward TH-6040. Journal of Economic Entomology 68:96.

176 J. E. WRIGHT AND E. VILLAVASO
National Academy of Sciences. 1975. An assessment of present and alternative technologies.
Cotton Pest Control, 139 p. Volume 3. National Academy of Sciences, Washington, DC.
Nilakhe, S. S., and N. W. Earle. 1976. Sperm production in normal vs sterile boll weevils. Journal of Economic Entomology 69:609-613.
Perkins, J. H. 1980. Boll weevil (Anthonomus grandis) eradication. Science
270:1044-1050.
Roberson, J., and J. E. Wright. 1982. Production of boll weevils. In Advances and Challenges
of Insect Rearing. E. G. King and N. Leppla, editors. U.S. Department of Agriculture, Technical Bulletin. [in press.]
Sikorowski, P. P., J. M. Wyatt, and 0. H. Lindig. 1977. Methods for surface sterilization of boll weevil eggs and
the determination of microbial contamination of adults. South- western Entomology 2:323-326.
Sterling, W. L., S. G. Wellso, P. L. Adkisson, and W. H. Dorough. 1965. A cotton seed meal diet for laboratory cultures of the
boll weevil. Journal of Economic Entomology 58:867-869.
Terry, P. H., S. C. Chang, and A. B. Borkovec. 1974. Chemosterilization by fumigation: Effects of temperature
and time of exposure. Journal of Economic Entomology 67:17-18.
D. G. McHaffey, and A. B. Borkovec. 1977. Uptake and residues of chemosterilants in boll weevils
treated by fumigation or dipping. Journal of Economic Ento- mology 70:427-430.
Vanderzant, E. S., and T. B. Davich. 1958. Laboratory rearing of the boll weevil: A satisfactory
larval diet and oviposition studies. Journal of Economic Entomology 51:288-291.
Villavaso, E. J. 1981. Field competitiveness of sterile male boll weevils re-
leased in the Boll Weevil Eradication Trial, 1979. Journal of Economic Entomology 74:373-375.
and N. W. Earle. 1976. Competitiveness of busulfan-fed sterile vs. native male
boll weevils. Environmental Entomology 5:279-280.

BOLL WEEVIL STERILITY 177
Villavaso, E. J., N. W. Earle, and D. D. Hollier. 1977. Boll weevils: Field and laboratory assessment of mating
ability and sperm content after irradiation with or without diflubenzuron treatments. Journal of Economic Entomology 70:562-564.
E. P. Lloyd, P. S. Lue, and J. E. Wright. 1980. Boll weevils: Competitiveness of sterile males in iso-
lated field plots. Journal of Economic Entomology 73:213-217.
S. S. Nilakhe, and W. L. McGovern. 1979. Field competitiveness of sterile male boll weevils. Journal of the Georgia Entomological Society 14:113-120.
Whitten, M. J., and G. G. Foster. 1975. Genetical methods of pest control. Annual Review of Entomology 20:461-476.
Wilkinson, J. D., R. K. Morrison, and P. K. Peters. 1972. Effect of calco oil red dye incorporated into a semiarti-
ficial diet of the imported cabbageworm, corn earworm, and cabbage looper. Journal of Economic Entomology 65:264-268.
Wright, J. E., J. W. Haynes, J. R. McCoy, and J. R. Dawson. 1979. Boll weevil: Mating ability, sterility, and survival of
irradiated and fumigated adults of different ages. South- western Entomologist 4:53-58.
J. R. McCoy, J. R. Dawson, J. Roberson, and P. P. Sikorowski.
1980a. Boll weevil sterility: Effects of different combinations of antibiotics, fumigation, and irradiation. Southwestern Entomologist 5:84-89.
R. Moore, J. McCoy, G. Wiygul, and J. Haynes. 1980b. Comparison of three sterilization procedures on the
quality of the male boll weevil. Journal of Economic Ento- mology 73:493-496.
and J. Roberson. 1981. Laboratory evaluation of the boll weevil sterility method. Journal of Economic Entomology 74:696-697.
and G. E. Spates. 1976. Reproductive inhibition activity of the insect growth
regulator TH6040 against the stable fly and the house fly: Effects on hatchability. Journal of Economic Entomology 69:365-368.
and B. R. Thomas. 1981. Determination of the components of the boll weevil
pheromone with a high pressure liquid Chromatographie method. Journal of Liquid Chromatography 4:1409-1416.


179
Chapter 8
PHEROMONES FOR SURVEY, DETECTION, AND CONTROL
E. P. Lloyd and G. H. McKibben Boll Weevil Eradication Research Agricultural Research Service U.S. Department of Agriculture Raleigh, NC 27607
J. E. Leggett Cotton Production Research Laboratory Agricultural Research Service U.S. Department of Agriculture Florence, SC 29503
A. W. Hartstack Pest Control Equipment and Methods
Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77843
ABSTRACT Since 1968, pheromones of the boll weevil (Anthonomus grandis Boheman), bollworm (Heliothis zea (Boddie)), and tobacco budworm (Heliothis virescens (F.)) have been isolated, identified, synthesized, formulated, and re- leased in the field with effective dispensing systems. The pheromone of the boll weevil is most effective when the four components (grand- lure) are prepared in polyethylene glycol and the mixture is impregnated at the desired dosage into polyester-wrapped cigarette fil- ters. Traps have evolved from sticky wing traps painted green to inverted 1-liter plastic soda cups painted fluorescent saturn yellow and fitted with a capture device on the top of the cups.
In field tests with grandlure-baited traps specially designed for use inside cotton fields, nearly all of the responding weevils were un- mated females. Subsequent tests in which phero- mone traps were combined with closely spaced applications of insecticide showed that the combination effectively prevented reproduction by emerged overwintered boll weevils. Results of field experiments indicated a 94% probability

180 E. P. LLOYD ET AL
of detecting the F progeny of a single over- wintered female weevil and a 100% probability of detecting the F« progeny when infield traps were spaced 64 m apart.
INTRODUCTION
The need to control the boll weevil (Anthonomus grandis Bohe- man) and other cotton insects, particularly the bollworm (Heliothis zea (Boddie)) and the tobacco budworm (il. virescens (F.)), has re- sulted in the development of highly effective insecticides that are applied during the fruiting period of the cotton plant to protect the developing crop from damaging pests. Cooke and Parvin (1982) estimated that the cost of the insecticides (does not include ap- plication costs) used on cotton was $235 million in 1978. Parencia et al. (1982) stated that estimated losses in yield due to the boll weevil from 1903 to 1950 averaged 10%. The National Cotton Council put the estimated costs of insecticide and application directed against boll weevils at $260 million annually for 1970-72 and noted that losses in yield in the southeastern United States were set at 11-23%.
The use of pheromones to suppress populations of pest insects or as part of an integrated management or eradication system may allow us to reduce our reliance upon insecticides while reducing costs and insect losses since pheromones evoke natural responses in insects. However, the pheromone must be used in such a way that it will elicit the desired response. Also, usage must be synchronized with development of the natural population and must be appropriate to the actual size of the population. In this chapter, we report on the efforts made to use pheromones to control Heliothis spp. and boll weevils in cotton.
PHEROMONES OF HELIOTHIS ZEA AND H. VIRESCENS
In 1962-63 in tests conducted at Oxford, NC (Gentry et al. 1964), to determine the sex attractant of the tobacco budworm, vir- gin females at least 4 days old were observed to attract males. Berger et al. (1965) subsequently found that extracts of adult fe- male _H. zea and _H. virescens elicited responses by males when con- ditions were carefully controlled. Mitchell et al. (1974), in later small-scale field tests with traps, found that extracts of calling Jl. virescens were attractive to males.
Roelofs et al. (1974) and Tumlinson et al. (1975) reported the isolation and identification of two components of the H. virescens pheromone, (_Z)-11-hexadecenal (Z^-11-HDAL) and (Z^)-9-teTradecenal (_Z-9-TDAL), better known as virelure (Hendricks et al. 1977). The ratio of the two components in extracts of live females was 16 parts ^-11-HDAL to 1 part ^-9-TDAL. Klun et al. (1979, 1980a, 1980b) concluded that H. zea females produce a mixture of (Z)-ll-

PHEROMONES FOR SURVEY, DETECTION, CONTROL 181
HDAL, (Z^)-9-hexadecenal, (_Z)-7-hexadecenal, and hexadecenal. Con- currentTy, Klun et al. (1980a, 1980b) concluded that the sex phero- mone of H. virescens contained these same four components plus Z-11-HDAL, tetradecenal, and Z^-9-TDAL. These recent identifi- "cations of the sex pheromones of H. zea and H. virescens may be useful in suppression programs. Additional research will be re- quired to determine their usefulness and the approach required for effective population suppression.
Dispensing System for Heliothis spp. Pheromone
Hendricks and Tumlinson (1974), working with the two compo- nents of H. virescens pheromone identified earlier, concluded that an ether e^xtract from the crude extract from females or 500 mg of the 16:1 ratio of the two synthesized components (virelure) in hex- ane were attractive to males for only 3-4 hours. They also con- cluded that higher doses (50 mg) of virelure dispensed from filter paper were not attractive for more than 6 hours during a single night and were completely unattractive by the second night.
Subsequently, Hendricks et al. (1977) found that mixing the two components of virelure with adhesive and laminating the mixture between two layers of vinyl polymer plastic sheets (formulated by Herculite Protective Fabrics Corporation, New York, NY 10010) pro- longed attractiveness to the point that 1.3- x 2.5-cm segments (dispensers) containing 20 mg of virelure were attractive at least 7 days and had minimum loss of attractiveness for 21 days. Also, Sparks et al. (1979a, 1979b) reported that in field tests the four-component H. zea pheromone and the seven-component H. vires- cens pheromone were highly attractive to males when they were dis- pensed from cigarette filters; however, these baits were attractive for only a few hours. They also reported that the formulation of the seven-component H. virescens pheromone was approximately three times as attractive as virelure when evaporated from cigarette filters.
On the other hand, Hartstac^ et al. (1980) found that lami- nated plastic dispensers (3.3 cm ) containing 20 mg of the seven- component pheromone were no more attractive than similar dispensers (6.7 cm ) containing virelure and that both were ca. 60-70% as effective as five virgin females. The laminated plastic baits were effective attractants for at least 14 days in the field.^ They also found that rubber septa containing 20 mg of virelure (Flint et al. 1979) were as attractive as the laminated plastic dispensers; how- ever, rubber septa containing the seven components were signifi- cantly less attractive. (The reason for the difference in attrac- tiveness of the seven-component H. virescens pheromone dispensed from cigarette filters and from laminated plastic dispensers is not known, but it probably involves the rate of dissipation of the various chemicals from the laminated plastic dispenser.) In addi- tion, the four-component Ö. zea pheromone (10 mg formulated in laminated plastic; 6.7 cm ) and rubber septa (20 mg of pheromone)

"•02 E. P. LLOYD ET AL
dispensers were both significantly less attractive than virgin fe- males (ca. 35 and 20%, respectively). The attractiveness of both these dispensers was increased when the amount of pheromone was reduced to 2.5 mg per dispenser. The laminated plastic baits were effective for at least 14 days.
Lopez et al. (1981) found that when they reduced the dosage of pheromone to 2.5 mg per bait (6.5 cm ) and used a thinner plastic, the four-component E. zea pheromone in a laminated plastic dispenser was very effective for at least 11-14 days.
Development of Heliothis spp. Pheromone Traps
For many years, major efforts have been made to develop and use traps to monitor or control populations of Heliothis spp. As the pheromones became available, the effort has taken another turn, partly because of the inconvenience of providing the electric power required by grid traps (Hollingsworth et al. 1963). One of the re- sulting pheromone traps, the pie plate or sticky trap (Snow and Copeland 1969), consists of two opposed 27.5-cm plastic plates joined centrally to maintain a 2.5-cm, 360** opening in a 4-cm-di- ameter x 6-cm-high cylinder shaped from 8- x 8-mesh hardware cloth. The upper surface of the lower plate is coated with Stikem. This trap is inexpensive and portable but relatively inefficient. A more efficient trap, the wind vane trap (Sparks et al. 1979a), was developed to catch male moths alive. It is made of plywood, sheet metal, and hardware cloth, and it turns so the entrance is always downwind, which means there is an opening for moths flying upwind toward the bait. Once through the opening, moths fly up and are trapped in a cage at the top of the trap. This trap, though effi- cient, is expensive to build and requires wind at all times for proper orientation.
The cone (or TP) trap with a pheromone source as bait that was designed and developed by Hartstack et al. (1979) has proved to be very efficient in capturing adult male Heliothis spp. Both the 50- 25 and 75-50 versions are similarly constructed of galvanized wire (hardware cloth, 8x8 mesh) and consist of two inverted wire cones, one inside the other. The outer cone has a base diameter and height of 50 or 75 cm, and the inner cone has the same diameter but a much flatter slope and no top. Thus, moths enter through the openings (50 and 25 cm for the 75-50 and 50-25 traps, respective- ly). The large trap will capture ca. 1.62 times as many moths as the 50-25 trap.
The performance of Heliothis spp. pheromone traps changes with time, primarily because of changes in the density of the popula- tion, which alters the competitiveness of the traps vis-a-vis na- tive females. In the spring and early summer, recovery of released marked moths was as high as 9.15%; at that time, nightly catches of ca. 10 moths per trap indicate field popujations estimated at one

PHEROMONES FOR SURVEY. DETECTION, CONTROL 183
moth per ha. In August, recovery was only 1.83% (Hartstack and Witz 1981).
Use of Pheromone for Control of Heliothis spp.
Methods of using pheromones to control populations of Helio- this spp. consist primarily of disruption and mass trapping. Some tests of disruption in small-scale field plots have shown reduction! in pheromone trap catches (Mitchell et al. 1975), and these may or may not reflect reductions in mating and thus some degree of con- trol. No control has yet been demonstrated in large-scale field tests. Possible causes for this failure are migration of moths in- to fields from outside and short life of the pheromone formula- tions. Within the next few years, researchers should develop much better formulations, and control by disruption may be possible.
No mass trapping experiments with Heliothis spp. pheromone traps have been conducted to date. However, now that efficient traps are available, this concept should be tested where popula- tions are low. For exampl.e, mass trapping in large areas in the spring when overwintering moths are emerging could reduce spring populations significantly. The resulting suppression could delay the onset of serious Heliothis spp. pressure and might mean that the crop would escape damaging infestations in August.
PHEROMONE OF THE BOLL WEEVIL
Seasonal Response of the Boll Weevil to Its Pheromone
Croso and Mitchell (1966) reported that male boll weevils feeding on cotton plants attracted female boll weevils at distances of 9.1 m or more. Hardee et al. (1969) observed that in the spring, traps baited with live males were more attractive to both sexes of overwintered boll weevils than weevil-free cotton plants. Mitchell and Hardee (1974) then reported that when pheromone-baited traps were placed in the field during the fruiting period of the cotton plant, nearly all of the field-generation weevils captured during July and early August were unmated females. However, Mit- chell et al. (1976) subsequently found that before squaring, more males than females were captured in traps; during midseason (July and August), many more female weevils were captured (280 to 24 males); and during September, as the crop matured, substantial num- bers of males were captured. Therefore, during emergence of the weevil population from overwintering in the spring before squaring, the boll weevil pheromone functions as an aggregating pheromone. Then, during the fruiting period of the cotton plant, it functions almost entirely as a sex pheromone. Finally, in the fall when wee- vil populations are often large, feeding and oviposition sites are limited in some fields, and weevils move from one field to another, the boll weevil pheromone again serves as an aggregating pheromone.

184 E. P. LLOYD ET AL
Isolation, Identification, and Synthesis of the Boll Weevil Pheromone
Keller et al. (1964) reported that a substance that was at- tractive to female boll weevils in the laboratory could be collect- ed by drawing air over males, passing the air through activated charcoal, and extracting the charcoal with chloroform. Hedin (1976) found that extractions of males with dichloromethane pro- duced a substance that was consistently attractive to females. The substance was even more attractive when it was steam distilled and solvent extracted. Tumlinson et al. (1969) found that when the frass of males (and later that of mixed sexes) was steam distilled, it too was highly attractive to males. Tumlinson et al. (1971) isolated, identified, and synthesized four terpenoid compounds ((+)-cis-2-isopropenyl-l-methlcyclobutaneethanol; (Z^)-3,3-dimethyl-
A-"^' cyclohexaneethanol; (Z^^-3,3-dimethyl-A cyclohexaneacetalde-
hyde; and (£)-3,3-dimethyl-A '^ cyclohexaneacetaldehyde) as the components of the pheromone of live male boll weevils. The synthe- sis and structural assignments of the four compounds are also re- ported by Tumlinson et al. (1971).
Later when McKibben et al. (1977) steam distilled frass of female boll weevils, they obtained an extract that was more attrac- tive to males than to females in the laboratory; and when these ex- tracts were purified by thin-layer chromatography, they were only attractive to males. The active components appeared to be alcohols and hydrocarbons. However, females were estimated to produce only 0.01 as much pheromone as the males, which explains the failure of earlier workers to find this pheromone.
Dispensing Systems for the Boll Weevil Pheromone
As noted, the pheromone produced by male boll weevils is a mixture of the two terpenoid alcohols and two aldehydes that were isolated, identified, and synthesized by Tumlinson et al. (1969). However, when the mixture of the synthesized components, grandlure, was dispensed in field tests by applying solutions to chromatog- raphy supports such as firebrick, these release systems were at- tractive for only a few hours. McKibben et al. (1971) therefore screened a number of polymers as controlled-release additives and found polyethylene glycol the most effective in controlling the re- lease of grandlure. Where grandlure was added to polyethylene glycol heated to ca. 40°C and then poured into empty gelatin cap- sules, the response of test weevils increased as the concentration of grandlure was increased. Hardee et al. (1972) then found that a formulation of grandlure containing glycerol, water, and methanol dispensed first from cotton dental rolls and then from cigarette filters was 80% as attractive for just under 7 days as live, caged males fed on cotton squares.
In 1972, a commercial gel formulation of grandlure was pre- pared by Zoecon Corporation (Hardee et al. 1974) and was used in

PHEROMONES FOR SURVEY, DETECTION, CONTROL 185
the Pilot Boll Weevil Eradication Experiment (PBWEE) then underway in southern Mississippi. Also, McKibben (1976) developed a cotton- seed oil base gel formulation of grandlure that was tested in the PBWEE. These dispensers were improvements over the cigarette fil- ter dispenser because they released grandlure for almost 2 weeks. However, Bull et al. (1973) were able to slow the rate of release of grandlure from the cigarette filter dispenser by placing it in an open glass vial (filter-in-vial system) that functioned as a physical barrier.
Bull (1976) later described a physical barrier formulation devised by A. Higgins, F. Boyd, and D. Whittam of the Animal and Plant Health Inspection Service, U.S. Department of Agriculture. A special paper cartridge was used instead of the glass vial to re- duce costs and facilitate aerial distribution. Also, McKibben (1972, 1974) devised and improved a device that made it possible for one operator to inject grandlure into 3000 cigarette filters in 1 hour. A laminated plastic dispensing system for grandlure devel- oped by Health-Chem Corporation was described by Hardee et al. (1975). Four thicknesses of these three-layer vinyl polymer lami- nates (ca. 0.5, 1.0, 1.5, and 2 mm thick) were placed in a clearing within a wooded area where no cotton was grown within 8 km, and laboratory-reared female boll weevils were released. The thicker dispensers appeared to retain grandlure longest in the field, but all were effective dispensers.
Because the polyethylene glycol filter-in-vial system was in- expensive and easily formulated, more than one million of these dispensers were used from 1973 to 1977. Dosage of grandlure per dispenser was 3 mg when effectiveness was needed for 1 week, 8 mg for 2 weeks, and 24 mg for 4 weeks. In Leggett traps, these grand- lure dispensers were placed on top of the inverted floral liner (various formulations were placed at several locations on the „ Leggett traps); in infield traps, they were placed in the 5-cm plastic capture box on the top. However, the capture box slowed the rate of release of the grandlure because it constituted an ad- ditional barrier and the rate of release from an unprotected filter was too rapid. McKibben et al. (1980) therefore conducted analyt- ical studies of several impervious coverings for the cigarette fil- ter to optimize the rate of release of grandlure. As a result, a commercially available polyester-wrapped cigarette filter became available in early 1978. In field tests conducted at five loca- tions, McKibben et al. (1980) found that a filter of this type placed inside the capture box was the most attractive of all grand- lure dispenser systems tested. It was used as the standard dispen- ser for the Boll Weevil Eradication and Optimum Pest Management Trials in 1978 and 1979.
Development of Boll Weevil Pheromone Traps
The first field tests with traps baited with male boll weevils were conducted by Cross et al. (1969) in Mexico and Florida and

186 E. P. LLOYD ET AL
near Mississippi State, MS. Of the 12 types of traps tested, a solid wing trap and an oblique funnel trap appeared most efficient when live males fed on fresh cotton squares were used as bait. However, the wing traps were coated with Stikem; weevils entering the oblique funnel traps were simply captured in a box that was not coated with an adhesive. Therefore, a Stikem-coated plywood wing trap painted dark green and baited with males fed on cotton squares or artificial diet was used in most field tests. Then in 1968, Cross et al. (1971) reported that white or bright-yellow traps were more attractive to boll weevils than the darker green traps. As a result, metal wing traps, 30 cm wide x 25.4 cm high, painted canary yellow but similar in construction to the plywood wing trap, were used in most 1969 field tests. Later, in 1968, Cross et al. (1976), in detailed studies with traps painted various colors, found that wing traps painted a fluorescent yellow (solar yellow) were the most attractive. Also, in 1970, Cross et al. (1971) de- vised a Stikem-coated wing trap made from plastic-impregnated, 2.1-liter paper cartons that were painted with a daylight fluores- cent saturn yellow paint. These traps were 19 cm wide x 25.5 cm high and were constructed by trimming two unformed cartons and sta- pling them together so that they formed a wing trap when unfolded. Subsequently, Leggett and Cross (1971) developed the so-called Leg- gett trap, a nonsticky trap constructed from a floral liner 29.21 cm high. It was painted daylight fluorescent yellow and capped with a screen cone (held just off the inverted liner with glass beads or other spacers) that had a small hole in the apex, which opened into a 5-cm plastic box. The completed trap was mounted on a stake ca. 1.22 m above the ground. A small cube of Vapona No- Pest strip placed in the box killed captured weevils. When Leggett et al. (1975) compared the Leggett trap with seven other designs, the Leggett trap captured significantly more weevils than the other traps. It also caught as many weevils when the screen cone and capture box were placed on top of the inverted floral liner (Leg- gett trap) as when the floral liner was coated with Stikem (Whittam trap).
Mitchell and Hardee (1974) designed an infield (Mitchell) trap that was similar to the Leggett trap but had a smaller base for use inside cotton fields. This trap was constructed of a 1-liter soda cup (inverted), painted saturn yellow, and topped with a screen cone that was fastened to the cup with brass fasteners. A 5-mm hole in the apex of the cone opened into a 5-cm plastic box. Ten or twelve triangular-shaped openings cut in the top of the cup al- lowed weevils to pass from the cup upward into the capture device. Dickerson et al. (1981) modified the Mitchell trap to reduce cost and facilitate mass production. The modified Mitchell trap ap- peared to be ca. 1.4 times more efficient than the original Mit- chell trap in capturing weevils.

PHEROMONES FOR SURVEY, DETECTION, CONTROL 187
Male-Baited Traps Around Cotton Fields for Suppression of Overwintered Boll Weevils
Because of the discovery that the male boll weevil emits a pheromone, many researchers made an effort to use male boll weevils to capture a sizable proportion of the overwintered population as it emerged in the spring. For example, Hardee et al. (1971), in an area with a very low population of native boll weevils, compared the following six treatments: (1) one wing trap per 0.4 ha, around the field; (2) one wing trap per 0.4 ha, in the field; (3) one-half wing trap per 0.4 ha, in the field and one-half wing trap per 0.4 ha, around the field; (4) one wing trap per 0.4 ha, around the field plus untreated trap plots of cotton; (5) one wing trap per 0.4 ha, around the cotton field, plus aldicarb-treated trap plots of cotton; and (6) one wing trap per 0.4 ha, around the field in three tiers. The wings of these traps were constructed of sheet metal, painted canary yellow, coated with Stikem, and mounted on reject hoe han- dles. The traps were baited with laboratory-reared boll weevils (weevils in transit from Mississippi State, MS, to the Crosbyton, TX, area were fed on artificial diet) that were provided with ar- tificial diet, seedling plants, squares, or bolls as available. During the spring trapping period, from May until August 22, the traps captured an average 0.64 weevils per trap from field popula- tions estimated at 0.08 to 0.21 weevils per 0.4 ha. Although all treatments therefore suppressed the boll weevil population by 62.6-100%, the small size of the populations made it difficult to ascertain the most effective treatment.
Boyd et al. (1973), in a large-scale trapping experiment con- ducted on 30,000 ha of cotton in the Rolling Plains area of west Texas in 1969, evaluated the effectiveness of Stikem-coated wing traps painted canary yellow, baited with male weevils, and placed around all of the cotton fields. Approximately 26,000 traps cap- tured 10,159 weevils between April 23 and July 23, and at least one weevil was captured in 14.7% of the traps. The fact that about 85% of the traps captured no weevils indicated that the location of in- dividual traps was very important. In addition, 85% of captured weevils were trapped between May 28 and June 25. As in the test reported by Hardee et al. (1971), the degree of suppression of the overwintered boll weevil population achieved was difficult to as- sess since the population was generally low and the area was very large, but infestations did develop in 19.2% of the fields by August 27.
Lloyd et al. (1972a), in an areawide trapping experiment, in- stalled approximately one trap per 0.4 ha around 1600 ha of cotton in Monroe County, MS, in the spring of 1969 following a voluntary

188 E. P. LLOYD ET AL
grower-sponsored reproduction-diapause control program conducted during the fall of 1968. Again, Stikem-coated wing traps painted canary yellow and baited with males were placed around the cotton fields in April. Traps were baited beginning on April 29 and were supplied with fresh males twice a week through August 22 except for 3 weeks when they were baited only once a week.
Because the reproduction-diapause control program was volun- tary and implemented by individual growers, effectiveness varied. The result was sizable boll weevil populations in some parts of the test area. These larger populations reduced the effectiveness of the trapping program, but they also provided an opportunity to as- sess the efficiency of the traps against populations of varying sizes. The results are shown in Table 1. Clearly, as the popula- tion increased, trap efficiency decreased.
As a part of the same experiment, trap densities of one, two, four, and eight traps per 0.4 ha were compared. Although the num- ber of weevils captured per trap did not differ, the total numbers of weevils trapped per 0.4 ha by one, two, four, and eight traps per 0.4 ha differed greatly, 7.16, 9.37, 17.73, and 34.60, respec- tively, and more weevils were observed in fields with more ty^ps. Thus more traps apparently attracted more weevils into the vicinity of the fields because more weevils were captured on the traps and more weevils were observed in the fields with the greatest numbers of traps. The most efficient arrangement in this experiment was two traps per 0.4 ha.
Roach and Ray (1972), in field experiments conducted in 1970 at the Pee Dee Experiment Station near Florence, SC, and Socastee, SC, used 10 traps per 0.2-ha cotton field (20 traps per 0.4 ha) to capture emerging overwintered boll weevils. These trap densities appeared to suppress the native overwintered population and pre- vented damage until the F^ field generation emerged in early
August.
Results of these large-scale field experiments with male- baited sticky traps placed around cotton fields therefore indicated that efficiency was inversely related to density of overwintered boll weevils. Traps suppressed low populations, but large popula- tions were not affected.
Pheromone-Baited Traps in Crops for Suppression of Overwintered Boll Weevils
Mally (1901) observed that in a given area, those cotton fields planted first were the first to be infested by the boll wee- vil and suggested that a few rows of an early-maturing variety should be planted so emerging weevils would gather there and be trapped. Hunter (1912) did not believe that weevils emerged early enough from overwintering quarters so that a trap plot would con- centrate appreciable numbers. Much later, Isley (1950) reported

PHEROMONES FOR SURVEY, DETECTION, CONTROL 189
c Cd
0) B u
i-l ON r-t v£) O OS
X^ »-I
^ « <U CO
4J «
C! >%
t % (U O > u o
Oí M-l O O u
c >^ o 4-» IS •H
CO •»
c ^ d) tkO
c 0)
eu Cd
u
Ö0
Ü
(U
w I I
(U r-I
Cd
H
0) 00 G Cd
Cd
d) 4-»
tí a r-< CO
Cd ^
4J «H O > +J (Ü
0)
t>0 Cd u-i Í-I o 0)
-N CO 0) i-H tí Où «H «H
tí > Cd <D T) M (D o;
tí ►^
tí Cd
,£3 g cu
tí tí •H
Cd O)
<u o >
O
(U rH
W) •H
tí > <U cd (U <Ü tí »-4 (U i-I 3 —^ 1^ Cd
4-»
^ u 73 O tí (U (U QJ •H
^ u JI! <u TJ ^ 4-1 <J- 0) tí tí 4J
•H O O
t>0 2 u Cd Oí (1) 1—1 }-i > a O o; o Ü > <:
O
u CO (U ^ ^ M-l .—1 s O <u 0 •H
^ M-l
C^
C3^
00
I I—I
CX)
I o C30
<!■ QN CN
I
tí -O
:^ o
C30 CX)
<t I
es <3N
1—I
en
CO
tí (U
T3
T3 O)
00 00
tí
JC Ö0
O <í r^
I CJN
00 00
o m
I o
CM
O
O <!■
00 I
CN m
I
CN
o 00
CN I
O
CJN
a I o (U
4-» o to 13 'Ö r-I
M-i V4 r-H (U O Q) OJ •H
XI -H M-l g g M-l 3 3 QJ CO tí <U ^ ^ 4-1 QJ W) 4-1
;tí tí tí 4-1 «iH tí •H
X) -H ÖO«H T3 tí > *J (U
•H «H tí > X> T? CU ^ •H CO CU > >^ 0) CO
•H ^ J-l JD T) a O
'O >^ cu 0) CO
^ > ^ tí •H O
T3 H O •H Oí (U iJ 4-i 4J T3 Cd ZJ T3 ■U O. CO Oí CO g a 4-1 <ü O Cd cd M-l o J-i g tí
4J -H •H
TJ iJ r-t M-l CO r-I
QJ O <D •H •H > M-l >^ M <D
O <U (U Q) tí 413 ;5
;C OJ g 4J -H í3 r-4
O tí rH tí 'H O
• H M-l CO ^ M-l 3
CO <L) r-< r-I
tí a Cd O -O tí
•H CL) CO 4-) 4-» > CU O Cd V-l Cd Cd
,-1 (U J-i 3 CO 4J M-l
O-^ O O O tí Cu «H ^s
o T3 • TJ m o; CO 0) V4 4J }^ CO (U tí 3 cu 4-) 3 4-» g tí O Ci. tí
•H o cd CO
'^ O CO }-i M-^ Cd 0) 0 V-( > <u >^ o ^1 X3 o
(U g tí M-l ^ tí CJ o g tí •H
U <u tí >. •H N ^ M-l
• H (U M-l CO JS CO QJ
4-» O- <u Cd tí. ^ >% V4 Cd 4J ,Û 4J u
4-1 M-l CO tí O Q) -H T3
4J QJ 0) Cd 73 4-» 4J g 0) Cd Cd 'H J-i r-I g 4J tí tí
•H CO 4J O 4J Qj a r-I CO Cd Cd W tí O U
o -^ .H CO "»^ .-H 4J r-I CN
'cd -H r-I >

"•90 E. P. LLOYD ET AL
that in Arkansas, trap plots 3-30 rows wide did concentrate over- wintered boll weevils where they could be destroyed more easily with insecticides; his suggestion resulted in "spot dusting" by small cotton producers on upland or hill soils.
After Cross and Mitchell (1966) discovered the response of fe- male boll weevils to the male pheromone, interest was renewed in attracting and concentrating overwintered boll weevils into trap crops where they could be killed without treating the entire field. The resulting reduction in insecticide usage became important when Ridgway et al. (1968) found that the systemic insecticide, aldi- carb, though it was highly effective in suppressing populations of overwintered boll weevils, did produce outbreaks of bollworms and tobacco budworms.
Bradley (1967) found that in Louisiana, overwintered boll wee- vils could be expected to congregate in early planted border strips provided that the difference in planting dates was sufficient to give a height advantage to the trap crop. Lloyd et al. (1972b), in a large-scale field experiment conducted in 1970 in Grenada County, MS, compared weevil populations in untreated fields and in fields treated with (1) traps only (sticky wing traps baited with males placed around cotton fields at the rate of one trap per 0.4 hec- tare); (2) traps plus aldicarb-treated strips of cotton (male- baited traps installed at the same rate but concentrated on two sides of the field adjacent to strips treated with aldicarb); (3) bait males (sterile males restricted by gluing wing covers to- gether) placed 7.6 m apart on rows adjacent to two sides of fields; and (4) restricted sterile males placed 7.6 m apart on the second of four untreated rows adjacent to aldicarb-treated strips on two sides of the fields.
The boll weevil population in the test area was low as a re- sult of a diapause control program in the fall of 1969 and a severe winter. The effectiveness of the treatments was therefore ap- praised by noting the dates on which fields exposed to each treat- ment needed insecticidal control for boll weevils.
Average number of days after July 21 before
Treatment application of insecticide J_/
Males only 16.42 a Untreated 19.28 a Traps only 23.28 ab Traps + strips 27.14 b Males + strips 30.00 b
\J Numbers followed by the same letter are not signifi- cantly different at the 5% level according to Duncan's multiple range test.

PHEROMONES FOR SURVEY, DETECTION, CONTROL 191
Scott et al. (1974) also evaluated the use of trap crops in fields (1) planted 2 weeks early to an early fruiting variety, Qua- paw, treated with aldicarb, and then baited with grandlure; (2) planted as in (1) but on the normal planting date and treated with aldicarb; and (3) planted on normal planting date, without grand- lure. Trap crops, with and without grandlure, proved to be much more attractive than the normal plantings until the normal planting began squaring, an indication that the early response of the boll weevil was to early fruiting as well as to the grandlure. However, after squaring began in the normal planting, trap crops with grand- lure were more attractive than those without grandlure.
Rummel et al. (1976), in 3 years of field tests on the Rol- ling Plains of Texas, failed to obtain consistent suppression of boll weevils with trap crops; however, they found that in Glascock County, TX, early planted, aldicarb-treated trap crops effectively suppressed populations of overwintered weevils and prevented the development of damaging infestations. Gilliland et al. (1976), in southern Mississippi, also found trap crops to be an effective means of concentrating and suppressing emerging boll weevil popula- tions if the trap crops were planted earlier than the main crops and if pheromone bait stations were used.
The use of trap crops has not had wide acceptance in the Cot- ton Belt because the northern parts have cool weather early in the season and growers in the warmer parts have such heavy demands on their time during the period when trap crops should be planted. However, in many sections, early planting of a trap crop of early fruiting varieties and subsequent baiting of traps with grandlure will attract a very high percentage of emerged overwintered weevils until squaring begins in normal plantings. Trap crops therefore represent a means of suppressing early-season boll weevils by treating or trapping only a small percentage of the crop. They should be considered one of several tools for improved management of cotton insects.
Infield Traps and Insecticides for Boll Weevil Suppression
Knipling (1979), on the basis of simulation models, advanced the following hypothesis: the total suppressive action of two methods of control that differ in efficiency when insect popula- tions are high or low may be substantially greater than the sum of the action of each method used alone. This would occur if at least one of the methods potentiates the action of the other method and if the efficiency of the first method is enhanced when the pest population is low. Also, some suppressive techniques may be mu- tually potentiating, resulting in an even greater increase in ef- ficiency. An example of the potentiating effect cited by Knipling would be the concurrent use of infield traps and the application of insecticides for boll weevil suppression. The insecticide

192 E. P. LLOYD ET AL
applications should achieve essentially the same degree of kill regardless of the density of the boll weevil population; however, the efficiency of a given number of pheromone traps should increase as the boll weevil population is reduced.
We therefore have examined the total effect of calculating the degree of suppression that might be expected from insecticide ap- plications alone, use of infield traps alone, and concurrent use of the two methods against a low level overwintered boll weevil popu- lation. The results are presented in Table 2.
The application of insecticides makes two important contribu- tions to total suppression. The assumed kill of 90% of the boll weevils reduces the number that can reproduce. It also reduces tenfold the number of males that will be competing with the traps for the attraction of the surviving females that are seeking mates. Therefore, the ratio of traps to competing males will increase from 10:1 when traps only are used to 100:1 when insecticides are also applied. Efficiency of trap capture should thus increase from 91 to 99%. If these methods of suppression were used independently and acted independently and if each achieved 90% control, regard- less of pest density, the suppression achieved by using the two concurrently would be only 99%. Theoretically, this degree of sup- pression would allow one successful mating and some potential prog- eny. In fact, the increasing effect of the traps as the population increases would seem to mean that no matings would be expected at the density assumed. The practical implications of such a high de- gree of suppression during a single generation are therefore re- flected by the relative number of progeny of survivors that can be expected when the techniques are used alone and when they are used concurrently. We thus have an example of the possibilities when the boll weevil pheromone is appropriately integrated with other suppressive measures.
The theoretical findings were subsequently tested in the field. Mitchell et al. (1976) combined twice weekly applications of azinphosmethyl (0.11 kg per 0.4 ha) from appearance of pinhead squares (June 13) until July 6 with 10 infield traps per 0.4 ha. Results are shown in Table 3.
Thus 10 infield traps per 0.4 ha plus twice weekly treatment with insecticide prevented reproduction during the treatment period (Phase II). In fact, infiltration of some mated females from other cotton growing areas cannot be precluded though the experiment was conducted in an area some 40 km from other cotton. (W. A. Dicker- son, personal communication, captured a marked boll weevil 104.6 km from the release point.) Therefore, complete elimination of repro- duction could not be achieved. The results obtained during Phase II, however, clearly indicate that concurrent application of insec- ticides and the operation of infield traps could be used against any isolated persisting infestations or new infestations should a beltwide eradication program be undertaken.
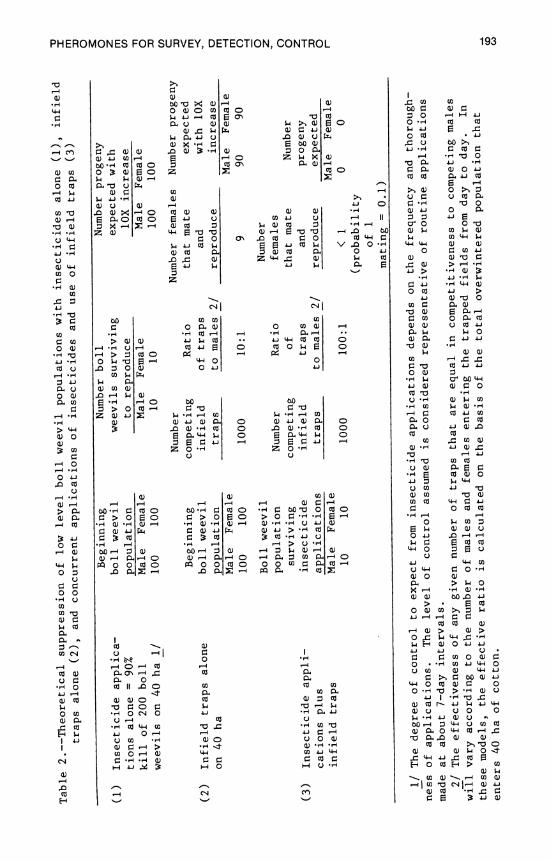
PHEROMONES FOR SURVEY, DETECTION, CONTROL 193
Q) CO
O Cd .-I u
Cd *j
CO 73 d) r-i
TJ <U •H 'H O "^^
•H C
O
CO O c
•H Q) CO
•H -O
Cd CO C CO O <D
4-) -H Cd o
r-l «H =J ^ Pu O O Oí Pu CO
tí
Q) O
O Cd o
rH «H O) r-l > P. Q) Pu
,-i Cd
> ti o tí ,-1 (U
O 3 O
tí tí o o
•H o
CO TJ (U tí í-i Cd Cu p. •*
CO C^J
Cd Q) o tí
•H O 4J r-H <U Cd U
es 0
Cd H
tí o;
o cu o
tí ^ cu 4-)
O ^
P* 'O
e PHO tí X -H
23 QJ
M 4-» <-H a o
Oí ^ V-1 Cu 4J
(D r-l
Cd <u (U Ë 4-> Ü <u Cd tí
tí O
.-• o Cd C3>
(U cd ^ rtí
cd
^1 00 tí
•H > cu
•H o > tí »-I X) tí o
u
-a ^ ^ >
O)
}-l
CO Q)
—I Cd o
CO CO p. Q) cd I—I j-i Cd
Cd j-j e tó M-l o o 4-1
.-• o Cd <-H
ÖO tí T3
•H I—I
0) «H Pu M-l e tí C->
ÖO'H tí tí > o
tío)*-» tí 15 Cd
•H r-H 60 r-l tí <U rH p4 « o o
^ Cu
I—I Cd o B o
fe
ÖO'H tí tí > o
•H O) «H tí <Ü 4-» tí ^ Cd
.-I o Cd o
OÛ.-I tí cu rH p4
pq o o ^ cu
o o o
rH cd o B o
fe
0 rH o cd o
Id ^ o ^
•H Èsç
.-H O .-H Cd p-a^ i-H X p. O Cd II ^ o
O) cu O -o tí O tí •H o CM Ü o .-1
•H Cd H-i CO 4J O rH O CO •H
CU tí rH > CO o rH <U tí -H •H CU M *-» ^ ^
CU Cd
Cd
o <1-
tí tí M O
CU x: CO CO t-l ÖO tí cu tí
'tí cd tí o rH M 4->
i^ P^ cu Ë O o -H Cd Cd
CU tí 4J cu U 4-) B ^ rO cu o fe o cd • 4J
bo cu Xí o t>0 >. j3 o P. 4-» -H tí Cd tí
IS í-i X pu cu
cu I—1
Cd S
O
rH -O a tí CU Cd Cd p
eti
to d
atio
/~s B r-^ rH >% cu O >. tí
>^ . o tí O Cd Cu 4-) O a .H 'tí o
CU cu •H cu 4-) o d. CO 4-J o rH II tí tí j-J e
u CU Cd tí •H —1 cr O o 'tí cu t—1 B T3 'tí rH ^ Ù0 CU i-i CO V-i cu
Xi Cd tí O Cd ^ tí u CO MH VJ
e 4-1 Cd j-i V ^ o •H MH MH cu cu J3 (U cd cu o 4J O tí CO 4->
^ MH jS cu u Cd <U cu 'tí tí 4-> M cu a ^ 0) > ,-1 .H
•H tí U O Cd ti
t i
fie
verw
-.»^ 4-1 0) -tí o ^1 CO tí
'O CU Pu CU e pu.-t
CO tí CO o cu Cd o CO (U rH CU cu Ü cd 4J
•H a^ .. cu ^4 u o 4-) MH Cd Cd O (U Pu tí 4-i 4-í
Cd o u B o T3 CU •H
tó 4-1 O 4J
f-^
CO tí -0
cu cu --• JS ^
Cd 4J 4-1 O CU tí
•H U cr öO'w 4-> CU <u tí o Cd 73 •H
Ö0 o «H CU >-l CO tí Tí •H CO u cu -H
V^ »H .-H CO .-1 tí Cd 4J CO
(U 4-> (U Pu o PH O tí Cd
JD CU •H Cd o CU o 4-» CU XI Ë Q,m »-I o Cd Cd tí e tí 4-1 i-H CO ;C CO cu 2; o •H <U .H 4-> CU ji:
o TJ rH U •H T^ CO Cd U CU 0- e tí
• H g Cd CU O •U tí U MH CJ CO 4J T3
CO cu CU CO 'O CU 1-4 CU tí r-* CO cd MH tí -U •H tí ÖCO o Cd o tí O Cd Cd > o tí •H «H B r-l •H .-H rH
CU •H «H O 4-> cu o V4 CO tí
cu 4-» > •H Cd fe ß u CU CU O :^ Cd «H
1-1 >
•U o u «H
0 u u tí
X »-I ^ e Cd Cd
r-l tí »-I (U rH (U <4H O tí e o r-i CU tí CO Cl rH O O tí O O CO tí CL Cd I—1 ■U MH CO
« a •H Cd 2 O »+-I tí o -H CU O cu CU > »-4 o X rH •H cu -H CU CU 00^ 4-» > B Cd O cu CO >^ tí ^1 4-» rH rH C tí
Cd Cd cu ,-1 <u > <u >
Zë ÍH MH ^ «H QJ O 4-> 4J
1 i-i 4-) O • •H tí tí CO O cu tí i-H O • •H CO 4-» MH o Cu O CO CU MH 4-»
eu tí >, tí bO CU u Cd CO MH O Cd CU tí o
CO p. O -H Xl > -rH (U o cu tí Cd 4-> 1 -H T3 ^
^O .-4 í-l CU Cd r-. 4-> Vu 4-> MH
•H CU 4J 0) o o o 0 (J V^ -H 4_) cu o •>
•H CO TJ ÖOrH 3 MH o CO Cd 4J C r-l CU Cu o MH Cd T-l X O O CU X) P.^ QJ <U Q) «H «H Cd Cd >^TJ o CO 4-) UH cu (U J-i o <^ tí Cd tí X '+^ 4-» 42 Cd e M U «H H o Cd H >
cu cu ^,.-i CO
CO u cu ^^ .-1 CO ■73 CN ^-1 cu 4J
m cu 2 ::í -^ tí N-^ tí B :^ 4-1 cu

194 E. P. LLOYD ET AL
Table 3.—Capture of boll weevils in infield traps (10 per 0.4 ha) on 43.2 ha of cotton near Havana, AR, 1974
Number of boll weevils captured
Date Males Females Total
Phase I 1 [no insecticide treatments) 171 103 274 336 205 541 364 277 641 114 156 270 158 217 375
1143 958 2101
*hase II (twice weekly insecticide treatments) 45 58 103 2 9 11
1/5 0
1/7 3
1/12 3
1 53
0 77
1 131
Phase III (no 1 insecticide treatments) 0 50 50 1 1
52 102
53 103
Phase IV (no i insecticide treatments) 1 83 84
10 37 47 2 13 15
10 45 55 87 353 440 92 364 456
132 239 371
May 21 May 27 June 3 June 10 June 13
Total
June 17 June 21 June 26 July 2 July 9
Total
July 12-19 July 23-31
Total
August 2-8 August 13 y August 19-23 August 26 September 4 September 10 September 17
Total 334 1134 1468
\J Sex of one weevil not determined. 2_/ An application of monocrotophos + azinphosmethyl (0.45 + 0.11
kg per 0.4 ha) applied for control of bollworms and boll weevils on August 14.
Infield Traps for Detection and Suppression
Detection of boll weevils when populations are low is essen- tial to any evaluation of containment and eradication programs. However, visual examination of a large number of plants for live insects or damage is laborious and costly and is also a relatively insensitive procedure when populations are low, particularly when progeny of reproducing females are closely clumped. Infield traps

PHEROMONES FOR SURVEY, DETECTION, CONTROL 195
baited with grandlure appeared to be a more sensitive method of de- tection. A field test was therefore conducted by Leggett et al. (1981) to determine the sensitivity of different densities of traps for the detection of the F, progeny that would be expected from a single overwintered female weevil reproducing naturally in a cotton field. A clump of 20 infested squares was placed in special weevil-free fields. In fields with one trap per 0.4 ha, the trap catches allowed detection of four of seven of the reproducing clumps in the F, generation; in fields with four traps per 0.4 ha, the trap catches allowed detection of seven of seven of the clumps.
In the initial special field tests of Leggett et al. (1981), one and four traps per 0.4 ha were spaced 64 m and 32 m apart, re- spectively, but the number of traps per hectare equaled the number of hectares times the desired trap density. In order to more fully understand the influence of trap densities and spacing on the prob- ability of detecting and controlling small isolated reproducing clumps of boll weevils, Lloyd et al. (1980) conducted a series of field experiments in 1979 in a noncotton area (Surry County, NC) that was more than 129 km from the nearest commercial cotton plant- ings. By using this site and creating known infestations, they eliminated competition from unknown competing males. From July 9 for 9 days, overwintered female weevils were allowed to deposit eggs in cotton squares at the Agricultural Research Service, Cotton Production Research Laboratory, U.S. Department of Agriculture, Florence, SC. The squares were then held on moist sand in an out- door insectary to simulate natural conditions as closely as pos- sible. Ca. 10-12 days later, the squares were transported to Surry County by automobile and placed near the center of 24 cotton plots (average 1.28 ha) laid out so they were 3.2 or more km apart. Sites for the squares were selected on the basis of the usual pattern for such clumps of progeny. Treatments were as follows: 20 infested squares per clump with traps 64, 45, 36, and 32 m apart; and 10 in- fested squares per clump with traps 45 and 32 m apart. The result- ing captures of emerged F, generation weevils are summarized in Table 4.
Knipling and McGuire (1966) and Knipling (1976, 1979) advanced the theory that the degree of control of insects obtained with pheromone-baited traps in a given area is governed by the relative attractiveness of the pheromone (competitive attraction) produced by calling insects present in the area. They set up theoretical models to estimate the potential value of traps for detection and for control. These simulation models and assumed parameters have proved useful in experimental design and interpretation of data ob- tained in these investigations.
The more important variables that govern efficiency of traps used for detection and control of boll weevils include: (1) the attractiveness of the pheromone emanating from the traps v. the at- tractiveness of the pheromone disseminated by the males; (2) the number of traps and the distribution of traps v. the number and

196 E. P. LLOYD ET AL
Table 4.—Summary of capture of F and F^ generation boll weevils with infield traps in test fields, Surry County, NC, 1979
Average number captured Distance (m) F F
^ "^2 ^^1. between tri ips
.4 ha)
20
emerging
(traps per 0. Males Females Total weevils from squares
infested squares per clump 64 (1.9) 2.0 17.5 77.0
45 (3.6) 0.2 3.0 15.2 78.0
36 (4.2) 1.0 5.5 9.5 73.0
32 (5.7) 10
0.7 6.7 5.7 infested squares per clump
63.0
45 (3.0) 0.2 2.7 5.0 77.0
32 (6.6) 0.5 3.0 2.0 76.0
distribution of the pheromone-producing males and the mate-seeking females; (3) the area within which a trap and competing males can elicit a response from mate-seeking females; and (4) the actual ef- ficiency of the traps in capturing responding females (and males in the case of overwintered populations). Then if all traps in a field are as attractive as males and are equally near to mate- seeking females, we can calculate the proportion of the available females that will be captured by assuming that all females respond- ing to the traps will be captured. When these calculations are related to actual capture, we can obtain clues to the type of ac- tion and the effectiveness of different densities of infield traps in relation to the distribution pattern of sexually active boll weevils. For example, the distribution of a given number of boll weevils entering a cotton field from hibernation will probably dif- fer greatly from the distribution of the same number of F^ progeny emerging in a restricted area from one or more egg-laying females. Also, if traps are uniformly distributed in a cotton field, over- wintered males and females will probably not aggregate because of the confusion (interference) produced by the synthetic pheromone emitted by the traps. On the other hand, F males emerging in a restricted area (clumps) would be expected to be more attractive than the synthetic pheromone to the emerging females in these clumps because the traps will usually be farther away than the emerging aggregated males.
To determine the theoretical capture rates for traps on the basis of the assumption that when traps and male boll weevils are uniformly distributed, traps are fully competitive in all respects with males for the attraction of mate-seeking females, we made the calculations presented in Table 5. Such estimates can then be com- pared with actual capture rates at given trap spacings and densi- ties to arrive at the relative effectiveness of traps and males in attracting females. Critical comparison of the data should then help in identifying and evaluating factors governing the action and

PHEROMONES FOR SURVEY, DETECTION, CONTROL 197
Table 5.—Daily probability of capturing emerging F female boll weevils on 0.4 ha if males and traps are equally attractive l_l
Probability of capturing F, femal es with indicated distance (m) between tr aps 2/
Day of 64 45 36 32
emergence 1/ (1.95) (3.59) (4.23) (5.67)
1 0.66 0.78 0.81 0.85
2 0.49 0.64 0.68 0.74
3 0.39 0.55 0.59 0.65
4 0.33 0.47 0.51 0.58
5 0.28 0.42 0.46 0.53
6 0.25 0.38 0.42 0.48
7 0.22 0.34 0.38 0.44
8 0.19 0.31 0.35 0.41
Total 2.81 3.89 4.20 4.68
1/ Assumed one female and one male emerging each day. ~2_l Number traps per 0.4 ha in parentheses.
effectiveness of pheromone' traps for the detection and control of small populations.
A brief description of the method of calculations based on the equal competitiveness assumption may be helpful. The data for days 1 and 2 and the density of 1.95 traps per 0.4 ha will be cited as examples. On day 1, the numerical ratio of traps to one competing male is 1.95:1 for the attraction of one mate-seeking female. The probability of female capture would be 0.66, and the probability of the female mating, therefore, would be 0.34. On day 2, the traps would be in competition with two males for the attraction of the one female seeking a mate on that day. The ratio of traps to males would be 1.95:2, and the probability of capturing the female would be 0.49.
Thus by the time of the emergence of the approximately eight males and eight females in each field, the traps will be more com- petitive than the males in capturing females. Theoretically, the 4.23 traps per 0.4 ha, spaced 36 m apart, should capture 4.2 fe- males per replicate. In fact, actual capture was 5.5 females per replicate. Also, the 5.67 traps per 0.4 ha, spaced 32 m apart, should capture 4.68 females per replicate; they actually captured 6.75. Plainly, the higher density of traps and the proximity of the traps to the clumps of F, progeny resulted in a concentration of pheromone that competed effectively with the pheromone produced by the males. Most females were eventually drawn to the traps rather than to the competing males.
These results are of great practical significance since they suggest that such trapping may be highly effective in suppressing

198 E. P. LLOYD ET AL.
and managing low populations of boll weevils. Not only is the ra- tio of traps to competing males a major factor in efficiency but so also is the greater volume of pheromone due to the shorter distance from the traps to females. For example, distribution appears to be so important that the system will likely be even more effective against scattered individual overwintered boll weevils as they enter cotton fields than it is against females emerging in the clumps. If that hypothesis is correct, this system of suppression deserves high research priority, specifically in regard to reduced cost of traps and pheromone, prolongation of effectiveness of pher- omone, and increased efficiency of actual capture.
In the test by Lloyd et al. (1980), infield traps spaced 64 m apart detected four of four clumps. However, the traps were so ar- ranged that the entire field was covered with a reasonably uniform array. (The first trap was installed approximately 4.6 m from one field corner, and the rest of the traps were 64 m apart.) The data indicate that 94% of the time, the F progeny of a single-mated overwintered female will be detected with such a spatial arrange- ment. As Table 4 shows, all arrangements detected F^ generation weevils. These results therefore support the conclusions reached by Leggett et al. (1981) that infield traps provide a highly sensi- tive method of detecting the existence of boll weevil infestations resulting from a single reproducing female in a cotton field of any size.
In addition, significant suppression of the F generation was achieved when traps were spaced 36 and 32 m apart. This reduction is reflected by the rate of capture of F« females at all trap den- sities (Table 4). If reproduction had been normal, the average F^ populations should have been similar in the four replicates of each trap spacing and density, and capture of F^ females should have been low at the lowest trap density and high at the highest trap density.
We estimate that 80% or more of the F. females were captured when traps were spaced 36 m apart and probably more than 90% of the F. females were captured when traps were 32 m apart. These re- sults indicate the potential value of grandlure for both detection and control. They also suggest that high density trapping can be used to manage populations of boll weevils that originate from low overwintered populations.

PHEROMONES FOR SURVEY, DETECTION, CONTROL 199
LITERATURE CITED
Berger, R. S., J. M. McGough, and D. F. Martin. 1965. Sex attractants of Heliothis zea and H_. virescens. Jour-
nal of Econonic Entomology 58:1023-1024.
Boyd, F. J., J. R. Brazzel, W. F. Helms, R. J. Moritz, and R. R. Edwards.
1973. Spring destruction of overwintered boll weevils in west Texas with wing traps. Journal of Economic Entomology 66:507- 510.
Bradley, J. R., Jr. 1967. Oriented movement of the boll weevil in response to trap
crop plantings, foliage color and sex pheromone. Ph.D. dis- sertation, Louisiana State University, Baton Rouge.
Bull, D. L. 1976. Formulations of grandlure. In Detection and Management
of the Boll Weevil with Pheromone. Texas Agricultural Experi- ment Station, Research Monograph No. 8, p. 5-8.
J. R. Coppedge, R. L. Ridgway, D. D. Hardee, and T. M. Graves.
1973. Formulations for controlling the release of synthetic pheromone (grandlure) of the boll weevil. 1. Analytical stud- ies. Environmental Entomology 2:829-835.
Cooke, F. T., Jr., and D. W. Parvin, Jr. 1982. The cost of insecticides used on cotton in the United
States. U.S. Department of Agriculture, Agriculture Handbook. [In press.]
Cross, W. H., D. D. Hardee, F. Nichols, H. C. Mitchell, E. B. Mitchell, P. M. Huddleston, and J. H. Tumlinson.
1969. Attraction of female boll weevils to traps baited with males or extracts of males. Journal of Economic Entomology 62:154-161.
J. E. Leggett, and D. D. Hardee. 1971. Improved traps for capturing boll weevils. U.S. Depart- ment of Agriculture, Cooperative Economic Insect Report 21:367- 368.
and H. C. Mitchell. 1966. Mating behavior of the female boll weevil. Journal of Economic Entomology 59:1503-1507.
H. C. Mitchell, and D. D. Hardee. 1976. Boll weevils: response to light sources and colors on
traps. Environmental Entomology 5:565-571.

200 E. P. LLOYD ET AL.
Dickerson, W. A., G. H. McKibben, E. P. Lloyd, J. F. Kearney, J. J. Lam, Jr., and W. H. Cross.
1981. Field evaluation of a modified infield boll weevil trap. Journal of Economic Entomology 74:280-282.
Flint, H. M., L. M. McDonough, S. S. Salter, and S. Walters. 1979. Rubber septa: a long lasting substrate for (Z^)-ll- hexadecenal and (Z^)-9-tetradecenal, the primary components of the sex pheromone of the tobacco budworm. Journal of Economic Entomology 72:798-800.
Gentry, C. R., F. R. Lawson, and J. D. Hoffman. 1964. A sex attractant in the tobacco budworm. Journal of Eco-
nomic Entomology 57:819-821.
Gilliland, F. R., Jr., W. R. Lambert, J. R. Weeks, and R. L. Davis. 1976. Trap crops for boll weevil control. In. Proceedings, Con-
ference on Boll Weevil Suppression, Management, and Elimination Technology, Memphis, TN, February 13-15, 1974. U.S. Department of Agriculture, ARS-S-71.
Hardee, D. D., W. H. Cross, and E. B. Mitchell. 1969. Male boll weevils are more attractive than cotton plants
to boll weevils. Journal of Economic Entomology 62:165-169.
T. M. Graves, G. H. McKibben, W. L. Johnson, R. C. Gueldner, and C. M. Olsen.
1974. A slow-release formulation of grandlure, the synthetic ph-eromone of the boll weevil. Journal of Economic Entomology 67:44-46.
0. H. Lindig, and T. B. Davich. 1971. Suppression of populations of boll weevils over a large
area in west Texas with pheromone traps in 1969. Journal of Economic Entomology 64:928-933.
a. H. McKibben, R. C. Gueldner, E. B. Mitchell, J. H. Tumlinson, and W. H. Cross.
1972. Boll weevils in nature respond to grandlure, a synthetic pheromone. Journal of Economic Entomology 65:97-100.
G. H. McKibben, and P. M. Huddleston. 1975. Grandlure for boll weevils: controlled release with a
laminated plastic dispenser. Journal of Economic Entomology 68:477-479.
Hartstack, A. W., Jr., J. D. Lopez, J. A. Klun, J. A. Witz, T. N. Shaver, and J. R. PI immer.
1980. New trap design and pheromone bait formulations for Helio- this. In Proceedings, Beltwide Cotton Production Research Con- ference, p. 132-136.

PHEROMONES FOR SURVEY, DETECTION, CONTROL 201
Hartstack, A. W., and J. A. Witz. 1981. Estimating field populations of tobacco budworm moths from
pheromone trap catches. Environmental Entomology 10:908-914.
J. A. Witz, and D. R. Buck. 1979. Moth traps for the tobacco budworm. Journal of Economic Entomology 72:519-522.
Hedin, P. A. 1976, Grandlure development. Ln Proceedings, Conference on Boll Weevil Suppression, Management, and Elimination Technology, Memphis, TN, February 13-15, 1974. U.S. Department of Agricul- ture, ARS-S-71.
Hendricks, D. E., A. W. Hartstack, and T. N. Shaver. 1977. Effect of formulations and dispensers on attractiveness of virelure to the tobacco budworm. Journal of Chemical Ecology 3:497-506.
and J. H. Tumlinson. 1974. A field cage bioassay system for testing candidate sex
pheromones of the tobacco budworm. Annals of the Entomological Society of America 64:547-552.
Hollingsworth, J. P., A. W. Hartstack, and J. M. Stanley. 1963. Electric insect traps for survey purposes. U.S. Depart- ment of Agriculture, ARS 42-3-1, 24 p.
Hunter, W. D. 1912. The boll weevil problem. U.S. Department of Agriculture,
Farmers' Bulletin No. 512, 43 p.
Isley, D. 1950. Trapping weevils in spots with early cotton. Arkansas Agricultural Experiment Station, Bulletin No. 496, p. 45-46.
Keller, J. C, E. B. Mitchell, G. McKibben, and T. B. Davich. 1964. A sex attractant for female boll weevils from males. Journal of Economic Entomology 57:609-610.
Klun, J. A., B. A. Bierl-Leonhardt, J. R. Plimmer, A. N. Sparks, M. Primiani, 0. L. Chapman, G. Lepone, and G. H. Lee.
1980a. Sex pheromone chemistry of the female tobacco budworm moth, Heliothis virescens. Journal of Chemical Ecology 6:177- 183.
J. R. Plimmer, B. A. Bierl-Leonhardt, A. N. Sparks, and 0. L. Chapman.
1979. Trace chemicals: the essence of sexual communication systems in Heliothis species. Science 204:1328-1330.

202 E. P. LLOYD ET AL.
Klun, J. A., J. R. Plimmer, B. A. Bierl-Leonhardt, A. N. Sparks, M. Primiani, 0. L. Chapman, G. H. Lee, and G. Lepone.
1980b. Sex pheromone chemistry of female corn earworm moth, Heliothis zea. Journal of Chemical Ecology 6:165-175.
Knipling, E. F. 1976. Biomathematical basis for suppression and elimination of
boll weevil populations. Iri Proceedings, Conference on Boll Weevil Suppression, Management, and Elimination Technology, Memphis, TN, February 13-15, 1974. U.S. Department of Agricul- ture, ARS-S-71.
1979. The basic principles of insect population suppression and management. U.S. Department of Agriculture, Agriculture Hand- book, No. 512 p. 571-576.
and J. U. McGuire, Jr. 1966. Population models to test theoretical effects of sex at-
tractants used for insect control. U.S. Department of Agri- culture, Agriculture Information Bulletin No. 308, 20 p.
Leggett, J. E., and W. H. Cross. 1971. A new trap for capturing boll weevils. U.S. Department
of Agriculture, Cooperative Economic Insect Report 21:773-774.
W. H. Cross, H. C. Mitchell, W. L. Johnson, and W. L. McGovern.
1975. Improved traps for capturing boll weevils. Journal of the Georgia Entomological Society 14:1444-1447.
E. P. Lloyd, and J. A. Witz. 1981. Efficiency of infield traps in detecting and suppressing
low population levels of boll weevils. Environmental Entomology 10:125-130.
Lloyd, E. P., G. H. McKibben, E. F. Knipling, J. A. Witz, A. W. Hartstack, J. E. Leggett, and D. F. Lockwood.
1980. Mass trapping for detection, suppression, and integration with other suppression measures against the boll weevil. jDn Proceedings, International Colloquium on the Management of In- sect Pests With Semio-chemicals, March 24-28, 1980, Gaines- ville, FL.
M. E. Merkl, F. C. Tingle, W. P. Scott, D. D. Hardee, and T. B. Davich.
1972a. Evaluation of male-baited traps for control of boll weevils following a reproduction-diapause program in Monroe County, Mississippi. Journal of Economic Entomology 65:552- 555.

PHEROMONES FOR SURVEY, DETECTION, CONTROL 203
Lloyd, E. P., W. P. Scott, K. K. Shaunak, F. C. Tingle, and T. B. Davich.
1972b. A modified trapping system for suppressing low-density populations of overwintered boll weevils. Journal of Economic Entomology 65:1144-1147.
Lopez, J. D., Jr., T. N. Shaver, and A. W. Hartstack, Jr. 1981. Evaluation of dispensers for the pheromone of Heliothis
zea. Southwestern Entomology 6(2): 117-122.
McKibben, G. H. 1972. A device for injecting grandlure into cigarette filters. Journal of Economic Entomology 65:1509-1510.
1974. An improved device for dispensing pheromone solutions. Journal of Economic Entomology 67:558.
1976. Composition for attracting the cotton boll weevil. U.S. Patent 3,803,303.
D. D. Hardee, T. B. Davich, R. C. Gueldner, and P. A. Hedin.
1971. Slow-release formulations of grandlure, the synthetic pheromone of the boll weevil. Journal of Economic Entomology 64:317-319.
P. A. Hedin, W. L. McGovern, N. M. Wilson, and E. B. Mitchell.
1977. A sex pheromone for male boll weevils from females. Journal of Chemical Ecology 3:331-335.
W. L. Johnson, R. Edwards, E. Kotter, J. F. Kearney, T. B. Davich, E. P. Lloyd, and M. C. Ganyard.
1980. A polyester-wrapped cigarette filter for dispensing grand- lure. Journal of Economic Entomology 73:250-251.
Mally, F. W. 1901. The Mexican cotton-boll weevil. U.S. Department of Agri-
culture, Farmers' Bulletin No. 130, p. 11-12.
Mitchell, E. B., and D. D. Hardee. 1974. Infield traps: a new concept in survey and suppression
of low populations of boll weevils. Journal of Economic Ento- mology 67:506-508.
E. P. Lloyd, D. D. Hardee, W. H. Cross, and T. B. Davich.
1976. Infield traps and insecticides for suppression and elimi- nation of populations of boll weevils. Journal of Economic Entomology 69:83-88.

204 E. P. LLOYD ET AL.
Mitchell, E. R., J. H. Tumlinson, W. W. Copeland, R. W. Hines, and M. M. Brennan.
1974. Tobacco budworm: production, collection, and use of natu- ral pheromone in field traps. Environmental Entomology 3:711- 713.
M. Jacobson, and A. H. Baumhover. 1975. Heliothis spp.: disruption of pheromonal communication with (Z^)-9-tetradecen--l-ol formate. Environmental Entom.ology 4:577-5"79.
Parencia, C. R., T. F. Pfrimmer, and A. R. Hopkins. 1982. Insecticides for control of cotton insects. U.S. Depart- ment of Agriculture, Agriculture Handbook. [in press.]
Ridgway, R. L., S. L. Jones, J. R. Coppedge, and D. A. Lindquist. 1968. Systemic activity of 2-methyl-2-(methylthio)propionalde- hyde 0^-(methylcarbamoyl)oxime (UC-21149) in the cotton plant with special reference to the boll weevil. Journal of Economic Entomology 61:1705-1712.
Roach, S. H., and L. Ray. 1972. Boll weevils captured at Socastee, South Carolina, in
1970, in wing traps placed around fields with and without growing cotton. Journal of Economic Entomology 65:599-560.
Roelofs, W. L., A. S. Hill, R. T. Carde, and T. C. Baker. 1974. Two sex pheromone components of the tobacco budworm moth, Heliothis virescens. Life Science 14:1555-1562.
Rummel, D. R., R. C. Mclntyre, and C. W. Neeb. 1976. Suppression of boll weevils with grandlure-baited trap
crops. In Detection and Management of the Boll Weevil With Pheromone. Texas Agricultural Experiment Station, Research Monograph No. 8, p. 53-61.
Scott, W. P., E. P. Lloyd, J. 0. Bryson, and T. B. Davich. 1974. Trap plots for suppression of low density overwintered
populations of boll weevils. Journal of Economic Entomology 67:281-283.
Snow, J. W., and W. W. Copeland. 1969. Fall armyworms: use of virgin female traps to detect males and to determine seasonal distribution. U.S. Department of Agriculture, Production Research Report No. 110.
Sparks, A. N., J. E. Carpenter, J. A. Klun, and B. G. Mullinix. 1979a. Field responses of male Heliothis zea (Boddie) to phero- monal stimuli and trap design. Journal of the Georgia Entomo- logical Society 14:319-325.

PHEROMONES FOR SURVEY, DETECTION, CONTROL 205
Sparks, A. N., J. R. Raulston, P. D. Lingren, J. E. Carpenter, J.
A. Klun, and B. G. Mullinix. 1979b. Field response of male Heliothis virescens (F.) to phero- monal stimuli and traps. Bulletin of the Entomological Society of America 25:268-274.
Tumlinson, J. H., D. D. Hardee, R. C. Gueldner, A. C. Thompson, P. A. Hedin, and J. P. Minyard.
1969. Sex pheromones produced by the male boll weevil: isola- tion, identification, and synthesis. Science 166:1010-1012.
R. C. Gueldner, D. D. Hardee, A. C. Thompson, P. A. Hedin, and J. P. Minyard.
1971. Identification and synthesis of the four compounds com- prising the boll weevil sex attractant. Journal of Organic Chemistry 36:2616-2621.
D. E. Hendricks, E. R. Mitchell, R. E. Doolittle, and M. M. Brennan.
1975. Isolation, identification, and synthesis of the sex pheromone of the tobacco budworm. Journal of Chemical Ecology 1:203-214.


207
Chapter 9
INSECT GROWTH REGULATORS WITH EMPHASIS ON THE USE OF BENZOYLPHENYL UREAS
D. L. Bull and J. R. Abies Cotton Insects Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77841
E. P. Lloyd Boll Weevil Eradication Research Agricultural Research Service U.S. Department of Agriculture Raleigh, NC 27607
ABSTRACT Insect growth regulators (iGR's) are highly selective chemical pesticides that interfere in different ways with the normal development of certain life stages of treated arthropods. Although many IGR's have been evaluated in laboratory and field experiments as alternatives to the conventional broad- spectrum insecticides currently used to control phytophagous pests of cotton, only a few sub- stituted benzoylphenyl ureas have shown suffi- cient promise for practical use. Of these, only diflubenzuron has been approved by the Environmental Protection Agency for use on cot- ton. This IGR is effective against the boll weevil (Anthonomus grandis Boheman) and appears to have little adverse effect on populations of beneficial species or other important pests associated with cotton.
The results of extensive field experimenta- tion suggest that diflubenzuron may be used to suppress low-to-moderate populations of the boll weevil. However, the chemical must be applied with an emulsifiable crop oil if it is to be ade- quately effective; applications must begin at the immediate onset of fruiting, that is, before over- wintered boll weevils have a chance to initiate oviposition; and applications must be repeated at 5- to 7-day intervals to reinforce coverage of boll weevils present in fields and to insure contact with insects that will continue to emerge from hi- bernation quarters well into the fruiting cycle.

208 D. L BULL ET AL
The most effective use of diflubenzuron or a related product against boll weevils will probably be as a component of an integrated pest management system wherein community-wide applications of the IGR will complement other suppression strategies such as the early and complete postharvest destruction of stalks, fall applications of pesticides to reduce the overwintering generation of boll weevils, the planting of early maturing varieties of cot- ton, and, if necessary, the judicious and stra- tegic use of conventional pesticides to reduce the numbers of overwintered weevils that emerge in early spring or the numbers that remain in fields after the applications of the IGR have been terminated.
INTRODUCTION
Many of the phytophagous pests found in cotton fields must be controlled if producers are to avoid unacceptable economic loss. However, the use of conventional insecticides against the so-called "primary" pests has tended to precipitate new problems. Primary pest species in cotton include the pink bollworm (Pectinophora gos- sypiella (Saunders)) and a lygus bug (Lygus hesperus Knight) in the western United States; the boll weevil (Anthonomus grandis Boheman) and the cotton fleahopper (Pseudatomoscelis seriatus (Reuter)) in the Southwest; and the boll weevil and the tarnished plant bug (Ly- gus lineolaris (Palisot de Beauvois)) in the Midsouth (Anonymous 1975TT These species are normally managed by using broad-spectrum insecticides. As a result, natural populations of entomophagous parasites and predators that normally hold populations of "second- ary" pests in check may be reduced to the extent that these pests too attain primary status. When this occurs, the producer may either suffer substantial crop loss or become locked into a costly, sometimes futile, season-long need to apply insecticides.
Presently, the most important of the secondary pests in cotton are the tobacco budworm (Heliothis virescens (F.)) and the bollworm (R, zee (Boddie)). In fact, these species now have the status of major pests across much of the Cotton Belt (Bottrell and Adkisson 1977). According to recent estimates (Anonymous 1980), they ac- count for as much as 35% of all losses attributable to insect dam- age to cotton. The problem is particularly severe in areas where populations of the tobacco budworm have developed resistance to organophosphorus insecticides and can be controlled only by using more costly materials.
Other potentially important secondary pests that may be re- leased from the control of their natural enemies by pesticides to the point that they inflict economic damage, though usually on a more limited scale, include the cabbage looper (Trichoplusia ni

INSECT GROWTH REGULATORS 209
(Hubner)), the saltmarsh caterpillar (Estigmene aerea (Drury)), the cotton leafperforator, (Bucculatrix thurberiella Busck), the beet armyworm (Spodoptera exigua (Hübner)), and different species of spider mites.
In view of the problem with Heliothis spp. and the potential for new problems of the same kind, efforts are underway to find se- lective materials and methods that could be alternatives to broad- spectrum insecticides in the management of cotton pests. Some of these methods are discussed elsewhere in this handbook. We are concerned here with the potential of chemicals that manifest their activity through disruption of normal insect growth.
BENZOYLPHENYL UREAS AS INSECT GROWTH REGULATORS
A broad array of synthetic chemicals are included among the insect growth regulators (lGR*s) that are considered the third generation of pesticides. The chemicals that make up this group, though diverse, have two characteristics in common. They manifest biological activity by interfering in different ways with the nor- mal development or functions of specific life stages of treated in- sects; and those that are approved or are being considered for use in pest control appear to be somewhat less detrimental to the en- vironment and to nontarget organisms than most conventional chemi- cal pesticides. Indeed, certain IGR's seem to offer the advantage of interspecific selectivity among some insect biocenoses to the extent that they favor natural enemies. The literature is replete with scientific reports and reviews of research on IGR's; for gen- eral reviews, the reader is referred to Menn and Beroza (1972) and Staal (1975).
Of the IGR's that have been evaluated for use in managing in- sect pests of cotton, only certain substituted benzoylphenyl ureas have shown sufficient promise for practical use. Currently, three of these materials (Figure 1) are being investigated intensively: ( 1 ) di f lubenzuron (N^-[ t (4-chlorophenyl)amino] carbonyl]-2 ,6-di- fluorobenzamide; Dimilin, TH 6040); (2) penfluron (2,6-difluro- N_-[[[4-(trifluoromethyl)phenyl]amino]carbonyl]benzamide; AI3- 63223); and (3) BAY SIR 8514 (2-chloro-N-[[[4-(trifluoromethoxy) - phenyl]amino]carbonyl]benzamide).
Diflubenzuron was introduced by Philips-Duphar B. V., Amster- dam, Holland, and developed in the United States by Thompson- Hayward Chemical Co., Kansas City, KS. Of the three benzoylphenyl ureas specified, only diflubenzuron is approved by the U.S. Envi- ronmental Protection Agency for commercial use in this country. It has conditional registration for use on cotton against the boll weevil and on forests for control of the gypsy moth (L3miantria dispar (L.) ).
Penfluron was originally synthesized by Philips-Duphar B. V. (Wellinga et al. 1973). Subsequently, Oliver et al. (1977)

210 D. L BULL ET AL
.0' '■^r"^
Ri
DIflubenzuron -CI 2,6-F2 Penfluron -CF3 2,6-F2 BAY SIR 8514 -OCF3 2-CI
Figure 1.—Structures of some benzoylphenyl urea compounds tested for use in cotton pest management.
synthesized penfluron and reported that it was considerably more active than diflubenzuron in inhibiting the hatch of boll weevil eggs. Because of this improved activity, penfluron may replace diflubenzuron as the chemical component of the combined treatment (irradiation and chemical) used to produce nonreproductive boll weevils for release in eradication experiments and programs. Pen- fluron is also being investigated for possible use as a foliar spray. In other studies (Chang 1979, Chang and Borkovec 1980), penfluron was more than twice as potent as diflubenzuron in inhib- iting the hatch of eggs of the house fly (Musca domestica L.).
BAY SIR 8514 was introduced by Bayer A. G., Leverkusen, Ger- many (Zoebelein et al. 1980) and is being developed in the United States by Mobay Chemical Corp., Kansas City, KS. This IGR has been reported to be more effective than diflubenzuron against the house fly (Chang 1979), the fall armyworm (Spodoptera frugiperda (J. E. Smith)) (Zoebelein et al. 1980), and a black fly (Simulium vittatum Zetterstedt) (Lacey and Mulla 1978). Preliminary results of field and laboratory tests at Florence, SC, by R. F. Moore and A. R. Hopkins (private communication) indicate that BAY SIR 8514 is as active as diflubenzuron against the boll weevil. These investiga- tors also observed that crop oil need not be added to BAY SIR 8514 to obtain effective insecticidal activity. (Diflubenzuron is es- sentially inactive without oil.)
DIFLUBENZURON
The preponderance of information concerning the use of the benzoylphenyl ureas in cotton pest management has been developed for diflubenzuron. Laboratory tests have shown that diflubenzuron is active against the pink bollworm when certain larval stadia are fed treated diet (Flint and Smith 1977), but it apparently is not effective against this pest in the field (Flint et al. 1978). Also, diflubenzuron has been generally ineffective in field tests

INSECT GROWTH REGULATORS 211
against Heliothis spp. (Taft and Hopkins 1975, House et al. 1978, Nemec 1978, Bull et al. 1979); the tarnished plant bug; the cotton fleahopper; the carmine spider mite (Tetranychus cinnabarinus (Boisduval)); and the cotton aphid (Aphis gossypii Glover) (Taft and Hopkins 1975, Nemec 1978). On the other hand, tests in field cages (Flint et al. 1977, 1978) indicate that diflubenzuron may provide good control of the cotton leafperforator, though informa- tion on practical control is apparently lacking. In addition, in both field and laboratory tests, diflubenzuron has been highly ef- fective against the boll weevil. Thus, after it had been estab- lished that diflubenzuron affected reproduction in the boll weevil (preliminary unpublished observations reviewed by Taft 1978), Moore and Taft (1975) conducted the first documented study and demon- strated conclusively that diflubenzuron inhibited the hatch of boll weevil eggs in the laboratory. On the basis of observations asso- ciated with subsequent intensive investigations of the effect of diflubenzuron on the boll weevil, we can conclude that the boll weevil is the only major cotton pest that is significantly affected by field applications of diflubenzuron.
Mode of Action of Diflubenzuron in the Boll Weevil
The biological activity of diflubenzuron against invertebrates is attributed to destructive physiological events that are trig- gered when the chemical disrupts the synthesis of chitin, an essen- tial structural component of insect cuticle (Mulder and Gijswijt 1973, Post et al. 1974, Verloop and Ferrell 1977, Hajjar and Casida 1979). Apparently, when adults of some species contact diflubenzu- ron or are fed the material, the eggs oviposited subsequently do not hatch. This result has been observed in the boll weevil (Moore and Taft 1975) and in several other species such as Spodoptera lit- toralis (Boisduval) (Ascher and Nemny 1974); the house fly (Gross- curt 1976); stable flies (Stomoxys calcitrans (L.)) and horn flies (Haemotobia irritans (L.)) (Wright and Harris 1976, Wright and Spates 1976); and several species of weevils that attack citrus (Schroeder et al. 1976, Lovestrand and Beavers 1980). The effect is caused by the ovicidal action of the IGR rather than by steriliza- tion of the treated adult (Grosscurt 1976, Verloop and Ferrell 1977, Ivie and Wright 1978). Typically, embryonic development in an egg from a treated female appears normal until the expected time for eclosión. Grosscurt (1976, 1978) concluded that affected em- bryos of the house fly and the Colorado potato beetle (Leptinotarsa decemlineata (Say)) developed into larvae that were unable to leave the egg. He attributed this failure to the effect of diflubenzuron in disrupting the formation of chitin in the larval cuticle and the resulting incapacitation of muscular function. Moore et al. (1978) made similar observations in their study of the boll weevil; they also found that larvae that were able to emerge from the eggs of treated females were usually moribund.
Ample evidence is available to prove that the primary action of diflubenzuron against the boll weevil is manifested in the adult

212 D. L BULL ET AL
females and that the chemical is effective when it is ingested or when it is administered as a contact treatment. There is also evi- dence that it is the secretion of unmetabolized diflubenzuron in the eggs of treated female boll weevils that apparently accounts for the ovicidal effects (Bull and Ivie 1980). The same is true for the house fly and the stable fly (ivie and Wright 1978, Chang and Borkovec 1980). Moreover, the females probably must be exposed to the chemical directly to obtain maximum results. Moore et al. (1978) concluded that treated males probably could not transfer sufficient diflubenzuron during copulation to cause a substantial inhibition of egg hatch. Though holding topically treated males continuously with females in very small cages did produce some in- hibition, such an indirect exposure of females to diflubenzuron seems unlikely to be of significant importance in the field.
14 In studies of the fate of C-labeled diflubenzuron after
topical application to adult virgin female boll weevils, Bull and Ivie (1980) demonstrated that the chemical was absorbed very slowly and that large amounts did not accumulate internally. For example, at 8 days posttreatment, ca. 64% of the applied diflubenzuron was unabsorbed, and internal radioactive material never exceeded 4% during the experimental period. Also, only ca. 3% of the applied diflubenzuron was metabolized by hydrolysis and conjugation reac- tions. Still and Leopold (1978) found no evidence that difluben- zuron was metabolized in boll weevils treated by immersion or injection or by feeding on treated cotton squares. On the other hand, Chang and Stokes (1979) reported that boll weevils metabo- lized and excreted as much as 19% of an injected dose of difluben- zuron in the form of water-soluble conjugates.
14 Bull and Ivie (1980) studied the secretion and fate of C-
labeled diflubenzuron in eggs after a single topical application (1 |ig per insect) to adult female boll weevils (Figure 2). At 4 days,posttreatment, eggs of treated females contained ca. 2.24 ppm of C-labeled diflubenzuron equivalents, but the amount declined progressively to ca. 0.4 ppm at 26 days posttreatment. This dimi- nution of secreted radiocarbon occurred despite the fact that di- flubenzuron was quite persistent on the treated females. Periodic analyses of the eggs revealed that all the radiocarbon in the eggs was unchanged diflubenzuron.
Daily observations of subsamples of the eggs from the treated females showed that hatch was inhibited in ca. 80% of the eggs through 10 days posttreatment. Thereafter, recovery was fairly rapid; and by 22 days posttreatment, 80-90% of the eggs hatched (Figure 2). At the 50% hatch level, which occurred at ca. 16 days posttreatment, the diflubenzuron content of eggs was ca. 0.6 ppm. Chang and Borkovec (1980) found a similar relationship between IGR secretion ( C-labeled diflubenzuron and penfluron) in eggs and hatch of house fly eggs. The study by Bull and Ivie (1980) thus supports and helps explain previous observations in the laboratory by McLaughlin (1976) and Moore et al. (1978), who found that the
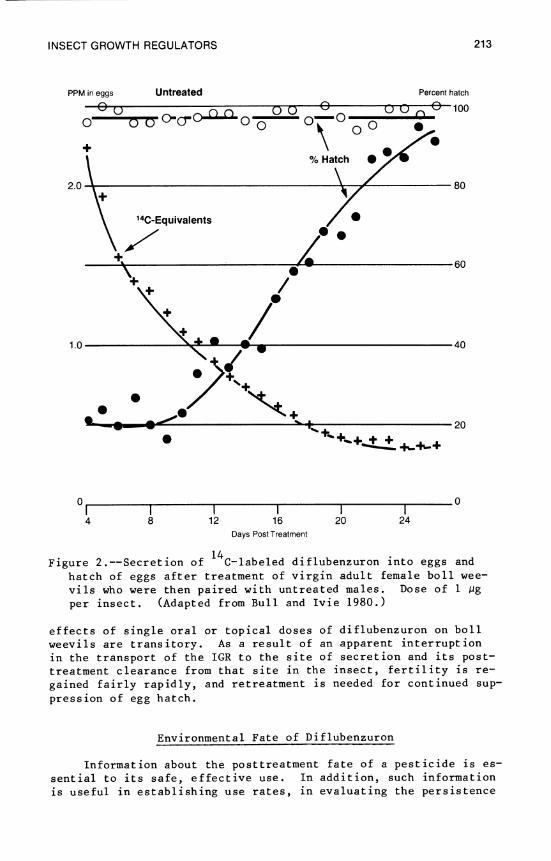
INSECT GROWTH REGULATORS 213
PPM in eggs Untreated Percent hatch
O o ^O 100
I 1 \ 1 1 \ 4 8 12 16 20 24
Days Post Treatment
14 Figure 2.—Secretion of C-labeled diflubenzuron into eggs and
hatch of eggs after treatment of virgin adult female boll wee- vils who were then paired with untreated males. Dose of 1 jLig per insect. (Adapted from Bull and Ivie 1980.)
effects of single oral or topical doses of diflubenzuron on boll weevils are transitory. As a result of an apparent interruption in the transport of the IGR to the site of secretion and its post- treatment clearance from that site in the insect, fertility is re- gained fairly rapidly, and retreatment is needed for continued sup- pression of egg hatch.
Environmental Fate of Diflubenzuron
Information about the posttreatment fate of a pesticide is es- sential to its safe, effective use. In addition, such information is useful in establishing use rates, in evaluating the persistence

214 D. L BULL ET AL
of posttreatment biological activity, and in assessing the poten- tial for detrimental effects of residues on nontarget organisms.
Available evidence (Bull and Ivie 1978, Mansager et al. 1979) indicates that whatever the method of foliar application of diflu- benzuron, little of the material (ca. 5% of the dose) is absorbed by the cotton plant and that residues of the IGR are quite persis- tent on foliar surfaces. In a ffgld study. Bull and Ivie (1978) found that ca. 90% of a dose of C-labeled diflubenzuron re- mained on treated leaf surfaces at 14 days posttreatment. In both the aforementioned studies, analyses revealed that unchanged diflu- benzuron accounted for > 98% of the unabsorbed and absorbed mate- rial that was recovered. All evidence indicates that the chemical is highly resistant to decomposition, either by photodegradation on foliar surfaces or by metabolism within the leaves of cotton. Sim- ilar results documenting the persistence of foliar residues and stability on plant material were reported by Verloop and Ferrell (1977) after application of diflubenzuron to soybeans, cabbage, corn, and apples.
In season-long studies of small plots of cotton treated at J^day intervals with multiple (six and ten) spray applications of C-labeled diflubenzuron (70 g active ingredient (Al) in 9.35
liters crop oil and 93.5 litçjs water per ha). Bull and Ivie (1978) found less than 0.01 ppm of C-labeled diflubenzuron equivalents in mature seed from cotton plants treated six times and only ca. 0.02 ppm in those from plants treated ten times. However, radio- active residues were rather high in some other parts of the plants, particularly in foliage (ca. 40-125 ppm) present during treatments.
Since diflubenzuron is quite persistent on cotton, it would probably be lost from treated foliage as a result of wind abrasion or rainwashing or because of the fall of senescent leaves. The recommended schedule of a maximum of six applications of difluben- zuron, with no more than 421 g AI per ha applied to a field during a season, should establish good coverage of plant foliage without producing hazardous residues in seed.
Soil can be contaminated with diflubenzuron directly by run- off of sprays or indirectly by contact with treated plant materi- als. In laboratory studies of soil treated directly with difluben- zuron, Metcalf et al. (1975) found that the material degraded very slowly, but Verloop and Ferrell (1977) and Mansager et al. (1979) reported rapid degradation (similar treatments but different soils). The differences can probably be attributed to the particle size of the diflubenzuron that was used in the studies since Ver- loop and Ferrell (1977) have demonstrated that the half-life of 10-ym particles in soil is 8-16 weeks, whereas that of 2-ym particles is 0.5-1 week. Particles in some of the earlier studies averaged ca. 10 ym; those in later (and current) preparations average ca. 2-5 ym. Bull and Ivie (1978) found that diflubenzuron was somewhat stable when it was applied to soil indirectly. For example, when treated cotton plants were shredded after harvest and cultivated

INSECT GROWTH REGULATORS 215
into the soil, residues of diflubenzuron did not begin to decline appreciably until the middle of the following summer, ca. 9 months later. During the 9 months, much of the residual material (ca. 80%) associated with the soil could be recovered by solvent extrac- tion, and the content was predominately (> 95%) diflubenzuron. Also, in laboratory tests (Bull and Shaver 1980) and field tests (Bull and Ivie 1978), diflubenzuron did not leach appreciably in different types of soil, and residues were concentrated near the surface (0-7.5 cm). Thus, the eventual decline of the postharvest residues in the field was probably caused by increased accessibil- ity of the molecule to different mechanisms of degradation in soil as a result of the progressive decay of the plant material with which it was initially associated. Fortunately, there is evidence (Bull and Ivie 1978) that crops (wheat, beans, collards, radish, native grass, and weeds) grown in rotation in fields previously planted to cotton that had been treated with the recommended dosage of diflubenzuron did not take up significant levels of residues.
Veech (1977, 1978a) has shown that application of difluben- zuron to cottonseed can provide protection from attack by the root- knot nematode (Meloidogyne incognita Kofoid & White), apparently by reducing the numbers of egg masses produced and by inhibiting the hatch of eggs through interference with the synthesis of chitin (which, in this organism, is found only within the egg shell). However, Veech (1978b) concluded that the levels of residues (maxi- mum of 0.1-0.3 ppm) that accumulate in soil when diflubenzuron is used for boll weevil control (Bull and Ivie 1978) probably would not provide practical control of several species of free-living nematodes.
Effects of Diflubenzuron on Natural Enemies
Because diflubenzuron appears to have little or no effect on other pest insects that often occur concurrently with the boll wee- vil, something other than an IGR may be needed to suppress major pests such as the Heliothis spp. That alternative may well be con- servation of the many natural enemies that prey on Heliothis spp. eggs and larvae and thus frequently retain these pests below eco- nomic injury levels (Whitcomb and Bell 1964, Ridgway and Lingren 1972, McDaniel and Sterling 1979).
Abies et al. (1977), Wilkinson et al. (1978), and House et al. (1980) tested diflubenzuron in the laboratory for harmful effects to several species of entomophagous arthropods. Spray applications (7.7 ppm AI) to pairs of adult convergent lady beetles (Hippodamia convergens Guerin-Meneville) reduced egg hatch and prevented larval development, but these harmful effects gradually subsided after treatments were terminated (Abies et al. 1977). Keever et al. (1977) reported similar effects on reproduction of H. convergens adults collected from diflubenzuron-treated (140 g AI per ha, 9-10 applications) cotton fields' and held in the laboratory. Wilkinson et al. (1978) reported that topical applications (concentrations

216 D. L BULL ET AL
of _< 10,000 ppm) of the IGR had no apparent adverse physical ef- fects on adult convergent lady beetles. Studies of the effects of diflubenzuron on the common green lacewing (Chrysopa carnea Stephens) indicated that egg hatch was inhibited when the adults were treated topically and that larval and pupal mortality were increased when larvae were fed topically treated prey (Abies et al. 1977). Wilkinson et al. (1978) also found that topical applica- tions of diflubenzuron to C^ carnea larvae significantly increased the mortality of pupae that developed from the treated larvae.
Laboratory tests showed that sprays of aqueous suspensions of diflubenzuron had no apparent effect on Geocoris punctipes (Say), Apanteles marginiventris (Cresson), and Trichogramma pretiosum Riley (Abies et al. 1977, Wilkinson et al. 1978). However, when applications of aqueous suspensions of diflubenzuron in oil for control of boll weevil were combined with inundative releases of Trichogramma for control of Heliothis spp. in the field. House et al. (1980) observed that the IGR treatments seemed to cause a transient reduction in the levels of parasitism of Heliothis spp. eggs by the parasites (5-44% decrease within 2-3 days postapplica- tion). These authors then conducted laboratory tests to study these effects and found (Table 1) that sprays of aqueous suspen- sions of diflubenzuron alone had no adverse effect on parasitism of Heliothis eggs, but the mixture of diflubenzuron and oil (Savol) and the oil alone significantly reduced the levels of parasitism when host eggs were treated before exposure to the parasites; there was no adverse effect when eggs were treated after they were para- sitized (Table 1). House et al. (1980) also demonstrated that treatments of host eggs with corn, soybean, and cottonseed oils had similar adverse effects on the subsequent levels of parasitism by Trichogramma.
Despite the relative consistency in results of the cited labo- ratory studies, extrapolation to actual field conditions is not ad- vised. Some of the topical applications probably produced greater direct exposure to the chemical. Also, many of the concentrations used were much higher than those that would be applied to cotton in the field. The effects of field applications of diflubenzuron on predaceous arthropods in cotton were studied in North Carolina by Keever et al. (1977) and in Texas by Abies et al. (1977) and Rummel et al. (1979). Keever et al. (1977) reported that diflu- benzuron (140 g AI in 3.8 liters of crop oil per ha, 9-10 applica- tions) caused a significant reduction in populations of G. punc- ^ipes and a reduction in the hatch of _H. convergens eggsT other predaceous species apparently were unaffected. Turnipseed et al. (1974) observed that populations of Geocoris spp. and Nabis spp. were reduced when diflubenzuron (281 or 562 g AI per ha) was applied twice to soybean foliage; however, they could not deter- mine whether the reduction was caused by the IGR, by a shortage of prey, or by a combination of these factors.
Studies by Abies et al. (1977, 1980b) in central Texas indi- cated that diflubenzuron had little or no impact on predaceous

INSECT GROWTH REGULATORS 217
Table 1.—Effect of different treatments on parasitism of tobacco budworm eggs by Trichogramma Ij
Test 1 2j 1' Test 2 h Al % % adult % % adult
No. para- emer- No. para- emer- Treatment eggs sitism gence eggs sitism gence
Untreated 629 60 a 95 a 685 72.7 a 95 a Diflubenzuron
+H 0 Savol + H^O Diflubenzuron
696 51 a 89 a 732 75.9 a 98 a 720 15 b 91 a 707 71.1 96 a
+ Savol + H^O 552 19 b 92 a 640 72.1 a 93 a
IJ Data taken from House et al. (1980). 2/ Eggs treated and then exposed to parasites. T/ Means followed by same letters are not significantly different
(P < 0.05) according to Duncan's multiple range test. Figures for % adult emergence are for eggs that were considered to have been parasitized. Data represent averages of eight or more replicates.
4/ Eggs exposed to parasites and then treated.
arthropods when sprays were applied six times at 5-day intervals at rates of 35, 70, or 140 g AI in 4.5 liters of crop oil plus 13.6 liters of water per ha. Predator populations were slightly higher in the untreated plots, but the plots receiving the highest dose of diflubenzuron had a greater abundance of predators than any of the other treated plots. (Of course, the observed differences may have been influenced by factors other than the IGR; for example, by plant phenology and prey abundance.) On the other hand, applica- tion of insecticide (methyl parathion plus toxaphene plus chlor- dimeform) severely reduced the numbers of predators (Abies et al. 1977). Also Rummel et al. (1979), in a similar study in west Texas, demonstrated that seasonal means for populations of preda- tors were higher in the plots treated with diflubenzuron (rate of 140 g AI per ha) than in plots receiving lower rates or in the un- treated control; meanwhile, applications of azinphosmethyl (336 g AI per ha) caused a major reduction in predators.
Thus, the available information suggests that diflubenzuron, and especially the emulsifiable crop oils with which the IGR is formulated, may have a detrimental impact on some of the entomopha- gous species associated with cotton. However, these effects are relatively slight and seem to diminish after treatments are termi- nated. It is clear that diflubenzuron is much more selective than most conventional insecticides. If the material is used in accord- ance with current recommendations (six applications at a rate of

218 D. L BULL ET AL
70 g per ha), many entomophagous arthropod species will be conserved.
Effects of Diflubenzuron on Field Populations of the Boll Weevil
Most of the available information obtained by applying diflu- benzuron against the boll weevil in field tests is summarized in Table 2, Almost all field testing has been done with a 25% wet- table powder (WP) formulation of diflubenzuron that has been milled to provide a mean particle size of 2-5 um.
Taft and Hopkins (1975) provided the first documentation of the effects of diflubenzuron against field populations of the boll weevil. Mixtures of the IGR with invert sugar-molasses baits caused dramatic reductions in weevil reproduction: adult emergence from egg-punctured squares was only ca. 2% in diflubenzuron-treated plots but was 89% in plots treated with conventional insecticides. Although this test involved relatively excessive amounts of diflu- benzuron (14 applications of 280-560 g AI per ha), the research represented a significant first step in the development of the product. Lloyd et al. (1977) subsequently evaluated formulations of diflubenzuron in the greenhouse and determined that mixtures of the IGR with certain oils, especially suspensions in raw cottonseed oil, were as effective as the invert sugar-molasses bait and poten- tially more convenient for practical application. Significantly, they also observed that simple mixtures of diflubenzuron and water were ineffective. When sprays of a suspension of diflubenzuron (280 g AI per ha, 16 applications) in raw cottonseed oil were then applied in the field, boll weevil reproduction was suppressed to the extent that no filial generations were detected until August 20, near the end of the season. (The late-season development of populations was attributed to incomplete control by diflubenzuron rather than to immigration of untreated boll weevils because the test site was somewhat isolated from potential sources of infestation.)
Ganyard et al. (1977) conducted an extensive study in which 141, 282, or 564 g AI per ha of diflubenzuron formulated in raw cottonseed oil were applied to small plots (0.07-0.09 ha) of cotton planted in an isolated, noncotton growing area of North Carolina. Test plots were infested artificially by releasing known popula- tions of overwintered weevils. All rates of the IGR (total of 12 applications throughout the season) provided > 99% suppression of reproduction. Meanwhile, boll weevils in the untreated (control) plots increased dramatically: as many as ca. 90,000 per ha were found; but only five insects were found in all treated plots. On the basis of these studies and those of others, Ganyard et al. (1977) concluded that diflubenzuron would have to be used regularly for prolonged periods to maintain suppression of boll weevil repro- duction because of the tendency of the insects to recover fertility after treatments were terminated.
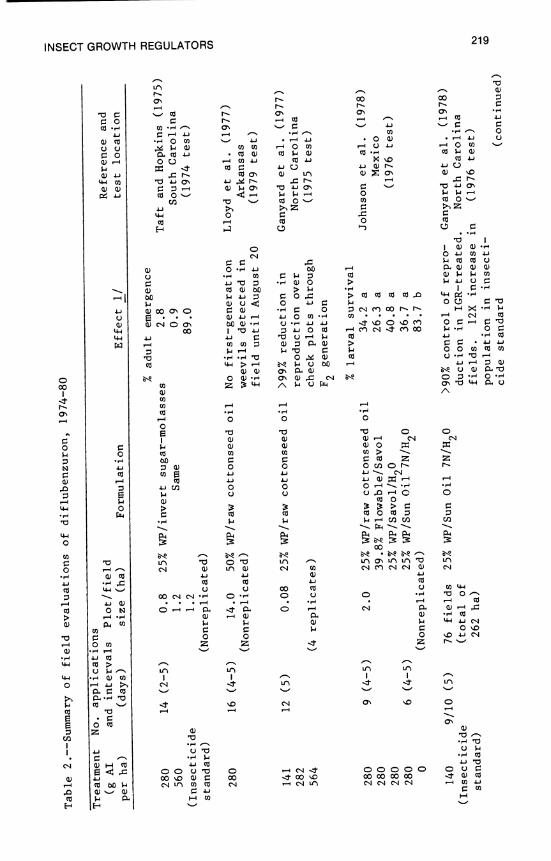
INSECT GROWTH REGULATORS 219
TJ c c= o CO •H
JJ <U CO O O C O Q) 1—1 U (D 4J
«4-4 CO (U (Ü Pi •U
O C30
I <!■ r>.
C O u N
0)
3
CO
CO >
Tí
CO
I œ
I I
CO
H
<U CO
4-t CÜ O N
PL, CD
CO >
<U CO y-i U P^ pu C CO
CO v-^ T3
. c o CO
13
<U M CO
e < ^ ■u CO Ö0 V-i (U >w/ (1)
H
c CO «H ^-N
C r-* 4J •H O CO
^ »-< 0) O. CO 4J O a
PC <i ^ r^ T? 4-» 0^ C P f-H
CO o ^^ CO
4-) IW
CO H
o c <D 00
<U C30 ON O g . . . (U CM O ON
00
CO
CO
O) CO
o
CO ÖO
CO g CO
U
X) ^,-^ /—N ^~s. (U r^ 00 00 p
^—s p->. r^ r^ c r^ <3N ON a\ •H
f^ 1—1 CO X-N 1-1 CO 4-)
Q«^ N«^ Ci >,_• 4J ^-^ P c 1—t /"N •H ^-s CO • H ^-N o v—' 4J , 1—1 4J • o (U . 1—1 4-1 o
CO CO 1—1 O CO r-l O 4-1 1-1 O CO "»-^ . CO CU CO u (U CO •H CO l-i (U
r-< CO 4-> CO 4-» X vO CO 4-»
CO C 4J O 4-t <U r-^ 4-t U CO C3^ <U in <D IS o> <U vD
4-t ^ r>. ^ r^ 1—1 ^ 1^
Qj v-i cr> n3 4.) ON c v-^ -O 4-t CJN
<3 '—t ^4 u o Í-I Í-I r-l
na ^w' CO o N.^ CO CO O ^-^
>% P^^ C >^Z O C X c
r-< CO O CO i-a O •->
1 o 1 QJ 0) •r-<
CN| O 4J CO 4J
c c rC ij CO CO O
o «H 4J 00 T—1 O- <U <U Q)
•i-i CO c v^ P CO <U 1-. Í-I CO
4_» T3 P •H 0) O > ÍJ 4J U c CO Cí 00 > u •H 1 C •H
U U 0 c o ^ > CO CO CO CO ,û M-l Pa «H <D O <í¡ o 4-) n: 1-1 O O c T? c <u •iH c o p CM CO 00 r-- r-- M X •H í^ 0) 4J rH XJ o CO • f-i CO • • • • • i-< Cvl CO
t>û <D 'H o •ri 4-1 4J <t vD o VO CO O C —t c •X3 1 TJ 4J P 4-> O CO i-< CO CNl <!- CO 00 P «H o c
4J C T3 CJ T—l Í-I CO 4-t •H CO
CO CO p CU P P- <u > c c • 4-t 4J
>-< i-< U 'O c VJ o o CO CO CO
•H .H TJ 0 M (D CO o •»-' T) 1—1
M-< > .-1 i< Í-I O Ö0 1—1 4J 1-4 p Q)
QJ 0) ON a <u Ö-o O (U ex T? O <U T-i ON 0) Jin CN 5-5 O ^ :r^ O •H
¡Z; ^ ^ /\ ^j o fu CJN TJ M-l a O
a; CO
o
s^ y^-S ^s ^-^ LO 13 O T) CM <U in cu
4-1 4-t CO CO o o
00 CM CM •H o »H r-4 • 1-H
O —• 1—1 p. <!■ a 0) r-H (U l-l Í-I c c o o !3 s
/"v
v_^
in in 1 1
CM 1
<u xi >^^ o 'O
•H Í-I 4J CO
O o O •Ü O 00 vO QJ C 00 c^^ in CO CO
4J CM
<U 0) CO C O
00 CO o o
•H o 1-H
P- eu Í-I
d) 1—1 OJ o CO >
C CO O CO 4-) -«-^ 4-> Q) O rH O ^
CO
CO O Í-I t-i \ pen
3 &^ 00
^5 • in ON CM CO
O
CM
O CNl
PC
o r-- Csl
tC I-« *-«>. «H 1-H c o > c CO p
CO CO
in in TJ CM CM 0)
Í-I c o
o CM
re ¡s
P CO
CO u-i TJ o /-^ 1-1 CO
0) 1-1 43 •H CO M-t j_» CM
O vD vD 4J CM
in I
1-1 CM <t <f 00 vO r-H CM in
CU TJ ^ •H T3 O U
•H CO 4J T3
O o O O O O U C 00 00 00 00 <!■ 0) CO
CM CM C^ CM 1—t CO 4-t

220 D. L BULL ET AL
na c c 0 íO •H
■u <u CO o O c o Q) 1—1
u Q) 4-»
U-i CO <D <U
(Xi ■U
o (30
I <t
ON
c o >-< D N
3
c TS o r-l ^^
•H <U CO ■U x: CO M-< Vw^
D •^ r-4 iJ <D
CO O N > r-H •H QJ
CO Cl^ CO
T) C 1—1 O CO 0) •H Í—1
•H ■U CO U-< CO >
O P /-N
«4-( •H Q) CO O 1—1 4-1 >>
a C CO >% fl- •r-i na >^ eo V-' CO X) ß • c ß o CO :3 :z,
1 4J • c /—s
CN <v M CO
B <d x: cu ■u
r—1 CO txO VJ X» <u 0)
CO ^ a H H
00 r^ o>
^-^ CO 4J
. CO l-< <ü CO
CO 4-1 CO X
4J VO <U Qj r^ H
CJ> <U ,-) CO v_^ p o ffi
c» 1^ o> i-H
CO
4-»
CO
1—1 0) CO CO 4J CO
X 4J r^ (U <U h-.
C3N H
<u r-i
CO S_^
P O ffi
1—1 CO CO X
d) 4J H Q)
1-4
3 pq
1—1 o '-^ CO 00 CO
C3> 4-»
4-1 —1 CO
O) >w/ 0) CO 4J CO
1—t 1—< X <U 0) vO (D Ë 6 f^ H Ë Ë ON 0 0 r-t
tó tó W
1-4 (U CO CO 4-> CO
X *j r>- (U (U r^ H
ON
O c a; W) »-• eu r
(U f^ es m ON m es r-l V£>
es 00
o C û; Ö0
<u o o e • • 0) o es
CO in
3 -a
CO
o c O)
0) o o e • • <U es vD
es vo
na CO
(D o c 0) ÖO
Q¿ o o o g . . . <u <j- o 1^
<!■ <1- es
3
CO
ä
o c QJ W)
o; m <}• r>- es e . . . . (u <!■ m ON cjN
es »-H vo
CO
O > CO
CO
o > CO
o > CO
C/3
O CM
CO
o es
O > CO
en
0^ 0^ in /-N in ^-N
es CO
<u 4-1
es CO
4J CO LO CO
es r>. es <t Ü •H
o es Ü
CM —t es o t—( CU
o o 0- <L)
CO OJ 4-> CO O
•H 1—1
O)
in /"-^ es CO
(U 4-> CO
in O • •IH 1—4 1—t
CU Cl> u
in es
O o
13 (U
a
o
I I lO
in o o o CO r- <j- in o o o
CO r-- <í- es o O o m r-- <f-

INSECT GROWTH REGULATORS 221
* CO 1—Í Q) CO
CO ■u CO
X ^^„^
4J r^ Q) o Q) r^ H 00 CO
ON ON CO i-H f—i r—1 X
<U ^w
0)
0)
0 r-) tó O
0) <u Ü o c c (U (U t>0 Ö0 M V4 <u in
0) <!■
<l; r^ CM Q) O
CO as o> c^ i-H vO <t
■u 4J r-l 1—1
P D TJ ^3 to CO
X) ^ (Ü O ^ (50
CO ON CO I-H «H r-t CO 1—1 T-t ^^ X P ^ <D
pq D CO H a 4J C CO 3 T3
O o o o 00 r^
O) Ü c 00
0) e en
00
CO
u Q) 00 u CO ^ C dJ
U-l f-H /->, O O CO :2 00 CO
ON X X) ,—1 <u c s-/ H CO
00 C
•H Xl
»-I CO m
Q) O P /—s 0) 00 CO CO CO 00 r-. 1 u ON vO CM 00 Q) r-H CN .-1 C^J
<f 00 •^»^ o r-l /—S
00 r^ CO CO CO Cï
0 ON 00 o CO <í TJ «—( vO KO \o O"
CO V--'
o CN
O > CO
O
o > CO
o es
O > CO
O CM
PC
o %
u è es
in
5>5 m CM
o o
'O in (U es /'-N /-^ 4-) /'-s CO CO CO <u (U 0) U 4J 4-» ■U
•H CO CO CO r-H in O r^ o in o CO a • •H • •H •H <u o r-t r-t »-• O 1-1 <f }-l <h ar«. D->d- (XCS c (U r—1 Q) (1) o u U U
13
es o o o in r-- «cf
r>. CO
in CO es
•H T) O Í-I
•H CO
o o O O C r^ r>. <U CO
CO CM CO r^ Q) • (1) 1—1 4-» in u
1 CO '-s. CO es o <f CJ • •rH • •H
<i .-1 <^ r-H
eu Pu Q) (U u »-• ^ es
^_^ ^-^ /"^ 1 VH
in CO »O C CO j-i CO
<U XI
CTN CO f—1 (U ^^ 4-)
CO in o • • H r—1 r-t
a 0) V-i
00 r^ <JN co r—1 (U V^ 4->
CO O u
•H •vi- r-l
a a> }^
CO o o in h- <í-
0) 4J O o; 4-1
i-H o .-H C o o o; >-< C CO
•H VJ
CO (U 0) 4-t W) 4-» CO Q) 4-» rH CO
<u <u g
«4-( CO •H CO
>% T3 ^ (U
•H T) IM CD •H ^ O O <U r-< a»-i CO o
U-l y-l o CO
4-> 1-t CO
CO Q > •H
> • • u u 4-i 0 o CO CO 4J Q;
CO 4J U r-i O 3 d)
M-( Ö0 Ö0 (D c
c u CO CO U QJ JC e *j Q)
f-i
1—( o Pu CO U •H C W) 4-> o 1—f
CO 4J 3 CO U g (U QJ CO CO CO
C 4J TH "c c CO <U II O CO C CU peí 3 í-i O O aM (U O u 4-1
CO CO 00 4J (U c CO U •H
TJ P ^3 4J U
•» Ü O T3 C o Q; 3 u *J a CO CO U 00 4->
•H 00 c »O 0) 0) c M
•H ;C (U 4-» U-l
0) .H U-i
>-• 1^ • H (U »O ^ CO
1^ (Ü >. ÍH r-t
4-t CO 4-i Pu D C <u cr CO o CO O X •H
w (= M-l o • H
^•^ 4-> C r-.|AJ Ö0
O O CO

222 D. L BULL ET AL
Because neither the invert sugar-molasses bait nor the raw cottonseed oil formulations were practical for large-scale research or commercial use, special field tests were conducted in Mexico during the spring of 1976 (in advance of the growing season in the United States) to assess the efficacy of mixtures of diflubenzuron with different commercial crop oils (Johnson et al. 1978). Results demonstrated that combinations of the IGR with Savol or Sun Oil 7N were as effective as mixtures with raw cottonseed oil: all sup- pressed boll weevil reproduction to some extent. Since that time, most field experimentation has been done with suspensions of diflu- benzuron and emulsifiable crop oil in water; in fact, the current registration label specifies that applications should include Dimoil at the rate of 4.68-9.35 liters per ha with at least two parts of water for each part of oil.
A large-scale test of the effects of diflubenzuron on cotton insects was conducted in 1976 (Ganyard et al. 1978) in Chowan County, NC, which is isolated by 40 km from other commercial cotton fields. The IGR was applied 9-10 times at a rate of 140 g per ha to the 76 fields (total of 262 ha) located in the county. Repro- duction by native boll weevils was reduced 90% while comparable initial populations (in fields 40 km west of the treated area) that were treated with conventional chemicals increased 12-fold (Ganyard et al. 1978). In addition, populations of Heliothis spp. in the fields treated with diflubenzuron were controlled during most of the growing season by natural enemies (Keever et al. 1977). (Chlordimeform had to be applied six times to these fields during August when population pressure was intense, but no broad-spectrum insecticides were used.) The study by Ganyard et al. (1978) thus established that: (1) areawide applications of diflubenzuron could effectively suppress boll weevils if treatments were initiated at the onset of fruiting and overwintered populations were low to moderate; (2) repetitive weekly applications had an accumulative effect; (3) most of the egg-punctured squares contained eggs that failed to hatch or newly eclosed larvae that died as a result of diflubenzuron treatments; they did not abort, but continued to de- velop normally; and (4) the crop seemed to mature earlier in fields treated with diflubenzuron than in fields treated with conventional insecticides. This last point has been noted by others (Harris 1980), but the cause has not yet been established. Possibly there is improved setting of the bottom crop of bolls as a result of ef- fective pest suppression, an avoidance of the lateness associated with the use of organophosphorus insecticides, or a physiological response by the plant. In any case, the effect would be advanta- geous if it is indeed real and consistent.
The studies of the field efficacy of diflubenzuron conducted in Texas differed from those conducted in the eastern region of the Cotton Belt in that all took place in areas where there was no iso- lation of treated fields from the potential influx of boll weevils from untreated areas. Also, in most of the tests made in Texas, the rates of diflubenzuron were more in line with those specified in the current registration. For example. House et al. (1978)

INSECT GROWTH REGULATORS 223
compared three rates of diflubenzuron (35, 70, and 140 g per ha, seven applications). They reported definite suppression of repro- duction by all treatments and a differential response to the different concentrations. The results of 2 years of testing in central Texas (Abies et al. 1980a, House et al. 1980) indicated that the effect of the diflubenzuron was greatest against the F, generation. They observed increases in the reproductive potential of the ensuing generations even though diflubenzuron was still be- ing applied. This effect was attributed to an influx of untreated boll weevils from adjacent fields, a conclusion in agreement with the conclusions of Ganyard et al. (1978) concerning the importance of areawide applications in achieving maximum impact on indigenous populations.
Preliminary results of large-scale replicated tests comparing the efficacy of aqueous suspensions of diflubenzuron (70 g AI per ha, six applications) with and without Dimoil (4.67 liters per ha) indicate that the IGR is somewhat less effective (ca. twofold) against the boll weevil when the oil is omitted (Table 2) (D. L. Bull unpublished information, 1980). Although both mixtures sup- pressed insect reproduction significantly from that in untreated fields or in fields treated with conventional insecticides, fewer adults emerged and more egg-punctured squares remained on the plant when diflubenzuron was applied in oil. This is the first field demonstration of diminished activity of diflubenzuron in the ab- sence of oil; and it substantiates the findings of Lloyd et al. (1977) in their greenhouse studies. Other results substantiate the report by Ganyard et al. (1978) of an apparent earliness in matura- tion of bolls in treated fields. For example, when open bolls were counted August 7, 1980, 74% were open in fields treated with IGR/ oil/water combinations, 46% were open in fields treated with IGR/ water mixtures, 14% were open in the check fields, and 17% were open in fields treated with standard insecticides.
In a study of the integrated control of codistributed popula- tions of boll weevils and Heliothis spp., Bull et al. (1979) demon- strated that treatment with diflubenzuron (70 g per ha, seven to nine applications) was compatible with augmentative releases of the egg parasite Trichogramma pretiosum and with applications of micro- bial pesticides. The biological methods effectively reduced heavy populations of Heliothis spp. (ca. mean of 140,000 eggs and 40,000 larvae per ha in the control) to levels below the economic damage threshold, and the IGR had no obvious adverse effects on the per- formance of the parasites. Even though the applications of diflu- benzuron were apparently initiated after overwintered boll weevils had entered the fields and begun ovipositing, the survival of the pest in egg-punctured squares was reduced substantially (seasonal mean of 22% adult emergence compared with 66% in untreated check). There was a distinct increase in the emergence of adults from egg-punctured squares when applications of the IGR were terminated: within ca. 3 weeks, the levels of reproduction in plots previously treated with diflubenzuron were similar to those in the check.

