
Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa
29
1037 Mycologia, 95(6), 2003, pp. 1037–1065. q 2003 by The Mycological Society of America, Lawrence, KS 66044-8897 Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa Ellen Larsson 1 Karl-Henrik Larsson Botanical Institute, Go ¨teborg University, P.O. Box 461, SE 405 30 Go ¨teborg, Sweden Abstract: Many homobasidiomycetes are character- ized by a combination of gloeocystidia and amyloid basidiospores. They display a great variation in basi- dioma morphology, including erect and effused forms and gilled and nongilled forms. Earlier studies have shown these taxa to be related, and the group has been named the russuloid clade. Phylogenetic re- lationships among russuloid basidiomycetes were in- vestigated using sequence data from the nuclear 5.8S, ITS2 and large-subunit rDNA genes. A dataset in- cluding 127 ingroup sequences representing 43 gen- era and ca 120 species were analyzed by maximum- parsimony and neighbor-joining methods. The sam- pling of taxa had an emphasis on nongilled taxa and two-thirds of the species possessed corticioid basi- diomata. Thirteen major well-supported clades were identified within the russuloid clade. All clades ex- cept one include corticioid species. Ten characters from basidioma morphology and cultured mycelium were observed and evaluated. Results suggest that gloeocystidia are a synapomorphy for taxa within the russuloid clade while the amyloidity of spores is in- consistent. The ornamentation of spores and type of nuclear behavior seems to be informative characters at genus level. The agaricoid genera Lactarius and Russula are nested in a clade with corticioid species at the basal position. The new combinations Boidinia aculeata, Gloeodontia subasperispora, Gloeocystidiopsis cryptacantha and Megalocystidium wakullum are pro- posed. Key words: amyloid spores, corticioid basidiom- ata, gloeocystidia, Homobasidiomycetes, nuclear rDNA, phylogeny, russuloid clade, sulfobenzaldehyde reaction INTRODUCTION The classification of macrofungi traditionally has re- lied almost entirely on macro- and microanatomical Accepted for publication April 2, 2003. 1 Corresponding author. E-mail: [email protected] features of the basidiome (e.g., Fries 1874, Patouil- lard 1900, Ju ¨lich 1981). However, anatomical simplic- ity, a scanty fossil record and high phenotypic plastic- ity make it difficult to separate the traces of ancestry from instances of parallel evolution. As a result, many fungal genera and families that we are familiar with now are shown to be highly artificial. A few examples are Coprinus, the ink-cap genus, (Redhead et al 2001), the coral fungi in Clavariaceae (Pine et al 1999) and Corticiaceae, the family partly at focus in this study (Hibbett and Thorn 2001). Along with morphological traits, staining reactions also have been widely applied in fungal systematics. Perhaps the best-known example is Melzer’s solution, which is used to detect some of the variable polysac- charide components of fungal cell walls. The active substance in Melzer’s is iodine. Some polysaccharides react with iodine to produce a bluish-violet (amyloid) coloration, while others turn reddish-brown (dextri- noid). Another chemical test involves sulfuric benz- aldehyde, usually in the form of sulfovanilline. When applied to fungal tissue, a dark violet coloration oc- curs in cells containing certain sesquiterpenes (Glu- choff-Fiasson and Ku ¨hner 1982). The reliability of these tests is subject to some uncertainty because col- or development is influenced by tissue age and con- dition and also by age and composition of the re- agents. Despite these uncertainties, both tests have been used extensively in fungal taxonomy for delim- iting species, genera and families (Boidin 1958, Le- mke 1964, Eriksson and Ryvarden 1975). A number of basidiomycetes combine an amyloid reaction of the basidiospore wall with a sulfo-positive reaction (SA1) of thin-walled, tubular or bladder-like cystidia, so-called gloeocystidia. These examples illus- trate the variety of fungal forms having such prop- erties: Russula Pers. and Lactarius Pers., known as important ectomycorrhizal partners in forest ecosys- tems (Gardes and Bruns 1996, Smith and Read 1997) and also widely recognized and collected for con- sumption; coral fungi in the genus Hericium Pers. growing on stumps and living hardwood trees and highly treasured as medical mushrooms in Asia (Ka- wagishi et al 1993, 1996); the aggressive parasite Het- erobasidion annosum, a polypore causing great eco- nomic losses to the forest industry (Stenlid 1986) and Echinodontium tinctorium, the hydnoid Indian paint
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa

1037
Mycologia, 95(6), 2003, pp. 1037–1065.q 2003 by The Mycological Society of America, Lawrence, KS 66044-8897
Phylogenetic relationships of russuloid basidiomycetes with emphasis onaphyllophoralean taxa
Ellen Larsson1
Karl-Henrik LarssonBotanical Institute, Goteborg University, P.O. Box 461,SE 405 30 Goteborg, Sweden
Abstract: Many homobasidiomycetes are character-ized by a combination of gloeocystidia and amyloidbasidiospores. They display a great variation in basi-dioma morphology, including erect and effusedforms and gilled and nongilled forms. Earlier studieshave shown these taxa to be related, and the grouphas been named the russuloid clade. Phylogenetic re-lationships among russuloid basidiomycetes were in-vestigated using sequence data from the nuclear 5.8S,ITS2 and large-subunit rDNA genes. A dataset in-cluding 127 ingroup sequences representing 43 gen-era and ca 120 species were analyzed by maximum-parsimony and neighbor-joining methods. The sam-pling of taxa had an emphasis on nongilled taxa andtwo-thirds of the species possessed corticioid basi-diomata. Thirteen major well-supported clades wereidentified within the russuloid clade. All clades ex-cept one include corticioid species. Ten charactersfrom basidioma morphology and cultured myceliumwere observed and evaluated. Results suggest thatgloeocystidia are a synapomorphy for taxa within therussuloid clade while the amyloidity of spores is in-consistent. The ornamentation of spores and type ofnuclear behavior seems to be informative charactersat genus level. The agaricoid genera Lactarius andRussula are nested in a clade with corticioid speciesat the basal position. The new combinations Boidiniaaculeata, Gloeodontia subasperispora, Gloeocystidiopsiscryptacantha and Megalocystidium wakullum are pro-posed.
Key words: amyloid spores, corticioid basidiom-ata, gloeocystidia, Homobasidiomycetes, nuclearrDNA, phylogeny, russuloid clade, sulfobenzaldehydereaction
INTRODUCTION
The classification of macrofungi traditionally has re-lied almost entirely on macro- and microanatomical
Accepted for publication April 2, 2003.1 Corresponding author. E-mail: [email protected]
features of the basidiome (e.g., Fries 1874, Patouil-lard 1900, Julich 1981). However, anatomical simplic-ity, a scanty fossil record and high phenotypic plastic-ity make it difficult to separate the traces of ancestryfrom instances of parallel evolution. As a result, manyfungal genera and families that we are familiar withnow are shown to be highly artificial. A few examplesare Coprinus, the ink-cap genus, (Redhead et al2001), the coral fungi in Clavariaceae (Pine et al1999) and Corticiaceae, the family partly at focus inthis study (Hibbett and Thorn 2001).
Along with morphological traits, staining reactionsalso have been widely applied in fungal systematics.Perhaps the best-known example is Melzer’s solution,which is used to detect some of the variable polysac-charide components of fungal cell walls. The activesubstance in Melzer’s is iodine. Some polysaccharidesreact with iodine to produce a bluish-violet (amyloid)coloration, while others turn reddish-brown (dextri-noid). Another chemical test involves sulfuric benz-aldehyde, usually in the form of sulfovanilline. Whenapplied to fungal tissue, a dark violet coloration oc-curs in cells containing certain sesquiterpenes (Glu-choff-Fiasson and Kuhner 1982). The reliability ofthese tests is subject to some uncertainty because col-or development is influenced by tissue age and con-dition and also by age and composition of the re-agents. Despite these uncertainties, both tests havebeen used extensively in fungal taxonomy for delim-iting species, genera and families (Boidin 1958, Le-mke 1964, Eriksson and Ryvarden 1975).
A number of basidiomycetes combine an amyloidreaction of the basidiospore wall with a sulfo-positivereaction (SA1) of thin-walled, tubular or bladder-likecystidia, so-called gloeocystidia. These examples illus-trate the variety of fungal forms having such prop-erties: Russula Pers. and Lactarius Pers., known asimportant ectomycorrhizal partners in forest ecosys-tems (Gardes and Bruns 1996, Smith and Read 1997)and also widely recognized and collected for con-sumption; coral fungi in the genus Hericium Pers.growing on stumps and living hardwood trees andhighly treasured as medical mushrooms in Asia (Ka-wagishi et al 1993, 1996); the aggressive parasite Het-erobasidion annosum, a polypore causing great eco-nomic losses to the forest industry (Stenlid 1986) andEchinodontium tinctorium, the hydnoid Indian paint

1038 MYCOLOGIA
fungus, also associated with aggressive decay of co-niferous trees (Thomas 1958); several thin inconspic-uous corticoid species of the genera GloeocystidiellumDonk, Boidinia Stalpers & Hjortstam and GloiotheleBres., which mainly live as saprobionts on differentkinds of deadwood (Eriksson and Ryvarden 1975).
Donk (1971) was the first to discuss a possible re-lationship between taxa such as those mentionedabove and other groups possessing a system of gloeo-plerous hyphae (gloeocystidia) and amyloid basidio-spores. His hypothesis was further expanded and de-veloped by Oberwinkler (1977), who also named theentire group the Russulales. Further arguments forrecognizing a unique russuloid lineage among thehomobasidiomycetes have come from recent molec-ular phylogenetic studies (Hibbett and Donoghue1995, Hibbett et al 1997). According to Hibbett andThorn (2001), the clade includes taxa formerlyplaced in the families Auriscalpiaceae Maas Geest.,Bondarzewiaceae Kotl. & Pouzar, ClavicoronaceaeCorner, Corticiaceae Herter sensu lato, Echinodon-tiaceae Donk, Hericiaceae Donk, LachnocladiaceaeDA Reid, Peniophoraceae Lotsy, Polyporaceae Fr. exCorda sensu lato, Russulaceae Lotsy, and StereaceaePilat and is estimated to hold about 1000 describedspecies (Hibbett and Thorn 2001 extrapolating fromHawksworth et al 1995).
Taxa with effused, corticioid basidiomata, a smoothhymenophore, and russuloid staining reactions firstwere placed together in Gloeocystidiellum Donk. Thehistory and scope of the genus was discussed thor-oughly by Donk (1956, 1964). Eriksson and Ryvarden(1975) regarded Gloeocystidiellum as unnatural andsuggested that the taxa known in Northern Europecould be divided into seven groups. All these groupssince have been segregated as genera (Hagstrom1977, Julich 1978, Hallenberg 1980, Hjortstam andStalpers 1982, Hjortstam 1987b, Boidin et al 1997a),and additional genera based on species from otherparts of the world have been added ( Julich 1982, Wu1995, 1996).
To elucidate evolutionary relationships within therussuloid clade, a dataset based on nuclear rDNA se-quence data (5.8S, ITS2 and 26S) was constructed.Taxa were selected with a particular emphasis on cor-ticioid species, but the range covers all major groupsof russuloid taxa except gasteroid and secotioidforms (Hibbett and Thorn 2001). We used this da-taset to (i) identify major clades among the russuloidfungi, (ii) explore how these clades correlate withprevious morphology based classifications and (iii)redefine corticioid genera within the russuloid clade.
MATERIALS AND METHODS
Sampling of taxa. The sampling (TABLE I) was guided byearlier classifications of Gloeocystidiellum in a wide sense and
by the discussions in Donk (1964), Julich (1981), Stalpers(1996) and Hibbett and Thorn (2001). Nomenclature ofthe species included follows Nordic Macromycetes vol 3(Hansen and Knudsen 1997), Hjortstam and Larsson(1995) and Hjortstam (1998), except in those cases wherewe suggest new taxonomic arrangements. We have adoptedthe convention for clade names introduced by Moncalvo etal (2002). Clade names are preceded by a slash, are spelledin lower-case letters and never italicized. We aimed to in-clude representatives from all groups with a combinationof amyloid basidiospores and gloeocystidia but also specieswith only the latter characteristic, providing they stain withsulfovanillin. The amyloid reaction is much more wide-spread and can be found in a number of genera, e.g., inthe corticioid Amyloathelia Hjortstam & Ryvarden, Aphan-obasidium Julich, and Melzericium Julich, in the poroid An-omoporia Pouzar, and in the agaricoid Catathelasma Lovej.,Hydropus (Kuhn) Singer, Melanoleuca Pat., Mycena (Pers. :Fr.) Roussel, and Panellus P. Karst. No comprehensive sam-pling was done from groups where basidiospore amyloidityis not combined with typical gloeocystidia.
A few genera with a gloeoplerous system were not avail-able for sequencing, viz. Acanthofungus Sheng H. Wu et al(three species), Amylofungus Sheng H. Wu (two), Amylos-porus Ryvarden (seven), Amylonotus Ryvarden (four), Dex-trinocystidium Sheng H. Wu (two), Dichantharellus Corner(two), Stecchericium D.A. Reid (13). We also wanted to in-clude Phlebiella paludicola Hjortstam & P. Roberts since itis the only species in Phlebiella with sulfo-positive gloeocys-tidia. The only modern material available is the holotype.The species forms thin basidiomata on dead leaves of marshplants, making it difficult to extract pure DNA. Since suc-cessful sequencing seemed questionable we decided not toput valuable type material at a risk.
All sequences first were aligned with an extensive datasetholding more than 600 taxa sampled from all major groupsof homobasidiomycetes (data not shown). This dataset iscontinuously expanded and used as an in-house tool forsequence quality control, approximate phylogenetic place-ment of new sequences and as a sampling guide. Trees aregenerated with the neighbor-joining method and the Has-egawa-Kishino-Yano 85 (HKY85) substitution model. Puta-tive russuloid taxa that did not cluster with the russuloidclade or showed ambiguous placement were excluded fromthe final dataset.
For 21 species two collections were sequenced to verifyresults but only one sequence was included in the final da-taset. In a few cases, two sequences with the same speciesname are included but then the nucleotide sequences di-verge and they might represent different species or origi-nate from different geographical areas.
Basidioradulum radula and Trichaptum abietinum were se-lected for rooting of trees because several molecular studiessuggest the hymenochaetoid clade as sister group to therussuloid clade (Binder and Hibbett 2001, Hibbett et al1997, 2000, Larsson 2001), although consistently with nosupport or weak support. Seven additional, more distantlyrelated species from Sistotrema Fr. and the heterobasidi-omycete genera Exidia Fr. and Auricularia Bull. were addedto the outgroup.

1039LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
TA
BL
EI.
Spec
ies
nam
es,
colle
ctio
n/c
ultu
rein
form
atio
nan
dG
enB
ank
acce
ssio
nn
umbe
rs
Spec
ies
Cla
deVo
uch
er/F
CU
Gn
rC
oun
try
Her
bari
uma
Gen
Ban
kac
cess
.n
o.b
Aca
nth
ophy
sellu
mliv
idoc
aeru
leu
m(P
.K
arst
.)Pa
rmas
tost
erea
les
NH
1076
4/20
01C
anad
a,B
.C.
GB
AF5
0640
0A
lbat
rellu
sco
nfl
uen
s(F
r.)K
otl.
&Po
uzar
alba
trel
lus
PV10
1-93
Cec
hos
lova
cia
GB
AF5
0639
3A
lbat
rellu
sov
inu
s(S
chae
ff.)
Kot
l.&
Pouz
aral
batr
ellu
sPV
22-8
9C
ech
oslo
vaci
aG
BA
F506
396
Alb
atre
llus
pes-
capr
ae(P
ers.
:Fr.)
Pouz
aral
batr
ellu
sPV
153-
95C
ech
oslo
vaci
aG
BA
F506
394
Alb
atre
llus
subr
ube
scen
s(M
urri
ll)Po
uzar
alba
trel
lus
PV15
4-95
Cec
hos
lova
cia
GB
AF5
0639
5A
leu
robo
trys
botr
yosu
s(B
urt)
Boi
din
ster
eale
sD
AO
M21
1598
Can
ada,
On
t.D
AO
MA
F506
398
Ale
uro
cyst
idie
llum
disc
ifor
me
(DC
:Fr.)
Telle
rıa
aleu
rocy
stid
iellu
mN
H13
003/
2690
Rus
sia
GB
AF5
0640
2A
leu
rocy
stid
iellu
msu
bcru
enta
tum
(Ber
k.&
M.A
.C
ur-
tis)
P.A
.L
emke
aleu
rocy
stid
iellu
mN
H12
874/
2615
Ger
man
yG
BA
F506
403
Ale
uro
disc
us
amor
phu
s(P
ers.
:Fr.)
J.Sc
hro
t.st
erea
les
KH
L42
40Sw
eden
GB
AF5
0639
7A
leu
rodi
scu
sce
russ
atu
s(B
res)
Hoh
n.
&L
itsc
h.
ster
eale
sN
H11
910/
2350
Den
mar
kG
BA
F506
399
Ale
uro
disc
us
dext
rin
oide
ocer
uss
atu
sM
anjo
n,
P.B
lan
co&
G.
Mor
eno
ster
eale
sE
L25
-97
Spai
n,
Ten
erif
eG
BA
F506
401
Am
ylos
tere
um
areo
latu
m(C
hai
llet:
Fr.)
Boi
din
amyl
oste
reac
eae
NH
8041
/108
0R
oman
iaG
BA
F506
405
Am
ylos
tere
um
chai
lleti
i(P
ers.
:Fr.)
Boi
din
amyl
oste
reac
eae
NH
8031
/103
5R
oman
iaG
BA
F506
406
Am
ylos
tere
um
laev
igat
um
(Fr.)
Boi
din
amyl
oste
reac
eae
NH
1286
3/25
90Sw
eden
GB
AF5
0640
7A
rtom
yces
pyxi
datu
s(P
ers.
:Fr.)
Julic
ham
ylos
tere
acea
eR
D81
8/23
02E
ston
iaG
BA
F506
380
Ast
eros
trom
ace
rvic
olor
(Ber
k.&
M.A
.C
urti
s)M
asse
epe
nio
phor
ales
KH
L92
39Pu
erto
Ric
oG
BA
F506
408
Ast
eros
trom
ala
xum
Bre
s.pe
nio
phor
ales
EL
33-9
9E
ston
iaG
BA
F506
410
Ast
eros
trom
am
usc
icol
a(B
erk.
&M
.A.
Cur
tis)
Mas
see
pen
ioph
oral
esK
HL
9573
Puer
toR
ico
GB
AF5
0640
9A
uri
cula
ria
mes
ente
rica
(Dic
ks.e
xS.
F.G
ray)
Pers
.E
L66
-97
USA
,N
.C.
GB
AF5
0649
2A
uri
scal
piu
mvi
llipe
s(L
loyd
)Sn
ell
&E
.A.
Dic
kau
risc
alpi
acea
eL
R42
783
Cos
taR
ica
OA
F506
478
Au
risc
alpi
um
villi
pes
auri
scal
piac
eae
LR
2421
8B
razi
lO
AF5
0647
7A
uri
scal
piu
mvu
lgar
eS.
F.G
ray
auri
scal
piac
eae
EL
33-9
5Sw
eden
GB
AF5
0637
5B
asid
iora
dulu
mra
dula
(Fr.)
Nob
les
NH
9453
Fin
lan
dG
B(A
F347
105)
Boi
din
iafu
rfu
race
a(B
res.
)St
alpe
rs&
Hjo
rtst
amru
ssul
ales
JS16
717
Nor
way
OA
F506
376
Boi
din
iagr
anu
lata
Shen
gH
.W
ugl
oeoc
ysti
diel
lum
IIW
u920
9-34
/264
9T
aiw
anG
B(A
F048
880)
Boi
din
iam
acro
spor
aSh
eng
H.
Wu
ster
eale
sW
u920
2-21
/279
1T
aiw
anN
MN
SA
F506
377
Boi
din
iapr
opin
qua
(H.S
.Ja
cks.
&D
eard
en)
Hjo
rts-
tam
&R
yvar
den
russ
ulal
esK
HL
1093
1Ja
mai
caG
BA
F506
379
Boi
din
iasp
.ru
ssul
ales
KH
L10
303
Puer
toR
ico
GB
AF5
0637
8B
onda
rzew
iabe
rkel
eyi
(Fr.)
Bon
dart
sev
&Si
nge
rbo
nda
rzew
iace
aeSL
M95
07U
SA,
NC
Priv
ate
(AF2
1856
3)C
onfe
rtic
ium
ochr
aceu
m(F
r.)H
alle
nb.
ster
eale
sN
H91
71/1
516
Rom
ania
GB
AF5
0638
3C
onfe
rtic
ium
ravu
m(B
urt)
Gin
ns
&G
.W.
Free
man
ster
eale
sN
H13
291
Est
onia
GB
AF5
0638
2C
onfe
rtob
asid
ium
oliv
aceo
albu
m(B
ourd
ot&
Gal
zin
)Ju
lich
pen
ioph
oral
esFP
9019
6U
SA,
GA
CFM
RA
F511
648
Con
fert
obas
idiu
msp
.pe
nio
phor
ales
TA
A15
9923
Aus
tral
iaT
AA
AF5
0638
4D
enti
pelli
sdi
ssit
a(B
erk.
&C
ooke
)M
aas
Gee
st.
her
icia
ceae
NH
6280
/581
Can
ada,
Que
GB
AF5
0638
6D
enti
pelli
sfr
agili
s(F
r.)D
onk
her
icia
ceae
KH
L65
69/1
755
Swed
enG
BA
F506
387
Den
tipe
llis
sp.
her
icia
ceae
KH
L11
015
Ven
ezue
laG
BA
F506
388

