
.APPELLATE CIVIL. MAUNG PO ZAW and others ^ E Mating for the ...
Upload
independentCategory
view
3download
0
Cloacal anatomy of paedomorphic female Ambystoma talpoideum (Caudata: Ambystomatidae), with comments on intermorph mating and sperm storage
STANLEY E. TRAUTH Department of Biological Sciences, Arkansas State University, State University, AR 72467, U. S. A.
DAVID M. SEVER Department of Biology, Saint Mary's College, Notre Dame, IN 46556, U. S. A.
AND
RAYMOND D. SEMLITSCH Division of Biological Sciences, University of Missouri, Columbia, MO 6521 1 , U. S. A.
Received January 26, 1994 Accepted August 12, 1994
TRAUTH, S. E., SEVER, D. M., and SEMLITSCH, R. D. 1994. Cloacal anatomy of paedomorphic female Ambystoma talpoideum (Caudata: Ambystomatidae), with comments on intermorph mating and sperm storage. Can. J. Zool. 72: 2 147 - 2 157.
The cloacal anatomy of paedomorphic female Ambystoma talpoideum was examined using light microscopy and histochemi- cal techniques. Three types of cloacal glands (anterior ventral glands, dorsal glands, and spermathecae) were observed. Glandular morphology and secretory activity were documented from seasonal samples collected in South Carolina. Maximal development of cloacal glands corresponded to a peak in reproductive activity. Sperm-storage glands, the spermathecae, con- tained sperm in specimens collected from November through February. Although greatly hypertrophied during the breeding season, the anterior ventral glands continue to release secretory material throughout the year. This suggests that these glands could function in capacities other than those related to reproduction. The rudimentary dorsal glands also contain sperm during the breeding season; however, the occurrence of sperm in these glands is considered to be incidental. The cloacal glands of paedomorphs are similar to those of metamorphs. Interbreeding between morphs may occur and thus facilitate the main- tenance of both morphs in nature.
TRAUTH, S.E., SEVER, D.M., et SEMLITSCH, R.D. 1994. Cloacal anatomy of paedomorphic female Ambystoma talpoideum (Caudata: Ambystomatidae), with comments on intermorph mating and sperm storage. Can. J. Zool. 72 : 2147-2157.
L'anatomie du cloaque des femelles pCdomorphes d'Ambystoma talpoideum a CtC CtudiCe par microscopie photonique et au moyen de techniques histochimiques. Trois types de glandes cloacales (glandes ventrales antkrieures, glandes dorsales et spermathkques) ont CtC observks. La morphologie des glandes et leur activitk sCcrCtrice ont CtC CtudiCes sur des Cchantillons saisonniers de Caroline du Sud. Le dCveloppement maximal des glandes cloacales correspond a une recrudescence de l'acti- vitC reproductrice. Les glandes de rCserve de spermatozoi'des, les spermathkques, contiennent du sperme chez les femelles recueillies entre novembre et la fin de fkvrier. Trks hypertrophiCes au cours de la saison de reproduction, les glandes ventrales antkrieures font une sCcrCtion continue durant toute I'annCe, ce qui indique que ces glandes ne sont pas strictement relikes a la reproduction. 11 y a Cgalement des spermatozoi'des dans les glandes dorsales rudimentaires pendant la saison de reproduc- tion, mais leur presence dans ces glandes parait &re accidentelle. Les glandes cloacales des individus pCdomorphes sont semblables a celles des salamandres qui se mCtamorphosent. Les salamandres pCdomorphes et les salamandres a metamor- phose peuvent se reproduire entre elles, ce qui assure la survie des deux formes en nature.
[Traduit par la RCdaction]
Introduction Ambystoma talpoideum is a common species of salamander
in the Atlantic Coastal Plain and Mississippi Valley of the United States, ranging from Virginia to northern Florida and west to Texas, Arkansas, and Tennessee (Shoop 1960; Conant and Collins 1991). It is a facultatively paedomorphic species with populations consisting of both terrestrial metamorphic adults and aquatic paedomorphic adults (Patterson 1978; Semlitsch 1985a). Both morphs utilize ponds for breeding; the terrestrial morphs migrate to sites in South Carolina during the winter months (November - January), whereas the paedo- morphic adults are present in ponds all year (Semlitsch 1985b). Courtship and oviposition by the terrestrial morph generally occur during December, January, and February (Semlitsch 1985a), but some paedomorphic individuals are known to breed earlier in some years (Scott 1993). Females oviposit up to 1000 eggs (F = 441), which are scattered singly across the pond bottom on leaf litter, twigs, and debris (Semlitsch 1985a), but geographic variation in egg-laying strategy has also been reported (Semlitsch and Walls 1990). Breeding population size at one pond ranged from 333 to 2596
over a 6-year period, with a nearly 1: 1 sex ratio each year (Semlitsch 1987). After the breeding season is finished in March, metamorphic adults migrate to surrounding terrestrial habitats (a 100- to 300-m distance), where they remain fos- sorial until the next reproductive season. Individuals can breed more than once in their lifetime (1 -6 times) and typically return to the same pond each season (Semlitsch et al. 1993).
