
Alternative male mating behaviour in the two-spotted spider mite: dependence on age and density
Upload
independentCategory
view
1download
0
Behav Ecot Sociobiol (1992) 31:63-71 Behavioral Ecology and Sociobiology © Springer-Verlag 1992
Extra-pair copulations in a lek: the secondary mating system of monogamous razorbills Richard H. Wagner*
Edward Grey Institute of Field Ornithology, South Parks Road, Oxford OXI 3PS, United Kingdom
Received October 24, 1991 / Accepted January 26, 1992
Summary. I studied the mating strategies of razorbills AIca torda, a monogamous colonial seabird, during 1987-1989 in Wales, U.K. The outstanding feature of the study population's mating system was the perfor- mance of most copulations in mating arenas outside the colony. Females visited the arenas and sometimes ac- cepted extra-pair copulations (EPCs), and males aggres- sively competed for EPCs. During the fertilizable period of most females, the arenas resembled leks because they contained no resources except mates, and males contrib- uted only sperm to extra-pair females. Typical of leks, the operational sex ratio was strongly male-biased, and male mating success was highly skewed. To examine the potential for sexual selection to operate in this system, I correlated male EPC success with variables that could be associated with male-male competition and/or female choice. The frequencies of three behaviors (interference of copulation attempts, fights won against other males, and EPC attempts), accounted for 62% of the variance in EPC success and were strongly intercorrelated, with interference explaining most of the variance in a multi- variate test. EPC success was not correlated with body size, age, paired status or either of two estimates of the time spent in the arena. The hypothesis that male EPC success was determined by active female choice was tested by examining the relative success (EPCs per at- tempt) of males. Relative success was not correlated with any of the three behaviors associated with absolute suc- cess, or with age, body size or attendance, suggesting that male-male competition, mainly in the form of copu- lation interference, is the principal correlate of EPC suc- cess. These findings in a monogamous species illustrate that EPC is a secondary mating system of razorbills in which sexual selection operates. The existence of this system outside the colony raises the possibility that lekk- ing for EPCs may also occur within the nesting territo- ries of other monogamous, colonial species but is "hid- den" by competition for nests and breeding partners.
* Present address." Department of Zoological Research, National Zoological Park, Smithsonian Institution, Washington, D.C. 20008, USA
Introduction
Sexual selection occurs most intensely in polygamous species (Darwin 1871), in which mating success is often highly skewed (Emlen and Oring 1977; Beehler and Fos- ter 1988). However, the mechanisms of sexual selection may also operate in monogamous species (Trivers 1972), especially when females cooperate in extra-pair copula- tions (EPCs) (Moller 1988; Hamilton 1990). While there is growing evidence that monogamous female birds pursue EPCs (common murre, Uria aalge, Hatchwell 1988; northern fulmar, Fulrnarus glacialis, Hatch 1987; barn swallow, Hirundo rustica. Moller 1988; black- capped chickadee, Parus atricapillus, Smith 1988) and that EPCs may account for a substantial percentage of fertilizations (Sherman and Morton 1988; Blakey 1989; Brooker et al. 1990; Gibbs et al. 1990; Westneat 1990), the significance of EPCs for sexual selection has not yet received much attention.
My study of razorbills (Alca torda) has presented an opportunity for viewing EPC as a distinct secondary mating system in which sexual selection may operate. Unique among colonial birds studied to date, razorbills in a marked population on Skomer Island, Wales, per- formed most of their copulations in mating arenas out- side their colony (Wagner 1992). During the prelaying period, the arenas effectively functioned as "colonies without nests" in which males and females in monoga- mous pairs attended synchronously and copulated. Dur- ing the laying period, however, the arenas' characteris- tics drastically change as female attendance decreases, resulting in a sharp male-biased skew in the sex ratio which coincides with aggressive male-male competition for EPCs. All paired females breeding in the study col- ony visited the arenas where 96% received EPC attempts and 33% accepted 1-7 EPCs, but only when their mates were absent (Wagner 1992); these EPCs were accepted voluntarily, as males required the cooperation of the female to achieve insemination (Wagner 1991a). As I report here, the clustering of males in competition for EPCs during the laying period and the pursuit of EPCs

64
by females have produced conditions similar to those which exist in leks.