1040 MYCOLOGIAT
AB
LE
I.C
onti
nue
d
Spec
ies
Cla
deVo
uch
er/F
CU
Gn
rC
oun
try
Her
bari
uma
Gen
Ban
kac
cess
.n
o.b
Den
tipr
atu
lum
bial
ovie
sen
seD
oman
ski
auri
scal
piac
eae
GG
1645
Fran
ceG
BA
F506
389
Dic
host
ereu
mdu
rum
(Bou
rd.
&G
alz.
)Pi
lat
pen
ioph
oral
esFu
ngi
Gal
lici
1985
Fran
ceG
BA
F506
429
Dic
host
ereu
mef
fusc
atu
m(C
ooke
&E
llis)
Boi
din
&L
anq.
pen
ioph
oral
esG
G93
0915
Fran
ceG
BA
F506
390
Dic
host
ereu
mgr
anu
losu
m(P
ers.
:Fr.)
Boi
din
&L
anq.
pen
ioph
oral
esN
H71
37/6
96C
anad
a,B
.C.
GB
AF5
0639
1D
icho
ster
eum
palle
scen
s(S
chw
ein
.)B
oidi
n&
Lan
q.pe
nio
phor
ales
NH
7046
/673
Can
ada,
B.C
.G
BA
F506
392
Dic
host
ereu
msp
.pe
nio
phor
ales
KH
L10
258
Puer
toR
ico
GB
AF5
0642
8E
chin
odon
tiu
mry
vard
enii
Ber
nic
chia
&Pi
gaL
R43
370
Ital
yO
AF5
0643
1E
chin
odon
tiu
mti
nct
oriu
m(E
llis
&E
verh
.)E
llis
&E
v-er
h.
bon
darz
ewia
ceae
NH
6695
/500
Can
ada,
B.C
.G
BA
F506
430
Exi
dia
glan
dulo
sa(B
ull.)
Fr.
EL
3-97
Swed
enG
BA
F506
493
Exi
dia
reci
sa(D
itm
.)Fr
.E
L15
-98
Swed
enG
B(A
F347
112)
Glo
eocy
stid
iellu
mac
ule
atu
mSh
eng
H.
Wu
russ
ulal
esW
u890
714-
52/2
647
Tai
wan
GB
AF5
0643
3G
loeo
cyst
idie
llum
aspe
llum
Hjo
rtst
amst
erea
les
LIN
625/
2644
Tai
wan
NM
NS
AF5
0643
2G
loeo
cyst
idie
llum
bisp
oru
mB
oidi
n,
Lan
q.&
Gill
esgl
oeoc
ysti
diel
lum
IK
HL
1113
5N
orw
ayG
B(A
Y048
877)
Glo
eocy
stid
iellu
mcl
avu
liger
um
(Hoh
n.
&L
itsc
h.)
Na-
kaso
ne
gloe
ocys
tidi
ellu
mI
JS16
976
Nor
way
GB
(AF3
1008
4)
Glo
eocy
stid
iellu
mcl
avu
liger
um
gloe
ocys
tidi
ellu
mI
NH
1315
9/27
31R
ussi
aG
B(A
F310
083)
Glo
eocy
stid
iellu
mcl
avu
liger
um
gloe
ocys
tidi
ellu
mI
NH
1118
5/21
59Sp
ain
,Te
ner
ife
GB
(AF3
1008
8)G
loeo
cyst
idie
llum
com
pact
um
Shen
gH
.W
ust
erea
les
Wu8
8061
5-21
/264
8T
aiw
anN
MN
SA
F506
434
Glo
eocy
stid
iellu
mcr
ypta
can
thu
m(P
at.)
Hjo
rtst
amst
erea
les
KH
L10
334
Puer
toR
ico
GB
AF5
0644
2G
loeo
cyst
idie
llum
form
osan
um
Shen
gH
.W
ust
erea
les
Wu9
404-
16/2
651
Tai
wan
NM
NS
AF5
0643
9G
loeo
cyst
idie
llum
poro
sum
(Ber
k.&
M.A
.C
urti
s)D
onk
gloe
ocys
tidi
ellu
mII
NH
1043
4/19
33D
enm
ark
GB
(AF3
1009
4)G
loeo
cyst
idie
llum
purp
ure
um
Shen
gH
.W
ugl
oeoc
ysti
diel
lum
IW
u931
0-45
Tai
wan
GB
(AF4
4133
8)G
loeo
cyst
idie
llum
sp.
gloe
ocys
tidi
ellu
mII
NH
1325
8/27
66T
urke
yG
B(A
F310
089)
Glo
eocy
stid
iellu
msp
.gl
oeoc
ysti
diel
lum
IIN
H12
972/
2679
Rus
sia
GB
(AF3
1009
0)G
loeo
cyst
idie
llum
suba
sper
ispo
rum
(Lit
sch
.)J.
Eri
kss.
&R
yvar
den
gloe
odon
tia
KH
L86
95N
orw
ayG
BA
F506
404
Glo
eocy
stid
iellu
mw
aku
llum
Bur
ds.,
Nak
ason
e&
Free
-m
anst
erea
les
Osl
o-93
0107
Tan
zan
iaO
AF5
0644
3
Glo
eocy
stid
iops
isfl
amm
ea(B
oidi
n)
Julic
hst
erea
les
AH
0002
19L
aR
eun
ion
GB
AF5
0643
8G
loeo
cyst
idio
psis
flam
mea
ster
eale
sC
BS3
24.6
6C
.A
fric
anR
ep.
LYA
F506
437
Glo
eocy
stid
iops
ishe
imii
(Boi
din
)Ju
lich
ster
eale
sC
BS3
21.6
6C
.A
fric
anR
ep.
LYA
F506
381
Glo
eodo
nti
aco
lum
bien
sis
Bur
tex
Bur
ds.
&L
omb.
gloe
odon
tia
NH
1111
8/21
33Sp
ain
,Te
ner
ife
GB
AF5
0644
4G
loeo
don
tia
disc
olor
(Ber
k.&
M.A
.C
urti
s)B
oidi
ngl
oeod
onti
aK
HL
1009
9Pu
erto
Ric
oG
BA
F506
445
Glo
eodo
nti
apy
ram
idat
a(B
erk.
&M
.A.
Cur
tis)
Hjo
rts-
tam
gloe
odon
tia
LR
1550
2C
olom
bia
OA
F506
446
Glo
eohy
poch
nic
ium
anal
ogu
m(B
ourd
ot&
Gal
zin
)H
jort
stam
NH
1214
0R
ussi
aG
BA
F506
447
Glo
eom
yces
gram
inic
ola
Shen
gH
.W
ust
erea
les
Wu9
210-
12/2
650
Tai
wan
GB
AF5
0644
8

1041LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
TA
BL
EI.
Con
tin
ued
Spec
ies
Cla
deVo
uch
er/F
CU
Gn
rC
oun
try
Her
bari
uma
Gen
Ban
kac
cess
.n
o.b
Glo
eope
nio
phor
ella
con
volv
ens
(P.
Kar
st.)
Boi
din
,L
anq.
&G
illes
russ
ulal
esK
HL
1010
3Pu
erto
Ric
oG
BA
F506
435
Glo
eope
nio
phor
ella
laxa
(Sh
eng
H.
Wu)
Boi
din
,L
anq.
&G
illes
russ
ulal
esW
u911
010-
8/26
45T
aiw
anN
MN
SA
F506
440
Glo
eope
nio
phor
ella
sp.
russ
ulal
esK
HL
1039
0Pu
erto
Ric
oG
BA
F506
436
Glo
iodo
nn
igre
scen
s(P
etch
)M
aas
Gee
st.
auri
scal
piac
eae
Des
jard
in72
87B
ali
GB
AF5
0645
0G
loio
don
stri
gosu
s(S
w.:F
r.)P.
Kar
st.
auri
scal
piac
eae
JS26
147
Nor
way
OA
F506
449
Glo
ioth
ele
irpi
sces
cen
sB
oidi
npe
nio
phor
ales
LR
3553
3Ve
nez
uela
OA
F506
452
Glo
ioth
ele
lact
esce
ns
(Ber
k.)
Hjo
rtst
ampe
nio
phor
ales
EL
8-98
Swed
enG
BA
F506
453
Glo
ioth
ele
lam
ello
sa(P
.H
enn
.)B
res.
pen
ioph
oral
esK
HL
1103
1Ve
nez
uela
GB
AF5
0645
4G
loio
thel
esp
.pe
nio
phor
ales
LR
4240
7Ve
nez
uela
GB
AF5
0645
1G
loio
thel
eto
rren
dii
(Bre
s.)
Boi
din
&H
.M
ich
elpe
nio
phor
ales
LY16
815
Fran
ceLY
AF5
0645
5H
eric
ium
abie
tis
(Wei
rex
Hub
ert)
K.
Har
riso
nh
eric
iace
aeN
H69
90/6
63C
anad
aG
BA
F506
456
Her
iciu
mal
pest
rePe
rs.
her
icia
ceae
NH
1324
0/27
54R
ussi
aG
BA
F506
457
Her
iciu
mam
eric
anu
mG
inn
sh
eric
iace
aeD
AO
M21
467
Can
ada,
On
t.D
AO
MA
F506
458
Her
iciu
mci
rrha
tum
(Per
s.:F
r.)N
ikol
ajev
ah
eric
iace
aeT
ubin
gen
F794
Ger
man
yT
UB
AF5
0638
5H
eric
ium
cora
lloid
es(S
copo
li:F
r.)Pe
rs.
her
icia
ceae
NH
282/
1229
Swed
enG
BA
F506
459
Her
iciu
mer
inac
eus
(Bul
l.:F
r.)Pe
rs.
her
icia
ceae
NH
1216
3/24
68R
ussi
aG
BA
F506
460
Het
erob
asid
ion
ann
osu
m(F
r.:F
r.)B
ref.
bon
darz
ewia
ceae
NH
1206
7/24
50R
ussi
aG
B(A
F347
096)
Lac
hnoc
ladi
um
sp.
pen
ioph
oral
esK
HL
1055
6Ja
mai
caG
BA
F506
461
Lac
tari
us
leon
isK
ytov
uori
russ
ulal
esSJ
9101
6Sw
eden
GB
AF5
0641
1L
acta
riu
ssu
ben
bora
tus
Lin
dgre
nru
ssul
ales
EL
10-0
0Sw
eden
GB
AF5
0641
2L
acta
riu
svo
lem
us
Fr.
russ
ulal
esK
HL
08-0
0Sw
eden
GB
AF5
0641
3L
auri
liasu
lcat
a(B
urt)
Pouz
arbo
nda
rzew
iace
aeK
HL
8267
Rus
sia
GB
AF5
0641
4L
axit
extu
mbi
colo
r(P
ers.
:Fr.)
Len
tzh
eric
iace
aeN
H51
66/1
350
Swed
enG
B(A
F310
102)
Len
tin
ellu
sau
ricu
la(F
r.)A
.St
rid
auri
scal
piac
eae
KG
N28
0994
Swed
enG
BA
F506
415
Len
tin
ellu
sca
stor
eus
Fr.
auri
scal
piac
eae
SJ93
067
Swed
enG
BA
F506
416
Len
tin
ellu
sco
chle
atu
s(F
r:F
r)P.
Kar
st.
auri
scal
piac
eae
KG
N96
-09-
28Sw
eden
GB
AF5
0641
7L
enti
nel
lus
omph
alod
es(F
r.)P.
Kar
st.
auri
scal
piac
eae
JJ20
77Sw
eden
GB
AF5
0641
8L
enti
nel
lus
urs
inu
s(F
r.:F
r.)K
uhn
erau
risc
alpi
acea
eE
L73
-97
USA
,N
.C.
GB
AF5
0641
9L
enti
nel
lus
vulp
inu
s(F
r.:F
r.)K
uhn
er&
Mai
reau
risc
alpi
acea
eK
GN
98-0
8-25
Swed
enG
B(A
F347
097)
Meg
aloc
ysti
diu
mch
elid
oniu
m(P
at.)
Boi
din
Lan
q.&
Gil-
les
ster
eale
sL
odge
SJ11
0.1
USA
,V
irgi
nIs
l.G
BA
F506
441
Meg
aloc
ysti
diu
mle
uco
xan
thu
m(B
res.
)Ju
lich
ster
eale
sH
K82
/191
5D
enm
ark
GB
AF5
0642
0M
egal
ocys
tidi
um
luri
dum
(Bre
s.)
Julic
hst
erea
les
JE24
636/
22Sw
eden
GB
AF5
9642
1M
egal
ocys
tidi
um
luri
dum
ster
eale
sK
HL
8635
Nor
way
GB
AF5
0642
2M
etu
lodo
nti
an
ivea
(P.
Kar
st.)
Parm
asto
pen
ioph
oral
esN
H13
108/
2712
Rus
sia
GB
AF5
0642
3P
enio
phor
aci
ner
ea(F
r.)C
ooke
ssp.
fagi
cola
Hal
len
b.&
E.
Lar
ss.
pen
ioph
oral
esN
H98
08/1
788
Spai
nG
BA
F506
424
Pen
ioph
ora
inca
rnat
a(P
ers.
:Fr.)
P.K
arst
.pe
nio
phor
ales
NH
1027
1/19
09D
enm
ark
GB
AF5
0642
5
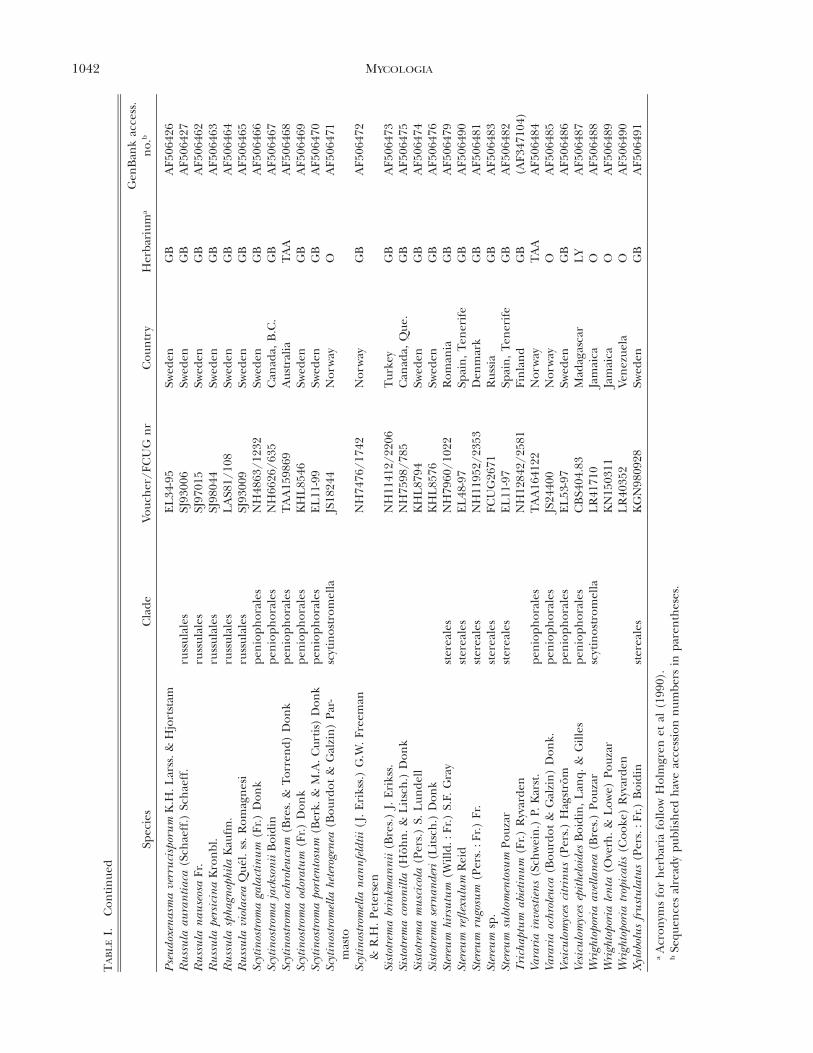
1042 MYCOLOGIA
TA
BL
EI.
Con
tin
ued
Spec
ies
Cla
deVo
uch
er/F
CU
Gn
rC
oun
try
Her
bari
uma
Gen
Ban
kac
cess
.n
o.b
Pse
udo
xen
asm
ave
rru
cisp
oru
mK
.H.L
arss
.&H
jort
stam
EL
34-9
5Sw
eden
GB
AF5
0642
6R
uss
ula
aura
nti
aca
(Sch
aeff
.)Sc
hae
ff.
russ
ulal
esSJ
9300
6Sw
eden
GB
AF5
0642
7R
uss
ula
nau
seos
aFr
.ru
ssul
ales
SJ97
015
Swed
enG
BA
F506
462
Ru
ssu
lape
rsic
ina
Kro
nbl
.ru
ssul
ales
SJ98
044
Swed
enG
BA
F506
463
Ru
ssu
lasp
hagn
ophi
laK
aufm
.ru
ssul
ales
LA
S81/
108
Swed
enG
BA
F506
464
Ru
ssu
lavi
olac
eaQ
uel.
ss.
Rom
agn
esi
russ
ulal
esSJ
9300
9Sw
eden
GB
AF5
0646
5Sc
ytin
ostr
oma
gala
ctin
um
(Fr.)
Don
kpe
nio
phor
ales
NH
4863
/123
2Sw
eden
GB
AF5
0646
6Sc
ytin
ostr
oma
jack
son
iiB
oidi
npe
nio
phor
ales
NH
6626
/635
Can
ada,
B.C
.G
BA
F506
467
Scyt
inos
trom
aoc
hrol
eucu
m(B
res.
&To
rren
d)D
onk
pen
ioph
oral
esT
AA
1598
69A
ustr
alia
TA
AA
F506
468
Scyt
inos
trom
aod
orat
um
(Fr.)
Don
kpe
nio
phor
ales
KH
L85
46Sw
eden
GB
AF5
0646
9Sc
ytin
ostr
oma
port
ento
sum
(Ber
k.&
M.A
.Cur
tis)
Don
kpe
nio
phor
ales
EL
11-9
9Sw
eden
GB
AF5
0647
0Sc
ytin
ostr
omel
lahe
tero
gen
ea(B
ourd
ot&
Gal
zin
)Pa
r-m
asto
scyt
inos
trom
ella
JS18
244
Nor
way
OA
F506
471
Scyt
inos
trom
ella
nan
nfe
ldti
i(J
.E
riks
s.)
G.W
.Fr
eem
an&
R.H
.Pe
ters
enN
H74
76/1
742
Nor
way
GB
AF5
0647
2
Sist
otre
ma
brin
kman
nii
(Bre
s.)
J.E
riks
s.N
H11
412/
2206
Tur
key
GB
AF5
0647
3Si
stot
rem
aco
ron
illa
(Hoh
n.
&L
itsc
h.)
Don
kN
H75
98/7
85C
anad
a,Q
ue.
GB
AF5
0647
5Si
stot
rem
am
usc
icol
a(P
ers.
)S.
Lun
dell
KH
L87
94Sw
eden
GB
AF5
0647
4Si
stot
rem
ase
rnan
deri
(Lit
sch
.)D
onk
KH
L85
76Sw
eden
GB
AF5
0647
6St
ereu
mhi
rsu
tum
(Will
d.:F
r.)S.
F.G
ray
ster
eale
sN
H79
60/1
022
Rom
ania
GB
AF5
0647
9St
ereu
mre
flex
ulu
mR
eid
ster
eale
sE
L48
-97
Spai
n,
Ten
erif
eG
BA
F506
490
Ster
eum
rugo
sum
(Per
s.:F
r.)Fr
.st
erea
les
NH
1195
2/23
53D
enm
ark
GB
AF5
0648
1St
ereu
msp
.st
erea
les
FCU
G26
71R
ussi
aG
BA
F506
483
Ster
eum
subt
omen
tosu
mPo
uzar
ster
eale
sE
L11
-97
Spai
n,
Ten
erif
eG
BA
F506
482
Tri
chap
tum
abie
tin
um
(Fr.)
Ryv
arde
nN
H12
842/
2581
Fin
lan
dG
B(A
F347
104)
Vara
ria
inve
stie
ns
(Sch
wei
n.)
P.K
arst
.pe
nio
phor
ales
TA
A16
4122
Nor
way
TA
AA
F506
484
Vara
ria
ochr
oleu
ca(B
ourd
ot&
Gal
zin
)D
onk.
pen
ioph
oral
esJS
2440
0N
orw
ayO
AF5
0648
5Ve
sicu
lom
yces
citr
inu
s(P
ers.
)H
agst
rom
pen
ioph
oral
esE
L53
-97
Swed
enG
BA
F506
486
Vesi
culo
myc
esep
ithe
loid
esB
oidi
n,
Lan
q.&
Gill
espe
nio
phor
ales
CB
S404
.83
Mad
agas
car
LYA
F506
487
Wri
ghto
pori
aav
ella
nea
(Bre
s.)
Pouz
arsc
ytin
ostr
omel
laL
R41
710
Jam
aica
OA
F506
488
Wri
ghto
pori
ale
nta
(Ove
rh.
&L
owe)
Pouz
arK
N15
0311
Jam
aica
OA
F506
489
Wri
ghto
pori
atr
opic
alis
(Coo
ke)
Ryv
arde
nL
R40
352
Ven
ezue
laO
AF5
0649
0X
ylob
olu
sfr
ust
ula
tus
(Per
s.:F
r.)B
oidi
nst
erea
les
KG
N98
0928
Swed
enG
BA
F506
491
aA
cron
yms
for
her
bari
afo
llow
Hol
mgr
enet
al(1
990)
.b
Sequ
ence
sal
read
ypu
blis
hed
hav
eac
cess
ion
num
bers
inpa
ren
thes
es.

