
Complexation of Cadmium and Copper by Fluvial Humic Matter and Effects on Their Toxicity
13
Annali di Chimica, 97, 2007, by Società Chimica Italiana 25 COMPLEXATION OF CADMIUM AND COPPER BY FLUVIAL HUMIC MATTER AND EFFECTS ON THEIR TOXICITY F. CORAMI 1 , G. CAPODAGLIO (°)1,2* , C. TURETTA 2 , M. BRAGADIN 1 , N. CALACE 3 , BM. PETRONIO 3 1 Dipartimento di Scienze Ambientali, Università Ca’ Foscari, Venezia; 2 Istituto per la Dinamica dei Processi Ambientali, CNR-IDPA, Venezia; 3 Dipartimento di Chimica, Università La Sapienza di Roma. Summary - The effects of humic acids and fulvic acids isolated from the River Arno (Italy) on the bioavailability and toxicity of cadmium and copper were assessed in relation to changes in their speciation. Measurements of the complexing capacity of solutions containing these organic ligands were carried out by a titration procedure followed by DPASV and toxicity tests were carried out using lysosomes isolated from rat liver. The complexing capacity of the physiological medium containing about 13 mg/L of humic acids, expressed as ligand concentrations, was 0.30 and 0.072 μmol/L for cadmium and copper respectively; the corresponding conditional stability constants were 4.2 x 10 11 and 1.3 x 10 8 (mol/L) -1 . The complexing capacities of the solution containing the same amount of fulvic acids were 0.33 and 0.164 μmol/L for cadmium and copper respectively, the conditional stability constants were 3.2 x 10 11 and 2.4 x 10 7 (mol/L) -1 . The humic acids reduced the toxicity of cadmium by about 5 times: the EC 50 changed from 4.4 to 20.4 µmol/L. The dose effect curve of copper presented a bi- sigmoid trend and two EC 50 values can be determined: The EC 50(1) in the presence of humic acids changed from 2.0 to 3.1 µmol/L, while the EC 50(2) increased from 22.3 to 45.3 µmol/L. The fulvic acids reduced the cadmium toxicity by about the same amount as humic acids, from 4.4 to 18.6 µmol/L, but they had no effect on copper toxicity. Analysing the chemical speciation of cadmium and copper in the presence of humic components and under toxicity test conditions we can say that the appreciable decrease of EC 50 is not related to changes in their speciation; we can hypothesize that this is due to different processes, as well as to blocking of the lysosomal membrane. On the basis of the shape of the dose-effect curves obtained for cadmium and copper respectively, we can say that the toxic effects of the two metals are different and we can hypothesize that copper could exercise its toxic activity by inhibiting the ATP-driven proton pump and the function of the Cl - selective channel. (°) Corresponding Author; E-mail: [email protected]
Transcript of Complexation of Cadmium and Copper by Fluvial Humic Matter and Effects on Their Toxicity

Annali di Chimica, 97, 2007, by Società Chimica Italiana 25
COMPLEXATION OF CADMIUM AND COPPER BY FLUVIAL HUMIC
MATTER AND EFFECTS ON THEIR TOXICITY
F. CORAMI1, G. CAPODAGLIO(°)1,2*, C. TURETTA2, M. BRAGADIN1, N. CALACE3, BM. PETRONIO3 1Dipartimento di Scienze Ambientali, Università Ca’ Foscari, Venezia; 2Istituto per la Dinamica dei Processi Ambientali, CNR-IDPA, Venezia; 3Dipartimento di Chimica, Università La Sapienza di Roma.
Summary - The effects of humic acids and fulvic acids isolated from the River Arno (Italy) on the bioavailability and toxicity of cadmium and copper were assessed in relation to changes in their speciation. Measurements of the complexing capacity of solutions containing these organic ligands were carried out by a titration procedure followed by DPASV and toxicity tests were carried out using lysosomes isolated from rat liver. The complexing capacity of the physiological medium containing about 13 mg/L of humic acids, expressed as ligand concentrations, was 0.30 and 0.072 µmol/L for cadmium and copper respectively; the corresponding conditional stability constants were 4.2 x 1011 and 1.3 x 108 (mol/L)-1. The complexing capacities of the solution containing the same amount of fulvic acids were 0.33 and 0.164 µmol/L for cadmium and copper respectively, the conditional stability constants were 3.2 x 1011 and 2.4 x 107 (mol/L)-1. The humic acids reduced the toxicity of cadmium by about 5 times: the EC50 changed from 4.4 to 20.4 µmol/L. The dose effect curve of copper presented a bi-sigmoid trend and two EC50 values can be determined: The EC50(1) in the presence of humic acids changed from 2.0 to 3.1 µmol/L, while the EC50(2) increased from 22.3 to 45.3 µmol/L. The fulvic acids reduced the cadmium toxicity by about the same amount as humic acids, from 4.4 to 18.6 µmol/L, but they had no effect on copper toxicity. Analysing the chemical speciation of cadmium and copper in the presence of humic components and under toxicity test conditions we can say that the appreciable decrease of EC50 is not related to changes in their speciation; we can hypothesize that this is due to different processes, as well as to blocking of the lysosomal membrane. On the basis of the shape of the dose-effect curves obtained for cadmium and copper respectively, we can say that the toxic effects of the two metals are different and we can hypothesize that copper could exercise its toxic activity by inhibiting the ATP-driven proton pump and the function of the Cl-
selective channel.
(°) Corresponding Author; E-mail: [email protected]

CORAMI and coworkers 26
INTRODUCTION Details of the mechanism controlling the cellular uptake of trace metals are not completely
known; however it is well known that the bioavailability and toxicity of metals are related to the chemical forms assumed by these elements. Studies to assess the influence of trace metal speciation on their toxicity have led to debatable results. There is evidence that processes able to reduce the ionic fraction of certain trace metals reduce both their bioavailability and toxicity.1-4 On the other hand, there is also evidence that organo-compounds of certain trace metals may be even more toxic than the free ionic fraction.5,6 In fact, the role of trace metal speciation on their bioavailability and their toxicity is closely linked to the mechanisms controlling their transport across the cellular membranes; however we do not know how metal speciation, when inside the cells, affects the action on biological structure.
Complexes of trace metals may be either organic or inorganic. Several organic compounds which are present in the environment may bind trace metals to form complexes; they may be biological products, such as exudates, proteins, peptides, polysaccharides, but they may also be detritical matter and humic substances. About 80% of the dissolved organic matter (DOM) present in natural waters, whose origin may be either terrestrial or marine, is formed by high molecular weight compounds, widely known as humic substances7-9. They are differentiated between humic acids and fulvic acids. Both of them, according to their chemical and structural properties, are able to form stable complexes with polyvalent cations.
