
Comparison between the macroinfauna of urbanized and protected beaches in Rio de Janeiro State,...
6
Short communication Comparison between the macroinfauna of urbanized and protected beaches in Rio de Janeiro State, Brazil Vale ´ria G. Veloso*, Elen S. Silva, Carlos H.S. Caetano, Ricardo S. Cardoso Departamento de Ciencias Naturais, Universidade Federal do Estado do Rio de Janeiro (UNIRIO), Av. Pasteur, 458, Sala 411, Urca, Rio de Janeiro, RJ, CEP 22290-240, Brazil ARTICLE INFO Article history: Received 9 July 2004 Received in revised form 16 September 2005 Accepted 20 September 2005 Available online 17 November 2005 Keywords: Sandy beaches Macroinfauna Trampling Human impact ABSTRACT The intertidal macroinfauna of five sandy beaches with similar morphodynamics condi- tions was studied to compare composition and structure between beaches near urbanized centers and protected beaches located in Rio de Janeiro State. The beaches were sampled in winter 1996 and summer 1997 according to a systematic design with stratification. A total of nine species were identified with the crustaceans being the most abundant and frequent animals. Species richness showed little variation between beaches while density of some species such as Emerita brasiliensis (Crustacea) and Phaleria testacea (Insecta) were lower at urbanized beaches when compared to protected ones. At the most urbanized beaches, Pseudorchestoidea brasiliensis was absent during both sampling periods. The neg- ative relationships between human recreational activities (e.g., trampling) and density of macroinfaunal species was recently gained support and in Barra da Tijuca beach these relation is very clear. In the developed and most-visited sector, Barra (Alvorada), the amphipod Pseudorchestoidea brasiliensis was never collected whereas in the protected por- tion, Barra (Reserva), the same species occurred in high densities. Thus, our results sug- gested that the amphipod Pseudorchestoidea brasiliensis is more vulnerable to trampling than are other species. Ó 2005 Elsevier Ltd. All rights reserved. 1. Introduction In recent decades, studies of beaches have focused on under- standing the influence of physical factors on communities (McLachlan, 1983, 1990; Defeo et al., 1992; McArdle and McLachlan, 1992; Jaramillo et al., 1993; Borzone et al., 1996; Barros et al., 2001) and populations (Dugan et al., 1994; Go ´ mez and Defeo, 1999; Defeo et al., 2001; Cardoso et al., 2003). For most species, seasonal changes in population density are nor- mal, and are related to many factors such as reproductive dynamics and changes in abiotic factors (Souza and Gianuca, 1995; Cardoso and Veloso, 1996, 2003; Veloso and Cardoso, 1999; Fonseca et al., 2000). According to the swash exclusion hypothesis (SEH), swash condition is the main factor control- ling the intertidal macrofauna communities (McLachlan et al., 1993, 1995). This hypothesis states that species diversity, total abundance, biomass increase from reflective towards dissipa- tive beaches and was corroborated by many subsequent works (Defeo et al., 1992; McLachlan et al., 1993; Borzone et al., 1996; Veloso et al., 2003). Despite the increase in rich- ness, diversity and biomass towards dissipative beaches, as suggested by the swash exclusion hypothesis, many species 0006-3207/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocon.2005.09.027 * Corresponding author: Tel.: +55 21 5877809. E-mail address: [email protected] (C.H.S. Caetano). BIOLOGICAL CONSERVATION 127 (2006) 510 – 515 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/biocon
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Comparison between the macroinfauna of urbanized and protected beaches in Rio de Janeiro State,...

B I O L O G I C A L C O N S E R VAT I O N 1 2 7 ( 2 0 0 6 ) 5 1 0 –5 1 5
. sc iencedi rec t .com
ava i lab le at wwwjournal homepage: www.elsevier .com/ locate /b iocon
Short communication
Comparison between the macroinfauna of urbanizedand protected beaches in Rio de Janeiro State, Brazil
Valeria G. Veloso*, Elen S. Silva, Carlos H.S. Caetano, Ricardo S. Cardoso
Departamento de Ciencias Naturais, Universidade Federal do Estado do Rio de Janeiro (UNIRIO), Av. Pasteur, 458, Sala 411, Urca,
Rio de Janeiro, RJ, CEP 22290-240, Brazil
A R T I C L E I N F O
Article history:
Received 9 July 2004
Received in revised form 16
September 2005
Accepted 20 September 2005
Available online 17 November 2005
Keywords:
Sandy beaches
Macroinfauna
Trampling
Human impact
0006-3207/$ - see front matter � 2005 Elsevidoi:10.1016/j.biocon.2005.09.027
* Corresponding author: Tel.: +55 21 5877809.E-mail address: [email protected].
