
Copper mediated neurological disorder: vision into Amyotrophic lateral sclerosis, Alzheimer and...
13
Click here to load reader
Transcript of Copper mediated neurological disorder: vision into Amyotrophic lateral sclerosis, Alzheimer and...

J
R
Cl
Aa
b
c
d
e
a
ARA
KCAAMN
C
h0
ARTICLE IN PRESSG ModelTEMB-25533; No. of Pages 13
Journal of Trace Elements in Medicine and Biology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Journal of Trace Elements in Medicine and Biology
jou rn al homepage: www.elsev ier .de / j temb
eview
opper mediated neurological disorder: Visions into amyotrophicateral sclerosis, Alzheimer and Menkes disease
nami Ahujaa,∗, Kapil Devb, Ranjeet S. Tanwarc, Krishan K. Selwald, Pankaj K. Tyagie
Department of Biotechnology, NIMS University, Jaipur, IndiaFaculty of Medicine in Hradec Kralove, University of Charles, Prague, Czech RepublicDepartment of Biotechnology, N.C. College of Engineering, Israna, IndiaDepartment of Biotechnology, Deenbandhu Chotu Ram University of Science and Technology, Murthal, IndiaDepartment of Biotechnology, Meerut Institute of Engineering and Technology, Meerut, India
r t i c l e i n f o
rticle history:eceived 30 December 2013ccepted 8 May 2014
eywords:
a b s t r a c t
Copper (Cu) is a vital redox dynamic metal that is possibly poisonous in superfluous. Metals can tradition-ally or intricately cause propagation in reactive oxygen species (ROS) accretion in cells and this may effectin programmed cell death. Accumulation of Cu causes necrosis that looks to be facilitated by DNA damage,followed by activation of P53. Cu dyshomeostasis has also been concerned in neurodegenerative disor-
opperlzheimer diseasemyotrophic lateral sclerosisenkes diseaseeurological disorders
ders such as Alzheimer, Amyotrophic lateral sclerosis (ALS) or Menkes disease and is directly related toneurodegenerative syndrome that usually produces senile dementia. These mortal syndromes are closelyrelated with an immense damage of neurons and synaptic failure in the brain. This review focuses oncopper mediated neurological disorders with insights into amyotrophic lateral sclerosis, Alzheimer andMenkes disease.
© 2014 Elsevier GmbH. All rights reserved.
ontents
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Copper metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Oxidative stress and copper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Copper in the brain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Copper and amyotrophic lateral sclerosis (ALS) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Copper–zinc superoxide dismutase (SOD1) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Oxidative stress . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
Mitochondrial dysfunction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Glutamate excitotoxicity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
Defects in axonal transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Copper and Alzheimer’s disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
Copper and Menkes disease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00ATP7A protein . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Copper-dependent localization of ATP7A and ATP7B . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
Please cite this article in press as: Ahuja A, et al. Copper mediated neurological disorder: Visions into amyotrophic lateral sclerosis,Alzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://dx.doi.org/10.1016/j.jtemb.2014.05.003
Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Conflict of interest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00Acknowledgment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 00
∗ Corresponding author. Tel.: +91 8901073412.E-mail addresses: [email protected], [email protected] (A. Ahuja).
ttp://dx.doi.org/10.1016/j.jtemb.2014.05.003946-672X/© 2014 Elsevier GmbH. All rights reserved.

IN PRESSG ModelJ
2 s in Medicine and Biology xxx (2014) xxx–xxx
I
ltpoCamnr[tfamctdftcbtstWCstctc[
C
toptlb[imoonC[
O
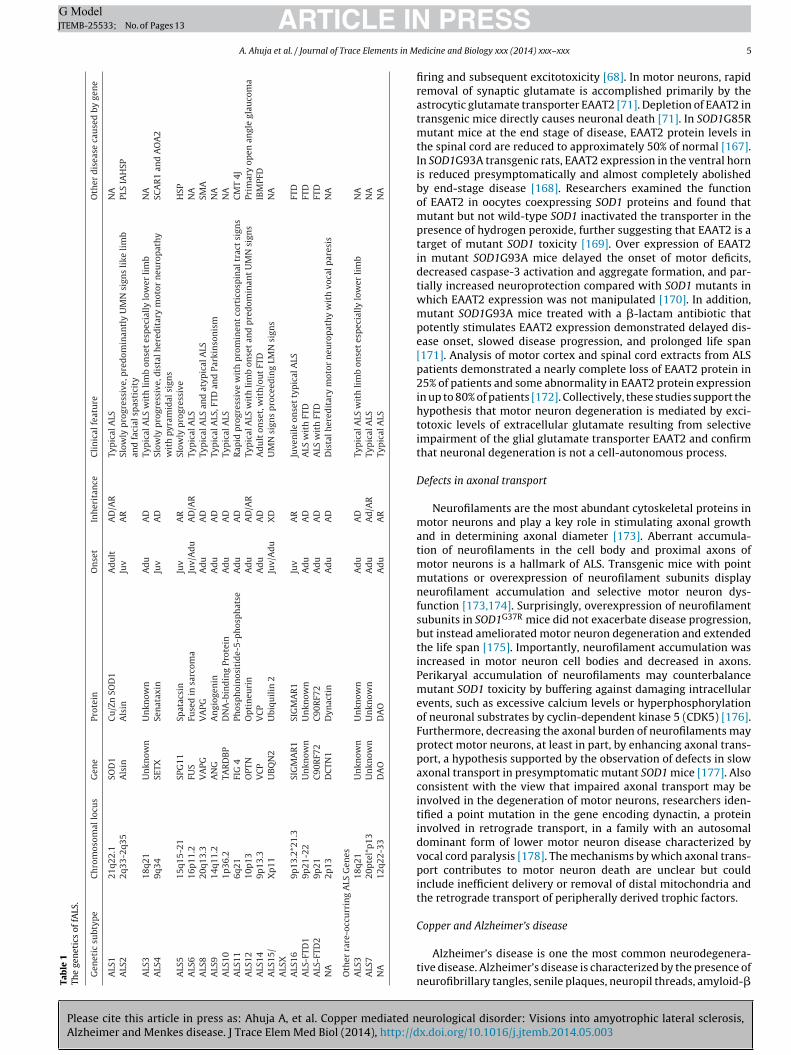
irsi(tcr
Fig. 1. Hypothetical model of ATP7A and ATP7B [43]. In this each molecule possesses
of copper transport [41,42]. Menkes disease is due to an inheriteddefect in copper absorption. Copper-transporting ATPase possesses
ARTICLETEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Element
ntroduction
Copper, an essential trace element responsible for diverse bio-ogical functions in human body [1]. Copper is present throughouthe brain, with most prominent concentration in basal ganglia, hip-ocampus, cerebellum, numerous synaptic membranes, cell bodiesf cortical pyramidal neurons and cerebellar granular neurons [2].opper is important for the normal development of the brainnd nervous system, playing a crucial role in the production andaintenance of myelin [43,124]. Beside this it is also involved in
eurotransmitter synthesis, for these reasons it plays an importantole in nerve impulse transmission and neuronal communication19,64,124,125]. At cellular level copper is required for respira-ion, peptide amidation, neurotransmitter biosynthesis, pigmentormation, and connective tissue strength [3–5,125]. Copper islso an essential component of several important enzymes; theost studied enzymes are superoxide dismutase (SOD), [9–13],
ytochrome-C oxidase, catalase, dopamine hydroxylase, uricase,ryptophan dioxygenase, lecithinase and other monoamine andiamine oxidases. Copper containing enzymes perform severalunctions in the body including oxidation–reduction reactions,ransport of oxygen and electrons and the protection of theell from oxygen radicals [14]. Role of oxidative stress haseen implicated in the progression of several neurodegenera-ive diseases like Alzheimer’s, Parkinson’s and amyotrophic lateralclerosis. Copper is also important for the prevention or modera-ion of certain neurodegenerative disease, including Parkinson’s,
ilson’s, Menkes, Alzheimer’s [15] and prion diseases [16–24].opper deficiency may induce abnormalities including anemia,keletal defects, degeneration of the nervous system, reproduc-ive failure, pronounced cardiovascular lesions, elevated bloodholesterol, impaired immunity and defects in the pigmenta-ion and structure of hair. Evidences suggest that changes inopper concentrations results in several neurological diseases3,6–8,76].
opper metabolism
Copper is readily absorbed from the diet across the small intes-ine (∼2 mg/day) and stored in the liver. The major excretory routef copper stored in liver is via the biliary pathway (∼80%) [25]. Cop-er is bound to either serum albumin or histidine and traffickedhrough the bloodstream for delivery to tissues or storage in theiver. Copper is imported into the hepatocytes via plasma mem-rane localized high-affinity human copper transporter (hCtr1)26]. HCtr1 also participates in the intracellular compartmental-zation of this metal. Once inside the cell, copper is escorted (i) to
etallothionein pool or (ii) to the mitochondria for cytochrome-Cxidase incorporation or (iii) for delivery to emerging Cu, Zn-SODr (iv) to the Wilson disease P-type ATP-ase in the trans-Golgietwork for subsequent incorporation to the ceruloplasmin [27].eruloplasmin contains about 95% of the copper found in serum37].
xidative stress and copper
Copper can induce oxidative stress by two mechanisms. Firstly,t can directly catalyze the formation of ROS via a Fenton-likeeaction [28,29]. Secondly, exposure to elevated levels of copperignificantly decreases glutathione levels [30]. Cupric and cuprous
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
ons can act in oxidation and reduction reactions. Cupric ion (CuII)), in the presence of superoxide anion radical or biological reduc-ants such as ascorbic acid or glutathione (GSH), can be reduced touprous ion (Cu (I)) which is capable of catalyzing the formation ofeactive hydroxyl radicals through the decomposition of hydrogen
six metal binding sites (Mbs) containing the GMTCxxC motif. Eight trans-membranedomains most likely form a channel for Cu (I) transport; one domain expresses a CPUmotif which is crucial for Cu (I) transport.
peroxide via the Fenton reaction [31–33].
Cu(II) + O2•− → Cu(I) + O2 (1)
Cu(I) + H2O2 → Cu(II) + •OH + OH− (Fentonreaction) (2)
The hydroxyl radical is extremely reactive and can further reactwith practically any biological molecules in the near vicinity, forexample via hydrogen abstraction leaving behind a carbon cen-tered radical, e.g. form a lipid radical from unsaturated fatty acids[37]. Copper is also capable of causing DNA strand breaks and oxi-dation of bases via ROS. Copper in both oxidative states (cupric orcuprous) was more active than iron in enhancing DNA breakageinduced by the genotoxic benzene metabolite 1,2,4-benzenetriol.DNA damage occurred mainly by a site-specific Fenton reaction[34,74].
Glutathione (GSH) a powerful antioxidant present in the cellis a substrate for several enzymes that removes ROS. It has multi-ple functions in intracellular copper metabolism and detoxification.Glutathione can suppress copper toxicity by directly chelating themetal [35], maintaining it in a reduced state and making it unavail-able for redox cycling [37]. Disruption of copper homeostasisresults in elevated cellular copper levels which may contribute toshift redox balance more toward oxidizing environment by deplet-ing glutathione levels [36,37,77]. The depletion of glutathione mayenhance the cytotoxic effect of ROS and allow the metal to be morecatalytically active, thus producing higher levels of ROS. The largeincrease in copper toxicity following GSH depletion clearly demon-strates that GSH is important cellular antioxidant acting againstcopper toxicity [38].