In the study of toxicity of dissolved pollutants, many animals, mussels particularly, have been widely used as bioindicators. In these species, exposure to toxicants gives rise to damage of cellular components such as lysosomes;10,11 however their toxic effects are mediated by their transport through the cellular membranes. The lysosomes are acidic sub-cellular structures, where the toxic compounds are accumulated by endocythosis with a consequent reduction of metabolic functional efficiency. Therefore, since the lysosomes are the preferential target of the toxic compounds, they can represent good indicators of environmental exposure to potentially toxic pollutants.
The internal pH of lysosomes is about 4.2. This derives from several processes: the Donnan effect, the presence of an ATP-driven proton pump and the partial reduction of internal potential through a selective Cl- channel activated by this ion. Under these conditions, pH reaches a value of about 4.2.12-14 Toxic effects proportionally increase the internal pH of this sub cellular particle reducing the efficiency of the above processes. Therefore, because the effects are not mediated by other biological structures, the tests using these particles allow for a more rapid and reproducible toxicity measurement.
The aim of this study was to assess the effects of complexation by natural organic ligands, humic acids and fulvic acids isolated from the River Arno (Italy), on the bioavailability and toxicity of both cadmium and copper. Complexation measurements were carried out by a titration procedure followed by DPASV and toxicity tests were carried out using lysosomes isolated from rat liver.
EXPERIMENTAL The samples were made by dissolution of purified humic acid (12.8 mg/L) or fulvic acid (12.6
mg/L) in a physiological medium (KCl 0.125 mol/L, MgCl2 0.010 mol/L, HEPES 0.010 mol/L); this concentration of humic matter is the normal concentration present in natural waters.15-16
The samples were analyzed to determine the initial cadmium and copper concentration in the solution by the HR-ICP-SFMS (Element2, Finnigan-MAT, Bremen, Germany) using the procedure previously described by Turetta et al.(2004)17. The instrument was installed in a dedicated laboratory and the sample introduction system was located inside a protected area under a laminar

Complexation of metals by humic matter and effects on their toxicity 27
flow cabinet. Cadmium was determined in low-resolution mode (m/∆m=400), while Cu was determined in medium resolution mode (m/∆m=4000). The sample introduction system was a µ-flow nebulizer (Elemental Scientific, 100 µl min-1) coupled with a desolvation unit (Aridus, Cetac Technologies, Omaha, NE, USA), coupled with an ASX-100 autosampler (Cetac Technologies, Omaha, NE, USA). Samples were diluted using ultrapure water (1:10 v:v) and acidified with UPA grade HNO3 and a solution of In, Sc, Y and W (final concentration 1 ng ml-1) as an internal standard covering the mass range examined. The quantification of trace elements was carried out by the matched calibration method; the mean metal concentrations (SD) in the sample containing the humic acid were 0.29 (0.03) µmol/L and 0.024 (0.003) µmol/L for Cd and Cu respectively, while the concentration of the same metals in the solution containing fulvic acid were 0.33 (0.03) µmol/L and 0.038 (0.004) µmol/L.
Extraction and separation of humic and fulvic acids.
The humic and fulvic acids used in this study were extracted from the water of the River Arno according to the method of Thurman and Malcolm.18 The humic substance was obtained from a 100 L water sample via adsorption on macromolecular anionic resin (XAD-8). After acidification (pH 2, with HCl) the non-filtered water sample was passed through at column filled with XAD-8; the humic substances were eluted by a counter-current flow of NaOH 0,1 M. The humic acids were separated from the fulvic acids by precipitation after acidification at pH 1 of the alkaline eluate. After 24 h, the humic acids were precipitated while the fulvic acids remained in solution. Humic acids were recovered after centrifugation (3000 rpm for 20 min) and were purified by dialysis (500 Dalton tubular membranes) against distilled water until free from chloride; they were then frozen and lyophilised (lyophilizer Labconco, -60°C, 5.10 mmHg). The solution of fulvic acids was purified from the inorganic components by ultra-filtration (semi-permeable membrane, 500 Dalton, Amicon ) under a pressure of N2 (3.5 atm), the solution was concentrated to a volume of 30 ml. The solution of fulvic acids was then concentrated to a final volume 10 ml, frozen and then lyophilized. This method allows great concentration and purification of samples removing up to 99% of dissolved salts.19
Humic and fulvic acids were then dissolved in NaOH 0.1 M and the solution was passed through a Chelex 100 to remove the metal ions. The solution was then buffered at pH 7.4 (Hepes 0.77 mol/L).
Cadmium and copper speciation
The complexing capacity of the solutions containing humic and fulvic acids for cadmium and copper, expressed as ligand concentration and conditional stability constant, were measured by titration of ligands with metals and following the titration by Differential Pulse Anodic Stripping Voltammetry (DPASV), according to the previously described procedure.20,21 The whole procedure, from the preparation of materials, including the cleaning of sample bottles and vessels, the preparation of the samples, and the DPASV analyses, was carried out in a clean room under a class 100 area.
The electrochemical equipment consisted of a Multipolarograph Analyzer EG&G mod. 384B operating in Differential Pulse mode; the polarograph was equipped with an electrochemical cell EG&G Rotel 2 for ultra – trace metal determination. The working electrode was a pre-formed thin mercury film deposited on a rotating Ultra Trace Graphite Tip (Metrohm, Switzerland). A platinum electrode and Ag/AgCl, KCl saturated electrode were used as auxiliary and reference respectively. The mercury film was prepared by electrolysis from a solution of Hg2+ 10-4 M as previously described.21 The electrochemical cell was rinsed with one aliquot of degassed physiological solution and the sample was placed in the analytical position; the titrimetric procedure was performed by multiple additions until complete saturation of the ligands. The procedure was totally automated by

CORAMI and coworkers 28
means of a personal computer controlling the electrochemical equipment interfaced with a robot (MasterLab System, Model 9000, Perkin Elmer, USA) performing the standard additions. The cleaning procedure for the working electrode has been thoroughly described elsewhere.20 The vessels are acid cleaned with 10-fold diluted Suprapur grade HCl (Merck) and rinsed several times with ultrapure water every time they are used. Cd and Cu standards are prepared in ultrapure water by serial dilution of 1000 mg/L standard solution (Merck Chemicals). The precision of the methodology is about 20% for determination of ligand concentration and 30% for the conditional stability constant, K’. Accuracy of the conditional stability constant obtained by this titrimetric methodology was previously tested using EDTA as a model ligand.22,23
The speciation was calculated by MINEQL+24 using the analytical concentration of inorganic species and the complexing capacity of organic ligands determined according to the procedure reported above. The stability constants of complexes were those present in the MINEQL+ database which were the best available at the moment of compiling (1998); the literature references used for the compilation of the database are reported in the program manual.24 In particular, the thermodynamic constant used for most inorganic complexes originally derives from the USEPA’s MINTEQA2 database.25
Toxicity tests
Lysosomes were prepared from rat liver according to Dell’Antone26, by centrifugation of a lysosomes enriched fraction through a Percoll gradient and resuspended in 20 mM HEPES-MMOPS buffer, pH 7.4, containing 125 mmol/L KCl and 20 mmol/L MgCl2. The protein concentration was monitored using the Lowry procedure.27
The internal pH of the lysosomal matrix was measured using the Acridine Orange (AO) dye. AO is a weak base which, following the uptake mechanism of weak permeant bases, accumulates in the matrix if the pH is acidic.13,14 This accumulation is associated with a chemical shift in the adsorbance maximum12,28 due to a metachromatic effect.26 Therefore, by operating at 492 nm, which is the maximum absorbance of AO in water, a decrease in absorbance indicates the accumulation of AO in the matrix and is a measure of the internal pH since the pH is the driving force for the accumulation of weak permanent bases.