A B S T R A C T
The intertidal macroinfauna of five sandy beaches with similar morphodynamics condi-
tions was studied to compare composition and structure between beaches near urbanized
centers and protected beaches located in Rio de Janeiro State. The beaches were sampled
in winter 1996 and summer 1997 according to a systematic design with stratification. A
total of nine species were identified with the crustaceans being the most abundant and
frequent animals. Species richness showed little variation between beaches while density
of some species such as Emerita brasiliensis (Crustacea) and Phaleria testacea (Insecta) were
lower at urbanized beaches when compared to protected ones. At the most urbanized
beaches, Pseudorchestoidea brasiliensis was absent during both sampling periods. The neg-
ative relationships between human recreational activities (e.g., trampling) and density of
macroinfaunal species was recently gained support and in Barra da Tijuca beach these
relation is very clear. In the developed and most-visited sector, Barra (Alvorada), the
amphipod Pseudorchestoidea brasiliensis was never collected whereas in the protected por-
tion, Barra (Reserva), the same species occurred in high densities. Thus, our results sug-
gested that the amphipod Pseudorchestoidea brasiliensis is more vulnerable to trampling
than are other species.
� 2005 Elsevier Ltd. All rights reserved.
1. Introduction
In recent decades, studies of beaches have focused on under-
standing the influence of physical factors on communities
(McLachlan, 1983, 1990; Defeo et al., 1992; McArdle and
McLachlan, 1992; Jaramillo et al., 1993; Borzone et al., 1996;
Barros et al., 2001) and populations (Dugan et al., 1994; Gomez
and Defeo, 1999; Defeo et al., 2001; Cardoso et al., 2003). For
most species, seasonal changes in population density are nor-
mal, and are related to many factors such as reproductive
dynamics and changes in abiotic factors (Souza and Gianuca,
er Ltd. All rights reservedbr (C.H.S. Caetano).
1995; Cardoso and Veloso, 1996, 2003; Veloso and Cardoso,
1999; Fonseca et al., 2000). According to the swash exclusion
hypothesis (SEH), swash condition is the main factor control-
ling the intertidal macrofauna communities (McLachlan et al.,
1993, 1995). This hypothesis states that species diversity, total
abundance, biomass increase from reflective towards dissipa-
tive beaches and was corroborated by many subsequent
works (Defeo et al., 1992; McLachlan et al., 1993; Borzone
et al., 1996; Veloso et al., 2003). Despite the increase in rich-
ness, diversity and biomass towards dissipative beaches, as
suggested by the swash exclusion hypothesis, many species
.

B I O L O G I C A L C O N S E R VAT I O N 1 2 7 ( 2 0 0 6 ) 5 1 0 –5 1 5 511
of sandy beaches can tolerate a wide spectrum of morphody-
namic variation, maintaining abundant populations in both
reflective and dissipative beaches.
Hydrodynamics and sediment remobilization continu-
ously alter beach profiles. Because their communities are
well adapted to these dynamics, the beaches do not suffer
significantly when small changes in the beach profiles occur.
Therefore, these ecosystems are not very vulnerable either
to natural changes or to human activities (Jaramillo et al.,
1996).
Pollution, exploitation of natural resources, and mainly
erosion are the most common problems in beach ecosys-
tems (McIntyre, 1995). Although urbanization and tourism
are increasing worldwide, studies on modifications caused
by landfills, recreation and cleaning are still rare. The dam-
age caused by large numbers of people trampling dune veg-
etation is better documented (Hosier and Eaton, 1980;
Brown and McLachlan, 1990; Rickard et al., 1994; Watson
et al., 1996). The first investigations of the effects of recre-
ation and trampling on the intertidal macroinfauna gave
conflicting results. Jaramillo et al. (1996), comparing a free
public access trampled area to a restricted public area on
the Chilean coast over a two-month period, found no influ-
ence of trampling. Contrariwise, trampling has been indi-
cated as the factor responsible for the disappearance of
amphipods from highly frequented beaches in Poland
(Welawski et al., 2000). Experiments done in South Africa
indicate that some species resist the impacts of trampling
better than others (Moffet et al., 1998). Barros (2001) com-
pared the numbers of burrows of ghost crabs, Ocypode cordi-
mana, between urban and non- urban beaches and verifies
that urban beaches presents numbers significantly lower
of burrows than non-urban ones at high shore levels. The
lack of information regarding the long-term effects of tram-
pling on the macroinfauna and the conflicting results so far
have revealed the importance of this subject and have
hinted that the consequences of trampling might be
irreversible.
