
Constraints on disordered locomotion A dynamical systems perspective on spastic cerebral palsy
26
ELSFWIER HUMAN MIMMENT StlENlCE Human Movement Science 15 (1996) 177-202 Constraints on disordered locomotion A dynamical systems perspective on spastic cerebral palsy Kemeth G. Holt a*b’ *, John P. Obusek “c, Sergio T. Fonseca a a Applied Kinesio1og.v Program, Sargent College of Allied Health Professions, Department of Physical Therapy, Boston University, 635 Commonwealth Ace., Boston, MA 02215. USA h Center for the Ecological Study of Perception and Action, Unioersiq of Connecticut, Storrs. CT, USA ’ Army Medical Specialist Corps, U.S. Army. Natick. MA, USA Abstract Upper motor neuron disorders @MN) result in abnormal movement patterns that are due to spasticity, weakness, cocontraction of muscles, inappropriate timing of muscle activations in relation to maximal mechanical advantage, and changes in the mechanical properties of muscles and connective tissues. Since many of the patterns observed in gait are reminiscent of immature patterns in non-disabled toddlers, prolonged abnormal patterns are often viewed as a failure of normal neural maturation. Since the 1960’s the emphasis of therapeutic interventions has been to ‘normalize’ the movement pattern through a variety of neurotherapeutic techniques. The efforts have met with limited success. Recently there have been a number of publications that have emphasized the possibility that the observed patterns may develop or be functional adaptations to a neural system that is not ‘normal’, and that the observed movement patterns in pathological populations may be normal and optimal for that system (Fetters, 1991; Holt, 1993; Latash and Anson, in press; Winter et al., 1990). Some authors have emphasized that alternative patterns develop and are facilitated by redundancy in the motor system (Latash and Anson, in press). Our emphasis has been to pursue the notion that pattern development and functional adaptations in locomotion are driven by the underlying dynamics of the task and the dynamic resources available to the individual. We use a model developed within the dynamical systems framework, the force-driven, pendulum-spring model of locomotion, to parse out the energy form that are needed for continued oscillations, and relate the energy forms to the dynamics that are available to a * Corresponding author. E-mail: holt@sarvxl .bu.edu. 0167-9457/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved SSDf 0167.9457(95)00043-7
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Constraints on disordered locomotion A dynamical systems perspective on spastic cerebral palsy

ELSFWIER
HUMAN MIMMENT StlENlCE
Human Movement Science 15 (1996) 177-202
Constraints on disordered locomotion A dynamical systems perspective on spastic
cerebral palsy
Kemeth G. Holt a*b’ * , John P. Obusek “c, Sergio T. Fonseca a
a Applied Kinesio1og.v Program, Sargent College of Allied Health Professions, Department of Physical
Therapy, Boston University, 635 Commonwealth Ace., Boston, MA 02215. USA
h Center for the Ecological Study of Perception and Action, Unioersiq of Connecticut, Storrs. CT, USA
’ Army Medical Specialist Corps, U.S. Army. Natick. MA, USA
Abstract
Upper motor neuron disorders @MN) result in abnormal movement patterns that are due to spasticity, weakness, cocontraction of muscles, inappropriate timing of muscle activations in relation to maximal mechanical advantage, and changes in the mechanical properties of muscles and connective tissues. Since many of the patterns observed in gait are reminiscent of immature
patterns in non-disabled toddlers, prolonged abnormal patterns are often viewed as a failure of normal neural maturation. Since the 1960’s the emphasis of therapeutic interventions has been to ‘normalize’ the movement pattern through a variety of neurotherapeutic techniques. The efforts
have met with limited success. Recently there have been a number of publications that have emphasized the possibility that the observed patterns may develop or be functional adaptations to a neural system that is not ‘normal’, and that the observed movement patterns in pathological
populations may be normal and optimal for that system (Fetters, 1991; Holt, 1993; Latash and Anson, in press; Winter et al., 1990). Some authors have emphasized that alternative patterns develop and are facilitated by redundancy in the motor system (Latash and Anson, in press). Our emphasis has been to pursue the notion that pattern development and functional adaptations in locomotion are driven by the underlying dynamics of the task and the dynamic resources available to the individual. We use a model developed within the dynamical systems framework, the
force-driven, pendulum-spring model of locomotion, to parse out the energy form that are needed for continued oscillations, and relate the energy forms to the dynamics that are available to a
* Corresponding author. E-mail: holt@sarvxl .bu.edu.
0167-9457/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved
SSDf 0167.9457(95)00043-7

178 K.G. Ho/r rr cd. / Humus Mowment Srienw 1.5 (19%) 177-202
disordered system, specifically, spastic cerebral palsy (CP). We then attempt to relate the available
dynamics to the observed gait patterns. In this way. w’e seek to understand ordered and disordered gaits as alternative functional solutions to the dynamical requirements for continued cycling of a
periodically forced oscillator. The implications of the theoretical model and experimental fIndings are discussed with respect to therapeutic intervention. and the dynamical systems approach.
PsyclNFO class~jication: 2330; 3290; 3291
Ke.word.st Cerebral palsy; Locomotion; Oscillatory models
1. Introduction
Locomotion serves a critical function in allowing animals and humans to explore their environment in search of food, shelter, and a mate. It may also be viewed as a means of dissipating energy in the capture-degrade-dissipate cycle required of thermodynamic, dissipative systems (Morowitz, 1978). For disabled individuals limitations in locomotor ability result not only in mobility and societal limitations, but also in entropic processes that exhibit themselves as pathophysiologies that include decreased cardiac function, musculoskeletal dis- orders, and other long-term medical complications. Constraints on functional locomotion may be multi-factorial - an individual may be limited by fatigue due to high metabolic cost, by instabilities that may lead to falls or fear of falling, by local muscular force and range of motion limitations, by pain, and/or by insufficient or inappropriate neurological control. The coordinated patterns of gait that do emerge may look quite different from those observed in the non-disabled. In previous papers we have investigated how metabolic, shock absorption, and stability constraints influence gait parameters. Our emphasis in this theoretical and data based paper is on the effects of limited muscle forcing in the gastrocnemius-soleus group, that has been observed in spastic cerebral palsy (CP), on gait. We hypothesize that the dynamic resources available to the disordered system, including cocontraction and changes in the mechanical properties of tissues, are utilized in the development of an optimal gait pattern in the absence of normal muscle forcing.
The cornerstone of the dynamical systems approach to understanding control and coordination is the inherent rhythmicity of biological processes (Turvey, 1990). It is claimed that thorough investigation of the physical properties of cooperating oscillatory processes will reveal the self-organizing capability for coordinated movement. The inherent rhythmicity of locomotion lends itself to dynamical description and prediction. We begin by reviewing the oscillatory

K.G. Halt er al. / Human Movement Science 15 (1996) 177-202 179
models that have been used to predict some of the gait characteristics of normal
and disordered walking, and the relationship of the models to gait patterns that minimize metabolic costs and maximize head stability. This overview will serve as a preface for our recent investigations that have sought to parse out the dynamic components of the models that contribute to the continued oscillations necessary for locomotion. The notion that it is the availability (or lack thereof) of the individual dynamic components that determine the observed kinematics of gait patterns is explored. Specifically, we will hypothesize the dynamic and kinematic consequences of constraints imposed on an oscillating system by inability of the muscles to produce sufficient force to maintain oscillations of the limb. We use empirical evidence on the gait characteristics of children with cerebral palsy to support the hypotheses. Implications of this work for rehabilita- tion of disabled populations are discussed.
2. The hybrid (pendulum and spring) harmonic oscillator
In our previous studies on locomotion. the leg has been modeled as a simple, frictionless pendulum with the axis at the hip joint while the composite stiffness of the muscles and soft tissues were represented by a global spring of variable stiffness (Holt et al., 1990a, b, 1991a, Holt et al., in press; Kugler and Turvey, 1987; Turvey et al., 1988). Energy for oscillation is conserved through the leg acting as a pendulum under the influence of gravity (the ‘gravitational compo- nent’), and through the muscle and soft tissues with their ‘elastic component’ acting as a spring. The major simplifying assumptions were: (1) in walking the energy contributions of the elastic and gravitational components of the leg pendulum are assumed to be equal; (2) preferred gait period is assumed to be at the resonant period of the hybrid model.
Full derivation of the hybrid model is given in Appendix A. A schematic representation of the hybrid mass-spring pendulum model may be found in Fig. 1. The multi-segmented lower extremity, a compound pendulum, is represented as a simple pendulum (Holt et al., 1990a, b, 1991a; Kugler and Turvey, 1987) consisting of rigidly linked thigh, shank and foot segments. The stiffness of the attached spring (k) represents the composite stiffness of all the passive soft tissues of the leg together with the active muscle tension. The distance from the axis of motion to the attachment of the composite spring on the lower extremity is denoted by 6. The total mass of the leg is given by m and it is defined as the sum of the leg mass, estimated from the total body mass using anthropometric data (Dempster given in Winter, 1990). The simple pendulum equivalent length