224 D. L BULL ET AL
probably because of the combined effects of recovery of fertility by treated insects and immigration of untreated insects.
Field tests of diflubenzuron in the cotton production area of the Texas Rolling Plains were made against extremely high popula- tions of overwintered boll weevils (> 8000 adults per ha) in 1976 (Rummel et al. 1979), and again in 1977 against somewhat lower pop- ulations (Rummel 1980). In 1976, the test fields were sprayed with malathion in an attempt to reduce the immense populations of over- wintered boll weevils to a manageable level; then applications of diflubenzuron (35, 70, and 140 g per ha, eight applications) were begun at the time when squares were in the pinhead stage. Even under these conditions, which were not at all optimum for the use of the IGR, treatments reduced reproduction to about half that in the untreated check. Yields of plots treated with the higher rates of diflubenzuron were significantly greater than those of the check and statistically equivalent to those of plots treated with azin- phosmethyl. Results of unreplicated tests in 1977 (Rummel 1980) on larger fields demonstrated that diflubenzuron was very effective against the lighter populations of boll weevils. All three rates (52.5, 70, and 140 g per ha; four applications) provided distinct levels of suppression compared with those in the untreated check, and there was a definite differential response to the different rates. The results obtained by Rummel (1980), like those in the earlier tests obtained by House et al. (1978) in Texas, thus sug- gest that a rate of 35 g of diflubenzuron per ha is insufficient to control this pest.
Large-scale evaluations in the Upper Gulf Coast area of Texas (Cole 1980) demonstrated that three or five applications of diflu- benzuron (70 g per ha) suppressed reproduction by light populations of the boll weevil to about half that observed in fields that were untreated or were treated with azinphosmethyl (Cole 1980). Cole (1980) also observed that survival of immature stages in egg- punctured squares in IGR-treated fields increased rapidly when treatments were terminated; recovery of reproductive potential be- gan about 1-2 weeks after the last application of the season.
In contrast to all the studies described thus far, tests in the Lower Rio Grande Valley of Texas over a 2-year period (1976-77) failed to demonstrate any differences in either the seasonal survi- val of boll weevils in egg-punctured squares or in yields in plots untreated or treated with diflubenzuron at rates ranging from 35 to 140 g per ha (Harding and Wolfenbarger 1980). Since the test area had a history of heavy populations of boll weevils, immigration from adjacent untreated areas may have been so extensive that the effects of the IGR were negated. There was no apparent suppression of predator populations by the diflubenzuron.

INSECT GROWTH REGULATORS 225
Potential for Use of Diflubenzuron in Pest Management
The evidence indicates that under certain conditions, diflu- benzuron (and most likely other benzoylphenyl ureas with comparable activity) can provide effective and selective control of the boll weevil. It is clear that the overwintered population of boll wee- vils is the key to effective use of diflubenzuron in a suppression program. Early-season scouting is critical because IGR treatments must begin before the overwintered generation has a chance to begin reproductive activity and become established in the field. Fortu- nately, in this country, commercial cotton is the only important host for the boll weevil; thus, distribution of the insect during the feeding/reproductive phase is reasonably predictable. The first application of diflubenzuron must be made at the time domi- nant plants in a potentially vulnerable field of cotton are produc- ing the primordial squares. Treatment should then be continued at no more than 7-day intervals because treated females tend to regain fertility after 7-10 days and because the spring emergence of over- wintered boll weevils from hibernation can continue for several weeks after fruiting begins (White and Rummel 1978). This need for repeated treatment might appear unusual since diflubenzuron is known to be quite persistent on foliar surfaces. However, this persistence may not have a major influence on efficacy because it is also well established that field populations of boll weevils tend to recover the reproductive function rapidly when treatments stop. Thus, for maximum efficacy, it may be necessary for the in- sects to have direct contact with the sprays or with recently sprayed surfaces.
If the eggs deposited in squares by IGR-treated females do not hatch, or if newly eclosed larvae die without feeding, most of the squares will continue to develop normally and will not abort (as- suming that growing conditions are favorable for plant develop- ment). This retention of superficially damaged squares is, of course, an advantage, but it complicates the assessment of current damage levels and estimates of impending populations (Ganyard et al. 1978). Traditional guidelines used in making control decisions are usually based on the results of examinations of cotton fields for the presence of adult boll weevils or for damaged fruit. As a typical example, the Texas Agricultural Experiment Station and Ex- tension Service recommend that, in fields where potentially damag- ing populations of overwintered boll weevils are found before the onset of fruiting and must be controlled, conventional insecticides should be applied beginning when squares are in the matchhead stage. The treatment is then repeated 3-5 days later if needed. Subsequently, decisions concerning the need for additional treat- ments are based upon the results of weekly examinations of 100 squares (at least one-third grown) selected at random from

226 D. L BULL ET AL.
representative areas in a field and from different parts of the plant. If boll weevil damage of the fruit reaches 15-25% during the time of squaring up to peak bloom, treatments with conventional insecticides should be initiated and repeated at 5-day (or less under heavy pressure) intervals for as long as necessary.
Obviously, such guidelines are not valid for IGR's. Decisions about launching this type of treatment will probably be based on such criteria as (1) the previous history of boll weevil activity in the area of concern, (2) the anticipated survival of the over- wintering generation, and (3) estimates of the actual survival and potential for infestation by emerging overwintered weevils based upon early-season monitoring of populations with pheromone traps or visual observations. Although all the technology needed to make such assessments is not yet in place, continued research will prob- ably lead to the development of mathematical models that will fa- cilitate decisions having to do with control strategies in cotton pest management, including the use of IGR*s.
Presently, there is no question but that diflubenzuron must be applied with a crop oil to be effective against boll weevils. Should continued research establish that other benzoylphenyl ureas (penfluron, BAY SIR 8514) with greater innate biological activity can be effective without oil, then the advantages that would accrue are obvious. The added cost of application and the apparent ad- verse action of the oil on certain nontarget organisms would be eliminated. However, we need yet to determine whether the benzoyl- phenyl ureas that seem to be more active than diflubenzuron against the boll weevil also have a more adverse effect on beneficial species.
Plainly, diflubenzuron is most effective when it can be used against low-to-moderate populations of boll weevils in large pro- duction areas where immigration of untreated insects is minimized or eliminated. Piecemeal use may well have only limited potential in commercial cotton production. However, IGR*s should serve well as components of a supervised integrated pest management (IPM) program.
An IPM program for suppressing the boll weevil while making maximum use of the natural enemies of codistributed pests such as Heliothis spp. might include: (1) destruction of cotton stalks as soon as possible after harvest to deprive the overwintering genera- tion of food, thus reducing the numbers that successfully enter diapause; (2) applications of insecticide in the fall to reduce the number of overwintering weevils; (3) the use of an early maturing variety of cotton, planted on schedule, to shorten the period of crop vulnerability to insect attack; (4) six applications of IGR (7-day intervals), beginning at the time of onset of fruiting, to suppress reproduction by overwintered insects and reduce the poten- tial for development of damaging populations in ensuing genera- tions; and (5) selective application of microbial pesticides or

INSECT GROWTH REGULATORS 227
other biological methods to enhance natural control of lepidopteran
pests.
Obviously, there are obstacles to the consistent, successful implementation of such an IPM program. For example, if populations of plant bugs must be controlled with insecticides early in the season, the natural control provided by parasites and predators will be disrupted, and problems with Heliothis spp. could increase. Also, the applications of an IGR might not adequately suppress the boll weevils. In this case, conventional controls could be needed to avoid unacceptable crop damage, and this need might develop at the time when the potential for creating a Heliothis spp. problem was greatest. Finally, populations of natural enemies might be insufficient to suppress infestations of Heliothis spp. Then aug- mentative biological control methods or perhaps conventional insec- ticides might be needed to manage these pests.
JUVENILE HORMONE MIMICS
The IGR class of pesticides includes a large number of chemi- cals that are categorized as juvenile hormone (JH) mimics (Menn and Beroza 1972). These chemicals, though they have widely different structures, have a common mode of action: they evoke a response in treated insects that mimics that induced by the natural JH (Staal 1975). Application of a JH mimic at the appropriate time can se- verely disrupt normal growth and development; the affected insect either dies or becomes incapacitated. There was much early enthu- siasm for the potential of JH mimics in the management of cotton pests, but none has yet shown sufficient potential for practical use.
The period of maximum vulnerability of lepidopteran species to JH treatment is during the last larval instar and early pupal sta- ges (Staal 1975). Cawich et al. (1974) conducted laboratory tests of one of the more promising JH mimics (Stauffer Chemical Co., R-20458 (£)-3-[5-(4-ethylphenoxy)-3-methyl-3-pentenyl]-2,2-di- methyloxirane) against the pink bollworm and found that relatively high doses of the chemical could induce both morphogenetic effects and termination of diapause. However, the cryptic feeding/develop- mental behavior of this species and the apparent lack of unusual sensitivity to JH applications are such that these chemicals do not appear to have significant potential for use in controlling the pest. Similarly, a large number of candidate JH mimics have been tested unsuccessfully against the tobacco budworm (Guerra 1970, Benskin and Vinson 1973, Guerra et al. 1973). Certain of the chem- icals induced typical JH responses such as supernumerary instars, extended larval development, and failure to pupate, but the doses required were excessive, and none of the chemicals offered suffi- cient promise for practical use. Although laboratory tests (Ben- skin and Vinson 1973) indicated that the extended development of tobacco budworm larvae also extended the feeding period and

228 D. L BULL ET AL.
resulted in oversized individuals, there is no evidence that such larvae would survive in the field or would cause increased crop damage. Because Heliothis spp. spend much of their larval develop- ment period feeding within the fruit of cotton and then burrow into the soil to pupate, they, like pink bollworm, offer an uncertain target for treatments with JH mimics, even if the chemicals have sufficient biological activity.
As a group, weevils are also considered relatively insensitive to JH mimics (Staal 1975). In the boll weevil, maximum vulnerabil- ity to JH treatment occurs during late prepupal and early pupal development (Whitten and Bull 1978). Although this pest is rela- tively sensitive to certain JH mimics (Hedin et al. 1972, Jacobson et al. 1972, Whitten and Bull 1978, Moore 1980), the immature stages develop within the cotton fruit and are usually protected during that time from any type of chemical treatment.
Bull et al. (1973) evaluated a large number of JH mimics rep- resenting a broad array of structural types for activity against such cotton pests as the tarnished plant bug, the cotton aphid, and the carmine spider mite, and also against some important predators of cotton pests including the common green lacewing, the big-eyed bug, and the convergent lady beetle. Their studies demonstrated that: (1) none of the JH mimics was effective against spider mites and only one chemical (hydroprene) had promising activity against aphids; (2) several JH mimics were active against the tarnished plant bug, but only at relatively high doses; and (3) the compounds that were effective against pests also had a deleterious effect on the three species of predators that were tested. They concluded that none of the JH mimics they evaluated were promising for prac- tical use in the field.
The evidence thus suggests that at present the available JH mimics hold little promise for use in controlling major pests of cotton. Those that have biological activity must be used at con- centrations that are too high for practical use. Moreover, the JH mimics have not demonstrated the unique selectivity associated with the aforementioned benzoylphenyl ureas; for example, JH mimics that are active against plant bugs have an equally adverse effect on beneficial hemipterans and other predators associated with cotton. The role of these compounds in pest management may well be limited to use in ecosystems where the target is well defined and readily accessible.

INSECT GROWTH REGULATORS 229
LITERATURE CITED
Abies, J. R., V. S. House, S. L. Jones, and D. L. Bull. 1980a. Effectiveness of diflubenzuron on boll weevils in central
Texas River Bottoms area. Southwestern Entomologist 5(Supple- ment Number 1):15-21.
S. L. Jones, and M. J. Bee. 1977. Effect of diflubenzuron on beneficial arthropods associa-
ted with cotton. Southwestern Entomologist 2:66-72.
S. L. Jones, V. S. House, and D. L. Bull. 1980b. Effect of diflubenzuron on entomophagous arthropods asso-
ciated with cotton. Southwestern Entomologist 5(Supplement Number 1):31-35.
Anonymous. 1975. Cotton pest control. In Pest Control: An Assessment of
Present and Alternative Technologies, Volume 3. National Acad- emy of Sciences, Washington, DC, 139 p..
1980. Annual Report, 33rd Cotton Insect Research and Control Conference, St. Louis, MO.
Ascher, K. R. S., and N. E. Nemny. 1974. The ovicidal effect of PH 60-40 (l-(4-chlorophenyl)-3-
(2,6-difluorobenzoyl)-urea) in Spodoptera littoralis Boisd. Phytoparasitica 2:131-133.
Benskin, J., and S. B. Vinson. 1973. Factors affecting juvenile hormone analogue activity in
the tobacco budworm. Journal of Economic Entomology 66:15- 20.
Bottrell, D. G., and P. L. Adkisson. 1977. Cotton insect pest management. Annual Review of Entomol-
ogy 22:451-481.
Bull, D. L., V. S. House, J. R. Abies, and R. K. Morrison. 1979. Selective methods for managing insect pests of cotton. Journal of Economic Entomology 72:841-846.
and G. W. Ivie. 1978. Fate of diflubenzuron in cotton, soil, and rotational
crops. Journal of Agricultural and Food Chemistry 26:515- 520.
and G. W. Ivie. 1980. Activity and fate of diflubenzuron and certain derivatives
in the boll weevil. Pesticide Biochemistry and Physiology 13:41-52.

230 D. L BULL ET AL.
Bull, D. L., R. L. Ridgway, W. E. Buxkemper, M. Schwarz, T. P. McGovern, and R. Sarmiento.
1973. Effects of juvenile hormone analogues on certain injurious and beneficial arthropods associated with cotton. Journal of Economic Entomology 66:623-626.
and T. N. Shaver. 1980. Fate of potassium 3,4-dichloro-5-isothiazolecarboxylate in
soil. Journal of Agricultural and Food Chemistry 28:982-985.
Cawich, A., L. A. Crowder, and T. F. Watson. 1974. Effects of a juvenile hormone mimic on the pink bollworm. Journal of Economic Entomology 67:173-176.
Chang, S. C. 1979. Laboratory evaluation of diflubenzuron, penfluron, and
BAY-SIR-8514 as female sterilants against the house fly. Journal of Economic Entomology 72:479-481.
and A. B. Borkovec. 1980. Effects of diflubenzuron and penfluron on viability of house fly eggs. Journal of Economic Entomology 73:285-287.
and J. B. Stokes. ,, 14^
1979. Conjugation: the major metabolic pathway of C-diflu- benzuron in the boll weevil. Journal of Economic Entomology 72:15-19.
Cole, C. L. 1980. Effectiveness of diflubenzuron in the upper Gulf Coast of
Texas. Southwestern Entomologist 5(Supplement Number l):22-26.
Flint, H. M., and R. L. Smith. 1977. Laboratory evaluation of TH-6040 against the pink boll-
worm. Journal of Economic Entomology 70:51-53.
R. L. Smith, D. Forey, and B. Horn. 1977. Diflubenzuron: evaluation for control of the pink boll- worm, cabbage looper, and cotton leafperforator in a field cage test. Journal of Economic Entomology 70:237-239.
R. L. Smith, J. M. Noble, D. Shaw, A. B. Demilo, and F. Khalil.
1978. Laboratory tests of diflubenzuron and four analogues against the pink bollworm and a field cage test with difluben- zuron and EL-494 for control of the pink bollworm and cotton leafperforator. Journal of Economic Entomology 71:616-619.
Ganyard, M. C, J. R. Bradley, J. F. Boyd, and J. R. Brazzel. 1977. Field evaluation of diflubenzuron (Dimilin) for control
of boll weevil reproduction. Journal of Economic Entomology 70:347-350.

INSECT GROWTH REGULATORS 231
Ganyard, M. C, J. R. Bradley, and J. R. Brazzel. 1978. Wide-area test of diflubenzuron for control of an indige-
nous boll weevil population. Journal of Economic Entomology. 71:785-788.
Grosscurt, A. C. 1976. Ovicidal effects of diflubenzuron on the housefly (Musca domestica). Mededelingen Rijksfaculteit Landbouwwetenschappen, Gent 41:949-963.
1978. Diflubenzuron: some aspects of its ovicidal and larvici- dal mode of action and an evaluation of its practical possibili- ties. Pesticide Science 9:373-386.
Guerra, A. A. 1970. Effect of biologically active substances in the diet on
development and reproduction of Heliothis spp. Journal of Eco- nomic Entomology 63:1518-1521.
D. A. Wolfenbarger, and R. D. Garcia. 1973. Activity of juvenile hormone analogs against the tobacco
budworm. Journal of Economic Entomology 66:833-835.
Hajjar, N. P., and J. E. Casida. 1979. Structure-activity relationships of benzoylphenyl ureas
as toxicants and chitin synthesis inhibitors in Oncopeltus fasciatus. Pesticide Biochemistry and Physiology 11:33-45.
Harding, J. A., and D. A. Wolfenbarger. 1980. Evaluation of diflubenzuron for boll weevil control in the Lower Rio Grande Valley of Texas. Southwestern Entomologist 5(Supplement Number l):27-30.
Harris, W. 1980. Dimilin status, cotton boll weevil. Ln Proceedings, 1980 Beltwide Cotton Production Mechanization Conference, St. Louis, MO, p. 78.
Hedin, P. A., L. R. Miles, A. C. Thompson, and R. C. Gueldner. 1972. Constituents of the boll weevil. V. Factors inhibiting
larval development. Journal of Economic Entomology 65:1232- 1235.
House, V. S., J. R. Abies, S. L. Jones, and D. L. Bull. 1978. Diflubenzuron for control of the boll weevil in unisolated
cotton fields. Journal of Economic Entomology 71:797-800.
J. R. Abies, R. K. Morrison, and D. L. Bull. 1980. Effect of diflubenzuron formulations on the egg parasite Trichogramma pretiosum. Southwestern Entomologist 5:133-138.

232 D. L BULL ET AL
Ivie, G. W., and J. E. Wright. 1978. Fate of diflubenzuron in the stable fly and house fly. Journal of Agricultural and Food Chemistry 26:90-94.
Jacobson, M., M. Beroza, D. L. Bull, H. R. Bullock, W. F. Chamberlain, T. P. McGovern, R. E. Redfern, R. Sarmiento, M. Schwarz, P. E. Sonnet, N. Wakabayashi, R. M. Waters, and J. E. Wright.
1972. Juvenile hormone activity of a variety of structural types against several insect species. Ln Insect Juvenile Hormones, p. 249-302. J. J. Menn and M. Beroza, editors. Academic Press, New York and London.
Johnson, W. L., D. L. Moody, E. P. Lloyd, and H. M. Taft. 1978. Boll weevil: egg hatch inhibition with four formulations
of diflubenzuron. Journal of Economic Entomology 71:179- 180.
Keever, D. W., J. R. Bradley, Jr., and M. C. Ganyard. 1977. Effects of diflubenzuron (Dimilin) on selected beneficial
arthropods in cotton fields. Environmental Entomology 6:732- 736.
Lacey, L. A., and M. S. Mulla. 1978. Biological activity of diflubenzuron and three new IGR's
against Simulium vittatum (Diptera: Simulidae). Mosquito News 38:377-381.
Lloyd, E. P., R, H. Wood, and E. B. Mitchell. 1977. Boll weevil: suppression with TH-6040 applied in cotton-
seed oil as a foliar spray. Journal of Economic Entomology 70:442-444.
Lovestrand, S. A., and J. B. Beavers. 1980. Effect of diflubenzuron on four species of weevils attack-
ing citrus in Florida. Florida Entomologist 63:112-115.
McDaniel, S. G., and W. L. Sterling. 1979. Predator determination and efficiency on Heliothis vires-
cens eggs in cotton using P. Environmental Entomology~Ti 1083-1087.
McLaughlin, R. E. 1976. Response of the boll weevil to TH-6040 administered by
feeding. Journal of Economic Entomology 69:317-318.
Mansager, E. R., G<i¿G. Still, and D. S. Frear. 1979. Fate of [ C]diflubenzuron on cotton and in soil. Pest-
icide Biochemistry and Physiology 12:172-182.
Menn, J. J., and M. Beroza. 1972. Insect juvenile hormones. Academic Press, New York and London, 341 p.

INSECT GROWTH REGULATORS 233
Metcalf, R« L., Lu Po-yung, and S. Bowlus.
1975. Degradation and environmental fate of l-(2,6-difluoroben- zoyl)-3-(4-chlorophenyl)urea. Journal of Agricultural and Food Chemistry 23:359-364.
Moore, R. F. 1980.^ Boll weevils: effect of insect growth regulators and ju- venile hormone analogues on adult development. Journal of the Georgia Entomological Society 15:227-231.
A. Leopold, and H. M. Taft. 1978. Boll weevils: mechanism of transfer of diflubenzuron from male to female. Journal of Economic Entomology 71:587-590.
and H. M. Taft. 1975. Boll weevils: chemosterilization of both sexes with bu-
sulfan plus Thompson-Hayward TH-6040. Journal of Economic En- tomology 68:96-98.
Mulder, R., and M. J. Gijswijt. 1973. The laboratory evaluation of two promising new insecti-
cides which interfere with cuticle deposition. Pesticide Science 4:737-745.
Nemec, S.
1978. How a consultant looks at Dimilin. Ln Dimilin: Break- through in Pest Control, p. 19-20. Agri-Fieldman and Consult- ant, Willoughby, OH.
Oliver, J. E., A. B. Demilo, R. T. Brown, and D. G. McHaffey. 1977. AI-63223: a highly effective boll weevil sterilant. Journal of Economic Entomology 70:286-288.
Post, L. C, B. J. DeJong, and W. R. Vincent. 1974. l-(2,6-disubstituted benzoyl)-3-phenylurea insecticides:
inhibitors of chitin synthesis. Pesticide Biochemistry and Physiology 4:473-483.
Ridgway, R. L., and P. D. Lingren. 1972. Predacious and parasitic arthropods as regulators of Heliothis populations. Ln Distribution, Abundance, and Control °^ Heliothis Species in Cotton and Other Host Plants. Southern Cooperative Service Bulletin No. 169, p. 48-56.
Rummel, D. R. 1980. Effectiveness of diflubenzuron on boll weevils in the Tex-
as rolling plains. Southwestern Entomologist 5(Supplement Num- ber 1):8-14.
G. R. Pruitt, J. R. White, and L. J. Wade. 1979. Comparative effectiveness of diflubenzuron and azinphos- methyl for control of boll weevils. Southwestern Entomologist 4:315-320.

234 D. L BULL ET AL
Schroeder, W. J., J. B. Beavers, R. A. Sutton, and A. G. Selhime. 1976. Ovicidal effect of TH-6040 in Diaprepes abbreviatus on
citrus in Florida. Journal of Economic Entomology 69:780-782.
Staal, G. B. 1975. Insect growth regulators with juvenile hormone activity. Annual Review of Entomology 20:417-460.
Still, G. G., and R. A. Leopold. 1978. The elimination of (N-[[(4-chlorophenyl)amino]carbonyl]-
2,6-difluorobenzamide) by the boll weevil. Pesticide Biochem- istry and Physiology 9:304-312.
Taft, H. M. 1978. Early tests with Dimilin on the boll weevil. In Dimilin: Breakthrough in Pest Control, p. 10-11. Agri-Fieldman and Con- sultant, Willoughby, OH.
and A. R. Hopkins. 1975. Boll weevils: field populations controlled by sterilizing
emerging overwintered females with a TH-6040 sprayable bait. Journal of Economic Entomology 68:551-554.
Turnipseed, S. G., E. A. Heinrichs, R» F. P. da Silva, and J. W. Todd.
1974. Response of soybean insects to foliar applications of a chit in synthesis inhibitor TH-6040. Journal of Economic Ento- mology 67:760-762.
Veech, J. A. 1977. A possible new approach to the chemical control of nema-
todes. Journal of Nematology 9:184-185.
1978a. The effect of diflubenzuron on egg formation by the root- knot nematode. Journal of Nematology 10:208-209.
1978b. The effect of diflubenzuron on the reproduction of free- living nematodes. Nematologica 24:312-320.
Verloop, A., and C. D. Ferrell. 1977. Benzoylphenyl ureas—a new group of larvicides interfering with Chitin synthesis. In. Pesticide Chemistry in the 20th Cen- tury, p. 237-270. J. Plimmer, editor. ACS Symposium Series Number 37, American Chemical Society, Washington, DC.
Wellinga, K., R. Mulder, and J. J. Van Daalen. 1973. Synthesis and laboratory evaluation of l-(2,6-disubsti-
tuted benzoyl)-3-phenylureas, a new class of insecticides. II. Influence of the acyl moiety on insecticidal activity. Journal of Agricultural and Food Chemistry 21:993-998.

INSECT GROWTH REGULATORS 235
Whitcomb, W. H., and K. Bell. 1964. Predaceous insects, spiders, and mites of Arkansas cotton
fields. Arkansas Agricultural Experiment Station, Bulletin No. 690, 83 p.
White, J. R., and D. R. Rummel. 1978. Emergence profile of overwintered boll weevils and entry
into cotton. Environmental Entomology 7:7-14.
Whitten, C. J., and D. L. Bull. 1978. In vivo effect of a juvenile hormone analogue on the non-
spec i"fTc""iest erase s of the boll weevil. Southwestern Entomolo- gist 3:226-231.
Wilkinson, J. D., K. D. Biever, C. M. Ignoffo, W. J. Pons, R. K. Morrison, and R. S. Seay.
1978. Evaluation of diflubenzuron formulations on selected para- sitoids and predators. Journal of the Georgia Entomological Society 13:227-236.
Wright, J. E., and R. L. Harris. 1976. Ovicidal action of Thompson-Hayward TH-6040 in the stable
fly and horn fly after surface contact by adults. Journal of Economic Entomology 69:728-730.
Wright, J. E., and G. E. Spates. 1976. Reproductive inhibition activity of the insect growth reg- ulator TH-6040 against the stable fly and the house fly: ef- fects on hatchability. Journal of Economic Entomology 69: 365-368.
Zoebelein, G., I. Hammann, and W. Sirrenberg. 1980. BAY SIR 8514, A new chitin synthetase inhibitor. Zeitschrift fuer Angewandte Entomologie 89:289-297.


237
Chapter 10
INSECTICIDES FOR CONTROL OF COTTON INSECTS
C. R. Parencia, Jr. and T. R. Pfrimmer Bioenvironmental Insect Control Laboratory Agricultural Research Service U.S. Department of Agriculture Stoneville, MS 38776
A. R. Hopkins Cotton Production Research Agricultural Research Service U.S. Department of Agriculture Florence, SC 29502
ABSTRACT Dust formulations of inorganic in- secticides such as arsenicals, sulfur, and cryolite were applied by airplane in the 1920's to control cotton insects. When the organo- chlorine and organophosphorus insecticides became available in the mid 1940's and late 1940's, they could be formulated as emulsifi- able concentrates that could be applied as emulsion sprays with low-pressure, low-volume spray equipment. A high percentage of the cot- ton acreage is presently treated by making aerial applications of insecticide sprays. However, many of the insect pests of cotton have developed or are developing resistance to insec- ticides. Therefore, researchers are searching for alternate methods of control. Use of these new methods to manage insect populations is re- ducing the number of insecticide applications needed to achieve control of target insects.
INTRODUCTION
Problems with cotton insects in the United States did not be- gin in 1892 with the introduction of the boll weevil (Anthonomus grandis Boheman) (Townsend 1895). The early colonists who culti- vated cotton had problems with other insects; and T. Glover, the first Government entomologist, mentions cotton insects in his first and second reports (Glover 1854, 1855). In those days, the cotton Leafworm (Alabama argillacea (Hübner)) and the bollworm (Heliothis sea (Boddie)) were the most important pests of cotton (Comstock L879, Riley 1885), and there are many references to other insects affecting cotton in the early entomological publications.

238 C. R. PARENCIA, JR., ET AL
Then the boll weevil entered Texas and at once became the most important pest of cotton in the rain belt portion of the Cotton Belt. Today the boll weevil is present from the eastern two-thirds of Texas and Oklahoma to the Carolinas, and it often causes exten- sive losses to cotton producers.
Dependable methods of controlling the boll weevil were not available until insecticides were developed in the 1920*s. How- ever, once they became available, large quantities of insecticides have been applied annually by growers to the point that estimated costs (insecticides plus application) now range from $250 million to $300 million a year. Indeed, even in 1976, the National Cotton Council of America put the price at $260 million annually for the period 1970-72, and by 1976, an estimated 64.1 million pounds of insecticide were being applied for control of cotton insects (Eichers et al. 1978). As a result, strains of boll weevil have developed that are resistant to the organochlorine insecticides used initially, which leaves the organophosphorus insecticides as the major class available for control. Meanwhile, the Heliothis spp. complex, which is present across the Cotton Belt, has become resistant to both organochlorine and organophosphorus insecticides.
In fact, cotton insect problems are interrelated and are com- plex. The boll weevil is the key pest in the rain belt, Lygus spp. in the Midsouth and far West, the cotton fleahopper (Pseudatomos- celis seriatus (Reuter)) in the Southwest, and the pink bollworm (Pectinophora gossypiella (Saunders)) in the far West. The Helio- this spp. complex now is a serious problem beltwide, in part be- cause natural populations of beneficial arthropods that provide control have been killed with insecticides applied for control of the other insect pests.
COTTON CROP LOSSES TO INSECTS
Any discussion of insecticides for control of cotton insects must include some details concerning grower losses. For a fuller discussion of the topic, please refer to Chapter 13 of this handbook.
Estimates of annual loss of cotton due to the boll weevil and to other insects for the period 1909-54 were prepared by the Bureau of Agricultural Economics and are shown here in Tables 1 and 2, respectively (taken from file of C. R. Parencia, Jr.).
The U.S. Department of Agriculture estimated that from 1942 to 1951 in States where the boll weevil occurs, 10.1% of the projected yield was lost because of boll weevils. The loss due to other cot- ton insects ranged from 1.6 to 8.9%. In States where the boll weevil does not occur, estimated losses to insects ranged from 2.9 to 10.1% (Anon3anous 1954). Also, the U.S. Department of Agricul- ture put the losses in cotton yield due to insects for the period

INSECTICIDES FOR CONTROL OF COTTON INSECTS 239
1951-60 at 19% and attributed 8% to the boll weevil (Anonymous 1965). More recently, the National Cotton Council estimated the annual average loss in yield to the boll weevil as 7.4% for the period 1970-72 (National Cotton Council 1973), and the Cotton Foun- dation put all insect losses to the cotton crop for 1974-76 at 6.6% annually (DeBord 1977). Even today with all the sophisticated con- trol measures that are available, the loss in yield to insects for the Cotton Belt for 1979 was put by the annual Conference on Cotton Insect Research and Control (Anonymous 1980) at 8.8% with 1.4% at- tributed to the boll weevil. (This group will develop figures an- nually hereafter.)
INORGANIC INSECTICIDES
The development of materials capable of controlling the boll weevil is of considerable historical interest. In the early part of the century, before the advent of calcium arsenate, Paris green and lead arsenate were the principal insecticides used to control outbreaks of the cotton leafworm. (The Paris green sulfur mixture became popular in the far West for control of hemipteran insects in the 1920*s.) Since Paris green was so effective against the cotton leafworm, it was tested against the boll weevil in 1896 but found to be ineffective. Also, a wettable powder of lead arsenate formu- lated as a spray was effective in controlling the bollworm and the cotton leafworm but was not effective against the boll weevil. Even when this material was formulated as a dust, control of boll weevils was erratic, so lead arsenate was not recommended by the Bureau of Entomology nor used extensively by growers. Sulfur, too, was sometimes used for control of the cotton fleahopper, and cryo- lite was used for control of the bollworm on cotton though it was not as effective in preventing damage as it was on such crops as lima beans.
The first significant breakthrough in chemical control came when Coad (1918) found that calcium arsenate was effective in field tests against the boll weevil, and machines that could be used for applying the dust were developed. The next step was the applica- tion of calcium arsenate dust from airplanes, a procedure suggested by the success the Ohio Agricultural Experiment Station achieved with aerial applications of lead arsenate for control of the catal- pa sphinx (Ceratomia caltalpae (Boisduval)) in 1921. The first aerial applications against cotton insects were made to control the cotton leafworm in 1922. This method of application rapidly became widespread, and commercial companies soon maintained fleets of air- planes for use in controlling the boll weevil and the cotton leaf- worm. Thereafter, for many years, that is, until 1945, arsenicals and sulfur were the principal insecticides used for control of cot- ton insects (Parencia 1978).
Although a mixture of calcium arsenate and Paris green was more effective against bollworms than calcium arsenate alone (Parencia 1978), calcium arsenate was the main weapon against the

240 C. R. PARENCIA, JR., ETAL.
CO u
mmr-HmvDr^r>o<T>vo CNON—tcoo>oo<ti^ON<f comoo
O 0^
vor^cscMcocnmr^ocn moNCSoovO'-d-cNcor-im vocsm .—I CM r-H -H r-H r-H
g <f CS «—t on C>J ^
O X H I—I I—II—I f—ICvICO«—<»-H i-HCM«-Hi-H
0)
o
o
I—t
•H
Cd
o
< ij
œ S
H
o
cNOi—•<j-mooo<tcMO insoinLocomooNCSoo r^^co»—•
.—I r-imCNCSCOCN«—' CNCOCOCMfO rH,-H -H r-H
OOOCS<l-vDvX>000>CS o>vDr»4vDcocsimcomcs <t<tCX5 r-iCMCN'-H CMCOCOCSCO—t .—tr-i ,—t
OOOOOOO-HCSIO O'-ir^ON—iCSOCMCOCM CMr-iCN
OOOOOr-HOOCsir^ OOOO-^Or-^CNOO ooo
a; u o CO
u pu
^
O«—•00<NOco-Hr>«.<l- ocNoocMcooov£><fo>aN <l-<i-o .—t —tcNcscM <tcncsicocnc^ r-t ^ ,—i
ooooooocoo^«—• ON.—im<}-r^mr>-mcjo<J- mr>-r-* r-1 r-im^-<tCOr-l .—lr-4 r-i
o
Pi5 I I
en oooooooooo cncor-^or^vDcs<i-r^m oocooo
CO
13
>
oooooooooo oo-^cocnr^oocovocM r-ir^oo .-H r-H .—I .—( es '-^
OOOOOOOOOO oooooooocNo <tcno
o pu CO o CO
U CO
ONO'-ícsicn^tiAvor^oo <7NO-H<Nm<fmvor>-oo ONO—' o.—I.—i—fr-irH.—i^Hr-Hi—I IF-ICSCNCSICSICSCSCMCMCM csicnco ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON ON

INSECTICIDES FOR CONTROL OF COTTON INSECTS 241
ino>r^oO'<i-inaN oovomoovDcnocof^iA
ir» so r^ o 00 CO
r^ CM vD in r^ CO ^ CM
cooN^t'^'—«cNim cocNii—iLncN<-Hcov£)-<j-<f in vD CO vo <]• CO CS CM ^
<í-O^^^CN<í <l-Csí00OC0C0C0'-HCr>O 00(T>00vO<tCN
.-ivD<fr^oouooo mr>-.-Hr^<t<fcMO<l-«Ñf cocs>d-<fc^cs
in I—I ON i-H lO VO ON oooc>40Nvx><faNcoaNco in r^ o vo 1^ »^ .—( CM -H
in in <i- ON CO <t <—• coocovoin<tcso\ON<i- r-H .—) CNl I—I f-H
r^ CO r-. -vt vo >:i CM CvJ
^H CM r-l ON <f- m o CS| ,-H i-H 1-H
oocNOcor-HiACJNr^or^ I—I i-H CM >—t r-H i-H rH
I—I CO ON vD O lA CO CO —I
ON00CM^«-Hi—ti—I CO«—It-Hi-Hi-H.—IrHCMi-HrH COONf^CM-víi-H
OOOOOOO OOOOOOOOOO o—i-HrHrHO
in ON 00 uo r^ r^ CO CM I-H 1-H .—I
<f<tcjNCOOoo<J-omr^ 1—I.—I«-H«-Hi-H i-HCM«—I
00 CM o ON in <!" •-H CM >-H i-H
CM 00 CM CM v£> o 00 CM —^. -—I -H f-H
>:tor-«.mmoo<i-r-iinco in in vD r^ >^ vO CO CM —H
in <i- »-H in m I—t VÛ oo<j-r^cMinvoomcjNOO CM -—I t—I i-H i-H I—I
ON •—I uo r^ ON vo CoJ CO
<1- 00 r^ CJN <}■ .-H vD f—I ^ CM
CO-—lor^ONinoNcovDvo CM '-H .-H rH
r^ >d- »-H ON in f^ cvi in <—I "H
CM r- <!■ CM —t o •—I rH -H CM
C^JCMCOvDvO«—icoincocM CO —I
CO CO VO 00 CM 00 CM VO rH .-H
CM CO -Í in VÛ r^ 00 CO CO CO CO CO CO CO ON ON ON ON ON ON ON
ONO'-HCMco<finvor^oo co<l-<í-<l-<f<l-<t<}-<f<í- ON ON ON ON ON ON ON ON ON ON
ON O ^^ Csl CO <!■ <}• in in in tn in ON ON ON ON ON ON

242 C. R. PARENCIA, JR, ETAL
> 0)
o
CO
O
(1)
C ^ O in
■u I
O O
O
T) I—I 0)
•H
o CO
u Pu
I I
U3 CO H
O Cl. CO O CO
U CO
1—i.-^mcocNcoc^cMcMCM m<f<i-csr^-d-cNcx5<fco CM»—••-<
—•CO<t<^'-'CX)CMCSCNCM CSlCSO^C0r>.CM«—tC30in»-t ,-ir-ti-(
csiv£>omcMcovocM<tco cnvDo^cncnoo^o<^oo<l■ cJi—tt—t
rocsvû<î">^vococNjcnco cor^mcnr^r^cnN^vovo mmcn
mcSvDCO'-icnOCM'-i'-i <tC0CMC0ON«-<CS<tC0CM ,—1,—ir-4
mcsvomi—t<i"'—<r->«.cs«—t cococscsvo»—••—»VOCMCM ,—i,—tr-i
CM»—tu^CS'-tt—ICMCM«—<Cs| r-<cOr-ii—100»—»OmCS--^ ,—I,-H,-(
oo«—'•—'»-'cn«—•c»4»-t.-H f—icscncNO«—•«—'r^cNr-4 ,_i,—(,_<
O«—'•—' OCM OO I—tOCMCMOCOOvOOiO LO.—i.—t
p^ocom«—'CîO-Nfco»—'CM <too<i-moo-d"Or^r^cM CMCO«—•
i-<»-i»-iCMi—11—11—ICM»—<CM CMCM<l-CO<l-'—I»—lOCMCM ,-HO'-^
r-HCM^H.—ICM«—ICM«—••—tt—I ,—iCMCM«—iCM»—II—100«—•«-< ^^r-ir-«
^^^^Cno»-ICMCMCO .—ICO«—iCMCMCMi—I—iinCM r-H.—tcO
O'-toom—looom O—ICM CM—IOOOO O^-^CM
ONO«—iCNicn<i-inNX>r^<30 ONO—icMcn-Ñfu^vor^oo o^o•—i O—4—ir-i—I—I—irHi-^—I —ICMCMCMCSCMCMCMC^CM CMCOCO O^CJ^<J^O^O^O^O^O^O^O^ O^0^0^C7^O^O^O^O^0^CJ^ 0^0^0^

INSECTICIDES FOR CONTROL OF COTTON INSECTS 243
•—icMvoooocN cMO>c3000covDrHvDvo r>»mr>>.r^r^r«-
cMcocs^cMCMco -H.—i<f<j-^HCMi-Hi—I.—ii—I i—icnm.—1^^.—I
cncncNvDcom^j- cSrHvocMcscs<icscNCN cNc>>j<|-cMCNim
mmcMom^oo -;t>ctor^<l-mvDmo-)m cnr^^fcocom
mcscsmcN«-Hcs CN»—ivocnf-Hi—icncsrH.—i .—icocs-—icscsj
CMi—ti—II—11—11—ICS I—11—ICSi—11—11—Ir—11—11—lO I—iCSCMi-Hf—11—I
Csj,-!,—ír-ICMi—ICS CSCNCS
CNi—<i—l'vtCn»—ICO Or-I»—I«—I»-HOI-HOI—li-H OCNCSi—IrHi—I
1—11—11—IVOCM^HVO I—lO«—ICn^HOO«—IOCS 1—IVD»—IrH.—ICS
CS,-<rHi—l^CM^ COCSCSCSCNi—iCMi-ICMrH CSCOi—I—iir>CM
cncSi—I—iCSi—ICM CSi—iCNJCSCSCSCSi—11—ICN CNl-^1—if-HCSCS
1—If—11—11—iCMi—HCS I—Ir-ICSf—11—If—11—It—If—11—I CMcni—<i—Ir-ICN
1—11—11—IrHCMi—ICM 1—lOi—II—11—I»—Ir-lf-Hf-Hi—I i-HCSCSt—Ir-ICS
1—11—lCMO«-Hi—I VOvOf—11—11—IrHrHi—lOO CSI<—im<—li—ICS
cMro^^unvor^oo ONO--icsm<l-mvor^cx) ONO—icvjco<i
0^0^0^0^C3^0^0^ O^O^CJ^O^O^O^O^O^O^O^ 0^0^0^0^0^0^

244 C. R. PARENCIA, JR., ET AL
boll weevil, bollworm, and cotton leafworm. However, when this material was used, there tended to be injurious infestations of the cotton aphid (Aphis gossypii Glover). For this reason, nicotine, the only available aphicide, was sometimes mixed with calcium ar- senate or formulated in hydrated lime. Also, mixtures of sulfur and calcium arsenate were widely used for control of combination infestations of boll weevils and cotton fleahoppers. Sulfur sup- pressed populations of cotton aphids, but sulfur used in combina- tion with arsenicals did not always prevent injurious infestations.
ORGANIC INSECTICIDES
The development of organochlorine insecticides in the mid- 1940' s brought about the first major change in insecticides used on cotton since the development of calcium arsenate (Ewing and Paren- cia 1948). Federal and State research workers concerned with cot- ton insects made haste to explore the possibilities of these new materials. Their general effectiveness then produced wide accept- ance by growers. As a result, by the late 1940's, considerably more cotton acreage was being treated with insecticides than ever before, and organochlorine compounds had largely replaced calcium arsenate though for a while it continued to be used to some extent in certain areas. Much of this changeover was based on research accomplished at U.S. Department of Agriculture research labora- tories and at many State agricultural experiment stations.
Among the new materials, BHC, aldrin, dieldrin, chlordane, and heptachlor were effective against boll weevils but were ineffective against bollworms, and mixtures of these materials with DDT gave effective control of combination infestations of these two impor- tant pests. Toxaphene and endrin were effective against both pests. Subsequently, organophosphorous compounds such as para- thion, methyl parathion, azinphosmethyl, demeton, and EPN and the carbamate, carbaryl, were developed, and these provided a broader range of chemical weapons with which to combat cotton insects.
INSECTICIDES APPLIED AS DUSTS
In the mid-1940's when the organochlorine insecticides were being introduced, the only equipment available for applications was that developed for application of calcium arsenate dust; thus, for several years, the new materials were applied only as dusts. There were, however, two major problems. One was the fact that weather conditions were critical. Dusts had to be applied during late afternoons, at night, or in early morning when the air was calm, or nearly so, to insure plant coverage and prevent excessive drift. In many areas, such conditions lasted only a short time and often did not occur at all so the acreage that could be treated in a day was limited. The second was the fact that use of the organochlo- rine insecticides often resulted in the buildup of populations of such secondary pests as spider mites (Tetranychus spp.), though

INSECTICIDES FOR CONTROL OF COTTON INSECTS 245
this particular difficulty was solved by including at least 40% sulfur in the dust formulations.
Entomologists at the Waco, TX, Cotton Insects Laboratory therefore took a new look at "presquare" control of boll weevils with the idea of developing a method of controlling early-season injurious insects such as thrips, cotton aphids, overwintered boll weevils, and cotton fleahoppers. Community-wide experiments in early-season cotton insect control with dust formulations of organ- ochlorine insecticides conducted in Wharton County, TX, in 1948, proved conclusively that an early-season control program was prof- itable for that area and would simplify the farmer's control prob- lem (Ewing and Parencia 1949a). However, the critical weather conditions necessary for application continued to limit the effec- tiveness of such control programs.
INSECTICIDES APPLIED AS SPRAYS
In 1948, the new organochlorine insecticides were found to be effective when they were formulated as emulsifiable concentrates and applied as emulsion sprays in low-pressure, low-volume (2 to 10 gallons per acre) spray equipment. Again, community-wide, early- season insect control experiments with ground machine sprays were conducted in central Texas. The results obtained in 1949 and 1950 were outstanding, and the problems experienced with dusts were con- siderably reduced (Ewing and Parencia 1949b and 1950). Moreover, the successes laid the groundwork for the concepts of total insect population suppression and integrated control that came to the forefront in the United States in the 1970's.
Spray applications of insecticides for control of late-season insect pests also were readily accepted by growers because of their several advantages over like application of dusts. For example, they fit well into a daytime farm operation, and weather conditions are less critical than with dusts. Today a high percentage of cot- ton acreage is treated with sprays applied by aircraft.
Meanwhile, however, problems of cotton producers with the spi- der mite had become somewhat more serious because sulfur could not be included in sprays of organochlorine insecticides. This diffi- culty was solved when organophosphorous insecticides such as para- thion, methyl parathion, and demeton became available as sprays.
Finally, in the 1960's, techniques were developed for applying technical materials as ultra-low volume sprays (0.5 gallon or less of total spray per acre) (Cleveland et al. 1966). Research results showed that most materials were at least equally effective when they were applied in this way as they were when the same dose was applied as low-volume (conventional) sprays. Only malathion, azinphosmethyl, endosulfan, methyl parathion, and a mixture of mal- athion plus methyl parathion are presently approved for ultra-low volume aerial applications to control certain cotton insects.

246 C. R. PARENCIA, JR., ET AL
SYSTEMIC INSECTICIDES
Systemic insecticides such as phorate and disulfoton (de- veloped in the mid-1950's) can be applied to cottonseed and will protect seedling plants for several weeks from such pests as thrips, the cotton aphid, and spider mites (Parencia et al. 1957). This method of control, however, has not been widely accepted be- cause the insecticides are expensive, do not give protection as long as desired early in the season, often reduce plant emergence, and may delay fruiting. Also, granular formulations of systemic insecticides can be applied in the seed furrow at planting and as side-dressings to established plants. For example, aldicarb (de- veloped in the 1960's) applied in the seed furrow at planting will kill overwintered boll weevils and control cotton fleahoppers for 8 weeks after planting. It will also kill boll weevils developing in cotton squares when it is applied to plants as a side-dressing at the time they are in the early squaring stage (Hopkins and Taft 1965); however, it is not used in this manner because the applica- tions often result in increased subsequent infestations of the bollworm.
RESISTANCE TO INSECTICIDES
In Louisiana, the boll weevil developed resistance to the or- ganochlorine insecticides in the mid 1950's (Roussel et al. 1957); in Arkansas and Mississippi and in other States where boll weevils occur, resistance developed somewhat later. Fortunately, an organ- ophosphorous compound, methyl parathion, was effective and avail- able. Also, azinphosmethyl and malathion had been tested and were soon available though they were more expensive, which limited their early use. Once azinphosmethyl was marketed competitively, it came to be considered the most effective "weevilcide" available. In ad- dition, the carbamate, carbaryl, was soon available, but it never gained full grower acceptance because it could be formulated only as a dust or a wettable powder and so was more difficult to apply than an emulsifiable concentrate. A mixture of toxaphene and DDT, though both are organochlorines, continued to be effective against resistant boll weevils. However, DDT was banned for use on cotton by the Environmental Protection Agency on December 31, 1972.
In sum, since 1947 when organic chemicals began to be widely used on cotton, 25 species of insects and spider mites that attack cotton have developed resistance to organochlorine insecticides, and there is reason to suspect several more species are now resistant. Moreover, at least one resistant species occurs locally in most cotton producing States from California to North Carolina. In ad- dition, four species of spider mites, the fall armyworm (Spodoptera frugiperda (J. E. Smith)), the bandedwing whitefly (Trialeurodes abut ilonea (Haldeman)), the bollworm, and the tobacco budworm (Heliothis virescens (F.)) are known to be resistant to organophos- phorus compounds (Anonymous 1980).

INSECTICIDES FOR CONTROL OF COTTON INSECTS 247
Development of resistance to both the organochlorine and or- ganophosphorus insecticides in the tobacco budworm and bollworm was an especially serious matter since very few effective insecticides were available (Lukefahr 1970). At first, the then recently devel- oped insecticides methomyl and chlordimeform were of some help in control of these resistant pests. However, methomyl proved to be phytotoxic and cannot be used in a regular schedule of applica- tions. Also, chlordimeform, which came into widespread use as an ovicide applied in combination with larvicides, was withdrawn from the market in 1976. (it became available again in 1978 on a limit- ed trial basis but only as a restricted closed system aerial appli- cation material, which eliminates exposure of loaders and pilots. It has been available under this restriction since that time.) Then the synthetic pyrethroids, permethrin and fenvalerate, were developed. These eased the problem of bollworm tobacco/budworm re- sistance (Hopkins et al. 1977), and the materials also suppress moderate infestations of boll weevils. Still other pyrethroids are presently being evaluated, but most researchers feel that the Heli- othis spp. complex may well develop resistance to these materials depending on the intensity of their use. A new organophosphorus compound, sulprofos, is available, but it is not effective against boll weevils.
REPRODUCTION-DIAPAUSE CONTROL
Cotton entomologists have long been aware that naturally oc- curring predators and parasites can sometimes keep infestations of the bollworm complex at subeconomic levels. They are also aware that insecticides applied for the control of pest insects such as the boll weevil, the bollworm complex, cotton fleahoppers, and plant bugs kill these beneficial insects. The difficulty is that in-season application of insecticides is necessary to protect the crop until it matures and is no longer attractive (Parencia 1978). The problem was brought into focus when the bollworm complex became increasingly difficult to control because it had developed resis- tance to previously available effective insecticides.
Because of this, in the late 1940*s, Ewing and Parencia (1949a, 1949b, and 1950) developed a system of providing early- season control of cotton insects that would meet the peculiar needs of the Texas Blacklands (Lincoln and Parencia 1977). Community- wide participation was stressed. Emphasis was placed on control of thrips and cotton fleahoppers, pests that delay early-season growth and fruiting, and on reduction of the population of overwintered boll weevils. In addition, applications of insecticides were ter- minated at least 30 days before the date when bollworms normally became a problem so as to allow populations of beneficial arthro- pods time to recover and contribute to the control of this pest. This was achieved by scheduling applications for entire communities encompassing several thousands of acres of cotton, thereby over- coming the deficiencies of the conventional scouting system in

248 C. R. PARENCIA, JR., ET AL
which one scout could adequately cover less than 2000 acres of cot- ton (Lincoln and Parencia 1977). Late-season treatments for con- trol of boll weevils and the bollworm complex often were not need- ed. The system worked well in the Texas Blacklands, but in the Midsouth and Southeast, it was not as effective where better hiber- nation sites were available to the boll weevils. In other areas, overwintered boll weevils emerged later, and the infestation of the first generation of the bollworm complex was greater than in Texas. Thus, there was often need for additional crop protection when the scheduled treatment period was over (communication with T. R. Pfrimmer and A. R. Hopkins).
Later, Brazzel and Newsom (1959) found that boll weevils must enter a physiological condition called diapause if they were to survive adverse environmental periods. (Diapause in the boll wee- vil is characterized by cessation of gametogenesis and atrophy of gonads, increase in fat content, decrease in water content, and de- crease in respiratory rate.) Brazzel and Hightower (1960) reported that in central Texas weevils with the diapause condition were first observed from late July through August in populations from selected fields. Lloyd and Merkl (1961) reported that in some fields in Mississippi such weevils were first observed as early as August 11; and in other fields, they were first observed as late as September 8. Subsequent studies by Lloyd et al. (1964) showed ini- tial entry of a segment of the weevil population into diapause was coincident with cessation of flowering by cotton plants in individ- ual fields, but in laboratory studies, Lloyd et al. (1967) associ- ated initiation of diapause with boll feeding by adult weevils, limited numbers of squares available to adult weevils, low night temperatures, exposure of immature stages to a short photophase, and larval feeding on bolls. In the field, these workers observed two periods of entry of the boll weevil into diapause. The first occurred in August or September (under Mississippi conditions) when fields ceased flowering and only bolls remained for food. The sec- ond period occurred during October when night temperatures de- creased and only squares were available for food.
During 1959, a large-scale boll weevil diapause control exper- iment (Brazzel et al. 1961) was conducted in the Big Bend area of Texas to determine whether weevils could be killed before they at- tained firm diapause. Four applications of methyl parathion were made at a rate of 0.5 pound per acre from September 30 to November 8 on a 525-acre isolated cotton planting. As a result, during the next crop season, only one of the ten treated fields had popula- tions of boll weevils that reached damaging levels before insecti- cide treatments were required for bollworm-budworm control. How- ever, when a similar test was made in Mississippi (Lloyd et al. 1964), insecticide treatments spaced a week or more apart in Sep- tember permitted sufficient deposition of boll weevil eggs so there were sizable boll weevi1 ,populations in mid- to late October when cooler temperatures reduced the effectiveness of insecticide treatments. Nevertheless, fall populations of boll weevils were significantly reduced.

INSECTICIDES FOR CONTROL OF COTTON INSECTS 249
Knipling (1963) suggested improved timing of insecticide treatments against the last reproducing generation (treatments spaced close together) so as to limit egg deposition. Harris et al. (1966) determined that boll weevil eggs laid after ca. Septem- ber 25 at Mississippi State, MS, would not develop into adult wee- vils before the first killing frost. Tingle and Lloyd (1969) ob- served that newly emerged adult weevils had to feed 2 to 4 weeks to attain firm diapause at temperatures simulating those of late Octo- ber. Lloyd et al. (1966), in field tests conducted in 1963-64, applied six insecticide treatments at 5-day intervals against the last reproducing generation of boll weevils during September and one insecticide treatment in October against weevils that had sur- vived or emerged following the September treatments. The results of this so-called "reproduction-diapause" program was to severely limit reproduction by the last reproducing generation of weevils in the late summer and to kill surviving and late emerging weevils from late September until frost if treatments were applied at 10- to 14-day intervals during this period. Adkisson et al. (1966) and Fye (1968) subsequently demonstrated that in Texas boll weevil pop- ulations could be suppressed 98% or more in the fall by applying ca. seven reproduction-diapause control treatments.
Reproduction-diapause control was used in 1972 in the Pilot Boll Weevil Eradication Experiment (13 insecticide treatments) in southern Mississippi. (This number was required because of the long growing season and the large boll weevil populations in that area.) Pheromone trap catches in the spring of 1973 indicated that few boll weevils survived (99% or better suppression) in the eradi- cation and first buffer areas (Parencia 1978).
Also, reproduction-diapause control applications were made in Year 1 (1978) of the Boll Weevil Eradication Trial (BWET) in both the evaluation and buffer areas. In Years 2 (1979) and 3 (1980), none were applied in the evaluation area. However, in Year 2, sterile boll weevils were released on all the evaluation acreage, and diflubenzuron was applied on 11% of this acreage. The evalua- tion area was judged free of native boll weevils at the end of the Trial (Cross et al. 1981). Likewise, in the Optimum Pest Manage- ment Trial (OPMT), four applications of methyl parathion were scheduled each year for control of diapausing boll weevils. The first application was applied 10 days after the grower made his last in-season application. The second was applied 10 days later and the last two at 15-day intervals. Trap captures in 1979 and 1980 in Panola County where this Trial was conducted were 74 and 94%, respectively, below captures in the control area (Pontotoc County) where diapause control was not used (Anonymous 1981).
In discussing the suppression of diapausing boll weevils as a pest management component, Rummel and Frisbie (1978) state that "The high level of organization and cooperation required is the greatest disadvantage in application of this control technique. However, it has been demonstrated in several areas that the neces- sary organization and cooperation can be achieved and an effective

250 C. R. PARENCIA, JR., ET AL
management program accomplished." The consensus is that, espe- cially in the heavily infested areas of the Cotton Belt, regulatory procedures will have to be adopted by the producers for a reproduc- tion-diapause control program to be effective.
AN APPRAISAL OF COTTON INSECT CONTROL WITH INSECTICIDES
Knipling (1979) discusses the merits and limitations of chemi- cal insecticides in suppressing insect populations. The merits are: (1) chemical insecticides are effective in controlling thou- sands of species that affect plants, animals, man, structures, and stored products; (2) insect control by chemicals is positive and rapid; (3) a single insecticide or a combination of two or more in- secticides can often be used to control several pest species on a given crop at the same time; (4) an individual property owner can take unilateral action as needed to protect his interests when pest problems arise; (5) suitable materials, formulations, and equipment are readily available for immediate use; and (6) one or more of the chemical insecticides already available can be evaluated quickly for use against new pest species.
He lists the following limitations: (1) broad-spectrum action of many insecticides creates hazards or risks of hazards to man and to nontarget organisms in the environment; (2) broad-spectrum in- secticides decimate the populations of natural biological agents that are so important in maintaining a balance between beneficial and harmful or potentially harmful pest species; (3) use of insec- ticides to suppress pests only when and where economic thresholds are reached does not provide any permanent solution to the pest problem and may still permit substantial losses; (4) continued de- pendence on insecticides for control, that is, year after year, is costly on a long-term basis; and (5) the ability of insects to de- velop insecticide-resistant strains limits the period a given mate- rial can be used.
It is therefore interesting to consider the actual impact of insecticides on cotton yield over the years. Three Agricultural Research Service laboratories—Florence, SC, in the Southeast; Tal- lulah, LA, in the South; and Waco, TX, in the Southwest—kept an- nual records of yields from treated and untreated plots for periods of 31, 37, and 32 years, respectively (Parencia 1978). The rec- ords, though they reflect crop losses to insects in these areas when insecticides were not applied, must be accepted for what they are. For example, the use of the "best" insecticides does not pre- vent all insect damage. Also, a certain amount of damage can be tolerated before use of an insecticide is economically reasonable because cotton plants can suffer some fruit loss or foliage damage before yield is reduced. Finally, more than one insect was usually monitored in the various tests, and no attempt was made to show how much of the loss in yield was attributable to a particular insect. Insects involved at all locations were boll weevils, bollworms, cotton aphids, and spider mites. In addition, lygus bugs were

INSECTICIDES FOR CONTROL OF COTTON INSECTS 251
involved at Tallulah and the cotton fleahopper, tobacco budworm, and thrips were involved at Waco.
Despite these limitations (and the obvious fact that insect infestations and weather conditions vary from year to year), rec- ords of this type made over a period of years are more meaningful than records for selected years. It is thus important that at Florence the average increase in yield in treated over untreated plots for the 31-year period 1928-58, was 40.6% (5.3% in 1942 to 217.9% in 1950); at Tallulah, for the 37-year period 1920-56, it was 31.2% (1.1% in 1924 to 112.8% in 1950); and at Waco, for the 32-year period 1939-70, it was 54.3% (9.9% in 1939 to 426.0% in 1961).
Obviously there has been considerable improvement in cotton insect control at the three locations since 1945 and the advent of the organochlorine, organophosphorus, and carbamate insecticides and the phasing out of inorganic insecticides. Table 3 summarizes improvement in insect control since the new insecticides have been used.
Table 3.—Increase in cotton yield over time
Study period
% increase in yield in treated over untreated plots
Location Average Before 1945 After 1945
Tallulah, LA Florence, SC Waco, TX
1920-56 1928-58 1939-70
31.2 40.6 54.3
26.4 23.6 34.0
41.3 53.9 77.6
Insecticides and miticides now used for control of cotton in- sects and spider mites are listed in Tables 4 and 5, respectively.

252 C. R. PARENCIA, JR., ETAL
Table 4.—Recommended dosages (pounds per acre technical of major Insecticides Boll Bollworm or Cotton Cotton
(pounds) weevil tobacco budworm aphid fleahopper
Acephate Ij Aldicarb 2/ 0. Azinphosmethyl 4^/0.1 Bacillus
0.75-1.0 6-1.0 2-0.5
0.3-0.5 0.6-1.0 0.25 1.0-0.25
thuringiensis 0.5-1.0 Baculovirus heliothis 5/ 0.12-0.25
Carbaryl 1.0-2.0 0. .25-1.4 Carbophenothion Chlor dime form jô/ 0.12-0.5
0.5-1.0
Chlorpyrifos Demeton
0.5-1.0 0.25-0.38
Dicrotophos 0.1-0.25 0. .1-0.25 Dimethoate 0.1-0.25 0. .1-0.25 Disulfoton 7/ 0.6-1.0 Endosulfan EPN 0.25-0.5 0.75-1.25 EPN + methyl
parathion 8/ 0.25+0.25 0.5-1.0+0.5-1, .0 Ethion 0.5 Fenvalerate 0.1-0.2 Malathion kj 0.5-1.0 0.5-1.25 0, .7-1.25 Methamidophos 0.5-1.0 Methomyl Methyl
parathion 4/ 0.25-0.5
0.45-0.67
1.0-2.0 0.25-0.5 0, .12-0.5 Monocrotophos 9^/ 0.6-1.0 0.6-1.0 Oxydeme t onme thy1 Parathion 0.25-1.0 1.0
0.25-0.38 0.2-0.5
Permethrin 0.1-0.2 0.1-0.2 Phenamiphos 3/ Phorate Ij 0.5-1.5 Sulprofos 0.75-1.0 Toxaphene Toxaphene + methyl
parathion 8/
2.0-4.0
1.0-2.0+
2.0-4.0
2.0-3.0+1.0-1 .5 0. .5-2.0 0.25-0.5
Trichlorfon 0. .25-1.0
J_/ For control of insects of lesser importance see Anonymous pound per hundredweight of cottonseed for thrips control. _3/ malathion, and methyl parathion may be applied ultra-low volume 0.5-0.75 pound per acre, respectively. _5/ 36-72 larval equiva- system. Jj In-furrow granule treatment or as seed treatment at and 1.3-1.5 pounds per hundredweight for phorate. 8^/ Chlor- for bollworm/tobacco budworm control. 9/ Monocrotophos may be of cottonseed for thrips control.

INSECTICIDES FOR CONTROL OF COTTON INSECTS 253
material) of the principal insecticides used for the control cotton insects 1/
Cotton Cotton leaf- Lygus bugs and Pink leafworm perforator other mirids bollworm Thrips
1.0 0.1-0.5 2.0 0.6-1.0 0.3-0.5
0.25-0.38 0.125-0.25 0.5-1.0 0.08-0.2
0.5-2.0 1.0-2.0 1.5-2.5 0.5-0.8
0.5
0.1-0.2 0.1-0.25 0.1-0.25 0.1-0.25
0.6-1.0 0.3-0.75
0.25+0.25-0.25+0.25
0.1 0.4-1.25 0.7-1.2 0.25-1.3
0.45-0.67
0.12-0.25
1.0
0.25-0.5 0.12-0.5 0.12-0.5 0.1-0.25 0.6-1.0 0.1-0.2
0.1-0.2 1.6-3.3 0.5-1.5
2.0-4.0 1.0-4.0 1.0-2.0
0.5-1.5
(1980). Ij Acephate may be applied as seed treatment at 0.5 In-furrow granule treatment at planting, kj Azinphosmethyl, as technical material at 0.125-0.25 pound, 0.5-1.2 pounds, and lents per acre. 6^/ Restricted to aerial application in a closed 0.25-0.5 pound per hundredweight of cottonseed for disulfoton dime form, methomyl, or chlorpyrifos may be added to the mixture applied as seed treatment at 0.25-1.25 pounds per hundredweight

254 C. R. PARENCIA, JR., ET AL
Table 5.—Dosages (pounds per acre technical material) of miti-
Mítícíde Carmine Desert McGregor Pacific
Aldicarb J^/ 0.6-1.0 0.6-1.0 0.6-1.0
Carbophenothion 0.25-0.75
Chlorpyrifos 0.25-1.0
Demeton 0.38 0.38 0.38 0.38
Dicofol 1.0-1.5 0.8-1.6 0.8-1.0
Dicrotophos 0.25-0.5 0.25-0.5 0.25-0.5
Disulfoton Ij 0.5-1.0 0.5-1.0
Ethion 0.25-1.5 0.25-0.75 0.25-1.0
Methidathion 1.0 1.0 1.0 1.0
Methyl parathion 0.6
Monocrotophos 0.5-1.0 0.5-1.0 0.5-1.0 0.5-1.0
Oxydemetonmethyl 0.25-0.5 0.25-0.5 0.25-0.5 0.25-0.5
Parathion 0.25-0.5 0.1-0.2
Phorate _3/ 0.5-1.5 0.5-1.5
Propargite 1.25 0.8-1.6
Sulfur 25-35
\J In-furrow granule treatment at planting. Ij In-furrow granule treatment at planting or 0.5 pound per _3/ In-furrow granule treatment at planting or 1.3-1.5

INSECTICIDES FOR CONTROL OF COTTON INSECTS 255
cides for the control of species of spider mites (Tetranychus)
T. ludeni Schoene Strawberry Tumid Two s pot ted Zacher
0.6-1.0 0.6-1.0
0.25-0.5 0.375
0.25-1.0
0.25-0.75
0.25-1.0
0.38 0.38
0.8-1.6
0.25-0.5
0.5-1.0
0. .38 0.38
0.8-1.6
0.25-0.5
0.5-1.0
0.38
0.25-1.0 0.25-1.0 0.25-1.5
1.0 1.0 0.5-1.0
0.6 0. .6 0.6
0.5-1.0 0.5-1.0 0. .5-1 -.0 0.5-1.0 0.5-1.0
0.25-0.5 0.25-0.5 0. ,25- ■0.5 - 0.25-0.5
0. .25- -0.5 0.125-1.0 0.2
0.5-1.5 0.5-1.5
0.8-1.6 0.8-1.6
25-35 20-50
hundredweight of planting seed. pounds per hundredweight of planting seed.

256 C. R. PARENCIA, JR., ET AL
INSECTICIDES IN AN INSECT PEST MANAGEMENT SYSTEM
Insecticides used for control of cotton insects beltwide in recent years will be discussed in other chapters of this handbook. However, methyl parathion and azinphosmethyl were chosen for con- trol of diapausing boll weevils in the OPMT and BWET, respectively. This choice was made, and the schedule of applications was drawn up (see Table 6), by the participating producers in areas that were in- volved in the OPMT and, after 1978, in the BWET. They were in ac- cordance with the choices and the schedules suggested in the Cotton Insect Control Guides for the respective States.
Table 6.—Insecticide use during the growing season in Optimum Pest Management Trial (OPMT) and Current Insect Control (CIC) areas, Boll Weevil Eradication Trial (BWET), and buffer areas 1977-80
Average no. insecticide applications for— Bollworm/
Lygus tobacco Boll ^^^^ Thrips bugs budworm weevil
OPMT 0.2 3.9 0 0.3 1.9 0 0.6 1.6 0 0.5 2.4 0.1
CIC 0 1.6 0 0 1.9 0 0 0.6 0 0 0.3 0.6
BWE 0 7.7 0.4 0 3.7 6.2 0 2.1 0.4 0 1.2 0
Buffer 0 0 0 0 11.3 0.7 0 7.8 5.1 0 4.4 3.9
1977 1/ 1978 1979 2/ 1980
1977 1978
i/
1979 2/ 1980
1977 1/ 1978 1979 1980
1977 1/ 1978 1979 1980
1, .5 0. .4 0. .6 0, .5
0. .3 0, .1 0. ,1 0. ,03
0 0 0 0
0 0 0 0
j_/ Prototype operation before Trials began. l_l Applications for boll weevil included in those for bollworm/
tobacco budworm.

INSECTICIDES FOR CONTROL OF COTTON INSECTS 257
Nevertheless, total reliance on insecticides is no longer ac- ceptable (and was not the pattern in these Trials) because of ad- verse effects of insecticides on the environment and on nontarget species. At the same time, the development of alternatives, though it is receiving much emphasis, is proving difficult and costly. Insecticides will therefore continue to be important in the control of cotton insects. This is why there is concern because compara- tively few promising candidate materials have recently been made available for testing.
In fact, the cost of developing and registering a compound also is expensive and continues to increase. Industry is thus tak- ing a hard look at compounds after initial screening to see whether their continued development is justified. As a result, few new compounds are being made available for testing programs, and only a small number of these are carried on to registration and ultimate use by growers. The resulting lack of new effective compounds is especially serious in regard to control of the Heliothis spp. com- plex that has developed resistance to the comparatively few availa- ble effective insecticides. Possibly some of the alternative tactics for insect control can be used in conjunction with insecti- cides in insect pest management systems that will reduce the crises produced because so few effective insecticides are available. Also, management systems that incorporate nonchemical tactics and strategies may reduce the number of insecticide applications needed to achieve control of cotton insects.

258 C. R. PARENCIA, JR., ET AL
LITERATURE CITED
Adkisson, P. L., D. R. Rummel, W. L. Sterling, and W. L. Owen, Jr. 1966. Diapause boll weevil control: a comparison of two meth-
ods. Texas Agricultural Experiment Station B-1054, lip.
Anonymous. 1954. Losses in agriculture. U.S. Department of Agriculture, ARS-20-1, 190 p.
1965. Losses in agriculture. U.S. Department of Agriculture, Agriculture Handbook No. 291, 120 p.
1980. 33rd annual conference report on cotton insect research and control. U.S. Department of Agriculture, Science and Edu- cation Administration, 77 p.
1981. Final Report, Optimum Pest Management Trial, Panola Coun- ty, MS. Cooperative Extension Service, Mississippi State Uni- versity, 35 p.
Brazzel, J. R., Jr., and L. D. Newsom. 1959. Diapause in Anthonomus grandis Boheman. Journal of Eco- nomic Entomology 52:603-611.
and B. G. Hightower. 1960. A seasonal study of diapause, reproductive activity, and
seasonal tolerance to insecticides in the boll weevil. Journal of Economic Entomology 53:41-46.
T. B. Davich, and L. D. Harris. 1961. A new approach to boll weevil control. Journal of Econom-
ic Entomology 54:723-730.
Cleveland, T. C., W. P. Scott, T. B. Davich, and C. R. Parencia, Jr.
1966. Control of the boll weevil on cotton with ultra-low volume (undiluted) technical malathion. Journal of Economic Entomol- ogy 59:973-976.
Coad, B. R. 1918. Recent experimental work on poisoning cotton boll weevils.
U.S. Department of Agriculture, Bulletin No. 731, 15 p.
Comstock, J. H. 1879. Report on cotton insects. U.S. Department of Agriculture,
315 p.

INSECTICIDES FOR CONTROL OF COTTON INSECTS 259
Cross, W. H., et al. 1981. Final Report, Biological Evaluation, Beltwide Boll Weevil
Cotton Insect Management Programs. Attachment C.
DeBord, D. V. 1977. Cotton insect and weed loss analysis. The Cotton Founda-
tion, 121 p. Memphis, TN.
Eichers, T. R., P. A. Andrilenas, and T. W. Anderson. 1978. Farmer's use of pesticides in 1976. U.S. Department of Agriculture, Agricultural Economic Report No. 418, 58 p.
Ewing, K. P., and C. R. Parencia, Jr. 1948. Control of boll weevil and cotton aphid with dusts con-
taining chlorinated camphene, benzene hexachloride or other new insecticides. Journal of Economic Entomology 41:558-563.
and C. R. Parencia, Jr. 1949a. Experiments in early-season application of insecticides
for cotton insect control in Wharton County, Texas during 1948. U.S. Department of Agriculture, Bureau of Entomology and Plant Quarantine E-772, 6 p.
and C. R. Parencia, Jr. 1949b. Early-season applications of insecticides for cotton in-
sect control. U.S. Department of Agriculture, Bureau of Ento- mology and Plant Quarantine E-792, 9 p.
and C. R. Parencia, Jr. 1950. Early-season applications of insecticides on a community- wide basis for cotton insect control in 1950. U.S. Department of Agriculture, Bureau of Entomology and Plant Quarantine E- 810, 8 p.
Fye, R. E. , C. L. Cole, F. C. Tingle, A. Stoner, D. F. Martin, and L. F. Curl.
1968. A reproductive-diapause control program for boll weevil in the Presidio, Texas-Ojinaga, Chihuahua Area, 1965-67. Journal of Economic Entomology 61:1660-1666.
Glover, T. 1854. The bollworm. Patent Office, Agricultural Report, 64 p.
1855. The bollworm. Patent Office, Agricultural Report, 99 p.
Harris, F. A., E. P. Lloyd, and D. N. Baker. 1966. Effects of the fall environment on the boll weevil in
northeast Mississippi. Journal of Economic Entomology 59: 1327-1330.

260 C. R. PARENCIA, JR., ET AL
Hopkins, A. R., and H. M. Taft. 1965. Control of certain cotton pests with a new systemic insec-
ticide, UC-21149. Journal of Economic Entomology 58:746-749.
H. M. Taft, and W. James. 1977. Tobacco budworm, bollworm, and boll weevil: effectiveness
of newly developed experimental insecticides on cotton in the southeast. Journal of Economic Entomology 70:723-726.
Knipling, E. F. 1963. An appraisal of the relative merits of insecticidal con-
trol directed at reproducing and diapausing boll weevils in efforts to develop eradication procedures. Entomology Research Division, ARS, USDA. (Mimeographed, 22 p.)
1979. The basic principles of insect population suppression and management. U.S. Department of Agriculture, Agriculture Hand- book No. 512, p. 243-280.
Lincoln, C, and C. R. Parencia, Jr. 1977. Insect pest management in perspective. Bulletin of Ento- mological Society of America 23:9-14.
Lloyd, E. P., and M. E. Merkl. 1961. Seasonal occurrence of diapause in the boll weevil in Mis-
sissippi. Journal of Economic Entomology 54:1214-1218.
M. L. Laster, and M. E. Merkl. 1964. A field study of diapause, diapause control, and popula-
tion dynamics of the boll weevil. Journal of Economic Ento- mology 57:433-438.
F. C. Tingle, J. R. McCoy, and T. B. Davich. 1966. The reproduction-diapause approach to population control
of the boll weevil. Journal of Economic Entomology 59:813-816.
F. C. Tingle, and R. T. Gast. 1967. Environmental stimuli inducing diapause in the boll wee-
vil. Journal of Economic Entomology 60:99-102.
Lukefahr, M. J. 1970. The tobacco budworm situation in the Lower Rio Grande Val-
ley and Northern Mexico. In Proceedings, Annual Texas Confer- ence on Insects, Plant Diseases, Weed and Brush Control, Texas A&M University, p. 140-145.
National Cotton Council. 1973. Boll weevil losses: value and location of losses caused
by the boll weevil. National Cotton Council Report, 52 p.

INSECTICIDES FOR CONTROL OF COTTON INSECTS 261
Parencia, C. R., Jr. 1978. One hundred twenty years of research on cotton insects in
the United States. U.S. Department of Agriculture, Agriculture Handbook No. 515, 75 p.
J. W. Davis, and C. B. Cowan, Jr. 1957. Control of early-season cotton insects with insecticides
employed as seed treatments. Journal of Economic Entomology 50:31-36.
Riley, C. V. 1885. Fourth Report of the U.S. Entomological Commission, being
a revised edition of Bulletin No. 3 and the final report on the cotton worm together with a chapter on the bollworm, 399 p.
Roussel, J. S., R. V. Bielarski, and D. F. Clower. 1957. The status of chlorinated hydrocarbon resistance in the boll weevil in Louisiana. Louisiana Agricultural Experiment Station, Department of Entomology Special Release No. 1.
Rummel, D. R., and R. E. Frisbie. 1978. Suppression of potentially overwintering boll weevils as a
pest management practice. The boll weevil: management strat- egies. Southern Cooperative Series Bulletin No. 228, p. 39-49.
Tingle, F. C, and E. P. Lloyd. 1969. Influence of temperature and diet on the attainment of
firm diapause in the boll weevil. Journal of Economic Entomol- ogy 62:596-599.
Townsend, C. H. T. 1895. Mexican cotton-boll weevil in Texas. Insect Life 7:295-
309.


SECTION III SUPPORT COMPONENTS


265
Chapter 11
MASS REARING BOLL WEEVILS
J. G. Griffin Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
P. P. Sikorowski Department of Entomology Mississippi State University Mississippi State, MS 39762
0. H. Lindig Boll Weevil Research Laboratory Agricultural Research Service U.S. Department of Agriculture Mississippi State, MS 39762
ABSTRACT Because large numbers of sterile boll weevils (Anthonomus grandis Boheman) were to be used as a component of weevil eradication programs, rearing and equipment development re- search was conducted for all phases of weevil production in a laboratory. This resulted in new and improved weevil rearing technology and adoption of certain sanitary practices. Since most of the production operations were mecha- nized, the manual labor needed for rearing was reduced by 85% or more. Also, mechanization meant more aseptic operation. In addition, sanitation practices were developed that prac- tically eliminated the microbial contamination that was so harmful to weevil rearing. The newly developed rearing technology was tested in mass production trials to evaluate its capa- bility of rearing large numbers (5-6 million per week) of high quality weevils. The first test disclosed that minor changes were needed. After these changes were made, a second test and a production trial proved that high quality weevils can be reared economically.

^^^ J. G. GRIFFIN ET AL
INTRODUCTION
The first of the developments that eventually made it possi- ble to mass rear boll weevils (Anthonomus grandis Boheman) in a laboratory was the formulation of a suitable artificial diet (Van- derzant and Davich 1958). This basic diet has since been modified CEarle et al. 1959, Vanderzant and Davich 1961, Gast and Davich 1966, Lmdig and Malone 1973), most recently by Lindig et al (1979), to make it better adapted for the insect, less expen¡ive, and more compatible with the equipment and procedures used for mass rearing.
The second of the developments was the progress made in de- signing mechanical equipment that could be used to rear larger numbers of weevils at lower cost (Gast 1961, 1965; Gast and Vardell iVbj;. Initially, use of the new equipment somewhat decreased the percentage of adults obtained from a given number of eggs, though
^QIA^^"^^^^ ^^^^ ^^^ weevil decreased much more (Gast and Davich 1966). Finally, by the midseventies to late seventies, the pro- duction of weevils was no longer limited by labor-intensive techniques.
The third development was the beginning of progress in con- trolling the diseases and microbial contamination encountered dur- ing the first efforts to accomplish laboratory rearing of weevils (Gast 1966).
As this progress was being made, the need arose to produce millions of sterile weevils per week for weevil eradication pro- grams. Improved or new mass rearing technology would be required if these insects were to be mass reared economically and in the desired quantities. Therefore, a new research and production facility was constructed, and efforts were increased to achieve these goals.
FACILITY
The^new structure is known as the Robert T. Gast Rearing Lab- oratory (commonly the Gast rearing facility) and is located on the campus of Mississippi State University. It was designed to accom- modate the rearing technology available at that time and was com- pleted and equipped by June 1972. McLaughlin (1970), Griffin et al. (1971), and Griffin and Lindig (1978) have described the lab- oratory in detail.
The facility was used during the summers of 1972 and 1973 to produce and sexually sterilize the weevils used in the sterile male release component of the Pilot Boll Weevil Eradication Experiment in south Mississippi and adjoining areas of Alabama and Louisiana. Production during these years was unsatisfactory because of prob- lems with contamination and because of the large amount of labor required. However, the information gained was valuable because it

MASS REARING BOLL WEEVILS 267
indicated that rearing procedures and equipment would have to be improved to achieve a successful mass rearing operation.
During the next few years, major efforts were made to design adequate rearing equipment and procedures. Thus by 1976, semiauto- matic or automatic equipment had been developed to handle many of the rearing chores that had previously been handled manually, which meant that sanitary conditions were easier to achieve. In addi- tion, labor requirements were reduced tremendously without reducing the percentage yield of weevils.
Therefore, in the summer of 1976, a 6-week mass trial was con- ducted at the Gast rearing facility to determine the production capability of the newly developed and modified rearing equipment and procedures. The goal was to produce 5 million weevils per week, and it was achieved, but the need for further changes in the operation became apparent. As a result, additional changes were made as follows: (1) High-Efficiency Particulate Air (HEPA) fil- ters were installed over outlets of the air conditioning supply in the egg planting and larval development rooms; (2) adult weevils were moved into rooms that were farther away from the egg planting and larval development areas; (3) the larval development area was expanded from one room to two adjoining rooms so there could be more space and thus better air circulation around the carts that held the trays of diet and the eggs and larval stages; (4) a cabi- net was installed for fumigating the plastic and Tyvek used in con- structing rearing trays; (5) antimicrobial agents were added to the sand used to cover the diet and eggs in the rearing trays (Sikorow- ski et al. 1980); and (6) the storage and processing operations re- lating to the diet were moved to a separate building to reduce the flow of microbes produced by these operations into the rearing areas. Most of these changes were made to provide better sanita- tion within the rearing facility because this had proved to be critical to control of microbial contamination during production.
Another 6-week production trial was conducted during the sum- mer of 1977. Once again the goal of 5 million weevils per week was attained, and this time there were fewer problems and less contami- nation. However, the need for other modifications was apparent so the following changes were made: (1) a rackveyor was developed (Griffin 1979b) for stacking, holding, and conveying the rearing trays; it replaced the cart system (Griffin et al. 1979); (2) corn- cob grit was mixed with the sand used to cover the diet and eggs in the rearing trays (S. Malone unpublished data); (3) a chamber was constructed for fumigating the rackveyors; (4) a high temperature (hot) chamber was constructed to kill the weevils remaining in rearing trays that were to be discarded and the adults in the egg producing colony that had become too old; (5) the sugar-starch planting solution (Griffin et al. 1979) was replaced with a solu- tion of Furcelleran (Bentonite 44/A) (S. Malone unpublished data); (6) emergence cubicles were developed and installed to replace the emergence boxes (Griffin and Roberson 1980); (7) the electrically operated mixers used in the egg harvesting operation were replaced

26Ö J. G. GRIFFIN ETAL
with pneumatically operated ones for better control of the mixing speed; (8) "escape" holes for emerging weevils were burned in the rearing trays so it would not be necessary to manually peel off the covering; and (9) new quality control and contamination monitoring operations were added. These changes saved labor, improved sanita- tion, reduced the numbers of live insects discarded to the outside environment, and helped in identifying some of the sources of contamination.
The equipment and techniques developed as a result of the 1976 and 1977 production trials were used to rear the weevils used in the North Carolina-Virginia Boll Weevil Eradication Trial program during the summer of 1979.
Thus by 1979, the Gast rearing facility included the following production areas: (1) diet sterilization, (2) pellet making, (3) oviposition, (4) egg harvesting and cleaning, (5) egg and planting solution sterilization, (6) sand and corncob grit (granular materi- als) preparation, (7) egg planting, (8) larval development (2 ad- joining rooms), (9) emergence, (10) general supply storage, (11) storage for dry ingredients of diet, and (12) dry ingredient pro- cessing areas. However, areas 11 and 12 are located in a separate building away from the mass rearing facility. The oviposition, larval development, and emergence rooms have specially controlled environments; the other areas have environments designed for human comfort.
Nevertheless, during production in 1979, several deficiencies in the facility became apparent. These will need to be improved, if possible, and should be avoided in designing future mass rearing facilities. For example, areas of the seamless floor covering peeled from the concrete base, which makes the floor surface rough and harder to clean and sanitize. Quarry tile of better quality or a better method of applying the seamless covering material is needed. Also, some of the interior partition walls covered with gypsum board have deteriorated as a result of exposure to cleaning and sanitizing materials and procedures and, in some instances, be- cause they were damaged when equipment was moved. Wall materials must be durable and easy to keep clean and must withstand the cleaning and sanitizing materials and application procedures. On the other hand, masonry (cinder) blocks or clay tile painted with epoxy enamel have given good service. Then drains were not pro- vided in the floors, and these are needed for better washing and cleaning of the floors and walls. In addition, although the light fixtures are recessed and have covers, adult weevils still can enter the fixtures. When they die there, they cause an unsanitary condition that can be eliminated only by frequent cleaning of the fixtures. Fixtures with better sealed enclosures that are properly installed should help eliminate this problem.
Moisture condenses on the inside surfaces of the exterior win- dows and walls, especially in the rooms that have special environ- mental controls, when outside temperatures are low. This excess

MASS REARING BOLL WEEVILS 269
moisture creates unsanitary conditions. The fewest and smallest possible insulated window units should be used, and the walls should be insulated to reduce or eliminate this problem. There is 3**-6**C differential in air temperature between the floors and ceil- ings in the rooms with environmental control. Proper type and lo- cation of the outlets that supply and return the conditioned air should eliminate or reduce the temperature differentials. Also, the use of insulation in the ceilings, walls, and floors would be helpful. Control of the environment is a separate problem that is particularly important in the areas where the culture is main- tained; e.g., the development, emergence, and oviposition rooms. Therefore, the equipment must be of the proper type and must be ad- equate and reliable. Backup equipment for the heating, cooling, and air moving operations must be available.
Finally, proper sanitation is the first concern in arranging work areas or rooms in a facility. Work and materials must flow from the cleanest to the less clean areas. Traffic must be re- stricted in all areas, but it must be even more restricted in the clean rooms; e.g., the egg planting, pellet making, and larval de- velopment rooms. (At the Gast facility, access between separate areas is by covered outside walks, but a central corridor would be satisfactory.) Also, dressing and shower accommodations and clean uniforms must be provided for workers entering these clean rooms, and their use must be mandatory.
DIETS
Ingredients and Proportions
Two diet formulations are presently used for mass rearing boll weevils; one is used for rearing larvae and the other is used with adults, both for feeding and as oviposition sites. The dry ingre- dients and proportions for both diets are given in Table 1. Other diet formulations have been tested recently (Lindig et al. 1979), but the ones shown are presently used as the standards. Ingredi- ents are normally obtained in quantities to last through a summer rearing season or a program. If necessary, the materials are stored in cool, dry, and easily accessible quarters that are as re- sistant to rodents and damaging insects as possible. Records are maintained on sources and dates of acquisition, and any changes of material are noted.
Processing
The ingredients of the vitamin mix used in both diets (Table 1) are mixed on-the-job in batches (Table 2) and blended to a homo- genous mixture in a twin shell blender.
Batches of the dry mix for both regular diets are prepared in advance by weighing out the ingredients in the quantities indicated
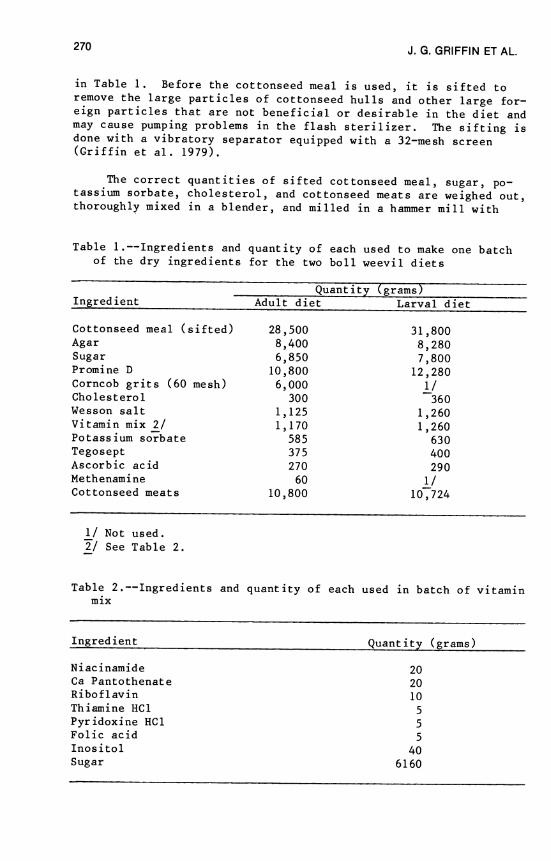
2'^° J. G. GRIFFIN ET AL.
in Table 1. Before the cottonseed meal is used, it is sifted to remove the large particles of cottonseed hulls and other large for- eign particles that are not beneficial or desirable in the diet and may cause pumping problems in the flash sterilizer. The sifting is done with a vibratory separator equipped with a 32-mesh screen (Griffin et al. 1979).
The correct quantities of sifted cottonseed meal, sugar, po- tassium sórbate, cholesterol, and cottonseed meats are weighed out, thoroughly mixed in a blender, and milled in a hammer mill with
Table 1.—Ingredients and quantity of each used to make one batch of the dry ingredients for the two boll weevil diets
Quantity (grams) Ingredient Adult diet Larval diet
Cottonseed meal (sifted) 28,500 31,800 Agar 8,400 8,280 Sugar 6,850 7,800 Promine D 10,800 12,280 Corncob grits (60 mesh) 6,000 1/ Cholesterol 300 360 Wesson salt 1,125 1,260 Vitamin mix 2/ 1,170 1,260 Potassium sórbate 585 630 Tegosept 375 400 Ascorbic acid 270 290 Methenamine 60 1/ Cottonseed meats 10,800 10,724
j_/ Not used. 2/ See Table 2,
Table 2.—Ingredients and quantity of each used in batch of vitamin mix
Ingredient Quantity (grams)
Niacinamide 20 Ca Pantothenate 20 Riboflavin 10 Thiamine HCl 5 Pyridoxine HCl 5 Folie acid 5 Inositol 40 Sugar 6160

MASS REARING BOLL WEEVILS 271
0.6-mm screen openings. (The ingredients milled with the cotton- seed meats absorb the oil from the meats, which would otherwise clog the mill screen.) The milled mixture and the other ingredi- ents are mixed in a blender until homogenous. Then the completed batch is sifted on the separator used to sift the cottonseed meal so as to remove lint and hull particles from the cottonseed meats and any other large particles or foreign material introduced in un- milled ingredients. Finally, the dry mix is placed in containers that are properly labeled and stored in the diet sterilizing room. During a production program, dry mixes are prepared 5 days per week so as to reduce storage time.
Processing of the dry mix produces considerable amounts of dust, which contains and supports microbes. The room used for this operation is therefore ventilated to keep the dust at acceptable levels (for the operator) and to prevent it from accumulating in the storage room(s). Because of the dust and the microbes, the storage and processing rooms are located in a separate building away from the main rearing facility.
Sterilizing
The diets used in mass rearing boll weevils are thermally sterilized by using a flash sterilizer (Cherry-Burell NO-Bac. Uni- therm IV) (Griffin et al. 1974, 1979) (Figure 1) in a separate room of the main rearing facility.
The diet is prepared for sterilizing by weighing out or meas- uring by volume each of the ingredients listed in Table 3. Then
Table 3.—Ingredients and quantity of each used in preparing 45.4 kg (water quantity) of the two boll weevil diets
Quantity Ingredient Larval diet Adult diet
Dry mix, kg (see Table 1) 8.5 8.6 Water, kg B,„ vitamins (0.01%), ml
45.4 45.4 9 12
Biotin (0.01%), ml 9 9 HCl, ml 40 40
these materials are placed in the product tank of the sterilizer and mixed to a homogenous mixture with a propeller-type mixer. The mixture is next pumped through the sterilizer, where it is heated to ca. 141 **C and then held at this temperature for a minimum of 30 seconds. Heated mixture to be used for larval and adult diets is cooled to ca. 40.5**C or 52*C, respectively, and then transferred through stainless-steel tubes to the points of use, the

272 J. G. GRIFFIN ETAL.
pellet machine in the case of the adult diet or the egg planting machine in the case of the larval diet (see sections on pellet mak- ing and egg planting). The sterilizers have heat exchangers made of coiled tubing inside a pressure-tight metal jacket and use steam for heating and regular tap water for cooling. The heating and cooling temperatures are automatically controlled with two propor- tional temperature controllers and motor-operated valves. Each sterilizer is rated at 170 liters per hour capacity, a size compatible with the capacity of the equipment the units are serv- ing. The sterilizer units are cleaned and sanitized after each use (Sikorowski 1975).
Figure 1.—Sterilizer and auxiliary equipment used to sterilize the diets.
EGG PRODUCTION
Source of Weevils
Boll weevils used in the Boll Weevil Eradication Trial were obtained by crossing weevils collected in the fields of North Caro- lina with an ebony strain (Bartlett 1967) that has been maintained at the Boll Weevil Research Laboratory for several years. This

MASS REARING BOLL WEEVILS 273
crossing, and careful selection for ebony characteristics, produces weevils that are easily identified when they are released in the field for eradication or for research work. Calco Red dye can also be incorporated into the larval diet to produce a marker (Gast and Landin 1966, Lindig et al. 1980).
Adults used for egg production, that is, the brood colony, are selected from those collected the first day of emergence from
the rearing trays.
Pellet Making
The pellets of adult diet that serve as both food and oviposi- tion sites for adult weevils in the brood colony are prepared in a pellet-making machine and then placed in a waxing machine where they are coated with a mixture of 40% beeswax and 60% paraffin (Griffin and Lindig 1974) (Figure 2) to prevent or reduce desicca- tion. (Also, the females prefer this cover for oviposit ion.) The procedure is as follows: Sterile diet is pumped from the sterili- zer to the pellet machine through stainless-steel tubes. It then flows through automatically operated control valves into the form- ing tubes, ca. 8-mm i.d., of the heat exchanger. The cold water flowing around these tubes causes the diet to solidify. As the re- sulting gelled rod is discharged from the tubes, a rotating drum with wires spaced around the circumference cuts it into ca.^U-mm sections. These sections fall into the vat of warm, ca. 68*'C, wax of the waxing machine. Then an inclined wire belt lifts the pel- lets out of the wax and discharges them over the edge so they fall into a catching container. As they fall, a blast of air from a fan causes the wax coat to harden. The finished pellets are ca. 9.5 (diameter) x 13 (long) mm. They are stored in sanitized covered
plastic pans until used.
The pellet making should be done in a separate clean-air room with an independent air conditioning system to hold microbial con- tamination of the diet to a minimum. The wax mixture used to coat the pellets is sterilized by boiling in a steam kettle and then held at the desired temperature in a wax warmer tank (Griffin et al. 1979). It is transferred from the tank to the vat as needed.
The equipment and space are cleaned and sanitized each day after operation (Sikorowski 1975).
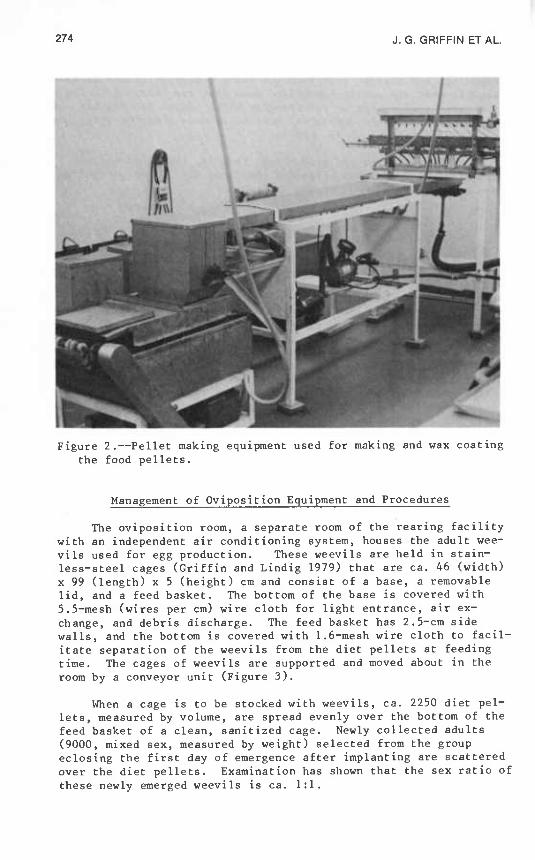
274 J. G. GRIFFIN ETAL
Figure 2.—Pellet making equipment used for making and wax coating the food pellets.
Management of Oviposition Equipment and Procedures
The oviposition room, a separate room of the rearing facility with an independent air conditioning system, houses the adult wee- vils used for egg production. These weevils are held in stain- less-steel cages (Griffin and Lindig 1979) that are ca. 46 (width) X 99 (length) x 5 (height) cm and consist of a base, a removable lid, and a feed basket. The bottom of the base is covered with 5.5-mesh (wires per cm) wire cloth for light entrance, air ex- change, and debris discharge. The feed basket has 2.5-cm side walls, and the bottom is covered with 1.6-mesh wire cloth to facil- itate separation of the weevils from the diet pellets at feeding time. The cages of weevils are supported and moved about in the room by a conveyor unit (Figure 3).
When a cage is to be stocked with weevils, ca. 2250 diet pel- lets, measured by volume, are spread evenly over the bottom of the feed basket of a clean, sanitized cage. Newly collected adults (9000, mixed sex, measured by weight) selected from the group eclosing the first day of emergence after implanting are scattered over the diet pellets. Examination has shown that the sex ratio of these newly emerged weevils is ca. 1:1.