Copper in the brain
The mechanism of copper transport and homeostasis in the Cen-tral nervous system is still a matter of intense investigation. Coppercorrelates positively with brain development [39,40], entering theCentral nervous system from the blood stream crossing the bloodbrain barrier (BBB). Copper-transporting ATPase were identified instudies on Menkes gene enabled us to understand the mechanism
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
six heavy metal-binding sites in the N terminal part of the molecule(Fig. 1) able to pump copper ions through physiological barriers (e.g.gastrointestinal mucosa and BBB).
After crossing the BBB, copper transport or distribution in gliaand neurons is controlled by ATP7A protein and thus the metal

ARTICLE ING ModelJTEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Elements in M
Fig. 2. Copper intracellular transport [48]. Atx1 = protein that binds copper in the1+ oxidation state and delivers it to the P-type ATPase Ccc2 for translocation acrossiim
irAecbc
tTooatwofp
issmC
C
nmcuoam5ptaAihna
ntracellular membranes; CCS = copper chaperone; Fet3 = multicopper oxidase local-zed to the cell surface that oxidizes Fe(II) to Fe(III) for transport across the plasma
embrane.
on is available to enzymes involved in the biosynthesis of neu-otransmitters, such as dopamine and norepinephrine [44]. WhenTP7A is inactivated by mutation, copper becomes trapped in thendothelial cells of the BBB and the brain becomes severely ion defi-ient, leading to the profound neurological symptoms manifestedy babies affected with Menkes disease, who usually die in earlyhildhood [41,42,45].
Copper, like other redox metal ions, is sequestered in nonreac-ive species. Multiple copper-binding equilibria exist in the cell.he copper binding proteins such as metallothionein, Atx1, andther are all cytosolic and must compete for available copper inrder to regulate cellular metal ion distribution through kineticnd thermodynamic factors. The scientists recently have beguno understand molecular details of intracellular copper traffickingith the identification of copper chaperones and metallochaper-
nes [46]. Metallochaperones differ from chaperones in that theormer protect and deliver metal ions, while the latter facilitaterotein folding [47].
The intracellular copper delivery pathway starts with metal ionmport by the Ctr membrane transporters (Fig. 2). Once in the cyto-ol, the chaperone Atx1 shuttles copper to the Cu-ATP [42] in theecretory pathway, and the copper chaperone for superoxide dis-utase (SOD) (hCCS in humans, yCCS in yeasts) delivers copper to
u–Zn-SOD.
opper and amyotrophic lateral sclerosis (ALS)
Amyotrophic lateral sclerosis (ALS) is a heterogeneous group ofeurodegenerative disorders characterized by progressive loss ofotor neurons of the primary motor cortex, brainstem and spinal
ord, consequently resulting in muscle weakness, paralysis andltimately the death [49,72]. ALS is characterized by selective deathf upper and lower motor neurons, causing progressive muscletrophy, weakness, and spasticity. Denervation of the respiratoryuscles is generally the fatal event and, in most cases, occurs within
years of disease onset. It progresses at a fast pace with most of theatients dying within 3–5 years of the onset [50]. ALS may be a mul-isystem neurodegenerative disease in which brain and brain stemre also affected [49–51]. For example, several cases suffered fromLS exhibit symptoms of cognitive impairment, which is patholog-
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
cally characterized by cytoplasmic ubiquitin positive inclusions inippocampus and neocortical neurons in addition to anterior horneurons [49–51]. ALS can be inherited in an autosomal dominant,utosomal recessive, or X-linked manner. 90% ALS are sporadic
PRESSedicine and Biology xxx (2014) xxx–xxx 3
cases with no clear genetic linkage. However, the remaining 10%of cases show familial inheritance [52,53]. Approximately 20% offamilial cases are caused by mutations in the Cu/Zn superoxidedismutase (SOD1) gene [81], in addition to superoxide dismutase 1(SOD1), mutations in the genes coding for TAR DNA-Binding Protein(TARDBP), fused in sarcoma (FUS), Ubiquilin 2 (UBQLN2), C9ORF72and several others are closely associated with typical clinical phe-notype of ALS (Table 1).
Copper–zinc superoxide dismutase (SOD1)
SOD1 is a vital antioxidant protein. Human SOD1 is a 32 kDaprotein that exists as a homodimer via an intermolecular disulfidebond. Each monomer consists of a beta-barrel with eight antipar-allel beta strands and one copper and one zinc binding site locatedjust outside the beta barrel. Three histidine residues and one aspar-tic acid residue coordinate the zinc site and is responsible for thestructural stability of SOD1. Copper is responsible for the catalyticactivity of the protein. In the reduced state, the copper ion is coor-dinated with three histidine residues but in the oxidized state, itis coordinated by a fourth bridging histidine residue that is sharedby the zinc ion. Fig. 3 depicts the secondary structure of SOD1 witha ribbon diagram and demonstrates the active site in the metalbinding region. SOD1 is ubiquitously expressed in all cell types,where it catalyzes the reaction that converts superoxide to molec-ular oxygen (O2) and hydrogen peroxide (H2O2) (reactions (3) and(4)).
Cu2+ZnSOD + O2•− → Cu + ZnSOD + O2 (3)
Cu+ZnSOD + O2− + 2H+ → Cu2+ZnSOD + H2O2 (4)
These two reactions are incredibly efficient, occurring at nearlydiffusion-limited rates (2 × 109 M−1 s−1) for enzymatic catalysts atphysiological pH [54].
Apart from its high catalytic efficiency, SOD1 is an extraor-dinarily stable protein, a large part of the protein’s stability isassociated with its intramolecular disulfide bond. In the reduc-ing environment of the cytosol, stable disulfide bonds are highlyunusual. The disulfide bond is formed by the copper chaperonefor SOD1 (CCS), which also delivers the copper ion to nascentSOD1. The high activity, stability and concentration emphasizethe importance of SOD1 in the cellular environment. Mutationsin SOD1 have a dramatic effect on cells however the exactmechanism by which SOD1 mutations lead to ALS pathology isunknown although numerous hypotheses have been proposed toexplain SOD1-mediated toxicity such as oxidative stress, mitochon-drial dysfunction, glutamate excitotoxicity and defects in axonaltransport.
Oxidative stress
Oxidative stress is a hallmark feature found in the spinal cordsand cerebrospinal fluid (CSF) of sporadic and familial ALS patients[55–57]. Oxidative stress is the result of an imbalance in the pro-duction of ROS and the cell’s ability to counteract ROS productionwith antioxidants like SOD1 [58]. Most cellular ROS arise from themitochondrial respiratory chain. In the mitochondria electrons are‘leaked’ by the incomplete reduction of molecular oxygen (O2) pro-ducing superoxide (O2
•−) and hydrogen peroxide (H2O2) duringoxidative phosphorylation.Both ONOO−, the product of superox-ide and nitric oxide, and OH, the dissociation of H2O2, are highlyreactive oxidizing agents capable of damaging unsaturated fatty
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
acids in the cell membrane, altering DNA and RNA species andchanging protein conformation [58]. It is unclear whether oxida-tive stress is a primary cause of ALS or a result of some othertoxic event, but it is a key component in the progression of the

ARTICLE IN PRESSG ModelJTEMB-25533; No. of Pages 13
4 A. Ahuja et al. / Journal of Trace Elements in Medicine and Biology xxx (2014) xxx–xxx
Fig. 3. A ribbon diagram of an SOD1 homodimer. A SOD1 homodimer showing the � antiparallel-sheets, and the copper (blue sphere) and zinc (orange sphere) binding siteswithin the protein. The metal binding region is expanded to show the histidine (His46, His48, His63, His71, His80 and His120) and aspartic acid (Asp124) residue responsiblef bondc pretatv
dihdcaiccittarWt(ttTitl
M
avipkf
pddms
or coordinating the metal ions and the amino acids that contribute to the hydrogenoordinating both the zinc and copper in the reduced state of the protein. (For interersion of this article.)
isease. One of the proposed gains of function for mutant SOD1s to become a pro-oxidant rather than an anti-oxidant [59]. It isypothesized that SOD1 can increase the number of ROS, possiblyue to aberrant copper chemistry, that have damaging and toxiconsequences for the living cell. For example, increased peroxidasectivity has been found in mutant SOD1 in which hydrogen perox-de (H2O2) [13,60], a product of superoxide disproportionation, isonverted to a hydroxyl radical by Cu+. Alternatively, SOD1 mutantsan interact with new substrates such as peroxynitrite (ONOO−) toncrease the rate of oxidative damage [73,75]. Several other fac-ors can contribute to the production of ROS that could lead tohe oxidative damage seen in ALS patients. ROS can increase withge, possibly through an increasingly ‘leaky’ mitochondrial respi-ation [61,79]. This could be related to the onset of ALS later in life.
ith a compromised mutant SOD1, the increase in ROS may beoo difficult to handle for other antioxidant proteins, such as SOD2manganese SOD) and glutathione peroxidases [58]. Additionally,he response of the endoplasmic reticulum (ER) to misfolded pro-eins, such as SOD1, can increase the rate of ROS production [62].he primary source of the elevated ROS and oxidative damagen ALS is unknown, though numerous factors could contribute tohe increase and lead to oxidatively modified proteins, DNA andipids.
itochondrial dysfunction
Mitochondria are the principal source of chemical energy within cell. Mitochondrial degeneration, manifested by mitochondrialacuolization, has been reported in sporadic ALS motor neurons ands an early pathological feature in mutant SOD1 transgenic mice,receding the onset of motor weakness. Mitochondrial function isnown to decline with increasing age, which is the most robust riskactor for neurodegenerative diseases.
Serial measurements of 8-hydroxy-2′-deoxyguanosine haverovided direct evidence of an age dependent increase in oxidative
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
amage to mitochondrial DNA [63]. Abnormalities of the mitochon-rial electron transfer chain, in addition to reduced or damageditochondrial DNA, have been detected in muscle and liver biop-
ies from patients with sporadic ALS [64,65].
ing network of the active site. His63 is the bridging histidine that is responsible forion of the references to color in this figure legend, the reader is referred to the web
Reduced concentrations of manganese dependent SOD (anothermember of the SOD enzyme family located on mitochondrial mem-branes) have been detected in the muscle of patients with sporadicALS. This enzyme is essential for ROS detoxification, and reducedconcentrations would tend to exacerbate oxidative stress [65,80].Impaired mitochondrial function may lead to a cellular energydeficit. Indeed, neuronal metabolism is particularly vulnerableto acute disturbances of aerobically generated chemical energy,as exemplified by poor tolerance of ischemia. It has been pre-dicted that the large size of motor neurons (axonal processes inhuman motor neurons can approach 1 m in length) relative to othercell types and most other neurons, and consequently their highenergy demands, may cause them to be particularly vulnerable tothe adverse effects of mitochondrial impairment [67]. Metaboli-cally compromised neurons may be unable to maintain membranepotential, resulting in opening of voltage dependent NMDA subtypeglutamate receptors and calcium influx. Therefore, a much lowerconcentration of extracellular glutamate may initiate the cascadeof events leading to cell death. Treatment of mutant SOD1 trans-genic mice with creatine, which buffers cellular energy values, hasbeen shown to be partially neuroprotective; this is consistent withthe hypothesis that neuronal energy depletion is pathologicallyrelevant in ALS [66].