180 K.G. Holtetal./HumanMo~~emer~tScirncr 15 (1996) 177-202
Fig. I, Schematic representation of a hybrid mass-spring pendulum model, where m = mass of the pendulum,
L, = simple pendulum equivalent length, h = distance from the axis of rotation to the spring attachment, and
k = composite global stiffness.
(L,) is the distance from the axis of motion, in this case the hip joint, to the point on the leg pendulum at which the entire mass exerts its effect.
If the oscillatory angle is small, the motion of the hybrid mass-spring pendulum is given by the following Newtonian equation of motion:
mL2,cj = mL,g+ i- kb2$, (1)
where C)J is the angular displacement of the leg pendulum from the vertical and 4 is its angular acceleration. rnL:$ is the moment of inertia of the leg, mL, ~C#J
is the restoring moment due to gravity, and kb*+ is the restoring moment due to stretch on soft tissues.
The natural period is derived as:
T=2T/,,,‘i,,ll’. (2)
‘In walking at the natural period the inertial and elastic forces are assumed equal (Kugler and Turvey, 1987; Walsh, 19921, thus:
kb2 - = 1. m&g
(3)
Inaced system the resonant period is identical to the natural period and is predictable from Eq. (2).

K.G. Holt et al./Human Mocement Science 15 (1996) 177-202 181
By substitution of Eq. (3) into Eq. (21, the equation used to predict preferred
walking period becomes:
The model is a minimal representation of the gait cycle in that it does not take into account the stance phase of gait that may be represented as an inverted pendulum (Mochon and McMahon, 1980). It does not account for the coriolis and other forces that influence the motion of a compound pendulum. Nor does it account for muscle forces necessary to overcome the frictional losses that are due to viscous damping in soft tissues and energy transfers between the stance leg (an inverted pendulum) and the swing leg (the modeled pendulum). Never- theless, this limited model of the gait cycle has proven robust in its ability to predict the preferred walking period of quadrupeds of varying leg lengths (Kugler and Turvey, 1987; Turvey et al., 19881, human adults (Halt et al., 1990a) and children (Holt et al., 1991b). Recent application of the model has shown that the natural frequency of the arms may be important in determining preferred walking frequency at lower speeds in some subjects (Wagenaar and Van Emmerik, 1994). Furthermore, the hybrid model has been shown to predict the walking period at which metabolic costs, variability in the head trajectory, and shock transfer from the feet to the head, are minimized (Halt et al., 199 1 a, b, Holt et al., in press; Ratcliffe and Holt, in press). The predicted frequency
16, , 0.09
0610.04 60 80 100 120 140
Percent of Predicted Frequency
Fig. 2. Metabolic cost and variability of head trajectory as a function of the walking frequency at a fixed
treadmill speed. Predicted frequency is calculated from Eq. (4) in text. Skewed curves suggest that the optimal
frequency is constrained by both variables (adapted from Holt et al., in press).

182 K.G. Halt et al. /Human Momment Science 15 (19%) 177-202
apparently emerges from two complementary constraints, metabolic cost and head stability (see Fig. 2 and legend for details).
Our interpretation for these findings has been that the criteria (metabolic cost, instability) that are minimized in gait serve as constraints on the adopted frequency. The mechanism for minimization of metabolic cost is best under- stood through the forcing in the hybrid model (Halt, 1993). In walking the oscillations would eventually be damped out by the viscous properties of the soft tissues. In a forced system, resonance is defined as the frequency at which the driving force required to maintain oscillations at a fixed amplitude is minimized. In a walking human the driving force is produced in large part by the gastrocnemius-soleus group at the ankle, and to a lesser extent by the quadriceps group at the knee (Winter, 1983). Muscle action is directly related to metabolic cost. Thus, minimal metabolic cost is expected at the natural (resonant) fre- quency. Hypotheses have been generated to account for the mechanism by which head stability may act as a constraint on the emergent movement patterns (Ratcliffe and Holt, in press). However, there remains much work to be done on the relationship between the dynamics of the hybrid model, stability constraints and the synergetics of emergent coordination patterns.
3. Stiffness in the control and coordination of movement
The assumed relationship between the elastic and inertial components in the hybrid model is indexed by the constant 2 applied to the gravitational compo- nent under the root in Eq. (4). In biped cantering and running in experienced humans the approximate integer constant that is needed to predict preferred running period is 5 (Holt et al., 1990b; Kugler and Turvey, 19871, in quadrupedal cantering the constant is 6, and in galloping it is 9 (Kugler and Turvey, 1987). If the hybrid model is correct, and all species operate at their natural (resonant) period, these findings suggest that the ratio kb2 : mL,g in Eq. (3) may vary as a function of the adopted gait pattern. Thus, Eq. (4) may be restated in the more general case
L ; 7=27Tr ,
i 1 ng (5)
where IZ is a constant indexing the relationship kb’ : mL, g. Since changes in the gravitational component mL, g would require rather complex configurational changes of the lower extremity, it might be argued that changes in the ratio may be more easily controlled by changes in the elastic component kb*.

KG. Holte~al./HumanMo~ementScience 15C1996) 177-202 183
The relative contribution of the elastic component to energy conservation has been differentiated across forms of gait. In walking the stance leg behaves like an inverted pendulum coupled to a free pendulum representing the swing leg. Potential (PE) and Kinetic (KE) energy exchanges in the center of mass are achieved through the gravitational effects on the limbs-as-pendulums and can be as high as 68% (McMahon, 1984). Less elastic energy is stored during both stance and swing phases when compared to running. Running has been likened to a bouncing ball in that much of the energy is conserved through storage and elastic recoil in the spring-like properties of the muscles and soft tissues (Cavagna et al., 1977). In current models elastic energy is stored in the stance leg which is coupled to the swing leg acting as a pendulum (McGeer, 1990). Very little energy is conserved through PE and KE exchanges that are the hallmark of walking. Nevertheless, since the body’s center of mass does not lie on the axis of rotation in the gravitational field, the gravitational component still
plays a conservative role. Since both running and walking have spring and pendulum components, the
hybrid model can be used to describe both. The gaits differ, however, in the relative contributions of elastic and inertial energy to total energy conservation. Since elastic energy is stored more effectively through a stiffer spring, it would be expected that the relative contribution of the stiffness and inertial terms would vary in gaits. If the hybrid model is correct, a greater kb’ : mL, g ratio would be expected in running, and the constant n in Eq. (5) would be greater. As noted in the previous paragraph, findings for animal and human walking and running support this claim. It must be recognized that the segmental configura- tions and the dynamics for storing elastic energy may also be different between the two gaits. In walking, energy is stored in the tendon and muscle of gastrocnemius-soleus group as the center of mass moves over the fixed foot of the stance leg, and to a lesser extent in the soft tissues that are being stretched in the swing leg. In running, energy is stored in the gastrocnemius-soleus group, the knee extensors and the hip extensors during weight acceptance following the flight phase. However, since the spring stiffness is considered a global measure, the mechanism is not critical. In anticipation of the model applications to be discussed, stiffness is obtained from the natural frequency and not directly (see Eq. (6)). The actual biological method for obtaining stiffness is immaterial to a hybrid model of a system operating at its natural frequency (although the potential neuromuscular and kinematic means are discussed below).
The hypothesized relationship between the elastic and the gravitational com- ponent suggests that stiffness is a controllable parameter. Is there evidence to support the hypothesis ? The regulation of muscle stiffness in the control of