1043LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
As standard mounting media for microscopic examina-tions of specimens, 2% KOH, Melzer’s reagent and sulfov-anillin have been used (Moser 1978).
Molecular techniques. DNA was isolated from herbariumspecimens and from cultured mycelia. Mycelia were grownat room temperature for 2 wk in 50 mL MYG liquid media(1% malt extract, 0.4% yeast extract, 1% glucose). Myceliawere harvested and dried between sheets of filter paper andca 50 mg was placed in microcentrifuge tubes. From her-barium specimens ca 3 3 3 mm of hymenium was used toextract DNA. DNA extractions were carried out using amodified 2% CTAB method (Savolainen et al 1995). Prep-arations from some of the herbarium specimens were fur-ther purified with Gene Clean (Bio 101 Inc.) to excludePCR inhibitors. The internal transcribed spacer 1 and 2(ITS 1 and 2) including the 5.8S region of nuclear rDNA,was amplified with primers ITS1F, ITS4B (Gardes and Bruns1993), ITS1 and ITS4 (White et al 1990). Approximately1200 bp of the 59 end of the large subunit of the rDNA(nuclear LSU) was amplified with LR0R and LR7 (Vilgalysand Hester 1990).
PCR amplifications were performed in 25 mL reactionsusing either Taq polymerase (Advanced Biotechnologies)together with reaction buffer IV or Ready To Goy PCRbeads (Amersham Pharmacia Biotech Inc.). The thermalcycling program included 4 min at 95 C, followed by 30cycles of 30 s at 95 C, 30 s at 52 C, 60 s at 72 C, and thenended by 8 min at 72 C. Fragments were examined on a1% SeaKem (FMC) agarose gel, and amplified productswere purified with QIAquick spin columns (QIAGEN).
Primers used for sequencing of both strands were ITS3,ITS4 (White et al 1990), LR5, LR21, LR3r (http://WWW.biology.duke.edu/fungi.html), and CTB6 (http://mendel.berkeley.edu/boletus.html). Cycle sequencing wascarried out using Thermosequenase flourescent labeledprimer cycle sequencing kit with 7-deaza-dGTP (AmershamPharmacia Biotech Inc.). Twenty-five ng of template DNAand 5 pmol Cy5-labeled primer were used per reaction. Se-quences were obtained using an ALFExpress (PharmaciaBiotech Inc.) automated sequencer. Sequences were editedand assembled using computer software ALF manager(Pharmacia Biotech) and Sequencher 3.1 (Gene CodesCorp.). Complete sequences were aligned manually usingthe data editor PAUP* 4.0 (Swofford 1999). Sequences weresubmitted to GenBank, and accession numbers are given inTABLE I.
The LSU sequence of Bondarzewia berkeleyi in this studywas taken from GenBank (AF218563). The aligned data ma-trices are available from the corresponding author uponrequest.
Phylogenetic analyses. Heuristic searches were performedusing PAUP* 4.0b8 (Swofford 1999) on a Power Macintoshcomputer. All transformations were considered unorderedand equally weighted. Variable regions with ambiguousalignment were excluded, and gaps were treated as missingdata. Heuristic searches with 1000 random-addition se-quence replicates, TBR branch swapping and MAXTREESset to 25 000 with restrictions to save 100 trees in each rep-licate, were performed.
Neighbor-joining analysis (NJ) was performed on thesame dataset using the Hasegawa-Kishino-Yano 85 (HKY85)substitution model.
Ten selected trees, with the best-likelihood score, fromthe initial heuristic searches were used as starting trees inrecurrent heuristic searches, with TBR and NNI branchswapping, under the maximum-likelihood criteria (ML) tosearch for more optimal trees. ML parameter settings cor-responded to HKY851I model, with the nucleotide substi-tution rate parameters estimated via ML. Searches usingTBR swapping were aborted before completion (after 24 h),due to the large dataset and computationally intensive al-gorithms. Trees with the best-likelihood scores were savedfor comparison.
To compare tree topologies of alternative phylogenetichypotheses for the dataset, several constrained analyseswere conducted. Constrained trees forcing monophyly of/peniophorales and keeping it as a sister clade to the restof the russuloid clade were constructed using Mac Clade4.0 (Maddison and Maddison 2000). All nodes within theclades were collapsed. Heuristic searches with 100 random-addition sequence replicate, enforcing constraints and sav-ing only trees compatible with constraint topologies wereperformed using the same taxa and settings as above. Con-strained analyses to test the support for monophyly of theG. porosum-clavuligerum complex were performed in thesame way.
Topological differences between the constrained phylo-genetic tree hypothesis and unconstrained trees were eval-uated with Kishino-Hasegawa (KH) maximum-likelihood ra-tio test (normal approximation, two-tailed test) and the Shi-modaira-Hasegawa (SH) test (using RELL bootstrap) imple-mented in PAUP*.
To improve the resolution within Stereum and allied gen-era, 27 sequences from the large dataset were realigned toinclude more characters from the variable ITS2 region.One sequence of Gloeocystidiellum aspellum was added, andGloeodontia pyramidata was selected as outgroup. In the par-simony analysis, all transformations were considered unor-dered and equally weighted. Variable regions with ambig-uous alignment were excluded, and gaps were treated asmissing data. Heuristic searches with 1000 random-additionsequence replicates, TBR branch swapping and MAXTREESset to auto-increase, were performed.
Relative robustness of clades was estimated by bootstrap-ping, with the following settings. For the large dataset byusing 1000 bootstrap replicates, with five random-additionsequences per replicate, TBR branch swapping and MAX-TREES set to 25 000 with restrictions to save 100 trees ineach replicate. For the Stereum-restricted dataset by using1000 bootstrap replicates, with 100 random-addition se-quences per replicate, TBR branch swapping and MAX-TREES set to auto-increase.
Morphological characters. Morphological features of thebasidiome, hymenophore and spore surface, presence ofclamps and skeletal hyphae, type of life strategy, nuclearbehavior and polarity (if available) have been observed orgathered from literature and compiled in TABLE II. Data onnuclear behavior and polarity are mainly taken from Boidin

1044 MYCOLOGIAT
AB
LE
II.
Sum
mar
yof
som
em
orph
olog
ical
and
cyto
logi
cal
char
acte
rs.
Tax
onor
der
follo
ws
the
stri
ctco
nse
nsu
str
ee(F
ig.
2).
Cur
ren
tly
acce
pted
gen
eric
posi
tion
sar
ein
dica
ted.
Spec
ies
Cla
deG
enus
Lif
eB
asb
Hym
cSp
dA
me
SAf
Skeg
PhN
uci
Cj
Vesi
culo
myc
esci
trin
us
Glo
ioth
ele
lact
esce
ns
Vesi
culo
myc
esep
ithe
loid
esG
loio
thel
eir
pisc
esce
ns
Glo
ioth
ele
lam
ello
saG
loio
thel
eto
rren
dii
Glo
ioth
ele
sp.
Scyt
inos
trom
apo
rten
tosu
mA
ster
ostr
oma
cerv
icol
orA
ster
ostr
oma
mu
scic
ola
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
Vesi
culo
myc
esG
loio
thel
eG
loio
thel
eG
loio
thel
eG
loio
thel
eG
loio
thel
eG
loio
thel
eSc
ytin
ostr
oma
Ast
eros
trom
aA
ster
ostr
oma
S S S S S S S S S S
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
S S S S S S S S S S
S S S S S S S S L L
Y Y Y Y Y Y Y Y Y Y
N Y Y Y Y Y Y Y N/Y
N
N N N N N N N Y Y Y
h h h h h IV h
SN SN SN SN SN SN*
SN*
a a a a a a a a
Ast
eros
trom
ala
xum
Scyt
inos
trom
aoc
hrol
eucu
mVa
rari
aoc
hrol
euca
Lac
hnoc
ladi
um
sp.
Scyt
inos
trom
aod
orat
um
Vara
ria
inve
stie
ns
Pen
ioph
ora
cin
erea
ssp.
fagi
cola
Pen
ioph
ora
inca
rnat
aD
ich
oste
reum
palle
scen
sD
ich
oste
reum
sp.
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
Ast
eros
trom
a? ? ? ? Va
rari
aP
enio
phor
aP
enio
phor
aD
icho
ster
eum
Dic
host
ereu
m
S S S S S S S S S S
Eff
Eff
Eff
Cla
vE
ffE
ffE
ffE
ffE
ffE
ff
S S S S S S S S S S
S S S S S S S S O O
Y N N Y N N/Y
N N Y Y
N Y N N N Y N Y Y Y
Y Y Y Y Y Y N N Y Y
h H h IV IV IV
SN HD
*
N N SN He
a a a a c c c c
Dic
host
ereu
mdu
rum
Dic
host
ereu
mgr
anu
losu
mD
icho
ster
eum
effu
scat
um
Scyt
inos
trom
aja
ckso
nii
Scyt
inos
trom
aga
lact
inu
mM
etu
lodo
nti
an
ivea
Con
fert
obas
idiu
mol
ivac
eoal
bum
Con
fert
obas
idiu
msp
.W
righ
topo
ria
trop
ical
isSc
ytin
ostr
omel
lan
ann
feld
tii
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
pen
ioph
oral
espe
nio
phor
ales
Dic
host
ereu
mD
icho
ster
eum
Dic
host
ereu
m? ? M
etu
lodo
nti
aC
onfe
rtob
asid
ium
Con
fert
obas
idiu
m? ?
S S S S S S S S S S
Eff
-Ref
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
S S S S S S S S P S
O O O S S S S S O O
Y Y Y N SP N Y Y Y Y
Y Y Y N/Y
Y Y Y Y Y Y
Y Y Y Y Y N Y Y Y Y
IV IV IV h IV IV h
He*
SN*
He
N N N
c c c c c c c c cG
loeo
hypo
chn
iciu
man
alog
um
Ech
inod
onti
um
ryva
rden
iiG
loeo
cyst
idie
llum
clav
ulig
eru
mG
loeo
cyst
idie
llum
purp
ure
mG
loeo
cyst
idie
llum
bisp
oru
mG
loeo
pen
ioph
orel
lala
xaG
loeo
pen
ioph
orel
laco
nvo
lven
sG
loeo
pen
ioph
orel
lasp
.L
acta
riu
sle
onis
Lac
tari
us
sube
nbo
ratu
s
gloe
ocys
tidi
ellu
mgl
oeoc
ysti
diel
lum
gloe
ocys
tidi
ellu
mru
ssul
ales
russ
ulal
esru
ssul
ales
russ
ulal
esru
ssul
ales
Glo
eohy
poch
nic
ium
? Glo
eocy
stid
iellu
mG
loeo
cyst
idie
llum
Glo
eocy
stid
iellu
mG
loeo
pen
ioph
orel
laG
loeo
pen
ioph
orel
laG
loeo
pen
ioph
orel
laL
acta
riu
sL
acta
riu
s
S S S S S S S S M M
Eff
Pil
Eff
Eff
Eff
Eff
Eff
Eff
Stip
Stip
S H S S S S S S L L
O O O O O O O O O O
N Y Y Y Y Y Y Y Y Y
Y Y N N/Y
Y Y Y Y Y N
N Y N N N N N N N N
? IV IV P
? N N HM
c b c c a a a a a