Glutamate excitotoxicity
A well-recognized mechanism of neuronal death is glutamateexcitotoxicity resulting from repetitive cell firing or the influx ofexcessive levels of calcium through permissive glutamate recep-tors [68]. Glutamate-mediated neurotoxicity was first proposedas a mechanism of motor neuron degeneration when increasedlevels of glutamate were discovered in the cerebrospinal fluid ofALS patients [69]. A follow-up study reported that increased cere-brospinal fluid glutamate levels were apparent in approximately
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
40% of nearly 400 patients with sporadic ALS and correlated withdisease severity; this was one of the largest pathogenesis-orientedhuman studies in ALS [70]. Clearance of glutamate from the synap-tic cleft by glutamate transporters is critical in preventing repetitive
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
ARTICLE ING ModelJTEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Elements in MTa
ble
1Th
e
gen
etic
s
of
fALS
.
Gen
etic
subt
ype
Ch
rom
osom
al
locu
s
Gen
e
Prot
ein
On
set
Inh
erit
ance
Cli
nic
al
feat
ure
Oth
er
dis
ease
cau
sed
by
gen
e
ALS
1
21q2
2.1
SOD
1
Cu
/Zn
SOD
1
Ad
ult
AD
/AR
Typ
ical
ALS
NA
ALS
2
2q33
-2q3
5
Als
in
Als
in
Juv
AR
Slow
ly
pro
gres
sive
, pre
dom
inan
tly
UM
N
sign
s
like
lim
ban
d
faci
al
spas
tici
tyPL
S
IAH
SP
ALS
3
18q2
1
Un
know
n
Un
know
n
Ad
u
AD
Typ
ical
ALS
wit
h
lim
b
onse
t
esp
ecia
lly
low
er
lim
b
NA
ALS
4
9q34
SETX
Sen
atax
in
Juv
AD
Slow
ly
pro
gres
sive
, dis
tal h
ered
itar
y
mot
or
neu
rop
ath
yw
ith
pyr
amid
al
sign
sSC
AR
1
and
AO
A2
ALS
5
15q1
5-21
SPG
11
Spat
acsi
n
Juv
AR
Slow
ly
pro
gres
sive
HSP
ALS
6
16p
11.2
FUS
Fuse
d
in
sarc
oma
Juv/
Ad
u
AD
/AR
Typ
ical
ALS
NA
ALS
8
20q1
3.3
VA
PG
VA
PG
Ad
u
AD
Typ
ical
ALS
and
atyp
ical
ALS
SMA
ALS
9
14q1
1.2
AN
G
An
giog
enin
Ad
u
AD
Typ
ical
ALS
, FTD
and
Park
inso
nis
m
NA
ALS
10
1p36
.2
TAR
DB
P
DN
A-b
ind
ing
Prot
ein
Ad
u
AD
Typ
ical
ALS
NA
ALS
11
6q21
FIG
4
Phos
ph
oin
osit
ide-
5-p
hos
ph
atse
Ad
u
AD
Rap
id
pro
gres
sive
wit
h
pro
min
ent
cort
icos
pin
al
trac
t si
gns
CM
T
4JA
LS12
10p
13
OPT
N
Op
tin
euri
n
Ad
u
AD
/AR
Typ
ical
ALS
wit
h
lim
b
onse
t
and
pre
dom
inan
t
UM
N
sign
s
Prim
ary
open
angl
e
glau
com
aA
LS14
9p13
.3
VC
P
VC
P
Ad
u
AD
Ad
ult
onse
t,
wit
h/o
ut
FTD
IBM
PFD
ALS
15/
ALS
XX
p11
UB
QN
2
Ubi
quil
in
2
Juv/
Ad
u
XD
UM
N
sign
s
pro
ceed
ing
LMN
sign
s
NA
ALS
16
9p13
.2*2
1.3
SIG
MA
R1
SIG
MA
R1
Juv
AR
Juve
nil
e
onse
t
typ
ical
ALS
FTD
ALS
-FTD
1
9p21
-22
Un
know
n
Un
know
n
Ad
u
AD
ALS
wit
h
FTD
FTD
ALS
-FTD
2
9p21
C90
RF7
2
C90
RF7
2
Ad
u
AD
ALS
wit
h
FTD
FTD
NA
2p13
DC
TN1
Dyn
acti
n
Ad
u
AD
Dis
tal h
ered
itar
y
mot
or
neu
rop
ath
y
wit
h
voca
l par
esis
NA
Oth
er
rare
-occ
urr
ing
ALS
Gen
esA
LS3
18q2
1
Un
know
n
Un
know
n
Ad
u
AD
Typ
ical
ALS
wit
h
lim
b on
set
esp
ecia
lly
low
er
lim
b
NA
ALS
7
20p
tel*
p13
Un
know
n
Un
know
n
Ad
u
Ad
/AR
Typ
ical
ALS
NA
NA
12q2
2-33
DA
O
DA
O
Ad
u
AR
Typ
ical
ALS
NA
PRESSedicine and Biology xxx (2014) xxx–xxx 5
firing and subsequent excitotoxicity [68]. In motor neurons, rapidremoval of synaptic glutamate is accomplished primarily by theastrocytic glutamate transporter EAAT2 [71]. Depletion of EAAT2 intransgenic mice directly causes neuronal death [71]. In SOD1G85Rmutant mice at the end stage of disease, EAAT2 protein levels inthe spinal cord are reduced to approximately 50% of normal [167].In SOD1G93A transgenic rats, EAAT2 expression in the ventral hornis reduced presymptomatically and almost completely abolishedby end-stage disease [168]. Researchers examined the functionof EAAT2 in oocytes coexpressing SOD1 proteins and found thatmutant but not wild-type SOD1 inactivated the transporter in thepresence of hydrogen peroxide, further suggesting that EAAT2 is atarget of mutant SOD1 toxicity [169]. Over expression of EAAT2in mutant SOD1G93A mice delayed the onset of motor deficits,decreased caspase-3 activation and aggregate formation, and par-tially increased neuroprotection compared with SOD1 mutants inwhich EAAT2 expression was not manipulated [170]. In addition,mutant SOD1G93A mice treated with a �-lactam antibiotic thatpotently stimulates EAAT2 expression demonstrated delayed dis-ease onset, slowed disease progression, and prolonged life span[171]. Analysis of motor cortex and spinal cord extracts from ALSpatients demonstrated a nearly complete loss of EAAT2 protein in25% of patients and some abnormality in EAAT2 protein expressionin up to 80% of patients [172]. Collectively, these studies support thehypothesis that motor neuron degeneration is mediated by exci-totoxic levels of extracellular glutamate resulting from selectiveimpairment of the glial glutamate transporter EAAT2 and confirmthat neuronal degeneration is not a cell-autonomous process.
Defects in axonal transport
Neurofilaments are the most abundant cytoskeletal proteins inmotor neurons and play a key role in stimulating axonal growthand in determining axonal diameter [173]. Aberrant accumula-tion of neurofilaments in the cell body and proximal axons ofmotor neurons is a hallmark of ALS. Transgenic mice with pointmutations or overexpression of neurofilament subunits displayneurofilament accumulation and selective motor neuron dys-function [173,174]. Surprisingly, overexpression of neurofilamentsubunits in SOD1G37R mice did not exacerbate disease progression,but instead ameliorated motor neuron degeneration and extendedthe life span [175]. Importantly, neurofilament accumulation wasincreased in motor neuron cell bodies and decreased in axons.Perikaryal accumulation of neurofilaments may counterbalancemutant SOD1 toxicity by buffering against damaging intracellularevents, such as excessive calcium levels or hyperphosphorylationof neuronal substrates by cyclin-dependent kinase 5 (CDK5) [176].Furthermore, decreasing the axonal burden of neurofilaments mayprotect motor neurons, at least in part, by enhancing axonal trans-port, a hypothesis supported by the observation of defects in slowaxonal transport in presymptomatic mutant SOD1 mice [177]. Alsoconsistent with the view that impaired axonal transport may beinvolved in the degeneration of motor neurons, researchers iden-tified a point mutation in the gene encoding dynactin, a proteininvolved in retrograde transport, in a family with an autosomaldominant form of lower motor neuron disease characterized byvocal cord paralysis [178]. The mechanisms by which axonal trans-port contributes to motor neuron death are unclear but couldinclude inefficient delivery or removal of distal mitochondria andthe retrograde transport of peripherally derived trophic factors.
Copper and Alzheimer’s disease
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
Alzheimer’s disease is one the most common neurodegenera-tive disease. Alzheimer’s disease is characterized by the presence ofneurofibrillary tangles, senile plaques, neuropil threads, amyloid-�

ING ModelJ
6 s in M
(otTtaugNfaowdwbiithaopfclhpoc
ctf
ARTICLETEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Element
A�) deposits and selective loss of neurons. Neuro-fibrillary tanglesccur in large numbers in the AD brain, in the entorhinal cor-ex, hippocampus, amygdala and the association cortices of lobes.he subunit protein of the paired helical filaments is the tau pro-ein, present in tangles as bundles of paired, helically wound tauggregates. These intra neuronal proteinaceous inclusions are oftenbiquitinated deposits, similar to those found in other neurode-enerative disorders, such as Parkinson and Lewy body disease.euritic plaques are spherical, multicellular lesions that are usually
ound in moderate or large numbers in limbic structures and in thessociation neocortex. They are composed of extracellular depositsf the amyloid A� protein with degenerating axons and dendritesithin and intimately surrounding the amyloid deposit. Amyloideposits are primarily composed of the A� amyloid protein ofhich the A�(1–40) isoform is the predominant soluble species in
iological fluids and the Ab1–42 isoform is the predominant speciesn developing plaque deposits [82–90]. The importance of coppern AD development depends on neurotoxic interaction betweenhe A� peptide and copper. A� peptide binds to copper with veryigh affinity (7 × 10−18 M) [91] reducing Cu (II) to Cu (I) with a cat-lytic generation of H2O2 and A� aggregation [92]. This high affinityf peptide A�(1–42) (abnormally cleaved from a larger amyloidrecursor protein (APP) for copper is due to its high �-sheet con-ormation (conformational pathologies), which favors high-affinityopper binding [93]. APP expression is a key issue in the modu-ation in neuronal copper homeostasis since APP knockout miceave increased copper levels in the brain [94]. High levels of cop-er were found in senile plaques [95] and in the cerebrospinal fluidf AD patients, concomitantly an increased copper transport viaeruloplasmin [96] have also been reported.
Several hypotheses have been proposed for the possible role ofopper in the progression of Alzheimer’s disease. However fromhe current understanding of the disease we can suggest that theollowing steps may be involved in progression of the disease.
1. Copper dyshomeostasis begins, with a decrease in the intracel-lular Cu+ pool [87,97].
2. Decreased release of Cu+ into the synaptic cleft resultsin decreased inhibition of N-methyl d-aspartate receptor(NMDAR) [88,98].
3. Decreased inhibition of NMDAR by Cu2+ results in increased A�release as compensation to avoid excitotoxicity [92,102].
4. Decreased intracellular Cu2+ results in a favoring of amyloidgenic processing of A�, with an increase in the A� (1–42):A�(1–40) ratio [126,127].
5. A� clearance is impaired [89,99] due to a decrease in neprilysin,resulting in a further increase in A� concentration in the synap-tic cleft.
6. Carnosine levels decrease [100], increasing the potential forA�:Cu2+ complex formation. The carnosine decrease alsoincreases the likelihood of advanced glycation end-productformation and an increase in oxidative stress at the synapse[90,100,131].
