
Costs and Benefits of Integrating Information Between the Cerebral Hemispheres: A Computational...
19
Neuropsychology 1998, Vol. 12, No, 3, 380-398 Copyright 1998 by the American Psychological Association, Inc. 0894-4105/98/S3.00 Costs and Benefits of Integrating Information Between the Cerebral Hemispheres: A Computational Perspective Aysenil Belger University of Illinois at Urbana-Champaign, Yale University School of Medicine, and Veterans Affairs Healthcare System, West Haven, Connecticut Marie T. Banich University of Illinois at Urbana-Champaign Because interaction of the cerebral hemispheres has been found to aid task performance under demanding conditions, the present study examined how this effect is moderated by computational complexity, the degree of lateralization for a task, and individual differences in asymmetric hemispheric activation (AHA). Computational complexity was manipulated across tasks either by increasing the number of inputs to be processed or by increasing the number of steps to a decision. Comparison of within- and across-hemisphere trials indicated that the size of the between-hemisphere advantage increased as a function of task complexity, except for a highly lateralized rhyme decision task that can only be performed by the left hemisphere. Measures of individual differences in AHA revealed that when task demands and an individual's AHA both load on the same hemisphere, the ability to divide the processing between the hemispheres is limited. Thus, interhemispheric division of processing improves performance at higher levels of computational complexity only when the required operations can be divided between the hemispheres. Despite the abundance of research on functional asymme- tries in the human cerebral hemispheres (e.g., Kimura, 1961, 1969; Nebes, 1978; Cohen, 1973) and their complementary modes of processing (e.g., Levy-Agresti & Sperry, 1968; Sergent, 1982), relatively little is known about their interac- tions and the effects of interhemispheric information process- ing on cognitive performance. Although the two hemi- spheres are functionally specialized, they are also strongly interconnected by over 200 million nerve fibers, which enables them to integrate information and produces strong bihemispheric activation during most tasks, including "later- alized" ones such as speaking (e.g., Lassen, Ingvar, & Skinhoj, 1978). In recent years, a significant effort has been dedicated to understanding the role of interhemispheric interaction on cognition. To clearly understand how the independent and specialized processing capacities of the hemispheres affect information processing, we must also Aysenil Belger, Department of Psychology and the Beckman Institute, University of Illinois at Urbana-Champaign; Department of Psychiatry, Yale University School of Medicine; and Cognitive Neuroscience Laboratory, Veterans Affairs Healthcare System, West Haven, Connecticut. Marie T. Banich, Department of Psychol- ogy and the Beckman Institute, University of Illinois at Urbana- Champaign. Preparation of this article was supported by a University of Illinois Graduate College Dissertation Research Grant and an American Psychological Association Dissertation Research Award. The research was conducted as partial fulfillment of the require- ments for a doctoral degree on the part of Aysenil Belger. We are grateful to Brian Meyer, Brigitte Hegemeier, Pam Becker, and Kathy Holler for assistance in data collection. Correspondence concerning this article should be addressed to Aysenil Belger, Psychiatry Service (116A), Veterans Affairs Health- care System, 950 Campbell Avenue, West Haven, Connecticut 06516. Electronic mail may be sent to [email protected]. understand how the hemispheres communicate, the costs and benefits of their interaction, and how the factors modulate the relative balance between these two outcomes. Hence, we must elucidate the implications of interhemi- spheric information processing for task performance. This study investigated the dynamics of interhemispheric integration of information and its effects on performance. We demonstrate that interhemispheric interaction increases the brain's information-processing capability during complex tasks by allowing the distribution of computations between the hemispheres. We further present evidence that the advan- tages of interhemispheric interaction can be constrained by effects of individual differences in asymmetric hemispheric activation and, in some very restricted cases, by the lateral- ization of the computations required to perform a task. From Hemispheric Autonomy to Interhemispheric Networks Early studies investigating neurologically intact individu- als and split-brain patients alike demonstrated that the hemispheres not only have their preferred mode of operation (left-verbal, right-spatial) but also rely upon independent resources (Brooks, 1968; Kinsbourne & Hicks, 1978, e.g., Kreuter, Kinsbourne, & Trevarthen, 1972) to enable them to perform certain operations simultaneously with relatively little interference. For example, simultaneous performance of two verbal tasks yields greater dual-task interference than performance of a verbal and a visuospatial task, which would rely upon distinct left- and right-hemispheric re- sources, respectively (Wickens, 1980; Wickens & Liu, 1988; Herdman & Friedman, 1985; Friedman & Poison, 1981; Poison & Friedman, 1988). This independence of resources raises the issue of how processing is coordinated and 380
Transcript of Costs and Benefits of Integrating Information Between the Cerebral Hemispheres: A Computational...

Neuropsychology1998, Vol. 12, No, 3, 380-398
Copyright 1998 by the American Psychological Association, Inc.0894-4105/98/S3.00
Costs and Benefits of Integrating Information Between the CerebralHemispheres: A Computational Perspective
Aysenil BelgerUniversity of Illinois at Urbana-Champaign,
Yale University School of Medicine, and Veterans AffairsHealthcare System, West Haven, Connecticut
Marie T. BanichUniversity of Illinois at Urbana-Champaign
Because interaction of the cerebral hemispheres has been found to aid task performance under
demanding conditions, the present study examined how this effect is moderated by
computational complexity, the degree of lateralization for a task, and individual differences in
asymmetric hemispheric activation (AHA). Computational complexity was manipulated
across tasks either by increasing the number of inputs to be processed or by increasing the
number of steps to a decision. Comparison of within- and across-hemisphere trials indicated
that the size of the between-hemisphere advantage increased as a function of task complexity,
except for a highly lateralized rhyme decision task that can only be performed by the left
hemisphere. Measures of individual differences in AHA revealed that when task demands and
an individual's AHA both load on the same hemisphere, the ability to divide the processing
between the hemispheres is limited. Thus, interhemispheric division of processing improves
performance at higher levels of computational complexity only when the required operations
can be divided between the hemispheres.
Despite the abundance of research on functional asymme-tries in the human cerebral hemispheres (e.g., Kimura, 1961,1969; Nebes, 1978; Cohen, 1973) and their complementarymodes of processing (e.g., Levy-Agresti & Sperry, 1968;Sergent, 1982), relatively little is known about their interac-tions and the effects of interhemispheric information process-ing on cognitive performance. Although the two hemi-spheres are functionally specialized, they are also stronglyinterconnected by over 200 million nerve fibers, whichenables them to integrate information and produces strongbihemispheric activation during most tasks, including "later-alized" ones such as speaking (e.g., Lassen, Ingvar, &Skinhoj, 1978). In recent years, a significant effort has beendedicated to understanding the role of interhemisphericinteraction on cognition. To clearly understand how theindependent and specialized processing capacities of thehemispheres affect information processing, we must also
Aysenil Belger, Department of Psychology and the Beckman
Institute, University of Illinois at Urbana-Champaign; Department
of Psychiatry, Yale University School of Medicine; and Cognitive
Neuroscience Laboratory, Veterans Affairs Healthcare System,
West Haven, Connecticut. Marie T. Banich, Department of Psychol-
ogy and the Beckman Institute, University of Illinois at Urbana-Champaign.
Preparation of this article was supported by a University of
Illinois Graduate College Dissertation Research Grant and an
American Psychological Association Dissertation Research Award.The research was conducted as partial fulfillment of the require-
ments for a doctoral degree on the part of Aysenil Belger. We are
grateful to Brian Meyer, Brigitte Hegemeier, Pam Becker, andKathy Holler for assistance in data collection.
Correspondence concerning this article should be addressed toAysenil Belger, Psychiatry Service (116A), Veterans Affairs Health-
care System, 950 Campbell Avenue, West Haven, Connecticut
06516. Electronic mail may be sent to [email protected].
understand how the hemispheres communicate, the costsand benefits of their interaction, and how the factorsmodulate the relative balance between these two outcomes.Hence, we must elucidate the implications of interhemi-spheric information processing for task performance.
This study investigated the dynamics of interhemisphericintegration of information and its effects on performance.We demonstrate that interhemispheric interaction increasesthe brain's information-processing capability during complextasks by allowing the distribution of computations betweenthe hemispheres. We further present evidence that the advan-tages of interhemispheric interaction can be constrained byeffects of individual differences in asymmetric hemisphericactivation and, in some very restricted cases, by the lateral-ization of the computations required to perform a task.
From Hemispheric Autonomy
to Interhemispheric Networks
Early studies investigating neurologically intact individu-als and split-brain patients alike demonstrated that thehemispheres not only have their preferred mode of operation(left-verbal, right-spatial) but also rely upon independentresources (Brooks, 1968; Kinsbourne & Hicks, 1978, e.g.,Kreuter, Kinsbourne, & Trevarthen, 1972) to enable them toperform certain operations simultaneously with relativelylittle interference. For example, simultaneous performanceof two verbal tasks yields greater dual-task interference thanperformance of a verbal and a visuospatial task, whichwould rely upon distinct left- and right-hemispheric re-sources, respectively (Wickens, 1980; Wickens & Liu, 1988;Herdman & Friedman, 1985; Friedman & Poison, 1981;Poison & Friedman, 1988). This independence of resourcesraises the issue of how processing is coordinated and
380

TASK COMPLEXITY AND INTERHEMISPHER1C INTERACTION 381
integrated across the hemispheres. Although having special-
ized processors enables various aspects of tasks to be
distributed and performed in parallel, there is also a costassociated with translating and integrating outputs of differ-
ent modules. Such costs associated with parallel informationprocessing in the brain have been described by Nelson andBowers (1990) as they apply to parallel computer maps. One
major obstacle to achieving optimal performance is commu-
nication overhead, which is the cost of integrating informa-
tion between a number of processing units. Such integration
is time consuming, and the characteristics of the connectionsmay degrade the resolution of the information to be commu-
nicated. Empirical evidence suggests that a certain amount
of information loss and degradation is also found duringinterhemispheric information processing (e.g., Banich &
Belger, 1991). Hence, interhemispheric processing mayeither improve or hinder performance depending on therelative benefit afforded by parallel computing across the
hemispheres as compared to the relative cost of the commu-nication overhead involved in coordinating the outputs of
the two hemispheres.Recent studies that addressed this networking issue by
examining the effects of interhemispheric interaction on per-
formance have yielded conflicting results; some showed thatdividing information between the hemispheres improvesperformance and others reported that interhemispheric pro-cessing hinders performance. Comparison of split-brainpatients to neurologically intact individuals provides a directevaluation of the relative costs or benefits of interhemi-spheric communication on cognitive performance, becausethese patients lack all interhemispheric communicationfibers. However, only some studies found that split-brainpatients perform better than neurologically intact individuals(Ellenberg & Sperry, 1979, 1980; Holtzman & Gazzaniga,1985), suggesting that (a) the two isolated hemispheres areindependent information processors and do not interferewith each other's performance, and (b) disconnection of thehemispheres increases overall processing capacity. Otherstudies, in contrast, found that neurologically intact individu-als perform better than split-brain patients (e.g., Kreuter etal., 1972; Holtzman & Gazzaniga, 1982), a finding that isattributed to the capability of neurologically intact individu-als to coordinate and to distribute information between thetwo hemispheres via the callosum.
As in studies of split-brain patients, studies of neurologi-cally intact individuals that examined the relative benefits ofinterhemispheric processing with tachistoscopic methodsalso yielded conflicting results. Although some studiesfound superior performance on across-hemisphere condi-tions (where the information is divided between the visualfields and hence projects to opposite hemispheres; Banich &Belger, 1990; Berger & Landolt, 1990; Davis & Schmit,1971, 1973; Dimond, 1971; Dimond & Beaumont, 1971,1972a, 1972b; Liederman, Merola, & Hoffman, 1986;Merola & Liederman, 1985, 1990; Miller, 1981, 1983;Norman, Jeeves, Milne, & Ludwig, 1992; Quinn & Geffen,1986; Sereno & Kosslyn, 1991), other studies found superiorperformance on within-hemisphere trials (where stimuli aredisplayed within one visual field and hence project to the
same hemisphere; e.g., Banich & Belger, 1990; Beaumont &Dimond, 1973, 1975; Dimond, 1969; Bradshaw, Nettleton,
& Patterson, 1973). The discrepancy in results across thesestudies suggests that the effects of dividing the processingbetween the hemispheres is not consistent but rather variesas a function of other factors.
Interhemispheric interaction appears to be a dynamicprocess that can support parallel and independent informa-tion processing in specialized modules through isolation butthat can also facilitate interaction and integration of the
modules' outputs to yield a single unified response. The aimof this study was to isolate some factors that affect theoutcome of this dynamic system by investigating the advan-tages of interhemispheric interaction and the factors thatmoderate this advantage.
Advantages and Limitations of Interhemispheric
Interaction: Costs and Benefits of Distributed
Processing During Complex Tasks
One factor that has been reported to increase the advan-tage of dividing processing between the hemispheres isinformation load. Banich (1985) observed that when all theinformation necessary for a decision is presented simulta-neously (0 ms interstimulus interval [ISI]), an across-hemispheres advantage emerges (Banich, 1985; Davis &Schmit, 1971, 1973; Liederman et al., 1986). On the otherhand, imposing an ISI between the presentation of the itemsyields a within-hemisphere advantage. Banicb (1985) con-ceptualized the presentation of information sequentially as adivision of the load over time, such that less informationneeded to be processed at once. Hence, she suggested thatdividing the information between the hemispheres enables adistribution of the load and increases parallel processing,which in turn improves overall performance.
In the Banich (1985) study, task load was manipulated byvarying ISI, but it can also be varied by manipulatingcomputational complexity, which can be conceptualized asthe number and sorts of transformations, operations, orcomputations that must be performed on an input before adecision can be reached. For example, if one is simply askedto decide if two numbers are physically identical (physical-identity task), the numerals must be perceptually encodedand then a comparison of their form must occur. However, ifone is asked to decide if one number is the square of theother, the items must not only be perceptually analyzed butthey must be identified and then one must be squared. Thus,the number-squaring task is computationally more complexthan the number-identity task as it requires not only theperceptual analysis operation but the squaring operation aswell. In a series of recent studies (Banich & Belger, 1990;Belger & Banich, 1992), we reported an across-hemisphereadvantage for relatively complex tasks with multiple opera-tions (ordinal and summation tasks) but with a within-hemisphere advantage for computationally simpler tasks(physical-identity task). Furthermore, by increasing compu-tational complexity at different stages of information process-ing, we found similar effects on the relative advantages ofinterhemispheric processing. Thus, a task in which the

