Comparative studies of the kinetic parameters of various algaenans and kerogens via open-system...
16
Pergamon Org. Geochem. Vol. 26, Nos. 11-12, pp. 705-720, 1997 © 1997 ElsevierScienceLtd. All rights reserved Printed in Great Britain PII: S0146-6380(97)00038-7 0146-6380/97 $17.00 + 0.00 Comparative studies of the kinetic parameters of various algaenans and kerogens via open-system pyrolyses DANIEL DESSORT ~, JACQUES CONNAW, SYLVIE DERENNE 2 and CLAUDE LARGEAU 2 ~Elf Aquitaine, CSTJF, 64018 Pau, Cedex, France and 2Laboratoire de Chimie Bioorganique et Organique Physique, UA CNRS 1381, ENSCP, 11 rue Pierre et Marie Curie, 75231 Paris, Cedex 05, France (Received 9 July 1996; returned to author for revision 14 October 1996; accepted 27 May 1997) A~traet--Kinetic parameters were determined for the first time, via open-system pyrolyses, on algae- hans (highly resistant biomacromolecules that are selectively preserved during kerogen formation) iso- lated from extant microalgae. Parallel studies were also carried out on 10 kerogens exhibiting, with one exception, a low level of maturity. These kerogens included samples chiefly derived from the selective preservation of the above algaenans and samples mainly, or almost exclusively, derived from the "natu- ral vulcanization" pathway. Important differences in activation energy (Ea) distributions were observed between the four algaenans investigated and correlated with their chemical structures. The kerogens predominantly derived from algaenan-selective preservation (Pula alginite, NE 70 and BJ 248 Torba- nites, Rundle Oil Shale) also exhibited pronounced differences in Ea distributions. These distributions provided: (i) information on the diversity of the source materials; and (ii) reflected the occurrence of im- portant differences in chemical structures and thermal behaviour between three of the tested kerogens, even though they are all classified as low maturity type I. The Kimmeridge Clay samples and the Lorca Oil Shale showed broad E~ distributions shifted to low energies when compared with the above algae- nans and kerogens. Such shifts reflect an important (or even almost exclusive for some of these kero- gens) contribution of materials originating from sulphur incorporation into various lipids during early diagenesis. Finally, the kinetic data derived for the nine low maturity fossil samples were extrapolated to a very low, geological heating rate of 3°C Ma -l and the generation rate curves and cumulative yield curves thus obtained were compared. © 1997 Elsevier Science Ltd Key words--open-system pyrolyses, algaenans, kerogens, kinetic parameters, activation energy distri- bution, selective preservation, natural vulcanization INTRODUCTION The kinetic parameters governing the thermal breakdown of kerogens have been extensively stu- died via laboratory experiments so as to derive values that can be subsequently used in basin mod- elling, for simulating kerogen maturation and pre- dicting the timing and extent of oil generation (e.g. Ungerer and Pelet, 1987; Hunt et al., 1991). To this end various heating methods have been used, including open-system pyrolyses (Braun et al., 1991; Tegelaar and Noble, 1994; Peters et al., 1995; Reynolds et al., 1995; Reynolds and Burnham, 1995; Schmidt, 1995). These experiments permit the calculation of activation energy distributions and generation curves for an assumed geological heating rate. Kerogens of different types have been studied in the last few years using such an approach. In some cases, the kinetic parameters thus derived were discussed in relation to the kerogen's chemical structure (Tegelaar and Noble, 1994), maceral com- position (Schmidt, 1995) or the influence of associ- ated mineral matter (Reynolds and Burnham, 1995). Recent studies (reviewed by de Leeuw and Largeau, 1993; Largeau, 1995; Largeau and de Leeuw, 1995) led to the recognition of a new type of biomacromolecule, characterized by an unusually high resistance to non-oxidative chemical degra- dation. Such non-hydrolysable macromolecular con- stituents were identified in the cell walls or the protective layers of a number of organisms, includ- ing microalgae. The resistant macromolecules, shown to comprise the outer walls occurring in nu- merous microalgal species, were termed "algaenans" (Tegelaar et aL, 1989). As reported in the three reviews mentioned above: (i) the chemical structure of several algaenans was elucidated via a combi- nation of spectroscopic and pyrolytic methods: (ii) high molecular weight lipids implicated as building blocks in algaenan formation were identified in a few cases; and (iii) parallel comparative studies with low maturity kerogens revealed an important role of algaenans in kerogen formation. In fact, this 705
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Comparative studies of the kinetic parameters of various algaenans and kerogens via open-system...

Pergamon Org. Geochem. Vol. 26, Nos. 11-12, pp. 705-720, 1997
© 1997 Elsevier Science Ltd. All rights reserved Printed in Great Britain
PII: S0146-6380(97)00038-7 0146-6380/97 $17.00 + 0.00
Comparative studies of the kinetic parameters of various algaenans and kerogens via open-system pyrolyses
D A N I E L D E S S O R T ~, J A C Q U E S C O N N A W , SYLVIE D E R E N N E 2 and C L A U D E L A R G E A U 2
~Elf Aquitaine, CSTJF, 64018 Pau, Cedex, France and 2Laboratoire de Chimie Bioorganique et Organique Physique, UA CNRS 1381, ENSCP, 11 rue Pierre et Marie Curie, 75231 Paris, Cedex 05,
France
(Received 9 July 1996; returned to author for revision 14 October 1996; accepted 27 May 1997)
A~traet--Kinetic parameters were determined for the first time, via open-system pyrolyses, on algae- hans (highly resistant biomacromolecules that are selectively preserved during kerogen formation) iso- lated from extant microalgae. Parallel studies were also carried out on 10 kerogens exhibiting, with one exception, a low level of maturity. These kerogens included samples chiefly derived from the selective preservation of the above algaenans and samples mainly, or almost exclusively, derived from the "natu- ral vulcanization" pathway. Important differences in activation energy (Ea) distributions were observed between the four algaenans investigated and correlated with their chemical structures. The kerogens predominantly derived from algaenan-selective preservation (Pula alginite, NE 70 and BJ 248 Torba- nites, Rundle Oil Shale) also exhibited pronounced differences in Ea distributions. These distributions provided: (i) information on the diversity of the source materials; and (ii) reflected the occurrence of im- portant differences in chemical structures and thermal behaviour between three of the tested kerogens, even though they are all classified as low maturity type I. The Kimmeridge Clay samples and the Lorca Oil Shale showed broad E~ distributions shifted to low energies when compared with the above algae- nans and kerogens. Such shifts reflect an important (or even almost exclusive for some of these kero- gens) contribution of materials originating from sulphur incorporation into various lipids during early diagenesis. Finally, the kinetic data derived for the nine low maturity fossil samples were extrapolated to a very low, geological heating rate of 3°C Ma -l and the generation rate curves and cumulative yield curves thus obtained were compared. © 1997 Elsevier Science Ltd
Key words--open-system pyrolyses, algaenans, kerogens, kinetic parameters, activation energy distri- bution, selective preservation, natural vulcanization
INTRODUCTION
The kinetic parameters governing the thermal breakdown of kerogens have been extensively stu- died via laboratory experiments so as to derive values that can be subsequently used in basin mod- elling, for simulating kerogen maturat ion and pre- dicting the timing and extent of oil generation (e.g. Ungerer and Pelet, 1987; Hunt et al., 1991). To this end various heating methods have been used, including open-system pyrolyses (Braun et al., 1991; Tegelaar and Noble, 1994; Peters et al., 1995; Reynolds et al., 1995; Reynolds and Burnham, 1995; Schmidt, 1995). These experiments permit the calculation of activation energy distributions and generation curves for an assumed geological heating rate. Kerogens of different types have been studied in the last few years using such an approach. In some cases, the kinetic parameters thus derived were discussed in relation to the kerogen's chemical structure (Tegelaar and Noble, 1994), maceral com- position (Schmidt, 1995) or the influence of associ-
ated mineral matter (Reynolds and Burnham, 1995).
Recent studies (reviewed by de Leeuw and Largeau, 1993; Largeau, 1995; Largeau and de Leeuw, 1995) led to the recognition of a new type of biomacromolecule, characterized by an unusually high resistance to non-oxidative chemical degra- dation. Such non-hydrolysable macromolecular con- stituents were identified in the cell walls or the protective layers of a number of organisms, includ- ing microalgae. The resistant macromolecules, shown to comprise the outer walls occurring in nu- merous microalgal species, were termed "algaenans" (Tegelaar et aL, 1989). As reported in the three reviews mentioned above: (i) the chemical structure of several algaenans was elucidated via a combi- nation of spectroscopic and pyrolytic methods: (ii) high molecular weight lipids implicated as building blocks in algaenan formation were identified in a few cases; and (iii) parallel comparative studies with low maturity kerogens revealed an important role of algaenans in kerogen formation. In fact, this
705

