
Evaluation of reproductive endocrine status in hornyhead turbot sampled from Southern California's...
12
Sources and Effects of Contaminants of Emerging Concern in Southern California Coastal Waters EVALUATION OF REPRODUCTIVE ENDOCRINE STATUS IN HORNYHEAD TURBOT SAMPLED FROM SOUTHERN CALIFORNIA’S URBANIZED COASTAL ENVIRONMENTS JESUS A. REYES, y DORIS E. VIDAL-DORSCH, z DANIEL SCHLENK,§ STEVEN M. BAY, z JEFFREY L. ARMSTRONG, k JOSEPH R. GULLY,# CURTIS CASH, yy MICHAEL BAKER, zz TIMOTHY D. STEBBINS,§§ GARY HARDIMAN, zz and KEVIN M. KELLEY*y yEnvironmental Endocrinology Laboratory, California State University, Long Beach, California, USA zSouthern California Coastal Water Research Project, Costa Mesa, California, USA §Department of Environmental Sciences, University of California, Riverside, California, USA kEnvironmental Laboratory and Ocean Monitoring Division, Orange County Sanitation District, Fountain Valley, California, USA #Sanitation Districts of Los Angeles County, Whittier, California, USA yyEnvironmental Monitoring Division, City of Los Angeles, Playa Del Rey, California, USA zzDepartment of Medicine, University of California, San Diego, California, USA §§City of San Diego Marine Biology Laboratory, Public Utilities Department, San Diego, California, USA (Submitted 31 May 2012; Returned for Revision 9 July 2012; Accepted 8 August 2012) Abstract —As part of a regionwide collaboration to determine the occurrence of contaminants and biological effects in coastal ecosystems offshore of urban southern California, the present study characterized the reproductive endocrinology of an indigenous flatfish, the hornyhead turbot (Pleuronichthys verticalis), and compared groups sampled from different study sites representing varying degrees of pollution to screen for potential endocrine disruptive effects. Turbot were sampled from locations near the coastal discharge sites of four large municipal wastewater treatment plants (WWTPs) located between Los Angeles and San Diego, California, USA, and were compared with fish sampled from three far-field reference locations in the region. Despite environmental presence of both legacy contaminants and contaminants of emerging concern and evidence for fish exposure to several classes of contaminants, both males and females generally exhibited coordinated seasonal reproductive cycles at all study sites. Patterns observed included peaks in sex steroids (17b-estradiol, testosterone, 11-ketotestosterone) in the spring and low levels in the fall, changes corresponding to similarly timed gonadal changes and plasma vitellogenin concentrations in females. Comparisons between fish captured at the different study sites demonstrated some regional differences in plasma levels of estrogens and androgens, indicative of location-associated effects on the endocrine system. The observed differences, however, could not be linked to the ocean discharge locations of four of the largest WWTPs in the world. Environ. Toxicol. Chem. 2012;31:2689–2700. # 2012 SETAC Keywords —Aquatic toxicology Effects-based monitoring Fish Endocrine disruptors Water quality INTRODUCTION The Southern California Bight (SCB) represents one of the most intensive interfaces in the world between a marine eco- system and a substantial human population. In the region encompassing Los Angeles, Orange, and San Diego Counties, a population of more than 20 million resides within 50 km of the coastline, in an area that includes extensive industrial, com- mercial, residential, and other activities. Servicing these urban centers are four of the largest wastewater treatment plant (WWTP) facilities in the world, which together with some smaller WWTPs release more than 1 billion gallons (3.8 bil- lion liters) of treated wastewater each day into the coastal marine environment. In addition to the inputs of treated waste- waters, numerous other point and nonpoint sources of contam- inants are present in the SCB ecosystem, including urban runoff, storm water, ports and harbors, and agriculture [1]. The impacts of southern California WWTP outfalls on environmental quality, benthic invertebrate and fish community structure, bioaccumulation of organic and metal contaminants in organisms, and toxicity to invertebrates and fish have been monitored and studied for more than 35 years by local munic- ipalities and sanitation districts and other agencies [2–6]. From this work, the fact that many locales in the SCB—both near to and far from WWTP outfalls—have been contaminated by a wide variety of human-derived chemicals, particularly offshore of Los Angeles, Orange County, and San Diego, has been well documented. A number of the contaminants known to be present in the SCB can act to disrupt endocrine and physiological systems in aquatic organisms such as fish. Such contaminants present in urban environments include chlorinated pesticides, industrial chemicals such as polychlorinated biphenyls (PCBs), or polycyclic aromatic hydrocarbons, but they also include so- called contaminants of emerging concern (CECs). The latter includes an increasing variety of pharmaceuticals and personal care products, current-use pesticides, various industrial and commercial compounds, as well as natural and synthetic hormones [7–10]. Steroid hormones, derived principally from excretion by humans and domestic animals, are well established as endocrine disruptors present in aquatic ecosys- tems worldwide [11], whereas many other CECs are beginning to be linked to effects in organisms. Because many CECs are designed to be active in animals at relatively low concentrations, concern exists that increasing presence of CECs in urban aquatic environment may be leading to disrupted animal health, with possible relevance to ecosystem-level impacts. Environmental Toxicology and Chemistry, Vol. 31, No. 12, pp. 2689–2700, 2012 # 2012 SETAC Printed in the USA DOI: 10.1002/etc.2008 * To whom correspondence may be addressed ([email protected]). Published online 15 September 2012 in Wiley Online Library (wileyonlinelibrary.com). 2689
Transcript of Evaluation of reproductive endocrine status in hornyhead turbot sampled from Southern California's...

Sources and Effects of Contaminants of Emerging Concern in Southern California Coastal Waters
EVALUATION OF REPRODUCTIVE ENDOCRINE STATUS IN HORNYHEAD TURBOTSAMPLED FROM SOUTHERN CALIFORNIA’S URBANIZED COASTAL ENVIRONMENTS
JESUS A. REYES,y DORIS E. VIDAL-DORSCH,z DANIEL SCHLENK,§ STEVEN M. BAY,z JEFFREY L. ARMSTRONG,kJOSEPH R. GULLY,# CURTIS CASH,yy MICHAEL BAKER,zz TIMOTHY D. STEBBINS,§§ GARY HARDIMAN,zz
and KEVIN M. KELLEY*yyEnvironmental Endocrinology Laboratory, California State University, Long Beach, California, USA
zSouthern California Coastal Water Research Project, Costa Mesa, California, USA
§Department of Environmental Sciences, University of California, Riverside, California, USA
kEnvironmental Laboratory and Ocean Monitoring Division, Orange County Sanitation District, Fountain Valley, California, USA
#Sanitation Districts of Los Angeles County, Whittier, California, USA
yyEnvironmental Monitoring Division, City of Los Angeles, Playa Del Rey, California, USA
zzDepartment of Medicine, University of California, San Diego, California, USA
§§City of San Diego Marine Biology Laboratory, Public Utilities Department, San Diego, California, USA
(Submitted 31 May 2012; Returned for Revision 9 July 2012; Accepted 8 August 2012)
Abstract—As part of a regionwide collaboration to determine the occurrence of contaminants and biological effects in coastalecosystems offshore of urban southern California, the present study characterized the reproductive endocrinology of an indigenousflatfish, the hornyhead turbot (Pleuronichthys verticalis), and compared groups sampled from different study sites representing varyingdegrees of pollution to screen for potential endocrine disruptive effects. Turbot were sampled from locations near the coastal dischargesites of four large municipal wastewater treatment plants (WWTPs) located between Los Angeles and San Diego, California, USA, andwere compared with fish sampled from three far-field reference locations in the region. Despite environmental presence of both legacycontaminants and contaminants of emerging concern and evidence for fish exposure to several classes of contaminants, both males andfemales generally exhibited coordinated seasonal reproductive cycles at all study sites. Patterns observed included peaks in sex steroids(17b-estradiol, testosterone, 11-ketotestosterone) in the spring and low levels in the fall, changes corresponding to similarly timedgonadal changes and plasma vitellogenin concentrations in females. Comparisons between fish captured at the different study sitesdemonstrated some regional differences in plasma levels of estrogens and androgens, indicative of location-associated effects on theendocrine system. The observed differences, however, could not be linked to the ocean discharge locations of four of the largest WWTPsin the world. Environ. Toxicol. Chem. 2012;31:2689–2700. # 2012 SETAC
Keywords—Aquatic toxicology Effects-based monitoring Fish Endocrine disruptors Water quality
INTRODUCTION
The Southern California Bight (SCB) represents one of themost intensive interfaces in the world between a marine eco-system and a substantial human population. In the regionencompassing Los Angeles, Orange, and San Diego Counties,a population of more than 20million resides within 50 km of thecoastline, in an area that includes extensive industrial, com-mercial, residential, and other activities. Servicing these urbancenters are four of the largest wastewater treatment plant(WWTP) facilities in the world, which together with somesmaller WWTPs release more than 1 billion gallons (3.8 bil-lion liters) of treated wastewater each day into the coastalmarine environment. In addition to the inputs of treated waste-waters, numerous other point and nonpoint sources of contam-inants are present in the SCB ecosystem, including urban runoff,storm water, ports and harbors, and agriculture [1].
The impacts of southern California WWTP outfalls onenvironmental quality, benthic invertebrate and fish communitystructure, bioaccumulation of organic and metal contaminantsin organisms, and toxicity to invertebrates and fish have beenmonitored and studied for more than 35 years by local munic-
ipalities and sanitation districts and other agencies [2–6]. Fromthis work, the fact that many locales in the SCB—both near toand far from WWTP outfalls—have been contaminated by awide variety of human-derived chemicals, particularly offshoreof Los Angeles, Orange County, and San Diego, has been welldocumented.