1045LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
TA
BL
EII
.C
onti
nue
d
Spec
ies
Cla
deG
enus
Lif
eB
asb
Hym
cSp
dA
me
SAf
Skeg
PhN
uci
Cj
Lac
tari
us
vole
mu
sR
uss
ula
viol
acea
Ru
ssu
laau
ran
tiac
aR
uss
ula
spha
gnop
hila
Ru
ssu
lan
ause
osa
Ru
ssu
lape
rsic
ina
Glo
eocy
stid
iellu
mac
ule
atu
mB
oidi
nia
pron
inqu
aB
oidi
nia
sp.
Boi
din
iafu
rfu
race
a
russ
ulal
esru
ssul
ales
russ
ulal
esru
ssul
ales
russ
ulal
esru
ssul
ales
russ
ulal
esru
ssul
ales
russ
ulal
esru
ssul
ales
Lac
tari
us
Ru
ssu
laR
uss
ula
Ru
ssu
laR
uss
ula
Ru
ssu
laB
oidi
nia
Boi
din
iaB
oidi
nia
Boi
din
ia
M M M M M M S S S S
Stip
Stip
Stip
Stip
Stip
Stip
Eff
Eff
Eff
Eff
L L L L L L S S S S
O O O O O O O O O O
Y Y Y Y Y Y Y Y Y Y
N Y Y N Y N Y Y Y Y
N N N N N N N N N Nh
N
a a a a a a a a cP
seu
doxe
nas
ma
verr
uci
spor
um
Glo
iodo
nst
rigo
sus
Glo
iodo
nn
igre
scen
sA
uri
scal
piu
mvu
lgar
eA
uri
scal
piu
mvi
llipe
sD
enti
prat
ulu
mbi
alov
iese
nse
Len
tin
ellu
som
phal
odes
Len
tin
ellu
sca
stor
eus
Len
tin
ellu
su
rsin
us
Len
tin
ellu
svu
lpin
us
auri
scal
piac
eae
auri
scal
piac
eae
auri
scal
piac
eae
auri
scal
piac
eae
auri
scal
piac
eae
auri
scal
piac
eae
auri
scal
piac
eae
auri
scal
piac
eae
auri
scal
piac
eae
Pse
udo
xen
asm
aG
loio
don
Glo
iodo
nA
uri
scal
piu
mA
uri
scal
piu
mD
enti
prat
ulu
mL
enti
nel
lus
Len
tin
ellu
sL
enti
nel
lus
Len
tin
ellu
s
S S S S S S S S S P
Eff
Eff
-Ref
Eff
-Ref
Stip
Pil
Cla
vPi
lPi
lPi
lPi
l
S H H H H H L L L L
O O O O O O O O O O
Y Y Y Y Y Y Y Y Y Y
Y Y Y Y Y Y Y Y Y Y
N Y Y Y Y N Y Y Y Y
IV IV IV IV IV IV IV
N N
c c b c c c c c c cL
enti
nel
lus
auri
cula
Len
tin
ellu
sco
chle
atu
sG
loeo
cyst
idie
llum
suba
sper
ispo
rum
Glo
eodo
nti
aco
lum
bien
sis
Glo
eodo
nti
adi
scol
orG
loeo
don
tia
pyra
mid
ata
Con
fert
iciu
moc
hrac
eum
Meg
aloc
ysti
diu
mlu
ridu
mM
egal
ocys
tidi
um
leu
coxa
nth
um
Glo
eocy
stid
iellu
mw
aku
llum
auri
scal
piac
eae
auri
scal
piac
eae
gloe
odon
tia
gloe
odon
tia
gloe
odon
tia
gloe
odon
tia
ster
eale
sst
erea
les
ster
eale
sst
erea
les
Len
tin
ellu
sL
enti
nel
lus
Glo
eodo
nti
aG
loeo
don
tia
Glo
eodo
nti
aG
loeo
don
tia
Con
fert
iciu
mM
egal
ocys
tidi
um
Meg
aloc
ysti
diu
mM
egal
ocys
tidi
um
S S S S S S S S S S
Pil
Pil
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
L L S H H H S S S S
O O O O O O S S S S
Y Y Y Y Y Y Y Y Y Y
Y Y Y Y Y Y N N N N
Y Y N N Y Y N N N N
h II II H H H H
N N HC
He
He
HC
c c c c c a c c ovA
leu
rodi
scu
sce
russ
atu
sA
leu
rodi
scu
sde
xtri
noi
deoc
eru
ssat
us
Glo
eom
yces
gram
inic
ola
Ale
uro
botr
ysbo
tryo
sus
Glo
eocy
stid
iellu
mfo
rmos
anu
mG
loeo
cyst
idie
llum
com
pact
um
Glo
eocy
stid
iellu
mas
pellu
mM
egal
ocys
tidi
um
chel
idon
ium
Boi
din
iam
acro
spor
aSt
ereu
mre
flex
ulu
m
ster
eale
sst
erea
les
ster
eale
sst
erea
les
ster
eale
sst
erea
les
ster
eale
sst
erea
les
ster
eale
sst
erea
les
? ? Glo
eom
yces
Ale
uro
botr
ys? ? ? ? ? St
ereu
m
S S S S S S S S S S
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
Eff
-Ref
S S S S S S S S S S
S S S O O O O S O S
Y Y N Y Y Y Y Y Y Y
Y Y Y Y Y Y Y Y Y 0
N N N N N N N N N N
II H II h hII
H
He
Hc
SN SN N HD
c c ov i c c c c

1046 MYCOLOGIAT
AB
LE
II.
Con
tin
ued
Spec
ies
Cla
deG
enus
Lif
eB
asb
Hym
cSp
dA
me
SAf
Skeg
PhN
uci
Cj
Ster
eum
subt
omen
tosu
mSt
ereu
mhi
rsu
tum
Ster
eum
sp.
Ster
eum
rugo
sum
Aca
nth
ophy
sellu
mliv
idoc
aeru
leu
mX
ylob
olu
sfr
ust
ula
tus
Con
fert
iciu
mra
vum
Glo
eocy
stid
iops
ishe
imii
Glo
eocy
stid
iellu
mcr
ypta
can
thu
mG
loeo
cyst
idio
psis
flam
mea
ster
eale
sst
erea
les
ster
eale
sst
erea
les
ster
eale
sst
erea
les
ster
eale
sst
erea
les
ster
eale
sst
erea
les
Ster
eum
Ster
eum
Ster
eum
Ster
eum
Xyl
obol
us
Xyl
obol
us
Glo
eocy
stid
iops
isG
loeo
cyst
idio
psis
Glo
eocy
stid
iops
isG
loeo
cyst
idio
psis
S S S S S S S S S S
Eff
-Ref
Eff
-Ref
Eff
-Ref
Eff
-Ref
Eff
Eff
Eff
Eff
Eff
Eff
S S S S S S S S S S
S S S S S S O O O O
Y Y Y Y Y Y Y Y Y Y
0 0 0 0 Y 0 Y Y Y Y
N N N N N N N N N N
H H H II H H H H
HC
HC
HC
As
HC
HC
HC
HC
v v v va r ov v vA
leu
rodi
scu
sam
orph
us
Art
omyc
espy
xida
tus
Am
ylos
tere
um
areo
latu
mA
myl
oste
reu
mla
evig
atu
mA
myl
oste
reu
mch
aille
tii
Glo
eocy
stid
iellu
msp
.G
loeo
cyst
idie
llum
poro
sum
Glo
eocy
stid
iellu
msp
.B
oidi
nia
gran
ula
taH
eric
ium
alpe
stre
ster
eale
sam
ylos
tere
acea
eam
ylos
tere
acea
eam
ylos
tere
acea
eam
ylos
tere
acea
egl
oeoc
ysti
diel
lum
gloe
ocys
tidi
ellu
mgl
oeoc
ysti
diel
lum
gloe
ocys
tidi
ellu
mh
eric
iace
ae
Ale
uro
disc
us
Art
omyc
esA
myl
oste
reu
mA
myl
oste
reu
mA
myl
oste
reu
mG
loeo
cyst
idie
llum
Glo
eocy
stid
iellu
mG
loeo
cyst
idie
llum
Glo
eocy
stid
iellu
mH
eric
ium
S S S S S S S S S P
Eff
Cla
vE
ff-R
efE
ffE
ffE
ffE
ffE
ffE
ffPi
l
S S S S S S S S S H
O S S S S O O O O O
Y Y Y Y Y Y Y Y Y Y
0 Y Y Y Y Y Y Y Y N
N N Y N Y N N N N N
IV IV IV IV II IV
N N N N N N
a c c c c c c cH
eric
ium
amer
ican
um
Her
iciu
mab
ieti
sH
eric
ium
erin
aceu
sH
eric
ium
cirr
hatu
mH
eric
ium
cora
lloid
esD
enti
pelli
sdi
ssit
aD
enti
pelli
sfr
agili
sD
enti
pelli
ssp
.L
axit
extu
mbi
colo
rA
lbat
rellu
sco
nfl
uen
s
her
icia
ceae
her
icia
ceae
her
icia
ceae
her
icia
ceae
her
icia
ceae
her
icia
ceae
her
icia
ceae
her
icia
ceae
her
icia
ceae
alba
trel
lus
Her
iciu
mH
eric
ium
Her
iciu
mH
eric
ium
Her
iciu
mD
enti
nel
lisD
enti
nel
lisD
enti
nel
lisL
axit
extu
mA
lbat
rellu
s
P S P P S S P S S M?
Pil
Pil
Pil
Pil
Pil
Eff
Eff
Eff
Eff
-Ref
Stip
H H H H H H H S S P
O O O S O O O O O S
Y Y Y Y Y Y Y Y Y Y
N N N N N N N N N N
N N N N N N N N N N
IV IV IV IV II IV IV
N N
b b c c c c c cA
lbat
rellu
spe
s-ca
prae
Alb
atre
llus
subr
ube
scen
sA
lbat
rellu
sov
inu
sA
leu
rocy
stid
iellu
mdi
scif
orm
eA
leu
rocy
stid
iellu
msu
bcru
enta
tum
Bon
darz
ewia
berk
elev
iH
eter
obas
idio
nan
nos
um
Lau
rilia
sulc
ata
Ech
inod
onti
um
tin
ctor
ium
Scyt
inos
trom
ella
hete
roge
nea
Wri
ghto
pori
aav
ella
nea
Wri
ghto
pori
ale
nta
alba
trel
lus
alba
trel
lus
alba
trel
lus
aleu
rocy
stid
iellu
mal
euro
cyst
idie
llum
bon
darz
ewia
ceae
bon
darz
ewia
ceae
bon
darz
ewia
ceae
bon
darz
ewia
ceae
scyt
inos
trom
ella
scyt
inos
trom
ella
Alb
atre
llus
Alb
atre
llus
Alb
atre
llus
Ale
uro
cyst
idie
llum
Ale
uro
cyst
idie
llum
Bon
darz
ewia
Het
ereb
asid
ion
Lau
rilia
Ech
inod
onti
um
Scyt
inos
trom
ella
? Wri
ghto
pori
a
M?
M M S S P P S P S S S
Stip
Stip
Stip
Eff
Eff
Pil
Eff
Eff
-Ref
Pil
Eff
Eff
Eff
P P P S S P P S H S P P
S S S O O O O O O O O O
N Y N Y Y Y N Y Y Y Y Y
N N N Y 0 Y Y N N Y 0 Y
N N N N Y Y Y Y Y Y Y Y
IV IV II II IV
He
He
N N
c a a c c c c c c c c

1047LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
←a Life 5 life strategy S 5 saprophytic. M 5 mycorrhizal; P 5 potentially parasitic.b Bas 5 basidiome morphology; Clav 5 clavarioid; Eff 5 effused; Eff-Ref 5 effused-reflexed; Pil 5 pileate; Stip 5 stipitate.c Hym 5 hymenophore configuration; H 5 hydnoid; L 5 lamellate; P 5 poroid; S 5 smooth.d Sp 5 basidiospore surface; L 5 smooth but lobed; O 5 ornamented; S 5 smooth.e Am 5 basidiospore wall reaction to Melzer’s; N 5 negative; Y 5 positive.f SA 5 reaction of gloeocystidia to sulfovanilline; N 5 negative; Y 5 positive; 0 5 gloeocystidia lacking.g Ske 5 presence of skeletal hyphae; N 5 skeletal hyphae lacking; Y 5 skeletal hyphae present.h P 5 sexual system (polarity); H 5 homothallic; h 5 heterothallic but polarity unknown; II 5 heterothallic bipolar; IV 5
heterothallic tetrapolar; P 5 parthenogenetic.i Nuc 5 nuclear behavior; As 5 astatocoenocytic; HC 5 holocoenocytic; HD 5 holodikaryotic; He 5 heterocytic; HM 5
holomonokaryotic; N 5 normal; SN 5 subnormal. An asterisk indicates that irregularities occur. For a description of thedifferent types of nuclear behavior see Boidin and Lanquetin (1984b).
j C 5 occurence of clamps in cultured mycelium; a 5 totally absent; b 5 clamps present but distribution not known; c 5constant, i 5 inconstant, clamps lacking at some septa; ov 5 opposite clamps at some septa but most septa simple; r 5 clampsrare; v 5 verticillate clamps present at subicular hyphae; va 5 variable, different septation at aerial and submerged hyphae.The terminology follows Boidin and Lanquetin (1984b).
and Lanquetin (1984b, 1997) and Boidin (1990). To visu-alize the distribution of observed characters, TABLE II showsspecimens in the order they occur in the strict consensustree (FIG. 2).
RESULTS
Initial analyses included sequences of Wrightoporiabracei (Murrill) Ryvarden, Dendrothele spp. and Dicho-pleuropus sp., which in preliminary tests with ourhomobasidiomycete inclusive dataset showed affinityto the russuloid clade. These taxa consistently gen-erated long branches because of extensive sequencedeviations in regions otherwise easily aligned. What-ever the cause of these differences, single taxa onlong branches might have negative effects on thephylogenetic analyses through so-called long-branchattraction (Felsenstein 1978). The problem can beaddressed best through the addition of taxa that canbreak up the long branches. Since we could not findsuch taxa, we preferred to exclude the above-men-tioned taxa from the present study. However, theyshould be given renewed consideration in futureanalyses with an expanded sampling of the russuloidclade. The final dataset included 127 ingroup se-quences representing 43 genera and ca 120 species,two-thirds of which (80 species) have effused basi-diomata and either a smooth or a hydnoid hymen-ophore. They would be considered by most mycolo-gists as corticioid and earlier were classified in thefamily Corticiaceae (FIG. 1).
The aligned data matrix covered 1993 base pairs(bp), starting at position 60 in the 5.8S region andincluding ITS2 and the 59 end of the LSU. Gaps forinsertion-deletion events were introduced to aid inthe alignment. The partial 5.8S region was easilyaligned through all sequences. The ITS2 region was
impossible to align over the entire dataset and, ex-cept for a small region of 12 base pairs, these char-acters were excluded. The 59 end of the LSU waseasily aligned in the conservative parts but moreproblematic in the variable domains D1–D3 (Hoppleand Vilgalys 1999). Most characters from ambiguousregions were excluded from the analysis. The finaldataset included 1240 aligned positions, of which 170were variable but uninformative and 364 were parsi-mony informative.
The phylogenetic analysis recovered 24 772 equallyparsimonious trees of 2468 steps (CI 5 0.3002, RI 50.7297). The trees were recovered from 263 islands,where the islands found in different replicates in factmight belong to the same island. A set of 2630 most-parsimonious (MP) trees (10 from each island) wereselected and kept for comparisons and analyses. FIG-URE 1 illustrates one of the MP trees presented as aphylogram to show the number of character statechanges per branch. The tree selected was one of theMP trees with the best-likelihood score. Species thathave been combined within Gloeocystidiellum aremarked with filled dots. FIGURE 2 illustrates the strict-consensus tree of all 24 772 MP trees.
The bootstrap analysis recovered 13 major sup-ported clades and six species on single branches.Each clade and its corresponding bootstrap value areindicated in the phylogram (FIG. 1) and the strict-consensus tree (FIG. 2). We have chosen names thatroughly correspond to the current concepts of or-ders, families and genera in fungal taxonomy (Hawk-sworth et al 1995). When competing names are avail-able, we have selected the oldest one. The clades are:/peniophorales, /amylostereaceae, /gloeocystidiel-lum I, /gloeocystidiellum II, /auriscalpiaceae,/gloeodontia, /aleurocystidiellum, /hericiaceae,

1048 MYCOLOGIA
FIG. 1. Phylogeny of the russuloid lineage based on unweighted parsimony analysis of nuclear rDNA sequence data. Oneof 24 772 equally parsimonious trees depicted as a phylogram (tree length 2468, CI 5 0.3002, RI 5 0.7297). Filled dotsindicate species that have been combined in Gloeocystidiellum. Bootstrap support is noted only for major clades discussed inthe text.

1049LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
FIG. 2. Strict-consensus tree of 24 772 equally parsimonious trees. Bootstrap values for discussed major clades are givenas numeric values. Asterisks indicate other bootstrap values above 80%. Names marked with a dot represent genus typespecies.