184 K.G. Halt et al. / Human Mooement Science 1.5 (19961 177-202
movements is one which has received considerable attention in the motor control literature (for review, see Latash, 1993). A muscle or group of muscles are considered to be adjustable springs, the stiffness of which is controlled by the level of muscle activation either through a single muscle or antagonist pair (e.g., Fel’dman, 1966, 1986; Kelso and Holt, 1980; Nichols and Houk, 1976; Polit and Bizzi, 1979). There is also support for the control of stiffness in locomotion as means to influence certain gait parameters. Dietz and colleagues (e.g., Dietz and
Berger, 1983) have claimed that gait control depends largely on the control of stiffness through interaction between muscle activation and the viscoelastic properties of the muscle and connective tissue fibers. Measured increases in leg stiffness based on mass-spring models of the leg have been related to increased hopping frequency (Farley et al., 1991), running frequency (Blickhan, 1989), and running velocity (McMahon and Greene, 1979).
Increased stiffness in locomotion can potentially be achieved in two ways: (1) by cocontraction of antagonist pairs (gastrocnemius-soleus and tibialis anterior, bieps femoris and rectus femoris), and/or (2) by increased isometric contraction of a single muscle opposing a moment due to gravity (e.g., rectus femoris during weight acceptance). The coordinative patterns that emerge facilitate the require- ment for increased stiffness in running. Running is characterized by mass flexion patterns in the actions of the lower extremity joints, while walking is characterized by reciprocal patterns. Mass flexion patterns lend themselves better to energy conservation through elastic storage (mass-spring), while recip- rocal patterns lend themselves better to energy storage through transfer (pendu- lar) mechanisms. Within the dynamical systems framework, it will be important in future research to understand how changes in kb* (and perhaps mL, g) are brought about by changes in the coordination patterns observed in the kinemat- ics even within a particular gait across, for example, different locomotor speeds.
In recent experiments, we investigated the control of stiffness in leg swinging at the hip in non-disabled individuals, modeling the leg as a simple undamped hybrid pendulum-spring (Obusek et al., 1995). The paradigm required the subject to stand on one leg and to swing the other at a comfortable rate under a no-load condition, and with three loads applied to the ankle of the swinging leg. The research was based on a peculiar finding from our previous work on walking (Halt et al., 1990a). In that experiment it was shown that as load was added to the ankle, there was an increasing divergence of the predicted period using a constant of 2 (Eq. (4)) from the observed period. That is, the preferred period could not be predicted by changes in the inertial components (mL,g, mL’,) alone. If the limb was moving at the natural period under all load conditions, from Eq. (2) it is seen that the only controllable variable that would

K.G. Halt et al./ Humun MoL,ement Science 15 (1996) I77-202 185
account for the divergence is kb*. Critical to the experimental estimates of stiffness is that the system is operating at its natural frequency. Leg swinging was chosen as the task because it can be shown that the acceleration and displacement of the limb are linearly related under all loads indicating that the leg is at its natural period. In addition, in this paradigm there are no major changes in the coordination patterns so that there are not changes in L, due to complex configurational changes. Data obtained from sagittal plane kinematics allowed for calculations of period under each of four load conditions. Since the limb is shown to be oscillating at its natural period, a value for limb stiffness can be obtained by rearranging Eq. (2):
I \
- mL,g. (6)
Results of the experiment indicated that kb’ increased linearly with increases in mL,g as load was added (Fig. 3). This finding suggests that stiffness is a controllable parameter in leg swinging. In addition, neural activation increases stiffness in a way that is scaled directly to the inertial properties of the limb suggesting a simplified control principle that might carry across limbs with different inertial properties. Perhaps more importantly, this mechanism could
0 subject3
0 subject6
A subject18
. subject5
0 50 100 150 200
mlg (N.m)
Fig. 3. Individual subject regressions of the elastic (stiffness) on gravitational components of leg swinging.
Note that increases in the load on the ankle produce linear increases in stiffness, but slopes vary across
individuals.

186 K.G. Halt et al. / Humun Mowment Science 15 (19%) 177-202
simplify control as limbs perform actions under differing orientations to gravity, diminishing the control problem due to this form of context-conditioned variabil- ity. Interestingly, the slopes of kb’ vs. mL,g differed for individual subjects. In learning the novel limb-to-mass scalings, individuals increase stiffness to a greater or lesser extent as the gravitational component increases. Some individu- als take greater advantage of the passive forces that are afforded by increases in mL,g, without great rate increases in stiffness that would require muscular cocontraction and larger driving forces (from the hip flexors and extensors) at an increased metabolic cost to the system. This mode of action defines skilled behavior in a dynamical sense in that the individual takes full advantage of the passive (reactive) forces and muscular input is minimized (Vereijken et al., 1992).
4. Sources of increased stiffness in disordered movement
In children with spastic hemiplegic and diplegic cerebral palsy, there are two potential causes for any observed changes in limb stiffness - (1) increases in muscle cocontractions or increased eccentric or isometric contractions against gravity, and/or (2) alterations in mechanical properties of the muscles and connective tissues. During the walking gait of children with spastic CP, there is a marked increase in cocontraction throughout the gait cycle. The physiological mechanisms of cocontraction in CP are not well understood, although several hypotheses have been forwarded.
Spasticity, defined as disinhibition of the stretch and flexor reflexes, can potentially lead to cocontraction as antagonist muscles contract in response to applied stretch (Bobath, 1980). The presence of excessive muscle coactivation and activity in non-stretched muscle through the gait cycle is seen as evidence that spasticity is not the sole cause of cocontraction (Knutsson, 1980). The pathological pattern of muscle activity with excessive cocontraction-contraction has also been attributed to impaired maturation of the locomotor pattern (Berger et al., 1982). The similarities between the pattern seen in the beginning of independent normal walking and that observed in cerebral palsy are claimed to support this assumption (Leonard et al., 1991).
Myklebust (1990) suggested that damage to supraspinal structures during maturation leads to the development of an abnormal spinal circuitry, causing ‘reciprocal excitation’ of agonist and antagonist muscles that is responsible for the abnormalities seen. Conversely, Berbrayer and Ashby (1990) found that

K.C. Halt et al. /Human Movement Science 15 (19961 177-202 187
reciprocal inhibition is normal in cerebral palsy. As an alternative, Brouwer and Ashby ( 199 1) proposed the idea that the lesion of the immature brain caused the development of altered corticospinal projections that are directed equally to motoneurons of both agonist and antagonist muscles.
A common feature of most theories proposed to explain the coactivation pattern in cerebral palsy is the assumption that the emerging movement deficit is caused by the failure of the progressive influence of the brain over the spinal circuits (Milner-Brown and Penn, 1979; Berger et al., 1982; Leonard et al., 1991; Myklebust, 1990; Brouwer and Ashby, 1991). The lack of descending control causes the use of primitive synergies, similar to those in supported gait and at the beginning of normal independent walking, as the main pattern of locomotion (Cahan et al., 1990; Gage, 1990).
The second source of increased stiffness in spastic cerebral palsy is through morphological changes in the muscle and connective tissues. Investigations on the development of muscle stiffness in gait have shown that the increased tension developed in the Achilles tendon of children with cerebral palsy and hemiplegic adults was related more to ankle joint angle than to the myoelectrical activity observed in the gastrocnemius muscle (Berger et al., 1982; Dietz and Berger, 1983). The authors proposed that muscle stiffness during locomotion in spastic patients is due to changed mechanical properties of muscle fibers and connective tissue (e.g., increased tendon-to-muscle length ratio). In addition, gait analysis after selective dorsal rhizotomy demonstrated that, despite the reduced spasticity, patterns of muscle activation during walking were not altered (Cahan et al., 1990). Several studies have demonstrated that the changes in mechanical properties of the muscle-tendon passive components are more related to the increased resistance to joint displacement than to the presence of an altered stretch reflex (Berger et al., 1982; DeSouza, 1987; Katz and Rymer,
1989; Price et al., 1991; Lin et al., 19941. Alterations of the connective and muscle tissues are well documented (Castle
et al., 1979; Hufschmidt and Mauritz, 1985; Tardieu et al., 1989; Lin and Brown, 1992). It has been suggested that the normal longitudinal muscle growth of children with cerebral palsy is altered due to a defective trophic regulation caused by inadequate muscle stretch or isotonic contraction (Tardieu et al., 1982b). This process leads to a bone growth that is not followed by the muscle, resulting in longer bones and tendons and shorter muscle bellies (Tardieu et al., 1982a, b). The result of this uneven structural growth may place the muscle in a different length-tension relationship that is less optimal for power generation and/or lead to increased resistance to a passive stretch (Hufschmidt and Mauritz, 1985; Tardieu et al., 1989; Lin and Brown, 1992).