706 D. Dessort et al.
g 7
- ~ - ~ . ~
g. -
g ~
v
©
o ~
~ - o
~ ..~.~
type of biomacromolecule appears also to exhibit an extremely high resistance to bacterial degra- dation and, as a result, they can escape diagenetic alterations to be retained nearly unaltered in kero- gens. Algaenan fossilization, therefore, occurs through the so-called "selective preservation" path- way and such macromolecules, thus, afforded a major contribution to a number of algal kerogens (Largeau and Derenne, 1993). It cannot be comple- tely excluded that algaenans undergo some altera- tions in chemical structure during the drastic treatments required for isolation from extant micro- algae and/or during the first steps of diagenesis. Nevertheless, the tight relationships based not only on bulk chemical features but also on extended comparative examinations of pyrolysis products observed between isolated algaenans and low maturity kerogens indicate that the occurrence of major alterations can be ruled out.
In spite of the important geochemical role of algaenans, no study dealing with the kinetic par- ameters of these non-hyrolysable diagenetically-re- sistant biomacromolecules has so far been reported to the best of our knowledge, In the present work, open-system pyrolyses were carried out on four algaenans isolated from extant microalgae and on kerogens known to be chiefly derived from the selective preservation of such highly resistant bio- macromolecules. For comparison, kerogens having only a relatively low (or even negligible) contri- bution of selectively-preserved components, whereas the "natural vulcanization" pathway played a sub- stantial to virtually exclusive role, were also exam- ined; namely five Upper Jurassic samples from the Kimmeridge Clay Formation and a Messinian kero- gen from the Lorca Basin. In recent years the so- called "natural vulcanization" pathway has been shown to play a major role in the formation of a number of kerogens (reviewed in Sinninghe Damst6 and de Leeuw, 1990). This pathway results in the formation of high molecular weight, diagenetically- resistant, sedimentary organic matter via the incor- poration of sulphur into various functionalized lipids.
The main purposes of the present study were: (i) to examine the relationships between the chemical structure of various algaenans and their kinetic par- ameters as obtained from open-system pyrolyses; (ii) to compare the above kinetic parameters with those of algaenan-derived kerogens so as to deter- mine the chemical features that control the thermal behaviour of these fossil materials; (iii) to compare the kinetic parameters of kerogen fractions derived from the selective preservation and the "natural vulcanization" pathways; and (iv) to examine the natural evolution of these kerogens via extrapol- ation of laboratory kinetic data to very low, geo- logical heating rates.

Kinetic parameters of algaenous and kerogens 707
EXPERIMENTAL
Samples
The studied algaenans were isolated from Botryococcus braunii and Scenedesmus quadricauda. Laboratory cultures of these green microalgae were grown under air-lift conditions (Casadevall et al., 1985); algaenan isolation via successive, drastic, base and acid hydrolyses on the lipid-free biomass was carried out as previously described (Berkaloff et al., 1983).
The bulk chemical features of the tested fossil samples are reported in Table 1. Three lacustrine organic-rich deposits chiefly composed of the ac- cumulation of fossil colonies of B. braunii, including a sample from the massive section of the Pula Oil Shale (Pliocene, Hungary), two Torbanites, NE 70 (Permian, Winkelhoak mine, Evander area, South Africa) and BJ 248 (Permian, Glen Davis, Sydney Basin, New South Wales, Australia) (Derenne et al., 1994) were used. A sample of the Rundle Oil Shale (Eocene, lacustrine, Kerosene Creek seam, Rundle, Queensland, Australia), known to be formed mainly by the accumulation of ultralaminae (Largeau et al., 1990), five samples from the Kimmeridge Clay Formation (Upper Jurassic, mar- ine, Marton 87 borehole, Pickering Vale, Cleveland Basin, Yorkshire, U.K.) previously studied by Boussafir et al. (1995a) and a sample of the Lorca Oil Shale (Messinian, marine, pre-evaporitic, Lorca, Murcia, Spain) (Derenne et al., 1995) were also used.
Kinetic studies
Previous pyrolysis (Pyromat) studies have shown generally good agreement between the kinetic par- ameters obtained from bitumen-free whole rocks and the corresponding isolated kerogens, provided that samples with TOC > 1% were examined (Tegelaar and Noble, 1994; Reynolds and Burnham, 1995). The present study was, therefore, carried out on pre-extracted samples (finely pow- dered material stirred at room temperature over- night in CH2CI2/MeOH, 2/1, v/v). Kinetic parameters were determined via open-system, pro- grammed temperature pyrolyses. These experiments were carried out under a flow of helium from 250 to 650°C, with a Pyromat II micropyrolyser (Lab Instruments, Kenwood, CA, U.S.A.). Kinetic data were obtained from multiple runs with at least three different constant heating rates between 1- 25°C min -1 for each sample. Heating rates below I°C were not used because they resulted in low sig- nal to noise ratios and to avoid problems related to the lower volatility of the pyrolysis products (Tegelaar and Noble, 1994). The bitumen-free samples (2-3 mg of organic matter), were loaded in a 3 mm i.d. quartz crucible. In situ temperature measurements were carried out in the centre of the
material with a calibrated thermocouple. The evol- ution of the total volatile products thus generated (except CO2 and H20) was measured using a flame ionization detector (FID).
These experiments afforded several profiles from a given kerogen or algaenan corresponding to the rate of evolution of pyrolysis products vs tempera- ture for the different heating conditions. Each pro- file was characterized by its Tmax value (temperature at which the maximum rate of evol- ution takes place) and its width at half-height (Reynolds et al., 1995). Kinetic parameters of gen- eration of volatile products were calculated for each sample from the above profiles using a discrete model. The multiplicity of the reactions implicated in the pyrolysis of such materials was replaced at each heating rate by 30 independent reactions. Further simplification was obtained by considering that all these parallel, non-isothermal, first-order pseudo-reactions have the same frequency factor (Ungerer and Pelet, 1987). Indeed, due to the com- plexity of pyrolysis reactions this factor would merely correspond to a mathematical optimization parameter bearing no relationship with molecular vibrational frequencies (Schaefer et al., 1990). The validity of the parallel reaction model with a single frequency factor was recently tested (Burnham et al., 1995), and such approximations were shown to be adequate except for very immature samples with high oxygen levels.
Thirty-one parameters were, therefore, considered and optimized by an iteration method to obtain the best-fit between the measured and calculated rate curves. The quality of the fit for these curves was judged from the difference between experimental and calculated Tmax values. A discrete distribution of activation energies (Ea), with a regular spacing of 1 kcal mol - l in the range 40 to 70 kcal mo1-1 and one frequency factor were thus obtained for each kerogen and algaenan. Calculations of kinetic par- ameters were carried out by using the regression analysis program KINETICS (Burnham et aL, 1987). The reproducibility was typically +2 kcal mo1-1 for Ea and +I°C for Tmax. Comparison of kinetic parameters can be biased when the frequency factors calculated for the stu- died samples are sharply different. This is due to possible compensation effects, since a low (high) fre- quency factor can compensate for low (high) acti- vation energies. To avoid such a problem, rate constants were calculated for each sample at 450°C (temperature close of the averaged Tmax observed in these laboratory heating experiments).
As stressed by Tegelaar and Noble (1994), com- parisons of the kinetic data for different samples are easier from cumulative yield curves (transform- ation ratios into volatile pyrolysis products vs tem- perature). These generation profiles were, therefore, established for the different laboratory heating rates

708 D. Dessort et al.
P R B A
i I % A = 7 . 1 E+15 s-1
80 . . . . . . . . . . . . .
60 . . . . . . . . . . . . . . . .
40 . . . . . . . . . . . . . . . .
20 . . . . . . . . . . . . .
(DqD
Kca l I m o l e
P R B B 00
% , A = 3 . 0 E + l T s - 1
80 . . . . . . . . . . . . . . . . . . . .
60 . . . . . . . . . . . . . . .
40 . . . . . . . . . . . . . . . . . . .
20 . . . . . . . . . . . . . . .
0 , . . , . % . , , , . . . , , , , , , , ,
Kca l / m o l e
P R B L 100
% A = 3.6 E+16 s-1
80
40
20
0
Kca l I m o l e
Fig. 1. Distribution of activation energies and frequency factor (A) of the algaenans of the three races of B. braunii.
used and extrapolated to a low geological value of 3°C Ma -1. The latter value was also used for simu- lating rate profiles under geological conditions.
RESULTS AND DISCUSSION
B. braunii algaenans and Botryococcus-derived kero- gens
B. braunii is characterized by a high efficiency for biosynthesizing very large amounts of lipids, and three distinct chemical races producing different types of hydrocarbons have been identified in this species (Metzger et al., 1991); namely A race (n- alkenes), B race (botryococcenes), L race (a lycopa-
diene). The algaenans isolated from the three races were termed PRB A, PRB B and PRB L, respect- ively.
The discrete distributions of activation energies and the frequency factors obtained for the non- hydrolysable macromolecular materials isolated from the different races of B. braunii are reported in Fig. 1. As an example, the experimental and calcu- lated rate profiles corresponding to the different heating rates used for determining E~ distribution are shown for PRB A in Fig. 2. The activation energy distributions of B. braunii algaenans are all characterized by one predominant energy in the high range. Such a predominance is especially pro- nounced for PRB A, for which the distribution is
1°C / m n 2 ° C / m n
6 ,. E x p e r i m e n t a l c u r v e s Jzt~f~/~ 5 °C / m n . . . . ca,ou,=eoc° os
5 ] ." .
2 -" . / . .
250 3so 45o 550 650
T ( ° C )
Fig. 2. Experimental and calculated generation rate curves obtained for PRB A at different laboratory heating rates.

