
Superstesaster promissor gen. et sp. nov., a new starfish ...
Upload
independentCategory
view
1download
0
Cloning and expression of a membrane-bound CMP-N-acetylneuraminicacid hydroxylase from the starfish Asterias rubens
Ilka Martensen, Roland Schauer and Lee Shaw
Biochemisches Institut, Christian-Albrechts-Universitat Kiel, Germany
The sialic acid N-glycolylneuraminic acid (Neu5Gc) is
synthesized by the action of CMP-Neu5Ac hydroxylase.
The enzyme from various mammals has been purified,
characterized and sequenced by cDNA cloning. Although
functional sequence motifs can be postulated from
comparisons with several enzymes, no global homologies
to any other proteins have been found. The unusual charac-
teristics of this hydroxylase raise questions about its
evolution. As echinoderms are phylogenetically the oldest
organisms possessing Neu5Gc, they represent a starting
point for investigations on the origin of this enzyme. Despite
many similarities with its mammalian counterpart, CMP-
Neu5Ac hydroxylase from the starfish A. rubens exhibits
fundamental differences, most notably its association with a
membrane and a requirement for high ionic strength. In order
to shed light on the structural basis for these differences, the
primary structure of CMP-Neu5Ac hydroxylase from
A. rubens has been determined by PCR and cDNA-cloning
techniques, using initial sequence information from the
mouse enzyme. The complete assembled cDNA contained
an ORF coding for a protein of 653 amino acids with a
molecular mass of 75 kDa. The deduced amino-acid
sequence exhibited a high degree of homology with the
mammalian enzyme, although the C-terminus was some 60
residues longer. This extension consists of a terminal hydro-
phobic region, which may mediate membrane-binding, and
a preceeding hydrophilic sequence which probably serves as
a hinge or linker. The identity of the ORF was confirmed by
expression of active CMP-Neu5Ac hydroxylase in E. coli at
low temperatures.
Keywords: sialic acid; echinoderm; Rieske oxygenase;
cytochrome b5; membrane binding.
Sialic acids constitute a family of more than 40 acidicsugars which generally occur as terminal residues in theglycan chains of cell surface and secreted glycoconjugates[1,2]. Due to their exposed position, sialic acids mayinfluence the physico-chemical properties of glycoconju-gates and they frequently serve as ligands for receptor-mediated intercellular or host cell-pathogen recognitionprocesses [3,4].
Although small amounts of sialic acid have been reportedin insects [5,6] and a gastropod [7], they are not foundgenerally in glycoconjugates from protostome animals [8,9].However, within the deuterostomate lineage of the animalkingdom, both the echinoderms and vertebrates possesssignificant amounts of sialylated glycoconjugates and
frequently display a considerable number of sialic acidtypes [8,10].
The structural variety among the sialic acids results froma number of enzyme-catalysed modifications of N-acetyl-neuraminic acid occurring at different stages of glycocon-jugate biosynthesis [11]. N-Glycolylneuraminic acid is oneof the most common sialic acids and formally results fromthe hydroxylation of the methyl group of the 5-N-acetylmoiety of Neu5Ac. Interestingly, no Neu5Gc has been foundin human and chicken tissues, although small amounts havebeen detected in tumours [12,13]. In echinoderms, Neu5Gcis frequently the predominant sialic acid and may serve as astarting molecule for some unusual oligosaccharidestructures unique to this phylum [14–19].
The biosynthesis of Neu5Gc is catalysed by the enzymeCMP-Neu5Ac hydroxylase yielding CMP-Neu5Gc (Fig. 1)[20–22]. The virtual lack of Neu5Gc in man is due to anexon deletion in the human gene that prevents the formationof functional enzyme [23,24]. Purification and characteriz-ation studies on the hydroxylase from mammals have shownthat it is a soluble, cytosolic oxygenase that is dependent oncytochrome b5 and cytochrome b5 reductase for delivery ofreducing equivalents from NADH [25–29]. This compart-mentalization is very unusual, as all other known cyto-chrome b5-dependent enzymes are membrane-bound andcolocalized with cytochrome b5 in the endoplasmicreticulum (ER) [30]. However, electron microscopic studiesprovide some evidence for an association of the hydroxylasewith intracellular membranes [31]. A further interestingfeature of CMP-Neu5Ac hydroxylase is the presence of aRieske iron–sulfur centre as demonstrated by electron spinresonance (ESR) spectroscopy and cDNA cloning, making
Correspondence to L. Shaw, Biochemisches Institut der Christian-
Albrechts-Universitat zu Kiel, Olshausenstr. 40, 24098 Kiel, Germany.
Fax: 1 49 431880 2238, Tel.: 1 49 431880 1679,
E-mail: [email protected]
(Received 25 May 2001, revised 7 August 2001, accepted 9 August
2001)
Enzymes: CMP-Neu5Ac hydroxylase (EC 1.14.13.45); cytochrome b5
reductase (EC 1.6.2.2).
Note: the nucleotide sequence reported in this paper has been submitted
to the EMBL data bank under accession no. AJ 308602.
Abbreviations: ARH, full-length enzyme; CSPD, chemiluminescent
alkaline phosphatase substrate; CMP-Neu5Ac, cytidine-50-
monophospho-N-acetylneuraminic acid; DIG, digoxygenin; DMG,
3,30-dimethylglutaric acid; IPTG, isopropyl thio-b-D-galactoside;
Neu5Ac, N-acetylneuraminic acid; Neu5Gc, N-glycolylneuraminic
acid; TMD, transmembrane domain
Eur. J. Biochem. 268, 5157–5166 (2001) q FEBS 2001

this the only known cytosolic Rieske protein in eukaryotes[32,33]. In Rieske [2Fe2S] centres, one iron is co-ordinatedby two cysteine residues and the other by the 1-nitrogenatoms of two histidine side chains. These liganding aminoacids form the highly characteristic sequence motifCXHX(15–20)CXXH present in all Rieske proteins [34].Activation of the hydroxylase by the addition of iron ions ledto the assumption that it contains an additional exchange-able iron cofactor, presumably a mononuclear iron centre.This combination of iron centres has only been observed in adiverse family of bacterial oxygenases [34], and three plantproteins functionally unrelated to CMP-Neu5Ac hydroxyl-ase [35–37]. The unusual distribution and enzymicproperties exhibited by CMP-Neu5Ac hydroxylase there-fore raise questions about its evolution.
In order to gain insight into the origins of this enzyme, adetailed study on the CMP-Neu5Ac hydroxylase from thestarfish A. rubens is being performed in this research group.As an echinoderm, it is one of the most primitive organismsknown to possess Neu5Gc and therefore serves as a startingpoint for such investigations. Previous results have revealedthat the starfish hydroxylase resembles its mammaliancounterpart in many respects. For instance, it is dependenton NADH as well as the cytochrome b5 system for activityand also possesses an essential exchangeable iron cofactor[38,39]. However, in contrast to the soluble mammalianenzyme, the hydroxylase from A. rubens is membrane-bound. Furthermore, it requires a relatively high ionicstrength for optimal activity [39], while the mammalianenzyme is potently inhibited by low salt concentrations [40].