Female A. talpoideum, like most salamanders, have sperm- storage glands in the walls of their cloaca (Duellman and Trueb 1986; Sever 199 la). Eggs are fertilized in the cloaca by sperm released from the spermathecae during oviposition (Boisseau and Joly 1975). Few observations are available on the duration of sperm storage both before and after mating. Because most ambystomatids migrate from terrestrial habitats to breeding ponds over a period of several weeks, a relatively short period of sperm storage probably occurs between mating and egg laying. Although retention of sperm in the sperma- thecae after oviposition has been reported for some sala- manders (Baylis 1939; Houck and Schwenk 1984; Verrell and Sever 1988; Massey 1990; Sever 1991 b; Sever and Kloepfer 1993), most reports have included no evidence of storage of
Printed in Canada 1 lrnprirne au Canada
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

2 148 CAN. J . ZOOL. VOL. 72, 1994
residual sperm between breeding seasons in other species of salamanders (Verrell and Sever 1988; Peccio 1992; Sever 1992a; Sever and Brunette 1993; Sever et al. 1995). Indeed, the spermathecal epithelium may be actively spermiophagic (Sever 1992a; Sever and Brunette 1993 ; Sever and Kloepfer 1993). Baylis (1 939), however, reported production of suc- cessive broods of young nearly 2 years apart by an isolated Salamandra salarnandra whose last mating was at least 2- 2.5 years before the birth of her second brood. Massey (1 990) also reported that female Notophthalmus viridescens retain sperm between successive breeding seasons, although the via- bility of such sperm was unknown. The only ambystomatid species in which the seasonal cycle of sperm storage has been studied is Ambystoma opacum (Sever and Kloepfer 1993; Sever et al. 1995). This species is unique in the family in that breeding occurs on land, and mating may occur before arrival to the pond and nesting area (Sever et al. 1995). Still, no evi- dence of mating > l month prior to oviposition has been pre- sented for this species, and sperm persist no longer than 6 months after oviposition (Sever et al. 1995).
The objectives of the present study were to describe the seasonal variation in cloacal anatomy of female paedomorphic A. talpoideum and to determine if paedomorphic females store sperm for more than one breeding season. The length of sperm storage for individual females was not determined; thus, the length of time sperm were present is a cumulative estimate for the population. Our test of sperm storage between breeding seasons is conservative and maximizes the opportunity for sperm storage because females were collected from the field and had had ample opportunity to mate several times or late in the breeding season.
Materials and methods Periodic monthly samples of paedomorphic Ambystoma talpoideum
were taken from October 1980 through November 1983 at four sites on the U.S. Department of Energy's Savannah River Site in Aiken and Barnwell counties, South Carolina (Gibbons and Semlitsch 1991). Two additional samples of salamanders were taken in August 1985. Collections were made at two permanent natural ponds (Ellen- ton Bay and Flamingo Bay) known to have large breeding populations of paedomorphic individuals (Patterson 1978; Semlitsch 1 9 8 5 ~ ) and at two permanent experimental ponds, A and B, also containing breeding populations of paedomorphic individuals originating from Ellenton Bay and Sun Bay, respectively (Sernlitsch and Wilbur 1989). Samples were taken using a dip net or unbaited wire minnow traps. salamanders were sacrificed in-dilute Chloretone solution in the fkld and immediately fixed in a solution of 10% neutral buffered formalin. Snout-vent length (SVL; from the tip of the snout to the anterior margin of the vent) of specimens was measured following preserva- tion and storage in 70% ethanol.
Cloacae from 29 females were prepared for histological examination using light microscopy. Blocks of cloacal tissue, including portions of the digestive and urogenital tracts, were dissected from the body of each female. The tissue blocks were then subjected to routine histotechniques as outlined in Humason (1979). In brief, tissues were dehydrated in a graded series of ethanol and cleared with xylene; this was followed by trimming of tissues and infiltration in paraffin for 10 h. Paraffin embedding in plastic molds allowed for the orientation of tissue masses to enhance both transverse and sagittal sectioning of paraffin blocks. Blocks were trimmed of excess paraffin, and ribbons of material, serially sectioned using a rotary microtome, were affixed to slides using Haupt's adhesive. Slides were then alternately stained with hematoxylin and eosin (H&E, for general cytology), Pollac trichrome stain (PTS, for connective tissues and mucins), and Alcian
blue at pH 2.5 (AB, for acid mucosubstances). Strong positive stain- ing reactions using PTS and AB on cloacal gland secretions appeared deep red for mucins and light blue for acid mucopolysaccharides, respectively, whereas alternate or neutral staining reactions were light blue and pink, respectively. Some sections from each specimen were stained with AB at pH 2.5 and then counterstained with periodic acid - Schiff's reagent (PAS, indicative of neutral carbohydrates). In this procedure, PAS-positive substances stain magenta, AB-positive substances stain turquoise-blue, and secretory material that is both PAS- and AB-positive is purple (Sheehan and Hrapchak 1980; Kiernan 1990).