A number of the contaminants known to be present in theSCB can act to disrupt endocrine and physiological systemsin aquatic organisms such as fish. Such contaminants present inurban environments include chlorinated pesticides, industrialchemicals such as polychlorinated biphenyls (PCBs), orpolycyclic aromatic hydrocarbons, but they also include so-called contaminants of emerging concern (CECs). The latterincludes an increasing variety of pharmaceuticals and personalcare products, current-use pesticides, various industrial andcommercial compounds, as well as natural and synthetichormones [7–10]. Steroid hormones, derived principallyfrom excretion by humans and domestic animals, are wellestablished as endocrine disruptors present in aquatic ecosys-tems worldwide [11], whereas many other CECs are beginningto be linked to effects in organisms. Because many CECsare designed to be active in animals at relatively lowconcentrations, concern exists that increasing presence of CECsin urban aquatic environment may be leading to disruptedanimal health, with possible relevance to ecosystem-levelimpacts.
Environmental Toxicology and Chemistry, Vol. 31, No. 12, pp. 2689–2700, 2012# 2012 SETAC
Printed in the USADOI: 10.1002/etc.2008
* To whom correspondence may be addressed([email protected]).
Published online 15 September 2012 in Wiley Online Library(wileyonlinelibrary.com).
2689

In southern California, information is severely lacking onCEC inputs from municipal WWTP discharges and on whetherassociated effects occur in resident organisms, which wouldappear to be likely given the substantial interface betweenhumans and the marine environment in this region ([12–15];J.A. Reyes, 2006, Master’s thesis, California State University,Long Beach, CA). Furthermore, limited study and understand-ing exist of the impacts of ocean-located WWTP outfallsworldwide on organism health [16].
In the present study, an indigenous flatfish species, thehornyhead turbot (Pleuronichthys verticalis), was investigatedto determine whether its residence in areas of WWTP dis-charges may be associated with disruptions in their reproductiveendocrine system. Four of the WWTPs located from LosAngeles southward to San Diego are some of the largestfacilities in the world and discharge into the coastal marineenvironment. These sites and three far-field study sites wereevaluated as part of a regionwide, multi-institution collabora-tion, which culminated in two chemistry studies measuringCECs and other contaminants present in WWTP effluentsand receiving water samples [17] and in sediments and fishtissue samples [18], and two biological studies centered onphysiological measures of reproduction [19] and on endocrinesystem assessment and its relationship to reproductive function(present study).
MATERIALS AND METHODS
Fish sampling and study design
All fish analyzed in the present study were caught by ottertrawling at specific coastal locations historically used in envi-ronmental monitoring in the SCB (Fig. 1), which included the
following seven sites: outfall site located within Santa MonicaBay in the city of Los Angeles, Bureau of Sanitation, Environ-mental Monitoring Division (LA); outfall site (PV) and far-fieldsite (PVF) located off the Palos Verdes Peninsula (Los AngelesCounty Sanitation Districts); outfall site (OC), far-field site(OCF), and a reference site near Dana Point (DP), all offshore ofthe Orange County coastline (Orange County Sanitation Dis-trict); outfall site (SD) located offshore of Point Loma, SanDiego (City of San Diego Public Utilities Department). All ofthe study sites were located at depths between 60 and 90m (i.e.,discharge depths of the WWTP outfalls) and exhibited temper-atures consistently between 10 and 128C. Trawling for fish ateach site was guided by differential global positioning systemand followed 500- to 600-m transects along the bottom at aspeed of 1 to 1.5m/s.
The reproductive seasonality in male and female hornyheadturbot was evaluated by quarterly sampling at five of the abovesites (PV, PVF, OC, OCF, and DP), in spring (May–June, 2006),summer (August 2006), fall (November 2006), and winter(January and February 2007). All seven study sites weresampled in May through June 2006 to assess potential site-associated differences in endocrine status across the entireregion. Each trawl catch was released into on-board racewayswith flow-through seawater, and adult hornyhead turbot rangingbetween 13 and 19 cm were selected for tissue sampling (thenumber of fish per each group is provided within tables andfigures); the remaining fish were returned to the environment.Mean body weight (measured to nearest g) and standard length(measured to nearest mm) indicated that fish age (both sexes)did not significantly differ across study sites and did notcorrelate with endocrine measures (see Forsgren et al. [19]for body size and age data). Blood was collected from the caudal
Fig. 1. Map of the Southern California Bight (SCB) indicating the locations of the study sites. Wastewater treatment plant outfall locations (represented by opencircles) includeCityofLosAngelesHyperionTreatmentPlant outfall inSantaMonicaBay (LA),LosAngelesCountySanitationDistrict outfall at thePalosVerdesPeninsula (PV), OrangeCounty SanitationDistrict outfall (OC), andCity of SanDiegoMetropolitanWastewater Treatment Plant outfall (SD). Far-field referencelocations (represented by filled circles) include sites at Dana Point (DP), northOrange County (OCF), and north Palos Verdes Peninsula (PVF). Off-coast currentsare predominantly northward in the inshore SCB regionwhere the study sites are located. [Color figure can be seen in the online version of this article, available atwileyonlinelibrary.com]
2690 Environ. Toxicol. Chem. 31, 2012 J.A. Reyes et al.

vein via a heparinized 22-gauge needle and syringe and imme-diately centrifuged for 5min at 5,000 rpm to pellet theblood cells; plasma was placed into heparinized microcentri-fuge tubes (Fisher Scientific), frozen on dry ice, andthen subsequently stored in a –808C freezer until analysis.The sex of each fish was determined by inspection of thegonads, followed by removal of the right-side gonad forweight measurement (to the nearest milligram). Gonad weightwas then used to calculate 0.5 gonadosomatic index (GSI¼right gonad wt (g)/body wt (g)� 100%), used as anindicator of relative reproductive status, as previously described[14,15]. Analyses of seasonal and site-related differencesin male and female gonadal histomorphology are reportedby Forsgren et al. [19]. Whole liver was removed,weighed to the nearest milligram, and used to calculate hep-atosomatic index (HSI; [liver wt (g)/bodywt (g)� 100%]), anindicator of relative hepatic fuel (glycogen, fat) storage infishes [20].
In the associated chemistry studies, a variety of CECsand other contaminants were measured in effluent, receivingwater, sediment, and in turbot liver composite samplesderived from the four WWTP outfall locations and the DPreference site [17,18]. Several pharmaceuticals and personalcare products, industrial and commercial compounds, andhormones were frequently detected in effluents from eachof the four WWTPs at relatively low microgram per literconcentrations, with only a small proportion detected inreceiving waters at much lower (400–1,000 times) concen-trations [17]. Sediment also contained a variety of CECs andother contaminants [18]. Although several steroid hormoneswere detected in effluents (17b-estradiol [E2], testosterone,progesterone, and estrone [E1]), they were not detected inwater [17] or sediment [18]; in fish liver samples the syntheticestrogen, 17a-ethinylestradiol, also was not detected. Thepharmaceutical and personal care product diazepam (tradename Valium) was the sole pharmaceutical and personalcare product detected in fish liver, present in 100% of thesamples at a mean concentration of 56mg/kg (wet wt), despiteits infrequent detection in sediment at low concentrations(<1mg/kg). The industrial and commercial compound4-nonylphenol was also frequently detected in fish tissue(mean concentration of 102mg/kg), in addition to the bromi-nated diphenyl ether congeners, (BDE)–47 (156mg/kg), BDE99 (69mg/kg), BDE 100 (47mg/kg), and BDE 154 (11mg/kg).Legacy contaminants were frequently detected in fish tissues,including PCB congeners 138 and 153 (111–132mg/kg) andnotably high levels of 4,40-DDE (12,200mg/kg). When com-paring across all study sites, generally lower concentrationsand frequencies of detection were found in sediment and fishtissues from the DP reference site as compared with theWWTP sites, which was particularly evident for polybromi-nated diphenyl ethers, PCBs, and organochlorine pesticides.The latter two classes (legacy contaminants) showed thehighest levels at the two northernmost outfalls (LA and PV),reflecting historic discharges from the Los Angeles region. Incontrast to these patterns, diazepam and 4-nonylphenol werepresent in liver of fish from the DP reference site at concen-trations comparable to those of fish from WWTP sites. Theresults from these studies, which serve as a backdrop tothe study sites used in the present study, generally indicatethat hornyhead turbot are exposed to a variety of CECs andlegacy contaminants at all sites, with the DP reference siteexhibiting relatively lower occurrences for several classes ofcontaminants [17,18].
Hormone measurements
Plasma concentrations of E2 and testosterone were measuredby radioimmunoassay (RIA) using 125I-labeled E2 or testoster-one and specific polyclonal rabbit antisera obtained from Diag-nostic Systems Laboratory (Beckman Coulter). Separation offree and bound antigen was achieved using a double-antibodysystem (goat anti-rabbit gamma globulin serum) and polyethy-lene glycol as a precipitating aid. Pelleted 125I was counted on aPerkin-Elmer Cobra II gamma counter (Packard Instruments),and hormone concentrations were calculated from a standardcurve of %B/Bo versus concentration of unlabeled ligand(0–750 pg/ml for E2; 0–25 ng/ml for testosterone), using SigmaPlot Version 11 software (SPSS). Intra-assay precision was 7.4and 7.5% (coefficients of variation [CV]¼ SD/mean� 100%)for the E2 and testosterone RIAs, respectively; interassayprecision was 8.6 and 8.4%, respectively.