382 BELGER AND BANICH
individual determines whether two of three numbers arephysically identical yields a within-hemisphere advantage, atask in which the individual determines whether two of fivenumbers are physically identical yields an across-hemi-sphere advantage, and a task in which the individual determineswhether two of five numbers have the same name yields aneven greater across-hemisphere advantage. Hence, both
increasing complexity at the perceptual level by increasingthe number of items to be processed (or the memory load ofthe task) and increasing complexity at the decision level by
requiring additional stages of processing (e.g., naming ascompared to physical identification) increases the advantageafforded by interhemispheric processing.
In this article, we propose that the enhanced performanceon across-hemisphere trials during the complex tasks in ourprior studies can be explained by the ability to distribute thecomputations over the two hemispheres and to performoperations in a parallel fashion. In support of this explana-tion, prior studies have provided evidence that interhemi-spheric interaction facilitates parallel processing of informa-tion between the hemispheres during performance of twoconflicting tasks (Merola & Liederman, 1990) or perceptu-ally interfering tasks (Liederman et al., 1986; Merola &
Liederman, 1985). We further propose that the advantages ofnetworking between the hemispheres are constrained by twofactors: (a) the lateralization of the computations required by
the task, and (b) the individual differences in hemisphericactivation and handedness.
Effects of Task Laterality on the Sizeof the Across-Hemisphere Advantage
We propose that the resource characteristics of eachhemisphere may in some cases limit the advantage thatinterhemispheric interaction can provide during the perfor-mance of complex tasks. The degree to which portions of atask can rely on each hemisphere's processing resourcesdetermines the extent to which distributing task computa-tions between the hemispheres is beneficial to performance.Most cognitive operations can be supported by both hemi-spheres, and hence during the performance of most complextasks the load can be divided between the hemispheres. Thishas been the case for all of the component operations of thetasks used in our previous studies. In contrast, when theprocessing resources of one hemisphere are critical formaking the final decision, then no matter how the input isdistributed between the hemispheres, the final outcome willhave to await the availability of this single processor.Consequently, if a significant portion of a computationallycomplex task can only be performed by one hemisphere,then no performance gain would be predicted for across-hemisphere trials relative to within-hemisphere trials.
One operation that was demonstrated to be very stronglylateralized in right-handers is grapheme-to-phoneme conver-sion (Rayman & Zaidel, 1991), which is a critical compo-nent of rhyming. The strong dependence of rhyming on theleft hemisphere (as indicated by split-brain patients' inabilityto perform this operation when stimuli are displayed in theleft visual field, projecting exclusively to the right hemi-
sphere; Zaidel & Peters, 1981) creates a situation similar tothe load imbalance problem described by Nelson and
Bowers (1990), which is considered to be one major sourceof inefficiency to parallel computing. Load imbalance resultsfrom a nonuniform distribution of the processing load, suchthat certain processors are overloaded as compared to others.
Thus, although computations in some processors are com-pleted quite rapidly, such as perceptual identification, the
final outcome has to be delayed until the slowest processorcompletes its computation, in this case the grapheme-to-phoneme conversion operation. Hence, in such a situation, itshould not much matter whether the critical information for
the task is divided across the hemisphere or just projected toone, since ultimate performance of the task is dependentupon the left hemisphere.
In contrast, the computations required by the name-identity and physical-identity tasks used in our prior studies
did not depend critically on the processing capabilities ofone hemisphere. Evidence suggests that the name-identitytask can be performed either by the left hemisphere usingphonemic codes or by the right hemisphere via a casetransformation (Boles & Eveland, 1983). This enabled thecomputations to be distributed and processed in parallelacross the hemispheres. Hence, we suggest that the absenceof a load imbalance in the tasks we have used in prior studiesis responsible for the across-hemisphere advantage we haveobserved. In contrast, for a rhyming task, although bothhemispheres can do the initial perceptual encoding of theinformation, the final decision has to await the left hemi-sphere's completion of the grapheme-to-phoneme conver-sion, creating a load imbalance. Despite the complexity ofthe rhyme task, we predict that the inability to divideprocessing at the decision level will obstruct the emergenceof an across-hemisphere advantage on the rhyme task. Thisstudy investigates this prediction by comparing performanceon within- and between-hemisphere trials on a tachisto-scopic rhyme task and two tasks previously shown to yieldacross-hemisphere advantages.
Effects of Individual Differences in HemisphericLaterality on InterhemisphericInformation Processing
Although numerous studies have demonstrated that indi-vidual differences in age, sex, and handedness modulatelaterality effects (e.g., Lake & Bryden, 1976), relatively littleis known about how these individual differences in brainorganization might affect interhemispheric processing. Wepropose that individual differences will also affect the abilityto divide the computations between the hemispheres as aconsequence of then" direct influence on lateralized process-ing. More specifically, in this experiment, we concentratedon two factors, asymmetric hemispheric activation (AHA)and handedness, and examined how they can affect interhemi-spheric information processing.
AHA. AHA, which is also referred to as hemispheric
arousal asymmetry or hemispheric reliance in the literature,corresponds to the tendency of an individual to systemati-cally engage one hemisphere relative to the other during task

TASK COMPLEXITY AND INTERHEMISPHERIC INTERACTION 383
performance, regardless of task demands (Levy, Heller,
Banich, & Burton, 1983a). Among right-handers, the degree
of AHA is closely related to the perceptual asymmetry on
both verbal and nonverbal tasks. For example, it has been
found that individuals biased toward right-hemisphere pro-
cessing exhibit a larger than average left visual field (LVF)
advantage on a right-hemisphere task, a LVF advantage on a
nonlateralized task, and a smaller than average right visual
field (RVF) advantage on a left-hemisphere task. Thus, for
all tasks, the individual exhibits greater than average involve-
ment of the right hemisphere. Likewise, individuals biased
toward left-hemisphere processing exhibit a larger than
average RVF advantage on a right-hemisphere task, a RVF
advantage on a nonlateralized task, and a smaller than
average LVF advantage on a left-hemisphere task (Kim &
Levine, 1991a, 1991b; Levine, Banich, & Kim, 1987;
Levine, Banich, & Koch-Weser, 1984; Levine, Banich, &
Koch-Weser, 1988; Levy et al., 1983a). This AHA results in
a positive correlation across an individual's perceptual
asymmetries on a series of lateralized tasks. Furthermore,
45.2% of between-subjects variations in asymmetry scores
has been reported to be attributable to individual differences
in AHA (Kim, Levine, & Kertesz, 1990). Although right-
and left-handers may vary in their patterns of hemispheric
asymmetry, the distribution of characteristic perceptual
asymmetries does not vary with handedness, such that AHA
accounts for the same amount of variance for dextrals and
sinistrals (Kim et al., 1990). Finally, AHA may be partly due
to baseline arousal asymmetries between the hemispheres
(e.g., Levy et al., 1983a; Green, Morris, Epstein, West, &
Engler, 1992).Although AHA is manifest on each and every lateralized
task, one method often used to classify individuals is the
Chimeric Face Test (Levy, Heller, Banich, & Burton,
1983b). This test has a high test-retest and split-half
reliability, has been validated on a sample ranging in age
from 5 to the late 70s (Levine & Levy, 1986), and has been
well normed on a sample of 111 right-handers and 111
left-handers (and more recently on a sample of over a
thousand by Heller, 1996). Furthermore, because this task
requires only a preference judgment, it avoids issues of
variations in level of performance across individuals. Finally,
the Chimeric Face Test has been found to predict perceptual
asymmetries on various tachistoscopic tasks (Levy et al.,
1983a; Levine et al., 1984, 1988), levels of performance on
nontachistoscopic tasks that are thought to engage process-
ing of one hemisphere more than the other (e.g., verbal
comprehension; Banich, Elledge, & Stolar, 1992), and asym-
metric aspects of emotional processing (Heller, Etienne, &
Miller, 1995).
Both the degree and the direction of AHA might affect the
efficiency of interhemispheric information processing. Indi-
viduals who exhibit a strong degree of AHA (regardless of
its direction) may be less efficient at integrating or distribut-
ing information between the hemispheres, because they may
have a tendency to let one hemisphere take over. In other
words, the strength of AHA could constrain performance to a
particular hemisphere and could create a load imbalance by
constraining processing to one hemisphere.
The direction of AHA could also modulate the advantages
of interhemispheric information processing by creating a
load imbalance. If an individual's bias is to activate the same
hemisphere that is recruited heavily by the critical opera-
tions of a particular task, the result will be a large load on a
single hemisphere. Because processing is so heavily depen-
dent on that hemisphere, the advantages afforded by inter-
hemispheric processing may be decreased. For example,
when a person performs a left-hemisphere task, an individual
bias toward left-hemisphere activation should restrict or re-
duce interhemispheric interaction, yielding no performance
advantage on across-hemisphere as opposed to within-
hemisphere trials. On the other hand, when a person performs
a right-hemisphere task, an individual bias toward left-
hemisphere activation would facilitate a division of process-
ing between the hemispheres to yield a greater across-
hemisphere advantage. Accordingly, individuals with greater
right-hemisphere activation should not benefit from inter-
hemispheric interaction when performing the physical- and
name-identity tasks that yield a right-hemisphere advantage,
because both their individual AHA and the demands imposed
by the task are biasing processing to the right hemisphere. In
contrast, individuals with greater left-hemisphere activation
should exhibit greater across-hemisphere advantages on the
physical- and name-identity tasks, because their individual
AHA complements the processing demands placed on the
right hemisphere by the task. In this article, we examined the
effects of the degree and direction of AHA on the relative
size of the across-hemisphere advantage individuals display
for the physical- and name-identity tasks, which yield
right-hemisphere advantages, and for the rhyme task, which
yields a left-hemisphere advantage.
Handedness. Evidence from a variety of clinical and
neuropsychological sources indicates that left-handers ex-
hibit less functional lateralization man right-handers (e.g.,
Bryden, 1965; Goodglass & Quadfasel, 1954; Rasmussen &
Milner, 1977; Satz, 1980). The greater functional equipoten-
tiality of the hemispheres of left-handers provides for the
possibility that patterns of interhemispheric interactions
might vary between left- and right-handers. However,
previous studies have not found any differences in interhemi-
spheric interaction (e.g., Banich, Goering, Stolar, & Belger,
1990; Piccirilli, Finali, & Sciarma, 1989; Beaumont &
Dimond, 1973, 1975; Dimond & Beaumont, 1972a), but
they utilized less lateralized tasks than we used in this study.
Hence, we examined whether handedness affects interhemi-
spheric interaction.
Therefore, the following three central issues were investi-
gated in this study: (a) We investigated the effects of compu-
tational complexity on the degree of the across-hemisphere
advantage by comparing performance on within- and across-
hemisphere trials during tasks of varying complexity, namely
a three-letter physical-identity task, a five-letter physical-
identity task, a three-letter name-identity task, and a five-
letter name-identity task. Accordingly, contrasting results
for the three- versus five-item displays enabled us to
examine the effects of increasing complexity at the percep-
tual level, comparison level, or both, whereas contrasting
physical- and name-identity tasks enabled us to examine the

