
Comparison of physico-chemical characteristics of four laccases from different basidiomycetes
11
Comparison of physico-chemical characteristics of four laccases from different basidiomycetes S.V. Shleev a, *, O.V. Morozova a , O.V. Nikitina a , E.S. Gorshina b , T.V. Rusinova b , V.A. Serezhenkov c , D.S. Burbaev c , I.G. Gazaryan d , A.I. Yaropolov a a Laboratory of Chemical Enzymology, A.N. Bach Institute of Biochemistry, Russian Academy of Sciences, Leninsky prospekt 33, 119071 Moscow, Russia b FGUP “GosNIISintezbelok” B. Kommunisticheskaya 27, 109004 Moscow, Russia c N.N. Semenov Institute of Chemical Physics, Russian Academy of Sciences, Kosigina 4, 119977 Moscow, Russia d Department of Chemical Enzymology, Faculty of Chemistry, M.V. Lomonosov Moscow State University, Lenin Hills, 119899, GSP-3 Moscow, Russia Received 7 March 2004; accepted 31 August 2004 Available online 21 September 2004 Abstract New strains of basidiomycetes producing extracellular laccases (Trametes ochracea 92-78, and Trametes hirsuta 56) have been found by screening of isolates of Trametes fungi. The laccases from T. hirsuta 56 and T. ochracea 92-78 as well as two laccases from previously found and described strains of basidiomycetes, namely Cerrena maxima and Coriolopsis fulvocinerea, were purified to homogeneity. The standard redox potentials of type 1 copper in the enzymes were determined and found to be 780, 790, 750, and 780 mV, respectively. The spectral and biochemical studies showed that the enzymes had no significant differences between the structures of their active sites (T1, T2, and T3). In spite of this fact, the basic biochemical properties as well as the redox potentials of the T1 sites of the enzymes were found to be different. The molecular weights of the laccases range from 64 to 70 kDa, and their pI values range from 3.5 to 4.7. The pH-optima are in the range 3.5–5.2. The temperature optimum for activity is about 50 °C. The thermal stabilities of the enzymes were studied. The catalytic and Michaelis constants for catechol, guaiacol, hydroquinone, sinapinic acid, and K 4 Fe(CN) 6 were determined. Based on these results as well as results obtained by comparing with published properties of several laccases, it could be concluded that T. hirsuta and Cerrena maxima laccases have some superior characteristics such as high stability, high activity, and low carbohydrate content, making them attractive objects for further investigations as well as for application in different areas of biotechnology. © 2004 Elsevier SAS. All rights reserved. Keywords: Basidiomycete; Laccase; Redox potential; T1 site; Active site 1. Introduction Laccase (benzenediol: oxygen oxidoreductases, EC 1.10.3.2) catalyses the oxidation of ortho- and para- diphenols, aminophenols, polyphenols, polyamines, lignins and aryl diamine as well as some inorganic ions coupled to the reduction of molecular dioxygen to water [1,2]. The enzyme exhibits a broad substrate specificity, which can be enhanced by addition of redox mediators. The efficiency of these mediators has been demonstrated in work presented in a number of publications [1,3]. Due to the broad variety of reactions catalysed by laccases, this enzyme holds great promise for many potential applications. Laccases are classified into two groups in accordance with their source, i.e. plant and fungal. However, diphenol oxi- dases (laccases) have also been identified in bacteria [4,5] and insects [6]. The enzyme is a copper protein and contains four metal ions classified into three types, referred to as T1, T2, and T3 [1,2]. The T1 copper is responsible for the blue color of the enzyme and has a characteristic absorbance around 610 nm. The T2 copper cannot be detected spectro- photometrically; however, it generates a characteristic EPR signal [2,7,8]. The bi-nuclear T3 copper is diamagnetic. It displays a spectral absorbance shoulder in the region of 330 nm and also displays a characteristic fluorescence spec- trum [9]. The key characteristic of laccase is the standard redox potential of the T1 site. The values of the redox potential of * Corresponding author. Tel.: +7-95-954-44-77; fax: +7-95-954-40-07. E-mail address: [email protected] (S.V. Shleev). Biochimie 86 (2004) 693–703 www.elsevier.com/locate/biochi 0300-9084/$ - see front matter © 2004 Elsevier SAS. All rights reserved. doi:10.1016/j.biochi.2004.08.005
Transcript of Comparison of physico-chemical characteristics of four laccases from different basidiomycetes

Comparison of physico-chemical characteristics of four laccasesfrom different basidiomycetes
S.V. Shleev a,*, O.V. Morozova a, O.V. Nikitina a, E.S. Gorshina b, T.V. Rusinova b,V.A. Serezhenkov c, D.S. Burbaev c, I.G. Gazaryan d, A.I. Yaropolov a
a Laboratory of Chemical Enzymology, A.N. Bach Institute of Biochemistry, Russian Academy of Sciences, Leninsky prospekt 33, 119071 Moscow, Russiab FGUP “GosNIISintezbelok” B. Kommunisticheskaya 27, 109004 Moscow, Russia
c N.N. Semenov Institute of Chemical Physics, Russian Academy of Sciences, Kosigina 4, 119977 Moscow, Russiad Department of Chemical Enzymology, Faculty of Chemistry, M.V. Lomonosov Moscow State University, Lenin Hills, 119899, GSP-3 Moscow, Russia
Received 7 March 2004; accepted 31 August 2004
Available online 21 September 2004
Abstract
New strains of basidiomycetes producing extracellular laccases (Trametes ochracea 92-78, and Trametes hirsuta 56) have been found byscreening of isolates of Trametes fungi. The laccases from T. hirsuta 56 and T. ochracea 92-78 as well as two laccases from previously foundand described strains of basidiomycetes, namely Cerrena maxima and Coriolopsis fulvocinerea, were purified to homogeneity. The standardredox potentials of type 1 copper in the enzymes were determined and found to be 780, 790, 750, and 780 mV, respectively. The spectral andbiochemical studies showed that the enzymes had no significant differences between the structures of their active sites (T1, T2, and T3). Inspite of this fact, the basic biochemical properties as well as the redox potentials of the T1 sites of the enzymes were found to be different. Themolecular weights of the laccases range from 64 to 70 kDa, and their pI values range from 3.5 to 4.7. The pH-optima are in the range 3.5–5.2.The temperature optimum for activity is about 50 °C. The thermal stabilities of the enzymes were studied. The catalytic and Michaelisconstants for catechol, guaiacol, hydroquinone, sinapinic acid, and K4Fe(CN)6 were determined. Based on these results as well as resultsobtained by comparing with published properties of several laccases, it could be concluded that T. hirsuta and Cerrena maxima laccases havesome superior characteristics such as high stability, high activity, and low carbohydrate content, making them attractive objects for furtherinvestigations as well as for application in different areas of biotechnology.© 2004 Elsevier SAS. All rights reserved.
Keywords: Basidiomycete; Laccase; Redox potential; T1 site; Active site
1. Introduction
Laccase (benzenediol: oxygen oxidoreductases, EC1.10.3.2) catalyses the oxidation of ortho- and para-diphenols, aminophenols, polyphenols, polyamines, ligninsand aryl diamine as well as some inorganic ions coupled tothe reduction of molecular dioxygen to water [1,2]. Theenzyme exhibits a broad substrate specificity, which can beenhanced by addition of redox mediators. The efficiency ofthese mediators has been demonstrated in work presented ina number of publications [1,3]. Due to the broad variety ofreactions catalysed by laccases, this enzyme holds greatpromise for many potential applications.