Identification and assignment of cloacal glandular clusters were according to Sever (199 l a , 1992b). For each specimen, 10 or more diameters of the largest alveolar portions of each gland were mea- sured (to the nearest 0. l pm) using an ocular micrometer. In the case of dorsal glands, which are structurally rudimentary and are of unknown function in many salamanders (Sever 1991a), only selected glandular clusters were examined or measured. The reproductive condition (presence or absence of vitellogenic ova) as well as the presence or absence of sperm within spermathecae and dorsal glands were recorded for each female.
Results
Gross anatomy We describe the cloacal anatomy of a 51 mm SVL paedo-
morphic female Ambystoma talpoideum collected on 25 Novem- ber 1981 from experimental pond A. A midventral incision (extending from the vent into the cloacal chamber and anter- iorly onto the chest cavity) exposed the preovulatory config- uration of the internal organs of this gravid specimen.
Upon lifting and reflecting the urinary bladder from its expanded position (approximately 10 mm anteriorly from its juncture with the cloaca), a highly constricted rectum of the large intestine was observed. The rectum (in this case mostly devoid of fecal material) was narrowly squeezed (and pushed ventrally) by enlarged vitellogenic ovarian follicles, whereas greatly expanded and convoluted glandular oviducts occupied the region distally on either side (posterolaterally). (All speci- mens from late November through February possessed vitello- genic ovarian follicles > 2 mm in diameter.) The most posterior segments of the oviducts are narrowed as they extend dorsally and medially to lie above the rectum before each empties into the cloacal tube through genital papillae. Mesonephric (opistho- nephric) kidneys reside along the dorsal body wall in close proximity to the oviducts.
Microscopic anatomy The major anatomical structures within the cloacal region
(as mentioned above) can also be viewed microscopically via a sagittal section as illustrated in Fig. 1, showing the positions of the three types of cloacal glands relative to one another. The following is a descriptive summary detailing the morphologi- cal features of the cloacal complex shown in Figs. 2 and 3 (from a late-November specimen, No. 58; Table l) , starting at the level of the juncture between the neck of the urinary bladder (Ub) and the large intestine (In).
As the Ub and In merge, the anteriormost clusters of anterior ventral glands (Av) are found squeezed between bands of abdominal musculature (Mu) and the abdominal cavity (Fig. 2A). Secretions fill the lumina of most of these glands (see Cloaca1 glands and their secretions). Dorsolateral to the Ub are the narrowed posterior segments of the oviducts (Ov*; Figs. 2A, 2D). The internal lining of these mostly circu-
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

TRAUTH ET AL.
FIG. 1. Cloaca1 anatomy (sagittal section) of a paedomorphic female Ambystoma talpoideum collected in late November, showing the cloacal glands at their maximum development (using Pollac trichrome stain). Av, anterior ventral glands; Ct, cloacal tube; Dg, dorsal glands; Gp, genital papillae; In, large intestine; Lf, lateral fold of the cloacal tube; Me, mesonephric kidney; Ov, oviduct (glandular portion); St, spermathecae; Ub, neck of urinary bladder; V, vent. The asterisk indicates the opening of the oviduct into the cloacal chamber.
lar tubes takes on a rugose configuration compared with the anterior-gland-laden portions of the oviducts (Ov). A ciliated columnar epithelium characterizes the lining of the rugae. Paired archinephric ducts (Ad) of the mesonephric kidney (Me), are positioned dorsally and dorsolaterally, respectively, to the Ov*. The intestinal epithelium exhibits a pseudostrati- fied columnar cell layer dominated by numerous round goblet cells.
In Fig. 2B, taken at a point 320 pm posterior to the section shown in Fig. 2A, the glandular oviducts have mostly disap-
peared within the abdominal cavity (Ac). The cloacal tube (Ct) lacks the profusion of goblet cells that characterizes the In, but retains the convoluted walls. The anterior ventral glands are now more conspicuous beneath the Ct.
The pair of genital papillae (Gp) hangs downward into the Ct (Figs. 2C -2F, starting approximately 100 pm posterior to the end of the section in Fig. 2B). The papillae open separately into the Ct; the left side (Fig. 2D) empties anterior to the right (Fig. 2F). The passageway between the Ov* and the Ct (as shown in Figs. 2D and 2E) is lined with a ciliated, pseudo-
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

2 150 CAN. J. ZOOL. VOL. 72, 1994
FIG. 2. Anterior to posterior transverse sections through the cloacal region of a paedomorphic female Ambystoma talpoideum collected in late November. See the text for an explanation of abbreviations and symbols. Scale bar (A-F) = 1 mm. (A) Section through the anterior region of the cloacal complex, showing the structures at the juncture between the urinary bladder and the large intestine. H&E stain. (B) Section showing the beginning of the cloacal tube. AB stain. (C) Section showing a genital papilla inside the cloacal tube. AB stain. (D) Section in the region of the genital papillae, showing a common pathway betwen the left oviduct and the cloacal tube. H&E stain. (E) Section stained with PTS, showing both genital papillae at the level of the anterior groove of the vent. (F) Section revealing the beginning of the cloacal chamber above the vent opening. AB stain.