To elucidate whether these differences are reflected in theprimary structure of the starfish enzyme, we have cloned andsequenced the cDNA encoding CMP-Neu5Ac hydroxylasefrom A. rubens. The amino-acid sequence deduced from theORF contained in the cDNA is compared with all otherknown CMP-Neu5Ac hydroxylase sequences. To confirmthe identity of the cDNA and to produce sufficient enzymefor further investigations, the cDNA was expressed inEscherichia coli. A preliminary report of these results hasbeen published as an abstract [41].
E X P E R I M E N T A L P R O C E D U R E S
Reagents and general methods
All chemicals were purchased in analytical grade or higherfrom Merck, Sigma and Biomol. CMP-[14C]Neu5Ac wasfrom Amersham Pharmacia Biotech. Oligonucleotideprimers were synthesized by Eurogentec. Taq polymeraseand dNTPs were also purchased from Eurogentec. Reversetranscriptase (SuperScript II), terminal desoxynucleotidyltransferase (TdT) and DNA polymerase I (Klenow) werefrom Life Technologies. Restriction enzymes were obtainedfrom Life Technologies and New England Biolabs. T4 DNAligase was from Promega and DNase I from RocheMolecular Biochemicals.
Plasmid DNA was isolated with the QIAprep plasmidminiprep kit (Qiagen). Sequencing was performed with theBigDye Ready Reaction Terminator Cycle Sequencing Kit(PE Biosystems), followed by analysis on an ABI Prism 310Genetic Analyser (Applied Biosystems).
All DNA and RNA manipulations were performedaccording to standard procedures as previously described[42].
Starfish material
Starfish were collected from the North Sea in March andApril. After killing the animals by osmotic shock, thegonads were immediately removed and frozen as well asstored in liquid nitrogen.
Construction of an oligonucleotide probe to screen acDNA library
Total RNA was extracted from starfish gonads by CsCldensity gradient centrifugation and the mRNA was isolatedwith Oligotex particles (Qiagen). RT-PCR was performedwith a first strand cDNA template, obtained by reversetranscription (SuperScript II) of 2 mg mRNA with an oligo-dT primer (50-T20V-30; V ¼A, G or C) in a final volumeof 20 mL, according to the manufacturer’s instructions.Degenerate primers were deduced from the mouse amino-acid sequence in the region of the Rieske motif that consistsof two boxes containing the liganding residues described inearlier. Their sequences were: 50-AAYCTNTGYAARCAYCARGG-30 (Ri1s) and 50-CCARTTRTGYTTNGTRCA-30
(Ri2as) (R ¼ A or G, Y ¼ T or C, N ¼ A, G,T or C). Areaction mixture containing 0.5 mL of cDNA, 25 pmol ofeach primer, 40 nmol dNTPs (10 nmol each) and 2 mM
MgCl2 in a final volume of 49 mL was heated to 95 8Cfor 5 min. One unit of Taq polymerase was added at 72 8C(hot start) and the mixture subjected to PCR in a‘Mastercycler Gradient’ (Eppendorf) with the followingprogram: 40 � (45 s 95 8C, 45 s 48 8C, 1 min 72 8C), 7 min72 8C. This PCR was repeated under the same conditions,using 1 mL of the first PCR reacton mixture as the template.The PCR product obtained was introduced into the pGEM-Tvector (Promega) by T/A cloning and subsequentlysequenced.
From the elucidated sequence, two nested primers werededuced for a 50-anchored PCR. Their sequences wereas follows: 50-CAATATCTTTAATAAACGTC-30 (Ri3as),50-GAATGCAGTTATCCCCATC-30 (Ri4as). The template
Fig. 1. Reaction catalysed by CMP-Neu5Ac hydroxylase.
5158 I. Martensen et al. (Eur. J. Biochem. 268) q FEBS 2001

cDNA was obtained by reverse transcription of 250 ngmRNA with 10 fmol of the primer Ri4as as previouslydescribed [43]. A poly(A) tail was added to the cDNA by theaction of terminal desoxynucleotidyl transferase (TdT).Subsequently, a hot-start PCR (35 cycles) was performed on0.5 mL of this cDNA with 60 pmol each of Ri4as and anoligo-dT primer (other reagents and reaction conditionswere as described above). One microliter of the first PCRsample served as the template in a second, nested PCR withRi3as and an oligo-dT primer (same conditions as firstPCR). The product obtained was cloned and sequenced asdescribed above.
For the production of a probe, a further primer wasdeduced from the sequence of the 50-anchored PCR product:50-GGAACAAGAAAGAGAAATCG-30 (Son2s). The probewas labelled with digoxigenin (DIG) by PCR (PCR DIGProbe Synthesis Kit, Roche Molecular Biochemicals) with0.5 mL cDNA and 25 pmol each Son2s and Ri3as underthe following conditions: 5 min 94 8C, 72 8C (hot start),10 � (45 s 94 8C, 45 s 55 8C, 1 min 72 8C), 20 � [45 s94 8C, 45 s 55 8C, 60 s (plus an additional 10 s per cycle)72 8C], 7 min 72 8C.
Screening of a cDNA library
For the construction of a cDNA library, double stranded, size-fractionated cDNA was produced from A. rubens mRNAwith the SuperScript Choice System (Life Technologies)and ligated to ZAP Express EcoRI-predigested vector arms(Stratagene). Phages were assembled by the addition ofGigapack III Gold Packaging Extract (Stratagene) and theresulting primary library was amplified as described by themanufacturer.
Phages were plated with E. coli XL-1 blue MRF0
(Stratagene), the resulting plaques transferred to HybondN1 nylon membranes (Amersham) and the filters hybrid-ized with the DIG-labelled probe (50 ng:mL21 in standardbuffer with 50% formamide, see ‘The DIG System User’sGuide for Filter Hybridization’, Roche Molecular Bio-chemicals) for 14–16 h at 38 8C. Subsequently, the filterswere washed twice for 5 min in 2 � NaCl/Cit, 0.1% SDS at38 8C, and twice for 15 min in 0.5 � NaCl/Cit, 0.1% SDS at58 8C. The DIG-labelled probe was detected with an anti-DIG Ig and the chemiluminescent alkaline phosphatasesubstrate, CSPD (both Roche Molecular Biochemicals), asdescribed in the DIG user’s guide. A positive plaque waspurified and a plasmid containing the cDNA (designatedpBCH1) was excised. Both strands of the cDNA insert weresequenced by primer walking.