7. A� reaches a critical concentration at the synapse where it canbegin to compete with remaining carnosine ligands for Cu2+,due to A�’s high affinity for Cu2+ [90,130].
8. Sub-stoichiometric Cu2+ binding to A� results in rapid Cu2+
induced A� fibril formation, where Cu2+ bound A� formed fib-rils twice as fast as apo-A�.
9. Once formed, these fibrils seed monomeric A� to result in fur-ther fibril formation. Additionally the results show that onceformed, fibrils can continue to bind Cu2+, as Cu2+ can diffuse
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
through the fibril structure to form a 1:1 stoichiometry with thefibers. Importantly, once Cu2+ bound A� aggregates are formed,metallothionein is unable to aid deaggregation of, or extractcopper from, the aggregates [91,101].
PRESSedicine and Biology xxx (2014) xxx–xxx
10. Newly released Cu+ or A� in response to NMDAR activationis ineffective as instead of binding to the NMDAR they aresequestered into the fibrils, resulting in prolonged NMDAR acti-vation, and further release of Cu+ and A� in response [128].
11. It was demonstrated that A�-Cu2+ fibers were significantlymore toxic to PC12 cells than apo-A� fibers or Cu2+ bound tothe soluble fragment A�(1–16). These fibers result in physicaldamage to the neuronal cell membrane [129].
12. Redox cycling of Cu2+ bound to A� and the low levels ofcarnosine result in increased oxidative stress at the synapse[130–132].
13. Inefficient inhibition of NMDAR by Cu+ and A� results in exci-toxicity [102].
14. Neuronal cell death and synapse loss begin to occur, resulting inolfactory dysfunction. Amyloid plaques spread from the olfac-tory bulb through the brain, eventually resulting in widespreadmemory loss and loss of brain function and severe AD pathology[127,133,134].
Copper and Menkes disease
Menkes disease (MD) is an X-linked multisystemic lethaldisorder of impaired copper metabolism characterized by neu-rodegenerative symptoms and connective tissue manifestation[103,135,136]. Menkes disease (MD) is a fatal neurodegenera-tive disorder characterized by symptoms like progressive cerebralatrophy, seizures, hypotonia, coarse hair and connective tis-sue abnormalities [103,104]. These pleiotropic symptoms can beattributed to the redudced activity of a range of copper depen-dent enzymes, which include cytochrome C oxidase, dopamine �hydroxylase, lysl oxidase, and tyrosinase [136]. Menkes patient suf-fer a systemic copper deficiency due to poor absorption of dietarycopper in small intestine and defective distribution within the body[105,106]. Affected individuals have reduced copper in brain, liverand serum, but elevated level in kidney and small intestine. Theserequirements of copper seem particularly acute during the initial12 months of life, when the velocity of brain growth and motorneurodevelopment is normally rapid [107]. Involvement of coppermetabolism in MD was first described by Danks [105] showing lowserum levels and defective intestinal absorption of the metal. Atfirst, copper malabsorption was thought to be the primary cause,but later investigations showing copper accumulation in extra-hepatic tissues, apart from the brain, indicated a multisystemicinvolvement [137,138].
The genes defective in menkes disesase (ATP7A;MNK) encodesa protein that has all the features of P-type ATPases, a fam-ily of protein that transfers cations through the lipid bilayer ofthe membranes using the energy derived from hydrolysis of ATP[2,19,108–119].
ATP7A protein
ATP7A is a member of a large family of P-type ATPases that areenergy-utilizing membrane proteins functioning as cation pumps(Fig. 1) [139]. They are called ‘P-type’ ATPases, as they form aphosphorylated intermediate during the transport of cations acrossa membrane. The super-family of P-type ATPases also includesthe Na+/K+ and H+/K+ pumps, as well as plasma membrane andsarcoplasmatic reticulum Ca2+ pumps [140]. ATP7A and ATP7Btransports copper across a membrane using the energy releasedby hydrolysis of ATP. This process involves domains specific for
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
binding and hydrolysis of ATP, and is similar in all P-type ATPases.The domains involved in the catalytic cycle of the protein are thenucleotide-binding domain (N-domain), phosphorylation domain(P-domain), and activation domain (A-domain). Transport and

ING ModelJ
s in M
tsawAsrattThcabttodrgrrFPTg
Fbrcbir
ARTICLETEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Element
ranslocation of copper furthermore requires special motifs andtructures for recognition, binding, and translocation of the metalcross a membrane. These motifs contain cysteine (C) residues,hich play an important role in copper binding. At the N-terminusTP7A has six metal-binding domains (MBD1–6) each with a con-ensus MTXCXXC motif. Copper binds to these domains in theeduced form, Cu (I). It is assumed that the two MBDs (MBD5nd MBD6) closest to the transmembrane domains are impor-ant for the functional activity of the protein, and at least one ofhese two sites is necessary for normal function of the protein.he first four metal-binding domains (MBD1–4) are thought toave a regulatory function. Interaction between ATP7A and theopper chaperone ATOX1 occurs through these domains. ATP7A isnchored to a membrane through eight hydrophobic transmem-rane domains, which form a channel for copper translocationhrough the membrane, and the CPC motif within TMD6 is assumedo play a direct role in the transfer of copper. The catalytic activityf ATP7A is likely mediated through a coordinated action of the N-omain, the P-domain, and the Adomain. The N- and P-domainseside between TMD6 and TMD7. N-domain binds ATP and the-phosphate of ATP is then transferred to the invariant aspartateesidue (D) in the DKTG motif, which resides in the P-domain. Thisesults in the formation of a transient phosphorylated intermediate.
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
ollowing translocation of the copper through the membrane, the-domain is dephosphorylated. The A domain is located betweenMD4 and TMD5, and includes the invariant TGE motif, where thelutamate residue (E) plays a key role in dephosphorylation of the
ig. 4. Schematic representation of copper-induced relocalization of ATP7A and ATP7B [oth cells, a copper enters through the copper transporter 1 (CTR1), and is then distributeise in copper concentrations,ATP7A and ATP7B relocalize from the TGN to the cell periphopper .The main difference is these two copper transport pathways lies in the direction.
asolateral side, whereas in the hepatocyte copper is excreted into the bloodstream at thnto the bile.The numbers indicate localization defects of ATP7A and ATP7B due to MD cesulting in constitutive localization at the TGN,(2) constitutive localization at the cell pe
PRESSedicine and Biology xxx (2014) xxx–xxx 7
phosphorylated intermediate. At the C-terminal ATP7A containsconserved di-leucine residues (LL), which is necessary for retrievalof the protein from the membranes (plasma membrane or vesicles).
ATP7A gene encodes a highly conserved copper translocat-ing P (1B)-type ATPase with orthologues present in eukaryotes,prokaryotes and archaea [121]. ATP7A mRNA is expressed in awide range of human tissues, but its expression is notably lowand sometimes even undetectable in the adult liver [118,120]. TheATP7A polypeptide contains several conserved domains requiredfor ATPase function and copper binding. It has been estimatedthat one-third of MD cases arise from de novo mutations [110].Over 200 MD-causing mutations have been identified, amongwhich small deletions/insertions, nonsense mutations, missensemutations, and splice site mutations are represented with equalfrequency [122,123]. Small deletions/insertions and nonsensemutations are found throughout the whole gene. In contrast, mis-sense mutations are almost exclusively distributed between thefirst trans-membrane domain and the stop codon. The relativelack of MD-causing missense mutations within the six metal-binding sites (MBSs) located in the amino terminal tail suggeststhat functional redundancy exists among these six MBSs. Splice-site mutations are mostly clustered between exons 6 and 8, whichencode a region just upstream of the first trans-membrane domain,
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
and between exons 21 and 22, a region encoding the last trans-membrane domain. These mutations lead to abnormal coppermetabolism which is in turn is responsible for the developmentof Menkes disease.
166]. Left side depicts an enterocyte, and right side represents and hepatocyte. Ind via the copper chaperone ATOX1 to ATP7A or ATP7B residing in the TGN. After aery, and in the case of ATP7A also the plasma membrane, to facilitate excretion of
In the enterocyte, ATP7A facilitates excretion of copper into the bloodstream at thee basolateral side whereas in the hepatocyte copper is excreted at the apical sideausing and AD causing mutations respectively: (1) lack of copper responsiveness,riphery, and (3) mislocalization at the ER, presumably due to misfolding.

ING ModelJ
8 s in M
ttldtisccded
C
twccbpwrtaAttt
TE
E
ARTICLETEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Element
ATP7A plays a significant role in the development and main-enance of the central nervous system. ATP7A serves as a copperransporter in vascular endothelial cells and retinal pigment epithe-ium forming the blood–brain barrier and is thus important forelivery of copper to the brain. This gene encodes a copper-ransporting P-type ATPase, it forms a covalently phosphorylatedntermediate from transfer of the gamma-phosphate of ATP to apecific aspartate residue at the catalytic site of the protein. Defi-iency of the ATP7A gene product results in abnormal cellularopper transport and decreased activities of numerous copper-ependent enzymes. Decreased activity of the copper dependentnzyme cytochrome-C oxidase is likely a major factor in the brainamage associated with Menkes disease.
opper-dependent localization of ATP7A and ATP7B
Under basal conditions, ATP7A and ATP7B are localized withinhe TGN (Fig. 4) [142–144,162]. This localization is consistentith their function in cuproenzyme biosynthesis, as several
uproenzymes are synthesized within the secretory pathway. Someontroversy exists about the localization of ATP7B, as it has alsoeen suggested that this protein resides in an endosomal com-artment [145,146]. In addition, a smaller isoform of ATP7B exists,hich has been localized to mitochondria [155]. However, the cur-
ent general agreement is that ATP7B is localized to the TGN, ashis has been confirmed by several independent groups, includingt ultrastructural resolution in human liver biopsies [143,147–153].
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
key mechanism in the regulation of the copper export func-ion of ATP7A and ATP7B became apparent from studies showinghat the subcellular distribution of both proteins was sensitiveo the concentration of copper to which the cell was exposed. In
able 2ffect of MD-causing missense mutations on function and regulation of ATP7A [166].
Mutation Cu proenzyme Biosynthesis Catalytic ATPaseactivity
Localization
A629P Reduced rescue Cc2 yeastS627L Reduced rescue Cc2 yeastR844H No signal observ
fibroblastG860V No signal observ
fibroblastL873R No rescue Cc2 yeast Increased formation
of acylphosphateintermediate
Cell periphery acopper response
G876R No signal observfibroblast
Q924R Partial in cell peC1000R No tyrosinase activity Normal, TGN, noA1007V Normal, TGNG1015D Normal, TGND1019D Reduced rescue Cc2 yeast Partial ER misloc
presentD1044G Normal, TGNK1282E Normal, TGNG1300E Normal, TGNG1302V Partial in cell peG1304S Reduced rescue Cc2 yeastN1304K Normal, TGND1305A Normal, TGNG1315RA1325V No signal observ
fibroblastA1362D Reduced rescue Cc2 yeastA1362V Normal, TGN, noA1369R No signal observ
fibroblastS1397F Normal, TGN
R, endoplasmic reticulum; TGN, trans-Golgi-network.