The AO (Merck) was purified before use according to Pal and Shubert.29 The uptake of AO by the lysosomes was monitored spectrophotometrically at room temperature, using a UV-Vis spectrophotometer (Perkin Elmer mod 320). Following the addition of the lysosomes and AO to the incubating medium (final concentration of lysosomes 0,3 ng/ml), the spectrophotometer was adjusted to zero absorbance.
Measurements were carried out by adding 50 µl of the suspension of lysosomes to 2.5 ml of physiological solution and Acridine Orange 8 µmol/L and the absorbance measurement was started. When the absorbance reached a stable value, 20 µl ATP 0.1 mol/L was added,30 the absorbance decreased until it reached a value corresponding to the internal value of pH 4.2. Subsequently 100 µl of humic acids or fulvic acids were added, and later, after about 20 min, standard additions of Cd or Cu were carried out. This increased the absorbance; the increasing speed is related to the toxic effect. The methodology was calibrated before each measurement cycle, the maximum slope, corresponding to 100% activity reduction, was measured by addition of cadmium or copper until a final concentration of 5x10-2 mol/L was reached. The percentage of reduction after each metal addition was obtained from the relative slope with respect to the calibration slope.
Preliminary tests were carried out to check the effect of copper on the selective Cl- channel; for experiments the physiological medium consisted of one solution of K2SO4 0.080 mol/L, MgSO4 0.010 mol/L and HEPES 0.010 mol/L, valinomicine was added to the solution as potassium carrier to compensate the membrane potential. The experimental procedure for tests was the same as that reported above.

Complexation of metals by humic matter and effects on their toxicity 29
Tests consisted in measuring the decrease of lysosomial activity in relation to the increasing metal concentration. The results for cadmium tests assumed the typical sigmoidal dose-effect curve. The value of EC50 and the corresponding slopes were obtained by fitting the experimental data to the Y values obtained by the following equation:
( ) slope/ElogX 50e1100Y −+
= (1)
where X represents the metal concentration and the slope is the sigmoidal curve slope at the EC50 value.
Toxicity tests for copper generated a double sigmoidal curve, thus determining two values of EC50 and slopes, the first curve decreased the lysosome activity to the value DM (see figure 2) and the second from DM to 100%. The reasons for this particular dose-effect curve are so far unknown, only one hypotheses can be made here, and will be the aim of further studies. The values of the two EC50 and slopes were obtained by fitting the experimental data to the following equation 2.
( ) ( ) 2)2(501)1(50 slope/ElogXslope/ElogX e1DM100
e1DMY −− +
−+
+= (2)
The non linear fitting was carried out by the Marquardt procedure.
RESULTS AND DISCUSSION
Complexing Capacity
Tables 1 and 2 report the complexing capacity of solutions containing the humic components for cadmium and copper respectively and the relative conditional stability constants. The mean cadmium concentration (SD) in the solution of humic acids was 0,29 (0.003) µmol/L while it was 0,33 (0.03) µmol/L in the solution containing fulvic acids; the concentration of copper in the same two solutions was 0.024 (0.003) and 0.038 (0.004) µmol/L respectively. Tables 3 and 4 report the cadmium and copper speciation calculated by the MINEQL+ at different metal concentrations, the initial metal content in the reagent and at the values corresponding to EC50 (see below).
The speciation varies significantly at the different metal concentrations in relation to the changes in the metal/organic ligands ratio. The complexing capacity, expressed as ligand concentrations, of the two solutions containing humic and fulvic acids for cadmium was 0.30 and 0.33 µmol/L respectively and the conditional stability constants were higher than 1011 (mol/L)-1; for both the solutions, when the cadmium concentration corresponded to the initial value (metal content in the physiological medium plus humic components), the fraction complexed by organic ligands ranged from 92 to 99% (see table 3). When the cadmium concentration increased to the value corresponding to EC50 (4.4 µmol/L) the organic form of metal contributed very little (about 7%) to the total metal concentration, while the chloride complexes were the more important species and constituted about 80% of the total. That was because the metal concentration was significantly higher than the complexing capacity of the two solutions.
.
CORAMI and coworkers 30
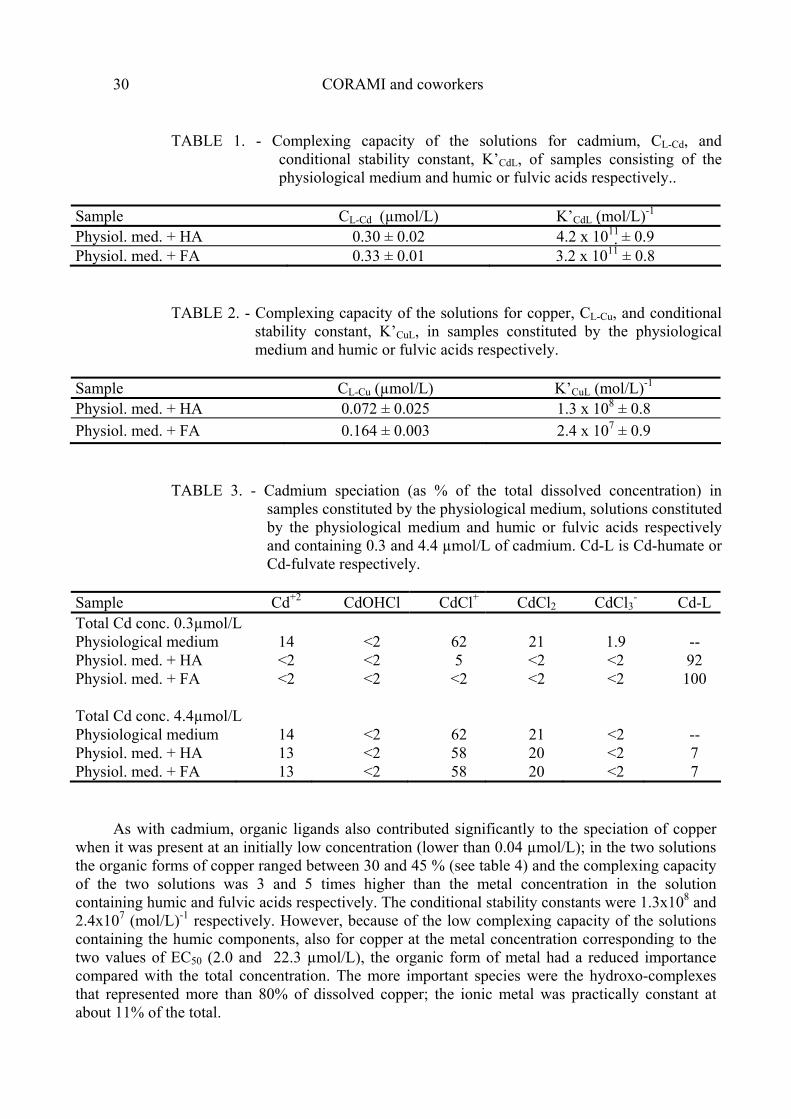
TABLE 1. - Complexing capacity of the solutions for cadmium, CL-Cd, and conditional stability constant, K’CdL, of samples consisting of the physiological medium and humic or fulvic acids respectively..