stratified columnar epithelium, as is most of the midventral exhibits a stratified cuboidal epithelium three or four cell floor of the Ct. In Fig. 2 E , the more lateral surfaces of the layers in thickness. Ct are mostly composed of nonciliated, low, simple columnar In Fig. 2F (a region approximately 0.5 mm posterior to the cells. On the other hand, the external surfaces of the Gp end of the section in Fig. 2D), the anterior cloacal chamber
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

TRAUTH ET AL. 2151
FIG. 3. Anterior to posterior transverse sections through the cloacal region of a paedomorphic female Ambystoma talpoideum. See the text for an explanation of abbreviations. Scale bar (A-E) = 1 mm. (A-E) Continuation of Fig. 2. (A) In this section the right archinephric duct empties into the cloacal tube. At the same time, the anterior ventral glands separate into lateral clusters within the cloacal lips. AB stain. (B) Section showing lateral diverticula and folds of the cloacal tube at the same level when spermathecae initially appear above the cloacal tube. PTS stain. (C) In this section the cloacal tube and cloacal chamber merge, resulting in a continuous growth from the vent to the openings of the spermathecal tubules. PTS stain. (D) Section taken in the posterior region of the cloaca, showing the anterior ventral glands opening into the cloacal chamber. AB stain. (E) Section near the posterior wall of the cloaca, showing large numbers of spermathecae and the lack of anterior ventral glands. Note the presence of a bulging mass of tissue within the cloacal chamber; this bulge possesses openings into the dorsal glands. H&E stain. (F) Sagittal section of the cloacal complex of a late-November specimen, showing the relative positions of the three types of cloacal glands. In this section, the dorsal glands contain sperm. PTS stain. Scale bar = 300 pm.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

2 152 CAN. J. ZOOL. VOL. 72, 1994
TABLE 1 . Seasonal variation in diameter (pm) of cloacal glands in paedomorphic female Ambystoma talpoideum
Spermathecae Dorsal glands Anterior ventral
Date of Specimen SVL Sperm glands Sperm collection No. (mm) Y + 2 SE present? (X f 2 SE) X f 2 SE present?
23 Jan. 1981 24 Jan. 1982 21 Feb. 1981 21 Feb. 1981 21 Feb. 1981 19 Apr. 1981 4 May 1983
15 May 1981 15 May 1981 9 June 1983 9 June 1983 9 June 1983
26 July 1983 26 July 1983 18 Aug. 1983 18 Aug. 1983 6 Sept. 1983 6 Sept. 1983 6 Sept. 1983
18 Oct. 1980 17 Nov. 1981 25 Nov. 1983 25 Nov. 1983 25 Nov. 1983 25 Nov. 1983 25 Nov. 1983
9 Dec. 1980 9 Dec. 1980
16 Dec. 1981
Yes Yes Yes Yes Yes No No No No -
No No No No No No No No No No No Yes Yes Yes Yes Yes Yes Yes Yes
Yes -
- -
-
- No -
-
- - -
- -
- - -
-
-
No Yes -
- Yes Yes Yes Yes Yes
(Cc) makes its first appearance just above the anterior margin of the vent (V). The Cc, lined with stratified squamous cells, receives secretions from the Av.
The right Ad can be seen emptying into the Ct in Fig. 3A (a continuation of Fig. 2 at a point approximately 450 pm posterior to Fig. 2F). In Fig. 3B, the first spermathecae (St) appear at around 180 pm posterior to Fig. 3A. The floor of the Ct has now begun to expand laterally to form distinct folds and recesses. When the latter occurs, as in Fig. 3C, the Av are divided into two separate lateral clusters within the cloacal lips. The Cc also exhibit conspicuous foldings as this cavity begins to push dorsally to meet the Ct (Fig. 3B). The Ct retains prominent lateral folds (Lf) as well as a mid-dorsal central groove (Cg), as shown in Fig. 3C. This region communicates with all of the St (also shown in Fig. 3D). The Ct and Cc merge soon after the appearance of the St.
The paired lateral folds of the cloacal tube (Lf; as viewed in Fig. 3D at a point 300 pm posterior to Fig. 3C) are retained laterally within the posterior cloacal walls. Each Lf continues to possess a ciliated, pseudostratified columnar epithelium, whereas both the Cg (lined with a stratified transitional epithelium of low columnar cells) and the Cc (lined with strati- fied squamous epithelium) lack cilia. Within the posterior cloacal chamber in Fig. 3E (approximately 300 pm posterior to Fig. 3D), the Cg has been lost and the grooves within the Lf have extended ventrally to surround an outpocketing of the posterior cloacal wall. This bulge can be observed lying directly beneath the Lf in Fig. 1. Moreover, this region is
marked by the presence of numerous ductal openings into dor- sal glands (Dg; see Fig. I). The Av, so prevalent within the anterior ventral cloacal wall and the anterior cloacal lips, have all but disappeared. Although the St are quite numerous where they are embedded in the roof of the posterior cloacal cham- ber, none of their ducts open into this region of the cavity. The separation between the Dg and the St is revealed in Fig. 3F; sperm are present in the Dg of Figs. 1, 3E, and 3F. The entire cloacal complex, measured from Figs. 2A through 3E, is roughly 2.2 mm in length.