Laccases are classified into two groups in accordance withtheir source, i.e. plant and fungal. However, diphenol oxi-dases (laccases) have also been identified in bacteria [4,5]and insects [6]. The enzyme is a copper protein and containsfour metal ions classified into three types, referred to as T1,T2, and T3 [1,2]. The T1 copper is responsible for the bluecolor of the enzyme and has a characteristic absorbancearound 610 nm. The T2 copper cannot be detected spectro-photometrically; however, it generates a characteristic EPRsignal [2,7,8]. The bi-nuclear T3 copper is diamagnetic. Itdisplays a spectral absorbance shoulder in the region of330 nm and also displays a characteristic fluorescence spec-trum [9].
The key characteristic of laccase is the standard redoxpotential of the T1 site. The values of the redox potential of
* Corresponding author. Tel.: +7-95-954-44-77; fax: +7-95-954-40-07.E-mail address: [email protected] (S.V. Shleev).
Biochimie 86 (2004) 693–703
www.elsevier.com/locate/biochi
0300-9084/$ - see front matter © 2004 Elsevier SAS. All rights reserved.doi:10.1016/j.biochi.2004.08.005

this site have been determined using potentiometric titrationswith redox mediators for a great number of different laccasesand they vary between 430 and 790 mV vs. NHE [10–15]. Ithas been shown for some laccases that T1 is the primarycenter at which electrons are accepted from reducing sub-strates [1,2,11]. Moreover, the catalytic efficiency (kcat/KM)for some reducing substrates depends on the redox potentialof the T1 copper [11,16]. This is the reason why laccases witha high redox potential T1 site are of special interest inbiotechnology, e.g., for different bleaching [17] and biore-mediation processes [18].
The primary structures of laccase isolated from severalorganisms have been determined (for example, see the Gen-Bank website: http://www.ncbi.nlm.nih.gov). It was shownthat two multiple forms of laccase from one origin (forinstance, basidiomycete T. hirsuta) with different biochemi-cal properties had very similar primary structures (Ref. [19]and also GenBank). On the other hand, it has also been shownthat while being similar biochemically, the isoenzymes oflaccase from the same origin (T. hirsuta) can have differentprimary structures [9]. Currently, it is not absolutely clearwhether basidiomycetes have several genes encoding lacca-ses, or whether the diversity of laccase isoenzymes is due topostranslational modification and/or proteolysis during cul-tivation and purification.
Laccase was first described by Yoshida as early as 1883[20]. This makes it one of the first enzymes ever described.However, during the last few years of work on laccase appli-cations has been extended. Reported applications includedevelopment of oxygen cathodes in biofuel cells [21], bio-sensors [22], green biodegradation of xenobiotics includingpulp bleaching [3,17], and labelling in immunoassays [23].Moreover, new interesting prospective directions for the ap-plication of the enzyme in biotechnology have been found:bioremediation [18], green organic synthesis [24], and evendesign of laccase fungicidal and bactericidal preparations[25]. These applications stimulate new waves of fundamentalresearch concerning this enzyme. Activities of current inter-est include screening of laccase sources, studying new lacca-ses [5,9,15,16,26,27], investigating the structure of the en-zyme [28–31], elucidating the mechanism of the intraproteinelectron transfer as well as the mechanism of oxygen reduc-tion to water [32,33], investigating the electrochemical prop-erties of laccases [34], and much more.
There is no doubt that biochemical and physico-chemicalproperties of laccase (activity, stability, carbohydrate con-tent, pH-optimum, etc.) comprise very important initial in-formation for both fundamental studies and for application oflaccases in biotechnology. The great differences in theseproperties even for different laccases from a single speciesmotivate the importance of broad biochemical characteriza-tion of any enzyme which will be the focus of further work.Thus, the goals of the present paper were (i) to screen newstrains of basidiomycetes for laccase secretion; (ii) to studyand thoroughly compare biochemical, spectral, kinetic, andelectrochemical properties of four laccases from strains se-
lected in the first step of our study; and (iii) to reveal thepossible nature of the differences in their key characteristicsin order to find new laccases as attractive objects for furtherinvestigations as well as for applications in different areas ofbiotechnology.
2. Materials and methods
2.1. Chemicals
DEAE Toyopearl 650M and Toyopearl HW-55 were fromTosoh (Tokyo, Japan); ampholine was from Pharmacia LKBBiotechnology (Bromma, Sweden); Coomassie G-250 andR-250, and markers for electrophoresis and chromatographywere from Serva (Heidelberg, Germany); 2-mercaptoethanolwas from Ferak (Berlin, Germany); Tris and glycine werefrom ICN (Costa Mesa, CA, USA); Servacel DEAE 52,acrylamide, N,N-methylenebisacrylamide and ammoniumpersulfate were from Reanal (Budapest, Hungary); catechol,guaiacol, and sinapinic acid were from Sigma (St. Louis,MO, USA); hydroquinone, 2,2′-biquinoline, NaCl,Na2HPO4, KH2PO4, K3Fe(CN)6 and K4Fe(CN)6 were fromMerck (Darmstadt, Germany); and glycerol, acetic acid,methanol, ethanol, MoO2, HCl, KCl, H3PO4, (NH4)2SO4 andNaOH were of the highest purity available from domesticsources. Buffers were prepared using water (18 MX) purifiedwith a Milli-Q system (Millipore, Milford, CT, USA). Themediators (K4[Mo(CN)8] and K3[Mo(CN)8]) were synthe-sised and purified according to previously published methods[35].
2.2. Material
Basidiomycetes Cerrena maxima (Fr.) Ryvarden and Co-riolopsis fulvocinerea (Murrill) Zmitr. were obtained fromthe Komarov Botanical Institute, Russian Academy of Sci-ences (St. Petersburg, Russia). Partially purified preparationsof extracellular laccases from these basidiomycetes havebeen kindly provided by Drs. O.V. Koroleva and V.P.Gavrilova in the framework of the joint research performedwith the support of INCO Copernicus Grant No. ICA2-CT-2000-10050.
Basidiomycetes Trametes hirsuta (Wulfen) Pilát andTrametes ochracea (Pers.) Gib. & Ryvarden were obtainedfrom the laboratory collection of the State Research Instituteof Protein Biosynthesis (Moscow, Russia). A list of strainswhich have been screened in the present study appears inTable 1 (top line).
2.3. Microbiological procedures
The Cerrena maxima and Coriolopsis fulvocinerea ba-sidiomycetes were grown as described in [8].
Basidiomycetes T. hirsuta and T. ochracea were grown bysubmerged cultivation using the following nutrient media
694 S.V. Shleev et al. / Biochimie 86 (2004) 693–703

(g/l): glucose, 10; peptone, 3; KH2PO4, 0.6; K2HPO4, 0.4;MgSO4, 0.5; ZnSO4, 0.001; FeSO4, 0.0005; MnSO4, 0.05;and CuSO4, 0.25. The cultivation was carried out in a fer-menter “Marubishi” (Japan) with a volume of 30 l under thefollowing conditions: pressure, 0.4 bar; temperature, 27 °C;aeration, 1.0 ppm; and agitation speed, 250 rpm. Duringcultivation, the pH of the solutions, as well as some bio-chemical parameters such as oxidase activity and biomass,were controlled. The nutrient media were inoculated withagar plugs (8 mm in diameter) of mycelium maintained on2% malt extract agar. The oxidase activity test was performedon the agar media by Bavendamm’s reaction in accordancewith a method presented in [36].