1050 MYCOLOGIA
/bondarzewiaceae, /albatrellus, /scytinostromella,/russulales and /stereales. Taxa on single branchesare Wrightoporia tropicalis, W. lenta, Scytinostromellanannfeldtii, Gloeohypochnicium analogum, Echinodon-tium ryvardenii and Pseudoxenasma verrucisporum.The bootstrap consensus-tree topology was almostidentical to the strict-consensus tree with one maindifference, a node uniting /stereales and /gloeodon-tia in the strict-consensus tree. The NJ analysis recov-ered the same 13 clades as the parsimony analysis butwith a major difference in the basal topology. The NJtree places /peniophorales as a sister clade to the restof the ingroup (see FIG. 3). None of the MP treesrecovered a similar topology. However, short incom-plete (nonconstrained) heuristic searches with thesame settings as in the parsimony analysis, occasion-ally recovered a similar topology as the NJ tree. Suchtrees were three steps longer (2471 steps) than themost-parsimonious trees. Constrained parsimonyanalyses simulating the basal topology of the NJ tree(/peniophorales forced as a sister clade to the restof the russuloid clade) recovered 4700 trees of 2473steps (five steps longer than the unconstrainedtrees). The constrained trees were found not to besignificantly worse than the unconstrained trees,based on the KH test (P 5 0.2982–0.6028) and theSH test (P 5 0.754–0.959).
Heuristic searches, forcing constraints for mono-phyly of the Gloeocystidiellum porosum-clavuligerumcomplex, recovered 3600 trees of 2474 steps (sixsteps longer than the unconstrained trees). Thesetrees were found not to be significantly worse thanthe unconstrained trees by the KH test (P 5 0.1020–0.3651) or the SH test (P 5 0.515–0.892).
Recurrent heuristic searches under maximum-like-lihood criteria with starting trees from the MP anal-ysis recovered several trees with a better-likelihoodscore than the starting tree. In general the TBRbranch swapping recovered better likelihood scoresthan the NNI branch swapping, despite the incom-plete analyses. The result might indicate the exis-tence of more optimal trees not recovered in theheuristic searches.
Phylogenetic analysis of the realigned dataset re-stricted to /stereales recovered 12 MP trees of 407steps, based on 118 parsimony-informative characters(CI 5 0.5971, RI 5 0.6840). FIGURE 4 presents thestrict-consensus tree with bootstrap values indicatedabove branches. Six clades with bootstrap supportabove 50% were identified: /gloeocystidiopsis, /ster-eum, /xylobolus, /chelidonium, /cerussatus and/megalocystidium. Conferticium ochraceum is placedon a single branch between /chelidonium and /cer-ussatus. Aleurobotrys botryosus and Aleurodiscus amor-
phus also were placed on single branches at the baseof the tree.
Morphological and cytological features for all taxaare summarized in TABLE II.
DISCUSSION
The phylogenetic analyses identify 13 major cladeswith strong or fair bootstrap support. These cladesare recovered always and appear resistant to changesin taxon sampling, character selection and methodchosen for analysis. Basal nodes are poorly resolvedand most basal branches collapse to a polytomy inthe strict consensus and bootstrap trees.
A basal division of our dataset in /peniophoralesand /eurussuloid was recovered in the NJ tree (FIG.3) while the parsimony analyses generally did notsupport such a topology. As discussed below there aresome morphological trends in /peniophorales thatsets this clade apart from the rest of the russuloidlineage and the constrained analysis does not rejectthe NJ topology. A denser sampling within /penio-phorales eventually might show if the NJ topologyreceives increased support.
All species with a gloeoplerous hyphal system thatwe tested with our large homobasidiomycete datasetturned out to cluster with the russuloid clade. Thisgives strong indications that possession of a gloeo-plerous hyphal system, often primarily observed astubular gloeocystidia, is a synapomorphy for the rus-suloid clade. This assumption, however, must be test-ed finally with a more inclusive dataset. In most taxathe gloeoplerous system gives a positive reaction withsulfobenzaldehyde, while in others it reacts positiveonly when fresh but loses this reaction after someperiod of storage (e.g., Asterostroma). A third groupis formed by those taxa that never give a positive re-action with sulfobenzaldehyde. However, sequenceanalysis and morphological features support the hy-pothesis that all gloeoplerous systems within the rus-suloid lineage, whether sulfo-positive or not, are ho-mologous (TABLE II).
In species having basidiomes with a dense texturedominated by thick-walled hyphae, the gloeopleroushyphae might be hard to detect, e.g., in Heterobasi-dion annosum and Bondarzewia montana, which hasled to conflicting views presented in the literature(Redhead and Norvell 1993). In both cases sulfo-pos-itive gloeocystidia develop in culture (Gluchoff-Fias-son et al 1983). In Stereum Pers., typical gloeopleroushyphae are lacking but it seems likely that the lactif-erous hyphae present in most Stereum species are ho-mologues.
The amyloid reaction of the spore wall is a secondcharacter often used to characterize russuloid spe-

1051LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
FIG. 3. Neighbor-joining distance based phylogenetic analysis of the nuclear rDNA sequence dataset. The NJ tree places/peniophorales as a sister clade to the rest of the ingroup, /eurussuloid.

1052 MYCOLOGIA
FIG. 4. Strict-consensus tree of 12 equally parsimonious trees, resulting from parsimony analysis of the realigned datasetof /stereales. Basal nodes with bootstrap values above 50% are indicated. Stippled line indicates uncertain support.
cies. However, amyloidity is a common phenomenonamong homobasidiomycetes and not a synapomor-phy for the russuloid clade. For example, MucronellaFr., possesses amyloid spores and previously was as-sociated with Hericiales based on basidiome mor-phology but the genus lacks gloeocystidia. In agree-ment with this, phylogenetic analysis of rDNA se-quence data places Mucronella outside the russuloidclade (data not shown). The genus Peniophora Cookelacks an amyloid reaction entirely while other genera,e.g., Scytinostroma Donk and Albatrellus S.F. Gray,contain both amyloid and non-amyloid species.
Most species in the russuloid clade have basidio-spores with an ornamented surface. The nature ofthis ornamentation has been extensively studied(e.g., Capellano and Keller 1978, Keller 1986, 1997).An outer layer, tectum, forms the ornamentation,and the same layer is responsible for the amyloid re-action. The tectum can be covered by up to threeadditional layers. Species with smooth basidiosporeslack a tectum, and in TEM no difference in sporewall structure between amyloid and non-amyloidspores can be detected (Keller 1997). We invariablyhave found the presence or absence of an ornamen-tation to be a generic character within the russuloid
clade (but see /hericiaceae below). The fine struc-ture of the spore wall as observed in TEM has beenstudied in a limited number of species only and itsuse as a phylogenetically informative character can-not be evaluated.
Corner (1932a, b) introduced hyphal analysis andthe concept of hyphal systems in fungal anatomy andtaxonomy. He defined skeletal hyphae as thick-walledto subsolid, nonseptate cells that retain ability forgrowth at the thin-walled tip. They arise from gen-erative hyphae. He also coined the terms monomiticfor species with only generative hyphae, dimitic forspecies with generative and skeletal hyphae, and trim-itic for species with generative, skeletal, and bindinghyphae. Corner’s ideas gained wide acceptance andhave greatly influenced basidiomycete systematics ingeneral and generic concepts in polypores in partic-ular. For an overview of the subject, see Pegler (1996)and Hibbett and Thorn (2001).
In the russuloid clade, skeletal hyphae occur inmany species and several genera have been intro-duced for species with a dimitic hyphal system, viz.Scytinostromella Parmasto, Wrightoporia Pouzar andConfertobasidium Julich. However, among the clades,we identify only /scytinostromella as completely dim-

1053LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
itic. Genera such as Scytinostromella and Wrightoporiaseem to wither when scrutinized by molecular meth-ods, and Confertobasidium has its closest relative inthe monomitic Metulodontia Parmasto. The clade/auriscalpiaceae includes clearly dimitic species suchas Gloiodon strigosum and Auriscalpium vulgare andthe monomitic Dentipratulum bialovicense. A similarpicture is found in /gloeodontia, where dimitic spe-cies such as Gloeodontia discolor cluster with the mon-omitic Gloeocystidiellum subasperisporum. We con-clude that skeletal hyphae as presently defined havebeen overestimated as a taxonomic character at thegeneric level. Our results indicate that dimitic hyphalsystems have evolved many times within the russuloidclade and support the views presented by Hibbettand Thorn (2001).
Recent studies have shown that the agaricoid gen-era Russula and Lactarius form a monophyletic cladetogether with their gasteroid and pleurotoid relatives(Miller et al 2001, Calonge and Martin 2000, Henkelet al 2000, Binder and Bresinsky 2002). We show thatthis clade is nested among corticioid taxa in what wecall /russulales. Bootstrap support for /russulales isgood (86%) but internal branching is mainly unre-solved. A notable exception is the basal node con-necting Boidinia furfuracea to the rest of the clade,where bootstrap support is 76%. Boidinia furfuraceais a wood-inhabiting fungus that forms thin, white,strictly resupinate basidiomes on decaying coniferwood. According to Nakasone (1990), it is capable ofproducing extracellular oxidases and hence causes awhite rot. There is no report of mycorrhizal activityconnected to Boidinia or Gloeopeniophorella, the sec-ond corticioid genus in /russulales. The result sug-gests that the ancestor to the agaricoid radiation inRussula and Lactarius had a corticioid basidiome andwas saprotrophic. This is in accordance with the con-clusions in Hibbett et al (2000) that evolution haswitnessed several independent shifts in nutritionalmode from saprotrophy to mycorrhizal associations.It also is consistent with an evolution from simple tocomplex fruiting bodies as put forward in Hibbettand Binder (2002).
CLADE CHARACTERISTICS AND TAXONOMY
/peniophorales (bootstrap support 95%). Our sam-pling in this clade was too restricted to allow a de-tailed phylogenetic analysis. Peniophora, Scytinostromaand Vararia P. Karst. together hold close to 150 de-scribed species but here are represented by onlyeight specimens. Within /peniophorales, only twosubclades are distinct enough to be recognized, viz./asterostromataceae and /metulodontia (FIG. 2).
There is no obvious morphological synapomorphy
for /peniophorales. The most striking differencefrom /eurussuloid is the predominance of smooth-walled basidiospores and the tendency toward non-amyloid spore-walls. In addition, species in this cladealmost invariably have basidiomes with a smooth hy-menophore, while hydnoid, poroid and lamellatebasidiomata occur frequently in most of the otherclades. Monophyly for the same group of taxa thatwe here call /peniophorales was detected already inseveral other studies (Hibbett et al 1997, 2000, Hib-bett and Donoghue 2001, Hibbett and Binder 2002).
The genera Asterostroma Massee, Dichostereum Pilat,Lachnocladium Lev., Scytinostroma and Vararia usuallyhave been referred to a separate family, Lachnocla-diaceae (Reid 1965). They are held together mor-phologically by the thick-walled, dextrinoid hyphaetermed astero-, dendro- or dichohyphidia. These hy-phidia are functionally equivalent to binding hyphaefound in many polypores. It is the dominating hyphaltype in many species, imparting them with a more orless tough consistency. Most species in Lachnocladi-aceae do not form a closed hymenium. Instead, ba-sidia are initiated deep among the hyphidia and pen-etrate to the surface just enough to freely shed theirspores. This kind of hymenium has been termed acatahymenium as opposed to the more familiar eu-hymenium with a closed palisade of basidia (Lemke1964). Basidiomes with a catahymenium are adaptedto resist periods of drought and to quickly resumesporulation when conditions become more favorable.
In our analyses Peniophora, Gloiothele and Vesiculo-myces Hagstrom are nested among the traditionalmembers of Lachnocladiaceae. No species in thesegenera have dichohyphidia or asterohyphidia, but afew species in Peniophora have dendrohyphidia. SincePeniophora is the oldest genus name, we prefer tohave that situation reflected in the clade name.
The four species of Scytinostroma in our dataset alloccur on separate branches that are distantly sepa-rated. They are morphologically quite different. Scy-tinostroma portentosum has simple-septate hyphae,amyloid subglobose spores and SA1 gloeocystidia, S.odoratum has simple-septate hyphae, non-amyloid el-lipsoid spores and SA2 gloeocystidia, S. galactinumhas nodose-septate hyphae, non-amyloid (except forthe suprahilar patch) ellipsoid spores and SA1 gloeo-cystidia, and S. jacksonii has nodose-septate hyphae,non-amyloid, ovoid spores and SA2 gloeocystidia. Inaddition, the two species representing Vararia do notcluster together and also are separated from the fourScytinostroma branches.
The taxonomic distinction between Scytinostromaand Vararia has been questioned (Boidin and Lan-quetin 1987, Boidin et al 1998, Hallenberg 1985, Stal-pers 1996). However, there has been general agree-

1054 MYCOLOGIA
ment that the two genera are closely related and thatthey together make up a natural group. Our resultsstrongly suggest that neither skeletal hyphae northeir branching patterns have any predictive powerin a phylogenetic context.
Scytinostroma was introduced for species with dex-trinoid, sparsely branched, skeletal hyphae, differingfrom the richly branched dichohyphidia characteris-tic of Vararia (Donk 1956). It is typified by Scytinos-troma portentosum, which in our analyses takes a po-sition close to Gloiothele. We estimate that less than10 species have characters corresponding to those ofthe type. However, a thorough molecular investiga-tion of Scytinostroma, Lachnocladium and Vararia isnecessary before the circumscription of these generacan be settled.
Five species of Dichostereum (40% of known speciesincluding the type D. durum) cluster together andappear to be a monophyletic group. However, in thebootstrap tree the clade collapses to a polytomy. LikeVararia, the genus has dichohyphidia but differs byglobose, coarsely ornamented and strongly amyloidspores.
The two Peniophora species sampled cluster togeth-er. This large genus with many closely related speciesis ecologically quite distinct. Species typically arefound in exposed situations on dead but still-attachedbranches. Most species in Peniophora have nodose-septate hyphae, SA1 gloeocystidia, and characteristicthick-walled incrusted cystidia (metuloids). Sporesare always smooth and non-amyloid. Basidiomata ap-pear well adapted to desiccation and have a dense,often strongly pigmented hyphal structure.
Hallenberg et al (1996) investigated the phyloge-netic relationships in Peniophora. Three speciesgroups were identified, and the genus was confirmedas uniform and well distinguished. In the MP tree,Peniophora is positioned together with Vararia inves-tiens as a sister group to Dichostereum. However, thisarrangement has no bootstrap support.
/asterostromataceae (99%). This clade includes As-terostroma, Scytinostroma portentosum (see above),Gloiothele and Vesiculomyces. The position of Gloiotheleand Vesiculomyces in /peniophorales was unexpected.Both genera are characterized by simple septate hy-phae, a monomitic hyphal system and globose tosubglobose, smooth, amyloid spores. Gloiothele lamel-losa, type species of the genus, has a variable hymen-ophore that can be odontioid, irpicoid or subporoid.Thus it is not surprising that the species appears un-der additional names as Gloeocystidiellum irpiscescensBoidin and Vesiculomyces epitheloides Boidin & Lanq.(Hjortstam 1987a). The three specimens included inour sampling represent some of the hymenophore
variation. Molecular data support the conclusion byHjortstam (1987a) that these names refer to one andthe same species.
Vesiculomyces citrinus was segregated from Gloeocys-tidiellum because gloeocystidia in basidiomata areSA2 (Hagstrom 1977). Boidin (1958) and Maekawaet al (1982) have reported that SA1 gloeocystidiacould be found in cultured mycelium of V. citrinus.However, we could not confirm this observation inour own cultures. The position of V. citrinus in thetree as a sister group to Gloiothele fits well with mor-phological characters such as simple septate hyphae,narrowly clavate basidia and smooth globose amyloidspores with a prominent apiculus. We suggest thatVesiculomyces be retained as a separate genus.
Asterostroma (type species Corticium apalum Berk& Broome 5 A. muscicola) appears monophyleticwith 71% bootstrap support. In addition, the genusis morphologically well characterized by unique as-terohyphidia. This type of hyphidium is short, has adense branching and develops from thin-walled hy-phae. Because long, skeletal-like hyphidia are lack-ing, Asterostroma has soft, fragile basidiomata andthey usually are found in moist, sheltered places. Ba-sidiospores are always globose and either smooth orornamented. However, the ornamentation does notseem to be homologous to the ornamentation seenin other russuloid species. In the latter case, orna-mentation is caused by material deposited in the out-er layer of the spore-wall, while in Asterostroma or-namentation seems to consist of lobes formed by sev-eral layers of the spore wall. Boidin et al (1997b)recognized 14 species in Asterostroma. Nine of themare very similar morphologically and separated main-ly by differences in spore size and ornamentation.Our sampling includes two of these morphotypes,and we found the sequences to be almost identical.Additional sequencing in this species complexshould be undertaken.
/metulodontia (100%). In all analyses there isstrong bootstrap support for /metulodontia as themost basal subclade in /peniophorales. Parmasto(1968) introduced Metulodontia to accommodatespecies with thick-walled incrusted cystidia, so-calledmetuloids. Eriksson and Ryvarden (1976) showedthat the type species, Metulodontia nivea, differs fromall other species originally assigned to Metulodontiaby having SA1 gloeocystidia, and they regarded Me-tulodontia as a monotypic genus of uncertain affinity.Here we show that Metulodontia, despite lacking anamyloid reaction of the spores, belong to the russu-loid clade.
Julich (1972) introduced Confertobasidium forathelioid species with brownish basal hyphae and

1055LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
with C. olivaceoalbum as the type. Hjortstam (1987a)showed that the type species had been misunder-stood already by Bourdot and that two species—onewith SA1 gloeocystidia and skeletal hyphae and onelacking these characters—were mixed into the spe-cies concept. The type specimen selected by Julichhas the gloeocystidia and skeletal hyphae. For speci-mens lacking those elements, the epithet fuscostratusBurt is available, now referred to Leptosporomyces Ju-lich.
Confertobasidium olivaceoalbum and Metulodontiashare an athelioid basidiome construction, small el-lipsoid smooth basidiospores and short, narrow SA1gloeocystidia confined to the hymenium. It is a mat-ter of taste if one wishes to combine the two speciesin one genus or keep them separate. Metulodontiahas inamyloid spores, a monomitic context and twokinds of cystidia, while Confertobasidium has amyloidbasidiospores, a dimitic context and only one kind ofcystidium.
Ginns and Lefebvre (1993) recently transferredCorticium olivaceoalbum to Scytinostromella. Theystressed the presence of gloeocystidia, skeletal hy-phae and an athelioid basidiome as the main reasonsfor the transfer. In our tree, Confertobasidium oliva-ceoalbum is not placed close to Scytinostromella heter-ogenea, the generic type of Scytinostromella.
/amylostereaceae (73%). The clade includes Amylos-tereum and Artomyces pyxidatus (5 Clavicorona pyxi-data [Pers. : Fr.] Doty). No obvious morphologicalcharacteristics indicate a close relationship betweenthese taxa, and no one before has suggested such arelation. Amylostereum has leathery, more or less re-flexed basidiomata with a smooth hymenophore,while Artomyces Julich has erect, clavarioid basidiom-ata with a characteristic branching pattern. Connec-tions can be found in micromorphology. They sharea constantly clamped hyphal system, a SA1 gloeo-plerous system and smooth, amyloid spores. Amylos-tereum usually is described as lacking gloeocystidia,but Eriksson and Ryvarden (1973) noticed thin-walled gloeocystidia-like organs in A. chailletii. In cul-ture, SA1 gloeocystidia regularly occur (Boidin andLanquetin 1984a, Nakasone 1990).
In the analyses by Hibbett et al (1997) and Pine etal (1999), Clavicorona pyxidata is nested in a cladewith Auriscalpium vulgare and Lentinellus spp. Therelation to Lentinellus is supported as well in laterstudies (Hibbett et al 2000, Hibbett and Donoghue2001). Our contradictory results prompted us to re-sequence the culture used to generate our sequenceof Artomyces pyxidatus, but the outcome was identical.However, it is possible that the culture actually rep-
resents something else. The position of Amylostereumis discussed with /bondarzewiaceae below.
Clavicorona Doty is typified by C. taxophila (Thom)Doty, a small, nonbranched species, quite differentfrom the large, multibranched C. pyxidata. Julich(1981) questioned the homogeneity of the genus andreferred C. pyxidata to a separate genus, Artomyces.Recent systematic and monographic studies in Arto-myces and Clavicorona support this arrangement(Lickey 2002).
/gloeocystidiellum I (92%) and II (66%). Gloeocysti-diellum porosum is the generic type for Gloeocystidiel-lum. In our tree, it clusters with Boidinia granulataand two unidentified specimens. Gloeocystidiellum cla-vuligerum was first regarded as a synonym of G. po-rosum but reinstated as a distinct species after a closerinvestigation of morphology and culture characteris-tics (Nakasone 1982). Larsson and Hallenberg(2001) found that molecular data support G. porosumand G. clavuligerum as distinct species and that eachof them also encompasses other closely related taxa.Our analyses confirm the results of Larsson and Hal-lenberg (2001). Gloeocystidiellum clavuligerum and G.porosum do not cluster together in any of our analysesirrespective of method, sampling or characters se-lected. However, constrained trees forcing monophy-ly of G. porosum/clavuligerum were found not to besignificantly worse than the MP trees.
One mitochondrial small-subunit rDNA sequenceof Gloeocystidiellum porosum was included in the anal-yses by Hibbett and Donoghue (2001), where it oc-curred nested with Laxitextum bicolor and Dentipellisseparans (Peck) Donk. The apparent deviation fromour results might be explained by the restricted sam-pling used by Hibbett and Donoghue (2001). In theirdataset, G. porosum simply had no closer relative toassociate with. It should be noted that Dentipellis se-parans is a younger synonym of Dentipellis leptodon(Mont) Maas Geest (Ginns 1986). However, most col-lections in American herbaria named D. separans infact belong to Dentipellis dissita (Ginns 1986). This isalso the correct name for the specimen and the sub-sequent culture collected by N. Hallenberg and byhim determined as D. separans. The culture (FCUG581) was the sequence source for both D. separansused by Hibbett and Donoghue (2001) and for D.dissita used in this study!
All species in Gloeocystidiellum sensu stricto arestrictly resupinate and have a smooth hymenophore.They are monomitic, have nodose-septate hyphae,gloeocystidia that are either SA1 or SA2, ellipsoidornamented amyloid spores and a heterothallic tetra-polar mating system. Gloeocystidiellum bisporum is a