Leks are assemblages of males which females visit for copulation; leks do not contain any resources except mates, and males do not provide parental care (Brad- bury and Gibson 1983). Lek systems are of interest :for their intensity of sexual selection (Beehler and Foster 1988) and because they enable researchers to identify traits which correlate with mating success without the confounding factors of additional male contributions (Bradbury and Gibson 1983).
Colonies of monogamous birds have been compared to leks because in some colonies males congregate and display to attract females (Draulans 1988). While the dense distributions of males and females in colonies may aid in acquiring breeding partners (Draulans 1988; Burger and Gochfeld 1990), high-density nesting may also increase the opportunity of males to attempt EPCs (MacRoberts 1973; Birkhead 1978; Beecher and Beecher 1979; Pierotti 1981; Moller 1985; Birkhead et al. 1987; Hatchwell 1988; Morton et al. 1990). For colonial birds, a more useful analogue to lek matings may be EPCs rather than pair-bonds because in monogamous species males can pair with only one female, whereas male mat- ing success can be highly skewed for EPCs (Westneat et al. 1990). Possible similarities between colonies and leks may not be obvious because of the occurrence of more than one resource for males to defend within the breeding territory, e.g. within colonies males may com- pete over territories as well as EPCs. The pursuit of EPCs by razorbills in arenas outside the colony excludes nesting-territory defense as a cause of male-male aggres- sion.
Lek studies often analyse male mating success in terms of the two modes of sexual selection identified by Darwin (1871), male-male competition and female choice, and the approach of many researchers is to iden- tify correlates of male mating success (e.g. H6glund and Lundberg 1987; Appolonio etal. 1989; McDonald 1989). Within a large body of literature on EPCs, several recent studies have identified such correlates of male EPC success as fighting ability (while ibis, Eudocimus aIbus; Frederick 1987), dominance rank (black-capped chickadee; Smith 1988), tail-length (barn swallows; Mallet 1988) and age (purple martins, Progne subis; Morton et al. 1990).
My objective in this paper is to describe the unusual feature of the razorbill mating system - the performance of copulations in extra-colonial arenas - to suggest an underlying similarity between leks and the colonies of other monogamous birds which pursue EPCs. More spe- cifically, my aim is treat razorbill EPCs performed in arenas as a promiscuous lek-type mating system in which sexual selection operates. In so doing, I correlate male EPC success with behavioral and morphological vari- ables which can be associated with male-male competi- tion and/or female choice.
Methods
I studied razorbills on Skomer Island, Wales, U.K. (51040 , N, 05°15' W) in 1987-1989 in the Basin colony. Behavioral observa- tions of ca. 100 color-banded individuals were made at two mating arenas (" lower" and "upper") which comprised horizontal ledges 3 m apart, which were located approximately 30 m outside the main breeding colony. The proximity of the two arenas allowed for their simultaneous observation. I observed the arenas every morning in 1988 (from 4 April to 14 May, and on 16, 19, 22 and 24 May) for a total of 100 h, and in 1989 (from 1 April to 20 May, and on 22 and 26 May) for a mean of 3 h (range 1-5 h) and a total of 125 h. I watched from a hide at a distance of 25 m with a 20-60x60 spotting scope, beginning at first light (ca. 0500 hours), usually before any razorbitls had arrived from roost- ing at sea and continuing until most or all razorbills had departed from the arenas, or until their mating activities had virtually ceased, usually before 0900-1000 hours. I recorded the time of arrival (or first sighting) of each marked bird and calculated mean arrival times (as the number of minutes since 0000 hours) for individuals in 1989 who attended on at least 10 days.