2.4. Isolation and purification procedure
The partly purified preparations of Cerrena maxima andCoriolopsis fulvocinerea laccases were received in the formsdescribed in [8,26].
After cultivation, the culture fluids of T. hirsuta 56 andT. ochracea 92–83 basidiomycetes containing extracellularlaccases were filtered to remove the mycelium. The usedpurification scheme included: precipitation of the enzymesfrom the cultural medium with saturated ammonium sulfate;FPLC ion-exchange chromatography on a Servacel DEAE52; rechromatography on a DEAE-Toyopearl 650M per-formed on standard equipment purchased from PharmaciaLKB Biotechnology; isoelectric focusing in the pH range3–5, then gel-filtration on Toyopearl HW-55 from Tohos.
The final purification step for all laccases including par-tially purified enzymes from Cerrena maxima and Coriolop-sis fulvocinerea was performed by means of HPLC on a TSKDEAE-2SW column from Pharmacia LKB Biotechnologyusing a “Stayer” system from Aquilon Ltd. (Moscow, Russia,web page: http://www.aquilab.ru/default_en.htm).
The enzyme preparations were homogeneous as judgedfrom SDS-PAGE [37]. The specific activity of each enzymepreparations was expressed in µM/min per mg of protein withcatechol as substrate. Homogeneous preparations of laccase(ca. 10 mg/ml) were stored in 0.1 M phosphate buffer, pH6.5, at –18 °C.
2.5. Biochemical characterization
Preparative isoelectric focusing was performed in accor-dance with [38] on a 440 ml Pharmacia LKB Biotechnologycolumn 8100 using a natural pH gradient 3–5 stabilized by agradient of glycerol (10–60%) The applied power was 30 Wat the beginning and 3 W at the end of the experiment.Fractions were collected at a flow rate of 4 ml/min.
Determination of carbohydrate content involved the de-rivatization of the monosaccharides released after acid hy-drolysis (incubation in 4 M CF3COOH for 3 h at 110 °C), andtheir subsequent analysis using a Biotronic LC 2000 analyzer(Maintal, Germany).
The molecular mass of laccase was determined by gel-filtration on TSK G 2000 SWXL column from Tosoh, cali-brated with a set of standard markers (15–92 kDa) fromServa. The assay of laccase-bound copper was performedwith 2,2′-biquinoline [39]. The protein was determined inaccordance with methodology presented in [40].
2.6. Spectral investigation
Absorbance spectra were recorded with a Hitachi-557 spectrophotometer (Tokyo, Japan) with different scanrates and slit widths. Fluorescent spectra were recorded witha Shimadzu spectrofluorophotometer RF-5301 (Tokyo, Ja-pan) with a scan rate of 120 nm/min and a bandwidth of 5 nm.EPR-spectra were registered with a Bruker EPR-spectrometer ESC-106 (Ettlingen, Germany) in the X-band.CD-spectra were recorded using a Jasco spectropolarimeterJ-715 (Tokyo, Japan) with a scan rate of 60 mm/min and abandwidth of 5 nm. The parameters of the secondary struc-tures were calculated using an in house written computerprogram “Belok” (Moscow, Russia). The program was de-signed by Dr. Vladimir V. Shubin (A.N. Bach Institute ofBiochemistry, e-mail: [email protected]).
2.7. Electrochemical studies
To determine the redox potentials of laccase T1 centers,the method of protein redox titration [41] was employed withpotassium octocyanomolibdate (IV and V) mediators. The
Table 1Dynamics of oxidase activity and biomass increase for different strains of Trametes fungi (T. hirsuta (Th) and T. ochracea (To))
Time Parameters Th NRB2 Th 86–43 Th 56 To 92-78 To 92-83 To 117 To M 103 To M 106 To 2397 days Oxidase activity (U/ml) 1.5 0.3 5.8 0.3 0.6 0.3 0.5 0.3 0.2
Biomass (mg/ml) 4.1 8.2 6.1 3.7 5.0 2,9 5.3 2.7 2.59 days Oxidase activity (U/ml) 2.5 0.8 4.6 2.0 0.9 0.4 0.9 0.4 0.8
Biomass (mg/ml) 5.3 8.5 7.8 4.9 6.6 5.9 8.3 5.0 3.812 days Oxidase activity (U/ml) 1.6 a 9,1 0.2 2,4 0.5 0.8 0.5 2.2
Biomass (mg/ml) 6.3 7.6 10.4 4.9 6.5 6.4 8.1 7.9 4.714 days Oxidase activity (U/ml) 0.6 0.5 6.4 0.6 0.7 0.6 0.8 a 1.7
Biomass (mg/ml) 7.0 6.7 12.8 4.9 5.8 7.3 7.79 9.2 5.1
Notes: Strains selected for further study are indicated in bold.a indicates that only traces of activity were observed. Oxidase activity is presented in units per ml of culture fluid (U/ml). One unit of activity is defined as the
amount of laccase oxidizing 1 µmol of catechol per minute under standard condition. The S.E.M. has been calculated from three independent experiments anddid not exceed 10%.
695S.V. Shleev et al. / Biochimie 86 (2004) 693–703
direct potential measurement on platinum electrodes wasperformed while simultaneously recording the absorbancespectra using a bioelectrochemical cell [42]. The potentialwas measured with an ionomer I-130.1 from APK Energos-ervis (St. Petersburg, Russia). Platinum wire with 1 mmdiameter was used as a working electrode, and an Ag/AgClelectrode EVL-1M3 from BioEko (Moscow, Russia) with E=201 ±2 mV was used as a reference electrode. Laccaseswere placed anaerobically into a cell containing a high con-centration of K3Mo(CN)8 in 0.1 M phosphate buffer, pH 6.0.Redox potentials were registered with platinum electrodesbefore and after the addition of enzymes. Further titrationwas performed with the reduced form of the mediator(K4Mo(CN)8) in 0.1 M phosphate buffer, pH 6.0. The absor-bance spectra within the range from 550 to 800 nm wererecorded after each addition after equilibrium was estab-lished. The calculations accounted for enzyme dilution,which never exceeded 10%. Anaerobic conditions were pro-vided by argon purified through a system of solutions con-taining VCl2 [43].
3. Results
As was mentioned in Section 1, one of the goals of presentwork was to study and thoroughly compare the properties oflaccases from various basidiomycetes. As a start, two en-zymes from different fungi, namely Cerrena maxima andCoriolopsis fulvocinerea, were kindly provided by our Rus-sian colleagues. To broaden the scope of the investigations,the first task was to select additional strains for laccaseproduction from our collection of Trametes fungi.