Plasma concentrations of the fish-specific androgen 11-ketotestosterone (11-KT) were measured by enzyme immuno-assay using an 11-KT-acetylcholinesterase conjugate as tracerand specific antisera obtained from Cayman Chemical.Antibody-bound tracer or endogenous hormones were boundto mouse monoclonal anti-rabbit immunoglobulin G andattached to microplate wells, washed, then reacted in Ellman’sreagent. Absorbance at 412 nm was measured in a microplatespectrophotometer (Spectramax R250, Molecular Devices), andhormone concentrations were calculated from a standard curveof %B/Bo versus concentration of unlabeled ligand (0–100 pg/ml)using Sigma Plot Version 11 software (SPSS). Intra-assayprecision was 11.2% (CV); interassay precision was 8.1%.
Vitellogenin measurement
Plasma concentrations of vitellogenin (VTG) were measuredby enzyme-linked immunosorbent assay as described in Rempelet al. [14] and Forsgren et al. [19], using rabbit anti-turbot VTGprimary antiserum from Cayman Chemical and alkaline phos-phatase-conjugated secondary antibody (goat anti-rabbit, Bio-Rad Labs). Absorbance at 405 nm was measured, and hormoneconcentrations in the samples were calculated from a standardcurve of %B/Bo versus concentration of unlabeled purifiedCalifornia halibut VTG as standard, as described previously,and expressed as milligrams per milliliter (females) or micro-grams per milliliter (males). In the paper by Forsgren et al. [19],location-related differences are reported, whereas in the presentstudy, correlative relationships with the endocrine system andoverall seasonality were reported.
Data analysis
Individual hormone concentrations (ng/ml, pg/ml), HSI (%),GSI (%), and VTG concentrations (ng/ml) were organizedaccording to sex, study site, and season, and used to calculatemean� standard error of the mean (SEM) values, as provided intables and figures. Potential differences in mean values (withineach sex) across the study sites were evaluated by one-wayanalysis of variance using Sigmaplot Version 11 software(SPSS). Differences in mean values according to season (fall,winter, spring, summer) were evaluated by analysis of varianceusing all quarterly sampled fish (i.e., from sites PVF, PV, OCF,OC, DP); analysis of only non-outfall quarterly sampled fish(from PVF, OCF, DP) did not elucidate any significant differ-ences (the former analysis is illustrated). Differences amonggroups were compared by the Holm-Sidek ad hoc multiplecomparisons test. The nonparametric Kruskall-Wallis analysisof variance on ranks test was used to evaluate datasets that were
Reproductive endocrine status in urban coastal marine fish Environ. Toxicol. Chem. 31, 2012 2691
not normally distributed. Spearman Rank Order correlationtests were carried out using the same software to examinerelationships between parameters (e.g., hormone concentrationvs GSI). Differences in mean�SEM values or in correlationcoefficient (r) were considered significant when p< 0.05.
RESULTS
Overall seasonal changes in reproductive endocrine status inmales
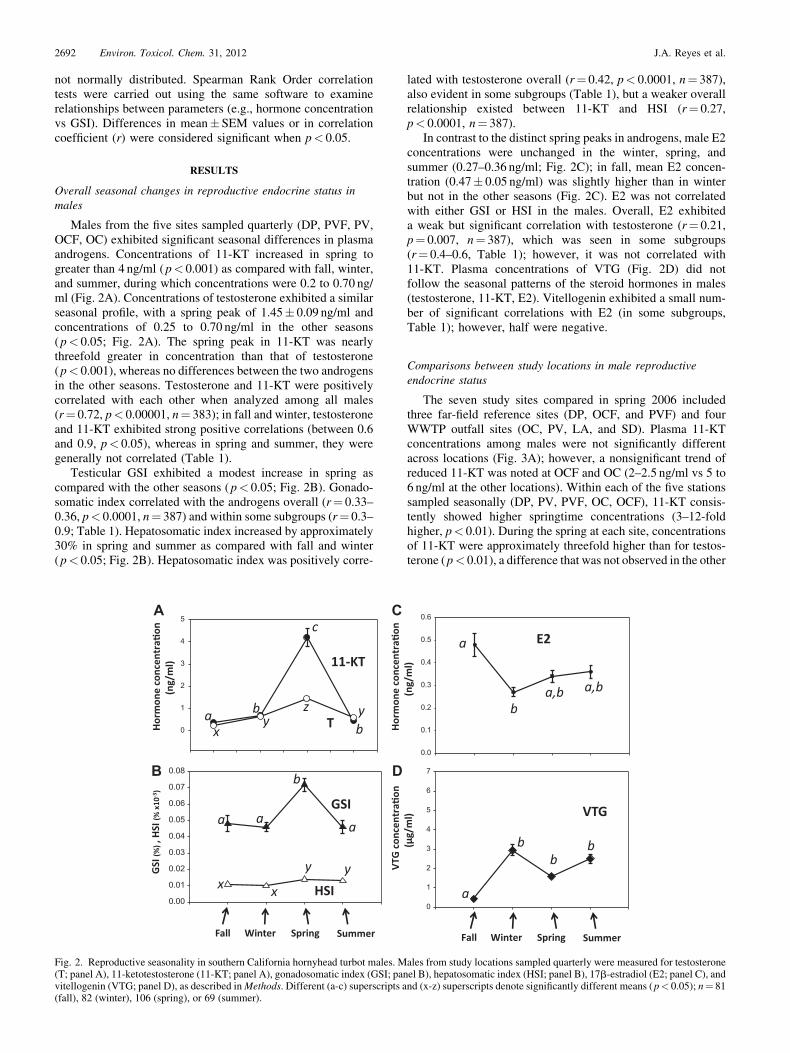
Males from the five sites sampled quarterly (DP, PVF, PV,OCF, OC) exhibited significant seasonal differences in plasmaandrogens. Concentrations of 11-KT increased in spring togreater than 4 ng/ml (p< 0.001) as compared with fall, winter,and summer, during which concentrations were 0.2 to 0.70 ng/ml (Fig. 2A). Concentrations of testosterone exhibited a similarseasonal profile, with a spring peak of 1.45� 0.09 ng/ml andconcentrations of 0.25 to 0.70 ng/ml in the other seasons(p< 0.05; Fig. 2A). The spring peak in 11-KT was nearlythreefold greater in concentration than that of testosterone(p< 0.001), whereas no differences between the two androgensin the other seasons. Testosterone and 11-KT were positivelycorrelated with each other when analyzed among all males(r¼ 0.72, p< 0.00001, n¼ 383); in fall and winter, testosteroneand 11-KT exhibited strong positive correlations (between 0.6and 0.9, p< 0.05), whereas in spring and summer, they weregenerally not correlated (Table 1).
Testicular GSI exhibited a modest increase in spring ascompared with the other seasons (p< 0.05; Fig. 2B). Gonado-somatic index correlated with the androgens overall (r¼ 0.33–0.36, p< 0.0001, n¼ 387) and within some subgroups (r¼ 0.3–0.9; Table 1). Hepatosomatic index increased by approximately30% in spring and summer as compared with fall and winter(p< 0.05; Fig. 2B). Hepatosomatic index was positively corre-
lated with testosterone overall (r¼ 0.42, p< 0.0001, n¼ 387),also evident in some subgroups (Table 1), but a weaker overallrelationship existed between 11-KT and HSI (r¼ 0.27,p< 0.0001, n¼ 387).
In contrast to the distinct spring peaks in androgens, male E2concentrations were unchanged in the winter, spring, andsummer (0.27–0.36 ng/ml; Fig. 2C); in fall, mean E2 concen-tration (0.47� 0.05 ng/ml) was slightly higher than in winterbut not in the other seasons (Fig. 2C). E2 was not correlatedwith either GSI or HSI in the males. Overall, E2 exhibiteda weak but significant correlation with testosterone (r¼ 0.21,p¼ 0.007, n¼ 387), which was seen in some subgroups(r¼ 0.4–0.6, Table 1); however, it was not correlated with11-KT. Plasma concentrations of VTG (Fig. 2D) did notfollow the seasonal patterns of the steroid hormones in males(testosterone, 11-KT, E2). Vitellogenin exhibited a small num-ber of significant correlations with E2 (in some subgroups,Table 1); however, half were negative.
Comparisons between study locations in male reproductiveendocrine status
The seven study sites compared in spring 2006 includedthree far-field reference sites (DP, OCF, and PVF) and fourWWTP outfall sites (OC, PV, LA, and SD). Plasma 11-KTconcentrations among males were not significantly differentacross locations (Fig. 3A); however, a nonsignificant trend ofreduced 11-KT was noted at OCF and OC (2–2.5 ng/ml vs 5 to6 ng/ml at the other locations). Within each of the five stationssampled seasonally (DP, PV, PVF, OC, OCF), 11-KT consis-tently showed higher springtime concentrations (3–12-foldhigher, p< 0.01). During the spring at each site, concentrationsof 11-KT were approximately threefold higher than for testos-terone (p< 0.01), a difference that was not observed in the other
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0
1
2
3
4
5
11-KT
T
c
a bbx
yyz
Horm
one
conc
entr
a�on
(n
g/m
l)
0.00
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
Fall Winter Spring Summer
GSI
HSI
a a a
b
x x
y yGSI
(%) ,
HSI
(% x
10-3
)
E2
a,b
a
b
Horm
one
conc
entr
a�on
(ng/
ml)
a,b
0
1
2
3
4
5
6
7
VTG
b
a
bb
VTG
con
cent
ra�o
n(µ
g/m
l)
Fall Winter Spring Summer
A C
DB
Fig. 2. Reproductive seasonality in southern California hornyhead turbot males. Males from study locations sampled quarterly were measured for testosterone(T; panel A), 11-ketotestosterone (11-KT; panel A), gonadosomatic index (GSI; panel B), hepatosomatic index (HSI; panel B), 17b-estradiol (E2; panel C), andvitellogenin (VTG; panel D), as described inMethods. Different (a-c) superscripts and (x-z) superscripts denote significantly different means (p< 0.05); n¼ 81(fall), 82 (winter), 106 (spring), or 69 (summer).