Sample CL-Cd (µmol/L) K’CdL ( mol/L)-1
Physiol. med. + HA 0.30 ± 0.02 4.2 x 1011 ± 0.9 Physiol. med. + FA 0.33 ± 0.01 3.2 x 1011 ± 0.8
TABLE 2. - Complexing capacity of the solutions for copper, CL-Cu, and conditional
stability constant, K’CuL, in samples constituted by the physiological medium and humic or fulvic acids respectively.
Sample CL-Cu (µmol/L) K’CuL (mol/L)-1
Physiol. med. + HA 0.072 ± 0.025 1.3 x 108 ± 0.8 Physiol. med. + FA 0.164 ± 0.003 2.4 x 107 ± 0.9
TABLE 3. - Cadmium speciation (as % of the total dissolved concentration) in
samples constituted by the physiological medium, solutions constituted by the physiological medium and humic or fulvic acids respectively and containing 0.3 and 4.4 µmol/L of cadmium. Cd-L is Cd-humate or Cd-fulvate respectively.
Sample Cd+2 CdOHCl CdCl+ CdCl2 CdCl3
- Cd-L Total Cd conc. 0.3µmol/L Physiological medium 14 <2 62 21 1.9 -- Physiol. med. + HA <2 <2 5 <2 <2 92 Physiol. med. + FA <2 <2 <2 <2 <2 100 Total Cd conc. 4.4µmol/L Physiological medium 14 <2 62 21 <2 -- Physiol. med. + HA 13 <2 58 20 <2 7 Physiol. med. + FA 13 <2 58 20 <2 7
As with cadmium, organic ligands also contributed significantly to the speciation of copper
when it was present at an initially low concentration (lower than 0.04 µmol/L); in the two solutions the organic forms of copper ranged between 30 and 45 % (see table 4) and the complexing capacity of the two solutions was 3 and 5 times higher than the metal concentration in the solution containing humic and fulvic acids respectively. The conditional stability constants were 1.3x108 and 2.4x107 (mol/L)-1 respectively. However, because of the low complexing capacity of the solutions containing the humic components, also for copper at the metal concentration corresponding to the two values of EC50 (2.0 and 22.3 µmol/L), the organic form of metal had a reduced importance compared with the total concentration. The more important species were the hydroxo-complexes that represented more than 80% of dissolved copper; the ionic metal was practically constant at about 11% of the total.
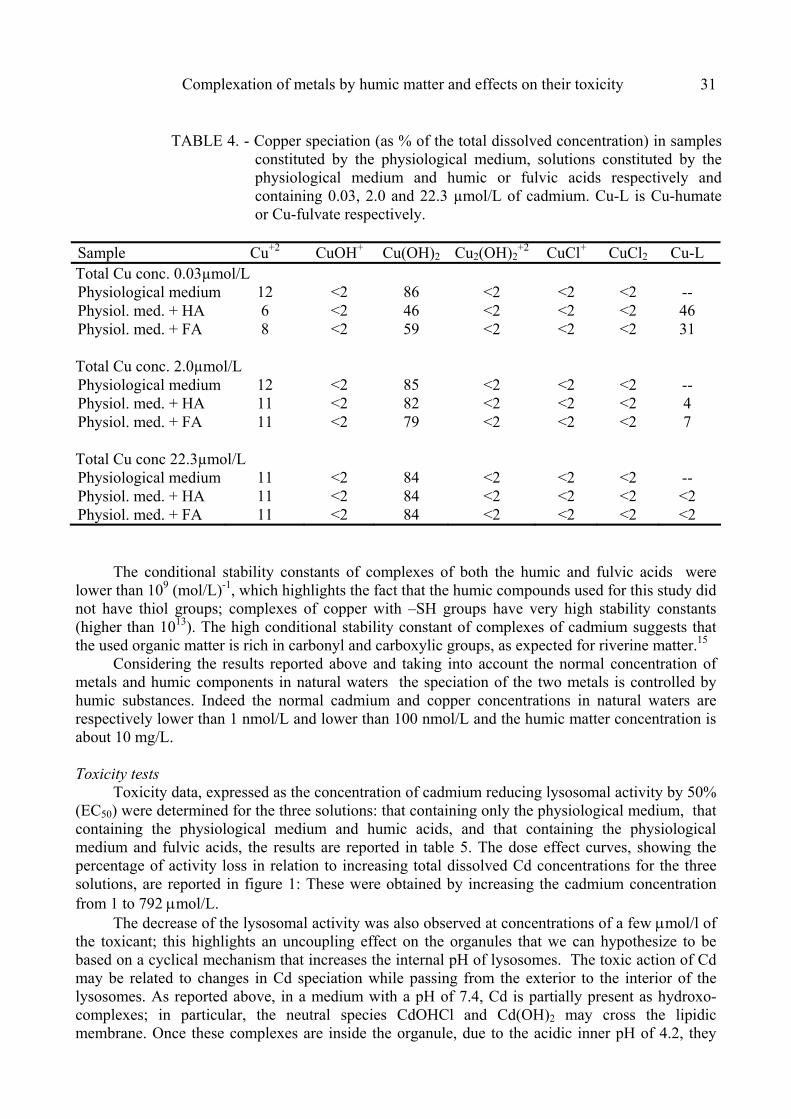
Complexation of metals by humic matter and effects on their toxicity 31
TABLE 4. - Copper speciation (as % of the total dissolved concentration) in samples constituted by the physiological medium, solutions constituted by the physiological medium and humic or fulvic acids respectively and containing 0.03, 2.0 and 22.3 µmol/L of cadmium. Cu-L is Cu-humate or Cu-fulvate respectively.