Kinetic parameters of algaenous and kerogens 709
almost exclusively (95%) represented by a single Ea value. The principal energy accounts for about 85 and 60% of the reaction in the case of PRB B and PRB L, respectively. For the latter algaenan, about one-third of the reaction is also described by rela- tively low activation energies in the 45- 60 kcal mol - l range. These kinetic parameters and their differences are correlated with the chemical structure of the algaenans.
The monoenergetic distribution of PRB A reflects an homogeneous structure based on a single type of building block exhibiting a relatively simple struc- ture, so that the thermolysis of the macromolecular structure would originate from the primary cleavage of the same type of bond with a similar environ- ment. Indeed, it was recently established (Gelin et al., 1994a) that the major building block of PRB A is an aliphatic polyaldehyde. Further condensation and/or reticulation of this soluble material leads to the formation of the insoluble and non-hydrolysable algaenan.
Similar bulk chemical features (H/C and O/C atomic ratios, FTIR and solid state 13C N M R spec- tra) were observed for PRB A and PRB B (Kadouri et al., 1988), and their pyrolysates are both domi- nated by n-alkane/n-alk- l -ene doublets originating from the thermal cleavage of polymethylene chains. In addition, recent studies on the high molecular weight lipids of the B race (Metzger et al., unpub- lished data) indicate that: (i) the aliphatic polyalde- hyde, mentioned above for PRB A, is also likely to play an important role in PRB B structure; and (ii) tetramethylsqualene is probably implicated, via ketal formation with these polyaldehyde units. Spectroscopic and pyrolytic examination of the algaenan of the L race (Derenne et al., 1989; Derenne et al., 1990) revealed substantial differences in bulk chemical features when compared with those of the A and B races. Isoprenoid chains with a C4o lycopane-type skeleton were, thus, shown to be implicated in the formation of PRB L, instead of unbranched chains for PRB A and PRB B. Some of these lycopane chains in PRB L are probably cross- linked by ether bridges (Gelin et al., 1994b; Behar et al., 1995). More recently, it was observed (Metzger et al., unpublished data) that: (i) the ali- phatic polyaldehyde also affords a substantial con- tribution to PRB L; and (ii) some polyaldehydic units are probably linked to lycopadiene chains via ketal formation. The involvement of the same poly- aldehyde in the formation of the algaenans of the three races of B. braunii is consistent with the pre- sence of principal energies around the same value in their corresponding activation energy distributions. The relatively broad Ea distributions observed for the algaenans of the B and L races likely reflects a more complicated macromolecular structure due to the occurrence of tetramethylsqualene and lyco- pane-type chains, respectively, and of ketal bridges.
MARTON U . I ~ MARTON I1-1 ~ ~ l ~ " - - ~ \ KWm~ndpT ~ ¥ / ~ A . ~ , • \ KImo~dg* • _ ~ - - ~ W " ~ L , i l l i p l ~ 111 \ Klmrneddge M J'w,~ -- J~lBm,-'-,w~,Ll~,~ \
~ , o, . , , , , ~ s ~ " ~ m r ~ P ~ j P ~ [ I . l r J H /7 /-80.0 RunaZe e3 l~S - ' l P m Y . , I g i l l m w ~ ~ l l l u ~ l [ i l j r I/- so.o
p . , . . ,o f . . .~ fP~v-~s~p . - -~ j / / -40 .0 -.... _ ~ ~ , d ~ ' ~ , , m ~ t l * m J F |/-20.0 PR s, ] ~ " - ~ ~ t m ~ " A d t , A g . ; PRB L I L W ~ a ~ P ~ - ~ ~ ....
k4S0
Fig. 3. Rate constants calculated at 450°C for the studied algaenans and fossil materials.
In addition, the presence of ether bridges between some lycopane chains was noted in PRB L and all the carbon atoms in the central part of the lycopane skeleton are implicated in the formation of such bridges (Behar et al., 1995). The PRB L structure, thus, comprises both tertiary and quaternary ether links and the latter would be expected to exhibit a lower thermal stability. The presence of these bridges could, therefore, account, at least in part, for the low energy range portion in the activation energy distribution of PRB L. The latter feature may also be related to the contribution of isopre- noid chains in this algaenan, since it is well docu- mented that such branched chains are more sensitive to thermal stress when compared with polymethylene chains. The lower thermal stability of PRB L is also illustrated by the higher reaction rate at 450°C (Fig. 3) when compared with those of PRB A and PRB B.
The lower stability of PRB L relative to PRB B was previously observed in artificial maturation ex- periments carried out in sealed gold tubes (Behar et al., 1993). While no kinetic parameters were de- rived from the above study it was found, for example, that conversion yields were of 48 and 16 wt% under isothermal conditions (300°C 3 h -1) from PRB L and PRB B, respectively. Interestingly, it was noted that the conversion yields calculated for the two algaenans from their energy distri- butions are lower than the experimental values obtained from the closed-system pyrolyses. This is likely due to the differences between open and closed pyrolyses, especially the nature of the detected products and the mechanism of the pri- mary cracking reactions.

710 D. Dessort et al.
Pule Alginite NE 70 Torbanite BJ 248 Torbanite
% / A=4.4E+15s-1 % A=3.9E+14s-1
80 t . . . . . . . . . . . . . . . . . . . . . 8o . . . . . . . . . . . . . . . . . 8o . . . . . . . . . . . . . . . . . . . . . . .
60 - 60. 60 . . . . . . . . . . . . .
, o . , o . , o . . . . . . . . . . . . . . . . . .
20 - 20 • 20
o
m
o . . . ~ . ~ . ; . . a . . . . . L . . . , , . . . . . o.
Kcal I mole Kcal / mole Kcal I mole
Fig. 4. Distribution of activation energies and frequency factor (A) of Botryococcus-derived kerogens.
Pyromat pyrolyses were also carried on Botryococcus-derived kerogens in order to compare their kinetic features with those of the algaenans discussed above. The first sample examined was the maar-type Pliocene oil shale from Pula. Previous palynologieal observations (Jambor and Solti, 1975) revealed an abundant contribution of fossil colonies of Botryococcus to Pula alginite and scanning elec- tron microscopy indicated excellent morphological preservation of the colonies in this low maturity type I material (Solti, 1985). Prolific growth of Botryoeoccus took place in the Pula crater and the total thickness of the organic-rich alginite deposit is about of 40m (Brukner-Wein et al., 1991). While the discrete distribution of activation energies obtained from the bitumen-free Pula alginite appears relatively broad (Fig. 4) it is dominated (75%) by a single value (60 kcal tool-l). These kin- etic parameters were compared with those obtained for the algaenans of the three races of B. braunii, taking into account the information derived from recent organic geochemical studies on the Pula deposit (Derenne et al., 1994, 1997). The kinetic features of the Pula sample, thus, appear to be fully consistent with these studies indicating that: (i) both the A and L races of Botryococcus were present, with the A race providing a predominant input; and (ii) that PRB A and (to a lesser extent) PRB L played a major role in the formation of this alginite by selective preservation.
The NE 70 Torbanite deposit is extremely rich in organic matter (ash content below 2%) composed of low maturity type I kerogen. Previous examin- ation by UV fluorescence microscopy showed that the bulk of this material corresponds to tightly packed colonies of Botryococcus cemented by a see- mingly amorphous organo-mineral matrix. Bulk spectroscopic examinations, identification of kero-
gen pyrolysis products and bitumen analyses (Largeau et al., 1984, 1986; Derenne et al., 1987, 1988, 1994; Kister et al., 1990) indicated that the NE 70 Torbanite was formed via the fossilization of the A and/or B race(s). The activation energy distri- bution from this sample (Fig. 4) is characterized by the presence of a single energy in the high value range, as observed for PRB A. However, the kinetic parameters of the NE 70 Torbanite could also be related to PRB B based on: (i) the reproducibility typically observed for Ea (_+2 kcal mol -]) in these experiments; and (ii) the possible occurrence of compensating effects originating from large differ- ences in frequency factors. Close k450 values are also observed for the Torbanite and the two resist- ant biopolymers (Fig. 3). Accordingly, the kinetic parameters do not allow clear discrimination between the two algaenans as the source of the Torbanite, although the presence of a single acti- vation energy makes an input from the A race more likely and indicates an overwhelming role for algae- nan selective preservation in the genesis of this extremely organic matter-rich deposit. Such a monoenergetic distribution was quite unexpected since kerogens usually exhibit a highly hetero- geneous structure, even when sharply dominated by a single source organism. Thus, micro-FTIR exam- ination of a set of low maturity Torbanites (Landais et al., 1993) (not including the NE 70) revealed a completely different composition of the organic matter in the cementing organo-mineral matrix when compared with the colonies. Furthermore, this matrix comprises two markedly distinct types of organic matter corresponding to degraded algal and bacterial constituents. In sharp contrast, based on the present kinetic observations, the degree of heterogeneity appears negligible in the NE 70 sample. Tegelaar and Noble (1994) also