Construction of expression plasmids
For the construction of a plasmid with the full-lengthhydroxylase sequence, pGEM-T containing the 50-anchoredPCR product was digested with BstU I and Pst I and thefragment containing the insert was ligated to pBCH1, whichhad previously been digested with Nhe I, filled in withKlenow-fragment and subsequently digested with Pst I.Inserts for expression plasmids were produced by PCR,using the resulting plasmid, pARHg.bc, as the template.Primers were designed that contained the required start andend points of the sequence to be expressed with additionalNde I restriction sites. The respective primer sequences for
the three expression constructs studied were as follows(restriction sites are underlined):
pARH.14: NdeIMet1, 50-GGAATGGCATATGGAACAAGAAAGAGAAATCG-30; StopNdeI, 50-TTGGTTCCATATGCTACTTTGACTCTGATTTCC-30. pARH:CD21.14: NdeI-Met1 (see above); Arg632NdeI, 50-CCAATTCCATATGCTATCGAAGTAATCGCTCGTAG-30. pARH:CD67.14:NdeIMet1 (see above); Lys586NdeI, 50-CCAATTCCATATGCTACTTCATGTCCCTTAAGAAACC-30. PCR amplifi-cation of the inserts was performed as follows. Reactionmixture: 7 ng pARHg.bc, 25 pmol of each primer, 40 pmoldNTPs (10 pmol each), 2 mM MgSO4, 2 U Platinum HiFiTaq polymerase (Life Technologies) in a final volume of50 mL of the manufacturer’s buffer; temperature profile:5 min 94 8C, 10 � [30 s 94 8C, 30 s 60 8C (minus 1 8C percycle), 2 min, 68 8C], 20 � [30 s 94 8C, 30 s 50 8C, 2 min(plus an additional 10 s per cycle) 68 8C], 7 min 68 8C. Theproducts were digested with Nde I, ligated to pET14b(Novagen), which had been previously cleaved with Nde I,and transformed into E. coli XL-1 blue MRF0. The orien-tation of the insert was checked by colony PCR. Plasmidswere isolated and the correctness of the insert-plasmidjunctions and coding sequence were confirmed bysequencing.
Expression of CMP-Neu5Ac hydroxylase in E. coli
E. coli BL21(DE3)/pLysS, transformed with the respectiveexpression plasmids, were grown at 37 8C to a D600 ¼ 0.5.The bacteria were further grown at the temperature assignedfor induction, namely 10, 15, 25 or 37 8C, until theD600 ¼ 0.65–0.75. Alternatively, cells were grown andinduced continuously at 15 8C. In all cases, expression wasinduced by the addition of isopropyl thio-b-D-galactoside(IPTG) to a final concentration of 0.5 mM. After 3 h ofinduction, cells were harvested at 4 8C and frozen at280 8C. In order to lyse the bacteria for enzyme tests, thepellets were thawed by resuspension in cold 50 mM
dimethylglutaric acid (DMG)/NaOH pH 6.4, containing1% Nonidet P-40 (2–10% of original culture volume).Dnase I (10 mg:mL21) and MgSO4 (10 mM) were addedand the lysates shaken on ice for at least 30 min Aftercentrifugation, the clear supernatants were desalted onNAP-10 columns (Amersham Pharmacia) equilibrated in50 mM DMG/NaOH pH 6.4 containing 1% Nonidet P-40and supplemented with glycerol and dithiothreitol togive final concentrations of 8.7% (v/v) and 2 mM,respectively.
Enzyme assay, protein determination and SDS/PAGE
CMP-Neu5Ac hydroxylase activity was assayed asdescribed by Gollub et al. [39]. Briefly, the reaction mixturecontained 16.5 mL desalted bacterial supernatant, 0.75%Nonidet P-40, 1 mM NADH, 0.5 mM FeSO4, 1 mM
dithiothreitol, 100 mM NaCl, 90 mg pig liver microsomalprotein and 2.5 mM (< 0.02 mCi) CMP-[14C]Neu5Ac in50 mM DMG/NaOH pH 6.4, giving a final volume of 28 mL.The incubations were performed at 25 8C and the reactionsstopped by the addition of 5 mL of 1 M trichloroacetic acid.After centrifugation (14 000 g, 5 min, room temperature)the supernatants were subjected to thin layer chromato-graphy (TLC) to separate the released [14C]sialic acids,
q FEBS 2001 A. rubens CMP-Neu5Ac hydroxylase (Eur. J. Biochem. 268) 5159

followed by analysis on an Automatic TLC-Linear Analyzer(Berthold). The difference between duplicate enzyme testswas less than 10%.
The protein content of samples was determined with theBCA protein assay (Pierce), as described by the manufac-turer, with bovine serum albumin as standard.
For analysis by SDS/PAGE, frozen cells were resus-pended in 8 M urea, 100 mM NaH2PO4, 10 mM Tris pH 8.0(10% of the original culture volume) and, after centrifu-gation, 10–15 mL of the clear supernatant was resolved onan 8% acrylamide gel.
R E S U L T S
Generation of an oligonucleotide probe to screen a cDNAlibrary from gonads of A. rubens
In order to design degenerate primers for an initial PCRamplification of the A. rubens hydroxylase sequence, analignment of the sequences both within and adjacent tothe Rieske motif of the CMP-Neu5Ac hydroxylase frommouse and several bacterial Rieske-type oxygenases was
performed (data not shown). With the exception of theliganding cysteine and histidine residues, no conservedamino acids were found. Therefore, the primers were basedsolely on the mammalian, in this case mouse, CMP-Neu5Achydroxylase sequence. RT-PCR carried out with thesedegenerate primers and a starfish cDNA template resulted inan 87-bp product (Fig. 2). When translated into amino acids,the newly acquired sequence shared < 70% homology withthe mouse enzyme.
This information enabled us to deduce specific primersfor an anchored PCR, which yielded a 480-bp product(Fig. 2). When translated, the 30-terminal portion of thisproduct exhibited 60% homology to the N-terminus of themouse sequence. In addition, a presumed 50-untranslatedregion (UTR) was found, in addition to several nucleotidesresulting from the action of the TdT (not depicted in Fig. 2).In the region homologous to the mouse hydroxylase, amethionine in the context of a Kozak sequence was observed[44]. This Met is therefore assumed to be the starting pointof translation (Fig. 2).
Due to the noticeable homology between the sequence ofthe mouse hydroxylase and the PCR amplified A. rubens
Fig. 2. Nucleotide and deduced amino-acid sequence of the cDNA encoding CMP-Neu5Ac hydroxylase from A. rubens. The products of the
RT-PCR and the 50-anchored PCR are indicated with an upper solid line and dotted underlining, respectively. The arrow marks the starting point of the
cDNA isolated from the library. Critical residues of the Kozak sequence surrounding the predicted start ATG are boxed. The liganding Cys and His
residues of the Rieske [2Fe2S]-cluster are printed bold. The postulated C-terminal membrane binding domain is underlined. The solid diamonds
indicate the end-points of the two truncated expression constructs (1, ARH:CD67; 2, ARH:CD21). The polyadenylation signal is printed in bold
italics.
5160 I. Martensen et al. (Eur. J. Biochem. 268) q FEBS 2001

cDNA, the latter was considered suitable for the generationof a specific probe to screen a library for cDNAs coding forthe A. rubens CMP-Neu5Ac hydroxylase.
