
Non-rhinovirus enteroviruses associated with respiratory infections in Peru (2005-2010)
Upload
independentCategory
view
0download
0
Veterinary Microbiology, 8 (1983) 321--361 321 Elsevier Science Publishers B.V., Amsterdam - -Pr in ted in The Netherlands
R e v i e w
C O N G E N I T A L I N F E C T I O N S W I T H N O N A R B O T O G A V I R U S E S
J.T. VAN OIRSCHOT
Central Veterinary Institute, Department of Virology, Houtribweg 39, 8221 RA Lelystad (The Netherlands)
(Accepted 31 January 1983)
ABSTRACT
Van Oirschot, J.T., 1983. Congenital infections with nonarbo togaviruses. Vet. Micro- biol., 8: 321--361.
The present review deals with the similarities and differences of selected aspects of pre- natal pestivirus infections of domestic animals and congenital rubella of man.
Hog cholera virus, bovine virus diarrhoea virus and border disease virus are antigenical- ly closely related, but unrelated to rubella virus. The nonarbo togaviruses are capable of producing congenital infections resulting in a wide spectrum of abnormalities. The in- fected foetus can die in utero, in the neonatal period, or it may be born with teratogenic defects. In addition, apparently healthy progeny can be delivered that develop a late on- set disease, months, or years after birth, or remain clinically normal for life. The ult imate outcome of a congenital infection is mainly determined by the stage of foetal develop- ment, at which infection occurs. Foetuses exposed to rubella virus raise an ant ibody re- sponse to the virus, whereas domestic animals frequently fail to respond immunological- ly to a congenital pestivirus infection.
In congenital rubella the virus usually disappears from the host 's body 1--2 years after birth. However, congenital pestivirus infections may be characterized by a lifelong and widespread persistence of virus in clinically healthy animals. Such animals are of signifi- cance in the epizootiology of bovine virus diarrhoea, border disease or hog cholera.
INTRODUCTION
T h e v i ruses t h a t c a u s e h o g c h o l e r a (HC, s y n o n y m : s w i n e f eve r ) b o v i n e v i ra l d i a r r h o e a ( B V D , s y n o n y m : m u c o s a l d i s e a s e ) o r b o r d e r d i s e a s e (BD, s y n o n y m : h a i r y s h a k e r d i s e a s e ) b e l o n g t o t h e g e n u s Pestivirus. R u b e l l a v i ru s ( R U V ) is t h e so le m e m b e r o f t h e g e n u s Rubiv irus . T h e p e s t i v i r u s e s a r e se ro - l o g i c a l l y r e l a t e d , b u t u n r e l a t e d t o R U V ( P o r t e r f i e l d e t al . , 1 9 7 8 ) . T h e t w o g e n e r a a re , t o g e t h e r w i t h t h e g e n e r a Alphav irus a n d Flavivirus, i n c o r p o r a t e d i n t o t h e f a m i l y o f T o g a v i r i d a e . A l p h a - a n d m o s t f l av iv i ruses a r e t r a n s m i t t e d b y a r t h r o p o d s . S i n c e t h e v i ru ses b e l o n g i n g t o t h e g e n e r a Pestivirus o r Rubi - virus d o n o t d e p e n d o n b i o l o g i c a l t r a n s m i s s i o n b y a r t h r o p o d s f o r t h e i r c o n - t i n u a n c e in n a t u r e , t h e c o l l e c t i v e t e r m " n o n - a r t h r o p o d - b o r n e " o r n o n a r b o t o g a v i r u s e s has b e e n i n t r o d u c e d f o r t h e s e v i ruses ( H o r z i n e k , 1 9 7 3 ) . L a c t i c d e h y d r o g e n a s e v i ru s ( L D V ) a n d e q u i n e a r t e r i t i s v i rus ( E A V ) a re a d d i t i o n a l
0378-1135/83/$03.00 © 1983 Elsevier Science Publishers B.V.

322
nonarbo togaviruses, outside the existing genera (Porterfield et al., 1978). After infecting a pregnant animal, the nonarbo togaviruses are capable of
crossing the placental barrier and invading the foetus. The congenital infec- tion can result in a wide spectrum of abnormalities varying from death of the foetus to a persistent, lifelong infection without clinical signs.
HC is a very contagious disease of pigs, that can be present as an acute, chronic or clinically inapparent infection. The mortali ty rate ranges from nil to one hundred percent depending largely on the virulence of the virus. In the early decades of this century, it was recognized that hog cholera virus (HCV) could cause congenital infections. Benner (1928) and Michal- ka (1931) described the occurrence of abortion, stillbirth and neonatal death after vaccination of pregnant sows by the simultaneous method. A teratogenic activity of HCV was strongly suggested by the birth of mal- formed piglets after vaccination of pregnant dams with modified live viruses (Sautter et al., 1953). The definite evidence that HCV was able to cross the placenta came from in vivo transmission experiments (Young et al., 1955). Congenitally-infected piglets usually are stillborn, or succumb shortly after farrowing, but Van Oirschot and Terpstra (1977) reported that prenatally infected piglets can survive for months after birth.
Two clinically-distinct diseases of cattle, BVD (Olafson et al., 1946) and mucosal-disease (Ramsey and Chivers, 1953} were eventually shown to be caused by bovine viral diarrhoea virus (BVDV). Most BVDV infections are inapparent, though acute and chronic forms of the disease exist. BVDV in- fections have been associated with abortion and neonatal death since the ini- tial description of the disease. It was not until 1967 that the virus was iso- lated from aborted foetuses (Gillespie et al., 1967). Experimental infections of pregnant cattle established that the virus can act as a teratogenic agent (Ward et al., 1969). Congenitally-infected calves do not always succumb shortly after birth, they may survive for years in a clinically healthy state, while remaining persistently infected (Coria and McClurkin, 1978; Strayer et al., 1983).
Infection of sheep with border disease virus (BDV) invariably remains sub- clinical. BD only becomes clinically manifest when a pregnant ewe encoun- ters the virus and consequently delivers a dead or clinically affected lamb. The existence of lambs with an abnormal hairy birthcoat, body tremors and a slow rate of growth in flocks of the Welsh border counties was first de- scribed by Hughes et al. (1959). The viral aetiology of the disease was es- tablished by in vivo transmission studies (Gardiner and Barlow, 1972). The teratogenic potential of BDV has already been indicated by the first descrip- tion of the disease. Recent reports describe in utero infected sheep which had been persistently infected for years, while remaining apparently healthy (Barlow et al., 1980b; Terpstra, 1981).
Rubella is generally a mild disease in humans, but once the foetus be- comes infected, serious damage may occur. Gregg (1941} initially recognized that maternal rubella in early pregnancy can lead to severe foetal malforma-

323
tions, of which the principal ones were those affecting the eye, heart and ear. It was not until 1963, that Selzer (1963) demonstrated the presence of RUV in a foetus, 10 days after the onset of rubella in the mother. In the wake of the great epidemic that swept the U.S.A. in 1964, a tremendous amount of new information on clinical manifestations, pathogenesis and epi- demiology of congenital rubella has been reported. These studies were facili- tated by the successful isolation of RUV in vitro in 1962 (Parkman et al., 1962; Weller and Neva, 1962). Persistent infections are common after birth in congenital rubella.
Most EAV infections take an inapparent course (McCollum and Bryans, 1973; MoraiUon and Moraillon, 1978). However, the virus may occasionally cause severe disease in horses (Bryans et al., 1957}. When a susceptible preg- nant mare is exposed to the virus, the foetus may become infected and abor- tion can occur. The virus was first isolated from an aborted foetus in 1957 (Doll et al., 1957). The significance of subclinical EAV infection with regard to abortion is not known.
LDV causes an asymptomatic persistent infection in mice. In the acute phase of the infection in pregnant mice LDV can cross the placental barrier and infect the offspring, without any serious effect (Georgii et al., 1964; Crispens, 1965).
There appear to be no serological relationships between the pestiviruses and RUV, EAV and LDV (Horzinek, 1981).
However, the viruses constituting the genus Pestivirus are antigenically and structurally closely related. HCV and BVDV cross-react in immunodiffu- sion (Darbyshire, 1960), immunofluorescence (Mengeling et al., 1963) and neutralization assays (Kumagal et al., 1962; Corthier et al., 1974}. Pigs ex- posed to certain strains of HCV or BVDV can develop neutralizing antibody to both HCV and BVDV (Fernelius et al., 1973b; Carbrey et al., 1976; Liess et al., 1976}. Immunization of pigs with BVDV can confer some protection against challenge with virulent HCV (Sheffy et al., 1961; Snowdon and French, 1976). BDV-immunized pigs also appear to be cross-protected against HCV infection (Laude and Gelfi, 1979). Sheep infected naturally or experimentally with BDV produce neutralizing antibody to all 3 pesti- viruses (Acland et al., 1972; Hamilton and Timoney, 1972; Osburn et al., 1973) and BDV antigen can be demonstrated in cell culture or tissue sections by immunofluorescence with a BVDV or HCV antiserum (Harkness et al., 1977, Terpstra, 1978). Sheep inoculated with a virus isolated from a lamb showing signs of BD produced higher neutralizing antibody titres to BVDV than to BDV (Barlow et al., 1979). In addition, BVDV and BDV may cross- infect the ovine and bovine species and induce comparable congenital de- fects (Ward, 1971; Gibbons et al., 1974; Terlecki et al., 1980). Laude (1979) has reported that the pestiviruses, when grown in the same cell system, have a similar buoyant density and sedimentation coefficient.
In view of the observations mentioned above, one may question whether it is justified to consider the pestiviruses, and BVDV and BDV in particular,

324
as distinct viral entities. However, for the sake of clarity the 3 pestiviruses are described here in relation to their "natura l" host, which means that HCV is related to swine, BVDV to cattle and that viruses isolated from sheep with signs of border disease or hairy shaker disease are termed BDV. A separate section will deal with the various cross-infections.
This paper is concerned mainly with the similarities and differences be- tween selected aspects of prenatal infection of domestic animals with pesti- viruses and additionally with congenital rubella infection of man.
CLINICO-PATHOLOGICAL FEATURES
Congenital infection with HCV, BVDV, BDV or RUV ranges from ex- tremely severe to clinically inapparent. The results are mainly dependent on the stage of foetal development at which infection takes place. In general, the risk for the foetus is highest during early pregnancy.
An in utero infection can result in prenatal death, foetal malformations and lesions, or death of the neonate or later on in life. In addition, apparent- ly healthy offspring can be delivered that develop a late onset disease months or years after birth, or remain clinically normal for life.
As a rule, clinical evidence of maternal disease is minimal or absent.
Prenatal death
Early embryonic death with subsequent complete resorption may occur after infection in the very early stages of pregnancy, before the skeleton has developed fully. This is suggested by the increase of barrenness of sows in- fected with HCV (Dunne and Clark, 1968; Stewart, 1969) and of ewes ex- posed to BDV (Manktelow et al., 1969; Porter et al., 1972). Several investi- gations have suggested that BVDV causes early embryonic death. Cows arti- ficially inseminated and subsequently given an intrauterine infusion with BVDV around breeding showed a significantly decreased pregnancy rate (Archbald and Zemjanis, 1977; Whitmore et al., 1981) and an abnormal development of preimplantation embryos (Archbald et al., 1979). On the other hand, cows infected oronasally 2 h after breeding have normal preg- nancy rates (Whitmore et al., 1981) and BVDV does not appear to cross the placenta before the 58th day of gestation (Kendrick, 1976). Thus, in view of the latter findings and because BVDV is rarely excreted in semen it is unlike- ly that, under natural conditions, BVDV is a major cause of early embryonic death. Whether RUV causes early embryonic death is not known.
Abortion is a common sequel to congenital infections with nonarbo toga- viruses, including EAV. Unfortunately, in many studies, virus isolation from the aborted foetus is not at tempted, or remained unsuccessful due to the ad- vanced stages of autolysis. In the field, abortion of animals may often go un- noticed. Thus, the real incidence of abortion is difficult to determine, but it seems to be more common in sheep and cattle than in man and swine. Abor-

325
tion is usually the consequence of infection during the first third of preg- nancy, but in sheep it can also be the result of maternal BVD infection around mid-pregnancy (Barlow and Gardiner, 1969). The period that elapses between virus exposure and abortion is variable and seems to be different for the various species. Obviously, it is dependent upon the mode of virus administration, but the minimum period required is usually 3 weeks. Spon- taneously-aborted foetuses are often autolyzed, which makes the presence of lesions difficult or impossible to observe. Occasionally, oedema of the subcutaneous tissues and excessive fluid in the pleural and peritoneal cavi- ties have been noted in aborted foals {Doll et al., 1957), piglets (Meyer, 1978) and calves (Gillespie et al., 1967; Casaro et al., 1971). In addition, the latter reports describe hyperplasia of reticulo-endothelium in spleen and lymph nodes in aborted calf foetuses.
Foetuses, which die in utero in early pregnancy, may become mummi- fied due to the resorption of fluid. In this case, the uterus is probably not properly stimulated to cause regression of the corpus luteum (Kendrick, 1971) and the conceptus consequently remains in utero. Mummies have been born from sows, cows and ewes after infection with pestiviruses.
Stillbirth is common after pestivirus infections, but seems to be an infre- quent complication of congenital rubella (Dudgeon, 1975). Pregnant sows infected with HCV seem to be more prone to delivering stillborn progeny than to producing aborted foetuses, whereas the reverse holds true for the ovine and bovine species after BDV and BVDV infections, respectively. Stillbirth is usually the consequence of foetal death in mid- and in the last third of pregnancy. In pigs, stillbirth occurs after maternal infection during the last third of gestation (Cowart and Morehouse, 1967; Meyer, 1978). At post-mortem examination, subcutaneous oedema, hydrops ascites and hydro- thorax are the most pronounced lesions in stillborn pigs. These excessive ac- cumulations of fluids are often associated with mottling or pitting of the liver or kidney (Fig. 1) (Cowart and Morehouse, 1967; Johnson et al., 1974; van Oirschot, 1979a). The mechanisms responsible for these lesions still re- main unexplained.
As a rule, prenatal death due to congenital nonarbo togavirus infections is more common when the maternal infection takes place earlier during gesta- tion.
Teratogenic effects
RUV, BVDV, BDV and HCV can act as teratogenic agents and when the foetus survives the initial infection a whole spectrum of abnormalities can develop. Manifestations of such deviant foetal development are malforma- tions, generalized or localized growth retardation, pathological lesions, which are not always clinically obvious, and more subtle biochemical or functional defects. In this context the term "malformat ion" is confined to cases where there are gross anatomical defects of one or more body components.
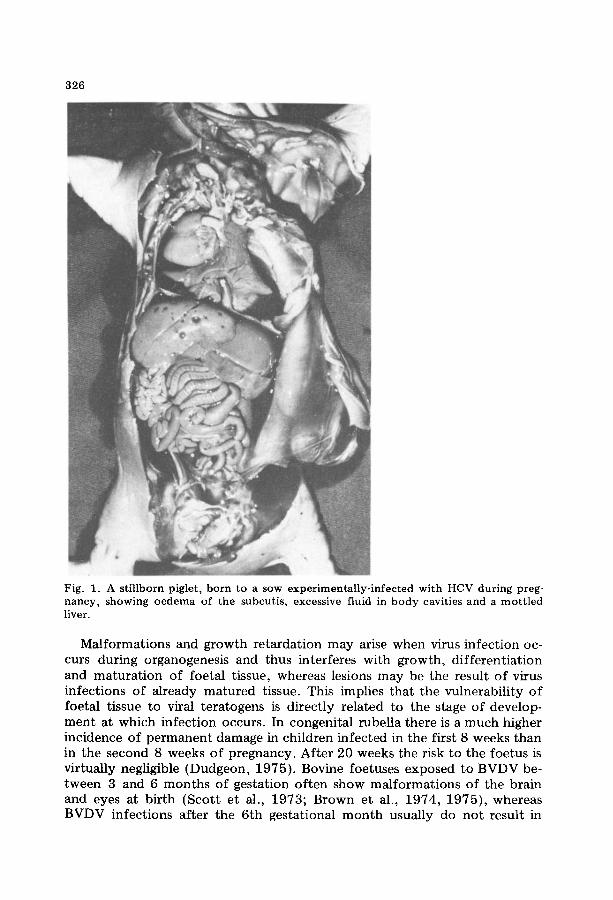
326
Fig. 1. A stillborn piglet, born to a sow experimentally-infected with HCV during preg- nancy, showing oedema of the subcutis, excessive fluid in body cavities and a mottled liver.
Ma l fo rma t ions and g rowth r e t a rda t ion m a y arise when virus in fec t ion oc- curs dur ing organogenes is and thus in terferes wi th g rowth , d i f f e ren t i a t ion and m a t u r a t i o n of foe ta l t issue, whereas lesions m a y be the resul t o f virus infec t ions o f a l ready m a t u r e d tissue. This implies t ha t the vulnerabi l i ty o f foeta l t issue to viral t e ra togens is d i rec t ly re la ted to the stage o f develop- m e n t a t which in fec t ion occurs . In congeni ta l rubel la the re is a m u c h higher incidence o f p e r m a n e n t d a m a g e in chi ldren in fec ted in the first 8 weeks t h a n in the second 8 weeks o f p regnancy . Af t e r 20 weeks the risk to the foe tus is vir tual ly negligible (Dudgeon , 1975) . Bovine foetuses exposed to B V D V be- tween 3 and 6 m o n t h s of ges ta t ion o f ten show m a l f o r m a t i o n s o f the bra in and eyes at b i r th (Sco t t e t al., 1973; Brown et al., 1974, 1975) , whereas BVDV infec t ions a f te r the 6 th ges ta t ional m o n t h usual ly do no t resul t in