The present study analyses the changes in the composi-
tion and structure of the macroinfauna in crowded urbanized
beaches in Rio de Janeiro City, Brazil. Comparisons between
beaches near urbanized centers and protected beaches were
made, and potential influence of human recreational activi-
ties such trampling was evaluated.
2. Study area
Five beaches located in the state of Rio de Janeiro, Brazil, were
selected for this study: Copacabana, Ipanema, Sao Conrado,
Barra da Tijuca and Grumari. Barra da Tijuca Beach presents
an area of high visitation frequency and a protected area
(environmental preservation area), respectively, named Barra
(Alvorada) and Barra (Reserva), that was independently
treated.
2.1. Copacabana and Ipanema
These beaches border crowded neighborhoods near the city
center of Rio de Janeiro, where many restaurants, bars and
hotels are located. In these areas, efficient public transporta-
tion provides easy access for visitors from the suburbs; there
are showers and toilets, and many peddlers. The beaches are
frequented year-round. The illumination enables people to go
to those beaches even at night, when they can practice foot-
ball or volleyball.
2.2. Barra (Alvorada) and Sao Conrado
Barra da Tjuca Beach is 18 km long. The urbanized first
6-km stretch is interrupted by a protected area (about
8 km), and the last 4 km are also urbanized. Barra (Alvorada)
constitute the first urbanized portion and together with Sao
Conrado beach are located farther from the urban center,
and have similar facilities to Copacabana and Ipanema.
Although ample infrastructure is provided, Barra (Alvorada)
and Sao Conrado beaches are less crowded in winter, espe-
cially this last beach which is a famous landing site for hang
gliders.
2.3. Barra (Reserva) and Grumari
Barra (Reserva) Beach is located on the same beach arc as Bar-
ra (Alvorada) Beach, but because is part of a protected area,
any kind of construction is forbidden. Also, there is no public
transportation to these beaches. Grumari Beach is 4 km long
and is an environmental preservation area.
3. Materials and methods
3.1. Sampling and laboratory procedures
The beaches were sampled in winter 1996 and summer
1997, during low spring tide. At each beach, two sectors
were established from the lower limit of the swash zone
to above the drift line. Next, five transects were marked
in each sector and 10 equally spaced sampling strata were
marked per transect, according to a systematic design with
stratification, defined: the first below the swash line (N1 –
30 cm water layer), the second last (N9) on the drift line,
and the last (N10) 10 m above the drift line. This systematic
design is necessary to describe the macroinfauna communi-
ties because of conspicuous and intensively recorded
biological zonation (Defeo et al., 1992; Jaramillo et al.,
1993; Borzone et al., 1996; James and Fairweather, 1996).
One sample was taken with a 0.04 m2 quadrat to a depth
of 25 cm from each sampling strata. The collected sediment
was washed through a 0.71 mm sieve, and the retained
material was taken to the laboratory, where the organisms
were sorted by species, counted and fixed in 5%
formaldehyde.
3.2. Physical characterization
Sediment samples for particle size analysis were taken with a
plastic corer of 3.5 cm diameter to a depth of 10 cm at strata
10 (supra), 6 (middle) and 2 (waterline). Samples were oven-
dried at 70 �C and passed through a series of sieves of �2.5,
�2.0, �1.0, 0.0, 0.5, 1.0, 1.5, 2.0, 2.5, 3.0 and 4.0 phi (phi = �log2mm) in order to determine mean particle size (Folk and Ward,
1957). Results were expressed in mm.

512 B I O L O G I C A L C O N S E R VAT I O N 1 2 7 ( 2 0 0 6 ) 5 1 0 –5 1 5
The beach-face slope of each transect was measured by
the height difference (Emery, 1961) between the drift line
and the waterline. Dean�s dimensionless parameter (X) (Short
andWright, 1983) was calculated for each beach as a measure
of its morphodynamic state: X = Hb/Ws. T, where Hb is the
breaker height in cm, Ws the sand settling velocity in cm s�1
(obtained from particle size and Gibbs et al., 1971) and T the
wave period in seconds.