Sequence of the isolated cDNA
Screening of the cDNA library from A. rubens gonads witha DIG-labelled probe resulted in the isolation of a positiveplaque from which a phagemid (pBK-CMV) with a cDNAinsert of 4075 bp was excised (pBCH1). Sequence analysisrevealed that the insert consisted of a 1869-bp ORF and a30-UTR of 2206 bp. The ORF was not complete at the 50-endbut overlapped with the 50-anchored PCR product, so thatthe whole cDNA sequence could be assembled (Fig. 2). Inthe 30-UTR, an AUUAAA polyadenylation signal waspresent, beginning 18 nucleotides upstream from thepoly(A) tail. Although this signal differed in one positionfrom the usual AAUAAA sequence, it neverthelessrepresents a common variant of similar activity [45].
The ORF of the cDNA encodes a protein of 653 aminoacids with a calculated molecular mass of 75 kDa. Analignment of the primary structure with other known CMP-Neu5Ac hydroxylase sequences is shown in Fig. 3. Thesequences of the mammalian hydroxylases are almostidentical, with a homology of more than 90%. The A. rubens
sequence exhibits a 60% identity with the mousehydroxylase. When conservatively exchanged residues areincluded, a homology of 70% is calculated. Despite thesesimilarities, several striking differences between the starfishand the mammalian sequences are apparent. Near theN-terminus, for example, the starfish sequence contains aninsertion of 12 residues starting at amino acid 40. Mostsignificantly, the C-terminus of the starfish enzyme is 60–70residues longer, depending on the mammalian sequenceused for comparison. This additional C-terminal sequencebegins with a region of < 40 amino acids exhibiting nounusual features and is followed by a stretch of 15hydrophobic residues. The final five C-terminal residues areall either charged or hydrophilic in nature. The possiblefunction of the C-terminus as a membrane anchor isaddressed in the Discussion.
Recombinant expression of the cDNA
To confirm the identity of the cloned cDNA, it was expressedin E. coli with a T7 promoter-driven, IPTG-inducibleexpression system consisting of the plasmid pET14b and theE. coli strain BL21(DE3)/pLysS. As aggregation frequentlycauses problems in the recombinant expression ofmembrane proteins, two constructs coding for truncated
Fig. 3. Comparison of the deduced amino-acid sequence of the A. rubens CMP-Neu5Ac hydroxylase with mammalian sequences. The
sequences were aligned with CLUSTALW and subsequently shaded with GENEDOC. Residues identical in at least five of the six sequences are shaded
grey, while homologous residues present in at least five of the six sequences are printed white on dark grey. The hamster sequence is incomplete at the
N- and C-termini. Box 1, binding site of the Rieske iron-sulfur centre; boxes 2 and 5, postulated binding sites of a mononuclear iron centre; box 3,
possible CMP-Neu5Ac binding site; box 4, possible site of interaction with cytochrome b5. Underlined amino acids indicate the postulated
transmembrane domain of the A. rubens hydroxylase. The GenBank accession numbers of the sequences used are: mouse, D21826; hamster,
AJ242835; pig, Y15010; chimpanzee, AF074481; macaque, AB013814.
q FEBS 2001 A. rubens CMP-Neu5Ac hydroxylase (Eur. J. Biochem. 268) 5161
forms of the hydroxylase were expressed in addition to thefull-length sequence (abbreviated to ARH for A. rubenshydroxylase). The first construct lacked the C-terminalstretch containing the hydrophobic and terminal hydrophilicregions (ending at Arg632; termed ARH:CD21). The secondlacked the complete C-terminal region and consisted of thepostulated catalytic domain (see Discussion), roughlyequivalent to the sequence of the mammalian enzyme(ending at Lys586; abbreviated to ARH:CD67). The insertswere ligated into pET14b in frame with an N-terminal His6
tag followed by a thrombin cleavage site, resulting in a2.2-kDa increase in the molecular mass of the recombinantprotein.
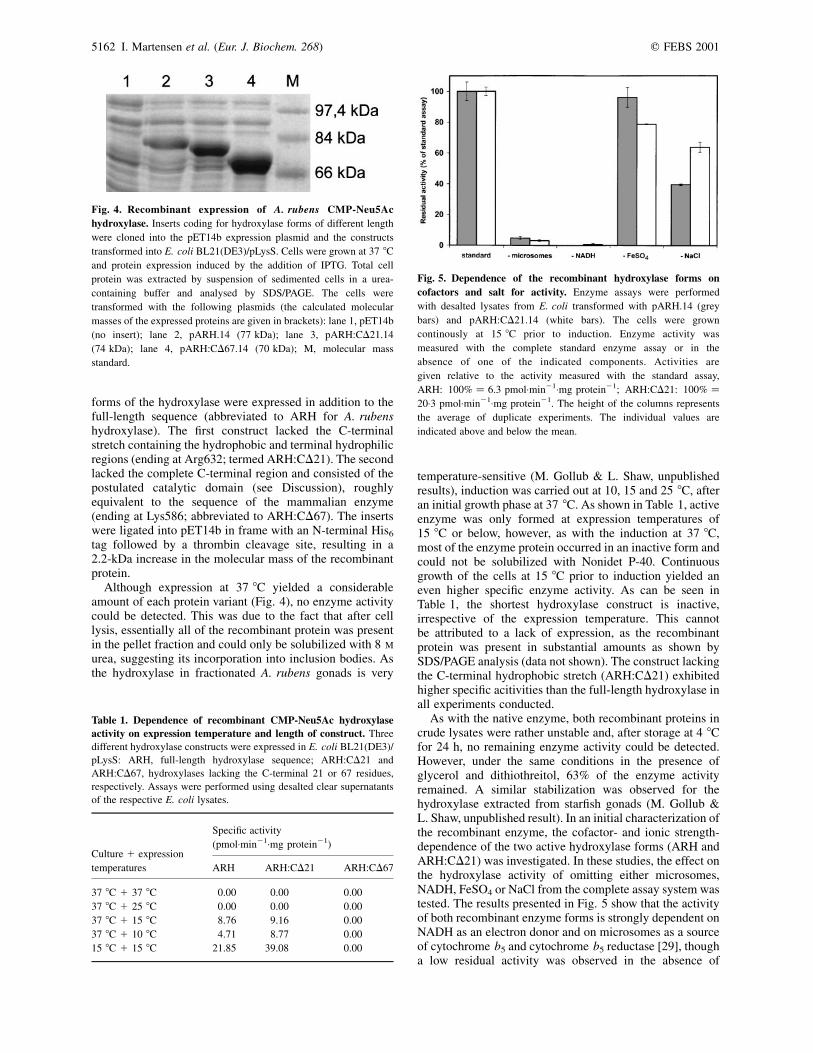
Although expression at 37 8C yielded a considerableamount of each protein variant (Fig. 4), no enzyme activitycould be detected. This was due to the fact that after celllysis, essentially all of the recombinant protein was presentin the pellet fraction and could only be solubilized with 8 M
urea, suggesting its incorporation into inclusion bodies. Asthe hydroxylase in fractionated A. rubens gonads is very
temperature-sensitive (M. Gollub & L. Shaw, unpublishedresults), induction was carried out at 10, 15 and 25 8C, afteran initial growth phase at 37 8C. As shown in Table 1, activeenzyme was only formed at expression temperatures of15 8C or below, however, as with the induction at 37 8C,most of the enzyme protein occurred in an inactive form andcould not be solubilized with Nonidet P-40. Continuousgrowth of the cells at 15 8C prior to induction yielded aneven higher specific enzyme activity. As can be seen inTable 1, the shortest hydroxylase construct is inactive,irrespective of the expression temperature. This cannotbe attributed to a lack of expression, as the recombinantprotein was present in substantial amounts as shown bySDS/PAGE analysis (data not shown). The construct lackingthe C-terminal hydrophobic stretch (ARH:CD21) exhibitedhigher specific acitivities than the full-length hydroxylase inall experiments conducted.