1056 MYCOLOGIA
haploid mitosporic derivative from G. clavuligerum,and its status as a distinct species can be questioned.
Our analyses imply that Gloeocystidiellum porosumand G. clavuligerum have a long history as separatetaxa, but the generally low resolution at basal nodesprevents definite conclusions about phylogenetic re-lationships and generic limits. Because morphologi-cal characteristics of the two clades overlap almostentirely, we think it would be impractical to introducea new genus for the taxa around G. clavuligerum.
/auriscalpiaceae (86%). This clade corresponds tothe family Auriscalpiaceae. At its creation, Maas Gees-teranus (1963) included the hydnoid genera Auris-calpium Gray and Gloiodon P. Karst. and the lamellateLentinellus P. Karst. Most of the species have a dimitichyphal system with clamps and gloeocystidia that givea SA1 reaction. Spores are subglobose to ellipsoid,ornamented, and with a strong amyloid reaction.
Analyses recovered two distinct subclades, with71% and 86% bootstrap support respectively. Thefirst subclade includes five species with a hydnoid hy-menophore, viz. Auriscalpium vulgare, A. villipes,Gloiodon strigosum, G. nigrescens and Dentipratulumbialoviesense. Auriscalpium vulgare is stipitate with areniform pileus. The abhymenial surface is hairy anddark brown. Auriscalpium villipes has a similar con-struction, but pilei have only a short stipe or are ses-sile and more or less broadly attached. Gloiodon spe-cies are resupinate or effused-reflexed, and Dentipra-tulum Domanski finally has a strongly reduced basi-diome, consisting of scarcely separate spinesconnected by a barely visible sterile mycelium. Theinclusion of Dentipratulum in /auriscalpiaceae sup-ports earlier results by Boidin et al (1998).
Gloiodon and Auriscalpium are very similar, as al-ready noted by several authors (e.g., Koski-Kotirantaand Niemela 1988). They are dimitic, and the gloeo-plerous hyphae are less obvious than in the othergenera in this clade. The only difference is the ef-fused-reflexed basidioma in Gloiodon as opposed tothe stipitate basidioma in Auriscalpium. It must bequestioned if Gloiodon should be maintained as anindependent genus.
The second subclade consists of the lamellate ge-nus Lentinellus and has a bootstrap support of 86%.The genus is characterized by sessile or stipitate, fla-belliform basidiomata with serrate lamellae. The pi-leus is convex to depressed and glabrous or tomen-tose-villose. Five or six species occurring in NorthernEurope, including the type species L. cochleatus, havebeen sequenced. Lentinellus ursinus and L. castoreusare almost identical and differ only by the distribu-tion of hair on the cap surface and preference forconifer versus deciduous wood. Some authors have
accepted them as separate species (Ryman and Hol-masen 1984, Watling and Gregory 1989), while oth-ers have treated them as synonyms (Miller and Stew-art 1971, Printz 1986, Stalpers 1996). In this study,the glabrous and tomentose variants were shown tohave identical nucleotide sequences. However, con-sidering their different ecology, it is still possible thatthey genetically behave as independent species.
/gloeodontia (78%). This clade includes GloeodontiaBoidin and Gloeocystidiellum subasperisporum. A rela-tionship between these taxa was shown by Boidin etal (1998) based on ITS sequence data. Nodose sep-tate hyphae, verrucose amyloid spores and SA1gloeocystidia unite the species. Gloeodontia specieshave hydnoid to irpicoid basidiomata, more or lessthick-walled to dimitic hyphae, encrusted hyphoidcystidia in the hymenium and ellipsoid to subglobosebasidiospores. Gloeocystidiellum subasperisporum has asmooth hymenophore, a monomitic hyphal system,no encrusted cystidia, and reniform basidiospores.Gloeodontia columbiensis and G. discolor are hetero-thallic unifactorial (Boidin and Lanquetin 1984b).Gloeocystidiellum subasperisporum is confirmed as het-erothallic, but factorial type is not identified. Allthree species have a normal nuclear behavior (Boidinand Lanquetin 1984b, Boidin et al 1997a).
Hjortstam and Ryvarden (1988) referred Gloeocys-tidiellum subasperisporum to Amylosporomyces S.S. Rat-tan, a genus described as lacking any kind of cystidia(Rattan 1977). We restudied type material of Amylos-poromyces echinosporus and found that it has numer-ous gloeocystidia and spores that are almost identicalto those in Gloeocystidiellum subasperisporum. Thetype collection is not in the best condition and thegloeocystidia do not react with sulfovanillin, whichmight explain why they have gone unnoticed. We re-gard Amylosporomyces echinosporus as a synonym ofGloeocystidiellum subasperisporum. In our tree, G. su-basperisporum occurs nested in Gloeodontia and wesuggest that G. subasperisporum be transferred toGloeodontia.
/aleurocystidiellum (99%). Aleurocystidiellum sub-cruentatum is a dimitic species with discoid basidiom-ata and large, minutely verrucose, amyloid spores.The skeletal hyphae terminate in the hymenium ascystidia-like, more or less encrusted elements. Gloeo-cystidia or gloeoplerous hyphae seem to be lacking.These features prompted Lemke (1964) to segregatesubcruentatum from Aleurodiscus Rabenh. ex J.Schrot., where the species had been placed by mostauthors because of the discoid basidiomata and theamyloid basidiospores. Boidin et al (1968) found thatAleurodiscus disciformis exhibited almost identical cul-ture characteristics, although basidiomata differed by

1057LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
being monomitic and by possessing SA1 gloeocysti-dia. Consequently they intended to make the appro-priate combination in Aleurocystidiellum Lemke butfailed to include a reference to the basionym. Tellerıa(1990) later provided a correct combination. Hallen-berg and Parmasto (1998) found that molecular datasupported a close relationship of the two species.They also studied basidiospores in SEM and showedthat the ornamentation in Aleurocystidiellum speciesis identical and clearly different from Aleurodiscusamorphus, type of Aleurodiscus. Both Aleurocystidiel-lum species are heterothallic tetrapolar and have aheterocytic nuclear behavior (Boidin et al 1968).They also have a similar ecology; they grow on thebark of living trees. In the tree presented by Wu etal (2001) Aleurocystidiellum is interpreted as belong-ing to the ingroup but actually is a sister group tothe rest of the ingroup. This position is still compat-ible with our tree, where Aleurocystidiellum has noclear connection to other clades.
/hericiaceae (87%). This clade includes three gen-era: Hericium with large coralloid, pileate or substip-itate basidiomata supporting a strongly hydnoid hy-menophore, Dentipellis Donk with effused-reflexedbasidiomata and a hydnoid hymenophore, and Lax-itextum Lentz with effused-reflexed basidiomata anda smooth hymenophore. They are all parasitic or sap-rotrophic and associated with a white rot. Morpho-logical characters uniting the clade are the monom-itic hyphal system with clamp connections, SA2gloeocystidia and relatively small subglobose to ellip-soid, slightly thick-walled and amyloid spores. Speciesseem to have a bifactorial mating system and are ei-ther heterothallic or amphithallic, and they have anormal nuclear behavior (Boidin and Lanquetin1984b, Boidin 1990).
Hericium cirrhatum usually has been referred to aseparate genus Creolophus P. Karst. One reason is thatbasidiospores have been interpreted as smooth. How-ever, when observed in SEM the spore surface ap-pears slightly rugose (Keller 1997). We cannot judgewhether this is an artifact caused by the preparationmethod or a true ornamentation. Since Hericium cir-rhatum in our tree occurs nested among Hericiumspecies that all have ornamented spores, we find itmost logical to regard Creolophus as a superfluous ge-nus.
The specimen called Dentipellis sp. is resupinatewith a smooth hymenophore. It is another strikingexample that basidiome morphology and hymenop-hore configuration are mostly of limited value as in-dicator of relationships.
/bondarzewiaceae (78%). This clade includes spe-cies with resupinate, effused-reflexed or pileate basi-
diomata and with smooth, poroid or hydnoid hymen-ophore configuration. All species cause a severewhite rot and several species, e.g., Bondarzewia ber-keleyi, Echinodontium tinctorium and Heterobasidionannosum, can attack living trees. The hyphal systemis characterized as di- or trimitic but, according toStalpers (1979), the skeletal hyphae are merely scler-ified generative hyphae with more distant septa thanusual. Donk (1964) did not regard Bondarzewia, Echi-nodontium, Heterobasidium and Laurilia as related,and he referred them all to different families, viz.Bondarzewiaceae, Echinodontiaceae, Polyporaceaeand Stereaceae. Stalpers (1979) suggested Bondar-zewiaceae sensu Donk also should include Heterobas-idion Bref., Echinodontium and Laurilia Pouzar. Hisview was further strengthened by the discovery thatboth Bondarzewia and Heterobasidion have a system ofSA1 gloeoplerous hyphae (Gluchoff-Fiasson et al1983). A summary of opinions on family arrange-ments can be found in Redhead and Norvell (1993).In the tree presented by Hibbett et al (1997), Bon-darzewia, Echinodontium and Heterobasidion belong tothe same superclade but not close together. In sub-sequent studies (Hibbett et al 2000, Hibbett andDonoghue 2001, Binder and Hibbett 2002, Hibbettand Binder 2002), Bondarzewia and Heterobasidioncluster together but always distinctly removed fromEchinodontium. Monophyly for Heterobasidion andBondarzewia also was detected by Bruns et al (1998).
Gross (1964) found the pileate-hydnoid Echinodon-tium and the resupinate-smooth Laurilia to be con-generic. They share constant presence of clamps,thick-walled encrusted cystidia, and a brown-coloredcontext. Molecular data presented here and by oth-ers (Hibbett and Donoghue 2001, Hibbett and Bind-er 2002) support this view, and we suggest that Laur-ilia be relegated to synonymy.
Amylostereum and Echinodontium cluster togetherwith high bootstrap support in the analysis by Hibbettet al (2000), Hibbett and Donoghue (2001), andBinder and Hibbett (2002). In our analyses, Amylos-tereum always is removed from Echinodontium insteadforming a moderately supported clade with Artomycespyxidatus. We have no explanation for this discrep-ancy; the rigorous four-gene analyses performed byBinder and Hibbett (2002) presents strong argu-ments for a phylogenetic connection between Amy-lostereum and Echinodontium. Similar morphologicalarguments can be found in the numerous conical,apically encrusted cystidia present in both genera.
Echinodontium ryvardenii is a recently describedspecies only found on Juniper on Corsica (Bernicchiaand Piga 1998). Despite having a hydnoid hymen-ophore, a dimitic hyphal system, amyloid, ornament-ed spores and SA1 gloeocystidia, it is not related to

1058 MYCOLOGIA
the other Echinodontium species. Its true affinities areunclear.
/albatrellus (100%). The four species of Albatrellusform a monophyletic group that includes the type A.ovinus. All species in the genus have stipitate basi-diomata with a poroid hymenophore. They form ec-tomycorrhiza and are associated with coniferous trees(Agerer et al 1996, Ryvarden and Gilbertson 1993).
Bruns et al (1998) found that Albatrellus syringae(Parmasto) Pouzar and A. peckianus were placed out-side their Russula-containing clade. A paraphyleticorigin for Albatrellus later was confirmed by Binderand Hibbett (2002) and Hibbett and Binder (2002).
The position of /albatrellus in the russuloid line-age is unclear, but morphological similarities existwith Hericium and Lentinellus. Both Albatrellus andHericium are monomitic with nodose septate hyphaeand have SA2 gloeocystidia. Albatrellus have amyloidhyphae in the context of the basidiomata, a characterfound also in Hericium and Lentinellus. In Albatrellusthe presence of clamp connections and amyloidity ofspores seem to be inconsistent characters, as A. sub-rubescens and A. ovinus lack clamps and only A. con-fluens and A. subrubescens have amyloid spores.
/scytinostromella (71%). Scytinostromella heterogenea,type of the genus, and Wrightoporia avellanea makeup this small clade. They share a dimitic hyphal sys-tem with nodose septate generative hyphae and atleast partly dextrinoid skeletal hyphae. They alsoshare small, ellipsoid, finely ornamented basidio-spores and a gloeoplerous system. However, supportfor the clade is not impressive and with only two ter-minal taxa the result must be regarded with caution.
Scytinostromella was introduced as a genus for re-supinate species with Gloeocystidiellum-like featuresbut with a dimitic instead of monomitic hyphal sys-tem (Parmasto 1968). Only two species originallywere included, the second one being Scytinostromellanannfeldtii (erroneously named subasperisporum byParmasto). In our tree, S. nannfeldtii is placed on asingle branch with no connection to the type species;its generic position remains unclear. We have se-quenced Scytinostromella cerina, and this species alsooccurs on a separate single branch (data not shown).
/russulales (86%). This clade includes corticioidwood-decaying species, e.g., Gloeopeniophorella convol-vens, Gloeocystidiellum aculeatum and Boidinia propin-qua and the whole family Russulaceae, with the ec-tomycorrhizal genera Russula and Lactarius and re-lated gasteroid and pleurotoid forms. Species in thisclade have a monomitic hyphal system, spores areglobose to subglobose often with a distinct ornamen-tation and a strong amyloid reaction, and gloeocys-
tidia are sulfo-positive. All species except Boidinia fur-furacea have simple septate hyphae. Spores are uni-nucleate and the secondary mycelium dikaryoticwhich indicates a normal nuclear behavior. Presum-ably most species are heterothallic (Boidin et al1997a). Gloeopeniophorella Rick differs from the otherspecies in the clade by the presence of incrusted cys-tidia (metuloids).
Boidinia furfuracea is placed at the first node in/russulales as a sister group to remaining taxa andmonophyly of species with simple septate hyphae issupported by a bootstrap value of 76%. Boidinia wassegregated from Gloeocystidiellum as a monotypic ge-nus (Hjortstam and Stalpers 1982). The authorsstressed basidiome structure and spore morphologyas the main diagnostic features. Later authors havepointed to the suburniform shape of basidia as anadditional characteristic ( Julich 1982, Hjortstam andRyvarden 1988, Ginns and Freeman 1994). Severalspecies subsequently have been placed in Boidinia,and three of them are included in our tree. Boidiniagranulata belongs in /gloeocystidiellum II, and Boi-dinia macrospora clusters in /stereales. Only Boidiniapropinqua belongs to /russulales, but its relation toBoidinia furfuracea could not be resolved. As notedabove, Boidinia propinqua differs from B. furfuraceaby simple-septate hyphae. Two other species with sim-ple septate hyphae, viz. Gloeocystidiellum aculeatumand Boidinia sp., also have their place in /russulales.Our results indicate that Boidinia still should be treat-ed as a monotypic genus. However, before a moredetailed view of the clade can be worked out we willuse the concept adopted by Hjortstam and Ryvarden(1988) and further developed by Wu and Buchanan(1998) and accept also species with simple-septate hy-phae in Boidinia.
Gloeopeniophorella was introduced with one speciesG. rubroflava Rick (Rick 1934) but since its introduc-tion has been largely ignored. Recently Boidin et al(1997) restudied the type specimen and concludedthat it shares similarities with Gloeocystidiellum con-volvens and G. laxum. They also made the appropri-ate combinations. In our tree Gloeopeniophorella con-volvens, G. laxa and one specimen closely related toG. laxa form a subclade in /russulales with weakbootstrap support (66%). The reintroduction ofGloeopeniophorella is a necessary step in the rear-rangement of Gloeocystidiellum. However, its relationto the nonclamped species now placed in Boidinianeeds further study.
/stereales (97%). This clade is strongly supported bybootstrap, and the same result was reported by Wuet al (2001). Species in this clade are saprotrophscausing a white rot on various deciduous and conif-