2692 Environ. Toxicol. Chem. 31, 2012 J.A. Reyes et al.

seasons. Seasonal patterns in testosterone, however, were morevariable, although testosterone was most often lowest in the falland relatively elevated in the spring (Fig. 3B). Spring concen-trations of testosterone were significantly lower in males fromthe OCF and PVF far-field locations as compared with the OCand SD outfall sites (p< 0.05; Fig. 3B). Fish at PVF and PVgenerally had relatively higher testosterone concentrations out-side of the spring season (Fig. 3B).
As reported in Forsgren et al. [19], mean testicular GSIvalues were not significantly different in fish across study sites,nor did GSI exhibit obvious seasonality at most stations.Hepatosomatic index also did not differ among the sites, withthe exception of one location (PV), in which it was slightlyincreased (to 1.7 from�1.2% in other groups, p< 0.05; data notillustrated). Hepatosomatic index did not exhibit any significantseasonal differences across sites, but in general tended to behigher in spring and summer at most sites (data not illustrated),consistent with the data shown in Fig. 2B.
Plasma concentrations of E2 in the males (Fig. 4A) werelowest at the OC and OCF sites (83–170 pg/ml, p< 0.05 vsother locations), intermediate in Santa Monica Bay (LA) andPalos Verdes (PVF and PV), and highest at the DP and SDlocations (575–800 pg/ml, p< 0.05 vs other locations). Amongthe seasonally sampled locations, DP tended to have the highest
E2 in males. No significant seasonal patterns were seen in maleE2 levels within any site (Fig. 4).
Overall seasonal changes in female reproductive endocrine status
Among females from the five quarterly sampled sites, over-all E2 concentrations ranged between 150 and 230 pg/ml andwere significantly higher in spring as compared with the otherseasons (p< 0.05; Fig. 5). Ovarian GSI similarly peaked inspring as compared with the other seasons (p< 0.05), and wasalso relatively elevated in summer as compared with fall andwinter (p< 0.05; Fig. 5B). Plasma concentrations of VTG wereconsiderably higher in females than in males (e.g., 1.7mg/ml vs1.6mg/ml, respectively, in spring; p< 0.0001) and followed aseasonal pattern similar to that observed for E2 and GSI(Fig. 5B). Concentrations of E2 showed overall positive corre-lations with both GSI (r¼ 0.27, p< 0.0001, n¼ 353) and VTG(r¼ 0.34, p< 0.0001, n¼ 353), and when evaluated within sub-groups (e.g., within season or site), some similar and strongercorrelations were observed (r¼ 0.4–0.7; Table 2). Gonadoso-matic index and VTG were strongly correlated with each other,both overall (r¼ 0.61, p< 0.00001, n¼ 353) and within amajority of subgroups (Table 2).
Hepatosomatic index in females exhibited a seasonal profilecomparable to that observed for E2, GSI, and VTG, with higher
Table 1. Correlation coefficients (r) and p values of the relationships among reproductive parameters measured in hornyhead turbot males sampled fromdifferent study sites
Males Orange County Palos Verdes San Diego Santa Monica Bay
Season Far-field DP Far-field OCF Outfall OC Far-field PVF Outfall PV Outfall SD Outfall LA
T vs 11-KT Fall 0.78c (23) 0.87c (22) 0.86c (18) 0.75d (6) 0.73b (15) – –Winter 0.82c (14) 0.74c (26) 0.74c (19) 0.56d (5) 0.77b (12) – –Spring NS (32) NS (5) 0.31d (24) NS (13) NS (31) NS (34) NS (23)Summer NS (20) 0.67b (19) NS (9) NS (8) NS (10) – –
All seasons 0.80c (89) 0.60c (72) 0.77c (70) NS (27) 0.69c (68) – –T vs GSI Fall NS (23) NS (22) NS (18) NS (6) 0.40a (15) – –
Winter 0.60a (14)e 0.39a (27) NS (19) NS (5) NS (13) – –Spring NS (32) 0.90d (5) NS (24) 0.35d (13)e NS (31) NS (34) 0.32d (23)Summer 0.41a (20) NS (20) NS (10) NS (8) NS (10) – –
All seasons 0.33b (89)e NS (74) NS (71) NS (27) 0.23a (69)e – –T vs HSIf Fall NS (23) 0.40a (22)f NS (18) NS (6) NS (15) – –
Winter NS (14) NS (27) NS (19) NS (5) NS (13) – –Spring 0.38a (32) NS (5) 0.67b (24) NS (13) NS (31) 0.40a (34) 0.31d (23)Summer NS (20) NS (20) 0.66a (10) NS (8) NS (10) – –
All seasons 0.30b (89) 0.31b (74) 0.41b (71) NS (27) 0.31b (69) – –T vs E2g Fall NS (23) �0.48a (22)i 0.38d (18) NS (6) 0.35d (15) – –
Winter 0.52a (14) 0.44a (27) 0.45a (19) NS (5) NS (13) – –Spring NS (32) NS (5) NS (24) 0.41d (13) NS (31) NS (34) 0.61b (23)Summer 0.35d (20) NS (20) NS (10) NS (8) NS (10) – –
All seasons NS (89) NS (74) NS (71) 0.31d (27) NS (69) – –E2 vs VTG Fall �0.36d (23) NS (23) NS (18) NS (6)h 0.51a (15)h – –
Winter NS (15) NS (28) NS (20) NS (5) NS (13) – –Spring �0.29d (32) 0.80d (5) NS (24) NS (13) NS (31) –0.45b (34) –0.39d (23)Summer NS (20) 0.45a (20) NS (10) 0.91c (8) NS (10) – –
All seasons �0.52b (90) NS (76) NS (72) NS (27) NS (69) – –
a Indicates significance level of correlation coefficient p¼ 0.05–0.01.b Indicates significance level of correlation coefficient p¼ 0.01–0.001.c Indicates significance level of correlation coefficient p< 0.001.d Indicates significance level of correlation coefficient p¼ 0.06–0.10.e 11-KT was also correlated with GSI in these groups (r¼ 0.24–0.62, p< 0.05).f 11-KT was not significantly correlated with HSI, with two exceptions (OCF-fall, r¼ 0.64; PV-all, r¼ 040, p> 0.05).g E2 was not significantly correlated with 11-KT, with one exception (OCF-fall, r¼�0.49, p< 0.05).h E2 was not significantly correlated with GSI, except in these groups.i 11-KT was also negatively correlated with E2 (r¼�0.52, p¼ 0.001, n¼ 23).DP¼Dana Point; OCF¼ north Orange County; OC¼Orange County Sanitation District outfall; PVF¼ north Palos Verdes Peninsula; PV¼Los AngelesCounty Sanitation District outfall at Palos Verdes Peninsula; SD¼City of San Diego Metropolitan Wastewater Treatment Plant outfall; LA¼City of LosAngeles Hyperion Treatment Plant outfall in Santa Monica Bay; T¼ testosterone; 11-KT¼ 11-ketotestosterone; GSI¼ gonadosomatic index; HSI¼hepatosomatic index; E2¼ 17b-estradiol; VTG¼ vitellogenin; NS¼ nonsignificant.
Reproductive endocrine status in urban coastal marine fish Environ. Toxicol. Chem. 31, 2012 2693

values in spring and summer (p< 0.05, Fig. 5C). Unlike inmales, HSI was positively correlated with GSI and plasma VTGin females, both overall (r¼ 0.54 and 0.44, respectively;p< 0.00001, n¼ 353) and within several subgroups (r¼ 0.4–0.8; Table 2). Hepatosomatic index was not correlated with E2
or 11-KT overall, but showed a weak overall relationship withtestosterone (r¼ 0.24, p¼ 0.005, n¼ 343).
Plasma concentrations of testosterone in females (Fig. 5D)were less than 50% of those observed in males (p< 0.05).Female testosterone concentrations were elevated in winter and
T conce
ntration
s (ng/m
l)
0
2
4
6
8
11-KT
concen
trations
(ng/ml)
0
2
4
6
8[A]
[B]
a,b a a,b b
a a,b
b
*
* *
* *
** * **
24 16 33 21 24 29 6 21 19 21 25 11 33 24 F W SP SU F W SP SU F W SP SU SP SP
1 6 14 9 16 14 32 11 F W SP SU F W SP SU
24 16 33 21 24 29 6 21 19 21 25 11 33 24 F W SP SU F W SP SU F W SP SU SP SP
1 6 14 9 16 14 32 11 F W SP SU F W SP SU
T co
ncen
tra�
on
(ng/
ml)
11-K
T co
ncen
tra�
on
(ng/
ml)
Orange County
Palos Verdes San Diego
Santa Monica Bay
Far-field Ou�all Far-field Far-field Ou�all Ou�alls DP OCF OC SD LA PVF PV
Fig. 3. Location-associated differences in androgen levels in male hornyhead turbot. Seven southern California locations were compared in spring 2006 and arerepresented as filled bars (City of Los Angeles Hyperion Treatment Plant outfall in SantaMonica Bay [LA], Los Angeles County Sanitation District outfall at thePalosVerdes Peninsula [PV],OrangeCounty SanitationDistrict outfall [OC], andCity of SanDiegoMetropolitanWastewater Treatment Plant outfall [SD], DanaPoint [DP], north Orange County [OCF], and north Palos Verdes Peninsula [PVF]; see map in Fig. 1). Open bars represent the other three seasons, which wereevaluatedonly at sitesDP,OCF,OC,PVF, andPV.Data are presented asmean� standard error ofmeanng/mlvalues for 11-ketotestosterone (11-KT; panelA) andtestosterone (T; panel B), with n shown below each bar. F¼ fall;W¼winter; SP¼ spring; SU¼ summer.Asterisk (�) indicates springmean is significantly higherthan in one or more of the other seasons at that site (p< 0.05). Different superscripts denote significantly different means between sites (p< 0.05).