384 BELGER AND BANICH
effects of varying complexity at the decision level, (b) We
investigated how interhemispheric interaction is affected by
the degree of lateralization of a task by comparing perfor-
mance on within- and across-hemisphere trials for the
weakly lateralized physical- and name-identity tasks and the
strongly left-hemisphere lateralized rhyme task. The rhyme
task is as computationally complex as the weakly lateralized
five-letter physical-identity task and the name-identity tasks,
with regard to perceptual requirements, the nature of the
decision process required, or both, but it differs in that it can
only be performed by the left hemisphere. Hence, we predict
that although the five-letter physical-identity task and the
name-identity tasks will yield an across-hemisphere advan-
tage, the rhyme task will not. (c) We investigated how
individual differences in AHA and handedness affect the
ability to distribute processing between the hemispheres
during computationally complex tasks. To do so, we corre-
lated the size of individuals' across-hemisphere advantages
on our various interhemispheric tasks with their degree of
AHA (as measured by the score on the Chimeric Face Test)
and investigated how across-hemisphere processing is af-
fected by handedness. We predicted that AHA limits the
advantages of interhemispheric processing for individuals
whose AHA is consistent with task demands. We also
predicted that to the degree that left-handedness moderates
the sole reliance on the left hemisphere for phonetic
processing, we should observe an increase on across-
hemisphere processing on this task relative to right-handers.
Method
Participants
Twenty-five right-handed (10 female and 15 male) and 28left-handed (13 female and 15 male) individuals participated in thisexperiment. Of the left-handers, 17 used a noninverted handposture for writing, while the other 11 utilized an inverted handposture (Levy & Reid, 1978). Handedness was assessed through aquestionnaire that determined lateral preference for items such ashammering or throwing a ball. Individuals were classified asright-handed if they performed seven of me eight items with theirright hand and wrote with their right hand and were classified asleft-handed if they performed six of the eight items with their lefthand and wrote with their left hand. Individuals were classified asinverters if they held their hand above the line of writing and heldthe point of the pencil pointed toward the bottom of the page. Allthe participants were University of Illinois students, who rangedbetween 18 and 27 years of age and had normal or corrected-to-normal visual acuity. None of the individuals had previouslyperformed any of the tasks.
Apparatus and Stimuli
Stimuli were presented on a medium-resolution Hewlett-Packard color graphics terminal, and trial presentations were underthe control of a Hewlett-Packard 310 computer, which alsorecorded reaction time.
Physical- and name-identity tasks. Four distinct weakly lateral-ized tasks were used in this study: a three-letter physical-identitytask, a five-letter physical-identity task, a three-letter name-identity
task, and a five-letter name-identity task. The stimuli for the
three-letter tasks consisted of a triangular array of letters with adigit at the center whose report was used to ensure central fixation.
The two top probe letters were presented 2.8° lateral from midlineand 1.4° above fixation. The bottom target letter appeared 1.4°
below the fixation point and 1.4° lateral from midline. Theseeccentricities should allow for initial receipt by each hemisphere,because most studies report little or no overlap in nasotemporalconnections (e.g., Haun, 1978) and any midline fusing mainlyserves the purpose of stereopsis (e.g., Harvey, 1978). Each letter
subtended a maximum of 0.85° horizontally and 1.17° vertically.The letters A, B, M, P, T, R, F, G, H, and Q were used as stimuli,because they appear least confusable on the computer screen that
we used. It should be noted that the three-letter name-identity taskas well as the three-letter physical-identity task used in thisexperiment are identical to those used by Banich and Belger(1990). The stimuli for the five-item tasks were identical exceptthat four probe letters appeared above the fixation point, two ineach visual field positioned one above the other, such that thecenter of each pair matched the location of the single letters in thethree-letter tasks. The bottom letter of each pair appeared 0.87°above and 2.8° lateral from the fixation point, while the top letter of
each pair appeared 1.9° above and 2.8° lateral from the fixationpoint. The sizes of the letters were identical to those in thethree-letter tasks, and the two letters were separated by 0.12°. Thevertical orientation of the two pairs of top probe letters was chosenover a horizontal orientation to help minimize the use of apostperceptual scanning habit induced by reading. For half thematch trials, the matching item was the lower of the top two letters,and for half the trials it was the upper of the two top letters. Thefive-letter name-identity arrays were similar to those used byBelger and Banich (1992). In the physical-identity tasks, all theletters were uppercase, whereas in the name-identity tasks, thebottom target letter was lowercase (see Figure 1).
For all four tasks, half of the 224 trials consisted of match trialsin which the bottom item matched one of the probe letters, and theother half were mismatch trials in which the bottom item did notmatch any of the probe letters. For half of the match trials (56trials), the bottom item matched the letter that appeared in the samevisual field, that is, on the same side of the fixation point.Henceforth, these will be referred to as within-field trials. For theother half of the match trials (56 trials), the bottom item appearedcontralateral to the letter it matched. Henceforth, these will bereferred to as across-field trials: For both within- and across-fieldtrials, the bottom item appeared in the LVF for half the trials and inthe RVF for the odier half. Thus, there were four different matchtrial types: within field with the bottom item in the RVF (withinRVF), within field with the bottom item in the LVF (within LVF),across field with the bottom item in the RVF (across b-RVF), andacross field with the bottom item in the LVF (across b-LVF).Furthermore, the mismatch trials were designed such that for halfof them the bottom item appeared in the RVF and for the other halfit appeared in the LVF.
Rhyme task. Whereas the four physical- and name-identitytasks can be performed by either hemisphere and hence areconsidered to be weakly lateralized, the grapheme-to-phonemeconversion required by the rhyme task can only be performed bythe left hemisphere and hence is strongly lateralized. The stimulipresented during the rhyme task consisted of triangular arrays ofletters with a digit at the center. The two top letters appeared 1.4°above and 2.8° lateral on either side of the center digit. The bottomthree letters formed a word (e.g., SEA) and were displayedvertically 1.4° lateral to the right or left of the central fixation (refer

TASK COMPLEXITY AND INTERHEMISPHERIC INTERACTION 385
MATCH TRIALS
Within LVF3-ITEM PHYSICAL IDENTITY TASKWithin RVF Across b-LVF Across b-RVF
A G2
A
Within LVF
Within LVF
5-tTEM PHYSICAL IDENTITY TASKWithin RVF Across b-LVF
3-ITEM NAME IDENTITY TASKWithin RVF Across b-LVF
A G
6A
Across b-RVF
AR
2A
GC
AR
GC
2C
AR
3C
GC
AR
GC
6A
Across b-RVF
A G2
Within LVF
Within LVF
S-ITEM NAME IDENTITY TASKWithin RVF Across b-LVF
RHYME TASKWithin RVF Across b-LVF
A G6
a
Across b-RVF
AR
2a
GC
AR
GC
2c
AR
3c
GC
AR
GC
6a
Across b-RVF
G A2
SEA
A G2
TEE
A G3
SEE
G A6
PEA
AR
SAMPLE MISMATCH TRIALSLVF RVF
GC
GR
GC
H H
Figure 1. Sample match trials for the 5 tasks used in this study: the 3-item physical-identity task,
the 5-item physical-identity task, the 3-item name-identity task, the 5-item name-identity task, and
the rhyme task. Some sample mismatch trials are also shown. Within LVF = within field with the
bottom item in the left visual field; within RVF = within field with the bottom item in the right visual
field; across b-LVF = across field with the bottom item in the left visual field; across b-RVF = across
field with ttie bottom item in the right visual field.
to :%ute 1), The top iattar ,of <he word .appeared 0.87° belowfixation, the «cond was located 2M" below fixation, and the third
wastocaadS-3° below fixation. -The letters spanned 0.78" horizon-
MSy and 1.17° vettie«%. !Ewo Mocks .of 28 .practice trials were
followed by four blocks of 56 itest toials. Each block was composedrf'half rhyme and half nonrbyming trials. Rhyme trials are those in•wSrich She -vertically presented word rhymes with either one of the
:two letters {e.g., P and TEE., or J and DAY). The rhyming trials
.consisted equally of RVF and LVF trials and within- and across-hemisphere trials. The resulting trial types correspond to those used
in the physical- and name-identity tasks. In the within-LVF trials,
the bottom word appeared in the LVF and rhymed with the letter
that appeared in the LVF above fixation; in the within-RVF trials,
the word in the RVF rhymed with me letter in the RVF abovefixation. For the across-b-LVF trials, the word presented in the LVF
rhymed with the letter in the RVF. Finally, for the across-b-RVF
trials, the word presented in the RVF rhymed with the letter in the
LVF (see Figure 1). All trials included a digit at the center whose
report was used to ensure central fixation. Stimuli were carefully
created such that the central digit did not rhyme with any of the