Kinetic parameters of algaenous and kerogens
PR sq 40 %
4O %
Rundle 0il shale
711
30 30
20 20
10 10
0 0
Kcal I mole Kcal I mole
Fig. 5. Distribution of activation energies and frequency factor (A) of the algaenan of S. quadricauda and of the Rundle Oil Shale.
noted a single activation energy for the Green River Shale kerogen and considered this to reflect a homogeneous polymethylenic structure. Likewise, narrow distributions consisting of only three adja- cent energies were observed from alginite-rich lacus- trine sediments (Horsfield et al., 1994). As emphasized by Tegelaar and Noble (1994), compari- son of kinetic parameters with literature data can only be made if the same sample is analysed. It is interesting to note, however, that a single, high acti- vation energy value was also previously obtained from an Australian low maturity Permian Torbanite via a discrete model (Tegelaar and Noble, 1994), whereas a substantially lower Ea value was calculated via a Gaussian model.
The second tested Torbanite, BJ 248, also appears to be composed of an accumulation of Botryococcus colonies when observed by UV fluor- escence microscopy and, based on kerogen and bitumen analyses (Derenne et al., 1987, 1988), the A and/or B race(s) of B. braunii was (were) impli- cated in its formation. In contrast to the NE 70 deposit, however, BJ 248 is mature enough to be in the oil window. Comparative spectroscopic and pyr- olytic studies of these two Torbanites (Derenne et al., 1987, 1988; Kister et al., 1990) showed that, as expected, thermal evolution resulted in: (i) a partial elimination of the oxygen groups; (ii) a progressive increase in aromaticity (aromaticity fraction, fa, of 0.58 determined by solid-state 13C NMR); and (iii) the shortening and release of some of the hydro- carbon chains. Nevertheless, as demonstrated by its spectroscopic features and by the study of its pyrol- ysis products the BJ 248 Torbanite retains a sub- stantial oil potential and long hydrocarbon chains still afford an important contribution to the kero- gen's macromolecular structure. The activation
energy distribution for BJ 248 (Fig. 4) extends from 45 to 67 kcal mo1-1 and the principal energy, at 56 kcal mol -~, accounts for about 60% of the reac- tion. The difference between the principal E~ of the BJ 248 and NE 70 Torbanites might not be as high as suggested by the calculated distributions and might reflect compensation due to frequency fac- tors. Indeed, it is well known that a lower acti- vation energy can be compensated by a lower frequency factor and both Ea and A values are much lower for the BJ 248 sample. It should, never- theless, be noted that: (i) the rate constants calcu- lated from these kinetic parameters for the two Torbanites are sharply different (Fig. 3); and (ii) the main point here is the pronounced difference in Ea distributions, that is, an extended range for BJ 248 instead of a single energy for the NE 70 sample. The latter feature cannot be accounted for by com- pensation effects. Comparison of activation energy distributions was recently carried out on bitumen- free source rock samples from the Lias e formation (Lower Toarcian, Germany) with different levels of maturity (Schaefer et al., 1990). Their study showed a selective cleavage of more thermally labile bonds during natural evolution with the mature samples exhibiting a narrower distribution and a shift to higher energy values. In sharp contrast, reverse trends were observed here on comparison of the BJ248 and NE 70 samples. Accordingly, the observed differences between the kinetic parameters of these two Torbanites should not chiefly reflect their relative levels of evolution but some intrinsic differences in their source materials. As stressed above, the NE 70 sample shows a quite unusual homogeneity and is almost exclusively composed of selectively preserved B. braunii algaenan. In con- trast, the BJ 248 Torbanite exhibits a higher degree

712 D. Dessort et al.
of heterogeneity (Landais et al., 1993). Various sources were, thus, probably important contributors along with PRB A and/or PRB B in the formation of the BJ 248 kerogen. Indeed, comparison of the fatty acids released on pyrolysis by the NE 70 and BJ 248 samples (Derenne et al., 1988) indicated that diagenetic incorporation of some degraded lipids occurred during the formation of the latter Torbanite.
S. quadricauda algaenan (PRsq) and Rundle Oil Shale
No predominantly single value was observed in the discrete distribution of activation energies obtained for the algaenan of the ubiquitous fresh- water microalga S. quadricauda (Fig. 5). This distri- bution extends from 46 to 67 kcal tool -~, and the principal energy at 60 kcal tool -1 only accounts for approximately 35% of the reaction. Based on such kinetic features, a rather heterogeneous structure can be inferred for PRsq. To date, the high molecu- lar weight lipids of S. quadricauda have not been examined; accordingly, no precise information is available on the building block of the a|gaenan. Nevertheless, spectroscopic and pyrolytic studies on PRsq (Derenne et al., 1991) indicated, as in the case of the algaenans of the A and B races of B. braunii, a highly aliphatic nature and a macromolecular structure based on long unbranched hydrocarbon chains, which are probably cross-linked at least in part by ether bridges. An important difference con- cerned nitrogen functions in PRB A and PRB B, on the one hand, and S. quadricauda algaenan, on the other hand. The latter, which contains 2.4 wt % of nitrogen instead of approximately 0.5% for the PRBs, comprises alkylamide groups. The thermal cleavage of these groups results in the formation of n-alkylnitriles with a bimodal distribution (Derenne et al., 1991, 1993). In sharp contrast, no nitrile pro- duction was observed, even in trace amounts, on pyrolysis of B. braunii algaenans. The presence of these amide functions should explain, in part, the far more heterogeneous structure of PRsq when compared with the latter algaenans, as reflected by its discrete Ea distribution.
The Rundle Oil Shale, when observed by trans- mission electron microscopy at high magnification, appears to be composed of accumulations of very thin lamellar structures associated into bundles. Such very thin (approximately 15nm) structures ("ultralaminae", Largeau et al., 1990) were also detected in a number of other kerogens. Parallel studies have shown that numerous extant microal- gae, among them S. quadricauda, contain very thin, algaenan composed outer walls (reviewed in Derenne et al., 1992a). In addition, examination of the algaenans isolated from some of the above species (Derenne et al., 1992b,c) revealed the same chemical features as in the case of PRsq, including
the formation of n-alkylnitriles on pyrolysis. Indeed, the occurrence of tight morphological and chemical correlations (Derenne et al., 1991, 1992b,c) showed that fossil ultralaminae were formed via the selective preservation of algaenan composed very thin outer walls of green microalgae like S, quadricauda. The kerogen from the Rundle Oil Shale is characterized by a broad distribution of activation energies from 45 to 67 kcal mol -t , with the principal energy only accounting for about one- third of the reaction (Fig. 5). Very close features were previously observed in studies of various Rundle Oil Shale samples (Tegelaar and Noble, 1994; Reynolds and Burnham, 1995) for the general shape of E a distributions, the principal activation energy (57 kcal mo1-1 instead of 58), the frequency factor (8.2 × 1014 S I instead of 1.8 x 1015) and Tm~ (481.5°C instead of 482 for a heating rate of 25"C rain-l).
Comparison of the Rundle Oil Shale sample with PRsq (Fig. 5) also showed close similarites in Ea distributions. This agreement is consistent with the above-described mode of formation of ultralaminae and with a major contribution of such structures in the kerogen of the Rundle Oil Shale. Comparisons of the activation energy distributions showed im- portant differences between the kerogen of the Rundle Oil Shale on the one hand, and Botryococcus-derived kerogens (Pula alginite and NE 70 Torbanite), on the other. Nevertheless, all these kerogens are characterized, based on bulk chemical composition, as low maturity type I. Accordingly, as already stressed by Tegelaar and Noble (1994), such similarities do not preclude large differences in chemical structures that are revealed by kinetic studies and should have major implications for the timing of oil formation.
Kimmeridge Clay kerogens
The Kimmeridge Clay is a low maturity marine deposit, composed of type II and I-S kerogens according to its bulk features and considered to be a lateral equivalent of the main source rocks of the North Sea. This formation is characterized by short-term, parallel, wave-shaped cyclic variations (microcycles), corresponding to about 30,000y, which are reflected in both kerogen quantity (TOC) and quality (hydrogen index) (Herbin et al., 1991, 1993). The origin of such variations was recently determined via a combination of morphological and chemical studies (Boussafir et al., 1995a). Kimmeridge Clay kerogens were, thus, shown to be chiefly composed of three distinct types of organic matter: (i) "orange OM" is nanoscopically amor- phous and originates from the sulphurization of lipids, mostly of algal origin; (ii) "brown OM" is chiefly composed of ultralaminae and, as discussed in the previous section, it originates from the selec- tive preservation of very thin resistant outer walls
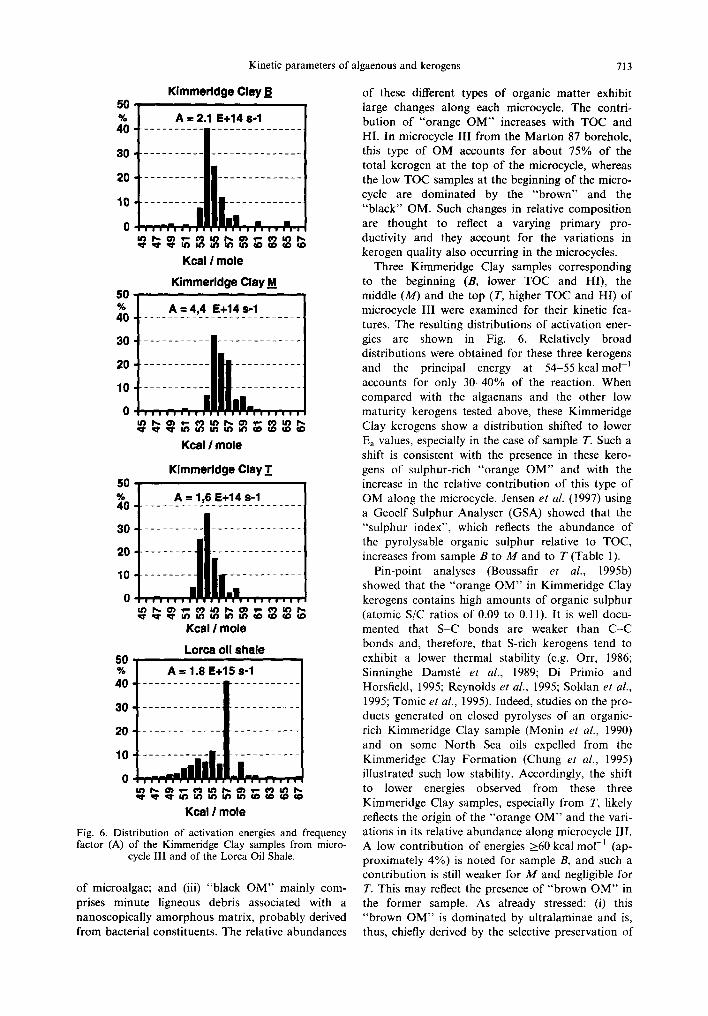
Kinetic parameters of algaenous and kerogens
Kimmeridge Clay B 50,
30
2O
10
0
Kcal / mole
Kimmeridge Clay M
40
30
20
10
0
Kcal I mole
Klmmerldge Clay ! 5O
~ o = "
30
20
10
0
5O % 40
30
20
Kesl I mole
Lores oli shale
A = 1.8 E+15 s-1 . . . . . . . . . . . . . . . . . . . . . . . . .
10
Kesl I mole
Fig. 6. Distribution of activation energies and frequency factor (A) of the Kimmeridge Clay samples from micro-
cycle III and of the Lorca Oil Shale.
of microalgae; and (iii) "black OM" mainly com- prises minute ligneous debris associated with a nanoscopically amorphous matrix, probably derived from bacterial constituents. The relative abundances
713
of these different types of organic matter exhibit large changes along each microcycle. The contri- bution of "orange OM" increases with TOC and HI. In microcycle III from the Marton 87 borehole, this type of OM accounts for about 75% of the total kerogen at the top of the microcycle, whereas the low TOC samples at the beginning of the micro- cycle are dominated by the "brown" and the "black" OM. Such changes in relative composition are thought to reflect a varying primary pro- ductivity and they account for the variations in kerogen quality also occurring in the microcycles.
Three Kimmeridge Clay samples corresponding to the beginning (B, lower TOC and HI), the middle (M) and the top (T, higher TOC and HI) of microcycle III were examined for their kinetic fea- tures. The resulting distributions of activation ener- gies are shown in Fig. 6. Relatively broad distributions were obtained for these three kerogens and the principal energy at 54-55kcalmol -~ accounts for only 30-40% of the reaction. When compared with the algaenans and the other low maturity kerogens tested above, these Kimmeridge Clay kerogens show a distribution shifted to lower Ea values, especially in the case of sample T. Such a shift is consistent with the presence in these kero- gens of sulphur-rich "orange OM" and with the increase in the relative contribution of this type of OM along the microcycle. Jensen e t al. (1997) using a Geoelf Sulphur Analyser (GSA) showed that the "sulphur index", which reflects the abundance of the pyrolysable organic sulphur relative to TOC, increases from sample B to M and to T (Table 1).
Pin-point analyses (Boussafir et al., 1995b) showed that the "orange OM" in Kimmeridge Clay kerogens contains high amounts of organic sulphur (atomic S/C ratios of 0.09 to 0.11). It is well docu- mented that S-C bonds are weaker than C-C bonds and, therefore, that S-rich kerogens tend to exhibit a lower thermal stability (e.g. Orr, 1986; Sinninghe Damst6 et al. , 1989; Di Primio and Horsfield, 1995; Reynolds et al. , 1995; Soldan et al. , 1995; Tomic et al., 1995). Indeed, studies on the pro- ducts generated on closed pyrolyses of an organic- rich Kimmeridge Clay sample (Monin et al., 1990) and on some North Sea oils expelled from the Kimmeridge Clay Formation (Chung e t al. , 1995) illustrated such low stability. Accordingly, the shift to lower energies observed from these three Kimmeridge Clay samples, especially from T, likely reflects the origin of the "orange OM" and the vari- ations in its relative abundance along microcycle III. A low contribution of energies >60 kcal tool -~ (ap- proximately 4%) is noted for sample B, and such a contribution is still weaker for M and negligible for T. This may reflect the presence of "brown OM" in the former sample. As already stressed: (i) this "brown OM" is dominated by ultralaminae and is, thus, chiefly derived by the selective preservation of