327
foe t a l a b n o r m a l i t i e s (Casaro e t al. , 1 9 7 1 ; K e n d r i c k , 1 9 7 1 ) . By e x p e r i m e n t a l - ly m a n i p u l a t i n g t h e t i m i n g o f B D V i n f e c t i o n d u r i n g p r e g n a n c y , n e u r o l o g i c a l a n d c u t a n e o u s l es ions c o u l d be s u p p r e s s e d a n d a r te r ia l l e s ions i n t r o d u c e d ( Z a k a r i a n e t al. , 1 9 7 5 ; D e r b y s h i r e a n d Ba r low , 1 9 7 6 ) . This m a y be i n d i c a t i v e o f t he c h a n g i n g ava i l ab i l i t y or s u s c e p t i b i l i t y o f t a r g e t cells as foe t a l t i s sue d i f f e r e n t i a t e s a n d m a t u r e s .
T h e n o n a r b o togav i ruses i n d u c e a va r iab le p a t t e r n o f foe ta l d i sorders . H o w e v e r , t h e de fec t s p r o d u c e d in t he r e spec t ive hos t s have several charac- te r i s t i cs in c o m m o n ( T a b l e I).
TABLE I
Unity and diversity in clinico-pathological features of congenital nonarbo togavirus infections
Hog Bovine Border Rubella cholera virus disease
diarrhoea
Abortion + ++ ++ + Foetal mummification + + + - Stillbirth ++ + + +
Intrauterine growth retardation - + ++ ++ Selective organ stunting + + + + Cerebral malformations + + + + Cerebellar hypoplasia + + + - Dysmyelination + + ++ + Chorioretinopathy - + - ++ Cataracts - + - + Skeletal defects + + + + Arthrogryposis + - + - Skin abnormalities + + ++ +
Neonatal morbidity and mortality ++ ++ ++ ++
Clinically inapparent persistently- + + + + infected individuals
++ = frequently occurring; + = recorded; - = not (yet) recorded.
I n t r a u t e r i n e g r o w t h r e t a r d a t i o n ( I U G R ) is a c o m m o n f e a t u r e o f c o n g e n i - ta l B V D V , BDV a n d R U V i n f e c t i o n s . I t m a y be d e f i n e d as h a v i n g t o o low a b i r t h w e i g h t for t h e g e s t a t i o n a l age. Se lec t ive o r ga n s t u n t i n g c a n also occu r . B D - a f f e c t e d l a m b s f r e q u e n t l y have a b o d y w e i g h t a n d c r o w n m a n u s l e n g t h b e l o w t h e n o r m a l r ange , t h e y possess f i ne r l o n g b o n e s (Te r l eck i e t al., 1 9 7 3 ) a n d have s i g n i f i c a n t l y r e d u c e d visceral o rgan a n d t h y m u s w e i gh t s ( R i c h a r d - s o n , 1 9 7 7 ) . L o w b i r t h we igh t is also very c o m m o n in c o n g e n i t a l r u b e l l a ( F o r r e s t a n d Mense r , 1975}. A s i g n i f i c a n t I U G R , d e f i n e d as h a v i n g a b i r t h w e i g h t b e l o w the 1 0 t h p e r c e n t i l e , was n o t e d in 59% of 92 i n f a n t s s t u d i e d ( D e s m o n d e t al. , 1 9 6 7 ) . In genera l , severe I U G R is o b s e r v e d in c h i l d r e n w i t h m u l t i p l e de fec t s , b u t i t is s e l d o m the sole m a n i f e s t a t i o n o f c o n g e n i t a l r u b e l l a

328
(Lejarraga and Peckham, 1974). In-utero infected children can have signifi- cantly lower organ weights than healthy children (Naeye and Blanc, 1965). Newborn calves, whose dams were exposed to BVDV at 100 days of preg- nancy showed evidence of IUGR (Done et al., 1980). The thymus weights were also significantly decreased. Kendrick (1975 ) r epo r t ed a reduction in foetal lung size after in utero BVDV infection. In congenital HC general IUGR has not been recorded, but a selective growth retardation of the lung has been observed (Young et al., 1955; Emerson and Delez, 1965).
Congenital nonarbo togavirus infections frequently interfere with the de- velopment of the central nervous system (CNS).
Intracranial malformations have been reported in association with pre- natal infection with pestiviruses. Hydranencephaly, which is defined as com- plete or near~omplete absence of cerebral hemispheres, with the space filled with cerebrospinal fluid, surrounded by thin, membraneous cerebral tissue (Cho and Leipold, 1977), was observed in lambs with apparently normal heads, whose dams were exposed to BDV at 54 days of gestation (Barlow, 1980). Hydranencephalic calves were born during field outbreaks of BVD (Markson et al., 1976; Axthelm et al., 1981; Badman et al., 1981) and after experimental BVDV infection (Done et al., 1980). Porencephalic cavitations of cerebrum and cerebellum can develop in foetal lambs infected with BDV (Barlow, 1980) and calves exposed to BVDV in utero (Markson et al., 1976; Axthelm et al., 1981).
Brain malformations are relatively uncommon in congenital rubella and HC, and are far overshadowed by retardation in growth and maturation of the brain, that may eventually lead to microencephaly (Rorke, 1973; John- son et al., 1974). Also in BD and BVD a decreased brain weight is a consistent morphological defect (Scott et al., 1973; Terlecki et al., 1973; Sweasey et al., 1979; Done et al., 1980).
Cerebellar hypoplasia is the most common brain defect in prenatal HCV and BVDV infections. It is often associated with congenital t remor in the newborn. Varying degrees of cerebellar hypoplasia can be induced by inocu- lating pregnant sows with field (Done and Harding, 1969; Vannier et al., 1981) or vaccine strains of HCV (Emerson and Delez, 1965; Johnson et al., 1974). The defect appears more severe the earlier sows are infected during gestation. After experimental infection of cattle with BVDV between 100 and 150 days of gestation cerebellar hypoplasia of the foetus can occur (Brown et al., 1973, 1974; Done et al., 1980). This cerebellar defect has also been noted in field outbreaks (Markson et al., 1976; Badman et al., 1981). Cerebellar hypoplasia is also part of the BD syndrome (Barlow, 1980). Hypoplasia refers to a lack of development, wi thout indicating how this arises. In BVD it appears to be the result of destruction of tissue formed previously rather than impaired cerebellar development. Severe folial oedema in conjunction with necrosis of cells in the external germinal layer appears to cause the cerebellar hypoplasia (Brown et al., 1974).
Dysmyelination Or hypomyelinogenesis is a disorder frequently seen after congenital nonarbo togavirus infections. In BDV infected lambs dysmyelina- tion of the CNS is a characteristic feature, and it may lead to congenital

329
tremor in the newborn lamb. The congenital defect is present in varying de- grees in the brain and spinal cord, the latter being constantly the most se- verely affected (Hughes et al., 1959). Histopathologically, the anomaly is characterized by a reduced affinity for myelin-specific stains, aggregation of interfascicular lipid droplets and the presence of numerous neuroglial cells, many of them abnormal (Patterson, 1975). Biochemically, there is evidence of a disturbed lipid metabolism and an enhanced macrophage enzyme activ- ity, which suggests a process of myelin degeneration (Storey and Barlow, 1972). Other studies suggest that BDV affects the precursors of myelin- forming oligodendrocytes and thus results in reduced myelin formation (Barlow and Storey, 1977). The above data support the concept that the dysmyelination is due to impaired myelination associated with concurrent destruction of newly formed myelin (Patterson, 1975; Terlecki, 1977). Re- cently, defective myelination of the spinal cord was demonstrated neuro- chemically in 8 calves prenatally-infected with BVDV (Done et al., 1980). Congenital t remor of pigs resulting from transplacental infection with HCV is, among other things, characterized by dysmyelination of the CNS. Again, the spinal cord shows the most severe disturbance of myelination, whereas the peripheral nerves are not significantly affected (Emerson and Delez, 1965; Done and Harding, 1969). Retardation of myelination of the CNS, in some case with a decrease in oligodendroglia, has also been described in con- genital rubella (Naeye and Blanc, 1965; Rorke and Spiro, 1967).
The developing eye appears to be very susceptible to the teratogenic ac- tion of RUV, and BVDV also can induce eye anomalies. Eye defects do not seem to have been described in congenital BDV and HCV infections. The most common ocular defect in congenital rubella is the typical pigmentary chorioret inopathy, which rarely impairs vision (Forrest and Menser, 1975). Infection of pregnant cows with BVDV at 150 days, but not between 65 and 115 days of pregnancy, can also lead to focal or total ret inopathy in the foetus. In the acute stage inflammation and necrosis of the retina are seen (Brown et al., 1975). Cataracts are frequently encountered in congenital rubella, usually involving both eyes (Dudgeon, 1976), and are often associ- ated with microphthalmos. Calves congenitally-infected with BVDV can also develop cataracts and microphthalmos (Bistner et al., 1970; Scott et al., 1973). However, the cataracts induced by the 2 infections seem to be histo- logically distinct (Bistner et al., 1970). Other ocular defects are less common than ret inopathy and cataracts. Severe eye defects may result in blindness.
In BD, BVD and rubella, abnormalities of the skeleton have been re- ported. These are skeletal growth arrest lines in BD, indicative of disturbed osteogenesis (Terlecki et al., 1973). Calves that died in utero after maternal BVDV infection showed growth arrest lines in long bones {Done et al., 1980). BD lambs possess finer limb bones and exhibit skull deformities. The bone of the skull is radiographically less dense (Terlecki et al., 1973}. De- formed heads have also been noted in congenital HC (Sautter et al., 1953) and BVD (Scott et al., 1973). Bone lesions in congenital rubella consisted of a large anterior fontanelle, long bone changes and demineralization of

330
the skull bones. There is a defect in bone formation, involving principally the laying down and calcification of osteoid (Rudolph et al., 1965). It is not clear whether the bone changes are secondary to a metabolic or nutritional disturbance or are a direct effect of viral infection of the bone, as suggested by Heggie (1977),
Defects in distal front legs, suggestive of arthrogryposis, have been ob- served in piglets exposed in utero to HCV (Emerson and Delez, 1965; Johnson et al., 1974). Arthrogryposis also appears as a rare sequel of a foetal BDV infection (Vantsis et al., 1980).
Skin abnormalities occur following congenital nonarbo togavirus infec- tions. A characteristic feature of BD is an abnormally hairy birthcoat, which is the result of an enlargement of primary follicles (Derbyshire and Barlow, 1976}. Alopecia may be the consequence of transplacental BVDV and HCV infections (Kendrick, 1971; Stewart et al., 1972). Achs et al. (1966} found dermatoglyphic abnormalities in children with severe congenital rubella. Some infants infected in utero with RUV show a rash at birth (Lindquist et al., 1965; Plotkin et al., 1965b).
The nonarbo togaviruses can cause additional foetal abnormalities. A consistent feature of congenital rubella is its multiplicity of defects, the only defects that commonly occur alone being deafness and retinopathy. Deafness is the most common defect in infants with congenital rubella, and it is usually of the sensoneurinal type (Forrest and Menser, 1975). Deafness is often not detected at birth, and in some cases it may deteriorate after birth (Peckham, 1972). Other major manifestations of the rubella syndrome are the structural cardiovascular defects, which mainly include persistent ductus arteriosus, pulmonary artery stenosis and ventricular septum defects (Dudgeon, 1976). The teratogenicity of RUV can cause numerous other anomalies and these are listed in several review articles (Dudgeon, 1976; Charles, 1980).
In addition to malformations nonarbo togaviruses can produce lesions in the foetus. Lesions are the result of infection of already fully-formed tis- sue, and the foetal immune response may contribute to their development. Various lesions have been observed after congenital infections with nonarbo togaviruses, among which are hepatitis and thrombocytopenia purpura in rubella (Dudgeon, 1975), hepatitis and petechiation in HC (Von Benten et al., 1980), periarteritis in BD (Gardiner et al., 1980) and mouth lesions in BVD (Scott et al., 1973).
Neonatal disease
After in utero infection with nonarbo togaviruses the clinical picture covers a broad spectrum of illness ranging from an apparently normal new- born to moribund individuals with extensive lesions or malformations. The affected newborn may die or recover from the disease. Neonatal mortal i ty is relatively high. Most of the prenatally-infected piglets succumb in the first days or weeks after birth, and BVD- or BD-affected offspring also have a

331
low viability. The mortali ty figures of children with congenital rubella vary considerably. When severe disease occurs in combinat ion with malformations it can lead to a high rate of death in early infancy (Cooper et al., 1965; Forrest and Menser, 1975).
Since nonarbo togavirus infections are widely disseminated throughout the foetus, the symptoms in the neonate may involve virtually every organ, either transiently, progressively or permanently. Low birth weight is a fairly constant feature in congenital rubella, BVD and BD and it is frequently as- sociated with a failure to thrive postnatally. Epidemiological studies show an abnormal incidence of long-term growth retardation in infants and adults, who have had congenital rubella (Menser et al., 1967a; Hardy, 1976). Lambs with BD are usually recognized in infected flocks by being small and s tocky, with a poor growth rate (Terlecki, 1977). Clinical manifestations of CNS disorders are common in the neonatal period. In BD, congenital t remor is one of the main characteristics, and is associated with dysmyelination. Af- fected lambs show a slight muscular t remor to rhythmic shaking movements; symptoms which may gradually subside in the course of the first months (Westbury et al., 1979; Terpstra, 1981). Congenital t remor has also fre- quently been noted after in utero HCV infection (Emerson and Delez, 1965; Done and Harding, 1969; Vannier et al., 1981). Calves with prenatally ac- quired BVD also can suffer from body tremor (Done et al., 1980; Binkhorst et al., 1983). Other nervous signs in pestivirus-infected animals include in- ability to rise and suck, ataxia and abnormal stance (Emerson and Delez, 1965; Terlecki, 1977; Done et al., 1980). The spectrum of neurological and neurosensory involvement in infants with congenital rubella is wide. These include lethargy, motor paresis, restlessness and developmental delay. Patho- logical changes in the CNS are vasculitis, leptomeningitis and encephalitis (Desmond et al., 1967; Rorke, !973). In addition to nervous signs and mal- formations numerous other symptoms of congenital rubella may be present in the newborn, notably thrombocytopenia purpura and hepatitis, frequent- ly with jaundice. These neonatal lesions often are transient. Other neonatal manifestations of congenital rubella are listed in several articles (Dudgeon, 1975; Forrest and Menser, 1975; Charles, 1980).
In addition to the signs referred to above, BD-affected lambs frequently show an abnormally hairy birthcoat. Many lambs with BD die shortly after birth, but they can survive for more than 100 days (Gard et al., 1976). At necropsy there may be evidence of intercurrent disease, particularly pneu- monia (Terlecki, 1977). Haemorrhages of skin and internal organs are the most pronounced lesions at necropsy in piglets which died in the neonatal period (Wrathall, 1975; Van Oirschot, 1979a; Meyer et al., 1981).
It is also possible that after prenatal nonarbo togavirus infections appa- rently healthy individuals may be born. Later such individuals either develop clinical signs or remain normal.
Late onset disease and long-term sequelae
An in utero infection may follow an asymptomatic course in the neo-