Sample Cu+2 CuOH+ Cu(OH)2 Cu2(OH)2
+2 CuCl+ CuCl2 Cu-L
Total Cu conc. 0.03µmol/L Physiological medium 12 <2 86 <2 <2 <2 -- Physiol. med. + HA 6 <2 46 <2 <2 <2 46 Physiol. med. + FA 8 <2 59 <2 <2 <2 31 Total Cu conc. 2.0µmol/L Physiological medium 12 <2 85 <2 <2 <2 -- Physiol. med. + HA 11 <2 82 <2 <2 <2 4 Physiol. med. + FA 11 <2 79 <2 <2 <2 7 Total Cu conc 22.3µmol/L Physiological medium 11 <2 84 <2 <2 <2 -- Physiol. med. + HA 11 <2 84 <2 <2 <2 <2 Physiol. med. + FA 11 <2 84 <2 <2 <2 <2
The conditional stability constants of complexes of both the humic and fulvic acids were
lower than 109 (mol/L)-1, which highlights the fact that the humic compounds used for this study did not have thiol groups; complexes of copper with –SH groups have very high stability constants (higher than 1013). The high conditional stability constant of complexes of cadmium suggests that the used organic matter is rich in carbonyl and carboxylic groups, as expected for riverine matter.15
Considering the results reported above and taking into account the normal concentration of metals and humic components in natural waters the speciation of the two metals is controlled by humic substances. Indeed the normal cadmium and copper concentrations in natural waters are respectively lower than 1 nmol/L and lower than 100 nmol/L and the humic matter concentration is about 10 mg/L. Toxicity tests
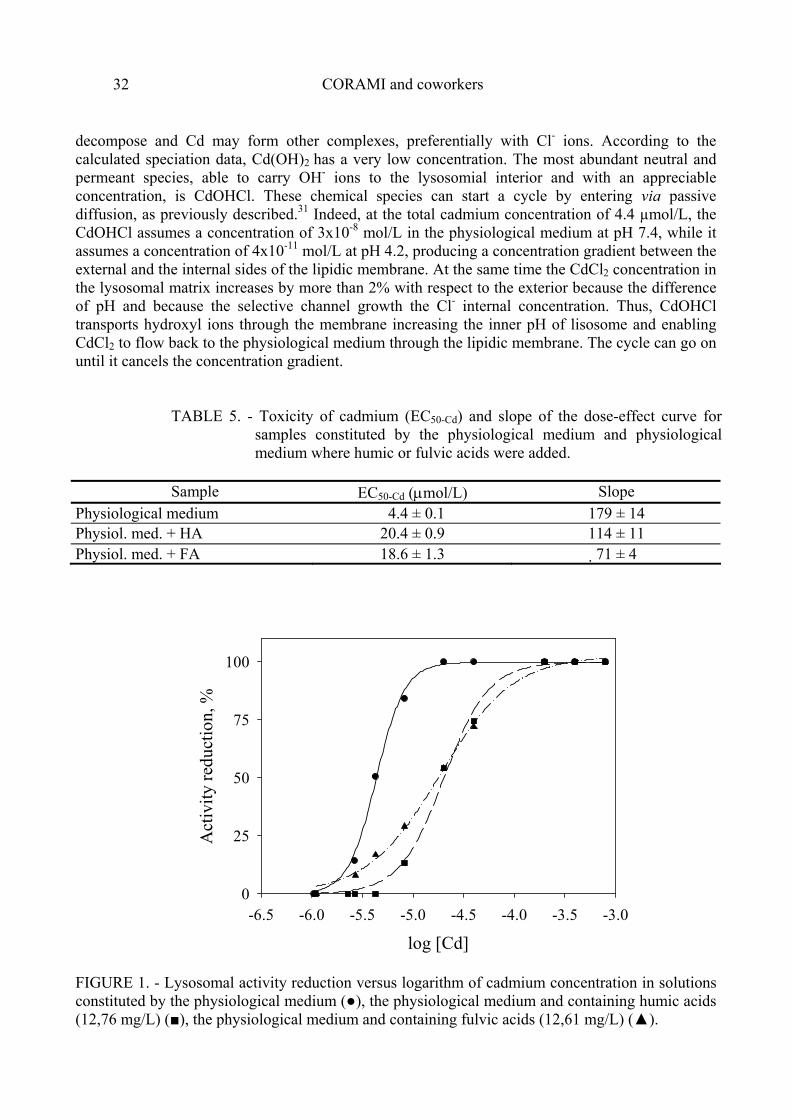
Toxicity data, expressed as the concentration of cadmium reducing lysosomal activity by 50% (EC50) were determined for the three solutions: that containing only the physiological medium, that containing the physiological medium and humic acids, and that containing the physiological medium and fulvic acids, the results are reported in table 5. The dose effect curves, showing the percentage of activity loss in relation to increasing total dissolved Cd concentrations for the three solutions, are reported in figure 1: These were obtained by increasing the cadmium concentration from 1 to 792 µmol/L.
The decrease of the lysosomal activity was also observed at concentrations of a few µmol/l of the toxicant; this highlights an uncoupling effect on the organules that we can hypothesize to be based on a cyclical mechanism that increases the internal pH of lysosomes. The toxic action of Cd may be related to changes in Cd speciation while passing from the exterior to the interior of the lysosomes. As reported above, in a medium with a pH of 7.4, Cd is partially present as hydroxo-complexes; in particular, the neutral species CdOHCl and Cd(OH)2 may cross the lipidic membrane. Once these complexes are inside the organule, due to the acidic inner pH of 4.2, they
CORAMI and coworkers 32
decompose and Cd may form other complexes, preferentially with Cl- ions. According to the calculated speciation data, Cd(OH)2 has a very low concentration. The most abundant neutral and permeant species, able to carry OH- ions to the lysosomial interior and with an appreciable concentration, is CdOHCl. These chemical species can start a cycle by entering via passive diffusion, as previously described.31 Indeed, at the total cadmium concentration of 4.4 µmol/L, the CdOHCl assumes a concentration of 3x10-8 mol/L in the physiological medium at pH 7.4, while it assumes a concentration of 4x10-11 mol/L at pH 4.2, producing a concentration gradient between the external and the internal sides of the lipidic membrane. At the same time the CdCl2 concentration in the lysosomal matrix increases by more than 2% with respect to the exterior because the difference of pH and because the selective channel growth the Cl- internal concentration. Thus, CdOHCl transports hydroxyl ions through the membrane increasing the inner pH of lisosome and enabling CdCl2 to flow back to the physiological medium through the lipidic membrane. The cycle can go on until it cancels the concentration gradient.