714 D. Dessort et aL
MARTON 11-16 MARTON 11-1 . . . . 60 60 % [ A:2.1E+13S-1 I % / A = s'8 E+13 s'l 5 0 . . . . . . . . . . . . . . . . . . . . . 5 0 . . . . . . . . . . . . . . . . . . . . . . . .
40 . . . . . . . . . . . . . . . . . . . . . . 40 . . . . . . . . . . . . . . . . . . . . . . . .
30 . . . . . . . . . . . . . . . . . . .
30 . . . . . . . . . . . . . . . . . . . . . . i . . . . . 20 . . . . . . . . . . . . . . . . . . . . . 20 . . . . . . . . . . . . . . . . . . . i
11 1 i - - ,
Kcal I mole Kcal I mole
Fig. 7. Distribution of activation energies and frequency factor (A) of the Kimmeridge Clay samples from microcycle II.
algaenan; and (ii) algaenans and related kerogens are characterized by high range values for Ea.
Previous kinetic studies of Kimmeridge Clay samples, exhibiting similar TOC values (Tegelaar and Noble, 1994; Reynolds and Burnham, 1995) when compared with T showed activation energy distributions and frequency factors similar to those observed here for the latter material.
Two additional Kimmeridge Clay samples corre- sponding to the beginning (Marton-1) and the top (Marton-16) of another microcycle, Marton-87-II, were also examined (Fig. 7). When compared with Marton-87-III, this microcycle is characterized by much higher TOC values (Table 1). The "sulphur index" of these two samples is 37 and approxi- mately 200, respectively, indicating that the "natu- ral vulcanization" pathway played a major role in the formation of the latter. Indeed: (i) the activation energy distribution of Marton-1 is similar to that observed for sample T; and (ii) the distribution of Marton-16 is still shifted to lower values and the principal energy at 51 kcalmol -~ accounts for ap- proximately 50% of the reaction.
Lorca Oi l Shale
This low maturity deposit, up to 100m thick, comprises calcareous organic-rich layers overlain by gypsiferous porous limestone partly filled with native sulphur (Benalioulhaj et aL, 1994). The acti- vation energy distribution of the Lorca sample (Fig. 6) is rather broad and the principal energy at 57 kcalmol -l accounts only for ca. 40% of the reaction. An important contribution of relatively low values is also noted, the energies <52 kcalmol -t describing 16% of the reaction. A previous examination based on bulk features (Permanyer et al., 1994) showed that the kerogen of the Lorca Oil Shale can be classified as a type I-S
material, as recently defined by Sinninghe Damst6 et al. (1993). In fact, the Ea distribution of the Lorca sample, with substantial contributions of relatively low energies recalls the distribution observed (Tegelaar and Noble, 1994) for the Ribesalbes Oil Shale, a typical Type I-S kerogen (Sinninghe Damst6 et al., 1993). Previous studies on the Lorca material (Derenne et al., 1995) demon- strated that sulphur incorporation into lipids over- whelmingly contributed to the formation of this kerogen, Such an origin was indicated by: (i) the exclusive presence of nanoscopically amorphous OM; (ii) an extremely high content of organic sul- phur (16.3 wt%, atomic S/C ratio of 0.1), whereas pyrite only occurs in trace amounts; and (iii) the nature of the pyrolysis products. Indeed, recent analyses (Jensen et al., 1997) revealed an extremely high "sulphur index" of approximately 290 (Table 1) for this oil shale. The Lorca sample, thus, provides an example of a kerogen formed virtually exclu- sively by the "natural vulcanization" pathway. As stressed above, this process leads to the formation of geomacromolecules exhibiting a relatively low thermal stability. This is consistent with: (i) recent observations on early release of bitumen from the Lorca Oil Shale, during artificial maturation (Landais et al., 1995); and (ii) the shift to lower ac- tivation energy values observed here for this shale when compared with the low maturity type I kero- gens (Pula alginite, NE 70 Torbanite, Rundle Oil Shale).
In agreement with previous pyrolytic studies on sulphur-rich kerogens (Baskins and Peters, 1992 and references therein) it, therefore, appears that the involvement of the "natural vulcanization" pathway in kerogen formation is associated with a decrease in activation energies. Nevertheless, the sulphur content is not the only factor that controls

Kinetic parameters of algaenous and kerogens
0.15 Thermal history: 3"C/MY 0.14 - - - Rundle Oil Shale 0.13 0.12 Pula Alginite 0.11
0.1 0.09 Lorca Oil Shale 0.08 ""
m 0.07 NE 70 Torbanite 0.06
--0.050.04 ~MARTONII-16 / I-" \~/',\ \
0.020"03 MARTON II-1 , " t " ¢ . " " / ~ , ~ '! t
0.01 ..... - 0
50 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200 210 T (°C)
Fig. 8. Simulated generation rate curves (geological heating rate of 3°C Ma -l) for the low maturity fos- sil samples. The curves corresponding to the Kimmeridge Clay samples from microcycle IIl, relatively
close to the curve obtained for the Marton II-1 sample, are not shown.
715
the extent of such a shift. Indeed, while the Lorca Oil Shale is characterized by a higher "sulphur index" than the Marton-16 sample, the shift to lower activation energies is less pronounced for the former material. Again, this may reflect compensat- ing effects associated with the large differences in frequency factors observed for these two samples. Nevertheless, in agreement with Ea distributions, the 450°C rate constant is markedly lower for the Lorca Oil Shale (Fig. 3). In fact, several factors suggest themselves to account for the relative ther-
mal stabilities of the Lorca and Marton-16 samples, such as." (i) the nature of the lipids where sulphur incorporation took place; (ii) the nature of the sul- phur groups implicated in lipid cross-linking, i.e. the relative importance of intramolecular (for- mation of moieties containing only one sulphur atom) and intermolecular links (formation of bridges that can contain a large number of sulphur atoms); and (iii) differences in the environment of the sulphur groups (influence of the neighbouring functions on their thermal stability).
1 Thermal history: 3°C/MY
°' I "un0eO "el O 0.8 / Pula Alginite ,=o., I 1 / . 7 / 0"61--'-- L°rca Oll Shale / / / / ,'/ / ~ 0 . 5 =: / NE'°r°r"n" r l " l . . - I I , 'O' 0"4 /--MARTONII'16 / flY.. I Io.3 } . - ' ] l . . J / ~0 .2
0 50 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200 210
T (°C)
Fig. 9. Simulated cumulative yield curves (geological heating rate of 3°C Ma -l) for the low maturity fossil samples. The curves corresponding to the Kimmeridge Clay samples from microcycle III relatively
close to the curve obtained for the Marton II-l sample, are not shown.