332
natal period, and clinical signs may not appear until the progeny are several months old. In the case of rubella, foetal infection after the first trimester of pregnancy is more subtle in its manifestations than infection during early pregnancy. Such children often appear normal at birth, but lesions continue to evolve and can give rise to disease months or years after delivery. Thus, rubella acquired in utero during the latter half of pregnancy produced active postnatal disease, including pneumonia and encephalitis, from 3 months of age (Marshall, 1973). Deafness, mental retardation, heart and blood vessel defects and other dysfunctions may not be recognized before the child enters school. Peckham (1972) described children with congenital rubella, who were apparently normal at 2 years of age, but were profoundly deaf by the age of 7 years. Another long-term effect is a poor intellectual per- formance. Children experience problems in perception, memory, sequencing and recall, which interferes with normal functioning. Such long,term seque- lae may not be uncommon (Hardy, 1976). Congenital rubella has been im- plicated in the evolvement of endocrinopathies like diabetes mellitus (Menser
1976; Floret et al., 1980) and growth hormone deficiency (Preece et al., 1977). A, probably uncommon, late effect has been recognized as "progres- sive rubella panencephalitis". It is a slowly progressive and fatal CNS dis- order of children in their second decade o f life that is characterized byh igh rubella ant ibody titres in the cerebrospinal fluid and the presence of virus in brain tissue (Townsend et al., 1975; Weil et al., 1975).
Swine exposed to HCV in utero, under natural conditions, looked healthy during the first months of life, but they invariably developed a runting-like syndrome, characterized by conjunctivitis, dermatitis, leucopenia and loco- mot ion disturbances and eventually died (Van Oirschot and Terpstra, 1977). This "late onset disease" could be reproduced experimentally, however, the infected piglets commenced to show symptoms 3 to 7 weeks after birth (Van Oirschot, 1979a; Meyer et al., 1981). These congenital, persistent HCV infections induced severe depletion of lymphocytes in the thymus and peri- pheral lymphoid organs and proliferation and reticuloendothelial tissue (Hermanns et al., 1981; Van der Molen ar, d VarrOirsc,hot, t981a) . I r rad- dition, vascular lesions, especially hydropic degeneration of endothelium, were a consistent finding in these pigs (Van der Molen and Van Oirschot, 1981b).
In contrast with congenital HC, prenatally-infected sheep and cattle that are born apparently healthy, may remain normal for years or even during their whole lifetime, in spite of the lifelong virus persistence in these ani- mals (Coria and McClurkin, 1978; Westbury et al., 1979; Terpstra, 1981, Strayer e ta l : ,~983) : -
Conceptuses congenitally-infected with BDV (Terpstra, 1981), BVDV (Braun et al., 1973), RUV (Phillips et al., 1965; Hardy et al., 1969) or HCV (Van Oirschot, 1979a) can eliminate the virus infection during the prenatal or newborn period and consequently grow up healthy.

3 3 3
Infections of other host species
Besides their "natural" host the nonarbo togaviruses can also infect and subsequently cross the placenta of other animal species (Table II).
Some strains of BVDV are highly pathogenic for the sheep foetus. After experimental infection of ewes by the intramuscular or intravenous route BVDV is capable of causing early embryonic death, abortion, foetal mummi- fication, stillbirth, or foetal malformations such as IUGR, dysmyelination, hydranencephaly and cerebellar hypoplasia. Lambs with signs typical for BD, or healthy lambs with neutralizing ant ibody in their pre-colostral serum, can occasionally be born (Ward, 1971; Snowdon et al., 1975; Parsonson et al., 1979; Barlow et al., 1980a; Terlecki et al., 1980). These BVDV infections caused a high rate of foetal mortal i ty in sheep.
T A B L E I I
C o n g e n i t a l n o n a r b o t o g a v i r u s i n f e c t i o n s o f s e ve r a l h o s t s p e c i e s
H o g B o v i n e v i ru s B o r d e r
c h o l e r a d i a r r h o e a d i s e a se
R u b e l l a
S w i n e + + ±b _
C a t t l e _a + ± _ S h e e p - + + - M a n - - - +
a N o t ( y e t ) r e c o r d e d . b F o e t a l d e f e c t s r e c o r d e d , b u t f o e t a l i n f e c t i o n n o t c o n f i r m e d .
There is circumstantial evidence that in utero infection of pigs with BVDV occurs naturally (Stewart et al., 1971; Fernelius et al., 1973a). After experi- mental infection by the oronasal route of 20 pregnant gilts, BVDV could be isolated from the foetuses of only one gilt. The foetuses showed nonsuppu- rative meningitis and chorioditis (Stewart et al., 1980). Thus, BVDV can cause congenital infections in swine, although it is probably not common.
Some observations indicate that BDV may infect calf, pig and goat foetu- ses. After intramuscular inoculation of 10 pregnant heifers at 50--51 days of gestation, 6 of the cows aborted and 3 showed evidence of foetal death. All foetuses were severely retarded in growth and 3 had cerebellar malforma- tions (Gibbons et al., 1974). Nine out of 19 piglets born to 3 sows inocu- lated intramuscularly with BDV at 34 days of pregnancy exhibited reduced cerebellum weights in the terminal stages of gestation (Wrathall et al., 1978). After inoculation of pregnant goats with BDV a placentitis devel- oped, foetal death and abort ion occurred, and some newborn kids showed neurological signs (Huck, 1973; Barlow et al., 1975). Regrettably, in these studies there is no conclusive evidence that BDV caused the congenital ab- normalities in calves, pigs and goats, because no mention is made of at- tempts to confirm foetal infection.

334
The author is unaware of any reports regarding the inoculation of preg- nant cattle or sheep with HCV, although HCV appears to propagate in calves, sheep and goats (Loan and Storm, 1968).
Thus under experimental conditions BVDV and BDV may cause congeni- tal infections in "non-natural" hosts. It may be assumed that such infections occasionally occur in the field, since BVDV can be transmitted to sheep by contact with infected cattle (French et al., 1974) and BD-affected lambs can be sources of infection for cattle (Barlow et al., 1980b) and pigs (Terpstra, 1981).
Unfortunately, there is no experimental animal model suitable for study- ing congenital rubella. Experimental vertical transmission of RUV has been achieved in monkeys (Parkman et al., 1965), rats (Cotlier et al., 1968), fer- rets (Rorke et al., 1968), rabbits (Kono et al., 1969) and mice (Matsuo, 1975). Occasionally, foetal defects and pathological changes, similar to those observed in humans, have been found in animal foetuses (Cotlier et al., 1968; Kono et al., 1969). However, none of these positive teratogenic ani- mal studies has been confirmed by other investigators.
PATHOGENESIS
Route o f infection
When a pregnant animal becomes infected, the virus can reach the foetus in a number of ways: (1) it can spread haematogenously and cross the pla- centa, (2) the virus can enter via the genital mucosa and pass the amnion, (3) the gamete in the ovary can be infected.
In nonarbo togavirus infections the transplacental route is the most im- portant. Cows bred by a persistently-infected bull, whose semen contained BVDV gave birth to uninfected normal calves (McClurkin et al., 1979). Con- versely, it was possible to produce BD-affected lambs by artificial insemina- tion of ewes with infected semen and by natural mating with BD rams (Gardiner and Barlow, 1981}. In addition, BDV antigen has been demon- strated in the germinal cells of the ovaries (Gardiner, 1980), which provides opportunities for transovarian passage to the foetus. However, there is no evidence that these modes of infection do occur under natural conditions. After a nonarbo togavirus infection of a susceptible pregnant mammal the virus disseminates by way of the bloodstream to the placenta and subse- quently to the foetus. This maternal infection usually runs a subclinical course. Immuni ty of the dam gained by prior infection Or vaccination pro- tects the foetus from becoming infected, but exceptions have been docu- mented in HC (Stewart et al., 1972), BD (Barlow et al., 1980a) and rubella (Eilard and Strannegard, 1974).
Placental infection
Little is known on the course of infection in the placenta, which presents a physical barrier to the transmission of viruses.

335
The histological structure of the placenta varies in different mammals. In the human haemochorial placenta the maternal blood is in direct contact with the chorion epithelium and is separated from foetal blood by 3 tissue layers. In the epitheliochorial placenta of ruminants and swine 6 tissue layers separate maternal from foetal blood. In addition, there are differences in placental structure at various stages of gestation.
Theoretically, a virus can leak across, be carried across or grow across the placenta. BDV (French et al., 1974) and BVDV (Snowdon et al., 1975) re- plicate to high titres in placentomes of sheep and RUV (Alford et al., 1964) and HCV (Van Oirschot, 1980) are regularly isolated from placental tissue. Thus, these viruses appear to grow across the placental barrier. The transmis- sion is of ten associated with the product ion of lesions in the placenta.
Histological work of TSndury (1975) on infected placentae and foetuses obtained by therapeutic interruptions of pregnancy after maternal rubella shows that necrosis occurs in the epithelial cells of the chorion and in the endothel ium of the chorionic vessels, the latter lesions being most promi- nent. Frequently, thrombi develop in the vascular areas, jDamaged endo- thelial cells are desquamated and consequently emboli are found in the branches of the umbilical vein. Emboli become fastened to a foetal vessel wall and induce foci of damage in the endothelium of arteries in various organs, resembling those in the chorionic vessels. These lesions are the first to be observed in the foetus. Thus, these observations suggest that RUV se- q u e n t i a t t y ~ f e c t s epithelium and endothelium of the chorion and spreads to the foe tus by way of infectecl emboii, consistihg o f necrotic vascular endothelial cells.
In cattle and sheep the mode of infection with pestiviruses is probably through the placentome, which comprises a foetal portion, the cotyledons, and a maternal portion, the caruncles. Pregnant ewes inoculated with BDV developed lesions, consisting of necrosis o f the placentomal septa, :within 10 days of exposure. The lesions either remained small and focal and showed a tendency to heal or extended to diffuse necrosis. The lesions apparently commenced in the maternal capillary walls with endothelial swelling and oc- clusion of the vessels; in a later stage the epithelium became involved. Ulti- mately, the cellular debris was discharged into the foeto--maternal space and ~ppare~tty ingested by the trophobl~st (Barlow, 1972). Af t e r experimental infection of sheep with strains of BVDV, thrombi in small b lood vessels or focal areas of haemorrhage and necrosis were seen at the junction of the foetal and maternal component of the placentome (Snowdon et al., 1975; Parsonson et al., 1979). These histological data suggest a passage of pesti- viruses to foetal tissue within cellular debris. However, other studies demon- strate that_BDV and BVDV can replicate in the placentome of sheep and cattle, respectively, wi thout producing lesions and yet readily cross the placenta (Casaro et al., 1971; French et al., 1974). Thus, it seems that detritus-free virus also may pass the placental barrier and infect the foetus.
A congenital infection with HCV may result in the farrowing of a com- pletely or a partially-infected litter. There is evidence that may indicate

336
that HCV passes the placenta at one or more sites and subsequently spreads from foetus to foetus (Van Oirschot, 1980) . Obviously, this does not ex- clude the possibility that all pig foetuses may also become infected simul- taneously.
A placental infection does not necessarily lead to a foetal infection, as in- dicated by a higher percentage of RUV (Alford et al., 1964), and BDV (French et al., 1974) isolations from placentae than from corresponding foetuses, The factors determining whether infection remains ccnfLucd to p ~ n ~ m l : ~ - 6 r - s p ~ d ~ t 6 ~ e f o ~ t ~ ar~ n~t well tmd~rsmod,(bu~ appear to be-related-to a certain.extent,-to gestationa! age~In B:~ mud BD thereis limited ~ c e t h a t : t h e : : ~ s = d o e s - n u t ~ v a ~ ; t h e ~ , , ~ , v . p ~ a ~ , ~ in the first month of pregnancy (Manktelow et a l , 1969; French et al., 1974; Whitmore et al., 1981). A possible explanation is that close contact between t rophoblast and well-developed maternal villi is required for vertical transmisaion_~ occux, and~ucha,~hridge£ ' ~eve!op.~ around . 3 O A a ~ ~ nancy (Barlow, 1972; Kendrick, 1976). Essentially, the mammalian placenta seems to be as p e ~ e a b I e to nonarbo t0gavirus~ during the firs~ :as during the last third of pregnancy (Kendrick, 1971; French et al., 1974; Van Oirschot, 1979a; Cradock-Watson et ~ . , !980) .
Foetal infection
Once the xdrus gainsaccess to the foetus-it,car~ spread sys temicat !y-byway of the bloodstream, or from cell to cell by continuity. The virus finds a favourab~ environment fo r replication in ~ e m etabol ica l~ ~_ctl.vefoet~_A ~!!s undergO~mg rap~d[mitoSiS ~nd [~ffe~f i t ia t iom ~ e [ u l t i ~ t ~ 5utcOni~ 'ofcon- g e n i * ~ ~ n s ...... -rml .... ~ ~.~ . . . . . . . . . . ~ . c l s - ~ . n u m b e ~ ~ , such ~ : ~ e d e v e I o p m e n ~ ~ e t ~ the genotype of the host: and' t he foetal ability tO repair damage, which is generally very high (Hicks, 1954; Willis, 1962). The age of the foetus deter-
tance for the extent of foetal damage. In general, when infection takes place before the onset of immune competence the virus is widely dissemmated in the foetus. RUV has been isolated from many organs (Monif et al., 1965; Rawts e t ate, t 965 ) and HGV ~ t i g e n (VanOi~sehot andT e rps t ~ a ,1977 )and BDV antigen (Terpstra, 1981) have been found to be widespread in foetuses. C0ngenjt~ virus infections ~ o u n d or a f a r t h e o n ~ t 0f immune c o m p e t ? n c e are more likely to be controlled or eliminated, due t o the development of a specific immune response by the foetus, In general, the risk of foetal damage is higher, the earlier infection occurs during pregnancy. However, a congeni- tal infection, even in the first month, does not necessarily lead to foetal ab- normalities.-Thi~ is-sho-wn, in the-ca~e-o~f ru~el la .by t~e~bi r th-of~pparent ly normal infants, who excrete virus, born to mothers with a history of rubella during early pregnancy (Rawls, 1968). In addition, Woods et al. (1966) de- tected foci of infected cells in tissues that were histologically normal. In the

337
case of pestiviruses, the incidence of foetal infection is clearly higher than the incidence of foetal malformations or lesions.
The specific properties of a virus strain have effects on the course of a foetal infection. Virus strains vary in their teratogenic potential. Pregnant sheep inoculated with BDV strain BP-77 gave birth to lambs with hypo- myelinogenesis, whereas strain H-77 produced hydranencephaly and arthro- gryposis in the same breed of sheep (Barlow et al., 1979). There are HCV strains that induce a high rate of congenital tremors in piglets infected in utero (Done and Harding, 1969; Vannier et al., 1981), whereas other strains do not (Van Oirschot, 1980). The NADL strain of BVDV did not produce severe abnormalities in foetal calves older than 100 days (Kendrlck, 1971; Braun et al,, 1973), as opposed to the Holmes strain, that gave rise to cere- bellar hypoplasia and ocular lesions (Brown et al., 1973, 1974). Japanese strains of RUV seem to be less teratogenic than American strains (Kono et al., 1969). This diversity may perhaps be explained by differences in inter- feron production. Japanese strains induced high levels of interferon in human placental cultures, whereas strains derived from the U.S.A. in- duced low levels (Potter et al., 1973). The virulence of a virus strain may influence the outcome of a congenital infection. Piglets prenatally-infected with a moderate-virulent HCV strain were more severely affected and died earlier than piglets exposed in utero to a low-virulent HCV strain (Van Oir- schot, 1980). Attenuated RUV (Vaheri et al., 1972; Ebbins et al., 1973) and HCV vaccines (Bran et al., 1971; Tesmer et al., 1973) can also transplacental- ly infect the foetus, but the risk for the conceptuses is virtually negligible. That the dose of virus influences the outcome is illustrated by Richardson et al. (1976) in the case of BD. The progeny of ewes inoculated with a 100 X higher dose were more severely affected than the progeny of ewes that re- ceived the low dose.
There is evidence indicating that virus strain--host genotype interactions affect the character of disease in the offspring. While pregnant ewes of 2 breeds inoculated with the H-77 strain delivered typical BD lambs, ewes of 2 other breeds inoculated with the same virus, gave birth to lambs without signs characteristics of BD (Barlow et al., 1979). Congenital rubella has rare- ly been noted in Japan even after extensive epidemics (Kono et al., 1969). An explanation for this phenomenon may be that, genetically, the Japanese are less prone to congenital rubella. An increased incidence of HLA antigens A1 and A8 has been found in cases of congenital rubella, and these antigens are seldom encountered among Japanese (Honeyman et al., 1975). However, there is also suggestive evidence that Japanese strains are of low teratogenic- ity (Koni et al., 1969).
Teratogenesis
Nonarbo togaviruses can induce abortion. It may be the result of maternal disease or foetal damage, or a combination of both. The general impression is that it is principally the result of foetal infection, since maternal infections