As with the native enzyme, both recombinant proteins incrude lysates were rather unstable and, after storage at 4 8Cfor 24 h, no remaining enzyme activity could be detected.However, under the same conditions in the presence ofglycerol and dithiothreitol, 63% of the enzyme activityremained. A similar stabilization was observed for thehydroxylase extracted from starfish gonads (M. Gollub &L. Shaw, unpublished result). In an initial characterization ofthe recombinant enzyme, the cofactor- and ionic strength-dependence of the two active hydroxylase forms (ARH andARH:CD21) was investigated. In these studies, the effect onthe hydroxylase activity of omitting either microsomes,NADH, FeSO4 or NaCl from the complete assay system wastested. The results presented in Fig. 5 show that the activityof both recombinant enzyme forms is strongly dependent onNADH as an electron donor and on microsomes as a sourceof cytochrome b5 and cytochrome b5 reductase [29], thougha low residual activity was observed in the absence of
Fig. 5. Dependence of the recombinant hydroxylase forms on
cofactors and salt for activity. Enzyme assays were performed
with desalted lysates from E. coli transformed with pARH.14 (grey
bars) and pARH:CD21.14 (white bars). The cells were grown
continously at 15 8C prior to induction. Enzyme activity was
measured with the complete standard enzyme assay or in the
absence of one of the indicated components. Activities are
given relative to the activity measured with the standard assay,
ARH: 100% ¼ 6.3 pmol:min21:mg protein21; ARH:CD21: 100% ¼
20:3 pmol:min21:mg protein21. The height of the columns represents
the average of duplicate experiments. The individual values are
indicated above and below the mean.
Fig. 4. Recombinant expression of A. rubens CMP-Neu5Ac
hydroxylase. Inserts coding for hydroxylase forms of different length
were cloned into the pET14b expression plasmid and the constructs
transformed into E. coli BL21(DE3)/pLysS. Cells were grown at 37 8C
and protein expression induced by the addition of IPTG. Total cell
protein was extracted by suspension of sedimented cells in a urea-
containing buffer and analysed by SDS/PAGE. The cells were
transformed with the following plasmids (the calculated molecular
masses of the expressed proteins are given in brackets): lane 1, pET14b
(no insert); lane 2, pARH.14 (77 kDa); lane 3, pARH:CD21.14
(74 kDa); lane 4, pARH:CD67.14 (70 kDa); M, molecular mass
standard.
Table 1. Dependence of recombinant CMP-Neu5Ac hydroxylase
activity on expression temperature and length of construct. Three
different hydroxylase constructs were expressed in E. coli BL21(DE3)/
pLysS: ARH, full-length hydroxylase sequence; ARH:CD21 and
ARH:CD67, hydroxylases lacking the C-terminal 21 or 67 residues,
respectively. Assays were performed using desalted clear supernatants
of the respective E. coli lysates.
Culture 1 expression
temperatures
Specific activity
(pmol:min21:mg protein21)
ARH ARH:CD21 ARH:CD67
37 8C 1 37 8C 0�.00 0�.00 0�.00
37 8C 1 25 8C 0�.00 0�.00 0�.00
37 8C 1 15 8C 8�.76 9�.16 0�.00
37 8C 1 10 8C 4�.71 8�.77 0�.00
15 8C 1 15 8C 21�.85 39�.08 0�.00
5162 I. Martensen et al. (Eur. J. Biochem. 268) q FEBS 2001

microsomes. While exogenous iron ions had little effect onthe ARH enzyme form, their omission led to a 20% decreasein the activity of ARH:CD21. Furthermore, both enzymetypes were more active in the presence of 100 mM NaCl, theARH and ARH:CD21 forms exhibiting a 2.5- and 1.5-foldincrease in activity, respectively.
D I S C U S S I O N
CMP-Neu5Ac hydroxylase sequence
Although the echinoderms emerged some 500 million yearsago, the amino-acid sequence of CMP-Neu5Ac hydroxylasefrom A. rubens exhibits an overall homology of < 70%to the primary structures of the mammalian enzymes. Notunexpectedly, the mammalian hydroxylases are highly relatedamong themselves, exhibiting more than 90% homology. Themajority of differences between the A. rubens and mamma-lian enzymes, as well as among the mammalian enzymesthemselves, are concentrated in the N- and C-terminalregions of their sequences, while the intervening portionsare particularly well conserved. The possible functional andstructural relevance of this pattern of sequence conservationis discussed in detail below.
N-Terminal region. The N-terminal region is defined here asthe first section of primary structure up to residue E141 ofthe A. rubens sequence. The start methionine residues of theA. rubens and the mammalian sequences do not coincide.However, the presence of the essential nucleotides of theKozak consensus sequence [44] in the starfish cDNA agreeswith the assigned starting point of translation (Fig. 2).
Although the first 60 amino acids exhibit some homologywith the mammalian enzymes, the starfish hydroxylasepossesses a 12 amino-acid insert (residues 40–51). Nospecific function can be ascribed to this insert. However, itshydrophilic character and the presence of many glutamateresidues exclude its involvement in membrane anchoring.
The presence of the Rieske sequence motif (Fig. 3, Box1)[34] is highly indicative for the presence of a Rieske [2Fe2S]centre in the A. rubens hydroxylase. Although the sequencesimmediately adjacent to the CXH and CXXH submotifsare conserved, the starfish hydroxylase does exhibit somesignificant differences in the intervening region. Accordingto crystal structure analyses of the a subunit of naphthalenedioxygenase [46], these residues interact with the Rieskecentre and may influence its redox properties. Although theCMP-Neu5Ac hydroxylase from mammals and A. rubenscatalyses the same reaction, the sequence differences mayreflect adaptations to the conditions and redox partnerswithin A. rubens cells.
Central region. The middle section of the hydroxylasesequences (i.e. the following 240 amino acids) is the mosthighly conserved. For the pig enzyme, some catalyticallyimportant primary structure regions were suggested to residein this area [32]. Residues potentially involved in thebinding of CMP-Neu5Ac (Fig. 3, box 3) and cytochrome b5
(Fig. 3, box 4) are both conserved between the starfish andmammalian hydroxylases. With respect to the binding sitefor a postulated mononuclear iron centre, two regions of thepig sequence (Fig. 3, boxes 2 and 5) were considered likelycandidates due to their similarity to an iron-binding motif
present in bacterial Rieske oxygenases. This motif generallyhas the sequence HX425H and, in all bacterial Rieskeoxygenases studied, is situated < 100 residues C-terminallyfrom the second histidine of the Rieske centre bindingsite [47,48]. Although both regions are conserved in themammalian and starfish CMP-Neu5Ac hydroxylases, box 2is favoured due to its position 120 amino acids C-terminal tothe Rieske-motif.