716 D. Dessort et al.
Table 2. Tmax and width at half height (IV) from the simulated generation rate curves (geological heating rate of 3°C Ma -1) for the low maturity fossil samples
Pula Alginite NE 70 Rundle Kimmeridge Kimmeridge Kimmeridge Kimmeridge Kimmeridge Lorca Oil Torbanite Oil Shale Clay B Clay M Clay T Clay II-1 Clay I1-16 Shale
Tmax 170 184 164 147 152 145 146 136 151 W (°C) 22 20 27 28 30 29 29 25 36
Extrapolation to geological conditions
The data obtained from the laboratory exper- iments can also be used for calculating kinetic fea- tures at very low heating rates, similar to those commonly assumed to occur in geological environ- ments. There are uncertainties in the extrapolation of such data to the much lower (approximately 1019-fold) heating rates experienced by kerogens during natural maturation (Schaefer et al., 1990; Tegelaar and Noble, 1994); howeve:, the general trends so predicted should be reliable. The kinetic results derived from open-system pyrolyses, for the low maturity fossil materials (Pula alginite, NE 70 Torbanite, Rundle Oil Shale, Kimmeridge Clay and Lorca Oil Shale) were extrapolated at a geological heating rate of 3°C Ma - l , with an initial tempera- ture of 10°C (Fig. 8). Width at half-height and Tmax were determined from the simulated generation rate curves thus obtained (Table 2). Cumulative yield curves (transformation ratio of kerogens into vol- atile pyrolysis products vs temperature) were also constructed (Fig. 9). Such curves are commonly referred to in the literature as "oil" or "hydro- carbon" generation profiles. It should, however, be noted that the information provided by Pyromat pyrolyses reflects the sum of the different types of volatile products swept away by the inert gas flow (including gaseous compounds and non-hydro- carbon constituents). Accordingly, the derived results cannot be considered, sensu stricto, as repre- sentative of oil or hydrocarbon generation.
It was previously noted from cumulative gener- ation curves simulating the transformation of oil into gas in reservoirs that the relative thermal stab- ility of some fossil materials can be inverted when curves corresponding to laboratory and geological heating rates are compared (Schenk and Horsfield, 1995). Such changes should reflect the effects of both activation energy distribution and frequency factors in reactivity control. In fact, at very low geological heating rates, the influence of Ea prevails and possible compensating effects due to differences in frequency factors are then negligible. In the pre- sent case no inversion was observed in the profiles corresponding to the nine tested samples on com- parison of the cumulative yield curves obtained at laboratory and geological heating rates. Such a fea- ture, therefore, suggests that the influence of Ea still prevails for the heating rates used in our laboratory
experiments. In agreement with the observations on 450°C rate constants, major compensation effects associated with frequency factors are, therefore, unlikely to occur in these experiments.
Comparison of the extrapolated kinetic features revealed substantial differences between the two Botryococcus-derived kerogens (Figs 8 and 9; Table 2). However, in agreement with a previous study (Tegelaar and Noble, 1994) showing gener- ation at higher temperatures for an Australian Torbanite when compared with other type I kero- gens, relatively high values of Tm~x were noted for both samples.
The five Kimmeridge Clay samples show rela- tively close features for simulated maturation (Figs 8 and 9; Table 2). Their Tmax is shifted to lower values when compared with Botryococcus-derived kerogens and to the Rundle Oil Shale. This shift appears especially pronounced in the case of the sulphur-rich Marton 11-16 sample. These obser- vations are consistent with previous studies (Tegelaar and Noble, 1994; Peters et al., 1995) indi- cating a shift of generation curves for sulphur-rich kerogens.
The Lorca Oil Shale, with an extremely high sul- phur content, also shows a lower value for Tmax when compared with the type I kerogens (Fig. 8). Although the Lorca Oil Shale kerogen, like the "orange OM" in the Kimmeridge Clay samples, originates from natural sulphurization the rate and generation profiles obtained for the former kerogen exhibit markedly different shapes. The rate profile of the Lorca material is unsymmetrical and shows both a pronounced shoulder, reflecting the release of an important part of its potential at low tem- peratures in the 90 to 120°C range, and a maximum at the relatively high value of 151°C. Accordingly, as already discussed in the previous section on acti- vation energy distributions, the kinetic differences between the Kimmeridge Clay samples and the Lorca Oil Shale are unlikely to be related merely to differences in the extent of "vulcanization" and in the abundance of organic sulphur. The maturation of the Lorca Oil Shale is characterized by a sub- stantially earlier onset of generation and a broader generation range (Figs 8 and 9). Catagenesis of this material should, thus, begin at a lower thermal stress and the oil window should correspond to a wider temperature range.

Kinetic parameters of algaenous and kerogens 717
CONCLUSIONS
The determination for the first time of the kin- etic paramaters of algaenans showed important differences between the four samples studied. The activation energies thus obtained were correlated with chemical structures. Two extremes were observed, first, the algaenan from the A race of B. braunii showed an E~ distribution virtually cor- responding to a single high value energy, reflect- ing an homogeneous structure with a relatively simple aliphatic polyaldehyde as building block. Secondly, on the contrary the algaenan from S. quadricauda showed a broad distribution and the principal Ea accounts for only one-third of the reaction, thus reflecting a more complex macro- molecular structure.
The E~ distributions of the four kerogens chiefly derived from the selective preservation of algaenan also exhibited pronounced differences. The kinetic features of the Pula Oil Shale are consistent with a contribution of both the A (pre- dominant) and the L races of B. braunii. The dis- tribution of the NE 70 Torbanite revealed an unusually homogeneous nature for a fossil ma- terial, with a major contribution of selectively preserved algaenan. The differences in kinetic par- ameters between the NE 70 and BJ 248 Torbanites do not chiefly reflect the different levels of maturity, but rather intrinsic differences due to the diversity of the materials implicated along with the selectively preserved algaenan in the formation of the latter Torbanite. The Rundle Oil Shale showed a broad distribution similar to the one obtained from S. quadricauda algaenan. Important differences in the chemical structure and thermal behaviour, not revealed by type classification since these three kerogens are classi- fied as low maturity type I are, therefore, reflected by the Pyromat-derived kinetic par- ameters of the Rundle, Pula and N E 70 samples.
The Kimmeridge Clay and Lorca samples showed broad Ea distributions shifted to low energies. Such shifts reflect a more or less important contribution of materials originating from sulphur incorporation into various lipids during early diagenesis. Nevertheless, as shown by comparison of the Lorca Oil Shale and of the sulphur-rich Kimmeridge Clay sample, the sulphur content is not the only factor that controls the shift to lower energies.
Finally, the kinetic data derived from Pyromat pyrolysis were extrapolated to a very low, geologi- cal heating rate of 3°C Ma -1 for the nine low maturity fossil samples. The simulated rate and gen- eration profiles showed: (i) substantial differences between the Botryococcus-derived kerogens, although they both exhibited relatively high Tmax; (ii) a shift of Tmax to lower values for the samples with a high contribution of the "vulcanizat ion" pathway; and (iii) a markedly different shape for
the profiles obtained from the Lorca sample, reflect- ing the release of an important part of its potential at low temperatures, probably due to differences in the nature and/or the environment of the cross-link- ing sulphur groups.
Associate Ed i to r - - J . S. Sinninghe Damste
Acknowledgements--This work was supported by Elf Aquitaine and by CNRS. Publication approval by Elf Aquitaine is also acknowledged.
REFERENCES
Baskins, D. K. and Peters, K. E. (1992) Early generation characteristics of a sulfur-rich Monterey kerogen. Bull. Am. Assoc. Pet. Geol. 76, 1-13.
Behar, F., Derenne, S., Largeau, C. and Rudkiewicz, J. L. (1993) Relative thermal stabilities of the resistant bioma- cromolecules of Botryococcus braunii, B and L races. Implications for the timing of oil generation from Botryococcus-derived kerogens. In Organic Geochemistry (Edited by Oygard, K.), pp. 254-258. Falch Hurtigtrykk, Oslo.
Behar, F., Derenne, S. and Largeau, C. (1995) Closed pyr- olyses, of the isoprenoid algaenan of Botryococcus brau- nii, L race. Geochemical implications for derived kerogens. Geochim. Cosmochim. Acta 59, 2983-2997.
Benalioulhaj, S., Schreiber, B. S. and Philp, R. P. (1994) Relationship of organic geochemistry to sedimentation under highly variable environments, Lorca Basin, Spain. In Sedimentology and Geochemistry of Modern and Ancient Saline Lakes (Edited by Renaut, R. W. and
Las t , W. M.), Vol. 50, pp. 315-324. Soc. Sed. Geol. spec. publication.
Berkaloff, C., Casadevall, E., Largeau, C., Metzger, P., Peracca, S. and Virlet, J. (1983) The resistant polymer of the walls of the hydrocarbon-rich alga Botryococcus braunii. Phytochemistry 22, 389-397.
Boussafir, M., Gelin, F., LaUier-Verg~s, E., Derenne, S., Bertrand, P. and Largeau, C. (1995a) Electron mi- croscopy and pyrolysis of kerogens from the Kimmeridge Clay Formation, U.K.: source organisms, preservation processes and origin of microcycles. Geochim. Cosmochim. Acta 59, 3731 3747.
Boussafir, M., Lallier-Verg~s, E., Bertrand, P. and Badaut-Trauth, D. (1995b) SEM and TEM studies on isolated organic matter and rock microfacies from a short-term organic cycle of the Kimmeridge Clay Formation (Yorkshire, G.B.). In Lectures Notes in Earth Science Series (Edited by Lallier-Vergrs, E., Tribovillard, N. P. and Bertrand, P.), Vol. 57, pp. 15 30. Springer-Verlag, Berlin.
Braun, R. L., Burnham, A. K., Reynolds, J. G. and Clarkson, J. E. (1991) Pyrolysis kinetics for lacustrine and marine source rocks by programmed micropyroly- sis. Energy and Fuels 5, 192-204.
Brukner-Wein, A., Het4nyi, M., and Solti, G. (1991) Organic geochemistry of alginite deposited in a volcanic crater lake. In Organic Geochemistry (Edited by Manning, D.), pp. 402-404. Manchester University Press, Manchester.
Burnham, A. K., Braun, R. L., Gregg, H. R. and Samoun, A. M. (1987) Comparison of methods for measuring kerogen pyrolysis rates and fitting kinetic parameters. Energy & Fuels l, 452-458.
Burnham, A. K., Schmidt, B. J. and Braun, R. L. (1995) A test of the parallel reaction model using kinetic measurements on hydrous pyrolysis residues. Org. Geochem. 23, 931-939.