188 K.G. Ho/t et al. /Human Mowment Science 15 f I9961 177-202
5. Measurement of limb stiffness in disordered movement
Walsh and his colleagues have shown changes in the resonant frequency of passively oscillated limbs in cerebral palsy and other disease states (Walsh, 1992). When driven by a torque motor at increasing frequency, individuals demonstrate a large increase in amplitude of oscillation at a particular frequency, indicating resonance. Essentially, disease states that induced hypotonia in mus- cles and soft tissue (e.g., early stages of stroke, hypermobile joints) produced a decrease in resonant frequency, while disease states that caused hypertonia and increased connective tissue stiffness (e.g., spastic hemiplegia, Parkinsonism) resulted in increased resonant frequency. In a previous study, we have shown increases in the natural frequency of a limb that is allowed to swing passively with induced delayed onset muscle soreness (DOMS) following high output eccentric contractions of the lower arm (Holt et al., 19871.
Of practical importance to physical therapists are the effects of upper motor neuron disorders on ,f~nctional activities such as walking and grasping objects, and the quantitative effects of interventions on those functions. In our experi- ments on children with spastic hemiplegic cerebral palsy, we have sought to use the natural frequency of the hybrid model to estimate the relative limb stiffness in individuals walking and active leg swinging from Eq. (5). By observation of the period of oscillation during overground walking, it is possible to calculate a value for rz if the system is at its natural frequency. The value y1 will index the limb stiffness relative to the gravitational component since the latter would change only with significant reconfiguration of the limb. In the preferred walking of non-disabled children, II = 2, in agreement with adults and quadrupeds (Holt et al., 1991a, b). In children with spastic hemiplegic CP, the mean value, n = 2.43, was significantly different from the non-disabled (Jeng et al., 1996). Olney et al. (1987) showed preferred walking frequencies of 105 strides per minute in the non-disabled and 131 strides per minute in spastic cerebral palsy, suggesting a 30% stiffness increase. These results indicate that either (a) there is an increase in stiffness that causes an increase in walking frequency and the natural frequency is maintained, or (b) that children with spastic CP do not walk at their natural frequency and that use of the model is therefore invalid.
A preliminary test of these alternatives in CP was made using the leg swinging at the hip paradigm, reported in Obusek et al. (1995). As is the case for the non-disabled, operation at the natural frequency was demonstrated through linearity in the acceleration vs. displacement curve of the spastic actively oscillated limb supporting the use of the hybrid model (Butcher, 1995).
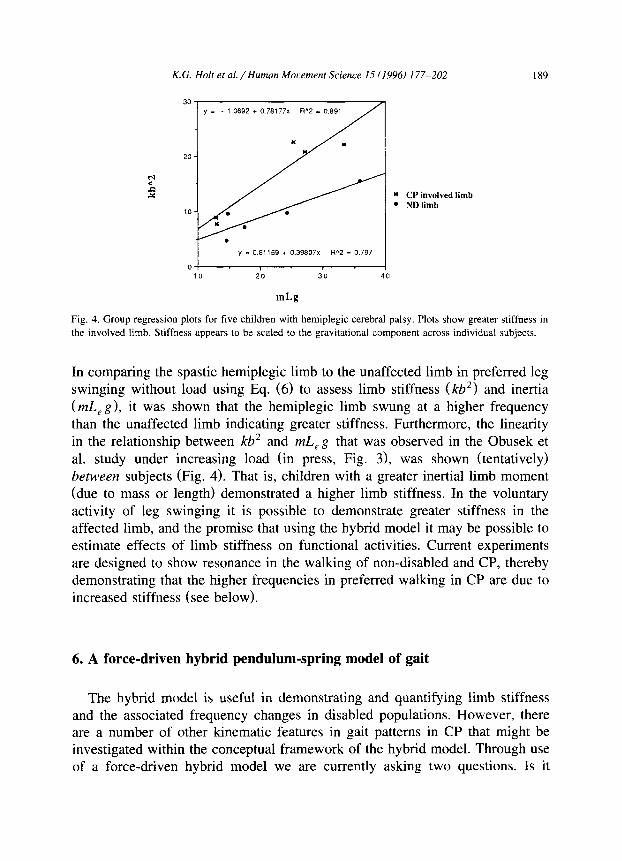
K.G. Holt et al./Human Mooement Science 15 (lYY6) 177-202 189
y = - 1.0692 + 0.78177x R"2 = 0.891
" CP involved limb . ND limb
y E 0.81159 + 039807x R"2 = 0.797
0,
10 20 30 40
mLg
Fig. 4. Group regression plots for five children with hemiplegic cerebral palsy. Plots show greater stiffness in
the involved limb. Stiffness appears to be scaled to the gravitational component across individual subjects.
In comparing the spastic hemiplegic limb to the unaffected limb in preferred leg swinging without load using Eq. (6) to assess limb stiffness (kb*) and inertia (mL, g), it was shown that the hemiplegic limb swung at a higher frequency than the unaffected limb indicating greater stiffness. Furthermore, the linearity in the relationship between kb’ and mL,g that was observed in the Obusek et al. study under increasing load (in press, Fig. 3), was shown (tentatively) between subjects (Fig. 4). That is, children with a greater inertial limb moment (due to mass or length) demonstrated a higher limb stiffness. In the voluntary activity of leg swinging it is possible to demonstrate greater stiffness in the affected limb, and the promise that using the hybrid model it may be possible to estimate effects of limb stiffness on functional activities. Current experiments are designed to show resonance in the walking of non-disabled and CP, thereby demonstrating that the higher frequencies in preferred walking in CP are due to increased stiffness (see below).
6. A force-driven hybrid pendulum-spring model of gait
The hybrid model is useful in demonstrating and quantifying limb stiffness and the associated frequency changes in disabled populations. However, there are a number of other kinematic features in gait patterns in CP that might be investigated within the conceptual framework of the hybrid model. Through use of a force-driven hybrid model we are currently asking two questions. Is it

190 K.G. Ho/t et al. /Human Mmvmmt Sciencr IS (IYY6I 177-202
a. b.
Fig. 5. Schematic representation of force-driven hybrid inverted pendulum spring model, where m = the mass
of the body acting at the body’s center of mass, F,, = the driving force generated by the push off leg,
c = damping, L = simple pendulum equivalent length of body and trunk. Fig. 5a represents the start of the half
cycle (heel strike) and Fig. Sb represents the end of the half cycle (weight acceptance on opposite leg). Solid
line represents the pendulum leg, dashed line represents the contralateral limb.
possible that increases in limb stiffness through cocontraction, increased muscle tone, increased connective tissue infiltration into muscle, and other mechanisms are utilized as sources of energy for locomotion in the face of inability to produce force in the muscles of the leg used for propulsion? Are the gait patterns that are observed in cerebral palsy an optimal configuration for the use of alternative sources of energy for continued locomotion?
To investigate these questions from a quantitative dynamical perspective, it is necessary to invoke a complete model that encompasses both conservative forces (as in the hybrid model) and a periodic forcing function. To this end we are investigating the Newton-Euler equation that determines the motion of a hybrid system that is periodically forced, and investigate more thoroughly the dynamics of walking.
During the stance phase of walking, the body may be treated as an inverted pendulum, in which the forward (weight accepting) foot acts as the axis over which the body center of mass rotates (Fig. 5a). The inverted pendulum travels through a half cycle and the oscillation is damped out by weight acceptance on the other leg (Fig. 5b). Some but not all of this energy is stored and returned to the original oscillating leg. Thus, energy for oscillation is lost through absorp- tion in the weight accepting leg and by energy losses that are due to viscous and