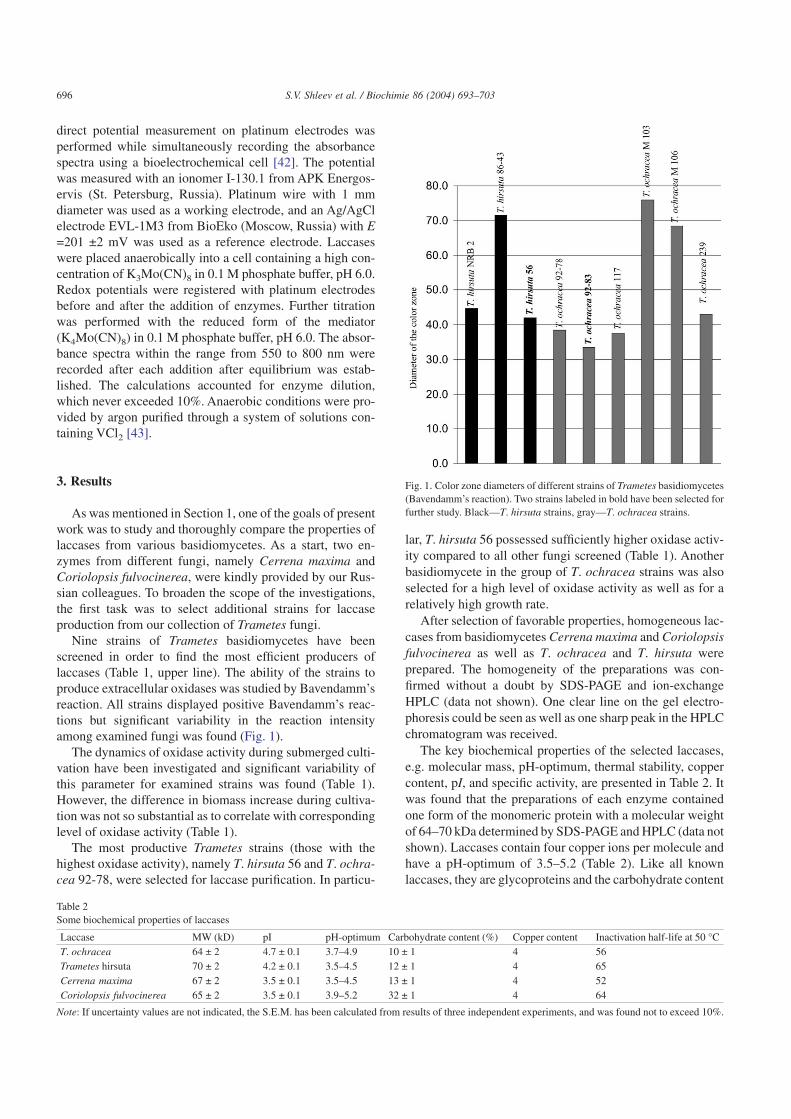
Nine strains of Trametes basidiomycetes have beenscreened in order to find the most efficient producers oflaccases (Table 1, upper line). The ability of the strains toproduce extracellular oxidases was studied by Bavendamm’sreaction. All strains displayed positive Bavendamm’s reac-tions but significant variability in the reaction intensityamong examined fungi was found (Fig. 1).
The dynamics of oxidase activity during submerged culti-vation have been investigated and significant variability ofthis parameter for examined strains was found (Table 1).However, the difference in biomass increase during cultiva-tion was not so substantial as to correlate with correspondinglevel of oxidase activity (Table 1).
The most productive Trametes strains (those with thehighest oxidase activity), namely T. hirsuta 56 and T. ochra-cea 92-78, were selected for laccase purification. In particu-
lar, T. hirsuta 56 possessed sufficiently higher oxidase activ-ity compared to all other fungi screened (Table 1). Anotherbasidiomycete in the group of T. ochracea strains was alsoselected for a high level of oxidase activity as well as for arelatively high growth rate.
After selection of favorable properties, homogeneous lac-cases from basidiomycetes Cerrena maxima and Coriolopsisfulvocinerea as well as T. ochracea and T. hirsuta wereprepared. The homogeneity of the preparations was con-firmed without a doubt by SDS-PAGE and ion-exchangeHPLC (data not shown). One clear line on the gel electro-phoresis could be seen as well as one sharp peak in the HPLCchromatogram was received.
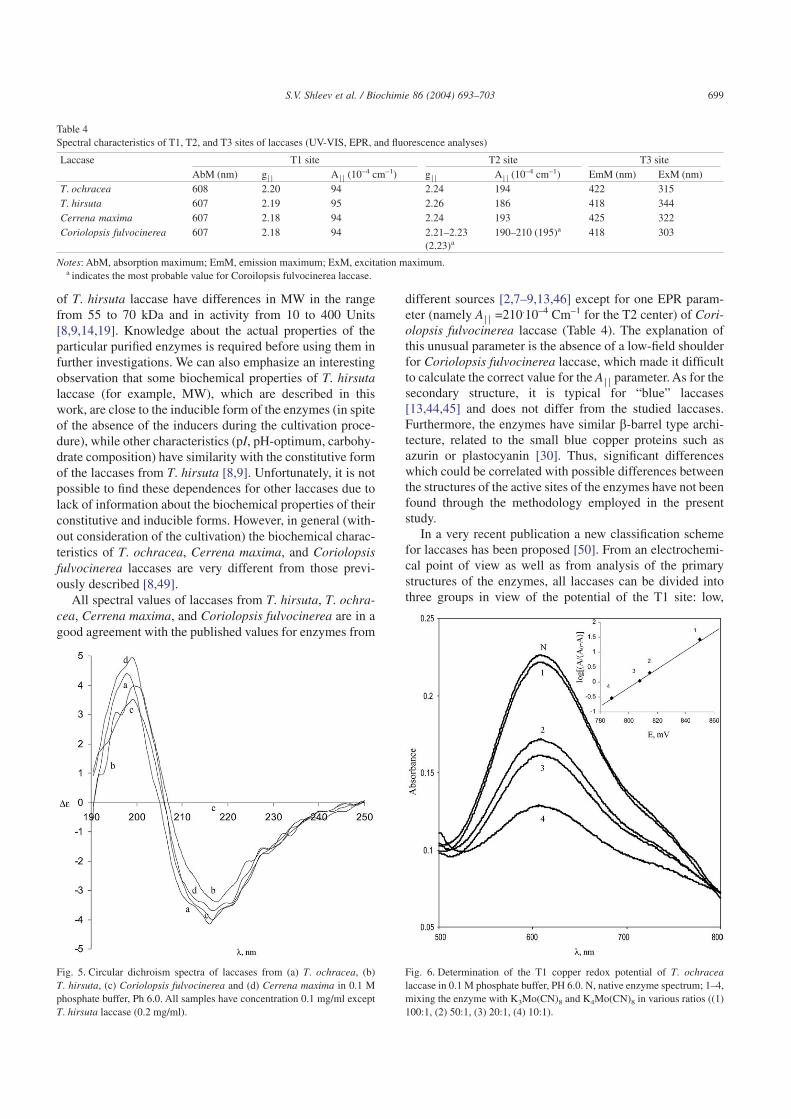
The key biochemical properties of the selected laccases,e.g. molecular mass, pH-optimum, thermal stability, coppercontent, pI, and specific activity, are presented in Table 2. Itwas found that the preparations of each enzyme containedone form of the monomeric protein with a molecular weightof 64–70 kDa determined by SDS-PAGE and HPLC (data notshown). Laccases contain four copper ions per molecule andhave a pH-optimum of 3.5–5.2 (Table 2). Like all knownlaccases, they are glycoproteins and the carbohydrate content
Fig. 1. Color zone diameters of different strains of Trametes basidiomycetes(Bavendamm’s reaction). Two strains labeled in bold have been selected forfurther study. Black—T. hirsuta strains, gray—T. ochracea strains.
Table 2Some biochemical properties of laccases
Laccase MW (kD) pI pH-optimum Carbohydrate content (%) Copper content Inactivation half-life at 50 °CT. ochracea 64 ± 2 4.7 ± 0.1 3.7–4.9 10 ± 1 4 56Trametes hirsuta 70 ± 2 4.2 ± 0.1 3.5–4.5 12 ± 1 4 65Cerrena maxima 67 ± 2 3.5 ± 0.1 3.5–4.5 13 ± 1 4 52Coriolopsis fulvocinerea 65 ± 2 3.5 ± 0.1 3.9–5.2 32 ± 1 4 64
Note: If uncertainty values are not indicated, the S.E.M. has been calculated from results of three independent experiments, and was found not to exceed 10%.