338
are often asymptomatic. In addition, hyperthermia, as such, does not pro- duce abortion in sows (Omtvedt et al., I 9 7 I ) . Moreover, virus ~ d lesions are found in aborted foetuses which are not autolyzed (Doll et al., I957; Gillespie et al., 1967; Meyer, 1978; Terpstra, 1981). The vascular lesions in the placenta, as has been reported for BDV (Barlow, 1972), BVDV (Snowdon et al., 1975) and rubella (TSndury and Smith, 1966; Ornoy et at., 1973) may contribute to foetal death and expulsion by, for instance, impairing the nutrit ion of the foetus. Also mummification and stillbirth are caused by foetal infection rather than by maternal illness.
For malformations to be produced, infection has to occur during periods of organogenesis, so that virus replication can interfere with growth, matura- tion and differentiation of foetal tissue. Foetal organs that are defined at the time of infection may escape gross anatomical damage. In addition, the virus must damage the foetus without killing it, thus it seems necessary that the virus is of sufficiently low virulence. The persistence of nonarbo togaviruses in the conceptuses is also an important factor in teratogenesis.
The most likely mechanisms for the teratogenic effects of nonarbo toga- viruses are (1) an inhibition or distortion of normal cellular development, (2) cell destruction as a direct consequence of virus infection, and (3) the production of vascular lesions. It is surprising that the teratological mecha- nisms have been studied better in rubella than in the animal nonarbo toga- virus infections. Most data on the pathogenesis of congenital rubella had to be based on studies of spontaneously or therapeutically-aborted foetuses.
Human foetal cells, infected in vitro with RUV, showed a marked depres- sion in mitotic activity (Plotkin et ah, 1965a). Carrier cultures, derived from tissues of infected foetuses, were found to have a reduced growth rate and appeared to have a shortened lifespan (Rawls and Melnick, 1966). In such carrier-cultures the virus does not cause obvious morphological changes. It is passed from parent to daughter cells during successive mitoses and this may give rise to clones of infected cells. In addition, RUV-infected cells ap- peared to produce a soluble growth-inhibitor, which might in part, account for the reduced rate of cell growth (Plotkin and Vaheri, 1967}. Children with congenital HC, BDV and BD. These viruses also persist in foetal tissues and basis for this is a subnormal number of cells in body organs (Naeye and Blanc, 1965). This defect is probably due to the inhibition of multiplication of RUV-infected cells. Since RUV is present in clones and cells in various organs (Woods et al., 1966) it seems probable that the slower rate of cell division of infected cells could also account for the development of specific organ defects. This is in keeping with changes of malformed organs that were interpreted as interrupted, retarded or disorganized maturation of tissue (Singer et al., 1967). Presumably, the above mechanism also plays a role in congenital HC, BDV and BD. These viruses also persist in foetal tissues and IUGR and selective organ stunting are frequently observed in these diseases. The pestiviruses are of low cytopathogenici ty in vitro and persistently-in- fected cultures are readily established (Mengeling et al., 1970; Terpstra,

339
1978). Such carrier cultures show no morphological alterations and infec- tion is maintained by distribution of virus from mother to daughter cells (Mengeling et al., 1970). HCV-infected cells also have a slightly extended period of DNA synthesis (Pirtle and Mengeling, 1970). It is not known, whether the infection of thyroid and other internally-secreting organs, as shown in BD (Terpstra, 1978), contribute to the general growth retardation.
Another possible teratogenic mechanism is an alteration in the cell differ- entiation process. In congenital rubella the lens cataract may be partially due to a disturbed differentiation of lateral epithelial cells of the lens into lens fibre cells (TSndury and Smith, 1966). BDV seems to affect the differ- entiation of oligodendrocytes from their precursors, thus leading to reduced myelin formation (Barlow and Storey, 1977). The typical abnormally coarse birthcoat in BD lambs appears to be the result of abberant differentiation of primary follicles (Derbyshire and Barlow, 1976).
In the case of rubella, there is histological evidence that the cytolyt ic ac- tion of the virus is operative in the production of malformations. Necrosis and cellular damage, without inflammation and fibrosis, have been found in the heart, developing lens, inner ear, teeth and other tissues of foetuses ob- tained after therapeutic interruption of pregnancy (TSndury and Smith, 1966). It has been suggested that BVDV has an affinity for mitotically ac- tive cells and causes destruction of the external germinal layer and other layers of the cortex of the cerebellum of foetal calves (Brown et al., 1973). Casaro et al. {1971) considered necrosis in lung, brain and skin in calf foe- tuses less than 150 days of age as potential causes of teratological defects. The early phases of CNS abnormalities produced by BDV are also associated with necrosis of the germinal layers (Barlow, 1980). Hence, the cytolytic activity of nonarbo togaviruses for cells of particular susceptibility appears to play a role in teratogenesis.
Foetal defects may also be a consequence, in an indirect way, of vascular lesions. Vasculitis has been noted in congenital rubella (TSndury and Smith, 1966}, predominantly in the brain (Rorke and Spiro, 1967; Singer et al., 1967). In these studies, vasculitis was considered to be the initial lesion leading to necrosis of surrounding tissue. The congenital heart malformations may also be initiated by vascular lesions (Overall, 1972). Also in congenital HC (Emerson and Delez, 1965; Van der Molen and Van Oirschot, 1981b) and BVD (Brown et al., 1973) substantial vascular damage occurs. In this context , it is notewor thy that all nonarbo togaviruses of domestic animals and RUV show a predilection for the vascular endothelium. The vascular disturbance is probably initiated by degenerative changes of the endothelial cells. Damage of endothelial cells may lead to thrombus formation, that can detach and circulate as emboli, giving rise to generalized vasculitis. At the same time, the blood~lot t ing mechanism can be triggered, directly or in- directly, by the virus refection, which may result in disseminated intravas- cular coagulation. The brain is among the organs most frequently involved in the disseminated intravascular coagulation syndrome. This process, which

340
may become manifest by clinical signs such as haemorrhagic diathesis and thrombocytopenia p u r p u r a , occurs in congenital rubella (McKay and Margaretten, 1967). Haemorrhagic diathesis is also a common finding in intrauterine-infected piglets (Van Oirschot, 1979a; Hermanns et al., 1981). The thrombosis and vasculitis of small blood vessels can produce partial to complete occlusion of these vessels, which may result in organ hypoxia, in turn leading to organ hypoplasia or necrosis. Hypoxia has been shown to in- duce specific malformations in laboratory animals (Wilson, 1973). It has been suggested that the cerebellar hypoplasia in newborn pigs congenitally- infected with HCV results from hypoxia due to vasculitis (Emerson and Delez, 1965). The cerebellar hypoplasia in BVD appears to be due to loss of vascular integrity by vasculitis, leading to folial oedema that in turn causes the-severe cerebettardestruct ion (Brown et al~, t973~, tn ~xtbetta~ ";a~ cular lesions are associated with ischaemic damage and retardation in growth and maturation of the brain (Rorke, 1973). The brain malformations in BD appear not to originate from vascular lesions, though, occasionally capillary thromboses are seen within the lesions (Barlow, 1980).
Chromosomal abnormalities have been described in children with congeni- tal rubella (Nusbacher et al., 1967; Ansari and Mason, 1977), and in cells in- fected with RUV in vitro (Plotkin et al., 1965a). Others have failed to find chromosomal damage (Mellman et al., 1965). Also in the case of HC there are contradictory findings on the induction of cytogenetic alterations in cells infected in vitro (Pirtle and Woods, 1968; Mengeling et al., 1970). Theoreti- cally, chromosomal damage in differentiating cells may cause malformations. However, there is no convincing evidence at hand for virus-induced chromo- somal changes as a mechanism of teratogenesis.
It remains speculative whether maternal disease affects foetal develop- ment. The changes in the growth-zone of the long bones may reflect a re- duced overall growth due to severe disturbances in placental function. It is unlikely, that maternal fever contributes to foetal malformations in the pig, since exposure of pregnant sows to high environmental temperatures appears to be without effect on foetuses (Omtvedt et al., 1971). Conversely, hyper- thermia has teratogenic effects in other animals, e.g., sheep (Berry et al., 1977).
Lesions, instead of true malformations, are produced in already fully- formed foetal organs according to principles that also apply to animals after birth, except that repair and regeneration are more important and immuno- logical and inflammatory responses are less important in foetal life.
Virus persistence
In general, the nonarbo togaviruses persist in the foetus after congenital infection, a condition which usually continues into postnatal life. In HC, BD and BVD, the virus persistence can be lifelong. In congenital rubella, however, the virus usually disappears within 1 to 2 years after birth. The

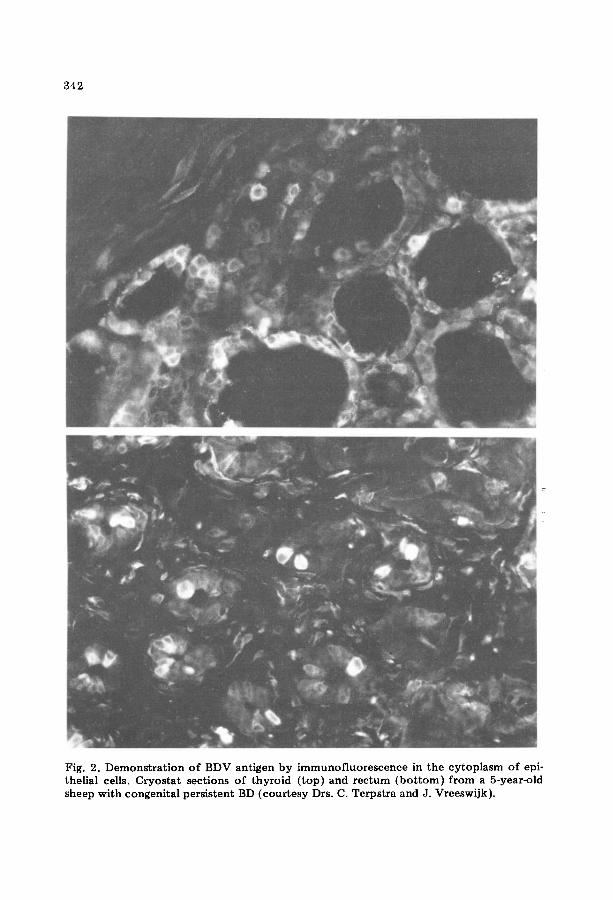
342
Fig. 2. Demonstration of BDV antigen by immunofluorescence in the cytoplasm of epi- theUal cells. Cryostat sections of thyroid ( top )and rectum (bo t tom)f rom a 5-year-old sheep with congenital persistent BD (courtesy Drs. C. Terpstra and J. Vreeswijk).

343
TABLE III
Factors o f (poss ib le ) importance in virus pers is tence in congeni ta l nonarho togav.iruses in fec t ions
Hog Bovine virus Border Rubella cholera diarrhoea disease
Foeta l age at t ime o f in fec t ion + + Low or no viral cytopathology + + Cell to cell spread + ? Little or no viral antigen in
cell membrane + ? Infec t ion of lymphoid tissue + + I m m u n e to lerance + + Defe c t s in i m m u n e system + ? Inadequate interferon response ? -
+ +
+ +
? +
?
+
? +
tively low or absent in foetuses exposed during the last tr imester (Casaro et al., 1971). In general, infants infected during the first third of pregnancy are more likely to shed virus for longer periods (Phillips et al., 1965; Cooper e t al., 1969) than children exposed during later stages (Hardy et al., 1969).
Several virus-specific factors have been proposed to be of importance in the initiation and maintenance of virus persistence. There is evidence that temperature-sensitive mutants (Williams et al., 1 9 8 1 ) a n d defective inter- fering particles are involved in the establishment of RUV persistence in cell culture (Norval, 1979). Cells infected with HCV appear to bear no, or very few, viral antigens on their surface, thereby allowing infected ceils to escape immunological attack (Van Oirschot, 1980). Most viruses that can produce persistent infections cause minimal cy topa thology (Mims, 1974) and this applies also to the nonarbo togaviruses. Many, if not all viruses that persist, infect lymphocytes or macrophages (Mims, 1974), and in this way elimina- t ion of virus by macrophages may be prevented. RUV (Simons and Jack, 1968) and HCV (Van Oirschot, 1979b) have been isolated from mononu- clear cells of the peripheral blood from persistently-infected infants and pigs, respectively. Cultures of mononuclear cells support the growth of RUV (Van der Logt et al., 1980}, BVDV (Truit t and Shechmeister, 1973; Atluru et al., 1979) and HCV (Korn and Lorenz, 1976; Van Oirschot, 1980). BDV has been isolated from thymus, spleen and lymph nodes (Terpstra, 1981). It is obvious that viruses which infect cells of the immune system may be able to induce defects in immunological functions. In fact, persistent nonarbo toga- virus infections are associated with specific and more generalized dysfunc- tions of the immune system. Such defects in immune response probably play an impor tant role in the establishment of nonarbo togavirus persistence.

344
IMMUNOLOGY
Effects on the immune system
Since the nonarbo togaviruses replicate in cells of the immune system, it is conceivable that they induce defects in the host defence mechanism, which in turn may favour virus persistence.
Congenital infections with RUV (Singer et al., 1969), BVDV (Braun et al., 1973) and HCV (Von Benten et al., 1980) may lead to precocious matura- tion of lymphoid tissue, i.e., germinal centre formation and plasma cell development, and this correlates well with elevations in serum levels of IgM and IgG. In infants with congenital rubella the IgM concentration may re- main above normal for the first 6 months of life (Cradock-Watson et al., 1976). Lambs, prenatally-infected with BVD have augmented IgG levels (Osburn et al., 1973). In congenital HC (Richter-l~eichhelm et al., 1980) and BVD (Braun et al., 1973; Brown et al., 1979) the elevated Ig levels have no or low specificity for the causative virus. These immunoglobulins may be directed to altered host tissue antigens {Brown et al., 1979) or they may be incomplete molecules of smaller size and low avidity (Richter-Reichhelm et al., 1980). In congenital rubella not all the extra IgM seems to be specific for RUV (Cradock-Watson et at., 1976),
There is evidence of lymphoid tissue damage in congenital infections with nonarbo togaviruses. Singer et al. (1969) observed a slight thymic involution in children with congenital rubella, who died in the first months after partu- rition. Lambs with BD have a significantly smaller thymus at birth than nor- mal lambs (Richardson et al., 1976; Sweasey et al., 1979). Necrosis of lymphoid tissue has been observed in congenitally-infected piglets, shortly after birth (Brack, 1971). In utero-infected pigs that survive for months after birth show severe morphological changes in lymphoid tissue. The most prom- inent lesions are an almost complete thymus atrophy, depletion and destruc- tion of lymphocytes , histiocyte hyperplasia with phagocytosis of lymphocyt ic debris and proliferation of reticulo~endothelial tissue (Hermanns, 1978; Van der Molen and Van Oirschot, 1981a). It is of interest to note that such pigs show a marked adrenal hyperptasia, and this may indicate an enhanced pro- duction of glucocorticosteroids. The adronncor~ical hormones might~ at i e ~ t partially, be responsible for the severe lymphocyte destruction in these per- sistently-infected pigs (Van der Molen and Van Oirschot, 1981a). In children with congenital rubella histiocytosis has also been observed (Claman et al., 1970).
Various abnormalities in humoral immunity can occur in congenital rubella. The most common aberration observed is a dysgammaglobulinaemia, with high IgM, low IgG, and low or undetectable IgA levels (Soothill et al., 1966; Singer et al., 1969; Stern and Forbes, 1975). The abnormality may be corrected as the child grows older, or the IgA deficiency may persist (South et al., 1975; Stern and Forbes, 1975). Transplacentally-infected piglets that