E2 con
centrat
ions (ng
/ml)
0.0
0.2
0.4
0.6
0.8
1.0
b
24 16 33 21 24 29 6 21 19 21 25 11 35 24 1 6 14 9 16 14 32 11 SP SP F W SP SU F W SP SU F W SP SU F W SP SU F W SP SU
d d
b,c
c
a
c
0.0
0.2
0.4
0.6
0.8
1.0
E2 c
once
ntra
�on
(ng/
ml)
Far-field Ou�all Far-field Far-field Ou�all Ou�alls DP OCF OC SD LA PVF PV
Orange County
Palos Verdes San Diego
Santa Monica Bay
Fig. 4. Location-associated differences in 17b-estradiol (E2) levels inmale hornyhead turbot. Seven southern California locationswere compared in spring 2006and are represented as filled bars (City of LosAngelesHyperionTreatment Plant outfall in SantaMonicaBay [LA], LosAngelesCounty SanitationDistrict outfallat the PalosVerdes Peninsula [PV], OrangeCounty SanitationDistrict outfall [OC], andCity of SanDiegoMetropolitanWastewater Treatment Plant outfall [SD],DanaPoint [DP], northOrangeCounty [OCF], and north PalosVerdes Peninsula [PVF]; seemap in Fig. 1).Openbars represent the other three seasons,whichwereevaluatedonly at sitesDP,OCF,OC,PVF, andPV.Data are presented asmean� standard error ofmeanng/ml values forE2,withn shownbeloweachbar. F¼ fall;W¼winter; SP¼ spring; SU¼ summer. Different superscripts denote significantly different means between sites (p< 0.05).
2694 Environ. Toxicol. Chem. 31, 2012 J.A. Reyes et al.

spring (>0.85 ng/ml) as compared with summer and fall(�0.4 ng/ml; p< 0.05). Concentrations of 11-KT (Fig. 5D)were lower (0.02–0.17 ng/ml; p< 0.05 in each season vs tes-tosterone values), but were also elevated in spring (p< 0.05 vseach other season) and in summer as compared with fall(p< 0.05). Testosterone, which is a precursor for E2 biosyn-thesis, was positively correlated to E2, both overall (r¼ 0.33,p< 0.0001, n¼ 343) and in several subgroups (r¼ 0.4–0.8),whereas 11-KT was mostly unrelated to E2 (Table 2). Testos-terone and 11-KT levels were positively correlated overall(r¼ 0.577, p< 0.0001, n¼ 342) and within several subgroups(r¼ 0.4–0.9; Table 2).
Comparisons between study locations in female endocrine status
Females exhibited spatial differences in plasma E2 concen-trations reminiscent of that seen in the males (Fig. 6A), with thehighest E2 concentrations at the DP and SD sites (p< 0.05 vs allsites except PV) and lowest values at the OC and OCF sites.Within each season except winter, E2 was consistently higher infemales from DP as compared with other sites. In contrast to theoverall spring peak in E2 (see Fig. 5A), seasonal patterns in E2concentrations were mostly not detectable when evaluatedwithin each location (Fig. 6A). Gonadosomatic index andVTG generally showed spring increases within each location,except OCF [19]. Correlations between these indices and E2were not detectable in some subgroups (e.g., OCF, DP) but weredetectable in others (e.g., OC, PV, PVF; Table 2).
Female testosterone concentrations were mostly not differ-ent between sites, except at the LA site at which relatively lowervalues were observed (Fig. 7A). At the seasonally sampled sites,testosterone showed higher levels in spring compared with oneor more other seasons, except at OCF (p< 0.05; Fig. 7A).Although present at an order of magnitude less concentration
than testosterone, 11-KT was also generally higher in spring-time across all sites; at OCF, spring 11-KT concentrations werelower than at several other sites (Fig. 7B). Testosterone and 11-KT showed weak overall correlations with plasma VTG andovarian GSI (r¼ 0.2–0.3, p< 0.001, n¼ 343–352), and selectedsubgroups also reflected these correlations (r¼ 0.4–0.8,p< 0.05; Table 2), except at OCF. Hepatosomatic index exhib-ited no seasonal or spatial patterns in females when evaluatedwithin or across study sites (data not illustrated). Consistentwith its overall relationship to GSI and VTG in females(reported above), HSI showed several positive correlations withGSI and VTG within site or season subgroups (Table 2);however, HSI had few relationships with E2 and testosteroneand no relationship with 11-KT in females, and there were nodiscernible patterns due to study site.
DISCUSSION
In the present study, hornyhead turbot residing offshoreof urban southern California were investigated for possibleenvironment-associated effects on their endocrine physiology,particularly in relation to the discharge locations of fourregional WWTP facilities. These four facilities are some ofthe largest in the world, and they use outfalls that dischargedirectly into the coastal ocean habitat of flatfish, includingturbot. Although the hornyhead turbot has a long history ofbeing monitored in southern California for contaminant accu-mulation and population measures by the discharge agencies[2–5] and by regional monitoring programs [8], this specieshas received only limited biological study, particularly withrespect to its endocrine physiology and reproduction ([15,21];L.D. Cooper, 1997, Master’s thesis, California State University,Long Beach, CA, USA).
0.00
0.05
0.10
0.15
0.20
0.25
0.30
E2
c
a,b b
a H
orm
one
conc
entr
a�on
(n
g/m
l)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
GSI
VTG a
b
d
c
w
x
y
z
VTG
con
cent
ra�o
n (m
g/m
l),
GSI
(%)
Fall Winter Spring Summer
1.1
1.2
1.3
1.4
1.5
1.6
1.7
HSI a
a,b b
b
HSI (
%)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
T
11-KT
b
a
b
a
xx,y y
z
Hor
mon
e co
ncen
tra�
on
(ng/
ml)
Fall Winter Spring Summer
A C
DB
Fig. 5. Reproductive seasonality in southern California hornyhead turbot females. Females from study locations sampled quarterly were measured for17b-estradiol (E2; panelA), gonadosomatic index (GSI; panelB), vitellogenin (VTG; panelB), hepatosomatic index (HSI; panelC), testosterone (T; panelD), and11-ketotestosterone (11-KT; panelD), as described inMethods.Different superscripts denote significantly differentmeans (p< 0.05);n¼ 71 (fall), 70 (winter), 90(spring), or 83 (summer).
Reproductive endocrine status in urban coastal marine fish Environ. Toxicol. Chem. 31, 2012 2695
Therefore, establishing how endogenous hormone levelschange throughout a given year in this species was of initialinterest. The results of the present study indicated that spring isthe season during which elevated sex steroid levels occur inboth male (testosterone and 11-KT) and female (E2 and andro-gens) hornyhead turbot, whereas fall was the principal seasonduring which the lowest hormone levels occurred. In both sexes,the spring peaks in these hormones were associated withcoincident changes in other reproductive parameters, includingGSI or plasma VTG concentrations in females.
Across the five seasonally sampled study sites (PV, PVF,OC, OCF, DP), distinct spring peaks in 11-KT were observed inmale hornyhead turbot at each site, and testosterone showedsimilar seasonality, although more variably. Preceding its
spring peak (in fall and winter), 11-KT concentrations werestrongly correlated with testosterone concentrations (r¼ 0.68–0.84); however, in spring and summer, the two androgens weregenerally not correlated. Because testosterone serves as theprecursor in the synthesis of 11-KT by the fish testis [22], thisfinding indicates that during the period of increasing androgensynthesis, 11-KT production is linked to testosterone produc-tion, whereas during the peak and post peak periods, thesteroidogenic pathways leading to 11-KT appear to decouple.Across all sites and seasons, significant differences in 11-KTwere not detected, although the androgen tended to be lower atthe OCF and OC sites. In contrast, testosterone concentrationswere lower at the OCF far-field site and to a lesser degree atPVF far-field site, as compared with WWTP outfall sites.