Cloaca1 glands and their secretions In terms of structural design, two of the three types of cloacal
glands (i.e., Av and St) of paedomorphic A. talpoideum are of the simple, tubuloalveolar, exocrine variety (Sever 19926); each exhibited tubules having an expanded secretory distal portion and a narrowed proximal duct. The Dg differ slightly from these two by generally lacking expanded alveolar regions. The method of secretion is similar in all three types; Av liberate in a more or less continuous manner (merocrine mode of secretion), and their secretions accumulate as globu- lar masses within the distal lumina. Secretions by the St are also of the merocrine type; however, these substances were discharged in a manner that resulted in distal segments having a thin-walled, evacuated, vesicular appearance (Figs. 1 and 4). When the St exhibited an extremely festooned morphology, their secretions may have been absent or may have only par- tially filled portions of the distal regions, yet their proximal
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.
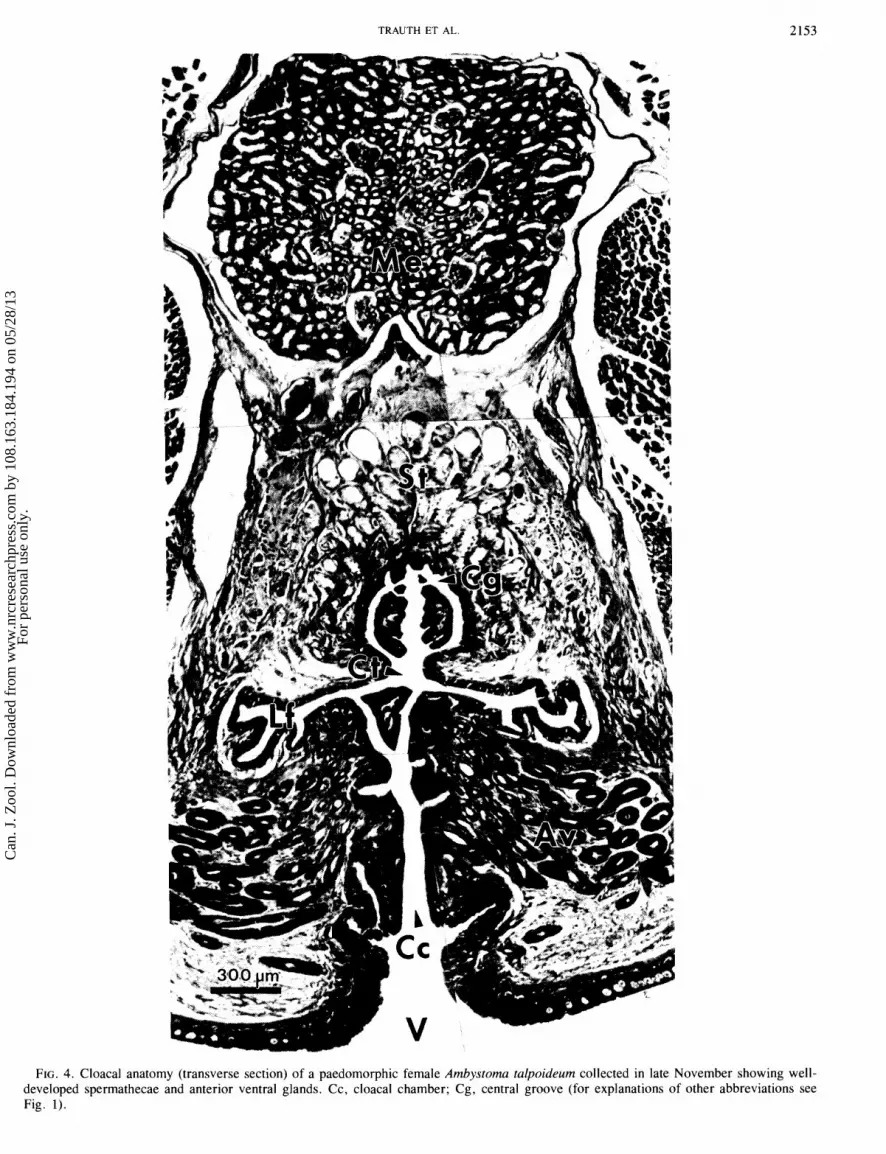
TRAUTH ET AL.
FIG. 4. Cloaca1 anatomy (transverse section) of a paedomorphic female Ambystoma talpoideum collected in late November showing well- developed spermathecae and anterior ventral glands. Cc, cloaca1 chamber; Cg, central groove (for explanations of other abbreviations see Fig. 1).
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

CAN. J . ZOOL. VOL. 72, 1994
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

TRAUTH ET AL. 2155
duct regions usually possessed secretory materials throughout their entire length. The Dg have no secretory products within the lumina.