3.3. Statistical analysis
The number of people passing the drift line was counted
every half hour (between 10.00 and 15.00) in 6 h of the peak
of abundance, to verify the trampling intensity. The number
of people was counted in a randomly chosen block of 50 m2.
ANOVA was employed to verify the trampling difference
(mean abundance related to the 6 h peak) between the
beaches.
The two-way ANOVAwas used to verify differences in the
macroinfauna density between beaches and seasons. The Tu-
key–Kramermultiple comparison test was used a posteriori to
assess significant differences. Density values were log(x + 1)
transformed to fulfill the assumptions of normality and hom-
ocedasticity of the data.
4. Results
4.1. Physical characterization
The morphodynamic states of the beaches of Ipanema, Sao
Conrado and Grumari were reflective while Barra (Alvorada)
and Barra (Reserva) were intermediate (Table 1). At Copaca-
bana beach, the morphodynamic state varied according to
the season, being intermediate in winter and reflective in
summer. The beaches classified as intermediate showed val-
ues near the reflective limits (X < 1). During this study, the
granulometry of most beaches varied from 0.37 to 0.50 (med-
ium sand) except at Ipanema beach, in winter, that presented
mean grain size of 0.68 mm (coarse sand).
Table 1 – Characterization of the beaches studied: w = winter;
Beaches Beachextension
(km)
Dean�sparameter
(X)
Intertidalslope (1/m)
M
Copacabana w 6 1.68 23.56
s 0.53 13.79
Ipanema w 5 0.63 9.44
s 0.82 12.69
Sao Conrado w 4 0.84 25.00
s 0.42 13.72
Barra (Alvorada) w 10 2.35 12.90
s 1.36 11.61
Barra (Reserva) w 8 1.15 12.45
s 1.72 11.20
Grumari w 4 0.77 11.38
s 0.72 11.86
Mean visitors = mean number of people in 6 h of the peak of abundance
a Frequency of visitors: 1 = high; 2 = medium; 3 = low.
The beaches of Copacabana and Ipanema are located in
more densely inhabited areas (Table 1) and received the high-
est mean number of visitors. These beaches did not differ sig-
nificantly from the others in regard to the number of visitors,
either in summer or winter (Table 2). The stretches of Barra
(Reserva) and Grumari beaches located within the protected
area were least trampled. Only during summer there was a
significant difference between the number of visitors at Gru-
mari and Barra (Reserva), as indicated by ANOVA. In spite of
being a protected area, Grumari receives more visitors during
summer because of the easy access to facilities and infra-
structure. Although Sao Conrado and Barra (Alvorada) bea-
ches are near a neighborhood area, they have fewer visitors,
mainly during winter, than Copacabana and Ipanema (Table
2).
4.2. Macroinfauna
A total of nine taxa were collected and identified during this
study. Crustaceans were the most abundant and frequent ani-
mals, represented by Emerita brasiliensis, Lepidopa richmondi
(Decapoda), Pseudorchestoidea brasiliensis (Amphipoda) and
Excirolana braziliensis (Isopoda). Molluscs were represented
by the bivalve Donax hanleyanus; and polychaetes by Hemipo-
dus olivieri, Pisionidens indica and unidentified species. At the
most urbanized beaches, Pseudorchestoidea brasiliensis was ab-
sent both in winter and summer. Despite their frequency,
Emerita brasiliensis and Phaleria testacea were present in lower
densities at urbanized beaches when compared to Grumari
and Barra (Reserva). Only one species, Excirolana brasiliensis,
was found at Copacabana, and was absent from the other
urbanized beaches except at Barra (Alvorada) Beach where it
was present in high densities during winter. Donax hanleyanus
and Hemipodus olivieri occurred in almost all the beaches, but
in low densities (Table 3).
The comparison of macroinfauna density between the
beaches and seasons revealed significant differences only be-
tween the beaches (Table 4). The Tukey–Kramer multiple
comparison test showed that more urbanized beaches
s = summer
ean grainsize (mm)
Populationnumber
(per district)
Visitorsabundance(means ± SD)
Frequency ofvisitorsa
0.45 >150,000,000 166.66 ± 20.73 1
0.40 323.50 ± 53.87
0.68 90–100,000,000 154.66 ± 16.68 1
0.45 333.50 ± 44.64
0.41 20–30,000,000 94.33 ± 13.29 1
0.38 203.16 ± 14.03
0.48 120–130,000,000 128.83 ± 20.12 1
0.41 251.33 ± 17.46
0.50 0 6.00 ± 4.00 3
0.43 31.00 ± 10.60
0.37 0 10.50 ± 4.50 2
0.42 93.33 ± 17.96
(persons/50 m2).