386 BELGER AND BANICH
letters or the word, and the top two letters were selected such thatthey would not rhyme with each other.
AHA: The Chimeric Face Test. Aside from the handedness test,a test of AHA, the Chimeric Face Test was also administered toeach participant (Levy et al., 1983b). This test consists of a 36-pagebooklet, with one test item per page. A test item consists of a pair ofchimeric faces, one at the top of the page and one at the bottom.Each face is composed of a smiling half and a neutral half, and thetwo faces appearing on each page are minor images of each other.
Procedure
Participants were tested over five sessions. One of the fivetachistoscopic tasks was administered in each session. Randomiza-tion of task presentation order was accomplished by counterbalanc-ing the presentation order of the two groups of tasks (the verbal taskand the nonverbal tasks) and then counterbalancing the order ofpresentation of the nonverbal tasks for each individual. Thus, halfthe individuals performed the verbal task (rhyme) first, and theother half performed the nonverbal tasks (physical and name iden-tity) first. The presentation order of the nonverbal tasks was coun-terbalanced across participants. Ten task orders were generated,and participants were randomly assigned to one of these orders.
All participants were given the handedness questionnaire and theChimeric Face Test in their first session, along with a vision test toensure that they had normal or corrected-to-normal vision. Bygiving these tasks in the first session, we were able to discontinuetesting of individuals who did not satisfy our criteria. Finally,participants were paid hourly for their participation.
The procedures of administration for each task were as follows:Physical-identity and name-identity tasks. During each ses-
sion, an individual's head was stabilized on achin rest, such that theeyes were located 32.9 cm from a graphics display screen. At thestart of each trial, a central fixation point appeared on the screen. Asingle button mounted on a grip, aligned with the vertical midlineof the screen, was located in front of the participant. Reaction time(RT) was the dependent measure. During the three-letter andfive-letter physical-identity tasks, individuals were required topress the button if the bottom letter matched any of the top probeletters (which never matched) and to refrain from pressing thebutton if it did not (go/no-go RT). For the name-identity tasks,individuals had to press the button if the bottom item had the samename as any of the top probe items and to refrain from pressing thebutton if it did not (go/no-go RT). All displays were presented for200 ms.
As a control over central fixation, participants were required toreport the center digit for each trial, and if they could not do so, thetrial was discarded. Stimulus duration was set at 200 ms to precludelateral eye movements and to ensure that the information presentedin each visual field was initially directed exclusively to the con-tralateral hemisphere. As an additional control over fixation, at thebeginning of a test session, the edges of each individual's left andright blindspots were located, and a peripheral dot was placed 0.5°from each edge. Thus, if an individual's eyes deviated more than0.5° from central fixation, the peripheral dots would be seen(Banich & Belger, 1990). Any trial in which the individual reportedseeing either of the peripheral dots was excluded fromsubsequent analysis.
Within each session, participants were first given two blocks of28 practice trials, followed by 224 test trials administered in four56-trial blocks. Within each block, half of the trials were match andhalf were nonmatch. Participants responded with the left hand fortwo blocks and with the right hand for two blocks. The order of theresponse hand was counterbalanced across individuals. Data from
trials were included in subsequent analyses only if (a) the correctbutton press was made, (b) the central item was reported correctly,and (c) the peripheral dots were not seen.
Rhyme task. During the rhyme test, participants were presentedwith two practice blocks of 28 trials, followed by four test blocks of56 trials. The stimuli appeared for 200 ms. On each trial,participants were instructed to press a button if the verticallyaligned word rhymed with either of the two top letters and to refrainfrom pressing the button if it did not (go/no-go RT). After makingthe rhyme decision, the participants were instructed to report thecenter fixation digit. For this task as well as the physical- andname-identity tasks, both RT and accuracy were recorded asdependent variables.
Chimeric Face Test. For each pair of 36 faces in the ChimericFace Test, participants were asked to decide which one lookedhappier, thus making a simple preference judgment. This test wasscored by calculating the number of pairs in which a rightwardattentional bias was shown (i.e., the face with the smile to theindividual's right was judged happier) minus the number of pairs inwhich a leftward attentional bias was shown (i.e., the face withsmile to the individual's left was judged happier), divided by thenumber of total pairs (36).
Results
The data analysis is presented in three sections, each of
which addresses one of the central issues of this study. The
first set of analyses examines how the manipulation of task
complexity at different stages of information processing
affects the pattern of performance on across-hemisphere
trials. We present evidence that the advantages of distribut-
ing processing between the hemispheres increases with task
complexity. The second set of analyses focuses on qualifica-
tions to this across-hemisphere advantage imposed by task
laterality. We present evidence that although across-
hemisphere trials yield superior performance during com-
plex bihemispheric tasks, such an advantage does not accrue
for a task that strongly relies on one hemisphere's processing
capabilities. Finally, the last set of data analyses focuses on
individual differences in AHA and provides evidence that
AHA affects the degree to which interhemispheric interac-
tion improves performance.
Effects of Manipulating Computational Complexityat Different Stages of Information Processing
RT analysis. An initial analysis of variance (ANOVA)
conducted on the mean RT data for the physical- and
name-identity tasks with handedness as the grouping factor
revealed no significant differences in the pattern of perfor-
mance of right- and left-handers. Therefore, handedness was
dropped as a between-subjects variable, and data from right-
and left-handers were collapsed into one group.
A repeated-measures ANOVA, with the factors of deci-
sion type (physical identity, name identity), perceptual load
(three items, five items), trial type (within right hemisphere
[within RH], within left hemisphere [within LH], across
right hemisphere [across RH], across left hemisphere [across
LH]), and response hand (left, right), revealed two maineffects that suggest that the manipulation of complexity was
effective. First, a highly significant main effect of decision
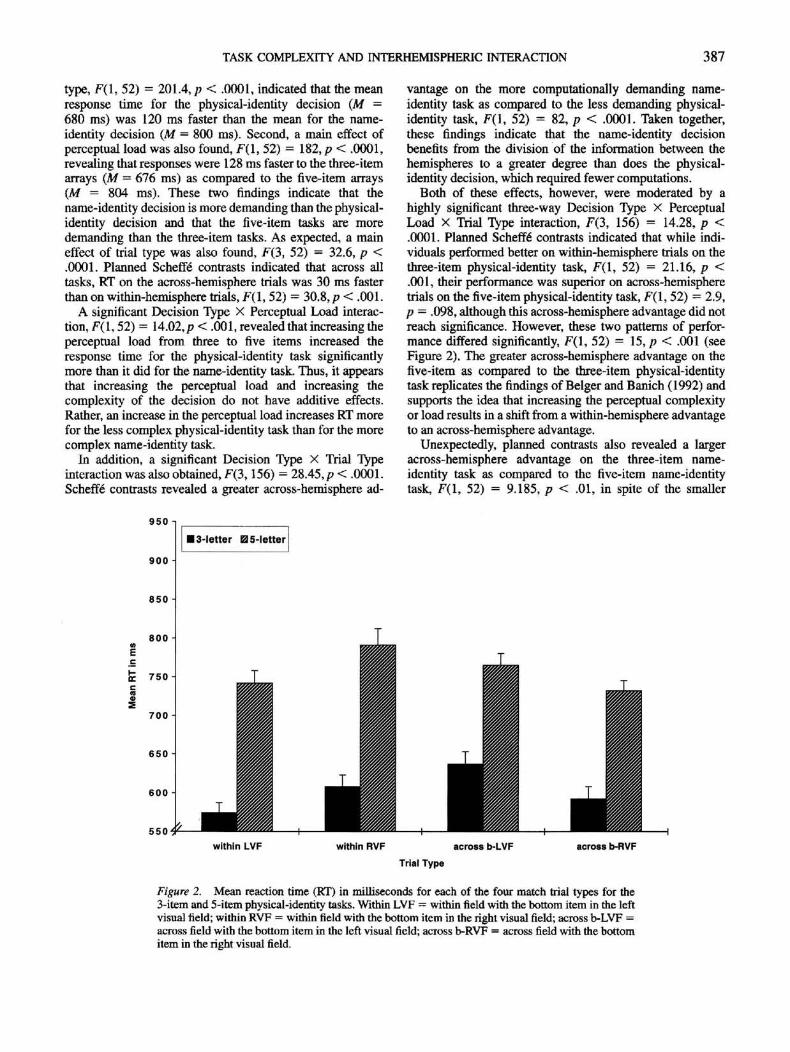
TASK COMPLEXITY AND INTERHEMISPHERIC INTERACTION 387
type, F(l, 52) = 201.4, p < .0001, indicated that the meanresponse time for the physical-identity decision (M -680 ms) was 120 ms faster than the mean for the name-identity decision (M — 800 ms). Second, a main effect ofperceptual load was also found, F(l, 52) = 182, p < .0001,revealing that responses were 128 ms faster to the three-itemarrays (M = 676 ms) as compared to die five-item arrays(M = 804 ms). These two findings indicate that thename-identity decision is more demanding than the physical-identity decision and that the five-item tasks are moredemanding than the three-item tasks. As expected, a maineffect of trial type was also found, F(3, 52) = 32.6, p <.0001. Planned SchefK contrasts indicated that across alltasks, RT on the across-hemisphere trials was 30 ms fasterthan on within-hemisphere trials, F(l, 52) = 30.8,p < .001.
A significant Decision Type X Perceptual Load interac-tion, F(l, 52) = 14.02,p < .001, revealed that increasing theperceptual load from three to five items increased theresponse time for the physical-identity task significantlymore than it did for the name-identity task. Thus, it appearsthat increasing the perceptual load and increasing thecomplexity of the decision do not have additive effects.Rather, an increase in the perceptual load increases RT morefor the less complex physical-identity task than for the morecomplex name-identity task.
In addition, a significant Decision Type X Trial Typeinteraction was also obtained, F(3,156) = 28.45, p < .0001.Scheffe contrasts revealed a greater across-hemisphere ad-
vantage on the more computationally demanding name-identity task as compared to the less demanding physical-identity task, F(l, 52) = 82, p < .0001. Taken together,diese findings indicate that the name-identity decisionbenefits from the division of the information between thehemispheres to a greater degree than does the physical-identity decision, which required fewer computations.
Both of these effects, however, were moderated by ahighly significant three-way Decision Type x PerceptualLoad X Trial Type interaction, F(3, 156) = 14.28, p <.0001. Planned Scheffe" contrasts indicated that while indi-viduals performed better on within-hemisphere trials on thethree-item physical-identity task, F(l, 52) = 21.16, p <.001, their performance was superior on across-hemispheretrials on the five-item physical-identity task, F(l, 52) = 2.9,p - .098, although this across-hemisphere advantage did notreach significance. However, these two patterns of perfor-mance differed significantly, F(l, 52) = 15, p < .001 (seeFigure 2). The greater across-hemisphere advantage on thefive-item as compared to the three-item physical-identitytask replicates the findings of Belger and Banich (1992) andsupports the idea that increasing the perceptual complexityor load results in a shift from a within-hemisphere advantageto an across-hemisphere advantage.
Unexpectedly, planned contrasts also revealed a largeracross-hemisphere advantage on the three-item name-identity task as compared to the five-item name-identitytask, F(l, 52) = 9.185, p < .01, in spite of the smaller
950 -i
900 -
850 -
800
750 -
700 -
650 -
600 -
5504
• 3-letter E35-letter
within LVF within RVF across b-LVF across b-RVF
Trial Type
Figure 2. Mean reaction time (RT) in milliseconds for each of the four match trial types for the3-item and 5-item physical-identity tasks. Within LVF = within field with the bottom item in the leftvisual field; within RVF = within field with the bottom item in the right visual field; across b-LVF =across field with the bottom item in the left visual field; across b-RVF = across field with the bottomitem in the right visual field.

388 BELGER AND BANICH
950 n
900
600
550
within LVF within RVF across b-LVF
Trial Type
across b-RVF
Figure 3. Mean reaction lime (RT) in milliseconds for each of the four match trial types for the3-item and 5-item name-identity tasks. Within LVF = within field with the bottom, item in the leftvisual field; within RVF — within field with the bottom item in the right visual field; across b-LVF =across field: with the bottom item in the left visual field; across b-RVF = across field with the bottomitem in the right visual field.
perceptual load on the three-item task (see Figure 3). Thethree-way interaction explains why we failed to obtain theexpected Perceptual Load X Trial Type interaction, F(3,156) = 0.794, p = .4991 (which we had anticipated becausewe expected the across-hemisphere advantage to be largerfor five-item than three-item displays). We attribute thisunforeseen fending to the unusually small size of theacross-hemisphere advantage obtained on the five-itemname-identity task in this experiment (5.7% of mean RT) ascompared to our previous studies (10% of mean RT; Banich& Belger, 1990; Belger & Banich, 1992). This fasterresponse on the five-item name-identity trials was notassociated with a speed-accuracy trade-off, hence we be-lieve it may be attributable to a decision deadline phenom-enon, such that participants responded without necessarilyreaching a correct decision. This proposal is supported bythe especially high error rates on these trials (26%).
Furthermore, the results of this Decision Type X Percep-tual Load X Trial Type interaction indicate that increasingtask, demands at the decision level and at the perceptual levelappear to have separable but not additive effects on theadvantages afforded by interhemispheric interaction. Pair-wise comparison indicates that the across-hemisphere advan-tage is significantly larger on the three-item name-identitytask as compared to the five-item physical-identity task, F(l,52) = 18.6, p < .001, indicating that distributing informa-tion between the hemispheres benefits performance to agreater extent when the complexity of a task is increased at
the decision level rather than the perceptual level. Yet theeffects are not additive, because the advantage afforded byacross-hemisphere processing on the five-item name-identity task is no greater than that observed on thethree-item name-identity task both in this study and in ourprior one (Belger & Banich, 1992).
Error rate analysis. The effects found in the erroranalysis were, in general, compatible with the findings formean RT. A repeated-measures ANOVA was conducted onthe error rate data, with the factors of decision type,perceptual load, trial type, and response hand (see Figures 2and 3). As with the RT data, participants were more accurateon the three-item displays than on the five-item displays asindicated by a main effect of perceptual load, F(l, 52) =74.2, p = .0001. Furthermore, a main effect of trial type alsoemerged, F(3, 52) = 17.3, p < .0001. Planned Scheffecontrasts indicated that participants responded more accu-rately overall on the aeross-hemisphere trials (errorrate = 14%) as compared to the within-heinisphere trials(error rate = 17%), F(l, 52) = 32, p < .0001.
The results also revealed a main effect of decision typeindicating that participants were overall more accurate onthe name-identity decision as compared to the physical-identity decision, F(l, 52) = 7.011, p = .01. Although thisfinding is discordant with the RT data, a correlationalanalysis between response time and accuracy indicated theabsence of a significant speed-accuracy trade-off in any ofthe cells of our ANOVA. This main effect of decision type