Table 2. Correlation coefficients (r) and p values of the relationships among reproductive parameters measured in hornyhead turbot females sampled fromdifferent study sites
Females Orange County Palos Verdes San Diego Santa Monica Bay
Season Far-field DP Far-field OCF Outfall OC Far-field PVF Outfall PV Outfall SD Outfall LA
E2 vs GSI Fall NS (7) NS (7) NS (12) NS (29) 0.58a (15) – –Winter NS (15) NS (1) NS (10) NS (25) NS (17) – –Spring NS (18) NS (9) 0.40d (26) NS (17) 0.41d (19) NS (16) 0.39a (27)Summer NS (10) NS (10) NS (20) 0.56a (22) NS (20) – –
All seasons NS (50) NS (28) 0.46c (68) 0.45a (93) NS (71) – –E2 vs VTG Fall NS (7) NS (7) NS (12) NS (29) 0.50a (15) – –
Winter NS (15) NS (1) 0.60a (10) 0.67c (25) 0.37d (17) – –Spring 0.56a (18) NS (9) 0.53a (26) 0.50a (17) 0.41d (19) NS (16) NS (27)Summer NS (10) NS (10) NS (20) 0.45a (22) NS (20) – –
All seasons NS (50) NS (28) 0.45c (68) 0.65c (93) 0.34b (71) – –GSI vs VTG Fall 0.82b (7) 0.61d (7) NS (12) 0.64c (29) 0.66b (15) – –
Winter 0.90c (15) NS (1) 0.67a (10) 0.40d (25) NS (17) – –Spring 0.57a (18) NS (9) 0.80c (26) NS (17) 0.60b (19) 0.42d (16) NS (27)Summer NS (10) 0.86c (10) 0.82c (20) 0.75c (22) 0.45a (20) – –
All seasons 0.80c (50) 0.50a (28) 0.87c (68) 0.38c (93) 0.40b (71) – –E2 vs T Fall NS (7) NS (7) 0.76b (11)e NS (29) 0.65b (15) – –
Winter 0.52a (13) NS (1) NS (10) 0.59b (25) 0.62b (17) – –Spring 0.42d (18) 0.73a (9)e NS (26) NS (17) 0.37d (19) 0.45d (16) 0.33d (27)Summer NS (10) NS (10) 0.55a (20) 0.63b (21) NS (20) – –
All seasons 0.40a (46) 0.40a (28) 0.46c (67) 0.40a (92) 0.69c (67)e – –T vs GSI Fall 0.75a (7) NS (7) NS (11) NS (29) 0.60a (15) – –
Winter 0.56a,g (13) NS (1) 0.69a,g (10) NS (25) NS (17) – –Spring NS (18) NS (9) NS (26)f 0.41d (17) NS (19)f NS (16) 0.47a (27)Summer NS (10) NS (10) NS (20) NSg (21) NS (20) – –
All seasons 0.37b,g (46) NS (28) 0.36b,g (67) �0.30a (92) NS (67)f – –T vs 11-KT Fall 0.93c (7) NS (7) 0.89c (11) 0.60b (29) 0.63a,g (15)
Winter 0.65a (13) NS (1) 0.67a (10) 0.38d (25) 0.61b (17)Spring NS (18) NS (9) 0.42a (26) NS (17) 0.50a (19) NS (16) NS (27)Summer NS (10) 0.62a (10) 0.73c (20) 0.57b (21) NS (20)
All seasons NS (46) 0.45b (28) 0.70c (67) 0.49c (92) 0.76c (66)HSI vs GSI Fall NS (7) 0.75a (7) NS (12) 0.44a (29) 0.60a (15) – –
Winter 0.78c (15) NS (1) 0.65a (10) NS (25) NS (17) – –Spring 0.64b (18) NS (9) 0.65c (26) NS (17) NS (19) 0.46d (16) 0.56b (27)Summer NS (10) 0.44d (10) 0.62b (20) 0.57b (22) 0.51a (20) – –
All seasons 0.55c (50) NS (28) 0.65c (68) NS (93) 0.37b (71) – –HSI vs VTG Fall NS (7) 0.64d (7) NS (12) NS (29) 0.52a (15) – –
Winter 0.80c (15) NS (1) 0.91c (10) 0.52b (25) NS (17) – –Spring 0.70c (18) NS (9) 0.73c (26) NS (17) NS (19) NS (16) NS (27)Summer NS (10) NS (10) 0.45a (20) 0.58b (22) NS (20) – –
All seasons 0.60c (50) 0.31d (28) 0.69c (68) 0.30b (93) 0.35b (71) – –
a Indicates significance level of correlation coefficient p¼ 0.05–0.01.b Indicates significance level of correlation coefficient p¼ 0.01–0.001.c Indicates significance level of correlation coefficient p < 0.001.d Indicates significance level of correlation coefficient p U 0.06–0.10.e E2 was also correlated with 11-KT in these three groups (r¼ 0.35–0.74, p< 0.05).f GSI was also correlated with 11-KT in these three groups (r¼ 0.53–0.54, p< 0.05).g T was also correlated with HSI in these groups (r¼ 0.39–0.68, p< 0.05), but not 11-KT.DP¼Dana Point; OCF¼ north Orange County; OC¼Orange County Sanitation District outfall; PVF¼ north Palos Verdes Peninsula; PV¼Los AngelesCounty Sanitation District outfall at Palos Verdes Peninsula; SD¼City of San Diego Metropolitan Wastewater Treatment Plant outfall; LA¼City of LosAngeles Hyperion Treatment Plant outfall in Santa Monica Bay; T¼ testosterone; 11-KT¼ 11-ketotestosterone; GSI¼ gonadosomatic index; HSI¼hepatosomatic index; E2¼ 17b-estradiol; VTG¼ vitellogenin; NS¼ nonsignificant.
2696 Environ. Toxicol. Chem. 31, 2012 J.A. Reyes et al.

Although the latter observation could imply an effect associatedwith the outfall discharges, testosterone levels were similar infish at the outfall sites and the DP reference site located farthestfrom any of the outfalls (and containing the lowest levels ofcontaminants among the study sites, as discussed further).Therefore, the findings do not support a linkage between the
spatial differences in testosterone concentrations in males andtheir presence at the WWTP discharge locations.
Although the testosterone and 11-KT concentrations in malehornyhead turbot were within normal ranges seen in males ofother fish species and also showed comparable seasonal changes([22–25]; J.A. Reyes, 2006, Master’s thesis, California State
0.0
0.2
0.4
0.6
0.8
E2 c
once
ntra
�on
(ng/
ml)
a,b b
a
b b
b
a
8 16 19 11 8 3 10 11 13 11 27 21 17 28 28 26 18 23 16 18 20 21 F W SP SU F W SP SU F W SP SU F W SP SU F W SP SU SP SP
Far-field Ou�all Far-field Far-field Ou�all Ou�alls DP OCF OC SD LA PVF PV
Orange County
Palos Verdes San Diego
Santa Monica Bay
‡ ‡
Fig. 6. Location-associated differences in 17b-estradiol (E2) levels in female hornyhead turbot. Seven southern California locations were compared in spring2006 and are represented as filled bars (City of Los Angeles Hyperion Treatment Plant outfall in SantaMonica Bay [LA], Los Angeles County Sanitation Districtoutfall at thePalosVerdesPeninsula [PV],OrangeCountySanitationDistrict outfall [OC], andCityofSanDiegoMetropolitanWastewaterTreatmentPlant outfall[SD],DanaPoint [DP], northOrangeCounty [OCF], andnorthPalosVerdesPeninsula [PVF]; seemap inFig. 1).Openbars represent the other three seasons,whichwere evaluated only at sites DP, OCF, OC, PVF, and PV. Data are presented as mean� standard error of mean ng/ml values for E2, with n shown below each bar.F¼ fall; W¼winter; SP¼ spring; SU¼ summer. The double dagger symbol (z) indicates mean is significantly lower than in other seasons at that site (p< 0.05).Different superscripts denote significantly different means between sites (p< 0.05).
0.0
0.2
0.4
0.6
0.8
1.0
0.0
0.5
1.0
1.5
2.0[A]
a,b
a a
a a,b
T co
ncen
tra�
on
(ng/
ml)
8 16 19 11 8 3 10 11 13 11 27 21 17 28 F W SP SU F W SP SU F W SP SU SP SP
30 26 18 23 16 18 20 21 F W SP SU F W SP SU
a
b
[B]
a,b
a a
b a,b
11-K
T co
ncen
tra�
on
( ng/
ml)
8 16 19 11 8 3 10 11 13 11 27 21 17 28 F W SP SU F W SP SU F W SP SU SP SP
30 26 18 23 16 18 20 21 F W SP SU F W SP SU
a
b
Orange County
Palos Verdes San Diego
Santa Monica Bay
Far-field Ou�all Far-field Far-field Ou�all Ou�alls DP OCF OC SD LA PVF PV
* *
*
*
*
**
Fig. 7. Location-associated differences in androgen levels in female hornyhead turbot. Seven southernCalifornia locationswere compared in spring 2006 and arerepresented as filled bars (City of Los Angeles Hyperion Treatment Plant outfall in SantaMonica Bay [LA], Los Angeles County Sanitation District outfall at thePalosVerdes Peninsula [PV],OrangeCounty SanitationDistrict outfall [OC], andCity of SanDiegoMetropolitanWastewater Treatment Plant outfall [SD], DanaPoint [DP], north Orange County [OCF], and north Palos Verdes Peninsula [PVF]; see map in Fig. 1). Open bars represent the other three seasons, which wereevaluatedonly at sitesDP,OCF,OC,PVF, andPV.Data are presented asmean� standard error ofmeanng/mlvalues for 11-ketotestosterone (11-KT; panelA) andtestosterone (T; panel B), with n shown below each bar. F¼ fall;W¼winter; SP¼ spring; SU¼ summer.Asterisk (�) indicates springmean is significantly higherthan in one or more of the other seasons at that site (p< 0.05). Different superscripts denote significantly different means between sites (p< 0.05).
Reproductive endocrine status in urban coastal marine fish Environ. Toxicol. Chem. 31, 2012 2697

University, Long Beach, CA, USA; D.M. Petschauer, 2008,Master’s thesis, California State University, Long Beach, CA,USA), plasma concentrations of E2 in male hornyhead turbotwere relatively high. This finding is in agreement with previouswork ([13,14]; J.A. Reyes, 2006, Master’s thesis, CaliforniaState University, Long Beach, CA, USA) demonstrating E2concentrations ranging between 300 and 1,400 pg/ml in horny-head turbot males sampled offshore of Los Angeles andOrange Counties, as compared with �50 pg/ml in males ofseveral other fish species tested. In the present study, plasmaE2 concentrations ranged between 200 and 900 pg/ml inmales, but did not show any seasonal patterns. The apparentlynormal seasonality in androgen levels in hornyhead turbotmales suggests that these E2 levels do not impair gonadalsteroid production or its seasonality. Whether the high E2levels in males of this species reflects a normal or abnormalcondition is not yet understood. In another species of flatfish,Platichthys flesus, Scott et al. [25] reported male plasma E2concentrations up to 220 pg/ml, which approaches the levelsobserved in this species; similarly, they found no seasonalityin E2 in male flounders, despite apparently normal levels andseasonality for 11-KT.