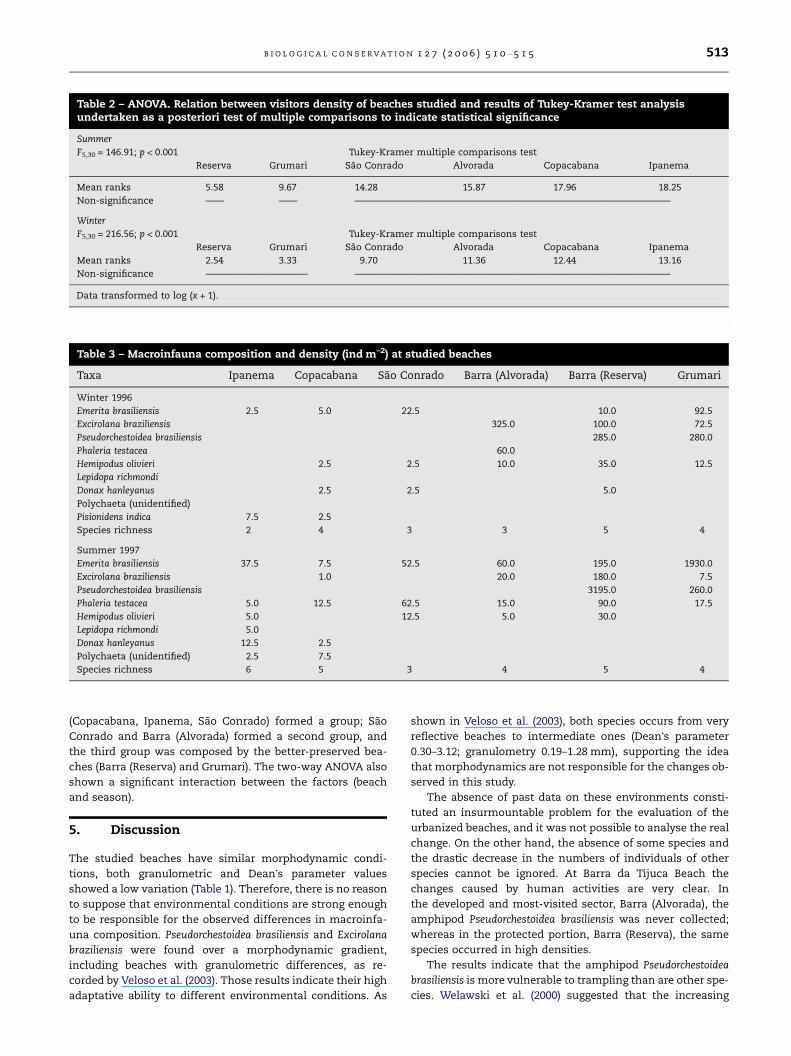
Table 2 – ANOVA. Relation between visitors density of beaches studied and results of Tukey-Kramer test analysisundertaken as a posteriori test of multiple comparisons to indicate statistical significance
Summer
F5,30 = 146.91; p < 0.001 Tukey-Kramer multiple comparisons test
Reserva Grumari Sao Conrado Alvorada Copacabana Ipanema
Mean ranks 5.58 9.67 14.28 15.87 17.96 18.25
Non-significance —— —— ——————————————————————————————————
Winter
F5,30 = 216.56; p < 0.001 Tukey-Kramer multiple comparisons test
Reserva Grumari Sao Conrado Alvorada Copacabana Ipanema
Mean ranks 2.54 3.33 9.70 11.36 12.44 13.16
Non-significance ——————————— ——————————————————————————————————
Data transformed to log (x + 1).