K. G. Halt et al. /Human Mooement Science 15 C 1996) 177-202 191
frictional damping in the joints and connective tissue. These losses are signified in the Newton-Euler equation by the damping term. To maintain the oscillations the center of mass is phasically forced over the axis by the push off leg as the
gastrocnemius-soleus group, and the quadriceps group concentrically contract against the ground. In addition, cycle energy is conserved through elastic and gravitational terms. When the system is at resonance, the force acts only to offset the damping losses and to maintain a given amplitude, and the elastic component determines only the frequency of oscillation. The general equation of motion describing the oscillation is:
mL2(8)=F,Lcos~+mLgsin8+kbsinBbcos8+c(8)bcos6, (7)
where mL2( l?) is the net moment of the center of mass, Fd L cos 8 is the driving moment produced by active muscle contraction, mL, sin 0 is the moment produced by gravity on the center of mass, kb sin 6b cos 0 is the moment produced by stretch on soft tissues, and c( 4 )b cos 8 is the moment produced by viscous properties of soft tissues and energy absorption in the opposite limb during foot contact and weight acceptance.
Potentially, the model could be used to estimate the ratio of different energy sources for locomotion. The critical issue in this respect is the assumption of resonance. We are currently performing experiments using this periodically forced, inverted hybrid model to investigate the dynamics of non-disabled walking over different stride frequencies and walking speeds. Although the relationship between the acceleration and displacement of the body’s center of mass is non- linear, preliminary results from frequency spectral analysis reveal the center of mass oscillates harmonically about the vertical with approximately 80% of the total power concentrated at the fundamental (stride) frequency. This finding remains constant, regardless of the stride frequency or speed of walking, indicating that the system oscillates at its natural frequency and resonance is established across all conditions. Furthermore, since this occurs without any significant change in the (measured) inertial characteristics of the body, the alteration in the natural frequency may only be attributed to a change in global
stiffness. * The ratio of torque contributions from forcing, inertial and elastic terms can
then be made. Assuming a constant damping coefficient, Eq. (6) may be used to calculate kb’ for any frequency or speed of walking. The inertial component
’ We investigated only changes in the inertial characteristics of the center of mass within the model.
Changes in the inertial characteristics of individual oscillators (arms and legs) acting in a complementary fashion across conditions could, potentially allow for this invariance (see Wagenaar and Van Emmerik, 1994).

(mL, g) can be directly calculated from an individual’s anthropometry. The net moment of the center of mass can be directly measured through motion analysis, and the forcing moment, which is equal to the losses due to damping in a system at resonance, can be calculated as the only unknown in Eq. (7). Thus, it is possible to estimate the ratio of different sources of energy for the continued oscillations of the system.
7. Pattern development and functional adaptations to altered dynamics
A significant problem in cerebral palsy gait is the inability to produce a sufficient amount of propulsive force at the right time in the gait cycle during the push-off phase of gait due to weakness in the gastrocnemius-soleus muscle
group (Berger et al., 1982; Olney et al., 1990). The Newton-Euler equation of motion for the forced hybrid model informs us that for any constant value of mL’( t?), oscillations (and locomotion) are maintained by a combination of forcing and the two forms of energy conservation, inertial and elastic. Is it possible that the observed increase in stiffness in spastic CP is an adaptation that allows greater energy conservation and continued locomotion in the face of an inadequacy in the power producing capability of muscle? Several pieces of research, clinical, and morphological evidence support this possibility.
Given the clinical presentation of children with spastic cerebral palsy, de- creased muscle forcing and increased stiffness, the predictions of the model would be as follows: At resonance, forcing is indexed by amplitude with little effect on frequency. In a system with decreased forcing the amplitude would decrease. Stiffness is indexed by frequency with no effect on amplitude. In a stiffer system, the frequency would be expected to increase. Experimental results show that children with CP tend to walk more slowly, with a lower stride amplitude, but at a higher frequency than their non-disabled counterparts (Jeng et al., 19961, supporting a decrease in forcing and an increase in stiffness.
Evidence is also to be found in clinical work that uses electrical muscle stimulation to invoke contractions in weak muscles. Diplegic CP is often characterized by a plantar-flexed foot at foot contact rather than the normally observed heel strike. Electrical stimulation has most usually been attempted on the anterior tibialis muscle to increase foot dorsi flexion and allow for normal heel strike (Atwater et al., 1991; Dubowitz et al., 1988). These attempts have been unsuccessful. However, in a recent case study on three children with spastic cerebral palsy, Carmick (1993) stimulated the major power producing

K. G. Holt et al. /Human Mor,ement Science I5 ( 19961 177-202 193
muscles, gastrocnemius-soleus group during push-off. The results were quite profound and counterintuitive. The observed gait pattern showed a more normal walking pattern that included heel strike despite stimulation of a muscle that might be expected to increase rather than decrease plantar-flexion. One interpre- tation of this finding is that the stimulation supplied the energy resource (forcing) that had been lacking, and the adaptation (increased plantar-flexion to allow for more spring-like action of the foot and ankle) was no longer necessary for continued oscillations.
Another observation that lends support to the concept of functional adaptation to the dynamic requirements of locomotion is also in the observed kinematic gait patterns of children with spastic diplegia. Diplegic walking gait has often been likened to running. There are a number of parameters of walking that distinguish it from running. While a flight phase may not truly represent running (McMahon and Cheng, 1990; McMahon et al., 19871, the two gaits may be distinguished by the exchanges of potential (PE) and kinetic energy (KE) (McMahon, 19841, by the phasic relationships between joint angles in the lower extremity (Deidrich and Warren, in press), and by the trajectory of the center of mass of the body. Cerebral palsy gait is characterized by a lack of energy exchanges between PE and KE (Olney et al., 19901, by in-phase flexion of the hip, knee, and ankle (Bruin et al., 1982; Gage, 1990; Strotzky, 1983; Leonard et al., 19911, and by increased vertical displacement of the center of mass (Strotzky, 1983). These factors are indicative of a gait pattern that more characterizes running than walking. The adoption of a running type of gait may be interpreted in the framework of the equations of motion. The CP may develop a running pattern to take greater advantage of the elastic properties of a running gait that cannot be
equalled in walking. The final piece of information relates to morphological changes in the
connective and contractile tissues in the calf. The ratio of tendon-to-muscle length is greater in CP than in non-disabled children and there is greater infiltration of connective tissue into muscle (Dietz and Berger, 1983; Katz and Rymer, 1989; Tardieu et al., 1982a, b). Our interpretation of this finding is that tendon and other connective tissues are a more economical way to store elastic energy than cocontraction, since they are essentially free of any metabolic cost. One would expect that if the form of locomotion relied on greater amounts of elastic energy to maintain its oscillations, the system would develop morpholog- ical profiles to allow for the efficient use of these resources.
In summary, morphological increases in connective tissue length, a plantar- flexed foot on ground contact, mass flexion patterns in lower extremity joints (crouch gait), and cocontraction are ideal patterns and/or adaptations for using