718 D. Dessort et al.
CasadevaU, E., Dif, D., Largeau, C., Gudin, C., Chaumont, D. and Desanti, O. (1985) Studies on batch and continuous cultures of Botryococcus braunii: hydro- carbon production in relation to physiological state, cell ultrastructure and phosphate nutrition. Biotechnol. Bioeng. 27, 286-295.
Chung, H. M., Claypool, G. E. and Waiters, C. C. (1995) Multiple origin of petroleum in the Viking Graben. In Organic Geochemistry: Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 500-504. A.I.G.O.A., Donostia-San Sebastian.
Derenne, S., Largeau, C., Casadevall, E. and Laupr&re, F. (1987) Structural analysis of two Torbanites at differ- ent evolutionary stages. Investigation of the quantitative reliability of f~ determination by 13C CP/MAS n.m.r. Fuel 66, 1084-1090.
Derenne, S., Largeau, C., Casadevall, E. and Connan, J. (1988) Comparison of torbanites of various origins and evolutionary stages. Bacterial contribution to their formation. Cause of the lack of botryococcane in bitu- mens. Org. Geochem. 12, 43-59.
Derenne, S., Largeau, C., Casadevall, E. and Berkaloff, C. (1989) Occurrence of a resistant biopolymer in the L race of Botryococcus braunii. Phytochemistry 28, 1137- 1142.
Derenne, S., Largeau, C., Casadevall, E. and Sellier, N. (1990) Direct relationship between the resistant bio- polymer and the tetraterpenic hydrocarbon in the lyco- padiene-race of Botryococcus braunii. Phytochemistry 29, 2187-2192.
Derenne, S., Largeau, C., Casadevall, E., Berkaloff, C. and Rousseau, B. (1991) Chemical evidence of kerogen for- mation in source rocks and oil shales via selective pres- ervation of thin resistant outer walls of microalgae: origin of ultralaminae. Geochim. Cosmochim. Acta 55, 1041-1050.
Derenne, S., Largeau, C., Berkaloff, C., Rousseau, B., Wilhelm, C. and Hatcher, P. (1992a) Non-hydrolysable macromolecular constituents from outer walls of Chlorella fusca and Nanochlorum eucaryotum. Phytochemistry 31, 1923-1929.
Derenne, S., Largeau, C. and Hatcher, P. G. (1992b) Structure of Chlorella fusca algaenan: Relationships with ultralaminae in lacustrine kerogens; species- and environment-dependent variations in the composition of fossil ultralaminae. Org. Geochem. 18, 417-422.
Derenne, S., Le Berre, F., Largeau, C., Hatcher, P., Connan, J. and Raynaud, J. F. (1992c) Formation of ultralaminae in marine kerogens via selective preser- vation of thin resistant outer walls of microalgae. Org. Geochem. 19, 345-350.
Derenne, S., Largeau, C. and Taulelle, F. (1993) Occurrence of non-hydrolysable amides in the macro- molecular constituent of Scenedesmus quadricauda cell wall as revealed by 15N NMR. Origin of n-alkylnitriles in pyrolysates of ultralaminae-containing kerogens. Geochim. Cosmochim. Acta 57, 851-857.
Derenne, S., Largeau, C. and Behar, F. (1994) Low po- larity pyrolysis products of Permian to Recent Botryococcus-rich sediments: first evidence for the con- tribution of an isoprenoid algaenan to kerogen for- mation. Geochim. Cosmochim. Acta 58, 3703-3711.
Derenne, S., Permanyer, A., Largeau, C., and Baranger, R. (1995) Relationships between ultrastructural and chemical features in the pre-evaporitic oil shale from the Lorca Basin (Upper Miocene, S. E. Spain). In Organic Geochemistry: Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 1041-1043. A.I.G.O.A., Donostia-San Sebastian.
Derenne, S., Largeau, C., Het6nyi, M., Brukner-Wein, A., Connan, J. and Lugardon, B. (1997) Chemical structure of the organic matter in a Pliocene maar-type oil shale. Implicated Botryococcus races and formation pathways. Geochim. Cosmochim. Acta 61, 1879-1889.
Di Primio, R. and Horsfield, B. (1995) Predicting the gen- eration of heavy oils in carbonate/evaporitic environ- ments using pyrolysis methods. In Organic Geochemistry: Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 410-412. A.I.G.O.A., Donostia-San Sebastian.
Gelin, F., de Leeuw, J. W., Sinninghe Damstb, J. S., Derenne, S., Largeau, C. and Metzger, P. (1994a) The similarity of chemical structures of soluble aliphatic polyaldehyde and insoluble algaenan in the green micro- alga Botryococcus braunii race A as revealed by analyti- cal pyrolysis. Org. Geochem. 21, 423-435.
Gelin, F., de Leeuw, J. W., Sinninghe Damst6, J. S., Derenne, S., Largeau, C. and Metzger, P. (1994b) Scope and limitations of flash pyrolysis-gas chromatography- mass spectrometry as revealed by the thermal behaviour of high-molecular-weight lipids derived from the green microalga Botryococcus braunii. J. Anal. Appl. Pyrol. 28, 183-204.
Herbin, J. P., Geyssant, J. R., MOiler, C., M61i6res, F. and Penn, I. E. (1991) Le groupe YORKIM: H6t6rog6n6it6 quantitative et qualitative de la mati6re organique dans les argiles du Kimm6ridgien du val de Pickering (Yorkshire, U.K.). Cadre sddimentologique et stratigra- phique. Rev. Inst. Fr. P~tr. 46, 1-39.
Herbin, J. P., MOiler, C., Geyssant, J. R., M61i6res, F., and Penn, I. E. (1993) Variation of the distribution of organic matter within a transgressive system tract: Kimmeridge Clay (Jurassic, England). In AAPG 1993 "Petroleum Source Rocks in a Sequence Stratigraphy Framework" (Edited by Katz, B. and Pratt, L.), pp. 67- 100.
Horsfield, B., Curry, D. J., Bohacs, K., Littke, R., Rullk6tter, J., Schenk, H. J., Radke, M., Schaefer R. G., Carroll, A. R., Isaksen, G. and Witte, E. G. (1994) Organic geochemistry of freshwater and alkaline lacus- trine sediments in the Green River Formation of the Washakie Basin, Wyoming, U.S.A. In Advances in Organic Geochemistry 1993 (Edited by Telnaes, N., van Graas, G. and Oygard, K.), Org. Geochem. Vol. 22, pp. 415-440. Pergamon Press, Oxford.
Hunt, J. M., Lewan, M. D. and Hennet, R. J.-C. (1991) Modeling oil generation with time-temperature index graphs based on the Arrhenius equation. Bull. Am. Assoc. Pet. Geol. 75, 795-807.
Jambor, A. and Solti, G. (1975) Geological conditions of the Upper Pannonian oil shale deposit recovered in the Balaton Highland and at Kemenesh~tt. Acta Miner. Petr. Szeged XXVII]I, 73-85.
Jensen, H. K. B., Connan, J., Bjoroy, M., Hall, K. and Wold, S. (1997) Using the Geoelf Sulphur Analyser to quantify thermally extractable and pyrolysable organic and mineral sulphur in source rocks. Org. Geochem. (in press).
Kadouri, A., Derenne, S., Largeau, C., Casadevall, E. and Berkaloff, C. (1988) Resistant biopolymer in the outer walls of Botryococcus braunii B Race. Phytochemistry 27, 551-557.
Kister, J., Guiliano, M., Largeau, C., Derenne, S. and Casadevall, E. (1990) Characterization of chemical structure, degree of maturation and oil potential of Torbanites (Type I kerogens) by quantitative FTIR spectroscopy. Fuel 69, 1356-1361.
Landais, P., Rochdi, A., Largeau, C. and Derenne, S. (1993) Chemical characterization of Torbanites by transmission rnicro-FTIR spectroscopy. Origin and