Extra-pair copulations. Both paired and unpaired males may in- crease their fitness by fertilizing the mates of other males (Trivers 1972). In this paper, the operational definition of an EPC is a potentially fertilizable copulation between any male and a paired female who is not his mate. EPCs were successful if the female permitted the male to achieve cloacal contact and, presumably, insemination. I considered EPCs to be potentially fertilizable if they were successful and occurred prior to egg-laying. Elsewhere, I have reported that females whose egg-laying dates were known only permitted EPCs to succeed prior to egg-laying, with all EPC attempts after laying failing to result in cloacal contact (Wagner 1991 a). Therefore, in this study I assume for a larger sample of females (with and without known egg-laying dates) that EPC at- tempts which were successful had occurred prior to egg-laying.
It was easy to distinguish EPCs from copulations between pair members. Mates were clearly identifiable because throughout most of the female's prelaying period the male and female often arrived simultaneously and consistently attended the arenas together, sit- ting side to side and allopreening, Mates also copulated frequently, with pairs averaging approximately 50 copulation attempts in the arenas during the 30 days prior to egg-laying (Wagner 1991 b). The identity of most pairs was also confirmed by observing the breeding colony, where males and females presumed to be mates in the arenas also shared nesting duties (Wagner, manuscript submitted).
Male-male competition. In order to evaluate the role of male-male competition in EPC success I recorded the frequencies of two forms of aggression and correlated them with the number of EPCs at- tained by males. "Interference" occurred when a male disrupted a copulation attempt, usually by lunging at the mounting male with his bill and forcing him to dismount from the female. "Fights" were rapid bouts of aggression which were initiated by one bird lunging at another with its bill. An individual was assigned a win if he displaced his opponent or if he succeeded in stabbing him with his bill without receiving retaliation. Interference was treated as a separate variable from fights. I also calculated an index of fighting ability as the percentage of fights won out of total wins and losses (Frederick 1987) and correlated this index with EPC success.
Female choice. If male EPC success is influenced by the preferences of females, then males with preferred traits should achieve a higher proportion of EPCs per attempt. I examined this by correlating relative success (EPCs divided by the total number of attempts) with male morphological and behavioral variables. Relative success was calculated for males who made 4 or more EPC attempts.
Laying dates. I recorded egg-laying dates from daily inspections of those breeding sites which were visible through a telescope. For
enclosed sites (e.g. those under boulders), I calculated laying dates by subtracting the mean incubation period of 34 days (Wagner 1991b) from the date of hatching; the hatching date was deter- mined during daily feeding watches in 1989 and was assumed to be the first day on which a fish was brought to the nest to feed the hatchling.
Male age. It is impossible to age adult razorbills except by banding them as nestlings and observing them in future years. The ages of some adults in the study population were known because band- ing has been carried out since 1969 by Dr. C.M. Perrins and his assistants• Razorbills do not commence breeding until the age of 4 years and usually not until 5 (Lloyd and Perrins 1977)• Thus, an individual banded as an adult may be assigned a minimum age of 4 years from the age of banding• However, razorbills are long-lived, with a maximum recorded age of 26 years (Mead and Clark 1988) and with several studies reporting annual survivorship of over 90% (Hudson 1985)• Due to this longevity, low minimum ages are not very informative, because males assigned a minimum age of 4 years have a high probability of being much older• To minimize this source of error I only analyzed the EPC success of males with relatively high minimum ages of 11 or more years and of males with known ages (which ranged from 4 to 14 years).
Morphometrics. When adults were trapped for color-banding I made 11 measurements of their body size. I used dial calipers for all linear measurements (to within 0•1 mm), except wing chord, for which I used a stop-ended wing rule. The following were meas- ured: (1) body mass: within 2 g using a 1000 g pesola spring bal- ance; (2) wing chord: unstretched, to within 1 mm; (3) bill length (culmen): from the end of feathering on the upper mandible to the tip of the bill; (4) bill depth: from the highest point of the upper mandible to the lowest mandible; (5) gape length: from th corner of the mouth to the tip of the upper mandible; (6) gonys (the lowermost ridge of the lower mandible): from end of feather- ing to the tip of the lower mandible; (7) eye-stripe: the white strips which runs from above the eye to the edge of the upper mandible; (8) head length: from the most distal point behind the head to the tip of the upper mandible; (9) head width: the widest part of the head, from the edges of a pair of supraorbital ridges; (10) tarsus length: distance between the patella and the tarsometatarsus (including the depth of the tarsometatarsus); (11) middle toe length: from where the toe meets the tarsometatarsus and up to and excluding the claw.