Table 3 – Macroinfauna composition and density (ind m–2) at studied beaches
Taxa Ipanema Copacabana Sao Conrado Barra (Alvorada) Barra (Reserva) Grumari
Winter 1996
Emerita brasiliensis 2.5 5.0 22.5 10.0 92.5
Excirolana braziliensis 325.0 100.0 72.5
Pseudorchestoidea brasiliensis 285.0 280.0
Phaleria testacea 60.0
Hemipodus olivieri 2.5 2.5 10.0 35.0 12.5
Lepidopa richmondi
Donax hanleyanus 2.5 2.5 5.0
Polychaeta (unidentified)
Pisionidens indica 7.5 2.5
Species richness 2 4 3 3 5 4
Summer 1997
Emerita brasiliensis 37.5 7.5 52.5 60.0 195.0 1930.0
Excirolana braziliensis 1.0 20.0 180.0 7.5
Pseudorchestoidea brasiliensis 3195.0 260.0
Phaleria testacea 5.0 12.5 62.5 15.0 90.0 17.5
Hemipodus olivieri 5.0 12.5 5.0 30.0
Lepidopa richmondi 5.0
Donax hanleyanus 12.5 2.5
Polychaeta (unidentified) 2.5 7.5
Species richness 6 5 3 4 5 4
B I O L O G I C A L C O N S E R VAT I O N 1 2 7 ( 2 0 0 6 ) 5 1 0 –5 1 5 513
(Copacabana, Ipanema, Sao Conrado) formed a group; Sao
Conrado and Barra (Alvorada) formed a second group, and
the third group was composed by the better-preserved bea-
ches (Barra (Reserva) and Grumari). The two-way ANOVA also
shown a significant interaction between the factors (beach
and season).
5. Discussion
The studied beaches have similar morphodynamic condi-
tions, both granulometric and Dean�s parameter values
showed a low variation (Table 1). Therefore, there is no reason
to suppose that environmental conditions are strong enough
to be responsible for the observed differences in macroinfa-
una composition. Pseudorchestoidea brasiliensis and Excirolana
braziliensis were found over a morphodynamic gradient,
including beaches with granulometric differences, as re-
corded by Veloso et al. (2003). Those results indicate their high
adaptative ability to different environmental conditions. As
shown in Veloso et al. (2003), both species occurs from very
reflective beaches to intermediate ones (Dean�s parameter
0.30–3.12; granulometry 0.19–1.28 mm), supporting the idea
that morphodynamics are not responsible for the changes ob-
served in this study.
The absence of past data on these environments consti-
tuted an insurmountable problem for the evaluation of the
urbanized beaches, and it was not possible to analyse the real
change. On the other hand, the absence of some species and
the drastic decrease in the numbers of individuals of other
species cannot be ignored. At Barra da Tijuca Beach the
changes caused by human activities are very clear. In
the developed and most-visited sector, Barra (Alvorada), the
amphipod Pseudorchestoidea brasiliensis was never collected;
whereas in the protected portion, Barra (Reserva), the same
species occurred in high densities.
The results indicate that the amphipod Pseudorchestoidea
brasiliensis is more vulnerable to trampling than are other spe-
cies. Welawski et al. (2000) suggested that the increasing

Table 4 – Two-way ANOVA. Comparison of macroinfauna density between the beaches and seasons (winter and summer)
Effect df Effect MS Effect df Error MS Error F p
Beach 5 6.56 108 0.33 19.70 0.0000
Season 1 1.30 108 0.33 3.92 0.5033
Interaction 5 0.92 108 0.33 2.77 0.0214
Tukey-Kramer multiple comparisons test
Copacabana Ipanema Sao Conrado Alvorada Grumari Reserva
Mean 0.29 0.41 0.58 0.95 1.51 1.64
Non-significance ———————————————————————————
———————————————
—————————————-
Results of Tukey-Kramer test analysis undertaken as a posteriori test of multiple comparisons to indicate statistical significance are also
showed. Data transformed to log (x + 1).
514 B I O L O G I C A L C O N S E R VAT I O N 1 2 7 ( 2 0 0 6 ) 5 1 0 –5 1 5
tourism on the Polish Baltic coast is one of the causes of the
decrease in the population density of the amphipod Talitrus
saltator. Historical data show a decrease in the occurrence of
this amphipod from 1965 to 1995. The areas where tourism
had most increased showed the greatest declines of this spe-
cies. Along 60% of the coast, the authors recorded 100 people
daily trampling a 1-m area. The method of cleaning was also
indicated as an important limiting factor for the occurrence of
this amphipod. The beaches of Copacabana and Ipanema are
visited year-round. During summer, they are overcrowded
and the space between people is minimal.
The swash zone filter-feeders, such as species of the genus
Emerita and Donax, are able to move to the infralittoral zone,
and their larvae have better power of dispersal and a good
chance to recolonize disturbed areas. For those reasons, they
should be less vulnerable to trampling. Our data showed that
these species occur in all the beaches, although in lower den-
sity in the more urbanized than in the most protected areas.