696 S.V. Shleev et al. / Biochimie 86 (2004) 693–703

varies from 10% to 32%. It was found that the carbohydratemoieties of the enzymes consisted of mannose andN-acetylglucosamine (for T. hirsuta, Cerrena maxima, andCoriolopsis fulvocinerea laccases), or mannose, N-acetyl-glucosamine, and galactose (for T. ochracea laccase). Thelaccases are all highly stable enzymes retaining 50% of theirinitial activity after incubation at 50 °C for about 60 h.Afterwards kinetic investigations of the laccases were per-formed and the catalytic constants (kcat and KM) measured forsome organic and inorganic compounds are presented inTable 3. As can be seen from Table 3 T. hirsuta laccasepossessed the largest catalytic constants with respect to allorganic substrates, but Cerrena maxima laccase showed thehighest oxidation rate for the inorganic substanceK4[Fe(CN)6]. Sinapinic acid is the best substrate for T. hir-suta, T. ochracea, and Coriolopsis fulvocinerea laccase,whereas K4[Fe(CN)6] is the best substrate for the enzymefrom Cerrena maxima basidiomycete.
In order to investigate the structure of the active sites ofthe enzymes, spectral studies have been performed. Absor-bance spectra presented in Fig. 2 are typical for “blue”laccases and contain signals correlated with T1 and T3 cop-
per centers (about 610 and 330 nm, respectively). Fluores-cence spectra, characteristic of the T3 site of the enzymes, arealso not so different for the examined laccases (Fig. 3). TheEPR spectra of the laccases under study are presented inFig. 4. They are also typical for “blue” laccases and exhibitsignals related to both copper centers (T1 and T2). Table 4summarizes all spectral parameters measured for the T1, T2,and T3 sites of laccases.
Secondary structures of laccases were investigated bycircular dichroism (CD) spectroscopy. CD spectra are pre-sented in Fig. 5. In the 190- to 250-nm wavelength range, thespectra were mathematically analyzed using the in-housedeveloped computer program “Belok”. It was shown that thelaccases contain about 10% a-helix, 30% b-sheet, 20% turns,along with 40% random coil. In the visible and near-infraredwavelength range, the CD spectra were similar to those ofTrametes versicolor, Neurospora crassa, and Rhus vernic-ifera laccases [44–46].
The potential of the T1 copper was measured by directredox titration using mediators. As an example, the redoxtitration of T. ochracea laccase is shown in Fig. 6. Thereduction of the T1 copper was accompanied by the disap-
Table 3Kinetic parameters of oxidation reactions
Substrate T. hirsuta T. ochracea Cerrena maxima Coriolopsis fulvocinereakcat (s–1) KM (µM) kcat (s–1) KM (µM) kcat (s–1) KM (µM) kcat (s–1) KM (µM)
Catechol 390 142 80 110 320 120 90 85
Guaiacol 430 63 90 90 300 160 95 70
Hydroquinone 450 61 110 74 290 100 110 68
Sinapinic acid 580 24 170 11 330 24 140 21
K4[Fe(CN)6] 400 180 150 96 450 115 130 170
Note: The S.E.M. was calculated from five independent experiments and did not exceed 10%.
697S.V. Shleev et al. / Biochimie 86 (2004) 693–703

pearance of the blue absorbance band at 610 nm. The redoxpotentials of the laccases were in the range from 750 to800 mV vs. NHE. The slope of the Nernst dependence(Fig. 6, inset) during the course of laccases redox titrations inlog[(A/(A0 – A)]/E coordinates varies from 32 mV for
T. ochracea laccase to 57 mV for Coriolopsis fulvocinerealaccase (Table 5).
4. Discussion
The well known Bavendamm reaction shows, in general,the ability of basidiomycetes to produce extracellular oxi-dases such as laccase and tyrosinase [36]. During the oxida-tion of gallic acid, zones with black color are formed. Thediameters of the color zones reflect the amount of oxidaseswhich are produced by fungi during their cultivation. How-ever, in our studies there was no direct positive correlationbetween Bavendamm’s reaction and the dynamics of oxidaseactivity during submerged cultivation of basidiomycetes. Inparticular, the most productive strains (those with the highestoxidase activities) showed the lowest levels of Bavendamm’sreaction. Our further investigations showed that the media aswell as conditions of cultivation strongly affect the oxidaseactivity of the strains. The differences are probably con-nected with the totally different media and conditions ofgrowth (submerged vs. agar plate medium cultivation proce-dures). Further investigations in this direction are necessary.
The other properties of these laccases, such as molecularweight, activity, and carbohydrate composition, differ to agreater or lesser extent from the properties previously re-ported for laccases from the same fungi (see Table 1 and alsoRefs. [8,9,14,19,26,47,48]). The importance of carrying outthe survey work reported here has been mentioned in Section1. As an example, we would like to stress that different forms
Fig. 2. Absorbance spectra of extracellular fungal laccases from (a)T. ochracea, (b) T. hirsuta, (c) Coriolopsis fulvocinerea and (d) Cerrenamaxima. The concentrations of laccases were about 1 mg/ml in 0.1 Mphosphate buffer, PH 6.0.
Fig. 3. Fluorescence spectra of laccases from (a) T. ochracea, (b) T. hirsuta,(c) Coriolopsis fulvocinerea and (d) Cerrena maxima in 0.1 M phosphatebuffer, PH 6.0. (I) Emission spectra, excitation wavelength 330 nm; (II)excitation spectra, emission wavelength 420 nm. The concentrations oflaccases were about 1 mg/ml.
Fig. 4. EPR-spectra of (a) T. ochracea, (b) T. hirsuta, (c) Coriolopsisfulvocinerea and (d) Cerrena maxima laccases in 0.1 M phosphate buffer PH6.0. X-band, microwave power 20 mW. Modulation amplitude 0.5 mT;amplification 1 × 104 (a), 5 × 104 (b), 1 × 104 (c), 1 × 105 (d); fouraccumulations for (a), (b), (c), and eight accumulations for (d). Temperature77°K. The concentrations of laccases were about 1 mg/ml.
698 S.V. Shleev et al. / Biochimie 86 (2004) 693–703
of T. hirsuta laccase have differences in MW in the rangefrom 55 to 70 kDa and in activity from 10 to 400 Units[8,9,14,19]. Knowledge about the actual properties of theparticular purified enzymes is required before using them infurther investigations. We can also emphasize an interestingobservation that some biochemical properties of T. hirsutalaccase (for example, MW), which are described in thiswork, are close to the inducible form of the enzymes (in spiteof the absence of the inducers during the cultivation proce-dure), while other characteristics (pI, pH-optimum, carbohy-drate composition) have similarity with the constitutive formof the laccases from T. hirsuta [8,9]. Unfortunately, it is notpossible to find these dependences for other laccases due tolack of information about the biochemical properties of theirconstitutive and inducible forms. However, in general (with-out consideration of the cultivation) the biochemical charac-teristics of T. ochracea, Cerrena maxima, and Coriolopsisfulvocinerea laccases are very different from those previ-ously described [8,49].