TASK COMPLEXITY AND INTERHEMISPHERIC INTERACTION 389
was moderated by a significant Decision Type X PerceptualLoad interaction, F(l, 52) = 71, p < .0001. Plannedcontrasts indicated that the higher accuracy on the name-identity task appears to be due to a significantly superiorperformance on the three-item name-identity task relative tothe three-item physical-identity task, F(l, 52) = 107.29,p <.001, which was significantly greater than the superioraccuracy for the five-item name-identity task relative to thefive-item physical-identity task, F(l, 52) = 10.23, p < .005(see Figures 2 and 3).
The lower accuracy on the physical-identity task ascompared to me name-identity task, despite faster responsetimes on the physical-identity task, provides evidence that itis indeed the computational complexity of a task and not justits difficulty as indexed by reaction time or error rate that islinked to die across-hemisphere advantage. The three-itemname-identity task is computationally more complex thanthe three-item physical identity task, but in this case it is notmore difficult, if difficulty is measured by accuracy level.Nonetheless, an across hemisphere advantage in RT wasobtained for the three-item name-identity task, whereas thethree-item physical-identity task yielded a within-hemi-sphere advantage. Furthermore, as will be discussed later,participants also demonstrated a significant across-hemi-sphere advantage in accuracy on the three-item name-identity task. Hence, the pattern of performance on thethree-item name-identity task strongly supports our hypoth-esis that computational complexity rather than some generallevel of difficulty causes the across-hemisphere advantage toemerge.
As with the RT results, these main effects were modifiedby an interaction of Decision Type X Trial Type, F(3,156) =5, p < .005. Planned contrasts indicated that the across-hemisphere advantage was greater for the name-identity taskas compared to the physical-identity task, F(l, 52) = 8.5,p = .005. Whereas individuals made 2% fewer errors on theacross- as compared to the within-hemisphere trials on thephysical-identity task, this difference was 5% for the name-identity task. Thus, dividing the information between thehemispheres benefited accuracy to a greater degree for thecomputationally more complex name-identity task than forthe less complex physical-identity task.
In addition, a highly significant Perceptual Load X TrialType interaction was also found, F(3, 156) = 9.226, p <.001. Planned contrasts revealed that the across-hemisphereadvantage for the five-item name task was greater than theacross-hemisphere advantage for the three-item task,F(l, 52) = 38.482, p < .001. Finally, the absence of adiree-way Trial Type X Decision Type X Perceptual Loadinteraction indicated that the across-hemisphere advantagewas comparably larger for the five-item as compared to thethree-item displays for both the physical- and the name-identity tasks.
Taken together, the results from the RT and error rateanalyses reveal that at higher levels of computationalcomplexity, dividing the processing between die hemi-spheres improves performance regardless of whether com-plexity is increased at the decision level or the perceptuallevel. Second, the results also indicate that increasing
complexity at different stages of information processingdoes not act additively to influence the advantage affordedby across-hemisphere processing. Third, the RT data suggestthat the across-hemisphere advantage appears to have aninherent ceiling effect. The relative size of the across-hemisphere advantage on the three-item arrays compared tothe five-item arrays for the name-identity task (2.5% and6.5%, respectively) was not significantly greater than therelative size of the across-hemisphere advantage on thethree-item arrays compared to the five-item arrays for thephysical-identity task. Finally, a combination of the RT anderror data suggests that the larger across-hemisphere advan-tage on the three-item as compared to the five-item name-identity task is an aberrant finding attributable to a decisiondeadline phenomenon.
Next, we examine the evidence that although interhemi-spheric interaction can enhance performance by dividingprocessing between the hemispheres, task characteristics canconstrain the degree of this advantage.
Effects of Task Lateralization
Because we predicted that me rhyme task would not yieldan across-hemisphere advantage despite being as complex assome of the tasks we have just discussed, it was necessaryfor us to conduct a within-subject comparison of the resultsobtained for the five tasks. Two right-handed participants and1 left-handed participant were excluded from this analysis,due to accuracies on the rhyme task that were below 60%.Thus, all the following analyses were conducted on data ob-tained from 23 right-handed and 27 left-handed individuals.
RT analysis. An initial ANOVA revealed no main effectsof handedness or interaction between this variable and anyothers. Hence, handedness was dropped as a between-subjects variable from the analysis, and analysis was con-ducted with the repeated measures of task, trial type, andresponse hand collapsed across the two handedness groups.
A significant effect of task, F(4, 49) = 393, p < .0001,indicated that the rhyme task was as demanding as theprevious four tasks, if not more so. Planned ScherK con-trasts revealed that the mean RT on me rhyme task wassignificantly slower than the mean RT on the four othertasks, F(l, 49) = 502, p < .0001. The ANOVA also revealeda highly significant effect of trial type and a highly signifi-cant Task X Trial Type interaction, F(3, 196) = 20, p <.0001, and F(12, 147) = 9.47, p < .0001, respectively.Planned Scheffi contrasts among the means for the within-and across-hemisphere trials on the five tasks indicated thatparticipants had a 23-ms within-hemisphere advantage forthe three-item physical-identity task, F(l, 49) = 19, p <
.001, a 20-ms marginally significant across-hemisphereadvantage for the five-item physical-identity task, F(l, 49) =3, p = .09, and significant across-hemisphere advantagesfor the three-letter and five-letter name-identity tasks,F(l, 49) = 136, p < .001, and F(l, 49) = 18, p < .001,respectively. However, the mean of within- and across-hemisphere trials for the rhyme task did not significantlydiffer, F(l, 49) = 0.418,p = .52 (see Figure 4).
The nonsignificant 7-ms across-hemisphere advantage on
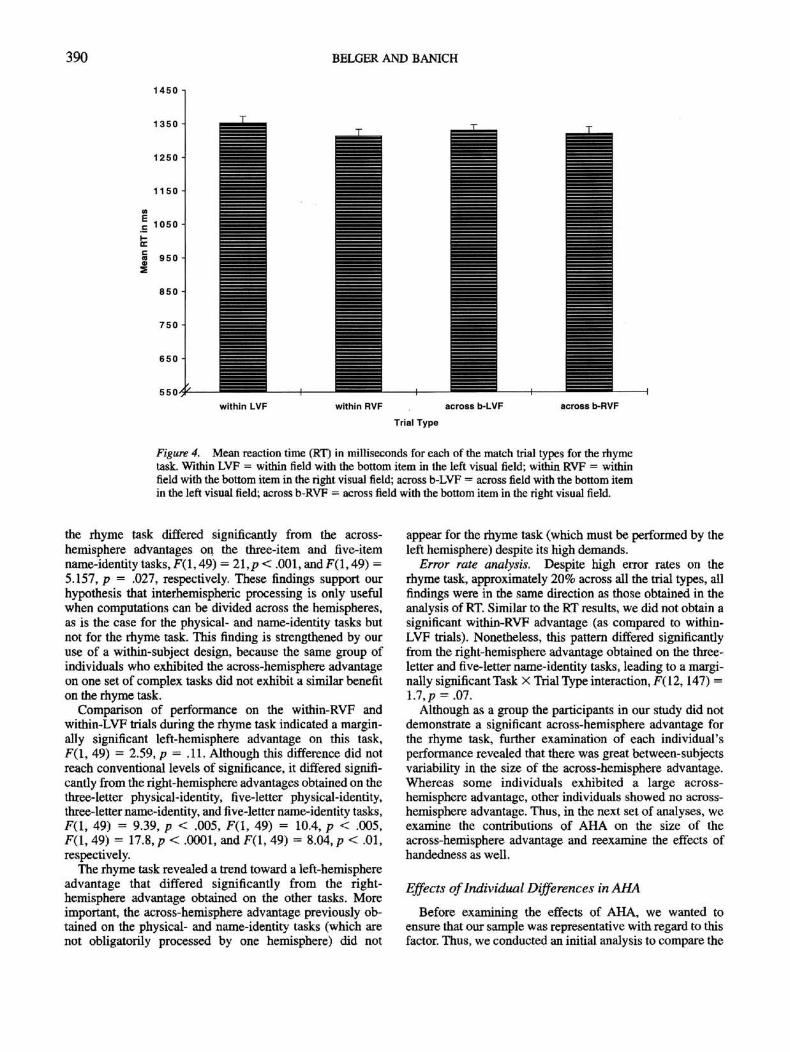
390 BELGER AND BAN1CH
1450 -,
1350 -
550-3within LVF within RVF across b-LVF
Trial Type
across b-RVF
Figure 4. Mean reaction time (RT) in milliseconds for each of the match trial types for the rhymetask. Within LVF = within field with the bottom item in the left visual field; within RVF = withinfield with the bottom item in the right visual field; across b-LVF = across field with the bottom itemin the left visual field; across b-RVF = across field with the bottom item in the right visual field.
the rhyme task differed significantly from the across-hemisphere advantages on the three-item and five-itemname-identity tasks, F(l, 49) = 2\,p < .001, and F(l, 49) =5.157, p = .027, respectively. These findings support ourhypothesis that interhemispheric processing is only usefulwhen computations can be divided across the hemispheres,as is the case for the physical- and name-identity tasks butnot for the rhyme task. This finding is strengthened by ouruse of a within-subject design, because the same group ofindividuals who exhibited the across-hemisphere advantageon one set of complex tasks did not exhibit a similar benefiton the rhyme task.
Comparison of performance on the within-RVF andwithin-LVF trials during the rhyme task indicated a margin-ally significant left-hemisphere advantage on this task,F(l, 49) = 2.59, p - .11. Although this difference did notreach conventional levels of significance, it differed signifi-cantly from the right-hemisphere advantages obtained on thethree-letter physical-identity, five-letter physical-identity,three-letter name-identity, and five-letter name-identity tasks,F(\, 49) = 9.39, p < .005, F(\, 49) = 10.4, p < .005,F(\, 49) = 17.8, p < .0001, and F(l, 49) = 8.04, p < .01,respectively.
The rhyme task revealed a trend toward a left-hemisphereadvantage that differed significantly from the right-hemisphere advantage obtained on the other tasks. Moreimportant, the across-hemisphere advantage previously ob-tained on the physical- and name-identity tasks {which arenot obligatorily processed by one hemisphere) did not
appear for the rhyme task (which must be performed by theleft hemisphere) despite its high demands.
Error rate analysis. Despite high error rates on therhyme task, approximately 20% across all the trial types, allfindings were in the same direction as those obtained in theanalysis of RT. Similar to the RT results, we did not obtain asignificant within-RVF advantage (as compared to within-LVF trials). Nonetheless, this pattern differed significantlyfrom the right-hemisphere advantage obtained on the three-letter and five-letter name-identity tasks, leading to a margi-nally significant Task X Trial Type interaction, F(12,147) =1.7,p = .07.
Although as a group the participants in our study did notdemonstrate a significant across-hemisphere advantage forthe rhyme task, further examination of each individual'sperformance revealed that there was great between-subjectsvariability in the size of the across-hemisphere advantage.Whereas some individuals exhibited a large across-hemisphere advantage, other individuals showed no across-hemisphere advantage. Thus, in the next set of analyses, weexamine the contributions of AHA on the size of theacross-hemisphere advantage and reexamine the effects ofhandedness as well.
Effects of Individual Differences in AHA
Before examining the effects of AHA, we wanted toensure that our sample was representative with regard to thisfactor. Thus, we conducted an initial analysis to compare the