345
survive after birth appear to have a depressed IgG synthesis concomitant with an enhanced IgM production (Hermanns, 1978). In view of the thymic atrophy found in HC as well as in rubella, the abnormal IgG:IgM ratio may be caused by a disturbance in the switch from IgM to IgG, that appears to be under some degree of thymus or T cell control (Roitt , 1980).
The antibody response to antigens unrelated to the causative virus does not seem to be decreased in animals infected in utero with nonarbo toga- viruses (Van Oirschot, 1977; Coria and McClurkin, 1978; Terpstra, 1981). except in the terminal stages of congenital HC (Van Oirschot, 1979b). In infants with congenital rubella the antibody response to unrelated antigens appears to depend on the state of illness. Children with active disease are deficient in antibody production when stimulated with specific antigens (Michaels, 1969, 1972), but children, who no longer excrete virus, respond normally (White et al., 1968; Michaels, 1972).
There are conflicting reports regarding responsiveness of peripheral blood lymphocytes (PBL) from infants, which acquired rubella in utero, to phyto- haemagglutinin (PHA). Some authors found a hypo- or unresponsiveness of PBL to PHA (Montgomery et al., 1967; Olson et al., 1968), whereas others have described a normal lymphocyte proliferation to PHA (White et al., 1968; Marshall et al., 1970). In a more recent paper (Buimovici-Klein et al., 1979) the PBL response to PHA was correlated with the foetal age at the time of infection. Children, aged 1--13 years, who had been infected with RUV in the first 2 months of gestation, and having clinical manifestations, showed a more severely depressed PHA response than children exposed after the third month of gestation. In another study, the cutaneous response of RUV-infected children to vaccination with vaccinia virus did not appear to be impaired, but there was a diminished in vitro response of PBL to vac- cinia virus (White et al., 1968). Thus, in a number of children with congeni- tal rubeUa a hyporesponsiveness of the immune system can be demonstrated, the severity probably depending on the foetal age at the time of infection and on the state of illness of the infant. In fact, most children may pass through a state of immune deficiency, from which they may recover, others may manifest permanent dysfunctions. However, these immune deficiencies rarely become clinically apparent (Forrest and Menser, 1975). PBL from pigs with congenital HC respond normally to PHA and pokeweed mitogen, ex- cept a few days before death (Van Oirschot, 1977, 1979b). No information is at hand on the effect of congenital BVDV or BDV infections on cell-medi- ated immune (CMI) functions. There are too few data available to draw con- clusions on immune func t ion in congenital pestivirus infections, but the re- suits obtained suggest that, apart from the specific immune tolerance, there is a normal immune responsiveness.
The mechanism involved in the immune dysfunct ion in congenital rubella is poorly understood. The defect is often considered to be a direct terato- genic effect of RUV on the not fully differentiated immune apparatus, al-

346
though the presence of virus in PBL does not appear to be required for im- pairment of the immune response (Buimovici-Klein et al., 1979). The im- mune dysfunction may also be the result of an inhibition of macrophage function by RUV (Van der Logt et al., 1980) or of antigenic competi t ion (Chandra and Soothill, 1971; Gershon et al., 1974).
Another factor of possible importance in viral persistence is the interferon response. Desmyter et al. (1967) reported a defect in the interferon-pro- ducing capacity of cells derived from a t~UV-infected human foetus. More- over, cell cultures persistently infected with RUV are not sensitive to inter- feron, whereas acutely infected cells are. In addition, no interferon could be detectectin- serum or urine -of infants with congenital ~-abella,~y~-en i n those who were actively excreting virus (Desmyter et al., 1967; Rawls, 1967). Thus, an inadequate interferon response might be involved in persistence of RUV. In contrast, foetal calves infected in utero with BVDV at Day 95 or Days 149--150 of pregnancy produce interferon to levels comparable to those in adult bovines (Rinaldo et al., 1976), indicating that a defective interferon response is not a factor involved in BVDV persistence.
Immune mechanisms, among which is macrophage function, that normal- ly terminate infection in postnatal life are absent or not fully developed in foetal life. In addition, the developing immune system appears to be highly susceptible to virus-induced defects. It is likely that these are the most im- portant factors involved in the establishment of virus persistence in congeni- tal nonarbo togavirus infections.
I m m u n e response to congenital infection
In man, maternal antibody of the IgG class is transmitted through the haemochorial placenta after the first trimester of pregnancy. In contrast, the epitheliochorial placenta of domestic animals is not permeable for mater- nal immunoglobulins during the whole period of pregnancy (Brambell, 1970). Instead, maternal antibodies are transferred to the neonate in the colostrum of the dam. Thus, antibodies present in the newborn animal be- fore sucking have been formed by the foetus itself.
Man and domestic animals gain immunocompetence during foetal life. The onset of immune reactivity in man, calf, lamb and pig is around mid-gesta- tion (Solomon, 1971; Schultz, 1973; Bourne et al., 1974; Fahey and Morris, 1978), indicating that, basically, foetuses are capable of producing ant ibody to nonarbo togaviruses. The human foetus commonly responds to a RUV infection with the production of specific IgM antibody, irrespective of the time of prenatal infection (Cradock-Watson et al., 1980). This antibody synthesis may continue for 6 months after birth, a period which seems to correlate with the period of virus shedding, which suggests that IgM antibody production results from viral persistence. IgG ant ibody titres to RUV in- crease during the first 3 months of life, and this response may have started during the last weeks of pregnancy (Cradock-Watson et al., 1976).

347
In BD, BVD and HC the age of the foetus at the time of infection appears decisive as to whether or not a specific ant ibody response will be elicited. A foetal infection before the onset of immune competence does not usually result in the production of specific antibody, whereas an infection during the late stages of gestation can lead to ant ibody formation. Although domestic animals can respond immunologically around mid-gestation BVD neutralizing ant ibody cannot be detected until about 200 days of gestation (Braun et al., 1973; Brown et al., 1979). Porcine foetuses may only produce neutralizing ant ibody to HCV, when infected after the 90th day of preg- nancy (Van Oirschot, 1979b; Frey et al., 1980). It is not clear, why in BVDV and HCV, such a long period elapses between the onset of immune reactivity and appearance of viral specific antibody.
A foetal immune response to pestiviruses is usually followed by virus elimination from the host~ Such non-persistent infections become evident by the birth of animals with specific virus-neutralizing ant ibody in the pre- colostral serum and from which no virus can be recovered (Braun et al., 1973; Frey et al., 1980; Terpstra, 1981). In addition, the offspring of sows infected with HCV may be born viraemic, but shortly after birth the virus is no longer detectable {Van Oirschot, 1979a). In this case, an ongoing immune response in conjunction with the ingested colostral ant ibody may effect the virus disappearance. The mechanism, by which the virus infection is termi- nated before parturition is not completely understood, but it is likely that the foetal immune response plays the most important role in curing the in- fection in domestic animals. In congenital rubella, however, the passively- acquired maternal virus-specific IgG ant ibody and the foetal IgM ant ibody often fail to eliminate the infection. This results in RUV persistence despite the presence of circulating ant ibody (Alford et al., 1965; Cooper et al., 1965). The virus is passed from parent to daughter cell thus enabling the virus to persist in the presence of neutralizing antibody. The ant ibody may only serve to limit further dissemination of the virus via the circulation. Yet, virtually all infants with congenital rubella, ultimately, eliminate the virus.
It is a well documented phenomenon that exposure to antigens during early foetal life can lead to immune tolerance to that particular antigen. It was therefore surprising to find that foetuses exposed to RUV in early pregnancy are able to mount a specific ant ibody response to the virus. In contrast, domestic animals often do not respond with ant ibody synthesis after infec- tion with nonarbo togaviruses during early or mid-gestational life (Gard et al., 1976; Van Oirschot, 1977; Coria and McClurkin, 1978; Terpstra, 1981). One of several plausible explanations for this discrepancy might be the pres- ence of maternal virus-specific IgG in the RUV-infected conceptuses, which limits the spread of virus and consequently facilitates the production of anti- body by the foetus. Since there is no passage of maternal immunoglobulins in swine, cattle or sheep, virus can disseminate unlimitedly in foetal tissues, resulting in an excessive load of viral antigen, during the development of

348
the immune system, which may lead to the induction of immunological tolerance. In congenital pestivirus infections immunological tolerance ap- pears neither absolute nor permanent. A few persistently-infected pigs developed low levels of precipitating antibody or antibody detectable by the indirect immunofluorescence test (Van Oirschot, 1977; Van Oirschot, 1979b). Persistently-infected sheep can produce a transient or persistent level of neutralizing antibody, postnatally (Barlow et al., 1980a; Terpstra, 1981).
Immune tolerance, generally, can be the result of deletion, destruction, in- activation or dysfunct ion_of specific antigen-~eac32~e ~mphQcytes. It can re- side at the level of B or T cells or both, or it may be a disturbance of the im- munoregulation mechanism. With regard to nonarbo togaviruses, it has been s h ~ w ~ h a ~ lymph~cytes f rom caive~chronicaily-infec~ed wRh BVDV had a decreased ability to secrete immunogtobutins (Muscoptat et al., 1973). In addition, BVDV appears to depress in vitro plasma cell development and subsequent synthesis of IgG and IgM after pokeweed mitogen stimulation (Atluru et al., 1979). A possible explanation for the immunological tolerance is that pestiviruses may act by infecting pestivirus-specific lymphocytes and in that way interfere with the production of specific antibody. The obser- vation that immune tolerance is neither absolute nor permanent suggests that the unresponsiveness is not due to a complete deletion or destruction o l ives- reac t ive lympho cytes.
It is not known whether tolerance of the CMI response exists in congeni- tal nonarbo togavirus infections. A few reports have dealt with the CMI re- sponse to RUV antigens. Simons and Fitzgerald (1968) found that lympho- cytes from infants with congenital rubella showed blastogenesis in the pres- ence of viral antigens. In another report (Buimovici-Klein et al., 1979) the lymphocyte response to RUV was found to be absent or relatively low in children with congenital rubella, when examined years after birth. Immune interferon production by stimulated lymphocytes showed a similar pattern. In addition, there was no detectable production of leucocyte migration in- hibition factor by RUV-stimulated lymphocytes. These authors observed a more severe impairment of the CMI in children infected in the first 2 months than in the latter stages of pregnancy. Fuccfllo et al. (1974) described that lymphocytes of infants with congenital rubella failed to lyse RUV-infected non-autologous target cells. The above data suggest that a hypo- or unres- ponsiveness of CMI might exist in some children with congenital rubella. Although some work has been done with respect to HC (Van Oirschot, 1979b), it is not known whether animals congenitally-infected with pesti- viruses mount a CMI response to the causative virus.
The specific unresponsiveness of the humoral immune system in swine, sheep and cattle after congenital pestivirus infection appears to be only par- tially responsible for the lifelong virus persistence. In vitro experiments have shown that HCV can spread from cell to cell through intercellular bridges (Pirtle, 1969b), and from mother to daughter cell in the presence of HC ira-

349
mune serum (Pirtle, 1969a). Ingestion of high-titre colostral ant ibody only transiently suppressed viraemia in pigs persistently-infected with HCV (Van Oirschot, 1979a). A lamb persistently-infected with BDV developed neutral- izing ant ibody at the age of 14 months, but the virus was still widely dis- seminated throughout the tissues when it was killed 7 months later (Terpstra, 1981). In congenital rubella the virus persists despite the presence of neutral- izing ant ibody (Cooper et al., 1965) and RUV carrier-cultures could not be cured by rubella ant ibody (Rawls and Melnick, 1966). Thus, neutralizing ant ibody by itself appears to be incapable of curing the virus infection. This is in keeping with the concept of Notkins (1974) that CMI plays a major role in terminating virus infections that spread from cell to cell. Unlike the pesti- viruses, RUV ultimately disappears in children with congenital rubella in spite of the inability of ant ibody to eliminate the infection and the possible existence of tolerance of the CMI system. A plausible explanation for the eventual cessation of RUV persistence is the clonal hypothesis (Rawls, 1974). In vitro, RUV-infected cells have a reduced growth rate and a limited doubling potential. If a similar virus--host relationship exists in vivo, clones of infected cells gradually die out, leaving the infant free of virus. Thus, ac- cording to this concept, the duration of virus persistence is largely deter- mined by the lifespan of the infected cell.
The discrepancy between congenital RUV and HCV infection with regard to immune response and virus persistence may theoretically be explained by a difference in maturation of these viruses. HCV appears to mature at intra- cytoplasmatic membranes (Scherrer et al., 1970; Rutili and Titoli, 1977) and induces few or no viral antigens in the cell membrane (Van Oirschot, 1980). Thus, a cell infected with HCV may not elicit a specific immune response and in addition may not be recognized by the immune system, a condition which obviously favours virus persistence. On the other hand, RUV develops by budding from plasma membranes, which results in antigenic changes of the cell's surface (Oshiro et al., 1969). The expression of new antigens in the cell's surface triggers a predominant CMI response, which in turn may con- tribute to the termination of RUV infection.
CONCLUDING REMARKS
Viruses, which produce congenital infections, belong to many families. In this respect the nonarbo togaviruses may be considered to be a unique group, since they all are able to cross the placenta, notwithstanding the di- versity in natural host species and the consequent variation in placental struc- ture. It is likely that one unifying mechanism underlies this capability. One pronounced biological feature the nonarbo togaviruses have in common is their predilection for vascular endothelial cells. Viral replication in these cells results in vascular lesions. Apparently, the placental endothelium is highly susceptible for these viruses, and this characteristic might well be respon- sible for the common ability of the nonarbo togaviruses to grow through the placenta.

350
Foeta l infec t ion with nona rbo togaviruses can resul t in a wide spec t rum o f congeni ta l abnormal i t ies , ranging f rom prenata l dea th to inapparen t in- fec t ion which persists for life. The nona rbo togaviruses, with the ex cep t i o n o f LDV, can all cause mild to severe deve lopmen ta l defects , which is remark- able since only a relat ively small p r o p o r t i o n o f viruses are proven tera togens. The te ra togenic nona rbo togaviruses usually are of low virulence and have low to zero cy topa thogen i c i t y . These features al low the foe tus to survive a n d consequen t ly the virus ~o persist and disseminate widely, which enables it to p roduce its de t r imenta l e f fec ts on the foetus . The mechanisms m o s t l ikely to be opera t ing in teratogenesis are an inhibi t ion, or d i s to r t ion , of normal cellular deve lopment , and the induc t ion o f vascular lesions, ini t ia ted by endothel ia l degenera t ion .
The age of the foe tus at the t ime o f exposure to the virus largely deter- mines the course and eventual o u t c o m e of infect ion. Virus pers is tence usu- ally fol lows a foetal in fec t ion with nona rbo togaviruses be fore or a round the onset o f i m m u n o c o m p e t e n c e . In congeni ta l rubel la the virus persists in spite o f the p r o d u c t i o n o f specific an t i body by the h u m a n foetus . The in u te ro infec ted foetuses o f domes t i c animals f r equen t ly are i m m u n o t o l e r a n t to the causative pestivirus. Virus pers is tence in congeni ta l pestivirus infec- t ions can be lifelong, bu t in congeni ta l rubel la it eventual ly ceases. Despite the widespread and con t inued presence o f virus in congeni ta l ly in fec ted catt le , sheep and swine they may appear hea l thy at bir th and remain clini- cally normal for m o n t h s or years. These animals shed virus co n t i n u o u s ly and act epizoot ica l ly like " t h e Tro jan ho r se " tha t surrept i t ious ly in t roduces an a rmy of pestiviruses into animal popula t ions . Such a vi rus--host in te rac t ion is vir tual ly never encoun t e r ed in any o the r viral infect ions o f man or animals.
F u r the r studies on the cellular level as well as the level of the macroor - ganism should con t inue to yield in fo rma t ion useful for unders tand ing the, in some respects un ique , congeni ta l nona rbo togavirus infect ions .
ACKNOWLEDGEMENTS
The a u tho r thanks Drs. J.G. van Bekkum, A.C. Hekker , C. Terps t ra and D. van Zaane for careful ly reading the manuscr ip t .
R E F E R E N C E S
Achs, R., Harper, K.G. and Siegal, M., 1966. Unusual dermatoglyphic findings associated with rubella embryopathy. New. Engl. J. Med., 274: 148--150.
Acland, H.M., Gard, G.P. and Plant, J.W., 1972. Infection of sheep with a mucosal disease virus. Aust. Vet. J., 48: 70--75.
Alford, Jr., C.A., Neva, F.A. and Weller, T.H., 1964. Virologic and serologic studies on human products of conception after maternal rubella. New. Engl. J. Med., 271: 1275-- 1281.
Alford, Jr., C.A., Neva, F.A. and Weller, T.H., 1965. Retrospective diagnosis of the rubella syndrome by virusisolation. J. Pediatr., 67: 983.