MASS REARING BOLL WEEVILS 275
Figure 3.—Oviposition cage, shaker, and conveyor for mass rearing boll weevils.
Once each day, the conveyor is used to move the oviposition cages to the feeding table. There they are lifted from the con- veyor, and old pellets are removed and replaced with fresh pellets. Then the conveyor moves the cages away from the feeding table as needed. Fresh pellets are provided at a rate of ca. four weevils per pellet.
The old pellets are later carried to the egg harvesting area for the harvesting operation.
Also, each day, new cages of young adults, the number needed to maintain the colony at the desired level, are prepared and placed in the oviposition room. Likewise, each day, cages contain- ing 14-day-old weevils (weevils are kept in the brood colony for egg production for 14 days) are removed and destroyed by placing them in a paper bag and holding them at ca. 66°C for over 20 hours.
Environment
The oviposition room is maintained at 31°±1°C and 50%±5% rela- tive humidity and in total darkness except during feeding and cleanup, usually a matter of 3-4 hours per day. Uniform air move- ment, ca. 30 m per minute, around the cages is necessary to main- tain the pellets at a moisture level that will give maximum egg

276 J. G. GRIFFIN ET AL.
production. Air velocity greater than ca. 50 m per minute around the cages will dry out the pellets, thereby reducing egg produc- tion.
EGG HARVEST AND PREPARATION
Harvest
After the females deposit their eggs at various depths in the diet pellets (beneath the wax coat), the eggs are harvested from the pellets mechanically (Griffin and Lindig 1977, Griffin et al. 1979) as follows:
Pellets collected from the oviposition cages during the adult feeding operation are dumped into the hopper of an electrically operated lift, transferred into a vibratory feeder hopper ((1) in Figure 4), and then sprayed with tap water as they are fed into wax crimping rollers (2). The rollers crack the wax coat, and the
Figure 4.—Equipment for harvesting the eggs from pellets.

MASS REARING BOLL WEEVILS 277
water, diet particles, and wax coating are discharged from the rollers into one compartment of a two-compartment separation tank (3) where the mixture is agitated by an air-operated, propeller- type mixer. When one compartment is filled, the flow of water and broken pellets is directed into the other compartment, and the mix- er in the first compartment is stopped. Wax particles then rise, and diet particles and eggs sink. The wax is skimmed off, washed, drained, placed in a steam kettle, and boiled to remove the remain- ing moisture and to sanitize it for re-use. The diet-eggs-water mixture is pumped (4) to disintegrators (choppers) (5) where the diet particles are further reduced in size so the eggs can float free. Then the mixture is discharged into the liquid-solid sepa- rator (6). One deck of the separator (top) consists of 10-mesh screening; the other (bottom) of 19-mesh screening. The larger particles of diet from the mixture remain on the top deck, and the eggs and smaller particles are deposited on the lower deck and then discharged into a basket covered with 20-mesh hardware cloth. The water and large particles of diet are deposited into and flushed down a trough into the waste disposal system.
Cleaning
The initial separation of the eggs from fine diet particles is done in saturated sodium chloride solution (brine) and a 57-liter separatory tank. First, the basket of eggs and fine particles is placed in brine in the tank, stirred thoroughly, and let set for ca. 5 minutes. Since the eggs float and the diet particles sink, the floating eggs can be skimmed from the surface. (The brine and diet particles are drained from the tank to make the tank ready for a new batch.) The final cleaning of eggs is accomplished by plac- ing the eggs in a 1000-ml separatory funnel and rinsing them with fresh brine. Then the clean eggs are rinsed in fresh water to re- move the brine and placed in graduated cylinders for measurement and transfer to the sterilizing area.
The egg harvesting and cleaning operation produces a moist unsanitary environment; therefore, these operations are performed in a separate, well-ventilated room. All equipment is cleaned daily after use, and a constant supply of brine is maintained in two 303-liter plastic tanks.
Surface Sterilizing
After the cleaning, the eggs are surface sterilized in a so- lution of formalin and water to kill any microbes that could con- taminate the diet during the rearing period (Sikorowski et al. 1977). A bottle, ca. 3.8 liter, containing 2000 ml fresh water (filtered by a millipore filter with 0.3 micron pore size) and 500 ml of 37% formalin solution can accommodate 200 ml of eggs. This bottle with the eggs and the solution is shaken on a laboratory

278 J. G. GRIFFIN ETAL
shaker for 20 minutes and then rinsed twice in filtered water by shaking on the shaker for 10 minutes.
Planting Solution
A sterile planting solution, prepared by mixing 2000 ml water and 10 g Furcelleran (S. Malone unpublished data), is placed in a regular planting bottle and shaken to dissolve the Furcelleran. The Furcelleran produces the necessary viscosity to hold the eggs in suspension. The bottle and contents are then sterilized in an autoclave and stored in a clean area at 21°C. Just before plant- ing, solution and eggs, 7:1 ratio, are placed in a sterile plant- ing bottle and mixed under a laminar flow clean air hood. The bot- tles of suspended eggs are then held in the clean hood until the actual egg planting operation.
REARING
Egg Planting
In 1973, plastic trays replaced the petri dishes used at the Gast rearing facility to mass rear boll weevils (Griffin 1978, Griffin et al. 1979, Harrell et al. 1977). As a result, major changes in equipment and methods were necessary. The rearing trays now used are formed from roll stock of ca. 17-mil-thick, high- impact polystyrene and are covered and sealed with a film of Tyvek lidding material, also from roll stock.
The egg planting operation is performed semiautomatically with an in-line form-fill-seal machine (FFSM). Auxiliary equipment con- sisting of a diet filler, diet cooler, egg planter, and granular material filler is located along the line between the forming and sealing heads of the FFSM (Griffin et al. 1979, Harrell and Griffin 1981) (Figure 5). The trays, 15 (width) x 28 (length) x 1.9 (depth) cm, are formed sequentially in a continuous ribbon and moved intermittently along the processing line by the FFSM.
The semiliquid filler machine accumulates the sterile larval diet from the sterilizer and meters and dispenses ca. 185 ml into each tray cavity. After the tray is filled, it passes through the diet cooler (Griffin 1979a) where the diet is solidified by cool- ing. Next, the trays move to the planter (Griffin 1979c), which meters and uniformly disperses 4 ml of the egg suspension (ca. 2100 eggs) on the surface of the diet. Then the granular material fill- er machine meters and dispenses ca. 48 ml of granular material (de- scribed later this section) into each tray. Now the tray is moved to the sealing head where it is covered and sealed with Tyvek. Once the trays have been sealed, the ribbon is sheared into sec- tions of two trays each. Each section of trays is then shaken by hand so as to spread the granular material evenly over the surface

MASS REARING BOLL WEEVILS 279
Figure 5.—Equipment for tray processing and egg planting.
of the diet and then stacked in the sanitized rackveyors (Griffin 1979b) (Figure 6). Each rackveyor holds 896 of these trays.
The FFSM with auxiliary equipment is located in front of a wall module of HEPA filters (rated 99.9% efficient on particles > 0.3 microns). This location is necessary to provide an environ- ment of clean air in the space where the sterile diet, egg suspen- sion, granular material, tray plastic, and Tyvek are exposed during the planting operation.
The rolls of plastic and Tyvek are received from the supplier in a clean but not a sterile condition. Therefore, before use, they are fumigated with paraformaldehyde gas in a special chamber. The procedure reduces contamination of the diet.
Earlier, when petri dishes were used in rearing, the diet sur- face was scarified and dried so the young larvae would not drown in the moisture from the planting solution (Gast and Davich 1966, Griffin and Lindig 1978). The granular material mentioned pre- viously absorbs the free moisture, provides a cover for the eggs

^0 J. G. GRIFFIN ETAL
Figure 6.--Rackveyor used for stacking and conveying planted trays.
and diet (Griffin and Lindig 1975), and serves as a base for the antibiotic agents used to control contamination (Sikorowski et al. 1980). Additionally, it provides an object for the small larvae to push against and allows them to begin eating from the surface of the diet, which they cannot do effectively unless the surface is scarified. The material used is a mixture of sand and corncob grits (8-16 mesh) in a ratio of 7:3 (volume) (S. Malone unpub- lished data) to which antibiotic agents are added.
The sand preparation local sand pits is dried sieve. It and the grits are combined; afterwards sterile containers in a mixing 8825 ml sand, 377 1.25 g neomycin sulfate (75% folpet) (Sikorowski be blended for 15 minute duce a homogenous mix. tainers until needed in
is as follows: washed, mason sand from and sifted through a 6.3-mesh screening are sterilized in an autoclave before they
, further handling or blending is done in clean air cubicle. A batch is prepared by 5 ml grit, 1.25 g streptomycin sulfate, (or the equal of these), and 6 g Phaltan 1982). One or two batches at a time can
s in a 0.03-m twin shell blender to pro- The mix is stored in closed 19-liter con- the egg planting operation.
As noted, the egg planting operation is performed in a sepa- rate room of the rearing facility. Entry to this room is restrict- ed and the room is maintained as nearly as possible at the clean- liness of a "class 100 clean room (3.5 particles per liter of

MASS REARING BOLL WEEVILS 281
air)." Therefore, the equipment involved in the planting operation is located in front of the clean air wall module. In addition, the conditioned air of the room is passed through HEPA filters because trays of diet that are contaminated at this point will be com- pletely destroyed by the end of the regular larval development
period.
Development
After the rackveyors are loaded with planted trays, they are rolled into the first of the two development rooms to remain for the first 9-10 days of the larval development period. Then they are rolled into the second development room where they remain for the remainder of the 13-day period. By the 13th day after the eggs are planted, ca. 10% of the weevils have developed to the adult stage and are ready to leave the trays.
Both development rooms are held at 3r±rc and 50%±5% relative humidity, with continuous light, and both are maintained as near the cleanliness of a class 100 clean room as possible. The sepa- rate rooms are necessary because the trays of newly planted pre- adult-stage weevils must be separated from the trays containing older weevils that might escape as adults and contaminate the newly planted trays.
The air temperature, relative humidity, and circulation in all areas of both development rooms must always be at the proper levels to obtain uniformity in development and the maximum yield of wee- vils (Griffin et al. 1979). As noted, the air in the rooms is kept clean by passing the entering conditioned air through HEPA filters, and entry to the rooms is restricted.
The rackveyors are arranged in the development rooms so there is adequate space and ventilation between adjoining units. This arrangement means that the rearing medium will produce maximum yields of weevils.
Adult Emergence
Weevils from eggs oviposited and planted the same day never develop so uniformly that all adult emergence occurs the same day (Griffin et al. 1979). Emergence will usually occur over a 6- to 7-day period (13-20 days after eggs are planted), but in the mass rearing facility, trays are held in the emergence area for only 5 days. By this time, 90-95% of the adults will have emerged, and any emerging later have proved to be of poor quality.
Thus, the 13th day after eggs are planted, the rackveyor of trays is rolled from the number 2 development room to the tray preparation room in the emergence area. Here, while the trays are still on the rackveyor, a hole, ca. 1.3 x 2.5 cm, is burned in one

282 J. G. GRIFFIN ETAL.
end of each tray with an electrically heated charcoal lighter to provide a way for the adult weevils to leave the trays. Immedi- ately afterwards the rackveyor of trays is rolled into an emergence cubicle (Griffin and Roberson 1980).
The emergence cubicle has inside dimensions of 2.0 (width) x 2.4 (length) x 1.9 (height) m. Each can accommodate three rackvey- ors of trays, the equivalent of one day's planting. Access into the cubicle is through a 1.1 (width) x 1.8 (height) m door located in one wall. This wall also has twenty-eight 6.4-cm-diameter holes arranged in four columns of seven holes each (Figure 7). Jar lids (100 mm) with 6.4-cm holes in their center are mounted over these holes, and 3.8-liter plastic jars with a hole cut into the bottoms are screwed into the lids to serve as collection containers for emerging adult weevils. The bottom holes are covered with 5.5-mesh hardware cloth to allow for air circulation but make the opening weevil proof. Since the only light in the cubicle enters through the jars and since boll weevils are positively phototactic, the weevils are attracted from the trays.
At least once each day (more frequently if adult emergence is heavy), the weevils are removed from the jars and weighed to
Figure 7.—Emergence cubicles.

MASS REARING BOLL WEEVILS 283
determine the number obtained. Later, they are distributed and used for colony maintenance or are sterilized. After the adults have been harvested, the rackveyors with trays are removed from the emergence cubicle and rolled into a small hot room maintained at ca. 66**C. They remain there for ca. 24 hours; that is, until any live weevils remaining in the trays have been killed. The trays of spent diet are then discarded.
The emergence cubicles are maintained at 31°±1**C and 50%±5% relative humidity. An exhaust fan exchanges the air inside the cu- bicles with the fresh air entering through the collecting jars. The cubicles also have an exhaust system vented above the roof of the building so the cubicles can be fumigated. The cubicles and rackveyors are cleaned and fumigated after each use.
A detailed list and a description of all production equipment used in the operations are given by Harrell and Griffin (1981).
MICROBIAL CONTAMINATION
One of the major problems encountered in mass rearing the boll weevil was microbial contamination, here defined as the harboring of or having contact with micro-organisms in the absence of a rela- tionship that may be considered commensal istic, mutualistic, or parasitic (Steinhaus and Martignoni 1970). The microbial contami- nants encountered are usually microbes that are otherwise nocuous. However, it is well-documented in the literature of animal pathol- ogy that micro-organisms normally considered nocuous can multiply extensively in the tissue of weakened hosts and so cause disease that may often be severe (Neter 1974). The boll weevil diet is a complex medium so it is subject to spoilage by many species of bac- teria and fungi. Then since spoilage is the result of metabolic activity associated with microbial growth, the spoiled diet causes catabolism and release of products of digestion. In other words, the biochemical changes produced by microbes alter the nutritional value of the medium. In addition, microbial products such as bac- terial toxins and myotoxins may interfere with weevil defense mech- anisms or may damage weevil cells.
The cost of rearing weevils was greatly reduced by establish- ing an environmental sanitation program (Sikorowski 1975). How- ever, much of the information pertinent to this aspect of labora- tory rearing of weevils has been obtained only recently and merits further study. Meanwhile, from present research results, sanita- tion is important in the mass rearing of boll weevils (and most other insects), and more attention will have to be given to strict enforcement of basic sanitary measures to achieve volume production of healthy insects.
The safety of employees must also be considered in designing environmental sanitation programs. A number of airborne micro- organisms such as Aspergillus, Pseudomonas, and Streptococcus,

284 J- G- GRIFFIN ET AL
which grow on almost any organic matter, are human pathogens too and constitute some degree of hazard to employees.
Sources of Contamination
The microbial contamination that causes problems in rearing laboratories is mostly generated by man and equipment. Humans are the primary source, and the level of contamination relates directly to the activity and density of personnel (Favero et al. 1966, 1968; Runkle and Phillips 1969). In fact, one healthy human body harbors millions of micro-organisms on the skin and in the mouth, respira- tory tract, genitourinary tract, and intestines. Jawetz et al. (1978) arranged the normal microbial flora of the human body into two groups: (1) the resident flora, which consists of fixed types of micro-organisms regularly found in a given area and in persons of a given age; and (2) the transient flora, which consists of non- pathogenic or potentially pathogenic micro-organisms that inhabit the skin and mucous membranes for hours, days, or weeks. (This second group is derived from the environment, does not produce dis- ease, and does not establish itself permanently on the surface.)
Our studies showed that freshly laundered and sterilized cot- ton uniforms do not stop the movement of bacteria through the cloth into the environment (Sikorowski 1975, Sikorowski and Thompson 1980) (Figure 8). Also, hairs from 50 persons (1 hair per person) each had from several to many bacteria; and cultures from hands always produce bacterial colonies, even shortly after normal wash- ing. Talking, coughing, and sneezing likewise produce numerous moisture droplets, many of which contain bacteria. Finally, the malfunction of equipment or incorrect sterilization of equipment has usually been associated with massive contamination (Sikorowski 1975).
Microbial Flora of Weevils
In nature, adult boll weevils are solitary insects. They are seldom found together except when mating or when the food supply becomes scarce (Gast 1966). Laboratory-reared weevils, on the other hand, are always crowded together. In addition, insectary conditions are generally different from conditions in nature, which also contributes to the problem of microbial contamination (Shapiro 1982).
Field-Collected Weevils
The newly emerged boll weevil larva is sterile, but it imme- diately becomes bacterially contaminated because it feeds on decom- posing cotton squares. Thus field-collected larvae examined in the laboratory have very high numbers of bacteria per larva. However, the composition of the bacterial flora is simple and usually
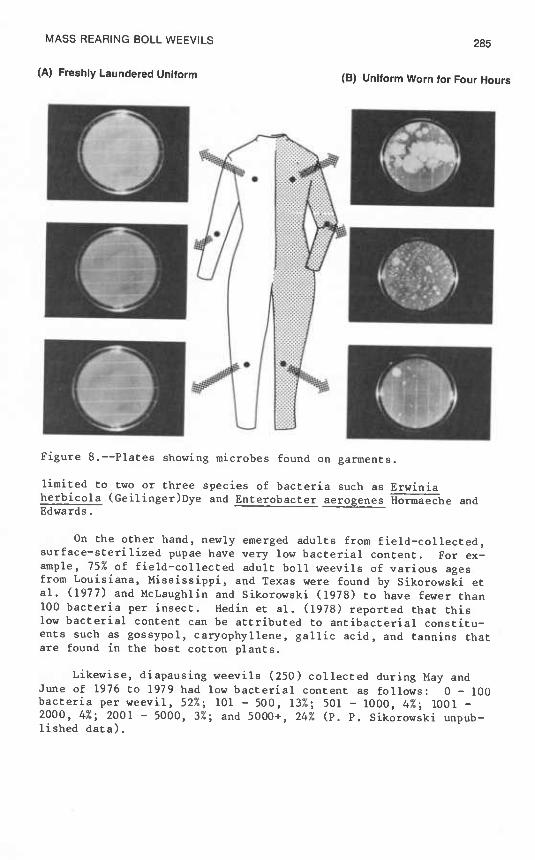
MASS REARING BOLL WEEVILS 285
(A) Freshly Laundered Uniform (B) Uniform Worn for Four Hours
Figure 8.—Plates showing microbes found on garments.
limited to two or three species of bacteria such as Erwinia herbicola (Geilinger)Dye and Enterobacter aerogenes Hormaeche and Edwards.
On the other hand, newly emerged adults from field-collected, surface-sterilized pupae have very low bacterial content. For ex- ample, 75% of field-collected adult boll weevils of various ages from Louisiana, Mississippi, and Texas were found by Sikorowski et al. (1977) and McLaughlin and Sikorowski (1978) to have fewer than 100 bacteria per insect. Hedin et al. (1978) reported that this low bacterial content can be attributed to antibacterial constitu- ents such as gossypol, caryophyllene, gallic acid, and tannins that are found in the host cotton plants.
Likewise, diapausing weevils (250) collected during May and June of 1976 to 197 9 had low bacterial content as follows: 0 - 100 bacteria per weevil, 52%; 101 - 500, 13%; 501 - 1000, 4%; 1001 - 2000. 4%; 2001 - 5000, 3%; and 5000+, 24% (P. P. Sikorowski unpub- lished data).

286 J. G. GRIFFIN ETAL
Insectary-Reared Weevils
Weevils reared in an insectary, unlike field-collected wee- vils, are frequently contaminated with diverse species of aerobic and anaerobic bacteria. Thus, Sikorowski (1982) isolated many taxonomically unrelated species from apparently healthy insectary- reared boll weevils as follows: from the family Micrococcaceae, Micrococcus luteus (Schroeter) Cohn and Staphylococcus aureus Rosenbach; Streptococcaceae, Streptococcus spp., Leuconostoc mesenteroides Cienkowski van Tieghem; Bacillaceae, Bacillus sphaericus Neide; Lactobacillaceae, Lactobacillus plantarum (Orla- Jensen) Bergey et al.; Pseudomonadaceae, Pseudomonas aeruginosa (Schroeter) Migula; Enterobacteriaceae, Enterobacter aerogenes; and Coryneform Group, Cornynebacterium humiferum Seliskar. Later McLaughlin and Sikorowski (1978) tested 35 bacterial cultures, mostly from the American Type Culture Collection, for their ability to grow on the boll weevil diet. They found that 5 of 15 human pathogens and 7 of 20 saprophytes developed on the diet.
Effects of Microbial Contamination
Effects of bacterial contamination on weevil development and on amino and fatty acids, pheromone, and histopathology were ex- amined by Sikorowski (1975), Gueldner et al. (1977), Sikorowski et al. (1977), Thompson et al. (1977), McLaughlin and Sikorowski (1978), Thompson and Sikorowski (1978), Sikorowski and Thompson (1979, 1980), and MacGown and Sikorowski (1980).
Weevil Development
The number of weevils that emerge from contaminated diets is influenced by the species of micro-organisms present, the stage of the insect at the time of contamination, the temperature, and other factors. However, the most conspicuous effects of microbial con- tamination are associated with the early developmental stages of the insect. For example, from 1972 to 1976 (the Gast rearing facility was completed in 1972), Aspergillus niger van Tieghem was present in the rearing medium for weeks or even months at a time. This contamination with A. niger, and occasionally with A. flavus Link, Pénicillium spp., yeast, and Rhizopus spp., was a serious problem when it occurred during early stages of larval development: larvae that were covered with conidia died, presumably from starva- tion. However, A. niger was normally present toward the end of the larval period when it did relatively little damage. Use of sand treated with Phaltan has since reduced A. niger contamination to almost zero (Sikorowski et al. 1980).
Bacterial contamination of the diet also will cause a reduc- tion in yield per tray. P. P. Sikorowski (unpublished data) found that boll weevil diet contaminated with ^. aureus or Streptococcus

MASS REARING BOLL WEEVILS 287
spp. at the time eggs were implanted produced 57 and 76%, respec- tively, as many weevils per petri dish as did uncontaminated diet. Similar values for the third and sixth days after hatch were 57 and 64% and 71 and 80%. In contrast, fast growing bacteria such as Leuconostoc spp. may overgrow an area of diet of about 700 mm dur- ing as little as 24 hours, meanwhile smothering all the larvae on the dish. Staphylococci (enterotoxin producers) also may have ad- verse effects on the larvae: the normal developmental time of 13 days from egg to adult weevil was extended to 14 or 16 days when diets were contaminated with such species of bacteria.
Amino and Fatty Acids, Pheromone, and Histopathology
Thompson et al. (1977) and Thompson and Sikorowski (1978) studied effects of Streptococcus sp., Micrococcus varians Migula, and E. aerogenes on the amino and fatty acid content of boll wee- vilsT They reported that all amino acids analyzed, except tyrosine and glutamine in females, were present in greater amounts in wee- vils free of bacterial contamination. The average reduction was 33% for males and 52% for females. The high levels of tyrosine and glutamine in contaminated female weevils were not explained. Comparison of individual fatty acids between groups showed a^de- crease of up to 76% in highly contaminated insects (2.1 x 10 bacteria per insect).
Gueldner et al. (1977), P. A. Hedin and P. P. Sikorowski (un- published data), and G. Wiygul and P. P. Sikorowski (unpublished data) examined the effects of bacterial contamination on pheromone production by the male boll weevil by measuring the amounts present in the feces and in the homogenate of the body. More pheromone was always isolated from the frass and homogenate of uncontaminated weevils than from the frass and homogenate of contaminated weevils. Then since pheromone production, and hence attractiveness of the sterile released weevil, is an essential characteristic, weevils that are bacterially contaminated would be expected to have greatly diminished value.
Also, McLaughlin and Sikorowski (1978), Sikorowski and Thomp- son (1979, 1980), and MacGown and Sikorowski (1980) examined the effects of bacterial contamination on the brush border of midgut epithelium of the boll weevil. In contaminated weevils, the brush border was frequently ulcerated and overgrown by bacteria. In ad- dition, MacGown and Sikorowski (1980) showed Jhat though bacterial contamination and irradiation (10 krads of Cs ) both cause rapid deterioration of midgut epithelium of adult boll weevils, S.aureus and Streptococcus spp. greatly accelerated the effects of irradia- tion"^ The result was destruction of the tissues within a few days and subsequent premature mortality. R. A. Leopold (personal com- munication) has detected similar histopathological changes in bac- terial ly contaminated boll weevils after irradiation.

288 J.G. GRIFFIN ETAL
Sanitary Measures
The principal objective of the sanitary methods used at the Gast rearing facility is to prevent entry and spread of microbes. The program includes maintaining a clean and sanitary environment by proper housekeeping, personal hygiene, proper waste disposal, proper use of gowns, gloves, and masks, and traffic control to minimize spread of microbes from area to area.
Numerous investigators have shown that the primary sources of microbial contamination within insectaries are: (1) human, (2) unsterilized diet, (3) insects, and (4) the facility itself. The significance of control of microbial contamination has been dis- cussed by Steinhaus (1953), McEwen and Hervey (1960), Beard and Walton (1965), Gast (1966), Gast and Davich (1966), Ignoffo (1966), Lyon and Flake (1966), Martin (1966), O'Dell and Rollinson (1966), Raun (1966), Magnoler (1970), McLaughlin (1970), Helms and Raun (1971), Howell (1971), Sutter and Miller (1972), Sikorowski (1975), Stewart et al. (1976), Baumhover et al. (1977), and Shapiro (1982). In addition, Sikorowski (1975) described microbial monitoring and sanitary measures that are used at the Gast rearing facility. Also see reviews by Shapiro (1982), Goodwin (1982), and Sikorowski (1982) for extensive treatments of this subject.
Housekeeping
Micro-organisms settle from the air (onto floors, tables, shelves, external surfaces of equipment) and rise into the air again as a result of any movement in the room. The general house- keeping routine for minimizing contamination from this source is mopping and disinfecting floors. Litsky and Litsky (1968) and many other workers in the field of microbial contamination have demon- strated that flooding and wet-vacuuming is the most effective method of doing this. Portable vacuum cleaners may be used only if the vacuum cleaner exhaust is filtered at least as well as the air in the room. Sweeping and dry-mopping tend to stir up micro- organisms and should be avoided (Nagasawa et al. 1970). Runkle and Phillips (1969) recommended elimination of sweeping by use of dry or wet pickup vacuum cleaners equipped with high efficiency exhaust air filters. All floors in the Gast facility are mopped twice daily, in the morning and again in the afternoon (1 hour be- fore the end of an 8-hour shift).
Personal Hygiene
Micro-organisms are usually shed with the human skin scales to which they are attached. Thus, personnel involved in handling in- sect diets and adult weevils and with planting eggs are required to wear caps and wash their hands frequently with medicated scrub soap. Workers handling weevil diets are also required to wear hair caps and face masks.

MASS REARING BOLL WEEVILS 289
Sterilization of Equipment
Sterilization means the destruction of all life, and heat is the most reliable and universally used method of sterilization. Whenever possible, this is the method used at the Gast rearing
facility.
Large equipment such as the apparatus for processing medium that is permanently attached to the walls is sanitized immediately before and just after use to prevent entry of microbes into the ap- paratus. (On one occasion, improper sterilization of this equip- ment caused massive contamination of diet that resulted in loss of time, money, and insects.) Small instruments that cannot be auto- claved are sanitized by immersion in sanitizing solution.
Contaminant Monitoring Program at Gast Rearing Facility
The purpose of contaminant monitoring at the facility was to evaluate the effectiveness of the sanitary measures used there. The tests are made to determine (1) air microbial content, (2) sterility of diet, (3) surface sterility of weevil eggs, (4) micro- organisms on surfaces, and (5) presence of micro-organisms in adult
weevils.
Air Microbial Content
The two methods used to monitor the microbial content of the air in the facility were as follows:
(1) Membrane filter method: The air was filtered through a membrane filter (0.45-micron pore size) supported in a filter as- sembly. The filter was then placed in a 12- x 54-mra plastic petri dish with an absorbent pad saturated with mycological or plate count broth and incubated at 32'C for 24 hours. Afterwards the filters were removed from the dishes, air dried, and stained with méthylène blue solution; the levels of contamination were recorded. A sample consisted of 100 liters of air drawn through the filter.
(2) Agar setting plate method: An uncovered, agar-filled petri dish was allowed to remain open on a particular surface for 5 minutes. When airborne particles containing viable micro- organisms formed bacterial or fungal colonies on the agar medium after incubation, the colonies were counted, and the level of con- tamination was recorded. Microbial content of the air was moni- tored in egg planting and larval holding areas.
Sterility of Larval Diet
During preparation of the larval diet, 20-cc samples of diet were collected aseptically in sterile petri dishes every 15

290 J. G. GRIFFIN ETAL
minutes. The dishes were examined after 24-48 hours of incubation at Sy^C, and levels of contamination were determined.
Sterility of Eggs
Eggs were removed from the diet pellets as described by Grif- fin et al. (1979), surface sterilized with 10% formaldehyde (Siko- rowski et al. 1977), and planted on the diet (Griffin et al. 1979). Five samples of 2000 eggs each (each bottle of egg suspen- sion contained 250 ml eggs plus 1750 ml planting solution) were planted in petri dishes with sterile trypticase soy agar and then incubated for 72 hours at 37*'C. A Gas Pak Anaerobic System (Bal- timore Biological Laboratory) was used for subsequent isolation of anaerobic bacteria. If a dish contained one or more colonies, con- tamination was considered to have been demonstrated.
Micro-organisms on Surfaces
Two methods were used for detection of bacteria on surfaces, the swab-rinse method and the agar contact method. In the first method, a cotton-tipped sterile swab was rubbed over the surface of the object to be sampled. Then the tip of the swab was broken into a tube containing sterile diluent, the tube was shaken, and the rinse fluid was plated on appropriate culture medium. Care was taken to avoid contaminating the swab.
In the second method, a rodac plate was used to monitor sur- face cleanliness. This plate was a modified petri dish that con- tained a raised nutrient agar bed. It was placed in contact with a surface; then the samples were usually incubated for 48 hours at 35**C before the colonies were counted. Trypticase soy agar with lecithin and polysorbate 80 medium was prepared for detection of surface contamination. (The lecithin and polysorbate 80 inacti- vated residual disinfectant collected with the specimen.)
Microbial contamination in the Gast rearing facility was found to vary depending on the area tested. In the egg planting and lar- val holding rooms where air was filtered through HEPA filters, the bacterial count was less than 10 colonies per plate. (The tests were made after wet cleaning and drying of the floor before any traffic was permitted.) The equipment, when properly cleaned and sanitized (Griffin et al. 1979), produced sterile medium. Formal- dehyde-sterilized eggs were almost free of microbial contamination, but eggs scored during extraction were occasionally infected with bacteria and fungi.

MASS REARING BOLL WEEVILS 291
Micro-organisms in Adults
The criteria used by Sikorowski (1975) to evaluate quality of adult boll weevils were presence of bacteria, protozoa, viruses, and yeast.
Presence of bacteria.—A sample of 25 adults was frozen at -20°C for 15-20 minutes, surface sterilized with 0.5% sodium hypo- chlorite for 5 minutes, washed in two changes of sterile water (5 minutes each), and blended into 40 cc of sterile water for 30 sec- onds. The homogenate (40 ml) was then poured into a flask contain- ing 30 cc of melted agar medium warmed to 45''C; then the mixture was poured into petri dishes. After incubation, the colonies were counted, and the level of contamination was determined.
Presence of protozoa (Microsporida).—A sample of 25 adults was blended into 25 cc of water. The suspension was then filtered through cheesecloth and centrifuged at 3000 r per min for 10 min- utes. The resulting pellet was resuspended in a small quantity of water, smeared on slideè, and examined under a phase contrast mi- croscope. For additional information, see Hazard and Oldacre (1975) and Poinar and Thomas (1978).
Presence of virus.—If a viral disease was suspected, the host tissue was examined with a light or electron microscope as re- quired. Inclusion bodies of occluded viruses (nuclear and cyto- plasmic polyhedroses, the granulöses, and the insect pox diseases) constituted one of the two broad groups of insect viruses and were easily detected with light or phase contrast microscopes. "Free"^ viruses, which are not embedded in a crystalline matrix and consti- tute the second broad group, can only be detected by using the electron microscope. M. E. Martignoni (personal communication) has since suggested an infectivity test as a more general and probably less expensive method of detecting nonoccluded viruses.
Presence of yeast.—When yeast contamination was suspected, the methods given by Poinar and Thomas (1978) were used.
Control
The importance of strict sanitation in controlling microbial contamination was recognized by Gast as early as 1961 when the mass rearing program was started at the Boll Weevil Research Laboratory (Gast 1966). However, only recently—with the use of HEPA filters and antibiotics and enforcement of sanitary measures—has microbial contamination been brought under control.

^^ J. G. GRIFFIN ETAL.
Use of Air Filters
High-efficiency particulate air (HEPA) filters were used to remove airborne contaminants generated or introduced by activities inside the facility. In fact, use of these filters in the egg planting and larval holding areas of the facility was a major factor m the final reduction of microbial contamination. For additional information see Griffin et al. (1979), Griffin (1982) and Roberson and Wright (1982). ^ ■^'>
Use of Antibiotics
Gast and Davich (1966) utilized methyl paraben and sorbic acid
(1966) tpl!r9n""TK-" -^ '•'^ ""^^ ^^^^^^ '^^^' ^y y^^^t- Gast (1966) tested 20 antibiotics for inhibition of Pseudomonas (tenta- tive identification), but only streptomycin sulfate inhibited growth of the bacterium, and it failed to control multiplication and spread Cast also tested antibiotics that might control Mat- 4#£ü |I|Edis McLaughlin but had no success. Childress and wñ- liams (1973) controlled L. mesenteroides contamination of boll wee- vil diets with erythromycin. Sikorowski et al. (1980) then used antibiotics to shield the growth medium of larvae of boll weevil from microbial contamination and found that they had no detectable adverse effects on insect development, egg production, hatch, and production of male pheromone. Today standard diet covered with
thin'sno K'^^" ^ '^''' ®^"^^^ """^y """^S^'^ ^^^^il« that have fewer than 500 bacteria per weevil compared with 18% obtained from diet not covered with medicated sterile sand.
PRODUCTION RESULTS
As far as is known, the Boll Weevil Research Laboratory (BWRL) was the largest producer of boll weevils before the Gast rearing
(M^rlh'I K'i i'^fo^o'""- ^°'' ^^^mle, during a 6-month period (March-August) m 1969. production at the BWRL was ca. 0.7 million weevils per week. Likewise, in 1970, 6-month production averaged
only 67 adults per petri dish planted with an average of 536 eggs (0. H. Lindig unpublished data).
Production was started in the Gast rearing facility during the summer of 1972, but problems were encountered because the rearing operation started before the facility was fully completed and while equipment was still being installed. The mechanical equipment in the building malfunctioned, personnel were new and inexperienced, and diet contamination was the rule. As a result, production reached only ca. 1.25 million weevils per week in 1972. and in ly/J, the rate was still disappointing. However, during June 1973 production reached ca. 1.8 million weevils per w^ek (O. H. Lindig unpublished data) despite continuing problems with diet contami- nation and the loss of a high percentage of planted dishes.

MASS REARING BOLL WEEVILS 293
Table 4.—Production data for Gast rearing facility
Parameters measured 1976 1977 ^979
Trays planted per week 14,468 17,880 7,027 Eggs placed per tray 2,595 2,415 2,100 Adults emerged per tray 346 349 643 Eggs per female per day 3.6 3.4 5.6 Percentage of production for
maintenance 32 25 14 Percentage of yield
(adults per eggs planted) 13.3 14.5 30.6 Percentage of trays with
microbial contamination 15.0 0.9 5.8
Finally, between 1973 and 1976, as noted, new and modified semiautomatic equipment and improved procedures replaced the man- ual equipment and inefficient procedures used during 1972 and 1973 (Griffin and Lindig 1974, 1975, 1977; Griffin 1978; Harrell et al. 1977).
Perhaps the most important equipment added to combat contami- nation losses was the in-line form-fill-seal machine plus auxil- iary equipment, which allowed the change from rearing in petri dishes to rearing in plastic trays. The fact that the planting operations were thereafter done semiautomatically by machinery meant that all manual handling was eliminated and contamination caused by workers was reduced.
When the changes were tested in the trial production opera- tions conducted in 1976 and 1977 and used in 1979 to rear the wee- vils for the Boll Weevil Eradication Trial (discussed earlier), production was as shown in Table 4 (J. Roberson unpublished data). V
Because the number of trays planted and of adults used in the brood colony depended on the number of adult weevils desired, max- imum production potential was never reached. However, the increase in percentage yield is revealing.
CONCLUSION
The progress made in mass rearing boll weevils has brought us to the point that a viable operational system is available and proved capable of providing up to ca. 10 million per week. How- ever, more effort is needed to refine and advance present rearing technology, buildings, and equipment so greater numbers of high quality weevils can be produced more economically. Important in- formation has already been gained regarding larval development, egg production, yields, contamination, and equipment needed when

294 j. G. GRIFFIN ETAL
producing weevils at a rate of 6-8 million per week, but if this number must be increased severalfold, care must be exercised. For example, problems might occur if some rooms and some equipment must be enlarged. In fact, difficulties were encountered when the rear- ing operation was increased from ca. 0.5 to 5-6 million weevils per week. Moreover, egg production and hatch tend to fluctuate from one batch of eggs to another so a uniform rate is difficult to maintain.
New production equipment must be designed and constructed so that It can be cleaned and sanitized with a minimum of effort and time. It must also withstand the required cleaning and sanitizing and still be available for continuous use.
The rearing facility must be divided into separated or isolat- ed production rooms or areas, first for better sanitation and then for greater efficiency in handling materials or work movement. Building materials and construction must be such that cleaning and sanitizing are easy; they also must be durable.
A good housekeeping program is essential for successful pro- duction. Traffic must be restricted in the critical areas; e.g. those used for egg planting, larval development, and preparation'of eggs and planting solution.
The major problem is plainly microbial contamination, which must be held to a minimum. In an operation of this type, microbial contamination may be described as the occurrence and persistence of microbes in an environment where they are not wanted. The dif- ficulty is that the weevil diets provide growth media for an almost infinite number of species of microbes. In addition, these micro- bial contaminants may have different effects upon the weevils de- pending upon vigor and age of the weevils and composition of the medium. Among the more frequent problems associated with microbial contamination are the following: (1) high mortality of early in- stars, (2) prolonged developmental time, (3) diminutive pupae and adults, (4) reduced production of pheromone, (5) reduced synthesis of ammo and fatty acids by the weevils, and (6) wide fluctuations m the quality of the insects.
The objective of an environmental sanitation program in an in- sectary is therefore elimination of the contamination that inter- feres with the rearing of healthy insects. Such a program must be planned carefully and based on research results. Then it must be carried out meticulously.
^ In the rearing operation at the Gast rearing facility, use of sanitation practices, HEPA (absolute) air filters, and antimicro- bial agents has essentially eliminated microbial contamination of diets and insects. Consequently, the laboratory now has the tech- nology to produce disease-free insects.

MASS REARING BOLL WEEVILS 295
LITEEIATURE CITED
Bartlett, A. C. 1967. Genetic marker in boll weevil. Journal of Heredity 58:
159-163.
Baumhover, A. H. , W. W. Cántelo, J. M. Hobgood, Jr., C. M. Knott,
and J. J. Lam, Jr. 1977. An improved method for mass rearing the tobacco hornworm.
U.S. Department of Agriculture, ARS-S-167.
Beard, R. L., and G. S. Walton. 1965. An Aspergillus toxin lethal to larvae of the house fly. Journal of Invertebrate Pathology 7:522-523.
Childress, D., and P. P. Williams. 1973. Control of bacterial contaminant of boll weevil diet.
Journal of Economic Entomology 66:554-555.
Earle, N. W., R. C. Gaines, and J. S. Roussel. 1959. A larval diet for boll weevils containing an acetone pow-
der of cotton squares. Journal of Economic Entomology 52:710-
712.
Favero, M. S., J. R. Puleo, J. H. Marshall, and G. S. Oxborrow. 1966. Comparative levels and types of microbial contamination detected in industrial clean rooms. Applied Microbiology 14:
539-551.
J. R. Puleo, J. H. Marshall, and G. S. Oxborrow. 1968. Comparison of microbial contamination levels among hospi-
tal operating rooms and industrial clean rooms. Applied Micro- biology 16:480-486.
Gast, R. T. 1961. Some shortcuts in laboratory rearing of boll weevils. Journal of Economic Entomology 54:395-396.
r96T. Methods for mass production of diet pellets for adult boll weevils. Journal of Economic Entomology 58:1024-1025.
1966. Control of four diseases of laboratory-reared boll wee- vils. Journal of Economic Entomology 59:793-797.
and T. B. Davich. 1966. Boll weevils. In Insect Colonization and Mass Production,
p. 405-419. C. N. Smith, editor. Academic Press, New York.
and M. Landin 1966. Adult boll weevil and eggs marked with dye fed in the lar- val diet. Journal of Economic Entomology 59:474-475.

296 j. G. GRIFFIN ET AL
Gast, R. T., and H. Vardell. 1963. Mechanical devices to expedite boll weevil rearing in the
laboratory. U.S. Department of Agriculture, ARS 33-89, 10 p.
Goodwin, R. W. 1982. Recognition and diagnosis of diseases in insectaries and
the effects of disease agents on insect biology. In Advances and Challenges in Insect Rearing. E. G. King and N. C. Leppla, editors. U.S. Department of Agriculture, Technical Bulletin. [In press.]
Griffin, J. G. 1978. A system for the egg planting operation in boll weevil mass rearing. Transactions of the ASAE 21:469-472.
1979a. Equipment for cooling larval diet in boll weevil mass rearing operation. U.S. Department of Agriculture, Science and Education Administration, Advances in Agricultural Tech- nology, Southern Series (ISSN 0193 3728) No. 1,3 p.
1979b. "Rackveyor" for use in mass rearing boll weevils. U.S. Department of Agriculture, Science and Education Administra- tion, Advances in Agricultural Technology, Southern Series (ISSN 0193 3728) No. 4, 3 p.
1979c. Egg planter for a boll weevil mass-rearing operation. U.S. Department of Agriculture, Science and Education Adminis- tration, Advances in Agricultural Technology, Southern Series (ISSN 0193 3728) No. 6, 5 p.
1982. Insect rearing facilities. ^Advances and Challenges in Insect Rearing. E. G. King and N. C. Leppla, editors. U.S. Department of Agriculture, Technical Bulletin. [in press.]
and 0. H. Lindig. 1974. Mechanized production of boll weevil diet pellets. Trans-
actions of the ASAE 17:15-16 & 19.
and 0. H. Lindig. 1975. Granular material used in implanting boll weevil eggs on
artificial diet. Journal of Economic Entomology 68:433-434.
and 0. H. Lindig. 1977. System for mechanical harvesting of boll weevil eggs from
diet pellets. Transactions of the ASAE 20:454-456.

MASS REARING BOLL WEEVILS 297
Griffin, J. G., and 0. H. Lindig. 1978. Pilot facility for mass rearing of boll weevils. jLn Fa-
cilities for Insect Research and Production, p. 75-78. N. C. Leppla and T. R. Ashley, editors. U.S. Department of Agricul- ture, Technical Bulletin No. 1576.
and 0. H. Lindig. 1979. Oviposition cage and auxiliary equipment for mass rearing
of boll weevils. U.S. Department of Agriculture, Science and Education Administration, Advances in Agricultural Technology, Southern Series (ISSN 0193 3728) No. 5, 5 p.
0. H. Lindig, and R. E. McLaughlin. 1974. Flash sterilizers: sterilizing artificial diets for in-
sects. Journal of Economic Entomology 67:689.
0. H. Lindig, R. E. McLaughlin, and 0. L. Malone. 1971. A facility for mass rearing of boll weevils: a pilot model. American Society of Agricultural Engineers, Paper No.
71-594.
0. H. Lindig, J. Roberson, and P. P. Sikorowski. 1979. System for mass rearing boll weevils in a laboratory. Mississippi Agricultural and Forestry Experiment Station, Technical Bulletin No. 95, 15 p.
and J. Roberson. 1980. New and improved techniques for mass rearing boll weevils.
Southwestern Entomologist 5:165-168.
Gueldner, R. C, P. P. Sikorowski, and J. M. Wyatt. 1977. Bacterial load and pheromone production in the boll wee-
vil, Anthonomus grandis. Journal of Invertebrate Pathology 29:
397-398.
Harrell, E. A., and J. G. Griffin. 1981. Facility for mass rearing boll weevils - engineering as
pects. U.S. Department of Agriculture, Science and Education Administration, Advances in Agricultural Technology, Southern Series ISSN 0193 3728) No. 19, 77 p.
W. D. Perkins, A. N. Sparks, and R. F. Moore. WiT, Mechanized techniques for adult boll weevil (Coleóptera: Curculionidae) production. Transactions of the ASAE 20:450-
453.
Hazard, E. I., and S. W. Oldacre. 1975. Revision of Microsporida (protozoa) close to Thelohania, with descriptions of one new family, eight new genera and thirteen new species. U.S. Department of Agriculture, Techni- cal Bulletin No. 1530, 104 p.

29^ J. G. GRIFFIN ETAL
Hedin, P. A., O. H. Lindig, P. P. Sikorowski, and M. Wyatt. 1978. Suppressants of gut bacteria in the boll weevil from the
cotton plant. Journal of Economic Entomology 71:394-396.
Helms, T. J., and E. S. Raun. 1971. Perennial laboratory culture of disease-free insects. In Microbial Control of Insects and Mites, p. 639-654. H. D. Burges and N. W. Hussey, editors. Academic Press, New York.
Howell, J. F. 1971. Problems involved in rearing the codling moth on diet in
trays. Journal of Economic Entomology 64:631-636.
Ignoffo, C. M. 1966. Insect viruses. Iri Insect Colonization and Mass Produc-
tion, p. 501-530. C. N. Smith, editor. Academic Press, New York.
Jawetz, E., J. L. Melnick, and E. A. Adelberg. 1978. Review of medical microbiology. Lange Medical Publica-
tions, Los Altos, CA, 550 p.
Lindig, 0. H., and 0. L. Malone. 1973. Oviposition of boll weevils fed diets containing germi-
nated puree or cottonseed meats puree. Journal of Economic Entomology 66:566-567.
J. Roberson, and J. E. Wright. 1979. Evaluation of three larval and adult boll weevil diets. Journal of Economic Entomology 72:450-452.
G. Wiygul, J. E. Wright, J. R. Dawson, and J. Roberson. 1980. Rapid method for mass-marking boll weevils. Journal of Economic Entomology 73:385-386.
Litsky, B. Y., and W. Litsky. 1968. Investigations on decontamination of hospital surfaces by
the use of disinfectant-detergents. American Journal of Public Health 58:534-543.
Lyon, L. R., and H. W. Flake, Jr. 1966. Rearing Douglas-fir tussock moth larvae on synthetic media. Journal of Economic Entomology 59:696-698.
McEwen, F. L., and G. E. R. Hervey. 1960. Mass rearing the cabbage looper, Trichoplusia ni, with notes on its biology in the laboratory. Annals of the Entomo- logical Society of America 53:229-234.

MASS REARING BOLL WEEVILS 299
MacGown, M. W., and P. P. Sikorowski. 1980. Histopathology of midgut of mass reared, irradiated boll weevils contaminated with Staphylococcus aureus and Streptococ- cus sp. Journal of Economic Entomology 73:81-87.
McLaughlin, R. E. 1970. Systems of mass-rearing disease-free insects. Their value
to studies of diseased natural populations, with particular reference to the boll weevil. In Proceedings, IV International Colloquium on Insect Pathology, College Park, MD, p. 255-261.
and P. P. Sikorowski. 1978. Observations of boll weevil midgut when fed natural food bacterially contaminated artificial diet. Journal of Inverte- brate Pathology 32:64-70.
Magnoler, A. 1970. A wheat germ medium for rearing of the gypsy moth, Lyman-
tria dispar L. Entomophaga 15:401-406.
Martin, D. F. 1966. Pink bollworms. In. Insect Colonization and Mass Produc-
tion, p. 355-366. C. N. Smith, editor. Academic Press, New York.
Nagasawa, S., T. Tsuchiya, and H. Nishimura. 1970. Hospital disinfection. In. Disinfection, p. 275-289.
M. A. Bernarde, editor. Marcel Dekker, Inc., New York.
Neter, E. 1974. Opportunistic pathogens: immunological aspects. In Op- portunistic Pathogens, p. 37-48. J. E. Prier and H. Freidman, editors. University Park Press, Baltimore.
O'Dell, T. M., and W. D. Rollinson. 1966. A technique for rearing the gypsy moth, Porthetria dispar
(L.), on an artificial diet. Journal of Economic Entomology 59:741-742.
^Poinar, G. 0., Jr., and G. M. Thomas. 1978. Diagnostic manual for the identification of insect patho-
gens. Plenum Press, New York and London, 218 p.
Raun, E. S. 1966. European corn borer. Iji Insect Colonization and Mass Pro-
duction, p. 323-338. C. N. Smith, editor. Academic Press, New York.
Roberson, J., and J. E. Wright. 1982. Production of boll weevils, Anthonomus grandis Boheman.
In Advances and Challenges in Insect Rearing. E. G. King and^ Ñ7 C. Leppla, editors. U.S. Department of Agriculture, Techni- cal Bulletin. [In press.].

300 J. G. GRIFFIN ET AL.
Runkle, R. S., and G. B. Phillips. 1969. Microbial contamination control facilities. Van Nostrand Reinhold Co., New York, 198 p.
Shapiro, M. 1982. Microorganisms as contaminants or pathogens. In Advances
and Challenges in Insect Rearing. E. G. King and N. C. Leppla, editors. U.S. Department of Agriculture, Technical Bulletin. [In press.]
Sikorowski, P. P. 1975. Microbiological monitoring in boll weevil rearing facil-
ity. Mississippi Agricultural and Forestry Experiment Sta- tion, Technical Bulletin No. 71, 21 p.
1982. Microbial contamination: occurrence, prevention and con- trol. J[n Advances and Challenges in Insect Rearing. E. G. King and N. C. Leppla, editors. U.S. Department of Agricul- ture, Technical Bulletin, [in press.]
A. D. Kent, 0. H. Lindig, G. Wiygul, and J. Roberson. 1980. Laboratory and insectary studies on the use of antibiotics
and antimicrobial agents in mass-rearing of boll weevils. Journal of Economic Entomology 73:106-110.
and A. C. Thompson. 1979. Effects of bacterial load on mass reared boll weevils Anthonomus grandis. Progress in invertebrate pathology. In Proceedings, International Colloquium on Invertebrate Pathol- ogy, September 11-17, 1978, Agricultural College Campus, Prague, CSSR.
and A. C. Thompson. 1980. Effects of internal bacterial contaminants on mass reared
boll weevils Anthonomus grandis Boheman. Mississippi Agricul- tural and Forestry Experiment Station, Technical Bulletin No. 103, 8 p.
J. M. Wyatt, and 0. H. Lindig. 1977. Method for surface sterilization of boll weevil eggs and
the determination of microbial contamination of adults. Southwestern Entomologist 2:32-36.
Steinhaus, E. A. 1953. Diseases of insects reared in the laboratory or insec-
tary. California Agricultural Experiment Station, Extension Series, 26 p.

MASS REARING BOLL WEEVILS 301
Steinhaus, E. A., and M. E. Martignoni. 1970. An abridged glossary of terms used in invertebrate pathol-
ogy. U. S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station, 38 p.
Stewart, F. D., M. R. Bell, A. J. Martinez, J. L. Roberson, and A. M. Lowe.
1976. The surface sterilization of pink bollworm eggs and spread of a cytoplasmic polyhedrosis virus in rearing containers. U.S. Department of Agriculture, APHIS 81-27.
Sutter, G. R., and E. Miller. 1972. Rearing the army cutworm on an artificial diet. Journal
of Economic Entomology 65:717-718.
Thompson, A. C., and P. P. Sikorowski. 1978. The effect of bacterial load on amino acids in the boll weevil, Anthonomus grandis. Journal of Invertebrate Pathology 32:388-389.
P. P. Sikorowski, and J. M. Wyatt. 1977. The effect of bacterial load on fatty acids in the boll weevil Anthonomus grandis (Coleóptera: Curculionidae). Jour- nal of Invertebrate Pathology 30:274-275.
Vanderzant, E. S., and T. B. Davich. 1958. Laboratory rearing of boll weevils: a satisfactory larval
diet and oviposit ion studies. Journal of Economic Entomology 51:288-291.
and T. B. Davich. 1961. Artificial diets for the adult boll weevil and techniques
for obtaining eggs. Journal of Economic Entomology 54:923-928.


303
Chapter 12
SAMPLING ARTHROPODS IN COTTON
J. W. Smith Bioenvironmental Insect Control Laboratory Agricultural Research Service U.S. Department of Agriculture Stoneville, MS 38776
W. A. Dickerson Boll Weevil Eradication Research Agricultural Research Service U.S. Department of Agriculture Raleigh, NC 27607
W. P. Scott Bioenvironmental Insect Control Laboratory Agricultural Research Service U.S. Department of Agriculture Stoneville, MS 38776
ABSTRACT This chapter deals first with arthropod sampling in general and addresses the problems of sampling locations, dura- tion, methods, size, and statistical treat- ment. Next, specific problems having to do with arthropod sampling in cotton are dis- cussed, and techniques are reviewed. Infor- mation is given about the efficiency of sev- eral often-used techniques.
Sampling for specific species of cotton pests and for beneficial arthropods is then reviewed.
The sampling methods actually used by the research teams associated with the Boll Weevil Eradication and Optimum Pest Management Trials are discussed in detail.
INTRODUCTION
Insect populations are usually sampled for two reasons: one is to determine whether an economic action level has been reached, and, hence, whether control measures should be initiated; the other is to obtain an estimate of the density of populations useful for

304 J. W. SMITH ET AL
research purposes. However, as Southwood (1966) points out, it is usually impossible to count all the vertebrates in a habitat; therefore sampling part of the population is necessary to obtain an estimate of a total population. Morris (1960) reminds us that "Sampling has no intrinsic merit but is only a tool which the ento- mologist should use to obtain certain information provided there is no easier way to get the information." Because several recent re- view publications (Sterling 1979, Kogan and Herzog 1980) have dis- cussed sampling for economic thresholds and decision making exten- sively, we are concerned here primarily with sampling for research purposes.
Population sampling may be either extensive or intensive (Mor- ris 1960, Strickland 1961). Extensive sampling is usually neces- sary when large areas are to be surveyed and tends to be concerned with such problems as the relative abundance and distribution of insects among fields, their relationship to crop damage, or the ef- ficacy of some areawide control measure. Intensive sampling, on the other hand, involves sampling of a specific population over time. The resulting data are often used to develop life tables, estimate changes in population growth parameters, or determine how a population is distributed within a field (Morris 1957, Southwood 1966, Harcourt 1969, Varley and Gradwell 1971).
ARTHROPOD SAMPLING FOR RESEARCH
The sampling methods selected by a researcher should be chosen according to the final goal. For example, methods used to deter- mine economic thresholds and the need for control procedures are often not adequate for estimating population densities or measuring inherent variability of insect populations from location to loca- tion. Sampling for insects in cotton has been particularly vulner- able in this respect (Byerly et al. 1978). We need to develop sampling theory and application that will avoid bias and maximize statistical efficiency. Important considerations include location, duration, method, and size of the sampling done.
Sampling Locations
Ideally, population sampling should be done at a number of randomly selected locations so as to obtain a statistical estimate of the variance between locations. Indeed, since variance reflects the environmental influences associated with different sample sites, a population model for a pest species cannot be developed by samp- ling in just one location (Watt 1966). Moreover, as Mott (1963) points out, the selected locations should include not only model values of the variables in question but the extreme values as well.
However, the pattern of sampling needed is dependent on the objective of each program. If the objective is to obtain estimates of mean density for preparing life tables, then it is desirable to

SAMPLING ARTHROPODS IN COTTON 305
minimize variance (Southwood 1966). In addition, unrestricted ran- dom sampling is not efficient in minimizing variance because most of the samples might be taken from one area of the field. The method of stratified random sampling is, therefore, preferred for most ecological work (Healy 1962). ^Here the area is divided into a number of equal-sized subdivisions or strata, and one or more sam- ples are randomly selected from each. This approach maximizes the accuracy of the estimate of the population, but exact estimates of sampling error can only be obtained if additional samples are taken from one strata.
Sampling Duration
When populations fluctuate irregularly, studies must continue long enough to permit observation of a sequence of increases and decreases so one can define the pattern of fluctuations. As col- lection of data proceeds, the sampling method should be tested and refined to ensure that deviations are accurately measured.
Sampling Methods
Watt (1966) suggested that each component of a biological sys- tem should be measured in such a way as to expose the real biologi- cal impact of the system. The problem is that the preliminary sam- pling and the analysis of the assembled data will provide a measure of many of the variables, but the actual decisions on technique must, in many cases, be a matter of judgment. As density changes, so will many of the statistical parameters, and a sampling method that is suitable at a higher density may be found inadequate as the population decreases. Therefore, sampling procedures must be flex- ible enough to deal with populations that vary in size.
Sampling Size
Obviously, the greater the sample size, the more reliable the estimates become. However, the cost per unit sampled may be sub- stantial, and the collection of large samples may be unnecessarily costly. Thus, the smallest sample size that assures the desired reliability of the estimate should be determined in advance. Karandiros (1976) refers to this as optimum sample size. He also defined practical reliability in three ways and derived formulas for the first two; that is, mean and variance (general case). He then derived the formulas for the Negative Binomial, the Poisson, and the Binomial distributions as special cases.
Waters and Henson (1959) suggest taking sampling units of at least two sizes. One sample should tend towards the smallest pos- sible limit, for example, a leaf or half a leaf, because a higher level of reproducibility is usually obtained (for the same cost) by taking more small units than by taking fewer large ones. In other cases, it may be necessary to increase the size of the sample unit

306 J. W. SMITH ET AL
(Anderson 1965) to overcome the disadvantage of sampling by small units the number of zeros that may result at low densities. (This truncation may make analysis difficult though moderate trun- cation can be overcome by suitable transformation.) However, the decision to take larger samples must be related to the density of the population. Small sampling units may also increase precision by distinguishing between favorable and unfavorable microhabitats.
Before sampling is begun, the investigator must know whether different regions of the sampling area should be sampled sepa- rately. If the distribution of the population throughout the habi- tats is biased towards certain subdivisions, errors will arise when sampling is done randomly. These errors can be overcome by arrang- ing the pattern so the number of samples taken from each subdivi- sion reproduces the gradient in the habitat or by regarding each part separately and correcting for it at the end. However, occa- sionally, such a large and constant proportion of the population occurs on one particular part of the plant that sampling may be restricted to this part. The taking of a certain number of samples randomly within a site, which is itself selected randomly from within a larger area, such as a field, is often referred to as nested sampling and may be on two, three, or more levels.
Morris (1955) laid down six criteria for a sample unit as follows:
(1) It must be such that all units of the universe have an equal chance of selection.
(2) It must have stability; if not, change should be easily and continuously measured.
(3) The proportion of the insect population using the sample unit as a habitat must remain constant.
(4) The sampling unit must lend itself to conversion to unit areas.
(5) The sampling unit must be easily delineated in the field.
(6) The sampling unit should be of such a size as to provide a reasonable balance between the variance and the cost.
Other Sampling Variables
The most serious biological problem in sampling arises when a part of the population has a behavioral pattern or habitat prefer- ence that is never sampled (MacLeod 1958). Less dangerous to the analysis (because the phenomenon will be recorded though it may well be misinterpreted) is the tendency for the behavior of the sampled insect to change. The behavior of insects is probably al- tered with age, weather, and even with the time of day, and these

SAMPLING ARTHROPODS IN COTTON 307
changes may lead to a confusion in interpreting data. For example, the time of day when sampling is done may affect quality consider- ably because the diurnal rhythm of many insects causes them to move from one part of the habitat to another. Also, during the day, quite a proportion of active insects may be airborne (Southwood 1966). Clark et al. (1967) suggest that many sampling problems can be overcome, or at least additional information gained, if work is done at dusk and dawn rather than during conventional working hours.
TECHNIQUES FOR SAMPLING ARTHROPODS IN COTTON VEGETATION
Sampling methods for estimating population densities fall into three general groups: absolute methods, relative methods, and pop- ulation indices (Roach et al. 1979). Population indices do not produce a count of insects; rather, they measure insect products or effects and are frequently used in making control decisions.
Absolute Methods
Accurate estimates of densities of arthropod populations are needed for almost all research-oriented sampling. Once an absolute method is devised, data obtained with relative methods can be com- pared and adjusted to provide estimates of density per unit of land area. Extensive reviews of absolute sampling methods and ad- ditional references can be found in Southwood (1978) and Ruesink and Kogan (1975).
An example would be the "clam shell" device (Leigh et al. 1970) suggested for estimating absolute insect populations on cotton. This method solved several problems: it prevented the es- cape of insects and allowed counting and identification in the lab- oratory. However, the plants must be removed from the study area, which is not desirable in most research situations.
A whole plant harvest technique was used to estimate adult ce- real leaf beetles (Oulema melanopus (L.)) on grain (Ruesink and Haynes 1973) and on soybeans (Mayse et al. 1978). J. W. Smith (un- published data 1972) used the same technique on cotton and compared it with the D-Vac and sweep net methods.
Smith et al. (1976) reported on an absolute unit area sampler that does not have the problems associated with the whole plant and clam shell methods. However, this sampler is not recommended as a sampling technique per se but rather as a method of establishing the relative efficiency of methods now being used.
Thus, absolute methods yield estimates such as density per unit of land area in the habitat, the type of data most often de- sired by population ecologists. However, such estimates must be

308 J- W. SMITH ET AL
made over time for nearly all studies of population dynamics of un- caged field populations.
Relative Methods
Most methods of sampling used today by cotton entomologists are relative methods that yield density per unit other than land area and must be converted to an absolute value. Often, this conversion cannot be done without correcting for the behavior of the insects or the effects of habitat, or both. For example, sweep nets, ground cloths, direct observations, and suction devices are widely used in sampling for insects in cotton. Direct observation can produce absolute (direct) population estimates for some spe- cies. The other methods constitute relative sampling, and the data need calibration against data obtained from reliable absolute samp- ling techniques before the results can be converted to estimates of density.
Another very important factor to consider when one is sampling arthropod populations in cotton is the intense interaction between the arthropods and the plants. Both the cotton plants and the arthropod populations change through the season (Byerly et al. 1978), and as the plants grow, they provide more places for arthro- pods to disperse. The combined result of plant growth and animal dispersion has great effect on the efficiency of most sampling methods.
Characteristically, cotton plants grow rapidly until the can- opy closes, or they suffer carbohydrate stress because of the de- mands of large numbers of bolls (Gutierrez et al. 1975). Then, late in the season, plants may bend from the weight of maturing fruit or be blown down by wind. This phenology is likely to in- fluence sampling efficiency in the following ways:
(1) When the plants are small and the canopy quite open, all sampling methods are likely to be quite efficient.
(2) As the canopy begins to close, some relative methods of sampling tend to sample only the populations on the upper part of the plant and so become less efficient.
(3) When the plants begin to bend, none of the methods of relative sampling are likely to provide adequate estimates of popu- lation size.
Whole Plant Survey
Whole plant examination is the oldest and most direct of all sampling methods for arthropods on cotton. Many entomologists have felt that it was the only accurate method. Recently, the method was used by Pieters and Sterling (1973) and Smith et al. (1976) to

SAMPLING ARTHROPODS IN COTTON 309
sample cotton arthropods. Also, Mayse et al. (1978) used this method to study arthropod communities in Illinois soybean fields.
Kogan and Pitre (1980) gave a detailed description of a method of whole plant examination. Initially, nonadjacent randomly selec- ted plants are scanned for large, often fast moving species of ar- thropods. Then both sides of each leaf on each of the plants are searched, as are the petioles, axils, and stems. The apical leaves are uncurled and searched for thrips, mites, and other small ar- thropods. Blossoms and fruiting forms are also searched. Numbers of each species, the stage of development, the relative position on the plant, and any other biological data of interest are recorded. The method can also be used to assess a population of a single spe- cies; in that case, the search strategy may be simplified by taking advantage of certain behavioral characteristics of the species of interest (Kogan and Pitre 1980).
Whole plant examination can be used early in the season when plants are small and tender without seriously damaging the plants. In addition, whole plant counts can be made for larger plants, though it becomes impossible to make them over a measured length of row. Mayse et al. (1978) found that they could survey the ar- thropod fauna on soybean from seedling emergence through physiolog- ical maturity by examining single plants; it was not necessary to remove any insects from the populations. Whole plant examination is the only method that allows correct positioning of the sampled organisms in the various strata of the canopy; and trophic rela- tionships such as prey eaten by predators can be also observed and recorded (Kogan and Pitre 1980).
On the other hand, whole plant examination is dependent on the sampler's personal ability and visual acuity, factors that make it difficult to compare data gathered by different samplers. Moreover, the perception of small moving objects is greatly affected by wind. (Most sampling methods may be affected by wind, but direct counts are virtually impossible in winds above 8 mi. per hour.) An addi- tional disadvantage is the chance for misidentification of certain species. Because the sampler quite often has only a glimpse of the sampled specimen, positive identification is often impossible (Kogan and Pitre 1980).
The simplest way to convert counts of arthropods on single plants to absolute populations is to multiply the counts per plant by the number of plants per unit of area. In crops like cotton and soybean, however, this cannot be done because the plants do not oc- cur as discrete units in the crop space. Their branches and leaves crisscross, and plants in the late vegetative growth stages usually form a continuous canopy, at least along the rows. For this reason, it is difficult to isolate plants for sampling purposes (Kogan and Pitre 1980). Workers in Mississippi have used hills of cotton in- stead of single plants because of the variability of plants within a stand of cotton. Unfortunately, there tend to be one to two dom- inant plants in hills of cotton, which can cause sampling problems

310 J.W.SMITH ETAL
if the counts for these plants are not averaged out with those for the less dominant plants.
Portable tape recorders have been used in several Mississippi studies for recording data. This procedure facilitates whole plant examinations by leaving both hands of the sampler free.
Ground Cloth
The ground cloth sampling method is also known as the beat cloth, drop cloth, shake cloth, or plant shake method. The arth- ropods are forcefully displaced from the plants onto a sheet spread on the ground between two adjacent rows of plants so the specimens can be collected or counted (Kogan and Pitre 1980).
The ground cloth cannot be used to sample insects that will escape rapidly although it is an adequate technique for species that drop to the ground when disturbed. It is an excellent method for most lepidopterans (caterpillars), Chrysomelidae, Pentatomidae (particularly nymphs), Nabidae, and other insects with slow escape reactions. The method also has limited application when plants are small and becomes rather inefficient when plants become senescent and shed their leaves. On the other hand, results obtained by dif- ferent researchers can be reasonably consistent because the main factor affecting the procedure is the rigor of the shaking action. Differences resulting from different samplers, however, should not be disregarded (Marston et al. 1979).
Sweep Net
For over a century, the sweep net has been the most widely used tool for sampling arthropods on small grain, forage, and many row crops. This is because no other method permits the capture of as many insects from vegetation per man hour at less cost and with- out damage to the crop (Ruesink and Kogan 1975). However, there are difficulties in quantifying and comparing data from sweep net sampling that stem from the many styles of sweeping though there are also some general procedures that may help one achieve a cer- tain degree of standardization (Ruesink and Haynes 1973, Jenson 1976).
Also, there are difficulties of other kinds. Delong (1932) was among the first to study the efficiency of the sweep net as a tool in insect population studies on some field crops. He recog- nized that environmental factors such as temperature, humidity, wind velocity, position of the sun, plant size, density of the canopy, and pubescence of leaves and stems are responsible for variability of results. If the foliage is wet with rain or dew, it becomes very difficult to operate the sweep net, although this is also true for most sampling techniques.

SAMPLING ARTHROPODS IN COTTON 311
Suction Net
Devices that use suction to remove arthropods from vegetation have been used for some time. One of the best known is the D-Vac suction net (Dietrick et al. 1959, Dietrick 1961).
Davis et al. (1966) compared the mean number of Miridae earlier sampled by visual in situ whole plant examination with the number collected by a D-Vac vacuum sampler and concluded that the visual technique was superior. Similar results were obtained by Shepard and Sterling (1972a) who were sampling for predaceous arthropods that were abundant on cotton. Callahan et al. (1966) concluded that the sweep net technique was superior to the D-Vac technique for collecting some insects that affect large plants.
On the other hand, Richmond and Graham (1969) compared methods of operating the D-Vac. Shepard and Sterling (1972b) showed that the D-Vac was the most economical method of obtaining reliable and repeatable results. Schuster and Boling (1974) used D-Vac sampling to determine phenology of both predatory and phytophagous cotton field insects.
J. W. Smith (unpublished data 1972) compared the results of visual whole plant and D-Vac sampling of cotton arthropods in Mis- sissippi. He concluded that no one method will give accurate popu- lation estimates for each of a diversified group of arthropods and that sampling techniques must be suited to the insect population (Pieters and Sterling 1973).
Studies by Harding and Dupnik (1976) and Fuchs and Harding (1976) conducted in the Lower Rio Grande Valley of Texas involved extensive D-Vac sampling.
Smith et al. (1976) included the D-Vac as one of the tech- niques compared in sampling populations of beneficial arthropods associated with cotton.
Pieters and Sterling (1973) compared the D-Vac and whole plant examination and stated that neither sampling method was more relia- ble nor more accurate than the other for coleopterans. However, the D-Vac nearly always provided a lower C.V. (Coefficient of Vari- ation) and a higher x (standard error of the mean) for Hemiptera and Homoptera than whole plant examination provided. They also found that the D-Vac method was more reliable than whole plant ex- amination for sampling arachnids.
Devices rather like the D-Vac but much larger, some mounted on tractors, have been used to sample cotton insects. Some of the early ones were developed originally as control measures, primarily against the boll weevil (Anthonomus grandis Boheman). However, Parencia (1968), in field tests conducted in 1944, found that a collecting machine (Webb) captured only 40% as many boll weevils as

312 J.W.SMITH ETAL
could be located visually; and M. E. Merkl, E. P. Lloyd, and E. C. Burt (unpublished data 1966) found that the Nisbet bug catcher, an almost identical machine, captured ca. 50% of the weevils that had been found and marked by a field crew. Later, McCoy and Lloyd (1975) developed the McCoy Insect Collector (a dual directional blower and vacuum system) for use in detecting low level infesta- tions of boll weevils. This device collected 130% more weevils than could be found by visual inspection.
Kogan and Pitre (1980) found that the suction net, like the sweep net, is operated in different ways by different researchers. Also, it is advisable to use the suction net in certain ways for specific conditions. Thus the interpretation of the results vary, just as with the sweep net. Since no one procedure is best for sampling all arthropods, preliminary tests should be made with the suction net to determine which procedure yields the most desirable results.
COMPARISON OF EFFICIENCY OF SAMPLING TECHNIQUES
Four sampling techniques were compared during one season (1978-79) of the Boll Weevil Eradication Trial (BWET) and the Opti- mum Pest Management Trial (OPMT). The unit area sampler was the standard for comparing the efficiency of the other three methods. The sweep net and the D-Vac methods were both included; the fourth method consisted of field observations by an entomologist with wide faunal experience who walked through a field and recorded numbers and species of all arthropods observed. Ten unit area samples were taken on each date in each sample field. D-Vac sampling (10 sam- ples in each field) was done with a 10-inch collecting cone by vac- uuming the plants from one side and from top to bottom while the operator moved along 10 feet of row.
Results are presented as ratios. For example.
Estimated numbers of arthropods per acre collected by D-vac = D/U Estimated numbers of arthropods per acre collectai
by unit area sampler
Variations in the ratios between months were greater in Mis- sissippi than in North Carolina, the two areas sampled. However, the season average of 0.50 was the same for both States.
The D/U*s for total important arthropods—after several numer- ous and less important species such as Thripidae had been removed from consideration—were as follows:

SAMPLING ARTHROPODS IN COTTON 313
Mi ssissippi North Carolina Number Number Number Number
Sampling of of of of period fields samples D/U fields samples D/U
June 3 30 0.69 6 60 0.60 July 6 60 0.31 6 60 0.44 August 6 60 0.47 3 50 0.50 Total or averag( e 15 150 0.49 15 170 0.51
The D/U*s of arthropods known to be predaceous or parasitic were as follows:
M ississippi North Carolina Number Number Number Number
Sampling of of of of period fields samples D/U fields samples D/U
June 3 30 0.70 6 60 0.99 July 6 60 0.70 6 60 0.78 August 6 60 0.63 3 50 0.63 Total or averag( B 15 150 0.68 15 170 0.80
The D-Vac sampler was thus more efficient as a sampling device for important species of predators and parasites. It was less ef- ficient when the entire insect population was considered.
Based on D/U's for Mississippi and North Carolina combined and on some extrapolations (numbers in parentheses) where less well sampled groups had similar behavior patterns with corresponding better sampled groups, the following ratios were developed:
Arthropod Group D/U Arthropod Group D/U
^ithonomus grandis Boheman Lathridiidae Conoderus vespertinus (F.) Systena spp. Chaetocnema Diabrotica undecimpunctata
howardi Barber
(0.50) Trialeurodes abutilonea (0.83) (Haldeman) 0.41 0.90 Aphidae 0.30 (0.48) Braconidae 0.50 (0.91) Chalcidoidea 0.36
Scelionidae (0.36) 0.34 Mjnmaridae (0.40)

314 J.W.SMITH ETAL
Arthropod Group D/U Arthropod Group D/U
Anthicidae 0.85 Coccinellidae 0.43 Phalacridae (0.43) Orthoperidae (0.83) Heliothis spp. (0.50) Pseudatomoscelis seriatus
(Reuter) (1.0) Lygus líneolarís (Palisot
de Beauvois) (1.0) Neurocolpus nubilus (Say) (1.0) Geocoris punctipes (Say) (1.0) Nabidae 0.66 Orius sp. 0.56 Fulgoroidea 0.44
Cicadellidae 0.36 Formicidae 0.94 Apis mellifera L. 0.08 Halictidae (0.17) Orthoptera (Saltatoria) (0.07) Thripidae 0.54 Psocoptera (0.40) Dolichopodidae 0.17 Chloropidae 0.17 Collembola (1.0) Araneida 1.0 Acariña (0.33) Other miscellaneous Arthropoda 0.47
Collections of beneficial arthropods from cotton with all four sampling methods are compared in Figure 1. The D-Vac sampler, in addition to providing an accurate estimate, provided an excellent system for collecting, counting, and careful identification of in- festing species.
SAMPLING MAJOR PEST SPECIES IN COTTON
Heliothis spp.
Heliothis zea (Boddie) and Heliothis virescens (F.) have be- come two of the most serious pest insects in the South, and both species now seriously limit the profitability of cotton production in many areas (Warren 1979).
Sampling of Heliothis spp. eggs on cotton, other row crops, or wild host plants is accomplished most efficiently through direct, visual examination. However, absolute density of eggs is difficult to determine because of the egg size and the large plant surface area available for oviposit ion. Relative estimates of egg abun- dance are, therefore, obtained by examination of the highly attrac- tive terminal foliage. However, numbers change rapidly depending on oviposit ion, prédation, and hatching, and these conditions should be considered in making any judgments (Stinner et al. 1980).
Sampling of Heliothis spp. larvae on cotton is most successful when the terminal area, squares, blooms, and bolls (single plant on linear row) are examined visually. Other methods such as sweep net, suction devices, and ground cloth are less satisfactory because the larvae are frequently inside the fruiting structure. Although, first-instar larvae are difficult to see and most estimates are biased toward the later instars, numbers of large larvae collected from squares and bolls usually provide good estimates of absolute numbers.
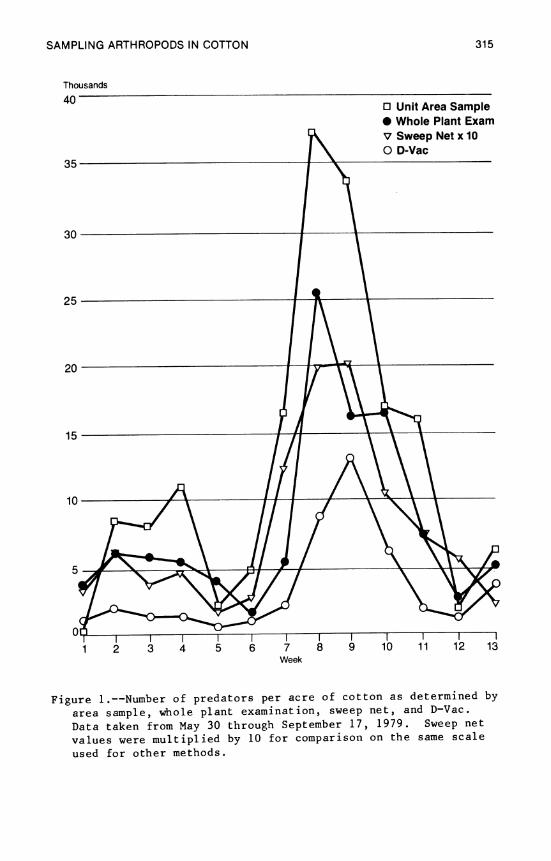
SAMPLING ARTHROPODS IN COTTON 315
Thousands
40 D Unit Area Sample • Whole Plant Exam V Sweep Net x 10 O D-Vac
Figure 1.—Number of predators per acre of cotton as determined by area sample, whole plant examination, sweep net, and D-Vac. Data taken from May 30 through September 17, 1979. Sweep net values were multiplied by 10 for comparison on the same scale used for other methods.

316 J.W.SMITH ETAL
Pupal sampling is difficult but has, nonetheless, been accom- plished by soil sifting, soil scraping, and estimating numbers of prepupae that have emerged from fruiting forms (Stinner et al. 1980). Soil sifting provides the most precise information but is quite expensive in both time and labor required. Another technique, determination of the emergence of adult moths from various crops, requires the use of soil covers or cages (Roach and Ray 1976). This method, though it provides a good assessment of the size and source of the moth generation, requires considerable labor and is, at best, an indirect measure of pupal abundance.
Continuous sampling of populations of adult Heliothis spp. during their active period may be one of the best ways to follow and forecast the time and size of egg and larval populations (Hart- stack et al. 1979). Adults may be sampled by making visual counts at night, by disturbing the moths during sampling, or by using "sugar lines" to attract moths. However, light and pheromone traps are used most commonly to survey for populations of adults. The use of traps has recently been extensively reviےwed by Hartstack et al. (1979). Two machines that have been used for area sampling of adult populations of Heliothis spp. are the Nisbet Insect Collector (Parencia 1968) and the McCoy Insect Collector (McCoy and Lloyd 1975) mentioned previously for control or sampling of boll weevils.
Southwood (1966) reviewed the mark, release, and recapture method used with traps in estimating populations and in dispersal studies. This method is now being employed in studies of 11. vir- escens dispersal in Mississippi.
Boll Weevil
The boll weevil, a key pest of cotton, has four stages: egg, larva, pupa, and adult. When conditions are favorable, a life cy- cle is completed in 2.5 or 3 weeks. Thus, in the extreme southern part of the Cotton Belt, there may be as many as seven generations during a crop season (Parencia 1978). In the spring, females lay eggs singly in cotton squares. Late in the season, eggs are laid both in squares and in young bolls. These eggs develop inside the square or boll and hatch in 3-5 days. Then the larvae feed 7-12 days inside the square or boll. After a 3- to 5-day pupal stage, adults cut their way out. The adults feed, mate, and begin depos- iting eggs in 3-7 days. This sequence continues until the plant is killed in the fall by cold weather. At about that time, adult wee- vils of the last generation move to ground trash or other suitable habitat where they overwinter in a physiological condition called diapause. Sampling for overwintering weevils is, therefore, nor- mally accomplished by collecting ground trash from the hibernation sites and sifting it through a series of sieves. The separated material containing the weevils is then placed on heat tables where observers collect the weevils as they become active and crawl from the trash.

SAMPLING ARTHROPODS IN COTTON 317
Overwintered weevils are most effectively detected by setting out pheromone traps in the spring. Sampling of subsequent genera- tions of adult weevils in cotton fields may be accomplished by the shag method (a modified ground cloth technique), by vacuum sweep- ing, or by visual inspection of the squares, blooms, and bolls. Most cotton entomologists prefer visual inspection of fruiting forms.
Also, the tractor-mounted collecting devices discussed earlier (Nisbet and McCoy Insect Collectors) have been used extensively in research situations for sampling adult weevils (Parencia 1968, McCoy and Lloyd 1975).
Eggs and larvae of the boll weevil can be sampled by collec- ting fruiting forms that show signs of oviposition punctures or that have been shed from the cotton plant. (in most surveys made to de- termine economic thresholds, indices such as feeding and oviposi- tion are used to assess levels of boll weevil populations.)
Plant Bugs
The tarnished plant bug (Lygus lineolaris (Palisot de Beau- vois)) is often considered a key pest in cotton because injurious numbers can occur during the early fruiting stage. The insecti- cides that are then applied for control may drastically reduce the numbers of predatory arthropods and thus leave the cotton crop vul- nerable to subsequent infestations of the bollworm and the tobacco budworm.
The results of special research conducted during the OPMT and the BWET in which five sampling methods (ground cloth, visual in- spection, sweep net, D-Vac, and unit area sampler) were compared in 1977, 1978, 1979, and 1980 showed that the D-Vac was an excellent device for collecting JL. lineolaris. Three times more of these plant bugs were collected by the D-Vac method than by sweep net, and five times more were collected by D-Vac than by visual inspection.
In general, tarnished plant bugs are similar in biology to the hemipteran predators that occur in cotton. Therefore, methods of sampling for these bugs are identical to those used to sample for other hemipteran species.
Secondary Pests
Sampling techniques for the secondary pests of cotton (thrips, leafhoppers, Aphididae, Aleyrodidae, and Tetranychidae) were dis- cussed by Kogan and Herzog (1980). The earlier discussions in this chapter on general arthropod sampling apply to this group of occa- sionally important cotton arthropods.

318 J.W.SMITH ETAL
SAMPLING BENEFICIAL ARTHROPODS IN COTTON
Cotton fields contain a substantial entomophagous fauna. Indeed, it would not be economical to produce cotton if parasitic and predaceous species did not suppress the pest complex (Reynolds 1976).
An estimated 400-500 arthropod species are routinely associ- ated with southern cotton fields. The surveys of predatory arthro- pods in Arkansas cotton made by Whitcomb and Bell (1964) revealed about 600 species associated with the crop, not including parasites.
Since the sampling of such arthropods requires basic knowledge of the biology and habitats of the parasites and predators involved, techniques must often be modified or devised.
Predators are free living throughout their life cycle and are capable of feeding on almost any prey they can capture. Also, some predators have both phytophagous and entomophagous habits. Para- sites are usually specific for one insect species or for a closely related group, and during their larval period, they are restricted to and dependent upon an individual host species. Thus, sampling for parasites may mean sampling the host(s) or sampling the para- site directly. Predators, however, must be sampled directly, re- gardless of the habitat or diversity of prey in the habitat.
Several workers have made extensive comparisons of sampling methods for predatory species in cotton (Leigh et al. 1970, Smith et al. 1976). Most of these techniques have been discussed earlier in this chapter under Techniques for Sampling Arthropods in Cotton Vegetation.
Experience in sampling for individual predators and groups of predators has shown that some techniques are especially useful.
Orius insidiosus (Say), described by Barber (1936) as the most important predator of Heliothis zea eggs, was the most numerous predator in a 1974 Mississippi study (Smith et al. 1976). It was also the most numerous predator found in the OPMT and BWET areas (Figure 2). However, the insect is so small, ca. 1.6 mm long, that sampling is difficult. Also, this predator typically lives in the terminal buds and is, therefore, not readily visible to direct ob- servation. However, the D-Vac machine has proved to be effective in sampling this insect because the plant terminal can be entirely covered by the collecting cone while the suction dislodges both im- mature and adult Orius. (Sweep nets and ground cloths are less ef- ficient.) Also, the D-Vac collected ca. 65% as many Orius as did the unit area sampler in the OPMT and BWET areas.
A big-eyed bug (Geocoris punctipes (Say)) was the most common predator in 3 of 4 years in the OPMT in Mississippi. Populations fluctuated from year to year, but peaks usually occurred from
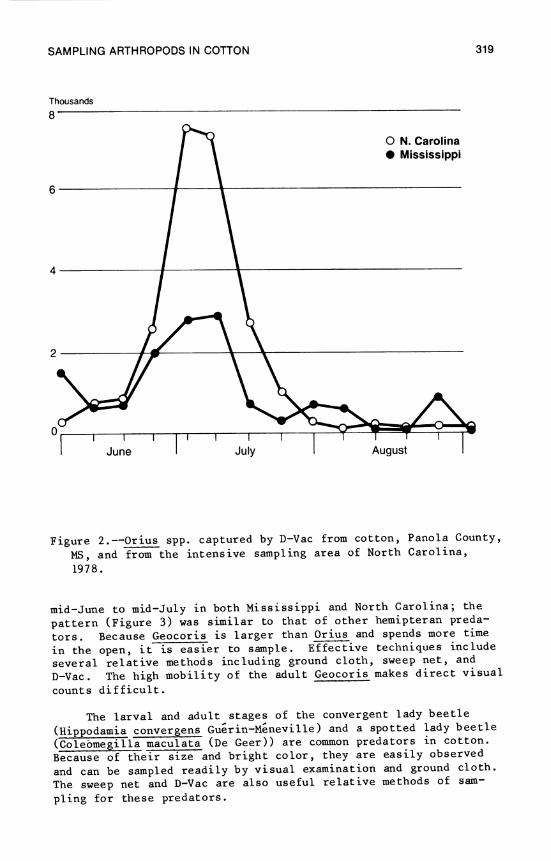
SAMPLING ARTHROPODS IN COTTON 319
O N. Carolina • Mississippi
Figure 2.—Orius spp. captured by D-Vac from cotton, Panola County, MS, and from the intensive sampling area of North Carolina,
1978.
mid-June to mid-July in both Mississippi and North Carolina; the pattern (Figure 3) was similar to that of other hemipteran preda- tors. Because Geocoris is larger than Orius and spends more time in the open, it is easier to sample. Effective techniques include several relative methods including ground cloth, sweep net, and D-Vac. The high mobility of the adult Geocoris makes direct visual counts difficult.
The larval and adult stages of the convergent lady beetle (Hippodamia convergens Guerin-Meneville) and a spotted lady beetle (Colebmegilla maculata (De Geer)) are common predators in cotton. Because of their size and bright color, they are easily observed and can be sampled readily by visual examination and ground cloth. The sweep net and D-Vac are also useful relative methods of sam- pling for these predators.

320 J. W. SMITH ET AL
Thousands
8
O N. Carolina # Mississippi
Figure 3.—Geocoris spp. populations collected by D-Vac from cot- ton, Panola County, MS, and from the intensive sampling area of North Carolina, 1978.
Adult Chrysopa spp. (primarily Chrysopa carnea Stephens) move into cotton quite early in the season (Smith et al. 1976), but re- production in cotton does not become evident until midseason. Sev- eral workers have documented the effectiveness of C. carnea adults and larvae as predators of Heliothis spp. (Quaintance and Brues 1905, Whitcomb and Bell 1964, Ridgway et al. 1967, van den Bosch et al. 1969).
Several nabid species (primarily Reduviolus roseipennis (Reuter)) are common in cotton and are excellent predators of small Heliothis spp. larvae. Both adult and nymphal nabids are sampled well with suction devices, sweep nets, and ground cloths.
All spiders are predaceous and, for the most part, show no discrimination in the taking of prey; thus, most are just as likely to prey on beneficial insects as on pest species. A striped lynx spider (Oxyopes salticus Hentz), so-called for its rapid chase of prey, is the most commonly observed species of spider on cotton. Spiders are quite easy to sample, and suction devices are usually

SAMPLING ARTHROPODS IN COTTON 321
satisfactory for collecting them. However, there are factors to consider such as time of day (many spiders are nocturnal) and time of year (many spider species are mature for only a few weeks out of the year, and immature spiders can seldom be identified as to spe- cies). Prey capture methods also differ. Some spiders are active hunters (Oxyopidae, Lycosidae, Pisauridae, Thomisidae, Salticidae); some are ambushers (Oxyopidae, Thomisidae), and many use a web to capture prey (Araneidae, Theridiidae, Dictyniodae, Linyphiidae, Tetragnathidae, Agelenidae). Vertical stratification likewise dif- fers (different species prefer different strata of the cotton can- opy). An excellent review of sampling techniques for spiders can be found in Kogan and Herzog (1980).
SAMPLING ARTHROPODS IN THE OPTIMUM PEST MANAGEMENT AND BOLL WEEVIL ERADICATION TRIALS
The North Carolina BWET and the Mississippi OPMT were con- ducted concurrently from 1978 to 1980. (Reports on these trials are found in Chapters 15 and 16 of this handbook.) In addition to the implementation of the BWET by the Animal and Plant Health In- spection Service and the OPMT by the Mississippi Cooperative Exten- sion Service, two research teams composed of personnel from Agri- cultural Research Service (ARS), U.S. Department of Agriculture collected extensive biological data on cotton and arthropods asso- ciated with cotton production in the test area. These data were then used in estimating and comparing the biological impact of the two programs and in evaluating the economic and environmental im- pacts. To collect representative data, the teams sampled ca. 193 cotton fields each year (219, 206, and 155 in 1978, 1979, and 1980, respectively) in North Carolina and 96 cotton fields each year in Mississippi. The number of fields sampled in North Carolina was greater because of the larger geographic area involved in the BWET. All fields were sampled at least once a week. A selected number of fields were sampled twice weekly to provide intensive monitoring data.
Because of the geographic separation and the large number of fields, accurate and consistent sampling procedures were a critical element in the evaluation of these Trials. Also, there was to be direct comparison between the Trial areas and nearby Current Insect Control practice areas. Therefore, the types of data collected and the sampling procedures were identical at all locations.
The primary methods of arthropod sampling were direct plant examinations and suction net (D-Vac). Plant examination was used to sample pest populations only. The suction net method was used to sample a broad spectrum of arthropods including pest, beneficial, and incidental species; it was chosen over other methods because the variability of sample quality due to sampler is less. This was important because so many individuals were involved in the sampling process.

322 J. W. SMITH ET AL
Pest Arthropods
Arthropod pests of cotton were sampled by examining the whole cotton plant or selected portions of the plant depending on plant size and the pest species being sampled. Numbers of boll weevils (including egg and oviposit ion punctures), bollworms/budworms, fleahoppers, plant bugs, aphids, spider mites, thrips, and white- flies were recorded. Other observed pests such as cutworms (Noctu- idae) and corn borers (Pyralidae) were also recorded. All species except aphids, spider mites, and thrips were counted and recorded as numbers found per row-foot. Populations of the excepted species were recorded as light—found on occasional plants; medium—found on numerous plants with slight damage; and heavy—found on numerous plants with damage easily observed.
The whole plant survey was the first pest sampling implemented during the season and was typically initiated 2 weeks before the cotton plant developed squares. Every cotton plant was examined in five randomly located 25 row-foot sections, a total of 125 row-feet per field. As plants grew larger and substantial square popula- tions developed, the whole plant survey was replaced with square, boll, and terminal surveys where insect pest activity was centered.
The terminal survey was conducted in the same way as the whole plant survey, in five randomly located 25 row-foot sections. How- ever, only terminals were examined. This technique was continued throughout the remainder of the season since actively growing ter- minals are an ideal site for bollworm/tobacco budworm oviposition and for buildup of aphid populations.
The square survey was initiated when squares reached approxi- mately 0.25 inch in diameter. A total of two hundred 0.25-inch or larger squares from all parts of the fields was examined. The top, middle, and lower portions of the plant were sampled. Since boll weevil egg and feeding punctures tend to be concentrated on squares, and bollworm/budworm eggs are frequently deposited on the outside of the bract (hatching larvae crawl inside and feed on the square, often at the protected base), the square survey was continued until the square population declined so much that collection of 200 rep- resentative squares from a field became difficult.
The boll survey was similar to the square survey: 200 bolls collected from throughout a field were examined. This survey was initiated shortly after blooming and continued throughout the season. Boll weevil oviposition and feeding punctures are found on young bolls as well as on squares. Bollworm/budworm larvae are especially important at the boll stage since the plant has less time to replace a lost or damaged boll than it has to replace dam- aged squares.

SAMPLING ARTHROPODS IN COTTON 323
Beneficial Arthropods
Beneficial arthropods were sampled with a suction net machine (D-Vac). Weekly samples were collected in each survey field from four randomly located 10 row-foot sections, and some fields were sampled twice weekly to provide more intensive data. In this case, the 10 row-foot section was marked, and the second collection was made two rows over along a row that had not been disturbed.
As the sampling of each 10 row-feet was completed, a location- date-time tag was dropped into the bag. The bag was then removed from the suction machine and closed with a rubber band. When the sampling had been completed, the bags were tied together and placed in an ice chest containing dichlorvos (Shell No-Pest Strip). The day's samples were transferred to a freezer and delivered to the laboratory for identification. The combination of dichlorvos and cold temperature prevented additional prédation in the sample bags.
A combined total of over 100 miles of cotton row was sampled by the suction net machine in the BWET and OPMT. The collected arthropods were all identified, and representative collections were established at both the Mississippi and North Carolina laboratories and at the Agricultural Research Service cotton insect collection at Mississippi State University in the Mississippi Entomological Museum. All specimens collected during 1980 were preserved for fu- ture reference and study.

324 J.W.SMITH ETAL
LITERATURE CITED
Anderson, F. S. 1965. The negative binomial distribution and the sampling of in-
sect populations. Proceedings, XII International Congress of Entomology, 395 p.
Barber, G. W. 1936. Method of rearing corn earworm larvae. Journal of Eco-
nomic Entomology 29:1175-1176.
Byerly, K. F., A. P. Gutierrez, R. E. Jones, and R. F. Luck. 1978. A comparison of sampling methods for some arthropod popu-
lations in cotton. Hilgardia 46:257-282.
Callahan, R. A., F. R. Holbrook, and F. R. Shaw. 1966. A comparison of sweeping and vacuum collecting certain in-
sects affecting forage crops. Journal of Economic Entomology 59:478-479.
Clark, L. R., P. W. Geier, R. D. Hughes, and R. F. Morris. 1967. The ecology of insect populations in theory and practice. Methven and Co., Ltd., London.
Davis, J. W., W. C. Watkins, Jr., C. B. Cowan, Jr., R. L. Ridgway, and D. A. Lindquist.
1966. Control of several cotton pests with systemic insecti- cides. Journal of Economic Entomology 59:159-162.
Delong, D. M. 1932. Some problems encountered in the estimation of insect pop-
ulations by the sweeping method. Annals of the Entomological Society of America 25:13-17.
Dietrick, E. J. 1961. An improved backpack motor fan for suction sampling of in-
sect populations. Journal of Economic Entomology 54:394-395.
E. I. Schlinger, and R. van den Bosch. 1959. New method for sampling arthropods using a suction collec-
ting machine and modified Berlese funnel separator. Journal of Economic Entomology 52:1085-1091.
Fuchs, T. W., and J. A. Harding. 1976. Seasonal incidence of arthropod predators in various habi-
tats in the Lower Rio Grande Valley of Texas. Environmental Entomology 5:288-290.
Gutierrez, A. P., L. A. Falcon, W. Loew, P. A. Leipzig, and R. van den Bosch.
1975. An analysis of cotton production in California: a model for Acala cotton and the effects of defoliators on its yields. Environmental Entomology 4:125-136.

SAMPLING ARTHROPODS IN COTTON 325
Harcourt, D. G. 1969. The development and use of life tables in the study of
natural insect populations. Annual Review of Entomology 14: 175-196.
Harding, J. A., and T. D. Dupnik. 1976. Associations among arthropods in a D-Vac sample from Rio Grande Valley cotton. Southwestern Entomologist 1:133-135.
Hartstack, A. W., D. E. Hendricks, J. D. Lopez, E. A. Stadelbacher, J. R. Phillips, and J. A. Witz.
1979. Economic thresholds and sampling of Heliothis species on cotton, corn, soybean, and other host plants. Southern Co- operative Series, Bulletin No. 231, p. 105-131. W. L. Ster-
ling, editor.
Healy, M. J. R. 1962. Some basic statistical techniques in soil zoology. l£ Progress in Soil Zoology, p. 3-9. P. W. Murphy, editor. But- terworths, Ltd., London.
Jenson, R. L. • o A ♦_ 1976. Are you doing a good job of sweeping the field? Agri
fieldman 32:31-32, 34.
Karandiros, M. G. ^ 1976. Optimum sample size and comments on some published formu
lae: pest management project. Bulletin of the Entomological Society of America 22:417-421.
Kogan, M., and D. C. Herzog, editors. 1980. Sampling methods in soybean entomology. Springer-Verlag,
Incorporated, New York.
and H. N. Pitre, Jr. 1980. General sampling methods for above-ground populations of
soybean arthropods. In Sampling Methods in Soybean Entomology. M. Kogan, and D. C. Herzog, editors. Springer-Verlag, Incor-
porated, New York.
Leigh, T. F., D. Gonzalez, and R. van den Bosch. 1970. A sampling device for estimating absolute insect popula-
tions in cotton. Journal of Economic Entomology 63:1704-1706.
McCoy, J. R., and E. P. Lloyd. ^ , n 1975. Evaluation of airflow systems for the collection of boll weevils from cotton. Journal of Economic Entomology 68:49-52.
MacLeod, J. r i • • 1958. The estimation of numbers of mobile insects from low inci-
dence recapture data. Transactions of the Royal Entomological Society of London 110:363-392.

326 J. W. SMITH ET AL
Marston, N. L., W. A. Dickerson, W. W. Ponder, and G. D. Booth. 1979. Calibration ratios for sampling soybean Lepidoptera: ef-
fect of larval species, larval size, plant growth stage, and individual sampler. Journal of Economic Entomology 72:110-114.
Mayse, M. A., M. Kogan, and P. W. Price. 1978. Sampling abundance of soybean arthropods: comparison of
methods. Journal of Economic Entomology 71:135-141.
Morris, R. F. 1955. The development of sampling techniques for forest insect
defoliators, with particular reference to the spruce budworm. Canadian Journal of Zoology 33:225-294.
1957. The interpretation of mortality data in studies of popula- tion dynamics. Canadian Entomologist 89:49-69.
1960. Sampling insect populations. Annual Review of Entomology 5:243-264.
Mott, D. G. 1963. The population model for the unsprayed area. Memoirs of
the Entomological Society of Canada 31:99-109.
Parencia, C. R., Jr. 1968. Control of cotton insects with an insect-collecting ma-
chine. Journal of Economic Entomology 61:274-279.
1978. One hundred twenty years of research on cotton insects m the United States. U.S. Department of Agriculture, Agriculture Handbook No. 515, 75 p.
Pieters, E. P., and W. L. Sterling. 1973. Inferences on the dispersion of cotton arthropods in Tex-
as, USA. Environmental Entomology 2:863-867.
Quaintance, A. L., and C. T. Brues. 1905. The cotton bollworm. U.S. Department of Agriculture, Bu-
reau of Entomology Bulletin No. 50, 155 p.
Reynolds, H. T. 1976. Insect pest management on cotton in the United States. Proceedings of US-USSR Symposium: Integrated Control of Ar- thropod Disease and Weed Pest of Cotton, Grain Sorghum and Deciduous Fruit. Texas Agricultural Experiment Station, Miscellaneous Publication No. 1276, p. 123-134.

SAMPLING ARTHROPODS IN COTTON 327
Richmond, C. A., and H. M. Graham. 1969. Two methods of operating a vacuum sampler to sample popu-
lation of the cotton fleahopper on wild hosts. Journal of Economic Entomology 62:525-526.
Ridgway, R. L., P. D. Lingren, C. B. Cowan, Jr., and J. W. Davis. 1967. Populations of arthropod predators and Heliothis spp. af-
ter applications of systemic insecticides to cotton. Journal of Economic Entomology 60:1012-1016.
Roach, S. H., J. W. Smith, S. B. Vinson, H. M. Graham, and J. A Harding.
1979. Sampling predators and parasites of Heliothis species on crops and native host plants. Southern Cooperative Series, Bulletin No. 231, p. 132-141. W. L. Sterling, editor.
and L. Ray. 1976. Pattern of emergence of adult Heliothis from fields
planted to cotton, corn, tobacco, and soybeans. Environmental Entomology 5:628-630.
Ruesink, W. G., and D. L. Haynes. 1973. Sweep net sampling for the cereal leaf beetle, Oulema melanopus. Environmental Entomology 2:161-172.
and M. Kogan. 1975. The quantitative basis of pest management: Sampling and measuring. Tn Introduction to Insect Pest Management, p. 309- 351. R. L. Metcalf and W. H. Luckmann, editors. John Wiley and Sons, New York.
Schuster, M. F., and J. C. Boling. 1974. Phenology of early mid-season predatory and phytophagous
insects in cotton in the lower Rio Grande Valley of Texas. Texas Agricultural Experiment Station, Miscellaneous Publica- tion No. 1133, 31 p.
Shepard, M., and W. L. Sterling. 1972a. Effects of early season applications of insecticides on beneficial insects and spiders in cotton. Texas Agricultural Experiment Station, Miscellaneous Publication No. 1045, 14 p.
and W. Sterling. 1972b. Incidence of parasitism of Heliothis spp. (Lepidoptera: Noctuidae) in some cotton fields of Texas. Annals of the Ento- mological Society of America 65:759-760.
Smith, J. W., E. A. Stadelbacher, and C. W. Gantt. 1976. Comparison of techniques for sampling beneficial arthropod
populations associated with cotton. Environmental Entomology 5:434-444.

328 J. W. SMITH ET AL
Southwood, T. R. E. 1966. Ecological methods, with particular reference to the study
of insect populations. Chapman and Hall, London EC4.
1978. Ecological methods. Halsted Press, New York.
Sterling, W. L., editor. 1979. Economic thresholds and sampling of Heliothis species on
cotton, corn, soybeans, and other host plants. Southern Coop- erative Series, Bulletin No. 231, 159 p.
Stinner, R. E., J. R. Bradley, and J. W. VanDuyn. 1980. Sampling Heliothis spp. on soybeans. Iii Sampling Methods
in Soybean Entomology, p. 407-421. M. Kogan and D. C. Herzog, editors. Springer-Verlag, Incorporated, New York.
Strickland, A. H. 1961. Sampling crop pests and their hosts. Annual Review of Entomology 6:201-220.
van den Bosch, R., T. F. Leigh, D. Gonzalez, and R. E. Stinner. 1969. Cage studies on predators of bollworm in cotton. Journal
of Economic Entomology 62:1486-1489.
Varley, G. C, and G. R. Gradwell. 1971. The use of models and life tables in assessing the role of
natural enemies. In Biological Control, p. 93-112. C. B. Huf- faker, editor. Plenum Press, New York.
Warren, L. 0. 1979. Economic thresholds and sampling of Heliothis species on
cotton, corn, soybeans, and other host plants. W. L. Sterling, editor. Southern Cooperative Series, Bulletin No. 231, lip.
Waters, W. E., and W. R. Henson. 1959. Some sampling attributes of the negative binomial distri- bution with special reference to forest insects. Forest Sci- ence 5:397-412.
Watt, K. E. 1966. System analysis in ecology. Academic Press, New York and London.
Whitcomb, W. H., and K. Bell. 1964. Predaceous insects, spiders and mites of Arkansas cotton
fields. University of Arkansas, Agricultural Experiment Station, Bulletin No. 690, 84 p.