TASK COMPLEXITY AND INTERHEMISPHERIC INTERACTION 391
performance of individuals in our study on the Chimeric
Face Test to those in the study of Levy et al. (1983a). The
mean asymmetry score for the right-handers in our study
was -0.280 (SD = 0.49), whereas for left-handers, it was
-0.103 (SD = 0.56). These scores are similar to the Levy et
al. (1983a) and Heller (1996) norms.
Rhyme task. To investigate the effects of the degree of
AHA, regardless of its direction, we divided right- and
left-handed individuals into two groups according to their
score on the Chimeric Face Test. Using the means and
standard deviations from the Levy et al. (1983b) study, we
assigned individuals to the strong activation (SA) group if
their score on the face booklet was either 1 SD or more
above the mean or below the mean for their respective
handedness group. They were assigned to the weak activa-
tion (WA) group if their score was within 1 SD of the mean
for their respective handedness group. Because the Levy et
al. (1983b) data showed that the means and standard
deviations were different for the right-handers (M = -0.303,
SD = 0.44) and left handers (M = -0.134, SD = 0.499),
right-handers were assigned to the SA group if their score
was below -0.743 or above 0.137 (n = 13) and to the WA
group if their score fell between -0.743 and 0.137 (n = 10).
Left-handed individuals were assigned to the SA group if
their asymmetry score was below -0.633 or above 0.365
(n = 16) and to the WA group if their score fell between
-0.633 and 0.365 (n = 11).
A repeated-measure ANOVA was then performed on the
mean RT data for the rhyme task with handedness (right,
left) and activation strength (SA, WA) as the between-
subjects grouping factors and trial type and response hand as
within-subject factors. This analysis revealed no difference
in the pattern of performance on the rhyme task for indivi-
duals in the SA and WA groups. Furthermore, mere were no
main effects of handedness, activation strength, or trial type
and none of the two-way or three-way interactions were sig-
nificant. A similar ANOVA performed on the error rate data
replicated the lack of any main effects and any interactions.
Thus, these analyses suggest that the degree of AHA does
not influence the degree of across-hemisphere processing.
To investigate the effects of direction of AHA on the size
of the across-hemisphere advantage, we divided dextrals and
sinistrals into two groups, using the data from Levy et al.
(1983b). Because the two handedness groups differed in
their mean asymmetry score, we conducted the analysis
separately for each group. Thus, right-handers with an
asymmetry score above —0.303 (the mean for right-handers)
were assigned to the left-hemisphere (LH) bias group
(n — 11), and those with a score below -0.303 were
assigned to the right-hemisphere (RH) bias group (n = 12).
Similarly, left-handers with an asymmetry score above
—0.134 (the mean for left-handers) were assigned to the LH
bias group (n — 14), and those with a score below —0.134
were assigned to the RH bias group (n = 13).
An ANOVA was conducted on the mean RT data for the
rhyme task for right-handers with bias (RH bias, LH bias) as
the grouping variable and trial type and response hand as the
within-subject variables (see Table 1). The analysis indi-
cated a significant Bias X Trial Type interaction, F(3,63) =
Table 1
Mean Reaction Times (ms) for Right- and Left-Handers in
the Left-Hemisphere {LH) Bias and Right-Hemisphere (RH)
Bias Groups on the Four Trial Types of the Rhyme Task
WithinLVF
WithinRVF
Acrossb-LVF
Acrossb-RVF
Group M error M error M error M error
Right-handersLHbias 1,333 22 1,326 21 1,369 23 1,396 23RHbias 1,386 17 1,328 20 1,308 19 1,312 21
Left-handersLHbias 1,391 24 1,349 21 1,370 26 1,344 23RHbias 1,295 20 1,254 21 1,255 13 1,261 15
Note. Within LVF = within field with the bottom item in the leftvisual field; within RVF = within field with the bottom item in theright visual field; across b-LVF = across field with the bottom itemin the left visual field; across b-RVF = across field with the bottomitem in the right visual field.
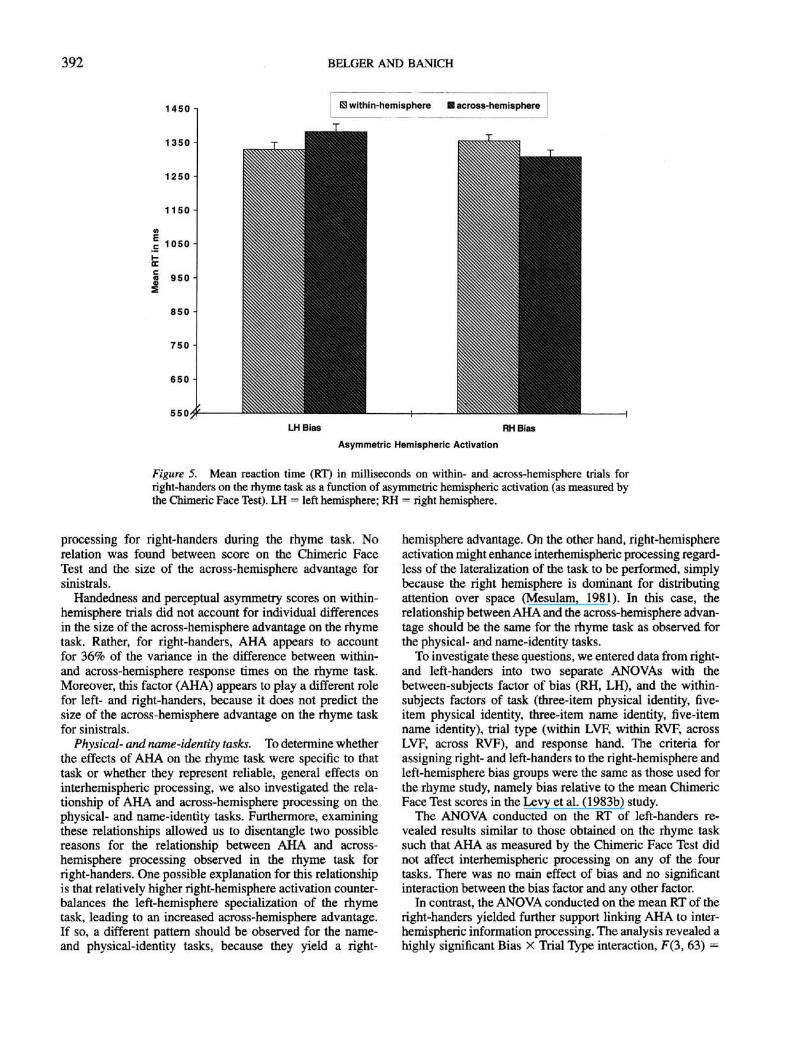
2.98, p < .04. Planned Scheffe contrasts indicated that
right-handers with an RH bias demonstrated a significant
across-hemisphere advantage, F(l, 11) = 4.92, p < .05, on
the rhyme task, whereas individuals with a LH bias demon-
strated a significant within-hemisphere advantage, F(l, 10) =
8.4, p < .02 (see Figure 5). A similar analysis conducted on
the left-handers indicated the absence of such a relation
between the direction of AHA and die across-hemisphere
advantage on the rhyme task. The error rate data were also
analyzed to examine whether the interaction of Bias X Trial
Type was significant for either handedness group, but it was
not. Thus, for right-handers, greater right-hemisphere activa-
tion yielded faster responses on across-hemisphere trials
during the rhyme task, whereas greater left-hemisphere
activation yielded faster responses on within-hemisphere
trials. In contrast, for left-handers, AHA did not predict the
degree to which interhemispheric interaction aided task
performance on the rhyme task.
In addition, we investigated whether both the magnitude
and the direction of AHA were related to the degree of the
across-hemisphere advantage. We computed, for each indi-
vidual, a measure of the size of the across-hemisphere
advantage (the RT difference between the within- and the
across-hemisphere trials relative to mean RT [(W - A)/
Mean RT]). A simple regression analysis was conducted to
determine if this score could be predicted by the score
obtained on the Chimeric Face Test. For right-handers, this
analysis revealed that the size of the across-hemisphere
advantage on the rhyme task is closely linked to the AHA, as
indicated by a highly significant correlation between the
Chimeric Face Test score and the size of the across-
hemisphere advantage on the rhyme task (r = -.599,
p < .005). The direction of the relationship indicates mat a
greater leftward bias on the face test, indicative of greater
RH activation, is associated with a greater across-hemi-
sphere advantage on the rhyme task. Because die leftward
bias is associated with a relative right-hemisphere activa-
tion, we can infer that a greater right-hemisphere activation
increases the efficiency of interhemispheric information
392 BELGER AND BANICH
1450
1350 -
E3 within-hemisphere H across-hemisphere
LH Bias RH Bias
Asymmetric Hemispheric Activation
Figure 5. Mean reaction time (RT) in milliseconds on within- and across-hemisphere trials forright-handei's on the rhyme task as a function of asymmetric hemispheric activation (as measured hythe Chimeric Face Test). LH = left hemisphere; RH = right hemisphere.
processing for right-handers during the rhyme task. Norelation was found between score on the Chimeric FaceTest and the size of the across-hemisphere advantage forsinistrals.
Handedness and perceptual asymmetry scores on within-hemisphere trials did not account for individual differencesin the size of the across-hemisphere advantage on the rhymetask. Rather, for right-handers, AHA appears to accountfor 36% of the variance in the difference between within-and across-hemisphere response times on the rhyme task.Moreover, this factor (AHA) appears to play a different rolefor left- and right-handers, because it does not predict thesize of the across-hemisphere advantage on the rhyme taskfor sinistrals.
Physical- and name-identity tasks. To determine whetherthe effects of AHA on the rhyme task were specific to thattask or whether they represent reliable, general effects oninterhemispheric processing, we also investigated the rela-tionship of AHA and across-hemisphere processing on thephysical- and name-identity tasks. Furthermore, examiningthese relationships allowed us to disentangle two possiblereasons for the relationship between AHA and across-hemisphere processing observed in the rhyme task forright-handers. One possible explanation for this relationshipis that relatively higher right-hemisphere activation counter-balances the left-hemisphere specialization of the rhymetask, leading to an increased across-hemisphere advantage.If so, a different pattern should be observed for the name-and physical-identity tasks, because they yield a right-
hemisphere advantage. On the other hand, right-hemisphereactivation might enhance interhemispheric processing regard-less of the lateralization of the task to be performed, simplybecause the right hemisphere is dominant for distributingattention over space (Mesulam, 1981). In this case, therelationship between AHA and the across-hemisphere advan-tage should be the same for the rhyme task as observed forthe physical- and name-identity tasks.
To investigate these questions, we entered data from right-and left-handers into two separate ANOVAs with thebetween-subjects factor of bias (RH, LH), and the within-subjects factors of task (three-item physical identity, five-item physical identity, three-item name identity, five-itemname identity), trial type (within LVF, within RVF, acrossLVF, across RVF), and response hand. The criteria forassigning right- and left-handers to the right-hemisphere andleft-hemisphere bias groups were the same as those used forthe rhyme study, namely bias relative to the mean ChimericFace Test scores in the Levy et al. (1983b) study.
The ANOVA conducted on the RT of left-handers re-vealed results similar to those obtained on the rhyme tasksuch that AHA as measured by the Chimeric Face Test didnot affect interhemispheric processing on any of the fourtasks. There was no main effect of bias and no significantinteraction between the bias factor and any other factor.
In contrast, the ANOVA conducted on the mean RT of theright-handers yielded further support linking AHA to inter-hemispheric information processing. The analysis revealed ahighly significant Bias X Trial Type interaction, F(3, 63) =
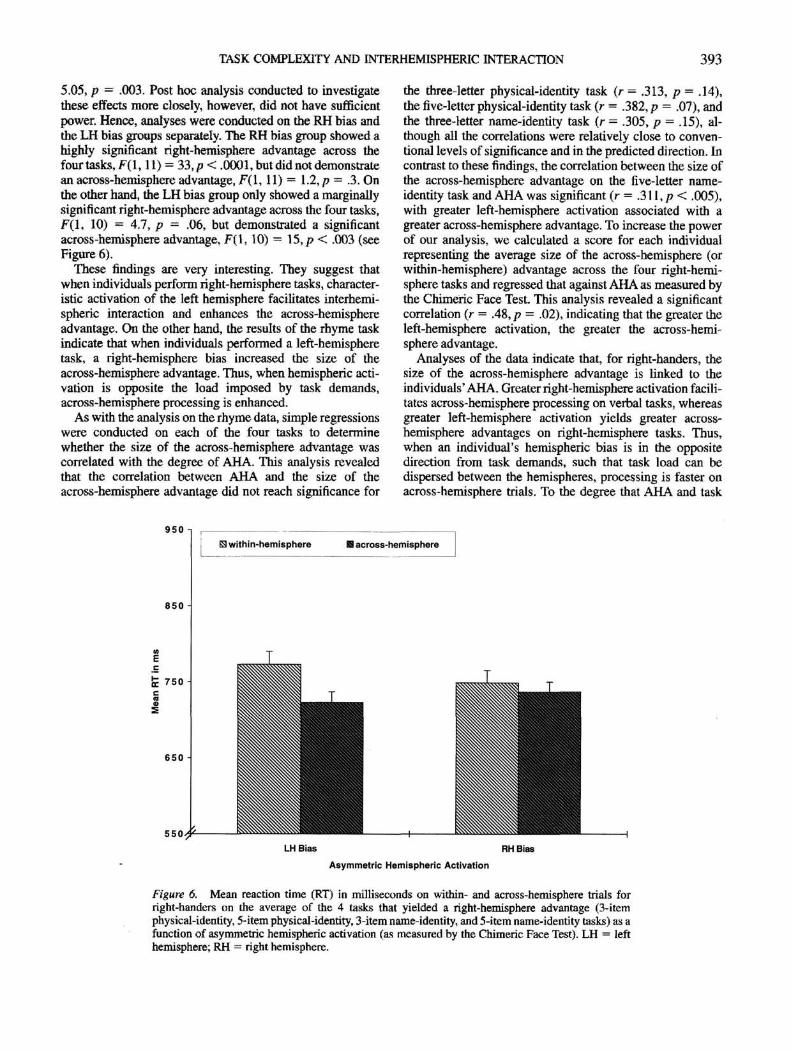
TASK COMPLEXITY AND INTERHEMISPHERIC INTERACTION 393
5,05, p = .003. Post hoc analysis conducted to investigatethese effects more closely, however, did not have sufficientpower. Hence, analyses were conducted on the RH bias andthe LH bias groups separately. The RH bias group showed ahighly significant right-hemisphere advantage across thefour tasks, F(l, 11) = 33,p < .0001, but did not demonstratean across-hemisphere advantage, F(l, 11) = 1.2, p - 3. Onthe other hand, the LH bias group only showed a marginallysignificant right-hemisphere advantage across the four tasks,F(l, 10) - 4.7, p = .06, but demonstrated a significantacross-hemisphere advantage, F(l, 10) = 15, p < .003 (seeFigure 6).
These findings are very interesting. They suggest thatwhen individuals perform right-hemisphere tasks, character-istic activation of the left hemisphere facilitates interhemi-spheric interaction and enhances the across-hemisphereadvantage. On the other hand, the results of the rhyme taskindicate that when individuals performed a left-hemispheretask, a right-hemisphere bias increased the size of theacross-hemisphere advantage. Thus, when hemispheric acti-vation is opposite the load imposed by task demands,across-hemisphere processing is enhanced.
As with the analysis on the rhyme data, simple regressionswere conducted on each of die four tasks to determinewhether the size of the across-hemisphere advantage wascorrelated with the degree of AHA. This analysis revealedthat the correlation between AHA and the size of theacross-hemisphere advantage did not reach significance for
the three-letter physical-identity task (r = .313, p = .14),the five-letter physical-identity task (r = .382, p = .07), andthe three-letter name-identity task (r — .305, p = .15), al-though all the correlations were relatively close to conven-tional levels of significance and in the predicted direction. Incontrast to these findings, the correlation between the size ofthe across-hemisphere advantage on the five-letter name-identity task and AHA was significant (r = .311, p < .005),with greater left-hemisphere activation associated with agreater across-hemisphere advantage. To increase the powerof our analysis, we calculated a score for each individualrepresenting the average size of the across-hemisphere (orwithin-hemisphere) advantage across the four right-hemi-sphere tasks and regressed that against AHA as measured bythe Chimeric Face Test. This analysis revealed a significantcorrelation (r = .48, p = .02), indicating that the greater theleft-hemisphere activation, the greater the across-hemi-sphere advantage.
Analyses of the data indicate that, for right-handers, thesize of the across-hemisphere advantage is linked to theindividuals'AHA. Greater right-hemisphere activation facili-tates across-hemisphere processing on verbal tasks, whereasgreater left-hemisphere activation yields greater across-hemisphere advantages on right-hemisphere tasks. Thus,when an individual's hemispheric bias is in the oppositedirection from task demands, such that task load can bedispersed between the hemispheres, processing is faster onacross-hemisphere trials. To the degree that AHA and task
950 -i
850 -
750
650 -
550^
E3 within-hemisphere B across-hemisphere
LH Bias RH Bias
Asymmetric Hemispheric Activation
Figure 6. Mean reaction time (RT) in milliseconds on within- and across-hemisphere trials forright-handers on the average of the 4 tasks that yielded a right-hemisphere advantage (3-itemphysical-identity, 5-item physical-identity, 3-item name-identity, and 5-item name-identity tasks) as afunction of asymmetric hemispheric activation (as measured by the Chimeric Face Test). LH = lefthemisphere; RH = right hemisphere.