Analyses. Before reporting the results of parametric tests, the resid- uals of linear analyses were plotted to ascertain that they approxi- mated a normal distribution. Variables in the form of percentages (e.g. sex ratio) were arcsine-transformed for parametric analyses•
Resul t s
Seasonal changes in sex ratio
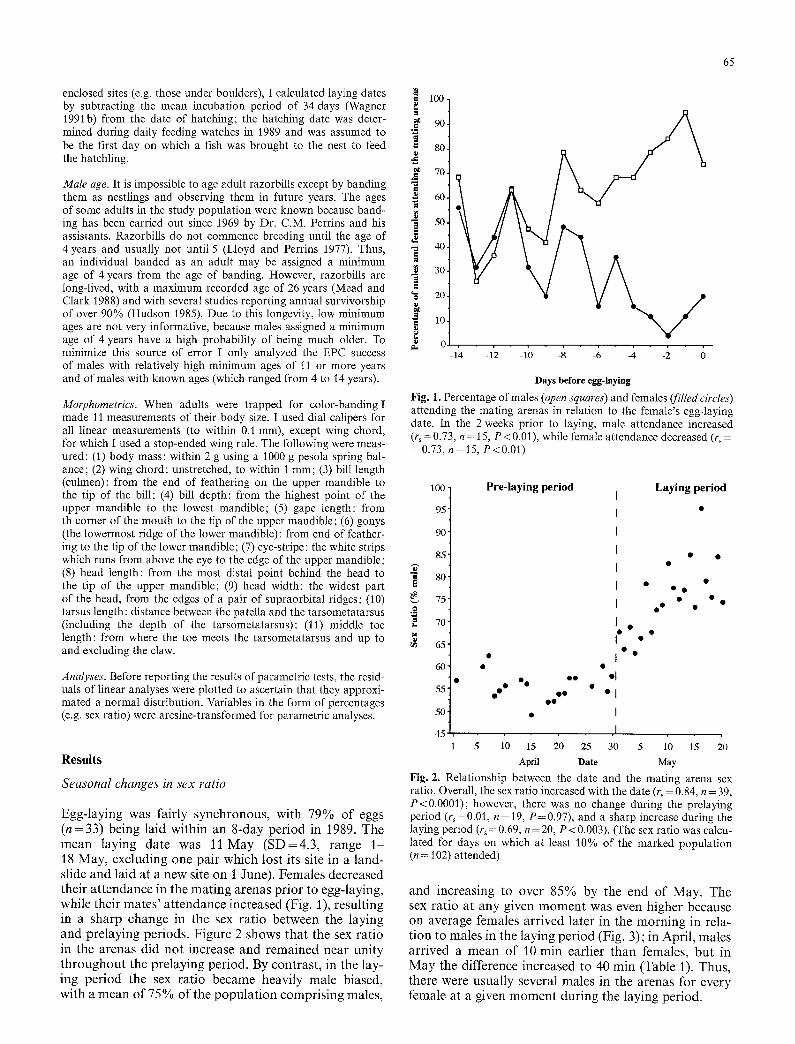
Egg-laying was fairly synchronous , with 79% of eggs ( n = 3 3 ) being laid wi th in an 8-day per iod in 1989. The m e a n laying date was 11 May ( S D = 4 . 3 , range 1 18 May, excluding one pair which lost its site in a land- slide and laid at a new site on 1 June). Females decreased their a t t endance in the ma t ing arenas pr ior to egg-laying, while their mates ' a t tendance increased (Fig. 1), result ing in a sharp change in the sex rat io between the laying and pre laying periods. Figure 2 shows that the sex rat io in the arenas did no t increase and remained near un i ty t h roughou t the prelaying period. By contrast , in the lay- ing per iod the sex rat io became heavily male biased, with a m e a n of 75% of the popu la t ion compr is ing males,
65
~- 100 o
90
$ 80
70 II e~ 60
i 50 ,~ 40
30 N
20
t0
0 g~ -14 -12 -10 -8 -6 4 -2 0
Days before egg-laying
Fig. 1. Percentage of males (oven squares) and females (filled circles) attending the mating arenas in relation to the female's egg-laying date. In the 2 weeks prior to laying, male attendance increased (r~ = 0.73, n = 15, P < 0.01), while female attendance decreased (r~ = -0.73, n= 15, P<0.01)
g
i00-
95-
90-
85"
80
75
70"
65"
60"
55"
50-
45
Pre-laying period Laying period
O • •
LD •
O0 O 0 O]
0 O0 O0 • [ oLD
• I
.I 5 l0 15 20 25 30 5 I0 15 20
April Date May
Fig. 2. Relationship between the date and the mating arena sex ratio• Overall, the sex ratio increased with the date (r~ = 0.84, n = 39, P<0.0001); however, there was no change during the prelaying period (r~=0.01, n= 19, P=0.97), and a sharp increase during the laying period (r~ = 0.69, n = 20, P < 0•003). (The sex ratio was calcu- lated for days on which at least 10% of the marked population (n = 102) attended)
and increasing to over 85% by the end of May. The sex ratio at any given m o m e n t was even higher because on average females arr ived later in the m o r n i n g in rela- t ion to males in the laying per iod (Fig. 3); in April , males arrived a m e a n of 10 mi n earlier than females, bu t in May the difference increased to 40 min (Table 1). Thus, there were usual ly several males in the arenas for every female at a given m o m e n t dur ing the laying period•

66
Pre-laying period Laying period
6 0 -
40 .