PRESSedicine and Biology xxx (2014) xxx–xxx
response to raised copper levels, ATP7A reversibly relocalizes toa peripheral vesicular compartment and to the plasma membrane[156,157]. In polarized cells, and in intestinal tissue sections, ATP7Aspecifically localizes toward the basolateral membrane upon cop-per exposure, consistent with its function in transferring copperacross the intestinal barrier [157–160]. ATP7A overexpressed inmouse liver tissue also localizes at the hepatocyte basolateralmembrane [161]. Specific targeting of ATP7A to the basolateralmembrane appears to be mediated through a putative PDZ bind-ing motif present in the carboxy terminal tail of ATP7A [158].Deletion of this motif results in targeting of ATP7A to the api-cal membrane in response to raised copper levels [158]. ATP7Bundergoes a similar copper-induced relocalization to a peripheralvesicular compartment. Although ATP7B has not been unequiv-ocally detected at the plasma membrane of polarized cells, theidea that the transporter rapidly recycles between the periph-eral vesicular compartment and the plasma membrane cannot beexcluded [143,162]. In polarized hepatocytic cell lines, exposureto high copper concentrations results in localization of ATP7B inthe proximity of the apical vacuoles, a structure reminiscent ofthe bile canaliculus [149,151,154,162]. This observation is consis-tent with its proposed function in excretion of copper from thehepatocyte via the bile. Small amounts of ATP7B localized in theproximity of the bile canaliculus have also been detected in humanliver tissues [141,151]. Signals mediating the specific targeting ofATP7B toward the apical region seem to be present within thefirst 63 amino acids, a region that is not present in ATP7A [154].
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
Taken together, these data indicate that differences in the traffick-ing destinations of ATP7A and ATP7B are not caused by generalcell-type specific differences in regulation of polarized membraneprotein localization, but rather are determined by intrinsic signals
Post-translationalModifications
Protein-2interactions
ed, protein absent in patient
ed, protein absent in patient
nd plasma membrane, no
ed, protein absent in patient
riphery copper response
alization, copper response Impaired glycosylation
riphery
ed, protein absent in patient
copper responseed, protein absent in patient

ARTICLE IN PRESSG ModelJTEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Elements in Medicine and Biology xxx (2014) xxx–xxx 9
F ation
[
ptithpitaitbaclsstCbbpsWcc[tnWdtm
ig. 5. Proposed flowchart to analyze MD causing mutations and WD causing mut166].
resent in their amino acid sequences. The difference in the direc-ionality of copper induced relocalization of ATP7A and ATP7Bllustrates why two distinct, but very similar, copper export pro-eins are required to ensure proper copper uptake and excretion inigher organisms. In addition, this difference, resulting in distincthysiological functions of these homologous proteins in maintain-
ng whole-body copper homeostasis, explains the opposed copperransport defects seen in MD and WD [78]. Several MD-causingnd WD-causing mutations are associated with defects in copper-nduced relocalization of ATP7A and ATP7B respectively, suggestinghat this is a key event that precedes cellular copper excretiony ATP7A and ATP7B.Therefore, such defects might possibly bessociated with the pathogenesis of MD and WD. In general, MD-ausing and WD causing mutations can result in three types ofocalization defects (Table 2, Fig. 5). The first type displays a normalteady-state localization of the protein within the TGN, but respon-iveness to copper is lost [148,163,164]. One mutation of interesthat has been shown to result in such a defect is the MD-causing1000R mutation in ATP7A, which disrupts the putative copper-inding CPC motif, suggesting that in this case impaired copperinding causes the lack of copper-induced trafficking to the celleriphery. This type of localization defect could potentially explainome of the biochemical symptoms seen in patients with MD or
D. For example although the G943S mutation in ATP7B prohibitsopper induced trafficking of ATP7B, this mutation still permitsuproenzyme biosynthesis in the �Ccc2 complementation assay148,165]. Although this correlation does not exist for all muta-ions causing this type of localization defect, it could explain theormal caeruloplasmin production found in some patients with
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
D. Unfortunately, serum caeruloplasmin levels have only beenescribed for one patient who was compound heterozygous forhe G943S mutation, and this was indeed found to be within nor-
al range [148]. Although this type of localization defect might
in ATP7A and ATP7B and to correlate the molecular defects to disease phenotype
still permit cuproenzyme biosynthesis in MD, resulting in allevi-ation of the clinical phenotype, this might not always be obviousin patients, owing to an impairment in copper translocation acrossthe mucosal barrier. More likely, this type of localization defectcould allow for the beneficial effects of copper replacement ther-apy in these patients. The second type of localization defects ison the opposite side of the spectrum. Several MD-causing andWD causing mutations result in constitutive peripheral localiza-tion of ATP7A or ATP7B, respectively [147,163]. Of interest is theMD causing L873R mutation in ATP7A, targeting a residue locatedadjacent to the conserved TGE motif. In this case, a constitu-tively peripheral localization of ATP7A as a result of this mutationhas been associated with hyper phosphorylation of ATP7A, sug-gesting that copper-induced relocalization is dependent on theATPase catalytic cycle. The third type of localization defect isprobably the most common and clinically important. MD-causingand WD-causing mutations result in mislocalization, or possiblyretention, of ATP7A and ATP7B within the endoplasmic reticulum[145,147,148,162,164,166]. ER mislocalization of proteins is oftendue to misfolding and associated with proteasomal degradation.A well-known example of this process in human disease develop-ment is the ER-associated degradation of the �F508 CFTR mutantin cystic fibrosis. In analogy, the H1069Q mutation in ATP7B indeedresults in an increased proteolysis rate. Whether the increased pro-teolysis resulting from the H1069Q mutation is due to defects in theconformation of ATP7B remains unclear, but the solution structureof the N domain of ATP7B containing the H1069Q mutation showsno folding defects.
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
Conclusion
The present information in the arena of metallo-biochemistryof oxidative tension shows that metal-induced/enhanced

ING ModelJ
1 s in M
fonmdpmaemba
C
A
R
ARTICLETEMB-25533; No. of Pages 13
0 A. Ahuja et al. / Journal of Trace Element
oundation of free radicals and additional reactive species can bebserved as a mutual issue that is responsible for metal-inducedoxiousness and carcinogenicity. Concluded ROS-mediated retorts,ost powerful action of metal that they causes “unplanned” DNA
amage followed by bio-protein modification and in the final lipideroxidation. Metal persuaded development of free radicals hasost significantly been showed aimed at copper. As sketched
bove, metal-induced oxidative tension is connected through sev-ral diseases and consequence partially from decayed antioxidantechanisms. Thus design of dual operative antioxidants, having
oth metal-chelating and ROS/RNS-scavenging activity should beimed.
onflict of interest
The authors have no conflicts of interest to report.
cknowledgment
This work was supported by the NIMS University, Jaipur.
eferences
[1] Angelova M, Asenova S, Nedkova V, Koleva KR. Copper in the human organism.Trakia J Sci 2011;9(1):88–98.
[2] Madsen E, Gitlin JD. Copper and iron disorders of the brain. Annu Rev Neurosci2007;30:317–37.
[3] Svetlana L, Ashima B, Ann LH. Copper handling machinery of the brain. Met-allomics 2010;2:596.
[4] Lech T, Sadlik JK. Copper concentration in body tissues and fluids in normalsubjects of southern Poland. Biol Trace Elem Res 2007;118(July (1)):10–5.
[5] Gybina AA, Tkac I, Prohaska JR. Copper deficiency alters the neurochemicalprofile of developing rat brain. Nutr Neurosci 2009;12(June (3)):114–22.
[6] Nischwitz V, Achim B, Bernhard M. Speciation analysis of selected metals anddetermination of their total contents in paired serum and cerebrospinal fluidsamples: an approach to investigate the permeability of the human blood-cerebrospinal fluid-barrier. Anal Chim Acta 2008;627:258–69.
[7] Basun H, Forssell LG, Wetterberg L, Winblad B. Metals and trace elementsin plasma and cerebrospinal fluid in normal aging and Alzheimer’s disease. JNeural Transm Park Dis Dement Sect 1991;3:231–58.
[8] Hershey CO, Hershey LA, Varnes A, Vibhakar SD, Lavin P, Strain WH. Cere-brospinal fluid trace element content in dementia: clinical, radiologic, andpathologic correlations. Neurology 1983;33(October (10)):1350–3.
[9] Hodgson EK, Fridovich I. The interaction of bovine erythrocyte superoxidedismutase with hydrogen peroxide: inactivation of the enzyme. Biochemistry1975;14(December (24)):5294–9.
[10] Blech DM, Borders Jr CL. Hydroperoxide anion, HO-2, is an affinity reagent forthe inactivation of yeast Cu, Zn superoxide dismutase: modification of onehistidine per subunit. Arch Biochem Biophys 1983;224(July (2)):579–86.
[11] Sinet PM, Garber P. Inactivation of the human Cu–Zn superoxide dismutaseduring exposure to O2 and H2O2. Arch Biochem Biophys 1981;212:411–6.
[12] Borders Jr CL, Fridovich I. A comparison of the effects of cyanide, hydrogenperoxide, and phenylglyoxal on eucaryotic and procaryotic Cu, Zn superoxidedismutases. Arch Biochem Biophys 1985;241(September (2)):472–6.
[13] Yim MB, Chock PB, Stadtman ER. Enzyme function of copper, zinc superox-ide dismutase as a free radical generator. J Biol Chem 1993;268(February(6)):4099–105.
[14] Harris E. Trace elements in health: a review of current issues. Woburn, MA,USA: Butterworthand Co. Publishers Ltd.; 1983.
[15] Strausak D, Mercer JFB, Dieter HH, Stremmel W, Multhaup G. Copper in disor-ders with neurological symptoms: Alzheimer’s, Menkes, and Wilson diseases.Brain Res Bull 2001;55:175–85.
[16] Slivarichová D, Mitrová E, Ursínyová M, Uhnáková I, Koscová S, Wsólová L.Geographic accumulation of Creutzfeldt–Jakob disease in Slovakia – envi-ronmental metal imbalance as a possible cofactor. Cent Eur J Publ Health2011;19(September (3)):158–64.
[17] Mitteregger G, Korte S, Shakarami M, Herms J, Kretzschmar HA. Role of copperand manganese in prion disease progression. Brain Res 2009;1292:155–64.
[18] Hodak M, Chisnell R, Lu W, Bernholc J. Functional implications of mul-tistage copper binding to the prion protein. Proc Natl Acad Sci USA2009;106:11576–81.
[19] Desai V, Kaler SG. Role of copper in human neurological disorders. Am J Clin
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
Nutr 2008;88(September (3)):855S–8S.[20] Viles JH, Klewpatinond M, Nadal RC. Copper and the structural biology of the
prion protein. Biochem Soc Trans 2008;36(Pt 6):1288–92.[21] Varela-Nallar L, González A, Inestrosa NC. Role of copper in prion diseases:
deleterious or beneficial? Curr Pharm Des 2006;12(20):2587–95.
PRESSedicine and Biology xxx (2014) xxx–xxx
[22] Quaglio E, Chiesa R, Harris DA. Copper converts the cellular prion protein intoa protease-resistant species that is distinct from the scrapie isoform. J BiolChem 2001;276:11432.
[23] Roger CP, Diane EG. Prions are copper-binding proteins. Trends Biochem Sci1998;23(6):197–8.
[24] Yamamoto N, Kuwata k. Difference in redox behaviors between copper-binding octarepeat and nonoctarepeat sites in prion protein. J Biol Inorg Chem2009;14(November (8)):1209–18.
[25] Linder MC, Hazegh-Azam M. Copper biochemistry and molecular biology. AmJ Clin Nutr 1996;63(May (5)):797S–811S.