394 BELGER AND BANICH
demands load on the same hemisphere, the advantageafforded by dividing processing between the hemispheres isreduced.
Discussion
Effects of Computational Complexity
on Interhemispheric Interaction
This study provides insights into both the extent to whichinterhemispheric interaction can aid task performance undercomputationally complex conditions and the constraints onsuch effects. One main issue addressed by this study waswhether increasing computational complexity at the percep-tual or comparison level would have similar effects oninterhemispheric interaction as does increasing computa-tional complexity at the decision level. Our results indicatethat both these manipulations yield a performance advantagewhen the critical information is divided between the hemi-spheres. This effect is illustrated by the presence of anacross-hemisphere advantage on the five-item physical-identity task but not on the three-item physical-identity taskand by an across-hemisphere advantage on the three-itemname-identity task that is lacking on the three-item physicaltask. Furthermore, increasing task demands at the decisionlevel and at the perceptual level appear to have separableeffects, which are not additive. Whereas the across-hemisphere advantage was approximately 5% of mean RTfor the five-item physical-identity task, it was 10% of meanRT for the three-item name-identity task. Thus, the effect oninterhemispheric interaction of increasing perceptual loadappears to be less than that imposed by increasing complex-ity at the decision level.
The absence of a greater across-hemisphere advantage onthe five-item name-identity as compared to the three-itemname-identity task indicates that the size of the between-hemisphere advantage does not increase constantly as afunction of computational complexity but rather reaches aceiling. The size of the across-hemisphere advantage in boththe three- and five-item name-identity tasks (as well as ourprevious studies using these tasks: see Experiment Id ofBanich & Belger, 1990; Belger & Banich, 1992) wasapproximately 10% of mean RT, which suggests that a pointis reached where the across-hemisphere advantage can notbe enlarged even if there is an accompanying increase incomputational complexity. This ceiling effect has beenobserved in prior studies. For example, in Banich & Belger(1990, Experiment 3), we reported that the size of theacross-hemisphere advantage was equivalent for the ordinaland summation tasks (8% and 10% of mean RT, respec-tively), even though mean RT was longer on the former thanon the latter. Similar to the present study, we had previouslyreported that the three-letter name-identity task (Banich &Belger, 1990) and the five-letter name-identity task (Belger& Banich, 1992) yield comparable across-hemisphere advan-tages (10% for each task) despite the longer RTs on the lattertask. The ceiling effect may be a reflection of the capacitylimits of each hemisphere. Dividing the processing loadbetween the hemispheres may improve performance be-
cause task demands are increased until the total load reachesor surpasses the sum of the hemispheres' processing capabilities.
It is important to note that the results obtained in this
study are consistent with a variety of other research in ourlaboratory, all of which indicate that the performance ofcomputationally complex tasks is aided by interhemisphericinteraction. Whereas we have consistently been unable toobtain an across-hemisphere advantage for simple physical-identity tasks, regardless of whether the stimuli were letters
(Banich & Belger, 1990, Experiment 1; Belger & Banich,1992), digits (Banich & Belger, 1990, Experiment 2), orshapes (Banich, Passarotti, & Chambers, 1994), we have
obtained an across-hemisphere advantage when steps aboveand beyond perceptual processing and identification are
required. For example, we have obtained an across-hemisphere advantage for a name-identity task (Banich &Belger, 1990, Experiment 1), for tasks requiring judgmentsabout the relative value of digits or their sum (Banich &Belger, 1990, Experiment 2), for a task requiring individualsto determine whether two letters are identical to those of a
target word (Banich et al., 1990), and for a task in whichindividuals make a categorization judgment about shapes(Banich et al., 1994). Furthermore, we have found sucheffects in the auditory and tactile modalities as well (Pass-arotti & Banich, 1998). Thus, repeatedly and consistently wehave demonstrated that more computationally complex tasksyield an across-hemisphere advantage.
We should also note that we believe this effect to be ageneral phenomenon, because some recent studies fromother laboratories have also demonstrated an across-hemisphere advantage under high-load conditions usingmethodologies different than ours. In one such study,Norman et al. (1992) showed that when presenting stimulibilaterally, superior performance is found at higher levels oftask difficulty when the load is divided between the hemi-spheres. Norman et al.'s stimuli consisted of dot patterns oftwo, four, or six dots. Individuals decided whether two-dotpatterns simultaneously presented either in the same visualfield or bilaterally to opposite visual fields were identical.Thus, to make a "same" decision, a simple templatejudgment was required. Their task-demand manipulationconsisted of increasing the number of dots in the display.Consistent with our findings, they observed that for samedecisions, an across-hemisphere advantage emerged at higherlevels of task demand.
A second main issue addressed by our study was thesource of this across-hemisphere advantage. Based on ourprevious findings, we posited that the across-hemisphereadvantage occurs because it allows for a division of process-ing between the hemispheres, lightening the load on each.We reasoned that if this were true, then if a computationallycomplex task relied exclusively on one hemisphere, anacross-hemisphere advantage would not be obtained. Toinvestigate this question, we tested right- and left-handedindividuals on a rhyme decision task, which requires agrapheme-to-phoneme conversion relying strongly on theleft hemisphere's processors (at least for right-handers)(Rayman & Zaidel, 1991), and tested these same individuals

TASK COMPLEXITY AND INTERHEMISPHERIC INTERACTION 395
on equally complex tasks that did not rely exclusively on one
hemisphere's processing capacity. The results indicate that
despite the complexity of the computations required by the
rhyme task, right-handers did not exhibit a performance
benefit on across-hemisphere trials as they did for the other
computationally complex tasks. Hence, these findings sug-
gest that interhemispheric information processing improves
performance to the extent that the computations required by
a particular task can be supported or carried out by either
hemisphere. It should be noted that there are very few tasks
that rely exclusively on one hemisphere. For most tasks,
both hemispheres are capable of performing them to some
degree, albeit to differing degrees of competence and in
different manners. Thus, we believe that for most computa-
tionally complex tasks (with phoneme-to-grapheme conver-
sion and die control of speech output being notable excep-
tions), a division of processing across the hemispheres will
aid task performance.
The third main issue investigated in this study was the
degree to which individual differences moderate the advan-
tages of across-hemisphere processing under computation-
ally complex conditions. We investigated two different
characteristics, AHA and handedness. The rationale for
investigating these factors was that each might affect the
ability to divide the processing load between the hemispheres.
Our results indicate that handedness did not affect the
ability to distribute the processing across the hemispheres
for complex tasks. Left-handers showed an equally large
across-hemisphere advantage on the five-letter physical-
identity, the three-letter name-identity, and the five-letter
name-identity tasks as right-handers. These findings were
consistent with earlier ones (Banich et al., 1990) in which no
differences in interhemispheric processing were found be-
tween right- and left-handers on a spelling-to-target task.
Further support for this position comes from the absence of a
correlation between the left-hemisphere advantage on the
rhyme task (as computed from die difference of within-RVF
and within-LVF trials) and the size of the across-hemisphere
advantage on this task. Such a finding suggests that lateral-
ization alone does not predict how advantageous interhemi-
spheric interaction is to task performance.
In contrast to the lack of a difference in the size of the
across-hemisphere advantage based on handedness, which
we suggest should index static organizational differences
between the hemispheres, AHA emerged as a highly reliable
factor for predicting the advantages of dividing processing
between the hemispheres, at least for right-handers. Our
results indicate that when Individuals performed right-
hemisphere tasks (in our study, the name- and the physical-
identity tasks), a bias toward left-hemisphere activation was
associated with a greater across-hemisphere advantage, at
least for speed of response. On the other hand, a bias toward
right-hemisphere activation diminished the size of the
across-hemisphere advantage for speed. The reverse pattern
was found for the rhyme task, because a right-hemisphere
bias increased the size of the across-hemisphere advantage,
whereas a left-hemisphere bias reduced it. For both the
rhyme and the name- and physical-identity tasks, the
relationship between hemispheric activation bias and across-
hemisphere advantage was a strong one, because a signifi-
cant correlation was found in both cases.
This effect, however, was limited to right-handers, be-
cause no relation was found between AHA and the size of
the across-hemisphere advantage on any of the tasks for
left-handers. This dissimilarity indicates mat there is a clear
difference between right- and left-handers in the role of this
factor and its relation to interhemispheric information pro-
cessing. The present investigation does not permit us to
pinpoint what might have caused the difference between
right- and left-handers with regard to this factor. However,
one could speculate that the consistent lateralization in
right-handers permits us to capture the dynamic influence of
AHA on interhemispheric interaction. In contrast, the vari-
ability of lateralization in left-handers in addition to the
dynamic characteristic of AHA may make it more difficult to
disentangle these effects. Note that this difference between
the effect on AHA on interhemispheric interaction in left-
handers and right-handers stands in contrast to the similari-
ties of AHA on perceptual asymmetries (i.e., lateralized
functioning) in these two handedness groups, which has
been reported to account for 45.2% of the between-subjects
variations in perceptual asymmetry scores (Kim et al., 1990).
The effects of AHA on the advantages afforded by
interhemispheric processing is not altogether surprising.
Although AHA is static for a given individual, its effects on
behavior are dynamic because they are moderated by task
demands (e.g., a right-hemisphere bias in AHA will increase
right-hemisphere processing on a right-hemisphere task but
will decrease left-hemisphere processing on a left-hemi-
sphere task). Similarly, our findings suggest that interhemi-
spheric information processing also appears to be dynamic,
because its effect varies with task demands (i.e., decreasing
performance for computationally simplex tasks but increas-
ing performance for computationally complex ones). Hence,
it is not surprising that the factor that most strongly predicts
the relative benefits of interhemispheric information process-
ing is also a measure of a dynamic aspect of hemispheric
processing rather than a more static characteristic such as
hemispheric specialization that is relatively consistent across
individuals (e.g., the left hemisphere is specialized for
speech in 95% of right-handers). The fact that the across-
hemisphere advantage is maximized when AHA is discor-
dant with task demands further supports the general prin-
ciple that performance is enhanced when task load is
distributed as equally as possible across the hemisphere.
This study and our previous results (Banich & Belger,
1990; Belger & Banich, 1992) suggest that the benefits of
interhemispheric information processing are affected by
task-related factors, such as complexity and task-specific
computations, and by individual differences, such as charac-
teristic individual differences in AHA. The conjoint consid-
eration of these two factors appears to reliably predict the
degree of across-hemisphere advantage observed in right-
handers. Dividing the processing between the hemispheres
during demanding tasks also appears to enhance perfor-
mance for left-handers. However, no single individual
difference factor appears to modulate and predict the size of
the across-hemisphere advantage during complex tasks for

396 BELGER AND BAN1CH
left-handers. Sinistrals appear to constitute a very heteroge-
neous group, and hence further studies with larger samples
should be conducted to decipher the effects of individual
differences on interhemispheric information processing in
left-handers.
A Dynamic Model of InterhemisphericInformation Processing
Based on the findings in this study, we propose a dynamic
model of interhemispheric processing. Earlier, we had
suggested a horse-race model of interhemispheric informa-
tion processing (Belger & Banich, 1992), in which, after
receipt of information, each hemisphere independently tries
to complete processing as fast as possible. Here, we expand
that conceptualization to include the effects of factors such
as attentional bias and each hemisphere's competence for
processing task-relevant information. We posit that increas-
ing the activation level of the minor or the less specialized
hemisphere for a particular task results in a greater involve-
ment of that hemisphere in the task, which in turn diminishes
the within-hemisphere advantage and enhances any advan-
tages associated with a distribution of the processing load
across the hemispheres. This distribution of the processing
load across the hemispheres would enable parallel and faster
processing of the load in a shorter time period. Hence, in
some sense, interhemispheric interaction seems to play a
regulatory role, by allowing the distribution of the computa-
tions across the hemispheres to yield more efficient informa-
tion processing. Through this mechanism, interhemispheric
interaction may also increase the amount of information that
can be processed, thus increasing the overall processing
capabilities of the brain. Conversely, the absence of a
mechanism for distributing load would result in poor
performance and decrements in overall attentional capabili-
ties at higher levels of task complexity, as observed with
split-brain patients (Kreuter et al., 1972).
This model conceptualizes interaction between the hemi-
spheres as a dynamic process that serves to transfer informa-
tion between the hemispheres and that has the emergent
characteristic of being able to influence the processing capa-
city of the brain. In this dynamic model, each hemisphere is
specialized to process information in a particular manner
(which leads to hemispheric differences depending on the
material to be processed), while at the same time possessing
a baseline level of activation, which varies among individu-
als. Although the functional specialization of the processors
shows great stability across right-handed individuals, the
baseline activation level shows strong consistency within an
individual across tasks but with great variability across indi-
viduals. Hence, during information processing, the load isdistributed such that the specific requirements of the task to
be performed and the asymmetry of an individual's hemi-
spheric activation levels interact to create a unique balance
point that would allow for optimum task performance with
the lowest level resource allocation on both hemispheres.
We speculate that this allocation of resources to particular
hemispheres is not controlled centrally but is controlled
automatically, such that most efficient processors are loaded
and complete their operations first. Thus, we are not positing
a model in which the cailosum acts as a humunculus,
overseeing the distribution of load. Rather, when the capac-
ity of a given hemisphere is being taxed, we postulate that it
serves to signal an opening of a callosal channel and the
shunting of information to the other hemisphere. Because
the efficiency of information processing in each hemisphere
is a function of both its functional specialization and its
activation level, the demands of a particular task and its
characteristics will produce a unique pattern of resource
allocation and distribution across the hemispheres.
This model interfaces well with what we know of
attention and resource allocation. Numerous studies have
demonstrated that although the human brain is a limited-
capacity information processor, these resources are not free
flowing but are task or processing specific (e.g., Friedman &
Poison, 1981; Friedman, Poison, Dafoe, & Gaskill, 1982).
By varying the nature and demands of a task (or a
combination of tasks), one can increase processing effi-
ciency without necessarily affecting overall processing
capacity. For example, although performance of a verbal
task will suffer during performance of a secondary verbal
task, it will be relatively undegraded by simultaneous
performance of a nonverbal spatial task specific (Friedman
et al., 1982). Thus, an increase in overall processing capacity
can be obtained by simply modifying the demands of the
tasks so that they rely on separate processors or by changing
activation that would increase (or decrease) the role of
different processors in task performance. Analogously, we
posit that because both hemispheres can process most
information, the brain has available, at the very least, two
alternative cognitive strategies for most any task and two
alternative processors. By dynamically redistributing the
processing load on each hemisphere depending on task
demands, one can increase the capacity of the brain. This
flexibility is somewhat constrained by individual differences
in AHA, which tend to consistently bias an individual to
invoke the processing strategy of one hemisphere over the
other. However, when such a bias is discordant with task
demands, processing can be more efficient.
References
Banich, M. T. (1985). The nature and time course of interhemi-spheric communication. Unpublished doctoral dissertation, Uni-versity of Chicago.
Banich, M. T., & Belger, A. (1990). Inter-hemispheric interaction:How do the hemispheres divide and conquer a task? Cortex, 26,77-94.
Banich, M. T., & Belger, A. (1991). Inter- versus intra-hemisphericconcordance of judgment in a non-explicit memory task. Brainand Cognition, 15, 131-137.
Banich, M. T., Elledge, V. C., & Stolar, N. (1992). Variations inlateralized processing among right-handers: Effects on patternsof cognitive performance. Cortex, 28, 273-288.
Banich, M. T., Goering, S., Stolar, N., & Belger, A. (1990).Interhemispheric processing in left- and right-handers. Interna-tional Journal oj'Neuroscience, 54, 197-208.
Banich, M. T., Passarotti, A., & Chambers, C. (1994). The role ofthe corpus cailosum in attentional processing. Poster presented