194 K.G. Holter al. / Human Moremmt Science 15 (19961 177-202
the soft tissues as a spring. The gait patterns in CP gait are optimal and functional for the task at hand given the available dynamic resources.
8. Consequences for rehabilitation
In clinical research, the quantification and effectiveness of clinical interven- tions on functional activities in upper motor neuron disease has been difficult. There are a number of important implications of the modeling approach in this regard. First, the modeling suggests that there may be rather simple clinical measures that can be used to estimate an inability to locomote effectively. Decreased stride length would suggest inadequate ability to produce force in the major power producing muscles of the leg. Increased frequency would suggest that the system has an increased stiffness. Second, the resonance model suggests that in preferred walking gait of the non-disabled the inertial and elastic components are equal (Eq. (4)). The determination of the ratio can be assessed using Eq. (5). This presents a ‘gold standard’ for comparisons to a norm by which the effectiveness of interventions might be assessed. A value of n greater than 2 would suggest unusually high stiffness, while a lower value might suggest some form of hypotonia. One advantage is that the norm is not based on population norms as is the case in most comparative gait analyses (e.g., Winter’s work), but on the individuals own anthropometry. Since Eq. (5) is based on simple anthropometrical measures and the observation of gait period (frequency), the model is simple to use in a clinical setting.
Of major concern to therapists is the plantar-flexed foot on ground contact (toe walking), cocontraction of agonist pairs, and a gait that has been likened to running. The way these abnormalities are interpreted will have consequences for rehabilitation. Let us take cocontraction as an example. Cocontraction may be a direct result of the disease process through a mechanism such as ‘reflex excitation’ (Myklebust, 1990). A classic interpretation is that cocontraction interferes with the normal gait pattern, and attempts should be made to therapeu- tically decrease or reduce the amount of cocontraction (to ‘normalize’ the gait pattern). The second interpretation is that the locomotor system by developing a gait pattern that takes advantage of this pathology stores and returns elastic energy. Cocontraction increases stiffness that in turn will allow greater energy return. Rehabilitation might be geared to taking full advantage of this energy source by actually teaching a running type of gait. In a more general sense, identification and training of a movement pattern that takes advantage of the available resources would become an integral part of the therapeutic regimen.
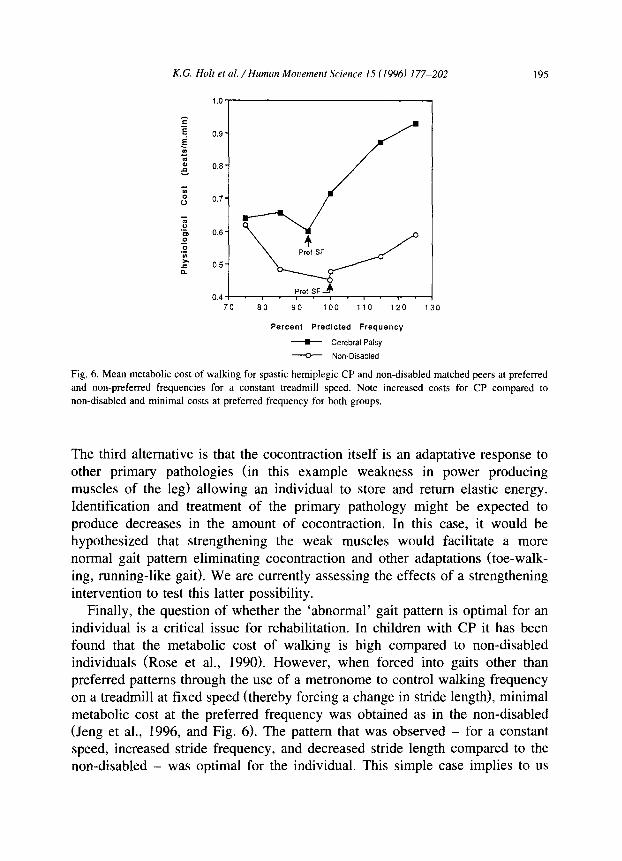
K.G. Holt et al. /Human Mowmeni Science 15 (1996) 177-202 195
i
0.9 -
0.8 -
0.7
;:&
Pref SF 4 0.4 . I I I . I I
70 80 90 100 110 120 1 0
Percent Predicted Frequency
--M- Cerebral Palsy
--O- Non-Disabled
Fig. 6. Mean metabolic cost of walking for spastic hemiplegic CP and non-disabled matched peers at preferred
and non-preferred frequencies for a constant treadmill speed. Note increased costs for CP compared to
non-disabled and minimal costs at preferred frequency for both groups.
The third alternative is that the cocontraction itself is an adaptative response to other primary pathologies (in this example weakness in power producing muscles of the leg) allowing an individual to store and return elastic energy. Identification and treatment of the primary pathology might be expected to produce decreases in the amount of cocontraction. In this case, it would be hypothesized that strengthening the weak muscles would facilitate a more normal gait pattern eliminating cocontraction and other adaptations (toe-walk- ing, running-like gait). We are currently assessing the effects of a strengthening intervention to test this latter possibility.
Finally, the question of whether the ‘abnormal’ gait pattern is optimal for an individual is a critical issue for rehabilitation. In children with CP it has been found that the metabolic cost of walking is high compared to non-disabled individuals (Rose et al., 1990). However, when forced into gaits other than preferred patterns through the use of a metronome to control walking frequency on a treadmill at fixed speed (thereby forcing a change in stride length), minimal metabolic cost at the preferred frequency was obtained as in the non-disabled (Jeng et al., 1996, and Fig. 6). The pattern that was observed - for a constant speed, increased stride frequency, and decreased stride length compared to the non-disabled - was optimal for the individual. This simple case implies to us

196 K.G. Holt et ul. / Humun Morenwnt Scirncr 15 (I 9%) 177-202
that rehabilitation should be directed at the cause of the abnormality (e.g., decreased capability of the muscles to produce sufficient force at the appropriate time in the gait cycle) and not the symptoms (e.g., coordination pattern, low speed, high frequency and short stride length). If nothing can be done about the cause, or it cannot be identified, then perhaps the pattern of gait should be allowed to take its own form as optimal for the individual.
9. Concluding remarks
Dynamical systems theory conceives locomotion as a behavior that arises from the contribution of several subsystems (Clark and Phillips, 1993). The organization of movement is achieved because biological systems are governed by dynamical laws, and by constraints that guide those dynamics (Kelso et al., 198 1). Biological systems are self-organizing entities in that movement patterns and order can emerge from the process of the interactions of the several components of the biological system and the environment, without the need for hierarchical control (Thelen, 1989). It has been suggested that the emergence of preferred patterns of movement is the result of task constraints (e.g., see van Van Emmerik and Newell, 1990) and constraints that are due to the tendency for biological systems to self-optimize (Halt, 19931. These constraints include but are not limited to; the minimization of metabolic cost, minimum variability of the head trajectory, and minimization of shock to the head (Halt et al., in press). Behavioral patterns emerge from the integration of mechanical and physiological variables that are organized in an optimal configuration to produce the observed performance at a minimal ‘cost’.
From a dynamical systems theoretical perspective, the stance outlined in this paper adds another implication for the details of self-organizing capability of biological systems in emerging movement patterns. It implies that observed coordinations of joints and muscle actions are driven in part by the underlying dynamics of both the task and the individual. The dynamics can be formalized in locomotion (and perhaps other tasks) through the force-driven hybrid oscillator model. Limitations or changes in the dynamics available to a particular biologi- cal system may drive it into a different organization. For example, development of a running-like gait in CP may be driven by limitations in the ability to produce sufficient force (amplitude1 or a stiffness (frequency) so high that it would prevent pendular (walking) movement at all. Similarly, changes in the task requirement may also bring about change in movement patterns. In a locomotory task that requires getting to a destination quickly, the inadequacy of

K.G. Holi et (11. /Human Morement Science 15 f 1996) 177-202 107
the dynamics in walking (pendular with limited spring), would necessarily be replaced by a dynamic organization of the pattern that would allow greater usage of elastic energy (spring with limited pendular), i.e., running.
In the terminology of the synergetics approach (e.g., Kelso and Schiiner, 1988), this discourse suggests that the control parameters that determine gait patterns are to be found in the dynamic equations of motion rather than in the frequency domain that is only an index of the underlying dynamics. It further points to the possibility that stability is only one constraint that is relevant for coordination. Physiological cost, shock absorption, dynamic and other as yet undiscovered constraints will help reveal why we observe the patterns of coordination that we do.
Appendix A. Derivation of equation of motion for hybrid mass-spring pendulum
To determine the simple pendulum equivalent length, individual segment moments of inertia are calculated using anthropometric data and segmental inertial estimates. The moments of inertial of the thigh, shank, and foot segments are given by:
where mseg is the mass of the segment estimated from the total body mass, pa_
is the estimated segment radius of gyration about its center of mass (Jensen, 1986), and lseg is the measured length of the segment in meters. The moment of inertia of the entire lower extremity about the hip axis is calculated using the parallel axis theorem according to the following:
3
zsyc= C(z;+m,nZ), (A9 i= I
where m, is the estimated segment mass, Di is the distance from the segment estimated center of mass to the axis of rotation at the hip.
The distance from the axis of motion at the hip joint to the center of mass of the lower extremity system (D_) is calculated by:
W)