[26] Zhou B, Gitschier J. hCTR1: a human gene for copper uptake identified bycomplementation in yeast. Proc Natl Acad Sci USA 1997;94:7481–6.
[27] Shim H, Harris ZL. Genetic defects in copper metabolism. J Nutr 2003;133(5(Suppl. 1)):1527S–31S.
[28] Prousek J. Fenton chemistry in biology and medicine. Pure Appl Chem2007;79(12):2325–38.
[29] Liochev SI, Fridovich I. The Haber–Weiss cycle – 70 years later: an alternativeview. Redox Rep 2002;7:55–7.
[30] Speisky H, Gomez M, Burgos-Bravo F, Lopez-Alarcon C, Jullian C, Olea-AzarC, Aliaga ME. Generation of superoxide radicals by copper-glutathione com-plexes: redox-consequences associated with their interaction with reducedglutathione. Bioorg Med Chem 2009;17(5):1803–10.
[31] Aruoma OI, Halliwell B, Gajewski E, Dizdaroglu M. Copper-ion-dependentdamage to the bases in DNA in the presence of hydrogen peroxide. BiochemJ 1991;273(Pt 3):601–4.
[32] Barbusinski K. Fenton reaction – controversy concerning the chemistry. EcolChem Eng 2009;16:347–58.
[33] Prousek J. Fenton reaction after a century. Chem List 1995;89(1):11–21.[34] Moriwaki H, Osborne MR, Phillips DH. Effects of mixing metal ions on oxida-
tive DNA damage mediated by a Fenton-type reduction. Toxicol In Vitro2008;22(February (1)):36–44.
[35] Mattie MD, Freedman JH. Copper-inducible transcription: regulation bymetal- and oxidative stress-responsive pathways. Am J Physiol Cell Physiol2004;286(February (2)):C293–301.
[36] Linder MC. Biochemistry of copper. New York: Plenum Press; 1991.[37] Jomova K, Valko M. Advances in metal-induced oxidative stress and human
disease. Toxicology 2011;283(May (2/3)):65–87.[38] Steinebach OM, Wolterbeek HT. Role of cytosolic copper, metallothionein and
glutathione in copper toxicity in rat hepatoma tissue culture cells. Toxicology1994;92(September (1/3)):75–90.
[39] El Hachimi KH, Chaunu MP, Brown P, Foncin JF. Modifications of oligo-dendroglia cells in spongiform encephalopathies. Exp Neurol 1998;154:23–30.
[40] Zatta P, Cervellin D, Favarato M, Gerotto M, Mattiello G. Microelementalconcentration in the ontogenesis of rat brain. Trace Elem Med 1994;11:143–7.
[41] Kiaei M, Bush AI, Morrison BM, Morrison JH, Cherny RA, Volitakis I, et al.Genetically decreased spinal cord copper concentration prolongs life ina transgenic mouse model of amyotrophic lateral sclerosis. J Neurosci2004;24(September (36)):7945–50.
[42] Kramer DR, Llanos RM, Mercer JFB. Molecular basis of copper transport: cellu-lar and physiological functions of Menkes and Wilson disease proteins (ATP7Aand ATP7B). In: Zatta P, editor. Metal ions and neurodegenerative disorders.Singapore/London: World Scientific; 2003. p. 207–44.
[43] Paolo Z, Frank A. Copper deficiency and neurological disorders in man andanimals. Brain Res Rev 2007;54(April (1)):19–33.
[44] Quian Y, Tiffany-Castiglioni E, Welsh J, Harris ED. Copper efflux from murinemicrovascular cells requires expression of the Menkes disease Cu-ATPase. JNutr 1998;128:1276–82.
[45] Kodama H. Recent developments in Menkes disease. J Inherit Metab Dis1993;16(4):791–9.
[46] Kumar N, Low PA. Myeloneuropathy and anemia due to copper malabsorp-tion. J Neurol 2004;251(June (6)):747–9.
[47] Bukau B, Deuerling E, Pfund C, Craig EA. Getting newly synthesized proteinsinto shape. Cell 2000;101:119–22.
[48] Rosenzweig AC. Copper delivery by metallochaperone protein. Acc Chem Res2001;34(February (2)):119–28.
[49] Appel SH, Zhao W, Beers DR, Henkel JS. The microglial-motoneuron dialoguein ALS. Acta Myol 2011;30:4–8.
[50] Pasinelli P, Brown RH. Molecular biology of amyotrophic lateral sclerosis:insights from genetics. Nat Rev Neurosci 2006;7:710–23.
[51] Czaplinski A, Yen AA, Simpson EP, Appel SH. Slower disease progression andprolonged survival in contemporary patients with amyotrophic lateral scle-rosis: is the natural history of amyotrophic lateral sclerosis changing? ArchNeurol 2006;63:1139–43.
[52] Maruyama H, Morino H, Ito H, Izumi Y, Kato H, Watanabe Y, et al. Mutationsof optineurin in amyotrophic lateral sclerosis. Nature 2010;465:223–6.
[53] Turner MR, Hardiman O, Benatar M, Brooks BR, Chio A, de Carvalho M, et al.Controversies and priorities in amyotrophic lateral sclerosis. Lancet Neurol2013;12:310–22.
[54] Fridovich I. Superoxide dismutases. Annu Rev Biochem 1975;44(1):147–59.
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
[55] Fitzmaurice PS, Shaw IC, Kleiner HE, Miller RT, Monks TJ, Lau SS. Evidence forDNA damage in amyotrophic lateral sclerosis. Muscle Nerve 1996;19:797–8.
[56] Ferrante RJ, Browne SE, Shinobu LA, Bowling AC, Baik MJ, MacGarvey U.Evidence of increased oxidative damage in both sporadic and familial amy-otrophic lateral sclerosis. J Neurochem 1997;69:2064–74.

ING ModelJ
s in M
ARTICLETEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Element
[57] Shibata N, Nagai R, Uchida K, Horiuchi S, Yamada S, Hirano A. Morphologicalevidence for lipid peroxidation and protein glycoxidation in spinal cords fromsporadic amyotrophic lateral sclerosis patients. Brain Res 2001;917:97–104.
[58] Barber SC, Shaw PJ. Oxidative stress in ALS: key role in motor neuron injuryand therapeutic target. Free Radical Biol Med 2010;48(5):629–41.
[59] Barnham KJ, Masters CL, Bush AI. Neurodegenerative diseases and oxidativestress. Nat Rev Drug Discovery 2004;3(3):205–14.
[60] Yim MB, Chock PB, Stadtman ER. Copper, zinc superoxide dismutase cat-alyzes hydroxyl radical production from hydrogen peroxide. Proc Natl AcadSci 1990;87(13):5006–10.
[61] Genova ML, Pich MM, Bernacchia A. The mitochondrial production of reac-tive oxygen species in relation to aging and pathology. Ann N Y Acad Sci2004;1011(1):86–100.
[62] Saxena S, Cabuy E, Caroni PA. Role for motoneuron subtype-selective ER stressin disease manifestations of FALS mice. Nat Neurosci 2009;12(5):627–36.
[63] Schulz JB, Lindenau J, Seyfried J. Glutathione, oxidative stress and neurode-generation. Eur J Biochem 2000;267:4904–11.
[64] Beal MF. Mitochondria and the pathogenesis of ALS. Brain 2000;123:1291–2.[65] Vielhaber S, Kunz D, Winkler K, Wiedemann FR, Kirches E, Feistner H, et al.
Mitochondrial DNA abnormalities in skeletal muscle of patients with sporadicamyotrophic lateral sclerosis. Brain 2000;123:1339–48.
[66] Klivenyi P, Ferrante RJ, Matthews RT, Bogdanov MB, Klein AM, AndreassenOA, et al. Neuroprotective effects of creatine in a transgenic animal model ofamyotrophic lateral sclerosis. Nat Med 1999;5:347–50.
[67] Shaw PJ, Eggett CJ. Molecular factors underlying selective vulnerability ofmotor neurons to neurodegeneration in amyotrophic lateral sclerosis. J Neu-rol 2000;247(Suppl. 1):17–27.
[68] Cleveland DW, Rothstein JD. From Charcot to Lou Gehrig: deciphering selec-tive motor neuron death in ALS. Nat Rev Neurosci 2001;2:806–19.
[69] Rothstein JD, Tsai G, Kuncl RW. Abnormal excitatory amino acid metabolismin amyotrophic lateral sclerosis. Ann Neurol 1990;28:18–25.
[70] Spreux-Varoquaux O, Bensimon G, Lacomblez L, Salachas F, Pradat PF, LeForestier N, et al. Glutamate levels in cerebrospinal fluid in amyotrophiclateral sclerosis: a reappraisal using a new HPLC method with coulometricdetection in a large cohort of patients. J Neurol Sci 2002;193:73–8.
[71] Rothstein JD, Dykes-Hoberg M, Pardo CA, Bristol LA, Jin L, Kuncl RW, et al.Knockout of glutamate transporters reveals a major role for astroglial trans-port in excitotoxicity and clearance of glutamate. Neuron 1996;16:675–86.
[72] Valentine JS, Hart PJ. Misfolded Cu Zn SOD and amyotrophic lateral sclerosis.Proc Natl Acad Sci USA 2003;100(7):3617–22.
[73] Hart PJ. Pathogenic superoxide dismutase structure, folding, aggregation andturnover. Curr Opin Chem Biol 2006;10(April (2)):131–8.
[74] Rivera-Mancía S, Pérez-Neri I, Ríos C, Tristán-López L, Rivera-Espinosa L,Montes S. The transition metals copper and iron in neurodegenerative dis-eases. Chem Biol Interact 2010;186(July (2)):184–99.
[75] Estévez AG, Crow JP, Sampson JB, Reiter C, Zhuang Y, Richardson GJ, et al.Induction of nitric oxide-dependent apoptosis in motor neurons by zinc-deficient superoxide dismutase. Science 1999;286:2498–500.
[76] Levenson CW. Trace metal regulation of neuronal apoptosis: from genes tobehavior. Physiol Behav 2005;86(October (3)):399–406.
[77] Burstein E, Ganesh L, Dick RD, van De Sluis B, Wilkinson JC, Klomp LW, et al.A novel role for XIAP in copper homeostasis through regulation of MURR1.EMBO J 2004;23(January (1)):244–54.
[78] Mufti AR, Burstein E, Csomos RA, Graf PC, Wilkinson JC, Dick RD, et al. XIAPIs a copper binding protein deregulated in Wilson’s disease and other coppertoxicosis disorders. Mol Cell 2006;21(March (6)):775–85.
[79] Pasinelli P, Belford ME, Lennon N, Bacskai BJ, Hyman BT, Trotti D, et al.Amyotrophic lateral sclerosis-associated SOD1 mutant proteins bind andaggregate with Bcl-2 in spinal cord mitochondria. Neuron 2004;43(July(1)):19–30.
[80] Higgins CM, Jung C, Ding H, Xu Z. Mutant Cu–Zn superoxide dismutase thatcauses motoneuron degeneration is present in mitochondria in the CNS. JNeurosci 2002;22(March (6)):RC215.
[81] Okado-Matsumoto A, Fridovich I. Amyotrophic lateral sclerosis: a proposedmechanism. Proc Natl Acad Sci USA 2002;99(June (13)):9010–4.
[82] Glenner GG, Wong CW. Alzheimer’s disease: initial report of the purificationand characterization of a novel cerebrovasculaar amyloid protein. BiochemBiophys Res Commun 1984;120(May (3)):885–90.