1059LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
erous trees. Many species, e.g., Stereum and Aleurod-iscus, decay dead but still attached branches and fruiton trunks or dead parts of living trees. Several specieshave adaptations for resisting periods of dry weather,e.g., a leathery consistency in Stereum or a catahy-menial organization of the basidioma in Aleurodiscus(Boidin et al 1985).
There are some striking similarities with /penio-phorales: Many species have smooth spores, manyspecies have simple septate hyphae, all species havea smooth hymenophore, and catahymenial organi-zation of the basidiomes is widespread. The restricteddataset of the clade supported six subclades, /gloeo-cystidiopsis, /xylobolus, /stereum, /chelidonium,/cerussatus and /megalocystidium (FIG. 4). It shouldbe noted that the NJ tree has a rather different to-pology within /stereales and both /gloeocystidiopsisand /chelidonium collapses.
/gloeocystidiopsis. A weakly supported clade (57%),consisting of two subclades, each with stronger sup-port. In one subclade, the generic type Gloeocystidiop-sis flammea groups with Gloeocystidiellum cryptacan-thum (99% bootstrap support). The other clade con-sists of Gloeocystidiopsis heimii and Conferticium ravum(81% bootstrap support). All species are monomiticwith simple septate hyphae. Gloeocystidia are long,tubular and always with a SA1 reaction, although itcan be weak in G. flammea (Boidin et al 1997a).Spores are ellipsoid, finely verrucose and stronglyamyloid. According to Boidin et al (1997a), all fourspecies are homothallic with binucleate spores andmultinucleate primary and secondary mycelium (hol-ocoenocytic). Clamps are absent from basidiomes,but in cultured mycelium opposite or verticillateclamps can be found.
Gloeocystidiopsis Julich was introduced as a genusfor Gloeocystidiellum-like species, with simple septatehyphae and a coenocytic nuclear behavior ( Julich1982). Two species, Gloeocystidiopsis flammea and G.heimii, originally were included. Wu (1996) movedheimii to Conferticium Hallenb. mainly because hethought that the dense hyphal structure in this spe-cies is typical for Conferticium but not present in thetype of Gloeocystidiopsis. Boidin et al (1997a) accept-ed that arrangement. Conferticium indeed is charac-terized by a dense, almost hard context and basidi-omes often are stratified. Species are monomitic withsimple septate hyphae and internal basidia repetition(Hallenberg 1980). Hallenberg included three spe-cies, viz. C. insidiosum (Bourdot & Galzin) Hallenb.(type), C. ochraceum and C. ravum (as C. karstenii).The type species and C. ochraceum are morphologi-cally indistinguishable but have different substratepreferences, growing on angiosperm and conifer
hosts respectively. In our tree, Conferticium ochraceumdoes not cluster with C. ravum but occurs on a singlebranch. We conclude that the similarities in basidi-oma structure and basidia regeneration seen amongspecies in Conferticium are of uncertain value for agenus characterization. Further arguments in sup-port of the results in the phylogenetic tree are thatConferticium insidiosum and C. ochraceum havesmooth spores, while C. ravum has ornamentedspores. Conferticium insidiosum and C. ochraceumhave similar nuclear behavior as species in /gloeo-cystidiopsis but differ by being totally devoid ofclamps in culture.
/xylobolus (73%). The type species of Xylobolus P.Karst., X. frustulatus, is externally quite characteristicwith its dark brown, strictly resupinate and stronglycracked, stratified basidiomata. In micromorphologyit has clear similarities to Stereum but differs by a neg-ative reaction in tests for phenoloxidase activity andby a white-pocket rot. Wu et al (2001) found thatXylobolus frustulatus and Acanthophysellum lividoca-eruleum cluster together with a relatively strong boot-strap support, and we can confirm their results.These two species share characters such as dark-col-ored basidiomata, densely interwoven and fused hy-phae, acanthohyphidia, narrowly clavate basidia, SA2gloeocystidia, and smooth amyloid spores. They dif-fer by the positive phenoloxidase reaction and nor-mal white rot in A. lividocaeruleum. It is interestingto note that a French specimen of A. lividocaeruleumstudied by Boidin and Des Pomeys (1961) first wasidentified as an undescribed species of Xylobolus. Lat-er Boidin et al (1968) added culture characteristicsand studied nuclear behavior, which confirmed thesimilarities with X. frustulatus.
The genus Acanthophysellum Parmasto was intro-duced for resupinate species with acanthohyphidiaand smooth non-amyloid basidiospores (Parmasto1967). Only two species originally were assigned tothe genus, viz. Corticium lividocoeruleum P. Karst.(type species) and Corticium cerussatum Bres. Thestatement about non-amyloid spores must be regard-ed as a typographical error. A year later Parmasto(1968) treated Acanthophysellum as a genus withintribe Aleurodisceae defined as having amyloid spores.
Cunningham (1963) restricted Aleurodiscus to spe-cies with more or less discoid basidiomata and adopt-ed Acanthophysium (Pilat) G. Cunn. for species witha strictly resupinate basidioma. With this circumscrip-tion the micromorphological variation within Acan-thophysium becomes almost identical to that seen inAleurodiscus sensu lato and includes a variety of sterileorgans, both smooth and ornamented spores, simple-septate and nodose-septate hyphal systems, and spe-

1060 MYCOLOGIA
cies with and without gloeocystidia. The type speciesAleurodiscus apricans Bourdot has, according to theprotologue, both smooth and finely asperulatespores. Boidin et al (1985) used SEM to study thespores in one of the original collections and foundonly smooth spores. This specimen was selected aslectotype (Galzin 4157). Nunez and Ryvarden (1997)studied another of the original collections and foundall spores to be ornamented. Boidin et al (1985) re-stricted Acanthophysium to species with smooth amy-loid spores and SA1 gloeocystidia. Wu et al (2001)regards Acanthophysium as a synonym of Xylobolusbased on similarities in nuclear behavior, the negativephenoloxidase reaction, and white-pocket rot (Boi-din et al 1968). As a consequence, some of the spe-cies placed in Acanthophysium were transferred toAcanthophysellum (see also Boidin and Gilles 2001).
The close phylogenetic and morphological con-nection between Xylobolus frustulatus and Acantho-physellum lividocaeruleum that have been demonstrat-ed implies that Acanthophysellum also should be re-garded as a synonym of Xylobolus. However, consid-ering that many species in Aleurodiscus, among themA. apricans, have yet to be studied by molecularmethods we do not want to initiate further taxonom-ical upheaval now.
/chelidonium (87%). This clade includes five mon-omitic species with nodose septate hyphae and longtubular gloeocystidia with an SA1 reaction. Thespores are ellipsoid to cylindrical, finely verrucoseand strongly amyloid. Three species, viz. Gloeocysti-diellum aspellum, G. formosanum and G. compactum,are very closely related and their sequences are al-most identical. Incompatibility between them hasbeen shown (Wu 1996), and they may be regardedas sibling species. Boidinia macrospora is closely relat-ed but can be distinguished by having longer spores.It should be noted that G. aspellum was describedfrom Africa while the culture we have used as a DNAsource originates from Taiwan. We have not com-pared the Taiwanese specimen with the type. Megal-ocystidium chelidonium is a species with similar char-acteristics, except that the spore wall is smooth, atleast as seen in a light microscope.
The relation of /chelidonium to other clades is notclear. Judging from our analyses, the group is fairlywell delimited and appears as a distinct taxon. Nogenus name is available for the group, but we thinkit is premature to introduce a new one at this stage.Similarly we find it unwise to make new combinationsjust to make all clade members belong to the samegenus.
/stereum (88%). This clade is strongly supported asmonophyletic. Species have leathery and often re-
flexed or pileate basidiomata. They have a monomi-tic hyphal system with clamps present only as whorlson certain subicular hyphae. The basidiome containsconducting hyphae that are responsible for the well-known bleeding reaction in certain species. These hy-phae, sometimes called pseudocystidia, are probablyhomologous to the gloeoplerous hyphae in the restof the russuloid lineage. The spores are cylindricalto narrowly ellipsoid, smooth and amyloid. All spe-cies tested have a holocoenocytic nuclear behavior(Boidin and Lanquetin 1984b and references there-in) and are presumably heterothallic bipolar (Boddyand Rayner 1982). Stereum is an old genus name thatearlier was used for all species with leathery, effused-reflexed basidiomata and a smooth hymenophore. Inits restricted sense it appears as a well-defined, natu-ral genus containing many closely related species.
/cerussatus (77%). Aleurodiscus cerussatus, A. dex-trinoideocerussatus and Gloeomyces graminicola clustertogether with good bootstrap support. However, thisis not surprising because the sequences are extremelysimilar, even within the hypervariable regions thatwere discarded before analyses. All three species havesmooth, thin-walled ellipsoid spores with an amyloidreaction and numerous SA1 gloeocystidia with amore or less moniliform outline. Aleurodiscus cerus-satus and A. dextrinoideocerussatus also are similarmorphologically and separated primarily by the dex-trinoid reaction of the acanthophyses in A. dextrino-ideocerussatus. Acanthophyses are sterile terminalcells with numerous protuberances. They appear inmany forms and can be found in most species inAleurodiscus but also in Stereum and Xylobolus. Gloeo-myces graminicola, type of Gloeomyces Sheng H. Wu,lacks acanthophyses and has simple septate hyphae,while the other two species have hyphae that are no-dose septate. Gloeomyces is characterized as dimitic be-cause species have terminal thick-walled andbranched cells embedded in the subhymenium. Theycould be compared to binding hyphae but are fewand do not lend any tough consistency to the basi-dioma. With these characteristics Gloeomyces looksquite different from the other two species in theclade. Our sequence of G. graminicola was generatedfrom cultured mycelium received from the culturecollection at NMNS (Taiwan). We also tried to se-quence DNA from the holotype basidioma but with-out success. Our results should be interpreted withcaution because it cannot be ruled out that the my-celium labeled as Gloeomyces graminicola representssomething else.
/megalocystidium (89%). This clade includes Megal-ocystidium leucoxanthum, M. luridum and a sequenceretrieved from a specimen determined as Gloeocysti-

1061LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
diellum wakullum. A connection between M. luridumand G. wakullum also was reported in the molecularstudy by Boidin et al (1998). Megalocystidium was oneof the earliest segregates from Gloeocystidiellum ( Ju-lich 1978). It originally included the two species se-quenced here and Gloiothele lactescens. Julich selectedM. leucoxanthum as type, but he gave no explanationas to why the genus was erected and how it was sup-posed to be circumscribed. Megalocystidium leucox-anthum and M. luridum have a monomitic hyphalsystem with nodose septate hyphae, SA1 gloeocysti-dia, and smooth, amyloid basidiospores. They are ho-mothallic with a holodikaryotic nuclear behavior(Boidin et al 1997). Gloeocystidiellum wakullum wasdescribed from subtropical areas in the United States(Burdsall et al 1981) and later also reported fromGuadeloupe (Boidin et al 1997a). Our specimenoriginates from Tanzania (Hjortstam and Larsson1995). We have not compared the Tanzanian speci-men with type material, but microscopical details fitthe description. The gloeocystidia originally were re-ported as sulfo-negative. However, Boidin et al(1997a) found, after an examination of the type, thatgloeocystidia are weakly but distinctly sulfo-positiveand we have seen the same reaction in our material.Wu (1996) moved Gloeocystidiellum wakullum to Ster-eum. The evidence he cited were presence of simpleseptate hyphae, smooth amyloid spores, acanthohy-phidia and culture characteristics.
Although Gloeocystidiellum wakullum possessessome features of Megalocystidium (resupinate basi-diomata, large smooth amyloid spores, SA1 gloeo-cystidia), there are some notable differences (acan-thohyphidia, simple septate hyphae). Consideringthe results from the phylogenetic analyses, we stillsuggest that the species should be transferred to Me-galocystidium.
Species on single branches. Aleurodiscus is a large andheterogeneous group of corticioid fungi that areloosely held together by some micromorphologicalcharacters, viz. large amyloid spores, catahymenial or-ganization of basidiomata, and various kinds of sterileorgans in the hymenium. Several new genera havebeen introduced in attempts to separate out morenatural groups (e.g., Lemke 1964, Oberwinkler 1965,Boidin et al 1985). Recently Wu et al (2001) pre-sented a molecular phylogenetic study of Aleurodis-cus, but no taxonomic changes were proposed. Thetype species, A. amorphus, is placed in our tree on asingle branch as a sister taxon to the rest of /stere-ales. This is in agreement with the results presentedby Wu et al (2001). The necessary rearrangements ofspecies in Aleurodiscus sensu lato must await a moredetailed molecular study that also includes a wider
sampling of the genera that we here demonstrate areclosely related.
Aleurobotrys botryosus was segregated from Aleurod-iscus based on the amyloid reaction of the botry-ophyses in combination with SA1 gloeocystidia, or-namented spores and a white-spore print (Boidin etal 1985). The phylogenetic analysis, with the restrict-ed dataset, confirms A. botryosus as distinct fromAleurodiscus, but because both are placed on singlebranches no further conclusions about relationshipscan be drawn. Wu et al (2001) found that Aleurobotrysbotryosus belongs in the vicinity of Aleurodiscus cer-ussatus, and they concluded that the phylogeneticimportance of the amyloid reaction of the botry-ophyses was doubtful.
Species in Wrightoporia share a poroid hymenop-hore, a dimitic context and amyloidity of the sporewall. Two other genera, viz. Amylonotus Ryvarden andAmylosporus Ryvarden, have similar features. Thescopes and limits of these three genera have beendebated (David and Rajchenberg 1985, 1987, Ryvar-den 1991). Three species are included here. Wrigh-toporia lenta (type species) occurs on a single branchbasal to /albatrellus (FIG. 1). Wrightoporia avellaneais included in /scytinostromella, which might be asister clade of /albatrellus, and W. tropicalis occurson a single branch with a basal position in /penio-phorales. For this study, but not included in the finaldata matrix, we also sequenced W. bracei and W. neo-tropica. In our preliminary analyses the former alwaysoccurred on a long, single branch with unclear po-sition. The latter species consistently appeared out-side the russuloid lineage.
Gloeohypochnicium analogum earlier was placed inHypochnicium J. Erikss. because of the thick-walledspores (Eriksson and Ryvarden 1976). Gloeohypochn-icium analogum has weakly ornamented but inamy-loid spores and SA1 gloeocystidia. Basidiomes arestrictly resupinate, and the hyphal system monomiticand nodose-septate. Gloeohypochnicium analogumcauses a white rot in deciduous wood. In our parsi-mony analyses, G. analogum often appears at themost basal node of the ingroup. However, in NJ treesit occurs inside /eurussuloid close to /amylosterea-ceae.
Pseudoxenasma verrucisporum has clear affinities to/russulales and occurs as a sister taxon to that cladein all our phylogenetic trees. However, the connec-tion is not supported by bootstrap. PseudoxenasmaKH. Larss. & Hjortstam was established as a mono-typic genus because it has pleurobasidia (lateral ba-sidia), a feature not known from any other genuswith ornamented amyloid spores and sulfo-positivecystidia. In some species with a dense, more or lessgelatinized hyphal texture, generative hyphae tend to

1062 MYCOLOGIA
grow horizontally in the hymenium and produce ba-sidia laterally. Both gelatinization of the hyphae andlateral production of basidia seem to facilitate quickresponse to favorable conditions for sporulation.Pseudoxenasma grows on dead but still attachedbranches of Picea abies and often is found fertile inmild periods of winter; its globose, coarsely orna-mented, and strongly amyloid spores fit well with thegeneral spore morphology in /russulales.
Scytinostromella nannfeldtii differs in many respectsfrom the type of the genus, and the only characterconnecting S. nannfeldtii to Scytinostromella is skeletalhyphae. In other respects, viz. ornamented amyloidspores and SA1 gloeocystidia, S. nannfeldtii fits manyof the genera in the russuloid lineage.
NEW COMBINATIONS
Boidinia aculeata (Sheng H. Wu) E. Larss. & K.H. Larss.comb. nov. Basionym: Gloeocystidiellum aculeatum ShengH. Wu, Mycotaxon 58: 29, 1996.
Gloeodontia subasperispora (Litsch.) E. Larss. & K.H.Larss. comb. nov. Basionym: Corticium subasperisporumLitsch. (Ann. Mycol. 39: 125, 1941).
Gloeocystidiopsis cryptacanthus (Pat.) E. Larss. & K.H.Larss. comb. nov. Basionym: Corticium cryptacanthumPat., Bull. Soc. Mycol. France 15: 210, 1899.
Megalocystidium wakullum (Burds., Nakasone & G.W. Free-man) E. Larss. & K.H. Larss. comb. nov. Basionym: Gloeo-cystidiellum wakullum Burds., Nakasone & G.W. Freeman,Syst. Bot. 6: 431, 1981.
ACKNOWLEDGMENTS
We are grateful to Drs Jacques Boidin, Sheng-Hua Wu, LeifRyvarden and Jogeir Stokland for loan of collections; to Er-ast Parmasto, Ronald H. Petersen, Lennart Andersson, NilsHallenberg and Roger Eriksson for valuable comments onearlier versions of the manuscript; to Andy Taylor for lin-guistic improvements and to David Hibbett and one anon-ymous reviewer for constructive comments. Financial sup-port for this study was received from the Royal SwedishAcademy of Sciences, Stiftelsen Anna-Greta och Holger Cra-foords fond, Kapten Carl Stenholms donationsfond, Annaoch Gunnar Vidfelts fond for biologisk forskning andKungl. Vetenskaps- och Vitterhets-Samhallet i Goteborg.
LITERATURE CITED
Agerer R, Klostermeyer D, Steglich W, Franz F, Acker G.1996. Ectomycorrizae of Albatrellus ovinus (Scutigera-ceae) on Norway spruce with some remarks on the sys-tematic position of the family. Mycotaxon 59:289–307.
Bernicchia A, Piga A. 1998. A new species of Echinidontiumfrom Italy. Mycotaxon 68:483–491.
Binder M, Bresinsky A. 2002. Deviation of a polymorphiclineage of Gasteromycetes from boletoid ancestors. My-cologia 94:85–98.
Boddy L, Rayner ADM. 1982. Population structure, inter-mycelial interactions and infection biology of Stereumgausapatum. Trans Brit Mycol Soc 78:337–351.
Boidin J. 1958. Essai biotaxonomique sur les Hydnes resu-pines et les Corticies. Rev Mycol Mem Hors-Ser 6.
———. 1990. Repertoire des donnees utiles pour effectuerles tests d’intercompatibilite ches les basidiomycetes VI.Aphyllophorales non porees (Premier supplement).Cryptogamie Mycol 11:175–188.
———, Des Pomeys M. 1961. Heterobasidiomycetes sapro-phytes et Homobasidiomycetes resupines. IX. De lutil-isation des criteres d’interfertilites et de polarite pourla reconnaissance objective des limites specifiques etdes affinites. Bull Soc Mycol France 77:237–261.
———, Gilles G. 2001. Basidiomycetes aphyllophorales del’ıle de la Reunion. XXIII Aleurodiscoideae. Bull SocMycol France 117:173–181.
———, Lanquetin P. 1984a. Le genre Amylostereum (Basid-iomycetes). Intercompatilibites partielles entre especesallopatriques. Bull Soc Mycol France 100:211–236.
———, ———. 1984b. Repertoire des donnees utiles poureffectuer les tests dintercompabilite chez les basidio-mycetes III. Aphyllophorales non porees. CryptogamieMycol 5:193–245.
———, ———. 1987. Le genre Scytinostroma Donk (Basid-iomycetes, Lachnocladiaceae). Bibl Mycol 114:1–130.
———, ———. 1997. Repertoire des donnees utiles poureffectuer les tests d’intercompatibilite ches les basid-iomycetes VII. Aphyllophorales non porees (Deuxiemesupplement). Cryptogamie Mycol 18:9–18.
———, ———, Gilles G. 1997a. Le genre Gloeocystidiellumsensu lato (Basidiomycotina). Bull Soc Mycol France113:1–80.
———, ———, ———. 1997b. Contribution a la connaiss-ance du genre Asterostroma Massee 1889 (Basidiomy-cotina). Bull Soc Mycol France 113:269–301.
———, ———, ———, Candoussau F, Hugueney R. 1985.Contribution a la connaissance des Aleurodiscoideae aspores amyloides (Basidiomycotina, Corticiaceae). BullSoc Mycol France 101:333–367.
———, Mugnier J, Canales R. 1998. Taxonomie moleculai-re des Aphyllophorales. Mycotaxon 66:445–491.
———, Terra P, Lanquetin P. 1968. Contribution a la con-naissance des caracteres myceliens et sexuels desgenres Aleurodiscus, Dendrothele, Laeticorticium et Vuil-leminia (Basidiomycetes Corticiaceae). Bull Soc MycolFrance 84:53–84.
Bruns TD, Szaro TM, Gardes M, Cullings KW, Pan JJ, Hor-ton TR, Kretzer A, Garbelotto M, Li Y. 1998. A se-quence database for identification of ectomycorrhizalbasidiomycetes by phylogenetic analysis. Molec Ecol 7:257–272.
Burdsall HH, Nakasone KK, Freeman GW. 1981. New spe-cies of Gloeocystidiellum (Corticiaceae) from the South-eastern United States. Syst Bot 6:422–434.
Calonge FD, Martin MP. 2000. Morphological and molec-ular data on the taxonomy of Gymnomyces, Martelliaand Zelleromyces (Russulales). Mycotaxon 76:9–15.
Capellano A, Keller J. 1978. Architecture de la paroi spo-