zo- I I
I
O.
-20. 1 5 10 15 20 25 1 5 10 15
April May Date
Fig. 3. Difference in mean arrival times (min) to the mating arenas between males and females on each day of attendance dur- ing the prelaying and laying periods. Posi- tive values indicate that males arrive first on average
Table 1. Arrival times of males and females to the mating arenas during the prelaying and laying periods
Period Sex No. of Mean SD Difference in arrivals arrival time a arrival times
Prelaying F 571 433.9 44.9 (April) M 683 424.0 44.6 9.9
Laying F 246 406.2 54.6 (May) M 736 366.5 49.8 39.7
a Expressed in numbers of minutes since 0000 hours
Seasonal changes in EPC frequency
The percentage of females receiving EPC attempts upon arrival at the arenas began to increase markedly at the end of April, several days before the commencement of egglaying (Fig. 4a), when female birds are most likely to be fertile (Birkhead et al. 1987). During the first half of April fewer than 20% of females were mounted upon arrival, but by the middle of May over 80% received EPC attempts. The EPC attempt frequency also in- creased with the sex ratio (Fig. 4b), while the sex ratio was also correlated with the date (Fig. 2). A partial ef- fects analysis showed that the frequency of EPC at- tempts increased significantly with the date when the sex ratio was held constant (t = 4.79, d f = 32, P < 0.0001) but did not increase when the date was held constant (t = 1.17, d f = 32, P = 0.25) suggesting that the EPC fre- quency is dependent upon the date.
Seasonal changes in aggression rates
The arrival of females at the arenas during the laying period usually resulted in fights between males who were attempting EPCs or guarding their mates against other males. Although females were sometimes involved in fights with males or other females, a large majority of
aggressive exchanges were intermale (89% of 1235 fights). The aggression rate (fights/individual h) re- mained low throughout most of the prelaying period but sharply increased around the time the first egg was laid (Fig. 5a). The rate peaked 1 day after the median laying date and decreased sharply thereafter, as a de- creasing number of females were fertilizable. The aggres- sion rate also increased with the sex ratio (Fig. 5b). However, the aggression rate increased significantly with the date when the sex ratio was held constant ( t= 2.39, d f = 38, P = 0.022), but not when the date was held con- stant ( t= 1.25, d f=38 , P=0.17) , suggesting that the ag- gression rate, like the EPC frequency, is determined by the date, which could serve as a general indicator of female fertility.