329
Chapter 13
LOSSES IN YIELD OF COTTON DUE TO INSECTS
P. H. Schwartz National Program Staff Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705
ABSTRACT Losses in yield of cotton due to insect pests despite best agricultural prac- tices have been estimated. These are compared with losses calculated from research studies that included control plots and treated plots.
The boll weevil (Anthonomus grandis Bohe- man), the bollworm (Heliothis zea (Boddie)), and the tobacco budworm (Heliothis virescens (F.)) have been the predominant pests of cotton during this century and account for the great- est losses. The boll weevil accounted for losses of cotton that were greater than the 1976 value of U.S. production for any one of over 55 other crops.
Losses from cotton pests, if not controlled, were calculated to be: aphids (Aphis spp.), 19%; boll weevil, 51%; cabbage looper (Trichoplusia ni (Hubner)), 92%; cotton fleahopper (Pseudato- moscelis seriatus (Reuter)), 34%; Heliothis spp., 63%; pink bollworm (Pectinophora gossypiella (Saunders)), 61%; and spider mites (Tetranychus spp.), 21%. When these pests are controlled, it was calculated that losses still occur as fol- lows: aphids, 8%; boll weevil, 21%; cabbage looper, 30%; cotton fleahopper, 12%; Heliothis spp., 15%; pink bollworm, 9%; and spider mites, 0.05%.
The amount of loss due to cotton insects is dynamic. Losses due to insects vary from year to year and from area to area. Losses from in- sect damage continue to occur at magnitudes that have not decreased appreciably over the past 30 years. Thus, the advances in insect control technology made during this period have made it possible for us to protect the cotton crop from greater losses while yields have increased sig- nificantly and production practices have changed dramatically.

330 P. H. SCHWARTZ
INTRODUCTION
The guidelines of the U.S. Department of Agriculture for the control of insect and mite pests (Hopkins et al. 1980) contain a list of 22 species of pests in cotton that may be controlled with chemical insecticides. Not all of these pests are seriously damag- ing to the crop at all times. However, experience has indicated that they may require some form of chemical control whenever a dam- aging population develops. My objective here is to bring together the information available in the literature on losses caused by these insect and mite pests (Table 1). This information is lim- ited to losses in either yield or quality and the cost of control- ling the pest. I will not deal with losses to society in general or with losses caused by pest control because of adverse environ- mental impacts.
The section on loss estimates by experts attempts to reflect the losses experienced under the best agricultural practice for the time periods when these estimates were made. The loss estimates calculated from field studies attempt to reflect the potential a pest population has in reducing yield under two regimes—with and without control.
LOSS ESTIMATES BY EXPERTS
One of the earliest reports on the losses caused by insects and mites to agricultural commodities in the United States was that of Marlatt (1904). Mr. Marlatt indicated that the principal insect pests on cotton were the boll weevil (Anthonomus grandis Boheman), the bollworm (Heliothis zea (Boddie)), and the cotton leafworm (Alabama argillacea (Hubner)). The bollworm was chiefly destruc- tive in those southwestern States that were then producing cotton and was damaging 2-60% of the crop with an overall estimated loss of 4% in those States. From 1899 to 1902, the boll weevil caused losses of cotton yield ranging from 40 to 50% in Texas as compared with the zero losses in Louisiana, which was still free from infestation. (Louisiana first experienced heavy losses due to boll weevils in 1903 (Cramer 1967).)
Other experts have also estimated losses in cotton yield due to pests despite best agricultural practices. For the boll weevil, these losses range from 1.4 to 21% (Table 1). Tlie values given by the Cotton Foundation (DeBord 1977) and the U.S. Department of Agriculture Conference Report on Cotton Insect Research and Control (1980) are based on survey data or the best estimates of knowledge- able experts. The estimates by the U.S. Department of Agriculture in 1954 and 1965 are based on statistical surveys and records and on estimates by experts.
However, when estimates are presented on a national basis, as they are in Table 1, they tend to provide an overall average which is somewhat lower than the actual in those States where the pest

LOSSES DUE TO COTTON INSECTS 331
has been a serious problem. For instance, in the U.S. Department of Agriculture Conference Report on Cotton Insect Research and Control (1980), the average for the boll weevil is given as a loss in yield of 1.4%, but during the year of the study (1979), the State of Georgia estimated a 16% loss; Alabama and Louisiana each estimated 4%; Mississippi and South Carolina, 3%; Florida and Texas, each 1.5%; and Arkansas and North Carolina, each 0.05%. Missouri, Tennessee, Virginia, and Oklahoma reported no losses. Thus, the southeastern United States apparently suffered much higher losses from the boll weevil in 1979 than the national average indicates. (it is interesting that North Carolina had only a 0.05% yield loss estimated for 1979 whereas surrounding States had much higher losses. This may be attributed to the boll weevil eradication program in that State.)
This portion of the data in Table 1 therefore indicates that the boll weevil and bollworm have been the predominately important pests on cotton since 1904 and have accounted for the greatest losses. This is not to imply that the other pests listed in Table 1 are not capable of causing serious damage, but sizable infesta- tions would appear to be more localized and sporadic and would thus require control less frequently.
A more detailed analysis of losses attributed by knowledgeable experts to the boll weevil for the various States where cotton is produced is presented in Table 2. Plainly these losses vary from State to State and from year to year (Table 2). Moreover, experts in all the States except Missouri estimate that the boll weevil has great potential for reducing yield if no chemical control is avail- able. In fact, in 1974-76 (DeBord 1977), the boll weevil caused an average annual loss of yield of 201.6 million bales valued at $56.3 million plus control costs of $89.2 million for a total annual loss of $145.5 million. This is equal to an average annual 5.7% of the value of the cotton produced during that period.
The loss attributed by the experts to the boll weevil on cot- ton ($145.5 million) is thus greater than the 1976 value of U.S. production of any one of the following crops:
apricots artichokes asparagus avocados bananas beets broccoli brussels sprouts bush berries cabbage cantaloupe carrots cauliflower celery
cherries coffee cranberries cucumbers dates dry beans eggplant escarole figs filberts flax seed garlic honeydew melon hops
lemon lima beans lime macadamia nuts maple syrup nectarines olives papayas pears peas pecans peppermint peppers persimmons
plums pomegranates popcorn rye seed crops snap beans spearmint spinach tangelos tangerines temples walnuts watermelon
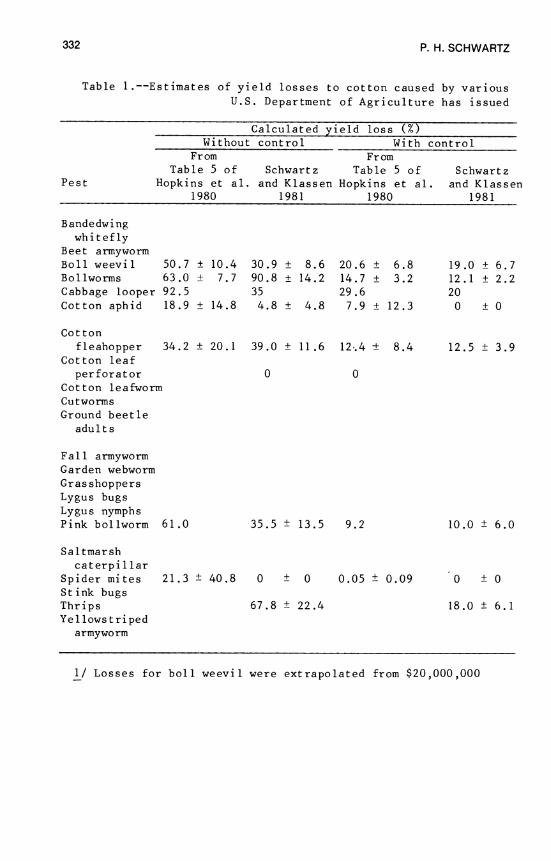
332 P. H. SCHWARTZ
Table 1.—Estimates of yield losses to cotton, caused by various U.S. Department of Agriculture has issued
Calculated y ield loss (%) Without control With control
From From Table 5 of Schwartz Table 5 of Schwartz
Pest Hopkins et al . and Klassen Hopkins et al and Klassen 1980 1981 1980 1981
Bandedwing whitefly
Beet armyworm Boll weevil 50.7 ± 10.4 30.9 ± 8.6 20.6 ± 6.8 19.0 ± 6.7 Bollworms 63.0 ± 7.7 90.8 ± 14.2 14.7 ± 3.2 12.1 ± 2.2 Cabbage looper 92.5 35 29.6 20 Cotton aphid 18.9 ± 14.8 4.8 ± 4.8 7.9 ± 12.3 0 ±0
Cotton fleahopper 34.2 ± 20.1 39.0 ± 11.6 12..4 ± 8.4 12.5 ± 3.9
Cotton leaf perforator 0 0
Cotton leafworm Cutworms Ground beetle
adults
Fall armyworm Garden webworm Grasshoppers Lygus bugs Lygus nymphs Pink bollworm 61.0 35.5 ± 13.5 9.2 10.0 ± 6.0
Saltmarsh caterpillar
Spider mites Stink bugs Thrips Yellowstriped
armyworm
21.3 ± 40.8 0 ± 0
67.8 ± 22.4
0.05 ± 0.09 0 ±0
18.0 ± 6.1
\_l Losses for boll weevil were extrapolated from $20,000,000

LOSSES DUE TO COTTON INSECTS 333
arthropod pests. (All arthropod species are included for which control guidelines (Hopkins et al. 1980))
Estimated loss (%) by experts despite best agricultural practices
Marlatt 1/ Hyslop USDA USDA USDA USDA DeBord USDA (1904) (1938) (1946) (1954) (1965) (1976) (1977) (1980)
0.2
6.7 10.9 8.7 10.1 8.0 8.0 3.0 1.4 4.0 2.0 4.0 4.0 3.3 3.0
2-50 0.2 1.4
0.2 0
0.7 1.4
0.3 0
0.2 0.7
7.5 13 0.2 0.3
loss figure presented in Marlatt (1904).

334 P. H. SCHWARTZ
o
O) U2
TU
(0 (U CO CO O
u CO 0)
Ù0 CO U
CO
(I)
O) 4-) CO 4-> CO
I I
CM
(U
■§ H
1-1 r^
O o :3 cjN o ^
CM|S£)
O r^ PQ <JN 0) rH
0) O M
o CO
Tí 0) 4J CO O
■H 'd tí
CO|.
CMJCM
T) I VJ o o r^
PQ 0^ Q) Q
CM
•H I o o tí r-^ 0 ON O ^
CM|VX>
TS I
o r- PQ ON CU o
Q C/3 ON
4J CO 4-> C/3
O O O C30 CM CM
o en Csl ON
ooooooooo
cM<i-cMcMcMmcnr^cn 00«—tooNv^cMcnoo^t »HrHr^>sDON<}-.—tcnr^
r>. o f*^ CM
CM CO i-H CO ON
m CM CO CO r^ CO r^ in CO <f r-
r-4 m 00 ^f CM r-» -st -H
r^ CM O in sO .-1 in C30
o o 00 ^ o lA CM ^
00 CO COONONCOOOONfH^t^t CM ,-1 rH
<l-in «^r^<i-co<i---H<i-<j-^ C^lr-l C^lv£)COO^D^^-^CMC^^
in ominoocNi o o vo <tOr-HvO<l*cOOOOcOO—lO
►Jís¿H-3<J<:cnOO!^OAX<í <¡'<PL,OIJS:SSZ;OCOHH>
o 00 ON
>-•
(U .^^ v^ <i ;3 r^ 4-> ON
I—< 1—1
tí ^-^ O
• H I—Í U • H Ö0 O
<ö tí tí M-l O O o 4J tí tí . o O) ^^ 4J a r^ 4J 4J r^ O v^ ON o CO r-H Pu v-^ 1—1
0) CO O td tí
IJ O . o -r^ CO FQ 4-1
Oi CO p Q ÎZ
.-H|CM|CO|

LOSSES DUE TO COTTON INSECTS 335
CALCULATED YIELD LOSS FROM FIELD STUDIES
Methods
The information concerning calculated yield losses in Table 1 must be used with caution. The losses were developed from field studies in which yield and pest density data from an untreated plot were compared with yield and pest density data from insecticide- treated plots. The losses calculated for plots treated with the best control reflect control with the best insecticide/miticide evaluated in research plots; these may not necessarily be the most effective chemicals in commercial use. Thus, the losses do not re- flect those that occur under best agricultural practices. Instead, they reflect the potential of a pest population to cause losses when no control is used and when the most effective chemical of those tested has been applied in research studies. It is important to bear in mind that the calculated losses in yield were generally obtained from studies in field plots chosen by researchers because of probable heavy pest infestation. Oftentimes these pest infesta- tions may have been heavier than those normally found in other areas; thus the somewhat higher estimates. However, the data are useful in that they provide information concerning the potential impact these pests can have on cotton and the relationship between density of a pest population and yield.
The methods used to calculate losses and determine the rela- tionship between pest density and yield loss were similar to those used by Schwartz and Klassen (1981). Thus, the Journal of Economic Entomology for the years 1945 to June 1980 and the Insecticide Acaricide Tests for 1976-80 were reviewed for studies containing data on cotton yield. Experiments were utilized wherein yield data were taken in conjunction with data on pest density and in which an untreated control was included.
However, various investigators have used different methods of measuring the density of a given pest and have sometimes used different methods of measuring yield. I therefore needed to trans- form the data for all studies involving a given pest to a common basis. For this, I used the equation
I = lOOT U
where I is equal to the index number for the yield or pest density; T is equal to the yield or pest density in the treated plots, and U is equal to the yield or pest density in the untreated plot.
With this equation, the yield for the untreated plot or the pest population equals 100. Then if the yield is greater in the treated plot than in the untreated control, the yield index is greater than 100; if the yield is less than that in the untreated control, the index is less than 100. Moreover, if the density of the pest population is greater in the treated plots than in the

336 P. H. SCHWARTZ
untreated plots, the pest population index is greater than 100. If the reverse is true, the pest population index is less than 100. The transformed data were then used in subsequent analyses to calculate losses. (The transformation did not affect the outcome of these calculations, but it did allow direct comparison of analyzed data across pests.)
For the purposes of this study, I assumed that a given pest population density would cause a corresponding loss in yield when the plant was attacked at a vulnerable stage in its growth. I also assumed that adequately reliable loss estimates could be made by treating this relationship as though it were linear. (Other work- ers have considered this relationship between the cotton plant and pests (DuRant 1979, Bailey et al. 1980).) Thus, as pest population densities increase, the losses in yield increase proportionally if the plant attacked by the pest is in a susceptible stage of growth. However, I recognize that under some circumstances there could be no cause and effect relationship or the relationship could be sig- moidal or very slightly curvilinear as Hartstack et al. (1978) found for damage to cotton by Heliothis spp. Therefore, data sets were only used when the correlation coefficient (r) was signifi- cantly different from 0 at the 5% level of probability. Schwartz and Klassen (1981) did not use a test for significance and summa- rized all their data (Table 1).
The relationship between pest density and yield or quality calculated by means of the linear equation:
was
Y + kP o
where Y is the yield index for any pest population density index P.
The regression coefficients were Y , which is the yield index when there is no pest infestation, and k, which reflects the change in Y per unit of P. (Also, the coefficient of determination (r ) was calculated.)
The coefficients Y and k were then used to calculate the losses when no treatments were applied (the untreated infestation) and when the most effective treatment in the study was used, as follows:
(1) % loss without controls = Y - Y,«^ ,^^ o 100 X 100 Y o
(2) % loss with best treatment = Y - Y ,^^ o X X 100
Y o
where Y is the yield index without any pest infestation, Y,^^ is the yield index when no control was applied (untreated check plot), and Y^ is the yield index for the most effective treatment.

LOSSES DUE TO COTTON INSECTS 337
Table 3.—Comparison of actual and transformed data
Insecticide
% boll weevil punctured squares 1/
Seed cotton _!./ Pest (pounds Yield index 2/ per acre) index 2/
A 59
B 29
C 27
D 57
E 66
Untreated 60
94
48
45
95
110
100
2346 122
2878 149
2678 139
2240 116
2204 114
1929 100
\_l Actual data from Table 4 of Roussel and Glower (1957), 2/ Transformation.
When the computed percentage loss was less than 0, it was rounded off to 0%; when it was greater than 100, it was rounded off to 100%. For each pest situation, a standard error of the mean times the t^ value at the 0.05 level of significance with n-1 de- grees of freedom was computed with three or more data sets.
The results obtained when these concepts were applied can be demonstrated by comparing the actual data obtained by Roussel and Glower (1957) and the transformations (Table 3). This study was one of the very few in my analysis of the 37 data sets which had a significant correlation and which was concerned only with the boll weevil. The information concerning this study is taken from Table 4 for the St. Joseph location in 1956, which involved 13 applica- tions of five insecticides and insecticide combinations. The specific data are shown in Table 3.
Two regression analyses were then conducted on these data, one by using the actual data and one by using the pest index. The cal- culations are shown in Table 4.
One can see that there are no differences between the index data and the actual data when an _n of 5 was used and the check for the actual data was excluded in calculating the index data. There is a slight difference, as would be expected, when the check is in- cluded because of the addition of another set of variables to the regression equation. However, elimination of the data from the check plots helps to lessen the impact of the influence of heavy pest populations on the prediction of percentage losses.

338 P. H. SCHWARTZ
Table 4.—Analysis of actual and transformed data
Actual data Actual for treated
Parameter Index data 1/ for and control calculated data treated plots plots
Y 0
(3198 165.83 .86 pounds)
3198.82 pounds 3263.94 pounds
k -0.48 -15.33 -17.81 2
r 0.90 0.89 0.78 % loss without
control 28.81 28.68 32.75 % loss with
control 12.96 12.94 14.74 n 5 5 6
IJ Actual data from Table 4 of Roussel and Glower (1957),
Results and Discussion
The data I developed concerning loss with and without control are given in Table 5, which shows the results of my analysis of 251 field experiments for the boll weevil reported in the literature since 1945. Of these, only 37 (15%) had correlation coefficients that were significantly different from 0. These are reported in Table 5 as are significant correlations for other pests.
The parameter Y is of particular interest because it gives the percentage of change in yield that results from an infestation and from an untreated plot when no pest population is present. Any value of Y^ that is 100 or less in Table 5 is an indication that the pest does not cause any reduction in yield. Conversely, any value of Y^ that is over 100 is an indication that the pest is capable of causing a reduction in yield.
The data in Table 5 indicate that the boll weevil has the greatest potential for reducing yield (an average 546% above an untreated check) when no control is used. However, the data for these 37 sets were highly variable; the range was 88.8-1002.8%. Heliothis spp. has the next greatest potential for causing losses in yield of cotton: the gain would be 456.1% over all untreated checks without an infestation. Also, the data for Heliothis spp. plots were much less variable than those for boll weevil plots since the range was 341.6-570.6.
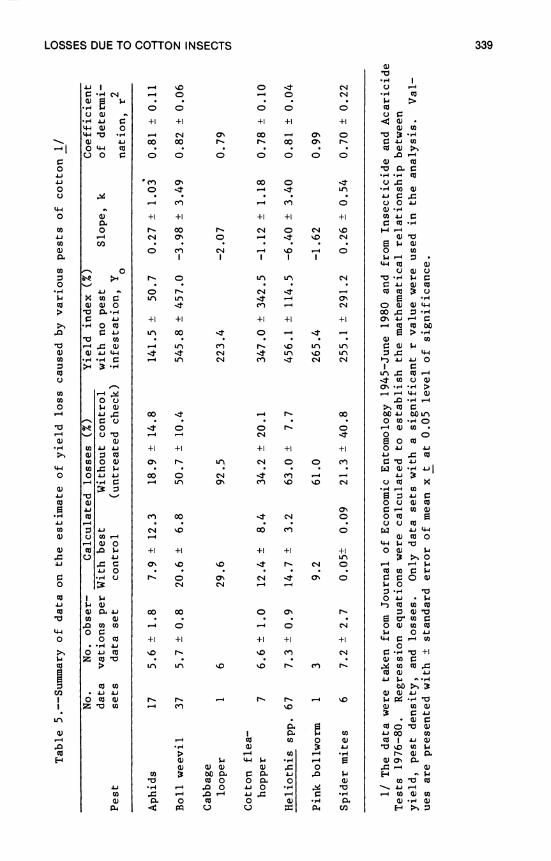
LOSSES DUE TO COTTON INSECTS 339
c o
■u u O O
Xi CO H
.î! a •H 4J M-l (U
eu O M-( U O
a o CO O) Q) CU
CO i—N
2 &«5 >^ o N—^
•H 4J A
M X CO C CO (U (U O > »o a •H
c 4-1 >^ •H o CO ^ c 4J
»o CO TJ 1—» ;c (U <ü <u 4J M-l CO • H •H C p Cd
>^ lí • H
CO t—1 ^ CO O CJ o u (ü
1-4 4J ^ /—s a O
TJ Ô-S o 1—I V--' o na
<U (U •H CO 4J 4J >^ <u 3 CO
CO O d) M-l CO 4= u O O 4J 4J
r-i •H c <U 12 3 J-J TJ Xw/
CO (U 6 4-»
•H CO 4J 1—1 CO D 4-t d) O CO r-4
1—1 <ü O 0) CO -Û J-l
JC u JJ 4J JC C
■u O C •H o O :2
CO 1 u 4J p <D CO a; CU AJ
T> CO 0) Xi CO CO
VM o C O O CO
•H •u >^ o J-l CO V4 ^ CO 73
> 1 CO CO
CO • 4-> 4-t 1 o CO <ü 1 Ä TJ CO
o +1 i-H
C30
O
'en o
CM
o o +1
CM C30
m
+1 C30 CJN
en I
0 in
+1 +1
m 00
1—1
00 <t
-:t 0 «—( f—1
+1 +1
ON r^
00 0 —t m
m
CM 1—4
C30
+1 +1
<JN vO
r^ 0 CM
00 00
1—1 0
+1 +1
vO r^
ON
O
O
CM I
cn CM CM
CM c^
CM
0 0
CM CM
0 0 0
+1 +1 +1
00 00
ON ON
0
00 i-H
0 in
f—I cn 0
+1 +1 +1
CM I-H
0 CM
1—1 1 1 1
0
in in CM
CM
cn f—1 1-H
1-H C3N CM
+1 +1 +1
0 1—1 <!■ 1—1
<!■
cn m
in
CM
in in CM
0 CvJ
r*- §
+1 +1 +1
CM 0 0 cn
cn cn VO
1—1
vO 1—(
CM
-«t CM <3N 0
+1
<!• CM 1—t
+1
1—1
CM
<7N
+1 m 0
0
0 CJN r^
1—1 0 CM
+1 +1 +1
VO cn CM
VO r-- cn r^
r^ r^ ^
CU a 1 a. E CO
r-l CO CO 0 0) •H (U ^ 4-1 > r-< CO 1—1 •H <U u M-l U •H 1-4 S (U 0) <u (U x: 0
CD 1^ bO CU C CU 4-) ^ »-< 13 CO 0 0 CU 0 <ü •H 1—1 Xï 0 4J 0 •H M T? JC rH Xi rH 4J 4= I—1 c •H CU 0 CO 0 Q) •H CU < PQ 0 0 ffi PLi cn
o; T> •H 1 0 1-4
•H CO U > CO U C
<3 0) • (U CO
73 ;^ • H C 4-1 CO CO Q) >.
Xi 1—1
(U CO 73 a c •H •H CO 0 ^
• H CO (ü 4-1 c ^ CJ 0 4-) <u •H CO 4-) C C CO •H M 1—1
<v 73 g u (U 0 CO U 1—1 P •
M-l CO Q) 0 (U 0
-o • H VJ C C 4J O) CO CO CO ^ 0
B • H 0 QJ <u M-l 00 JC 3 •H ON 4-1 1—1 c
CO CO ÖO g > •H Q) CO C (U u 3 ,c M-l
•-0 ■U 4J 0
tn X CO 1—1
<!• CO 0 <U CJN •H •H > 1—1 r-l M-l 0)
Xi • H 1-4
>. CO C to 4-» öom 0 CO •H 0
1—1 <u CO 0 0 g 0 CO 0 4-) 4J 4-» ^ CO C 73 4-t u (U • H ^1
4-» > 0 CO X
•H 1—1 CO 6 3 *J c 0 U <D CO C! 1-1 CO 0) 0 CO g U 0 CO U 4J M-l
(U CO 0 M-l VJ 73 0 (U U
^ >% 0 1—1 1—1 VJ
CO CO a Wi C C 0 <u }-< 0 3 •H 73 0 4-1 • Wi
•-) CO CO CO 0 Q) 73 g er CO C
0 <u CO CO u 0 4-1
M-l c¡ 0
1—1 CO
C •H »o +1 (U CO c ^ CO CO JC
CO (U 4J 4-1 »-I •* •H
tJ) >N :J (U <u 4-1 VJ peí •H 73 (U CO <U ^ C 4-t • (1) C CO 0 73 <ü
4-1 00 CO CO 4J (1)
73 vO CO u r-- (Ü a
<U ON CU JC 1—1 (U H n VJ
CO 73 CO •v^ 4-1 1—1
1—1 CO QJ CO (U •H 0) H f»^ 3

340 P. H. SCHWARTZ
My results appear to be consistent with data developed by other workers (Table 6). For example, a comprehensive report on cotton losses was presented by Cramer (1967). He indicated that the boll weevil accounted for two-thirds of the 12% loss in yield due to insects attacking cotton during the period 1942-51. For the period 1951-60, he reported that the proportion of the loss due to the boll weevil dropped to 42%, whereas bollworms were responsible for 20%; the remaining losses were attributed to the infestation of all other species such as plant bugs (Lygus spp.), thrips (Thripi- dae), and spider mites (Tetranychus spp.). He based his figures on data in Losses in Agriculture (U.S. Department of Agriculture 1954, 1965). During these two periods, it was estimated that in- sect pests on cotton caused losses amounting to 12 and 19.0%, respectively.
Cramer (1967) developed an equation for estimating the losses that would be obtained in the absence of any control measures as follows:
X + (52x) = 696 100
where 52 is equal to the percentage increase when control measures are used, and 696 is the average nationwide yield per unit area.
From this, cotton production during the period 1951-60 could have been increased by approximately 20% provided modern control measures against insects had been applied to the entire acreage. Instead Cramer estimated from his equation that the equivalent re- duction was 34% of the potential yield per area.
Other workers too have undertaken to develop experimental data to determine losses from cotton pests. Their results are shown in Table 6. Fye et al. (1962) published results of experiments under- taken from 1928 to 1958 in South Carolina in their study of the ef- fects of insecticides on cotton yields and on control of cotton insect pests. The boll weevil and the bollworm were emphasized. Between 1928 and 1945 when the test plots were being treated with inorganic insecticides, average yield was 1225 pounds of seed cot- ton per acre each year for treated plots and 955 pounds for un- treated plots, or an increase of 23.6%. Between 1946 and 1958, the test plots were treated with inorganic insecticides, and average yield was 1441 pounds of seed cotton per acre for treated plots and 953 pounds for untreated plots, or an increase of 53.9%. However, analysis of the results for the different years showed that the differences between the yields of the treated and untreated plots were especially large in years when infestations were above aver- age. For example, in 1929, 1938, 1941, 1946, 1949, and 1950, years of severe boll weevil infestation, the average yield was 1146 pounds per acre in treated plots and 756 pounds per acre in un- treated plots, or an increase of 52%. In those years when boll weevil infestations were below average, the average yield was 1363 pounds per acre for treated plots and 1211 pounds per acre for un- treated plots, or an increase of only 12.5%.
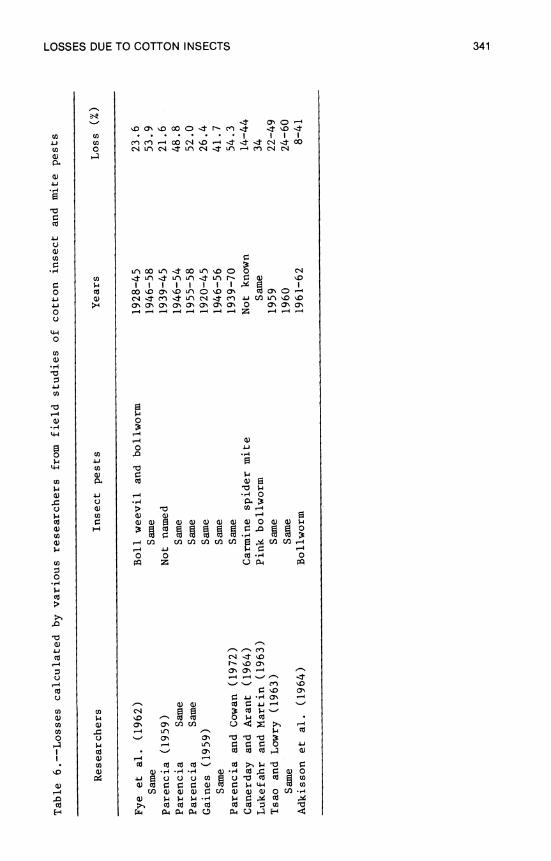
LOSSES DUE TO COTTON INSECTS 341
o
u Cd
vD ON ^ 00 o <i <f 0> O ^
CO <!■ <!■ vD <J- I III
CNimcM<l-mcSv^in.—tcocNcs
Ü u to (1) CO <u
moom<l'oou^^oo
I I I I I I I I C30vO(T>^LOOvDCrN
0^C3^0^C^^0^0^0^0^
ÛJ es
cd CTN O ---I en m vo vr>
0> 0^ CTN
O > r-< 1—1 O
CO ^ CO T3 0) c cu cd
o •H (U > t3 CO <ü 9á C (U d) B M ;^ IS
r-í en 1—t XJ O 0 pq !Zi
<ü (D Q) 0) di
c3 Cd cd Cd vd en C/3 C/3 CO C/3
0) 0 'T3 M •H O
CU ^ CO rH
t-4 0 QJ O (U dj U
1 1 O 1^
0 ^ CO en r-t »^ C r-H Cd -H O Ü P^ PQ
CM <t vO r^ vo o> o^ o\ ^^
/-S Si 9¿ es 0 0 vO /--N cd cd CJN <y> en en
•u 0 o; cd
cj o o c: c! c
CÜ
C 4J -H cd C3 4J ¿ cd }-4 ' o j-4 Cd o <: :s: ■xj TJ 'd
g g S cd cd Cd
cd t»^ }-i TJ
vO ON
en <ü <D Qj }J VJ J-i -H cd cd cd cd
Pu pi4 eu O
Cd 42 . o "TO cd cd
cö a v-< M-i C en <ü (u 0)
>-• {= ^ Cd Cd ;3
PM u HJ H
o en «H cd ^ CD T3

342 P. H. SCHWARTZ
C. R. Parencia, Jr., (1959) obtained similar results in a study carried out from 1939 to 1958 in Texas. A total of 320 com- parative experiments was undertaken. Data were obtained from 787 untreated plots covering an overall area of 2177 acres and from 1385 treated plots covering a total area of 6102 acres. Over the 20-year period, the untreated plots produced an average of 740 pounds per acre, and the treated plots yielded 1049 pounds per acre, equivalent to an increase of 41.5%. From 1939 to 1945, the test plots were treated with arsenical compounds from 1946 to 1954 with chlorinated hydrocarbons, and from 1955 to 1958 with organo- phosphates. The average increase in yield of treated plots, as reflected by losses in Table 6 for these three periods, was as follows: 1939-45, 21.6%; 1946-54, 48.8%; 1955-58, 52%.
Gaines (1959) reported a similar study made in Louisiana from 1920 to 1956. The use of insecticides increased the annual yield of seed cotton an average 26.4% between 1920 and 1945 and an aver- age 41.7% between 1946 and 1956. Parencia and Cowan (1972), in a review of the comparative yields of cotton in treated and untreated plots in central Texas for the period 1939-70, found that in plots treated with an average 6.1 applications of insecticides, yield was 54.3% greater (366 more pounds of seed cotton per acre) than in untreated plots. During this period, the insects of primary impor- tance were the boll weevil, bollworm, and tobacco budworm (Helio- this virescens (F.)). However, the tobacco budworm became consid- erably more important over time and often was the predominant spe- cies, especially after it developed resistance to several of the recommended insecticides. The cotton fleahopper (Pseudatomoscelis seriatus (Reuter)) also increased in importance as it developed re- sistance to the organochlorine insecticides.
Canerday and Arant (1964) showed that yield losses of seed cotton caused by the carmine spider mite (Tetranychus cinnabarinus (Boisduval)) ranged from 14 to 44% and were directly related to population density and duration of infestation.
Lukefahr and Martin (1963) evaluated the damage to lint and seed cotton caused by the pink bollworm (Pectinophora gossypiella (Saunders)). The data indicated that the value of the cotton crop was reduced 34% when at least 60% of the bolls were infested and that reductions in staple length, strength, and fineness or imma- turity of fiber were directly related to the levels of infestation. In artificially infested bolls, the degree of damage was related to age of boll and number of larvae per boll. Tsao and Lowry (1963) estimated losses in yield due to the pink bollworm by using the equation:
Y = lOOy/p and loss = Y - y, where y equals actual yield, p equals percentage of locks pickable, and Y equals potential yield.
In the application of this formula, they found that potential yield was 1379 pounds per acre from a potential check plot yield of 696.9 pounds per acre. The loss per acre caused by the pink

LOSSES DUE TO COTTON INSECTS 343
bollworm (potential yield minus actual yield) was 681 pounds for the check plot and 296 for the treated plot. In 1959 and 1960, the loss per acre was 738 pounds in the check and 238 in treated plots. Thus, the data of Tsao and Lowry (1963) indicate that the potential loss in yield from untreated plots was 49% in 1959 and 60% in 1960; that in treated plots was 22% in 1959 and 24% in 1960.
In other studies not reported above, Brazzel and Gaines (1956) likewise studied the effect of three levels of pink bollworm infes- tations and an insect-free check on yield and quality of cotton. Their data indicated that heavy infestations caused large reduc- tions in the value of cotton. If 100% of the bolls were infested with an average 6.4 or 9.6 larvae per boll, most of the loss in the value of cotton was the result of reduced quality. When the infes- tation reached 12.9 larvae per boll, the reduced yield accounted for more of the loss than the reduced quality. Subsequently, Brazzel and Gaines (1957) further evaluated cotton yield and qual- ity losses caused by various levels of pink bollworm infestations. They found that when ca. 50% of the bolls were infested at a rate of less than two larvae per boll, the value of the cotton decreased little. Yields per acre of both lint and seed decreased slightly with increased levels of insect infestations in both of their experiments. However, these differences were not significant though a trend to decreased yields with increased bollworm in- festations was indicated.
DuRant (1979) studied the effectiveness of selected insecti- cides used in combination against three species of insects on cot- ton. He found that highly significant correlations existed between the degree of Heliothis spp. control and yield but no correlation existed between beet armyworm (Spodoptera exigua (Hübner)) control and yield. On the basis of his data, Heliothis spp. and beet army- worms caused an average yield reduction of 81% compared with the un- treated check.
Adkisson et al. (1964) studied the effect of the bollworm on the quality of cotton. They found that losses in cotton due to at- tack of the bollworm are more likely to be in yield than in qual- ity. Moreover, relatively small numbers of bollworms may cause significant yield losses. Under the conditions of their experi- ment, significant yield losses were caused when infestations aver- aged 8-10 bollworm larvae per 100 plants or more than 2 per foot of row. On the basis of their data, significant yield loss did not occur until damage counts exceeded 3% of the squares or 3% of the bolls. Severe yield loss occurred when more than 5% of the bolls were damaged for prolonged periods. Thus, light, medium, and heavy infestations of bollworms caused losses of 8, 23, and 41%, respectively.
Bailey et al. (1980) studied 16 cotton genotypes and the ef- fect of insect populations on yield. They found that significant correlations existed between lint yield and insect populations for untreated plots infested with adult tarnished plant bugs (Lygus

344 P. H. SCHWARTZ
lineolaris (Palisot de Beauvois)), tarnished plant bug nymphs, and the total number of tarnished plant bugs in the plots. Also, in the treated plots, lint yield and populations of these pests as well as whiteflies showed significant correlations. Under the con- ditions of these tests, correlations were insignificant for yield and populations of Heliothis spp. larvae, Heliothis spp. eggs, and cotton fleahoppers. Untreated plots yielded an average 33% less lint than plots treated with insecticide.
Hartstack et al. (1978) studied the damage done to cotton by the bollworm and tobacco budworm. Their data showed relationships between the number of larvae present and the percentage and number of damaged squares and bolls. A highly significant relationship was also shown between the number of larval days (number of larvae counted times number of days observed) and percentage of lint lost. They showed that an average of 2% damaged squares and bolls and 60,000-70,000 larval days could be tolerated without loss. They also found that the percentages of lint loss v. damaged squares, damaged bolls, and total number of larval days were curvilinear re- lationships. Yield losses ranged from 0 to 72% with larval infes- tation levels of an average of 1-12 per acre. Therefore, with 25% damaged squares, a loss in yield of about 70% can be anticipated with 25% damaged bolls. With 10% damaged squares or bolls, yield loss was calculated to be about 30%.
Lloyd et al. (1962) studied the effect of four levels of boll weevil infestations on yield and quality of cotton. Small plots of cotton fields were caged as soon as the first young squares became visible, and different numbers of boll weevils were introduced into the cages with the intention of obtaining infestation levels of 25, 50, and 75%. The exact infestation levels actually attained were not those desired. Their data indicated an inverse relationship between total yield and percentage of squares damaged throughout the season. However, infestation levels of approximately 27% re- sulted in 30 and 15% yield reductions for 1959 and 1960, respec- tively; and infestation levels of 40 and 48% resulted in reduced yield levels of 44 and 18%, respectively. At infestation levels of 62 and 67%, reductions of 59 and 52% were obtained.
Lloyd and Merkl (1966) made a 2-year replicated field cage study of the population dynamics of the boll weevil. Their data indicated that first-generation progeny from boll weevil popula- tions estimated at 0, 14, 25, 50, and 100 weevils per acre damaged 0, 28, 46, 66, and 83% of the squares, respectively. Second-gener- ation weevils damaged from 84 to 96% of the squares. Seed cotton harvested from the respective treatments averaged 2802, 1875, 1678, 1380, and 479 pounds per acre; yields were reduced 33, 40, 53, and 70% with 28, 46, 66, and 83% damage of the squares, respectively. They also detected differences in lint quality between plots in- fested with 100 overwintered weevils (83% damaged squares) and those infested at the other four levels. No other differences in quality were observed.

LOSSES DUE TO COTTON INSECTS 345
The general conclusion therefore is that cotton insect loss data are dynamic; they change from year to year in geographical areas. Losses due to insect damage continue to occur at magnitudes that have not decreased appreciably over the past 30 years, even with the increased intensity of cotton plantings. While some cot- ton pests may cause increased losses now because of the increased importance of one pest species over another pest species due to resistance, climatological factors, or cultural practices, the mag- nitude of the losses has not shifted downward to any extent. Also, the boll weevil and the bollworm continue to be the insects that cause major losses to cotton. This means that we are barely stay- ing abreast in the control of these insect pests by the research, development, and implementation of pest management programs. There are times when cotton insects can tip the scales in their favor as evidenced by the atypical year Georgia had in 1977. In that year, the State of Georgia reported severe losses to many crops (Todd and Suber 1981). There was an extended drought which favored insect development, making infestations more severe than normal. Cotton and corn were heavily damaged. In 1977, Georgia harvested 170,000 acres of cotton at a production value of approximately $18.5 mil- lion. The control and yield reduction costs for that crop amounted to $26.7 million, a loss of 144%.
There is a continual need to apply pest control technologies to cotton pests and to maintain a strong research program directed toward a reduction in losses caused by these pests. In addition, research should be directed toward the refinement of the cost of application of these technologies to maximize cotton yields by re- ducing pest control costs to a minimum.

346 P. H. SCHWARTZ
LITERATURE CITED
Adkisson, P. L., C. F. Bailey, and R. L. Hanna. 1964. Effect of the bollworm, Heliothis zea, on yield and qual-
ity of cotton. Journal of Economic Entomology 57:448-450.
Bailey, J. C, B. W. Hanny, and W. R. Meredith, Jr. 1980. Combinations of resistant traits and insecticides: ef-
fect on cotton yield and insect populations. Journal of Eco- nomic Entomology 73:58-60.
Brazzel, J. R., and J. C. Gaines. 1956. The effect of pink bollworm infestations on yield and
quality of cotton. Journal of Economic Entomology 49:852-854.
and J. C. Gaines. 1957. Cotton yield and quality losses caused by various levels
of pink bollworm infestations. Journal of Economic Entomology 50:609-613.
Canerday, T. D., and F. S. Arant. 1964. The effect of spider mite populations on yield and quality
of cotton. Journal of Economic Entomology 57:553-556.
Cramer, H. H. 1967. Plant protection and world crop production. Pflanzen-
schutz-Nachrichten 20:524-527.
DeBord, D. V. 1977. Cotton insect and weed loss analysis. The Cotton Founda-
tion. Mimeographed, 122 p.
DuRant, J. A 1979. Effectiveness of selected insecticides and insecticide
combinations against the bollworm, tobacco budworm, and beet armyworm on cotton. Journal of Economic Entomology 72:610- 613.
Fye, R. E., A. R. Hopkins, and W. P. DuBose. 1962. Cotton yields with recommended and experimental insecti-
cides. U.S. Department of Agriculture, Technical Bulletin No^ 1264, 41 p.
Gaines, R. C. 1959. Ecological investigations of the boll weevil, Tallulah,
LA, 1915-1958. U.S. Department of Agriculture, Technical Bul-+ let in No. 1208.
Hartstack, A. W., R. L. Ridgway, and S. L. Jones. 1978. Damage to cotton by the bollworm and tobacco budworm.
Journal of Economic Entomology 71:239-243.

LOSSES DUE TO COTTON INSECTS 347
Hopkins, A. R., R. F. Moore, Jr., and C. R. Parencia, Jr. 1980. Cotton pests. U.S. Department of Agriculture, Agriculture
Handbook No. 571, p. 50-69.
Hyslop, J. A. 1938. Losses occasioned by insects, mites, and ticks in the United States. U.S. Department of Agriculture, Bureau of Ento- mology and Plant Quarantine, E-444, 57 p.
Lloyd, E. P.i and M. E. Merkl. 1966. A field-cage study of population dynamics of the boll wee-
vil. Journal of Economic Entomology 59:83-86.
M. E. Merkl, and B. G. Crowe. 1962. The effect of boll weevil infestations on yield and qual-
ity of cotton. Journal of Economic Entomology 55:225-227.
Lukefahr, M. J., and D. F. Martin. 1963. Evaluation of damage to lint and seed of cotton caused by
the pink bollworm. Journal of Economic Entomology 56:710-713.
Marlatt, C. L. 1904. The annual loss occasioned by destructive insects in the United States. U.S. Department of Agriculture, Yearbook of Agriculture, p. 461-472.
National Cotton Council. 1974. Boll weevil losses: value and location of losses caused by the boll weevil. Mimeographed, 52 p.
Parencia, C. R., Jr. 1959. Comparative yields of cotton in treated and untreated
plots of insect control experiments in central Texas, 1939- 1958. Journal of Economic Entomology 52:757-758.
and C. B. Cowan, Jr. 1972. Comparative yield of cotton in treated and untreated plots
in insect-control experiments in central Texas, 1939-1970. Journal of Economic Entomology 65:480-481.
Roussel, J. S., and D. F. Clower. 1957. Resistance to the chlorinated hydrocarbon insecticides in
the boll weevil. Journal of Economic Entomology 50:463-468.
Schwartz, P. H. , and W. Klassen. 1981. Estimate of losses caused by insects and mites to agricul-
tural crops. In Handbook of Pest Management in Agriculture, Vol. 1. D. Pimentel, editor. CRC Press, Inc., Boca Raton, FL. p. 15-77.

348 P. H. SCHWARTZ
Todd, J. W., and E. F. Suber. 1981. Losses due to insect damage and costs of control in Geor-
gia in 1977. Research Report, Division of Entomology, Univer- sity of Georgia, Special Committee on Insect Survey and Losses. [In press.]
Tsao, C. H., and W. L. Lowry. 1963. Control of the pink bollworm and a method for estimating
losses in cotton yield. Journal of Economic Entomology 56:158- 160.
U.S. Department of Agriculture. 1946. Examples of estimated losses caused by insects. U.S. De-
partment of Agriculture, Bureau of Entomology and Plant Quaran- tine, Typewritten tables, 2 p.
1954. Losses in agriculture. A preliminary appraisal for re- view. U.S. Department of Agriculture, ARS 20-1, 190 p.
1965. Losses in agriculture. U.S. Department of Agriculture, Agriculture Handbook No. 291, 120 p.
1976. Cotton and tobacco insect control. U.S. Department of Ag- riculture, Agricultural Research Service, NRP 20230. Mimeo- graphed, 42 p.
1980. Thirty-third Annual Conference Report on Cotton Insect Re- search and Control. Agricultural Research (Southern Region), Science and Education Administration, U.S. Department of Agri- culture, New Orleans, LA. Mimeographed, 77 p.

LOSSES DUE TO COTTON INSECTS 349
SELECTED REFERENCES
Bacheler, J. S., and J. S. Wilkins. 1980a. Cotton bollworm control, 1977. Insecticide and Acaricide
Tests, Volume 5, Paper No. 235, p. 132.
and J. S. Wilkins. 1980b. Cotton bollworm and tobacco budworm control, 1978. In-
secticide and Acaricide Tests, Volume 5, Paper No. 236, p. 132- 133.
Becnel, I. J., H. S. Mayeux, and J. S. Roussel. 1947. Insecticide tests for the control of cotton boll weevil
and cotton aphids in 1946. Journal of Economic Entomology 40: 508-513.
Bell, M. R., and C. L. Romine. 1980. Tobacco budworm field evaluation of microbial control in
cotton using Bacillus thuringiensis in a nuclear polyhedrosis virus with a feeding adjuvant. Journal of Economic Entomology 73:427-430.
Bottger, G. T., A. J. Chapman, R. L. McGarr, and C. A. Richmond. 1958. Laboratory and field tests with Sevin against cotton in-
sects. Journal of Economic Entomology 51:236-239.
Bull, D. L. 1980. Fate and efficacy of sulprofos against certain insects as-
sociated with cotton. Journal of Economic Entomology 73:262- 264.
V. S. House, J. R. Abies, and R. K. Morrison. 1979. Selective methods for managing insect pests of cotton. Journal of Economic Entomology 72:841-846.
Calhoun, S. L., and E. W. Dunnam. 1953. Heptachlor and other insecticides for control of cotton
pests. Journal of Economic Entomology 46:157-158.
and W. R. Smith. 1950. Control of boll weevil, bollworm, and cotton aphid with
organic insecticides applied as concentrated sprays. Journal of Economic Entomology 43:606-610.
Cowan, C. B., Jr., and J. W. Davis. 1963. Control of several late-season cotton pests in field ex-
periments in 1962. Journal of Economic Entomology 56:790-793.
and J. W. Davis. 1968. Field tests for the conventional low volume or ultra-low volume sprays for control of the boll weevil, budworm, and to- bacco budworm on cotton in 1967. Journal of Economic Entomol- ogy 61:1115-1116.

350 P. H. SCHWARTZ
Cowan, C. B., Jr., J. W. Davis, and C. R. Parencia, Jr. 1957. Control of the boll weevil and bollworm with chlorinated hydrocarbon and phosphorus insecticides in 1956. Journal of Economic Entomology 50:663-666.
J. W. Davis, and C. R. Parencia, Jr. 1960. Field experiments against several late-season cotton in-
sects in 1959. Journal of Economic Entomology 53:747-749.
C. R. Parencia, Jr., and J. W. Davis. 1956. Late-season control of the boll weevil and the bollworm with new insecticides in 1955. Journal of Economic Entomoloev 49:783-785.
C. R. Parencia, Jr., and J. W. Davis. 1959. Field experiments for control of late-season infestations
of several cotton insects. Journal of Economic Entomoloev 52: 975-977. -^
C. R. Parencia, Jr., and J. W. Davis. 1961. Control of several cotton insects in field experiments in
1960. Journal of Economic Entomology 54:1011-1014.
Davis, J. W., and C. B. Cowan, Jr. 1972. Field evaluation of three formulations of aldicarb for
control of cotton insects. Journal of Economic Entomoloev 65- 231-232. ^
. and C. B. Cowan, Jr. 1974. Early season insects on cotton: control with two systemic
insecticides. Journal of Economic Entomology 67:130-131.
C. B. Cowan, Jr., and C. R. Parencia, Jr. 1957. Thrips control on cotton with phosphorus insecticides. Journal of Economic Entomology 50:676-678.
C. B. Cowan, Jr., and C. R. Parencia, Jr. 1962. Field experiments with insecticides on cotton for control
of the boll weevil, bollworm, and cotton leafworm in 1961. Journal of Economic Entomology 55:688-692.
C. B. Cowan, Jr., W. C. Watkins, Jr., P. D. Lingren, and R. L. Ridgway.
1966a. Experimental insecticides applied as sprays to control thrips and the cotton fleahopper. Journal of Economic Entomol- ogy 59:980-982.
C. R. Parencia, Jr., and C. B. Cowan, Jr. 1958. Control of thrips and the cotton fleahopper with sprays
in central Texas in 1957. Journal of Economic Entomoloev 51: 489-490.

LOSSES DUE TO COTTON INSECTS 351
Davis, J. W., C. R. Parencia, Jr., and C. B. Cowan, Jr. 1961. Field experiments for control of thrips, cotton leafhop-
pers, and overwintered boll weevils in 1960. Journal of Eco-
nomic Entomology 54:966-970.
W. C. Watkins, C. B. Cowan, Jr., R. L. Ridgway, and
D. A. Lindquist. 1966b. Control of several cotton pests with systemic insecti-
cides. Journal of Economic Entomology 59:159-162.
Dean, H. A., and J. C. Gaines. 1950. Comparison of dust and sprays for cotton insect control. Journal of Economic Entomology 43:225-226.
Dunnam, E. W., and S. L. Calhoun. 1948. Benzene hexachloride to control boll weevil and cotton
aphid. Journal of Economic Entomology 41:22-25.
Enkerlin, S. D. 1959. Different concentrations of thiodan for the control of cotton insects. Journal of Economic Entomology 52:1068-1069.
Ewing, K. P., and C. R. Parencia, Jr. 1945. DDT for control of cotton fleahopper. Journal of Economic
Entomology 38:495-496.
C. R. Parencia, Jr., and E. E. Ivy. 1947. Cotton-insect control with benzene hexachloride, alone or
in mixture with DDT. Journal of Economic Entomology 40:374-
381.
Fenton, J. A. 1950. Spraying and dusting for cotton insect control. Journal
of Economic Entomology 43:292-294.
Fife, L. C, and R. L. Walker. 1954. Comparative effectiveness of various phosphorus and chlo-
rinated hydrocarbon insecticides for control of cotton pests.
Journal of Economic Entomology 47:803-807.
R. L. Walker, and S. F. Bondy. 1949. Boll weevil control with several organic insecticides dur-
ing 1948. Journal of Economic Entomology 42:682-684.
Fye, R. E., A. R. Hopkins, and R. L. Walker. 1961. Field experiments on control of overwintered boll weevils.
Journal of Economic Entomology 54:622-624.
Gaines, J. C, and H. A. Dean. 1948. Tests of insecticides for the control of several insects.
Journal of Economic Entomology 41:48-54.

352 p. H. SCHWARTZ
Gaines, J. C., and H. A. Dean. 1949. Insecticide tests for boll weevil control during 1948. Journal of Economic Entomology 42:795-798.
H. A. Dean, and R. Wipprecht, 1948. Control of thrips on cotton. Journal of Economic Entomol-
ogy 41:510-512.
H. A. Dean, and R. Wipprecht. 1951. Test of insecticides for control of insects during 1950. Journal of Economic Entomology 44:367-372.
and R. L. Hanna. 1948. Comparison of diluents in insecticide mixtures for cotton
insect control. Journal of Economic Entomology 41:811-812.
Gaines, R. C, M. T. Young, and G. L. Smith. 1947. Calcium arsenate and nicotine to control bpll weevil and
cotton aphid, 1939-1946. Journal of Economic Entomology 40: 600-603.
Gard, I. E. 1980. Chemical control of Heliothis spp. on cotton. Insecticide
and Acaricide Tests, Volume 5, Paper No. 237, p. 133-134.
Hanna, R. L. 1954. Application schedules for control of cotton insects. Journal of Economic Entomology 47:1129-1131.
1958. Insecticidal seed treatments for cotton. Journal of Eco- nomic Entomology 51:160-163.
and J. C. Gaines. 1950. Tests of insecticides for control of cotton insects during
1949. Journal of Economic Entomology 43:288-292.
and J. C. Gaines. 1951. Lime-free calcium arsenate mixed with organic insecticides
for cotton insect control. Journal of Economic Entomology 44: 430-432.
and J. C. Gaines. 1952. Evaluation of dusting schedules for control of cotton in-
sects. Journal of Economic Entomology 45:549-550.
and W. J. Mistric, Jr. 1953. Effect of different treatment schedules for control of
cotton insects. Journal of Economic Entomology 46:641-644.

LOSSES DUE TO COTTON INSECTS 353
Hopkins, A. R., and H. M. Taft. 1964. Field experiments for control of the boll weevil, bollworm
spp., and the cotton aphid on cotton in 1960-62. Journal of Economic Entomology 57:509-511.
and H. M. Taft. 1965. Control of certain pests with a new systemic insecticide, UC-21149. Journal of Economic Entomology 58:746-749.
H. M. Taft, and W. James. 1967. Comparison of spray nozzles for ground applications of ultra-low-volume (undiluted) technical insecticides. Journal of Economic Entomology 60:561-565.
H. M. Taft, and W. James. 1979. Comparison of spray nozzles for ground applications for
control of cotton insects and spider mites. Journal of Economic Entomology 72:180-183.
Ivy, E. E., J. R. Brazzel, A. L. Scales, and D. F. Martin. 1955. Two new phosphate insecticides for cotton insect control. Journal of Economic Entomology 48:293-295.
C. R. Parencia, Jr., and K. P. Ewing. 1947. A chlorinated camphene for control of cotton insects. Journal of Economic Entomology 40:513-517.
C. R. Parencia, Jr., R. W. Moreland, and K. P. Ewing. 1945. DDT for bollworm control during 1944. Journal of Economic Entomology 38:534-537.
Kinzer, R. E., L. A. Bariola, R. L. Ridgway, and S. L. Jones. 1976. Systemic insecticides and a nuclear polyhedrosis virus for control of the budworm and tobacco budworm on cotton. Journal of Economic Entomology 69:697-701.
Kulash, W. M. 1948. New insecticides for cotton insect control. Journal of Economic Entomology 41:986-987.
Lentz, G. L. 1980. Bollworm and tobacco budworm control, 1978. Insecticide
and Acaricide Tests, Volume 5, Paper No. 238, p. 134.
Loden, H. D., and H. O. Lund. 1948. Chlorinated camphene and parathion to control the cotton
boll weevil and cotton aphid. Journal of Economic Entomology 41:581-583.
Luttrell, R. J., W. C. Yearian, and S. Y. Young. 1979. Laboratory and field studies on the efficacy of selected
chemical insecticide-Elcar (Baculovirus heliothis) combinations against Heliothis spp. Journal of Economic Entomology 72:57- 60.

354 p. H. SCHWARTZ
McGarr, R. L.
1953. Combinations of insecticides for control of the pink boll- worm and other cotton insects in the lower Rio Grande Valley. Journal of Economic Entomology 46:1103-1105.
1955. Weekly applications of insecticides for control of the pink bollworm and the boll weevil. Journal of Economic Ento- mology 48:95-96.
1957. Tests with insecticides for control of cotton insects in the lower Rio Grande Valley. Journal of Economic Entomology 50:632-634.
and A. J. Chapman. 1963. Control of three important cotton insects in the lower Rio Grande Valley in 1960. Journal of Economic Entomology 56:902- 903.
A. J. Chapman, and W. L. Lowry. 1961. Field tests with insecticides for pink bollworm control in
1958. Journal of Economic Entomology 54:972-974.
H. T. Dulmage, and D. A. Wolfenbarger. 1972. Field tests with HD-1 ôendotoxin of Bacillus thuringien- £is, and with chemical insecticides for control of the tobacco budworm and the bollworm in 1970. Journal of Economic Entomol- ogy 65:897-899.
and D. A. Wolfenbarger. 1970. Insecticides for control of four cotton insects in 1968. Journal of Economic Entomology 63:1324-1325.
Parencia, C. R., Jr., and C. B. Cowan, Jr. 1960. Increased tolerance of the boll weevil and cotton fleahop-
per to some chlorinated hydrocarbon insecticides in central Texas in 1958. Journal of Economic Entomology 53:52-56.
C. B. Cowan, Jr., and J. W. Davis. 1958. Field tests with the systemic insecticides Thimet and Bayer 19639 as cottonseed treatments in 1957. Journal of Eco- nomic Entomology 51:872-875.
C. B. Cowan, Jr., and J. W. Davis. 1960. Control of several early-season cotton pests with insecti-
cides. Journal of Economic Entomology 53:1051-1054.
J. W. Davis, and C. B. Cowan, Jr. 1957. Further field tests with systemic insecticides employed
as seed treatments. Journal of Economic Entomology 50:614-617,

LOSSES DUE TO COTTON INSECTS 355
Parencia, C. R. , Jr., and K. P. Ewing. 1948. Control of the cotton fleahopper by chlorinated camphene,
DDT, and sulphur. Journal of Economic Entomology 41:735-738.
and K. P. Ewing. 1950. Late-season control of boll weevil and bollworm with dust
and sprays. Journal of Economic Entomology 43:593-595.
E. E. Ivy, and K. P. Ewing. 1946., Control of bollworm and cotton fleahopper by DDT. Jour-
nal of Economic Entomology 39:329-335.
Pfrimmer, T. R. 1958. Insecticide tests against the boll weevil and the bollworm
at Tallulah, LA, in 1956. Journal of Economic Entomology 51: 41-44.
1979. Heliothis spp.: control on cotton with pyrethroids, car- bamates, organophosphates, and biological insecticides. Journal of Economic Entomology 72:593-598.
R. E. Furr, and E. A. Stadelbacher. 1971. Materials for control of boll weevils, bollworms, and to- bacco budworms on cotton at Stoneville, Mississippi. Journal of Economic Entomology 64:475-478.
and R. C. Gaines. 1956a. Field tests with insecticides against cotton insects at
Tallulah, LA, in 1954. Journal of Economic Entomology 49:72- 74.
and R. C. Gaines. 1956b. Field tests with new materials against cotton insects at
Tallulah, LA, during 1955. Journal of Economic Entomology 49: 712-713.
E. P. Lloyd, M. E. Merkl, and R. E. Furr. I960. Field experiments with several insecticidal sprays against
the boll weevil and bollworm. Journal of Economic Entomology 53:711-714.
Pieters, E. P., and J. D. Boyette. 1977. Evaluation of insecticides to control Heliothis spp. on
cotton in Mississippi, 1976. Insecticide and Acaricide Tests, Volume 2, Paper No. 121, p. 93.
P. W. Mullins, and VÎ. T. Halford. 1978. Chemical control of Heliothis spp. on cotton, 1977. In-
secticide and Acaricide Tests, Volume 3, Paper No. 176, p. 127.

356 P. H. SCHWARTZ
Pieters, E. P., and D. L. Pitts. 1979. Efficacy of chemicals for control of cotton bollworm and
tobacco budworm, 1978. Insecticide and Acaricide Tests, Volume 4, Paper No. 219, p. 136-137.
Price, R. G., J. H. Young, D. Bogel, and B. Ree. 1978. Cotton bollworm and tobacco budworm control, 1977, Insec-
ticide and Acaricide Tests, Volume 3, Paper No. 177, p. 127.
J. H. Young, B. Ree, and F. Guilavogui. 1979. Cotton bollworm and tobacco budworm control, 1978. Insec-
ticide and Acaricide Tests, Volume 4, Paper No. 220, p. 137- 138.
Richmond, C. A. 1956. Tests with phosphorus insecticides for control of pink
bollworm and some other cotton pests, 1955. Journal of Eco- nomic Entomology 49:874-875.
Ridgway, R. L., H. J. Walker, R. L. Hanna, and W. L. Owen. 1967. Fertilizers impregnated with systemic insecticides for
control of cotton insects. Journal of Economic Entomology 60: 592-594.
Schwartz, P. H., and D. R. Hamel, editors. 1980. Guidelines for control of insect and mite pests of foods,
fibers, feeds, ornamentals, livestock, households, forests, and forest products. U.S. Department of Agriculture, Agriculture Handbook No. 571, 796 p.
Shipp, 0. E., and R. W. Earhart. 1967. Comparative seasonal abundance of Heliothis larvae in cot-
ton fields in Texas and Arkansas during summer 1964. Journal of Economic Entomology 60:893-898.
Smith, C. E., and I. J. Becnel. 1947. Effectiveness of different forms of nicotine in cotton
aphid control. Journal of Economic Entomology 40:536-539.
Smith, W. R., and S. L. Calhoun. 1950. Comparative effectiveness of aldrin applied at different
times of the day for boll weevil control. Journal of Economic Entomology 43:598-601.
and S. L. Calhoun. 1951. Spraying for early-season control of boll weevil. Journal
of Economic Entomology 44:919-920.
Taft, H. M., and A. R. Hopkins. 1967. Control of cotton pests with low-volume insecticides ap-
plied with a low-volume mist sprayer. Journal of Economic En- tomology 60:607-610.

LOSSES DUE TO COTTON INSECTS 357
Walker, J. K., Jr., A. R. Hopkins, and R. E. Fye. 1958. Effectiveness of several insecticides against the boll wee-
vil, bollworm, and cotton leafworm. Journal of Economic Ento- mology 51:783-786.
W. J. Mistric, Jr., and D. F. Martin. 1954. Evaluation of interval and dosage in bollworm control.
Journal of Economic Entomology 47:824-826.
Watson, T. F. 1976a. B. thurberiella control in cotton, 1975. Insecticide and AcaricTde Tests, Volume 1, Paper No. 124, p. 92-93.
1976b. P. gossypiella control in cotton, 1975. Insecticide and AcaricTde Tests, Volume 1, Paper No. 125, p. 93.
Watts, J. G. 1948. Cotton insect control with organic insecticides. Journal
of Economic Entomology 41:543-547.
Weaver, J. B., Jr., J. N. All, D. B. Weaver, and E. P. Hornyak. 1979. Influence of various insecticides on yield parameters of
two cotton genotypes. Journal of Economic Entomology 72:119-
123.
Williams, B. R., D. F. Glower, and K. L. Koonce. 1979. Evaluation of five insecticides for control of the banded- wing whitefly in cabbage and soybean loopers in cotton, 1977. Insecticide and Acaricide Tests, Volume 4, Paper No. 224, p. 140-141.
R. L. Rogers, D. F. Glower, and K. L. Koonce. 1979. Cotton insecticide efficacy on H. zea and H. virescens on
loessial hills of northeast Louisiana, 1977. Insecticide and Acaricide Tests, Volume 4, Paper No. 222, p. 139.
Wolfenbarger, D. A., R. L. McGarr, R. R. Longoria, and J. B. Nosky. 1970. Toxicity of EPN, accothion, and certain chlorinated hy-
drocarbons to certain cotton insects. Journal of Economic En- tomology 63:1568-1573.
Young, M. T. 1948. Control of boll weevil and cotton aphid with benzene hexa-
chloride and chlorinated camphene in 1947. Journal of Economic Entomology 41:401-403.
and R. C. Gaines. 1950. Tests of insecticides to control boll weevil, cotton
aphid, and twospotted spider mite. Journal of Economic Ento- mology 43:727-729.

358 P. H. SCHWARTZ
Young, M. T., and R. C. Gaines. 1953a. Tests of insecticides for control of cotton insects.
Journal of Economic Entomology 46:133-136.
and R. C. Gaines. 1953b. Control of insects and spider mites on cotton in 1952.
Journal of Economic Entomology 46:6^ -696.
and R. C. Gaines. 1954. Control of insects and spider mites on cotton in 1953.
Journal of Economic Entomology 47:753-756.
G. L. Smith, G. L. Garrison, and R. C. Gaines. 1945. Effect of nicotine applied with calcium arsenate at dif-
ferent times of day upon cotton aphids and yields. Journal of Economic Entomology 38:383-384.

359
Chapter 14
MODELS FOR COTTON INSECT PEST MANAGEMENT
A. W. Hartstack and J. A. Witz Pest Control Equipment and Methods Research Agricultural Research Service U.S. Department of Agriculture College Station, TX 77843
ABSTRACT Systems science techniques such as computer modeling are helping scientists take a new look at insect control, a major problem in cotton production. Models relating to the cot- ton plant, to important cotton insect pests, and, more recently, to beneficial insects have been developed. MOTHZV, a model for the bollworm (Heliothis zea (Boddie)) and tobacco budworm (Heliothis virescens (F.)), has been used suc- cessfully for 5 years in a statewide extension program in Texas. Updates to this model, in- cluding an improved cotton fruiting model, SIM- PLECOT, and a Heliothis spp. damage model, have recently made new research studies possible.
INTRODUCTION
Dramatic changes have been made in the control of cotton in- sect pests during the last 10-15 years because of developing in- sect resistance to insecticides, the increasing cost of energy, and concern over environmental quality. As a result, the cotton industry has been searching for alternative methods of producing a satisfactory crop, and several new systems have been proposed and tested. For example, cotton varieties having early, more rapid fruiting characteristics are being examined as are more efficient use of fertilizer and water, judicious use of insecticides, and maximum reliance on naturally occurring insect control agents. However, as this work has progressed, it has become clear that no single method of control can be expected to provide an acceptable solution to all cotton insect problems. The entire cotton produc- tion system must be examined, and many factors such as ecology and behavior must be considered in deciding what techniques are likely to be feasible and practical against a given pest (Knipling 1979). As a result, a new discipline, integrated pest management, based on the philosophy of total system consideration and multiple con- trol techniques, has developed.

360 A. W. HARTSTACK AND J. A. WITZ
In this chapter, after a discussion of the basic concepts un- derlying the systems approach, we describe several of the larger computer models that have been developed in the United States to study cotton pest management. A specific example, the cotton- Heliothis model, MOTHZV, is then described in some detail. We conclude with a discussion of the Texas Agricultural Extension Service program in which the MOTHZV model is used.
THE SYSTEMS APPROACH
Systems modeling has emerged from man's need to study and con- trol large and complex systems. The National Aeronautics and Space Administration moon exploration program is a prime example. Multi- discipline teams with specialized knowledge worked together with the aid of sophisticated computers and equipment to solve complex problems (Witz 1973). DeMichele and Bottrell (1976) discuss in considerable detail how the same approach can be used in cotton pest management to unify and guide research and produce a better understanding of interactions and the consequences.
The systems, or modeling, approach usually begins with simple mental models that a person formulates on the basis of his experi- ence (witz 1973):
"Watch out for the July 4th bollworm egglay." "Full moon occurs on July 10 so there will be large Heliothis spp. egglays about 2 to 4 days after the 10th."
"These summer showers will really bring on the boll weevil."
If these mental models are transformed to a flow chart, a set of equations, or a computer program, they become tangible and can be communicated to and used by other people. Several such tangible models may then be combined into more complex models through the use of computer and systems analysis techniques (Witz 1973).
The investigation of complex systems may result in the devel- opment of multiple models, each at different levels of detail. The level used in a given situation must be determined on the basis of both the intended purpose and the current knowledge avail- able. The study of an animal or plant, for example, can be subdi- vided into basic physiological processes, which, in turn, relate to various components or organs. Each organ can then be studied and modeled at the cellular level, the biochemical level, or, possibly, the molecular and atomic levels. Models developed at one level can often be simplified and made useful for applications at levels that require less precision.

MODELS FOR COTTON INSECT PEST MANAGEMENT 361
REVIEW OF COTTON PEST MANAGEMENT MODELS
The cotton plant (Gossypium hirsutum L.) is usually an inte- gral part of any modeling study of a cotton insect pest. On the one hand, the behavior and population dynamics of a given pest are affected by the plant; on the other hand, the insect affects the growth of the plant. For this reason it is important to our pur- pose here to review the development of several cotton plant models.
The first digital computer simulation of cotton growth was re- ported by Stapleton (1970) and Stapleton et al. (1973). It was used to study the effect of machinery design alternatives on the yield of cotton. SIMCOT, another early model, was developed by Duncan (1971) to calculate daily production and distribution of photosynthate as the cotton plant developed. McKinion et al. (1974, 1975) then used the same approach as Duncan (1971) in devel- oping Simcot II but included many additional factors that affect plant growth and maturity, among them the location and condition of individual fruiting points on the plant.
GOSSYM (Baker et al. 1977) is a more recent cotton plant model that stemmed from the SIMCOT research program. Many of the func- tional relationships are carried over from SIMCOT II, but a new plant growth dynamics algorithm is used, and a highly detailed model of soil, water, and plant-root relationships has been added.
A simpler, less detailed approach was reported by Wilson et al. (1972). By simulating the typical fruiting pattern of the cot- ton plant, they are able to adjust the rate of fruiting parametri- cally to simulate different varieties or different environments. In addition, the indeterminate behavior of cotton is modeled with a simple feedback mechanism, so the model can simulate regrowth subsequent to programmed removal of fruit. Thus, it has sufficient precision to be useful in many pest management applications.
In the early 1970's, SIMCOT II was revised by Gutierrez et al. (1975) to fit the California environment and cotton varieties. Calculation of fruit was converted from a single, average plant to a total field basis. Gutierrez et al. (1975, 1977a, 1977b) then incorporated several simple insect models into this revised version so they could study plant interactions with cotton defoliators such as the beet armyworm (Spodoptera exigua (Hübner)) and cabbage loop- er (Trichoplusia ni (HÜbner)) and fruit attackers such as Lygus hesperus Knight, Heliothis spp., and the pink bollworm (Pectinoph- ora gossypiella (Saunders)).
At about the same time, Stinner et al. (1974) described a Heliothis model that included a spatial grid of fields and crop types. Each crop was modeled on a field basis by using a measure

362 A. W. HARTSTACK AND J. A. WITZ
of the attractance of that crop for Heliothis spp. No prediction of yield was possible, but the approach allowed study of the move- ment of the insect population.
Jones (1975) and Jones et al. (1977) reported a detailed simulation model of the influence of weather, crop status, and management practices on populations of the boll weevil (Anthonomus grandis Boheman). This model has since been combined with a modified version of the SIMCOT II cotton model (Jones et al. 1980) and with a Heliothis spp. model derived from MOTHZV (Hartstack et al. 1976). It is being used to study improved pest management strategies (Brown et al. 1979) and is discussed in detail in Chapter 17 of this handbook.
Most recently, a cotton crop-boll weevil model has been devel- oped that is concerned with natural and induced mortality of the boll weevil in the field (Curry et al. 1981). Row spacing and ori- entation are used to calculate light penetration through the crop canopy so as to achieve a more accurate measure of the microclimate under the plant where the immature boll weevil develops. Damage to the plant is estimated based on the size of the adult population, and yield can be predicted. This model is currently being field tested and will be used to test alternative control strategies.
THE HELIOTHIS POPULATION MODEL: MOTHZV
The model for Heliothis spp. designed by Hartstack et al. (1973), as shown by flow diagram in Figure 1, was not complex. However, Hartstack and Hollingsworth (1974) converted it to mathe- matical equations and used a computer to predict the number and time of emergence of the subsequent generation of adults. A de- gree-day concept was used with development times to calculate the total length of each generation (adult to adult). For example, rates of increase were input as constants for each day by using light trap data collected at College Station, TX. Moonlight was the only other factor considered to affect rate of increase. Timing and size of future generations were accurate for College Station, TX, but this regression-type model lacked versatility and could not be used at other locations.
The 1975 Version: MOTHZV-2
MOTHZV-2, a descendant of the original MOTHZV, was described by Hartstack et al. (1976). It incorporated considerably more de- tailed population and physiology algorithms (Figure 2). It also included three simple crop models—corn (Zea mays L.), cotton, and sorghum (Sorghum bicolor (L.) Moench). The FORTRAN computer pro- gram for MOTHZV-2 consisted of a main computer program and 16 subroutines.

MODELS FOR COTTON INSECT PEST MANAGEMENT 363
Input Catch Per Trap Corn, Cotton,
etc. Day "A"
Convert Trap Catch to Moths Per Acre
I Calculate Generation Length Expected fron^i
Day "A"
I M Store Day "B" Next Generation
Day No.
Calculate Rate of Increase Expected from Day "A" to Day "B" for
Each Crop H (Rate of Increase in Each Crop) x (No. Moths/
Acre in Crop)
Sum Projected Populations in
Each Crop
I M I
(Projected Population) x (% of Crop in Area)
Calculate Moonlight Effect and Adjust
Projected Population
I Calculate Longevity
of Projected Population and Store Accordingly
Output Population Projected
and Day "B"
Stop j Figure 1.—Flowchart of original MOTHZV concept (Hartstack et al.
1973).

364 A. W. HARTSTACK AND J. A. WIT2
Mk/UL.1
C
D
S n
V® ¡ GENLN
V© ;\© i VS Ï V2 Ä 0 TEMP 50
EGGS 0 TEMP 50 0 TEMP. 50 0 TEMP. 50 0 TEMP. 50
1-3 INSTAR 4-5 INSTAR PUPA PREOVIPOSITION O D
S 0
D TEMP. 50 OVIPOSITION
0 0 y ^''^^'^S ' LARVAE
CANNIBALIS
CANBAL
-u °' NAT MOF
365 1TALITY
Figure 2.—Flowchart of MOTHZV-2 (Hartstack et al. 1976).

MODELS FOR COTTON INSECT PEST MANAGEMENT 365
With MOTHZV-2, predictions concerning the timing and size of future generations of Heliothis spp. were made by using expected (long-term average) temperatures and projected crop phenology. Simulations were initialized with numbers of eggs or moths and were carried through the number of generations that occur at a given locality during one season. A bookkeeping system was used to re- cord the number of eggs, first- to third-instar larvae, fourth- and fifth-instar larvae, pupae, preovipositing adults, and ovipositing adults for each day of the simulation. Each stage of the insect could be advanced one calendar day at a time by using the degree- day concept so daily mortality (natural, insecticide, parasites, and predators) could be considered. The output of MOTHZV-2 con- sisted of tables or graphs showing the numbers of Heliothis spp. of each stage present on each day.
The three crop models, like MOTHZV-2, were temperature de- pendent, and the degree-day concept was used to predict crop phe- nology (e.g., first square, boll, and open boll). These events were then used to predict the relative attraction of the crop to Heliothis spp. females, since this attraction has a considerable effect on oviposition. No prediction of yield was possible, and a separate run of the model was required for each crop. However, the numbers of adult Heliothis zea (Boddie) predicted to migrate out of corn or sorghum were held in a vector that could be used as input for a subsequent run with the cotton model.
Rather than attempt an in-depth mathematical description of the model, we refer readers to the reports of Hartstack et al. (1973, 1976).
The 1978 Version: MOTHZV-3
In 1978, MDTHZV-2 was revised by replacing the original cotton model with a more dynamic crop model and by adding a Heliothis dam- age model. This revision allowed the user to study changes in cot- ton yield caused by Heliothis. Since this latest version has not been reported, the changes are described here in detail.
The Cotton Submodel: SIMPLECOT
The submodel for cotton fruiting behavior, SIMPLECOT, was de- rived from the earlier model developed by Wilson et al. (1972). The number of new fruit-per-plant per degree-day (FRT) was given
by: FRT=(V+0.00288xFRTMX)x(l-0.0003xFRTPTxTOTBOL) (1)
where V = Variety factor: 0.054 - 0.072 for Delta varieties, e.g., Stoneville 213 0.072 - 0.100 for short-season varieties, e.g., TAMCOT-
SP37

366 A. W. HARTSTACK AND J. A. WITZ
FRTPT = Accumulated fruiting points per plant - a measure of the size of the plant
TOTBOL = Total number of bolls (small, large, and open) on plant
FRTMX = The smaller of the values FRTPT and 41.38xYIELD YIELD = The expected yield in bales per acre
The FRTPT in combination with the TOTBOL thus cut off square pro- duction when all the photosynthate production had been used for maintenance of the plant and its fruit load. If the calculation of fruiting points became negative, it was set to zero.
Natural shedding of fruit was based on the load of green bolls and temperature The probability of fruit survival was calculated as:
PRSUR = (0.9995-0.0001xTOTGRB)xDDAY (2)
where TOTGRB = Total green bolls. (With average conditions, about 75% of
the total fruit produced would be shed.) DDAY = Degree-days for each day. (Degree-days were calculated
by using 12.6 C and 33.3 C as lower and upper thresholds, respectively, as described by Hartstack et al. (1976) for Heliothis development.)
Although soil type, fertility, and moisture influence the rate of production and retention of fruit, they were not included in this model. Rather, they were assumed to be adequate to produce the expected yield.
The plant model was converted to a per acre basis since all the insect data in MOTHZV-3 were calculated on that basis. Multi- plying the fruit production by the expected yield per acre for av- erage conditions gave the accumulated fruit per acre as:
FRUIT = FRTPTx5000xYIELD. (3)
The model thus must show production of about 150,000 open bolls for each 1.0 bale of predicted yield.
The SIMPLECOT routine was not called up by MOTHZV unless squar- ing had commenced. It was assumed that 378 degree-days were re- quired from planting to first square, 317 degree-days from first square to first boll, 173 degree-days from first boll to large green bolls, and 448 degree-days from large green boll to open boll. The rate of production of fruit was adjusted by expected yield in bales per acre and the variety factor. Each cohort of fruit was moved from one stage to the next as a group (no distribu- tion). The model tabulated the age and number of each fruit cohort as it developed.
Fruiting curves generated by the model for two cotton varie* ties are shown in Figure 3. In this case, the variety factor (V)

MODELS FOR COTTON INSECT PEST MANAGEMENT 367
Relative number of fruit
Squares
1 140
I 150
1 160
1 170
1 180
1 1 190 200
Day no.
1 210
1 220
"H T" 230 240
—I 1 250 260
Figure 3.—Comparison of fruiting curves produced by SIMPLECOT for TAMCOT-SP37 and Stoneville 213 cotton varieties.
was set at 0.054 to simulate a traditional Delta variety, e.g., Stoneville 213, and set at 0.1 to simulate a newer, faster fruit- ing, more determinant variety, e.g., TAMCOT-SP37. The yield was projected at 2 bales per acre for each.
The simulation produced by the submodel is compared in Figure 4 with actual field counts of squares and bolls produced by Stone- ville 213 at two locations (Figures 4a and 4b) and by TAMCOT-SP37 at College Station, TX (Figure 4c).

368 A. W. HARTSTACK AND J. A. WITZ
100000
"X Field \ Counts Model
\ ° Sq. \ ■ Gr. Bolls
\ —— Op. Bolls
1 1 I 230 240 250
Figure 4a.—Comparison of field counts of cotton fruit with output from SIMPLECOT, Stoneville 213, Pearsall, TX, 1973.
800000
600000
400000
200000
-| ^ I I I I I I I ^ I
140 150 160 170 180 190 200 210 220 230 240 Day no.
Figure 4b.—Comparison of field counts of cotton fruit with output from SIMPLECOT, Stoneville 213, College Station TX, 1974.
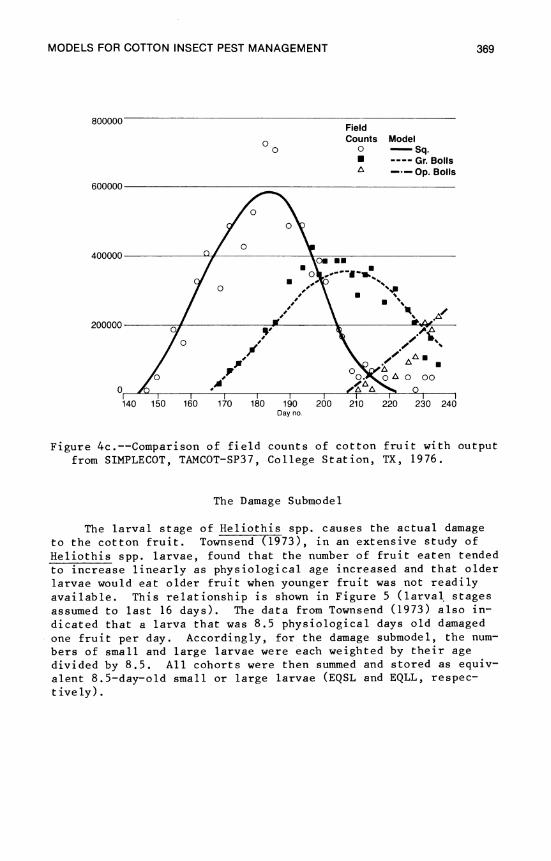
MODELS FOR COTTON INSECT PEST MANAGEMENT 369
800000" Field
0 Counts Model o o Sq.
■ Gr. Bolls A ——Op. Bolls
600000-
400000
200000
140 150
Figure 4c.—Comparison of field counts of cotton fruit with output from SIMPLECOT, TAMCOT-SP37, College Station, TX, 1976.
The Damage Submodel
The larval stage of Heliothis spp. causes the actual damage to the cotton fruit. Townsend (1973), in an extensive study of Heliothis spp. larvae, found that the number of fruit eaten tended to increase linearly as physiological age increased and that older larvae would eat older fruit when younger fruit was not readily available. This relationship is shown in Figure 5 (larval stages assumed to last 16 days). The data from Townsend (1973) also in- dicated that a larva that was 8.5 physiological days old damaged one fruit per day. Accordingly, for the damage submodel, the num- bers of small and large larvae were each weighted by their age divided by 8.5. All cohorts were then summed and stored as equiv- alent 8.5-day-old small or large larvae (EQSL and EQLL, respec- tively) .
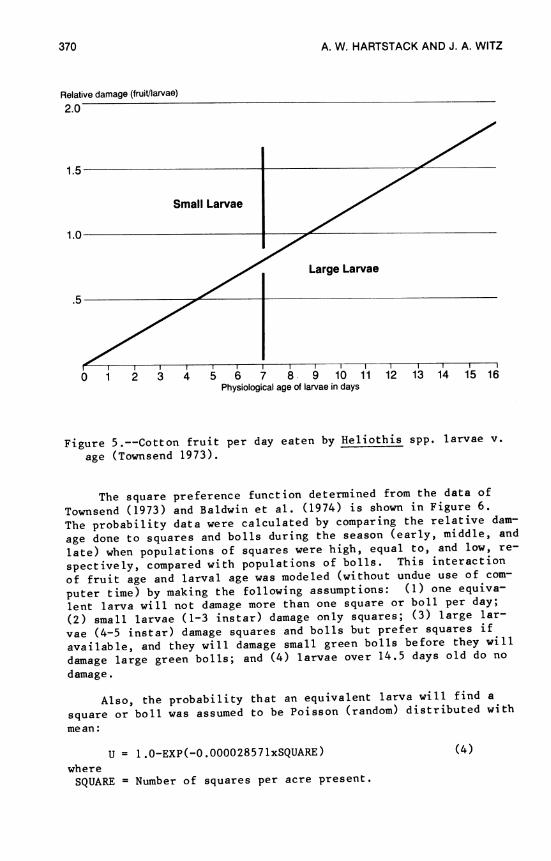
370 A. W. HARTSTACK AND J. A. WITZ
Relative damage (fruit/larvae)
2.0
n 1 1 1 r 5 6 7 8 9 10 11
Physiological age of larvae in days
T 1 1
13 14 15 16
Figure 5.—Cotton fruit per day eaten by Heliothis spp. larvae v. age (Townsend 1973).
The square preference function determined from the data of Townsend (1973) and Baldwin et al. (1974) is shown in Figure 6. The probability data were calculated by comparing the relative dam- age done to squares and bolls during the season (early, middle, and late) when populations of squares were high, equal to, and low, re- spectively, compared with populations of bolls. This interaction of fruit age and larval age was modeled (without undue use of com- puter time) by making the following assumptions: (1) one equiva- lent larva will not damage more than one square or boll per day; (2) small larvae (1-3 instar) damage only squares; (3) large lar- vae (4-5 instar) damage squares and bolls but prefer squares if^ available, and they will damage small green bolls before they will damage large green bolls; and (4) larvae over 14.5 days old do no
damage.
Also, the probability that an equivalent larva will find a square or boll was assumed to be Poisson (random) distributed with
U= 1.0-EXP(-O.000028571xSQUARE)
where SQUARE = Number of squares per acre present.
(4)
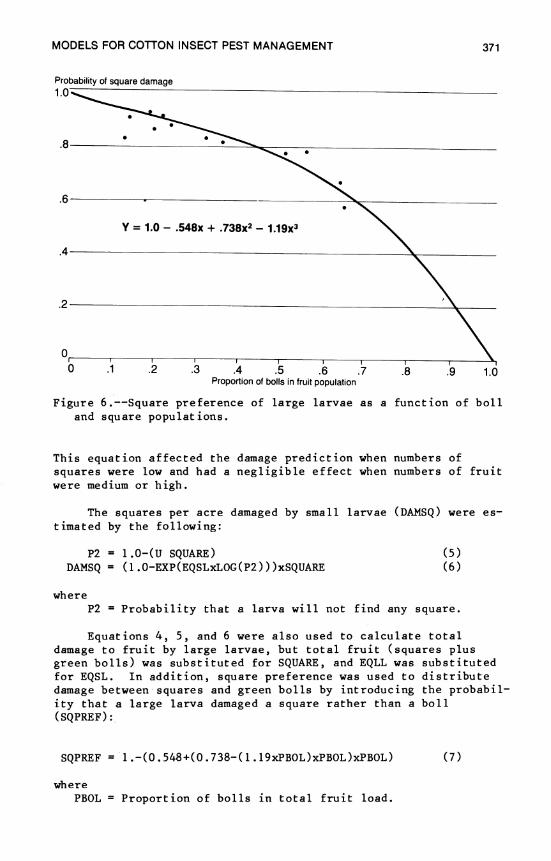
MODELS FOR COTTON INSECT PEST MANAGEMENT 371
Probability of square damage
1.0
-T —I \ 1 1 -r •3 .4 .5 .6 .7 .8 .9 1.0
Proportion of bolls in fruit population
Figure 6.—Square preference of large larvae as a function of boll and square populations.
This equation affected the damage prediction when numbers of squares were low and had a negligible effect when numbers of fruit were medium or high.
The squares per acre damaged by small larvae (DAMSQ) were es- timated by the following:
P2 = 1.0-(U SQUARE) (5) DAMSQ = (1.0-EXP(EQSLxLOG(P2)))xSQUARE (6)
where P2 = Probability that a larva will not find any square.
Equations 4, 5, and 6 were also used to calculate total damage to fruit by large larvae, but total fruit (squares plus green bolls) was substituted for SQUARE, and EQLL was substituted for EQSL. In addition, square preference was used to distribute damage between squares and green bolls by introducing the probabil- ity that a large larva damaged a square rather than a boll (SQPREF):
SQPREF = l.-(0.548+(0.738-(1.19xPBOL)xPBOL)xPBOL) (7)
where PBOL = Proportion of bolls in total fruit load.

372 A. W. HARTSTACK AND J. A. WITZ
The total damage to squares and green bolls thus calculated was used to reduce each cohort of fruit to complete the damage algorithm.
Results of tests made to validate the damage model are shown in Table 1. The values compare favorably with the data of Townsend (1973) and Baldwin et al. (1974). Moreover, Quaintance and Brues (1905) found that on the average, one larva would damage eight squares, 1 2/3 bolls, and one flower (considered a boll), precisely what the model predicted.
The addition of a dynamic cotton crop model,, SIMPLECOT, and the Heliothis spp. damage model made possible new studies with MOTHZV-3 that were not previously possible. The cotton model can be adjusted for a particular simulation by varying the two input parameters, variety factor, and yield. A base run with no damage can be made. Then the effect on yield of various insect popula- tions or pest management decisions can be studied by comparing sub- sequent runs with the base run or to each other.
A STATEWIDE EXTENSION PROGRAM: BUGNET
BUGNET, a computerized pest management delivery system, was developed for use in Texas by entomologists of the Texas Agricul- tural Extension Service and research cooperators. It consists of making computer models such as MOTHZV and other BUGNET programs available to the personnel of the Extension Service and thereby to producers across the State for use in making decisions about pest control.
A statewide pest management plan of this type was first pro- posed for Texas (Frisbie and Adkisson 1975) in 1974 and was tested in pilot programs in 1973, 1974, and 1975. Results of these tests were so encouraging that the Texas Association of Cotton Producer Organizations asked the Texas Agricultural Extension Service, the Texas Agricultural Experiment Station, and the Texas Department of Agriculture to develop and implement such a plan. This was done in 1976.
Table 1.—Validation of the damage model
Test Squares Bolls No. damaged per larva season per acre per acre Squares Bolls
Early 100,000 Mid 270,000 Late 200,000
0 10.5 n.a. 70,000 9.5 1.3
200,000 8.0 2.8

MODELS FOR COTTON INSECT PEST MANAGEMENT 373
The statewide plan consisted of 10 subplans developed by indi- vidual subcommittees. Nine of the plans were local, that is, they were tailored for the nine geographic areas of Texas that are in- volved in cotton and sorghum production. The tenth plan, TAMU- BUGNET, was designed to investigate the potential of a computer- based system for forecasting populations of insect pests and crop yields. The goal of all 10 plans was to aid farmers in making decisions concerning insect control and crop management.
Phase One
The intent of BUGNET was to make models such as MOTHZV avail- able to the county extension entomology specialists. Phase One of BUGNET was operated from a headquarters in College Station, TX, and the central computer at Texas A&M University was used to run the models.
Necessary information was obtained by installing insect traps in each of the areas and having the specialist monitor the number ^^ Heliothis spp. taken in each daily catch. The traps were in- stalled as early as possible in the spring so as to monitor the emergence of the diapausing (overwintered) moths. The time the traps were put out ranged from February for the Rio Grande Valley to May for the Rolling Plains area. Traps were located within early-season hosts of H_. zea (corn or sorghum or both), depending on which was the major crop in the area. When cotton began to square or when the corn or sorghum became mature, the traps were moved into cotton.
Additional input data gathered by the specialist for MOTHZV were as follows: (1) minimum and maximum daily temperature; (2) planting date, emergence date, and date of first fruit (silk, bloom, square, boll) for each of the three crops (corn, sorghum, cotton) present in the area; and (3) the percentage of each of the three crops in each area (one or multiple counties).
The first batch of data was available for use about 60 days after the traps were installed. Data were mailed to the headquar- ters by the extension entomologist. Thereafter, data were for- warded whenever it seemed advisable to update a forecast, usually every 7-14 days.
As soon as the data arrived at College Station, they were punched on computer cards, and MOTHZV (stored online on disk at the Texas A&M Data Processing Center) was run. The output, that is, the forecast, was then mailed to the extension entomologist and other interested personnel. The forecast included the time of oc- currence and the relative size of the expected egg and larval pop- ulations of 11. zea and H. virescens for the entire season. Recipi- ents were cautioned that the numbers indicated the anticipated trend and should not be used as predictions of actual numbers in specific fields.

374 A. W. HARTSTACK AND J. A. WITZ
In 1976, three light traps (Hollingsworth and Hartstack 1972) were installed at each of five test areas. The egg peaks fore- cast by MOTHZV-2 on the basis of the data obtained at these loca- tions predicted peaks that actually occurred within 3 days of the prediction. Figure 7 shows an example of the predicted and actual egg counts of tl. zea for one location. The successful results of this first year were reported by Hartstack et al. (1977).
In 1977, the BUGNET-MOTHZV program was expanded from 5 areas to a total of 15. In addition, two or three electric grid phero- mone traps (Wolf et al. 1972) baited with virelure (Tumlinson et al. 1975, Hendricks et al. 1977), a synthetic attractant for ji. vires- cens, were installed at each location. The catches of these phero- mone traps were used as inputs to MOTHZV-2 for predictions of E, virescens oviposition.
In 1978, MOTHZV-2 was replaced with MOTHZV-3, which included the more dynamic cotton model, SIMPLECOT. The next year the grid traps were replaced with cone traps (Hartstack et al. 1979) that required no electricity and thus were much more versatile. The program has continued to operate in about 15 areas since 1977.
Relative number of eggs
— Predicted June 2,1976—MOTHZV-2 Actual Egg Counts (Hillsboro Area)
—r~ 21
—T" 11
.-./-y^ April May
31 20 June
Figure 7.—A comparison of the June forecast of Heliothis zea ovi- position with the actual field egg counts, Hillsboro, TX, 1976.

MODELS FOR COTTON INSECT PEST MANAGEMENT 375
Phase Two
The second phase of the overall BUGNET program was begun in 1977. Computer power was placed directly in the hands of the area and county extension specialists by purchasing and installing IBM 5100 minicomputers at three locations. In 1979, MOTHZV-4, a simplified version of MOTHZV-3 written in APL, was sent to the field for use on these minicomputers. The version lacked much of the versatility of MOTHZV-3 but could successfully predict timing of Heliothis spp. oviposit ion. Phase Two was expanded in 1980 by adding four new computers and upgrading the original three so that seven IBM 5110 computers are now available to area and county extension personnel at seven locations. Other software packages made available to BUGNET users include a Cotton Budget Generator, an Insecticide Compatibility Chart, Cotton Production Games, an IBM Statistics Pack, and a Boll Weevil Model.
Five Years of BUGNET: The Results
Forecasts during the first year of BUGNET were of limited use to farmers, both because there was doubt about reliability and be- cause populations were low and of little economic importance except for a few isolated locations. However, confidence in the program has grown steadily each year. Over the 3 years, 1977-79, the pre- dictions on the timing of peak egg production were within 4 days of the actual field peaks ca. 70% of the time, this despite the fact that in some areas and in some years, Heliothis spp. populations were low, and there actually were no significant peaks in the field. When populations were low, the probability of error was as great in the field data as in MOTHZV. Thus, some misses might not have occurred if more accurate egg peaks could have been determined from field data.
Although MOTHZV has never been recommended as the exclusive basis for a decision to apply insecticide for control of Heliothis spp., the advance information concerning possible infestations alerted producers who then could intensify scouting efforts. Also, in areas where crops were irrigated, early warning of Heliothis outbreaks has been valuable to those who need this knowledge to time irrigations properly (Slosser 1980). This was because the rapid plant growth and lush foliage that resulted from irrigation were extremely conducive to Heliothis spp. egg lay. Thus it was advisable to avoid irrigation when adult insect populations were high.
Finally, in areas where other insect pests such as the boll weevil and the cotton fleahopper (Pseudatomoscelis seriatus (Reu- ter)) were problems early in the season before Heliothis spp. in- festations occurred, any application of pesticides for control of these early-season pests must be terminated several weeks before Heliothis spp. populations build up so populations of beneficial insects can recover enough to help with their control. Advance

376 A. W. HARTSTACK AND J. A. WITZ
knowledge of when Heliothis spp. were likely to build helped determine the best cutoff date.
The Texas Agricultural Extension Service released the following statement in 1979:
"In 1979, approximately 5,740 cotton producers in 91 counties used this tool (MOTHZV) to help manage 4,069,300 acres of cotton. It is conserva- tively estimated that these producers realized a combined net benefit of $4,967,000. In addition to this, there are intangible benefits (i.e., peace of mind from knowing you*re doing the right thing) that cannot be measured in dollars. It is reasonable to assume that net benefits comparable to this have also been realized in 1977 and 1978, as MOTHZV has enjoyed the same widespread use over the past three years."
The Future for MOTHZV and BUGNET
Although the value of the systems approach to pest management in cotton has not been fully tested and evaluated, research gaps and priorities have been identified. The model itself needs im- provement; it is especially lacking in regard to the impact of beneficial insect species and the effects of rainfall and soil moisture. In addition, early-season trapping data are not as pre- cise as they should be.
At present, natural insect predators and parasites are being studied to determine their occurrence and their ability to control pest species; however, little basic biological data are available concerning these beneficial species. Such information will be re- quired if these native beneficial insects are to be managed and augmented by distribution of reared beneficial species. Also, im- proved methods of sampling beneficial populations to determine their occurrence and number will be necessary before management and releases of beneficiáis will become practicable on a large scale.
MOTHZV-3 does not consider the effect of rainfall and soil moisture on the growth of the crop despite the obvious effects of soil moisture on lushness and the number of eggs laid by Heliothis spp. moths. Moisture also affects mortality of eggs and larvae. However, before these factors can be incorporated in the model, methods of monitoring rainfall and soil moisture over a wide area are needed. Remote sensing might be useful in this application.
The matter of early-season trap catches is another problem. Light traps (Hartstack et al. 1971, 1973) have proved useful in monitoring numbers of Heliothis spp. adults, but there are problems. Such traps must have an electrical power source, which often inter- feres with field operations. Moreover, the catches are sometimes

MODELS FOR COTTON INSECT PEST MANAGEMENT 377
difficult to identify. In addition, experience with MOTHZV-BUGNET has shown that at least 3 and perhaps 5-10 traps must be operated within each area to achieve adequate data. That would be expen- sive, time consuming, and troublesome so numbers have usually been sacrificed for convenience. Finally, the matter of locating the traps in the proper crop and at the proper place can be diffi- cult because of power requirements. These problems, plus irregu- lar servicing, have contributed substantially to prediction errors.
The pheromone traps (Hartstack et al. 1979) and synthetic sex pheromones (Klun et al. 1979) that have recently been developed for H. zea and H. virescens may provide a more convenient and effective way to monitor Heliothis spp. adults. The traps are highly effi- cient, simple to construct, inexpensive, and portable (requiring no power). Also, only a single species of Heliothis is attracted, which makes identification and counting quick and easy. Since these pheromones are now commercially available, the desired array of 5-10 traps per area can be located within the proper crops, and monitoring should not be difficult. Additionally, with pheromone traps, spring emergence of moths can be detected 2-3 weeks earlier than with light traps.

378 A. W. HARTSTACK AND J. A. WITZ
LITERATURE CITED
Baker, D. N. , J. R. Lambert, C. J. Phene, and J. M. McKinion. 1977. GOSSYM: a simulator of cotton crop dynamics. Tn Compu-
ters Applied to the Management of Large Scale Agricultural En- terprises. Ln Proceedings, United States-Union of Soviet So- cialist Republics Seminar, Moscow, Riga, Kishinev, p. 100-133.
Baldwin, J. L., J. K. Walter, J. R. Gannaway, and G. A. Niles. 1974. Bollworm attack on experimental semidwarf cottons. Texas A&M University. Texas Agricultural Experiment Station Bulletin No. 1144, 11 p.
Brown, L. G., R. W. McClendon, and J. W. Jones. 1979. Computer simulation of the interaction between the cotton
crop and insect pests. Transactions of the ASAE 22:771-774.
Curry, G. L., P. J. H. Sharpe, D. W. DeMichele, and J. R. Cate. 1981. Towards a management model of the cotton boll weevil eco-
system. In IIASA Pest Management Network, Volume 6. [in press.]
DeMichele, D. W., and D. G. Bottrell. 1976. Systems approach to cotton insect pest management. Iri In-
tegrated Pest Management, p. 197-232. Plenum Press, New York,
Duncan, W. G. 1971. SIMCOT: a simulator of cotton growth and yield. In Pro-
ceedings, Workshop on Tree Growth Dynamics and Modeling, Duke University, October 11-12, 1971, p. 115-118.
Frisbie, R. E., and P. L. Adkisson. 1975. Summary: statewide pest management plan. Texas Agricul-
tural Extension Service, Texas A&M University System, College Station.
Gutierrez, A. P., G. D. Butler, Jr., Y. Wang, and D. Westphal. 1977a. The interaction of pink bollworm (Lepidoptera: Gelichi-
idae), cotton, and weather: a detailed model. Canadian Ento- mologist 109:1457-1468.
L. A. Falcon, W. Loew, P. A, Leipzig, and R. van den Bosch.
1975. An analysis of cotton production in California: a model for Acala cotton and the effects of defoliators on its yield. Environmental Entomology 4:125-136.
T. F. Leigh, Y. Wang, and R. D. Cave. 1977b. An analysis of cotton production in California: Lygus hesperus (Heteroptera: Miridae) injury—an evaluation. Cana- dian Entomologist 109:1375-1386.

MODELS FOR COTTON INSECT PEST MANAGEMENT 379
Hartstack, A. W., Jr., J. L. Henson, J. A. Witz, J. A. Jackman, J. P. Hollingsworth, and R. E. Frisbie.
1977. The Texas program for forecasting Heliothis spp. infesta- tions on cotton. ^E Proceedings Beltwide Cotton Production Research Conference (National Cotton Conference, Memphis, TN), p. 151-154.
and J. P. Hollingsworth. 1974. A computer model for predicting Heliothis populations. Transactions of the ASAE 17:112-115.
J. P. Hollingsworth, R. L. Ridgway, and J. R. Coppedge. 1973. A population dynamics study of the bollworm and the tobac-
co budworm with light traps. Environmental Entomology 2:244-
252.
J. P. Hollingsworth, R. L. Ridgway, and H. H. Hunt. 1971. Determination of trap spacings required to control an in-
sect population. Journal of Economic Entomology 64:1090-1100.
J. A. Witz, and D. R. Buck. 1979. Moth traps for the tobacco budworm. Journal of Economic
Entomology 72:519-522.
J. A. Witz, J. P. Hollingsworth, R. L. Ridgway, and
J. D. Lopez. 1976. MOTHZV-2: a computer simulation of Heliothis zea and Heliothis virescens population dynamics. Users manual. U.S. Department of Agriculture, ARS-S-127, 55 p.
Hendricks, D. E., A. W. Hartstack, Jr., and T. N. Shaver. 1977. Effect of formulations and dispensers on attractiveness
of virelure to the tobacco budworm. Journal of Chemical Ecol- ogy 3:497-506.
Hollingsworth, J. P., and A. W. Hartstack, Jr. 1972. Effect of components on insect light trap performance.
Transactions of the ASAE 15:924-927.
Jones, J. W. 1975. A simulation model of boll weevil population dynamics as
influenced by the cotton crop status. Ph.D dissertation. North Carolina State University, Raleigh, 254 p.
H. D. Bowen, R. E. Stinner, J. R. Bradley, Jr., and J. S. Bacheler.
1977. Simulation of boll weevil populations as influenced by weather, crop status, and mangement practices. Transactions of the ASAE 20:121-125, 131.

380 A. W. HARTSTACK AND J. A. WITZ
Jones, J. W., L. G. Brown, and J. D. Hesketh. 1980. COTCROP: a computer model for cotton growth and yield.
Chapter 10. _In Predicting Photosynthesis for Ecosystems Mod- els. CRC Press, West Palm Beach.
Klun, J. A., J. R. Plimmer, B. A. Bierl-Leonhardt, A. N. Sparks, and 0. L. Chapman.
1979. Trace chemicals: the essences of sexual communication systems in Heliothis spp. Science 204:1328.
Knipling, E. F. 1979. The basic principles of insect population suppression and management. U.S. Department of Agriculture, Agriculture Hand- book No. 512, 659 p.
McKinion, J. M., D. N. Baker, J. D. Hesketh, and J. W. Jones. 1975. SIMCOT II: a simulation of cotton growth and yield. In
Computer Simulation of a Cotton Production System. Users man- ual. U.S. Department of Agriculture, ARS-S-5^, p. 27-82.
J. W. Jones, and J. D. Hesketh. 1974. Analysis of SIMCOT: photosynthesis and growth. Tn Pro-
ceedings, Beltwide Cotton Production Research Conference (National Cotton Conference, Memphis, TN), p. 117-124.
Quaintance, A. L., and C. T. Brues. 1905. The cotton bollworm. U.S. Department of Agriculture, Bu-
reau of Entomology Bulletin No. 50, 155 p.
Slosser, J. 1980. Irrigation timing for bollworm management in cotton. Journal of Economic Entomology 73:346-349.
Stapleton, H. N. 1970. Crop production in system simulation. Transactions of the ASAE 13:110-113.
D. R. Buxton, F. L. Watson, D. J. Nolting, and D. N. Baker.
1973. COTTON: a computer simulation of cotton growth. Arizona Agricultural Experiment Station, Technical Bulletin No. 206, 124 p.
Stinner, R. E., R. L. Rabb, and J. R. Bradley. 1974. Population dynamics of Heliothis zea and H. virescens in North Carolina: a simulation model. Environmental Entomology 3:163-168.
Townsend, J. R., Jr. 1973. Economic threshold studies of Heliothis spp. on cotton. M.S. thesis. Department of Entomology, Mississippi State Uni- versity, Mississippi State, 72 p.