At the peak of reproductive activity, which corresponds to maximal cloacal gland development, a gravid January speci- men (Table 1; No. 2) possessed Av that extended through about 2.3 mm of cloacal tissue. Approximately 80- 100 glands composed the anteriormost Av cluster of distal segments (Fig. 2D); the majority of these segments contained a globular secretion (H&E neutral, PTS- and PAS-positive, AB-negative), whereas the lumina of the proximal ducts were devoid of materials. Droplets within the cytoplasm of the simple cuboidal epithelium lining the distal segments revealed a combination of staining reactions from faintly red to mostly blue-green, whereas the ductal epithelium appeared purple with PTS. The 100- 120 St with sperm amassed within their secretions (H&E neutral, PTS- and PAS-negative, AB-positive) were observed emptying into the Cg. Both the cuboidal epithelium and the secretions of the St were blue, whereas in the same region Av secretions were light pink. Within the posterolateral wall of the cloaca, there was a sharp demarcation line between the narrow proximal ducts of the St and the Dg based upon their staining reactions. About 100 St resided above the 75 -90 Dg. Although the Dg also exhibited sperm, their epithelial linings were PTS-positive, PAS-positive (slight), and AB-negative.
The PAS reaction in Av was very strong. During the November - February period, when the glands were hyper- trophied, the apical cytoplasm as well as the lumen contained much secretory material, and the luminal masses were in the form of globules (Fig. 5). But from March to August there was a less intense reaction in the apical cytoplasm (a spotty appear- ance), whereas the globules were absent; the lumina were nar- row. The St were intensely AB-positive in specimens collected from November to February, with positively staining materials in either the cytoplasm or the lumen (Fig. 5). From March to July, only a slight AB-positive reaction occurred in the apical cytoplasm. The Dg gave a scant (slight) reaction with PAS around the luminal borders. There was very little change in secretory activity of the Dg throughout the year.
Discussion The overall cloacal anatomy of paedomorphic Ambystoma
talpoideum was similar to that of a metamorphosed female briefly described by Sever (1992b), except that the central groove in the paedomorphs extended farther into the cloacal chamber. The staining reactions and cytology of the glands were also similar, suggesting similar functions. Licht and Sever (1991) reported no difference in cloacal anatomy between metamorphosed and paedomorphic Ambystoma gracile and concluded that the two morphs of that species should be cap- able of interbreeding. Our data suggest that this is true for A. talpoideum also, and that reproductive isolation between
morphs does not occur on the basis of cloacal anatomy. More- over, this suggests that any reproductive isolation can occur only through other mechanisms such as assortative mating or temporal isolation. The ability to interbreed also has important ecological implications if offspring fitness is increased by occasional interbreeding of morphs. For example, if a paedo- morphic female produces more metamorphic offspring by mating with a metamorphic male rather than with a paedo- morphic male, she increases the probability that offspring will survive in a drying pond during a drought year. Thus, similar- ity in cloacal anatomy enhances the possibility of interbreeding of morphs under certain evironmental conditions and could facilitate the maintenance of both morphs through alternating selection on mate choice.
The continuation of secretory activity in the anterior ventral glands may indicate their importance in activities other than breeding. Sever (1988) hypothesized that the ventral glands secrete a pheromone and that this may be the ancestral func- tion of the cloacal glands. We speculate that the anterior ven- tral glands of A. talpoideum may be involved in territorial marking or individual recognition throughout the year. The continued secretory activity, even at a reduced level, may be what is expected of glands of general importance in daily activity, but could have added significance (as when they hypertrophy) during a certain time of the year. Recent behav- ioral studies of aggression and defense of resident sites (e.g., refugia, burrows) by metamorphic ambystomatid salamanders suggest that chemical cues may be important (Ducey and Ritsema 1988; Walls 1990). Walls and Roudebush (1991) also demonstrated kin recognition in A. opacum, which also sug- gests that secretory substances might be important for behav- ioral interactions of larvae during foraging.
We found that some sperm also occur in the dorsal glands. The only other report of sperm occurring in cloacal glands other than spermathecae was by Hardy and Dent (1986), who found sperm in the ventral glands of the newt N. viridescens (which lacks dorsal glands). They proposed that sperm move thigmotactically along the cloacal epithelium and, upon encountering the openings of a gland, are carried through these pores by a continuation of the thigmotactic response. Few sperm were found in the ventral glands of N. viridescens, however, possibly because their lumina were occluded with secretory products (Hardy and Dent 1986). In A. talpoideurn, the dorsal glands were largely posterior to the spermathecae, and some sperm would encounter the openings of these glands before those of the spermathecae. Entry into the dorsal glands might be restricted, however, by their more minute pores and narrow lumina. No one has provided evidence that the sperma- thecae secrete a substance that attracts sperm.