Due to the overall homology between the hydroxylasesfrom A. rubens and mammals, sequence comparisons aloneprovide no further validation of the significance of thesecentral primary structure regions.
C-terminal region. The C-terminus of the mammalianhydroxylase displays significant species-dependent differ-ences, suggesting that this region is not functionallyessential. However, the C-terminus of the A. rubenshydroxylase is < 60 residues longer than that of themammalian enzyme. A closer inspection of these additionalamino acids reveals a stretch of 15 consecutive hydrophobicresidues (amino acids 634–648; Figs 2 and 3). This part ofthe starfish hydroxylase presumably serves as the membraneanchor, as anticipated from the association of the enzymeextracted from the gonads of A. rubens with a particulatefraction [39]. The preceeding amino acids could form somekind of linker or hinge, connecting the membrane anchorwith the catalytic domain, which is expected to be roughlythe same size as the mammalian enzyme. The location of thepossible transmembrane domain (TMD) adjacent to theC-terminus suggests that the hydroxylase is a tail-anchoredprotein with a type IV membrane topology [49].
Because the cytochrome b5 form known to interactwith enzymes such as cytochrome P450 and fatty aciddesaturases is associated with the cytosolic face of theendoplasmic reticulum [30], it can be assumed that CMP-Neu5Ac hydroxylase is similarly located. This is supportedby the fact that targeting of tail-anchored proteins to theER is mediated by relatively short TMDs of less than 20amino acids [50,51]. As CMP-Neu5Ac hydroxylase infractionated tissues of other echinoderms is also membrane-bound [39], it is probable that the possession of such aC-terminal TMD is a common feature of this enzyme inother members of this phylum. The membrane-boundhydroxylase in echinoderms is thus a more typicalcytochrome-b5-dependent enzyme than its soluble mam-malian counterpart. The question therefore arises as towhen, in the course of evolution, CMP-Neu5Ac hydroxylaselost the C-terminal domain observed in the enzyme fromA. rubens and thus its ability to associate with a membrane.Furthermore, the functional consequences of this unexpectedchange in compartmentalization also requires elucidation.
Recombinant expression of A. rubens CMP-Neu5Achydroxylase
The recombinant expression of the complete ORF of theA. rubens cDNA in E. coli yielded CMP-Neu5Ac hydroxyl-ase activity. This result confirms unequivocally that thecDNA cloned encodes the hydroxylase, as suggested by thesignificant homology to the mammalian enzymes.
Influence of temperature and length of construct. An optimalexpression of active full-length enzyme (ARH) could only
q FEBS 2001 A. rubens CMP-Neu5Ac hydroxylase (Eur. J. Biochem. 268) 5163

be achieved at low cultivation and induction temperatures.This may reflect the temperature sensitivity observed for theenzyme isolated from fractionated gonads of A. rubens,though expression artefacts may be involved. The factthat under all the expression conditions tested the majorityof the protein could only be extracted under denaturingconditions suggests that E. coli is not able to correctly foldthis enzyme. The higher expression levels inducedby temperatures above 15 8C may therefore intensify thisproblem. The higher specific activity of lysates frombacteria expressing ARH:CD21 may be due to differences inexpression levels and the formation of correctly foldedenzyme.
The lack of activity of the hydroxylase variant with alength similar to that of the mammalian enzyme(ARH:CD67) suggests that the 47-residue linker domainmissing in this mutant protein may assist folding in E. coli ormay contain further structural and functional elements.Interestingly, the linker connecting the catalytic andN-terminal membrane-binding domain of cytochrome P4502C2 also seems to be essential for the correct folding of thisenzyme [52]. It remains to be investigated which residues inparticular are necessary to retain the activity of the starfishCMP-Neu5Ac hydroxylase.
Preliminary characterization of the recombinant hydroxyl-ase. As was shown for the enzyme from mouse, pig andA. rubens, optimal CMP-Neu5Ac hydroxylase activitydepends on the presence of NADH, the cytochromeb5/cytochrome b5 reductase system and Fe(II) ions[28,29,39]. Furthermore, the starfish hydroxylase requiresthe presence of 100 mM NaCl for maximum acitvity [39].An initial characterization of the active recombinant formsof the starfish enzyme showed that both proteins exhibitedqualitatively similar characteristics. Therefore, cytochromeb5/cytochrome b5 reductase, added in the form of pig livermicrosomes, and NADH are essential for hydroxylaseactivity. The trace of activity detectable in the absence ofmicrosomes may be due to nonspecific electron transferfrom another redox protein present in E. coli.
The influence of iron ions on the activity of the recom-binant hydroxylase is not as pronounced as expected fromstudies on the enzyme extracted from A. rubens. While thehydroxylase solubilized from starfish gonads is activated upto fivefold upon addition of FeSO4 [39], the omission ofFe(II) ions from the standard assay only led to a small, ifany, reduction in the activity of both recombinant enzymeforms. This may be due to different amounts of enzyme-bound iron in the respective extracts.
Although the activity of both recombinant forms isenhanced by 100 mM NaCl (Fig. 5), the extent of activationis lower than that of the enzyme from starfish gonads, whoseactivity is increased by 3.5-fold under these conditions [39].This may result from differences in the conformation of therecombinant enzymes or could be due to the presence of theHis6 tag and thrombin cleavage site. The different levels ofsalt activation observed for the complete hydroxylase andthe ARH:CD21 mutant suggest that the membrane-bindingdomain may be involved in governing the ionic-strengthdependence of this enzyme.
Despite these differences, the enzymic properties of bothrecombinant forms are qualitatively similar to those of theenzyme extracted from A. rubens and, as such, are suitable
for further investigations on the structure and function of theCMP-Neu5Ac hydroxylase from this species. Together withstudies on the mammalian hydroxylase, these investigationsmay shed light on the evolution and catalytic mechanism ofthis unique enzyme.
A C K N O W L E D G E M E N T S
The authors would like to thank Dr Marcus Gollub for many helpful
discussions regarding the properties of CMP-Neu5Ac hydroxylase from
A. rubens. This work was supported by the Deutsche Forschungsge-
meinschaft (grant Scha 202/29–1).
R E F E R E N C E S
1. Schauer, R., Kelm, S., Reuter, G., Roggentin, P. & Shaw, L. (1995)
Biochemistry and role of sialic acids. In Biology of the Sialic Acids
(Rosenberg, A., ed.), pp. 7–67. Plenum Press, New York.
2. Varki, A. (1992) Diversity in the sialic acids. Glycobiology 2,
25–40.