MODELS FOR COTTON INSECT PEST MANAGEMENT 381
Tumlinson, J. H., D. E. Hendricks, E. R. Mitchell, R. E. Doolittle, and M. M. Brennan.
1975. Isolation, identification and synthesis of the sex phero- mone of the tobacco budworm. Journal of Chemical Ecology 1: 203-214.
Wilson, A. G., R. D. Hughes, and N. Gilbert. 1972. The response of cotton to pest attack. Bulletin of Ento- mological Research 61:405-414.
Witz, J. A. 1973. Integration of systems methodology and scientific re-
search. Agricultural Science Review 11:37-48.
Wolf, W. W., H. H. Toba, A. N. Kishaba, and N. Green. 1972. Antioxidant to prolong the effectiveness of cabbage looper
sex pheromone in the field. Journal of Economic Entomology 65: 1039-1041.


SECTION IV ALTERNATIVE PROGRAMS


385
Chapter 15
OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI
J. L. Hamer, G. L. Andrews, R. W. Seward, D. F. Young, Jr., and R. B. Head Mississippi Cooperative Extension Service Mississippi State, MS 39762
ABSTRACT The 3-year Optimum Pest Management Trial (OPMT) provided a thorough test of avail- able technology for suppression of the boll weevil (Anthonomus grandis Boheman) during the period when it enters diapause in the late sum- mer and fall and for in-season management of other insect and arthropod pests of cotton. The number of in-season insecticide applications was lowered to an average of 3.3 by the end of the Trial. In 1978, the total insect control cost for 3.3 applications including the scouting fee was $20.47 per acre. During 1979, the total in- sect control cost for 3.41 treatments was $19.36 per acre. In 1980, the total insect control cost for an average of 3.01 applications was $17.50 per acre. In addition, the presence with- in the Trial area and efforts of well trained Extension pest management specialists who worked closely with producers, private consult- ants, aerial applicators, county Extension per- sonnel, and others achieved a unified effort by all cooperators. These cooperative efforts resulted in an expansion of cotton acreage from 32,075 to ca. 39,000 acres from 1978 to 1980. Participation by growers in the scouting pro- gram increased each year of the Trial—98.7, 99.6, and 99.7% of the cotton acreage was in- cluded in 1978, 1979, and 1980, respectively. Yields of lint were increased an average of 84 pounds per acre during the Trial period and an average 34 pounds per acre above the 10-year average.
Through the use of correctly timed pin- head square and diapause control applications and precise monitoring of boll weevil activity with pheromone traps, significant boll weevil suppression was attained. Modified Mitchell traps were used to monitor weevils in both

386 J. L HAMERETAL
Pontotoc, a Current Insect Control Practice Area, and Panola County, the test area. Dur- ing the fall of 1978, 1979, and 1980, up to four diapause applications were applied on most of the participating acreage in Panola County. As a result, in the spring of 1979 and 1980, weevil captures were 78 and 94% lower, respec- tively, in Panola County than in Pontotoc Coun- ty. Also, weevil captures in line traps in- stalled at 2-mile intervals extending into four counties in north, south, east, and west direc- tions from Panola County indicated higher wee- vil numbers near or outside the boundaries of Panola County. The need for pinhead square treatment in the test area was limited because the areawide diapause control treatments proved highly effective. Only 57 acres received a pin- head square insecticide treatment during the the Trial, and that was in 1980.
Participation in the Trial required that producers have their acreage scouted. Private consultants scouted 75% of the acreage and pro- ducers, 15%; the remaining 10% was monitored by the Panola County Pest Management Society.
Other benefits of the program to the pro- ducers included improved understanding of the role of beneficial arthropods, economic thresh- olds, and timing of insecticide treatments in the control of cotton insect pests. In addi- tion, there was improved understanding of re- lated agronomic practices such as matching plant densities to soil types and the advan- tages of new methods of fertilization and weed control.
INTRODUCTION
In 1976, States in the Cotton Belt were invited by the U.S. Department of Agriculture Boll Weevil Interagency Working Group to submit plans for a State pest management program that would include all implementable technological advances. This pest management program received the title "Optimum Pest Management (OPM)." Re- sults would be compared with those of the Boll Weevil Eradication Trial (BWET), which was to be carried out concurrently in North Carolina. After extensive evaluation of the responses submitted, the Panola-Pontotoc area of Mississippi was selected as the area where the program would be conducted because it was similar to the

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 387
area in North Carolina where the BWET was to be conducted. Both trials took place during the same 3-year period and were evaluated as alternative methods of improving cotton insect management throughout the Cotton Belt. Panola County was the site of the Op- timum Pest Management Trial (OPMT); Pontotoc County, a comparable area in Mississippi, was the control (Current Insect Control; CIC), that is, the producers in this county used their normal procedures. Panola County is located ca. 50 miles south of Memphis, TN, in the edge of the Delta foothills. About 65% of the acreage in Panola County is hill or river bottom; the Mississippi Delta makes up the rest of the acreage. In 1977, 32,075 acres of cotton were planted in Panola County. However, this acreage can be expanded (to more than 40,000 acres) if profit margins are such as to make cotton a more desirable crop. The average field size is ca. 25 acres with a range from 1 to 100 acres per field. Before the Trial, organized scouting was conducted in the county by private agricultural con- sultants, producers, business firms, and Extension Service personnel.
Past experience has shown that in Mississippi effective use of pest management strategies will improve net profits for producers (Collins et al. 1976). For example, prior to 1972, some Missis- sippi cotton producers relied heavily on insecticides to protect their crops: they made as many as 18 scheduled applications with- out regard to thresholds or scouting (Moody et al. 1973). Other producers did not treat at all and so had lower yields. With the introduction of pest management concepts and their gradual adoption since the pilot pest management project in 1972, insecticide usage has been reduced significantly, and yields have increased. Similar benefits have been derived from other pest management programs throughout the Cotton Belt (Smith 1969, Carruth and Moore 1973, Barnes et al. 1974, Black and Burton 1974).
The experience of such successful pest management programs and support from advisory committees and others helped in planning and implementing the OPMT.
Cotton in the OPMT area is infested by the same insect pests that are present in other hill areas of the State. Populations of boll weevils (Anthonomus grandis Boheman) often reach treatment level during early season and will seriously damage fields that do not receive proper management. The bollworm (Heliothis zea (Boddie)) usually requires more frequent midseason and late-season insecticide applications than other pests. This is especially true in fields where it has been necessary to make repeated applications for early-season boll weevils. Control measures for plant bugs (Lygus spp.) may be required in some fields during early season. Whiteflies (Aleyrodidae), spider mites (Tetranychus spp.), and aphids (Aphis spp.) occasionally reach damaging levels and must be controlled.

^^^ J. L HAMERETAL
OBJECTIVES
The objectives for the OPMT were;
(1) To develop and demonstrate the technological and opera- tional capability to implement a community-wide optimum cotton in- sect management program to maintain boll weevils and other insects below treatment levels through voluntary participation of producers.
(2) To make available to evaluation teams biological, envi- ronmental, and program cost data collected in the operation of the Trial as part of the information necessary in estimating costs and benefits of a beltwide cotton insect management program.
GOALS
To achieve the objectives for the OPMT, several goals were es- tablished. These were:
(1) Maintenance of boll weevils and other cotton pests below treatment levels as recommended by the Mississippi Cotton Insect Control Guide.
(2) Higher net returns for participating growers than for nonparticipating growers.
(3) Ninety percent grower participation in the demonstration area at the end of the first year, 95% by the end of the second year, and 100% by the third year.
(4) Complete cooperation of all agencies and personnel in- volved in the Trial.
(5) Implementation of other pest management practices by par- ticipants in the Trial. Agronomic and other pest control practices would include those currently recommended by the Mississippi Coop- erative Extension Service (MCES) to increase cotton profits.
PERSONNEL
Operations
During early planning for the OPMT, personnel required for ac- tual operations were projected on the basis of land area, potential acreage to be included, number of participating clientele expected, and resource personnel located in the program area. The minimum necessary was determined to include an operations chief and five unit employees. The unit employees were used in operating traps collecting field data, and scouting (after summer scouts returned

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 389
to school); they also had other duties as assigned by the operations chief. The operations chief assisted in supervision of employees, helped analyze data, and worked closely with aerial applicators, consultants, producers, chemical suppliers, and Extension person- nel. He had primary responsibility for maintenance of an effec- tive, well-coordinated program. An Area Pest Management Specialist was in charge of the Trial and worked closely with the county Ex- tension agent, associate agent, and other Extension personnel in conducting the program. He also worked closely with all clientele in Panola County to insure operational success. Backup support was provided by the Extension cotton entomologist and pest management specialist at Mississippi State University. Personnel relation- ships are illustrated in Figure 1.
Advisory
All aspects of the OPMT were under the supervision of support- ive advisory committees. Both State and regional advisory person- nel participated in scheduled meetings during the 3-year Trial pe- riod. The State Pest Management Advisory Committee, established in 1972, functioned in an advisory capacity by assisting in Trial planning and guidance. The Regional Extension Educational Advisory Committee consisted of one representative from each of the 11 boll weevil-infested States. Twice each year this committee reviewed the progress of the Trial and offered suggestions for improvements. The State Working Group included representatives from Extension, research and regulatory groups, private consultants, and the chemi- cal industry. This group attended a meeting held monthly in the Trial area. Public attendance was also encouraged.
PUBLIC AWARENESS AND EDUCATION
The collection of baseline data in 1977 by members of the sev- eral teams that would be evaluating the OPMT alerted many producers to the fact that the Trial would probably begin in Panola County in 1978. In addition, during the fall of 1977 and throughout 1978, educational efforts were undertaken by 0PM personnel. County meet- ings with producers, programs for civic clubs and ginning groups, and informal meetings with producers were means of communication used in informing growers and the public of the OPMT program. News releases describing the program were submitted to regional and na- tional magazines and local newspapers.
The operations chief, a person knowledgeable in pest manage- ment, and the five unit employees, local individuals holding B.S. and M.S. degrees, were hired in the spring of 1978. These people carried out the operational components of 0PM and gained the confi- dence of the farm community. In addition, 0PM personnel met in- formally with producers, consultants, aerial applicators, and others and discussed the potential role of the OPMT as it related to their particular interests. These informal communications

390 J. L HAMERETAL
Regional Extension
Educational Advisory
Committee
Dr. David F. Young, Jr. Leader, Extension
Entomology
Dr. Jim Hamer Pest Management
Specialist
State Pest Managernent
Advisory Committee Other State
Cooperatives
Dr. Gordon Andrews Area Pest Management Specialist
Dr. H. C. Mitchell (Deceased)
Dr. Robert Head Extension Entomologist
Joe Hensley County Agent Panola County
Government Cooperators (ASCS, MDA-DPI, Evaluation Teams)
Producers Aerial Applicators
Chemical Suppliers
Figure 1.—Optimum Pest Management Trial Cooperators: ASCS = Agri- cultural Stabilization and Conservation Service. MDA-DPI = Mis- sissippi Department of Agriculture and Commerce, Division of Plant Industry.

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 391
assisted in solving specific problems with producers. Also one-on-
one meetings were frequently used to inform growers and other in- terested individuals of insect problems throughout each growing season and to help maintain effective interactions between program personnel and growers. Interest increased as producers observed
reduced costs and excellent yields on their own farms.
Scout training received emphasis throughout the 3-year Trial. At least three scout training workshops were conducted for scouts employed by the county Extension personnel or by private consult- ants. Training sessions were held at night so farmers could at- tend. Follow-up scout training continued throughout the season as Extension personnel visited with scouts and consultants to work out recommendations for special situations that arose.
Because of the educational activities undertaken by the 0PM staff and the county Extension personnel in Panola County, the lev- els of participation by growers in the OPMT were high. In 1978, 1979, and 1980, participating growers accounted for 98.7, 99.6, and
99.7%, respectively, of all cotton acreage.
CONTRACTUAL AGREEMENTS
Funds allocated for the OPMT covered chemical and application costs for up to one pinhead square application (if needed) and four diapause control applications. Contractual agreements for disburs- ing these funds were necessary to improve efficiency in payment and
record keeping, to insure effective use of chemicals, and to make all parties legally bound to a contract. The MCES made contracts with aerial applicators and producers, and the contracts were used to inform the operations chief of the aerial applicator selected by
each producer.
At the beginning of each season, bids were solicited for the cost of the chemicals and the applications (on a per acre basis). Bids were then averaged, and the average cost became the fixed price for the season for all cooperators in the county. Pajmients for chemicals were made directly to the producer. Payments for ap- plications were made to the aerial applicator chosen by the pro- ducer. This procedure did not disrupt the regular channel of trade of the producer and proved to be a sound approach for a voluntary
program.
MAPPING
Mapping of cotton acreage is a prerequisite to an effective scouting program. However, locating and mapping approximately 39,000 acres of cotton in an area as large as Panola County re- quired considerable effort and so could not be done entirely by 0PM personnel. Private consultants and county Extension personnel

392 J. L HAMERETAL
cooperated by specifying the acreage they were scouting. Only the acreage scouted by producers was actually mapped by OPMT personnel.
Sixteen orthophotoquad maps were used to map the acreage in the county. With these maps, any point in the county could be lo- cated with longitude-latitude coordinates. 0PM personnel were thus able to pinpoint fields and traps and to assign them unique coordi- nates for identification. In addition, computer storage of data and retrieval of data in map form (Figure 2) made it possible to survey the entire county each week to determine trap captures.
The accurate data from precise locations produced from the mapping effort were essential to decision making in the program. They should also be analyzed carefully before other larger programs are initiated. Weevil populations in the county can be precisely located, and such information can be invaluable in determining how much isolation is needed for a diapause program.
Components used during the 3-year period of the 0PM Trial were as follows:
(l) Trapping
(A) Pheromone traps
(1) Modified Mitchell trap (about 1500 peripheral field traps monitored weekly from April 15 to July 15 and September 11 to December 1.
(2) Leggett traps at different distances from the county lines (72 monitored).
(3) Cone Heliothis spp. traps.
(B) Blacklight traps (three traps used to detect Heliothis spp. activity during the soason).
(II) Planting dates (producers encouraged to plant in as short an interval as possible once planting initiated).
(Ill) Pinhead square application (paid from 0PM funds, if needed).
(IV) Scouting.
(V) In-season control of boll weevils, bollworms, plant bugs, spider mites, and other pest complexes based on scouting reports (costs of chemical and application borne by producers).

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 393
fftt^-fttt^ ^ '^ ^V '^^l ^^'^'^l -f f- -ft^f^ 1^2^'^^^'^''^^ ^ ^ 1 ^ -f i^ttttti^t't i^ \r t i^ t t l i^
-^-^ ^ T t r t t i\i^ t t t t t
t^-rt] rr-firi'-f'f \ -f -f r l^'^ t t t ^\2
^ * 1- 1 1 1- ^ 1 ^ ^/i 1 ^ 1
l'^llll'^'^lll"^'^!!''^'^'^'^'^^ 1 ^1 /l ^ ^ ^ 2
1 2•^ / 1214^ -^3 11 + 1-
- 2212^M ^M -f + + 1 'f',_-..-...^ ^1^1^ 2 1 * /--i _^...^ ^-2"
\4 3 1 ...1 i-^-^--^~iJr 1 1 -2^23 \-"2 2332^^ r 1^ t t -r
\22233 2 1^ 1^ 1^ 1- 1 X 1 1 2 2 '^ 'i' ^ 31 34411 ^ 2 52 325M 1 M ^ 1 M 1 1 3 \ M M 1 1 1 1 1 1 \ M12^^2^^^2 \ ^ i~±-i—■ '
Figure 2.—A 1979 computerized map of Panola County showing average captures in pheromone traps during the week (5/29/78-6/4/78) of peak spring emergence of boll weevils.

394 j. L HAMERETAL
(Vl) Diapause control of boll weevils in late summer and fall with insecticide (three or four treatments). Diapause con- trol treatments applied at no expense to producers.
(VII) Stalk destruction.
TECHNOLOGY USED IN OPMT
Trapping
As noted, Mitchell and Leggett pheromone traps were used in the 0PM program to monitor weevil populations. The modified Mit- chell pheromone traps (McKibben et al. 1971, Mitchell and Hardee 1974, Frisbie 1976, Mitchell et al. 1976, Dickerson et al. 1981) were distributed at a ratio of one trap per 20 acres of cotton in Panola County. In order to compare the size of boll weevil popula- tions in the OPMT and the CIC areas, the research team collecting biological evaluation data used the same trapping density in sam- pling fields in Pontotoc County. Each spring, these field-oriented pheromone traps were placed on the periphery of the fields that had been planted in cotton the preceding year and were used to deter- mine the need for a pinhead square application of insecticide for the control of overwintered boll weevils. The detection of five weevils per week in May was a signal to alert consultants and pro- ducers. Row-foot counts were then undertaken in fields that were threatened. The treatment threshold recommended was 25 weevils per acre.
In addition, a line of traps radiating from 8 miles within the county to 2 miles outside the county in all compass directions was operated at ixentical sites throughout the 3 years of the 0PM pro- gram to monitor movement of weevils into the Trial area. These trap lines consisted of three Leggett traps at trapping stations that were positioned at 2-mile intervals (Leggett and Cross 1971, Leggett et al. 1975, Leggett 1979). Since there were no buffer zones in this program, trap lines were used to evaluate suppres- sion of normally managed weevil populations at various distances outside the county.
The three blacklight traps and the cone pheromone traps (Roach 1975, Hollingsworth et al. 1978, Hartstack et al. 1979) were used to monitor populations of adult bollworms and to alert producers, consultants, and scouts of possible bollworm activity.
The pheromone trapping program of the OPMT was the largest pheromone trapping program for boll weevils ever conducted. It provided 0PM personnel with current information on the selective density of populations throughout the county. Figure 3 shows the

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 395
Average weevils/trap/week
5
Ponotoc '80
Panola '80
Ponotoc '79
Panola '79
May June
Figure 3.---Captures of boll weevils in peripheral traps, Optimum Pest Management Trial, spring 1979-80.

396 J. L HAMER ETAL
trend of catches in the peripheral traps in Panola and Pontotoc Counties during the spring emergence of weevils from ground trash. In both areas, the severe winter of 1977 had reduced boll weevil populations to a low level. Then in the fall of 1978 and 1979, insecticides for diapause control were applied to participating acreage in Panola County. Figure 3 shows the difference between boll weevil populations in the two counties as the 0PM program progressed. In the spring of 1979 and 1980, weevils were 78 and 94% lower, respectively, in Panola County than in Pontotoc County.
The gradients of weevil captures in line traps during the spring trappings of 1978, 1979, and 1980 are shown in Figure 4.
Average weevils/trap
1.575
.175
.025 1 \ I \ r 2 0 2 4 6
Outside Panola County I Inside Panola County
Station positions (miles)
Figure 4.—Captures of boll weevils in line traps during spring emergence. Optimum Pest Management Trial, 1978-80.

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 397
Fewer weevils were taken in traps outside the county than in traps inside Panola County in 1978, the first year of the program. How- ever, the numbers taken outside increased each spring thereafter. Meanwhile, traps 4 miles or more inside Panola County captured fewer weevils in 1979 and 1980 after fall diapause control treatments.
Planting Dates
Uniform planting dates were part of the effort to improve in- sect pest management in Panola County during the OPMT since fields of cotton that are at two stages of growth frequently offer an in- sect population a chance to develop in one field and then move to the later-planted field, which now may be in a more attractive stage. Moreover, in Panola County, producers must plant as soon as soil temperatures are suitable so full benefit can be received from the short growing season. However, weather, equipment, and avail- able labor determine how quickly the cotton crop can be planted.
In all 3 years of the OPMT, cotton was planted by cooperating growers as soon as soil temperatures would allow.
Pinhead Square Application
The procedures used in the OPMT were designed to prevent dam- aging infestations of boll weevils from occurring before the onset of large Heliothis spp. populations made it necessary to apply in- secticides. Applications early in the season destroy beneficial insects, thus releasing other insect pests, principally bollworms, which then must be controlled with insecticides. Therefore, a pin- head square application of insecticide was used only when popula- tions of overwintering weevils were large enough to produce a dam- aging first generation of boll weevils. Also, the application was timed to kill overwintering weevils before egg laying and at a time when it would not disrupt the development of the beneficial insect populations needed to suppress bollworms in late July and August.
During the 3 years of the 0PM program, more than 105,000 acres of cotton were planted in Panola County, and it was only necessary to use the pinhead square application on ca. 57 acres of cotton (in 1980). This cotton was near other cotton that was planted at the county border and so had not received diapause control treatments in 1979.
Scouting
Scouting is an important and difficult component of any cotton pest management program, but necessary control decisions can only be made if fields are thoroughly scouted. The most useful data are obtained by scouts who are highly motivated, well trained.

^^Ö J. L. HAMERETAL
experienced, and observant. When such scouts are available, data collected from each day of scouting can be used with confidence though previously collected data, the stage of the crop, current growing conditions, past history of the crops, potential yields, and previous crop input must also be considered in making an accurate control decision.
The only requirement for participation in the OPMT was that the producer have his cotton scouted and that a weekly scouting re- port be filed with the operations chief throughout the season. The Extension agricultural agent for Panola County encouraged growers of large acreages to hire private consultants. The Panola County Pest Management Society worked with producers who had not previ- ously scouted their acreage. There was some concern at the begin- ning of the program that scouting done by such diverse groups would result in a diversity of recommendations. However, formal and in- formal communications between the Trial personnel and other scouts resulted in early identification of localized insect problems. Careful consideration of the problems by all groups then resulted in general agreement concerning recommendations to be made to growers.
During the 3 years of the OPMT, the percentage of the acreage scouted remained fairly constant. Approximately 75, 15, and 10% were scouted by private consultants, producers, and the Panola County Pest Management Society, respectively. There were 1250, 1339, and 1469 scouted fields in 1978, 1979, and 1980, respec- tively. Although the number of acres scouted by the Panola County Extension Pest Management Society was relatively low, this service was provided at actual cost to producers with small farms, a deci- sion that improved overall management of cotton insects in the Trial area significantly. Figures 5 and 6 show the percentage of squares damaged by boll weevils and bollworms, respectively, as re- ported by the 0PM scouts. Plainly the populations of both pests fluctuated from year to year and were, on the average, below threshold levels.
In-Season Control
Natural factors such as disease and beneficial insects (Dink- ins et al. 1970) are important in management of insect pests. They are especially effective in suppressing the first and second gener- ations of bollworms in cotton when disruptive insecticides are not applied. As noted, if one is to avoid early use of insecticide, boll weevils must be managed by using pinhead square applications or by diapause control. When weevil infestations can be held below the economic threshold, fewer applications of insecticide will be needed to control bollworms though there may still be a need for occasional control of infestations of plant bugs, spider mites, whiteflies, or aphids before onset of heavy bollworm pressure. However, the damage caused by these pests usually does not affect as much acreage as the damage done by boll weevils and bollworms.

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 399
Percent
20 1978 1979 1980
15-
6-11 6-25 7-09 7-23 8-06 Week
8-20 9-03 9-17
Figure 5.—Percentage of squares damaged by boll weevils reported by Optimum Pest Management Trial scouts, 1978-80.
Percent
20 1978- 1979- 1980
15-
10-
Figure 6.—Percentage of squares damaged by bollworms reported by Optimum Pest Management Trial scouts, 1978-80.

400 J. L HAMERETAL
In 1978, weevil populations were maintained below economic threshold during the season so insecticides did not have to be ap- plied to control bollworms until the first of August or later. Only a few fields required treatment for plant bugs. Then four fall diapause control applications were made to most of the cotton acreage to lower the weevil population before overwintering.
In the spring of 1979, weevil emergence was low, and popula- tions were insufficient to justify a pinhead square application. Then during the season, extremely wet conditions prevailed so the cotton had difficulty in fruiting. Also, wild hosts remained at- tractive to plant bugs longer so relatively large populations were ready to move into cotton when it was fruiting. As a result, plant bugs required control in most fields in 1979, but treatments were usually not applied until after the first generation of boll- worms was reduced by beneficial arthropods. Also, the bollworm population did not reach a damaging level until late in the growing season and thus caused few problems. Boll weevils remained below treatment level during the season in almost all fields. Three dia- pause applications and a partial fourth application were made to all acreage in the Trial area. (An early freeze was the main fac- tor in the decision to cease the fourth application to some acreage.)
In 1980, populations of boll weevils increased greatly over much of the hill acreage outside the Trial area in Mississippi. This resurgence made it necessary to make more frequent in-season applications to control this pest in the surrounding counties and also resulted in larger numbers of boll weevils near the Panola County line. It was for this reason that the pinhead square appli- cation had to be used on 57 acres of cotton near the border of the Trial area. Nevertheless, in 1980, infestations of boll weevils in the Trial area were maintained below the economic threshold and did not require treatment during most of the season. A small percent- age of the acreage was infested by spider mites and required treat- ment. The plant bug was of little significance as a cotton insect pest in 1980.
Producers in the OPMT therefore benefited from reduced weevil populations because they had to make fewer applications of insecti- cide. For example, in 1978, the average number of foliar applica- tions was 3.3 on fields selected for evaluation, and total cost of insect control, including scouting, was about $20.47 per acre. In 1979, the average number of applications was 3.4, and the cost was $19.36 per acre. In 1980, the average was 3.0 applications, and the cost was $17.50 per acre. In addition, yields in the Trial area were higher than yields in all surrounding counties where more or the same number of insecticide applications were applied in 1980 (Figure 7). In the 3 years of the OPMT area, yield of lint cotton increased by an average 84 pounds per acre per year, which equalled an average increase of 34 pounds of lint per acre compared with the 10-year average yield (Tables 1 and 2).

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 401
Figure 7.—Estimated yields (x), insecticide applications (y), and cotton acreage (z) for representative counties in north central Mississippi in 1980. Estimates based on survey of county agents, fall 1980. Data based on Current Insect Control programs, which included organized pest management programs.
Diapause Control
As noted, as many as four diapause control treatments were provided at no cost to the producer and with little interference to his normal routine. The producer could choose his aerial applica- tor and could purchase chemicals from the dealer he normally used.
All diapause applications were applied aerially to insure ad- herence to the schedule. The first application was scheduled 10 days after the producer had ended in-season control. The second followed the first by 10 days. The third and fourth were applied 15 days after the preceding application. (Most producers included a defoliant in one of the applications.) Methyl parathion was se- lected for diapause control and was used at a rate of 0.5 pound of active ingredient per acre. In 1978, all applications were made at a rate of 1 gallon total volume per acre, but the rate was in- creased to 2 gallons per acre in 1979 and 1980 to achieve better plant coverage. The schedule for the first two applications was adhered to closely. Only routine weather problems such as rain and

402 J. L HAMERETAL
Table 1.—Lint yields and averages for 3 years of Optimum Pest Man- agement Trial y V. 10-year average 2/
Year
1978 1979 1980 Three-year average Ten-year average Difference between
10- and 3-year averages
Lint yie Ids (pounds per acre :)
Panola Pontotoc County County
630 533 582 485 582 436 598 485 514 422
84 63
]_l Yield estimates obtained from Biological Evaluation Team of OPMT. A value of 485 pounds lint per bale was used as a conversion factor.
2./ Ten-year average yield data taken from Mississippi Crop and Livestock Reporting Service reports.
wind caused short delays. Later in the season during the period when the last two applications were scheduled, temperature became a critical factor and caused some delays. For example, if predic- tions did not indicate that the temperature would reach 70**?, ap- plications were delayed. All diapause applications were halted after a killing frost or if the stalks were destroyed in a field.
Table 2. -Lint yields and averages for 3 years of Optimum Pest Man- agement Trial and 10-year averages 1/
Year
1978 1979 1980 Three-year average Ten-year average Difference between
10- and 3-year averages
Lint yie Ids (pounds per acre i)
Panola Pontotoc County County
557 500 602 411 485 348 548 420 514 422
+34
J_/ Data taken from Mississippi Crop and Livestock Reporting Ser- vice reports.

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 403
Areawide diapause control treatments were applied for the first time in the Trial area in 1978. Since the crop had matured early, the initial treatment was applied beginning September 10. Because of the warm fall and this early starting date, all four diapause applications were needed except in a few fields where the stalks were destroyed. In 1979, the late maturing crop and bad weather put off the first diapause application to September 20. That year an early freeze stopped applications in the middle of the fourth application. In 1980, diapause applications were started September 10, and all four applications were made except on 4034 acres where stalks had been destroyed before the completion of the treatments. The catch of overwintered weevils in pheromone traps reported previously shows the effectiveness of the diapause control applications.
Stalk Destruction
Stalk destruction is a method of eliminating the food that weevils must have to attain firm diapause, but those planning fu- ture management programs should take into account the time avail- able for destruction after the crop of cotton is produced and the harvest is completed. In 1978, cotton matured early in the OPMT area, and harvesting was in progress by the middle of September. As a result, about 80% of the stalks were destroyed before the first killing frost. In 1979, harvest did not begin until late September, and the early frost that occurred during harvest made stalk destruction impractical. In 1980, about 70% of the stalks had been destroyed by the time of the first killing frost.
Stalk destruction in the fall was also affected adversely by two problems: (1) rented land was leased on a year-to-year basis, and (2) producers were not sure how much cotton they would plant the next season because of fluctuating prices.
Stalk destruction must take place before the first killing frost, but during most years, producers in Panola County are still harvesting when the frost occurs. The technique is therefore of little value in many areas of the Cotton Belt.
TRIAL SUPPORT
Regulatory
The Mississippi Department of Agriculture and Commerce, Divi- sion of Plant Industry (MDAC-DPI), a cooperating agency, is charged with State regulatory activities. No regulatory functions were in- volved in the OPMT, but this agency provided support by insuring "up-to-standard" pesticides and usage in accordance with labeling. Other responsibilities included periodic checks of aerial applica- tion equipment with emphasis on drift, dosage, and application

404 J. L HAMERETAL
procedures. The MDAC-DPI issues all new and renewed licenses for aerial applicators.
This agency provided much assistance during the 3 years of the Trial and should continue to be included in future programs.
Research
Special projects and recurring State pest management programs are strengthened by a productive relationship between research and Extension entomologists. This relationship has been effective in Mississippi throughout the years and was equally effective during the OPMT.' The MCES sought the advice of and was fully supported by research personnel.
Various agencies and groups that were associated with the Trial and their activities were as follows:
Boll Weevil Research Laboratory
ARS, USDA Starkville, MS
Bioenvironmental Laboratory
ARS, USDA Starkville, MS
Mississippi Agricultural and Forestry Experiment Station
Mississippi State University
Computer Center Mississippi State
University
Animal and Plant Health Inspection Service, USDA
(1) Overall and biological evaluation
(2) Weevil trapping
(3) Coordination of data re- trieval and storage
(4) Research (a) Host plant resistance (b) Weevil trapping (c) Cotton insect ecology
(1) Biological evaluation
(2) Research (a) Bollworm pheromone trapping (b) Cotton insect ecology
(1) Economic evaluation
(2) Research - cotton insect ecology
(1) Data storage
(2) Data summarization
(3) Modeling
(1) Environmental monitoring and evaluation

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 405
Mississippi Cooperative (1) Program education Extension Service
Panola County, MS (2) Cotton scouting
(3) Advice and support
(4) Chemical demonstrations
EVALUATION
In 1979, a questionnaire was mailed by the operations group of the OPMT to cotton producers who had participated in the Trial. The intent was to help operations personnel evaluate their efforts and to obtain information pertinent to the overall evaluation of the trial. Significant statements derived from analysis of the re- turned questionnaires were:
(1) Producers shifted from no routine scouting to some form of regular scouting.
(2) Private consultants were used by a higher percentage of producers, and the percentage of fields scouted has increased by 25% since 1975.
(3) Producers decreased the total number of insecticide ap- plications and the number directed specifically against boll weevils.
(4) Of the producers responding, 91% indicated they will con- tinue the program after public funding is withdrawn. Producers responding "No," indicated that the reason was the location of their farms; that is, they were near a county where infestations were not to be controlled or they farmed acreage in two counties.
Other methods of evaluation of the program included summaries of weekly infestation records from each field. This information was compiled annually and in a final report at the end of the 3- year trial period. These reports will be available to evaluation teams and to others interested in the OPMT.

406 J. L HAMERETAL
LITERATURE CITED
Barnes, G., W. P. Boyer, J. J. Kimbrough, H. R. Sterling, and M. L. Wall.
1974. Cotton pest management program. Arkansas Agricultural Ex- tension Service, Leaflet No. 52 (Revised), 12 p.
Black, J. H., and V. E. Burton. 1974. Pest management, what it really is and how it is working
in California cotton. Agrichemical Age, September, p. 6-9.
Carruth, L. A., and L. Moore. 1973. Cotton scouting and pesticide use in eastern Arizona. Journal of Economic Entomology 66:187-190.
Collins, H. L., R. J. Moody, C. F. Sartor, D. F. Young, and J. L. Hamer.
1976. Results of a three-year cotton pest management program in the hill section of Mississippi. U.S. Department of Agricul- ture, Animal and Plant Health Inspection Service, Plant Protec- tion and Quarantine Programs, p. 81-86.
Dickerson, W. A., G. H. McKibben, E. P. Lloyd, J. F. Kearney, J. J. Lam, Jr., and W. H. Cross.
1981. Field evaluations of a modified in-field boll weevil trap. Journal of Economic Entomology 74:280-282.
Dinkins, R. L., J. R. Brazzel, and C. A. Wilson. 1970. Seasonal incidence of major predaceous arthropods in Mis-
sissippi cotton fields. Journal of Economic Entomology 63:814- 817.
Frisbie, R. E. 1976. Role of pheromone traps in survey, detection, program
evaluation, and related biological studies. Texas Agricultural Experiment Station, Research Monograph No. 8, p. 32-36.
Hartstack, A. W., J. A. Witz, and D. R. Buck. 1979. Moth traps for the tobacco budworm. Journal of Economic Entomology 72:519-522.
Hollingsworth, J. P., A. W. Hartstack, D. R. Buck, and D. E. Hen- dricks.
1978. Electric and non-electric moth trap baited with synthetic sex pheromone of the tobacco budworm. U.S. Department of Agri- culture, ARS-S-173, 13 p.
Leggett, J. E. 1979. Boll weevil: some new concepts in trap design and evalua-
tion of trap efficiency. Environmental Entomology 8:70-72.

OPTIMUM PEST MANAGEMENT TRIAL IN MISSISSIPPI 407
Leggett, J. E., and W. H. Cross. 1971. A new trap for capturing boll weevils. U.S. Department of Agriculture, Cooperative Economic Insect Report No. 21:773-774.
W. H. Cross, and H. C. Mitchell. 1975. Improved traps for capturing boll weevils. Journal of the Georgia Entomological Society 10:52-61.
McKibben, G. H., D. D. Hardee, and T. B. Davich. 1971. Slow-release formulation of grandlure, the synthetic pher-
omone of the boll weevil. Journal of Economic Entomology 64: 317-319.
Mitchell, E. B., and D. D. Hardee. 1974. In-field traps: a new concept in survey and suppression
of low populations of boll weevils. Journal of Economic Ento- mology 67:506-508.
E. P. Lloyd, D. D. Hardee, W. H. Cross, and T. B. Davich.
1976. In-field traps and insecticides for suppression and elimi- nation of populations of boll weevils. Journal of Economic En- tomology 69:83-88.
Moody, R. J., H. Collins, J. Hamer, C. F. Sartor, and D. F. Young. 1973. Mississippi cotton pest management first annual report, Mississippi Cooperative Extension Service, Mississippi State.
Roach, S. H. 1975. Heliothis zea and JH. virescens: moth activity as measured by blacklight and pheromone traps. Journal of Economic Ento- mology 68:17-21.
Smith, R. F. 1969. The importance of economic injury levels in the develop- ment of integrated pest control programs. Qualitas Plantarum et Materiae Vegetabiles 17:81-92.


409
Chapter 16
ANALYSIS OF TECHNOLOGY AVAILABLE FOR ERADICATION OF THE BOLL WEEVIL
E. F. Knipling Agricultural Research Service U.S. Department of Agriculture Beltsville, MD 20705
ABSTRACT The technology for eradication of the boll weevil (Anthonomus grandis Boheman) has been demonstrated. It has focused on the highly vulnerable overwintering generation. Properly scheduled insecticide applications after the regular growing season reduced popu- lations by next spring to an average of a few per acre. Grandlure traps along field borders further suppressed survivors and also identi- fied fields where boll weevils were most preva- lent. Diflubenzuron was applied, if necessary, where field populations seemed higher than can be effectively suppressed by sterile boll wee- vil releases. The sterile boll weevils at high sterility-to-fertile ratios are counted on to inhibit reproduction by most of the low sur- viving population. However, infield traps at the rate of one or two per acre will assure early detection of any infestations that may occur. Infield and border field traps have such high technical capability that possible reintro- duction can be discovered before infestations increase to high levels and become widespread.
INTRODUCTION
The technology employed in the Boll Weevil Eradication Trial (BWET) conducted in North Carolina during 1978-80 involved the use of chemical insecticides and defoliants, pheromone of the boll wee- vil (Anthonomus grandis Boheman), and the release of sterile boll weevils. Limited use was also made of the growth regulating chemi- cal, diflubenzuron (Dimilin). The general plan was to suppress the boll weevil population during Year 1 to a very low level by the use of insecticides and cultural measures and then eliminate the re- duced population during Year 2 by the use of at tractants, sterile boll weevil releases, and limited use of chemical insecticides.

410 E. F. KNIPLING
In terms of numbers of pests destroyed, chemical insecticides have their greatest efficiency when the pest population is high; the attractant and the sterile boll weevil release components have their greatest efficiency when the population is low. Therefore, the two control procedures are highly complementary. The suppres- sive pressure was directed against the overwintering generation, the most vulnerable period during the annual cycle of the boll weevil.
The basic technology, then still in the early stages of de- velopment, was first tested in the Pilot Boll Weevil Eradication Experiment (PBWEE), which took place in southern Mississippi during 1971-73. The results of the PBWEE were evaluated in reports of the Technical Guidance Committee (Knipling 1976b) and the Entomological Society of America (ESA) Review Committee on the Pilot Boll Weevil Eradication Experiment (Eden 1976).
The Technical Guidance Committee for the PBWEE concluded that it was technically and operationally feasible to eliminate the boll weevil as an economic pest in the United States by the use of tech- niques that are ecologically acceptable. This conclusion seemed justified in view of the extremely high boll weevil population in the experimental area and the difficult circumstances under which the experiment was conducted. Even though the techniques used were still under development, the experiment demonstrated that by meth- ods then available a population could be suppressed below the level of detection in all 170 cotton fields that were 25 miles or farther from heavily infested cotton. However, the Committee also recog- nized the need for improvements in the techniques and identified the following research needs:
(1) Improved mass rearing procedures to assure the capability of producing adequate numbers of high-quality boll weevils for sterilization and release.
(2) Improved sterilization techniques to assure the attainment of maximum (and consistently high) levels of sterilization with minimum detrimental effect on the vigor and mating competitiveness of the males.
(3) New methods of sterilizing both sexes of the boll weevil so as to obviate the cost of separating sexes and reduce costs and logistic problems associated with feeding boll weevils for 6 days before release.
(4) Continued investigation of grandlure (boll weevil phero- mone) to develop the most effective and least costly method of using the attractant for: (a) suppression, (b) detection and popu- lation assessment, and (c) monitoring progress in population sup- pression.
The ESA Review Committee for the PBWEE did not agree on wheth- er technical feasibility of eradication of boll weevil populations

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 411
was demonstrated. It recognized the same research needs specified by the Technical Guidance Committee. It concluded that an attempt to eradicate the boll weevil should be a sociopolitical decision. It recommended that all members of the ESA inform themselves as to the long-range environmental and economic benefits that would re- sult from a successful eradication program and weigh the benefits against the costs involved.
Following completion of the PBWEE and before initiation of the BWET in North Carolina, major technical advancements were made which significantly improved the probability of success of the BWET while reducing costs as well. The major improvements, all dis- cussed in this handbook, are:
(1) Improved rearing procedures (Griffin et al. Chapter 11) resulted in production of large numbers of quality boll weevils for sterilization.
(2) Improved procedures for sterilizing the boll weevil (Wright and Villavaso Chapter 7) resulted in a highly sterile wee- vil which is fairly competitive.
(3) The insect growth regulator, diflubenzuron, was developed as an additional suppressive component. When applied as a foliar spray, the chemical prevents hatch of eggs deposited by females while not interfering with release of sterile weevils (Bull et al. Chapter 9).
(4) A highly sensitive detection system that consisted of as few as one infield trap per acre was developed. It has a 94% prob- ability of detecting F, progeny and a 100% probability of detecting F- progeny of a single female weevil (Lloyd et al. Chapter 8).
No other eradication or management procedure for an insect has ever been put to such elaborate and demanding tests as a prelude to possible wide-scale practical application. However, no other pre- viously proposed eradication program has involved the same tech- niques and basic principles of suppression that were applied in the two trial programs.
The purpose of this chapter of the handbook is to describe and analyze in detail the nature and function of the methods of suppression employed and to point out what the several suppressive measures can contribute individually and collectively to eradica- tion, in the event a national effort to eradicate the boll weevil is undertaken. While the techniques and principles are applicable to eradication, they could be equally relevant to effective and ecologically acceptable boll weevil population management programs.
The magnitude of the boll weevil problem has caused Federal, State, and private institutions to spend millions of dollars in ef- forts to develop effective and acceptable solutions to the problem.

412 E. F. KNIPLING
The boll weevil ranks high among the nation's most damaging pests and, without question, is the most objectionable from the stand- point of the adverse effect it has had on the quality of our agri- cultural environment, because of the necessity of using a variety of ecologically disruptive chemicals for boll weevil control year after year for almost a half century. This effect takes into ac- count the impact such use of insecticides has had on Heliothis spp. and other insect problems.
SOME FUNDAMENTAL PRINCIPLES OF INSECT POPULATION SUPPRESSION AS IT RELATES TO THE BOLL WEEVIL
The application of insecticides, together with the use of cul- tural measures in some areas, has been relied upon for boll weevil control since the development of arsenicals. Before chemicals be- came available, the fast-maturing cotton varieties were grown to obtain reasonable yields before overwintered populations increased to highly damaging numbers. From the mid-1940's to the present, a dozen or more highly effective insecticides came into use (Parencia et al. Chapter 10 in this handbook). These insecticides have served their purpose reasonably well when viewed from the stand- point of profitable cotton production. However, as used, insecti- cides do not provide a satisfactory solution to the boll weevil problem. Annual costs for control, inevitable losses due to grower neglect or miscalculations, unavoidable variations in boll weevil abundance due to weather and other causes, risks of insecticide re- sistance, and the ecological impact of broad-spectrum insecticides all contribute to a continuation of one of the nation's most costly and obnoxious insect pests. Indirectly, its presence makes it more difficult to manage Heliothis spp. and other pests on cotton or even in adjacent crop ecosystems because of the ecological disrup- tion resulting from broad-spectrum insecticides used for its control.
It has long been my contention that the boll weevil has been a costly pest year after year, not because of a lack of effective control measures but rather because of the way the available con- trol measures have been used. In 1958, I began studies involving the use of simulation boll weevil population models in an effort to gain a better understanding of the dynamics of the boll weevil in quantitative terms. Such information is necessary to explain why the pest continues to be so costly and destructive despite the availability and intensive use year after year of highly effective boll weevil insecticides by most growers in areas where the prob- lem is severe. The most important conclusion reached from these studies was that the boll weevil will continue to be a major pest indefinitely unless the total population is adequately suppressed in an organized and systematic manner. As noted (Knipling 1960), the dynamics of boll weevil populations are such that if only 10% of the farmers in a cotton community fail to control the pest, enough boll weevils can develop by generation 3 to infest all bolls and squares throughout the community even if 90% of the farmers

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 413
apply insecticides and obtain near 100% control on their cotton. In contrast, on the basis of the postulated dynamics of the pest, if all growers applied insecticides diligently and moderately and obtained 90% control each generation, there would be 100 times fewer boll weevils in the community by the end of one season and at the beginning of the next. More convincingly, if the insect is not controlled on 1% of the cotton in a community, 10 times more boll weevils will likely exist than in a similar community where all cotton is subjected to moderate and consistent boll weevil suppression.
This simple basic principle of insect population suppression in relation to the dynamics of the pest was largely ignored for many years by research workers, by growers, and by pest management advisers in developing systems of management for the pest. How- ever, it convinced me that an organized and fully coordinated ap- proach to the management of a key and dynamic pest like the boll weevil will be necessary to maintain populations at levels of lit- tle or no economic consequence. This fundamental principle holds regardless of the nature of the suppressive measures used.
Reliance on insecticides alone to achieve eradication or rigid management of the boll weevil could lead to ecological problems and possibly intensify insecticide resistance. Hence, supplemental suppression measures that avoid ecological disruption and reduce chances of resistance are necessary before one can advocate area- wide eradication or management practices. These goals have now been reached. Thus, on the basis of current technology, there are two alternatives to the boll weevil problem: (1) continue to rely on ecologically disruptive chemicals for an indefinite period of time, or (2) attack the pest on an areawide basis by making optimum use of alternative suppressive techniques.
USE OF SIMULATION MODELS AND INDIRECT CALCULATIONS AS AN AID TO EVALUATION OF BOLL WEEVIL SUPPRESSION MEASURES
In eradication, containment, or rigid pest population manage- ment programs, the pest numbers generally reach such low levels that detection of those remaining becomes a major problem. In the PBWEE in southern Mississippi, boll weevils could not be found in fields 25 miles or more from infested cotton with the detection methods then available. However, because of limitations in the sensitivity of the detection methods, complete absence of reproduc- tion could not be assured with a high degree of confidence. Simi- lar detection problems have been encountered in many other insect eradication or containment programs conducted in the past. The lower the density of the target pest the more difficult it becomes to establish presence or absence of a pest in specific areas or to make valid determinations of actual numbers of insects remaining. Consequently, research workers and pest managers are confronted with the problem of providing "scientific proof" of success to those who must endorse a new approach to a problem.

414 E. F. KNIPLING
The difficulty of obtaining reliable, quantitative data to support pest management procedures is a problem that must be given adequate attention and analysis by entomologists because it is relevant to the concepts of pest population eradication and con- tainment and, in some cases, may be equally relevant to rigid pest population management. However, it should be emphasized at the outset that absolute "proof" of eradication of any pest population is not scientifically possible. It is no more possible to prove complete absence than to prove that a chemical is completely free of hazards. This is also a matter of grave concern to toxicolo- gists concerned with the use of chemicals as pesticides.
The outstanding recent advances in research on insect sex pheromones have made possible great progress in the sensitivity of detection methods for a wide range of insect pests, but better in- terpretation of the data in quantitative terms is still needed. As progress continues, this should lead to less uncertainty concerning the validity of results obtained in eradication or pest management programs. This surely should be the case for the boll weevil. The advances that have been made in detection methods for this pest are reported in the section on the role of the sex and aggregating at- tractant as a suppressive measure (Lloyd et al. Chapter 8).
As noted, the conventional methods used to detect many insect pests have had limited reliability. This has made it difficult to determine in quantitative terms the degree of success of various suppressive measures under consideration. Accordingly, I have made extensive use of simulation population models based on assumed num- bers of insects present in an effort to determine the mechanism of action of various techniques of insect control and to estimate in numerical terms the results to expect from various methods of sup- pression. Such models have been used to estimate the impact on insect populations subjected to suppression by the use of insecti- cides, insect attractants, programmed release of parasites, growing resistant plant varieties, and the release of sterile or geneti- cally altered insects. They have also been used to show the great advantage of concurrent use of two or more compatible or complemen- tary methods of control. The various procedures used to make such a determination have been described in a recent publication (Knip- ling 1979).
Entomologists have not yet made extensive use of, and many do not have confidence in, indirect modeling procedures as an aid to the evaluation and interpretation of data obtained in suppression experiments or programs. They still prefer or expect direct quan- titative data that are virtually impossible or impractical to ob- tain after pest populations reach a certain minimum. In my view, simulation models are almost essential to fully appraise and ex- plain how and why different suppression techniques achieve certain results. In the final analysis, eradication, or even rigid manage- ment of pest populations, involves a game of numbers, which often defies accurate determination by conventional collection of data. Therefore, in discussing the various techniques of suppression that

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 415
may be used in boll weevil eradication programs and what they can contribute to suppression, I will make maximum use of simulation models that are based on reasonably well-established parameters.
INFLUENCE OF BOLL WEEVIL MOVEMENT ON ERADICATION
The movement of insects can account for the failure or appar- ent failure of many eradication experiments, even though the tech- niques used may have been capable of achieving eradication of iso- lated populations. It is axiomatic that eradication of a pest cannot be achieved unless the total population in the area of per- manent survival is subjected to suppression. Entomologists have consistently underestimated, or in some cases have not taken into account, the extent of insect movement. When the rate of infiltra- tion is below the level of detection and when insects still remain, scientists almost invariably attribute such presence of insects to a failure of the technology tested. The controversy over the suc- cess or lack of success of the PBWEE in southern Mississippi was in large measure due to the presence of some boll weevil reproduction in portions of the originally defined eradication area. Actually, infestations were detected only in cotton fields that were within 25 miles of heavily infested areas. The entire core area was assumed to be adequately isolated, but this subsequently proved not to be the case. In fact, the extent of movement of the boll weevil has been a major subject of investigation in connection with the different eradication trials. However, eradication, or even highly effective management of boll weevil populations, will never be ac- complished unless the areas under suppression are adequately iso- lated or are large enough to avoid direct infiltration of large numbers of the insects into substantial portions of the area under suppression. In the planning for the North Carolina trial program, movement of boll weevils 45 miles or more was considered a proba- bility because records of this extent of movement had been ob- tained; therefore, a barrier zone approximately 85 miles deep was established to prevent or minimize the possibility of infiltration into the eradication zone. Present movement studies now indicate some movement for up to 65 miles, so the possibility that some boll weevils will disperse a greater distance cannot be ruled out unless the population at the source is strongly suppressed. The introduc- tion of boll weevils as hitchhikers or on infested commodities was, of course, recognized.
CHARACTERISTICS AND FUNCTION OF VARIOUS SUPPRESSION COMPONENTS CONTRIBUTING TO ERADICATION
The techniques currently envisioned for the use in boll weevil eradication programs differ in suppressive action. They also vary in efficiency, depending on the density and distribution of the in- sects in the natural population. This makes it possible to achieve greater efficiency by employing two or more techniques concurrently or sequentially instead of relying on only one method. Indeed,

416 E. F. KNIPLING
techniques that are potentially effective and highly desirable from an ecological standpoint, such as the use of sex pheromones or genetically altered insects, may have relatively little practical value for eradication or management of well-established insects unless they are employed as supplements to insecticides, cultural and sanitary measures, or host plant resistance, methods of control with essentially the same efficiency at all levels of pest density. In order to clarify the practical significance and the potential value of different techniques of suppression when appropriately in- tegrated, I will discuss in detail the characteristics, action, and function of each of the several methods used for boll weevil sup- pression in the North Carolina trial program.
Insecticide Suppression Component
Insecticides are highly effective for the control of boll wee- vils when used at approved dosages on proper spray schedules. When eradication or a high degree of suppression is the objective, it is obvious that applications must be thorough. In the PBWEE, it was found during Year 1 that aerial applications alone did not produce a high degree of suppression because of natural and man-made obsta- cles such as tall trees, power lines, and others. However, supple- mental sprays applied with ground equipment resulted in a very high degree of suppression. It was also found that highly effective suppression with insecticides cannot be expected unless the behav- ior of the pest is taken into account and applications are appro- priately timed to disrupt boll weevil development.
Brazzel and Newsom (1959) determined that adult boll weevils enter a state of diapause in the late summer and early fall. In this state, they feed on squares and bolls for a period of 10-14 days or longer to build up fat reserves. They then leave the cot- ton fields for overwintering quarters. In view of this behavior, Brazzel (1961) and Brazzel et al. (1961) proposed the diapause spray schedule and demonstrated that four appropriately timed ap- plications of spray at about 10- to 14-day intervals in the fall would reduce overwintered populations by ca. 90%. This substantial control reduced the number of in-season applications required the following season. However, reproducing boll weevils are also present in the late summer and early fall. Therefore, spray inter- vals of 10 days or more permit considerable reproduction because the preoviposition period of the boll weevil may be as short as 5-6 days, even in the fall. Accordingly, on the basis of results of earlier simulation control models, I proposed modifications to the diapause spray schedule so as to limit reproduction by the last generation that produces most of the diapausing population (Knip- ling 1968). I also proposed that applications should be continued long enough to kill most of the surviving progeny before they leave the cotton fields for hibernating quarters. It was calculated that if four spray applications at intervals of 10-14 days to kill dia- pausing boll weevils will achieve about 90% suppression, seven applications properly scheduled to also limit reproduction of the

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 417
last reproducing generation will achieve higher than 99% suppres- sion. The general validity of the calculations was subsequently verified in boll weevil suppression programs undertaken by regula- tory agencies in the Presidio and High Plains areas in Texas (Adkisson et al. 1966, Fye et al. 1968).
Although most of the diapausing boll weevils are produced in the late summer and early fall, a certain percentage of the popula- tion may diapause and leave cotton fields for hibernation quarters before the time that a reproduction diapause spray schedule is initiated. This seemed to be the case with the high populations present during Year 1 of the PWBEE. (Many of the growers did not carry out a good in-season control program, and boll weevil popula- tions were very high.) Considerable numbers of boll weevils ap- parently entered hibernation when squares and bolls became scarce before the normal diapause period. Therefore, during Year 2, all cotton was properly monitored and sprayed during the growing season since a good in-season spray program protects the crop and also assures that few, if any, boll weevils will enter diapause and leave the field for overwintering sites during July and August. This in-season program was then followed by properly timed sprays to limit the rate of reproduction of the last reproducing genera- tion and to kill most of the greatly reduced population of dia- pausing progeny before it could complete development and leave the fields for hibernating quarters. The result was a drastic reduc- tion in the overwintered population.
The BWET conducted in North Carolina was carried out under close supervision by U.S. Department of Agriculture and cooperating State personnel. In the first year (1978), there was critical mon- itoring of cotton insect infestations, and in-season applications of insecticides were made for the control of Heliothis spp. and other pests, as well as for control of the boll weevil. However, the boll weevil population was abnormally low because the two pre- vious winters had been severe and because insecticides had been used intensively in 1977 for control of the bollworm (Heliothis zea (Boddie)); thus the 1978 in-season program included an average of 3.7 applications for bollworm control and 2.2 for boll weevil control. These treatments further reduced the boll weevil popula- tion. Nevertheless, the regular in-season program was supplemented by five sprays properly timed for maximum suppression of both re- producing and diapausing boll weevils. Defoliants were also ap- plied to limit available food for diapause development.
In the spring of 1979, only seven overwintered boll weevils were captured in 16,676 survey traps around North Carolina cotton fields totaling 12,485 acres of cotton. However, an accurate esti- mate of the total surviving overwintered population cannot be made because it is not known what proportion of the overwintered survi- vors was captured in the border field traps before the first re- leases of sterile boll weevils. After sterile boll weevil releases are made, much of the competitiveness of the traps is lost because of the competing pheromone produced by released males. However,

418 E. F. KNIPLING
it is probable that up to 50% of the overwintered boll weevils had emerged from hibernating quarters by the time the first sterile boll weevil releases were made. E. P. Lloyd (personal communica- tion) estimated that pheromone traps placed along field borders in the earlier PBWEE program captured 25% of the overwintered boll weevils.
If it is conservatively assumed that seven captured boll wee- vils equaled 25% of the total overwintered population in the eradi- cation area, the total number of overwintered survivors would have been of the order of 28 in an area having 15,222 acres of cotton during the 1979 season. While this estimate may not be highly accurate, it is accurate enough to allow the conclusion that vir- tual annihilation of the boll weevil population had been achieved by the insecticide applications and defoliant treatments during 1978.
It has long been my view that during the first year of a pro- gram designed to eradicate the boll weevil, the overwintered popu- lations should be reduced to an average level of one or two boll weevils per acre. I believe that such a low population can be readily eliminated the next season by the use of pheromone traps, sterile male releases, and strategic use of insecticides where needed. The very low capture rate in the spring of 1979 in North Carolina indicated that the overwintered population there had been reduced far below a level of one per acre. If the surviving over- wintered population was of the order of 28, as estimated, this would mean the survival of only one boll weevil on about 445 acres of cotton, (it should be pointed out that if the population had been 100 times higher but a comparable reduction had been attained, the overwintered surviving population of 2800 would still have been much lower than the average of one or two boll weevils per acre which I have arbitrarily set as the desired degree of suppression during Year 1.) This analysis indicates in numerical terms the degree of success of the first year of suppression.
As noted earlier, the boll weevil population in the BWET area was abnormally low. Therefore, it would be presumptuous to assume that similar low populations will be encountered in all areas if an eradication program is extended. For this reason, it seems impor- tant to consider the results to expect from the use of insecticides when the boll weevil population during Year 1 is much higher.
The population in the Trial area was abnormally low when the eradication treatments were started, but the actual number of boll weevils present is not known. A total of 1009 boll weevils were captured in 2293 traps operating in 1978 during the fall insecti- cide treatment period. W. H. Cross (personal communication) esti- mated 2% capture of available boll weevils with the density of traps employed. On this basis, the total population was of the order of 51,000, or 4-5 per acre.

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 419
With these data, we can establish numbers that will be reason- ably representative of maximum densities likely to be encountered in areas of severe boll weevil infestation and then calculate the results expected of a well-monitored in-season control program fol- lowed by a thorough, well-timed spray schedule designed to reduce the overwintered population to a practical minimum. This can be done by the use of simulation models based on conservative parame- ters such as the level of kill that will result from insecticide treatments.
In-season boll weevil applications are usually started when the square puncture level is ca. 10%. Therefore, it is assumed that this level of infestation can be maintained in areas planned for eradication if a well-conducted in-season control program is implemented. When the crop has largely matured and squares are relatively scarce, as few as 200 boll weevils per acre can account for a square puncture rate of ca. 10%. Therefore, this is assumed to be representative of the level of density when actual eradica- tion treatments are started. It is also assumed that each insecti- cide treatment will kill 95% of the weevils present at the time of each treatment, both reproducing and diapausing forms. The in- crease rate for highly favorable boll weevil reproduction is esti- mated to be tenfold. Therefore, each reproducing pair can account for 20 diapausing boll weevils. The diapausing boll weevils are estimated to have 90% natural mortality from the time of emergence in the fields until they enter the fields the next spring. These basic parameters were discussed with J. R. Brazzel and E. P. Lloyd who participated directly in both the North Carolina trial and in the PBWEE. These two authorities (personal communication) consider the basic parameters as being on the conservative side since they are of the opinion that thorough insecticide treatment of the type applied will achieve higher than 95% kill of weevils present. How- ever, it seems desirable to use parameters that are conservative.
On the basis of the assumed number of boll weevils per acre and the effectiveness of the spray treatments, I estimated the results to expect when a population is more representative of nor- mal density than that encountered in the North Carolina trial. For these calculations, a cotton growing area consisting of 25,000 acres is assumed. Then, in order to clearly indicate the expected impact of the insecticide and cultural control components during the first year, I developed a basic model that reflects an otherwise uncontrolled population after a well-executed in-season program has been followed. (This hypothetical population is not subjected to suppression during the diapausing period, which is characteristic of boll weevil management in many areas.) The model can then be used to project the effect of the insecticide application on a sim- ilar population when the applications are directed against the last reproducing generations and the surviving diapausing progeny. On the basis of the parameters given, the calculations show the rela- tive number of overwintered boll weevils per acre for an uncon- trolled and a suppressed population that would enter hibernation

420 E. F. KNIPLING
and emerge to enter cotton fields the next spring. A natural winter mortality of 90% is assigned to each population.
Uncontrolled population (per acre)
Reproducing boll weevils during generation 4 (F^) = 200
Number of diapausing progeny = 2000
Winter mortality = 90%
Overwintered boll weevils per acre entering cotton fields the next spring = 200
Suppressed population (per acre)
Reproducing boll weevils during generation 4 (F^) = 200
Reproducing boll weevils surviving treatment = 10
Number of diapausing progeny = 100
Number of diapausing progeny surviving treatments = 5
Winter mortality = 90%
Overwintered boll weevils per acre entering cotton fields the next spring = 0.5
This estimated effect of Year 1 suppression is based on realis- tic numbers of boll weevils likely to be present in areas under good boll weevil management. The assigned rate of increase seems reasonable. The results assumed after thorough, well-timed insec- ticide applications are probably conservative. The natural mortal- ity also seems reasonable. Therefore, the calculations should indicate in numerical terms the minimum effect expected from the insecticide treatment phase during the first year of an eradication program. This estimated average overwintered population of 0.5 boll weevils per acre would represent a drastic reduction, but it would still mean some 12,500 boll weevils in the hypothetical area, and these would require additional suppression during Year 2 in order to achieve eradication. It is obvious that the number of survivors per unit area will vary greatly in different cotton fields, both in the fall and spring. However, it is the function of the detection component to identify fields where surviving popula- tions are above average so suitable adjustments can be made in the

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 421
intensity and type of control measures to apply. For example, in fringe areas where populations are very low, fewer applications of insecticides should be necessary to reduce overwintered populations to a very low level. Also, if some fields have higher than average overwintered survivors, supplemental insecticide applications may be necessary. In any event, the model depicts the drastic accumu- lative impact of the suppressive procedure used and indicates that prehibernation î)opulations 100 times as high as those occurring in the trial program should be reduced to less than one overwintered
boll weevil per acre.
It might be mentioned that if each insecticide application
kills 98% of the boll weevils, as J. R. Brazzel and E. P. Lloyd contend, the surviving overwintered population would be reduced to
a level of 0.08 boll weevils per acre instead of 0.5 per acre. This would mean an average of one overwintered boll weevil on 12 acres of cotton or about one boll weevil in an average-size cotton field; nevertheless, in an area consisting of 25,000 acres of cot- ton, this would still mean the survival of some 2000 boll weevils that must be killed or whose reproduction must be inhibited if
eradication is to be obtained.
Suppressive Measures Directed Against the Overwintering Generation Versus In-Season Generations
In the previous section, a simulation model was presented to depict the effect of properly scheduled insecticide treatments to reduce the number of diapausing boll weevils that can survive to enter hibernating quarters. It was stated that the diapausing progeny of the last reproducing generation in the fall constitutes a highly vulnerable portion of the population of the pest. The model depicted the effect of in-season suppression of an in-season population to a level of 200 boll weevils per acre until the last reproducing generation in the fall and showed that 95% control of reproduction plus 95% control of the diapausing progeny produced overall suppression of 99.75%. In addition, natural mortality of 90% would limit the number of overwintering survivors. However, it is difficult to fully appreciate the accumulative impact of the factors that cause such high mortality by considering percentages alone. Therefore, it is important to convert percentages to repre- sentative numbers. If a boll weevil population is uncontrolled, one can expect an overwintering population of 1000 per acre. If this potential overwintered population were reduced by 99.75%, the overwintered population would be reduced to an average of 2.5 boll weevils per acre. But the fact is that in most areas, cotton grow- ers today achieve reasonably good control, especially under present supervised integrated pest management (IPM). Under such programs, an overwintering population is not likely to exceed 200 per acre, and 99.75% suppression would mean a reduction to 0.5 boll weevils
per acre, as previously discussed.

422 E. F. KNIPLING
Unless appropriate simulation models are established, it is difficult to visualize the relative effect of different strategies for suppressing the boll weevil. The traditional method of con- trolling the boll weevil has been to attack the in-season genera- tions. Nevertheless, from the standpoint of the dynamics and behavior of the boll weevils, suppressive measures applied during this period are much less efficient than suppressive measures di- rected against the overwintering generation. Indeed, the advantage of the attack on the overwintered generation, as proposed for erad- ication programs, is so impressive that it seems important to ana- lyze the relative efficiency of the two approaches to boll weevil suppression.
We assume, as before, that each application of insecticide kills 95% of the boll weevils present at the time of treatment. We also assume, as before, that reproducing survivors will increase tenfold per generation. The hypothetical overwintering population starts with 200 boll weevils per acre, which is considered repre- sentative of a normal population in an area with a severe infesta- tion. Insecticide applications are programmed at 5-day intervals through five generations, including the F« generation, which is assumed to be the last reproducing generation that produces the diapausing progeny.
This model thus assumes an intensive attack during the growing season but no control of the diapausing population. If we allow 21 days per generation and if insecticide applications are necessary at 5-day intervals to achieve 95% control of a generation, a total of about 18 applications of insecticides will be required to achieve the results shown in Table 1.
Table 1.—Accumulative impact on the dynamics of a boll weevil population subjected to intensive suppression of the pest with insecticides during the regular growing season
Gener- ation
Boll weevils per acre (No.)
Degree of
control (%)
Repro- ducing boll
weevils (No.)
Progeny (No.)
Year 1 1 1/ 200 95 10 100
1.25
2 100 3 50 4 25
95 5 50 95 2.5 25 95 1.25 2/ 12.5 ar 2 3/
Ij Overwintered. 1/ Diapausing. 3_l Natural winter mortality = 90%.

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 423
Therefore, the intensive attack against all the reproducing generations would greatly reduce the overwintered population. How- ever, there would be 2.5 times as many overwintered survivors dur- ing Year 2 as there would be after the application of about six or seven properly scheduled insecticide applications directed against the overwintering population in the fall. (There is also an eco- logical advantage in concentrating suppression against the popula- tion after the cotton crop is made.)
Obviously, the difference in effectiveness of the two ap- proaches to boll weevil suppression cannot be readily visualized and would be difficult and costly to measure in conventional field trials because of the low numbers of survivors per unit area that would be involved under either method. However, appropriate use of simulation suppression models that depict populations in numerical terms clearly indicates the advantage of one suppressive procedure over the other. This is an excellent example of the way simulation models can be used to guide research and development and opera- tional strategies.
Pheromone Trapping Component
Role of the Sex and Aggregating Attractant for Detection
The identification and synthesis of the pheromone of the male boll weevil, grandlure, and the demonstration of the value of grandlure for detection are among the major developments in boll weevil research. This use of grandlure has great practical signif- icance to the concept of boll weevil eradication and to management. The history of the research is reviewed by Lloyd et al. in Chapter 8 of this handbook.
Grandlure provides us with a method of detection that has such a high degree of sensitivity, particularly when it is used in in- field traps, that the problem of determining whether and where low level infestations still exist is no longer a matter of major con- cern in eradication and containment programs. There is little question that technology is available for eliminating incipient boll weevil infestations if it is known where they exist. It is also highly important from the standpoint of optimum efficiency in the conduct of an eradication program to know with a high degree of certainty where the boll weevils no longer exist within an eradica- tion area. In fact, detecting boll weevils in cotton fields was still a major problem when results of the PBWEE were interpreted. This was one reason many pest management authorities were reluctant to accept the results of the Mississippi experiment as a valid dem- onstration of the feasibility of eradicating the boll weevil. The methods then used were based on visual detection of adult boll wee- vils or detection of fertile egg punctures in squares.
The sensitivity of the border and infield traps now used for detection is discussed by Lloyd et al. in Chapter 8 of this

424 E. F. KNIPLING
handbook and will not be discussed in detail here. However, it is important to note that though even one infield trap per acre will detect reproduction by a single female by the F^ generation, an average of slightly under two infield traps per acre (30,244 traps in 15,222 acres) were used in 1979 in the North Carolina trial. No reproduction was detected during at least three generations of boll weevil development during Year 2. Thus, it can be assumed with virtually 100% certainty that the boll weevil population was eradi- cated from the eradication area by Year 2, rather than Year 3, which was the original goal. In the absence of the newly available detection technology, this conclusion could not have been reached during Year 2 and with such high degree of confidence.
One reproducing clump of boll weevils was discovered in Sep- tember 1980 (Year 3) in one field. However, the failure to detect the presence of boll weevils in infield traps during at least three generations in 1979 and at least two generations in 1980 plus the failure to detect boll weevils in border traps in the spring and fall of 1979 and the spring and summer of 1980 makes it possible to attribute the infestation discovered in Year 3 to long-range dispersal from heavily infested cotton or accidental movement from an infested area. In view of the probability of detecting the presence of boll weevils during each of these generations and peri- ods, the chances are of the order of 1,000,000:1 that the infesta- tion was the result of an introduction rather than a surviving population.
The capability of detecting the absence of or existence of reproduction in cotton fields at the lowest reproducing level offers prospects for optimizing the effectiveness and efficiency of the various procedures available for boll weevil suppression.
Role of the Sex and Aggregating Attractant as a Suppressive Measure
The potential role of pheromone traps for the control of very low overwintered boll weevil populations has not yet been fully ex- ploited. However, available information and general observations indicate that border field traps used at a rate of one per acre and subsequent infield traps used at a rate of three to four per acre after cotton fruiting begins would make a major contribution to suppression of very low populations.
On the basis of the competitive theory of attraction (Knipling and McGuire 1966), the efficiency of pheromone traps baited with the synthetic replica of a natural pheromone is inversely correla- ted with the. density of the insects that produce the competing nat- ural pheromone. The validity of this theory has been confirmed in principle by studies conducted by Lloyd et al. (1972). Thus, the lower the population of overwintered boll weevils, the greater will be the suppressive action of a given density of traps. Knipling (1976a) developed models to estimate the effect of infield traps

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 425
used against low-level overwintered populations. The calculations indicate that at a trap density of 10 per acre, a high degree of suppression can be expected when the overwintered population has been reduced as low as 10 per acre. It was also calculated that ca. 98% control could be expected when the overwintered population was as low as two per acre. Since a properly executed Year 1 pro- gram of suppression of the boll weevil can be expected to reduce an overwintered population well below an average of two boll weevils per acre and since the lower the overwintered population, the more effective a given number of traps will be, the prospects seem ex- cellent that a border and infield trapping system could have a vital role in both detection and suppression of a low density boll weevil population.
The validity of the theoretical models has not been fully con- firmed, but Lloyd et al. (1980) obtained encouraging data concern- ing the effectiveness of infield traps used against simulated "clumped" F, progeny from single reproducing females in small cot- ton fields of about three acres. Although these studies were de- signed to determine the effectiveness of infield traps for detec- tion, the results also indicated that four infield traps per acre will capture more than 90% of an average population of ca. eight female boll weevils emerging in each cotton field. These data, while preliminary, suggest that the parameters used in early calcu- lations of trap density may have been conservative and that the probability is high that pheromone traps could make a major contri- bution to the elimination of overwintered boll weevils when densi- ties average less than one per acre. I will consider this possi- bility later in connection with a discussion of opportunitier for further improvement in eradication technology.
Sterile Boll Weevil Release Component in Eradication
Although the number of overwintering boll weevils surviving the insecticide suppression component of an eradication program may be at a very low level, reproduction by the few survivors must be inhibited. When suppression measures that are equally effective at all pest densities are employed, it may take as much effort and cost to kill 1% of a surviving population as it requires to sup- press the first 99-99.9%. Nevertheless, even when a boll weevil population reaches an average density of one per acre, there are thousands of survivors in an ecosystem consisting of thousands of acres of cotton. These survivors must be eliminated to achieve eradication, but reliance on insecticides to achieve this purpose would be inefficient and could be objectionable from an ecological standpoint.
In contrast, the sterile insect technique becomes increasingly efficient as a population declines. Also, boll weevil rearing has reached such efficiency that the cost of rearing is not likely to exceed $2-$3 per thousand (Griffin et al. Chapter 11 in this hand- book). Therefore, when natural populations reach an average level

426 E. F. KNIPLING
of one or fewer boll weevils per acre, an overwhelming ratio of sterile to native boll weevils can be achieved at low cost. This is important because sterilized males may not approach the competi- tiveness of the native males. However, when native boll weevils can be outnumbered by a factor of several hundred, release of wee- vils that are only 5-10% as competitive will still prevent repro- duction in virtually all cotton fields.
As previously noted, only seven native overwintered boll wee- vils were captured in the North Carolina eradication zone. It was postulated that the total native boll weevil population in 15,222 acres of cotton was likely of the order of 28. Since a total of about 11 million sterile boll weevils was released, the sterile to fertile ratio achieved was of the order of several hundred thou- sands to 1. When the native population is at such a low level, there is no way to make a statistical measurement of the effect of the sterile boll weevils. Neither can one measure the effect of sterile boll weevil releases on the basis of overall ratios since that depends on the ratio of fully competitive sterile weevils to fertile weevils in the specific habitats of the native boll wee- vils. Even so, the release of sterile boll weevils provides an efficient system for final elimination that is ecologically safe and highly complementary with other suppression techniques.
Sterile boll weevils in cotton fields may do more than inter- fere directly with reproduction of native females when the native population is greatly reduced. Their presence should prevent the normal aggregation of the few native boll weevils remaining. It is not possible to obtain statistical data concerning this effect that are adequate for analysis. Yet, by rationalization as well as by observation, aggregation of overwintered boll weevils is a natural behavior of boll weevils. It undoubtedly evolved to in- crease opportunities for mating in natural low and widely scattered populations. In view of the high degree of attractiveness of the pheromone produced by the boll weevil, mate finding may not be a serious problem to successful reproduction, even when a single male and female are present in an average-size cotton field. However, in the presence of several hundred sterile boll weevils, it seems highly unlikely that so few native males and females could aggre- gate because of the competing pheromone produced by the released males. This same effect could result from high density trapping. For example, if a native population averages one male and one fe- male per 10 acres and if a thousand or more sterile boll weevils are present that produce the pheromone, matings may seldom occur, even if the males have a low level of mating competitiveness. Ran- dom distribution predicts the presence of two males only in about 25% of the cotton fields, two females in 25% of the fields, and one male and one female in ca. 50% of the cotton fields. (Such calcu- lations ignore the probability of more or fewer boll weevils in some fields.) However, even in fields with several of each sex, the probability of encounters between native males and females may be nil in the presence of several hundred sterile males that would interfere with the normal aggregation of low density populations.

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 427
These effects have not yet been investigated, but they are likely to be of very great practical significance in the final elimination of boll weevil populations.
Finally, it should be pointed out that the sterile boll wee- vils may reduce the opportunity for reproduction of any native boll weevils that have survived exposure to insecticides because of an insecticide-resistant gene or genes. In the absence of competing sterile boll weevils, a few survivors carrying genes for resistance would have the opportunity to reproduce, which would immediately increase the frequency of resistant genes. This will not be the case if the few surviving weevils with resistant genes mate with sterile boll weevils.
Diflubenzuron Component in Eradication Programs
The development of the growth regulating chemical, difluben- zuron, as a control measure for the boll weevil added a new compo- nent that could be valuable in boll weevil eradication (Bull et al. Chapter 9 in this handbook). The initial plans were to make sub- stantial use of the material as a supplementary suppressive measure in individual cotton fields where trapping data indicated the presence of enough overwintered survivors so sterile males might not control reproduction adequately. In fact, the capture of na- tive boll weevils in survey traps was so low there was little need for supplemental diflubenzuron applications. Nevertheless, about 900 acres of cotton were treated with diflubenzuron as a security measure in areas where the few overwintered boll weevils were captured.
Although the need for diflubenzuron did not materialize in the North Carolina trial, the potential value of this chemical to any eradication program should not be discounted. Diflubenzuron has relatively little effect on beneficial insects, a feature of great importance in cotton insect management or eradication, since broad- spectrum insecticides destroy the beneficial insects that hold such secondary pests as Heliothis spp. in check. Thus, when supple- mental chemical treatments were needed during the growing season, diflubenzuron would be preferred. This growth regulating chemical has little adverse effect on adult boll weevils, but females cannot produce progeny for a week or longer after exposure to the mate- rial. Therefore, a series of applications superimposed on releases of sterile males would have a compounding effect and result in greater suppression than can be obtained by either technique used alone.
Diflubenzuron also is important in producing sterilized boll weevils for release (Wright and Villavaso Chapter 7 in this hand- book). Irradiation alone at an acceptable dosage level does not assure near 100% sterility of female boll weevils. However, ir- radiation plus diflubenzuron in the adult diet assures virtually 100% sterility of both females and males.

428 E. F. KNIPLING
OPPORTUNITIES FOR IMPROVEMENTS IN BOLL WEEVIL ERADICATION TECHNOLOGY
New technology in any field of science is never fully perfect- ed until all aspects of the technology are thoroughly investigated and until adequate experience is gained to permit optimal use of technology that may be available. Therefore, it would be reason- able to assume that the technology used in the North Carolina trial can be made more effective and more efficient through additional research and development. Although there is every reason to have full confidence in our ability to eradicate the boll weevil on the basis of available technology and experience, it is my view that the probability is very high that substantial improvements can be realized through further research and operational experience. The same opportunity should exist for the improvement of management procedures. It would seem important that the possibilities for improvements in technology be kept in mind by those who will have the responsibility for making a decision on boll weevil management or eradication. However, my analysis here deals primarily with eradication technology.
I have already alluded to several opportunities for increased efficiency. For example, the degree of suppression in terms of survivors during Year 1 in the North Carolina trial was of the or- der of 100 times as high as would have been necessary to reduce the population to a level that could readily have been eliminated by the suppressive components available for Year 2. Therefore, sub- stantial savings in cost, especially in areas with low boll weevil density, can in all likelihood be realized by reducing the number of insecticide applications.
There are opportunities for increasing the vigor and competi- tiveness of sterile boll weevils which could mean greater effi- ciency or less cost of the sterile male component. For example, research is underway at the ARS Metabolism and Radiation Research Laboratory, Fargo, ND, in cooperation with scientists of the Uni- versity of Minnesota, in an effort to select a strain of boll wee- vil that will survive longer after the application of a sterilizing dosage of irradiation. The preliminary results are encouraging.
Then modified cotton culture and host plant resistance, as discussed by Namken et al. (Chapter 4 in this handbook), could con- tribute to eradication in certain areas. In particular, appropri- ate cultural measures designed for the southern portions of the boll weevil area could contribute materially to eradication and the maintenance of eradication. It was mentioned earlier that infield traps may be used to eliminate low density boll weevil populations and at the same time provide highly sensitive detection. Indeed, four infield traps per acre will capture more than 90% of the F females from a single reproducing overwintered female. If this de- gree of efficiency against clumped F. boll weevils can be obtained by so few traps per acre, it is probable that an even higher cap- ture efficiency can be achieved by the same number of traps

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 429
operating against a low density overwintered population. For ex- ample, at least some males are likely to respond to infield traps as they do to border traps. Also, boll weevils entering trapped cotton fields from hibernation are likely to enter in a distributed pattern rather than a clumped pattern because the pheromone emanat- ing from the traps would interfere with normal aggregation. The data indicate that at a density of four well-distributed traps per acre, each trap baited with the optimum amount of pheromone will equal or exceed a competing male in attracting females. On the basis of these preliminary results, which will be given more inten- sive study, it may be possible to use infield traps as a highly sensitive method of detecting overwintered boll weevils and as a method of providing control when densities are reduced to levels below an average of one boll weevil per acre. The use of infield traps may therefore obviate the need for releases of sterile boll weevils, especially in areas where boll weevil populations are
normally low.
In view of the excellent prospects of making greater use of infield traps for both detection and elimination of very low sur- viving overwintered populations, a theoretical suppression model has been developed as depicted in Table 2. The model is based on the assumption that by the time cotton fruiting begins, overwinter- ing survivors will average 0.4 boll weevil per acre or four weevils per 10-acre field. This is the approximate population level calcu- lated for the controlled population described in my first model. Insecticides would be applied when squares are first subject to infestation in order to destroy all early emerging boll weevils not
Table 2.—Theoretical control of 40 overwintered boll weevils en- tering a 100-acre cotton field in which 400 infield traps are operating. Parameters are discussed in the text
Proba- Proba- bility bility
No . of No. of Ratio of of native of
boll weevils traps in traps to female female
Day Male
2
Female operation males matings capture
1 2 400 200:1 0.005 1.995
2 4 2 400 100: 0.010 1.990
3 6 2 400 67: 0.015 1.985
4 8 2 400 50 0.020 1.980
5 10 2 400 40 0.025 1.975
6 12 2 400 33 0.030 1.970
7 14 2 400 29 0.034 1.966
8 16 2 400 25 0.040 1.960
9 18 2 400 22 0.046 1.954
10 20 2 400 20 0.050 1.950
Accumulative probability 0.275 19.725

430 E. F. KNIPLING
captured in border field traps. The infield traps would be used at the rate of four per acre, or 400 traps on 100 acres. The larger acreage is assumed in order to deal with larger numbers of boll weevils and minimize the use of fractions in making calculations. To further simplify calculations without significantly changing the results, I have assumed that emergence will occur at a given rate during each of 10 days. The model shows the high ratio of traps to competing native males for the attraction of the relatively few fe- males as the males and females enter the trapped fields. Although this ratio will decrease as the native males accumulate, the total theoretical effect will be to prevent reproduction in about 99% of the fields. This would mean that no reproduction would occur in most cotton fields. However, even if occasional females in an area consisting of thousands of acres of cotton are successful in mat- ing, which would be expected, the traps would, during the period of F. emergence, identify the sites where reproduction had occurred and, at the same time, provide a high degree of control. In any event, if F, progeny are captured in any of the fields, supple- mental treatments with an appropriate insecticide could eliminate the infestation. It should be noted that the suppression model assumes that none of the males would be captured. If a significant number of males are captured, the ratio of traps to competing males would be higher which, in turn, should result in the capture of a higher proportion of the females.
TECHNOLOGY FOR MAINTAINING AREAS FREE OF BOLL WEEVILS
Some question the rationale of undertaking boll weevil eradi- cation on the grounds that even if successful, the pest will become reestablished after the eradication program terminates and that another eradication program of a similar magnitude and cost would again be necessary. It may also be argued that a continuing pro- gram of surveillance and eradication to prevent reestablishment of the pest in vulnerable areas would be so costly that little would be gained from eradication. These are highly speculative assump- tions, of course. Nevertheless, such questions and implications may well be a matter of major concern to agricultural administra- tors, budgetary officials, and others in decision-making roles. Therefore, it seems highly important to analyze this possibility. Likewise, it would be unfortunate if boll weevil eradication pro- grams were discouraged on the basis of a continuing cost. Such cost might well amount to several million dollars annually, but such a sum is a very small fraction of the continuing annual cost of $200 million to $300 million that can be expected if eradication is not undertaken, to say nothing of the adverse ecological effects of wide-scale, intensive use of chemical insecticides for the in- definite future.
For maintenance, a good monitoring program will obviously have to be instituted in the Rio Grande Valley for an indefinite period to serve as a guide to the nature and intensity of

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 431
suppressive measures that will be required to keep the pest at a low level. This cotton growing area is in direct flight range of boll weevils developing on cotton (and perhaps on wild host plants) in Mexico or in some areas in south Texas. The suppressive meas- ures that would be used, if required, should make optimum use of available technology, including the use of insecticides that have minimum effect on nontarget organisms, the use of pheromoiie traps, the application of cultural measures, and the growing of short- season varieties of cotton. The release of sterile boll weevils on a routine basis may also assure the degree of suppression necessary to prevent infiltrating boll weevils from reaching high levels. The objective would be to maintain infestations at a practical min- imum, partly to protect crops in the Valley region from excessive damage but primarily to minimize the possiblity of boll weevil spread to cotton growing areas north of the Valley. Fortunately, cotton-free ranch land extends for a distance of about 50 miles from Valley cotton to other cotton growing areas in southern Texas, and this natural barrier and a low boll weevil population at the source of possible spread should assure minimum dispersal. Even so, an adequate surveillance program will, no doubt, have to be maintained throughout south Texas to detect any boll weevil repro- duction that may result from dispersal from infested cotton grown in the Valley and in Mexico border regions or possibly from wild host plants. Any infestations that might be detected north of the Rio Grande Valley would require prompt elimination.
Also, pheromone traps should be used for at least several years in all cotton growing areas in order to detect infestations that may occur because of shipment of infested commodities or hitchhiking boll weevils. Only experience will tell us how long and to what extent surveillance will be necessary in the various cotton growing regions. However, the cost of maintaining at least one trap per cotton field in uninfested areas is not likely to be excessive and could be made a part of a general IPM monitoring
program for cotton and other crops.
In sum, it is by no means certain that season-long suppression of boll weevils on the total acreage of cotton grown in the Rio Grande Valley region adjacent to Mexico would be necessary. How- ever, on the basis of available information about the flight range of the boll weevil, it must be assumed that the pest will continue to infiltrate into the Valley in sufficient numbers to require con- tinuing suppressive efforts so long as uncontrolled populations exist in cotton or wild host areas within 50 miles or more of the border area. A well-organized, well-executed boll weevil suppres- sion program designed to maintain populations at a very low level so as to minimize the chances of spread may.cost little, if any, more than the average losses due to the pest under present manage- ment practices. In fact, an organized program designed to minimize reinfestation and spread of the pest from Mexico might actually reduce costs. It seems apparent that in the event the boll weevil is pushed back to the border region, a joint boll weevil management

432 E. F. KNIPLING
effort by Mexico and the United States would be considered; and this might minimize the possibility of continuous infestations in the Valley.
Some argue that the boll weevil is likely to become widespread before any reintroduced populations are detected and that the need for costly eradication will recur frequently. The experience of the North Carolina trial is highly reassuring in both respects. The single infestation discovered near the North Carolina/Virginia line in 1980 was detected before the population had reached a high level and spread. The discovery of only one established reintro- duction during two seasons in some 40,000 acres of cotton indicates that reintroductions are not likely to be frequent. This is espe- cially reassuring because there was heavily infested cotton within 100 miles of the eradication zone. In any case, use of the highly sensitive detection methods now available should virtually assure that infestations will be detected before they reach high levels and become widespread. Finally, as to possible concerns about the difficulty of eliminating reestablished infestations, if we have the technology to eliminate well-established infestations on mil- lions of acres of cotton, we surely have the capability of elimi- nating any possible incipient reintroduced infestations. The key to preventing the reestablishment of any pest that has been erad- icated is to maintain adequate surveillance and to take prompt action to eliminate any reintroduced infestations before they be- come widespread.
GENERAL SUMMARY AND CONCLUSIONS
Technology for possible use to eradicate an insect has never before been tested so thoroughly as was the technology proposed for use against the boll weevil in the event an eradication program is undertaken. On the basis of highly favorable results obtained in North Carolina, it can be stated without reservations that adequate technology for boll weevil eradication is available. It has the following components: Year 1 : (1) a well-monitored, well- executed, in-season boll weevil management program; (2) an inten- sive insecticide and cultural spray schedule involving an average of about six or seven applications of chemical following the in- season control program that is directed against the hibernating population. Year 2: (1) the use of pheromone traps for detection and to further suppress the low number of surviving overwintered boll weevils; (2) when cotton starts fruiting, release of sterile boll weevils at a rate of about 100 per acre per week for a period of ca. four weeks; (3) the use of infield boll weevil traps follow- ing the termination of sterile boll weevil releases, in order to detect and subsequently eliminate any boll weevil reproduction that may persist; (4) the supplemental use of chemical insecticides as needed.
In view of the abnormally low boll weevil population that ex- isted at the beginning of the trial program in North Carolina, the

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 433
procedure used in the trial program during Year 1 virtually elimi- nated the population by the use of chemicals alone. However, sim- ulation models were developed to estimate the degree of suppression that might be expected from available technology when the boll wee- vil population is representative of normal numbers likely to be encountered in severely infested boll weevil areas. The simulation models suggest that if a boll weevil population were 100 times higher than that in the North Carolina trial, a thorough in-season management program followed by about five to seven properly timed insecticide treatments would still have reduced the overwintered population to an average of less than one boll weevil per acre. Such low populations should be readily eliminated by the pheromone traps, sterile boll weevil releases, and strategic application of insecticides used during Year 2.
The high degree of effectiveness of the several suppressive measures, when properly integrated, is attributed in large measure to the strategy of concentrating suppressive efforts against the overwintering generation before and after the insects leave the hi- bernating quarters. The overwintered population is also subject to high natural mortality due to winter hazards, which adds greatly to the suppressive efforts. The compounding effect of the several mortality factors can reduce very high potential populations to near zero.
Although available technology for boll weevil eradication has been well demonstrated, the possibilities are excellent for further improvements and reduced costs. There are possibilities for making optimal use of insecticides and cultural measures during Year 1. There are also possibilities for improving the competitiveness and quality of sterile boll weevils. The use of pheromone traps, both for detection and as a suppressive measure, offers unusual opportu- nities for possible application against surviving populations that may average less than one boll weevil per acre.
The unusually high boll weevil detection capability of phero- mone traps placed around borders of cotton fields and inside the cotton fields is considered one of the most important developments in optimizing the efficiency of boll weevil eradication procedures and for detecting and making possible prompt elimination of any re- introduced infestations that may occur in areas free of boll wee- vils. The experience in the trial program indicates that reintro- ductions into areas free of boll weevils are likely to be infre- quent. Only one reintroduction was discovered in the eradication zone during two seasons despite the existence of normally infested cotton within 100 miles of the eradication zone.

434 E. F. KNIPLING
LITERATURE CITED
Adkisson, P. L., D. R. Rummel, W. L. Sterling, and W. L. Owen, Jr. 1966. Diapause boll weevil control: a comparison of two meth-
ods. Texas Agricultural Experiment Station, Bulletin No. 1054, 11 p.
Brazzel, J. R. 1961. Destruction of diapause boll weevils as a means of boll weevil control. Texas Agricultural Experiment Station, Miscel- laneous Publication No. 511, 22 p.
T. B. Davich, and L. D. Harris. 1961. A new approach to boll weevil control. Journal of Eco- nomic Entomology 54:723-730.
and L. D. Newsom. 1959. Diapause in Anthonomus grandis. Journal of Economic Entomology 52:603-611.
Eden, W. G. 1976. Report of Entomological Society of America Review Com- mittee on the Pilot Boll Weevil Eradication Experiment. In Proceedings, Conference on Boll Weevil Suppression, Manage ment, and Elimination Technology, Memphis, TN, February 13-15, 1974. U.S. Department of Agriculture, ARS-S-71, 126 p.
Fye, R. E., C. L. Cole, F. C. Tingle, A. Stoner, D. F. Martin, and L. F. Curl.
1968. A reproduction-diapause control program for the boll weevil in the Presidio, Texas - Ojinaga, Chihuahua area, 1965- 67. Journal of Economic Entomology 61:1660-1666.
Knipling, E. F. 1960. Use of insects for their own destruction. Journal of Economic Entomology 53:415-420.
1968. Technically feasible approaches to boll weevil eradica- tion. Ln Proceedings, Beltwide Cotton Production-Mechanization Conference, Hot Springs, AR, January 11-12, 1968. National Cotton Council of America, Memphis, TN.
1976a. Biomathematical basis for suppression and elimination of boll weevil populations. In Proceedings, Conference on Boll Weevil Suppression, Management, and Elimination Technology, Memphis, TN, February 13-15, 1974. U.S. Department of Agricul- ture, ARS-S-71, p. 130-148.

TECHNOLOGY FOR ERADICATION OF BOLL WEEVIL 435
Knipling, E. F. 1976b. Report of Technical Guidance Committee for the Pilot Boll Weevil Eradication Experiment. _In Proceedings, Conference on Boll Weevil Suppression, Management, and Elimination Tech- nology, Memphis, TN, February 13-15, 1974. U.S. Department of Agriculture, ARS-S-71, p. 122-125.
1979. The basic principles of insect population suppression and management. U.S. Department of Agriculture, Agriculture Hand- book No. 512, 623 p.
_^ and J. U. McGuire, Jr. 1966. Population models to test theoretical effects of sex
attractants used for insect control. U.S. Department of Agri- culture, Agriculture Information Bulletin No. 308, 20 p.
Lloyd, E. P., G. H. McKibben, E. F. Knipling, J. A. Witz, A. W. Hartstack, J. E. Leggett, and D. F. Lockwood.
1980. Mass trapping for detection, suppression, and integration with other suppression measures against the boll weevil. Pre- sented at an international colloquium on the management of insect pests with semio-chemicals, Gainesville, FL.
M. E. Merkl, F. C. Tingle, W. P. Scott, D. D. Hardee, and T. B. Davich.
1972. Evaluation of male-baited traps for control of boll weevils following a reproduction-diapause program in Monroe County, MS. Journal of Economic Entomology 65:552-555.


437
Chapter 17
COTTON AND INSECT MANAGEMENT SIMULATION MODEL
L. G. Brown Department of Industrial Engineering Mississippi State University Mississippi State, MS 39762
R. W. McClendon Department of Agricultural and
Biological Engineering Mississippi State University Mississippi State, MS 39762
J. W. Jones Department of Agricultural Engineering University of Florida Gainesville, FL 32611
ABSTRACT The cotton crop model COTCROP was interfaced with the boll weevil model CIM-BW and the Heliothis spp. model CIM-HEL to form the Cotton and Insect Management (CIM) model. The effects of beneficial insects and insecti- cides were also included. The CIM model was structured to aid in the development and evalu- ation of insect pest management strategies.
Simulated results with selected popula- tions of boll weevils (Anthonomus grandis Boheman), Heliothis spp., and predators, with and without insecticide applications, are presented.
The CIM model starts on the day of emer- gence of the cotton plant and advances through the season. The status of the crop and that of the insect populations are updated each day.
INTRODUCTION
Current approaches to cotton insect pest management, like past ones, rely heavily on insecticides to prevent pests from reducing yield and economic value of the crop. This type of management has allowed producers to satisfy the demand and to realize profits, but new methods are needed today. Insecticides have harmful side ef- fects on nontarget organisms, and extended use of an insecticide

438 L. G. BROWN ET AL.
may produce resistance in the target species. As an example, Heliothis virescens (F.) has evolved as a major cotton pest in some areas because of its increased resistance to organophosphate insec- ticides (Harris 1972). In addition, costs of the fossil fuels used in the production of many insecticides are increasing rapidly.
Two pest management systems (see chapters 15 and 16 of this handbook).have been proposed as alternatives to current methods of controlling insects in cotton. One was tested in the Boll Weevil Eradication Trial (BWET) and the other in the Optimum Pest Manage- ment Trial (OPMT). Both Trials were regional because the mobility of the target insects is such that traditional field plot experi- ments would not provide sufficient information. Results of the two approaches were then evaluated to determine the advantages and dis- advantages of each, and, in particular, the impacts on yield, in- secticide usage, and net return above insect management costs.
Three methods of determining the effects of the Trials on yield and insecticide usage were studied: direct experimental method, regression analysis, and simulation. Replicated experi- ments, of course, can be designed to produce a direct comparison of yield for a range of soils, weather, and insect and management practices. However, the variations in climate and soil, the dif- ferences in time of development and in the magnitude of insect populations, and the differences in management practices in the two Trial areas were so great that the required degree of experi- mental control was difficult to achieve. Regression analysis involves use of historical data to explain the factors that produce variations in yield of a crop and thus requires a large amount of historical data; however, such a data base was unavailable.
For simulation, a model is developed that is capable of mimick- ing the behavior of the real system. First, the objective is de- fined; then the system is divided into relevant components. Next, the experimental method is used to study the components and to quantify the effects of important variables on each. Mathematical descriptions of the processes that change the state of the compo- nents are developed. These mathematical relationships are then combined and solved by computer simulation so as to study the be- havior of the entire system.
In this case, the objective was to determine the effects of insect management strategies on insecticide usage and yield of cot- ton grown on different soil types and exposed to different weather and levels of insect populations. The system was therefore the cotton field, and the components were the crop and the populations of boll weevils (Anthonomus grandis Boheman), Heliothis zea (Bod- die), H. virescens, and predators. Each component would be affect- ed by weather conditions and by management strategies and tactics. The crop integrates, through time, the effects of weather, insects, and management tactics on growth and yield. Simulation appeared feasible because of earlier efforts to develop and interface cotton and insect models (Jones et al. 1974, 1977; McKinion et al. 1975;

COTTON INSECT MANAGEMENT SIMULATION MODEL 439
Brown et al. 1976, 1979; Hartstack et al. 1976; Gutierrez et al. 1977; McClendon et al. 1977).
In this chapter, we describe the Cotton and Insect Management (CIM) simulation model that was developed by interfacing three com- ponent models, the crop model (COTCROP), the boll weevil model (CIM-BW), and the Heliothis spp. model (CIM-HEL). A sensitivity analysis of the CIM model is presented to give the reader an idea of the responsiveness of the model to changes in critical factors. Although this model is discussed relative to specific insect man- agement strategies (BWET, OPMT, and the Current Insect Control (CIC)), it has potential for evaluation of other strategies.
COTTON CROP COMPONENT MODEL
The cotton crop component model, COTCROP, was developed by an interdisciplinary research team of engineers, plant physiologists, economists, and entomologists (Jones et al. 1980; Brown et al. 1982) to study insect and other management strategies including ir- rigation and nitrogen fertilization. Subsequently, COTCROP was modified, integrated into the more comprehensive system model (CIM), and tested for use in insect management studies.
Considerable research and several single (average) plant mod- els were developed prior to COTCROP; and these provided much of the background for the current model. Two such models were developed in the late 1960*s and early 1970's: SIMCOT (Duncan 1972; Hesketh et al. 1972a, 1972b) for southeastern conditions, and COTTON (Sta- pleton et al. 1973) for western production under irrigation. SIM- COT was later revised and became SIMCOT II (Jones et al. 1974; McKinion et al. 1975). These models simulated single plants by maintaining a carbohydrate balance, and later a nitrogen balance was included. They predicted the production of discrete numbers of fruit by an average plant; therefore, to estimate production by plant populations, they multiplied numbers of fruit by the plant population. However, to include insect feeding on fruit and floral buds, one must vary the number of fruit and floral buds per area in a continuous manner rather than discretely with large increments. Brown et al. (1976) and Gutierrez et al. (1977) therefore restruc- tured SIMCOT II for interfacing with insect models in studies of insect management strategies.
In COTCROP, crop growth is calculated for plants growing on 1 m of ground area. It maintains carbohydrate and nitrogen bal- ances for the plants and water and nitrogen balances for the soil. Each day's demands (or sink strengths) for carbohydrates and nitro- gen are calculated on the basis of organogénesis and growth rates of the plants and organs. The available carbohydrate is then de- termined on the basis of carbohydrate reserves in the plant and photosynthate produced. Available nitrogen is determined from plant uptake on the basis of depth of roots and distribution of nitrogen in the soil. If there is insufficient carbohydrate or

440 L G. BROWN ET AL.
nitrogen to meet daily demands, the rate of organ initiation and the growth rates of existing organs are reduced until the demand for nutrients is equal to the amount available. A surplus of either nitrogen or carbohydrate is stored in the crop for later use; a shortage of either nitrogen or carbohydrate causes fruit of different ages to be abscised after a certain period. Water stress also causes the abscission of fruit and retards the development of new organs for several days.
Model Structure
The mathematical structure of the model is described by vec- tors of continuous state variables plus a discrete time transition matrix as,
\ ' \ \-i * \ til
where
X = a state variable vector; A = the transition matrix for day t; and b = the growth and organogénesis rate vector for day t.
The vector x represents several column vectors describing the state of the crop on day t, and each of these has m elements, where m is the maximum number of days of the simulation. For ex- ample, FR is the (mxl) vector of fruit densities and fr (i) is the vector element representing the fruit density on day t for fruit "t-i" days old since they were initiated on day i. In addition, on day t the mass of the fruit that are "t-i" days old is maintained in the ith element of vector FRW . Likewise, the nitrogen content of the fruit is maintained in the vector NFR . Similar variables are maintained for leaves and stems although root mass is a scalar quantity.
Growth rates of existing organs of the plant depend on poten- tial growth rates (which vary with temperature and the age of the organs) and supplies of carbohydrate and nitrogen. Insect damage to fruit reduces the number and mass of growing fruit and therefore reduces total boll growth or fruit sink strength on a given day. The balance between the supply of and demand for carbohydrate and nitrogen throughout the season alters the initiation of crop organs and the growth rates and thus affects both the fruit density and yield. Therefore, insect damage to fruit will have different mag- nitudes of effect, depending on the time of the year and growing conditions.
Because of the physiological detail in COTCROP, simulation is required to solve the model and describe the growth response of the

COTTON INSECT MANAGEMENT SIMULATION MODEL 441
cotton crop to variations in temperature, insect damage, rainfall, solar radiation, soil characteristics, and nitrogen fertilization. Focus is on processes; for example, organ initiation, organ growth, photosynthesis, respiration, and nitrogen uptake. A schematic of the subroutines that are structured around the processes included in COTCROP is presented in Figure 1. For a description of each subroutine, see Jones et al. (1980) and Brown et al. (1982).
Soil Model
The soil model in COTCROP maintains water and nitrogen bal- ances. The soil is divided into L homogeneous 10-cm deep layers, each having distinct volumes of water and concentrations of nitro- gen. Availability of water and nitrogen to the plant depends on root depth. Root depth increases in time to a maximum depth, which is an input to the model. The emphasis of this soil water balance is on the overall soil volume; thus, all boundary values are calcu- lated first.
The overall soil water balance is
W, = W^_^ 'h-\- E,t - ^t " \ [2]
where
W = water volume in the root zone on day t;
I = the irrigation volume on day t;
D = the drainage volume on day t;
E = the soil evaporation on day t;
E = the plant transpiration on day t; and
\ the rainfall infiltration on day t (all in cm water per cm soil area).
Redistribution of water in the one dimensional soil profile is calculated on the basis of an approximate finite difference (x 10 cm) solution to the water flow equation
a0(x,t) = ^[D(0) 9e(x,t) + k ] -S [3] 3t 8x 3x ® ^
where
0(x,t) = volumetric soil water content on day t at depth x;
D( 0) = the soil water diffusivity;
k = the unsaturated hydraulic conductivity; and
S = the rate of uptake by roots.
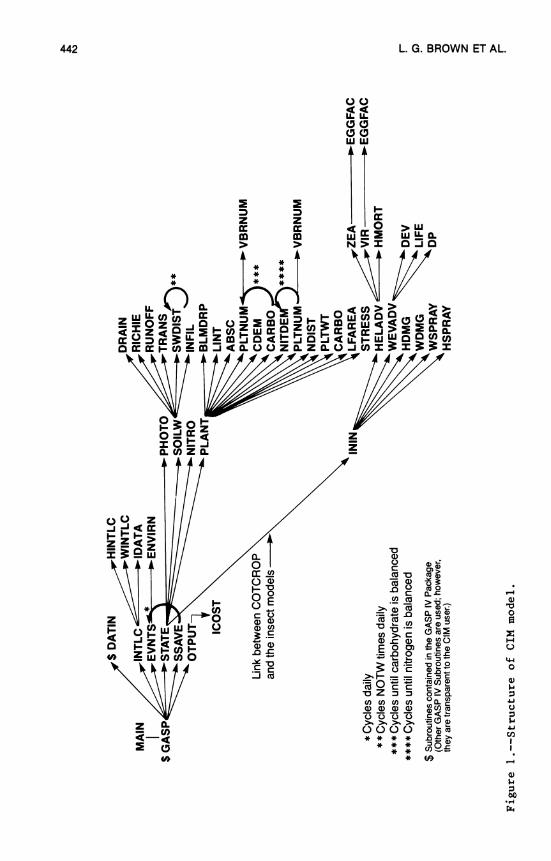
442 L G. BROWN ET AL
o 6
u O
u u w I I
60

COTTON INSECT MANAGEMENT SIMULATION MODEL 443
The equation is solved with boundary conditions of (1) drainage as calculated by the method of Jackson and Whisler (1970) or (2) an upward flow from a wetter zone. The boundary conditions depend on the soil conditions and on soil evaporation and plant transpiration as calculated by the model of Ritchie (1972). _!_/ Rainfall infil- tration is calculated by the Soil Conservation Service method (Schwab et al. 1966). The model, excluding redistribution, was tested by Jones et al. (1976) for Leeper sandy loam soil in Mississippi.
Nitrogen in the soil is assumed to be nitrate in solution. Nitrogen uptake by the plants is proportional to the mass flow of the transpiration stream. Redistribution of nitrogen in the soil is by mass flow; for this, a dispersion coefficient as outlined by Gardner (1965) is used.
Crop Growth Model
Considerable data on crop growth were available for arriving at estimates for rates of photosynthesis (Baker et al. 1972), organ initiation (Jones et al. 1974), and growth rates (Hesketh et al. 1972a, 1972b). However, several relationships such as the effects of carbohydrate, nitrogen, and water stresses on fruit abscission and organ initiation were not available. Therefore, carbohydrate and nitrogen stresses were defined as the ratios of supply to de- mand of carbohydrate and nitrogen, respectively. Water stress was considered a nonlinear function of average soil-water potential (H. D. Bowen unpublished data).
The fraction of fruit of each age that would abort given the severity of predicted stresses was based on data of Bruce and Romkens (1965). In their 1961 experiments on Leeper sandy loam, one of the treatments, AAA, consisted of frequent irrigations that maintained soil water potential above -0.3 bar throughout the sea- son. Another treatment, ADD, consisted of maintaining the same water level until plants reached the small boll stage; then when soil dried to a water potential of -2.4 bar, the plots were irri- gated. (Rainfall was excluded by rainfall shelters.) The result- ing data on soil-water status, plant development, and weather pro- vided an excellent data set for model calibration. (Nitrogen stress probably did not occur because of the high rate of nitrogen fertilization (300 pounds per acre).)
A trial and error procedure was therefore used to calibrate the abscission parameters. Three comparisons of simulated results with experimental data were used in the calibration: number of fruiting sites, number of fruit, and final yield. Both AAA and ADD treatments were simulated. Actual yields were 3.18 and 2.46 bales
\J Appreciation is expressed to L. A. Smith, ARS, U.S. Depart- ment of Agriculture, Mississippi State, MS, for assistance in im- proving the solution of soil-water redistribution in the soil profile.