All spectral values of laccases from T. hirsuta, T. ochra-cea, Cerrena maxima, and Coriolopsis fulvocinerea are in agood agreement with the published values for enzymes from
different sources [2,7–9,13,46] except for one EPR param-eter (namely A� � =210.10–4 Cm–1 for the T2 center) of Cori-olopsis fulvocinerea laccase (Table 4). The explanation ofthis unusual parameter is the absence of a low-field shoulderfor Coriolopsis fulvocinerea laccase, which made it difficultto calculate the correct value for the A� � parameter. As for thesecondary structure, it is typical for “blue” laccases[13,44,45] and does not differ from the studied laccases.Furthermore, the enzymes have similar b-barrel type archi-tecture, related to the small blue copper proteins such asazurin or plastocyanin [30]. Thus, significant differenceswhich could be correlated with possible differences betweenthe structures of the active sites of the enzymes have not beenfound through the methodology employed in the presentstudy.
In a very recent publication a new classification schemefor laccases has been proposed [50]. From an electrochemi-cal point of view as well as from analysis of the primarystructures of the enzymes, all laccases can be divided intothree groups in view of the potential of the T1 site: low,
Table 4Spectral characteristics of T1, T2, and T3 sites of laccases (UV-VIS, EPR, and fluorescence analyses)
Laccase T1 site T2 site T3 siteAbM (nm) g� � A� � (10–4 cm–1) g� � A� � (10–4 cm–1) EmM (nm) ExM (nm)
T. ochracea 608 2.20 94 2.24 194 422 315T. hirsuta 607 2.19 95 2.26 186 418 344Cerrena maxima 607 2.18 94 2.24 193 425 322Coriolopsis fulvocinerea 607 2.18 94 2.21–2.23
(2.23)a190–210 (195)a 418 303
Notes: AbM, absorption maximum; EmM, emission maximum; ExM, excitation maximum.a indicates the most probable value for Coroilopsis fulvocinerea laccase.
Fig. 5. Circular dichroism spectra of laccases from (a) T. ochracea, (b)T. hirsuta, (c) Coriolopsis fulvocinerea and (d) Cerrena maxima in 0.1 Mphosphate buffer, Ph 6.0. All samples have concentration 0.1 mg/ml exceptT. hirsuta laccase (0.2 mg/ml).
Fig. 6. Determination of the T1 copper redox potential of T. ochracealaccase in 0.1 M phosphate buffer, PH 6.0. N, native enzyme spectrum; 1–4,mixing the enzyme with K3Mo(CN)8 and K4Mo(CN)8 in various ratios ((1)100:1, (2) 50:1, (3) 20:1, (4) 10:1).
699S.V. Shleev et al. / Biochimie 86 (2004) 693–703

middle and high potential laccases. The low potential groupincludes enzymes from trees, e.g. R. vernicifera laccase witha T1 site potential of about 430 mV vs. NHE [10]. The middlegroup includes laccases from basidiomycetes like Myce-liophthora thermophila [11], basidiomycete C30 [15],Rhizoctonia solany [11], and Coprinus cinereus [13]. Theseenzymes have T1 site potentials ranging from 470 to 710 mVvs. NHE. All the high potential laccases (e.g. those fromT. hirsuta, T. versicolor, and Trametes villosa) have T1 sitepotentials of about 780 mV vs. NHE [10,11]. It was foundthat the redox potentials of examined laccases were in therange from 750 to 800 mV vs. NHE; thus, the isolatedenzymes were seen to belong to a group of middle and highredox potential laccases (Table 5).
It was previously suggested that the value of the redoxpotentials of the copper-containing oxidases depends on theligands of the T1 copper and on the amino acids which formthe T1 pocket [19,51]. Evidently, as can be seen from Table 6,all high redox potential laccases have absolutely similaramino acid sequences in the section of polypeptide chaincontaining the main ligands to the T1 copper and forming theT1 pocket. However, site-directed mutagenesis studies haveshown only minor correlation between T1 Cu-ligating resi-dues and E0 of the T1 site [12,27,51]. Based on the recentlyacquired structural data of the native high potential laccase(T. versicolor), a new mechanism was suggested by whichlaccase, and possibly other redox metalloenzymes, can in-
crease their redox potentials [30]. Such a mechanism as-sumes a reduction of electron density contribution at themetal and the ligating amino acid. In the case of the highpotential laccase the distance between Cu and one of thehistidines is longer compared with the distance between thesame moieties in the middle potential group of the enzyme,for example Coprinus cinereus laccase [30,52]. The hydro-gen bond between His-479 and Ser-113 (114 in the case ofT. villosa laccase) seems to be responsible for this (Table 6,Fig. 7; see detailed explanation in [30]).
It is obvious that the great differences between redoxpotentials for laccases from different sources are not yet fullyunderstood, and further investigations in this direction areneeded. However, our data show that the T1 site of T. hirsutalaccase has a redox potential of about 780 mV vs. NHE,which is reasonably justified by comparison to the sequencesof other high potential laccases (Table 6). We can also sug-gest that knowledge about the redox potential of the T1 sitecan give an idea about the structure of the T1 site of laccases.Thus, analyzing the redox potentials of the T1 sites for thestudied laccases, we can here speculate that the amino acidcomposition of polypeptide chain constituting the mainligands to the T1 coppers and forming the T1 pocket forT. ochracea and Coriolopsis fulvocinerea laccases are veryclose to that of the T. hirsuta and T. versicolor enzymes,while the Cerrena maxima laccase has a structure of theT1 site close to that of the enzyme from Marasmius querco-
Table 5Some electrochemical characteristics of the T1 site of laccases
Laccase E0′, T1 (mV) b (mV) Ratio of concentrations (mediator/laccase)T. ochracea 790 ± 10 32 ± 2 50:1T. hirsuta 780 ± 10 49 ± 2 100:1Cerrena maxima 750 ± 5 56 ± 1 120:1Coriolopsis fulvocinerea 780 ± 10 57 ± 2 100:1
Note: All potentials are given vs. NHE.
Table 6Comparison of the redox potentials, E0′, and the amino acid sequence alignment of the T1 site for oxidases from different sources
Oxidase Sequence aliment E0′, T1 (mV)Trametes hirsuta G H S F L... H... H C H I D F H L E A G F 780a
[19]Trametes versicolor G H S F L... H ... H C H I D F H L E A G F 780
GenBank Accession number A35883 [10]Trametes villosa (pinsutus) G N S F L... H ... H C H I D F H L E A G F 780
GenBank Accession number JC5356 [11]Marasmius quercophilus C30 (I) G H A F L... H ... H C H I D F H L E A G F 730
GenBank Accession number AAM10738 [15]Rhizoctonia solani G P A F V... H ... H C H I D F H L E A G L 710
GenBank Accession number S68120 [11]Coprinus cinereus G H A F L... H ... H C H I E F H L M N G L 550
GenBank Accession number AAD30964 [13]Rhus vernicifera G T N F I... H ... H C H F E R H T T E G M 430
GenBank Accession number BAB63411 [10]Zucchini ascorbate oxidase G E T F F... H ... H C H I E P H L H M G M 340
GenBank Accession number A51027 [12]
Notes: Underlined residues ligands to T1 copper; italicized—amino acids which might affect the electron density contribution at the metal and the ligating aminoacid (in [30]);
a —this paper.
700 S.V. Shleev et al. / Biochimie 86 (2004) 693–703

philus basidiomycete (see the GenBank website:http://www.ncbi.nlm.nih.gov and also Ref. [10,15]).