1063LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
rique des Auriscalpiacees et des Hericiacees. Persoonia9:511–514.
Corner EJH. 1932a. The fruit-body of Polystictus xanthopusFr. Ann Bot 46:71–111.
———. 1932b. A Fomus with two systems of hyphae. TransBrit Mycol Soc 17:51–81.
Cunningham GH. 1963. The Thelephoraceae of Australiaand New Zealand. N Z Dep Sci Industr Res Bull 145:1–359.
David A, Rajchenberg M. 1985. Pore fungi from French An-tilles and Guiana. Mycotaxon 22:285–325.
———, ———. 1987. A reevaluation of Wrightoporia andAmylonotus (Aphyllophorales, Polyporaceae). Can J Bot65:202–209.
Donk MA. 1956. Notes on resupinate Hymenomycetes III.Fungus 26:3–24.
———. 1964. A conspectus of the families of Aphyllophor-ales. Persoonia 3:199–324.
———. 1971. Progress in the study of the classification ofthe higher Basidiomycetes. In: Petersen RH, ed. Evo-lution in the higher Basidiomycetes. Knoxville. p 3–25.
Eriksson J, Ryvarden L. 1973. The Corticiacae of North Eu-rope. Vol. 2. Oslo: Fungiflora. p 61–285.
———, ———. 1975. The Corticiacae of North Europe.Vol. 3. Oslo: Fungiflora. p 286–548.
———, ———. 1976. The Corticiacae of North Europe.Vol. 4. Oslo: Fungiflora. p 549–884.
Felsenstein J. 1978. Cases in which parsimony and compat-ibility methods will be positively misleading. Syst Zool27:401–410.
Fries EM. 1874. Hymenomycetes Europaei. Berling, Upp-sala.
Gardes M, Bruns TD. 1993. ITS primers with enhancedspecificity for basidiomycetes-application to the identi-fication of mycorrhizas and rusts. Molec Ecol 2:113–118.
———, ———. 1996. Community structure of ectomycor-rizal fungi in a Pinus muricata forest: above- and below-ground views. Can J Bot 74:1572–1583.
Ginns JH. 1986. The genus Dentipellis (Hericiaceae). Win-dahlia 16:35–45.
———, Freeman GW. 1994. The Gloeocystidiellaceae (Ba-sidiomycota, Hericiales) of North America. Bibl Mycol157:1–118.
———, Lefebvre MNL. 1993. Lignicolous corticioid fungiof North America. Systematics, distribution and ecolo-gy. Mycologia Mem 19:1–247.
Gluchoff-Fiasson K, Kuhner R. 1982. Le principe respons-able du bleuissement au reactif sulfo-vanillique des cys-tides ou laticiferes de divers Homobasidiomycetes: in-teret taxonomique. C R Acad Sc Paris 294:1067–1071.
———, David A, Dequatre B. 1983. Contribution a l’etudedes affinites entre Heterobasidion annosum (Fr.)Bres.et les Bondarzewiaceae. Cryptogamie Mycol 4:135–143.
Gross HL. 1964. The Echinodontiaceae. Mycopath MycolAppl 24:1–26.
Hagstrom E. 1977. Vesiculomyces gen. nov. segregated fromGloeocystidiellum (Corticiaceae). Bot Notiser 130:53–54.
Hallenberg N. 1980. New taxa of Corticiaceae from N. Iran(Basidiomycetes). Mycotaxon 11:447–475.
———. 1985. The Lachnocladiaceae and Coniophoraceaeof North Europe. Oslo: Fungiflora. 96 p.
———, Larsson E, Mahlapuu M. 1996. Phylogenetic studiesin Peniophora. Mycol Res 100:179–187.
———, Parmasto E. 1998. Phylogenetic studies in speciesof Corticiaceae growing on branches. Mycologia 90:640–654.
Hansen L, Knudsen H. 1997. Nordic macromycetes. Vol. 3.Copenhagen, Denmark: Nordsvamp.
Hawksworth DL, Kirk PM, Sutton BC, Pegler DN. 1995. Dic-tionary of the fungi. 8th ed. Wallingford, UK: CAB In-ternational.
Hibbet DS, Binder M. 2002. Evolution of complex fruiting-body morphologies in homobasidiomycetes. Proc R SocLond B 269:1963–1969.
———, Donoghue MJ. 1995. Progress toward a phyloge-netic classification of the Polyporaceae through parsi-mony analyses of mitochondrial ribosomal DNA se-quences. Can J Bot 73:853–861.
———, Gilbert L-B, Donoghue MJ. 2000. Evolutionary in-stability of ectomycorrhizal symbioses in basidiomy-cetes. Nature 407:506–508.
———, Pine EM, Langer E, Langer G, Donoghue MJ. 1997.Evolution of gilled mushrooms and puffballs inferredfrom ribosomal DNA sequences. Proc Nat Acad SciUSA 94:12002–12006.
———, Thorn RG. 2001. Basidiomycota: Homobasidiomy-cetes. In: McLaughlin DJ, McLaughlin, EG, Lemke, PA,eds. The Mycota VII part B, Systematics and evolution.Springer-Verlag. Heidelberg, Berlin. p 121–168.
Hjortstam K. 1987a. A check-list to genera and species ofcorticioid fungi (Hymenomycetes). Windahlia 17:56–85.
———. 1987b. Studies in tropical Corticiaceae (Basidio-mycetes) VII. Mycotaxon 28:19–37.
———. 1998. A check-list to genera and species of corti-cioid fungi (Basidiomycotina, Aphyllophorales). Win-dahlia 23:1–54.
———, Larsson K-H. 1995. Annotated check-list to generaand species of corticioid fungi (Aphyllophorales, Basid-iomycotina) with special regards to tropical and sub-tropical areas. Windahlia 21:1–75.
———, Ryvarden L. 1988. Notes on the Corticiacae ofnorthern China. Acta Mycol Sinica 7(2):77–88.
———, Stalpers J. 1982. Notes on Corticiaceae (Basidio-mycetes) XI. Boidinia, a new genus segregated fromGloeocystidiellum. Mycotaxon 14:75–81.
Holmgren PK, Holmgren NK, Barnett LC. 1990. Index her-bariorum. Part I: the herbaria of the world. New York:New York Botanical Garden. 693 p.
Hopple JS Jr, Vilgalys R. 1999. Phylogenetic relationships inthe mushroom genus Coprinus and dark-spored alliesbased on sequence data from the nuclear gene codingfor the large ribosomal subunit RNA: divergent do-mains, outgroups and monophyly. Mol Phyl Evol 13:1–19.
Julich W. 1972. Monographie der Atheliae (Corticiaceae,Basidiomycetes). Willdenowia, Beih. 7:1–283.
———. 1978. Studies in resupinate basidiomycetes V. Per-soonia 10:137–140.

1064 MYCOLOGIA
———. 1981. Higher taxa of Basidiomycetes. Bibl Mycol 85:1–485.
———. 1982. Studies in resupinate basidiomycetes VII. IntJ Myc Lich 1:27–37.
Kawagishi H, Ando M, Shinba K, Sakamoto H, Yoshida S,Ojima F, Ishiguroy, Ukai N, Furukawa S. 1993. Chro-mans, hericenones F, G and H from the mushroomHericium erinaceum. Phytochemistry 32(1):175–178.
———, Shimada A, Hosokawa S, Mori H, Sakamoto H, Ish-iguro Y, Sakemi S, Bordner J, Kojima N, Furukawa S.1996. Erinacines E, F, and G, stimulators of nervegrowth factor (NGF)-synthesis, from the mycelia of Her-icium erinaceum. Tetrahedron Letters 37(41):7399–7402.
Keller G. 1986. Ultrastructure des parois sporiques de qu-elques Aphyllophorales. Mycologia Helvetica 2:1–34.
———. 1997. Atlas des Basidiomycetes vus aux microscopeselectroniques. Union des Societes Suisses de Mycolo-gie, Neuchatel.
Koski-Kotiranta S, Niemela T. 1988. Hydnaceous fungi ofthe Hericiaceae, Auriscalpiaceae and Climacodonta-ceae in northwestern Europe. Karstenia 27:43–70.
Larsson E, Hallenberg N. 2001. Species delimitation in theGloeocystidiellum porosum-clavuligerum complex in-ferred from compatibility studies and nuclear rDNA se-quence data. Mycologia 93:907–914.
Larsson K-H. 2001. The position of Poria mucida inferredfrom nuclear ribosomal DNA sequences. Harvard Pa-pers in Botany 6:131–138.
Lemke PA. 1964. The genus Aleurodiscus (sensu stricto) inNorth America. Can J Bot 42:213–282.
Lickey E. 2002. Systematic and monographic studies in Ar-tomyces Julich and Clavicorona Doty [Thesis]. Knoxville:Univ. Tennessee.
Maas Geesteranus RA. 1963. Hyphal structures in Hydnums.II. Proc Kon Ned Akad Wetensch Series C, 66 part 5:426–430.
Maddison WP, Maddison DR. 1992. MacClade, version 3.Analysis of phylogeny and character evolution. Sunder-land, Massachusetts: Sinauer Associates Inc.
Maekawa N, Arita I, Hayashi Y. 1982. Corticiaceae in Japan.Rep Tottori Myc Inst 20:33–41.
Miller OK, Stewart L. 1971. The genus Lentinellus. Mycol-ogia 63:333–369.
Miller SL, McClean TM, Walker JF, Buyck B. 2001. A mo-lecular phylogeny of the Russulales including agari-coid, gasteroid and pleurotoid taxa. Mycologia 93:344–354.
Moncalvo J-M, Vilgalys R, Readhead SA, Johnson JE, JamesTY, Aime MC, Hofstetter V, Verduin SJW, Larsson E,Baroni TJ, Thorn RG, Jacobsson S, Clemencon H, Mill-er Jr OK. 2002. One hundred and seventeen clades ofeuagarics. Mol Phyl Evol 23:357–400.
Moser M. 1978. Kleine Kryptogamenflora Bd. IIb/2. DieRohrlinge und Blatterpilze. Stuttgart, New York: GustavFischer Verlag.
Nakasone KK. 1982. Cultural and morphological studies ofGloeocystidiellum porosum and Gloeocystidiellum clavuli-gerum. Mycotaxon 14:316–324.
———. 1990. Cultural studies and identification of wood-
inhabiting Corticiaceae and selected Hymenomycetesfrom North America. Mycologia Mem 15:1–412.
Nunez M, Ryvarden L. 1997. The genus Aleurodiscus (Ba-sidiomycotina). Oslo: Fungiflora.
Oberwinkler F. 1965. Primitive Basidiomyceten. Revision ei-ner Formenkreis von Basidienpilze mit plastischer Bas-idie. Sydowia, Ann Mycol Ser II 19:1–77.
———. 1977. Das neue System der Basidiomyceten. In: FreyW, Hurka H, Oberwinkler F, eds. Beitrage zur Biologieder niederen Pflanzen. Stuttgart, New York: Gustav Fi-scher Verlag. p 59–104.
Parmasto E. 1967. Corticiaceae U. R. S. S. IV. Descriptionestaxorum novorum. Combinationes novae. Eesti NSVTead Akad Toimet Biol 16:377–394.
———. 1968. Conspectus Systematis Corticiacearum. Insti-tutum zoologicum et botanicum Academiae Scientia-rum R.P.S.S. Estonicae.
Patouillard N. 1900. Essai taxonomique sur les familles etles genres des hymenomycetes. Lons-le-Saunier,France.
Pegler DN. 1996. Hyphal analysis of basidiomata. Mycol Res100:129–142.
Pilat A. 1926. Monographie der mitteleuropaischen Aleu-rodiscineen. Ann Mycol 24:203–230.
Printz P. 1986. Savbladhatte slaegten Lentinellus i Danmark.Svampe 14:59–63.
Rattan SS. 1977. The resupinate Aphyllophorales of theNorth Western Himalayas. Bibl Mycol 60:1–427.
Redhead SA, Norvell LL. 1993. Notes on Bondarzewia, Het-erobasidion and Pleurogala. Mycotaxon 48:371–380.
———, Vilgalys R, Moncalvo J-M, Johnson J, Hopple JS.2001. Coprinus Pers. and the disposition of Coprinusspecies sensu lato. Taxon 50:203–241.
Reid DA. 1965. A monograph of the stipitate stereoid fungi.Nova Hedwigia, Beih. 18:1–382.
Rick J. 1934. Monographia Thelephoracearum resupinatar-um Riograndensium. Broteria 3:31–48.
Ryman S, Holmasen I. 1984. Svampar. Stockholm: Interpub-lishing AB.
Ryvarden L. 1991. Genera of polypores. Nomenclature andtaxonomy. Synopsis Fungorum 5. Oslo: Fungiflora.
———, Gilbertson RL. 1993. European polypores. Part 1Abortiporus—Lindtneria. Oslo: Fungiflora.
Savolainen V, Cuenoud P, Spichiger R, Martinez MDP,Crevecoeur M, Manen JF. 1995. The use of herbariumspecimens in DNA phylogenetics: evaluation and im-provment. Pl Syst Evol 197:87–98.
Smith SE, Read DJ. 1997. Mycorrhizal symbiosis. San Diego:Academic Press.
Stalpers JA. 1979. Heterobasidion (Fomes) annosum and theBondarzewiaceae. Taxon 28:414–417.
———. 1996. The aphyllophoraceous fungi II. Keys to thespecies of the Hericiales. Stud Mycol 40.
Stenlid J. 1986. Biochemical and ecological aspects of theinfection biology of Heterobasidion annosum. Swedishuniversity of agricultural siences, Uppsala.
Swofford DL. 1999. PAUP*: phylogenetic analysis using par-simony (* and other methods) version 4.0. Sunderland,Massachusetts: Sinauer Associates Inc.
Tellerıa MT. 1990. Annotated list of the Corticiaceae sensu

1065LARSSON AND LARSSON: PHYLOGENY OF RUSSULOID BASIDIOMYCETES
lato (Aphyllophorales, Basidiomycotina), for Peninsu-lar Spain and Balearic Islands. Bibl Mycol 135:1–152.
Thomas GP. 1958. The occurrence of the Indian paint fun-gus, Echinodontium tinctorium E. & E., in British Co-lumbia. Studies in Forest Pathology 18:3–28.
Vilgalys R, Hester M. 1990. Rapid genetic identification andmapping of enzymatically amplified ribosomal DNAfrom several Cryptococcus species. J Bacteriol 172:4238–4246.
Watling R, Gregory NM. 1989. Crepidotaceae, Pleurotaceaeand other pleurotoid agarics. Royal Botanic Garden,Edinburgh: British Fungus Flora 6.
White TJ, Bruns T, Lee S, Taylor JW. 1990. Amplificationand direct sequencing of fungal ribosomal RNA genes
for phylogenetics. In: Innis MA, Gelfand DH, SininskyJJ, White TJ, eds. PCR protocols: a guide to methodsand applications. New York: Academic Press Inc. p 315–322.
Wu SH. 1995. Two new genera of corticioid basidiomyceteswith gloeocystidia and amyloid basidiospores. Mycolo-gia 87:886–890.
———. 1996. Studies on Gloeocystidiellum sensu lato (Basid-iomycotina) in Taiwan. Mycotaxon 58:1–68.
———, Buchanan PK. 1998. Species of Boidinia (Basidio-mycotina) with simple septate hyphae. Mycotaxon 67:123–128.
———, Hibbett DS, Binder M. 2001. Phylogenetic analysesof Aleurodiscus s. l. and allied genera. Mycologia 93:720–731.




