444 L G. BROWN ET AL.
per acre for treatments AAA and ADD, respectively. Corresponding simulated yields (after calibration) were 3.22 and 2.69 bales per acre.
The percentages of fruit assumed to abscise per day were 11, 2.5, and 2.5 for young, medium age, and old squares, respectively, and 13 and 9 for bolls less than 10 days old and bolls between 10 and 20 days old, respectively. These percentages are the maximum per day; they occur only with severe stress conditions such as zero supply of either carbohydrate or nitrogen. It was assumed that older bolls do not abscise. Most fruit abscission occurs when the fruit are young squares and young bolls according to experimental data of Jones (1975).
Discussion
A subsequent experiment was conducted in 1977 at the site of the Bruce and Romkens experiments (J. D. Hesketh unpublished data 1977). However, there were two major differences between the two experiments. First, in 1977, the rate of nitrogen application was 100 pounds per acre. Second, water stress was allowed to occur earlier during the season. Also, four water treatments (0, 3, 5, and 8 irrigations) were established for each of four varieties of cotton. The results with two varieties, Stoneville 213 and Delta- pine 16, were averaged for comparison with simulated results.
Comparisons of the results of zero and three irrigations in the 1977 experiments and in the simulation showed that the plants responded differently if water stress occurred after flowering. Despite early water stress, organ initiation proceeded at a normal rate; however, if water stress occurred for the first time after flowering, organ initiation was sharply curtailed. Therefore, the effect of stress on organ initiation before the boll period was modified in the model. The abscission parameters were not further modified.
The simulated versus actual yield data are shown in Figure 2 for both experiments. The results demonstrate the responsiveness of COTCROP to both nitrogen and water variables. In 1977, the eight-irrigation treatments resulted in approximately 55% of the yield produced with the frequent irrigation treatment of 1961. The nitrogen application rates were 100 and 300 pounds per acre for 1977 and 1961 experiments, respectively. In addition, simulated yields compared well with the actual yields for the different water stress treatments. In 1977, the simulated yield for five irriga- tions was the same as that for eight irrigations, which was similar to the observed field response.
The COTCROP model was designed for interfacing with insect models. The age distribution of the fruit is included because of the preference for fruit of different ages by boll weevils (Lloyd et al. 1961; Jones et al. 1975) and Heliothis spp. (Nicholson

COTTON INSECT MANAGEMENT SIMULATION MODEL 445
Simulated yield, bales/acre
5
Í 2 3
Actual yield, bales/acre
Figure 2.—Comparison or simulated and actual yields for 1961 and 1977 irrigation treatments, and the 1978 Panola County, MS, field data (from Brown et al. 1982).
1975). In addition, the crop response to fruit damage depends on the age of fruit damaged.
BOLL WEEVIL COMPONENT MODEL
The boll weevil component model used is the population dynam- ics Cotton and Insect Management-Boll Weevil (CIM-BW) model, which is a modification of the Boll Weevil Simulation (BWSIM) model de- veloped by Jones et al. (1977). For our purposes, BWSIM was

446 L. G. BROWN ET AL
simplified to reduce the computer run time and was restructured to facilitate coupling with the cotton crop model (Brown et al. 1979).
The boll weevil population is closely related to the growth dynamics of the cotton crop as shown in Figure 3. The model is initiated with emergence of overwintering adult weevils into the cotton field. »This may occur when the crop is young and has no fruit (bolls or squares). Since the boll weevil must have cotton fruit for reproduction, no reproduction occurs until the crop be- gins to produce squares. As the female adult encounters fruit, she oviposits"into them. As the larvae develop, the fruit are abscised from the plants. This fruit loss causes a response in the growth and development processes of the plants. Both the development and survival of the larvae depend on temperature and quality of food.
Many of the relationships used in the model to describe feed- ing, oviposit ion, development, and mortality have been reported (Bacheler et al. 1975a, 1975b; Jones 1975; Jones et al. 1975, and Jones et al. 1977). Following is a summary of the structure and processes used in the CIM-BW model.
Model Structure
The state variables in the model consist of vectors of popula- tion densities for cohorts of each life stage (egg, larva, pupa, adult). Thus, age structure of each life form is explicitly con- sidered. The vectors EGG (a), LARV^(a), PüPA^(a), and ADULT (a) represent the densities of boll weevils on day t of age a in days. A discrete time step of one day is used to solve the following pop- ulation dynamics equations for each stage
Nj.(a) = N^_j(a-1) (1-6^) (l-P^(a)) I^ (4]
where
N (a) = the density of one boll weevil life stage on day t, age a;
6 = the probability of insect death on day t;
P (a) = the probability that an insect of age "a" will make a transition to the next stage of development on day t; and
I = the survival of insects on the day of an insecticide application.
Initial and boundary conditions for solving these equations consist of initial population influx and transitions between life stages. Figure 4 is a schematic that shows the subroutines and the calling sequence of the CIM-BW model.
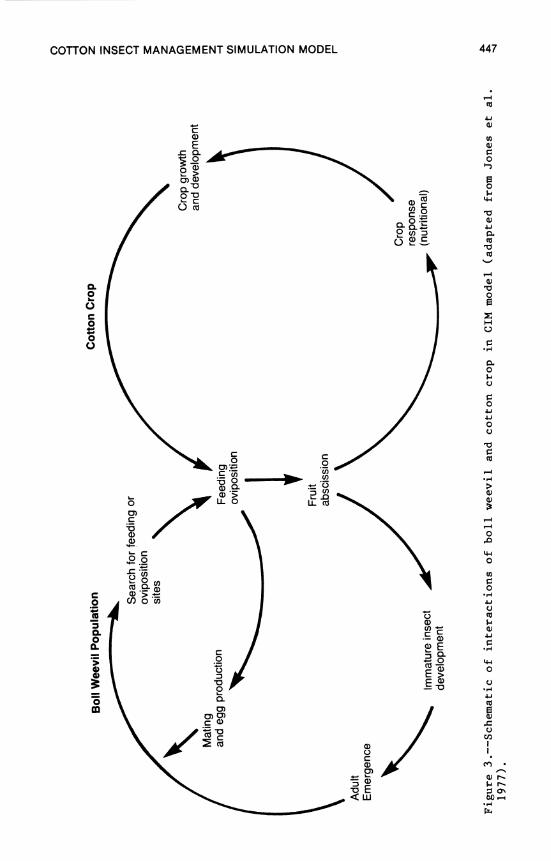
COTTON INSECT MANAGEMENT SIMULATION MODEL 447
no Q) U Cu CO
CO
no O B S M
C
O
o
c o
o o
»TO c CO
o 43
CO
§ O
CO
m
3 ON &0.-H
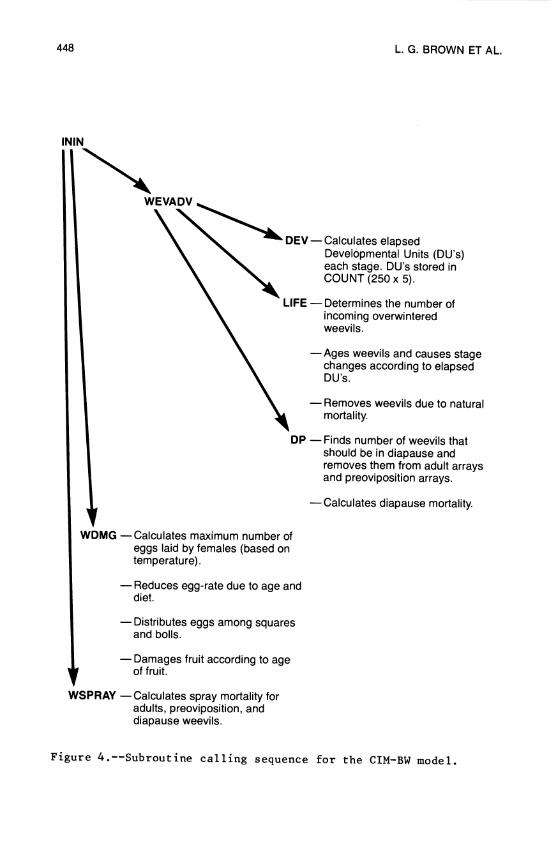
448 L G. BROWN ET AL
ININ
DEV — Calculates elapsed Developmental Units (DU's) each stage. DU's stored in COUNT (250x5).
LIFE — Determines the number of incoming overwintered weevils.
—Ages weevils and causes stage changes according to elapsed DU's.
— Removes weevils due to natural mortality.
DP -Finds number of weevils that should be in diapause and removes them from adult arrays and preoviposition arrays.
-Calculates diapause mortality.
WDMG —Calculates maximum number of eggs laid by females (based on temperature).
— Reduces egg-rate due to age and diet.
— Distributes eggs among squares and bolls.
— Damages fruit according to age f of fruit.
WSPRAY —Calculates spray mortality for adults, preoviposition, and diapause weevils.
Figure 4.—Subroutine calling sequence for the CIM-BW model.

COTTON INSECT MANAGEMENT SIMULATION MODEL 449
Development Times and Variances
Development, described by P (a) in equation 4, is a nonlin- ear function of temperature. It was derived from developmental data for the various stages (egg, larva, and egg to adult) from several sources (see Figure 5). For example, data of Bacheler et al. (1975b) were used to estimate the coefficients for describing the average development time for egg, larva, and pupa; and coeffi- cients for adult longevity were based on data from Isley (1932). The form of the equation is
L((t)) = C^ exp (C^/cl)) for (|)>14.4'C [5]
where
L((1)) = the length of a stage for a given temperature; and C,, C« = the parameters given in Table 1.
Since rate of development is the reciprocal of L((|)), the de- velopmental rate for variable temperatures is integrated through time to calculate developmental units. Transitions from one stage to the next are initiated when the development unit value reaches 0.83 and are completed when it reaches 1.17. Half of the stage transitions occur as developmental units increase from 0.83 to 1.0. P (a) describes the proportion of insects of age a that makes a transition on day t based on a Gaussian distribution (Jones et al. 1977).
Table 1.—Development time parameters (Jones et al. 1977)
Stage n (r^)
Egg Larva Pupa Adult
45.9 39.8 40.1 58.0
0.55 1.80 1.63 3.46
(0.97) (0.98) (0.94)
Fecundity and Feeding
Damage to the cotton crop due to boll weevil feeding and ovi- position is calculated by subroutine WDMG (see Figure 4). It is assumed that damage done by the female weevil during oviposition is affected by the average daily temperature, insect age, and diet. (Boll weevils prefer squares for feeding and oviposition, but young bolls (less than 19 days old) can be substituted if squares are scarce.) Oviposition is reduced 80% when the diet consists solely of small bolls, but there is no reduction if the diet consists only of squares provided the square density is adequate. The fecundity reduction factor due to square availability is given by

450 L G. BROWN ET AL
Temperature, °C
34 ^
30-
26-
i' Temperature, °C
34
30-
> Larva
22- i 26- Í
18- \
22- V
♦ T? XT
\
I 1 1 1 1 0 2 4 6 8
Days
18 -ö ^
I 1 1 1 1 1 1 I 0 4 8 12 16 20 24 28
Days
O Bacheler et al. (1975a) X Isley (1932) > Cole (1970) ^ Hunter and Pierce (1912)
Temperature, °C
34 *«^^
Y Egg to Adult
30 yp^
18- -xo-
1 10
1 15
1 20
1 25
r 30
Days
1 35
1 40 45
Figure 5.—Developmental time for boll weevil eggs, larva, and egg to adult (from Jones 1975).

COTTON INSECT MANAGEMENT SIMULATION MODEL 451
g^ = 0.25 I ?.. + 0.05 I (1-Pi.) [6] ^ i = t-3 ^^ i = t-3 ^^
where
P,. = fraction of potential eggs that are laid in squares.
In the more detailed BWSIM (Jones 1975), selection of sites for oviposit ion and feeding are based on detailed models of search- ing and egg laying or feeding processes. For the CIM-BW model, re- sults from that work were used to generate a family of curves based on densities of bolls and squares. Figure 6 is an example of the type of curves used to partition eggs to squares and bolls, thus defining P-. according to densities of squares and bolls on day i.
It is also assumed that female weevils oviposit one egg per fruit form before moving to another fruit. No discrimination is made against fruit with previous boll weevil damage (Jenkins et al. 1975). Density of new eggs on day t is calculated by
EGG (1) = 0.5g q I ADULT, (j) E.((t)) tj = 1 t J
[7]
where
g = reduction in egg production on day t due to the in- sect diet during the last 4 days;
q = probability that an egg will be viable if it is in an optimum environment; and
E.((|>) = potential number of eggs produced per day by females of age j at temperature (j). J
In the CIM-BW model, the maximum number of eggs laid per day per female is assumed to range from 3 (at an average temperature of IS'^C) to 12 (at 30'*C). After the female is 8 days old, the rate is allowed to decline with age (Jones 1975). The feeding damage by males, preoviposition females, and diapausing weevils is calculated as one fruit per insect every 2 days.
Mortality
In CIM-BW, daily mortality of each boll weevil stage is as- sumed to be constant (0.28, 0.074, 0.028, and 0.001 for egg, larva, pupa, and adult, respectively). Adult mortality due to longevity is included in the transition probabilities. Insecticide-induced mortality for treatment with EPN-methyl parathion was assumed to be 0.90 for adults (i.e., I = 0.10) and 0.0 for all immature stages (i.e., I = 1.0) on days when insecticide was applied (Jones et al. 1977).
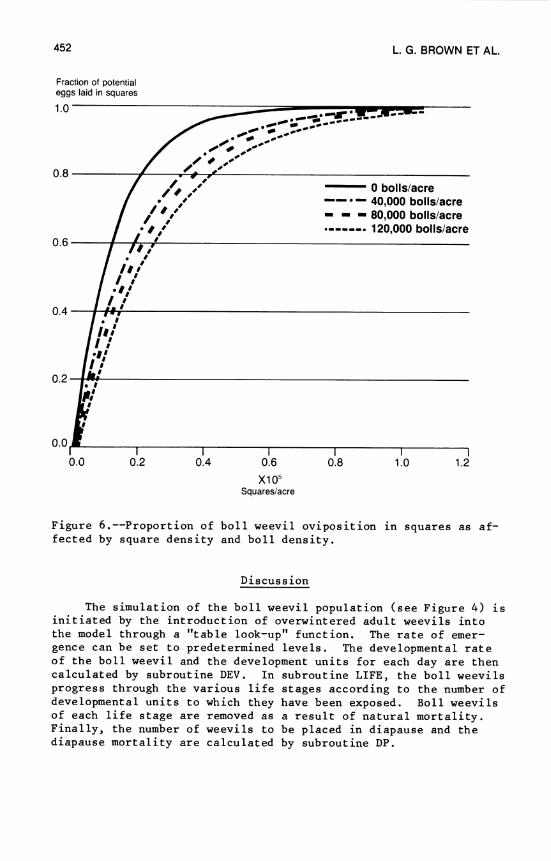
452 L G. BROWN ET AL
Fraction of potential eggs laid in squares
1.0
X10' Squares/acre
Figure 6.—Proportion of boll weevil oviposit ion in squares as af- fected by square density and boll density.
Discussion
The simulation of the boll weevil population (see Figure 4) is initiated by the introduction of overwintered adult weevils into the model through a "table look-up" function. The rate of emer- gence can be set to predetermined levels. The developmental rate of the boll weevil and the development units for each day are then calculated by subroutine DEV. In subroutine LIFE, the boll weevils progress through the various life stages according to the number of developmental units to which they have been exposed. Boll weevils of each life stage are removed as a result of natural mortality. Finally, the number of weevils to be placed in diapause and the diapause mortality are calculated by subroutine DP.

COTTON INSECT MANAGEMENT SIMULATION MODEL 453
Simulated results from the original BWSIM model were compared with experimental results for three North Carolina cotton fields (Jones et al. 1977). The simulated and actual fruit damage due to populations of adult boll weevils is presented in Figure 7a, b, and c.
Also, simulations obtained with the CIM-BW model were compared with those obtained with BWSIM to ensure that the modifications had not significantly changed the results. In this case, CIM-BW was used to simulate boll weevil population dynamics in two producer fields in North Carolina, and the results were compared with simu- lated results for the same two fields from BWSIM (Figure 8a and b). Results were in close agreement though computer time and memory were reduced by factors of 10 and 3, respectively, for CIM-BW. The effects of the detailed oviposit ion and feeding behavior submodels were incorporated by using simulated results to establish curves for fruit selection for the simplified model.
Damaged squares/acre on plants (xlO^)
250
No Insecticide ^\ o Observed Simulated (BWSIM)
T 205 215 225
Time, Julian day number
Figure 7a—Comparison of simulated and observed damaged squares for an assumed initial population of 1,977 weevils per ha, Raleigh, NC, 1973 (Jones et al. 1977).

454 L G. BROWN ET AL
Insecticide Spray Dates
iU i i Ui \ Damaged squares/acre on plants (XIO^)
100
o Observed — Simulated
(BWSIM)
160 I
200 220 Time, Julian day number
Figure 7b.—Comparison of simulated and observed damaged squares in producer's field 1, Rich Square, NC, 1973 (Jones et al. 1977).
Insecticide Spray Dates ^^
11 i li il 11 Damaged squares/acre (XI0^)
75
O Observed •— Simulated
(BWSIM)
200 220 Time, Julian day number
260
Figure 7c.—Comparison of simulated and observed damaged squares in producer's field 2, Rich Square, NC, 1973 (Jones et al. 1977).
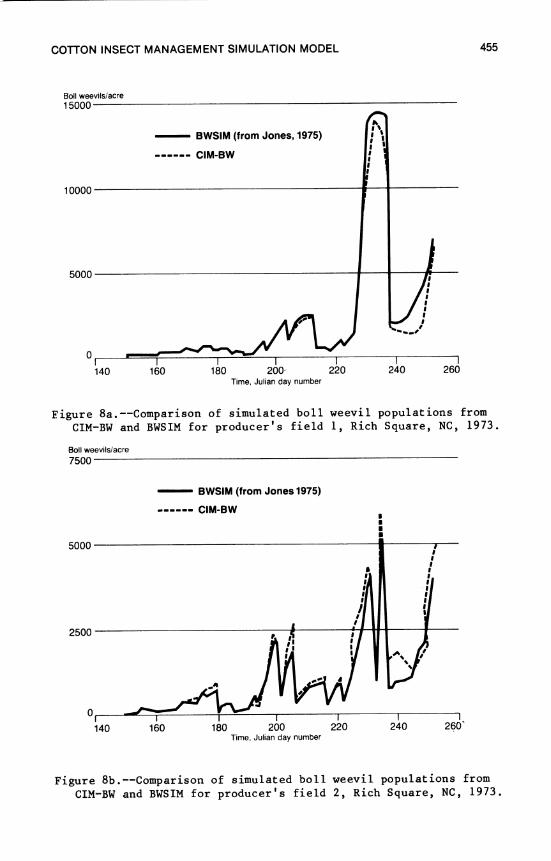
COTTON INSECT MANAGEMENT SIMULATION MODEL 455
Boll weevils/acre 15000
BWSIM (from Jones, 1975)
CIM-BW
10000
5000
180 200' 220 Time, Julian day number
260
Figure 8a.—Comparison of simulated boll weevil populations from CIM-BW and BWSIM for producer's field 1, Rich Square, NC, 1973.
Boll weevils/acre
7500
BWSIM (from Jones 1975)
CIM-BW
5000
2500
140 T 160
1 r 180 200 220
Time, Julian day number 240 260
Figure 8b.—Comparison of simulated boll weevil populations from CIM-BW and BWSIM for producer's field 2, Rich Square, NC, 1973,

456 L. G. BROWN ET AL.
HELIOTHIS SPP. COMPONENT MODEL
"^^^ Helíothís spp. component model CIM-HEL was developed by L. G. Brown and D. B. Hogg (unpublished, Mississippi State Uni- versity) specifically for interfacing with the cotton model COT- CROP and the boll weevil model CIM-BW. The model advances time during the season in one-day increments and updates the status of populations each day. The subroutines of the CIM-HEL component model, their purposes, and the calling sequence are shown in Figure 9.
Since H. zea and E. virescens have different rates of develop- ment and oviposit ion and react differently to insecticide, they are kept separate in the model. Each species is also separated in the model by cohorts. The following eight arrays are used to maintain the number in each cohort: ZEGGS, ZLARV, ZPUPAE, ZADULT, VEGGS VLARV, VPUPAE, and VADULT.
Degree days above 12.6^*0 (Hartstack et al. 1973) are used to calculate physiological time. Hourly temperatures are used to calculate the degree days; that is,
25 Degree days = I Max (0, (i).-12.6)/24 [8]
i = 2 ^
where the <t>^ is hourly temperature. The temperature at midnight the previous day is represented by (|)- .
Since the number of degree days that have accumulated from the start of the simulation is calculated and stored each day as PT(t), calendar age and physiological age of each cohort can be deter- mined. Then as the model moves through the season in daily incre- ments, cohorts age (in degree days) until they make the transition to the next life stage. However, it is necessary to account for the variation in developmental times within a cohort. Therefore, the transition of the members of a cohort to the next life stage is distributed over several days. Each day, the percentage of the in- sects within each cohort that advances to the next life stage is a function of the physiological age of the cohort and the degree days accumulated that day.
Developmental Times
The time required for eggs to hatch is estimated to be 50 de- gree days from data reported by Butler and Hamilton (1976). In fact, the variation reported in early studies has been sufficiently small so that all eggs are assumed to hatch (move to the ZLARV or VLARV array) on the day that 50 degree days have been accumulated.
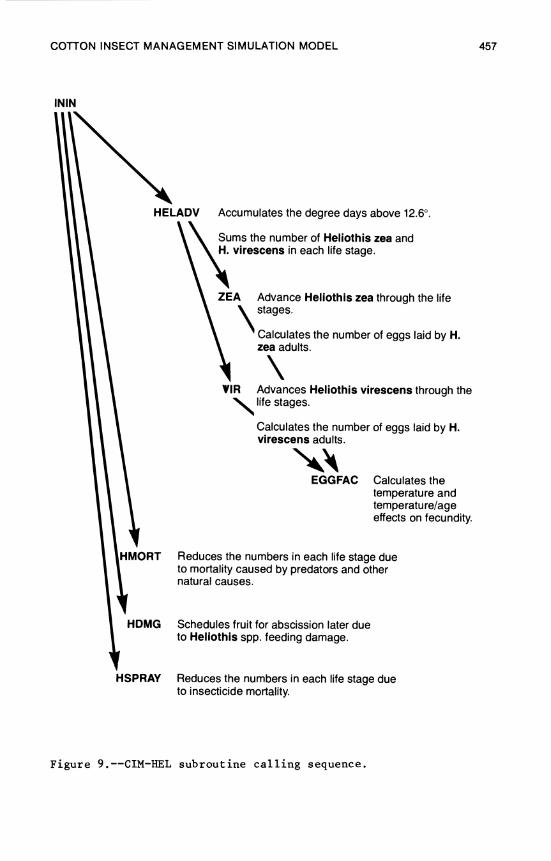
COTTON INSECT MANAGEMENT SIMULATION MODEL 457
ININ
HELADV Accumulates the degree days above 12.6°
Sums the number of Heliothis zea and H. virescens in each life stage.
ZEA Advance Heliothis zea through the life stages.
\ Calculates the number of eggs laid by H. zea adults.
\ VIR Advances Heliothis virescens through the
'V^ life stages.
Calculates the number of eggs laid by H. virescens adults. x\
EGGFAC Calculates the temperature and temperature/age effects on fecundity.
|HMORT Reduces the numbers in each life stage due to mortality caused by predators and other natural causes.
HDMG Schedules fruit for abscission later due to Heliothis spp. feeding damage.
HSPRAY Reduces the numbers in each life stage due to insecticide mortality.
Figure 9.—CIM-HEL subroutine calling sequence.

458 L G. BROWN ET AL
Thus, the number of eggs hatching on day t is calculated from
t-1 ZLARV(t) = I ZEGGS(i) ô^(i,t) [9]
i = 1 ^ t-1
and VLARV(t) = I VEGGS(i) 5^(i,t) [10] i = 1 ^
where
to.O if T(i,t)< 50 degree days ÖE(i,t) =
(l.O ifT(i,t)>^50 degree days and
T(i,t) = elapsed degree days from day i (the day the co- horts were established) to the current day t.
Thus T(i,t) represents physiological age of cohorts established on day i, that is, PT(t)-PT(i).
Developmental times assumed for the larvae and pupae in CIM- HEL are based on field data collected by Hogg and Calderón (1981). They reported that the developmental time for the larval stage of Ü* ^^^ ^^s significantly longer than for that of _H. virescens. They also found that developmental times of female pupae of both species were shorter than those of males. Since the developmental rates of the females control the rates of population growth (Hogg and Gutierrez 1981), they are used in the model rather than de- velopmental rates for males. The developmental times for H. zea and JH. virescens pupae are separated in the model (though the means are not significantly different) because JI. virescens took slightly longer to develop.
Developmental times for Heliothis spp. larvae are shown in Figure 10. The number of larvae pupating on day t is calculated from
t-1 ZPüPAE(t) = I ZLARV(i)ô (i,t) [11]
i = 1 ^
t-1 and VPUPAE(t) = I VLARV(i)ô (i,t) [12]
i = 1 ^
where
ô^(i,t) = (P^(i,t) - P^(i,t-1))/ (1.0 - P^(i,t-1)) and
P_(i,t) = fraction of larvae that will have pupated by physiological age T(i,t).

COTTON INSECT MANAGEMENT SIMULATION MODEL 459
Fraction of larvae that have pupated— PL(T)
1.0
190 210 230 T
250 270 Physiological age—r
330
Figure 10.—Cumulative developmental times of Heliothis spp. larvae (Hogg and Calderón 1981).
When larvae are moved into pupal arrays, they are removed from the larval arrays.
Developmental times for Heliothis spp. pupae are shown in Fig- ure 11. The number of adults emerging on day t is calculated from
t-1 ZADULT(t) = I ZPUPAE(i) ô (i,t)
i = 1
[13]
t-1 and VADULT(t) = I VPUPAE(i) <S (i,t)
i = 1
[14]
where
Ô (i,t) = (P (i,t)-P (i,t-l)/(1.0-P (i,t-l)) and p P P P
P (i,t) = fraction of pupae that will have emerged by physiological age T(i,t).

460 L G. BROWN ET AL
Fraction of pupea that have emerged —Pp (r)
1.0
210 230 Physiological age—r
250 270
Figure 11.—Cumulative developmental times of Heliothis spp. pupae (Hogg and Calderón 1981).
When pupae are moved into adult arrays, they are removed from the pupal arrays.
Fecundity
In CIM-HEL, fecundity is a function of temperature and adult age. The functions that reduce fecundity due to temperature and temperature/age effects are from MOTHZV-2 (Hartstack et al.

COTTON INSECT MANAGEMENT SIMULATION MODEL 461
1976). Thus, total eggs laid by the population on day t is calcu- lated by:
t-1 EGGS(t) = 0.5 I ADULTS(i) PI P2 EP [15]
where
i = 1
EP = potential number of eggs laid per female per day (400 for H. zea; 300 for ii. virescens);
Pi = reductTon in fecundity due to temperature (Hartstack et al. 1976); and
P2 = reduction in fecundity due to temperature/age effect (Hartstack et al. 1976).
Mortality
In the model, mortality can be caused by insecticide, by pred- ators, and by other natural causes. The mathematical relationships for the mortality of eggs and larvae were developed from field data collected by D. B. Hogg in 1978 and 1979 (unpublished data). The co- efficients for CIM-HEL were adjusted after the results of a simula- tion were compared with Hogg's data and with field data collected by G. L. Andrews (unpublished data).
The proportion of eggs that are destroyed daily by predators (PME) is thus given by
PME = (number of predators) 1.325x10 . [16]
The proportion of larvae within each cohort that are destroyed daily by predators (PML) varies with the age of the cohort as
PML = PME EXP(-1.011 AGE ^'^^^) [17]
where
AGE = age of the cohort in degree days.
The proportion of the larvae within each cohort that dies as a re- sult of other natural causes (XML) is given by
XML= 1.0-0.9915(^^(^>-^^^'-^^^ [18]

462 L G. BROWN ET AL
The mathematical relationships for the survival of adults were developed from data for caged adults collected by D. B. Hogg. The adult survival functions are:
XMAZ = 1.0-EXP((AGEY/118.54)^-^^-(AGE/118.54)^-^^) [19] for ji. zea and
XMAV = l,0-EXP((AGEY/134.93)^-^^-(AGE/134.93)^-^^) [20] for jl. virescens
where
AGEY = age in degree days yesterday (that is, (i,t-l)); XMAZ = proportion of H. zea adult cohorts surviving day t;
and XMAV = proportion of li. virescens adult cohorts surviving
day t.
Figure 12 illustrates these survival functions.
Proportion surviving
0 20 40 60 80 100 120 140 160 180 200 Age in degree days
Figure 12.—Survival of H. zea and H. virescens.
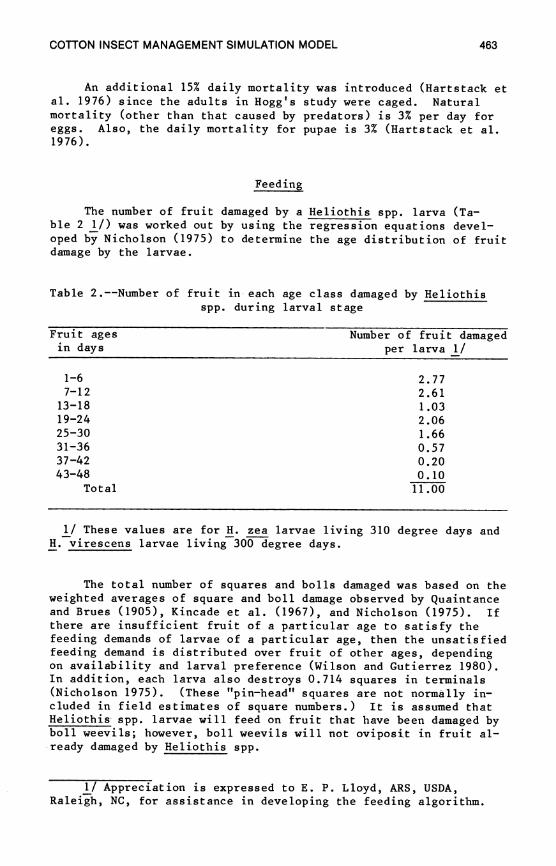
COTTON INSECT MANAGEMENT SIMULATION MODEL 463
An additional 15% daily mortality was introduced (Hartstack et al. 1976) since the adults in Hogg's study were caged. Natural mortality (other than that caused by predators) is 3% per day for eggs. Also, the daily mortality for pupae is 3% (Hartstack et al. 1976).
Feeding
The number of fruit damaged by a Heliothis spp. larva (Ta- ble 2 _1/) was worked out by using the regression equations devel- oped by Nicholson (1975) to determine the age distribution of fruit damage by the larvae.
Table 2.—Number of fruit in each age class damaged by Heliothis spp. during larval stage
Fruit ages Number of fruit damaged in days per larva _1_/
1-6 2.77 7-12 2.61
13-18 1.03 19-24 2.06 25-30 1.66 31-36 0.57 37-42 0.20 43-48 0.10
Total 11.00
\J These values are for H. zea larvae living 310 degree days and li. virescens larvae living 300 degree days.
The total number of squares and bolls damaged was based on the weighted averages of square and boll damage observed by Quaintance and Brues (1905), Kincade et al. (1967), and Nicholson (1975). If there are insufficient fruit of a particular age to satisfy the feeding demands of larvae of a particular age, then the unsatisfied feeding demand is distributed over fruit of other ages, depending on availability and larval preference (Wilson and Gutierrez 1980). In addition, each larva also destroys 0.714 squares in terminals (Nicholson 1975). (These "pin-head" squares are not normally in- cluded in field estimates of square numbers.) It is assumed that Heliothis spp. larvae will feed on fruit that have been damaged by boll weevils; however, boll weevils will not oviposit in fruit al- ready damaged by Heliothis spp.
IJ Appreciation is expressed to E. P. Lloyd, ARS, USDA, Raleigh, NC, for assistance in developing the feeding algorithm.

464 L G. BROWN ET AL.
CIM MODEL
Integration of Component Models
The objective in developing the CIM model was to provide a method of evaluating the effect of insect pest control strategies on yield, insecticide usage, and cost of insect management. As noted, it was formed by using three component models, COTCROP, CIM- HEL, and CIM-BW.
In the CIM model, the interaction, between the cotton crop and the insect pests occurs through the fruit. The crop damage done by the insect pests is calculated each day and transferred to the crop component model. Also, status of the fruit is updated daily and transferred to the component models of the two insect pests.
Insect pest management strategies in the composite CIM model are concerned largely with insecticide applications. These appli- cations are performed either on a predetermined fixed schedule or in response to the percentage of square damage or insect pest den- sities or both. The interval between simulated scouting reports of damage and density can also be varied to simulate different strate- gies. The insecticide used in the simulations, EPN-methyl para- thion, differs in degree of effectiveness against Heliothis spp., depending on the temperature and the age and species of the insect. The insecticide-induced mortality factors (Tables 3 and 4) are from laboratory data collected by McDaniel (1976). Each insecticide ap- plication affects Heliothis spp. for up to 3 days. The same appli- cation kills 90% of the adult boll weevils on the day of the appli- cation. For boll weevils it is assumed that the insecticide has no residual effect or temperature dependency.
The data required as input to the CIM model are the following: (1) Daily weather information
(a) Maximum and minimum temperatures (b) Rainfall (c) Solar radiation (d) Pan evaporation
(2) Agronomic data (a) Day of crop emergence (b) Day of harvest (c) Soil parameters - sample soil parameters that must be
specified by the user for the particular soil type considered are shown in Table 5
(d) Day and amount of nitrogen fertilizer applications (3) Insect populations
(a) Initial Heliothis spp. population (b) Initial boll weevil population (c) Predator population for the year
(4) Insect control strategies (a) Boll weevil (b) Heliothis spp.

COTTON INSECT MANAGEMENT SIMULATION MODEL 465
Table 3.—Daily kills of instars of Heliothis spp. larvae due to insecticide 1/
Da ily fraction of each age group kill ed^/ 1st instar 2nd instar 3rd instar
Day TJ H. zea H. vir- escens H. zea
H. vir- escens H. zea
H. vir- escens
0 1 2
0.95 0.32 0.16
0.96 0.48 0.38
0.90 0.59 0.25
0.9: 0.36 0.18
0.70 0.41 0.18
0.72 0.39 0.13
Daily fraction of each age group killed _3/
4th instar 5th instar 6th instar H. vir- H. vir- H. vir-
H. zea escens H. zea escens H. zea escens
0.51 0.46 0.25 0.21 0.06 0.01 0.32 0.33 0.16 0.17 0.05 0.04 0.16 0.20 0.13 0.06 0.06 0.03
]J From McDaniel 1976. Ij Days after insecticide application. _3/ These values are reduced by 30% in the insecticide mortality
calculations in the model to simulate inefficiencies under actual field conditions.
The output from the CIM model includes a daily record of crop status and daily levels of insects in each life stage. At the end of the growing season, a summary report (Table 6) is produced that gives the value of some of the initial parameters as well as final yield, number and cost of insecticide applications, and monetary returns above costs of insect control.
Table 4.—Temperature effect on mortality of Heliothis spp. larvae exposed to insecticide J_/
% effectiveness relative to effectiveness at 25°C
Temperature (*C) H. zea H. virescens
20 95.4 86.1 25 100.0 100.0 30 105.0 103.0
1/ From McDaniel 1976.

466 L G. BROWN ET AL
Table 5.—Estimated soil parameters for a hypothetical silty loam soil
COTROP soil parameters Description
Texture
THETAS THETAR THETFC
HB ETA
HYDROC XN
U COEFF
PORGAN
QO
RDEP SOILNI
Estimated value
12% sand 75% silt 13% clay
3 3 Saturated water content (cm per cm ) Residual water content (cm per cm ) Field capacity water content (cm per cm ) Bubbling pressure (cm) Brooks & Corey moisture tension constant Hydraulic conductivity (cm per day) Runoff curve number for hydrologie soil
cover complexes Cumulative evaporation (mm) Slope of cumulative evaporation curve
(mm per day * ) Fraction of organic matter content in
the soil Initial drainage rate from bottom of
profile at saturated condition (cm per day) Rooting depth (cm) Residual nitrogen (g per m )
0.3964 0.1019 0.2938
60.0 2.8080 6.0
65.0
9.0 4.04
0.01
4.2
90.0 12.61
Sensitivity Analysis
A sensitivity analysis was undertaken to demonstrate the re- sponsiveness of the CIM model and to show how a simulation might be useful in evaluating cotton insect management strategies. This study is analogous to a field experiment in which certain variables are held constant and others are varied over a range of interest.
The variables selected for study were densities of Heliothis spp., boll weevils, and predators. Two insect control strategies were used for each pest density. Strategy 1 (Sl) consisted of us- ing no insecticide. Strategy 2 (S2) consisted of scouting twice a week (every 4 days in the model) starting on June 1 and continuing through September 15 with insecticide being applied according to the following thresholds:
(1) Boll weevil - treat when oviposit ion-punctured squares reach 5% (does not include feeding punctures).
(2) Heliothis spp. - treat before first bloom whem damaged squares reach 25%; after first bloom and until first treatment, treat at 5% square damage; after first treat- ment, treat when 4% of the terminals contain larvae and there are eggs present (in the CIM model, 4% terminal infestation is estimated to be 1500 larvae per acre).

COTTON INSECT MANAGEMENT SIMULATION MODEL 467
Table 6.—Sample CIM model output
COTTON INSECT MANAGEMENT (CIM) MODEL
8/13/80
WEATHER DATA USED - STONEVILLE 1972 DAY OF EMERGENCE - 125 PLANT POPULATION - 40000 PLANTS/ACRE ROOTING DEPTH OF SOIL - 35.4 IN RESIDUAL NITROGEN ON DAY OF EMERGENCE - 112.5 LB/ACRE RESIDUAL WATER ON DAY OF EMERGENCE - .36 IN**3 NITROGEN FERTILIZATION
TOTAL - IRRIGATION
DAY LB/ACRE 125 80.0 145 30.0
110.0 DAY INCHES 0 .0 0 .0 TOTAL
TOTAL RAINFALL - 16.8 IN TOTAL SOLAR RADIATION - 85090.00 LANGLEYS TOTAL PHYSIOLOGICAL DAYS - 164.5 DAY OF HARVEST - 300 REMARKS - INITIALIZE WITH EGGS
GROSS CROP VALUE YIELD - 889.8 LB/ACRE
GROSS LINT VALUE OF $.65 COTTON - GROSS SEED VALUE OF $.06/LB SEED - GROSS CROP VALUE (LINT+SEED VALUE) -
INSECT CONTROL COSTS
NUMBER OF INSECTICIDE APPLICATIONS
578.35 $/ACRE 82.75 $/ACRE
661.10 $/ACRE
COST OF EACH INSECTICIDE APPLICATION - 4.96 $/ACRE TOTAL COST OF INSECTICIDE APPLICATIONS - 29.76 $/ACRE COST OF TWICE WEEKLY SCOUTING - 3.75 $/ACRE TOTAL COST OF INSECT CONTROL - 33.51 $/ACRE
RETURNS ABOVE COST OF INSECT CONTROL
PRICE $/LB .50 .55 .60 .65 .70 .75 .80 RETURNS $/ACRE 494.12 538.61 583.10 627.59 672.08 716.56 761.05

468 I- G. BROWN ET AL.
The soil and crop variables were held constant. The soil is described in Table 5. The plant population was always 40,000 plants per acre, the date of crop emergence was May 5, and the har- vest date was October 27. Nitrogen was applied to the crop at planting in the amount of 80 pounds per acre and as side-dressing in the amount of 30 pounds per acre on May 25. Two years of weath- er data from Stoneville, MS, (1967 and 1972) were used to demon- strate the impact of weather variability on simulated results. Simulations were also performed for both years without insect dam- age to provide a basis for comparison.
Three populations (low, medium, and high) of predators, Helio- this spp., and boll weevils were used in the analysis. These popu- lations were identified on the basis of consultations with J. W. Smith and E. P. Lloyd (personal communication).
The seasonal progression in the three predator populations is shown in Figure 13. The low, medium, and high populations peaked at 7,400, 11,800 and 15,700 predators per acre, respectively. As mentioned earlier, the predator population is treated as an exog- enous variable. However, this population is reduced by insecticide applications.
The simulated Heliothis spp. populations with the 1972 weather data and the medium predator population are shown in Figure 14a. Heliothis spp. larvae of the third generation (September 1 to Sep- tember 20) reached maximums of 8,500, 16,000, and 63,700 per acre for the low, medium, and high populations, respectively. (To initi- ate the CIM-HEL model, we enter adults in the model early in the year to mimic adults entering a field. Then, to simulate the dif- ferent populations, we vary the number and timing of the adults en- tered in the CIM-HEL model.) These adults were assumed to be 60% Heliothis virescens and 40% Heliothis zea. The number and timing of the adults entered in the CIM-HEL model were identical for 1967 and 1972; however, the resulting populations were different due to differences in weather.
The simulated boll weevil populations with the 1972 weather data and with no Heliothis spp. or predators present are shown in Figure 15a. The low, medium, and high populations reached maximums of 19,800, 33,600, and 60,900 adults per acre, respectively. (These populations are affected by the diet of the adults and num- ber of oviposition sites available. Since the number of Heliothis spp. present alters the crop, the simulated boll weevil populations would be different if Heliothis spp. were present.) The number of adults and the time they entered the field were identical for 1967 and 1972; however, the resulting populations were different because of differences in weather and crop development.
The simulated yield and number of insecticide applications for each scenario in each year are shown in Table 7. The simulated yields with 1967 and 1972 weather with no insects were 501 and 971 pounds, respectively. The rainfall during the growing season was

COTTON INSECT MANAGEMENT SIMULATION MODEL 469
-I
(0 H
^ ÍH O- <ü
col
rol
-O -O
_ H o -H ÏM a
u cs|
OvOCMmO^O^OCOr^O^Ol/^O^mc^lOCOOOOvOCO
.—ivocsmcs«—tvocsovofooovocvicsi—tONi—tfoeso
O^O^00vOO^a^CJ^00P>■»J^a^O^00^^00O^O^O^O^^««.00
oooocsmf>ooo<t-<tvoooor^mcsovo
or^eN<j-p^cscNirorOr-imooi^ooeN>d-<fcs<l-v00o
'-lCSOO^C^lmmo<J"O^^C^I^OCNlO>d•CT^^/^<î■OOr^ 00^0^«d■0^0000^^<í■0^^»>^CSP^O^G^^^OOO^CSfO Ln<t<t>^<l■<^•<í•-*<í■c^J<t<í■<^<í■>^«^<^<t■<i■*í<í
OOOOfOmr^OOOCSrOvOOOOr^m^OvD
KÍ S K hJ S W
a S S S S S
,j s PC iJ s K
^SK^SÄSS
ssssssss
5
-lO)
►J C PQ o
^^ -H ->^ CM|JJ cn|
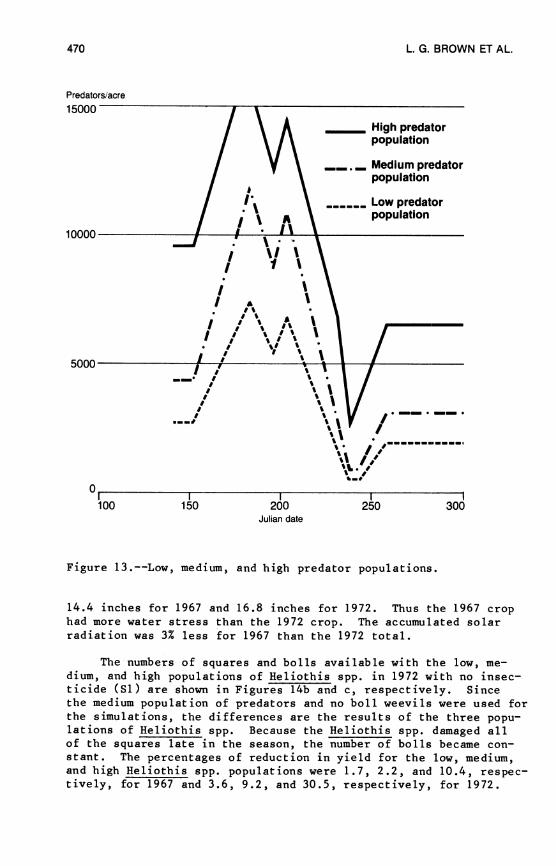
470 L G. BROWN ET AL
Predators/acre
15000
10000
5000
250 100 150 200 Julian date
300
Figure 13.—Low, medium, and high predator populations.
14.4 inches for 1967 and 16.8 inches for 1972. Thus the 1967 crop had more water stress than the 1972 crop. The accumulated solar radiation was 3% less for 1967 than the 1972 total.
The numbers of squares and bolls available with the low, me- dium, and high populations of Heliothis spp. in 1972 with no insec- ticide (Sl) are shown in Figures 14b and c, respectively. Since the medium population of predators and no boll weevils were used for the simulations, the differences are the results of the three popu- lations of Heliothis spp. Because the Heliothis spp. damaged all of the squares late in the season, the number of bolls became con- stant. The percentages of reduction in yield for the low, medium, and high Heliothis spp. populations were 1.7, 2.2, and 10.4, respec- tively, for 1967 and 3.6, 9.2, and 30.5, respectively, for 1972.
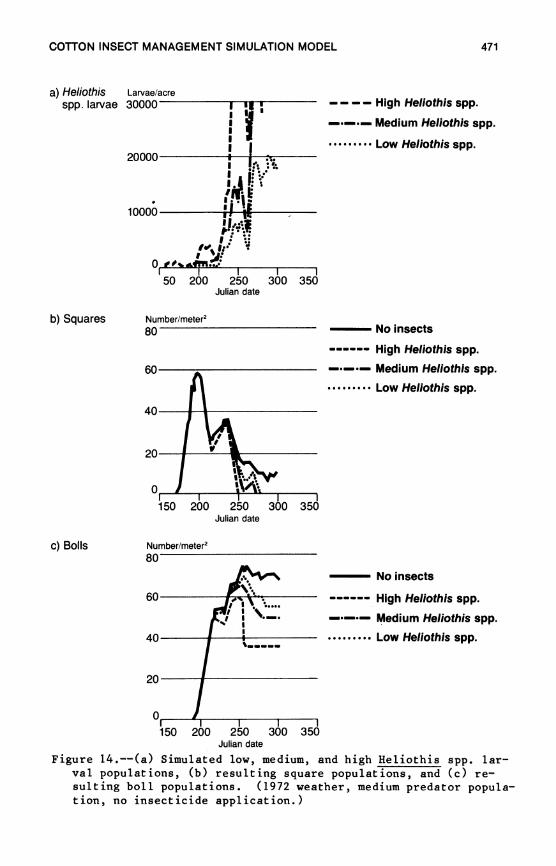
COTTON INSECT MANAGEMENT SIMULATION MODEL 471
a) HeliOthis Larvae/acre spp. larvae 30000
b) Squares
c) Bolls
20000-
I II ■
-^ 1—^
High HeliOthis spp.
Medium HeliOthis spp.
Low HeliOthis spp.
10000- íf4i- I
/< I:
¡ff^
\ 50 200 250 300 350
Julian date
Number/meter^ 80 No insects
High HeliOthis spp.
Medium Heliothis spp.
Low HeliOthis spp.
200 250 Julian date
350
Number/meter^ 80
No insects
High HeliOthis spp.
Medium Heliothis spp.
Low HeliOthis spp.
250 300 350 Julian date
Figure 14.—(a) Simulated low, medium, and high Heliothis spp. lar- val populations, (b) resulting square populations, and (c) re- sulting boll populations. (1972 weather, medium predator popula- tion, no insecticide application.)

472 L. G. BROWN ET AL.
For each Heliothis spp. population, the percentage of reduction in yield was much less in 1967 than in 1972.
In the simulations with insect control strategy S2, there were five applications with the medium H. spp. population and seven ap- plications with the heavy K. spp. population for 1967 and 1972. For the low Heliothis spp. population, three applications were nec- essary in 1967 and two in 1972. The percentages of reduction in yield with insect control strategy S2 were approximately the same for both years for all three Heliothis spp. populations.
The effects of the boll weevil populations on the numbers of squares and bolls are shown in Figures 15b and c. With insect con- trol strategy SI, the percentages of reduction in yield for the low, medium, and high boll weevil populations were 6.3, 11.3, and 40.1, respectively, for 1967 and 15.3, 21.7, and 46.9 for 1972. Also, with insect control strategy S2, the number of insecticide applications was less in 1967 for the low and medium populations of boll weevils. For the high population, the number of insecticide applications was equal in 1967 and 1972.
The effects of the predator populations on the medium popula- tion of Heliothis spp. in 1972 with insect control strategy SI are shown in Figure 16a. With the low, medium, and high predator popu- lations, the medium Heliothis spp. population reached maximums of 33,300, 16,000, and 4,700 larva per acre, respectively, during the third generation (September 1 to September 20). The effects of these Heliothis spp. populations on the squares and bolls are shown in Figures 16b and c, respectively. With insect control strategy SI, the percentages of reduction in yield for the low, medium, and high predator populations were 5.8, 2.2, and 1.2, respectively, for 1967 and 23.5, 9.2, and 1.0 for 1972.
With S2, the number of insecticide applications was the same for 1967 and 1972 for both the low and the medium predator popula- tions (seven and five, respectively). With the high predator popu- lation, one insecticide application was made for 1967 and two for 1972. With S2, the percentage of reduction in yield was less for 1972 than for 1967.
Simulations were also performed with all three insect popula- tions at the medium level. In 1967, the reductions in yield were 14.6% with insect control Si and 12.8% with insect control S2. In 1972, the reductions were 25.6 and 8.3 for SI and S2, respectively.
This sensitivity analysis illustrates how the simulated yield of the CIM model is affected by different levels of three insect populations, when an insecticide is or is not used. The results also indicate the usefulness of the CIM model in evaluating insect pest management strategies. Because the simulated experiments can be run under controlled conditions, the factor of interest can be isolated, and the experiment can be replicated over several years.
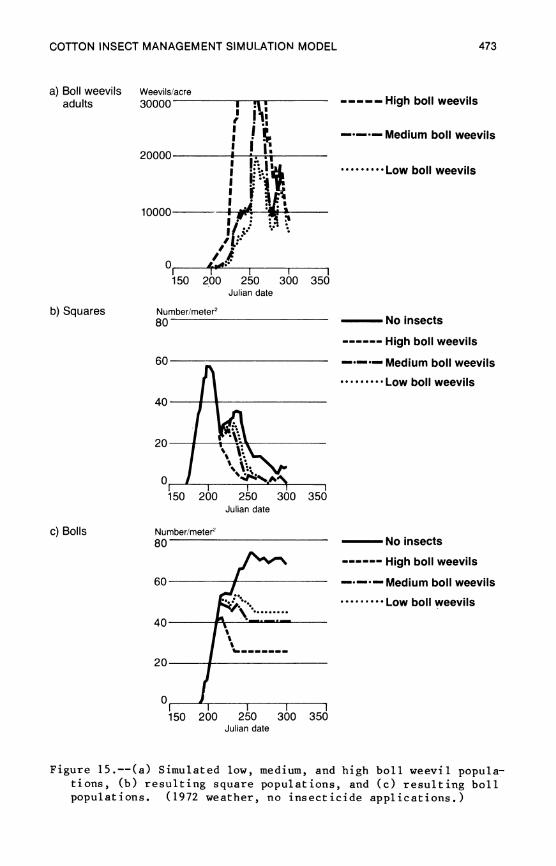
COTTON INSECT MANAGEMENT SIMULATION MODEL 473
a) Boll weevils adults
b) Squares
c) Bolls
Weevils/acre
30000
20000-
I i I I
""■fT
! \ 1rrr
-----High boll weevils
——— Medium boll weevils
10000- ! J If • 'Low boll weevils
W 150 200 250 300
Julian date 350
Number/meter^ 80 • No insects
' High boll weevils
' Medium boll weevils
' Low boll weevils
I \ r 150 200 250 300 350
Julian date
Number/meter^
80 • No insects
High boll weevils
■ Medium boll weevils
* Low boll weevils
n I r 150 200 250 300 350
Julian date
Figure 15.—(a) Simulated low, medium, and high boll weevil popula- tions, (b) resulting square populations, and (c) resulting boll populations. (1972 weather, no insecticide applications.)

474 L G. BROWN ET AL
a) Heliothis Larvae/acre spp. larvae 30000
20000
-■ •sai
; •• •• l'-'i :• iJ
10000-
I— 150 200
Mí
1 1— 250 300
Julian date
-- High predators
— Medium predators
• • • Low predators
350
b) Squares Number/meter^ 80 No insects
High predators
Medium predators
Low predators
150 200 250 300 350 Julian date
c) Bolls Number/meter^ 80 No insects
High predators
Medium predators
Low predators
150 200 n 1
250 300 350 Julian date
Figure 16.—Simulated effects of low, medium, high predator popu- lations on (a) medium Heliothis spp. larval population, (b) squares, and (c) bolls"! (No insecticide application; (1972
weather.)

COTTON INSECT MANAGEMENT SIMULATION MODEL 475
In addition, once the model has been developed, the time and cost of each experiment is small compared with that of a field experi- ment. The simulation of one season with the CIM model requires ap- proximately 2 minutes on the UNIVAC 1100/80 at Mississippi State University.

476 L. G. BROWN ET AL.
LITERATURE CITED
Bacheler, J. S., J. W. Jones, and J. R. Bradley. 1975a. The effect of temperature on development and mortality of
boll weevil immature stages. Environmental Entomology 4:808- 810.
J. W. Jones, J. R. Bradley, Jr., and H. D. Bowen. 1975b. Influence of temperature on abscission of cotton squares
infested with boll weevil eggs. Journal of Economic Entomol- ogy 68:298-300.
Baker, D. N., J. D. Hesketh, and W. G. Duncan. 1972. Simulation of growth and yield in cotton. Gross photosyn-
thesis, respiration and growth. Crop Science 12:432-435.
Brown, L. G., J. W. Jones, and F. A. Harris 1976. A simulation study of insect pest management alternatives
by integration of a Heliothis spp. model and a cotton crop model. American Society of Agricultural Engineers, Paper no. 76-5025, St. Joseph, MI.
J. W. Jones, J. D. Hesketh, J. D. Hartsog, F. D. Whis- ler, and F. A. Harris.
1982. COTCROP: Computer simulation of a cotton production sys- tem. Users manual. Mississippi Agriculture and Forestry Ex- periment Station, Mississippi State, MS.
R. W. McClendon, and J. W. Jones. 1979. Computer simulation of the interaction between the cotton
crop and insect pes^-s. Transactions of the ASAE 22:771-774.
Bruce, R. R., and J. M. Romkens. 1965. Fruiting and growth characteristics of cotton in relation
to soil moisture tension. Agronomy Journal 57:135-140.
Butler, G. D., Jr., and A. G. Hamilton. 1976. Developmental time of Heliothis virescens in relation to
constant temperature. Environmental Entomology 5:759-760.
Cole, C. L. 1970. Influence of certain seasonal changes on the life history
and diapause of the boll weevil. Anthonomus grandis Boheman. Ph.D. dissertation. Texas A & M University, College Station, TX.
Duncan, W. G. 1972. SIMCOT. A simulation of cotton growth and yield. Pro-
ceedings, Workshop Modeling Tree Growth. C. M. Murphy and J. D. Hesketh, editors Oak Ridge National Laboratory, Oak Ridge, TN.

COTTON INSECT MANAGEMENT SIMULATION MODEL 477
Gardner, W. R. 1965. Movement of nitrogen in soil, Ln Soil Nitrogen. W. V. Bartholomew, and F. W. Clark, editors. American Society of Agronomy, Madison, WI, 557 p.
Gutierrez, A. P., T. F. Leigh, Y. Wang, and R. D. Cave. 1977. An analysis of cotton production in California: Lygus hesperus injury - an evaluation. Canadian Entomologist 109: 1375-1386.
Harris, F. A. 1972. Resistance to methyl parathion and toxaphene-DDT in boll- worm and tobacco budworm from cotton in Mississippi. Journal of Economic Entomology 65:1193-1194.
Hartstack, A. W., Jr., J. P. Hollingsworth, R. L. Ridgway, and J. R. Coppedge.
1973. A population dynamics study of the bollworm and the tobac- co budworm with light trays. Environmental Entomology 2:244- 252.
J. A. Witz, J. P. Hollingsworth, R. L. Ridgway, and J. D. Lopez.
1976. MOTHZV-2: a computer simulation of Heliothis zea and Heliothis virescens population dynamics. Users manual. U.S. Department of Agriculture, ARS-S-127, 55 p.
Hesketh, J. D., D. N. Baker, and W. G. Duncan. 1972a. Simulation of growth and yield; respiration and the car-
bon balance. Crop Science 11:394-398.
D. N. Baker, and W. G. Duncan. 1972b. Simulation of growth and yield in cotton. II. Environ- mental control of morphogenesis. Crop Science 12:436-439.
Hogg, D. B., and M. Calderón. 1981. Field developmental times of Heliothis zea and H. vires-
cens (Lepidoptera: Noctuidae) larvae and pupae in cotton. En- vironmental Entomology 10:177-179.
and A. P. Gutierrez. 1981. A model of the flight phenology of the beet armyworm
(Lepidoptera: Noctuidae) in central California. Hilgardia 48:1-36.
Hunter, W. D., and W. D. Pierce. 1912. The Mexican boll weevil: a summary of the investigations
of this insect up to December 31, 1911. U.S. Senate Document 305, 188 p.

478 L. G. EÎROWN ET AL
Isley, D. 1932. Abundance of the boll weevil in relation to summer weather
and food. Arkansas Agriculture Experiment Station, Bulletin No. 271.
Jackson, R. D., and F. D. Whisler. 1970. Equations for approximating the vertical nonsteady-state
drainage of soil columns. Soil Science Society of America, Proceedings 34:715-718.
Jenkins, J. N., W. L. Parrott, and J. W. Jones. 1975. Boll weevil oviposition behavior: multiple punctured
squares. Environmental Entomology 4:861-867.
Jones, J. W. 1975. A simulation model of boll weevil population dynamics as
influenced by the cotton crop status. Ph.D. disseration, North Carolina State University, Raleigh, NC.
H. D. Bowen, R. E. Stinner, J. R. Bradley, Jr., and J. S. Bacheler.
1977. Simulation of boll weevil population as influenced by weather, crop status, and management practices. Transactions of the ASAE 20:121-125, 131.
H. D. Bowen, R. E. Stinner, J. R. Bradley, Jr., R. S. Sowell, and J. S. Bacheler.
1975. Female boll weevil oviposition and feeding process: a simulation model. Environmental Entomology 4:815-821.
L. G. Brown, and J. D. Hesketh. 1980. COTCROP: a computer simulation model for cotton growth
and yield. In Predicting Photosynthesis for Ecosystem Models, Vol. 2, p. 209-241. CRC Press, Boca Raton, FL.
J. D. Hesketh, E. J. Kamprath, and H. D. Bowen. 1974. Development of a nitrogen balance for cotton growth
models: a first approximation. Crop Science 14:541-546.
F. D. Whisler, and J. D. Hartsog. 1976. Soil water model for crop level simulations. In Proceed-
ings, Beltwide Cotton Production Research Conference, p. 116-118.
Kincade, R. T., M. L. Laster, and J. R. Brazzel. 1967. Damage to cotton by the tobacco budworm. Journal of Eco- nomic Entomology 60:1163-1164.
Lloyd, E. P., J. L. McMeans, and M. E. Merkl. 1961. Preferred feeding and egg laying sites of the boll weevil
and the effect of weevil damage on the cotton plant. Journal of Economic Entomology 54:979-984.

COTTON INSECT MANAGEMENT SIMULATION MODEL 479
McClendon, R. W., L. G. Brown, J. W. Jones, J. D. Hesketh, J. Hartsog, F. A. Harris, and D. W. Parvin.
1977, Modeling crop-insect pest ecosystems for studying con- flicting alternatives caused by species interactions. Iri Pro- ceedings, International Federation of Automatic Control~Sympo- sium on Cotton Mechanisms in Bio and Ecosystems, p. 121-131.
McDaniel, S. G. 1976. Rate, temperature, Heliothis spp. and developmental stage
effect on methyl parathion efficacy in cotton. M.S. thesis, Mississippi State University, Mississippi State, MS.
McKinion, J. M., D. N. Baker, J. D. Hesketh, and J. W. Jones. 1975. SIMCOT II: a simulation of cotton growth and yield. In Computer Simulation of a Cotton Production System. Users man- ual. U.S. Department of Agriculture, ARS-S-52, p. 27-82.
Nicholson, W. F., Jr. 1975. Feeding of Heliothis virescens (F.) and ^. zea (Boddie) on
cotton with emphasis and development of a simulation model of larval feeding. Ph.D. dissertation, Mississippi State Univer- sity, Mississippi State, MS.
Quaintance, A. L., and C. T. Brues. 1905. The cotton bollworm. U.S. Department of Agriculture,
Bureau of Entomology Bulletin No. 50.
Ritchie, J. T. 1972. Model for predicting evaporation from a row crop with in-
complete cover. Water Resources Research 8:1204-1213.
Schwab, G. 0., R. K. Frevert, T. W. Edminster, and K. K. Barnes. 1966. Soil water conservation engineering, 103 p. John Wiley &
Sons, New York.
Stapleton, H. N., D. R. Buxton, F. L. Watson, D. J. Nolting, and D. N. Baker.
1973. Cotton: a computer simulation of cotton growth. Univer- sity of Arizona, Agricultural Experiment Station Technical Bulletin No. 206.
Wilson, L. T., and A. P. Gutierrez. 1980. Fruit prédation submodel: Heliothis larvae feeding upon
cotton fruiting structures. Hilgardia 48:24-36.


481
Chapter 18
IMPACT OF ALTERNATIVE COTTON INSECT MANAGEMENT STRATEGIES ON PRODUCER INCOME IN MISSISSIPPI
E. H. Simpson, III, and D. W. Parvin, Jr. Department of Agricultural Economics Mississippi State University Mississippi State, MS 39762
ABSTRACT A simulation model was used to esti- mate the biological impacts of each of four al- ternative management strategies used against cotton insects in Mississippi. The results were then used as input in linear programming models to estimate the economic impacts of each of the alternative strategies on producer in- come. Soil constraints were found to limit acreage shifts to less than expected levels.
INTRODUCTION
Methods of controlling cotton insects have changed so radi- cally over time that they can be grouped by period; for example, the pre-boll weevil period, the pre-calcium arsenate period, the calcium arsenate period, and the synthetic organic insecticide period (Newsom 1974). We now appear to be entering a period during which new synthetic organic and biological insecticides will be components of integrated management strategies for controlling cot- ton insects. Since many of these strategies are based on a commun- ity or regional approach, a large proportion of the cotton acreage in a given treatment area must be included in a program. There- fore, producers require information about how new or proposed strategies will impact on their incomes and how the impact will differ from that of current methods of control.
THE PROBLEM
Cotton producers in many regions of the Cotton Belt have re- cently had to pay excessively high costs to control damaging in- sects. Thus, various strategies have been devised to aid them in reducing these costs. Two major alternatives are presently receiv- ing considerable attention as a result of the trials conducted in North Carolina and Mississippi, the so-called Boll Weevil Eradica- tion Trial (BWET) and the Optimum Pest Management Trial (OPMT). Eradication implies that the boll weevil will be eliminated from

482 E. H. SIMPSON III AND D. W. PARVIN, JR.
the environment. 0PM implies that boll weevil populations will be reduced to levels that do not require in-season treatment.
STRATEGIES TO BE EVALUATED
In this chapter, we report the results obtained wVien we used a simulation model in an attempt to estimate the impact on producer income of four alternative strategies developed from the two major strategies. It was assumed that each of the four had been imple- mented successfully and that the desired impact on the target pest(s) had been stabilized. These are compared with a fifth strategy, current normal insect control (CIC), that is, the ap- proach currently used by cotton producers. (CIC is not constant within or among regions of the Cotton Belt.) We do not address the transition periods so the massive costs during these periods are not considered. Additionally, these alternatives may impact the ecosystem in different and subtle ways and thereby have economic consequences for producers (a classic example is insecticide re- sistance), but these are not considered.
The alternative strategies considered are as follows:
(1) CIC—The approach to cotton insect control currently used by producers. CIC is constant neither within nor among regions.
(2) MOPM—Modified Optimum Pest Management. This alterna- tive assumes an increased level of producer education such that most acreage (field by field) is managed by using the best available technology applicable.
(3) 0PM—This alternative is equivalent to MOPM but com- munity or regional activities are included; these are designed to maintain subtreatment levels of boll weevils during most of the growing season.
(4) CIC-BWE—This alternative assumes that boll weevil eradication (BWE) begins with CIC already in place. Then after completion of BWE, producers manage the re- maining cotton insects, both those that are beneficial and those that are damaging, by using CIC techniques.
(5) OPM-BWE—This alternative assumes that BWE begins with an 0PM program already in place. Then after completion of BWE, producers manage the remaining cotton insects by using 0PM techniques.
COTTON PRODUCING REGIONS IN MISSISSIPPI
The Mississippi Agricultural and Forestry Experiment Station (MAFES) considers that there are five cotton producing regions in

PRODUCER INCOME AND MANAGEMENT STRATEGIES 483
Mississippi (Cameron 1978, Seale 1979) (Figure 1). However, admin- istratively, the Department of Entomology of the Mississippi Co- operative Extension Service (MCES) considers that there are only four. The difference stems from the MAFES delineation, which is primarily related to soil types and farm size; the MCES delineation is determined on the basis of insect problems. However, MAFES Re- gions IV and V are equivalent to one of the four MCES regions, and the other three MAFES regions are consistent with the other three MCES regions. The size of the five MAFES regions in terms of crop- land acreage and the historical relationship among cotton and al- ternative crops are summarized in Table 1.
METHODOLOGY
The analysis progressed through two distinct phases. The first was concerned with obtaining estimates of the biological im- pacts of the alternative strategies on per acre yield of cotton lint and the number of insecticide applications. The second phase was concerned with obtaining estimates of the economic impact of the biological changes on producers.
Method of Estimating Biological Impacts
The computer simulation model (CIM) was used in the experi- ment to determine the specific changes in per acre yield and num- ber of insecticide applications for each alternative strategy and for CIC. Readers interested in the details of this model are re- ferred to earlier publications (Brown et al. 1970, 1976, 1979; McClendon et al. 1977, Escarra 1979; Jones et al. 1979; Murphey 1980).
The experimental simulations were run on the Mississippi State University Univac 1100/80 computer. Each of the five strategies was simulated for four levels of infestation of three insect spe- cies. Also, 4 years of weather data were provided for each of the four MCES regions. The three insect groups were boll weevils (Anthonomus grandis Boheman), the bollworm/tobacco budworm complex (Heliothis spp.), and beneficial insects (predators and parasites). The four levels of infestation were zero, light, moderate, and heavy. However, the probability of occurrence of each level varies among these insect groups so the probabilities were provided by the Biological Evaluation Team (BET) of Beltwide Boll Weevil/Cotton In- sect Management Programs. The BET also provided estimates of the numbers of insects associated with each level of infestation.
The experimental design implied the need to execute CIM some 192 times for each strategy (4 infestation levels x 3 insect groups X 4 years x 4 regions = 192). Then since it was intended to simu- late five strategies, 960 executions of CIM would have been re- quired. However, CIC, the strategy used as the basis for compari- son, consists of as many as four substrategies in some regions
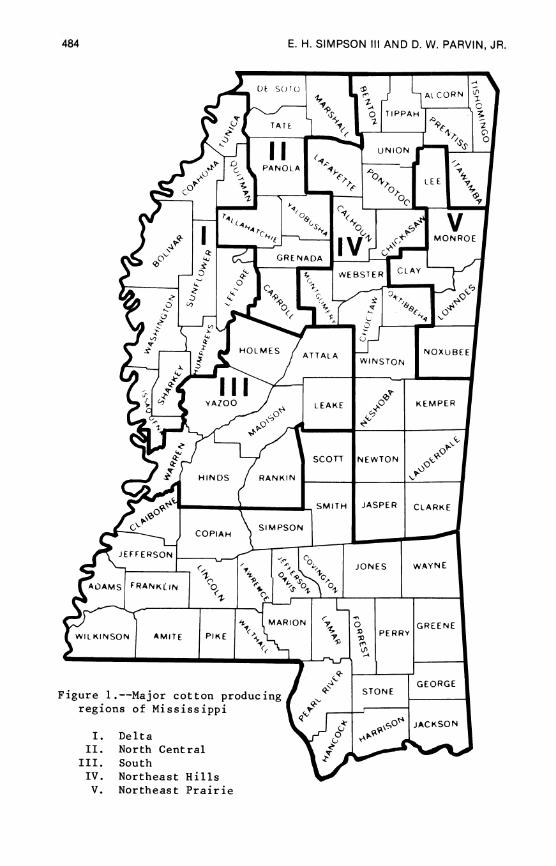
484 E. H. SIMPSON III AND D. W. PARVIN, JR.
Figure 1.—Major cotton producing regions of Mississippi
I. Delta II. North Central
III. South IV. Northeast Hills V. Northeast Prairie
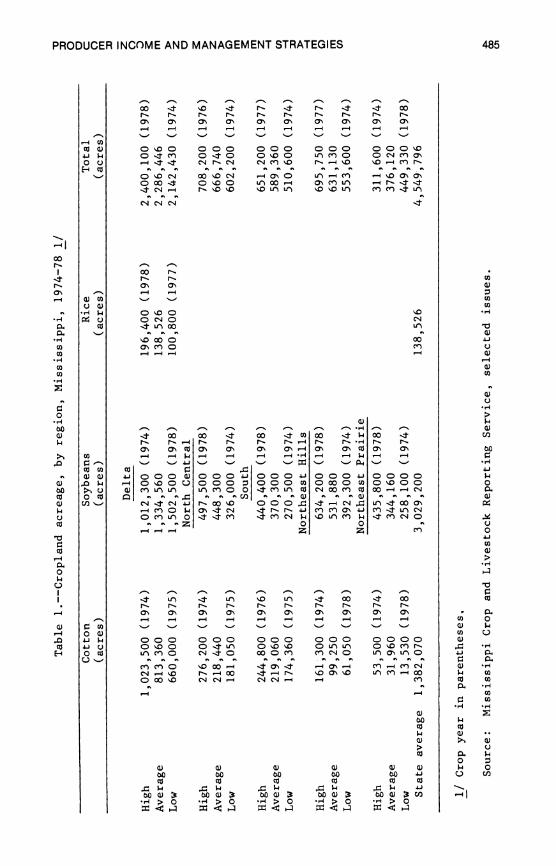
PRODUCER INCOME AND MANAGEMENT STRATEGIES 485
I
Pu Ou
C O
•H ÖO
u
ÖO CO 0) u o CO
c CO .-< cu o u
1-4 CO CO 0) U u O o H CO
0) CO
Pij Ü CO
CO C ^ CO CO 0) (U
>-, O O CO
CO v-^
CÜ C CO r-l O Q) ^ ■U }^
CO 4-» o H o CO
o w
CX)
CJN ON ON 1-H
<JN 1—1
ON i-H
<JN
<!■
ON i-H
ON
00
ON •—1
o vO O <t
O O o CNl
O O o CM
O o CM
o CO
O o VO
O m
o en r-l
O O
O o VO
O es
O vD CO ON CO r^
o vO o 00 ^ CM
C30 O
CM O in
ON 00 in
O
m m ON CO
CO in m
1—1
1—1
CO
vO
CO
CJN ON
<!■ m •» CN CM CM <!•
o VO o O CM O •Ñf in 00
vO 00 o <3N CO o
VO es in
00 CO
00 r^ .-I CJN CO rH »-I N-' U
tí O O <ü VO o U in in
<|- CM 4J CO O >-l CO m O
CJN
00
CJN C3N
O O O 4J o o o o o o 3 O O O in CO o o <f CO in
r^ 00 vo o o o ON <f es Nj- r^ r^ •-Í -^ CO <j- CO es
00 <f >-4 r^ r«* «H ON ON CO •—< r-) M
O O O +J O 00 O CO CM 00 CO CO
r. « « Oí ■<f —I es ^ CO CO ON 4J vo in CO }-i
o
o o o o o VO o o 00 »-H rH CM
in <í- 00 cjN CO "ñJ- m es <!• CO es o
m in vO in 00 00 r^ f^ r-- r^ r^ r^ r>. r^ r^ r^ ON
f—<
ON 1—1
ON 1—1
ejN 1-H
CJN t—t
C7N 1—1
<7N 1—t
CJN 1—1
C3N (3N f—)
O o o o O O O o o O o o O o o o O vO o o <t in o VO VO o in m o vO CO 1^ in CO o CM •> o 00 o CO CO es o in C5N in o
CO CO o CM rH VO o 00 vO
vO 00 -H r^ rH 00 es es rH
-* ON <t <t »H r^ es es 1—1
»H ON 1—1 VO <7N vO
CO iH CO es m CO ^ 00
Ö0 CO
'^ ^ ^ •H > o ffi <: hj
(U Q) (ü W) 00 Ö0 CO CO CO
-JC ^ ^ -^ ^ ^ ^ v^ bO Q) ^ txO Qí > Ö0 (U
•H > O •H > O •H > ce -í ^J ffi <Í HJ ffl <í
QJ 00 CO U Q) > CO
<ü Q) txO u CO CO
X U ■u W) Cü ^ œ
•H > O !X! < HJ
-o
<u o
•H > Wi 0) w
W) tí •H ■M M O Ou <U tó
,i¿ O O •M CO (U >
•H hJ
^3 tí CO
^ fX CO O 0) U CO O <u ^ •H 4J CU tí 0- O) •H V^ CO CO CO Ou •H
CO tí CO •H •H
S V^ CO CU >% (U
o a u o 0 )-4 o u C/3

486 E. H. SIMPSON III AND D. W. PARVIN, JR.
(once a week scouting, twice a week scouting, once a week scouting plus an occasional spot check, and a calendar spray program), and each of these would require as many executions of CIM as would a single strategy without substrategies. Consequently, an excessive number of possible executions were projected. Therefore, situa- tions determined to have an extremely low probability of occurrence were not simulated, and a panel of entomologists uniquely familiar with cotton production and insect problems in Mississippi was called upon to adjust the results of the experiment to account for the situations that were not simulated.
Method of Estimating Economic Impacts
Five regional linear programming optimization models were used in the experiment to estimate the impact of the alternative strate- gies on producer income. The constraint for each of the linear programming models was the soil resource. Yields of cotton and of the alternative crop (soybean) by soil resource for each of the re- gions are reported in Table 2.
The models were structured in such a way as to select the crop (cotton or soybeans) for a given soil that would maximize returns above specified expenses (direct and fixed costs). Cost estimates (inputs) used are appropriate for 1979 (Hamill et al. 1979a, 1979b, 1979c; Parvin et al. 1979a, 1979b). Output prices (producer re- turns) selected were the average prices received by Mississippi farmers in 1979 (Mississippi Crop and Livestock Reporting Service, selected issues). The prices were: cotton lint—$0.607 per pound; cotton seed—$0.062 per pound; soybeans—$6.45 per bushel; and rice—$3.60 per bushel. These prices were assumed to be constant because numerous factors such as weather, domestic consumption, and export demand have a far greater impact on commodity prices than does the particular strategy selected for control of insect pests.
Each of the linear programming models was executed once for CIC and once for each of the four alternative strategies. The ef- fect of each alternative strategy was built into the linear pro- gramming models by modifying the CIC cotton yield and that portion of the direct cost that would be associated with insect control for that strategy. Each insecticide application was assumed to cost $4.96 per acre (including aerial application charges). This amount reflects the 1979 cost of application of the standard rate of EPN plus methyl parathion.
Since each alternative strategy could impact on producer in- come by increasing per acre returns for cotton or increasing cotton acreage, the linear programming models were designed to capture both effects, subject to the appropriate constraints. The linear programming models were solved by using the Functional Mathematics Programming System—a standard linear programming solution algo- rithm (Anonymous 1977, Cameron 1978, Cameron et al. 1981, Seale 1979).
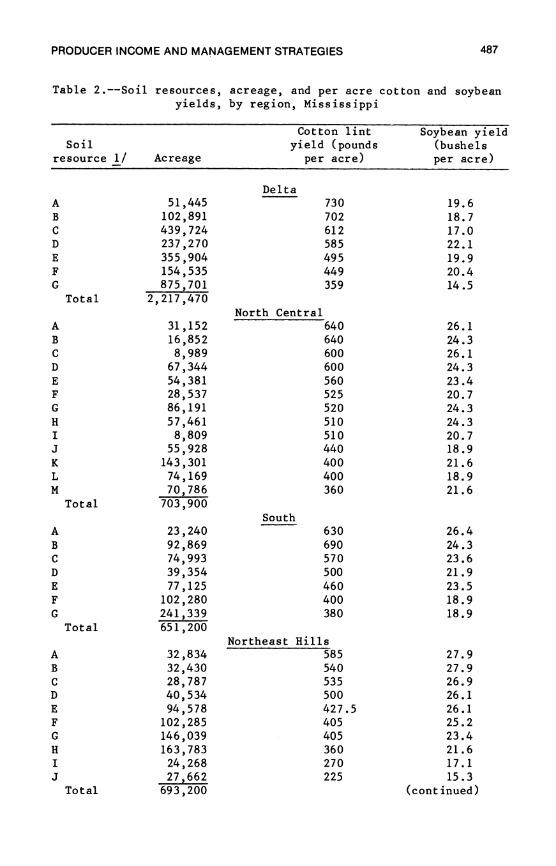
PRODUCER INCOME AND MANAGEMENT STRATEGIES 487
Table 2.—Soil resources, acreage, and per acre cotton and soybean yields, by region, Mississippi
Cotton lint Soybean yield Soil yield (pounds (bushels
resource \J Acreage per acre) per acre)
Delta A 51,445 730 19.6 B 102,891 702 18.7 C 439,724 612 17.0 D 237,270 585 22.1 E 355,904 495 19.9 F 154,535 449 20.4 G 875,701 359 14.5
Total 2,217,470 North Central
A 31,152 640 26.1 B 16,852 640 24.3 C 8,989 600 26.1 D 67,344 600 24.3 E 54,381 560 23.4 F 28,537 525 20.7 G 86,191 520 24.3 H 57,461 510 24.3 I 8,809 510 20.7 J 55,928 440 18.9 K 143,301 400 21.6 L 74,169 400 18.9 M 70,786 360 21.6
Total 703,900 South
A 23,240 630 26.4 B 92,869 690 24.3 C 74,993 570 23.6 D 39,354 500 21.9 E 77,125 460 23.5 F 102,280 400 18.9 G 241,339 380 18.9
Total 651,200 Northeast Hills
A 32,834 585 27.9 B 32,430 540 27.9 C 28,787 535 26.9 D 40,534 500 26.1 E 94,578 427.5 26.1 F 102,285 405 25.2 G 146,039 405 23.4 H 163,783 360 21.6 I 24,268 270 17.1 J 27,662 225 15.3
Total 693,200 (continued)

488 E. H. SIMPSON III AND D. W. PARVIN, JR.
Table 2 (continued).—Soil resources, acreage, and per acre cotton and soybean yields, by region, Mississippi
Cotton lint Soybean yield Soil yield (pounds (bushels
resource J^/ Acreage per acre) per acre)
Northeast Prairie
A 14,046 550 25.6 B 14,120 530 22.6 C 42,019 505 26.0 D 95,464 470 26.8 E 154,929 445 26.6 F 36,331 400 21.7 G 88,091 380 20.7
Total 445,000
State total 4,710,770
\J A-M represent broad soil classifications identified by Cameron 1978; Collins 1980; Seale 1979; and U.S. Department of Agriculture, selected issues.
RESULTS
Biological Impact
The results of the adjusted CIM simulation of the four alter- native strategies and of CIC are presented in Table 3. The major changes in yield and in the number of insecticide applications were related to the level of the boll weevil infestation, and the impact of this level was greatest in the South Region where the level of infestation is highest. Since a large portion of the Delta Region is not infested by boll weevils, estimated impacts of the alter- native strategies were not as great in the Delta Region as in other regions.
In the four regions where boll weevils are a problem, MOPM was estimated to result in an 8- to 10-pound increase per acre in yield and a 0.9-1.0 decrease in the number of insecticide applications. 0PM produced an estimated 18- to 22-pound increase in yield and a 2.2-2.4 decrease in the number of insecticide applications. CIC- BWE produced an estimated increase in yield of 19-24 pounds and a 2.4-2.6 decrease in the number of insecticide applications. 0PM- BWE had the largest impact (Table 3).
In the Delta Region, which is relatively free of boll weevils, both MOPM and 0PM slightly outperformed CIC-BWE. This difference is attributed primarily to improved early-season insect control re- sulting from the increased level of producer education.

PRODUCER INCOME AND MANAGEMENT STRATEGIES 489
Table 3.—Estimated increases per acre in yield of cotton lint and decreases per acre in applications of insecticide by region and by strategy, Mississippi \_l
Region CIC MOPM 0PM CIC-BWE OPM-BWE
Delta Base +6(-0.6) +10("0.9) +4(-0.4) +7(-0.9) North Central Base +10(-0.9) +22(-2.2) +23(-2.4) +29(-2.9) South Base +10(-0.9) +22(-2.4) +24(-2.6) +31(-3.1) Northeast Hills Base +8(-1.0) +18(-2.4) +19(-2.5) +24(-3.0) Northeast Prairie Base +8(-1.0) +18(-2.4) +19(~2.5) +24(-2.0)
1/ Decrease in parentheses.
Note: CIC = Current Insect Control; MOPM = Modified Optimum Pest Management; 0PM = Optimum Pest Management; CIC-BWE = Current Insect Control-Boll Weevil Eradication; OPM-BWE = Optimum Pest Management-Boll Weevil Eradication.
Economic Impact
The results of the attempt to determine economic impacts are presented in Tables 4-8 for each of the regions so each reader can determine how acccurately the linear programming models simulate CIC and can compare the impact of the alternative strategies on the various regions.
In fact, the linear programming models tracked actual acreage extremely well; thus, they can be viewed as providing reasonable estimates of the impacts of alternative strategies relative to CIC. For example, the estimated cotton acreage for CIC in the Delta Re- gion is consistent with the 1974-78 Delta Region average (Table 1).
Table 4.—Impact of alternative cotton insect control strategies on producer returns (above specified costs) and on cotton acreage. Delta Region of Mississippi
Strategy
Total Returns above cotton
specified expenses acreage (million dollars) (acres)
CIC MOPM 0PM CIC-BWE OPM-BWE
133.572 831,330 139.558 831,330 139.558 831,330 137.554 831,330 141.370 831,330
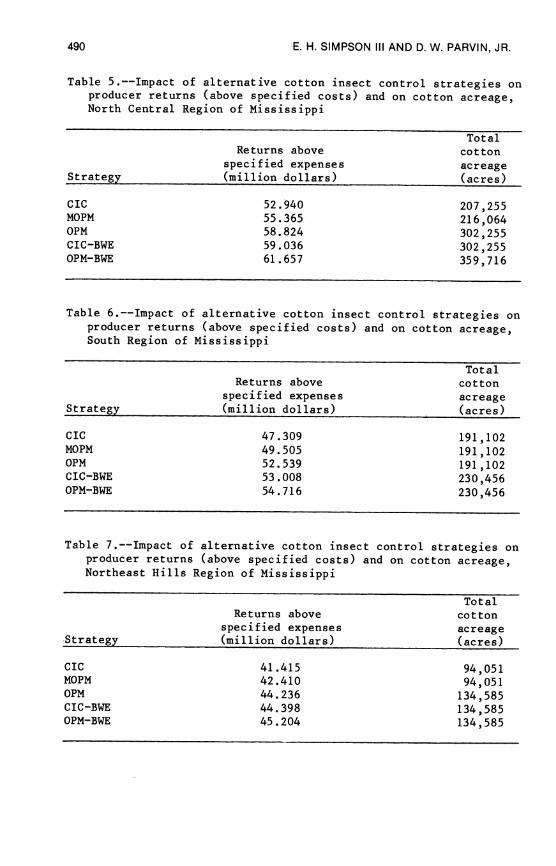
490 E. H. SIMPSON III AND D. W. PARVIN, JR.
Table 5.—Impact of alternative cotton insect control strategies on producer returns (above specified costs) and on cotton acreage, North Central Region of Mississippi
Total Returns above cotton
specified expenses acreage Strategy (million dollars) (acres)
CIC 52.940 207,255 MOPM 55.365 216,064 0PM 58.824 302,255 CIC-BWE 59.036 302,255 OPM-BWE 61.657 359,716
Table 6.—Impact of alternative cotton insect control strategies on producer returns (above specified costs) and on cotton acreage. South Region of Mississippi
Total Returns above cotton
specified expenses acreage Strategy (million dollars) (acres)
CIC 47.309 191,102 MOPM 49.505 191,102 0PM 52.539 191,102 CIC-BWE 53.008 230,456 OPM-BWE 54.716 230,456
Table 7.—Impact of alternative cotton insect control strategies on producer returns (above specified costs) and on cotton acreage. Northeast Hills Region of Mississippi
Strategy
CIC MOPM 0PM CIC-BWE OPM-BWE
Total Returns above cotton
specified expenses acreage (million dollars) (acres)
41.415 94,051 42.410 94,051 44.236 134,585 44.398 134,585 45.204 134,585

PRODUCER INCOME AND MANAGEMENT STRATEGIES 491
Table 8.—Impact of alternative cotton insect control strategies on producer returns (above specified costs) and on cotton acreage, Northeast Prairie Region of Mississippi
Strategy
Total Returns above cotton
specified expenses acreage (million dollars) (acres)
CIC 33.835 28,166 MOPM 34.133 28,166 0PM 34.527 28,166 CIC-BWE 34.561 28,166 OPM-BWE 34.884 70,185
Also, none of the alternatives caused an increase in cotton acreage there relative to CIC. Likewise, cotton acreage estimated for CIC in the North Central Region was consistent with actual acreage. MOPM increased acreage, OTPM and CIC-BWE increased it more, and OPM- BWE had an even greater impact. In the South Region, CIC cotton acreage was estimated to be 191,102 acres, somewhat lower than the actual average (Table 1). There was no increase in cotton acreage with either MOPM or 0PM, but both CIC-BWE and OPM-BWE increased acreage. Also, in the Northeast Hills Region, cotton acreage esti- mated for CIC was somewhat lower than the actual acreage, and again MOPM did not result in an increase in acreage. However, 0PM, CIC- BWE, and OPM-BWE all increased cotton acreage. CIC cotton acreage estimated for the Northeast Prairie Region likewise was a little low. MOPM, 0PM, and CIC-BWE did not result in an increase in cot- ton acreage. However, OPM-BWE essentially doubled the average for 1974-78 and more than doubled the acreage for CIC. The dominant factors that influenced the results were the constraints on acreage and the relative yields of cotton and soybean associated with the various soil groups.
The estimated returns above specified expenses for each region and the State are reported in Table 9.
Table 9.—Estimated total returns (millions of dollars) above spec- ified expenses for five insect control strategies and five re- gions in Mississippi
Region CIC MOPM 0PM CIC-BWE OPM-BWE ilVtT 133.572 139.558 139.558 137.554 141.370 North Central 52.940 55.365 58.824 59.036 61.657 South 47.309 49.505 52.539 53.008 54.716 Northeast Hills 41.415 42.410 44.236 44.398 45.204 Northeast Prairie 33.835 34.133 34.527 34.561 34.884 State 309.071 320.971 329.684 328.557 337.831
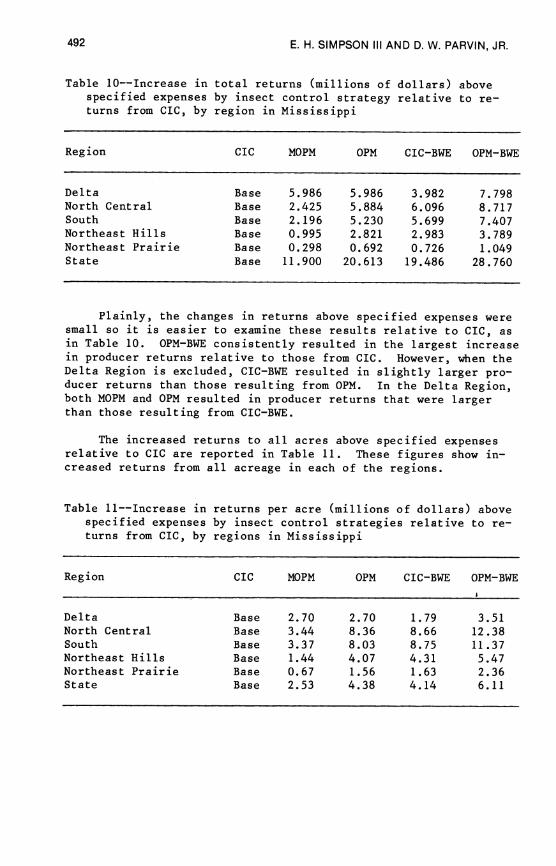
492 E. H. SIMPSON III AND D. W. PARVIN, JR.
Table 10—Increase in total returns (millions of dollars) above specified expenses by insect control strategy relative to re- turns from CIC, by region in Mississippi
Region CIC MOPM 0PM CIC-BWE OPM-BWE
Delta Base 5.986 5.986 3.982 7.798 North Central Base 2.425 5.884 6.096 8.717 South Base 2.196 5.230 5.699 7.407 Northeast Hills Base 0.995 2.821 2.983 3.789 Northeast Prairie Base 0.298 0.692 0.726 1.049 State Base 11.900 20.613 19.486 28.760
Plainly, the changes in returns above specified expenses were small so it is easier to examine these results relative to CIC, as in Table 10. OPM-BWE consistently resulted in the largest increase in producer returns relative to those from CIC. However, when the Delta Region is excluded, CIC-BWE resulted in slightly larger pro- ducer returns than those resulting from 0PM. In the Delta Region, both MOPM and 0PM resulted in producer returns that were larger than those resulting from CIC-BWE.
The increased returns to all acres above specified expenses relative to CIC are reported in Table 11. These figures show in- creased returns from all acreage in each of the regions.
Table 11—Increase in returns per acre (millions of dollars) above specified expenses by insect control strategies relative to re- turns from CIC, by regions in Mississippi
Region CIC MOPM 0PM CIC-BWE OPM-BWE t
Delta Base 2.70 2.70 1.79 3.51 North Central Base 3.44 8.36 8.66 12.38 South Base 3.37 8.03 8.75 11.37 Northeast Hills Base 1.44 4.07 4.31 5.47 Northeast Prairie Base 0.67 1.56 1.63 2.36 State Base 2.53 4.38 4.14 6.11

PRODUCER INCOME AND MANAGEMENT STRATEGIES 493
IMPLICATIONS
The figures presented in Tables 9, 10, and 11 indicate that in some regions, and in the State, the producer returns associated with the various alternative strategies did not differ substan- tially. Therefore, the "preferred" strategy for Mississippi cotton producers may be related to factors and costs that were not con- sidered in this study, for example, the producer share of the cost of eradication during the transition period and the producer share of the cost of treating diapausing boll weevils after the 0PM pro- gram has been implemented and the ecosystem stabilized. Apparently a careful examination of these two items must be undertaken.
LIMITATIONS
In this study, we examined alternative cotton insect manage- ment strategies from the producer standpoint assuming no change in product prices. Policy makers who must select the "best" strategy must consider numerous other factors such as public costs, pesti- cide resistance, impact on the environment, and regional shifts in cotton acreage among States. Also, the results of this analysis are specific for the regions of Mississippi. Extrapolation to other portions of the Cotton Belt would not be appropriate.

494 E. H. SIMPSON III AND D. W. PARVIN, JR.
LITERATURE CITED
Anonjnnous. 1977. Functional mathematical programming system (FMPS). Pro-
gram reference, Sperry Rand Corporation.
Brown, L. G., J. W. Jones, and F. A. Harris. 1976. A simulation study of insect pest management alternatives
by integration of a Heliothis spp. model and a crop model. Presented at the Annual ASAE Meeting, Lincoln, NE, Paper No. 76-5025.
R. W. McClendon, and J. W. Jones. 1970. Complete simulation of the interactions between the cotton
crop and insect pests. Transactions of the ASAE 22.
R. W. McClendon, and J. W. Jones. 1979. An integrated simulation of cotton and insect pests. Pre-
sented at the Joint National Meeting of the Institute of Man- agement Science and Operations Research Society of America, New Orleans, LA.
Cameron, D. M. 1978. An economic assessment of production alternatives avail-
able to representative farms of the Mississippi Delta. Unpub- lished M.S. thesis, Department of Agricultural Economics, Mississippi State University.
D. W. Parvin, Jr., and F. T. Cooke, Jr. 1981. A survey of farm resources and production activities in the Yazoo-Mississippi Delta. Mississippi Agricultural and Fores-
try Experiment Station, AECR. R. No. 120.
Collins, G. S., III. . 1980. An econometric simulation model for evaluating aggregate
economic impacts of technological change on major U.S. field crops. Unpublished dissertation. Department of Agricultural Economics, Texas A&M University.
Escarra, B. 1979. Estimates of the effects of alternative insect management
programs on cotton yield and insect control cost in the State of Mississippi. Unpublished thesis. Department of Industrial Engineering, Mississippi State University.
Hamill, J. G., D. W. Parvin, Jr., F. T. Cooke, Jr., D. M. Cameron, and R. D. Seale.
1979a. Cost of production estimates for the black belt of north- east Mississippi, 1979. Special Edition of Research High- lights, Mississippi Agricultural and Forestry Experiment Sta- tion, Mississippi State University.

PRODUCER INCOME AND MANAGEMENT STRATEGIES 495
Hamill, J. G., D. W. Parvin, Jr., F. T. Cooke, Jr., R. D. Seale, and G. Simpson.
1979b. Estimated costs and returns, row crops, northern brown loam area of Mississippi, 1979. Special Edition of Research Highlights, Mississippi Agricultural and Forestry Experiment Station, Mississippi State University.
R. D. Seale, D. W. Parvin, Jr., and F. T. Cooke, Jr. 1979c. Estimated costs and returns, row crops, central brown
loam area of Mississippi, 1979. Mississippi Agricultural and Forestry Experiment Station, Mississippi State University, AEG M.R. No. 91.
Jones, J. W., L. G. Brown, and J. D. Hesketh. 1979. COTCROP, a complete model for cotton growth and yield. In Predicting Photosynthesis for Ecosystem Models. CRC Press, West Palm Beach, FL.
McClendon, R. W., L. G. Brown, and J. W. Jones. 1977. Investigation of integrated pest management strategies utilizing computer simulation of the cotton crop insect inter- actions. Presented at the Annual ASAE Meeting, NC Paper No. .77-5019.
Mississippi Crop and Livestock Reporting Service, selected issues.
Murphey, S. M. 1980. Use of simulation to evaluate alternative cotton insect
pest control strategies. Unpublished M.S. thesis. Department of Industrial Engineering, Mississippi State University.
Newsom, L. D. 1974. Pest management: history, current status and future prog-
ress. In Proceedings, Summer Institute on Biological Control of Plan~Insects and Diseases, Mississippi State University, p. 64.
Parvin, D. W., Jr., J. G. Hamill, F. T. Cooke, Jr., D. M. Cameron, and E. H. Simpson III.
1979a. Cost of production estimates, major crops, sand clay hills of Mississippi, 1979. Special Edition of Research High- lights, Mississippi Agricultural and Forestry Experiment Sta- tion, Mississippi State University.
J. G. Hamill, F. T. Cooke, Jr., S. H. Holder, Jr., and D. M. Cameron.
1979b. Budgets for major crops. Delta of Mississippi, 1979. Special Edition of Research Highlights, Mississippi Agricul- tural and Forestry Experiment Station, Mississippi State Uni- versity.

^^^ E. H. SIMPSON III AND D. W. PARVIN, JR.
Seale, R. D.
1979. The economic impact of optimum pest management on cotton production in Mississippi. Unpublished thesis, Department of Agricultural Economics, Mississippi State University.
U.S. Department of Agriculture. Soil Conservation Service and Mississippi Agricultural and Forestry Experiment Station, Soil Survey, selected issues.

497
Chapter 19
ECONOMIC EVALUATION OF THE BOLL WEEVIL ERADICATION TRIAL IN NORTH CAROLINA, 1978-80
G. A. Carlson and L. F. Suguiyama Department of Economics and Business North Carolina State University Raleigh, NC 27650
ABSTRACT Biological, economic, and environ- mental variables were monitored in the Boll Weevil Eradication Trial (BWET) after opera- tions began in 1978. The objective of the BWET was to determine the feasibility of its imple- mentation across the boll weevil-infested por- tion of the Cotton Belt. In this chapter, we tabulate the economic effects of the BWET on the trial zone and on a comparison zone by examining data drawn from a random sample of farmers. The economic evaluation of the Trial centered on measurement of changes in the costs of production, in cotton yields, and in public expenditures. The major economic impact of the program was the reduction in the use of insec- ticides in the trial zone over time and the even greater reduction in the use of insecti- cides in comparison with a nontrial zone. There was an indication that the reduction in cotton acreage that occurred in the Trial zone was attributable to farmer eradication fees in the first year of the trial. The direct ef- fects of the trial on yields were difficult to assess; however, there was evidence that small increases in yields were an indirect effect of the trial. In order to insure complete removal of the boll weevil from the trial area, moni- toring of the project was very thorough and, therefore, public costs were high.
INTRODUCTION AND METHODS OF ANALYSIS
The major purpose of the Boll Weevil Eradication Trial (BWET) in North Carolina was to determine whether eradication of the boll weevil (Anthonomus grandis Boheman) from a large area is techni- cally feasible. Therefore, the Trial was planned so that biologi- cal, environmental, and economic variables might be monitored. Information on these three features is essential to any decision

^9Ö G. A. CARLSON AND L SUGUIYAMA
about the desirability of attempting eradication across the boll weevil-infested portion of the Cotton Belt in the United States.
This economic evaluation of the BWET centers on the measure- ment of changes in costs of cotton production, in cotton yields, and in public expenditures relative to current insect control (CIC) practices.
Comparisons with CIC in the same years as the BWET (1978-80) were possible by examination of data from a randomly selected set of 60 farmers in three counties outside the trial area (control zone, CZ). The information was also obtained from a random sample of 133 farmers in four counties of the trial area (eradication zone, EZ). It was also possible to compare CIC in the eradication zone itself by examining the costs and yields in the 4-year period prior to the implementation of the Trial (1974-77).
In this chapter we present data for the base period 1974-77 and for the 3 years of the BWET (1978-80). Data on pesticide use, cot- ton yields, and public expenditures are actual figures obtained in the personal and telephone surveys of about 200 randomly selected farmers. This information has been supplemented by information from the Animal and Plant Health Inspection Service (APHIS), BWET per- sonnel, spray group directors, the Biological Evaluation Team of the State Extension and Crop Reporting Service offices, and a sample of pesticide applicators and pesticide dealers.
The regions covered by the study are the EZ and the CZ. The EZ includes Edgecombe, Halifax, Northampton, and Nash Counties; that is, about 95% of the APHIS evaluation zone. The CZ includes Robeson, Scotland, and Cleveland Counties. A limited amount of cotton (less than 20% of the State acreage) is grown in other counties of North Carolina, but these areas were not included in the analysis.
There are known differences and similarities in cotton produc- tion between the CZ and the EZ. Table 1 gives a summary of differ- ences and similarities of characteristics for cotton farms in the CZ and EZ (Grube and Carlson 1978). Major differences include smaller cotton acreages, lower yields, lower scouting costs, more use of spray groups, and a larger share of pest control efforts for boll weevils in the EZ than in the CZ. Similarities include the number of trips across the fields, years of farming experience, farmer age, boll weevil and bollworm (Heliothis zea (Boddie)) treatment thresholds, number of treatments with boll weevil as the primary target, number of diapause treatments per season (1974-76), and average length (days) of the pest control season. The histori- cal differences in pest control between the two zones are well known, so comparisons during the eradication period can be made.
Cotton acreages and yields for the EZ and the CZ for 1970-80 are shown in Figures 1 and 2. Cotton acreages for both zones show a distinct declining trend over the 10-year period. Cotton yields
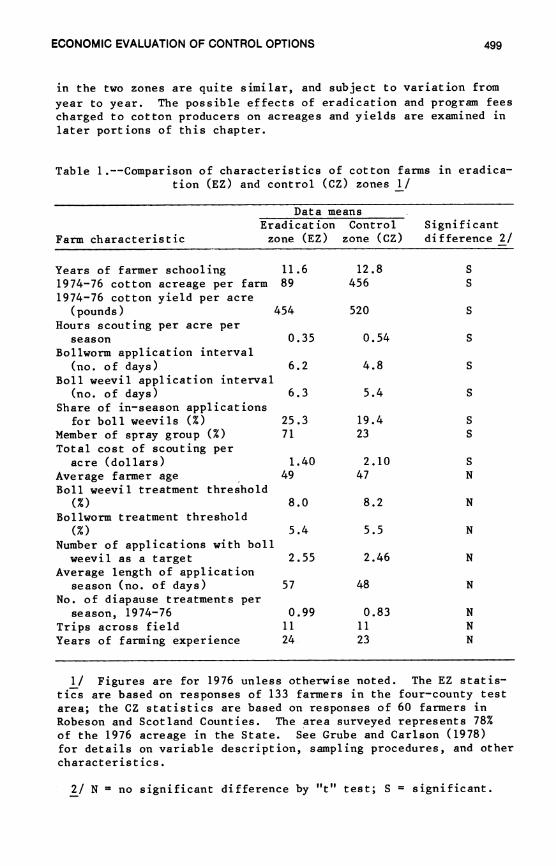
ECONOMIC EVALUATION OF CONTROL OPTIONS 499
in the two zones are quite similar, and subject to variation from year to year. The possible effects of eradication and program fees charged to cotton producers on acreages and yields are examined in later portions of this chapter.
Table 1.—Comparison of characteristics of cotton farms in eradica- tion (EZ) and control (CZ) zones J^/
Data means Eradication Control Significant
Farm characteristic zone (EZ) zone (CZ) difference Ij
Years of farmer schooling 11.6 12.8 S 1974-76 cotton acreage per farm 89 456 S 1974-76 cotton yield per acre
(pounds) 454 520 S Hours scouting per acre per
season 0.35 0.54 S Bollwotm application interval
(no. of days) 6.2 4.8 S Boll weevil application interval
(no. of days) 6.3 5.4 S Share of in-season applications
for boll weevils (%) 25.3 19.4 S Member of spray group (%) 71 23 S Total cost of scouting per
acre (dollars) 1.40 2.10 S Average farmer age 49 47 N Boll weevil treatment threshold
(%) 8.0 8.2 N Bollworm treatment threshold
(%) 5.4 5.5 N Number of applications with boll
weevil as a target 2.55 2.46 N Average length of application
season (no. of days) 57 48 N No. of diapause treatments per
season, 1974-76 0.99 0.83 N Trips across field 11 11 N Years of farming experience 24 23 N
\j Figures are for 1976 unless otherwise noted. The EZ statis- tics are based on responses of 133 farmers in the four-county test area; the CZ statistics are based on responses of 60 farmers in Robeson and Scotland Counties. The area surveyed represents 78% of the 1976 acreage in the State. See Grube and Carlson (1978) for details on variable description, sampling procedures, and other characteristics.
2/ N = no significant difference by "t" test; S = significant.
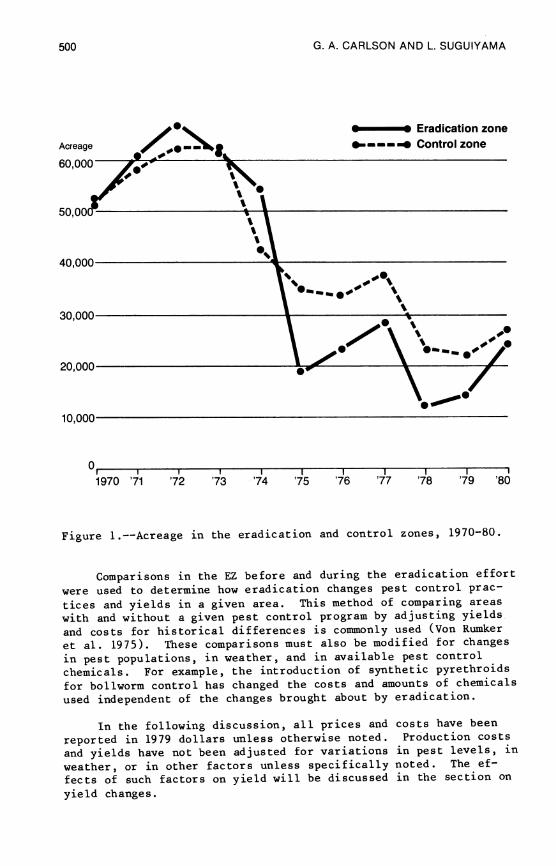
500 G. A. CARLSON AND L SUGUIYAMA
• Eradication zone Acreage
60,000
20,000 u-O^ 10,000-
0 t 1 1 1 1 1 1 1 1 \ 1 1970 71 72 73 74 75 76 77 78 79 '80
Figure 1.—Acreage in the eradication and control zones, 1970-80.
Comparisons in the EZ before and during the eradication effort were used to determine how eradication changes pest control prac- tices and yields in a given area. This method of comparing areas with and without a given pest control program by adjusting yields and costs for historical differences is commonly used (Von Rumker et al. 1975). These comparisons must also be modified for changes in pest populations, in weather, and in available pest control chemicals. For example, the introduction of synthetic pyrethroids for bollworm control has changed the costs and amounts of chemicals used independent of the changes brought about by eradication.
In the following discussion, all prices and costs have been reported in 1979 dollars unless otherwise noted. Production costs and yields have not been adjusted for variations in pest levels, in weather, or in other factors unless specifically noted. The ef- fects of such factors on yield will be discussed in the section on yield changes.