Concentrations of E2 in male hornyhead turbot exhibitedsignificant location-associated differences. In accordance withReyes et al. ([13]; J.A. Reyes, 2006, Master’s thesis, CaliforniaState University, Long Beach, CA, USA), male E2 concen-trations were lowest in fish from Orange County (OC and OCF)and relatively higher in fish from Palos Verdes (PV and PVF)and Santa Monica Bay (LA). However, higher E2 concentra-tions were observed in males from the DP reference site and atthe SD outfall site. The basis for these environment-associateddifferences is not clear, but again they did not coincide withresidence of fish at WWTP discharge locations. Despite thedifferent E2 levels by location, no detectable correlations withtesticular GSI were seen, nor did plasma VTG concentrationsexhibit any consistent relationships with E2. In a few subgroups,E2 was negatively correlated with VTG, whereas in othersubgroups the opposite was observed. The latter findingsmay suggest that either estrogenic or androgenic effects maypredominate at certain locations in the SCB, presumablydependent on the specific environmental constituents presentat each site. In addition, the spatial patterns in E2 and 11-KTwere similar in that their levels were relatively lower in OrangeCounty and highest at DP and SD. These two steroids showed asignificant positive correlation overall, and both use testoster-one as a precursor in their biosynthesis. Therefore, the spatialdifferences observed in the present study are possibly associatedwith factors that influence steroidogenic pathways, leading toconversion of testosterone into 11-KT as well as into E2.
Female hornyhead turbot exhibited a similarly timed repro-ductive cycle as compared with males, characterized by a springpeak and a fall decline in the reproductive indices measured. Inaddition to an overall spring peak in E2 concentrations, femalefish showed significant springtime increases in ovarian GSI aswell as in plasma VTG concentrations. Gonadosomatic indexand VTG were strongly correlated with each other, and theywere also positively correlated with E2. These findings point toa coordinated seasonal female reproductive system regulated byE2. When each of the study sites was evaluated separately,however, seasonality in E2 concentrations was not evident.Instead, the androgens showed more prominent seasonality,overall and within study sites. Although at significantly lowerconcentrations than in males, testosterone levels were signifi-cantly increased in winter and peaked in spring, and 11-KT
showed a similar pattern. Both androgens showed some modestpositive correlations with ovarian GSI. Testosterone and 11-KTwere correlated with each other in the females, which wasparticularly strong in the fall and winter seasons. Recent workby Forsgren et al. [26] indicates a potential role for androgens inregulating ovarian follicular development in salmon. Increasinglevels of testosterone and 11-KT in the winter-to-spring periods inhornyhead turbot (Fig. 5) would be consistent with such actions.
Across the study sites, mean E2 levels in female hornyheadturbot exhibited significant location-associated differences. Thehighest E2 levels were observed in females from DP and SD,and relatively lower levels were seen in females from OrangeCounty (OC and OCF) and PVF, a spatial pattern similar to thatobserved in the males E2 levels. The spatial pattern in bothsexes did not coincide with the locations of WWTP outfalldischarges, but the findings do appear to point to commonenvironmental or biological factor(s) influencing gonadal ster-oidogenesis and E2 production in both sexes. Correlationsbetween E2 and GSI and VTG in females were detectable insome subgroups (e.g., PV sites) but not detectable in others(e.g., OCF, DP), indicating possible spatial differences in E2action (e.g., on VTG production). Further research will berequired to understand these differences.
Although no significant differences were seen in age amongall study sites for both female and male hornyhead turbot,Forsgren et al. [19] noted smaller mean length and lowerspringtime gonadal maturity stage in fish sampled from theOCF study site as compared with fish from the other sites, whichall had similar springtime maturity (true in both sexes). There-fore, the sample of fish collected at the OCF site may have beenslightly less mature, which may help to explain the relativelylower springtime concentrations of E2 in females and testos-terone and E2 in males from this site. With the exception of theOCF group, however, endocrine measures did not correlate withfish size and age, likely a result of the limited range of fish sizechosen for evaluation in the study. Forsgren et al. [19] specu-lated that the proximity of petroleum-drilling platforms nearOCF and relatively higher levels of polycyclic aromatic hydro-carbons could be related to reduced maturity, which wouldrequire additional studies. Possibly the turbot population agestructure is different in the area where the OCF site is located, ascompared with other study sites.
As part of their annual cycle, both male and female horny-head turbot exhibited increases in HSI in spring and summer.Although this may represent seasonal differences in foodavailability, and therefore relatively increased hepatic fuelstorage [20] during these months, testosterone concentrationswere significantly correlated with HSI in both sexes. In females,GSI and VTG concentrations were also correlated with HSI andtestosterone. These findings suggest a role of androgens inadapting hepatic metabolism during the reproductive seasonin hornyhead turbot, actions that have been documented instudies of other fish species [27,28].
A large number of published studies have reported that fishresiding in WWTP-impacted environments may exhibit repro-ductive abnormalities, including altered levels of sex steroidsand VTG, as well as gonadal changes such as ovo-testisformation or aberrant expression of steroidogenic genes [29–35]. Although most research has been carried out in relativelyconfined aquatic locations (e.g., rivers or streams), the presentstudy was undertaken in an open coastal marine environment.Matthiessen [16] previously noted that endocrine disruptioneffects are likely to be more difficult to detect in marineenvironments, given that dispersion and dilution rates are
2698 Environ. Toxicol. Chem. 31, 2012 J.A. Reyes et al.

typically greater (e.g., of WWTP effluents). Offshore of OrangeCounty, previous research has been unable to demonstrateunequivocal effects of the Orange County Sanitation District(OSCD) outfall on plasma VTG or reproductive steroid levels,GSI, or in gonadal histology in flatfish ([13–15,35]; J.A. Reyes,2006, Master’s thesis, California State University, Long Beach,CA, USA; D.M. Petschauer, 2008, Master’s thesis, CaliforniaState University, Long Beach, CA, USA). Yet, fish sampledfrom the OCSD outfall area are well known to exhibit highertissue concentrations of several kinds of contaminants (e.g.,PCBs) typical at the discharge site.
In the paper by Maruya et al. [18], hornyhead turbot livercomposite samples were measured for a variety of contami-nants, in parallel to our biological measures of hornyhead turbotfrom the same study sites. Their results indicate that hornyheadturbot experience exposures to contaminants such as PCBs,dichlorodiphenyltrichloroethane, and polybrominated diphenylethers, which are greater in sediments and tissue of fish sampledfrom WWTP discharge locations as compared with the DPreference location. As described earlier, the observed spatialdifferences in the endocrine system of hornyhead turbot did notreflect this pattern. Although present in effluents, E2, E1,testosterone, or 17a-ethinylestradiol were not detectable inreceiving waters or sediments [17,18], and therefore environ-mental delivery of steroids to the fish (leading to alteredendogenous E2 or androgens) is not supported by the data.Among the list of analytes measured in liver, the only pharma-ceutical and personal care product detectable in hornyheadturbot was diazepam (trade name Valium), which was presentin 100% of the samples at a mean concentration of 56mg/kg[18]. The industrial and commercial compound 4-nonylphenolwas also frequently detected, at a mean concentration of102mg/kg. Based on previous experimental studies, both dia-zepam [36] and 4-nonylphenol [37] may impair gonadal ster-oidogenesis. However, the liver concentrations in hornyheadturbot from the DP reference location were comparable to thosefrom all four of the WWTP sites. Therefore, although someenvironment-associated differences in endocrine function weredocumented in hornyhead turbot in this study, the degree towhich environmental contaminants or WWTP discharges in thecoastal habitat are related is not clear. Further research will beneeded, particularly evaluations that can combine measure-ments of contaminant exposures and effects in the same sub-jects, to enable direct linkages to be made.
Acknowledgement—The present study was supported by the NationalSea Grant College Program, National Oceanic and AtmosphericAdministration, U.S. Department of Commerce, under University ofSouthern California Sea Grant number NA06OAR4170012, CFDA No.11.417, project CE-17, and by the California State Resources Agency. Theviews expressed herein do not necessarily reflect the views of NationalOceanicandAtmosphericAdministrationoranyof its sub-agencies.TheU.S.Government is authorized to reproduce and distribute copies forgovernmental purposes. The present study was also supported in part byfunds provided by City of Los Angeles, Sanitation Districts of Los AngelesCounty, Orange County Sanitation District, City of Oxnard and City of SanDiego. The authors would like to thank the numerous staff members from thedifferent agencies who helped in sampling the offshore fish populations,including from Orange County Sanitation District, City of Los AngelesBureau of Sanitation Environmental Monitoring Division, Los AngelesCounty Sanitation Districts, the City of San Diego Public UtilitiesDepartment, and the Southern California Coastal Water Research Project.We also thank K. Sak, D. Petschauer, K.R.E. Hagstrom, and A. Hamilton(California State University, Long Beach, CA, USA) for help in thelaboratory work. To the best of our knowledge, the authors do not have anyinterests or relationships, financial or otherwise, that might be considered apotential source of conflict of interest relating to the present study and itsdissemination.
REFERENCES
1. Lyon G, Stein E. 2009. How effective has the Clean Water Act been atreducing pollutant mass emissions to the Southern California Bight overthe past 35 years? Environ Monit Assess 154:413–426.
2. Orange County Sanitation District, 2009–2010. Orange CountySanitation District Ocean Monitoring Program Annual Report 2009-2010, Fountain Valley, California, USA.
3. City of Los Angeles, EnvironmentalMonitoringDivision. 2010.MarineMonitoring in Santa Monica Bay: Biennial Assessment Report for thePeriod January 2009 through December 2010. Department of PublicWorks, Bureau of Sanitation, Hyperion Treatment Plant, Playa del Rey,California, USA.