198 K.G. Halt et al. /Human Morement Science 15 C I9961 177-202
where mi is the estimated segment mass, and m is the estimated total mass of
the leg. The simple pendulum equivalent of the leg can then be calculated from the
system moment of inertia (L.,,), the estimated center of mass ( Ds,,), and the estimated total mass (m) according to the following:
*sys L, = -
Dsys m W)
(Kugler and Turvey, 1987; Holt et al., 1990a, b). The motion of the hybrid mass-spring pendulum is given by the following
Newtonian equation of motion:
mL?,$=mL,gsin$+kb’sin~cos$, (W
where 4 is the angular displacement of the leg pendulum from the vertical and 4 is its angular acceleration. If the amplitude of oscillation is small (4 < 20 degrees), it may be assumed that sin C$ = 4 and cos C)J = 1. Therefore, the motion of the hybrid pendulum may be given by the following linear approxima- tion of Eq. (A%:
mLi;f; = mL,g+ f kb2$. Gw
Eq. (A61 identifies the two contributions to the motion of the hybrid pendulum - the moment due to gravity (mL, g), and the moment due to the elastic component ( kb2) which is provided by the attached spring system. These two forces act in the same direction in the model and their forms are identical, therefore their combined effect may be taken as their sum.
In this experiment, we assume that the frequency of oscillation is determined by the following linear equation of motion:
jl= -&& (‘47)
where x is the horizontal displacement of the pendulum, .Y is the horizontal acceleration, and w is the natural frequency. Since the model consists of a single, rigid link between the axis of motion and the pendulum mass, we may assume:
+ -(&. W)
The negative sign indicates direction, thus the magnitude is given as I - 0~1.
Harmonic motion of this type implies that both the forcing and damping functions influence amplitude but have a negligible effect on the frequency of oscillation and therefore are not considered.

K.G. Holt et d/Human Mocement Science 15 (1996) 177-202 199
This assumption further allows the specification of the following relationship
between the cycle period and frequency:
2?r 7= -, @9) w
where G- is the measure of cycle period. By solving Eq. (A6) for 4 and substituting into Eq. (A8), we obtain the
following expression for the natural frequency:
w= [‘““:;L:*b2]:. (*lo)
By applying the relationship given in Eq. (A9), the natural period of the hybrid mass-spring pendulum may be defined as:
Solving Eq. (Al 1) for the moment due to stiffness ( kb2):
- mL,g. bw
References
Atwater, SW., M.E. Tatarka, J.E. Kathrein and S. Shapiro, 1991. Electromyography-triggered electrical
muscle stimulation for children with cerebral palsy: A pilot study. Pediatric Physical Therapy 3, 190-199.
Berbrayer, D. and P. Ashby, 1990. Reciprocal inhibition in cerebral palsy. Neurology 40, 653-656.
Berger, W., J. Quintern and V. Dietz, 1982. Pathophysiology of gait in children with cerebral palsy.
Electroencephalography and Clinical Neurology 53, 538-548.
Blickhan, R., 1989. The mass-spring model for running and hopping. Journal of Biomechanics 22, 1217-1227.
Bobath, B., 1980. Abnormal postural reflex activity caused by brain lesions (2nd Ed.). London, England:
William Heinemann Medical Books Ltd.
Brouwer, B. and P. Ashby, 1991. Altered corticospinal projections to lower limb motoneurons in subjects with
cerebral palsy. Brain 114, 1395-1407.
Bruin, D.D., P. Eng, J. Russell, J.E. Latter and J.T.S. Sadler, 1982. Angle-angle diagrams in monitoring and quantifications for children with cerebral palsy. American Journal of Physical Medicine 6 I, 176- 192.
Butcher, R., 1995. Comparisons of limb stiffness between non-disabled children and children with spastic
cerebral palsy in limb swinging using a force-driven mass-spring pendulum model of the leg. MS thesis,
Boston University, Sargent College.

200 K.G. Halt et ~11. / Humcm Moremerrt Science 15 (19%) 177-202
Cahan, L.D., J.M. Adams, J. Perry and L.M. Beeler, 1990. Instrumented gait analysis after selective dorsal
rhizotomy. Developmental Medicine and Child Neurology 32, 1037- 1043.
Carmick, J., 1993. Clinical use of neuromuscular electrical stimulation in children with cerebral palsy.
Physical Therapy 73, 505-5 13.
Castle. M.E.. T.A. Reyman and M. Schneider, 1979. Pathology of spastic muscle in cerebral palsy. Clinical
Orthopedics and Related Research 42, 223-233.
Cavagna. G.A., N.C. Heglund and C.R. Taylor, 1977. Mechanical work in terrestrial locomotion: Two basic
mechanisms for minimizing energy expenditure. American Journal of Physiology 233. R243-R26l.
Clark, J.E. and S.J. Phillips, 1993. A longitudinal study of intralimb coordination in the first year of
independent walking: A dynamical systems analysis. Child Development 64, 1143-l 157.
Deidrich, F.J. and W.H. Warren. in press. Why change gaits‘? Dynamics of the walk-run transition. Journal of
Experimental Psychology: Human Performance and Perception.
DeSouza. L.H.. 1987. The measurement and assessment of spasticity. Clinical Rehabilitation I. 89-96.
Dietz, V. and W. Berger. 1983. Normal and impaired regulation of muscle stiffness in gait: A new hypothesis
about muscle hypertonia. Experimental Neurology 79. 680-687.
Dubowitz, L.. N. Finnie, S.A. Hyde, 1988. Improvement in muscle performance by chronic electrical
stimulation in children with cerebral palsy. Lancet, March 12. 587-588
Farley, C.T.. R. Blickhan,, J. Saito and R. Taylor, 1991. Hopping frequency in human: A test of how springs
set stride frequency in bouncing gaits. Journal of Applied Physiology 71, 2 127-2 132.
Fel’dman, A.G., 1966. Functional tuning of the nervous system with control of movement or maintenance of a
steady posture: Il. Controllable parameters of the muscles. Biophysics I I, 565-578.
Fel’dman, A.G., 1986. Once more on the equilibrium point hypothesis (I model) for motor control. Journal of
Motor Behavior 18. 17-53
Fetters. L.. 199 I. Measurement and treatment in cerebral palsy: An argument for a new approach. Physical
Therapy 7 I. 243-247.
Gage, J.R.. 1990. Surgical treatment of knee dysfunction in cerebra1 palsy. Clinical Orthopedics and Related
Research 253, 45-54.
Holt, K.G.. 1993. Toward general principles for research and rehabilitation of disabled populations. Physical
Therapy Practice 2(4). I - 18.
Holt. KG.. P. Clarkson. R.O. Andres and 1. Tromblay. 1987. Assessment of muscle damage using a non-linear
mass-spring model. Proceedings Institute of Electronic and Electrical Engineers (IEEE). Engineering in
Medicine and Biology, Boston.
Holt. K.G.. J. Hamill and R.O. Andres, 1990a. The force-driven harmonic oscillator as a model for human
locomotion. Human Movement Science 9. 55-68.
Holt, K.G., J. Hamill and M.M. Slavin, 1990b. Running at resonance: Is it a learned phenomenon’?
Proceedings of the Canadian Society for Biomechanics, Quebec: Organizing Committee, CSB.
Holt, K.G.. J. Hamill and R.O. Andres, 199la. Predicting the minimal energy costs of human walking.
Medicine and Science in Sports and Exercise 23, 491-498.
Holt, K.G., S.F. Jeng and L. Fetters, 199lb. Walking cadence of 9-year olds is predictable as the resonant
frequency of a force-driven harmonic oscillator. Pediatric Exercise Science 3. 12 I - 128.
Holt, K.G.. S.F. Jeng, R. Ratcliffe and J. Hamill, in press. Energetic cost and stability in preferred human
walking. Journal of Motor Behavior.
Hufschmidt. A. and K.A. Mauritz. 1985. Chronic transformation of muscle in spasticity: A peripheral contribution to increased tone. Journal of Neurology, Neurosurgery, and Psychiatry 48, 676-685.
Jeng, S.F.. K.G. Holt, L. Fetters and C. Certo, 1996. Self-optimization of walking in non-disabled children and
children with spastic hemiplegic cerebral palsy. Journal of Motor Behavior 28, 15-27.
Jensen. R.K.. 1986. Body segment mass, radius and radius of gyration proportions of children. Journal of Biomechanics 19. 359-36X.
Katz. R.T. and Z. Rymer, 1989. Spastic hypertonia: Mechanism and measurement. Archives of Physical Medicine and Rehabilitation 70, 141-155.