ECONOMIC EVALUATION OF CONTROL OPTIONS 501
Average yield per acre (in lbs. of lint)
V-. //'^v^-.../! 400 ^ ^^.^^ ^
300 —
200- ^ Eradication zone
•-----* Control zone 100-
'I 1 1 1 1 1 1 1 1 1 1 1970 71 72 73 74 75 76 77 78 79 '80
Figure 2.—Average yield per acre in the eradication and control zones, 1970-80.
Source: North Carolina Crop Reporting service.
CURRENT INSECT CONTROL
Current insect control is defined as those practices that the average grower uses to control cotton insects. This is the base set of conditions to which eradication is compared for each of the two zones.
Some of the features of historical insect control in the EZ and the CZ are shown in Table 2 and Figure 3. County averages and the acreage-weighted zone averages for yield, numbers of insecti- cide applications, and total insect control cost are given. This information is based on grower surveys asking for recall back to 1974. Items (2) and (4) in Table 2 are zone averages for only those growers who continued to grow cotton in 1978 or 1979. These averages are computed since there is a considerable change in grow- ers between these two periods. Historically, in-season treatments have numbered ca. 2.1 treatments per season more in the CZ than in the EZ (11.13 treatments per acre in the CZ and 9.03 in the EZ). The number of diapause treatments has been slightly less than one treatment per season in both zones (0.92 in the EZ and 0.82 in the CZ). Total insect control costs have been slightly higher in the CZ than in the EZ, about 12% higher for all growers ($56.83 in the CZ and $50.81 in the EZ). These costs include scouting costs at
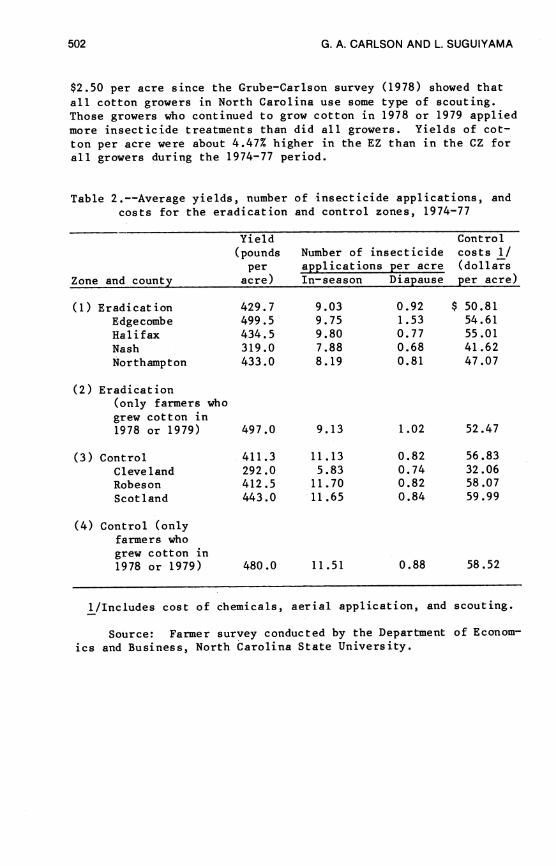
502 G. A. CARLSON AND L SUGUIYAMA
$2.50 per acre since the Grube-Carlson survey (1978) showed that all cotton growers in North Carolina use some type of scouting. Those growers who continued to grow cotton in 1978 or 1979 applied more insecticide treatments than did all growers. Yields of cot- ton per acre were about 4.47% higher in the EZ than in the CZ for all growers during the 1974-77 period.
Table 2.—Average yields, number of insecticide applications, and costs for the eradication and control zones, 1974-77
Yield Control (pounds Number of insecticide costs 1/
per applica tions per acre (dollars Zone and county acre) In-season Diapause per acre)
(1) Eradication 429.7 9.03 0.92 $ 50.81 Edgecombe 499.5 9.75 1.53 54.61 Halifax 434.5 9.80 0.77 55.01 Nash 319.0 7.88 0.68 41.62 Northampton 433.0 8.19 0.81 47.07
(2) Eradication (only farmers who grew cotton in 1978 or 1979) 497.0 9.13 1.02 52.47
(3) Control 411.3 11.13 0.82 56.83 Cleveland 292.0 5.83 0.74 32.06 Robeson 412.5 11.70 0.82 58.07 Scotland 443.0 11.65 0.84 59.99
(4) Control (only farmers who grew cotton in 1978 or 1979) 480.0 11.51 0.88 58.52
1/Includes cost of chemicals, aerial application, and scouting.
Source: Farmer survey conducted by the Department of Econom- ics and Business, North Carolina State University.
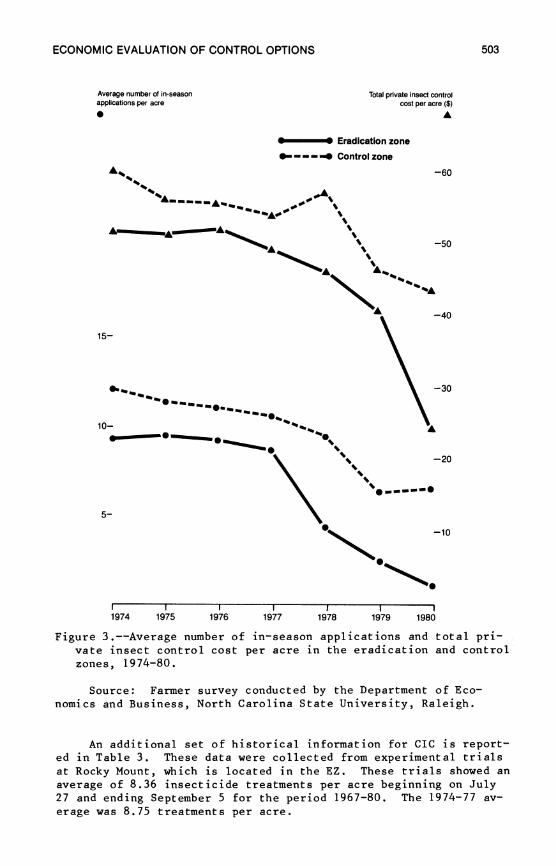
ECONOMIC EVALUATION OF CONTROL OPTIONS 503
Average number of in-season applications per acre
Total private Insect control cost per acre ($)
▲
Eradication zone
Controi zone
-60
1974 1975 1976 1977 I
1978 1979 1980
Figure 3.—Average number of in-season applications and total pri- vate insect control cost per acre in the eradication and control zones, 1974-80.
Source: Farmer survey conducted by the Department of Eco- nomics and Business, North Carolina State University, Raleigh.
An additional set of historical information for CIC is report- ed in Table 3. These data were collected from experimental trials at Rocky Mount, which is located in the EZ. These trials showed an average of 8.36 insecticide treatments per acre beginning on July 27 and ending September 5 for the period 1967-80. The 1974-77 av- erage was 8.75 treatments per acre.
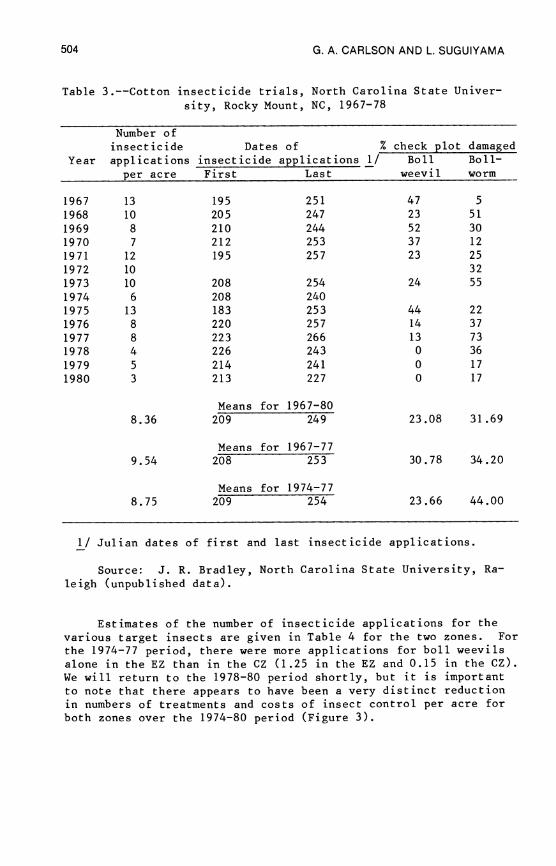
504 G. A. CARLSON AND L SUGUIYAMA
Table 3.—Cotton insecticide trials, North Carolina State Univer- sity, Rocky Mount, NC, 1967-78
Number of insecticide Dates of % check plot damaged
Year applications insecticide applications 1/ Boll Boll- per acre First Last weevi 1 worm
1967 13 195 251 47 5 1968 10 205 247 23 51 1969 8 210 244 52 30 1970 7 212 253 37 12 1971 12 195 257 23 25 1972 10 32 1973 10 208 254 24 55 1974 6 208 240 1975 13 183 253 44 22 1976 8 220 257 14 37 1977 8 223 266 13 73 1978 4 226 243 0 36 1979 5 214 241 0 17 1980 3 213 227 0 17
Means for 1967-80 8.36 209 249 23.08 31.69
9.54
8.75
Means for 1967-77 208 253
Means for 1974-77 209 254
30.78 34.20
23.66 44.00
1/ Julian dates of first and last insecticide applications.
Source: J. R. Bradley, North Carolina State University, Ra- leigh (unpublished data).
Estimates of the number of insecticide applications for the various target insects are given in Table 4 for the two zones. For the 1974-77 period, there were more applications for boll weevils alone in the EZ than in the CZ (1.25 in the EZ and 0.15 in the CZ). We will return to the 1978-80 period shortly, but it is important to note that there appears to have been a very distinct reduction in numbers of treatments and costs of insect control per acre for both zones over the 1974-80 period (Figure 3).

ECONOMIC EVALUATION OF CONTROL OPTIONS 505
Table 4.—Insecticide applications by target insect
Target Average no. appl ications insect 1974-77 1978 1979 1980
Eradication zone Boll weevil 1.25 0.00 0.00 0.00 Bollworm-boll
weevil 1/ 7.66 0.00 0.00 0.00 Bollworm 0.10 4.40 2.43 1.13 Boll weevil-
diapause 0.92 5.70
Control zone
0.00 0.00
Boll weevil 0.15 0.00 0.09 0.65 Bollworm-boll
weevil 1/ 9.40 0.15 0.52 2.73 Bollworm 0.76 8.91 5.42 2.87 Boll weevil-
diapause 0.82 0.05 0.59 0.37 Other:
Spider mites 0.00 0.21 0.11 0.20 Thrips 0.00 0.06 0.05 0.00
1/ The bollworm-boll weevil category refers to bollworms as the major pest and boll weevils as the secondary pest.
Source: Farmer survey conducted by the Department of Econom- ics and Business, North Carolina State University, Raleigh.
Table 5 gives estimates for CIC during 1978-80 for the CZ and the EZ. 1/ Farmer surveys provided information on CIC practices in the CZ. These estimates are given in the final three columns of Table 5 for each of the 3 years of the BWET (1978-80). The most direct comparison over time for CIC is for the CZ. For example. Table 2 shows annual insect control costs for 1974-77 of $56.83 per acre; the comparable figures for total private costs per acre (item 6 in Table 5) are $57.53 in 1978, $46.68 in 1979, and $43.92 in 1980. Numbers of in-season treatments declined from 11.13 in the base period (Table 2) to 9.33 in 1978, 6.19 in 1979, and 6.45 in 1980 (Table 5). These figures reflect both the relatively low pest infestations of 1978 and 1979 and the slight increase in boll weevil pressure in 1980 (Table 4). In addition, the dry August weather of 1978 and 1979 and the sharp increase in the use of syn- thetic pyrethroid insecticides tended to reduce the need to make as many applications.
j_/ The CZ as defined in this chapter is not directly analogous to any particular Delphi Region. Adjustments have been made by using weighted averages of appropriate Delphi Regions. (U.S. De- partment of Agriculture, 1981.)

506 G. A. CARLSON AND L SUGUIYAMA
•H CO
»-< I 0) o Q ^
00 r>. O
es r>. lo vo o (jN vo es ^ m
es in r«. ^l- vO o
O o^ .-H O es
:5i -I---I en o NÍ in in
■<1-| VO
35-:: I '-' I
. 1 0) o W Q -H
cnl
t^ o 00
■•^-^ ^>». "^^
•<i-|in|vo|
CO ^1 c
o (0 0) -u
CO CO 0) -u
(U .u ^1 4J
(U (U
3 CO
p > CO r-< i-t
<U CO M Q o
•HO 4J 4-1 H w
rH I CO «H V4 r-l 0.1-4 e.H.-H«4-l PUO a to M Q Ou O -o
^ U3 3 3 a Ou
m o > 4-> c • o p .H o 00 CO 0) 'O •u WC XJ o (U <U o .u 4J C O-
14-1 X 43 o W 3
o.
CO Wl 4J 3 CO
r-< u CO
0) 4J > CO CO e
i Q
J= O o-z
(U (U a
u CJ 4J CO CO CO
PQ o P o (U . p-^ '-•
o CO g Vu (1) o AJ ta M e •H IM o o o
•H 73
CO M-l o
• 4J 4-> > CO CO CO «H (U o s M
■u u «H o.
I c
« s o. 6 4J g X 3 X 3 M CO M CO
^|es|^ cn|>i-fn|kO|

ECONOMIC EVALUATION OF CONTROL OPTIONS 507
Current insect control practices were not used in the EZ since all cotton acreage there was involved in the eradication effort. However, experts were able to estimate the number of insecticide applications, rates, and costs per acre for both zones (Table 5). These estimates are based on 10-year mean insect populations and 1979 chemicals assumed available. The expert estimate of insecti- cide costs of $53.42 per acre for the CZ is very close to the 1978 estimate obtained by the survey but about 22% higher than the 1980 estimate.
The same experts estimated that assuming long-run pest infes- tations, total private insect control costs per acre were $5.97 (or 12.50%) per acre higher in the CZ than in the EZ. This esti- mated difference is very similar to the historical 11.85% differ- ence in insect control costs during the base period 1974-77 ($56.83 to $50.81) shown in Table 2.
Finally, the public expenditures shown in Table 5 were com- puted by allocating extension budgets to the two zones based on the proportion of extension time and effort spent in each zone and di- viding them by the respective cotton acreages. The cost of scouting was estimated by contacting commercial scouts in the CZ and using extension estimates for the EZ.
ERADICATION
A description of the cost of eradication in the EZ and the cost of a hypothetical eradication program for the CV based on the same technology used in the EZ for 1978-80 is shown in Table 6. Most of the information was obtained directly from the eradication program personnel. (Target pests are listed in Table 4.) Only the insecticides actually used in Nash, Edgecombe, Northampton, and Halifax Counties are included and used in the calculation of the cost of control by chemicals. (This excludes the insecticides used in the buffer zone and about 1000 acres in other counties in the APHIS evaluation zone. A complete description of the insecticides used in the EZ in 1978-80 is available.)
In 1978, the eradication program included the chemical control of the boll weevil and also of other pests. Cost of this program (both insecticides and application) in the four eradication coun- ties was $42.25 per acre. Though this expenditure varied by area, the cost was evenly shared by all producers in the BWET area; that is, a charge of $46.61 per acre was assessed each producer, which was 50% of all APHIS costs. Total public (APHIS) plus private costs were $93.22; there was another $1.03 expended by the Exten- sion Service for a total of $94.25. The APHIS budget included some additional costs for research and capital items which were neces- sary for the program but were not charged to the producers. Total costs decreased considerably to $67.03 in 1979 and $40.63 in 1980.
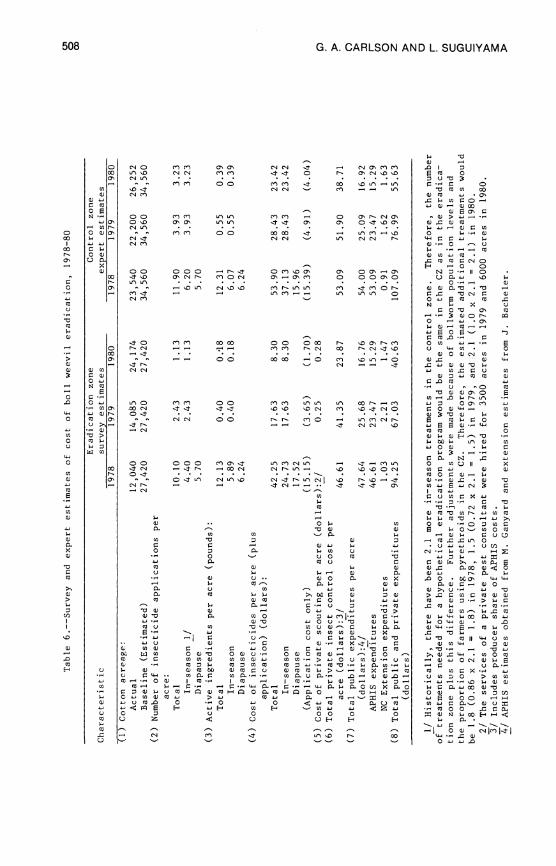
508 G. A. CARLSON AND L SUGUIYAMA
<N O ro ro o> ON CM CM <J- ^ es CJN OO OO Xi 1 3 m vo csl OO OO <l; <!■ o ON es vO v£> B tO O cNi m D o T3 5
m 00 o o 00 00 <t 00 vO in in C c vD <}■ es CN 00 in T) CO CO cN m
■u
CO
eu CO
0)
C O 0) 00 e ON
o o en OO in m OO OO o CJN r^ es C7N <u > 4-1 1—1 o v£> o> o> in in <|- >J- as ON o •vJ- v£) ON <U -c CU CO (N lO U 0) c
en OO O o cô 00 <J- in OO vO o VJ TH
CN <}■ OM csl in CM es r>~ U-t c c ■u <N ro
CO CO
o
■u CD
CO •
O O o o o r~v •<1- O OO vO ON ON o <3N ON M D •<t vO ON csl P^ OO O CM o ON OO O o O ON o O CL 4-) in in O
vO in Cv| vO v£> OO r^ in in OO <!■ 00 O r^ ¿ (U a x) • 00 •4- in OO in m in O c x: •^3 es csi en o
N c
•rH l CO
X ID
CU O
<t o f^ es
in o .00 es O <i-
o o <}■ csl O <t
00 00 o o OO OO
o 00 r>> CM 00
v£> ON 1^ 00 r>< CM <f v£>
o o 00 00 ^ o OO es
vO in '-• o
OO o r-H
•—' <}■ .—I ro in \D vX5 ^ o es
es <j- r>s. un CM I <t CM ^ ^ ..'
c a
P 4J CO -iH 4-1 o o .. XI CO U cu '-v c
c u cu^^ 4-1 cu X)
C 4-1 o o cu o 'H
C 4-> r-l <U X <í-|-.
cu .-I X3 C c o
OJ CO c o
I .-• y, ^ c • C D cu J-i U o 4J
4-1- U 4J- <d I
Q cu CO M Q o CL
0) CO CO >-i CO CL o D. I CO .r^
r-H C •'-' -I U-l CO M Q ex O
CO "O • CO CU
C r-í CO •rH ^ .-I CU <U ^ r-H
>-l CU 3 .-I 4-1 D .-I CLM D-OCO X CLO
U -O M W X3 I—I CO rH ^
CO CL, U CO 4-1 4-1 <¡ Z 4-1
o .-I 4J Jj (U r-H CO -4 —I 4-" e O e W C CO XI -rH C o CO 4-1 .—I • r-l o M-l CO •
O) o cu es CO CU x; <u X -u cu cu XJ J-1
X) cu cu CJN XI u r^
3 o C7N O cu U-J ^ :% X3 cu
CO »-" c e e 0) TH
»-I > ,_4 • rH cu c CL (SI X 4-1 o CO u II X CO e 4-) <U cu CO o C cu M CU CU x; OJ X) CO 1 CO
e 4-1
CM > c CO
c a CO c X 4-1 D •iH c XJ
XJ es CO 1-1 cu CO -O CO r^ 4-1 eo CO t-l u CO XI 4-1 Í>N
OJ O D CO c u O CO O CO CU u C o o
CO x: X in O O 4-1 4-t O C/D
es M (U M 2 4-1 P »-< 4-1 PC
C cu li, >. CO CL. cu -c aoo (U < cu 4-1 r^ CL P X o MON 14-1 U-l
0- CU c CU O cu >. o •H 4J XI > x: c CO C CO CU cu CO OJ 3 > u c X CO »-I CO
cu CO >-• x: CO cu >-i l4-< }-l 00 Cl- CO 4-1 >-l o u-< 5¿ X
>. cu , r-H (U
: ^ CO »I O
f—I '0 4-1
X o o
r-t «r^ vO
ex 4-1 00 >-i QJ CU cu XI CO D CO
■u Ë (U o o --^ M CO 4-t C CLs-^ cu o ÍC
•rH CO O O X C PH H M <: í-i 00
Du . C -H
^1 O <U CM |0O !<!• I

ECONOMIC EVALUATION OF CONTROL OPTIONS 509
In 1979 and 1980, APHIS expended eradication money only for boll weevil suppression and eradication. Scouting services for bollworm control were provided to farmers, and an additional Exten- sion Service employee was hired to assist farmers in their de- cisions regarding bollworm treatment, but EZ producers paid the costs of controlling bollworms. These expenditures averaged $17.63 per acre in 1979 and $8.30 in 1980, considerably less than the his- torical insect control cost (1974-77) of $50.91 per acre (Table 2). The APHIS assessment to each EZ cotton farmer for boll weevil con- trol was $23.47 per acre in 1979 and $15.29 in 1980. The Extension Service expended $2.21 per acre in the EZ in 1979 and $1.47 in 1980. Thus public plus private costs for eradication of the boll weevil plus bollworm control totaled $94.25 in 1978, $67.03 in 1979, and $40.63 in 1980. These amounts include the cost of pri- vate scouting since all spray groups did not charge for private provision of this service.
The costs of insecticides plus application for a hypothetical eradication program for the CZ were estimated at $53.09 per acre. Total private costs per acre (including 50% of total APHIS eradica- tion costs) were estimated to be $53.09 in 1978, $51.90 in 1979, and $38.71 in 1980. The CZ costs were higher than the EZ costs be- cause of the estimated 2.1 additional bollworm treatments per acre (Tables 2 and 6) (11.13-9.03 = 2.1). Total costs per acre were es- timated to be $107.09 in 1978, $76.99 in 1979, and $55.63 in 1980. These estimates are higher than the actual costs in the EZ and also higher than the most recent APHIS eradication estimates of $75.55 per acre for a 2-year eradication program (Brazzel 1980).
ACREAGE ADJUSTMENTS TO ERADICATION
One important feature that complicates comparisons over time and space is the movement of land in and out of cotton production as a result of implementation of an eradication program. Cotton producers have production inputs and profitable alternative crops to cotton. They therefore may elect not to participate in eradi- cation by switching their land out of cotton production. Rodriguez (1980) showed that 42.3% of all cotton farmers in the EZ stopped producing cotton between 1977 and 1978, the first year of eradica- tion. The comparable figure for the CZ was only 17.7%.
Analysis of cotton acreage in the EZ from 1956 to 1980 was conducted in an attempt to refine and further quantify the effect of eradication on acreage. A statistical and economic model was designed to reflect the options of the farmers to switch crops and thus avoid paying fees for boll weevil control. The fees per acre of cotton planted were $50.50 in 1978, $24.00 in 1979, and $20.00 in 1980. 2/ The fee in 1978 covered both boll weevil and bollworm control; it did not include bollworm control in 1979 and 1980.
2_/ These were fees charged at the time of planting decision. Re- bates were given at the end of each crop year. A separate analysis in which fees less rebates were used produced very similar results.

510 G. A. CARLSON AND L SUGUIYAMA
The model for the eight major cotton counties in North Caro-
lina was: _3/
A. = a + X.d. + a,P^. + a«P^. + ^-{^4*- It o .11 1 et 2 st J It 1
where A. = natural logarithm of county cotton acreage in
year t, in county i,
d. = dummy variable, 1 for county i and 0 for other counties,
P = expected cotton price or deflated futures price at cotton planting time in year t.
St expected soybean price as a competing crop or deflated futures price at planting time.
Y. = average cotton yield for county i in previous 4 years,
RPC = price risk as reflected by percent deviation of futures price from actual cotton price.
It = rainfall index in planting season (rain days,
inches of rainfall).
E. = eradication payment in county i and year t as given, and
e. = random disturbance term It
The model was evaluated by ordinary-least-squares and by generalized-least-squares by using the Fuller-Battese method. The model was also evaluated for the period 1956-78 so as to measure only the effect of the first year of eradication.
The result of this analysis showed that the direct fee for eradication was statistically significant (1% level) in reducing the acreage of cotton over the 3-year period. The effect in the first year of eradication was especially large: about half the acreage reduction occurring between 1977 and 1978 was associated with eradication. The other half reflected the effects of yields, prices, rainfall, and other factors. Consequently, if eradication fees were raised by $10 per acre, each county in the EZ would re- duce acreage by about 15% (from the geometric mean of 12,600 acres) for the 3-year period. In fact, without the large negative effect
3/ A complete justification of the model and statistical methods is^given in Rodriguez (1980). However, his estimates were for 1978
only.

ECONOMIC EVALUATION OF CONTROL OPTIONS 511
of the first year of eradication, the fee still had a negative ef- fect in the last 2 years of the eradication trial. The temporary effect of eradication fees on acreage can be seen in a general way in the time trends of Figure 1.
Further analysis was conducted to determine what characteris- tics were associated with farmers who participated in eradication. It was found that farmers with more cotton pickers, farmers who had been members of community cotton insect control groups, and farmers with higher cotton yields were most likely to participate in eradi- cation (Rodriguez 1980).
There are two major policy implications of the acreage shifts due to eradication fees. First, charging farmers directly for the program can reduce the number of acres requiring eradication, thereby reducing variable costs of eradication below those based on average cotton acreage. Second, the potential for temporarily ad- justing acreage out of cotton will differ among farmers and lead to lower private costs of insect control during the eradication imple- mentation period. This is a temporary gain that probably would not have any considerable effect on long-run costs and returns of eradication.
YIELD CHANGES
Yield effects of the eradication option relative to CIC were difficult to assess. Cotton yields vary greatly from year to year in North Carolina (Table 7; Figure 2). For example, the standard deviations of yields for 1970-80 were 91.20 and 73.80 pounds per acre for the EZ and CZ, respectively. Yields per acre in the per- iod 1974-77 were higher in the EZ, though they were higher in the CZ for 1965-75. Weather, particularly rainfall and frost, affects yields. Also, in recent years, there appears to be a sizable ef- fect of farmers with lower yields who discontinue cotton produc- tion. This discontinuance is shown as a 15.6% yield advantage (1974-77) for those farmers in the EZ who continued cotton produc- tion in 1978 or 1979 (Table 2). The equivalent figure for the con- trol zone is 16.7%.
Biologically, one could expect a gain in yield from boll wee- vil eradication from: (1) reduced use of organophosphate insecti- cides to control early-season boll weevils; this leads to earlier maturity of cotton, permitting avoidance of frost and late-season rain effects; (2) reduced boll damage from bollworms that are con- trolled by populations of natural enemies which are not reduced by early-season boll weevil treatments; and (3) reduced direct damage to bolls from boll weevils.
Analyses of various data sets have not provided any direct measure of effects of reduced early-season boll weevil populations on yields. One indirect measure has been the effect of delayed applications of insecticide on yields. Table 8 shows the results
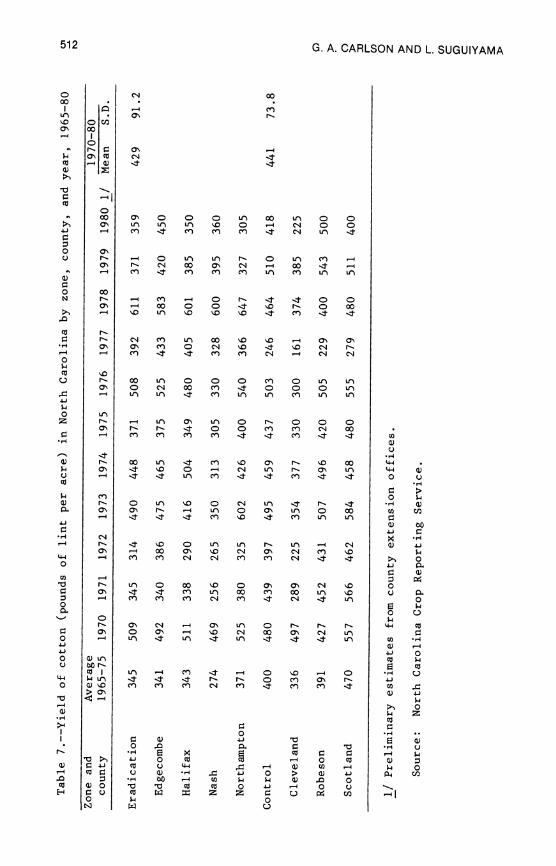
512 G. A. CARLSON AND L SUGUIYAMA
o 00
I LO VO 0^
U CO
C «O
tí o N
td tí
o u Cd
o
U O Cd
u 0) cu
-o tí O
o o
O
T3 1—1 (ü
•H PH
I
r>.
O) rH Xi
Cd H
CM
0^
Cd I u m > ON
<3 —I
tí 4J Cd tí
<D O tí O O
ON o o m CO
o VO CO
in o CO
00 1—1
m CN CN
o o m o o
r—1
CO
o 00 CO
m ON CO
CM CO
o 1—1
in
m 00 CO
CO
in in
PH
t—1 vO
CO 00 o
o o vO vo
VO CO
o o o 00
es ON CO
CO CO o
00 CN CO
vo vO CO
VO
CM
1—)
vO 1—1
ON CN CM
CJN
CM
00 o CM o 00
o CO CO
o m
CO o m
O o CO
in o in
in m in
CO CO
ON
<!■ CO
in o CO
o o CO o CO CO
o CN
o 00
00 o m
CO 1—1
CO
vO CN
ON in
CO
vO ON
00 in
O ON 1—1
o in CO
CN o vO
in ON m
CO O in
00 in
t-H CO
00 CO
o ON CM
in vO CM
m CM CO
ON CO
m CN CM
1—1
CO CN vO
uo
CO
o CO
00 CO CO
vO in CM
o 00 CO
<3N CO
CJN 00 CN
CN m
VO VO in
ON o UO
CM CJN <1-
rH
UO
CJN in CM in
o 00 <7N CN m
in
CO
i-H
CO
CO
CO CM
rH
CO
o 5
VO CO CO
ON CO
o
tí <v 4J 'O O Xi a tí T3
•H B X Cd tí tí 4-i o Cd Cd I—t o Cd Cd o M-l Xi rH 0) CO 1—1 O Q) •H ^ 4-> O > <D 4J
• H W) rH CO U U Q) Xí O TJ Ta Cd Cd O 4-> 1—1 O O
Cd w ffi Ä Ä tí U çxi CO u O w o
Q) ü
•H M-t , 4-1 <D O O
•H tí > O ^1
•H 0) en œ tí Q) Ö0 4J tí X •H OJ 4J
»-1 Í>> O 4J CU tí cu 3 tó O o cu
o B u o o u
<-M cd tí CO •H Q) 1—1 4J O Cd u e Cd
•H o •U CO x 0) ■u
u >. o ^1 ÍS Cd tí
•H • • B <u
•H u I—1 u <u tí »-i o
CíA CO
^1

ECONOMIC EVALUATION OF CONTROL OPTIONS 513
Table 8.—Effects of biological and economic variables on per acre cotton yields, Rocky Mount, NC, experimental data, 1967-78
Independent variables Coefficient ^/ t-Statistic Mean
Intercept 8776.63 2.35 Pounds of insecticides 18.18 4.03 13.97 Julian date of first
treatment 21.33 3.61 210.00 Julian date of last
treatment -46.32 -3.76 252.00
Chlordimeform use 512.66 2.92 0.24
August rainfall (inches) (August rainfall)
325.47 1.95 4.37 -29.55 -1.75 22.40
Check plot % damage ^ (Check plot % damage)
-53.99 -2.81 40.50 0.42 1.82 1922.00
Dependent variable: Seed R^ = 0.75
cotton yield per acre 1328.00
Sample size = 38 plots
1/ The pounds of lint cotton change per unit change in each inde- pendent variable.
Source: Data in Table 3 plus experimental plot data provided by J. R. Bradley, North Carolina State University, Raleigh.
obtained by analyzing 38 experiments at Rocky Mount, NC, during the period 1967-78 (Table 3). In this case, the data for the experi- mental plots were adjusted for five other factors by multiple regression.
The analyses showed that each additional day of delay in in- secticide treatment, from a mean date of July 26 (Table 3), added about 21.3 pounds of seed cotton to yield; that is, yield was in- creased by about 1.6% per day (mean yield per acre 1328). There- fore, a 10-day delay in the initiation of treatment would increase yields by about 70 pounds of lint. The Delphi panel estimated a gain of 21 pounds of lint due to eradication for northern North Carolina (U.S. Department of Agriculture 1981); therefore, this would require a delay in the initiation of bollworm treatments of only 3 days on the average.
However, it has not been shown in a statistically valid manner that boll weevil eradication will delay the date when the popula- tion of bollworms reaches a treatable level in North Carolina. The best indirect information seems to be the approximately 15-day de- lay in bollworm treatments for about 50% of the EZ cotton fields in 1978-80. This is further complicated by the fact that most boll- worm populations arrive in North Carolina cotton fields by mass migration from other crops.

514 G. A. CARLSON AND L SUGUIYAMA
The use of new types of insecticides in recent years has also complicated yield assessments. For example, in the Rocky Mount plots, the use of chlordimeform was associated with increases in yield of 513 seed pounds of cotton. However, it is very possible that the use of chlordimeform may have been correlated with un- observed variables in the experimental data. Pyrethroid insecti- cides have probably had similar unidentified effects.
Actual yields in the EZ were 611 pounds per acre in 1978, 371 in 1979, and 359 in 1980 (Table 7). Producers who grew cotton in 1978 or 1979 had a 1974-77 average yield of 497 pounds (Table 2). Only the 1978 yield in the EZ figure is statistically higher than the base year mean yield. However, the weather was especially favorable for cotton production in 1978. Also, insect scouting was especially good, and chemical selections for bollworm control were especially effective in the EZ in 1978. These factors were not specifically a part of the eradication program. In any case, when one compares the 1978 yields in the EZ and the CZ, it is clear that cotton yields can be increased by effective bollworm control. The 1978 yields were 114 pounds per acre above the 1974-77 yields for farmers growing cotton in 1978 or 1979 in the EZ, but in the CZ they were 16 pounds per acre less.
SUMMARY AND POLICY IMPLICATIONS
Some of the major characteristics of the two North Carolina cotton zones are listed in Table 9, which reports the economic im- pact of the BWET on cotton producers in 1978-80. Both zones showed a trend to reduced producer insect control cost; however, the re- duction was larger in the EZ than in the CZ. Both zones also showed lower insect control costs in these 3 years than in the pre- vious period (Table 2).
Public insect control costs were much higher in the EZ than in the CZ. This is the nature of an investment in which early costs are high and future costs are low. For example, if the average 1979 and 1980 insecticide (and application) cost saving between the EZ and the CZ is a permanent benefit, this is a $30.70 saving per acre of production costs. Even if there was a $5 per acre main- tenance cost (for buffer zone, monitoring, and boll weevil suppres- sion), there would be a net gain of about $25 per acre iñ perpetu- ity. A $25 net gain per acre would return the investment cost of $127.62 expended for eradication in 1978-80, plus a return on the investment of about 20%. If the costs of eradication were only $75 per acre as estimated by Brazzel for a 2-year program (Brazzel 1980), then the rate of return would be 33%. These estimates assume no changes in yield, acreage, or price from the eradication program.
The yields for both zones are compared in Tables 7 and 9. Yields were highly variable in both zones, and it was not possible to detect any changes due to the eradication program.

ECONOMIC EVALUATION OF CONTROL OPTIONS 515
00 +J tí r-4
•H tí CO »O »H 0)
ONCSCMcn<fcom CN CM 00 cs tí ÏE!
O (30 ON
^moooNr-iincs ON
^ csi • « vO es CM 00 CO ^
vO CO <t CM
O -H C3N
CO cs CO
1—1 CD
O V4 -u tí (ü tí
•H O CO
na (^ CO O
0) u u ^ CO cu tí
W) c
•H u
o OOOOCMOOOO 00 CS o ON QJ CO
Om-I 1—1 1—1
M CO CO
Q) B Jtí >^-H 4-> JQ tí
ON
O ON U -H 4J a o
f-iO«^vOCM<l-0 ^ ^ es
•» vO CO m 00 CO vD (N CO <t CM
CO —1 • m
1—)
CM
CO 00 CM
CO o <í O V-i 13 00 (U o TJ <ü II ON o
o }^
Cl.
moooooocoo CO -I >;f 1—1 tí í-t CO CO CA
r-l CO Pd
1—1
(30
o\ 1—4
.-H<l-cococor^m m ,-H in • • • • •
•» cjN o CO '-t €^ r^ CO >—< -^t •—< "^ CM
ON v£>
O 00
CM
u o
M-l
<ü 1-1 CO c Q) tí ■u o
> u tí (U
1—1 1-1 (U (U o CO r-4 o
•H o CO tí 00 }-t XI CO Tí 00 H ^ o
•2 a JJ CO CÜ )-i
ON<i-cooooooooN r^ vD CJN m 0) ^1
X! cu tí- 4J >H
tí Jtí M-l 4J o O «H CO
Tí 0)
13 tí CO
C3^
O C30 ON 1—í
<ü
mr^<—<•—ivDr-^cMcM oo cs ^
«r-nOvOi—<Om CO ^ ^ CM cs
r^ in . CO
vD 1—1
o CNI
ON cs
•H O tí !-• tí C3N
"^ ^ O o dJ o 1—1 CO CX
1-1 4-i •H > M-l <ü O
N í-l pu 4J
C O C3^
4J <3N CO ^ O
comcoooouomr^ m 00 -H ON 4J X u o
M-4
CO 4J tí
OOOO'^'^fONvOCM'd- CO .-HO *
•»csococooco »-H <!■ r-H CM <f 1—1
. CO in cs
vO
00 ON 1—1
tí Q> "TJ o Oí o CO CO
•H CO -tí ptí <ü (U H CO
5 « •H
CO
no CO tí CO
1—1 •
(U O
1-1 CO r—t d)
U ONOOcovoin 1—1 «—• <!■ ^ in U CO CO
tí CD CO •H «H 1^
4J
r>* O >^
pq CO
00
ON
r-^COrHi-l^l-'-l ^ ^ ^ o • • • •
•« o es 1—1 in vD vO
vO —1
1—t cs •—!•—• CO 1—1 <f <l"
-1 <f in
CO CO «H 0) 4-» > M CO «H O O 4J CO
00
4-1 O O CJN
M-l O
CO
CO V4
ß - 9Í 1—1
U
O
CO
.g
^ ^1 to ^^ /^ CO
T3 CO CO 1-1 C! ÍH »-I i-t
CO e cu U CO Ö0 í-l -H o öOvO
u o
M-l
p CO CO /-^ O V^ M o • CO
O r-l 1—1 CO 13 (U cu V4 vO 4J tí d,^ ^ J-l ^X CL, V^ p-<t
O •H
\^^ o o to <U 4J tí <o- Q)
OJ no TJ «-^ ••-> ■*-* CU ^ " "^1
O J-l O
o § Ü
VH 0) ^-^ ^«-^ rH CO CO CO Q) cs •H d) M-l
O Í-4 o o o J-» TJ CO 4-) ^ o vO CO O <L) <U T) O O CO tí tí ^-^ CO 4J vD
cO }-i }-t ^»^ o tí .H CO o o (U J-i O O 0/3 1—1 o o )-l •H 00 (U (U V^ «0 CO (1) M O cu U CO T3 CO V4 i-I
CU (U í-i pí }-i rH N--' QJ 1-1 CO tí CO (U 1 i ÇX, u u u pu -u
CO O) <u CO <3 tí O o .-• V-i 1-1 X
1 u <u tí o (U CU u • <3^ c CO o, ei4 o
o J-í J-i M-l o tí o o w QJ ^-N < cu o
CO
<ü CO •HC-t-»-»-»<l>0 ^"^ o ^^ CO Wl -^ '::! •H Ü
}^ 4J(UC0C0PLI 4JC0 o CO cu CO 4-) O > •H
CO
H OoOO'OOO-MV^CÜíO *J CO 0)
4-t (U CO •« 4J CO <u
4J ;3C0«H<U COCOCOi-4 u 1-4 ex > O 00 tí cu C/3 tí 0 TjOir-HÍ-lQítíO^tíT-t cu rH O o r- o V4 • r- Oi-iO-OOTJOOCO'HO CO o T) ^ CJN U CO tí o
v^ J^ O O. tí 'H «H Tí tí T? r-l CO 1-1 1—1 o 4J
Puco CO'H o-U W)í-< V-i^^-^ •r-l CO tí <U 0)
M-ltí'+^CU-MO'HOOQÍ
•H v_x O) o CO U
tí •!-> tí C w 4J
o
4->
O
ci 000>0*H4-»3íí>-t •H M J-l tí !^ PQ 9^ Cl
4J •H(Ur-l';3'T3'T30 1—1 o 4-» tí o ^^ 1—1 cu ^■^
, U .+JCOOuOOOCO u:i <o tí 4-» O ^l'::^ 2i CO CM
é OOOOtíO^O»-i>-i > ^Ua<ÍM<C/3Cl^fX,
tí -H ^ o .c Xi J-»
tí M

^"•^ G. A. CARLSON AND L SUGUIYAMA
In the EZ, the presence of eradication personnel and the ab- sence of need to treat for boll weevils reduced insecticide use relative to use in earlier periods and to use in the CZ. Also, eradication personnel tried to keep eradication costs and farmer insect control costs low. Partly as a result, few bollworm treat- ments were necessary compared with those seen as necessary in pre- vious years (Table 4). In addition, farmers in the EZ became more aware of the need for scouting and prescriptive treatment. This awareness is growing in other parts of North Carolina but at a slower rate.
Finally, the tabulations and comparisons of cotton insect con- trol costs discussed in this chapter relate to a particular region of the Cotton Belt. They must be viewed cautiously in respect to other areas. Then, too, the farmers who participated in the BWET normally have higher yields and more specialized cotton equipment than the average northern North Carolina farmer. The insecticide use changes were applicable to particular weather and insect popu- lation situations. Economic evaluation must thus be supplemented by environmental and biological evaluations.

ECONOMIC EVALUATION OF CONTROL OPTIONS 517
LITERATURE CITED
Brazzel, J. 1980. Boll weevil eradication plan, third draft. U.S. Depart- ment of Agriculture, Animal and Plant Health Inspection Ser- vice, Raleigh, NC, November 1980.
Grube, A., and G. Carlson. 1978. Economic analysis of cotton insect control in North Caro-
lina. North Carolina State University, Department of Economics and Business, Economics Information Report No. 52, 54 p.
Rodriguez, R. 1980. Acreage response to government pest control programs: the
case of boll weevil eradication. Unpublished Ph.D. dissertation. Department of Economics and Business, North Carolina State University, Raleigh.
U.S. Department of Agriculture. 1981. The Delphi: insecticide use and lint yields. Beltwide boll weevil/cotton insect management programs. USDA Economics and Statistics Service Staff Report, March, 142 p.
Von Rumker, R., G. A. Carlson, R. D. Lacewell, R. B. Norgaard, and D. W. Parvin, Jr.
1975. Evaluation of pest management programs for cotton, pea- nuts, and tobacco in the United States. Contract Report for Environmental Protection Agency and Council on Environmental Quality, 589 p.


SECTION V FUTURE PROSPECTS


521
Chapter 20
OPPORTUNITIES FOR IMPROVING COTTON INSECT MANAGEMENT PROGRAMS AND SOME CONSTRAINTS ON BELTWIDE IMPLEMENTATION
R. E. Frisbie Integrated Pest Management Coordinator Texas A&M University College Station, TX 77843
J. R. Phillips Entomology Department University of Arkansas Fayetteville, AR 72701
W. R. A. Lambert Entomology Department University of Georgia Tifton, GA 31794
H. B. Jackson Plant Pest Regulatory Services Clemson University Clemson, SC 29631
ABSTRACT Cotton pest management programs that have been demonstrated as effective in Texas, Arkansas, and the Southeast include, respec- tively: (1) a short-season management system for cotton production, (2) community-wide boll- worm management, and (3) improved cotton insect management. Regional strategies developed for each of these geographic areas consist of com- binations of techniques that have proved to be economically and environmentally sound. An updated review of these three systems is presented.
Specific tactics for suppression of boll weevils (Anthonomus grandis Boheman) such as early-season insecticidal control of spring emerging weevils, fall diapause control pro- grams, the role and efficacy of diflubenzuron (Dimilin), the role of alternate hosts, and the impact of in-season application of phosphate-type insecticides on beneficial

522 R. E. FRISBIEETAL
arthropods are reviewed. The merits of these individual tactics are compared with those of tactics employed in the Boll Weevil Eradi- cation Trial conducted in North Carolina and Virginia, 1978-80.
Logistical constraints to beltwide imple- mentation of a boll weevil eradication or man- agement program are discussed. Constraints considered are cost of implementation, person- nel needs, the role of independent consulting entomologists, conduct of an areawide insecti- cide spray program, maintenance of weevil-free areas and containment zones, and Federal and State regulatory requirements for such programs.
INTRODUCTION
In this chapter, pest management programs that differ slightly from the programs of the Boll Weevil Eradication Trial (BWET) and Optimum Pest Management Trial (OPMT) of 1978-80 or that are alter- natives to those programs are discussed. The emphasis is placed on three: (1) a short-season management system for cotton production that has been tried in Texas, (2) a community-wide bollworm manage- ment system for cotton production that was tested in Arkansas, and (3) an improved cotton insect management system for cotton produc- tion that could be used in the Southeast. All are considered to be improvements on insect management programs currently available in these areas. In addition, discussion of these systems makes it possible to formulate other possible alternatives. Many of the tactics set forth in the three systems were used in the OPMT con- ducted in Panola County, MS, and in the BWET conducted in North Carolina and Virginia. It is not assumed that all are applicable to all regions of the United States where cotton is produced. How- ever, review of the tactics broadens one's perspective on other candidate management systems designed for optimal pest suppression.
The second section consists of certain biological inquiries relative to beltwide implementation of the programs used in the BWET and OPMT. These inquiries are not meant in any way to indi- cate criticism or to supersede observations and conclusions of the agencies and personnel involved in these trials nor the report of the Biological Evaluation Team, the Environment Evaluation Team, and the Economic Evaluation Team. All are to be heartily congrat- ulated for the insight and thoroughness with which they carried out their tasks, sometimes under difficult conditions. However, it is of utmost importance that an indepth evaluation be made of the techniques developed, particularly those that would be used for eradication, as they relate to the various cotton production areas that are infested with boll weevils (Anthonomus grandis Boheman).

MANAGEMENT PROGRAMS AND IMPLEMENTATION 523
For example, optimum pest management is essentially defined by research, extension, and farmers within a certain State or region, and each such group must assess cotton growth conditions and pest situations so an optimum combination of management techniques can be developed. In other words, optimum pest management must vary State by State. In contrast, the techniques used for eradication as in the BWET would have to function as well in all cotton pro- ducing areas. Comments and discussion are therefore provided con- cerning earlier experiences encountered beltwide in testing the various tactics tested in the BWET for beltwide boll weevil eradi- cation. It is recognized that such research tests did not expose boll weevil populations to the areawide suppression achieved in the BWET. Nevertheless, studies of the strengths and weaknesses of the various tactics in the several cotton producing areas are still helpful in gaining more complete knowledge of the suppressive values. The fact that these tactics were not employed sequentially and over a large geographic area is not in itself sufficient reason to reject the information. For example, it is necessary to estab- lish a complete profile of all studies of such tactics as early- season insecticidal control of spring emerging weevils, fall dia- pause control, the role and efficacy of diflubenzuron (Dimilin), and the role of alternate hosts. (Since many of these studies are discussed in previous chapters of this handbook, efforts have been made to minimize duplication.)
Finally, a section on the logistical constraints on implemen- tation of a beltwide program is provided as a means of reviewing the strengths and shortcomings of the draft plan for boll weevil eradication prepared by Plant Protection and Quarantine, Animal and Plant Health Inspection Service (APHIS), U.S. Department of Agri- culture (USDA), in September 1980. It is recognized at the outset that this plan is in the draft stage and subject to alteration. It is hoped that the discussions will provide insight and direction regarding the proposed use and outcome of the boll weevil eradica- tion plan and needs for improved cotton insect management.
ALTERNATE MANAGEMENT SYSTEMS
Short-Season Management System for Texas Cotton Production
Although the role of breeding for insect resistance, modified cotton culture, and the value of short-season cotton production were discussed in a previous chapter (Chapter 4 of this handbook), pertinent literature and evaluations of short-season production systems in Texas are highlighted in this chapter.
The decade of the 1970's is notable as the period when the mystery of the importance of the "short-season effect" of cotton production relative to boll weevil suppression was described and

524 R. E. FRISBIEETAL
finally resolved (Parker et al. 1980). Only after a painstakingly careful review of past research inquiries, trial and error, and an exhaustive study of the intricate relationship between the fruiting pattern of cotton and the population dynamics of the boll weevil were principles set forth that define and elucidate the breeding of cotton cultivars to enhance escape from boll weevil populations. The basic information concerning this relationship arose out of studies by Walker and Niles (1971). These scientists, who were working in recent times, had an advantage that only history can provide re the confrontation of U.S. cotton producers with the boll weevil since the turn of the 19th century. Although Howard (1896) provided early information on the biology, ecology, and control of the boll weevil, Mally (1901), the first entomologist of the Agri- cultural and Mechanical College of Texas, was assigned the task of solving the boll weevil problem after the boll weevil invaded Texas and devastated the cotton plantings in that State. This scientist reasoned quite correctly that it was the cotton plant itself and its growth characteristics that made the crop vulnerable to the attack of the boll weevil. Cottons of this period fruited in a slow and prolonged fashion that allowed a progressive increase in the numbers of boll weevils in succeeding generations and extended the period of plant susceptibility to weevil damage. After study- ing the matter during several production seasons, Mally noted that the plant and its fruiting characteristics were the keys to man's successful cohabitation with the boll weevil. He stressed a need for early maturing cotton varieties and for areawide destruction of cotton stalks at the end of the season so as to disrupt the normal behavior of boll weevils preparing for winter. Mally's recommenda- tions were considered radical in view of the relative unavailabil- ity of early maturing cotton varieties. Also, mechanical equipment capable of dealing with the large numbers of acres requiring stalk destruction was then in a primitive state. In addition, several months were required then to hand harvest the cotton crop.
Mally's sage advice was ignored at the time; only after some years did cotton breeders finally begin to select cotton genotypes that possessed an early fruiting characteristic (Ware 1925). In fact, it was the development of cotton varieties that possessed this unique ability to commence fruiting earlier and to develop larger numbers of fruit in a shorter time that allowed cotton pro- duction to survive in the weevil-infested areas of thé Cotton Belt. Growers could stay in business though production was limited to the first few weeks of fruiting before the large increase in numbers of weevils.
Still, something else was needed because the effective produc- tion period was limited to early fruit set. The chlorinated hydro- carbon insecticides that came into use after World War II seemed to be the answer. With them cotton now could be produced by planting long-season indeterminate varieties and could be protected from boll weevil attack by using these broad-spectrum insecticides. This alteration in the type of varieties grown plus increased fer- tility plus irrigation dramatically increased yields. Cotton

MANAGEMENT PROGRAMS AND IMPLEMENTATION 525
production throughout the rain belt was therefore limited only by the length of the growing season and not by how much could be pro- duced before boll weevil populations reached damaging numbers.
The respite was nevertheless short-lived. By the 1950's, the boll weevil had developed resistance to chlorinated hydrocarbons (Roussel and Glower 1955). Growers responded by using the newer organophosphate insecticides, sometimes mixed with organochlorines. Again, the weevils were neatly controlled. However, the tobacco budworm (Heliothis virescens (F.)) developed resistance to these compounds in the I960's (Adkisson and Nemec 1966). It was also during this time that the cost of insect control and the uncertain- ty concerning effectiveness of compounds created great concern among Texas cotton producers. As a result, the stage was set for a reordering of the management of insect pests in Texas cottons.
At this point Walker and Niles (1971) elucidated the value of rapidly fruiting short-season cottons for managing the boll weevil since use of these cottons precludes the necessity of treating season long, which enhances the potential for population increase of the bollworm (Heliothis zea (Boddie)) and the tobacco budworm by virtue of the insecticidal destruction of beneficial arthropods. The principles set forward were essentially two and are as follows: "1. Precocity and rapidity of fruiting increases the probability for setting an acceptable fruit load before weevil infestations reach a level that will reduce yield. 2. Early and rapid crop ma- turity permits early disposal of crop residue, thereby reducing host plant support for diapausing weevils and contributing to a re- duction in overwintering populations of the pest. The crop may be harvested and stalks destroyed before the environment forces the weevil to diapause. This would be disastrous to the boll weevil and might relegate the insect to the role of a minor pest."
Although these principles were developed around an experimen- tal cotton genotype, it was only a short time before commercial varieties were available that possessed a rapidly fruiting, short- season characteristic. In fact, the Texas A&M University-Multiple Adversity Resistant (TAM-MAR) cottons were released as commercial varieties in the early 1970's. Subsequently other commercial cot- tons were developed and released that possessed a rapidly increased fruiting characteristic. These cottons provided a distinct advan- tage in the management of the boll weevil. The question was why. The stage was set for a study of the reasons for this advantage.
Although it was generally accepted that the newly termed "short-season cotton production system" was economically advanta- geous, factors other than cotton genotype entered into this manage- ment system. Parker et al. (1980), from work in the late 1970's, finally pieced together the last part of the puzzle as to why the early, rapid fruiting of cotton served to limit damage by the boll weevil. In a detailed set of experiments in Falls, Frio, and Uvalde Counties of Texas in 1975 and 1976, Parker et al. (1980) studied the relationship between the fruiting pattern of short-season

526 R. E. FRISBIEETAL
cotton varieties or indeterminate types and susceptibility to boll weevil damage. The objective of this study was to develop a de- scription of cotton production in the presence of the boll weevil and to achieve an acceptable lint yield (500-750 pounds per acre) without the necessity for late-season application of insecticides. Their results showed clearly that if damaging populations of boll weevils were suppressed long enough to allow for 30 days of bloom- ing, acceptable yields could be expected. Also, study of the sus- ceptibility of bolls to boll weevil damage revealed that a popula- tion of 10-day-old and older bolls had a high probability of escap- ing damage. Parker et al. (1980) demonstrated that where numbers of boll weevil punctured squares were kept below 1000-1500 per acre during the period the cotton was in the prebloom stage, a signifi- cant delay in subsequent population buildup could be achieved, thereby allowing the critical 30 days of blooming to occur and as- suring an adequate yield. The salient point here is that if a fast fruiting cotton is allowed to fruit without encountering an unrea- sonable level of damage from boll weevils through about 30 days of blooming, bolls will have developed to the point (10 days or older) where they are relatively- safe from the attack of later developing weevil generations. In this experiment, treatments with one and three insecticide applications were applied near the time of ap- pearance of the first one-third grown square to keep populations of overwintering boll weevils low during the critical blooming period.
There are several earlier examples of elaborations of short- season cotton production systems in various parts of Texas (Namken and Heilman 1973, Sprott et al. 1976, Heilman et al. 1977). These also involved alterations in plant varieties and row configuration, reduced fertility, and reduced irrigation rates to minimize vege- tative growth and prolong the fruiting cycle, all of which served to expand the short-season effect and make the system more robust.
Today, short-season cotton production, as it relates to inte- grated pest management (IPM) in Texas, consists of two major strate- gies. The first, the suggested and commonly used practice in the Central Blacklands, Central Texas Coastal Plains, Winter Garden, and Lower Rio Grande Valley, includes: (1) early and uniform plant- -ing of a short-season cotton variety for optimum yield; (2) reduced application of nitrogen and irrigation where appropriate; (3) in- tensive field sampling (scouting, pheromone trapping, Heliothis spp. M0TH-ZV3 forecasting) for early-season pests such as the cot- ton fleahopper (Pseudatomoscelis seriatus (Reuter)) and overwinter- ing boll weevils so they can be controlled if damaging levels are anticipated; (4) continued intensive field scouting of key insects and application of insecticides based upon appropriate action levels; (5) use of a harvest aid chemical for desiccation or de- foliation; (6) early harvest; and (7) complete and areawide stalk destruction soon after harvest. This system of short-season cotton production has, in effect, eliminated the need for midseason and late-season insect control in most areas.

MANAGEMENT PROGRAMS AND IMPLEMENTATION 527
The second major strategy involves a cotton production system used primarily in the Rolling Plains of Texas. The spring emer- gence profile of the boll weevil in this region is predictable from year to year (White and Rummel 1978). Thus uniform delay of plant- ing means that cotton is only marginally suitable for feeding and is not suitable for reproduction for the majority of weevils emerg- ing from diapause and overwintering habitats in the spring. As a result, a high degree of suicidal emergence results. The proce- dure has received wide acceptance in the Rolling Plains (Boring 1974, Slosser 1978). As a result, uniformly delayed planting, in- tensive field sampling, selective use of insecticides on small acreages when one-third grown squares first appear, and occasional use of a fall boll weevil suppression program in areas of intense weevil activity generally constitute the short-season cotton pro- duction system used in the Rolling Plains. Thus the uniformly de- layed planting, in effect, acts as a cultural early-season control measure comparable to the early-season insecticide control used in the southern regions of the State.
Extensive economic and environmental evaluations have been conducted to determine the impact of the strategy of the sh.ort- season cotton production system used in the southern part of the State (Namken and Heilman 1973, Sprott et al. 1976, Heilman et al. 1977, Lacewell et al. 1977, Frisbie et al. 1978, Masud et al. 1981). All have shown common threads of consistency. For example, insecticide use has diminished greatly, and yields have increased. A basic philosophy of the management of cotton insects has there- fore emerged from this short-season production system. Essential- ly, insect control relies on varietal and cultural control measures as primary inputs, intensive field sampling, and selective applica- tion of insecticides (primarily in the early fruiting stage), and areawide, postharvest stalk destruction. For the most part, farm- ers generally make a maximum effort not to engage in midseason to late-season insecticide applications for the suppression of insect pests.
Perhaps the best example of areawide implementation of the short-season production system is found in the Coastal Bend of Tex- as. A decline in cotton acreage in San Patricio and Nueces Count- ies that was observed beginning in 1970 (104,900 harvested acres) bottomed out in 1975 (50,400 harvested acres) (Masud et al. 1981). Then with the introduction of rapidly fruiting and early maturing cotton varieties (TAMCOT-SP37, TAMC0T-SP21, and others) in the mid- 1970 's, a turnabout in harvested acreage was observed (236,500 har- vested acres in 1979). Presently, in excess of 80% of the cotton varieties planted in the Coastal Bend are the short-season type. Today, the use of short-season varieties in combination with the IPM program previously described has produced a current areawide annual benefit of $29 million with a potential increase of $41 mil- lion for the Coastal Bend area (Masud et al. 1981), in part because of decreased costs of production and increased yields.

Ö28 R. E. FRISBIE ET AL
In the case of the second strategy, delayed uniform planting,, the reasoning once again is sound. Weevils that emerge from over- wintering in the spring must find a suitable food source and ovi- position sites. When the cotton plant has not begun to fruit, nutrition is generally poor, and reproduction is not possible; thus, the boll weevil population declines dramatically. The few .weevils that survive do not increase their numbers until late in the sea- son after the crop is substantially made.
Results from a Fisher County, TX, uniform delayed planting demonstration were quite revealing (Finley and Boring 1978). Prior to the initiation of the study, that is, from 1974 to 1976, an aver- age of 2.1 more applications of insecticide were required in 10 fields with a history of heavy boll weevil infestation than in 10 fields with a history of light boll weevil infestation. However, immediately after the study began in 1977, only 0.3 more applica- tions were required in the fields with a history of heavy weevil occurrence (1.3 application) than in the fields with normally light infestations (1.0 applications). Then in 1978 only 0.2 more appli- cations were needed in the heavily infested fields (0.2 compared with 0). No insecticide applications were required in 1979 for either category of field. Yield comparisons indicated that before implementations of the program, boll weevils reduced yields by an average of 170 pounds of lint per acre (29% in 1974, 32% in 1975, and 31% in 1976) (Boring and Fuchs 1980). After the implementation of uniform planting, the cotton fields in the heavily infested areas eventually experienced increased yields over the lightly in- fested area of 6.4% (697 pounds lint per acre-heavy v. 655 pounds per acre-light) in 1977 and 72% in 1978 (387 pounds lint per acre-heavy v. 225 pounds per acre-light).
Since uniform delayed planting presumably impacts on about 25% of the Rolling Plains acreage heavily infested with boll weevils, producer savings in yield and insecticide costs were estimated to range from $3.38 to $5.45 million. In addition, producers have in- creased acreage 51.1%, from 703,700 harvested acres in 1976 to 1,051,250 in 1979 (Boring and Fuchs 1980). In sum, uniform delayed planting plus a short-season management system has been a major factor in acreage increase at a time when other regions of the Cot- ton Belt are going through an acreage stabilization or decline.
The two short-season cotton production systems that have been discussed are well suited to Texas. For example, several short- season varieties (CAMD, Cascot, G&P, Lankart, and Paymaster) are available for commercial use in Texas. The majority of cotton in Texas is grown on upland soils, not on highly fertile delta or river bottom soils. Low rainfall (20-35 inches) restricts the length of the cotton production season. Finally, with approximate average yields of 340 pounds lint cotton per acre, production in- puts must be kept at a minimum. As a result, integrated pest man- agement systems have been developed to control insect pests without increasing production cost because the vast majority of Texas farmers cannot afford to engage in a season-long insect control

MANAGEMENT PROGRAMS AND IMPLEMENTATION 529
program involving multiple insecticide applications. However, the two short-season cotton production systems that were designed pri- marily to control the boll weevil have a spin-off of major impor- tance. Consistent insecticide applications for bollworm and tobac- co budworm control are needed on less than 7% of the acreage grown on fertile river bottom or on irrigated production systems (Parker et al. 1980). Natural outbreaks of these pests, that is, outbreaks not induced by insecticidal destruction of beneficial arthropods, now occur only occasionally.
The broad implementation of short-season cotton production systems and other factors have caused a dramatic reduction in in-^ secticide use in Texas (Thomas 1979). Insecticide use on cotton in Texas was at its highest, nearly 20 million pounds, in 1964, de-^ clined to 11.5 million pounds in 1966 and to 9.6 million pounds in 1971, and was estimated at just under 2.5 million pounds in 1976 (Thomas 1979). During this time the total pounds of insecticide applied to cotton (and the percentage of total use on U.S. cotton) remained relatively constant (1964-76) in the Southeast, increased sharply in the Delta, and decreased sharply in the Southern Plains. Thus, insecticide use during these years was reduced 88.2% in Texas with near constant acreage, decreased 13.4% in the Southeast with decreasing acreage, and increased 38.8% in the Delta with only a minimum increase in acreage (Thomas 1979). Cooke and Parvin (Chap- ter 2 in this handbook) provide more extensive and recent informa- tion on reduced insecticide costs in Texas as compared with costs in other regions in the United States.
It should be clearly understood that cotton producers in Texas utilize insecticides as the last alternative for control of insect pests. Their primary inputs into insect control are varietal and cultural. As a result of the short-season cotton production sys- tems used on the majority (95%) of the acres in Texas, an ecological stability has been created in a relatively insecticide-independent environment. The greatest spin-off of this system, with a few iso- lated exceptions, has been that the bollworm and tobacco budworm, which are presently considered the key pests of U.S. cotton, have been reduced to occasional pests in Texas.
Community-Wide Bollworm Management in Arkansas
The concept of insect population management within a cotton ecosystem is not recent in Arkansas. As early as 1924, Isely and Baerg (1924) discussed the value of population surveillance and spot dusting in managing the boll weevil. They found that auto- matic early-season applications of calcium arsenate failed to ef- fectively control weevils or increase yields, whereas scouting and treating as needed proved quite effective. From this early work, an insect pest management system was developed. Its components were scouting, spot dusting, and an early maturing cotton crop. Later, Lincoln and his coworkers (Lincoln et al. 1963), though fol- lowing the Isely concept, incorporated changes dictated by the

530 R. E. FRISBIEETAL
availability of more versatile insecticides, changes in mechaniza- tion, land clearing and cleaning, and further quantification and simplification of the scouting system.
The availability of more versatile insecticides that were of- ten relatively inexpensive provided a means of economically pro- ducing a "top crop" of cotton. These tactics added a month to the production season, thereby sacrificing earliness for yield.
Mechanization made it possible to clean out small overwinter- ing sites for boll weevils and led also to more nearly uniform planting dates, which usually obviated the need for spot dusting. The dispersed population of boll weevils, changed cultural prac- tices, and improved mechanization thus diluted the weevil popu- lation to the extent that it was below treatment thresholds until late in the season.
As noted, the dilution of the boll weevil populations meant that scouting had to be substituted for spot dusting. However, quantification and simplification of scouting procedures was neces- sary before the procedure was generally accepted. The modified system is referred to as "Point Sample" (Lincoln et al. 1963). It was adopted because the individual farm was basic to the Arkansas system. Thus the weevil emerged from diapause and infested only a few fields initially. However, as the season progressed, addi- tional fields were infested until all or most all fields within an area were infested. As a result, boll weevil control was practiced on a field-by-field basis.
Still, by 1949, the bollworm had become an annually occurring pest. The scouting system was therefore modified slightly and used to monitor bollworm populations, and insecticide use was recom- mended on a similar basis, i.e., field by field. Population stud- ies by Arkansas cotton entomologists during the 1950*s and I960's indicated that populations of the bollworm increased steadily dur- ing the season with infestations becoming quite heavy by August and September. It was also observed that bollworm populations were different in different seasons and in different regions of the State.
Under the auspices of a multistate IPM research project (Huf- faker Project), an effort was begun in Arkansas to determine why such population increases occur and to develop an improved system of managing the bollworm. The area chosen for the initial study is 25 miles east of Little Rock, has an excellent cotton production record, and is among the most threatened by the bollworm. Research conducted in the area included sampling techniques, surveillance techniques, damage threshold studies, modified life table studies, and a short-range bollworm forecasting program.
By the end of the third year of the study, data had been obtained that indicated the following: (1) long-range bollworm migration did not contribute to bollworm outbreaks during the

MANAGEMENT PROGRAMS AND IMPLEMENTATION 531
3 years of the program; (2) the June populations of bollworms in cotton were sufficient to produce the population levels that were subsequently measured; (3) treatment thresholds could be adjusted upward; (4) considerable progress had been made in developing a bollworm forecasting network; and (5) it was possible to utilize as much as 12,000 acres of cotton as a single field for a bollworm suppression system. Conclusions from the data were that the com- munity, through mismanagement of insect control procedures, had been largely responsible for the bollworm problems encountered.
The data also seemed to indicate that the July population of bollworms might be tolerated without treatment, thus reducing the risk of a damaging bollworm infestation in August. Although there was an element of risk in not treating the July population, it was decided that such a tactic should be proposed to the producers and that with their concurrence the community-wide program should be undertaken. It should be pointed out that from the point of view of researchers, there was considerable reservation about the under- taking. No replication of a 12,000-acre experiment was possible; so data analysis, reporting to the scientific community, and assur- ance that cause and effect were being measured were of concern. The experiment was completely vulnerable to the public eye.
Nevertheless, local farm leaders and Cooperative Extension Service personnel arranged for a meeting with the growers in Febru- ary 1976. The proposed control program was outlined, and a poll was taken to determine grower interest in starting a community-wide program. The response was much better than expected: over 90% of the producers contacted were in favor of the community-wide program.
The principal tactic employed in 1976 was that of treating the entire community of cotton fields as a single field. Scouting pro- cedures were changed to support the tactic as were treatment thresholds. For example, the scouting procedure included a whole plant search for eggs and larvae (which were categorized by size) and a total fruit damage count. The treatment threshold was based on community-wide averages for eggs, larvae, and damage. A prede- termined threshold was used that will be refined before publishing the results.
When the predetermined treatment threshold was reached, the community growers were so advised at a meeting arranged by the Ex- tension Service. There the growers were appraised of the situation and told which treatments were recommended though they had the option of using conventional materials Treatment was to begin as quickly as possible. The assessment of results would be made as soon as reentry regulations permitted. In short, a "hard" insecti- cide was applied very quickly over a rather extensive acreage that was infested by the target pest (Phillips et al. 1980).
The program in 1976 was very successful. Insecticide usage for bollworm control was reduced by 80% (a 10-year average of nine

532 R. E. FRISBIE ET AL
applications reduced to 2). Control was excellent, and no unex- pected insect control problems were encountered. Treatments were made on August 6 and August 27. It required approximately 3 days for the community to be treated. Check fields subsequently showed that failure to treat at the recommended times resulted in popula- tions of at least 16,000 large bollworm larvae per acre, which caused economic loss (Phillips et al. 1979).
Sampling data taken during the course of the study indicated that the magnitude of the July bollworm population depended largely on the density of the June population and how well it was synchron- ized with the phenological stage of the cotton plant. When cotton had reached a fruiting level of approximately 50,000 flower buds per acre and the larval population was in synchrony, larval estab- lishment was good. If the population was large enough (usually > 1500 larvae per acre), treatments were required in July. Thus, the June population was adequate to produce a subsequent population within the community that would warrant action by the growers. In other words, there are times when a suppressive treatment is neces- sary in June.
It was concluded that if the June population could be reduced by as much as 50%, it might be possible to avoid any further con- trol program against the bollworm until late August. Obviously, when the June population was quite low, as has sometimes been the case, control action would be delayed until the July population reached a preset treatment level.
Materials that are suitable for a June treatment must be spe- cific so as to protect the beneficial insects that provide natural control of the Heliothis spp. complex. Of the materials currently registered for use on cotton, those that most nearly meet the re- quirement for an environmentally nondisruptive substance are Bacil- lus thuringiensis (Bactur, Dipel, Thurcide) and Baculovirus helio- this (Elcar). Neither directly affects the naturally occurring control components in the cotton ecosystem. The efficacy of these materials against the Heliothis spp. complex in cotton is poorly understood despite some years of research. However, they clearly do not measure up to an effective conventional insecticide in re- gard to immediate crop protection. Indeed, neither material pro- vides sufficient levels of protection to warrant serious considera- tion for use against moderate to heavy Heliothis spp. infestations (Yearian et al. 1980).
On the other hand, the key to an early-season Heliothis spp. control program is population suppression. Action is usually taken at a stage of crop phenology when the need for Heliothis spp. pro- tection, if any, is considerably less than in midseason or late season. Thus, some of the disadvantages of B^. thuringiensis and B^. heliothis are not important in early season. For example, the ex- tended time required for mortality to occur is not as critical be- cause the objective is to remove insects from the population, not

MANAGEMENT PROGRAMS AND IMPLEMENTATION 533
to produce immediate kill. Although improved efficacy would be desirable, 40-50% kill plus the action of the natural enemy complex might be adequate to produce the desired level of suppression.
The program in 1977 therefore included the use of a virus against the second generation of the bollworm (the first-generation population in cotton was too low, < 500 per acre). The virus was applied July 12 on approximately 10,000 acres of cotton in the com- munity at the rate of 2 ounces of the formulated product per acre.
By August 12, the bollworm population had increased to a level that required treatment with a broad-spectrum material. As a re- sult, the community was treated with EPN-methyl parathion or with methomyl. This was the only such application needed except that a second treatment was needed on approximately 1500 acres of late planted, irrigated cotton.
Generally, the data obtained in 1977 support the premise that the use of a material with characteristics similar to those of Elcar will have a sufficient impact on the bollworm population so that a much reduced level of insecticide will be required for the balance of the season. For example, data taken during 1973-75 showed an approximate 15-fold increase in the population from July to late August. In 1977 when the virus was used, the increase was only fourfold (Phillips et al. 1979). Obviously other factors could have been involved, and the effects of the virus were still not known with certainty. It was felt, however, that there was a solid basis for trying a "nondisruptive" material early in the season as part of an areawide bollworm management program (Phillips et al. 1979).
The early-season suppression concept was carried out again in 1978. By late June (June 20), the bollworm population had reached an average of approximately 1200 larvae per acre community-wide. The decision was made to apply the virus against the June popula- tion, and 2 ounces per acre of formulated virus were again used. Following treatment, larvae were collected from the fields within the community and held for mortality studies. On the basis of these collections, about 25% of the larvae died from a virus.
Subsequent weather conditions in 1978 precluded the taking of data that would be comparable with those taken in 1977. A severe drought began in early July. Obviously, in such a situation, there was considerable disparity of moisture in different locations and attendant disparity in plant growth and fruiting characteristics. By August 1, community integrity had been lost, and a return had to be made to a field-by-field control program. Also, irrigated cot- ton required more application of insecticides than did unirrigated cotton. However, the community averaged only five applications of insecticide for the season, and the July to August increase in the population of the bollworm was only fivefold, considerably less than the 15-fold reported for 1973-75.

534 R. E. FRISBIE ET AL
In spite of the adverse conditions encountered in 1978, the results of the community program were superior to the results of the field-by-field program in a neighboring area. The community averaged five applications of insecticides; the check area averaged 11 applications. Weather conditions and yields were comparable in the two areas.
Unquestionably, the experiment in Arkansas demonstrated the value of the community-wide approach to bollworm management of rain-grown cotton. Insecticide usage was reduced, and yields were maintained or improved. The concept of community control has spread and the procedure is now used on over 75,000 acres of cotton in four areas of the State of Arkansas.
Improved Cotton Insect Management in the Southeast
In the Southeast where the average annual rainfall is approxi- mately 50 inches, high yields of quality cotton are a realistic ex- pectation. However, little cotton acreage in this region can be irrigated, so growers must depend on the vagaries of the rainfall pattern. This dependence means that a full-season production sys- tem must be used to achieve high yields and desired quality.
A full-season approach to cotton production puts heavy demands on management resources, and management of insect pests is particu- larly important. Cotton begins fruiting in May and continues to fruit until harvest, which provides pests an extended opportunity to cause economic damage.
The Heliothis spp. complex, i.e., tobacco budworm and bollworm, poses the greatest threat to cotton in the Southeast. Either H. zea or E. virescens is present throughout the cotton fruiting season, often in large numbers. Populations increase on alternate hosts and move to cotton. The boll weevil too is a major pest of southeastern cotton, especially in more southern areas where win- ters are often mild. Small field size and abundant overwintering habitat enhance the potential for economic boll weevil infesta- tions.
The tarnished plant bug (Lygus lineolaris (Palisot de Beau- vois)), although only an occasional pest, can be devastating to in- sect management programs when conditions favor its movement into cotton. Other occasional pests include thrips (Thysanoptera), aphids (Aphididae), cutworms (Noctuidae), beet and fall armyworms (Spodoptera exigua (HÜbner) and S. frugipera (J. E. Smith)), white- fly (Aleyrodidae), spider mites TXetranychidae), European corn borer (Ostrinia nubilalis (Hubner)), loopers (Geometridae), cotton fleahoppers, and others.
Cotton insect management in the Southeast can be divided into two categories: preventive measures and corrective actions. Pre- ventive measures include such procedures as variety selection.

MANAGEMENT PROGRAMS AND IMPLEMENTATION 535
rotation, and fertility management; corrective actions include detection of outbreaks through field scouting, setting of action thresholds, and judicious use of insecticides. Areawide pest moni- toring with traps and other survey techniques facilitates selection and implementation of both preventive and corrective management techniques.
Insect Pest Surveys
Blacklight traps have previously been used extensively for monitoring the activity of adult bollworms. A promising alterna- tive is the pheromone-baited trap, which is effective in capturing both bollworm and budworm moths. As trap design and pheromone for- mulations improve, this survey method should be incorporated into all organized cotton pest management programs in the Southeast.
The "sugar line" survey is another method of determining activity. It involves applying a sugar-water solution to cotton plants in late evening to arrest moths and make identification and counting possible. This procedure is simple and inexpensive, but must be done several nights each week of the fruiting season by someone capable of identifying the various species that are collected.
Grandlure-baited boll weevil traps have been used primarily to monitor movement of overwintered boll weevils into cotton fields in the spring and, to a lesser extent, late-season migration and emi- gration to hibernation sites in the fall. Because of time con- straints in late spring, pheromone trapping of boll weevils is the preferred method of timing the pinhead square applications of insecticide.
Other survey techniques are being developed for cotton insect pests including plant bugs, fall armyworms, and others. These techniques will not replace field-by-field inspections, but will alert growers and scouts to potential outbreaks.
Preventive Insect Management
Total crop management is the term used to describe a produc- tion system that includes insect pest management. The goal of cur- rent cotton production systems is to maximize profits by obtaining optimum yields and quality with minimum essential inputs. There- fore, insect control costs are reduced by incorporating certain management practices into the production system. Since production systems vary in different States and even within States, insect management strategies vary similarly.
Cotton varieties are being developed that incorporate in- sect resistant characters; for example, frego bract, glabrous stems and leaves, red color, and nectariless leaves and fruit.

536 R. E. FRISBIE ET AL
Currently, only the nectariless character has been used in a com- mercially grown variety, and it is widely planted in areas where plant bugs may be damaging. Determinate and semideterminate vari- eties are gaining acceptance because they can reduce the fruiting period and thereby reduce the length of time that cotton is suscep- tible to insect pests. However, the need for the flexibility of a full-season cotton limits the use of the more determinate varieties to situations involving double-cropping systems and irrigated fields.
Fertility management is used to minimize the attractiveness of cotton to pests and to reduce the fruiting period. Excessive nu- trients (especially nitrogen) produce rank growth, poor fruiting, and increased insect pest problems. Yet cotton producers are in- clined to apply more fertilizer than necessary to insure an ade- quate supply under extreme environmental conditions. Nitrate moni- toring through leaf petiole sampling and analysis enables growers to apply minimal rates of nitrogen to the soil and to supplement nitrate requirements with foliar applications of urea according to test results.
Planting dates for cotton in the Southeast depend on several factors including weather, alternate crops, and anticipated insect problems. Uniform, early planting reduces the threat from increas- ing late-season populations of boll weevils and Heliothis spp. Later planting can be used in an insect management program based on a shortened fruiting period, but then fertility and water manage- ment are critical.
Stand density and row pattern can be adjusted to reduce pest pressure according to the anticipated pest pressures and climate. Where thrips and other seedling pests are easily managed but plant bugs are a potential threat, lower seeding rates can be used to discourage Lygus spp. infestations. Higher rainfall areas that ex- perience excessive vegetative growth can utilize modified skip-row patterns. Since the plant canopy tends to remain open later in the season when skip-row patterns are used, competition for light and other requisites slows growth, thereby reducing boll rot and increasing penetration of insecticide sprays.
Trap crops have also proved effective in managing the boll weevil but have met limited acceptance by growers. Such strips of cotton are planted several weeks before the remainder of the field. As a result, emerging overwintered boll weevils are concentrated in the strips, and only a small portion of the field must be sprayed, thereby sparing the majority of the population of beneficial insects.
Use of pinhead square sprays is another effective way to re- duce populations of overwintered boll weevils. These consist of low rates of organophosphate insecticide applied to cotton prior to

MANAGEMENT PROGRAMS AND IMPLEMENTATION 537
the initiation of fruiting to prevent the oviposition that would produce subsequent generations during the fruiting period. Pinhead square sprays are most effective when peak emergence of over- wintered boll weevils occurs prior to fruiting.
Diapause boll weevil control involves reducing the number of boll weevils that enter hibernation by applying insecticides after cotton matures and regular sprays are terminated. The diapause control method is not successful when practiced on an areawide basis. The primary obstacle to this management tool is grower un- certainty concerning planting intentions the following spring.
Corrective Actions
Despite preventive management, pest outbreaks regularly occur in southeastern cotton. However, the severity of outbreaks varies considerably, and the necessity for corrective actions must be de- termined for each situation. The decisions are based on the knowledge of pest conditions gained through field scouting and a familiarity with natural mortality factors and applied controls.
Biological control of cotton insect pests in the Southeast de- pends largely on conserving natural enemies. Parasites and preda- tors play a significant role in suppressing infestations of major pests such as tobacco budworm and bollworm when insecticides are used judiciously.
Action thresholds (economic thresholds) have been developed for most cotton insect pests. These thresholds serve as a starting point for decisions on the need for insecticide applications. Thresholds must be adjusted by considering plant growth stage, weather, beneficial insect populations, pest species, value of crop, and cost of control.
The use of chemical insecticides is the primary method of cor- recting pest outbreaks. Since chemical applications often cause secondary pest problems, sprays should only be applied as needed to prevent certain economic loss. The use of selective insecticides and of minimum rates that are adequate against specific pests and infestation levels reduces costs and is less likely to disrupt natural control systems.
Regular inspection of fields is the only proven method of de- termining the need for corrective actions for all the potential in- sect pests of cotton. Field scouting is best done by specially trained individuals employed by cotton producers. Scouting serv- ices are offered by individuals, private consulting firms, and grower-owned organizations. The southeastern land-grant universi- ties provide training and technical support through the Cooperative Extension Services and Experiment Stations.

538 R. E. FRISBIE ET AL
BIOLOGICAL INQUIRIES AND CONSTRAINTS TO BELTWIDE IMPLEMENTATION OF BOLL WEEVIL ERADICATION AND OPTIMUM PEST MANAGEMENT
Biological Evaluation of Techniques
A discussion of the techniques employed in the implementation of beltwide boll weevil eradication and optimum pest management is presented here. However, the mass rearing, release, and efficacy of sterile boll weevils as a control component are reviewed in Chapter 11 of this handbook and consequently will not be discussed. It is very important that the biological strengths and weaknesses of the various techniques used in the BWET and OPMT and in other relevant studies be evaluated on the basis of the experience gained. Eradication assumes elimination; i.e., reduction to zero of the boll weevil in the infested portion of the Cotton Belt. Anything less would simply allow the problems of the past to recur. Eradi- cation places an unusual responsibility on one who might chart such a course, a responsibility not met by achieving 95% or 98% population suppression. Therefore, it is important that any weak- ness in the proposed technology for eradication be discussed. For our basis of reference we will use the specific control techniques proposed in the preliminary draft of the boll weevil eradication plan prepared by Plant Protection and Quarantine, APHIS, USDA, Sep- tember 1980. Again, it is recognized that this is a draft and that alterations can be made, but the point of the following discussion is to comment on the biological impact and feasibility of the fun- damental suppression techniques suggested. In addition, the strengths and limitations of optimum pest management will be dis- cussed with the view that this strategy is designed to suppress and maintain boll weevils below economically damaging levels.
Fall Diapause Insecticide Program
The application of insecticides in the fall for the control of prehibernating weevils (diapause control) has proved effective in suppressing weevil populations in the subsequent season, thus mini- mizing the necessity for in-season insect control. This method of boll weevil suppression has been most widely used in west Texas and was refined there as a result of the largest diapause boll weevil control program undertaken in the United States, the High Plains Boll Weevil Suppression Program. In 1964, an areawide fall boll weevil control program was initiated in eight counties on the southern and eastern edge of the High Plains. The objective was to prevent the westward spread of the boll weevil into the single largest cotton growing area in the United States, the High Plains of Texas. Excellent program organization, coordination, and cost sharing between Plains Cotton Growers, Inc., the Texas A&M Univer- sity, the USDA, the Texas Department of Agriculture, and the Texas Technological University provided an organizational framework and financial base for the implementation of this large-scale program. In addition, there was somewhat restricted availability of

MANAGEMENT PROGRAMS AND IMPLEMENTATION 539
favorable overwintering habitats and high boll weevil mortality re- sulting from low winter temperatures along the western edge of the test area. Approximately 295,000 acres of cotton were treated an average of 3.9 times at 10-14 day intervals with 16 fluid ounces of ultra-low-volume malathion in 1964. (Rummel et al. 1975).
In 1965, the program was revised into a two-phase program that was continued in 1966 and 1967. Due to budgetary restrictions, fields with light or moderate populations received limited treat- ment and in some instances no treatment, and fields were not treat- ed until the survey indicated that a certain infestation level had been reached (Rummel et al. 1975). This selective treatment was continued until 1974 when it was realized that adequate suppression was not being achieved. Since that time, all infested acreage within the suppression zone has been receiving insecticide applica- tions. Meanwhile it has been shown that the boll weevil is capable of surviving on the High Plains. Nevertheless, the large-scale ap- plication of boll weevil diapause control has successfully pre- empted establishment of the boll weevil in the High Plains of Tex- as, and as a result, substantial losses of High Plains cotton have been prevented.
Also, in the St. Lawrence Valley area of West Texas, which is well isolated from other infested cotton, a fall boll weevil con- trol program has been used effectively since 1968 (Neeb and Cole 1973). This program was designed to be responsive to the fluctua- tions in boll weevil populations between the various production seasons. One hundred percent of 20,000 acres was treated for diapause control in 1968-70. As a result, the boll weevil was suc- cessfully controlled, the number of acres requiring fall treat- ment dropped to 1200 in 1971, and in-season control could be virtually eliminated (the overall goal of the program). However, the boll weevil possesses the unique ability to resurge and in- crease in abundance. The weevil resurged in 1973-75, and the en- tire acreage had to be treated in 1975 and 1976.
This occurrence, like similar occurrences in the High Plains Boll Weevil Suppression Program, was thought to be primarily the result of successive mild winters in 1974 and 1975. Still, the St. Lawrence Valley program has demonstrated that despite fluctuations in weevil populations from year to year and the tremendous poten- tial of the population to rebound, a technically well designed pro- gram with 100% grower participation can effectively manage the boll weevil by utilizing this technique.
An important factor in both of the Texas diapause control pro- grams was their cost-sharing features. The High Plains program was cost-shared by growers and USDA-APHIS. The St. Lawrence program was cost-shared by growers and the Texas Department of Agriculture. Such cost-sharing provided a strong incentive for program implemen- tation and continued maintenance.

540 R. E. FRISBIEETAL
In contrast, the several fall boll weevil control programs conducted in south Texas have been relatively limited in success. In the programs conducted in 10 counties in 1969, 5 counties in 1970, 4 counties in 1971, and 1 county in 1976-80, nearly all had a cost-sharing incentive. However, the environmental parameters of south Texas, i.e., mild winters and abundant overwintering habitat, are more favorable to successful overwintering by weevils than the harsh conditions of west Texas. In addition, the warm spring and summer temperatures mean that the weevil has a higher biotic poten- tial in this area than in other cotton producing areas of Texas (Rummel and Frisbie 1978). Indeed, the south Texas conditions seem to be somewhat similar to the conditions in the more southern cot- ton producing areas in the United States. For example, freezes in south Texas and generally throughout the South do not occur until well into winter, so it is necessary to supplement a fall diapause control program with complete and areawide postharvest stalk de- struction. Also, inclement fall weather disrupted several diapause control programs in south Texas.
Fall boll weevil control has received limited acceptance belt- wide to date. Producer acceptance in Texas has been far greater than in other States, though this control method has been used in some areas outside of Texas. Lloyd et al. (1972) reported on a suc- cessful grower-sponsored program conducted in Monroe County, MS, in 1969. By utilizing a "reproduction diapause" control technique as the major component of the program in this Midsouth area, the grow- ers achieved a high level of population suppression. However, the greatest use of the fall diapause technique occurred in the early 1970's when an estimated 1,754,739 acres of cotton in several southern States received fall treatments of insecticide (Thomas 1974). The treatment was used mostly in Arkansas, Louisiana, and Texas though there was limited use in Alabama, North Carolina, South Carolina, and Tennessee, but most programs except those in Texas and South Carolina consisted of a single treatment of insec- ticide applied with a defoliant. Cooperative Extension Service en- tomologists reported (personal communication) that little benefit was realized from a single application of insecticide applied with a defoliant. Since then, the number of fall programs for the sup- pression of the boll weevil has, by and large, decreased. However, a well-coordinated fall program was conducted in the south Missis- sippi Pilot Boll Weevil Eradication Experiment in the early 1970's, and in the BWET, the fall program was a suppression component.
It is suggested, the experience gained with this method of boll weevil management shows its strength. However, in milder and more moist areas of the South, this control technique has limita- tions, even under highly organized pest management programs.
Insecticidal Control of Boll Weevils Emerging in the Spring
The use of insecticides to suppress the population of over- wintering boll weevils that enters cotton when plants are in the

MANAGEMENT PROGRAMS AND IMPLEMENTATION 541
squaring stage was first suggested by Ewing and Parencia (1949). Such treatment of a 10 -mile area in North Carolina did, in fact, delay the development of the boll weevil population (Mistric and Covington 1968, Mistric and Mitchell 1968). Sprott et al. (1976), in a review of Walker's short-season, narrow-row cotton production system, subsequently discussed the use of spring insecticide appli- cations to delay the development of subsequent boll weevil popula- tions. Parker et al. (1980) utilized this system in a short-season production scenario to delay boll weevil increase and to allow suf- ficient time for the safe development and maturation of fruit. The use of insecticide applications directed at overwintering adults entering cotton has proved to be an effective tool in Texas and in other areas of the Cotton Belt that are infested with boll weevils (personal communication. Extension entomologists 1980). Also, this so-called "pinhead" square application had limited use in the 0PM Trial. Rummel et al. (1981) have now developed an index whereby grandlure-baited pheromone traps are used to obtain an indication of which fields require treatment. By utilizing the catches in these traps as an index, Rummel suggests that it may not be neces- sary to treat an entire area; only selected fields need be treated as indicated by the trapping index.
The suppression of overwintering and spring emerging adults has proved to be beneficial in both short-season and long-season cotton production. However, care should be taken in applying early-season insecticide to avoid eliminating beneficial arthropods that subsequently suppress bollworm/tobacco budworm populations. If insecticides are applied within 10-14 days of the bloom stage, there is a high probability that beneficial insects will be de- stroyed, and cotton will be left vulnerable to bollworm/tobacco budworm attack. The draft of the boll weevil eradication plan sug- gests that four applications of diflubenzuron (Dimilin) be made (7- day intervals) to fields infested as indicated by trap catches. These applications would then be followed, if necessary, by a sin- gle application of azinphosmethyl (Guthion) as a cleanup treatment, but this treatment would occur during the bloom period. Although the plan states that the diflubenzuron and azinphosmethyl treat- ments will be applied in an effort to reduce, "insofar as possi- ble," the probability of destroying beneficial insects and induc- ing a bollworm outbreak, such a procedure would still be extremely hazardous. It might have less impact in those cotton producing areas that normally treat for bollworm/tobacco budworm infesta- tions; however, those areas that routinely do not apply insecti- cides for bollworm and tobacco budworm, but rely primarily on beneficial arthropods for control, would be placed in jeopardy. For example, throughout most of Texas where the short-season, low insecticide input systems are used, a serious problem could be cre- ated that could force farmers to apply additional insecticide to control the Heliothis spp. complex. The economic situation in these production areas is not sufficiently robust to allow for this additional expenditure, a fact well understood by farmers of the area.

542 R. E. FRISBIEETAL
Role and Efficacy of Diflubenzuron
The role and efficacy of diflubenzuron is discussed in Chapter 9 of this handbook. However, a brief summary of pertinent litera- ture is presented here along with a discussion of the utility of this compound in weevil control programs. This insect growth regu- lator has shown activity against the boll weevil; for example, it has been reported to be effective in suppressing boll weevil popu- lations in tests conducted in North Carolina, South Carolina, and Arkansas (Taft and Hopkins 1975, Ganyard et al. 1977, Lloyd et al. 1977, Ganyard and Bradley 1978). Also, studies by Abies et al. (1980b) and Keever et al. (1977) indicated that Dimilin is general- ly less detrimental to beneficial arthropods than conventional insecticides.
However, tests of the effectiveness of Dimilin in suppressing boll weevil populations conducted by Rummel (1980) in the Rolling Plains, Abies (1980b) in the Central Texas River Bottoms, Cole (1980) in the Upper Gulf Coast, and Harding and Wolfenbarger (1980) in the Lower Rio Grande Valley showed varying degrees of effective- ness. Generally, the highest level of reproductive suppression occurred with higher application levels directed against light wee- vil populations. The poorest results were reported in the Lower Rio Grande Valley study: there was no significant difference in reproduction suppression between treated and untreated plots having moderate to heavy populations (Harding and Wolfenbarger 1980). Later in the season after 35% of the squares were damaged, migra- tion may have accounted for the poor results; however, results were not significantly better before the onset of migration. Likewise, in the Upper Gulf Coast study, average adult emergence in the treated fields never fell below 43% under light boll weevil popula- tion pressure (Cole 1980). On the other hand, under heavy boll weevil infestations in west Texas and with rates of 1.0 and 2.0 ounces active ingredient (Al) per acre, average adult emergence in treated plots was about 40% compared with an average emergence of 73% in untreated plots (Rummel 1980). Under a light infestation in the same area, the average adult emergence reported was only 13.4% for the 10 ounce AI rate per acre and 9.2% for the 2.0 ounce AI rate. Also, in the test conducted in the Central Texas River Bot- tom, substantial suppression of reproduction in treated fields was noted (Abies et al. 1980b): weevil mortality in fields treated with Dimilin at rates of 0.5, 1.0, and 2.0 ounces AI per acre aver- aged 46.4, 67.4, and 77.3%, respectively.
Thus suppression of reproduction was obtained with difluben- zuron in the Texas studies, but the degree of suppression achieved generally did not approach the degree reported from tests in other States. In general, the heavier boll weevil infestations encoun- tered in Texas could have accounted for the difference in suppres- sion (Rummel 1980). However, the results of the Texas studies indicate that one might have difficulty in controlling even light to moderate infestations of boll weevils with diflubenzuron. Therefore, special management practices are indicated for the

MANAGEMENT PROGRAMS AND IMPLEMENTATION 543
effective use of this material. It is suggested that diflubenzuron is best applied at the pinhead square stage of cotton, or earlier, before weevil oviposit ion begins. Also, diflubenzuron would have to be applied at regular intervals, every 5-7 days, for an extended period to maintain control. In years of moderate to heavy boll weevil population pressure, it would be necessary to supplement diflubenzuron applications with applications of a conventional insecticide.
Because of erratic results obtained with diflubenzuron in the several multiple-year studies that have been made, the material is not recommended for use in pest management systems at present (Ex- tension entomologists, personal communication). (An optimistic but speculative view of its use is presented in Chapter 9 where the authors suggest the utility of diflubenzuron as a multiple applica- tion, early-season boll weevil suppressor in a short-season cotton production system.) Even if diflubenzuron were effective, it is questionable whether producers would be willing to pay the exces- sive costs of material and application in view of the lower costs of conventional materials applied fewer times. Further refinement and study are necessary to determine the role of diflubenzuron in a pest management program (Extension entomologists, personal communication).
In-Season Application of Phosphate-type Insecticides for Boll Weevil Control
The preservation of entomophagous arthropods is central to a balanced IPM program. Therefore, the use of phosphate-type insec- ticides directed at the boll weevil in-season, that is, beginning near the bloom period, imposes an element of risk. As noted in Chapter 5 in this handbook, the risk of destruction of beneficial arthropods that play a key role in the suppression of Heliothis spp. and other pests is ever present. The point here is that the risk entailed in artificially releasing Heliothis spp. from the biological control of beneficial arthropods must be carefully assessed. We thus come again to the risk involved, particularly in large acreages of Texas cotton, where great care is taken to mini- mize the use of phosphate-type and other insecticides just prior to and around the bloom period. Indeed, one of the special advantages of short-season cotton production systems is that they are basi- cally geared not to become involved in insecticide treatments for Heliothis spp. populations; thus insecticide applications are care- fully applied so as to vastly minimize risk of Heliothis spp. out- break. A question arises: What is the risk and how detrimental will it be if an application of azinphosmethyl is required after diflubenzuron in applied near the bloom period? The matter is ser- ious since it impacts greatly on approximately 3.75 million acres of cotton produced in Texas and on additional acreage in other cot- ton producing States where efforts to conserve natural enemies of the bollworm are a high priority.

544 R. E. FRISBIEETAL
Impact of Alternate Hosts on Eradication
Cross et al. (1975) provided an extensive review of the liter- ature on host plants of the boll weevil. Malvaceous species that have been implicated as possible hosts of the boll weevil in south Florida (Gossypium hirsutum L. var. punctatum (Shum. and Thonn.) Roberty, Thespesia populnea (L.) Soland.x Correa, Cienfuegosia yucatanensis Mills p., and Malvaviscus drummondii Torr. & Gray) are presumed to be sufficiently isolated from present populations of boll weevil so they have low probability of becoming infested and sustaining an incipient boll weevil population. Thespesia populnea was found by Luke fahr (1956) to be infested in Browns- ville, but this ornamental tree may be only a sporatic host of the boll weevil. Cross et al. (1975) concluded that Sphaeralcea angus- tifolia (Cav.) G. Don. and other related species of the genus may only occasionally serve as boll weevil hosts in desert and semi- desert areas of Texas and the western States. Hibiscus syriacus L., a frequent and popular dooryard planting in the South and Southeast, has only been recorded as being infested when grown ad- jacent to infested cotton (Parrot et al. 1966). The boll weevil has been reported as feeding on, but not reproducing on, the flow- ers of yellow woolywhite (H3mienopappus flavescens Gray) during May and June in the Rolling Plains of Texas (Rummel et al. 1978).
It appears that the most important persistent alternate host for the boll weevil in the United States is Cienfuegosia drummondii (Gray) Lewton, which is distributed throughout the Coastal Bend region of Texas. This plant apparently has the ability to sustain low level, incipient boll weevil populations at some distance from cotton. Therefore, it would be wise to consider suppressing the boll weevil in this alternate host if eradication is attempted.
LOGISTICAL CONSTRAINTS ON BELTWIDE IMPLEMENTATION OF BWE AND 0PM
Logistical problems frequently create the most nagging con- straints on the implementation of large-scale projects. One such problem, cost of eradication, should therefore be seriously consid- ered. Although the draft plan for boll weevil eradication contains a carefully considered analysis of the costs involved in the eradi- cation of the boll weevil, special care should be taken so that cost estimates (particularly in personnel, production of mass- reared sterile weevils, maintenance of weevil-free areas, and the maintenance of a containment zone to prevent boll weevil reentry) are calculated as precisely as possible.
Cost of Implementation
Cost estimates in the draft boll weevil eradication plan are based on the 1979 cost of the BWET. However, boll weevil infesta- tions in both the BWET and OPMT areas were considered very light in 1977 and 1978, and light in 1979 (Science and Education Adminis- tration 1981). This biological occurrence is favored in the

MANAGEMENT PROGRAMS AND IMPLEMENTATION 545
evaluation zones in regard to costs. A question then arises: would there be a cost increase in eradication if weevil populations were moderate or heavy? More specifically, are costs under- estimated for beltwide implementation because they are based on cost estimates for a situation characterized by very light weevil populations?
Cost for rearing and sterilizing weevils has been estimated at $2 per thousand with additional dispersal costs of $2 per acre. Sterile boll weevils used for release in the BWET were reared at the USDA-ARS Gast Rearing Laboratory at Mississippi State, MS. A question arises concerning the increase in demand for sterile wee- vils needed for an areawide eradication plan. Will the Boll Weevil Laboratory be able to rear a sufficient number of weevils to accom- modate the eradication? If not, will an additional facility(ies) be required, and what additional cost and time will be involved in constructing this new facility(ies)?
Reference must be made to the establishment of a containment zone at the Mexico-United States border or running north-south along the western boundaries of Arkansas and Louisiana and the eastern edge of Oklahoma and Texas in a relatively cotton-free area. If technology is available to develop an effective barrier zone to prevent spread of the boll weevil into uninfested cotton, careful consideration must be given to the cost of the establish- ment and maintenance of this area. Costs of a containment program would continue, essentially, into perpetuity. The agencies and others responsible for encumbering these costs must be identified.
Should an eradication plan be implemented, the question arises as to the benefits that would be realized by the farmers involved in the program as it is implemented, presumably from east to west. Those farmers who have been involved in an eradication phase will have a distinct economic advantage over those who are still re- quired to control the boll weevil by conventional means. This will place farmers in the weevil-free area at a distinct advantage over those in infested areas (L. D. Anderson, past President, Texas Asso- ciation of Cotton Producers Organizations, Lubbock, TX, personal communication). It is anticipated that after eradication and in the absence of the boll weevil, production costs for insect control would be decreased. However, the time frame for beltwide eradica- tion is estimated at approximately 9 years during which a favorable economic position would be enjoyed by a certain percentage of the U.S. farmers in the weevil-free area.
Second, and possibly of greater significance, an increase in acreage could come about as a result of government cost-sharing of control costs during the implementation phase. Increased planted acreage resulting in an increase in supply of cotton, depending on market conditions, could drive the market price of cotton down (W. Hart, Executive Officer, Southern Texas Cotton and Grain Producers Association, Victoria, TX, personal communication). Further, if the containment zone should be established along the eastern edge

546 R. E. FRISBIE ET AL
of Texas and Oklahoma instead of on the United States-Mexico bor- der, farmers in Texas and Oklahoma could be placed at a long-range economic disadvantage because of the need to deal independently with weevil control costs.
Indeed, it is not clear how a containment zone in Texas and Oklahoma would function. If it is assumed that eradication is to take place over the entire weevil-infested area, why would this area be selected? What would be the cost associated with control- ling the weevil over the large acreages of Texas and Oklahoma if reinfestation occurred? Biologically, what would prevent reinfes- tation of the weevil from Mexico into Texas and Oklahoma, a buildup there in numbers, and an overrun of the containment zone? The only reasonable location for a containment zone would be along the United States-Mexico border.
Then there is the matter of utilizing available Cooperative Extension Service personnel and others from land-grant universities to provide technical and supervisory support for an eradication program. Costs associated with involvement of the time and facil- ities of universities cooperating with beltwide implementation should be calculated as part of the total cost of the program. Frequently, similar hidden costs that are not included, but as- sumed, do not appear in an overall cost estimate. These should be identified and included as legitimate costs of implementation.
Beltwide boll weevil eradication costs have been estimated by USDA-APHIS. The most recent estimate is $483 million though costs are increased to $569 million if an optimum pest management program precedes the beginning of eradication. It is interesting that a Stanford Research Institute study contracted for by Cotton Incor- porated to estimate costs of beltwide boll weevil eradication in the early 1970*s, that is, following the south Mississippi Boll Weevil Eradication Trial, gave a high cost of $2.46 billion and a low cost of $1.11 billion when the figures were corrected for in- flation and undiscounted (Peter Stent and Andrew Korsack. Refined Cost Estimate for Beltwide Eradication of the Boll Weevil. Stan- ford Research Institute Project No. 2372. Unpublished report, August 31, 1973).
This discrepancy between the cost projected by an independent study group and by USDA-APHIS gives rise to several questions. USDA-APHIS responds by saying that technology for eradication has improved and that diflubenzuron is now available. However, cost estimates were higher about 1.5 years ago before it was shown that diflubenzuron was not as effective as originally assumed. Since then, cost estimates have decreased. This discrepancy leads one to question the origin and accuracy of the USDA-APHIS figures. They appear to be alarmingly low, and if so, are misleading rela- tive to the cost of this massive proposed program.

MANAGEMENT PROGRAMS AND IMPLEMENTATION 547
Personnel Needs
The need for qualified professional and paraprofessional per- sonnel is always a problem when one is implementing large-scale management programs. As an example, there has always been a need for well-trained and qualified supervisory and scouting personnel in the ongoing pest management programs that have been underway throughout the boll weevil infested area of the Cotton Belt. In personal communications with Cooperative Extension Service ento- mologists, it was found that the number of available scouts fre- quently limited the number of acres that could be included in the particular State's pest management program. Particularly in Texas, implementation of a beltwide program would require excessively large numbers of scouts. The identification, training, and super- vision of these scouts by qualified personnel is viewed quite real- istically as an identifiable constraint. Further, even if such paraprofessionals (scouts) could be identified, the Cooperative Extension Services frequently have a problem because of the diffi- culty of retaining scouts long enough to participate in the entire program. In any beltwide effort, a minimum of 4-6 months would be required to cover the spring, in-season, and fall activities. The most reliable scouts have been found to be college age students (Extension entomologists, personal communication), but because of conflicting schedules, students oftentimes have to return to class before the end of the production season; and in the southern lati- tudes, the ability of college students to participate in a scouting program in the spring is hampered by their need to complete spring semester classes. Nothing is more important to an areawide program than highly qualified paraprofessionals who can collect the perti- nent field information season long. Without this vital ingredient, any program is severely handicapped.
Role of Independent Consulting Entomologists
The role of the independent consulting entomologist should be considered in any beltwide implementation plan. Consulting ento- mologists who participate in pest management programs in their ca- pacity as business consultants should have a clearly defined role in the plan. It has been estimated that approximately 1000 inde- pendent consulting entomologists are in business within the boll weevil infested area of the Cotton Belt (E. S. Raun, personal com- munication). These consultants have been valuable in the dissemi- nation of vital, high quality information to farming clientele. No other private group has had more impact on the implementation of IPM than the independent consulting entomologist. High priority should be placed on developing an agreement with independent con- sultants, possibly a contractual agreement for their services, as an integral part of any plan.