Comparing the properties of several laccases from differ-ent sources, also including multiple forms of laccases fromthe same species, it can be concluded that T. hirsuta andCerrena maxima laccases have some superior characteristicssuch as high stability, high activity, and low carbohydratecontent, making these laccases attractive objects for furtherinvestigations as well as applications. Moreover, we believethat detailed comparative studies of laccases from differentsources can open up new opportunities for their use in differ-ent aspects of biotechnology as well as reveal the true mecha-nism of their function. Such studies (e.g., electrochemicalinvestigation of the enzymes, creation of biosensors, studiesof laccase mediator systems, achievement of laccase-basedgreen organic synthesis) building on the survey results pre-sented in this paper are being carried out in our laboratories[14,24,50].
Acknowledgements
The authors thank Drs. O.V. Koroleva and V.P. Gavrilovafor the preparation of Coriolopsis fulvocinerea and Cerrenamaxima laccases. The authors also thank Dr. V.V. Shubin forthe computer program for CD spectra calculation, and Dr.Curt T. Reimann for critical reading and helpful suggestions.
The work was supported by INCO-Copernicus Grant(contract No.ICA2-CT-2000-10050), the Russian Founda-tion for Basic Research (project No. 02-04-48885 and No.03-04-48937), and by the FNTP of the Russian Ministry ofScience and Technology (project No.43.073.1.1.2505).
References
[1] A.I. Yaropolov, O.V. Skorobogat’ko, S.S. Vartanov, S.D. Varfolom-eyev, Laccase. Properties, catalytic mechanism, and applicability,Appl. Biochem. Biotechnol. 49 (1994) 257–280.
[2] E.I. Solomon, U.M. Sundaram, T.E. Machonkin, Multicopper oxi-dases and oxygenases, Chem. Rev. 96 (1996) 2563–2605.
[3] H.P. Call, I. Mucke, History, overview and applications of mediatedlignolytic systems, especially laccase-mediator-systems (Lignozym-process), J. Biotechnol. 53 (1997) 163–202.
[4] A. Givaudan, A. Effosse, D. Faure, P. Potier, M.L. Bouillant, R. Bally,Polyphenol oxidase in Azospirillum lipoferum isolated from ricerhizosphere: evidence for laccase activity in nonmotile strains ofAzospirillum lipoferum, FEMS Microbiol. Lett. 108 (1993) 205–210.
[5] O. Martins Ligia, M. Soares Claudio, M. Pereira Manuela, M. Teix-eira, T. Costa, H. Jones George, O. Henriques Adriano, Molecular andbiochemical characterization of a highly stable bacterial laccase thatoccurs as a structural component of the Bacillus subtilis endosporecoat, J. Biol. Chem. 277 (2002) 18849–18859.
[6] F.M. Barrett, in: K. Binnigton, A. Retnakaram (Eds.), Physiology ofthe Insect Epidermis CISRO1991, pp. 194–196, East Melbourne,Australia.
Fig. 7. Schematic drawing illustrating (A) the distance between significant amino acids for redox potentials of laccases, and (B) the movement of a helicalsegment in the enzymes (according to [30]).
701S.V. Shleev et al. / Biochimie 86 (2004) 693–703

[7] A.A. Leontievsky, T. Vares, P. Lankinen, J.K. Shergill, N.N. Pozdnya-kova, N.M. Myasoedova et al., Blue and yellow laccases of ligni-nolytic fungi, FEMS Microbiol. Lett. 156 (1997) 9–14.
[8] O.V. Koroljova-Skorobogat’ko, E.V. Stepanova, V.P. Gavrilova,O.V. Morozova, N.V. Lubimova, A.N. Dzchafarova et al., Purificationand characterization of the constitutive form of laccase from thebasidiomycete Coriolus hirsutus and effect of inducers on laccasesynthesis, Biotechnol. Appl. Biochem. 28 (1998) 47–54.
[9] K.-S. Shin, Y.-J. Lee, Purification and characterization of a newmember of the laccase family from the white-rot basidiomyceteCoriolus hirsutus, Arch. Biochem. Biophys. 384 (2000) 109–115.
[10] B.R.M. Reinhammar, Oxidation–reduction potentials of the electronacceptors in laccases and stellacyanin, Biochim. Biophys. Acta 275(1972) 245–259.
[11] F. Xu, W. Shin, S.H. Brown, J.A. Wahleithner, U.M. Sundaram,E.I. Solomon, A study of a series of recombinant fungal laccases andbilirubin oxidase that exhibit significant differences in redox poten-tial, substrate specificity, and stability, Biochim. Biophys. Acta 1292(1996) 303–311.
[12] F. Xu, A.E. Palmer, D.S. Yaver, R.M. Berka, G.A. Gambetta,S.H. Brown et al., Targeted mutations in a Trametes villosa laccase,axial perturbations of the T1 copper, J. Biol. Chem. 274 (1999)12372–12375.
[13] P. Schneider, M.B. Caspersen, K. Mondorf, T. Halkier, L.K. Skov,P.R. Ostergaard et al., Characterization of a Coprinus cinereus lac-case, Enz. Microb. Technol. 25 (1999) 502–508.
[14] S.V. Shlev, E.A. Zaitseva, E.S. Gorshina, O.V. Morozova,V.A. Serezhenkov, D.S. Burbaev, B.A. Kuznetsov, A.I. Yaropolov,Spectral and electrochemical study of laccases from basidiomycetes,Moscow University Chem. Bull. 44 (2003) 35–39.
[15] A. Klonowska, C. Gaudin, A. Fournel, M. Asso, J. Le Petit, M. Giorgi,T. Tron, Characterization of a low redox potential laccase from thebasidiomycete C30, Eur. J. Biochem. 269 (2002) 6119–6125.
[16] F. Xu, J.J. Kulys, K. Duke, K. Li, K. Krikstopaitis, H.J. Deussen et al.,Redox chemistry in laccase-catalyzed oxidation of N-hydroxy com-pounds, Appl. Environ. Microbiol. 66 (2000) 2052–2056.
[17] M. Balakshin, C.-L. Chen, J.S. Gratzl, A.G. Kirkman, H. Jakob,Biobleaching of pulp with dioxygen in laccase-mediatorsystem—effect of variables on the reaction kinetics, J. Mol. Catal. B:Enz. 16 (2001) 205–215.
[18] A.M. Mayer, R.C. Staples, Laccase: new functions for an old enzyme,Phytochemistry 60 (2002) 551–565.
[19] Y. Kojima, Y. Tsukuda, Y. Kawai, A. Tsukamoto, J. Sugiura,M. Sakaino et al., Cloning, sequence analysis, and expression ofligninolytic phenoloxidase genes of the white-rot basidiomyceteCoriolus hirsutus, J. Biol. Chem. 265 (1990) 15224–15230.
[20] H. Yoshida, Chemistry of lacquer (urushi), J. Chem. Soc. 43 (1883)472–486.
[21] S.C. Barton, H.-H. Kim, G. Binyamin, Y. Zhang, A. Heller, The“Wired” laccase cathode: high current density electroreduction of O2
to water at +0.7 V (NHE) at pH 5, J. Am. Chem. Soc. 123 (2001)5802–5803.
[22] R.S. Freire, N. Duran, L.T. Kubota, Effects of fungal laccase immo-bilization procedures for the development of a biosensor for phenolcompounds, Talanta 54 (2001) 681–686.
[23] B.A. Kuznetsov, G.P. Shumakovich, O.V. Koroleva, A.I. Yaropolov,On applicability of laccase as label in the mediated and mediatorlesselectroimmunoassay: effect of distance on the direct electron transferbetween laccase and electrode, Biosens. Bioelectron. 16 (2001)73–84.