351
Ansari, B.M. and Mason, M.K., 1977. Chromosomal abnormality in congenital rubella. Pediatrics, 59: 13--15.
Archbald, L.F. and Zemjanis, R., 1977. Intrauterine infusion of the virus of bovine virus diarrhea and artificial insemination in the cow at estrus. Vet. Med. Small Anim. Clin., 72: 221--225.
Archbald, L.F., Fulton, R.W., Seger, C.L., A1-Bagdadi, F. and Godke, R.A., 1979. Effect of the bovine viral diarrhea virus on preimplant~tion bovine embryos. Theriogenoiogy, 11: 81--89.
Atluru, D., Notowidjojo, W., Johnson, D.W. and Muscoplat, C.C., 1979. Suppression of in vitro immunoglobulin biosynthesis in bovine spleen cells by bovine viral diarrhea virus. Clin. Immunol. Immunopathol. , 13: 254--260.
Axthelm, M.K., Leipold, H.W., Jayasekara, U.M. and Phillips, R.M., 1981. Congenital microhydranencephalus in cattle. Cornell Vet., 71 : 164--174.
Badman, R.T., Mitchell, G., Jones, R.T. and Westbury, H.A., 1981. Association of bovine viral diarrhoea infection to hydranencephaly and other central nervous system lesions in perinatal calves. Aust. Vet. J., 57: 306--307.
Barlow, R.M., 1972. Experiments in border disease. IV. Pathological changes in ewes. J. Comp. Pathol., 72: 151--157.
Barlow, R.M., 1980. Morphogenesis of hydranencephaty and other intracranial malforma- tions in progeny of pregnant ewes infected with pestiviruses. J. Comp. Pathol., 90: 87--98.
Barlow, R.M. and Gardiner, A.C., 1969. Experiments in border disease. I. Transmission, pathology and some serological aspects of the experimental disease. J. Comp. Pathol., 79: 397--405.
Barlow, R.M. and Storey, I.J., 1977. Myelination of the ovine CNS with special reference to border disease. II. Quantitative aspects. Neuropathot. Appl. Neurobiol., 3: 255-- 265.
Barlow, R.M., Rennie, J.C., Keir, W.A., Gardiner, A.C. and Vantsis, J.T., 1975. Experi- ments in border disease. VIII. The disease in goats. J. Comp. Pathol., 85: 291--297.
Barlow, R.M., Vantsis, J.T., Gardiner, A.C. and Linklater, K.A., 1979. The definition of border disease: problems for the diagnostician. Vet. Rec., 104: 334--336.
Barlow, R.M., Rennie, J.C., Gardiner, A.C. and Vantsis, J.T., 1980a. Infection of preg- nant sheep with the NADL strain of bovine virus diarrhoea virus and their subsequent challenge with border disease IIB pool. J. Comp. Pathol., 90: 67--72.
Barlow, R.M., Vantsis, J.T., Gardiner, A.C., Rennie, J.C., Herring, J.A. and Scott, F.M.M., 1980b. Mechanisms of natural transmission of border disease. J. Comp. Pathol., 90: 57--65.
Benner, J.W., 1928. Hog cholera in the young pig. J. Am. Vet. Med. Assoc., 73: 39--54. Berry, C.L., Beveridge, W.I.B., Done, J.T., Edwards, M.J., Hendrix, A.G., Milouchine,
V.N., Priester, W.A. and Tuchmann-Duplessis, H., 1977. Non-Mendelian developmental defects: animal models and implications for research into human disease. Bull. W.H.O., 55: 475--487.
Binkhorst, G.J., Journ~e, D.L.H., Wouda, W., Strayer, P.J. and Vos, J.H., 1983. Neurolog- ical disorders, virus persistence, hypomyeliuation due to intra-uterine infection with bovine virus diarrhoea virus. I. Clinical signs and morphological lesions. Vet. Q., in press.
Bistner, S.I., Rubin, L.F. and Saunders, L.Z., 1970. The ocular lesions of bovine viral diarrhea--mucosal disease. Pathol. Vet., 7: 275--286.
Bourne, F.J., Curtis, J., Johnson, R.H. and Collings, D.F., 1974. Antibody formation in porcine fetuses. Res. Vet. Sci., 16: 223--227.
Brack, M., 1971. Pathologische Ver~inderungen der europ~/ischen Schweinepest bei Feten und Neugeborenen. Zbl. Vet. Med. B, 18: 749--760.
Brambell, F.W.R., 1970. The transmission of passive immunity from mother to young. In: Frontiers of Biology. Vol. 18. A. Neuberger and E.L. Tatum (Editors), North- Holland, Amsterdam, 385 pp.

352
Bran, L., Mihaita, S., Popa, M. and Totorcea, N., 1971. Trans-uterine and transplacentar transmission of attenuated rabbit adapted swine fever virus strains in pregnant sows. Arch. Vet. Bucuresti, 11--20.
Braun, R.K., Osburn, B.I. and Kendrick, J.W., 1973. Immunologic response of the bovine fetus to bovine viral diarrhea virus. Am. J. Vet. Res., 34: 1127--1132.
Brown, T.T., DeLahunta, A., Scott, F.W., Kahrs, R.F., McEntee, K. and Gillespie, J.H., 1973. Virus induced congenital anomalies of the bovine fetus. II. Histopathology of cerebellar degeneration (hypoplasia) induced by the virus of bovine viral diarrhea-- mucosal disease. Cornell Vet., 63: 561--578.
Brown, T.T., DeLahunta, A., Bistner, S.I., Scott, F.W. and McEntee, K., 1974. Patho- genetic studies of infection of the bovine fetus with bovine viral diarrhea virus. I. Cerebellar atrophy. Vet. Pathol., 11 : 486--505.
Brown, T.T., Bistner, S.I., DeLahunta, A., Scott, F.W. and McEntee, K., 1975. Patho- genetic studies of infection of the bovine fetus with bovine viral diarrhea virus. II. Ocular lesions. Vet. Pathol., 12: 394--404.
Brown, T.T., Schultz, R.D., Duncan, J.R. and Bistner, S.I., 1979. Serological response of the bovine fetus to bovine viral diarrhea virus. Infect. Immun., 25: 93--97.
Bryans, J.T., Doll, E.R., Crowe, M.E.W. and McCollum, W.H., 1957. The blood picture and thermal reaction in experimental viral arteritis of horses. CorneU Vet., 47 : 42--52.
Buimovici-Klein, E., Lang, P.B., Ziring, P.R. and Cooper, L.Z., 1979. Impaired cell-medi- ated immune response in patients with congenital rubella: correlation with gestational age at time of infection. Pediatrics, 64: 620--626.
Carbrey, E.A., Stewart, W.C., Kresse, J.I. and Snyder, M.L., 1976. Bovine viral diarrhea infection in pigs and its differentiation from infection with hog cholera strains of low virulence. C.E.C. Agricultural research seminar, diagnosis and epizootiology of classi- cal swine fever. Amsterdam, 1975. EUR 5486, Luxembourg, pp. 126--158.
Casaro, A.P.E., Kendrick, J.W. and Kennedy, P.C., 1971. Response of the bovine fetus to bovine viral diarrhea--mucosal disease virus. Am. J. Vet. Res., 32 : 1543--1562.
Chandra, R.K. and Soothill, J.F., 1971. Suppression of developed spleen haemolytic plaque formation by neonatal antigen stimulus in mice. Immunology, 20: 175--183.
Charles, D., 1980. Viral infections. In: Infections in Obstetrics and Gynecology, Saun- ders, Philadelphia, PA, pp. 103--112.
Cho, D.Y. and Leipold, H.W., 1977. Congenital defects of the bovine central nervous system. V_et. Bull., 47: 489--503.
Claman, H.N., Suvatte, V., Githens, J.H. and Hathaway, W.E., 1970. Histiocytic reaction in dysgammaglobulinaemia and congenital rubella. Pediatrics, 46: 89--96.
Cooper, L.Z., Green, R.H., Krugman, S., Giles, J.P. and Mirick, G.S., 1965. Neonatal thrombocytopenic purpura and other manifestations of rubella contracted in utero. Am. J. Dis. Child, 110: 416--427.
Cooper, L.Z., Ziring, P.R., Ockerse, A.B., Fedun, B.A., Kiely, B. and Krugman, S., 1969. Rubella. Clinical manifestations and management. Am. J. Dis. Child., 118:18--29.
Coria, M.F. and McClurkin, A.W., 1978. Specific immune tolerance in an apparently healthy bull persistently infected with bovine viral diarrhea virus, J. Am. Vet. Med. Assoc., 172: 449--451.
Corthier, G., Aynaud, J.M., Galicher, C. and Gelfi, J., 1974. Activit4 antig~nique com- parse de deux togavirus: le virus de la peste porcine et le virus de la maladie des muqueuses. Ann. Rech. Vet., 5: 373--393.
Cotlier, E., Fox, J., Bohigian, G., Beaty, C. and Du Pree, A., 1968. Pathogenic effects of rubella virus on embryos and newborn rats. Nature, 217 : 38--40.
Cowart, W.O. and Morehouse, L.G., 1967. Effects of attenuated hog cholera virus in pregnant swine at various stages of gestation. J. Am. Vet. Med. Assoc., 151: 1788-- 1794.
Cradock-Watson, J:E., Ridehaigh, M:K.S. and Chantter, S., 1976. Specific immunoglobu- lins in infants with the congenital rubella syndrome. J. Hyg., 76: 109--123.

353
Cradock-Watson, J.E., Ridehalgh, M.K.S., Anderson, M.J., Pattison, J.R. and Kangro, H.O., 1980. Fetal infection resulting from maternal rubella after the first trimester of pregnancy. J. Hyg., 85: 381--391.
Crispens, C.G., 1965. On the transmission of the lactic dehydrogenase agent from mother to offspring. J. Natl. Canc. Inst., 34: 331--336.
Darbyshire, J.H., 1960. A serological relationship between swine fever and mucosal dis- ease of cattle. Vet. Rec., 72: 331.
Derbyshire, M.B. and Barlow, R.M., 1976. Experiments in border disease. IX. The patho- genesis of the skin lesion. J. Comp. Pathot., 86 : 557--570.
Desmond, M.M., Wilson, G.S., Melnick, J.L., Singer, D.B., Zion, T.E., Rudolph, A.J., Pineda, R.G., Ziai, M.H. and Blattner, R.J., 1967. Congenital rubella encephalitis. Course and early sequelae. J. Pediatr., 71 : 311--331.
Desmyter, J., Rawls, W.E., Melnick, J.L., Yow, M.D. and Barrett, F.F. , 1967. Interferon in congenital rubella: response to live at tenuated measles vaccine. J. Immunol. , 99: 771--777.
Doll, E.R., Bryans, J.T., McCollum, W.H. and Crowe, M.E.W., 1957. Isolation of a filter- able agent causing arteritis of horses and abort ion by mares: its differentiation from the equine abort ion (influenza) virus. Cornell Vet., 47: 3--41.
Done, J.T. and Harding, J.D.J., 1969. An experimentally-induced congenital tremor of pigs. In: A. Bertelli and L. Donati (Editors), Teratology. Int. Congress Series No. 173, Proc. Italian Soc. Exp. Teratology, Como, 1967, pp. 112--118.
Done, J.T., Terlecki, S., Richardson, C., Harkness, J.W., Sands, J.J., Patterson, D.S.P., Sweasey, D., Shaw, I.G., Winkler, C.E. and Duffell, S.J., 1980. Bovine virus diarrhea-- mucosal disease virus: pathogenici ty for the fetal calf following maternal infection. Vet, Rec., 106: 473--479.
Dudgeon, J.A,, 1975. Congenital rubella. J. Pediatr., 87: 1078--1086. Dudgeon, J.A., 1976. Infective causes of human malformations. Br. Med. Bull., 32: 77--
83. Dunne, H.W. and Clark, C.D., 1968. Embryonic death, fetal mummification, stillbirth
and neonatal death in pigs of gilts vaccinated with at tenuated live-virus hog cholera vaccine. Am. J. Vet. Res., 29: 787--796.
Ebbins, A., Wilson, M.G., Chandor, S.B. and Wehrle, P.F., 1973. Inadvertent rubella im- munisation in pregnancy. Am. J. Obstet. Gynecol., 117: 505--512.
Eilard, T. and Strannegard, O., 1974. Rubella reinfection in pregnancy followed by transmission to the fetus. J. Infect. Dis., 129: 594--596.
Emerson, J.L. and Delez, A.L., 1965. Cerebellar hypoplasia, hypomyelinogenesis and congenital tremors of pigs, associated with prenatal hog cholera vaccination of sows. J. Am. Vet. Med. Assoc., 147: 47--54.
Fahey, K.J. and Morris, B,, 1978. Humoral immune responses in foetal sheep. Immunol- ogy, 35: 656--661.
Fernelius, A.L., Amtower, W,C., Lambert , G., McClurkin, A.W. and Matthews, P.J., 1973a. Bovine viral diarrhea virus in swine: characteristics of virus recovered from naturally and experimentally infected swine. Can. J. Comp. Med., 37 : 13--20.
Fernelius, A.L., Amtower, W.C., Malmquist, W.A., Lambert , G: and Matthews, P.J., 1973b. Bovine viral diarrhea virus in swine: neutralizing ant ibody in naturally and experimentally infected swine. Can. J. Comp. Med., 37: 96--102.
Floret , D., Rosenberg, D., Hage, G.N. and Monnet, P., 1980. Hyperthyroidism, diabetes mellitus and the congenital rubella syndrome. Acta Paediatr. Scand., 69: 259--261.
Forrest, J.M. and Menser, M.A., 1975. Recent implications of intxauterine and postnatal rubella. Aust. Paediat. J., 11: 65--75.
French, E.L., Hore, D.E., Snowdon, W.A., Parsonson, I.M. and Uteri, J., 1974. Infection of pregnant ewes with mucosal disease virus of ovine origin. Aust. Vet. J., 50: 45--54.
Frey, H.R., Liess, B., Richter-Reichhelm, H.B., Von Benten, K. and Trautwein, G., 1980. Experimental transplacental transmission of hog cholera virus in pigs. I. Virological and serological studies. Zbl. Vet. Med, B, 27: 154--164.