548 R. E. FRISBIE ET AL
Conduct of an Areawide Insecticide Spray Program
Perhaps the most experience in an areawide, intensive insecti- cide spray program was gained in the High Plains Boll Weevil Sup- pression Program (Rummel et al. 1975). The earlier discussion of the fall diapause insecticide program explained how this large- scale program effectively stopped the westward spread of the boll weevil into the High Plains of Texas. In this program, the primary technical component was the areawide application of insecticides to reduce the numbers of potentially overwintering weevils in several hundred thousand acres of cotton. The intensive organization and administration of this project by USDA-APHIS, Plains Cotton Grow- ers, Inc., and Texas A&M University clearly showed the logistical magnitude of such an operation. However, the perennial con- straints. . .problems with intensive field mapping, trapping, field inspection, contracting for chemicals and aerial application, and precise application of insecticide on target fields...were overcome in this project. Much is to be learned from this experience as it relates to any areawide insecticide spray program.
A matter of critical importance when an insect species is to be eliminated from identified target fields is application of the appropriate rate of chemical at the appropriate time with a guaran- tee of adequate coverage of the target area. Wind speed, aircraft height, nozzle pressure and arrangement, and working conditions are all factors that must be considered. Excellent experience in this was gained in the High Plains Boll Weevil Suppression Program and in the North Carolina BWET. As a result, a prototype of coordina- tion and logistics has been worked out.
Maintenance of Weevil-Free Areas and Containment Zones
The very rapid extension of the range of the boll weevil dur- ing the first years of its invasion of the United States is strong testimony to the dispersal ability of this insect. Long-range, late-season dispersal, frequently termed migration, has been the major reason for the expansion of the boll weevil into previously uninfested cotton (Bottrell et al. 1972). This behavorial. phenome- non is cited as probably the most powerful weapon that the boll weevil possesses. Certainly the ability to migrate comparatively long distances has allowed the insect to continually invade new territories and to reinvade old territories that have been tempo- rarily cleared of this pest (Bottrell et al. 1972). Some evidence of the long-range mobility of the insect was provided by a catch in male-baited wing traps located between Juarez and Chihuahua, Mexi- co, in 1968: 15 boll weevils were captured on 49 of these traps though the traps were 25-45 miles distant from cultivated cotton (Davich et al. 1970). Also, after weevils were released in South Carolina during midseason, the F, through succeeding generations were trapped at least 19 miles away from the point of release (Roach and Ray 1972). In addition, a weevil marked and released at McBee, SC, in late June 1980 was captured in Candor, NC, a distance

MANAGEMENT PROGRAMS AND IMPLEMENTATION 549
of approximately 65 miles (W. A. Dickerson, USDA-ARS, personal com- munication). Such late-season dispersal of the boll weevil repre- sents a potential constraint on the establishment and maintenance of weevil-free areas and containment zones.
Finally, relatively little attention has been focused on the requirements for the development and maintenance of boll weevil containment zones though experience gained from previously discus- sed boll weevil suppression programs should provide some clues. Presently, it is suggested that in a containment zone there should be a fall diapause program supplemented by intensive trapping and survey. These procedures would then be followed by a regulatory program to prevent movement of potentially infested items, as in the program in effect against the pink bollworm (Pectinophora gos- sypiella (Saunders)). These tactics may quite adequately keep the weevil from moving into uninfested cotton; however, if elimination of the weevil is the goal, consideration should be given to gaining a greater understanding of requirements of the containment zone. It is assumed that the weevil will be substantially suppressed through the implementation of the eradication strategies and that procedures in the containment zone should suffice to suppress the weevils to such low numbers that reinfestation will be unlikely. However, historically, fall boll weevil suppression programs used in conjunction with survey traps have not been sufficient to elimi- nate or contain the weevil from season to season (Rummel et al. 1975).
In view of the reproductive potential of the boll weevil and its long-range migrating ability, consideration should also be given to other suppressive techniques that could be utilized within a containmment zone. These include uniform planting and posthar- vest destruction dates, use of short-season cotton varieties, and early-season suppression of spring emerging adults. The question arises: how effective will these tactics be singly and cumula- tively in the maintenance of a containment zone? Except for past experience with a limited number of these tactics, little informa- tion exists on the cumulative effect of such strategies in contain- ing boll weevil populations. Probably additional information should be developed to more clearly define the needs and limita- tions of a boll weevil containment zone. These comments reflect no intention to minimize any past efforts at boll weevil suppression; they are only meant to assist in a better description of the technical components necessary to maintain a containment zone.
But there is still another matter. Assuming elimination of the boll weevil, the maintenance of weevil-free areas will pose a problem. The plan for implementation suggests that should spot in- festations occur directly after implementation within a particular zone, populations will be eliminated immediately by a special group of personnel involved in the implementation effort. However, if spot infestations occur after the completion of the implementation phase, individual States will presumably be responsible for elimi- nating these infestations. It is therefore recommended that a

550 R. E. FRISBIEETAL
series of techniques should be developed to deal with this poten- tial problem and the associated costs. This has not been done. It is of critical importance that weevil-free areas be maintained and spot infestations be identified before weevil populations can build and expand into uninfested cotton. A great deal of attention should be placed on contingency plans for dealing with such an occurrence.
Federal and State Regulatory Requirements for Boll Weevil Eradication
The first basic regulatory requirement for any suppression, containment, or eradication program involving an economic pest is to enact legislation that establishes authority for conducting the program. Such legislation is essential at the Federal and State level for the enhancement of cooperative efforts. The secondary requirement is passage of a referendum specific to the boll weevil program.
Legislation already exists at the Federal level and in some States, namely, North Carolina, South Carolina, Texas, and Vir- ginia, regarding the boll weevil beltwide program. All other States in the weevil-infested areas of the Cotton Belt must enact similar legislation before a program can be initiated. These States are Alabama, Arkansas, Georgia, Louisiana, Mississippi, Missouri, Oklahoma, and Tennessee.
Special regulatory requirements, previously outlined in the respective action plans for the BWET and the OPMT, and the neces- sity for them prior to and during the 3-year life of the program in a given area, were explained as follows (National Cotton Council Technical Subcommittee 1973):
"(1) Authority to establish suppression and elimina- tion zones for the boll weevil. Because of the complexity and size of the area involved, costs, logistics, etc., it is more feasible and practical to implement the program in sections rather than covering the entire infested acreage simultaneously. As the program progresses, certain sections will encompass acreage in more than one state.
"(2) Access and entry authority. During normal pro- gram operations such as treatment applications, monitor- ing, trapping, etc., it will be necessary for Federal, State, and sometimes contractural personnel, to enter cotton fields and adjoining properties in suppression and elimination zones.
"(3) Authority to execute the program on 100% of the cotton acreage. Cooperation and participation by all cotton producers in the treatment zones are essential

MANAGEMENT PROGRAMS AND IMPLEMENTATION 551
ingredients to the success of the boll weevil program.
This authority would insure that necessary activities could be be conducted effectively and efficiently.
"(4) Authority to require reporting of cotton acre- age by the grower to insure all acreage is included in
the program. Accurate information on the size and loca- tion of all cotton fields, including small patches grown as ornamentals for aesthetic purposes, is paramount in
an eradication program.
"(5) Authority to purchase and destroy cotton which may pose an undue hazard to program objectives because of difficulty in program execution. Field location, size, accessibility and other factors may prevent certain pro- gram functions from being conducted. In these instances, it would be better to destroy the cotton and/or indemnify the landowner than risk the possibilities of reinfesta- tion of adjacent areas and ultimate program.failure.
"(6) Authority to prohibit planting of noncommercial
cotton, and prevent volunteer cotton and alternate host plants from jeopardizing program objectives. Strict authority is needed to prohibit ornamental and other noncommercial cotton from being planted. The same holds true for volunteer cotton or alternate host plants to maintain program objectives and continuity."
These same requirements are essential for the conduct of any other boll weevil program. Authority must be provided for in the basic act and set forth in quarantines or regulations promul- gated thereunder. A previous report covers the necessity of each requirement in more detail (National Cotton Council Technical
Subcommittee 1973).
Additional requirements that must be considered are: (1) the interstate and intrastate movement of regulated articles; i.e., the boll weevil, cotton, used harvesting equipment, and alternate host plants, (2) authorization for compliance agreements, (3) authoriza-
tion for penalties for violations, and so forth, that are usually standard conditions and requirements under any quarantine invoked for an economic pest. These requirements are also essential at the onset of the boll weevil program, but would have greater influence once elimination of the weevil from a given sector is imminent.
Those States that have enabling legislation must approve, by referendum, a program authorized by the legislation specific to the eradication requirements. Those States that do not have enabling legislation must both adopt such legislation and pass à referendum. Most State legislatures meet biennially. There must therefore be considerable advance preparation for the development and implemen-
tation of the appropriate necessary legislation.

552 R. E. FRISBIE ET AL
LITERATURE CITED
Abies, J. R., S. L. Jones, V. S. House, and D. L. Bull. 1980a. Effect of diflubenzuron on entomophagous arthropods
associated with cotton. jLn Effectiveness of Diflubenzuron (Dimilin) for Boll Weevil Suppression in Texas. Southwestern Entomologist 5 (Supplement l):31-35.
S. L. Jones, V. S. House, and D. L. Bull. 1980b. ^Effectiveness of diflubenzuron in Central Texas River
Bottoms. In Effectiveness of Diflubenzuron (Dimilin) for Boll Weevil Suppression in Texas. Southwestern Entomologist 5: 15-21.
Adkisson, P. L., and S. J. Nemec. 1966. Comparative effectiveness of certain insecticides for killing bollworms. Texas Agricultural Experiment Station, Bul- letin No. 1048, 4 p.
Boring, E. P. 1974. Planting date and early maturity importance in boll weevil
control. Texas Agricultural Extension Service, News Release, February 12, 2 p.
and T. W. Fuchs. 1980. Delayed uniform planting - a new weapon in the boll weevil
war. Texas Agricultural Extension Service, 1980 Annual Report, 4 p.
Bottrell, D. G., D. R. Rummel, and P. L. Adkisson. 1972. Spread of the boll weevil into the High Plains of Texas. Environmental Entomology 1:136-140.
Cole, C. L. 1980. Effectiveness of diflubenzuron in the Upper Gulf Coast of
Texas. In Effectiveness of Diflubenzuron (Dimilin) for Boll Weevil Suppression in Texas. Southwestern Entomologist 5: 22-26.
Cross, W. H., M. J. Lukefahr, P. A. Fryxell, and H. R. Burke. 1975. Host plants of the boll weevil. Environmental Entomology
4:19-26.
Davich, T. B., D. D. Hardee, and M. Acala. 1970. Long-range dispersal of boll weevils determined with wing
traps baited with males. Journal of Economic Entomology 63: 1706-1708.
Ewing, K. P., and C. R. Parencia, Jr. 1949. Early-season applications of insecticides for cotton in-
sect control. U.S. Department of Agriculture, Bureau of Ento- mology and Plant Quarantine E-792, 9 p.

MANAGEMENT PROGRAMS AND IMPLEMENTATION 553
Finley, M. D., and E. P. Boring. 1978. In Texas Statewide Pest Management Program Annual Report
1977-T978. R. E. Frisbie et al., editors. Texas Agricultural Extension Service, Mimeograph, p. 16-22.
Frisbie, R. E., M. A. Rothrock, W. E. Buxkemper, and G. C. Moore. 1978. Texas statewide pest management program annual report
1977-1978. Texas Agricultural Extension Service, Mimeograph, 23 p.
Ganyard, M. C., and J. R. Bradley, Jr. 1978. Wide-area field test of diflubenzuron for control of an
indigenous boll weevil population. Journal of Economic Ento- mology 71:785-788.
J. R. Bradley, Jr., F. J. Boyd, and J. R. Brazzel. 1977. Field evaluation of diflubenzuron (Dimilin) for control of
boll weevil reproduction. Journal of Economic Entomology 70: 347-350.
Harding, J. A., and D. A. Wolfenbarger. 1980. Evaluation of diflubenzuron for boll weevil control in the Lower Rio Grande Valley of Texas. Ln Effectiveness of Diflu- benzuron (Dimilin) for Boll Weevil Suppression in Texas. Southwestern Entomologist 5:27-30.
Heilman, M. D., M. J. Lukefahr, L. N. Namken, and J. W. Norman. 1977. Field evaluation of a short-season production in the Lower
Rio Grande Valley of Texas. In_ Proceedings Beltwide Cotton Production Research Conference, Atlanta, GA, January 1977, p. 80-83.
Howard, L. 0. 1897. Insects affecting the cotton plant. U.S. Department of Agriculture, Office of Experiment Station, Bulletin No. 33, 328 p.
Isely, D., and W. J. Baerg. 1924. The boll weevil problem in Arkansas. Arkansas Agricul-
tural Experiment Station, Bulletin No. 462, 29 p.
Keever, D. W., J. R. Bradley, Jr., and M. C. Ganyard. 1977. Effects of diflubenzuron (Dimilin) on selected beneficial
arthropods in cotton fields. Environmental Entomology 6:732- 736.
Lacewell, R. D., J. E. Casey, and R. E. Frisbie. 1977. An evaluation of integrated pest management programs in
Texas: 1964-1974. Texas Agricultural Experiment Station, De- partment Technical Report No. 77-4, 215 p.

554 R. E. FRISBIE ET AL
Lincoln, C, G. C. Dowell, W. P. Boyer, and R. C. Hunter. 1963. The point sample method of scouting for boll weevils. Arkansas Agriculture Station, Bulletin No. 666, 31 p.
Lloyd, E. P., M. E. Merkl, F. C. Tingle, W. P. Scott, D. D. Hardee, and T. B. Davich.
1972. Evaluation of male-baited traps for control of boll weevils following a reproduction-diapause program in Monroe County, Mississippi. Journal of Economic Entomology 65:552- 555.
R. H. Wood, and E. B. Mitchell. 1977. Boll weevil: suppression with TH-6040 applied in cotton-
seed oil as a foliar spray. Journal of Economic Entomology 70:442-444.
Lukefahr, M. 1956. A new host of the boll weevil. Journal of Economic Ento- mology 49:877-878.
Mally, F. W. 1901. The Mexican boll weevil. U.S. Department of Agriculture,
Farmers* Bulletin No. 130, 30 p.
Masud, S. M., R. D. Lacewell, C. R. Taylor, J. H. Benedict, and L. A. Lippke.
1981. Economic impact of integrated pest management strategies for cotton production in the coastal bend region of Texas. Southern Journal of Agriculture Economics. [in press.]
Mistric, W. J., Jr., and B. M. Covington. 1968. A preventive boll weevil control program applied to a
ten-square-mile area within an untreated county. Journal of Economic Entomology 61:186-190.
and E. R. Mitchell. 1968. A preventive boll weevil control program applied to a
ten-square-mile area within a treated county. Journal of Eco- nomic Entomology 61:179-186.
Namken, L. N., and M. D. Heilman. 1973. Determinate cotton cultivars for more efficient cotton
production in the Lower Rio Grande Valley. Agricultural Journal 65:953-956.
National Cotton Council Technical Subcommittee. 1973. Overall plan for national program to eliminate the boll weevil from the United States. Technical Report, 73 p.
Neeb, C. W., and A. W. Cole. 1973. Boll weevils whipped with diapause programs. Texas Agri-
cultural Experiment Station, Report No. 644, 19:1.

MANAGEMENT PROGRAMS AND IMPLEMENTATION 555
Parker, R. D., J. K. Walker, G. A. Niles, and J. R. Mulkey. 1980. The short-season effect in cotton and escape from the boll weevil. Texas Agricultural Experiment Station, Bulletin No. 1315, 45 p.
Parrott, W. L., F. G. Maxwell, and J. N. Jenkins. 1966. Feeding and oviposit ion of the boll weevil, Anthonomus grandis (Coleóptera: Curculionidae) on rose-of-sharon, an alternate host. Annals of the Entomological Society of America 59:547-550.
Phillips, J. R., P. L. Adkisson, and A. P. Gutierrez. 1980. General accomplishments toward better insect control in
cotton, rn New Technology of Pest Control, Chapter 5, Carl Huffaker, editor. John Wiley, New York.
W. C. Yearian, R. G. Luttrell, and W. F. Nicholson, Jr. 1979. Use of commercial formulations of Bacillus thuringiensis
and nuclear polyhedrosis virus for control of Heliothis spp. in cotton. Ln Proceedings, Beltwide Cotton-Mechanization Confer- ence, p. 39-41. Beltwide Cotton Production Research Confer- ence. Phoenix, AZ, January 7-11, 1979.
Roach, S. H., and L. Ray. 1972. Boll weevils captured at Socastee, South Carolina, in
1970, in wing traps placed around fields with and without growing cotton. Journal of Economic Entomology 65:559-560.
Roussel, J. S., and D. F. Glower. 1955. Resistance to chlorinated hydrocarbon insecticides in the
boll weevil (Anthonomus grandis Boheman). Louisiana Agricul- tural Experiment Station, Circular No. 41, 9 p.
Rummel, D. R. 1980. Effectiveness of diflubenzuron on boll weevils in the Texas Rolling Plains. Iri Effectiveness of Diflubenzuron (Dimilin) for Boll Weevil Suppression in Texas. Southwestern Entomologist 5 (Supplement 1):8-14.
D. G. Bottrell, P. L. Adkisson, and R. C. Mclntyre. 1975. An appraisal of a 10-year effort to prevent the westward
spread of the boll weevil. Bulletin of the Entomological Society of America 21:6-11.
and R. E. Frisbie. 1978. Suppression of potentially overwintering boll weevils as
a pest management practice. Ln The Boll Weevil: Management Strategies. Southern Cooperative Series, Bulletin No. 228, p. 39-49.

556 R. E. FRISBIE ET AL
Rummel, D. R., J. R. White, S. C. Carroll, and G. R. Pruitt. 1980. Pheromone trap index system for predicting need for over- wintered boll weevil control. Journal of Economic Entomology 73:806-810.
J. R. White, and G. R. Pruitt. 1978. A wild feeding host of the boll weevil in west Texas.
Southwestern Entomologist 3:171-175.
Science and Education Administration 1981. Biological evaluation of alternative beltwide boll weevil/
cotton insect management programs. Overall Evaluation Appendix A. SEA Staff Report, April.
Slosser, J. E. 1978. The influence of planting date on boll weevil management. Southwestern Entomologist 3:241-246.
Sprott, J. M., R. D. Lacewell, G. A. Niles, J. K. Walker, and J. R. Gannaway.
1976. Agronomic, economic, energy, and environmental implications of short-season, narrow-row cotton production. Texas Agricul- tural Experiment Station, MP-1250C, 23 p.
Taft, H. M., and A. R. Hopkins. 1975. Boll weevils: field populations controlled by sterilizing
emerging overwintered females with TH-6040 sprayable bait. Journal of Economic Entomology 68:551-554.
Thomas, J. G. 1974. The all-out war against the boll weevil. Texas Agricul-
tural Extension Service Report. In 1974 Cotton International 41:52-53, 220-222.
Thomas, J. G., chairman. 1979. Present and future pest management strategies of cotton
and sorghum pests in Texas. Pest Management Strategies, Vol- ume II, Working Papers. Office of Technical Assessment, Con- gress of the united States, 75 p.
Walker, J. K., and G. A. Niles. 1971. Population dynamics of the boll weevil and modified cotton
types. Texas Agricultural Experiment Station, Bulletin No. 1109, 14 p.
Ware, J. 0. 1925. Cotton variety experiments. Arkansas Agricultural Experi- ment Station, Bulletin No. 197, 27 p.
White, J. R., and D. R. Rummel. 1978. Emergence profile of overwintered boll weevils and entry
into cotton. Environmental Entomology 7:7-14.

MANAGEMENT PROGRAMS AND IMPLEMENTATION 557
Yearian, W. C, R. G. Luttrell, A. L. Stacey, and S. Y. Young. 1980. Efficacy of Bacillus thuringiensis and Baculovirus^ heliothis - chlordimeform spray mixture against Heliothis spp. on cotton. Journal of Georgia Entomological Society 15:260-
271.


INDEX
559
Abdul Matin, A. S. M., 164, 170
Abies, J. R., 103, 114, 120, 121, 144, 207, 215, 216, 217, 223, 229, 231, 349, 552
Acala, M., 552 Acariña, 62 ACNPV (nuclear polyhedrosis
virus of alfalfa looper) has shown promise for control of Heliothis spp., 138-141
Adams, C. H., 105, 121 Adams, J. R., 146 Adelberg, E. A., 298 Adkisson, P. L., 4, 19, 24; 25,
117, 121, 124, 176, 229, 249, 258, 341, 343, 346, 378, 417, 434, 525, 552, 555
Agricultural and Mechanical College of Texas, now Texas A&M Univ., 524
Alabama argillacea, cotton leafworm, 4, 5, 6, 65, 237, 253, 330, 332
Aldicarb, 190, 191 Alfalfa looper, Autographa
californica, 137, 138 Alkylating agents, apholate,
aphomide, and aphoxide, 154-155
All, J. N., 357 Allen, G. E., 138, 139, 143 Anderson, F. S., 306, 324 Anderson, R. E., 100 Anderson, T. W., 51, 259 Andow, D., 124 Andres, V., Jr., 171 Andrews, G. L., 4, 19, 148, 385 Andrilenas, P. A., 32, 34,
37, 48, 49, 51, 259
Angus, T. A., 133, 143, 145 Animal and Plant Health
Inspection Service (APHIS), 4, 185, 321, 498, 507, 508, 509, 515, 523
Anonymous, 8, 12, 19, 30, 31, 32, 51, 208, 229, 238, 239, 246, 249, 252, 258, 486, 494
Anthicidae, 63 Anthocoridae (minute pirate
bugs), 106 Anthonomus grandis, boll
weevil, see Boll weevil
Apanteles marginiventris, 216
Aphididae, 62 Aphids, Aphis spp., 64, 329
estimated cotton yield losses from, 339
Aphis gossypii, cotton aphid, 6, 244
cotton losses due to, 329, 332
diflubenzuron ineffective against, 211
insecticide control, 246, 252
Apholate, 154-155 Aphomide, 154 Aphoxide, 154 Apis mellifera, honey bee,
63, 129 Araneida (spiders), 62, 64 Araneidae (orb weavers), 106 Arant, F. S., 346 Arle, H. F., 150 Armyworm, Spodoptera spp.,
105 ARS Metabolism and Radiation
Research Laboratory, Fargo, N. D., 428
Arthropods (cotton): beneficial, 7, 62, 63, 66,
67, 103-108, 110, HI, 115, 120, 216, 314, 315, 318-320
boll weevil, Anthonomus grandis, see Boll weevil
bollworm, Heliothis zea, see Heliothis spp.
cabbage looper, Trichoplusia ni, 105, T4r,~l:08,' 329, 332, 339, 361

560 INDEX
cotton aphid. Aphis gossypii, 6, 211, 244, 246, 252, 329, 332
cotton fleahopper, Pseuda- tomoscelis seriatus, 8, 85, 208, 211, 238, 252, 314, 329, 332, 333, 339, 342, 375, 526
cotton leafperforator, Bucculatrix thurberi- ella, 142, 209, 253, 332
cotton leafworm, Alabama argillacea, 4, 5, 6, 65, 237, 253, 330, 332
entomophagous: 7, 62, 63, 66, 67, 103-108, 110- 111, 115
approaches to use of, in integrated pest man- agement, 118-120
conservation of, 119 efficacy of predators in
suppression of Heliothis spp., 109- 111
factors affecting preda- tor efficacy, 111-114
modeling populations of, and Heliothis spp., for IPM decision making, 114-118
populations of key preda- tors of Heliothis spp., 106-111
fall armyworm, Spodoptera frugiperda, 210, 246
major groups in cotton fields, 62
pink bollworm, Pectinoph- ora gossypiella, 3, 4, 5, 7, 15, 104, 105, 119, 120, 136, 137, 138, 208, 210, 227, 238, 246, 247, 329, 332, 333, 339, 341, 342, 361, 549
predators, 7, 62, 63, 66, 67, 103-108, 110-111, 115
sampling: beneficial arthropods,
318-320
comparison of efficacy of techniques, D-Vac, field observations, sweep method, and unit area 312-315
for pests: major:
boll weevils, 316- 317 Heliothis spp,, 314-316 plant bugs, Lygus, 317
secondary, 317 population densities:
absolute methods, 307 relative methods, 308-
311 variables, sampling
duration, locations, size, and others, 303-307
scouting, 6, 64, 68, 225, 385, 392, 397, 398, 399, 400, 486
spider mites, Tetranychus spp., 329, 340
tarnished plant bug, Lygus lineolaris, 63, 74, 208, 211, 314, 317, 343, 534
tobacco budworm, Heliothis virescens, see Heliothis spp.
Ascher, K. R. S., 211, 229 Aspergillus niger, bacterial
contaminant in boll weevil rearing facil- ity, 286
Autographa californica, alfalfa looper virus (ACNPV) infects pink bollworm larvae, 137, 138
Azinphosmethyl, 48, 49, 82, 92, 94, 192, 217, 224, 252, 256
Azodrin, 119
Bacheler, J. C., 349, 379, 446, 449, 450, 476, 478
Bacillus sphaericus, 286

INDEX 561
jB. thuringiensis, 129, 130, 133, 136, 137, 139, 141, 532
registered for use against Heliothis spp., 138
Bacterial blight, a seedling disease caused by Xanthomonas malva- cearum, 77
Baculovirus heliothis, a nuclear poly- hedrosis virus (NPV) from Heliothis zea, infects only Heliothis spp., 129, 130, 131, 132, 139, 532
registered for use against Heliothis spp., 138
Baerg, W. J., 23, 553 Bailey, C. F., 121, 346 Bailey, J. C., 86, 96, 98,
336, 343, 346 Baker, D. N., 259, 361, 378,
380, 443, 476, 477, 479
Baker, S., 100 Baker, T. C, 204 Baldwin, J. L., 370, 372, 378 Ballmer, G. R., 126 Bandedwinged whitefly,
Trialeurodes abuti- lonea, 62, 64, 246
Barber, G. W., 318, 324 Bariola, L. A., 96, 150, 353 Barker, C. H., 26 Barnes, G., 387, 406 Barnes, K. K., 479 Bartlett, A. C., 155, 162,
170, 272, 295 Batzer, 0. F., 146 Baumhover, A. H., 204, 288,
295 BAY SIR 8514 (insect growth
regulator), a benzoylphenyl urea that has been as effective as diflu- benzuron against boll weevil, 209-210
Beard, R. L., 288, 295 Beauveria bassiana, boll
weevil susceptible to, 136
Beavers, J. B., 232, 234 Becnel, I. J., 349, 356 Bee, M. J., 229 Beet armyworm, Spodoptera
exigua, a secondary pest of cotton, 141, 209, 361
Bell, A. A., 100 Bell, J. v., 134, 143 Bell, K. 0., Jr., 70, 110
121, 126, 235, 328 Bell, M. R., 129, 137, 138,
139, 140, 143, 148, 150, 301, 349
Benedict, J. H., 554 Beneficial insects, 7, 62, 63,
66, 67, 103-108, 110, HI, 115, 120, 216, 314, 315, 318-320
Benskin, J., 227, 229 Benzene hexachloride, 6, 244 Benzoylphenyl ureas (insect
growth regulators), 207-226, 228
BAY SIR 8514, as effective as diflubenzuron against boll weevil, 210
diflubenzuron, only ben- zoylphenyl urea approved for use on cotton, 207, 209
penfluron, was more active than diflubenzuron in inhibiting the hatch of boll weevil eggs, 210
structures and activity of BAY SIR 8514, diflubenzuron, and penfluron, 210
Berger, R. S., 180, 199 Beroza, M., 232 Bibow, W. R., 172 Bielarski, R. V., 261 Bierl-Leonhardt, B. A., 201,
202, 380 Biever, K. D., 235 Bigeyed bug, Geocoris
punctipes, 62, 114, 216
Biological Evaluation Team, 402, 483, 498, 522
Bird, L. S., 12, 19

562 INDEX
Bisazir (chemosterilant), 157 Black fly, Simulíum vittatum,
210 Black, J. H., 387, 406 Blake, H., 51 Bogel, D., 356 Bohmfalk, G. T., 149 Boling, J. C, 125, 327 Boll weevil, Anthonomus
grandis, 3 alternative cotton insect
management strategies, impact on prodüber income in Mississippi, 481-493
alternative host plants in the United States, 53
bacterial control of, 133, 135-136
biology, behavior, and ecology, 54-61
Boll Weevil Eradication Trial (BWET), see under main entry
Boll Weevil Research Lab- oratory, 164, 272, 292
BWSIM (boll weevil simula- tion model), 445
BWSIM model compared with CIM-BW model, 453, 455
simulated results from BWSIM compared with experimental results, 453-454
cause of substantial damage to cotton in Southeast Region, 76
CIM (cotton and insect management simulation model), integration of component models, boll weevil model (CIM-BW), cotton crop model (COTCROP), and Heliothis spp. model (CIM-HEL), into CIM, 464, 466, 469, 472- 473, 475
CIM-BW (cotton and insect management-boll weevil simulation model):
model compared with earlier boll weevil simulation model (BWSIM), 453, 455
schematic of boll weevil and cotton interac- tions, in CIM model, 447
structure, 446 weevil development,
fecundity, feeding, and mortality, 449- 452
subroutine calling sequence, 448
community-wide bollworm management system in Arkansas, an alter- nate to BWET and OPMT, 529-534
control, new directions and technologies for, 7-18
cotton culture in weevil- infested areas, 74, 76, 78
cotton production regions for boll weevil evaluation in 1979, 41
diflubenzuron, effect on, 156-157, 165-166, 209-213, 218-224, 225-226, 411
distribution of, in United States, 55
entomophagous arthropods for control, 104
eradication: biological inquiries and
constraints to belt- wide implementation:
cost, 544-545 federal and state regulatory require- ments, 550-551
maintenance of weevil- free areas and containment areas, 548-549
personnel needs, 547 role of independent
consulting entomolo- gists, 547

INDEX 563
Boll weevil (eradication) components of suppression:
attractant, sex and aggregating, 424-425
diflubenzuron, 427 insecticide, 416-421 pheromone trapping, 423-424
sterile boll weevil release, 159, 425-427
suppression of over- wintering generations, 421-423
economic evaluation of BWET in North Carolina, 497-516
improvements in, 428-430 movement of boll weevil
and its effect on, 415
population suppression, fundamental principles of, 412
simulation models, use of to evaluate suppres- sion measures, 413-414
technology: employed in BWET, 409 for maintaining boll weevil-free areas, 430-433
High Plains Boll Weevil Suppression Program, 539, 548
initial years of infestation in United States, 5
insecticides: an appraisal of cotton
insect control with, 250-251
beginning of use of syn- thetic organic insecticides against, 6-7
cost of insecticides used on cotton, 29-50
for control in Optimum Pest Management Trial (OPMT), Current Insect Control (CIC), and Boll Weevil Eradica- tion Trial (BWET), 256
inorganic, 4, 5, 7, 239
organic, 244 as dusts, 244-245 as sprays, 245
recommended dosages for control of major cotton insects, 252-253
reproduction-diapause control with methyl parathion, 248, 249
resistance to, 246 systemic, 246
juvenile hormone mimics, not effective against, 227
mass rearing: adult emergence, 281-283 as component of weevil
eradication, 265 control of microbial
contamination, 266 development room, 281 diets :
artificial, first suitable formula- tion, 266
Gast diet used for sterile weevils reared for BWET and OPMT, 162
larval and adult, 270 egg harvest and prepara-
tion, 276-277 egg production, 272 emergence cubicles, 282 Gast rearing facility,
266-269 production data for,
293 microbial contamination,
283-292 amino or fatty acid
content, effect on, 287
cell tissues, effect on, 287
control of, 266 detection of, methods
for, 289-290 of insectary-reared weevils, with di- verse aerobic and anaerobic bacteria, 286

564 INDEX
mass rearing (contamination) pheromone production,
effect on, 287 production problems
caused by, 292 sanitary measures
against, 288-289, 291-292
sources of contamina- tion, 284
weevil development, effect on, 286
oviposition: cage, shaker, and
conveyor for, 275 oviposition equipment
and procedures, management of, 274
oviposition room, environment of, 275
pellet-making equipment, 275
pellet-making of adult diet, 273
rackveyor for stacking and conveying planted trays, 280
sterilization of prepared diets, 272
tray processing and egg planting, equipment for, 279
microbials for control of, 133
olfactory and visual stimuli, response to, 57
Optimum Pest Management Trial (OPMT), see under main entry
pheromone: components, isolation,
identification, and synthesis of, 184
dispensing systems, 184- 185
production of, 165 seasonal response of boll
weevil to, 183 pheromone traps:
daylight fluorescent paint (lemon yellow) color on traps attracts the greatest number of weevils, 61
infield traps and insec- ticides for detection and suppression, 191-194
weevil capture in in- field traps: in Arkansas, 194 in North Carolina, 196
male-baited traps around cotton fields for boll weevil suppres- sion, 187-188
pheromone-baited traps for capturing over- wintered weevils, 188-191
Pilot Boll Weevil Eradica- tion Experiment (PBWEE), see under main entry
rapid fruiting of cotton, impact on boll weevil populations, 91
resistance in cotton: frego bract, 81-83 primitive cottons, 84 red leaf, 84-85
sampling for, 316-317 sampling procedures
identical for OPMT and BWET, 321
sterilization: chemosterilants, 154-155,
156 gamma irradiation,
154-157, 425-427 irradiation source,
164 new methods of,
156-157 sterile males:
competitiveness of sterilized weevils, 158
evaluation of sterile weevils released in BWET in 1979, 165- 168
release of sterile males in BWET, 159
requirements for using sterile insects for eradication, 168

INDEX 565
Boll weevil (continued) yield losses in cotton due
to: calculated from field
studies, 335-345 estimated, 329-334
Boll Weevil Eradication Trial (BWET) in North Carolina: 3, 13, 15, 53, 61-62, 110, 154, 162, 164, 185, 249, 432-433, 438, 481, 522-523, 538, 540, 544, 545
area: control zone - Robeson,
Scotland, and Cleve- land counties, 498
eradication zone - Edge- combe, Halifax, Northampton, and Nash counties, 498
components of BWET: areawide diapause boll
weevil control with insecticides, 11, 256, 416-417
education and technical assistance to growers, 11
field scouting, 11, 225 insect growth regulator
(diflubenzuron), selective use of, 225, 427
mandatory participation based on grower referendum, 11
pheromone traps, monitor- ing and suppression of boll weevils with, 11, 423-425
quarantine procedures, 11, 413
sterile boll weevils, release of, 11, 159, 425-427
evaluation of: economic : changes in cotton pro-
duction, 498-501, 509-511
changes in cotton yields, 498-501, 511-514
insecticide applica- tions, 501-506
producer income, 514- 516
public expenditures, 507-509
released sterile boll weevils in eradica- tion trial in 1979, 165-168
purpose of trial, designed to test new methods for insect control, 12
sampling techniques, four compared: field observations by entomologist, D-Vac, sweep net, and unit area sampler, 312-323
Bollworm, Heliothis zea, see Heliothis spp.
Bondy, F. F., 351 Bonell, N. P., 21 Booth, G. D., 326 Boring, E. P., 527, 528, 552,
553 Borkovec, A. B., 154, 156,
157, 170, 173, 175, 176, 230
Bottger, G. T., 349 Bottrell, D. G., 4, 19, 20,
70, 105, 121, 208, 229, 378, 548, 552, 555
Bourland, F. M., 19 Bowen, H. D., 379, 476, 478 Bowlus, S., 233 Boyd, F. J., Jr., 187, 199,
230, 553 Boyer, W. P., 406, 554 Boyette, J. D., 355 Bracon kirkpatricki, a poten-
tial parasite of boll weevil, 104
J.* mellitor, has parasitized boll weevil larvae, 83, 105
Braconidae, 62, 64 Bradley, J. R., Jr., 21, 190,
199, 230, 231, 232, 328, 379, 380, 476, 478, 553
Bradshaw, R. D., 100

566 INDEX
Brazzel, J. R., 7, 19, 21, 24, 121, 123, 137, 143, 199, 230, 231, 248, 258, 343, 346, 353, 406, 416, 434, 478, 514, 517, 553
Brennan, M. M., 204, 205, 381 Bridge, R. R., 99 Briggs, R. W., 174 Brown, L. G., 12, 19, 21, 362,
378, 380, 437, 439, 441, 445, 446, 476, 478, 479, 483, 494, 495
Brown, R. G., 99 Brown, R. T., 233 Bruce, R. R., 443, 476 Brues, C. T., 124, 326, 380,
479 Bryson, J. 0., 204 Bucculatrix thurberiella,
cotton leafperforator, 142, 209, 253, 332
Buck, D. R., 201, 379, 406 Buford, W. T., 84, 96, 97 BUGNET, a computerized pest
management system developed for use in Texas utilizing com- puter models such as MOTHZV in making decisions about pest control, especially for Heliothis spp., 372-377
Bull, D. L., 131, 139, 144, 146, 157, 166, 170, 185, 199, 207, 211, 212, 213, 214, 215, 220, 223, 228, 229, 230, 231, 232, 235, 349, 552
Bullock, H. R., 131, 137, 144, 232
Burgerjon. A., 134, 144 Burke, H. R., 69, 552 Burks, M. L., 171 Burton, V. E., 406 Bush, D. L., 19 Busulfan, a chemosterilant,
155, 158 Butler, G. D., Jr., 378, 456,
476 Buxkemper, W. E., 230, 553
Buxton, D. R., 380, 479 BWSIM (boll weevil simula-
tion model), see also Models, 445, 453-455
Byerly, K. F., 304, 308, 324
Cabbage looper, Trichoplusia ni, 105, 141, 208, 329, 332, 339, 361
Caillavet, D. F., 52 Calcium arsenate, 4, 5-7, 239 Calco oil red N1700 dye, used
in boll weevil larval diet as marker for released sterile adults, 163-165
Calderón, M., 477 Calhoun, S. L., 349, 351, 356 Callahan, R. A., 311, 324 Cameron, D. M., 483, 486,
488, 494, 495 Cameron, J. W. M., 133, 144 Canerday, T. D., 341, 342,
346 Cántelo, W. W., 295 Carabidae, 111 Carde, R. T., 204 Carlson, G. A., 497, 517 Carmine spider mite,
Tetranychus cinna- barinus, 211
earner, G. R., 149 Carpenter, J. E., 204, 205 Carrillo, J. L., 124 Carroll, S. C., 556 Carruth, L. A., 387, 406 Carson, A. D., 149 Carson, R., 8, 19 Casey, J. E., 554 Casida, J. E., 231 Cassidy, T. P., 20 Cate, J. R., 378 Catolaccus grandis, 104 Cave, R. D., 378, 477 Cawich, A., 227, 230 Chaetocnema sp., 62, 313 Chalcidoidea, 62, 64 Chamberlain, W. F., 232 Chambers, H., 163, 170 Chan, B. G., 100 Chang, S. C., 176, 210, 212,
230 Chapman, A. J., 23, 140, 144,
146, 349, 354

INDEX 567
Chapman, O. L., 201, 202, 380 Chemosterilants, 153
apholate, 154 aphomide, 154 aphoxide, 154 bisazir, 156, 157 busulfan, 155 hempa, 155, 156
Cheraosterilization, 153-159 Chesnut, T. L., 121 Chiang, H. C., 171 Childress, D., 172, 292, 295 Chlordimeform, 48, 49, 222 Chloropidae, 62 Chrysopa carnea, common green
lacewing, both adults and larvae are effec- tive predators of Heliothis spp., 106, 110, 120, 216, 320
jC. oculata, goldeneye lace- wing, a known preda- tor of Heliothis spp., 106
—' rufilabris, predator of Heliothis spp., 106
Chrysopa spp., 111 Chrysopidae, 106 Cicadellidae, 62 Cienfuegosia drummondii,
most persistent of the alternate hosts for boll weevil in United States occurs in Coastal Bend region of Texas, 54, 544
C^. yucatanensis, implicated as possible host of boll weevil in Florida, 544
CIM (cotton and insect manage- ment simulation model), see also Models, 437-475, 481- 493
CIM-BW (cotton and insect management-boll weevil simulation model), see also Models, 445
calling sequence for, 448 compared with BWSIM, 454-
455 schematic interactions in,
447 structure, 446
insect development, feed- ing, fecundity, and mortality, 449-452
CIM-HEL (cotton and insect management-Heliothis^ spp. simulation model), see also Models, 456-463, 468
calling sequence for, 457 structure, 456 Heliothis virescens and
_H. zea separated in model because of different rates of development, 456
insect development, feeding, fecundity, and mortality, 456-463
Clark, L. R., 307, 324 Cleveland, T. C, 148, 245,
258 Clower, D. F., 26, 261, 347,
357, 555 Coad, B. R., 5, 6, 19, 20,
22, 239, 258 Coccinellidae, 62, 64, 111 Cody, C. P., 173 Cole, A. W., 555 Cole, C. L., 221, 224, 230,
259, 434, 450, 476, 542, 552
Coleomegilla maculata, 106 Coleomegilla spp.. Ill Coleóptera, 111 Collembola, 63 Collins, H. L., 387, 406, 407 Collins, S. G., Ill, 488,
494 Collops spp., 107, 111 Colorado potato beetle,
Leptinotarsa decemlineata, 211
Common green lacewing, Chrysopa carnea, 106, 110, 120, 216, 320
Comstock, J. H., 5, 20, 237, 258
Cone trap, for control of Heliothis spp., 182
Conoderus vespertinus, tobacco wireworm, 63
Cooke, F. T., Jr., 29, 32, 33, 34, 35, 37, 42, 47, 49, 51, 180, 199, 494, 495

568 INDEX
Cooksey, K. E., 134, 144 Copeland, W. W., 204 Coppedge, J. R., 12, 20, 199,
204, 379, 477 Com (Zea mays) :
constitutes the largest single crop pesticide market in the world and the United States, 31-32
MOTHZV-2 simulation model includes simple crop model for corn, 362- 365
Corylophidae, 62 Corynebacterium humiferum, 286 COTCROP (cotton crop compo-
nent model), see also Models, 440-444
Cotton: aphid, see also Arthropods Belt, 4, 33, 34, 35, 36,
37, 43, 73, 111, 191, 208, 222, 238, 250, 386-387, 481, 482, 493, 524
constitutes the second largest single crop pesticide market in the world and the United States, 31-32
cultivars: bloom patterns for several
cotton strains and cultivars, 87-88
characteristics of those grown in boll weevil- infested areas, 80
current cultivars in boll weevil-infested areas of Southeast, Midsouth, and South- west, 76-79
fruiting and fiber characteristics of more recently released cultivars, 86-91
lint yields of selected lines, 90
rapid fruiting, effect on boll weevil popula- tion, 91-92
field animal community of, 61-65
fleahopper, see Arthropods insect management: alternatives to boll
weevil eradication: community-wide boll- worm management, Arkansas, 529-534
improved cotton insect management for South- east, 534-535
short-season system, Texas, 92-94, 521- 529
beltwide constraints on implementation of BWE and 0PM, 538-551
beneficial insects, 7, 62, 63, 66, 67, 106, 107, 110, 111, 120, 216, 314, 315, 318- 320
Boll Weevil Eradication Trial (BWET) in North Carolina, see under main entry
economic evaluation of BWET in North Carolina, production cost changes, cotton yield, and public expenditures, 497- 516
evolution of, in the United States, from two divergent para- digms:
integrated pest manage- ment , 9
total population management, 9
insect growth regulators (IGR) for, the benzoylphenyl ureas, especially difluben- zuron, 156-157, 159, 165-167, 207-228, 411, 427
insecticides: 3, 5, 6, 32, 34, 36, 48, 49, 50, 82, 92, 119, 192, 217, 224, 237, 238, 239, 244, 245, 246, 247, 250, 252, 253, 254, 255, 256

INDEX 569
Cotton (insecticides) microbial, 130-131 resistance to, 3, 7,
238, Ik^-lkl, 251 integrated pest management,
9-18, 82, 103, 111, 114, 116, 118-120, 208, 226, 359
mismanagement of cotton ecology, a case history, 65-68
Optimum Pest Management Trial (OPMT) in Mississippi, see under main entry
pathogens for control, general character- istics:
bacteria, 134 fungi, 144 protozoa, 135 viruses, 130-132
Pilot Boll Weevil Eradi- cation Experiment (PBWEE) in Mississippi, see under main entry
producer income, impact of alternative insect management strategies on, in Mississippi, 481-493
leafperforator, see Arthropods leafworm, see Arthropods losses due to: arthropod pests, 180, 238,
319-332, 339 calculated by experts,
330-334 calculated from field
studies, 335-338 boll weevil from 1909-
1954, 240-241 insects other than boll
weevil from 1909-1954, 242-243
loss models (simulation), see also Models:
CIM, 439, 442, 447, 464-475, 483-493
COTCROP, 439-445 COTTON, 439 cotton-Heliothis spp.
model, MOTHZV, MOTHZV- 2, and MOTHZV-3, 362-376
GOSSYM, 361 SIMCOT, 361, 439 SIMCOT II, 361, 439 SIMPLECOT, 365-369,
372, 374 production:
acreage average by state, 1974-1978, 75
regions: cotton producing areas
in Mississippi, 483-485
Delphi Region and cotton production subregion, estimated harvested acres for selected years, 42-43
for boll weevil evalua- tion, 40
for cotton cost surveys, 39
for pesticide use survey, 38
resistance: to boll weevils:
in frego bract, 81-83 in primitive cottons,
84 other potential sources
of, 83-84 to Heliothis spp. com-
plex, 84-85 to tarnished plant bugs,
85-86 Courtney, K. D., 171 Covington, B. M., 554 Cowan, C. B., Jr., 25, 125,
261, 324, 327, 347, 349, 350, 351, 354
Cramer, H. H., 330, 340, 346 Cross, W. H., 53, 54, 57, 69,
70, 98, 99, 104, 121, 123, 165, 170, 175, 183, 185, 186, 190, 199, 200, 202, 203, 249, 406, 407, 544, 552
Crowder, L. A., 230 Crowe, B. G., 347 Curculionidae, 54 Curl, L. F., 259, 434 Current Insect Control (CIC),
the approach to cotton insect

570 INDEX
control currently used by producers compared with approach used in BWET and OPMT areas:
control zone (CZ) for BWET- North Carolina compared with eradication zone (EZ):
changes in cotton produc- tion, 498-501, 509- 511
changes in cotton yields, 498-501, 511-514
number of insecticide applications, 501-506
producer income, 514-516 public expenditures, 507-
509 control zone for OPMT-
Mississippi compared with test area:
changes in cotton produc- tion, 491
changes in per acre yield, 488-489
number of insecticide applications, 488- 489
producer income, 489-492 Curry, G. L., 21, 362, 378 Cushman, R. A., 124 Cycloneda, 111
Darden, E., 173 da Silva, R. F. P., 234 Daum, R. J., 136, 144, 148 Davich, T. B., 69, 154, 155,
158, 170, 171, 172, 173, 175, 176, 200, 201, 202, 203, 204, 258, 260, 295, 301, 407, 434, 435, 548, 552, 554
David, W. A. L., 130, 131, 144
Davis, J. W., 25, 101, 125, 261, 311, 324, 327, 349, 350, 351, 354
Davis, R. L., 200 Dawson, E. M., 99 Dawson, J. R., 173, 174, 177,
298 DDT, 6, 7, 8, 48, 49 Dean, H. A., 21, 351, 352
DeBord, D. V., 239, 259, 330, 331, 333, 334, 346
DeJong, B. J., 233 Delong, D. M., 310, 324 DeMichele, D. W., 12, 20, 21,
360, 378 Demilo, A. B., 230, 233 Dempster, J, P., 123 Diabrotica undecimpunetata
howardi, a minor pest of cotton, 63
Diapause control, 11, 249, 256, 391, 394, 401, 416, 417
Dickerson, W. A., 186, 200, 303, 326, 394, 406
Dicrotophos, 48, 49 Dietrick, E. J., 311, 324 Diflubenzuron, an insect
growth regulator and insecticide:
analysis of, in treated boll weevils released in BWET, 166
approved for control: of gypsy moth, 209 only benzoylphenyl urea
approved for use on cotton, 207, 209
boll weevil is only major cotton pest signifi- cantly affected by, 211
component of integrated pest management system, 208
effect on: environment, posttreat-
ment, 213-214 field populations of boll
weevils, 218-224 natural enemies, 215-217 treated sterile males
released in BWET, 165, 411, 427
irradiation plus difluben- zuron to sterilize boll weevils, 153, 156-157, 159
mixed with invert sugar- molasses baits or oil, 218

INDEX 571
mode of action in boll weevil, 211-213
reduced parasitism of tobacco budworm eggs by Trichogramma after eggs treated with, 216-217
structure of, 210 Dilday, R. H., 101 Dillier, J. H., 21 Dimethoate, 48, 49 Dimilin, see Diflubenzuron Dinkins, R. L., 108, 121,
398, 406 Dolichopodidae, 62 Donahoe, M. C, 113, 122, 124 Doolittle, R. E., 205, 381 Dorough, W. H., 176 Dowell, G. C, 554 DuBose, W. P., 346 Dulmage, H. T., 12, 20, 133,
140, 144, 145, 147, 354
Duncan, W. G., 361, 378, 439, 476, 477
Dunlap, T. R., 8, 20 Dunn, H. A., 54, 69 Dunnam, E. W., 349, 351 Dupnik, T. D., 325 DuRant, J. A., 336, 343, 346 Dyson-Hudson, R., 124
Earhart, R. W., 356 Earle, N. W., 12, 20, 156,
157, 159, 160, 165, 166, 171, 173, 176, 177, 266, 295
Earnheart, A. T., 97 Eaton, J., 172 Ebony marker, a genetic semi-
dominant trait in boll weevil which by crossing and back- crossing, provided 99% identifiable weevils for the sterile releases in the BWET, 162-163, 165
Ecology, of cotton insects, 53-68
Economic Evaluation Team, 522 Economics and Statistics
Service, 20
Ectatomma tuberculatum, potential parasite of boll weevil, 104
Eden, W. G., 13, 20, 168, 171, 410, 434
Edminster, T. W., 479 Edwards, R. R., 199, 203 Ehler, L. E., 105, 122 Eichers, T. R., 32, 34, 35,
37, 48, 49, 51, 238, 259
Ekbom, B. S., 122 Elcar, see Heliothis nuclear
polyhedrosis virus Elliger, C. A., 100 Ellman, G. L., 163, 171 Endrin, 48, 49 Enkerlin, S. D., 351 Enterobacter aerogenes, found
on insectary-reared weevils, 286
amino and fatty acids reduced in contami- nated weevils, 286, 287
Entomophagous arthropods, see Arthropods
Environment Evaluation Team, 522
Environmental Protection Agency, 130
EPN, 46, 48, 49, 464 EPN-methyl parathion, 464 Eradication, see Boll weevil
eradication Escarra, B., 483, 494 Eveleens, K. G., 122 Ewing, K. P., 6, 20, 21, 23,
244, 245, 247, 259, 351, 353, 355, 541, 553
Falcon, L. A., 27, 126, 134, 140, 141, 145, 324, 378
Fall armyworm, Spodoptera frugiperda, 210, 246
Favero, M. S., 284, 295 Fawcett, D., 52 Featherstone, R. M., 171 Fenton, J. A., 351 Fenvalerate, a synthetic
pyrethroid insecti- cide effective against Heliothis spp., 46, 48, 49

572 INDEX
Ferrell, C. D., 234 Ferron, P., 134, 145 Field surveys, see Scouting Fife, L. C., 351 Finley, M. D., 528, 553 Flake, H. W., Jr., 298 Fletcher, R. K., 110, 122 Flint, H. M., 154, 158, 164,
172, 181, 200, 210, 211, 230
Forey, D., 230 Formicidae, minor secondary
pests of cotton, 63
Foster, G. G., 177 Fox, A., 51 Frear, D. S., 232 French, B. L., 26 Frevert, R. K., 479 Fricke, H., 159, 172 Fried, M. , 158, 172 Frisbie, R. E., 22, 261, 372,
378, 379, 394, 406, 521, 527, 553, 554, 556
Fryxell, P. A., 69, 552 Fuchs, T. W., 311, 324, 552 Fulgoridea, minor primary
cotton pests, 63 Functional Mathematics
Programming System, 486
Furr, R. E., 149, 355 Fusarium oxysporum f.
vasinfectum, the cause of Fusarium wilt in cotton, in Southeast, 76
Fusarium spp., 77 Fye, R. E., 60, 69, 249, 259,
340, 341, 346, 351, 357, 417, 434
Gaines, J. C., 6, 21, 346, 351, 352
Gaines, R. C., 295, 341, 342, 346, 352, 355, 357, 358
Gallahan, D., 124 Gamma irradiation, see
Irradiation Gannaway, J. R., 100, 378,
556 Gantt, C. W., 327
Ganyard, M. C., 4, 12, 21, 203, 218, 219, 222, 223, 225, 230, 231, 232, 542, 553
Garcia, C., 146 Garcia, R. D., 231 Gard, I. E., 352 Gardner, W. R., 443, 477 Garrison, G. L., 358 Garst, G., 52 Gassner, G., 156, 172 Gast, R. T., 69, 136, 145,
160, 163, 171, 172, 260, 266, 273, 279, 284, 288, 291, 292, 295, 296
Gausman, H. N., 24 Geier, P. W., 324 Genetics, of ebony body boll
weevil, 162 Genotypes, and phenotypes of
boll weevil progeny from crosses of ebony characteristic x wild- type, 163
Gentry, C. R., 180, 200 Geocoris punctipes, bigeyed
bug: beneficial insect, 62, 216 first instar feeds almost
exclusively on tobacco budworm eggs, 114
Geocoris spp.: beneficial insect in
eastern parts of Cotton Belt, 64
predator of Heliothis spp. larvae, 111
Gerard, C. J., 86, 96 Gijswijt, M. J., 233 Gilbert, N., 381 Gilliland, F. R. , Jr., 149,
191, 200 Glass, E. H., 171 Glodt, R. E., 100 Glover, T., 237, 259 Glugea gasti, a protozoan
pathogen that infects Heliothis spp. and pink bollworm, 136
Gonzalez, D., 27, 107, 122, 126, 325, 328
Good, J. M., 11, 21

INDEX 573
Goodenough, J. L., 103 Goodwin, R. W., 288, 296 Gorham, D. P., 104, 122 Gorzycki, L. J., 175 Gossypium anomalum, a cotton
with some resistance to boll weevil, 84
G. arboreum, reduced boll ~ weevil oviposition
about 15%, 84 G. barbadense, SI Seaberry "" strain slightly
reduced oviposition of weevils, 84
G. herbaceum, a cotton with "" fewer anthers than
regular type reduced boll weevil oviposi- tion, 84
G'. hirsutum, 54 ~ some lines show low levels
of resistance to boll weevil, 84, 85
G. hirsutum var. punetatum, implicated as a possible host of boll weevil in Florida, 544
Gradwell, G. R., 328 Graham, H. M., 4, 8, 21, 98,
145, 146, 327 Grandlure, the pheromone of
boll weevil, has been isolated, identified, and synthesized as four terpenoid com- pounds, 184
Grandlure-baited traps: development of, 185-186 dispensing systems for,
184-185 Granulosis virus, one of two
types of viruses that can infect insects, 131
Graves, G. N., 137, 145 Graves, J. B., 7, 21 Graves, T. M., 199, 200 Green, J. M., 81, 96 Greenslade, R. M., 26 Gregory, B. G., 143 Griffin, J. G., 160, 163,
172, 173, 265, 266, 267, 270, 271, 274,
276, 278, 279, 280, 281, 282, 290, 293, 296, 297
Grosscurt, A. C., 211, 231 Grube, A., 498, 499, 517 Gueldner, R. C., 165, 172,
175, 200, 203, 205, 231, 286, 287, 297
Guerra, A. A., 227, 231 Guilavogui, F., 356 Gutierrez, A. P., 12, 21,
25, 308, 324, 361, 378, 439, 477, 479, 555
Gypsy moth, Lymantria dispar, diflubenzuron has been approved for use on, 209
Haemotobia irritans, horn fly, 211
Hagen, K. S., 26, 27, 126 Hajjar, N. P., 211, 231 Halford, W. T., 355 Halictidae, 62 Hall, P. K., 85, 96 Hamel, D. R., 356 Hamer, J. L., 385, 406, 407 Hamill, J. G., 486, 494, 495 Hamilton, A. G., 476 Hammann, I., 235 Hampea sp., a host for the
boll weevil, but not native to the United States, 54
Handy, R. B., 22 Haney, P. L., 15, 22 Hanna, R. L., 27, 121, 346,
352, 356 Hanny, B. W., 96, 98, 346 Harcourt, D. G., 304, 325 Hardee, D. D., 54, 69, 144,
170, 171, 183, 184, 185, 187, 199, 200, 202, 203, 205, 407, 435, 552, 554
Harding, J. A., 221, 224, 231, 311, 324, 325, 327, 542, 553
Harrell, E. A., 278, 283, 293, 297
Harris, F. A., 249, 259, 438, 476, 477, 479, 494

574 INDEX
Harris, L. D., 258, 434 Harris, R. L., 235 Harris, W., 222, 231 Hart, E. J., 172 Hartsog, J. D., 476, 478, 479 Hartstack, A. W., 12, 22, 103,
115, 116, 117, 122, 175, 179, 181, 182, 183, 200, 201, 202, 203, 316, 325, 336, 344, 346, 359, 362, 363, 364, 365, 366, 374, 376, 377, 379, 394, 406, 435, 439, 456, 460, 463, 477
Hassell, M. P., 112, 122 Haynes, D. L., 171, 327 Haynes, J. W., 27, 154, 155,
156, 158, 159, 165, 172, 173, 175, 177
Hazard, E. I., 291, 297 Head, R. B., 385 Healy, M. J. R., 305, 325 Heath, J., 26 Hedin, P. A., 154, 173, 184,
201, 203, 205, 228, 231, 285, 298
Heilman, M. D., 12, 22, 73, 92, 93, 94, 96, 99, 526, 527, 553, 554
Heimpel, A. M., 134, 145 Heinrichs, E. A., 234 Heiser, R. F., 175 Heliothis nuclear polyhedrosis
virus, is host-specific to Heliothis spp,, 129-131, 138, 139
Heliothis spp. (including H. virescens (tobacco budworm) and H. zea (bollworm)): 2'9,~93, 387, 397, 418, 437, 444, 498, 526, 536
as cotton pests: development of several gen-
erations on cotton with- in growing season, 65
midseason or late season, 108
most important of second- ary pests, 63, 64., 208
Bacillus thuringiensis, regis- tered for use against, 129-130, 138-142, 532
Baculovirus heliothis, regis- tered for use against, 129-131, 138-142, 532
Boll Weevil Eradication Trial (BWET) in North Carolina, see under main entry
bollworm and cotton leafworm were the most import- ant pests of cotton before 1885, 237
Chrysopa carnea adults and larvae are effective predators of, 320
CIM (cotton and insect man- agement model, formed from three component models, COTCROP, CIM-HEL, and CIM-BW, 464-475
CIM-HEL, Heliothis spp. component simulation model, 456-463
insect growth regulators for: diflubenzuron, is ineffec-
tive against, 211, 427 juvenile hormone mimics,
reaction to, 227-228 insecticides for control:
frego bract cotton allows delay in use of insecticides, thus protecting beneficial insects that contrib- ute to control of bolIworm/budworm complex, 82-83
insecticides used for boll weevil often kill beneficial insects that control Heliothis spp. and create a serious pest problem beltwide, 68, 104, 105, 238, 412, 541, 543
Paris green more effec- tive than calcium arsenate against bollworm in early years, 239
recommended dosages of principal insecticides for control, 252

INDEX 575
resistance: budworm/bollworm complex
has developed resist- ance to the compara- tively few available effective insecticides, 257
to DDT, 7 to the organochlorine
insecticides in Louisiana, 246-247
role of insecticides in control of, 238-239, 244-246
shift to use of pyrethroid insecticides, 46
toxaphene and endrin effective against bollworms, 244
losses from, in cotton, 329-333, 336, 338-345
capable of being problem in irrigated fields of Lower Rio Grande Valley, 81
management: BUGNET, a computerized
management system, see BUGNET
community-wide bollworm management system in Arkansas, 521-522, 529-534
entomophagous arthropods for, 103, 118-120
insecticides, effect upon, 119
predator and prey density, distribution, and preferences, 112-114
some known predators of, 106
five alternative strate- gies including current insect control used by cotton producers (CIC), for control of bollworm/tobacco budworm complex and boll weevil, were compared to determine the specific changes in acre yield and
number of insecticide applications for each alternative strategy and for CIC, 483
modified cotton produc- tion system for early maturity to escape late season infesta- tion with, 95
Optimum Pest Management Trial (OPMT) in Mississippi, see under main entry
significant advances in control of, 4
modeling populations of entomophagous arthro- pods and Heliothis spp. with a cotton- Heliothis simulation model (MOTHZV) and also, revised models, 359-377
MOTHZV-2, 114-117 pheromone:
dispensing system for pheromones used as attractants, 181-182
disruption with, for control of, 183
identification of components of pheromone of H. virescens (virelure) and H. zea, 180-181
traps, development of, 182 plant resistance to, in
glabrous and nectari- less cottons, 84
sampling: in BWET and OPMT, whole
plant, terminal squares, and bolls, 322
of eggs, larvae, pupae, and adult populations, 314-316
Helms, T. J., 288, 298 Helms, W. F., 199 Hemiptera, 111 Hempa, a chemosterilant, 155,
158 Hendricks, D. E., 180, 181,
201, 205, 325, 374, 379, 381, 406

576 INDEX
Henneberry, T. J., 143, 150 Henson, J. L., 22, 379 Henson, W. R., 328 Hervey, G. E. R., 298 Herzog, D. C., 325 Hesketh, J. D., 380, 439, 443,
476, 477, 478, 479, 495
Hibiscus sp., 54 H. syriacus, 544 High Plains Boll Weevil
Suppression Program, 538, 539, 548
High pressure liquid chroma- tography, 166
Hightower, B. G., 27, 258 Hill, A. S., 204 Hill, R., 170 Hillhouse, T. L., 124 Hinds, W. E., 6, 22, 23 nines, R. W., 204 Hippodamia convergens, lady
beetle, 106, 215 Hippodamia sp.. Ill Hobgood, J. M., Jr., 295 Hoffman, J. D., 200 Hoffmann, C. H., 12, 22 Hogg, D. B., 458, 459, 460,
477 Holbrook, F. R., 324 Holder, S. H., Jr., 495 Hollier, D. D., 177 Holling, C. S., 114, 122 Hollingsworth, J. P., 22,
122, 182, 201, 374, 379, 394, 406, 477
Hood, C. E., 124 Hood, J. E., 19 Hooker, P.A., 170 Hopkins, A. R., 70, 204, 234,
237, 246, 247, 260, 330, 332, 333, 346, 347, 351, 353, 356, 357, 556
Horn, B., 230 Horn fly, Haemotobia irritans,
211 Hornyak, E. P., 357 Hostetter, D. L., 146 Houghtaling, J. E., 98 House, V. S., 144, 211, 215,
216, 217, 220, 222, 223, 224, 229, 231, 349, 552
House fly. Musca domestica, 210
Howard, L. 0., 5, 22, 524, 553
Howell, J. F., 288, 298 Huddleston, P. M., 69, 170,
171, 199, 200 Huffaker, C. B., 9, 11, 12,
22, 109, 122 Hughes, R. D., 324, 381 Hunt, H. H., 379 Hunter, D. K., 150 Hunter, R. C., 81, 96, 554 Hunter, W. D., 5, 22, 23,
188, 201, 450, 477 Hymenopappus flavescens, 544 Hyslop, J. A., 333, 347
Ignoffo, C. M., 12, 23, 131, 133, 136, 137, 138, 139, 144, 145, 146, 147, 148, 235, 288, 298
Index of prices, the prices paid by farmers for agricultural chemi- cals, 1961-1979, 36
Insect growth regulator (s): BAY SIR 8514, 209-210, 226 diflubenzuron, 153, 156-157,
159, 165-166, 208-226 juvenile hormone (JH)
mimics, 227-228 penfluron, 156-157, 209-
210, 226 Insecticides :
aldicarb, 190, 191, 246, 252
application of: aerial, 237, 252 as dusts, 5, 6, 239 as low volume sprays,
245, 252-253 quantities applied by
region, 46, 48-50 with infield traps, 192
appraisal of merits and limitations of, in suppressing insect populations, 250-251
azinphosmethyl, 48, 49, 82, 92-94, 192, 217, 224, 252, 256

INDEX 577
insecticide used against diapausing boll weevils in BWET, 256
azodrin, 119 benzene hexachloride, 6, 244 calcium arsenate, 5, 239,
244 chlordimeforra, 222, 247,
252 control of cotton insects
improved with advent of organochlorine, organophosphorus, and carbamate insecticides, and the phasing out of inorganic compounds, 251
cost: of developing and register-
ing, 257 to producer for use on
cotton, 32, 34, 36, 42-50, 238
cotton cultivar-insecticide interaction, 82
DDT, 6, 48, 246 deleterious effect of conven-
tional insecticides to entomophagous arthro- pods, 108, 119
dosage: of miticides for control
of spider mites, 254-255
recommended dosages of principal insecticides for major cotton insects, 252-253
inorganic, calcium arsenate, lead arsenate, Paris green, and sulfur, 5, 6, 339, 244
insect pest management system, place in, 256
malathion, 224, 252 methyl parathion, 217, 244,
249, 252 insecticide used against
diapausing boll weevils in OPMT, 256
organic: carbamate, carbaryl, 244 organochlorines, develop-
ment of, aldrin,
benzene hexachloride, chlordane, dieldrin, endrin, heptachlor, and toxaphene, 244
organophosphates, develop- ment of, azinphos- methyl, demeton, EPN, malathion, methyl parathion, parathion, and sulprofos, 244-247
pyrethroids, permethrin and fenvalerate, 247
systemlos, aldicarb, disulfoton, and phorate can be applied to cotton seed or in the seed furrow, 246
pesticide use survey, 32, 34, 35, 37-40
reproduction-diapause con- trol with, 248-249
resistance to: organochlorines:
in boll weevil, 246 in 25 species of
insects and spider mites, 246
organochlorine and organophosphorus insecticides in Heliothis spp. com- plex, 238, 247
organophosphorus insecti- cides in 4 species of spider mites, fall armyworm, and banded- winged whitefly, 246
Integrated pest management, 9-13, 82, 103, 111, 114, 116, 119, 226, 359
Irish, M. A., 124 Irradiation, for boll weevil
sterilization, 153-159 source, cobalt-60 and
cesium-137, 164 Isley, D., 6, 23, 83, 96,
188, 201, 449, 450, 478, 529, 553
Ivie, G. W., 157, 170, 173, 211, 212, 229, 231, 232
Ivy, E. E., 6, 20, 21, 23, 351, 353, 355

578 INDEX
Jackman, J. A., 22, 379 Jackson, H. B., 521 Jackson, R. D., 443, 478 Jacobson, M., 204, 228, 232 Jacobson, S. N., 124 James, W., 260, 353 Jaques, R. P., 131, 147 Jawetz, E., 284, 298 Jay, D. L., 150 Jenkins, J. N., 73, 81, 82,
84, 85, 86, 96, 97, 98, 99, 100, 162, 173, 451, 478, 555
Jenkins, R., 51 Jenson, R. L., 310, 325 Johnson, E., 20, 97 Johnson, W. L., 104, 123, 200,
202, 203, 219, 222, 232
Jones, J. E., 83, 85, 97 Jones, J. S., 81, 97 Jones, J. W., 19, 362, 378,
379, 380, 437, 438, 439, 441, 443, 444, 445, 446, 447, 449, 450, 451, 453, 454, 455, 476, 478, 479, 483, 494, 495
Jones, R. E., 324 Jones, S. L., 121, 122, 123,
204, 229, 231, 346, 353, 552
Juvenile hormone (JH) mimics, insect growth regula- tors that can severely disrupt normal insect growth and develop- ment, 227-228
Kamprath, E. J., 478 Kanavel, R. F., 143 Karandiros, M. G., 305, 325 Kearney, J. F., 200, 203, 406 Keever, D. W., 215, 216, 222,
232, 542, 553 Keller, J. C, 148, 170, 184,
201 Keller, K. R., 30, 51 Kennett, C. E., 122 Kent, A. D., 300 Khalil, F., 230 Kido, K., 126 Kimbrough, J. J., 406 Kinard, H. C., 124
Kincade, R. T., 463, 478 King, E. G., 124 Kinzer, R. E., 20, 353 Kiritani, K., 109, 123 Kishaba, A. N., 381 Kittock, D. L., 150 Klassen, W., 155, 173, 174,
347 Klun, J. A., 180, 181, 200,
201, 202, 204, 205, 377, 380
Knipling, E. F., 11, 12, 13, 20, 113, 115, 123, 154, 155, 168, 169, 174, 175, 191, 195, 202, 249, 250, 260, 359, 380, 409, 410, 412, 414, 416, 424, 434, 435
Knott, C. M., 295 Kogan, M., 304, 309, 310,
312, 317, 321, 325, 326, 327
Koonce, K. L., 357 Kotter, E., 203 Krenz, R., 32, 37, 45, 46, 52 Kroop, S. F., 124 Kuhn, T. S., 13, 23 Kulash, W. M., 353
LaBrecque, G. G., 154, 174 Lacewell, R. D., 517, 527,
554, 556 Lacey, L. A., 210, 232 Lactobacillus plantarum,
sometimes found contaminating insec tary-reared weevils, 286
Labren, O. K., 172 Lam, J. J., Jr., 200, 295,
406 Lambert, J. R., 378 Lambert, L., 85, 97, 99 Lambert, W. R. A., 200, 521 Laminated plastic baits, as
dispensing systems for Heliothis spp. pheromone, 181
Landin, M., 172, 295 Langsford, E. L., 32, 34, 35,
52 Laster, M. L., 12, 23, 29,
478

INDEX 579
Lathridiidae, 62 Latson, L. N., 85, 98 Lawrence, R. K., 114, 123 Lawson, F. R., 200 Lead arsenate, in early part
of century was one of the principal insecti- cides used to control cotton leafworm, 5, 239
Lee, G. H., 201, 202 Leggett, J. E., 175, 179, 186,
195, 198, 199, 202, 394, 406, 407, 435
Leigh, T. F., 27, 96, 122, 126, 307, 318, 325, 328, 378, 477
Leipzig, P. A., 324, 378 Lentz, G. L., 353 Leopold, R. A., 171, 175, 233,
234 Lepone, G., 201, 202 Leuconostoc mesenteroides,
isolated from insect- ary-reared weevils, 286
Liapis, P. S., 105, 123 Lickerish, L. A., 26 Lin, Y. N., 32, 34, 35, 43,
48, 49, 52 Lincoln, C., 81, 96, 98, 247,
248, 260, 529, 530, 554
Lindig, 0. H., 160, 161, 162, 163, 164, 172, 173, 174, 175, 176, 200, 265, 266, 269, 273, 296, 297, 298, 300
Lindquist, A. W., 154, 174, 175
Lindquist, D. A., 170, 171, 204, 324, 351
Lingren, P. D., 25, 110, 123, 124, 125, 205, 233, 327, 350
Linstone, H. A., 44, 52 Lippke, L. A., 554 Litsky, B. Y., 288, 298 Litsky, W., 298 Little, V. A., 64, 69 Lloyd, E. P., 3, 11, 12, 15,
23, 24, 54, 69, 155, 167, 175, 177, 179, 187, 190, 195, 198,
200, 202, 203, 204, 207, 218, 219, 223, 232, 248, 249, 259, 260, 261, 325, 344, 347, 355, 406, 407, 424, 425, 435, 444, 478, 540, 542, 554
Lockwood, D. F., 175, 202, 435
Loden, H. D., 353 Loew, W., 324, 378 Longoria, R. R., 357 Lopez, J. D., Jr., 22, 110,
122, 123, 182, 200, 203, 325, 379, 477
Lovestrand, S. A., 211, 232 Lowe, A, M., 301 Lowry, W. L., 348, 354 Luck, R. F., 324 Lue, P. S., 177 Lukefahr, M. J., 22, 69, 84,
96, 98, 100, 125, 247, 260, 341, 342, 347, 544, 552, 553, 554
Lund, H. 0., 353 Lu Po-yung, 233 Luttrell, R. G., 140, 147,
151, 353, 555, 557 Lygaeidae (big-eyed bugs),
predators known to attack Heliothis spp., 106
Lygus hesperus, lygus bug, a primary pest species of cotton in Western United States, 208, 361
L. lineolaris, tarnished plant bug, 63, 74, 208, 211
collection by D-Vac, 317 insect management programs
can be devastated by, 534
lint yield affected by, in untreated fields, 343
pest of cotton (primary) in Midsouth, 208
Lygus spp., 8, 64, 65, 85- 86, 104, 119, 238, 253, 256, 340, 387, 536

580 INDEX
Lymantría dispar, gypsy moth, diflubenzuron is registered for use against, 209
Lyon, L. R., 288, 298
MacGown, M. W., 286, 287, 299 MacGregor, J. T., 26 MacLeod, J., 306, 325 Madelin, M. F., 134, 148 Magnoler, A., 288, 299 Malathion, 224 Mally, F. W., 188, 203, 524,
554 Malone, 0. L., 174, 297, 298 Malvaviscus drummondii, a
possible alternate host of boll weevil in south Florida, but its isolation makes it unlikely, 544
Mansager, E. R., 214, 232 Marking with calco oil red
N1700 dye for identi- fication of mass- reared sterile adult boll weevils, 163-164
Marlatt, C. L., 330, 333, 347
Marshall, J. H., 295 Marston, N. L., 310, 326 Martignoni, M. E., 150, 301 Martin, D. F., 23, 27, 69, 98,
143, 146, 199, 259, 288, 299, 347, 353, 357, 434
Martinez, A. J., 301 Martinez, E., 145 Martouret, D., 144 Mass rearing of sterile boll
weevils for release in BWET, procedures for, 155-162
Masud, S. M., 527, 554 Mating, of sterile male boll
weevils and native females, theoretical possibilities, 166- 167
Mattesia grandis, a protozoan pathogen that infects Heliothis spp. and pink bo11worm, 136
Mattix, E., 173
Maxwell, F. G., 96, 97, 98, 99, 100, 125, 555
Mayer, M. S., 175 Mayeux, H. S., 349 Mayse, M. A., 307, 309, 326 McCarty, J. C., Jr., 84, 96,
97, 98, 99, 173 McClendon, R. W., 19, 378,
437, 439, 476, 479, 483, 494, 495
McCommas, D. W., Jr., 121 McCoy, C. W., 134, 147 McCoy, J. R., 27, 156, 175,
177, 260, 312, 316, 317, 325
McDaniel, S. G., 105, 110, 118, 123, 215, 232, 464, 465, 479
McDonough, L. M., 200 McEwen, F. L., 288, 298 McGarr, R. L., 139, 147, 148,
349, 354, 357 McGough, J. M., 199 McGovern, T. P., 230, 232 McGovern, W. L., 83, 98, 99,
121, 123, 165, 173, 175, 177, 202, 203
McGuire, J. U., Jr., 123, 202, 435
McHaffey, D. G., 156, 170, 175, 176, 233
Mcintyre, R. C., 204, 555 McKibben, G. H., 163, 165,
170, 171, 173, 175, 179, 184, 185, 200, 201, 202, 203, 394, 406, 407, 435
McKinion, J. M., 361, 378, 380, 438, 439, 479
McLaughlin, J. R., 137, 147 McLaughlin, R. E., 133, 135,
136, 139, 144, 147, 148, 172, 173, 212, 232, 266, 285, 286, 287, 288, 297, 299
McLeod, P. J., 131, 148 McMeans, J. L., 478 McNeil, G. L., 20 Melnick, J. L., 298 Meloidogyne incognita, root-
knot nematode, is prevalent in cotton fields in Southeast Region, 76

INDEX 581
Melville, D. R. , 149 Menn, J. J., 209, 227, 232 Meredith, W. R., Jr., 85, 86,
96, 98, 99, 346 Merkl, M. E., 69, 171, 175,
202, 260, 347, 355, 435, 478, 554
Metarrhizium anisopliae, boll weevils are suscepti- ble to infection by this fungus, 136
Metcalf, R. L., 214, 233 Methomyl, 46, 48, 49, 67 Methyl parathion, 7, 29, 46,
48, 49, 67 for reproduction-diapause
control : in large-scale experiment
in Big Bend area of Texas, 248
insecticide used in OPMT, 401
Meyer, J. R., 98 Michael, C., 52 Microbial agents, 129-142
Heliothis nuclear polyhe- drosis virus and Bacillus thuringiensis can both suppress moderate to low popu- lations of Heliothis zea and H. virescens, Î29"
nuclear polyhedrosis virus from alfalfa looper (ACNPV) infects pink bollworm larvae, 137
Serratia marcescens, a bacterium, can kill the boll weevil, 133
two protozoan pathogens, Mattesia grandis and Glugea gasti, infect the bollworm, the tobacco budworm, and pink bollworm, 136
Micrococcus luteus, bacteria infecting boll weevils, 286
M. varians, bacterial con- ~" taminant of boll
weevils, 287 Miles, L. R., 231 Miller, A. E., 21
Miller, E., 301 Miller, P. A., 73 Minyard, J. P., 205 Miridae, 111 Mississippi Cooperative
Extension Service, 4 Mississippi Crop and Live-
stock Reporting Service, 485, 486, 495
Mistric, W. J., Jr., 352, 357, 541, 554
Mitchell, E. B., 24, 69, 158, 170, 171, 172, 175, 183, 186, 192, 199, 200, 201, 203, 232, 394, 407, 554
Mitchell, E. R., 12, 24, 180, 183, 204, 205, 381, 554
Mitchell, H. C, 54, 57, 69, 70, 81, 99, 121, 123, 199, 202, 407
Mitlin, L. L., 54, 70 Mitlin, N., 70, 173 Models (simulation), for
cotton insect pest management:
BWSIM (J>oll weevil simula- tion model), 445, 453-455
CIM (cotton and insect management simula- tion model), formed from the boll weevil model (CIM-BW), the Heliothis spp. model (CIM-HEL), and the cotton crop model (COTCROP), was structured to aid in development and evaluation of insect pest control strate- gies on yield, insecticide usage, and cost of insect management, 464-475, 481-493
CIM-BW, based on BWSIM, 445-455
CIM-HEL, 456-463 COTCROP, based on SIMCOT
and COTTON, 439-445

582 INDEX
cotton growth model, 439-440
soil model, 441-445 CIM-BW (cotton and insect
management-boll weevil simulation model:
model compared with BWSIM, from which it was modified, 453, 455
schematic of boll weevil and cotton inter- actions, 447
structure, 446 weevil development,
fecundity, feeding, and mortality within CIM-BW model, 449- 452
subroutine calling sequence, 448
CIM-HEL (cotton and insect management-Heliothis spp. simulation model), with 11. virescens and Ü* ^^^ separated because of different rates of development, 456, 458-460
calling sequence for, 457 data on insect development,
fecundity, feeding, and mortality, 456-463
structure, 456 COTCROP (cotton model),
developed to study insect and other management strategies including irrigation and nitrogen fertili- zation and composed of a crop growth model and soil model, 439-445
COTTON, a cotton model developed for western cotton production under irrigation, 439
GOSSYM, a more recent cotton model than SIMCOT but derived from it with a highly detailed model of soil, water.
and plant-root rela- tionships and plant growth dynamics algorithm, 361
MOTHZV (Heliothis popula- tion model), 362
flowchart for, 363 MOTHZV-2, a more detailed
population model than MOTHZV, that included simple crop models for corn, cotton, and sorghum; the FORTRAN computer program for MOTHZV-2 consisted of a main computer program and 16 subroutines, 362, 365
flowchart for, 364 use in BUGNET program in
Texas, 374 MOTHZV-3:
produced by addition of a Heliothis damage sub- model, 369-372, 374-377
revision of MOTHZV-2 with replacement of original cotton model with SIMPLECOT, 365-369
MOTHZV-4, a simplified version of MOTHZV-3 which lacked versa- tility of MOTHZV-3 but could predict timing of Heliothis spp. oviposition, 375
use with BUGNET, 375 SIMCOT, an early cotton
model developed to calculate daily production and dis- tribution of photo- synthate of cotton, under southeastern conditions, 361, 439
SIMCOT II, a revision of SIMCOT to interface with insect models to fit California environment and cotton varieties, 361, 439

INDEX 583
SIMPLECOT, was derived from SIMCOT by adding a submodel for cotton fruiting behavior that interfaced with MOTHZV- 3, 365-369
Modified Optimum Pest Manage- ment (MOPM), 482
Monocrotophos, 46, 48, 49 Montoya, E. L., 139, 148 Moody, D. L., 232 Moody, R. J., 387, 406, 407 Moore, G. C., 553 Moore, L. , 406 Moore, R. F., Jr., 27, 156,
157, 175, 177, 211, 212, 228, 233, 297, 347
Moreland, R. W., 353 Moritz, R. J., 199 Morris, R. F., 304, 306, 324,
326 Morrison, R. K., 144, 177,
229, 231, 235, 349 Moss, A. M., 124 Mott, D. G., 304, 326 Mulder, R., 211, 233, 234 Mulkey, J. R., 99, 555 Mulla, M. S., 232 Mullinix, B. G., 204, 205 Mullins, P. W., 355 Murphey, S. M., 483, 495 Musca domestica, house fly,
154, 210 Mjonaridae, 62
Nabidae (damsel bugs), beneficial insects, 63, 64, 106, 111
Nabis spp., 216 Nagasawa, S., 288, 299 Nail, B. J., 173 Namken, L. N., 12, 22, 24,
73, 86, 87, 96, 99, 526, 527, 553, 554
National Academy of Sciences, 168, 176
National Cotton Council, 180, 239, 260, 334, 347
National Cotton Council Technical Subcommittee, 550, 551, 554
Neeb, C. W., 204, 539, 555 Nemec, S. J., 7, 24, 211,
233, 552
Nemny, N. E., 229 Neter, E., 283, 299 Neurocolpus nubilus, clouded
plant bug, can be a pest of cotton, 63-65
Newsom, L. D., 4, 13, 24, 97, 119, 123, 258, 434, 481, 495
Nichols, F., 69, 199 Nicholson, W. F., Jr., 444,
463, 479, 555 Nilakhe, S. S., 20, 165, 171,
176, 177 Niles, G. A., 27, 99, 100,
378, 555, 556 Nishimura, H., 299 Noble, J. M., 230 Noble, L. W., 8, 24 Nolting, D. J., 380, 479 Nomuraea rileyi, a fungus
pathogenic to boll weevils, 136
Norgaard, R. B., 517 Norland, J. F., 173, 174 Norman, J. W., 22, 96, 553 North Carolina Crop Reporting
Service, 501 Nosky, J. B., 357 Notoxus spp., predators of Heli-
othis spp., 107, 111 Nuclear polyhedrosis virus:
ACNP, from alfalfa looper, 141
Baculovirus heliothis, 129- 131, 138, 139
Nucleopolyhedrosis, a disease caused by a nuclear polyhedrosis virus, 131
O'Dell, T. M., 288, 299 Oldacre, S. W., 297 Oliver, B. F., 84, 85, 99 Oliver, J. E., 209, 233 Olsen, C. M., 200 Oman, P., 171 Optimum Pest Management (OPM),
482 Optimum Pest Management-Boll
Weevil Eradication, 482 Optimum Pest Management Trial
(OPMT) in Mississippi, 3, 13, 15, 53, 61-62, 110, 159, 185, 249, 481-482, 522, 538, 544

584 INDEX
components of OPMT: diapause control (with full
reimbursement to producers for costs):
insecticide costs per acre for 1978, 1979, and 1980, 385
insecticide usage, 249, 256-257, 391, 394, 401
inseason control of cotton insects based on scout- ing reports, 392, 398, 400
pinhead square application of insecticides, 392, 397
planting dates, 392, 397 scouting, 392, 397-399 mapping, 391, 393
stalk destruction, 394, 397
trapping: blacklight tra^s for Heliothis spp., 392
pheromone traps: cone Heliothis spp.
traps, 392-397 Leggett traps, 392-397 modified Mitchell
traps, 392-397 contractual agreements, 391 location of: Panola County, Mississippi,
site of trial, 387 Pontotoc County,
Mississippi, control site where normal procedures were followed (Current Insect Control (CIC)), 387
objectives and goals, 388 Orius insidiosus, most
numerous predator in area, 318
personnel : advisory, 389 for operations, 388-390
public awareness and education, 389, 391
sampling techniques, four compared:
field observations by entomologist, 312-323
D-Vac, 312-323 sweep net, 312-323 unit area sampler, 312-323
support for: Mississippi Department of
Agriculture and Commerce, Division of Plant Industry, 403- 404
research groups, 404-405 yield and insecticide usage
determined by regres- sion analysis, repli- cated experiments and simulation, 438
Orius insidiosus, predator of Heliothis spp., 106
Orius spp., 62, 64, 111, 114 —' tristicolor, predator of
Heliothis spp., 106 Orthoptera (Saltatoria), 62 Owen, W. L., Jr., 258, 356,
434 Oxborrow, G. S., 295 Oxyopes salticus, a predator
of Heliothis spp., 106
Parathion, 48, 49 Parencia, C. R., Jr., 4, 21,
24, 86, 99, 180, 204, 237, 239, 246, 247, 249, 250, 258, 259, 261, 311, 316, 317, 326, 341, 343, 347, 350, 351, 353, 354, 355, 553
Paris green, 5, 239 Parker, R. D., 91, 95, 99,
524, 525, 526, 529, 541
Parrott, W. L., 83, 85, 96, 97, 98, 99, 100, 173, 478, 544, 555
Parvin, D. W. , Jr., 23, 29, 51, 52, 199, 479, 481, 486, 494, 495, 517
Pate, T. L., 143 Patti, J. H., 139, 149 Pectinophora gossypiella,
pink bollworm, 3, 4, 5, 7, 15, 104, 105, 119, 120, 136, 137

INDEX 585
138, 208, 210, 227, 238, 246, 247, 329, 332, 333, 339, 341, 342, 361, 549
Penfluron, insect growth regu- ator, 156-157, 209-210, 226
Pénicillium spp., 286 Pentatomidae (Asopinae), Heli-
othis predator. 111 Percy, R. G., 19 Perkins, J. H., 4, 9, 11, 13,
24, 168, 176 Perkins, W. D., 297 Permethrin, 46, 48, 49 Pesticide use, see also
Insecticides, 30 survey and results, 37, 38,
40, 44 Pest management strategies,
implementation, 16 Peters, P. K., 177 Pfrimmer, T. R., 139, 149,
204, 237, 355 Phalacridae, 63 Phaltan, 286 Phene, C. J., 378 Phillips, G. B., 300 Phillips, J. R., 4, 15, 21,
25, 325, 521, 532, 533, 555
Ph3miatotrichum omnivorum, causes root rot of cotton, 81
Pickett, A. D., 6, 25 Pierce, W. D., 5, 23, 25, 104,
124, 477 Pieters, E. P., 140, 149, 308,
311, 326, 355, 356 Pilot Boll Weevil Eradica-
tion Experiment, Mississippi, 11, 15, 153, 155, 185, 249, 266, 410, 411, 413, 415, 416, 417, 418, 419, 423, 540
Pilot pest management project, 389
Pimentel, D., 119, 124 Pink bollworm, Pectinophora
gossypiella, see Arthropods
Pinnell, R. E., 123 Pitre, H. N., Jr., 107, 122,
124, 325
Pitts, D. L., 356 Plimmer, J. R., 200, 201,
202, 380 Poinar, G. 0., Jr., 291, 299 Pomonis, G., 172 Ponder, W. W., 326 Pons, W. J., 235 Populations of insects:
boll weevil, effect of short season cottons on, 91-94
differences in predator populations :
by cotton varieties, 107 by location, 107
modeling populations of entomophagous arthropods and Heliothis spp. for IPM decision making, 114-118
suppression, of Heliothis spp., efficacy of predators, 109-120
fundamental principles of, 410-412
Post, G. B., 6, 25 Post, L. C., 211, 233 Predators, of cotton insects,
particularly Heliothis spp., 106- 120
Price, P. W., 105, 112, 114, 124, 326
Price, R. G., 356 Primiani, M., 201, 202 Production indexes, cost of,
36 Protozoa, Mattesia grandis
and Glugea gasti infect Heliothis spp., 136
Pruitt, G. R., 233, 556 Pryor, N. W., 144 Pseudatomoscelis seriatus,
cotton fleahopper, 8, 63, 64, 85, 208, 211, 238, 252, 314, 329, 332, 333, 339, 342, 375, 526
Pseudomonas aeruginosa, 286 Psocoptera, 63 Puleo, J. R., 295 Pythium spp., 77

586 INDEX
Quaintance, A. L., 110, 124, 320, 326, 372, 380, 463, 479
Rabb, R. L., 9, 13, 25, 380 Ramsey, D. A., 122 Ranney, C. D., 99 Raulston, J. R. , 205 Raun, E. S., 288, 298, 299 Ray, L., 70, 204, 327, 555 Reddy, M. S., 100 Redfern, R. E., 232 Red imported fire ant,
Solenopsis invicta, 104
Reduviidae, 111 Reduviolus alternatus,
a predator of Heliothis spp., 106
_R. americoferus, a Heliothis spp. predator, 106
R. roseipennis, 114 Ree, B., 356 ~" Reeves, S. A., 96 Regional Extension Educational
Advisory Committee, 389
Reproduction-diapause control, 188, 247, 249-251, 256-257, 391, 394, 401
Resistance, to boll weevil in cotton, 81-85
Reynolds, H. T., 4, 25, 119, 124, 171, 318, 326
Reynolds, W. T., 126 Rhizoctonia solani, cause of
seedling disease in cotton, 77
Rhizopus spp., 286 Rhyne, C., 98 Rice:
acreage in Mississippi, 485 yield, 487
Richmond, C. A., 311, 327, 349, 356
Ridgway, R. L., 3, 4, 6, 8, 12, 15, 20, 22, 25, 26, 103, 106, 108, 113, 117, 119, 121, 122, 123, 124, 125, 144, 190, 199, 204, 215, 230, 233, 320, 324, 327, 346, 350,
351, 353, 356, 379, 477
Riley, C. v., 104, 125, 237, 261
Ripper, W. E., 6, 26 Ritchie, J. T., 443, 479 Roach, S. H., 54, 70, 188,
204, 307, 316, 327, 3.94, 407, 548, 555
Roberson, J. L., 163, 173, 174, 176, 177, 292, 297, 298, 299, 301
Roberts, D. W., 134, 149 Rodriguez, R., 510, 511,
517 Roelofs, W. L., 180, 204 Rogers, R, L., 357 Rogers, S., 52 Rollinson, W. D., 299 Romine, C. L., 143, 349 Romkens, M. J. M., 476 Root-knot nematode,
Meloidogyne incognita, 76, 215
Root rot, of cotton, caused by Phymatotrichum omnivorum, 81
Rothrock, M. A., 553 Roussel, J. S., 7, 21, 26,
246, 261, 295, 337, 338, 347, 349, 525, 555
Ruesink, W. G., 307, 310, 327 Rummel, D. R., 191, 204, 216,
217, 220, 221, 224, 233, 235, 249, 258, 261, 434, 539, 540, 541, 542, 544, 548, 549, 552, 555, 556
Runkle, R. S., 284, 288, 300
Salter, S. S., 200 Salticidae (jumping spiders),
106 Saltmarsh caterpillar,
Estigmene aerea, 209 Sampling, arthropods in
cotton, 303-323 Sarmiento, R., 230, 232 Sartor, C. R., 406, 407 Scales, A. L., 175, 353 Scelionidae, 62 Schlinger, E. I., 324 Schreiner, I., 124

INDEX 587
Schroeder, W. J., 211, 234 Schuster, M. F., 83, 85, 97,
99, 100, 107, 125, 311, 327
Schwab, G. 0., 443, 479 Schwartz, P. H., 329, 332,
335, 336, 347, 356 Schwarz, M., 230, 232 Scott, H. A., 148 Scott, W. P., 191, 202, 203,
204, 258, 303, 435, 554
Scouting, 6, 68, 385, 386, 392, 397, 399, 486
a key component of present- day pest management strategies, 6
control decisions based on examination of cotton fields for presence of adult boll weevils or damaged fruit, 225
data gathered by scouts include in addition to insect populations, stage of cotton crop, current growing condi- tions, past history, and potential yield, 398
percentage of cotton squares damaged by bollworms reported by OPMT scouts, 1978-1980, 399
scout training workshops conducted for OPMT, 391
whiteflies, thrips, aphids, Lygus, fleahoppers, Heliothis spp., clouded plant bugs, and boll weevils are the pests the trained scout looks for and records for eastern part of Cotton Belt, 64
Scymnus, 111 Seale, R. D., 483, 486, 488,
494, 495, 496 Seay, R. S., 235 Secondary cotton pests, most
important, 208 Selhime, A. G., 234
Serratia marcescens, a bacterium that some- times kills boll weevils, 133, 136
Seward, R. W., 385 Shapiro, M., 284, 288, 300 Sharpe, J. H., 378 Shaunak, K. K., 203 Shaver, T. N., 200, 201, 203,
230, 379 Shaw, D., 230 Shaw, F. R., 324 Shepard, H. A., 6, 26 Shepard, M., 107, 108, 124,
125, 311, 327 Shieh, T. R., 133, 139, 149 Shipp, 0. E., 356 Sikorowski, P. P., 160, 171,
176, 177, 265, 267, 272, 273, 277, 280, 283, 284, 285, 286, 287, 288, 290, 291, 292, 297, 298, 299, 300, 301
Simmons, L. A., 20, 171 Simpson, E. H., Ill, 481, 495 Simpson, G., 495 Simulium vittatum, black fly,
BAY SIR 8514 effective against, 210
Sirrenberg, W., 235 Skeith, R., 21 Slatten, B. H., 136, 149 Sloan, C. E., 173 Slosser, J. E., 375, 380,
527, 556 Smith, C. E., 356 Smith, D. B., 100, 146 Smith, G. L., 352, 358 Smith, J. W., 108, 125, 303,
307, 308, 311, 318, 320, 327
Smith, K. M., 131, 149 Smith, R. F., 22, 25, 26,
124, 387, 407 Smith, R. L., 230 Smith, W. R., 349, 356 Snow, J. W., 182, 204 Solenopsis invicta, red
imported fire ant, a potential predator of boll weevil, 104
Solomon, M. E., 112, 125 Sonnet, P. E., 232

588 INDEX
Sorghum bicolor, sorghum, crop model included within MOTHZV-2, 362, 373
Southwood, T. R. E., 304, 305, 307, 316, 328
Sowell, R. S., 478 Soybean:
cropland acreage in Mississippi, 485
soil resources, acreage, and per acre soybean yields, 487-488
Sparks, A. N., 181, 182, 201, 202, 204, 205, 297, 380
Spates, G. E., 177, 235 Sphaeralcea angustifolia,
possibly an occasional host for boll weevil in Texas, 544
Spider mites, Tetranychus spp., secondary pests of cotton, 6, 244, 255, 329, 332, 333, 339, 340
Spiders, 111 Spodoptera exigua, beet army-
worm, a cotton defoliator, 141, 209, 361, 534
-§-• frugiperda, fall army worm, is resistant to organophosphorus insecticides, 210, 246, 534
1' l^ttoralis, 211 Spodoptera spp., 105 Sprott, J. M., 526, 527, 541,
556 Staal, G. B., 209, 227, 228,
234 Stable flies, Stomoxys
calcitrans, 211 Stacey, A. L., 140, 149, 151,
557 Stadelbacher, E. A., 125, 149,
325, 327, 355 Stairs, G. R., 133, 150 Stanley, J. M., 201 Staphylococcus aureus, 286 Stapleton, H. N., 361, 380,
439, 479 Starbird, I. R., 8, 26, 32,
37, 45, 52
Starler, N. E., 26 State Extension and Crop
Reporting Service (North Carolina), 498
Staten, R. T., 150 State Pest Management
Advisory Committee (Mississippi), 389
Stehr, F. W., 109, 125 Steinhaus, E. A., 130,^131,
150, 283, 288, 300, 301 Sterilization, of boll
weevils, 153, 154- 157
Sterling, H. R., 406 Sterling, W. L., 12, 26, 105,
123, 125, 149, 160, 176, 232, 258, 304, 326, 327, 328, 434
Stern, V. M., 7, 26, 27, 126 Stewart, F. D., 288, 301 Still, G. G., 212, 232, 234 Stinner, R. E., 126, 314,
316, 328, 361, 379, 380, 478
Stipanovic, R. D., 85, 100 Stokes, J. B., 230 Stoner, A., 259, 434 Streptococcus spp., 286 Strickland, A. H., 304, 328 Suber, E. F., 348 Suguiyama, L. F., 497 Sulfur, was more effective
than calcium arsenate in control of late- season cotton insects, 6, 239
Surveys, of pesticide use and sales in the world and in the United States in the last decade, 30-50
Sutter, G. R., 288, 301 Sutton, R. A., 234 Systena spp., minor pests of
cotton, 63
Taft, H. M., 70, 175, 211, 218, 219, 232, 233, 234, 260, 353, 356, 542, 556
Tañada, Y., 135, 150

INDEX 589
Tarnished plant bug, Lygus líneolarís, see Arthropods
Taylor, C. R., 554 Tejada, L. 0., 110, 126 Telford, A. D., 126 Terry, P. H., 156, 170, 176 Tetranychus cinnabarinus,
diflubenzuron effective against, 211
Tetranychus spp., 6, 244, 255, 329, 332, 333, 339-340
Texas A&M University, 120, 373, 524, 548
Texas Agricultural Extension Service, development of BUGNET, a computer- ized pest management delivery system, 372-377
Texas Pest Management Association, 15
TH-6040, see Diflubenzuron Thespesia populnea, possible
host of boll weevil in south Florida, 544
Thespesia sp., 54 Thomas, B. R., 177 Thomas, F. L., 122 Thomas, G. M., 299 Thomas, J. G., 529, 540, 556 Thomisidae (crab spiders), 106 Thompson, A. C., Jr., 173,
205, 231, 286, 287, 300, 301
Thompson, T. E., 124 Thripidae, Thrips, 62, 64, 340 Aeolothrips, bandedwinged
thrips, 64 cotton with high gossypol
has increased susceptibility to thrips, 85
Tifton, K. W., 97 Tingle, F. C, 69, 202, 203,
249, 259, 260, 261, 434, 435, 554
Tinney, J. C., 26 Toba, H. H., 381 Tobacco budworm, Heliothis
virescens, see Heliothis spp.
Todd, J. W., 234, 345, 348 Toscano, N. C, 126
Total population management (TPM), 9-13
Townsend, C. H. T., 5, 26, 237, 261
Townsend, J. R., Jr., 369, 370, 372, 380
Toxaphene, 29, 48, 49, 67 extensive use of this
insecticide dimin- ished with the advent of the pyrethroid insecticides, 46
Trap crops, for suppression of boll weevils, 188, 191
Trialeurodes abutilonea, bandedwinged whitefly, 62, 64, 246
Trichogramma pretiosum, egg parasite of Heliothis spp., 216, 223
Trichogramma spp., 115, 120, 217
Trichoplusia ni, cabbage looper, 105, 141, 208, 329, 332, 339, 361
Tsao, C. H., 341, 342, 343, 348
Tsuchiya, T., 299 Tumlinson, J. H., 69, 180,
184, 199, 200, 201, 204, 205, 374, 381
Turnipseed, S. G., 216, 234 Turoff, M., 52
Ultraviolet radiation, of boll weevils, 131
University of California, Riverside, 120
U.S. Congress, 8, 26 U.S. Department of Agricul-
ture, 4, 12, 15, 27, 32, 36, 37, 44, 45, 52, 330, 331, 333, 334, 340, 348, 488, 496, 505, 513, 517, 523
U.S. Department of Agricul- ture Marketing service office areas, 78
Vail, P. v., 137, 142, 150 Van Daalen, J. J., 234

590 INDEX
van den Bosch, R., 8, 26, 27, 105, 108, 110, 113, 119, 122, 126, 320, 324, 325, 328, 378
Vanderzant, E. S., 160, 176, 266, 301
VanDuyn, J. W., 328 Van Steenwyk, R. A., 105, 113,
119, 126 Vardell, H., 296 Varley, G. C., 304, 328 Veal, S. D., 148 Veech, J. A., 215, 234 Verloop, A., 211, 214, 234 Verticillium dahliae, a prev-
alent fungal pathogen of cotton, in the Midsouth, 76, 77
Villavaso, E. J., 153, 157, 158, 159, 165, 166, 171, 176, 177
Vincent, W. R., 233 Vinson, S. B., 26, 229, 327 Vinzant, B. G., 124 Virelure, synthetic pheromone
of Heliothis vires- cens, 180-181
Viruses: chilo irridescent virus,
infectious to boll weevil, 138
Heliothis spp. nuclear polyhedrosis virus, is host specific to Heliothis spp,, 131
nuclear polyhedrosis virus, ACNPV, isolated from alfalfa looper, infects pink bollworm larvae, 137
Von Rumker, R., 500, 517
Waddle, B. A., 81, 86, 96, 98, 100
Wade, L. J., 233 Waiss, A. C., Jr., 85, 100 Wakabayashi, N., 232 Walker, A. B., 171 Walker, H. J., 356 Walker, J. K., Jr., 7, 12, 27,
54, 70, 86, 91, 95, 99, 100, 125, 357, 524, 525, 555, 556
Walker, R. L., 351
Wall, M. L., 406 Walter, J. K., 378 Walters, S., 200 Walton, G. S., 295 Wang, Y., 21, 378, 477 Ware, J. 0., 524, 556 Warren, L. 0., 314, 328 Waters, R. M., 232 Waters, W. E., 305, 328 Watkins, W. C., 324, 350, 351 Watson, F. L., 380, 479 Watson, T. F., 123, 145,
230, 357 Watt, K. E. F., 114, 126,
304, 305, 328 Watts, J. G., 357 Weaver, D. B., 357 Weaver, J. B., Jr., 83, 97,
100, 357 Weeks, J. R,, 200 Weiser, J., 135, 150 Wellinga, K., 209, 234 Wellso, S. G., 176 Westphal, D., 378 Whisler, F. D., 476, 478 Whitcomb, W. H., 64, 70,
105, 109, 110, 121, 126, 215, 235, 318, 320, 328
White, J. R., 225, 233, 235, 527, 556
White, W. H., 97 Whitefly, bandedwinged,
Trialeurodes abuti- lonea, 62, 64, 246
Whitten, C. J., 228, 235 Whitten, M. J., 168, 177 Wilkins, J. S., 349 Wilkinson, J. D., 163, 177,
215, 216, 235 Williams, B. R., 357 Williams, P. P., 295 Wilson, A. G., 361, 365, 381 Wilson, C. A., 121, 406 Wilson, F. D., 150 Wilson, L. T., 463, 479 Wilson, N. M., 203 Wilson, R. J., 6, 27 Wilson, R. L., 150 Wipprecht, R., 21, 352 Witz, J. A., 22, 122, 175,
200, 201, 202, 325, 359, 360, 379, 381, 406, 435, 477

INDEX 591
Wiygul, G., 27, 172, 174, 177, 298, 300
Wolf, W. W., 374, 381 Wolfenbarger, D. A., 86, 101,
147, 231, 354, 357, 553
Wood, R. H., 24, 232, 554 Woods, C. W., 170 Wright, J. E., 12, 27, 153,
156, 157, 158, 163, 165, 166, 170, 173, 174, 175, 176, 177, 211, 232, 235, 298, 299
Wyatt, J. M., 176, 297, 298, 300, 301
Xanthomonas malvacearum, the cause of bacterial blight of cotton, 77
Yearian, W. C, 12, 27, 139, 140, 146, 147, 148, 149, 151, 353, 532, 555, 557
Yendol, W. G., 149 Yield (cotton):
increase with new insecti- cides, 251
reduction, due to insects, 240-243, 329-345
Yokayama, V. Y., 108, 127 Young, D. F., Jr., 64, 70,
385, 406, 407 Young, J. H., 356 Young, M. T., 352, 357, 358 Young, S. Y., 27, 131, 139,
146, 147, 148, 149, 151, 353, 557
U)-9-tetradecenal (Z;-9-TDAL) and (Z^)-ll-hexadecenal U-11-HDAL) are two components of Heliothis virescens pheromone, 180
Zelus renardii, assassin bug, predator of pests in cotton, 114
Zoebelein, G., 210, 235