TABLE 5. - Toxicity of cadmium (EC50-Cd) and slope of the dose-effect curve for samples constituted by the physiological medium and physiological medium where humic or fulvic acids were added.
Sample EC50-Cd (µmol/L) Slope
Physiological medium 4.4 ± 0.1 179 ± 14 Physiol. med. + HA 20.4 ± 0.9 114 ± 11 Physiol. med. + FA 18.6 ± 1.3 71 ± 4
log [Cd]-6.5 -6.0 -5.5 -5.0 -4.5 -4.0 -3.5 -3.0
Act
ivity
redu
ctio
n, %
0
25
50
75
100
FIGURE 1. - Lysosomal activity reduction versus logarithm of cadmium concentration in solutions constituted by the physiological medium (●), the physiological medium and containing humic acids (12,76 mg/L) (■), the physiological medium and containing fulvic acids (12,61 mg/L) (▲).

Complexation of metals by humic matter and effects on their toxicity 33
However, Bragadin et al suggested that other charged species, such as CdOH+ and Cd2OH+3, might start analogous cycles, given that they can cross the lysosomal membrane by an active mechanism without the presence of transmembrane carriers being required.32 Anyway, whatever the species involved, it is to be remarked that, in each cycle, there is transport of OH- from the exterior to the lysosomal matrix.
As shown in the curves obtained from the experimental data (figure 1), the presence of organic ligands reduces cadmium toxicity. In the presence of humic acids the EC50 value is 20.4 µmol/L and the slope is about 1.6 times lower than that observed for the curve in the absence of ligands. In the presence of fulvic acids the EC50 value is 18.6 µmol/L, which is not significantly different to that observed in the presence of humic acids. However there is a significant difference in the curve slope: in the solution containing fulvic acids it is about 1.6 times lower than that observed for the solution containing humic acids. This means that in the presence of fulvic acids Cd toxicity at low concentrations is higher than in the presence of humic acids. The difference of slope is obviously more pronounced between the solution containing fulvic acids and that containing only the physiological medium; the slope ratio is about 2.5.
As observed above (table 3), organic matter does not change the metal speciation significantly when the metal assumes a concentration corresponding to EC50. It can therefore be excluded that differences of metal speciation are responsible for changes in toxicity. Thus, the toxicity variation might not be linked to any specific chemical species and the toxicity effect might be linked to other factors that are not directly traceable to organic complexation. For example it may derive from the blocking of the membrane. This supports the hypothesis cited above that activity reduction is related to an uncoupling process by a cyclical mechanism.
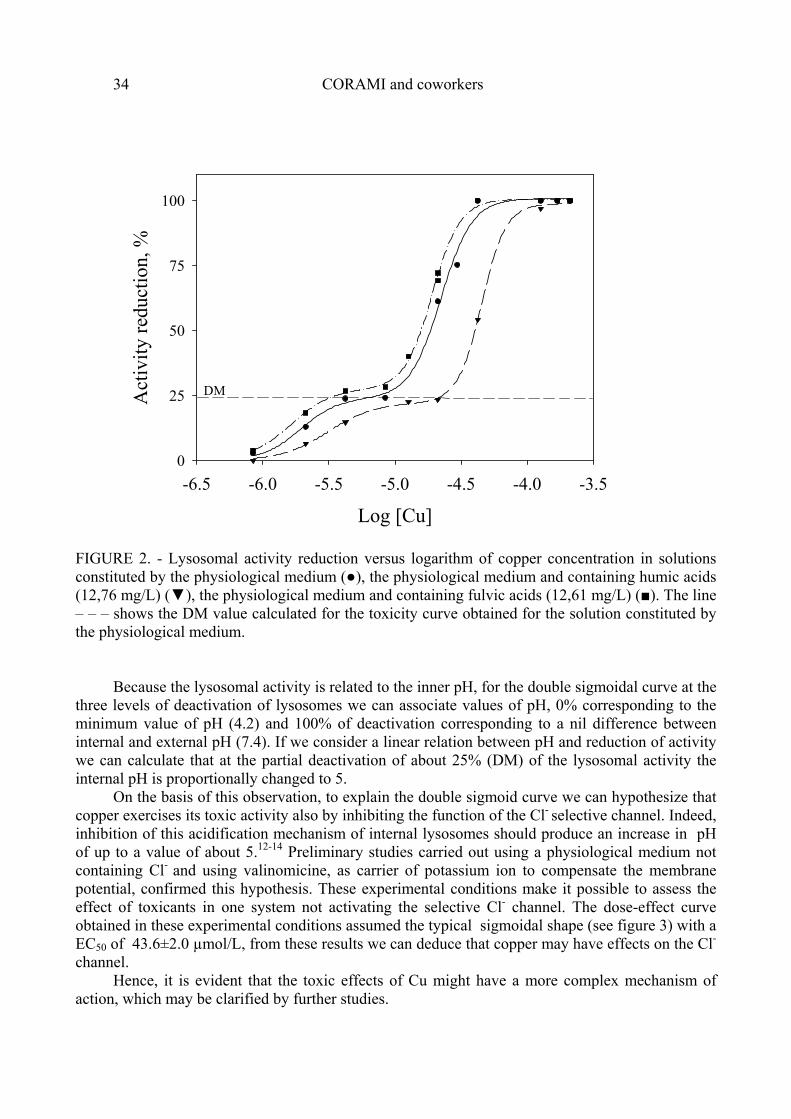
Lysosomal activity in relation to Cu concentrations is shown in fig. 2, where the copper concentration ranges between 0.9 and 844 µM. The two EC50 values and the corresponding slopes of the dose effect curve relative to samples constituted by the physiological medium and the physiological medium containing humic acids or fulvic acids are reported in table 6.
The decrease in lysosomal functions, as reported above, follows a double sigmoid trend. The first at the lowest concentration decreases lysosomal activity by about 25% (DM). The complete inactivation of the lysosomes was observed at concentrations almost one order of magnitude higher. Also for copper, as observed for Cd, the presence of humic acids reduces Cu toxicity, while the fulvic acids seem not to affect it. However, unlike Cd toxicity, the trend of Cu toxicity is always fully explained by a double sigmoid.