TASK COMPLEXITY AND INTEKHEMISPHERIC INTERACTION 397
at the inaugural meeting of the Cognitive Neuroscience Society,San Francisco, CA.
Beaumont, J. G., & Dimond, S. J. (1973). Transfer between the
cerebral hemispheres in human learning. Ada Psychologica, 37,87-91.
Beaumont, J. G., & Dimond, S. J. (1975). Interhemispheric transferof ngural information in right and non-right-handed subjects.Acta Psychologica, 39, 97-104.
Belger, A., & Banich, M. T. (1992). Interhemispheric interactionaffected by computational complexity. Neuropsychologia, 30,923-929.
Berger, J.-M., & Landolt, M. (1990). Bilateral processing ofnumbers: Parallel processing versus functional unity. Cortex, 26,555-566.
Boles, D. B., & Eveland, D. C. (1983). Visual and phonetic codesand the process of generation in letter matching. Journal ofExperimental Psychology: Human Perception and Performance,9, 657-675.
Bradshaw, J. L., Nettleton, N. C., & Patterson, K. (1973). Identifi-cation of mirror-reversed and non-reversed facial profiles insame and opposite visual fields. Journal of Experimental Psychol-ogy, 99,42^t8.
Brooks, L. (1968). Spatial and verbal components of the act ofrecall. Canadian Journal of Psychology, 22, 349-368.
Bryden, M. P. (1965). Tachistoscopic recognition, handedness andcerebral dominance. Neuropsychologia, 3, 1-8.
Cohen, G. (1973). Hemispheric differences in serial vs. parallelprocessing. Journal of Experimental Psychology, 97, 349-356.
Davis, R., & Schmit, V. (1971). Timing the transfer of informationbetween the hemispheres in man. Acta Psychologica, 35,335-346.
Davis, R., & Schmit, V. (1973). Visual and verbal coding in theinterhemispheric transfer of information. Brain, 95, 347-356.
Dimond, S. J. (1969). Hemisphere function and immediate memory.Psychonomic Science, 16, 111-112.
Dimond, S. J. (1971). Hemisphere function and word registration.
Journal of Experimental Psychology, 87, 183-186.Dimond, S. J., & Beaumont, J. G. (1971). Use of two cerebral
hemispheres to increase brain capacity. Nature, 232, 270-271.Dimond, S. J., & Beaumont, J. G. (1972a). Hemisphere function
and color naming. Journal of Experimental Psychology, 96,87-91.
Dimond, S. J., & Beaumont, J. G. (1972b). Processing in perceptualintegration between and within the cerebral hemispheres. BritishJournal of Psychology, 63, 509-514.
EUenberg, L., & Sperry, R. W. (1979). Capacity for holdingsustained attention following commissurotomy. Cortex, 15,421-138.
EUenberg, L., & Sperry, R. W. (1980). Lateralized division ofattention in the commissurotomized and intact brain. Neuropsy-chologia, 18, 411^118.
Friedman, A., & Poison, M. C. (1981). The hemispheres asindependent resource systems: Limited-capacity processing andcerebral specialization. Journal of Experimental Psychology:Human Perception and Performance, 7, 1031-1058.
Friedman, A., Poison, M. C., Dafoe, C. G., & Gaskill, S. (1982).Dividing attention within and between hemispheres: Testing amultiple resources approach to limited-capacity informationprocessing. Journal of Experimental Psychology, 8, 625-650.
Goodglass, H., & Quadfasel, F. A. (1954). Language laterality inleft-handed aphasics. Brain, 77, 521-548.
Green, J., Morris, R. D., Epstein, C. M., West, P. D., Engler, H. F.(1992). Assessment of the relationship of cerebral hemispherearousal asymmetry to perceptual asymmetry. Brain and Cogni-tion, 20, 264-279.
Harvey, L. O., Jr. (1978). Single representation of the visualmidline in humans. Neuropsychologia, 16, 601-610.
Haun, F. (1978). Functional dissociation of the hemispheres usingfoveal input. Neuropsychologia, 16, 725-733.
Heller, W. (1996). [Scores on the Chimeric Face Test from a sampleof over 1000]. Unpublished raw data.
Heller, W., Etienne, M. A., & Miller, G. A. (1995). Patterns ofperceptual asymmetry in depression and anxiety: Implications
for neuropsychological models of emotion and psychopathology.Journal of Abnormal Psychology, 104, 327-333.
Herdman, C. M., & Friedman, A. (1985). Multiple resources individed attention: A cross-modal test of the independence of
hemispheric resources. Journal of Experimental Psychology:Human Perception and Performance, II, 40-49.
Holtzman, J. D., & Gazzaniga, M. S. (1982). Dual task interactionsdue exclusively to limits in processing resources. Science, 218,1325-1327.
Holtzman, J. D., & Gazzaniga, M. S. (1985). Enhanced dual taskperformance following corpus commissurotomy in humans.
Neuropsychologia, 25(3), 315-321.Kim, H., & Levine, S. C. (1991a). Inferring patterns of hemispheric
specialization for individual subjects from laterality data: Atwo-task criterion. Neuropsychologia, 29, 93-105.
Kim, H., & Levine, S. C. (1991b). Sources of between-subjectsvariability in perceptual asymmetries: A meta-analytic review.Neuropsychologia, 29, 877-888.
Kim, H., Levine, S. C., & Kertesz, S. (1990). Are variations amongsubjects in laterality asymmetry real individual differences orrandom error in measurement?: Putting variability in its place.Brain and Cognition, 14, 220-242.
Kimura, D. (1961). Cerebral dominance and the perception ofverbal stimuli. Canadian Journal of Psychology, 15, 166-171.
Kimura, D. (1969). Spatial location in left and right visual fields.Canadian Journal of Psychology, 23, 445-458.
Kinsbourne, M., & Hicks, R. E. (1978). Functional cerebral space:A model for overflow, transfer and interference effects in humanperformance: A tutorial review. In J. Requin (Ed.), Attention andperformance VII (pp. 345-362). Hillsdale, NJ: Erlbaum.
Kreuter, C., Kinsbourne, M., & Trevarthen, C. (1972). Aredeconnected cerebral hemispheres independent channels? Neuro-psychologia, 10, 453-461.
Lake, D. A., & Bryden, M. P. (1976). Handedness and sexdifferences in hemispheric asymmetry. Brain and Language, 3,266-282.
Lassen, N., Ingvar, D., & Skinhoj, E. (1978). Brain function andblood flow. Scientific American, 239, 62-71.
Levine, S. C., Banich, M. T., & Kim, H. (1987). Variations inarousal asymmetry: Implications for face processing. In D.Ottoson (Ed.), Duality and unity of the brain (Wenner-GrenInternational Symposium Series, Vol. 47, pp. 207-222). London:Macmillan Press.
Levine, S. C., Banich, M. T., & Koch-Weser, M. (1984). Variationsin patterns of lateral asymmetry among dextrals. Brain andCognition, 3, 317-334.
Levine, S. C., Banich, M. T, & Koch-Weser, M. (1988). Facerecognition: A general or specific right hemisphere capacity?Brain and Cognition, 8, 303-325.
Levine, S. C., & Levy, J. (1986). Perceptual asymmetry forchimeric faces across the life span. Brain and Cognition, 5,291-306.
Levy, J., Heller, W., Banich, M. T, & Burton, L. A. (1983a). Arevariations among right-handed individuals in perceptual asymme-tries caused by characteristic arousal differences between hemi-spheres? Journal of Experimental Psychology: Human Percep-tion and Performance, 9, 329-359.

398 BELGER AND BANICH
Levy, J., Heller, W., Banich, M. T., & Burton, L. A. (1983b).Asymmetries of perception in free viewing of ehimeric faces.Brain and Cognition, 2, 404-419.
Levy, J., & Reid, M. (1978). Variations in cerebral organization as afunction of handedness, hand posture in writing, and sex.Journal of Experimental Psychology: General, 107, 119—144.
Levy-Agresti, J., & Sperry, R. W. (1968). Differential perceptualcapacities in major and minor hemispheres. Proceedings of theNational Academy of Sciences, USA, 61, 1151.
Liederman, J., Merola, J., & Hoffman, C. (1986). Longitudinal dataindicate that hemispheric independence increases during earlyadolescence. Developmental Neuropsychology, 2, 183-201.
Merola, J. L., & Liederman, J. (1985). Developmental changes inhemispheric independence. Child Development, 56, 1184-1196.
Merola, J. L., & Liederman, J. (1990). The effects of task difficultyupon the extent to which performance benefits from between-hemisphere division of inputs. International Journal ofNeurosci-ence, 51, 35^t4.
Mesulam, M.-M. (1981). A cortical network for directed attentionand unilateral neglect. Annals of Neurology, 10, 309-325.
Miller, L. K. (1981). Perceptual independence of the hemifields inchildren and adults. Journal of Experimental Child Psychology,32, 298-312.
Miller, L. K. (1983). Hemifield independence in the left-handed.Brain and Language, 20, 33-43.
Nebes, R. D. (1978). Direct examination of cognitive function inthe right and left hemispheres. In M. Kinsbourne (Ed.), Asym-metrical function of the brain (pp. 99-140). New York: Cam-bridge University Press.
Nelson, M. E., & Bowers, J. M. (1990). Brain maps and parallelcomputers. Trends in Neuroscience, 13,403-408.
Norman, W. D., Jeeves, M. A., Milne, A., & Ludwig, T. (1992).Hemispheric interactions: The bilateral advantage and taskdifficulty. Cortex, 28, 623-642.
Passarotti, A., & Banich, M. T. (1998). A generalized role ofinterhemispheric interaction under attentionally-demanding con-ditions: Evidence from the auditory and tactile modalities.Manuscript in preparation.
Piccirilli, M., Finali, G., & Sciarma, T. (1989). Negative evidenceof differences between right- and left-handers in interhemi-spheric transfer of information. Neuropsychologia, 27,1023-1026.
Poison, M. C., & Friedman, A. (1988). Task-sharing within andbetween hemispheres: A multiple-resources approach. HumanFactors, 30(5), 633-643.
Quinn, K., & Geffen, G. (1986). The development of tactile transferof information. Neuropsychologia, 24, 793-804.
Rasmussen, T., & Milner, B. (1977). The role of early left-braininjury in determining lateralization of cerebral speech functions.In S. J. Dimond (Ed.), Annals of the New York Academy ofSciences: Vol. 299. Evolution and lateralization of the brain (pp.355-369). New York: New York Academy of Sciences.
Rayman, J., & Zaidel, E. (1991). Rhyme and right hemisphere.Brain and Language, 40(1), 89-105.
Satz, P. (1980). Incidence of aphasia in left-handers: A test of somehypothetical models of cerebral speech organization. In J.Herron (Ed.), Neuropsychology of left handedness. New York:Academic Press.
Sereno, A. B., & Kosslyn, S. M. (1991). Discrimination within andbetween hemifields: A new constraint on theories of attention.Neuropsychologia, 29(7), 659-675.
Sergent, J. (1982). The cerebral balance of power: Confrontation orcooperation. Journal of Experimental Psychology: Human Per-ception and Performance, 8, 253—272.
Wickens, C. D. (1980). The structure of attentional resources. In R.Nickerson (Ed.), Attention and performance VIII (pp. 239-257).Hillsdale, NJ: Erlbaum.
Wickens, C. D., & Liu, Y. (1988). Codes and modalities in multipleresources: A success and a qualification. Human Factors, 30,599-616.
Zaidel, E., & Peters, A. M. (1981). Phonological encoding andideographic reading by the disconnected right hemisphere: Twocase studies. Brain and Language, 14, 205-234.
Received February 24, 1997
Revision received September 22, 1997
Accepted September 22, 1997 •




