Kinetic parameters of algaenous and kerogens 719
extent of compositional heterogeneities. Geochim. Cosmochim. Acta 57, 2529-2539.
Landais, P., Permanyer, A., Schlegela, M. and Mansuy, L. (1995) Artificial maturation of a sulphur-rich source- rock from the Lorca Basin, Spain. In Organic Geochemistry: Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 1054-1057. A.I.G.O.A., Donostia-San Sebastian.
Largeau, C., Casadevall, E., Kadouri, A. and Metzger P. (1984) Formation of Botryococcus braunff kerogens. Comparative study of immature Torbanite and of the extant alga Botryococcus braunii. In Advances in Organic Geochemistry 1983 (Edited by Schenck, P. A., de Leeuw, J. W. and Lijmbach, G. W. M.), Org. Geochem. Vol. 6, pp. 327-332. Pergamon Press, Oxford.
Largeau, C., Derenne, S., Casadevall, E., Kadouri, A. and Sellier, N. (1986) Pyrolysis of immature Torbanite and of the resistant biopolymer (PRBA) isolated from extant alga Botryococcus braunii. Mechanism of formation and structure of Torbanite. In Advances in Organic Geochemistry 1985 (Edited by Leythaeuser, D. and Rullk6tter, J.), Vol. 10, pp. 1023-1032. Pergamon Press, Oxford.
Largeau, C., Derenne, S., Casadevall, E., Berkaloff, C., Corolleur, M., Lugardon, B., Raynaud, J. F. and Connan, J. (1990) Occurrence and origin of "ultralami- nar" structures in "amorphous" kerogens of various source rocks and oil shales. In Advances in Organic Geochemistry 1989 (Edited by Durand, B. and Behar, F.), Vol. 16, pp. 889-895. Pergamon Press, Oxford.
Largeau, C. and Derenne, S. (1993) Relative efficiency of the selective preservation and degradation recondensa- tion pathways in kerogen formation. Source and en- vironment influence on their contributions to type I and II kerogens. Org. Geochem. 20, 611-615.
Largeau, C. (1995) Formation of refractory organic matter from biological precursors. In The Role of Non-living Organic Matter in the Earth's Carbon Cycle (Edited by Zepp, R.), Dahlem Conference Reports, pp. 275-292. Springer, Berlin.
Largeau, C. and de Leeuw, J. W. (1995) Insoluble, non- hydrolysable, aliphatic macromolecular constituents of microbial cell walls. In Advances in Microbial Ecology, Vol. 14, pp. 77-117. Plenum, New York.
de Leeuw, J. W. and Largeau, C. (1993) A review of macromolecular organic compounds that comprise liv- ing organisms and their role in kerogen, coal and pet- roleum formation. In Organic Geochemistry Principles and Applications (Edited by Engel, M. H. and Macko, S. A.), pp. 23-72. Plenum, New York.
Metzger, P., Largeau, C. and Casadevall, E. (1991) Lipids and macromolecular lipids of the hydrocarbon-rich microalga Botryococcus braunii. Chemical structure and biosynthesis--geochemical and biotechnological import- ance. In Progress in the Chemistry of Organic Natural Products (Edited by Herz, W., Kirby, G. W., Steglich, W. and Tamm, C.), Vol. 57, pp. 1-70. Springer Verlag, New York.
Monin, J. C., Connan, J., Oudin, J. L. and Durand, B. (1990) Quantitative and qualitative experimental approach of oil and gas generation: Application to the North Sea source rocks. In Advances in Organic Geochemistry 1989 (Edited by Durand, B. and Behar, F.), Vol. 16, pp. 889-895. Pergamon Press, Oxford.
Orr, W. L. (1986) Kerogen/asphaltene/sulfur relationships in sulfur-rich Monterey oils. In Advances in Organic Geochemistry 1985 (Edited by Leythaeuser, D. and Rullkftter, J.), Vol. 10, pp. 499-516. Pergamon Press, Oxford.
Peters, K. E., Cunningham, A. E., Waiters, C. C., Jigang, J. and Zhaoan, F. (1995) Unusual petroleum systems in
the Dangyang-Jiangling area, Jianghan Basin, China. In Organic Geochemistry: Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 475-477. A.I.G.O.A., Donostia-San Sebastian.
Permanyer, A., Baranger, R. and Lugardon, B. (1994) Oil shale characterization in Messinian pre-evaporitic sedi- ments from the Lorca basin (south-east Spain). Bull. Centres Rech. Explor.-Prod. Elf Aquitaine 18, 135-149.
Reynolds, J. G. and Burnham, A. K. (1995) Comparison of kinetic analysis of source rocks and concentrates. Org. Geochem. 23, 11-19.
Reynolds, J. G., Burnham, A. K. and Mitchell, T. O. (1995) Kinetic analysis of California petroleum source rocks and concentrates. Org. Geochem. 23, 109-120.
Schaefer, R. G., Schenk, H. J., Hardelauf, H. and Harms, R. (1990) Determination of gross kinetic parameters for petroleum formation from Jurassic source rocks of different maturity levels by means of laboratory exper- iments. In Advances in Organic" Geochemistry 1989 (Edited by Durand, B. and Behar, F.), Vol. 16, pp. 115- 120. Pergamon Press, Oxford.
Schenk, H. J. and Horsfield, B. (1995) Simulating the con- version of oil into gas in reservoirs: The influence of fre- quency factors on kinetic predictions. In Organic Geochemistry." Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 1102-1103. A.I.G.O.A., Donostia-San Sebastian.
Schmidt, B. J. (1995) Relationships between maceral com- position, kerogen classification and kinetic parameters. In Organic Geochemistry: Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 532-534. A.I.G.O.A., Donostia-San Sebastian.
Sinninghe Damstr, J. S., Eglinton, T. I., de Leeuw, J. W. and Schenck, P. A. (1989) Organic sulphur in macromolecular organic matter. I. Structure and origin of sulphur-containing moieties in kerogen, asphaltenes and coals as revealed by flash pyrolysis. Geochim. Cosmochim. Acta 53, 873-889.
Sinninghe Damstr, J. S. and de Leeuw, J. W. (1990) Analysis, structure and geochemical significance of orga- nically-bound sulphur in the geosphere: State of the art and future research. In Advances in Organic Geochemistry 1989 (Edited by Durand, B. and Behar, F.), Vol. 16, pp. 1077-1101. Pergamon Press. Oxford.
Sinninghe Damstr, J. S., de las Heras, F. X. C., van Bergen, P. F. and de Leeuw, J. W. (1993) Characterization of Tertiary catalan lacustrine oil shale: discovery of extremely organic sulphur-rich type I kero- gens. Geochim. Cosmochim. Acta 57, 389-415.
Soldan, A. L., Rangel, H. D., Telles, A. C. S. Jr., De Grande, S. M. B. and Henz, G. I. (1995) Kinetic varia- bility in the Lagoa Feia Formation, palaeoenvironmen- tal influences. In Organic Geochemistry: Developments and Applications to Energy, Climate, Environment and Human History (Edited by Grimalt, J. O. and Dorronsoro, C.), pp. 128-130. A.I.G.O.A., Donostia- San Sebastian,
Solti, G. (1985) Prospection and utilization of alginite and oil shale in Hungary. In Neogene Mineral Resources in the Carpathian Basin (Edited by Hala, J.), pp. 503-517, Hungarian Geological Survey, Budapest.
Tegelaar, E. W., de Leeuw, J. W., Derenne, S. and Largeau, C. (1989) A reappraisal of kerogen formation. Geochim. Cosmochim. Acta 53, 3103-3106.
Tegelaar, E. W. and Noble, R. A. (1994) Kinetics of hydrocarbon generation as a function of the molecular structure of kerogen as revealed by pyrolysis-gas chromatography. In Advances in Organic Geochemistry 1993 (Edited by Telnaes, N.. van Graas, G. and

720 D. Dessort et al.
Oygard, K.), Vol. 22, pp. 543-574. Pergamon Press, Oxford.
Tomic, J., Behar, F., Vandenbroucke, M. and Tang, Y. (1995) Artificial maturation of Monterey kerogen (Type II-S) in a closed system and comparison with
Type II kerogens: implications on the fate of sulfur. Org. Geochern. 23, 647-660.
Ungerer, P. and Pelet, R. (1987) Extrapolation of kinetics of oil and gas formation from laboratory experiments to sedimentary basins. Nature (London) 327, 42-44.