As reported above the regulation of interior pH involves more membrane proteins, the Cl- selective channel and ATP pumps. It is also well known that trace metals may compete for the same cofactor site in different enzymatic systems, for example inhibition of the ATP pumps, though that is observed at higher metal concentrations. We can therefore say that copper, as well cadmium, has an uncoupling effect that tends to homogenize the external and internal pH by an analogous mechanism related to the change of speciation with pH. The prevalent species at a copper concentration of 2 or 22 µmol/L and pH of 7.4 are Cu2+ (11% of the total) and complexes with OH-, Cu(OH)2 (more than 80%) (see table 4); the CuCl2 concentration is about 10-9 mol/L (0.1%). With pH at 4.2 the concentration of Cu(OH)2 is 2x10-11 (<<1%) and that of CuCl2 is 10-8 mol/L (5%). Therefore, these differences in the speciation produce the concentration gradients necessary to activate the cyclic mechanism.
CORAMI and coworkers 34
Log [Cu]-6.5 -6.0 -5.5 -5.0 -4.5 -4.0 -3.5
Act
ivity
redu
ctio
n, %
0
25
50
75
100
DM
FIGURE 2. - Lysosomal activity reduction versus logarithm of copper concentration in solutions constituted by the physiological medium (●), the physiological medium and containing humic acids (12,76 mg/L) (▼), the physiological medium and containing fulvic acids (12,61 mg/L) (■). The line – – – shows the DM value calculated for the toxicity curve obtained for the solution constituted by the physiological medium.
Because the lysosomal activity is related to the inner pH, for the double sigmoidal curve at the three levels of deactivation of lysosomes we can associate values of pH, 0% corresponding to the minimum value of pH (4.2) and 100% of deactivation corresponding to a nil difference between internal and external pH (7.4). If we consider a linear relation between pH and reduction of activity we can calculate that at the partial deactivation of about 25% (DM) of the lysosomal activity the internal pH is proportionally changed to 5.
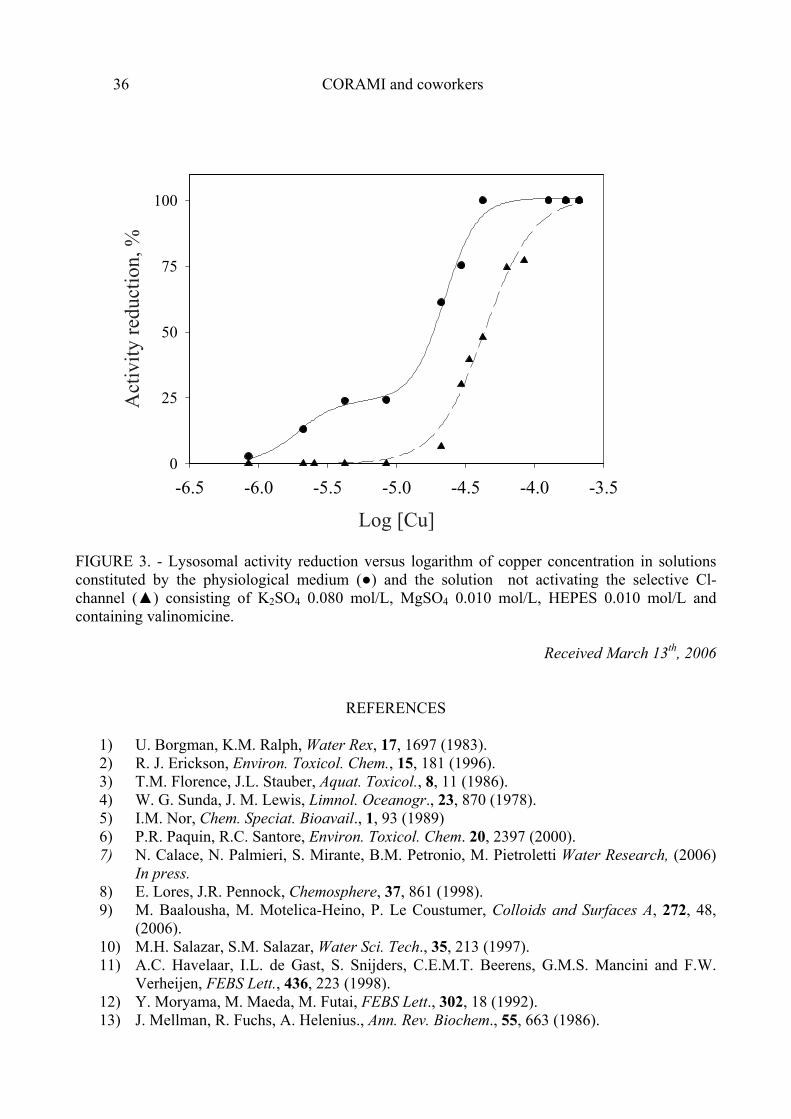
On the basis of this observation, to explain the double sigmoid curve we can hypothesize that copper exercises its toxic activity also by inhibiting the function of the Cl- selective channel. Indeed, inhibition of this acidification mechanism of internal lysosomes should produce an increase in pH of up to a value of about 5.12-14 Preliminary studies carried out using a physiological medium not containing Cl- and using valinomicine, as carrier of potassium ion to compensate the membrane potential, confirmed this hypothesis. These experimental conditions make it possible to assess the effect of toxicants in one system not activating the selective Cl- channel. The dose-effect curve obtained in these experimental conditions assumed the typical sigmoidal shape (see figure 3) with a EC50 of 43.6±2.0 µmol/L, from these results we can deduce that copper may have effects on the Cl- channel.
Hence, it is evident that the toxic effects of Cu might have a more complex mechanism of action, which may be clarified by further studies.

Complexation of metals by humic matter and effects on their toxicity 35
TABLE 6. - Toxicity of copper and slope of the dose-effect curves for samples constituted by the physiological medium and physiological medium containing humic or fulvic acids, the solution constituted of K2SO4, MgSO4, HEPES buffer and containing valinomicine. The two values of EC50.Cu and slopes are the values relative at the two sigmoidal parts of the curves and the DM value is the reduction, in %, of activity corresponding to the first sigmoidal curve.
Sample EC50-Cu(1) Slope (1) DM EC50-Cu(2) Slope (2)
Physiologic medium 2.0 ± 0.6 43 ± 21 24 ± 6 22.3 ± 1.7 204 ± 47 Physiol. med. + HA 3.1 ± 0.5 33 ± 9 22 ± 2 45.3 ± 1.4 244 ± 103 Physiol. med. + FA 1.6 ± 0.2 19 ± 1 28 ± 2 19.2 ± 0.5 191 ± 22 Physiol. med. + valinomicine -- -- -- 43.6 ± 2.0 139 ± 16
CONCLUSIONS The results highlight the fact that coupling lysosome tests and speciation studies can help to
clarify mechanisms controlling trace metal toxicity. The measurements emphasize that in samples containing cadmium and copper at concentrations normally detected in natural waters the humic and fulvic components extracted from river waters may control their speciation. Indeed, when the metal concentration of cadmium and copper is lower than the complexing capacity of these organic ligands more than 95 % of cadmium and more than 30-45% of copper are present in organic form. However, when the metal assume concentrations reducing lysosome activity by 50%, EC50, the organic ligands do not significantly affect the metal speciation; the organic fraction always represents less than 10% of the total metal concentration. The appreciable decrease in the toxicity of cadmium and copper in the presence of humic acid is not related to changes in their speciation; therefore this is probably due to different processes, for example the blocking of the lysosomal membrane. Although fulvic acids form strong complexes with Cd and Cu, the toxicity texts showed that fulvic acids behave differently when complexing Cu rather than Cd. As a matter of fact, the dose effect curve of Cu does not show any significant difference either in the presence or in the absence of fulvic acids. Cd toxicity, on the other hand, is appreciably reduced in the presence of both humic and fulvic acids.