4. Schiff K, Allen JM, Zeng EY, Bay SM. 2000. Southern California.MarPollut Bull 41:76–93.
5. City of San Diego. 2011. Annual Receiving Waters MonitoringReport for the Point Loma Ocean Outfall, 2010. City of San DiegoOcean Monitoring Program, Public Utilities Department, Environ-mental Monitoring and Technical Services Division, San Diego,CA.
6. AllenMJ,Mikel T,CadienD,Kalman JE, Jarvis ET, SchiffKC,DiehlD,Moore SL, Walther S, Deets G, Cash C, Watts S, Pondella DJ II,Raco-Rands V, Thomas C, Ralph Y, Gartman R, Sabin L, Power W,Groce AK, Armstrong JL. 2007. Southern California Bight 2003Regional Monitoring Program. IV. Demersal Fishes and MegabenthicInvertebrates. Southern California Coastal Water Research Project,Costa Mesa, CA, USA.
7. Gravel A, Vijiyan MM. 2006. Salicylate disrupts interrenal steroido-genesis and brain glucocorticoid receptor expression in rainbow trout.Toxicol Sci 93:41–49.
8. Carr JA, Patino R. 2011. The hypothalamus-pituitary-thryoid axis inteleosts and amphibians: Endocrine disruption and its consequences tonatural populations. Gen Comp Endocrinol 170:299–312.
9. Alvarez-Cohen L, Sedlak DL. 2003. Emerging contaminants in water.Environ Eng Sci 20:387–388.
10. Snyder SA. 2008. Occurrence, treatment, and toxicological relevance ofEDCs and pharmaceuticals in water. Ozone Sci Eng 30:65–69.
11. Rempel MA, Schlenk D. 2008. Effects of environmental estrogens andantiandrogens on endocrine function, gene regulation, and health in fish.Int Rev Cell Mol Biol 267:207–252.
12. Roy LA, Armstrong JL, Sakamoto K, Steinert S, Perkins E, Lomax DP,Johnson LL, Schlenk D. 2003. The relationships of biochemicalendpoints to histopathology and population metrics in feralflatfish species collected near the municipal wastewater outfall ofOrange County, California, USA. Environ Toxicol Chem 22:1309–1317.
13. Reyes JA, GalimaMM, SakK, Schlenk D, Kelley KM. 2002. Endocrineandmetabolic parameters of stress andgrowth in two southernCaliforniaflatfish species subjected to varying environmental levels of municipalwaste. Integr Comp Biol 42:1300.
14. Rempel MA, Reyes J, Steinert S, HwangW, Armstrong J, Sakamoto K,Kelley KM, Schlenk D. 2006. Evaluation of relationships betweenreproductive metrics, gender and vitellogenin expression in demersalflatfish collected near the municipal wastewater outfall of OrangeCounty, California, USA. Aquatic Toxicol 77:241–249.
15. Deng X, Rempel MA, Armstrong J, Schlenk D. 2007. Seasonalevaluation of reproductive status and exposure to environmentalestrogens in hornyhead turbot at the municipal wastewater outfall ofOrange County, CA. Environ Toxicol 22:464–471.
16. Matthiessen P. 2003. Endocrine disruption in marine fish. Pure ApplChem 75:2249–2261.
17. Vidal-Dorsch DE, Bay SM, Maruya K, Snyder SA, Trenholm RA,Vanderford BJ. 2012. Contaminants of emerging concern in municipalwastewater effluents andmarine receivingwater.Environ Toxicol Chem31:2674–2682 (this issue).
18. Maruya KA, Vidal-Dorsch DE, Bay SM, Kwon JW, Xia K, ArmbrustKL. 2012. Organic contaminants of emerginc concern in sediments andflatfish collected near outfalls discharging treated wastewater effluent tothe Southern California Bight. Environ Toxicol Chem 31:2683–2688(this issue).
19. Forsgren KL, Bay SM, Vidal-Dorsch DE, DengX, Lu G, Armstrong JL,Gully JR, Schlenk D. 2012. Annual and seasonal evaluation ofreproductive status in hornyhead turbot atmunicipal wastewater outfallsin the Southern California Bight. Environ Toxicol Chem 31:2701–2710(this issue).
20. Haigwood JT, Flores RM, Mazloumi R, Ngan G, Kelley KM. 2000.Diabetogenic role of insulin’s counterregulatory hormones in theisletectomized, diabetic goby. Endocrine 13:273–280.
Reproductive endocrine status in urban coastal marine fish Environ. Toxicol. Chem. 31, 2012 2699

21. Goldberg SR. 1982. Seasonal spawning cycles of two Californiaflatfishes,Pleuronichthys verticalis (Pleuronectidae) andHippoglossinastomata (Bothidae). Bull Marine Sci 32:347–357.
22. Kusakabe M, Kobayashi T, Todo T, Lokman PM, Nagahama Y,Young G. 2002. Molecular cloning and expression during spermato-genesis of a cDNA encoding testicular 11b-hydroxylase (P45011b)in rainbow trout (Oncorhynchus kisutch). Mol Reprod Dev 62:456–469.
23. Garcia-Lopez A, Fernandez-Pasquier V, Couto E, Canario AVM,SarasqueteC,Martinez-RodriguezG. 2006.Testicular development andplasma sex steroid levels in cultured male Sengalese sole, Soleasenegalensis Kaup. Gen Comp Endocrinol 147:343–351.
24. Gemmell B, Pleskach K, Peters L, Palace V,Wautier K, Park B, DarlingC, Rosenberg B, McCrindle R, Tomy GT. 2011. Toxicokinetics oftetrabromoethylcyclohexane (TBECH) in juvenile brown trout (Salmotrutta) and effects on plasma sex hormones. Aquatic Toxicol 101:309–317.
25. Scott AP, Katsiadaki I, Kirby MF, Thain J. 2006. Relationship betweensex steroid and vitellogenin concentrations in flounder (Platichthysflesus) sampled fromanestuarycontaminatedwith estrogenicendocrine-disrupting compounds. Environ Health Perspect 114:27–31.
26. Forsgren KL, Young G. 2012. Stage-specific effects of androgens andestradiol-17beta on the development of late primary and early secondaryovarian follicles of coho salmon (Oncorhynchus kisutch) in vitro. BiolReprod DOI 10.1905/bioreprod.111.098772.
27. Holloway AC, Sheridan MA, Van der Kraak G, Leatherland JF. 1999.Correlations of plasma growth hormone with somatostatin, gonadalsteroid hormones, and thyroid hormones in rainbow trout during sexualrecrudescence. Comp Biochem Phys B 123:251–260.
28. Sangiao-Alvarellos S, Polakof S, Arjona FJ, Garcia-LopezA,Martin delRioMP,Martinez-RodriguezMP,Miguez JM,Mancera JM,Soengas JL.2006. Influence of testosterone administration on osmoregulation andenergy metabolism in gilthead sea bream Sparus auratus. Gen CompEndocr 149:30–41.
29. JoblingS,WilliamsR, JohnsonA,TaylorA,Gross-SorokinM,NolanM,Tyler C, van Aerle R, Santos E, Brighty G. 2006. Predicted exposures tosteroid estrogens in U.K. rivers correlate with widespread sexualdisruption in wild fish populations. Environ Health Perspect 114:32–39.
30. Hassanin A, Kuwahara S, Nurhidayat B, Tsukamoto Y, Ogawa K,Hiramatsu K. 2002. Gonadosomatic index and testis morphology ofcommon carp (Cyprinus carpio) in rivers contaminated with estrogenicchemicals. J Vet Med Sci 64:921–926.
31. Woodling JD, Lopez EM,Maldonado TA, Norris DO, Vajda AM. 2006.Intersex and other reproductive disruption of fish in wastewater effluentdominated Colorado streams. Comp Biochem Phys C 144:10–15.
32. Scott AP, Sanders M, Stentiford GD, Reese RA, Katsiadaki I. 2007.Evidence for estrogenic endocrine disruption in an offshore flatfish, thedab (Limanda limanda L.). Marine Environ Res 64:128–148.
33. Vajda AM, Barber LB, Gray JL, Lopez EM, Woodling JD, Norris DO.2008. Reproductive disruptions in fish downstream from an estrogenicwastewater effluent. Environ Sci Technol 42:3407–3714.
34. Vajda AM,Barber LB,Gray JL, Lopez EM,BoldenAM, ShoenfussHL,Norris DO. 2011. Demasculinization of male fish by wastewatertreatment plant effluent. Aquatic Toxicol 103:213–221.
35. BakerME, Ruggeri B, Sprague LJ, Eckhardt-Ludka C, Lapira J,Wick I,Soverchia L, Ubaldi M, Polzonetti-Magni AM, Vidal-Dorsch D, Bay S,Gully JR, Reyes JA, Kelley KM, Schlenk D, Breen EC, Sasik R,Hardiman G. 2009. Analysis of endocrine disruption in southernCalifornia coastal fishusing an aquaticmultispeciesmicroarray.EnvironHealth Perspect 117:223–230.
36. CalvoDJ,CamposMB,CalandraRS,Medina JH,RittaMN.1991.Effectof long term diazepam administration on testicular benzodiazepinereceptors and steroidogenesis. Life Sci 49:519–525.
37. Kortner TM, Arukwe A. 2007. The xenoestrogen, 4-nonylphenol,impaired steroidogenesis in previtellogenic oocyte culture of Atlanticcod (Gadus morhua) by targeting the StAR protein and P450sccexpressions. Gen Comp Endocr 150:419–429.
2700 Environ. Toxicol. Chem. 31, 2012 J.A. Reyes et al.




