Our results show that sperm are present in the spermathecae of all females during the breeding season (November - March) and that sperm storage may occur for a period up to 5 months. No females contained sperm during the nonbreeding period
FIG. 5. Histosections showing seasonal variation in the spermathecae (A-E) and anterior ventral glands (F-J) in paedomorphic female Ambystoma talpoideum. Scale bar in A and F are the same for A -E and F -J, respectively. (A) Late-November specimen exhibiting masses of sperm (Sp) within tubules. Mc, melanocyte. PTS stain. (B) Relatively few sperm (Sp) are present in a mid-February specimen. AB stain. (C) Regressed glands of an early May specimen; no sperm are present. PTS stain. (D) Hypertrophy of epithelial cell nuclei in a September specimen. H&E stain. (E) Glandular activity in a specimen collected in mid-October reveals cytological preparation for the reception of sperm. AB stain. (F) Maximal secretory activity in late-November specimen, showing globular masses (arrowheads) within the lumina of glands. PTS stain. (G) Highly regressed condition of the glands in early May. H&E stain. (H) Total regression in August specimen; numerous melanocytes (Mc) are interspersed among the glands. H&E stain. (I) Recrudescence of glands in September; epithelial cell hypertrophy is evident. PTS stain. (J) September specimen exhibiting increased cytological activity within the glands. AB stain.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

2 156 CAN. J . ZOOL. VOL. 72, 1994
(May -October). This is a maximal estimate for sperm storage because individual females were not followed over the breed- ing season. We suspect that the actual time for which individ- uals can store sperm is considerably less. A recent study of a closely related species, A. opacum, indicates that sperm begin to degenerate within a month, and none persist for more than 6 months after oviposition (Sever et al. 1995).
The presence of sperm storage for even a short time has direct implications concerning sperm competition and sexual selection. Sperm competition is highly probable if females (Parker 1970; Houck and Schwenk 1984) (i) mate more than once before the eggs are fertilized, (ii) store inseminated sperm, (iii) retain inseminated and viable sperm for longer than one breeding season, or (iv) use sperm sparingly. For birds (Birkhead and Moller 1992), it has been proposed that the presence of sperm storage may compensate for the lack of synchrony between copulation and fertilization in the mating system, or may simply be the phylogenetic consequence of a particular ancestry (i.e., reptilian in the case of birds). For salamanders, the presence of sperm storage could be advan- tageous when the probability of finding a mate each year is low, as in fossorial, nonmigrating species (i.e., some pletho- dontid salamanders). Sperm storage in A. talpoideum and other migrating species is probably not necessary to ensure fertilization each year because of the high concentration of individuals at breeding ponds. We suggest that short-term sperm storage like that found in ambystomatid salamanders may be advantageous in allowing environmentally dependent oviposition in aquatic habitats. Once inseminated, females can wait until aquatic conditions are optimal to maximize the sur- vival of embryos and larvae in the pond. For instance, ovi- positing too early in the season can result in the loss of eggs due to pond freeze (Harris 1980; R.D. Semlitsch, unpublished data). The decoupling of insemination from oviposition facili- tates reproductive success in variable aquatic environments such as ponds.
However, as has been found in other species, any sperm storage increases the probability of sperm competition (Birk- head and Moller 1982). At present, it is unknown whether female A. talpoideum pick up more than one spermatophore or mate with more than one male, thereby producing clutches fer- tilized by more than one male (but see Arnold 1977). Research on these and related topics of sperm release from the sperma- thecae or dorsal glands should increase our understanding of the mating system and sexual selection in this facultatively paedomorphic species.
Acknowledgements We thank J.W. Gibbons for his help and permission to col-
lect specimens at Flamingo Bay and Ellenton Bay. Collection of salamanders was supported by Department of Energy con- tract DE-AC09-76SR00-8 19 with the University of Georgia's Savannah River Ecology Laboratory. Support was provided to S.E.T. by the Department of Biological Sciences, Arkansas State University, and to D.M.S. by National Science Founda- tion grant DEB-90249 18.
Arnold, S.A. 1977. The evolution of courtship behavior in New World salamanders with some comments on Old World sala- manders. In The reproductive biology of amphibians. Edited by D.H. Taylor and S.I. Guttman. Plenum Press, New York. pp. 141 -183.
Baylis, H .A. 1939. Delayed reproduction in the spotted salamander. Proc. Zool. Lond. Sect. A, 109: 243 -246.
Birkhead, T.R., and Moller, A.P. 1982. Sperm competition in birds: evolutionary causes and consequences. Academic Press, London.
Boisseau, C., and Joly, J. 1975. Transports and survival of sper- matozoa in female Amphibia. In The biology of spermatozoa: transport, survival and fertilizing ability. Edited by E. S.E. Hafex and C.G. Thibault. Karger, Basel. pp. 94- 104.
Conant, R., and Collins, J.P. 1991. A field guide to reptiles and amphibians of eastern and central North America. Houghton Mifflin Co., Boston.
Ducey, P.K., and Ritsema, P. 1988. Intraspecific aggression and responses to marked substrates in Ambystoma maculatum (Caudata: Ambystomatidae). Copeia, 1988: 1008 - 10 13.
Duellman, W.E., and Trueb, L. 1986. Biology of amphibians. McGraw-Hill, New York.
Gibbons, J.W., and Semlitsch, R.D. 1991. Guide to the reptiles and amphibians of the Savannah River Site. University of Georgia Press, Athens.
Hardy, M.P., and Dent, J.N. 1986. Transport of sperm within the cloaca of the female red-spotted newt. J. Morphol. 190: 259 -270.
Harris, R.N. 1980. The consequences of within-year timing of breed- ing in Ambystoma maculatum. Copeia, 1980: 7 19 - 922.
Humason, G.L. 1979. Animal tissue techniques. 4th ed. W.H. Freeman, San Francisco.
Houck, L.D., and Schwenk, K. 1984. The potential for long-term sperm competition in a plethodontid salamander. Herpetologica, 40: 410-415.