On the other hand, in most midlittoral species of peracarids
with direct development (such the amphipod Pseudorchestoi-
dea brasiliensis and isopod Excirolana braziliensis), the juveniles
are recruited into the same environment as the adult popula-
tion. Other differences includes: a thinner ‘‘carapace’’ than
Emerita and Donax ; and localization in the most-trampled
central midlittoral zone. Different susceptibility was observed
by Moffet et al. (1998), who attempted experimentally to verify
the effects of different intensities of recreational activity on
the survival rate of four species of the macroinfauna. The re-
sults showed a small change in abundance when trampling
occurred at low intensity. However, there was substantial
damage when trampling increased. The most affected species
were those with the more fragile carapaces, such as mysida-
ceans and juvenile bivalves. Contradictory results were ob-
tained by Jaramillo et al. (1996), who observed no significant
changes between a recreational area and an area restricted
to visitors over a two-month period.
Although several experimental studies have attempted to
show lethal effects of trampling and/or recreational use of
the beaches, the chronic effects on aspects of life cycles such
as fecundity rate, recruitment, growth rate, or the length of
ovigerous females are still unknown. The danger that certain
species may be eliminated from these ecosystems is far from
being understood, considering the sparse knowledge about
energy flow within and between the adjacent ecosystems.
Specific studies should be developed to evaluate the composi-
tion and structure of the macroinfauna related to intense hu-
man activity. Such studies will form a fundamental basis for
developing management and preservation policies.
Acknowledgment
We thank all participants in the field work for their valuable
effort. Thanks also to Dr. Janet W. Reid for English language
revision. This study was supported by CNPq (Conselho Nac-
ional de Desenvolvimento Cientıfico e Tecnologico) and FA-
PERJ (Fundacao de Amparo a Pesquisa do Estado do Rio de
Janeiro).
R E F E R E N C E S
Barros, F., 2001. Ghost crabs as a tool for rapid assessment ofhuman impacts on exposed sandy beaches. BiologicalConservation 97, 399–404.
Barros, F., Borzone, C.A., Rosso, S., 2001. Macroinfauna ofsix beaches near Guaratuba bay, southern Brazil.Brazilian Archives of Biology and Technology 44 (4),351–364.
Borzone, C.A., Souza, J.R.B., Soares, A.G., 1996. Morphodynamicinfluence on the structure of inter and subtidal macrofaunalcommunities of subtropical sandy beaches. Revista Chilena deHistoria Natural 69, 565–577.
Brown, A.C., McLachlan, A., 1990. Ecology of Sandy Shores.Elsevier, Amsterdam.
Cardoso, R.S., Veloso, V.G., 1996. Population biology andsecondary production of the sandhopper Pseudorchesto-idea brasiliensis (Amphipoda: Talitridae) at Prainhabeach, Brazil. Marine Ecology Progress Series 142,111–119.
Cardoso, R.S., Veloso, V.G., 2003. Population dynamics andsecondary production of the wedge clam Donax hanleyanus(Bivalvia: Donacidae) on a high-energy, subtropical beach ofBrazil. Marine Biology 142, 153–162.
Cardoso, R.S., Veloso, V.G., Caetano, C.H.S., 2003. Life history ofEmerita brasiliensis (Decapoda: Hippidae) on two beaches withdifferent morphodynamic characteristics. Journal of CoastalResarch 35, 392–401 (Special issue).
Defeo, O., Gomez, J., Lercari, D., 2001. Testing the swash exclusionhypothesis in sandy beach populations: the mole crab Emerita

B I O L O G I C A L C O N S E R VAT I O N 1 2 7 ( 2 0 0 6 ) 5 1 0 –5 1 5 515
brasiliensis in Uruguay. Marine Ecology Progress Series 212,159–170.
Defeo, O., Jaramillo, E., Lyonnet, A., 1992. Communitystructure and intertidal zonation of the macroinfauna inthe Atlantic coast of Uruguay. Journal of Coastal Research8, 830–839.
Dugan, J.E., Hubbard, D.M., Wenner, A.M., 1994. Geographicalvariation in the life history of the sand crab Emerita analoga(Stimpson) on the California coast: relationships toenvironmental variables. Journal of Experimental MarineBiology and Ecology 118, 255–278.
Emery, K.O., 1961. A simple method of measuring beachesprofiles. Limnology and Oceanography 6, 90–93.
Folk, R.L., Ward, W.C., 1957. Brazos River bar, a study insignificance of grain size parameters. Journal of SedimentaryPetrology 27, 3–26.