3. Kelm, S. & Schauer, R. (1997) Sialic acids in molecular and
cellular interactions. Int. Rev. Cytol. 175, 137–240.
4. Varki, A. (1992) Selectins and other mammalian sialic acid-binding
lectins. Curr. Opin. Cell Biol. 4, 257–266.
5. Roth, J., Kempf, A., Reuter, G., Schauer, R. & Gehring, W.J. (1992)
Occurrence of sialic acids in Drosophila melanogaster. Science
256, 673–675.
6. Malykh, Y.N., Krisch, B., Gerardy-Schahn, R., Lapina, E.B., Shaw,
L. & Schauer, R. (1999) The presence of N-acetylneuraminic acid
in Malpighian tubules of larvae of the cicada Philaenus spumarius.
Glycoconj. J. 16, 731–739.
7. Staudacher, E., Burgmayr, S., Grabher-Meier, H. & Halama, T.
(1999) N-glycans of Arion lusitanicus and Arion rufus contain
sialic acid residues. Glycoconj. J. 16, S114.
8. Warren, L. (1963) The distribution of sialic acids in nature. Comp.
Biochem. Physiol. 10, 153–171.
9. Corfield, A.P. & Schauer, R. (1982) Occurence of sialic acids. In
Sialic Acids – Chemistry, Metabolism and Function (Schauer, R.,
ed.), pp. 5–50. Springer Verlag, New York.
10. Schauer, R. (1982) Chemistry, metabolism, and biological
functions of sialic acids. Adv. Carbohydr. Chem. Biochem. 40,
131–234.
11. Schauer, R. & Kamerling, J.P. (1997) Chemistry, biochemistry and
biology of sialic acids. In Glycoproteins II (Montreuil, J.,
Vliegenthart, J.F.G. & Schachter, H., eds), pp. 243–402. Elsevier
Science B.V., Amsterdam.
12. Higashi, H., Hirabayashi, Y., Fukui, Y., Naiki, M., Matsumoto, M.,
Ueda, S. & Kato, S. (1985) Characterization of N-glycolyl-
neuraminic acid-containing gangliosides as tumor-associated
Hanganutziu–Deicher antigen in human colon cancer. Cancer
Res. 45, 3796–3802.
13. Kawai, T., Kato, A., Higashi, H., Kato, S. & Naiki, M. (1991)
Quantitative determination of N-glycolylneuraminic acid expres-
sion in human cancerous tissues and avian lymphoma cell lines as a
tumor-associated sialic acid by gas chromatography-mass spec-
trometry. Cancer Res. 51, 1242–1246.
14. Bergwerff, A.A., Hulleman, S.H., Kamerling, J.P., Vliegenthart,
J.F., Shaw, L., Reuter, G. & Schauer, R. (1992) Nature and
biosynthesis of sialic acids in the starfish Asterias rubens.
Identification of sialo-oligomers and detection of S-adenosyl-
L-methionine:N-acylneuraminate 8-O-methyltransferase and CMP-
N-acetylneuraminate monooxygenase activities. Biochimie 74,
25–37.
15. Kamerling, J.P., Schauer, R., Vliegenthart, J.F. & Hotta, K. (1980)
Identification of the sialic acids from the egg jelly coat of the sea
5164 I. Martensen et al. (Eur. J. Biochem. 268) q FEBS 2001

urchin Pseudocentrotus depressus (Okayama). Hoppe Seylers
Z. Physiol. Chem. 361, 1511–1516.
16. Klein, A., Diaz, S., Ferreira, I., Lamblin, G., Roussel, P. & Manzi,
A.E. (1997) New sialic acids from biological sources identified by a
comprehensive and sensitive approach: liquid chromatography-
electrospray ionization-mass spectrometry (LC-ESI-MS) of SIA
quinoxalinones. Glycobiology 7, 421–432.
17. Kubo, H., Irie, A., Inagaki, F. & Hoshi, M. (1990) Gangliosides
from the eggs of the sea urchin, Anthocidaris crassispina.
J. Biochem. (Tokyo) 108, 185–192.
18. Muralikrishna, G., Reuter, G., Peter-Katalinic, J., Egge, H.,
Hanisch, F.G., Siebert, H.C. & Schauer, R. (1992) Identification of
a new ganglioside from the starfish Asterias rubens. Carbohydr.
Res. 236, 321–326.
19. Smirnova, G.P., Kochetkov, N.K. & Sadovskaya, V.L. (1987)
Gangliosides of the starfish Aphelasterias japonica, evidence for a
new linkage between two N-glycolylneuraminic acid residues
through the hydroxy group of the glycolic acid residue. Biochim.
Biophys. Acta 920, 47–55.
20. Shaw, L. & Schauer, R. (1988) The biosynthesis of N-glycoloyl-
neuraminic acid occurs by hydroxylation of the CMP-glycoside
of N-acetylneuraminic acid. Biol. Chem. Hoppe Seyler 369,
477–486.
21. Shaw, L. & Schauer, R. (1989) Detection of CMP-N-acetylneur-
aminic acid hydroxylase activity in fractionated mouse liver.
Biochem. J. 263, 355–363.
22. Schauer, R., Malykh, Y.N., Krisch, B., Gollub, M. & Shaw, L.
(1999) Biosynthesis and Biology of N-glycolylneuraminic Acid. In
Sialobiology and Other Novel Forms of Glycosylation (Inoue, Y.,
Lee, Y.C. & Troy, F.A. II, eds), pp. 17–27. Gakushin Publishing
Co., Osaka.
23. Chou, H.H., Takematsu, H., Diaz, S., Iber, J., Nickerson, E.,
Wright, K.L., Muchmore, E.A., Nelson, D.L., Warren, S.T. &
Varki, A. (1998) A mutation in human CMP-sialic acid hydroxylase
occurred after the Homo-Pan divergence. Proc. Natl Acad. Sci.
USA 95, 11751–11756.
24. Irie, A., Koyama, S., Kozutsumi, Y., Kawasaki, T. & Suzuki, A.
(1998) The molecular basis for the absence of N-glycolylneur-
aminic acid in humans. J. Biol. Chem. 273, 15866–15871.
25. Kawano, T., Kozutsumi, Y., Kawasaki, T. & Suzuki, A. (1994)
Biosynthesis of N-glycolylneuraminic acid-containing glycoconju-
gates. Purification and characterization of the key enzyme of the
cytidine monophospho-N-acetylneuraminic acid hydroxylation
system. J. Biol. Chem. 269, 9024–9029.
26. Schneckenburger, P., Shaw, L. & Schauer, R. (1994) Purification,
characterization and reconstitution of CMP-N-acetylneuraminate
hydroxylase from mouse liver. Glycoconj. J. 11, 194–203.
27. Schlenzka, W., Shaw, L., Schneckenburger, P. & Schauer, R. (1994)
Purification and characterization of CMP-N-acetylneuraminic acid
hydroxylase from pig submandibular glands. Glycobiology 4,
675–683.
28. Kozutsumi, Y., Kawano, T., Yamakawa, T. & Suzuki, A. (1990)
Participation of cytochrome b5 in CMP-N-acetylneuraminic acid
hydroxylation in mouse liver cytosol. J. Biochem. (Tokyo) 108,
704–706.