Kiernan, J .A. 1990. Histological and histochemical methods. 2nd ed. Pergamon Press, Oxford, U.K.
Licht, L.E., and Sever, D.M. 1991. Cloaca1 anatomy of metamor- phosed and neotenic salamanders. Can. J. Zool. 69: 2230 -2233.
Massey, A. 1990. Notes on the reproductive energy of red-spotted newts (Notophthalmus viridescens). Copeia, 1990: 106 - 107.
Parker, G . A. 1970. Sperm competition and its evolutionary conse- quences in the insects. Biol. Rev. Camb. Philos. Soc. 45: 525 - 568.
Patterson, K.K. 1978. Life history aspects of paedogenic populations of the mole salamander, Ambystoma talpoideum. Copeia, 1978: 649 - 655.
Peccio, A. 1992. Insemination and egg laying dynamics in the smooth newt, Triturus vulgaris, in the laboratory. Herpetol. J. 2: 5 -7.
Scott, D.E. 1993. Timing of reproduction of paedomorphic and metamorphic Ambystoma talpoideum. Am. Midl. Nat. 129: 397- 402.
Semlitsch, R.D. 1985~. Reproductive strategy of a facultatively paedomorphic salamander Ambystoma talpoideum. Oecologia, 65: 305-313.
Semlitsch, R.D. 1985b. Analysis of climatic factors influencing migrations of the salamander Ambystoma talpoideum. Copeia, 1985: 477-489.
Semlitsch, R.D. 1987. Relationship of pond drying to the reproduc- tive success of the salamander Ambystoma talpoideum. Copeia, 1987: 61 -69.
Semlitsch, R.D., and Walls, S.C. 1990. Geographic variation in the egg-laying strategy of the mole salamander, Ambystoma tal- poideum. Herpetol. Rev. 21: 14.- 15.
Semlitsch, R.D., and Wilbur, H.M. 1989. Artificial selection for paedmorphosis in the salamander Ambystoma talpoideum. Evolu- tion, 43: 105- 112.
Semlitsch, R.D., Scott, D.E., Pechmann, J.H.K., and Gibbons, J. W. 1993. Phenotypic variation in the arrival time of breeding salamanders: individual repeatability and environmental influ- ences. J. Anim. Ecol. 62: 334-340.
Sever, D.M. 1988. The ventral gland in female salamander Eurycea bislineata (Amphibia: Plethodontidae). Copeia, 1988: 572 -579.
Sever, D.M. 1991a. Comparative anatomy and phylogeny of the cloacae of salamanders (Amphibia: Caudata) . I. Evolution at the family level. Herpetologica, 47: 165 - 193.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.

TRAUTH ET AL. 2157
Sever, D.M. 1991b. Sperm storage and degradation in the sperma- thecae of the salamander Eurycea cirrigera (Green). J . Morphol . 210: 71-84.
Sever, D. M. 1992a. Spermiophagy of the spermathecal epithelium of the salamander Eurycea cirrigera. J. Morphol. 212: 28 1 -290.
Sever, D.M. 1992b. Comparative anatomy and phylogeny of the cloacae of salamanders (Amphibia: Caudata) . VI. Ambystomatidae and Dicamptodontidae. J. Morphol. 212: 305 -322.
Sever, D.M., and Brunette, N.S. 1993. Regionalization of eccrine and spermiophagic activity in the spermathecae of the salamander Eurycea cirrigera (Amphibia: Plethodontidae). J. Morphol. 217: 161 - 170.
Sever, D.M., and Kloepfer, N .M. 1993. Spermathecal cytology of Ambystoma opacum (Amphibia: Ambystomatidae) and the phylo- geny of sperm storage in female salamanders. J. Morphol. 217: 115- 127.
Sever, D.M., Krenz, J.D., Johnson, K.M., and Rania, L.C. 1995.
Morphology and evolutionary implications of the annual cycle of secretion and sperm storage in spermathecae of the salamander Ambystoma opacum. J. Morphol. 223: 35-46.
Sheehan, D.C., and Hrapchak, B.B. 1980. Theory and practice of histotechnology . 2nd ed: Battell Press, Columbus, Ohio.
Shoop, C.R. 1960. The breeding habits of the mole salamander, Ambystoma talpoideum (Holbrook), in southeastern Louisiana. Tulane Stud. Zool. 8: 65 -82.
Verrell, P.A., and Sever, D.M. 1988. The cloaca and spermatheca of the female smooth newt, Triturus vulgaris (Amphibia: Sala- mandridae). Acta Zool. 69: 65 - 70.
Walls, S.C. 1990. Interference competition in postmetamorphic salamanders: interspecific differences in aggression by coexisting species. Ecology, 71: 307-314.
Walls, S .C., and Roudebush, R.E. 199 1. Reduced aggression towards siblings as evidence of kin recognition in cannibalistic salamanders. Am. Nat. 138: 1027 - 1038.
Can
. J. Z
ool.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
108.
163.
184.
194
on 0
5/28
/13
For
pers
onal
use
onl
y.
Copyright © 2022 FDOKUMEN




