Fonseca, D.B., Veloso, V.G., Cardoso, R.S., 2000. Growth, mortalityand reproduction of Excirolana braziliensis Richardson, 1912(Isopoda: Cirolanidae) on the Prainha beach, Rio de Janeiro,Brasil. Crustaceana 73, 535–545.
Gibbs, R.J., Matthews, M.D., Link, D.A., 1971. The relationshipbetween sphere size and settling velocity. Journal ofSedimentary Petrology 41, 7–18.
Gomez, J., Defeo, O., 1999. Life history of the sandhopperPseudorchestoidea brasiliensis (Amphipoda) in sandy beacheswith contrasting morphodynamics. Marine Ecology ProgressSeries 182, 209–220.
James, R.J., Fairweather, P.G., 1996. Spatial variation of intertidalmacrofauna on a sandy ocean beach in Australia. Estuarine,Coastal and Shelf Science 43, 81–107.
Jaramillo, E., Contreras, H., Quijon, P., 1996. Macroinfauna andhuman disturbance in a sandy beach of south-central Chile.Revista Chilena de Historia Natural 69, 655–663.
Jaramillo, E., McLachlan, A., Coetzee, P., 1993. Intertidal zonationpatterns of macrofauna over a range of exposed sandybeaches in south-central Chile. Marine Ecology Progress Series101, 105–118.
Hosier, P.E., Eaton, T.E., 1980. The impact of vehicles ondune and grassland vegetation on a south-eastern NorthCarolina barrier beach. Journal of Applied Ecology 17,173–182.
McArdle, S.B., McLachlan, A., 1992. Sand beach ecology: swashfeatures relevant to the macrofauna. Journal of CoastalResearch 8, 398–407.
McIntyre, A.D., 1995. Human impacts on the oceans: the 1990sand beyond. Marine Pollution Bulletin 31, 147–151.
McLachlan, A., 1983. Sandy beach ecology: a review. In: McLachlan,A., Erasmus, T. (Eds.), Sandy Beaches as Ecosystems. W. JunkPublishers, The Netherlands, pp. 321–380.
McLachlan, A., 1990. Dissipative beaches and macrofauna com-munities on exposed intertidal sands. Journal of CoastalResearch 6, 57–71.
McLachlan, A., Jaramillo, E., Donn, T.E., Wessels, F., 1993. Sandybeach macrofauna communities and their control by thephysical environment: a geographical comparison. Journal ofCoastal Research 15, 27–38 (Special issue).
McLachlan, A., Jaramillo, E., Defeo, O., Dugan, J., De Ruyck, A.,Coetzee, P., 1995. Adaptations of bivalves to different beachtypes. Journal of Experimental Marine Biology and Ecology187, 147–160.
Moffet, M.D., McLachlan, A., Winter, P.E.D., De Ruyck, A.M.C.,1998. Impact of trampling on sandy beach macrofauna.Journal of Coastal Conservation 4, 87–90.
Rickard, C.A., McLachlan, A., Kerley, G.I.H., 1994. The effects ofvehicular and pedestrian traffic on dune vegetation in SouthAfrica. Ocean and Coastal Management 23, 225–247.
Short, A.D., Wright, L.D., 1983. Physical variability of sandybeaches. In: McLachlan, A., Erasmus, T. (Eds.), Sandy Beachesas Ecosystems. W. Junk Publishers, The Netherlands, pp. 133–144.
Souza, J.R.B., Gianuca, N.M., 1995. Zonation and seasonal varia-tion of the intertidal macrofauna on a sandy beach of ParanaState, Brazil. Scientia Marina 59, 103–111.
Veloso, V.G., Cardoso, R.S., 1999. Population biology of the molecrab Emerita brasiliensis (Decapoda: Hippidae) at Fora beach,Brazil. Journal of Crustacean Biology 19, 147–153.
Veloso, V.G., Caetano, C.H.S., Cardoso, R.S., 2003. Composition,structure and zonation of intertidal macroinfauna in relationto physical factors in microtidal sandy beaches at Rio deJaneiro State, Brazil. Scientia Marina 67 (4), 393–402.
Watson, J.J., Kerley, G.I.H., McLachlan, A., 1996. A Humanactivity and potential impacts on dune breeding birds in theAlexandria Coastal Dunefield. Landscape Urban Planning 34,315–322.
Welawski, J.M., Stanek, A., Siewert, A., Beer, N., 2000. Thesandhopper (Talitrus saltator, Montagu 1808) on the PolishBaltic Coast. Is a victim of increased tourism? OceanologicalStudies 29, 77–87.



