29. Shaw, L., Schneckenburger, P., Schlenzka, W., Carlsen, J.,
Christiansen, K., Jurgensen, D. & Schauer, R. (1994) CMP-
N-acetylneuraminic acid hydroxylase from mouse liver and pig
submandibular glands. Interaction with membrane-bound and
soluble cytochrome b5-dependent electron transport chains. Eur.
J. Biochem. 219, 1001–1011.
30. Vergeres, G. & Waskell, L. (1995) Cytochrome b5, its functions,
structure and membrane topology. Biochimie 77, 604–620.
31. Malykh, Y.N., Krisch, B., Shaw, L., Warner, T.G., Sinicropi, D.,
Smith, R., Chang, J. & Schauer, R. (2001) Distribution and
localization of CMP-N-acetylneuraminic acid hydroxylase and
N-glycolylneuraminic acid-containing glycoconjugates in porcine
lymph node and peripheral blood lymphocytes. Eur. J. Cell Biol.
80, 48–58.
32. Schlenzka, W., Shaw, L., Kelm, S., Schmidt, C.L., Bill, E.,
Trautwein, A.X., Lottspeich, F. & Schauer, R. (1996) CMP-
N-acetylneuraminic acid hydroxylase: the first cytosolic Rieske
iron-sulphur protein to be described in Eukarya. FEBS Lett. 385,
197–200.
33. Kawano, T., Koyama, S., Takematsu, H., Kozutsumi, Y., Kawasaki,
H., Kawashima, S., Kawasaki, T. & Suzuki, A. (1995) Molecular
cloning of cytidine monophospho-N-acetylneuraminic acid
hydroxylase. Regulation of species- and tissue-specific expression
of N-glycolylneuraminic acid. J. Biol. Chem. 270, 16458–16463.
34. Mason, J.R. & Cammack, R. (1992) The electron-transport proteins
of hydroxylating bacterial dioxygenases. Annu. Rev. Microbiol. 46,
277–305.
35. Rathinasabapathi, B., Burnet, M., Russell, B.L., Gage, D.A., Liao,
P.C., Nye, G.J., Scott, P., Golbeck, J.H. & Hanson, A.D. (1997)
Choline monooxygenase, an unusual iron-sulfur enzyme catalyzing
the first step of glycine betaine synthesis in plants: prosthetic group
characterization and cDNA cloning. Proc. Natl Acad. Sci. USA 94,
3454–3458.
36. Caliebe, A., Grimm, R., Kaiser, G., Lubeck, J., Soll, J. & Heins, L.
(1997) The chloroplastic protein import machinery contains a
Rieske-type iron-sulfur cluster and a mononuclear iron-binding
protein. EMBO J. 16, 7342–7350.
37. Gray, J., Close, P.S., Briggs, S.P. & Johal, G.S. (1997) A novel
suppressor of cell death in plants encoded by the Lls1 gene of
maize. Cell 89, 25–31.
38. Schlenzka, W., Shaw, L. & Schauer, R. (1993) Catalytic properties
of the CMP-N-acetylneuraminic acid hydroxylase from the starfish
Asterias rubens: comparison with the mammalian enzyme.
Biochim. Biophys. Acta 1161, 131–138.
39. Gollub, M., Schauer, R. & Shaw, L. (1998) Cytidine monophos-
phate-N-acetylneuraminate hydroxylase in the starfish Asterias
rubens and other echinoderms. Comp. Biochem. Physiol.
B. Biochem. Mol. Biol. 120, 605–615.
40. Shaw, L., Schneckenburger, P., Carlsen, J., Christiansen, K. &
Schauer, R. (1992) Mouse liver cytidine-50-monophosphate-
N-acetylneuraminic acid hydroxylase. Catalytic function and
regulation. Eur. J. Biochem. 206, 269–277.
41. Martensen, I., Gollub, M., Schauer, R. & Shaw, L. (2000) Cloning
and expression of the cDNA encoding CMP-N-acetylneuraminic
acid hydroxylase from the starfish Asterias Rubens. In Abstracts of
the 20th International Carbohydrate Symposium, C-191. (Thiem,
J., ed.), p. 92, LCI Publisher GmbH, Hamburg.
42. Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman,
J.G., Smith, J.A. & Struhl, K. (2001) Current Protocols in
Molecular Biology. John Wiley & Sons, New York.
43. Dorit, R.L. & Ohara, O. (1987) cDNA amplification using one-
sided (anchored) PCR. In Current Protocols in Molecular Biology
(Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman,
J.G., Smith, J.A. & Struhl, K., eds), John Wiley & Sons, Inc., New
York.
44. Kozak, M. (1989) The scanning model for translation: an update.
J. Cell Biol. 108, 229–241.
45. Wickens, M. (1990) How the messenger got its tail: addition of
poly(A) in the nucleus. Trends Biochem. Sci. 15, 277–281.
46. Kauppi, B., Lee, K., Carredano, E., Parales, R.E., Gibson, D.T.,
Eklund, H. & Ramaswamy, S. (1998) Structure of an aromatic-ring-
hydroxylating dioxygenase 2 naphthalene-1,2-dioxygenase. Struc-
ture 6, 571–586.
47. Butler, C.S. & Mason, J.R. (1997) Structure–function analysis of
the bacterial aromatic ring-hydroxylating dioxygenases. Adv.
Microb. Physiol. 38, 47–84.
48. Neidle, E.L., Hartnett, C., Ornston, L.N., Bairoch, A., Rekik, M. &
Harayama, S. (1991) Nucleotide sequences of the Acinetobacter
calcoaceticus benABC genes for benzoate-1,2-dioxygenase reveal
q FEBS 2001 A. rubens CMP-Neu5Ac hydroxylase (Eur. J. Biochem. 268) 5165

evolutionary relationships among multicomponent oxygenases.
J. Bacteriol. 173, 5385–5395.
49. Spiess, M. (1995) Heads or tails 2 what determines the orientation
of proteins in the membrane. FEBS Lett. 369, 76–79.
50. Yang, M., Ellenberg, J., Bonifacino, J.S. & Weissman, A.M. (1997)
The transmembrane domain of a carboxyl-terminal anchored
protein determines localization to the endoplasmic reticulum.
J. Biol. Chem. 272, 1970–1975.
51. Honsho, M., Mitoma, J.Y. & Ito, A. (1998) Retention of
cytochrome b5 in the endoplasmic reticulum is transmem-
brane and luminal domain-dependent. J. Biol. Chem. 273,
20860–20866.
52. Chen, C.D., Doray, B. & Kemper, B. (1997) Efficient assembly of
functional cytochrome P450 2C2 requires a spacer sequence
between the N-terminal signal anchor and catalytic domains. J. Biol.
Chem. 272, 22891–22897.
5166 I. Martensen et al. (Eur. J. Biochem. 268) q FEBS 2001
Copyright © 2022 FDOKUMEN




