354
Fuccitlo, D.A., Stule, R.W., Hensen, S.A., Vincent, M.M., Hardy, J.B. and Bellanti+ J ,A , 1974. Impaired cellular immunity to rubella virus in congenital rubella. Infect. Immun., 9: 81--84.
Gard, G.P., Acland, H.M. and Plant, J.W., 1976. A mucosal disease virus as a cause of abortion, hairy birth coat and unthriftiness in sheep. 2. Observations on lambs sur- viving for longer than seven days. Aust. Vet. J., 52: 64--68.
Gardiner, A.C., 1980. The distribution and significance of border disease viral antigen in infected lambs and foetuses. J. Comp. Pathol., 90: 513--518.
Gardiner, A.C. and Barlow, R.M., 1972. Experiments in border disease. III. Some epi- demiological considerations with reference to the experimental disease. J. Comp. Pathol., 82: 29--35.
Gardiner, A.C. and Barlow, R.M., 1981. Vertical transmission of border disease infection. J. Comp. Pathol., 91: 467--470.
Gardiner, A.C., Zakarian, B. and Barlow, R.M., 1980. Periarteritis in experimental border disease of sheep. III. Immunopathotogical observations. J. Comp. Pathol., 90: 469-- 474.
Georgii, A., Lenz, I. and Zobel, H., 1964. Penetration of the placental barrier by the lactate dehydrogenase-elevating virus (Riley) and its behaviour in mouse cultures following infection in utero. Proc. Soc. Exp. Biol. Med., 117: 322--326.
Gershon, R.K., Gery, I. and Waksman, B.H., 1974. Suppressive effects of in vivo im- munisation on PHA rcsponses-~i t ro ,~I , !mmunv!., 1 1 ~ 2 1 5 ~ 2 2 1 ~ - . . . .
Gibbons, D.F., Winkler, C.E., Shaw, I.G., Tertecki, S., Richardson, C. and Done, J.T., 1974. Pathogenicity of the border disease agent for the bovine foetus. Br. Vet. J., 130: 357--361. : :
Gillespie, J., Bartholomew, P., Thomson, R. and McEntee, K., 1967. The isolation of noncytopathic virus diarrhea' virus from two aborted fetuses. Cornell Vet., 57 : 564-- 571.
Gregg, N.M., 1941. Congenital cataracts following German measles in the mother. Trans. Ophthalmol. Soc. Aust., 3: 35--46.
Hamilton, A. and Timoney, P.J., 1972. BVD virus and "Border disease". Vet. Rec., 91: 468.
Hardy, J.B., 1976. Immediate and long-range effects of maternal viral infection in preg- nancy. Birth Defects, 12: 23--31.
Hardy, J.B~, McCracken, G.H., Jr., Gilkeson, M.R. and Sever, J.L., 1969. Adverse fetal outcome following maternal rubella after the first trlmesier of pregnancy. J. ~m: 1VVed. Assoc., 207: 2414--2420.
Harkness, J.W., King, A.A., Terlecki, S. and Sands, J.J., 1977. Border disease of sheep: isolation of the virus in tissue culture and experimental reproduction of the disease. Vet. Rec., 100: 71--72.
Heggie, A.D., 1977. Growth inhibition of human embryonic and fetal rat bones in organ culture by rubella virus. Teratology, 15: 47--56.
Hermanns, W., 1978. Immunopathologie der experimentellen, diaplazentaren Infektion mit dem Virus der Europ~ischen Schweinepest beim neugeborenen Schwein, Disser- tation, Hannover, 183 pp.
Hermanns, W., Trautwein, G., Meyer, H. and Liess, B., 1981. Experimental transplacental transmission of hog cholera virus in pigs. V. Immunopathological findings in newborn pigs. Zbl. Vet. Med. B., 28: 669--683.
Hicks, S.P., 1954. Mechanism of radiation anencephaly, anophthalmia and pituitary anomalies. Repair in the mammalian embryo. Arch. Pathol., 57: 363--378.
Honeyman, M.C., Dorman, D.C., Menser, M.A., Forrest, J.M., Guiman, J.J. and Clark, P., 1975. HL-A antigens in congenital rubella and the role of antigens 1 and 8 in the epi- demiology of natural rubella. Tissue Antigens, 5: 12--18.
Horzinek, M.C., 1973. Comparative aspects of togaviruses. J. Gen. Virol., 20: 87--103. Horzinek, M.C., 1981. Nonarbo togavirus infections of animals: comparative aspects and
diagnosis. Comparative Diagnosis of Viral Disease. Vol. IV, Academic Press, London, pp. 441--475.

355
Huck, R.A., 1973. Transmission of border disease in goats. Vet. Rec., 92: 151. Hughes, L.E., Kershaw, G.F. and Shaw, I.G., 1959. "B" or border disease. An unde-
scribed disease of sheep. Vet. Rec., 71 : 313--317. Johnson, K.P., Ferguson, L.C., Byington, D.P. and Redman, D.R., 1974. Multiple fetal
malformations due to persistent viral infection. I. Abortion, intrauterine death, and gross abnormalities in fetal swine infected with hog cholera vaccine virus. Lab. Invest., 30: 608--617.
Kendrick, J.W., 1971. Bovine viral diarrhea---mucosal disease virus infection in pregnant cows. Am. J. Vet. Res., 32: 533--544.
Kendrick, J.W., 1975. Laboratory diagnosis of bovine abortion. C.A. Kirkbridge (Editor), Am. Assoc. Vet. Lab. Diagnost., Madison, WI, U.S.A., pp. 1--6.
Kendrick, J.W., 1976. Bovine viral diarrhea--virus induced abortion. Theriogenology, 5: 91--93.
Kono, R., Hayakawa, Y., Hiki, M. and Ishii, K., 1969. Experimental vertical transmission of rubella virus in rabbits. Lancet, i: 343--347.
Korn, G. and Lorenz, R.J., 1976. Untersuchungen zur Vermehrung des Virus der euro- p~'ischen Schweinepest in Makrophagen- und Lymphozyten-kulturen yon normalen und immunen Tieren. Zbl. Bakt. Hyg. I. Abt. Orig. Reihe A, 236: 150--162.
Kumagai, T., Morimoto, T., Shimizu, T., Sasahara, S. and Watanabe, M., 1962. Antigenic relationship between hog cholera virus and bovine viral diarrhea virus as revealed by cross-neutralisation. Natl. Inst. Anita. Health Q., 2: 201--206.
Laude, H., 1979. Nonarbo-togaviridae: comparative hydrodynamic properties of the pestivirus genus. Arch. Virol., 62: 347--352.
Laude, H. and Gelfi, J., 1979. Properties of border disease virus as studied in a sheep cell line. Arch. Virol., 62: 341--346.
Lejarraga, H. and Peckham, C.S., 1974. Birthweight and subsequent growth of children exposed to rubella infection in utero. Arch. Dis. Child., 49: 50--54.
Liess, B., Frey, H.R., Buksch, U., Raeder, B. al.d Frost, J., 1976. Hog cholera viral anti- gens and some recent results of the relationship to envelope antigens of bovine viral diarrhea--mucosal disease virions. C.E.C. Agricultural research seminar, studies on virus replication. Brussels, 1974. EUR 5451, Luxembourg, pp. 333--346.
Lindquist, J.M., Plotkin, S.A., Shaw, L., Gilden, R.V. and Williams, M.L., 1965. Congeni- tal rubella syndrome as a systemic infection. Studies of affected infants born in Phila- delphia, U.S.A. Br. Med. J., 2: 1401--1406.
Loan, R.W. and Storm, M.M., 1968. Propagation and transmission of hog cholera virus in nonporcine hosts. Am. J. Vet. Res., 29: 807--811.
Manktelow, B.W., Porter, W.L. and Lewis, K.H.C., 1969. Hairy shaker disease of lambs. N.Z. Vet. J., 17: 245--248.
Markson, L.M., Winkler, C.E. and Terlecki, S., 1976. A report of an outbreak of en- cephalic malformation in calves. Proc. 9th Congr~s International sur les Maladies du Bgtail, Paris, 1 : 211--215.
Marshall, W.C., 1973. Intrauterine infections. In: K. Elliott and J. Knight (Editors), Intrauterine Infections. Ciba Fnd. Syrup., No. 10. Associated Science Publishers, Amsterdam, pp. 3--22.
Marshall, W.C., Cope, W.A., Soothill, J.F. and Dudgeon, J.A., 1970. In vitro lymphocyte response in some immunity deficiency diseases and in intrauterine virus infections. Proc. R. Soc. Med., 63: 351--354.
Matsuo, S., 1975. Experimental vertical infection with rubella virus in mice. Trop. Med., 17 : 47--54.
McClurkin, A.W., Coria, M.F. and Cutlip, R.C., 1979. Reproductive performance of ap- parently healthy cattle persistently infected with bovine viral diarrhea virus. J. Am. Vet. Med. Assoc., 174: 1116--1119.
MeCollum, W.H. and Bryans, J.T., 1973. Serological identification of infection by equine arteritis virus in horses of several countries. Proc. 3rd. Int. Conf. Equine Inf. Dis. Paris, 1972. Karger, Basel, pp. 256--263.

McKay, D.G. and Margaretten, W., 1967. Disseminated intravascutar coagulation in virus diseases. Arch. Intern. Med., 120: 129--152.
Mellman, W.J., Plotkin, S.A., Moorhead, P.S. and Hartnett, E.M., 1965. Rubella infection of human leucocytes. Am. J. Dis. Child., 110: 473--476.
Mengeling, W.L., Gutekunst, D.E., Fernelius, A.L. and Pirtle, E.C., 1963. Demonstration of an antigenic relationship between hog cholera and bovine diarrhea viruses by im- munofluorescence. Can. J. Comp. Med., 27 : 162--164.
Mengeling, W.L., Pirtle, E.C. and Packer, R.A., 1970. Persistent infection of an estab- lished porcine kidney cell line with selected strains of hog cholera virus. Am. J. Vet. Res., 31: 2019--2024.
Menser, M.A., Dods, L. and Harley, J.D., 1967a. A twenty-five year follow-up of con- genital rubella. Lancet, ii: 1347--1350.
Menser, M.A., Harley, J.D., Hertzberg, R., Dorman, D.C. and Murphy, A.M., 1967b. Per- sistence of virus in the lens for three years after prenatal rubella. Lancet, ii: 387--388.
Menser, M.A., Forrest, J.M., Honeyman, M.C. and Burgess, J.A., 1974. Diabetes, HL-A antigens, and congenital rubella, Lancet, ii: 1508--1509.
Meyer, H., 1978. Experimentelle diaplazentare Infektion yon Schweinefeten mit dem Virus der europ~iischen Schweinepest. Virologische und serologische Untersuchungen. Dissertation, Hannover, 128 pp.
Meyer, H., Liess, B., Frey, H.R., Hermanns, W. and Trautwein, G., 1981. Experimental transplacental transmission of hog cholera virus in pigs. IV. Virological and serological studies in newborn piglets. Zbl. Vet. Med. B., 28: 659--668.
Michaels, R.H., 1969. Immunologic aspects of congenital rubella.Pediatrics, 43: 339--350. Michaels, R.H., 1972. Suppression of antibody response in congenital rubella. J. Pediatr.,
80 : 583--588. Michalka, J., 1931. t~ber die Auswirkungen der Simultan-impfung gegen Schweinepest
in Zucht- und Mastbest~nden. Zugleich ein Beitrag zur Frage der Virustr~ger bei der Schweinepest. Arch. Wiss. Prakt. Tierheilkd., 63: 529--542.
Mires, C.A., 1974. Factors in the mechanism of persistence of viral infections. Progr. Med. Virol., 18: 1--14.
Monif, G.R.G., Avery, G.B., Korones, S.B. and Sever, J.L., 1965. Postmortem isolation of rubella virus from three children with rubella-syndrome defects. Lancet, ii: 723-- 724.
Montgomery, J.R., South, M.A., Rawls, W.E., Melnick, J.L., Olson, G.B., Dent, P.B. and Good, R.A., 1967. Viral inhibition of lymphocyte response to phytohaemagglutinin. Science, 157: 1068--1070.
Moraillon, A. and Moraillon, R., 1978. Results of a serological survey of viral arteritis in France and in several European and African countries. Proc. IVth Int. Conf. Eq. Inf. Dis., J.T. Bryans and H. Gerber (Editors), Lyon, 1976, pp. 467--474.
Muscoplat, C.C., Johnson, D.W. and Teuscher, E., 1973. Surface immunoglobutin of cir- culating lymphocytes in chronic bovine diarrhea: abnormalities in cell populations and cell functions. Am. J. Vet. Res., 34: 1101--1104.
Naeye, R.L. and Blanc, W., 1965. Pathogenesis of congenital rubella. J. Am. Med. Assoc., 194: 109--115.
Nieburg, P.L and Gardner, L.I., 1976. Thyroiditis and congenital rubella syndrome. J. Pediatr., 88: 156.
Norval, M., 1979. Mechanism of persistence of rubella virus in LLC-MK: cells. J. Gen. Virol., 43: 289--298.
Notkins, A.L., 1974. Immune mechanisms by which the spread of viral infections is stopped. Cell. Immunol., 11: 478--483.
Nusbacher, J., Hirschhorn, K. and Cooper, L.Z., 1967. Chromosomal abnormalities in congenital rubella. New Engl. J. Med., 276: 1409--1413.
Olafson, P., McCallum, A. and Fox, F., 1946. An apparently new transmissible disease of cattle (BVD). Cornell Vet., 36: 205--213.

357
Olson, G.B., Dent, P.B., Rawls, W.E., South, M.A., Montgomery, J.R., Melnick, J.L. and Good, R.A., 1968. Abnormalities of in vitro lymphocyte responses during rubella virus infection. J. Exp. Med., 128: 47---68.
Omtvedt, I.T., Nelson, R.E., Edwards, R.L., Stephens, D.F. and Turman, E.J., 1971. Influence of heat stress during early, mid and late pregnancy of gilts. J. Anita. Sci., 32: 312--317.
Ornoy, A., Segal, S., Nishmi, M., Simcha, A. and Polishuk, W.Z., 1973. Fetal and pla- cental pathology in gestational rubella. Am. J. Obstet. Gynecol., 116: 949--956.
Osburn, B.I., Clarke, G.L., Stewart, W.C. and Sawyer, M., 1973. Border disease like syndrome in lambs: antibodies to hog cholera and bovine viral diarrhea viruses. J. Am. Vet. Med. Assoc., 163: 1165--1167.
Oshiro, L.S., Schmidt, N.J. and Lennette, E.H., 1969. Electron microscopic studies on rubella virus. J. Gen. Virol., 5: 202--210.
Overall, Jr., J.C., 1972. Intrauterine virus infections and congenital heart disease. Am. Heart J., 84: 823--833.
Parkman, P.D., Buescher, E.L. and Artenstein, M.S., 1962. Recovery of rubella virus from army recruits. Proc. Soc. Exp. Biol. Med., 111: 225--230.
Parkman, P.D., Phillips, P.E., Meyer, Jr., H.M., 1965. Experimental rubella virus infec- t ion in pregnant monkeys. Am. J. Dis. Child., 110: 390--394.
Parsonson, I.M., O'Halloran, M.L., Zee, Y.C. and Snowdon, W.A., 1979. The effect of bovine viral diarrhoea--mucosal disease (BVD) virus on the ovine foetus. Vet. Micro- biol., 4: 279--292.
Patterson, D.S.P., 1975. Border Disease. Vet. Annu., 15: 83--86. Patterson, K., Chandra, R.S. and Jenson, A.B., 1981. Congenital rubella, insulitis and
diabetes mellitus in an infant. Lancet, i: 1048--1049. Peckham, C.S., 1972. Clinical and laboratory study of children exposed in utero to
maternal rubella. Arch. Dis. Child., 47 : 571--577. Phillips, C.A., Melnick, J.L., Yow, M.D., Bayatpour, M. and Burkhardt, M., 1965. Per-
sistence of virus in infants with congenital rubella and in normal infants with a history of maternal rubella. J. Am. Med. Ass., 193: 111--113.
Pirtle, E.C., 1969a. In vitro spread of hog cholera viral infection from cell to cell. Demon- stration of viral antigen in dividing cells. Am. J. Vet. Res., 30: 1909--1912.
Pirtle, E.C., 1969b. In vitro spread of hog cholera viral infection from cell to cell. Demon- stration of viral antigen in cytoplasmatic bridges. Am. J. Vet. Res., 30: 1913--1919.
Pirtle, E.C. and Mengeling, W.L., 1970. Deoxyribonucleic acid synthesis in cultured por- cine kidney cells infected with hog cholera virus. Am. J. Vet. Res., 31 : 1795--1799.
Pirtle, E.C. and Woods, L.K., 1968. Cytogenic alteration in swine kidney cells persistent- ly infected with hog cholera virus and propagated with and without serum in the medium. Am. J. Vet. Res., 29: 153--164.
Plotkin, S.A. and Vaheri, A., 1967. Human fibroblasts with rubella virus produce a growth inhibitor. Science, 156: 659--661.
Plotkin, S.A., Boue, A. and Boue, J.G., 1965a. The in vitro growth of rubella virus in human embryo cells. Am. J. Epidem., 81: 71--85.
Plotkin, S.A., Oski, F.A., Hartnett, E.M., Hervada, A.R., Friedman, S. and Gowing, J., 1965b. Some recently recognized manifestations of the rubella syndrome. J. Pediatr., 67: 182--191.
Porter, W.L., Lewis, K.H.C. and Manktelow, B.W., 1972. Hairy shaker disease of lambs: further studies on acquired immunity, abortion and effects on foetal growth. N.Z. Vet. J., 20: 4--7.
Porterfield, J.S., Casals, J., Chumakov, M.P., Gaidamovich, S.Y., Hannoun, C., Holmes, I.H., Horzinek, M.C., Mussgay, M., Oker-Blom, N., Russell, P.K. and Trent, D.W., 1978. Togaviridae. Intervirology, 9: 129--148.
Potter, J.E., Banatvala, J.E. and Best, J.M., 1973. Interferon studies with Japanese, and U.S. rubella virus strains. Br. Med. J., i: 197--199.