[83] Haass C, Schlossmacher MG, Hung AY, Vigo-Pelfrey C, Mellon A, OstaszewskiBL, et al. Amyloid beta-peptide is produced by cultured cells during normalmetabolism. Nature 1992;359(September (6393)):322–5.
[84] Masters CL, Multhaup G, Simms G, Pottgiesser J, Martins RN, Beyreuther K.Neuronal origin of a cerebral amyloid: neurofibrillary tangles of Alzheimer’sdisease contain the same protein as the amyloid of plaque cores and bloodvessels. EMBO J 1985;4(November (11)):2757–60.
[85] Masters CL, Simms G, Weinman NA, Multhaup G, McDonald BL, Beyreuther K.Amyloid plaque core protein in Alzheimer disease and Down syndrome. ProcNatl Acad Sci USA 1985;82:4245–9.
[86] Roher AE, Chaney MO, Kuo YM, Webster SD, Stine WB, Haverkamp LJ, et al.Morphology and toxicity of Abeta-(1-42) dimer derived from neuritic and vas-
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
cularamyloid deposits of Alzheimer’s disease. J Biol Chem 1996;271(August(34)):20631–5.
[87] Roher AE, Palmer KC, Yurewicz EC, Ball MJ, Greenberg BD. Morphologicaland biochemical analyses of amyloid plaque core proteins purified fromAlzheimer disease brain tissue. J Neurochem 1993;61:1916–26.
PRESSedicine and Biology xxx (2014) xxx–xxx 11
[88] Seubert P, Vigo-Pelfrey C, Esch F, Lee M, Dovey H, Davis D, et al. Isolationand quantification of soluble Alzheimer’s beta-peptide from biological fluids.Nature 1992;359:325–7.
[89] Shoji M, Golde TE, Ghiso J, Cheung TT, Estus S, Shaffer LM, et al. Production ofthe Alzheimer amyloid beta protein by normal proteolytic processing. Science1992;258(October (5079)):126–9.
[90] Vigo-Pelfrey C, Lee D, Keim P, Lieberburg I, Schenk DB. Characterizationof beta-amyloid peptide from human cerebrospinal fluid. J Neurochem1993;61:1965–8.
[91] Atwood CS, Perry G, Zeng H, Kato Y, Jones WD, Ling KQ, et al. Coppermediates dityrosine cross-linking of Alzheimer’s amyloid beta. Biochemistry2004;43(January (2)):560–8.
[92] Opazo C, Huang X, Cherny RA, Moir RD, Roher AE, White AR, et al.Metalloenzyme-like activity of Alzheimer’s disease beta-amyloid Cu-dependent catalytic conversion of dopamine, cholesterol, and biologicalreducing agents to neurotoxic H2O2. J Biol Chem 2002;277(October(43)):40302–8.
[93] Atwood CS, Scarpa RC, Huang X, Jones WD, Fairlie DP, Tanzi RE, et al.Characterization of copper interactions with Alzheimer A� peptides iden-tification of an attomole affinity copper binding site on A�1-42. J Neurochem2000;75:1219–33.
[94] White AR, Reyes R, Mercer JF, Camakaris J, Zheng H, Bush AI, et al. Copper levelsare increased in the cerebral cortex and liver of APP and APLP2 knockout mice.Brain Res 1999;842(September (2)):439–44.
[95] Lovell MA, Robertson JD, Teesdale WJ, Campbell JL, Markesbery WR. Copper,iron and zinc in Alzheimer’s disease senile plaques. J Neurol Sci 1998;158(July(1)):47–52.
[96] Vassiliev V, Harris ZL, Zatta P. Ceruloplasmin in neurodegenerative diseases.Brain Res Brain Res Rev 2005;49(November (3)):633–40.
[97] Deibel MA, Ehmann WD, Markesbery WR. Copper, iron, and zinc imbalances inseverely degenerated brain regions in Alzheimer’s disease: possible relationto oxidative stress. J Neurol Sci 1996;143(November (1/2)):137–42.
[98] Schlief ML, Craig AM, Gitlin JD. NMDA receptor activation mediates cop-per homeostasis in hippocampal neurons. J Neurosci 2005;25(January(1)):239–46.
[99] Mawuenyega KG, Sigurdson W, Ovod V, Munsell L, Kasten T, Morris JC, et al.Decreased clearance of CNS beta-amyloid in Alzheimer’s disease. Science2010;330(December (6012)):1774.
[100] Fonteh AN, Harrington RJ, Tsai A, Liao P, Harrington MG. Free amino acidand dipeptide changes in the body fluids from Alzheimer’s disease subjects.Amino Acids 2007;32(February (2)):213–24.
[101] Chung RS, Howells C, Eaton ED, Shabala L, Zovo K, Palumaa P, et al. The nativecopper- and zinc-binding protein metallothionein blocks copper-mediated Abeta aggregation and toxicity in rat cortical neurons. PLoS ONE 2010;5(August(8)):e12030.
[102] Snyder EM, Nong Y, Almeida CG, Paul S, Moran T, Choi EY, et al. Regulationof NMDA receptor trafficking by amyloid-beta. Nat Neurosci 2005;8(August(8)):1051–8.
[103] Menkes JH, Alter M, Steigleder GK, Weakley DR, Sung JH. A sex-linked reces-sive disorder with growth retardation, peculiar hair, and focal cerebral andcerebellar degeneration. Pediatrics 1962;29(May):764–79.
[104] Kaler SG. Menkes disease. Adv Pediatr 1994;41:263–304.[105] Danks DM, Campbell PE, Walker-Smith J, Stevens BJ, Gillespie JM, Blom-
field J, et al. Menkes’ kinky hair syndrome. Lancet 1972;1(May (7760)):1100–2.
[106] Danks DM, Cartwright E, Stevens BJ, Townley RR. Menkes’ kinky hair disease:further definition of the defect in copper transport. Science 1973;179(March(4078)):1140–2.
[107] Berg BO. The neurologic examination. In: Berg BO, editor. Child neurology: aclinical manual. Philadelphia: JB Lippencott; 1994. p. 1–26.
[108] Cox DW, Moore SD. Copper transporting P-type ATPases and human disease.J Bioenerg Biomembr 2002;34(October (5)):333–8.
[109] Rinaldi AC. Meeting report: copper research at the top. Biometals2000;13:9–13.
[110] Kaler SG. Metabolic and molecular bases of Menkes disease and occipital hornsyndrome. Pediatr Dev Pathol 1998;1:85–98.
[111] Liu PC, McAndrew PE, Kaler SG. Rapid and robust screening of the Menkes dis-ease/occipital horn syndrome gene. Genet Test 2002;6(Winter (4)):255–60.
[112] Kaler SG, Buist NRM, Holmes CS, Goldstein DS, Miller RC, Gahl WA. Early cop-per therapy in classic Menkes disease patients with a novel splicing mutation.Ann Neurol 1995;38:921–8.
[113] Kaler SG, Das S, Levinson B, Goldstein DS, Holmes CS, Patronas NJ, et al.Successful early copper therapy in Menkes disease associated with amutant transcript containing a small in-frame deletion. Biochem Mol Med1996;57:37–46.
[114] Kaler SG. Menkes disease mutations and response to early copper histidinetreatment. Nat Genet 1996;13:21–2.
[115] Liu PC, Chen YW, Centano J, Quesado M, Lem KE, Kaler SG. Down regulationof myelination, energy, and translational genes in Menkes disease brain. MolGenet Metab 2005;85:291–300.
[116] Kaler SG, Holmes CS, Goldstein DS, Tang J, Godwin SC, Donsante A,
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
et al. Neonatal diagnosis and treatment of Menkes disease. N Engl J Med2008;358:605–14.
[117] Kaler SG, Gallo LK, Proud VK, Percy AK, Mark Y, Segal NA, et al. Occipital hornsyndrome and a mild Menkes phenotype associated with splice site mutationsat the MNK locus. Nat Genet 1994;8:195–202.

ING ModelJ
1 s in M
ARTICLETEMB-25533; No. of Pages 13
2 A. Ahuja et al. / Journal of Trace Element
[118] Chelly J, Tumer Z, Tonnesen T, Petterson A, Ishikawa-Brush Y, Tommerup N,et al. Isolation of a candidate gene for Menkes disease that encodes a potentialheavy metal binding protein. Nat Genet 1993;3:14–9.
[119] Mercer JF, Livingston J, Hall B, Paynter JA, Begy C, Chandrasekharappa S, et al.Isolation of a partial candidate gene for Menkes disease by positional cloning.Nat Genet 1993;3:20–5.
[120] Vulpe C, Levinson B, Whitney S, Packman S, Gitschier J. Isolation of a candidategene for Menkes disease and evidence that it encodes a copper transportingATPase. Nat Genet 1993;3:7–13.
[121] Arnesano F, Banci L, Bertini I, Ciofi-Baffoni S, Molteni E, Huffman DL, et al.Metallochaperones and metal-transporting ATPases: a comparative analysisof sequences and structures. Genome Res 2002;12(February (2)):255–71.
[122] Tumer Z, Moller LB, Horn N. Mutation spectrum of ATP7A, the gene defectivein Menkes disease. Adv Exp Med Biol 1999;448:83–95.
[123] Hsi G, Cox DW. A comparison of the mutation spectra of Menkes disease andWilson disease. Human Genet 2004;114:165–72.
[124] Govindaraju M, Shekar HS. Role of copper in the clinics of neurological disor-ders. IJRPC 2011;1(3).
[125] Culotta VC, Gitlin JD. The metabolic molecular basis of inherited disease. 8thed. New York, NY: McGraw-Hill Companies; 2001. p. 3115–8.
[126] Suzuki N, Cheung TT, Cai XD, Odaka A, Otvos Jr L, Eckman C, et al. Anincreased percentage of long amyloid betaprotein secreted by familial amy-loid beta protein precursor (beta APP717) mutants. Science 1994;264(May(5163)):1336–40.
[127] Claire JS (Ph.D. thesis) The copper-amyloid-beta-peptide complex ofAlzheimer’s disease: affinity, structure, fibril formation and toxicity. Univer-sity of London; 2010.
[128] Lesne S, Ali C, Gabriel C, Croci N, MacKenzie ET, Glabe CG, et al. NMDArecep-tor activation inhibits alpha-secretase and promotes neuronal amyloid betaproduction. J Neurosci 2005;25:9367–77.
[129] Meloni G, Sonois V, Delaine T, Guilloreau L, Gillet A, Teissié J, et al. Metalswap between Zn7-metallothionein-3 andamyloid-beta-Cu protects againstamyloid-beta toxicity. Nat Chem Biol 2008;4(June (6)):366–72.
[130] Kohen R, Yamamoto Y, Cundy KC, Ames BN. Antioxidant activity of carnosine,homocarnosine, and anserine present in muscle and brain. Proc Natl Acad SciUSA 1988;85(May (9)):3175–9.
[131] Horning MS, Blakemore LJ, Trombley PQ. Endogenous mechanisms of neuro-protection: role of zinc, copper, and carnosine. Brain Res 2000;852(January(1)):56–61.
[132] Lanza V, Bellia F, D’Agata R, Grasso G, Rizzarelli E, Vecchio G. New glycosidederivatives of carnosine and analogs resistant to carnosinase hydrolysis: syn-thesis and characterization of their copper(II) complexes. J Inorg Biochem2011;105(February (2)):181–8.