The toxicity tests showed that Cd and Cu have an uncoupling effect activated by changes in the inorganic speciation due to the difference in pH between the external and internal lysosomal membranes. They may enhance the permeability of the lysosomal membrane to OH- by a cyclic mechanism, so they reduce the internal acidification and decrease the membrane potential, that damages the regular metabolic processes. The dose-effect curve shows that Cu acts on two different sites - the deactivation of the ATP pumps and probably the Cl- selective channels - while for Cd we can hypothesize one single site.
According to the values of the conditional stability constants of complexes with Cu and Cd we can say that the humic and fulvic acids extracted from fluvial environments showed, as expected, a reduced number of sulphidrile functional groups (-SH); they also showed that most of them are composed of carboxylic and carbonylic groups.
Studies of speciation and toxicity, also in relation to differences in the properties of the complexing agents, should be carried out on estuarine and coastal waters containing these natural organic ligands to verify if the reported results are also valid for these natural waters that are frequently subject to pollution from metals.
CORAMI and coworkers 36
Log [Cu]-6.5 -6.0 -5.5 -5.0 -4.5 -4.0 -3.5
Act
ivity
redu
ctio
n, %
0
25
50
75
100
FIGURE 3. - Lysosomal activity reduction versus logarithm of copper concentration in solutions constituted by the physiological medium (●) and the solution not activating the selective Cl- channel (▲) consisting of K2SO4 0.080 mol/L, MgSO4 0.010 mol/L, HEPES 0.010 mol/L and containing valinomicine.
Received March 13th, 2006
REFERENCES
1) U. Borgman, K.M. Ralph, Water Rex, 17, 1697 (1983). 2) R. J. Erickson, Environ. Toxicol. Chem., 15, 181 (1996). 3) T.M. Florence, J.L. Stauber, Aquat. Toxicol., 8, 11 (1986). 4) W. G. Sunda, J. M. Lewis, Limnol. Oceanogr., 23, 870 (1978). 5) I.M. Nor, Chem. Speciat. Bioavail., 1, 93 (1989) 6) P.R. Paquin, R.C. Santore, Environ. Toxicol. Chem. 20, 2397 (2000). 7) N. Calace, N. Palmieri, S. Mirante, B.M. Petronio, M. Pietroletti Water Research, (2006)
In press. 8) E. Lores, J.R. Pennock, Chemosphere, 37, 861 (1998). 9) M. Baalousha, M. Motelica-Heino, P. Le Coustumer, Colloids and Surfaces A, 272, 48,
(2006). 10) M.H. Salazar, S.M. Salazar, Water Sci. Tech., 35, 213 (1997). 11) A.C. Havelaar, I.L. de Gast, S. Snijders, C.E.M.T. Beerens, G.M.S. Mancini and F.W.
Verheijen, FEBS Lett., 436, 223 (1998). 12) Y. Moryama, M. Maeda, M. Futai, FEBS Lett., 302, 18 (1992). 13) J. Mellman, R. Fuchs, A. Helenius., Ann. Rev. Biochem., 55, 663 (1986).

Complexation of metals by humic matter and effects on their toxicity 37
14) P. Dell’Antone, M. Bragadin, P. Zatta, Biochim. Biophys. Acta, 1270, 137 (1995). 15) E. Lores, J.R. Pennock., Chemosphere, 38, 293, 1999. 16) F. Mercier, V. Moulin, N. Barrè., F. Casanova., P. Toulhoat, Analytica Chimica Acta, 427,
101, (2001). 17) C. Turetta, G. Cozzi, C. Barbante, G. Capodaglio, P. Cescon. Anal. Bioanal. Chem., 380,
258 (2004). 18) E.M. Thurman, R.L. Malcom, Environ. Sci. Technol., 15, 463 (1981). 19) N. Calace, F. De Paolis, F. Minniti, B.M. Petronio, Talanta, 47, 803 (1998). 20) G. Capodaglio, C. Turetta, G. Toscano, A. Gambaro, G. Scarponi, P. Cescon., Int. J.
Environ. Anal. Chem., 71, 195 (1998). 21) G. Scarponi., G. Capodaglio, C. Barbante, P. Cescon in “Element speciation in
bioinorganic chemistry”, S. Caroli, ed.: J. Wiley & Sons, New York, 1996, Chapt. 11, pp. 363-418.
22) G. Capodaglio, G. Toscano, C. Barbante., A. Gambaro, G. Scarponi, P. Cescon Ann. Chim. (Rome), 92, 163 (2002).
23) K.H. Coale, K.W. Bruland, Limnol. Oceanogr., 33, 1084 (1988). 24) W.D. Schecher, D.C. McAvoy MINEQL+. A Chemical Equilibrium Modelling System,
Version 4.0, Environmental Research Software, Hallowell, USA, 1998. 25) J.W. Boll, D.K. Nordstrom, User’s Manual for WATEQ4F, with Revised Thermodynamic
Data Base and Test Cases for Calculating Speciation of Major Trace, and Redox Elements in Natural Waters. U.S. Geo. Sur. Open-File Report 91-183, Menlo Park, CA, 1991.
26) P. Dell’Antone, R. Colonna, C. Azione,M. Bragadin, Env. J. Biochem., 94, 553 (1972). 27) O.H. Lowry, N.J. Rosenbrough, A.L. Farr, R.J. Randall., J. Biol. Chem., 193, 265 (1951). 28) S. Ohkuma, Y. Mariyama, T. Takano, J. Biochem., 94, 1935 (1983). 29) M.K. Pal, M. Schubert J. Am. Chem. Soc., 84, 4384 (1962). 30) X. Xiao-Song et al. J. Biol. Chem., 264, 18870 (1989). 31) A.P. Dawson, M.J. Selwyn, Biochem. J., 138, 349 (1974). 32) M. Bragadin, G. Perin, A. Giacometti, D. Bottos., S. Manente, Ecology, Environ. Conserv.,
(2006) in press.




