358
Preece, M.A., Kearney, P.J. and Marshall, W.C., 1977. Growth-hormone deficiency in congenital rubella. Lancet, ii: 842--844.
Ramsey, F.K. and Chivers, W.H., 1953. Mucosal disease of cattle. North. Am. Vet., 34: 629--633.
Rawls, W.E., 1967. Congenital rubella: a chronic viral disease. Pediatr. Digest. Sept., pp. 41--49,
Rawls, W.E., 1968. Congenital rubella: the significance of virus persistence. Prog. ivied. Virol., 10: 238--285.
Rawls, W.E., 1974. Viral persistence in congenital rubella. Prog. Med. Virol., 18: 273-- 288.
Rawls, W.E. and Melnick, J.L., 1966. Rubella virus carrier cultures derived from congeni- tally infected infants. J. Exp. Med., 123: 795--816.
Rawls, W.E., Melnick, J.L., Rosenberg, H.S. and Bayatpour, M., 1965. Spontaneous virus carrier cultures and postmortem isolation of virus from infants with congenital rubella. Proc. Soc. Exp. Biol. Med., 120: 623--626.
Rawls, W.E., Phillips, C.A., Melnick, J.L. and Desmond, M.M., 1967. Persistent virus in- fection in congenital rubella. Arch. Ophthalmol., 77: 430--433.
Richardson, C., 1977. Morphological parameters of intrauterine growth retardation in the newborn lamb. Vet. Rec., 101: 151--152.
Richardson, C., Hebert, C.N. and Done, J.T., 1976. Experimental border disease in sheep: dose-response effect. Br. Vet. J., 132: 202--207.
Richter-Reichhelm, H.B., Trautwein, G., Von Benten, K., Liess, B. and Frey, H.R., 1980. Experimental transplacental transmission of hog cholera virus in pigs. II. Immuno- pathological findings in the fetus. Zbl. Vet. Med. B, 27 : 243--252.
Rinaldo, Jr., C.R., Isackson, D.W., Overall, Jr., J.C., Glasgow, L.A., Brown, T.T., Bistner, S.I., Gillespie, J.H. and Scott, F.W., 1976. Fetal and adult bovine interferon produc- tion during bovine viral diarrhea virus infection. Infect. Immun., 14: 660--666.
Roitt, I.M., 1980. Essential Immunology. Blackwell, Oxford, 358 pp. Rorke, L.B., 1973. Nervous system lesions in the congenital rubella syndrome. Acta
Otolaryngol., 98: 249--251. Rorke, L.B. and Spiro, A.J., 1967. Cerebral lesions in congenital rubella syndrome. J.
Pediatr., 70: 243--255. Rorke, L.B., Fabiyi, A., Elizan, T.S. and Sever, J.L., 1968. Experimental cerebrovascular
lesions in congenital and neonatal rubella-virus infection of ferrets. Lancet, ii: 153-- 154.
Rudolph, A.J., Singleton, E.B., Rosenberg, H.S., Singer, D.B. and Phillips, C.A., 1965. Osseous manifestations of the congenital rubella syndrome. Am. J. Dis. Child., 110: 428--433.
Rutili, S. and Titoli, F., 1977. Ultrastructural studies on some organs from pigs with swine fever. FAO/CEC expert consultation on the eradication of classical and African swine fever, Sept. 6--11, 1976. Hannover, EUR 5904 EN, Brussels-Luxembourg, pp. 40--51.
Sautter, J.H., Young, G.A., Luedke, A.J. and Kitchell, R.L., 1953. The experimental production of malformations and other abnormalities in fetal pigs by means of attenu- ated hog cholera virus. Proc. Am. Vet. Med. Assoc., Toronto, pp. 146--150.
Scherrer, R., Aynaud, J.M., Cohen, J. and Bic, E., 1970. Etude au microscope ~lectro- nique du virus de la peste porcine classique (hog cholera) dans des coupes ultra-fines de cellules infect~es in vitro. C.R. Acad. Sei., Paris, 271 : 620--623.
Schiff, G.M. and Dine, M.S., 1965. Transmission of rubella from newborns. Am. J. Dis. Child., 110: 447--451.
Schultz, R.D., 1973. Comments on the immune response of the fetus and the neonate. J. Am. Vet. Med. Assoc., 163: 804--806.
Scott, F.W., Kahrs, R.F., DeLahunta, A., Brown, T.T., McEntee, K. and Gillespie, J.H., 1973. Virus induced congenital anomalies of the bovine fetus. I. CerebeUar degenera-

359
tion (hypoplasia), ocular lesions and fetal mummification following experimental in- fection with bovine viral diarrhea--mucosal disease virus. Cornell Vet., 63 : 536--560.
Selzer, G., 1963. Virus isolation, inclusion bodies and chromosomes in rubella infected human embryos. Lancet, ii: 336--337.
Sheffy, B.E., Coggins, L. and Baker, J.A., 1961. Protection of pigs against hog cholera with virus diarrhea of cattle. U.S. Livestock San. Assoc., 65: 347--353.
Simons, M.J. and Fitzgerald, M.G., 1968. Rubella virus and human lymphocytes in cul- ture. Lancet, ii: 937--940.
Simons, M.J. and Jack, I., 1968. Lymphocyte viraemia in congenital rubella. Lancet, ii: 953--954.
Singer, D.B., Rudolph, A.J., Rosenberg, H.S., Rawls, W.E. and Boriuk, M., 1967. Pathol- ogy of the congenital rubella syndrome. J. Pediatr., 71 : 665--675.
Singer, D.B., South, M.A., Montgomery, J.R. and Rawls, W.E., 1969. Congenital rubella syndrome. Lymphoid tissue and immunologic status. Am. J. Dis. Child., 118: 54--61.
Snowdon, W.A. and French, E.L., 1976. The role of mucosal disease virus in the epizo- otiology and control of swine fever virus, Amsterdam, 1975. EUR 5486, Luxembourg, pp. 159--183.
Snowdon, W.A., Parsonson, I.M. and Broun, M.L., 1975. The reaction of pregnant ewes to inoculation with mucosal disease virus of bovine origin. J. Comp. Pathol., 85: 241-- 251.
Solomon, J.B., 1971. Foetal and neonatal immunology. In: A. Neuberger and E.L. Tatum, (Editors), Frontiers of Biology, 20. North-Holland, Amsterdam, 381 pp.
Soothill, J.F., Hayes, K. and Dudgeon, J.A., 1966. Immunoglobulin in congenital rubella. Lancet, i :1385--1386.
South, M.A., Montgomery, J.R. and Rawls, W.E., 1975. Immune deficiency in congenital rubella and other viral infections. Birth Defects, 11: 234--238.
Stern, L.M. and Forbes, I.J., 1975. Dysgammaglobulinaemia and temporary immune paresis in a case of congenital rubella. Aust. Paediatr. J., 11: 38--41.
Stewart, W.C., 1969. Transplacental infection determined by isolation of hog cholera virus in neonatal pigs. M.Sc. Thesis, Iowa State University, IA, 117 pp.
Stewart, W.C., Carbrey, E.A., Jenney, E.W., Brown, C.L. and Kresse, J.I., 1971. Bovine viral diarrhea infection in pigs. J. Am. Vet. Med. Assoc., 159:1556--1563.
Stewart, W.C., Carbrey, E.A. and Kresse, J.I., 1972. Transplacental hog cholera infection in immune sows. Am. J. Vet. Res., 33: 791--798.
Stewart, W.C., Miller, L.D., Kresse, J.I. and Snyder, M.L., 1980. Bovine viral diarrhea in- fection in pregnant swine. Am. J. Vet. Res., 41 : 459--462.
Storey, I.J. and Barlow, R.M., 1972. Experiments with border disease. VI. Lipid and en- zyme histochemistry. J. Comp, Pathol., 82:163--171.
Strayer, P.J., Journ~e, D.L.H. and Binkhorst, G.J., 1983. Neurological disorders, virus persistence, hypomyelination due to intra-uterine infection with bovine virus diarrhoea virus. II. Virology and epizootiology. Vet. Q., in press.
Sweasey, D., Patterson, D.S.P., Richardson, C., Harkness, J.W., Shaw, I.G., and Williams, W.W., 1979. Border disease: a sequential study of surviving lambs and an assessment of its effect on profitability. Vet. Rec., 104: 447--450.
Terlecki, S., 1977. Border disease: a viral teratogen of farm animals. Vet. Annu., 17: 74--79.
Terlecki, S., Hebert, C.N. and Done, J.T., 1973. Morphology of experimental border dis- ease of lambs. Res. Vet. Sci., 15: 310--317.
Terlecki, S., Richardson, C., Done, J.T., Harkness, J.W., Sands, J.J., Shaw, I.G., Winkler, C.E., Duffell, S.J., Patterson, D.S.P. and Sweasey, D., 1980. Pathogenicity for the sheep foetus of bovine virus diarrhoea--mucosal disease virus of bovine origin. Br. Vet. J.. 136: 602-~11 .
Terpstra, C., 1978. Detection of border disease antigen in tissues of affected sheep and in cell cultures by immunofluorescence. Res. Vet. Sci., 25: 350--355.
Terpstra, C., 1981. Border disease: virus persistence, antibody response and transmission studies. Res. Vet. Sci., 30: 185--191.

360
Tesmer, S., Urbaneck, D., Kaden, V., Wittmann, W. and Hahnefeld, H., 1973. Zur Wirkung yon Schweinepest-Lebendvakzine aus dem Impgvirusstamm "C" bei tragen- den Sauen und deren Nachzucht. Monatsh. Veterinaermed., 28: 251--254,
TSndury, G., 1975. Pathogenese angeborener Missbildungen. P~diatr. Fortbildungskurse Prax., 41: 94--109.
TSndury, G. and Smith, D.W., 1966. Fetal rubella pathology. J. Pediatr., 68: 867--879. Townsend, J.J., Baringer, J.R., Wolinsky, J.S., Malamud, N., Melnick, J.L., Panitch, H.S.,
Scott, R.A.T., Oshiro, L.S. and Cremer, N.E., 1975. Progressive rubella panencephal- itis. Late onset after congenital rubella. N. Engl. J. Med., 292: 990--993.
Truitt, R.L. and Shechmeister, I.L., 1973. The replication of bovine viral diarrhea--muco- sal disease virus in bovine leukocytes in vitro. Arch. Ges. Virusforsch., 42: 78--87.
Vaheri, A., Vesikari, T., Oker-Blom, N., Seppala, M., Parkman, P.D., Veronelli, J. and Robbins, F.C., 1972. Isolation of attenuated rubella-vaccine virus from human prod- ucts of conception and uterine cervix. N. Engl. J. Med., 286: 1071--1074.
Van der Logt, J.T.M., Van Loon, A.M. and van der Veen, J., 1980. Replication of rubella virus in human mononuclear blood cells. Infect. Immun., 27: 309--314.
Van der Molen, E.J. and Van Oirschot, J.T., 1981a. Congenital persistent swine fever (hog cholera). I. Pathomorphological lesions in lymphoid tissues, kidney and adrenal. Zbl. Vet. Med. B, 28: 89--101.
Van der Molen, E.J. and Van Oirschot, J.T., 1981b. Congenital persistent swine fever (hog cholera). II. Further pathomorphological observations with special emphasis on epithelial, vascular and nervous tissues. Zbl. Vet. Med., B, 28: 190--204.
Vannier, P., Plateau, E. and Tillon, J.P., 1981. Congenital tremor in pigs farrowed from sows given hog cholera virus during pregnancy. Am. J. Vet. Res., 42: 135--137.
Van Oirschot, J.T., 1977. A congenital persistent swine fever infection. II. Immune re- sponse to swine fever virus and unrelated antigens. Vet. Microbiol., 2: 133--142.
Van Oirschot, J.T., 1979a. Experimental production of congenital persistent swine fever infections. I. Clinical, pathological and virological observations. Vet. Microbiol., 4: 117--132.
Van Oirschot, J.T., 1979b. Experimental production of congenital persistent swine fever infections. II. Effect on functions of the immune system. Vet. Microbiol., 4: 133-- 147.
Van Oirschot, J.T., 1980. Persistent and inapparent infections with swine fever virus of low virulence. Their effects on the immune system. Ph.D. Thesis, State University of Utrecht, 211 pp.
Van Oirschot, J.T. and Terpstra, C., 1977. A congenital persistent swine fever infection. I. Clinical and virological observations. Vet. Microbiol., 2 :121--132.
Vantsis, J.T., Barlow, R.M., Gardiner, A.C. and Linklater, K.A., 1980. The effects of challenge with homologous and heterologous strains of border disease virus on ewes with previous experience of the disease. J. Comp. Pathol., 90: 39--45.
Von Benten, K., Trautwein, G., Richter-Reichhelm, H.B., Liess, B. and Frey, H.R., 1980. Experimental transplacental transmission of hog cholera virus in pigs. III. Histopatho- logical findings in the fetus. Zbl. Vet. Med., B, 27 : 714--724.
Ward, G.M., 1971. Experimental infection of pregnant sheep with bovine viral diarrhoea-- mucosal disease virus. Cornell Vet., 61 : 179--191.
Ward, G.M., Roberts, S.J., McEntee, K. and Gillespie, J.H., 1969. A study of experimen- tally induced bovine viral diarrhea--mucosal disease in pregnant cows and their pro- geny. Cornell Vet., 59: 525--539.
Weil, M.L., Itabashi, H.H., Cremer, N.E., Oshiro, L.S., Lennette, E.H. and Carnay, L., 1975. Chronic progressive panencephalitis due to rubella virus simulating subacute sclerosing panencephalitis. N. Engl. J. Med., 292: 994--998.
Weller, T.H. and Neva, F.A., 1962. Propagation in tissue culture of cytopathic agents from patients with rubella-like illness. Proc. Soc. Exp. Biol. Med., 111 : 215--225.
Westbury, H.A., Napthine, D.V. and Straube, E., 1979. Border disease: persistent infec- tion with the virus. Vet. Rec., 104: 406--409.

361
White, L.R., Leiken, S., Villavivencio, O., Abernathy, W., Avery, G. and Sever, J.L., 1968. Immune competence in congenital rubella. Lymphocyte transformation, delay- ed hypersensitivity, and response to vaccination. J. Pediatr., 73: 229--234.
Whitmore, H.L., Zemjanis, R. and Olson, J., 1981. Effect of bovine viral diarrhea virus on conception in cattle. J. Am. Vet. Med. Assoc., 178: 1065--1067.
Williams, M.P., Brawner, T.A., Riggs, Jr., H.G. and Roehrig, J.T., 1981. Characteristics of a persistent rubella infection in a human cell line. J. Gen. Virol., 52: 321--328.
Willis, R.A., 1962. The Borderland of Embryology and Pathology. Chap. 13. Butter- worths, London, 676 pp.
Wilson, J.G., 1973. Environment and Birth Defects. Academic Press, New York. Woods, W., Johnson, R.T., Hostetler, D.D., Lepow, M.L. and Robbins, F.C., 1966. Im-
munofluorescent studies on rubella infected tissue cultures and human tissues. J. Im- munol., 96: 253--260.
Wrathall, A.E., 1975. Reproductive disorders in pigs. Review series, 11. Commonwealth Agricultural Bureaux, Farnham Royal, Slough, 313 pp.
Wrathall, A.E., Bailey, J., Done, J.T., Shaw, I.G., Winkler, C.E., Gibbons, D.F., Patterson, D.S.P. and Sweasey, D., 1978. Effect of experimental border disease infection in the pregnant sow. Zbl. Vet. Med. B, 25: 62--69.
Young, G.A., Kitchell, R.L., Luedke, A.J. and Sautter, J.H., 1955. The effect of viral and other infections of the dam on foetal development in swine. I. Modified live hog cholera viruses. Immunological, virological and gross pathological studies. J. Am. Vet. Med. Assoc., 126: 165--171.
Zakarian, B., Barlow, R.M. and Rennie, J.C., 1975. Periarteritis in experimental border disease of sheep. I. The occurrence and distribution of the lesion. J. Comp. Pathol., 85 : 453--460.
Copyright © 2022 FDOKUMEN




