[133] Urbanc B, Cruz L, Le R, Sanders J, Ashe KH, Duff K, et al. Neurotoxic effectsof thioflavin positive amyloid deposits in transgenic mice and Alzheimer’sdisease. Proc Natl Acad Sci USA 2002;99(October (22)):13990–5.
[134] Meyer-Luehmann M, Spires-Jones TL, Prada C, Garcia-Alloza M, de CalignonA, Rozkalne A, et al. Rapid appearance and local toxicity of amyloid-betaplaques in a mouse model of Alzheimer’s disease. Nature 2008;451(February(7179)):720–4.
[135] Horn N, Tonnesen T, Tulmer Z. Menkes disease: an X-linked neurologicaldisorder of the copper metabolism. Brain Pathol 1992;2:351–62.
[136] Danks DM. Disorders of copper transport. In: Scriver JR, Beaudet AL, Sly WS,Valle D, editors. The metabolic basis of inherited disease. New York: McGraw-Hill; 1995. p. 2211–35.
[137] Keydorn K, Damsgaard E, Horn N, Mikkelsen M, Tygstrup I, Vestemark S, et al.Extra-hepatic storage of copper: a male foetus suspected of Menkes’ disease.Humangenetik 1975;29(2):171–5.
[138] Horn N, Heydorn K, Damsgaard E, Tygstrup I, Vestermark S. Is Menkessyndrome a copper storage disorder? Clin Genet 1978;14(September (3)):186–7.
[139] Lutsenko S, LeShane ES, Shinde U. Biochemical basis of regulation ofhuman copper-transporting ATPases. Arch Biochem Biophys 2007;463(July(2)):134–48.
[140] Palmgren MG, Axelsen KB. Evolution of P-type ATPases. Biochim Biophys Acta1998;1365(June (1/2)):37–45.
[141] Tumer Z, Møller LB. Menkes disease. Eur J Human Genet 2010;18:511–8.[142] Yamaguchi Y, Heiny ME, Suzuki M, Gitlin JD. Biochemical characterization
and intracellular localization of the Menkes disease protein. Proc Natl AcadSci USA 1996;93:14030–5.
[143] Hung IH, Suzuki M, Yamaguchi Y, Yuan DS, Klausner RD, Gitlin JD. Biochemicalcharacterization of the Wilson disease protein and functional expression inthe yeast Saccharomyces cerevisiae. J Biol Chem 1997;272:21461–6.
[144] Petris MJ, Mercer JF, Culvenor JG, Lockhart P, Gleeson PA, Camakaris J. Ligand-regulated transport of the Menkes copper P-type ATPase efflux pump from theGolgi apparatus to the plasma membrane: a novel mechanism of regulatedtrafficking. EMBO J 1996:156084–95.
[145] Harada M, Sakisaka S, Terada K, Kimura R, Kawaguchi T, Koga H, et al. Role ofATP7B in biliary copper excretion in a human hepatoma cell line and normalrat hepatocytes. Gastroenterology 2000;118:921–8.
[146] Harada M, Kawaguchi T, Kumemura H, Terada K, Ninomiya H, Taniguchi E,
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
et al. The Wilson disease protein ATP7B resides in the late endosomes withRab7 and the Niemann-Pick C1 protein. Am J Pathol 2005;166:499–510.
[147] Huster D, Hoppert M, Lutsenko S, Zinke J, Lehmann C, Mossner J, et al. Defec-tive cellular localization of mutant ATP7B in Wilson’s disease patients andhepatoma cell lines. Gastroenterology 2003;124:335–45.
PRESSedicine and Biology xxx (2014) xxx–xxx
[148] Forbes JR, Cox DW. Copper-dependent trafficking of Wilson disease mutantATP7B proteins. Hum Mol Genet 2000;9:1927–35.
[149] Roelofsen H, Wolters H, Van Luyn MJ, Miura N, Kuipers F, Vonk RJ. Copperinduced apical trafficking of ATP7B in polarized hepatoma cells provides amechanism for biliary copper excretion. Gastroenterology 2000;119:782–93.
[150] Schaefer M, Roelofsen H, Wolters H, Hofmann WJ, Muller M, Kuipers F, et al.Localization of the Wilson’s disease protein in human liver. Gastroenterology1999;117:1380–5.
[151] Schaefer M, Hopkins RG, Failla ML, Gitlin JD. Hepatocyte-specific localizationand copper-dependent trafficking of the Wilson’s disease protein in the liver.Am J Physiol 1999;276:G639–46.
[152] Mercer JF, Barnes N, Stevenson J, Strausak D, Llanos RM. Copper-inducedtrafficking of the Cu-ATPases: a key mechanism for copper homeostasis.Biometals 2003;16:175–84.
[153] La Fontaine S, Theophilos MB, Firth SD, Gould R, Parton RG, Mercer JF. Effectof the toxic milk mutation (tx) on the function and intracellular localizationof Wnd, the murine homologue of the Wilson copper ATPase. Hum Mol Genet2001;10:361–70.
[154] Guo Y, Nyasae L, Braiterman LT, Hubbard AL. NH2-terminal signals in ATP7BCu-ATPase mediate its Cu-dependent anterograde traffic in polarized hepaticcells. Am J Physiol Gastrointest Liver Physiol 2005;289:G904–16.
[155] Lutsenko S, Cooper MJ. Localization of the Wilson’s disease protein productto mitochondria. Proc Natl Acad Sci USA 1998;95:6004–9.
[156] Petris MJ, Mercer JF. The Menkes protein (ATP7A; MNK) cycles via the plasmamembrane both in basal and elevated extracellular copper using a C-terminaldi-leucine endocytic signal. Hum Mol Genet 1999;8:2107–15.
[157] Nyasae L, Bustos R, Braiterman L, Eipper B, Hubbard A. Dynamics of endoge-nous ATP7A (Menkes protein) in intestinal epithelial cells: copperdependentredistribution between two intracellular sites. Am J Physiol Gastrointest LiverPhysiol 2007;292:G1181–94.
[158] Greenough M, Pase L, Voskoboinik I, Petris MJ, O’Brien AW, Camakaris J. Sig-nals regulating trafficking of Menkes (MNK; ATP7A) copper-translocatingPtype ATPase in polarized MDCK cells. Am J Physiol Cell Physiol2004;287:C1463–71.
[159] Monty JF, Llanos RM, Mercer JF, Kramer DR. Copper exposure induces traffick-ing of the Menkes protein in intestinal epithelium of ATP7A transgenic mice.J Nutr 2005;135:2762–6.
[160] Ravia JJ, Stephen RM, Ghishan FK, Collins JF. Menkes copper ATPase (Atp7a) isa novel metal-responsive gene in rat duodenum, and immunoreactive proteinis present on brush-border and basolateral membrane domains. J Biol Chem2005;280:36221–7.
[161] Ke BX, Llanos RM, Wright M, Deal Y, Mercer JF. Alteration of copper physiologyin mice overexpressing the human Menkes protein ATP7A. Am J Physiol RegulIntegr Comp Physiol 2006;290:R1460–7.
[162] Cater MA, La Fontaine S, Shield K, Deal Y, Mercer JF. ATP7B mediates vesicularsequestration of copper: insight into biliary copper excretion. Gastroenterol-ogy 2006;130:493–506.
[163] Ambrosini L, Mercer JF. Defective copper-induced trafficking and localiza-tion of the Menkes protein in patients with mild and copper-treated classicalMenkes disease. Hum Mol Genet 1999;8:1547–55.
[164] Kim BE, Smith K, Petris MJ. A copper treatable Menkes disease mutation asso-ciated with defective trafficking of a functional Menkes copper ATPase. J MedGenet 2003;40:290–5.
[165] Forbes JR, Cox DW. Functional characterization of missense mutations inATP7B: Wilson disease mutation or normal variant? Am J Hum Genet1998;63:1663–74.
[166] de Bie P, Muller P, Wijmenga CL, Klomp WJ. Molecular pathogenesis of Wilsonand Menkes disease: correlation of mutations with molecular defects anddisease phenotypes. J Med Genet 2007;44:673–88.
[167] Bruijn LI, Becher MW, Lee MK, Anderson KL, Jenkins NA, Copeland NG, et al.ALS-linked SOD1 mutant G85R mediates damage to astrocytes and pro-motes rapidly progressive disease with SOD1-containing inclusions. Neuron1997;18:327–38.
[168] Howland DS, Liu J, She Y, Goad B, Maragakis NJ, Kim B, et al. Focal lossof the glutamate transporter EAAT2 in a transgenic rat model of SOD1mutant mediated amyotrophic lateral sclerosis (ALS). Proc Natl Acad Sci USA2002;99:1604–9.
[169] Trotti D, Rolfs A, Danbolt NC, Brown Jr RH, Hediger MA. SOD1 mutants linkedto amyotrophic lateral sclerosis selectively inactivate a glial glutamate trans-porter. Nat Neurosci 1999;2:427–33.
[170] Guo H, Lai L, Butchbach ME, Stockinger MP, Shan X, Bishop GA, et al. Increasedexpression of the glial glutamate transporter EAAT2 modulates excitotoxicityand delays the onset but not the outcome of ALS in mice. Hum Mol Genet2003;12:2519–32.
[171] Rothstein JD, Patel S, Regan MR, Haenggeli C, Huang YH, Bergles DE, et al. Beta-lactam antibiotics offer neuroprotection by increasing glutamate transporterexpression. Nature 2005;433:73–7.
[172] Rothstein JD, Van Kammen M, Levey AI, Martin LJ, Kuncl RW. Selective loss ofglial glutamate transporter GLT-1 in amyotrophic lateral sclerosis. Ann Neurol1995;38:73–84.
[173] Xu Z, Cork LC, Griffin JW, Cleveland DW. Increased expression of neurofil-
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
ament subunit NF-L produces morphological alterations that resemble thepathology of human motor neuron disease. Cell 1993;73:23–33.
[174] Lee MK, Marszalek JR, Cleveland DW. A mutant neurofilament subunit causesmassive, selective motor neuron death: implications for the pathogenesis ofhuman motor neuron disease. Neuron 1994;13:975–88.

ING ModelJ
s in M
[
[
[177] Williamson TL, Cleveland DW. Slowing of axonal transport is a very earlyevent in the toxicity of ALS-linked SOD1 mutants to motor neurons. Nat
ARTICLETEMB-25533; No. of Pages 13
A. Ahuja et al. / Journal of Trace Element
175] Couillard-Després S, Zhu Q, Wong PC, Price DL, Cleveland DW, Julien JP. Pro-tective effect of neurofilament heavy gene overexpression in motor neuron
Please cite this article in press as: Ahuja A, et al. Copper mediated nAlzheimer and Menkes disease. J Trace Elem Med Biol (2014), http://d
disease induced by mutant superoxide dismutase. Proc Natl Acad Sci USA1998;95:9626–30.
176] Nguyen MD, Lariviere RC, Julien JP. Deregulation of Cdk5 in a mouse modelof ALS: toxicity alleviated by perikaryal neurofilament inclusions. Neuron2001;30:135–47.
PRESSedicine and Biology xxx (2014) xxx–xxx 13
eurological disorder: Visions into amyotrophic lateral sclerosis,x.doi.org/10.1016/j.jtemb.2014.05.003
Neurosci 1999;2:50–6.[178] Puls I, Jonnakuty C, LaMonte BH, Holzbaur EL, Tokito M, Mann E, et al. Mutant
dynactin in motor neuron disease. Nat Genet 2003;33:455–6.




