K.G. Holt et al. / Human Molxement Science 15 ( I9961 177-202 201
Kelso. J.A.S. and K.G. Holt, 1980. Exploring a vibratory systems analysis of human movement production.
Journal of Neurophysiology 43. 1183-I 196
K&o, J.A.S., K.G. Holt, P. Rubin and P.N. Kugler, 1981. Patterns of human interlimb coordination emerge
from the properties of non-linear, limit cycle oscillatory processes: Theory and data. Journal of Motor
Behavior 13, 226-26 1.
Kelso, J.A.S. and G.S. Schiiner. 1988. Self-organization of coordinative movement patterns. Human Move-
ment Science 7, 27-46.
Knutsson, E., 1980. ‘Restraints of spastic muscle in different types of movement’. In: R.G. Feldman, R.R.
Young and W.P. Koella (Eds.1, Spasticity: Disordered motor control. Chicago, IL: Year Book Medical
Publishers Inc.
Kugler, P.N. and M.T. Turvey, 1987. Information. natural law, and the self-assembly of rhythmic movement.
New Jersey: Erlbaum.
Latash, M.L., 1993. Control of human movement. Champaign, IL: Human Kinetics Publishers.
Latash, M.L. and J.G. Anson, in press. What are ‘normal movements’ in atypical population? Behavioral and
Brain Sciences.
Leonard, CT., H. Hirschfeld and H. Forssberg, 1991. The development of independent walking in children
with cerebral palsy. Developmental Medicine and Child Neurology 33, 567-577.
Lin, J-P. and J.K. Brown, 1992. Peripheral and central mechanisms of hindfoot equinus in childhood
hemiplegia. Developmental Medicine and Child Neurology 34, 949-965.
Lin, J-P., J.K. Brown and R. Brotherstone, 1994. Assessment of spasticity in hemiplegic cerebral palsy I:
Proximal lower-limb reflex excitability. Developmental Medicine and Child Neurology 36, 116- 129.
McGeer, T., 1990. Passive dynamic walking. International Journal of Robotics Research 9, 62-82.
McMahon, T.A., 1984. Muscles, reflexes, and locomotion. Princeton. NJ: Princeton University Press.
McMahon T.A. and P.R. Greene, 1979. The inlluence of track compliance on running. Journal of Biomechan-
its 12, 893-904
McMahon, T.A. and G.C. Cheng, 1990. The mechanics of running. How does stiffness couple with speed?
Journal of Biomechanics 23, 65-78.
McMahon, T.A.. G. Valiant and E.C. Frederick, 1987. Groucho Running. Journal of Applied Physiology 62,
2326-2337.
Milner-Brown, H.S. and R.D. Penn, 1979. Pathophysiological mechanisms in cerebral palsy. Journal of
Neurology, Neurosurgery, and Psychiatry 42, 606-618.
Mochon. S. and T.A. McMahon, 1980. Ballistic walking: An improved model. Mathematical Biosciences 52,
24 I-260.
Morowitr. H.J., 1978. Foundations of Bioenergetics. New York: Academic Press.
Myklebust, B.M.. 1990. A review of myotatic reflexes and the development of motor control and gait in
infants and children: A special communication. Physical Therapy 70, 188-203
Nichols T.R. and J.C. Houk, 1976. Improvement in the linearity and regulation of stiffness that results from
actions of the stretch reflex. Journal of Neurophysiology 39, 1 19-142
Obusek, J.. K.G. Holt and R. Rosenstein, 1995. The hybrid mass-spring pendulum model of leg swinging:
Stiffness as a determinant of cycle period. Biological Cybernetics 73, 139- 147.
Olney, S.J., P.A. Costigan and D.M. Hedden, 1987. Mechanical energy patterns in gait of cerebral palsied
children with hemiplegia. Physical Therapy 67, 1348- 1354.
Olney, S.J., H.A. MacPhail and D.M. Hedden, 1990. Work and power in hemiplegic cerebral palsy. Physical
Therapy 70, 43 l-438.
Polit A. and E. Bizzi, 1979. Characteristics of motor programs underlying arm movements in monkeys.
Journal of Neurophysiology 42, 1 X3- 194
Price, P., K.F. Bjornson, J.F. Lehmann, J. F.McLaughlin and R.M. Hays, 1991. Quantitative measurement of
spasticity in children with cerebral palsy. Developmental Medicine and Child Neurology 33, 585-595.
Ratcliffe, R. and K.G. Holt. in press. Shock absorption in human walking. Gait and Posture.
Rose, J., J.G. Gamble. A. Burgos. J. Medeiros and W.L. Haskell, 1990. Energy expenditure index of walking

202 K.G. Holt et al. /Human Movement Science 15 (19961 177-202
for normal children and for children with cerebral palsy. Developmental Medicine and Child Neurology
32, 333-340.
Strotzky, K., 1983. Gait analysis in cerebral palsied and nonhandicapped children. Archives of Physical
Medicine and Rehabilitation 64, 291-295.
Tardieu, G., E. Huet de la Tour, M.D. Bret and C. Tardieu, 1982a. Muscle hypoextensibility in children with
cerebral palsy I. Clinical and experimental observations. Archives of Physical Medicine and Rehabilitation
63, 97- 102.
Tardieu. G.. C. Tardieu, P.C. Justin and A. Lespargot, 1982b. Muscle hypoextensibility in children with
cerebral palsy II. Therapeutic implications. Archives of Physical Medicine and Rehabilitation 63, 103- 107.
Tardieu, C., A. Lespargot, C. Tabary and M.D. Bret, 1989. Toe-walking in children with cerebral palsy:
Contributions of contracture and excessive contraction of triceps surae muscle. Physical Therapy 69,
656-662.
Thelen. E., 1989. ‘Self-organization in developmental processes: Can systems approaches work?’ In: M.
Gunnar and E. Thelen (Eds.), Systems and development: The Minnesota Symposium on Child Psychology
(22, pp. 77-l 17). Hillsdale: Lawrence Earbaum Associates Inc.
Turvey, M.T., 1990. Coordination. American Psychologist 45, 938-953.
Turvey, M.T., R.C. Schmidt, L.D. Rosenblum and P.N. Kugler, 1988. On the time allometry of co-ordinated
rhythmic movements. Journal of Theoretical Biology 130, 285-325.
Van Emmerik, R.E.A. and K.M. Newell, 1990. The effects of task constraints on intralimb and pen kinematics
in a drawing task. Acta Psychologica 73, 171-190.
Vereijken, B., R.E.A. van Emmerik, H.T.A. Whiting and K.M. Newell, 1992. Free(z)ing degrees of freedom
in skill acquisition. Journal of Motor Behavior 24, 133-142.
Wagenaar, R.C. and R.E.A. van Emmerik, 1994. Dynamics of pathological gait. Human Movement Science
13.441-471.
Walsh, G.E.. 1992. Muscles, masses and motion. New York: Cambridge University Press.
Winter, D.A., 1983. Biomechanical motor patterns in nondisabled walking. Journal of Motor Behavior 15,
302-330.
Winter, D.A., 1990. Biomechanics and motor control of human movement. New York: Wiley and Sons Inc.
Winter. D.A.. G.K. Ruder and CD. MacKinnon. 1990. ‘Control of balance of upper body during gait’. In:
J.M. Winters and S.L.-Y. Woo (Eds.), Multiple muscle systems. Biomechanics and movement organization
(pp. 534-541). New York: Springer-Verlag.




