[24] A.V. Karamyshev, S.V. Shleev, O.V. Koroleva, A.I. Yaropolov,I.Y. Sakharov, Laccase-catalyzed synthesis of conducting polyaniline,Enz. Microb. Technol. 33 (2003) 556–564.
[25] C. Johansen, PCT Int. Appl, Novo Nordisk A/s, Den, 1996, pp. 52,Wo.
[26] O.V. Koroleva, I.S. Yavmetdinov, S.V. Shleev, E.V. Stepanova,V.P. Gavrilova, Isolation and study of some properties of laccase fromthe basidiomycetes Cerrena maxima, Biochemistry (Moscow) 66(2001) 618–622.
[27] A.E. Palmer, R.K. Szilagyi, J.R. Cherry, A. Jones, F. Xu,E.I. Solomon, Spectroscopic characterization of the Leu513His vari-ant of fungal laccase: effect of increased axial ligand interaction on thegeometric and electronic structure of the type 1 Cu site, Inorg. Chem.42 (2003) 4006–4017.
[28] M. Antorini, I. Herpoel-Gimbert, T. Choinowski, J.-C. Sigoillot,M. Asther, K. Winterhalter, K. Piontek, Purification, crystallizationand X-ray diffraction study of fully functional laccases from twoligninolytic fungi, Biochim. Biophys. Acta 1594 (2001) 109–114.
[29] N. Hakulinen, L.-L. Kiiskinen, K. Kruus, M. Saloheimo, A. Paananen,A. Koivula, J. Rouvinen, Crystal structure of a laccase from Melano-carpus albomyces with an intact trinuclear copper site, Nat. Struct.Biol. 9 (2002) 601–605.
[30] K. Piontek, M. Antorini, T. Choinowski, Crystal structure of a laccasefrom the fungus Trametes versicolor at 1.90 Å resolution containing afull complement of coppers, J. Biol. Chem. 277 (2002) 37663–37669.
[31] F.J. Enguita, L.O. Martins, A.O. Henriques, M.A. Carrondo, Crystalstructure of a bacterial endospore coat component: A laccase withenhanced thermostability properties, J. Biol. Chem. 278 (2003)19416–19425.
[32] S.-K. Lee, S. DeBeer George, W.E. Antholine, B. Hedman,K.O. Hodgson, E.I. Solomon, Nature of the intermediate formed in thereduction of O2 to H2O at the trinuclear copper cluster active site innative laccase, J. Am. Chem. Soc. 124 (2002) 6180–6193.
[33] A.E. Palmer, L. Quintanar, S. Severance, T.-P. Wang, D.J. Kosman,E.I. Solomon, Spectroscopic characterization and O2 reactivity of thetrinuclear Cu cluster of mutants of the multicopper oxidase Fet3p,Biochemistry 41 (2002) 6438–6448.
[34] D.L. Johnson, J.L. Thompson, S.M. Brinkmann, K.A. Schuller,L.L. Martin, Electrochemical characterization of purified Rhus ver-nicifera laccase: voltammetric evidence for a sequential four-electrontransfer, Biochemistry 42 (2003) 10229–10237.
[35] C. Gercog, K. Gustav, I. Shtrele, Manual on Inorganic Synthesis, Mir,Moscow, 1985, pp. 386, (in Russian).
[36] R. Roesch, Detection of phenoloxidases by Bavendamm’s reactionusing the ring dish test, Zentralblatt fuer Bakteriologie, Parasiten-kunde, Infektionskrankheiten und Hygiene 127 (1972) 555–563.
[37] M.P. Tombs, D.P. Akroy, Acrilamide gel electrophoresis, ShandonInstr. Appl. 18 (1967) 1–16.
[38] O. Vesterberg, H. Svensson, Isoelectric fractionation, analysis, andcharacterization of ampholytes in natural pH gradients. IV. Furtherstudies on the resolving power in connection with separation ofmyoglobins, Acta Chem. Scand. A 20 (1966) 820–834.
[39] G. Felsenfeld, The determination of cuprous ion in Cu proteins, Arch.Biochem. Biophys. 87 (1960) 247–251.
[40] B. Ehresmann, P. Imbault, J.H. Weil, Spectrophotometric determina-tion of protein concentration in cell extracts containing tRNA andrRNA, Anal. Biochem. 54 (1973) 454–463.
[41] P.L. Dutton, Redox potentiometry: determination of midpoint poten-tials of oxidation–reduction components of biological electron-transfer systems, Methods Enzymol. 54 (1978) 411–435.
[42] S.V. Shleev, S.V. Kuznetsov, A.F. Topunov, A cell for bioelectro-chemical studies with parallel spectrophotometric monitoring of thestudied substance, Appl. Biochem. Microbiol. (Moscow) 36 (2000)354–358.
[43] Z. Galus, Theoretical Bases of the Electrochemical Analysis, Mir,Moscow, 1974, pp. 189, (in Russian).
[44] S.-P.W. Tang, J.E. Coleman, Y.P. Myer, Conformational studies ofcopper proteins. Pseudomonas blue protein and Polyporus laccase, J.Biol. Chem. 243 (1968) 4286–4297.
[45] K.E. Falk, B. Reinhammar, Visible and near-infrared circular dichro-ism of some blue copper proteins, Biochim. Biophys. Acta 285 (1972)84–90.
702 S.V. Shleev et al. / Biochimie 86 (2004) 693–703

[46] A. Messerschmidt, Multi-Copper Oxidases1997, pp. 465, Singapore.[47] A.L. Gindilis,Y.A. Baranov, E.O. Zhazhina, V.P. Gavrilova, V.V. Verz-
ilov, A.I. Yaropolov, Laccase from the basidial fungus Cerrenamaxima: some properties and kinetic mechanism of action, Biochem-istry (Moscow) 55 (1990) 315–322.
[48] A.L. Gindilis, E.O. Zhazhina, Y.A. Baranov, A.A. Karyakin,V.P. Gavrilova, A.I. Yaropolov, Isolation and properties of laccasefrom the basidial fungus Coriolus hirsutus (Fr.), Quel, Biochemistry(Moscow) 53 (1988) 735–739.
[49] S.A. Smirnov, O.V. Koroleva, V.P. Gavrilova, A.B. Belova, N.L. Kly-achko, Laccases from basidiomycetes: physicochemical characteris-tics and substrate specificity towards methoxyphenolic compounds,Biochemistry (Moscow) 66 (2001) 774–779.
[50] A. Christensson, N. Dimcheva, E. Ferapontova, L. Gorton, T. Ruzgas,L. Stoica, S. Shleev, Y.A.D. Haltrich, R. Thorneley, S. Aust, Directelectron transfer between ligninolytic redox enzymes and electrodes,Electroanalysis 6 (2004) 1074–1092.
[51] F. Xu, R.M. Berka, J.A. Wahleithner, B.A. Nelson, J.R. Shuster,S.H. Brown et al., Site-directed mutations in fungal laccase: effect onredox potential, activity and pH profile, Biochem. J. 334 (1998)63–70.
[52] V. Ducros,A.M. Brzozowski, K.S. Wilson, S.H. Brown, P. Ostergaard,P. Schneider et al., Crystal structure of the type-2 Cu depleted laccasefrom Coprinus cinereus at 2.2 Å resolution, Nat. Struct. Biol. 5 (1998)310–316.
703S.V. Shleev et al. / Biochimie 86 (2004) 693–703




