Variation in male EPC success
The number of EPCs obtained by males ranged from 0 to 7 in 1988 and 0 to 5 in 1989. The distribution of male EPC success was highly skewed, with a minority of males obtaining a majority of matings. On the lower mating arena in 1988, 16males obtained a total of 16 EPCs, with the 2 (13%) most successful males obtain- ing 12 (75%), while 10 (63%) of the males did not obtain any (Fig. 6a). The pattern was similar in 1989 when 29 males obtained 25 EPCs, but with the top 2 males (7%) obtaining 10 (40%) of the matings. To investigate this variation, I correlatd EPC success with behavior, body size, age and mating status.
Behavior. I correlated male EPC success with six behav- ioral variables (Table 2). The frequencies of three of these behaviors were strongly correlated with the number of EPCs males obtained. EPC success increased with (1) the number of times males interfered with the copulation attempts of other males, (2) the number of EPC attempts made and (3) the number of fights males won ("wins") . It was apparent that most fights were

"E
=
~4o
10o
80
60
( a )
40
2 0
0 •
1
• •
• • • •
O • O O
5 10 15 20 25 30 5 10 15 20
April Date May
.'!
es 0
=
.¢
..~
?
100
80
60
40
20
(b)
S
• • • •
IO
o •
0 • • • • • , . , . , . , . , . , . ,
45 50 55 60 65 70 75 80
Sex r a t i o (% male)
Fig. 4 a, b. Relat ionship between the percentages o f females receiv- ing extra-pair copula t ion (EPC) a t tempts upon arrival to the arenas and (a) the date (rs=0.93, n = 3 4 , P<0 .0001) and (b) the sex ratio (rs = 0.84, n = 33, P < 0.0001). The percentages of females receiving EPC at tempts were calculated for days with a min imum of 10 fe- male arrivals
Table 2. Correlat ions o f male EPC success with six behavioural variables
Behavioural variable r P d.f.
Number o f copulat ions interfered with 0.758 < 0.0001 41 Number o f fights won 0.606 < 0.0001 41 Number o f EPC at tempts 0.681 < 0.0001 41 Percentage of fights won a 0.280 = 0.088 38 Number o f days o f arena a t tendance 0.196 =0.21 41 Mean arrival t ime at arena 0.144 =0.38 39
Arcsine t ransformed
67
8] .~ .6
~ . 4
.2
O- 9
(a)
" "o o • O o O o 0
• o •
• M e d i a n
l a y i n g d a t e
¢ 30 5 10 15
May 5 10 15 20 25
April Date
O •
20
1.2 (b)
1
.8 E
"~ .6
,< .4
.2 • •
o O d b o • • • o o
0 - ' " • " • "
45 50 55 60
O O
• 0 •
, . , . , . , . , . , . , . ,
65 70 75 80 85 90 95 100
Sex r a t i o
Fig. 5a, b. Change in the aggression rate (fights/individual h) in the mat ing arenas with (a) the date (rs=0.76, n = 3 9 , P<0 .0001) and (b) the sex ratio (r~=0.73, n = 3 9 , P<0 .0001)
in competition over EPCs rather than mate-guarding (i.e. driving other males away from their mates), because 1490 (74%) of 2014 fights occurred in the absence of both males' mates. Males interfered with copulations as a way to obtain EPCs, in that they followed interfer- ence with an EPC attempt after 33% of 60 disruptions.
If males fight and interfere with copulations in order to obtain EPCs, then the frequency of these two behav- iors might correlate with the number of EPC attempts and with each other. Wins correlated significantly with interferences (r=0.61, df=41, P<0.0001), as did wins with attempts (r = 0.66, df= 41, P < 0.0001) and interfer- ences with attempts (r=0.58, df=41, P<0.0001). The three variables together explained 62% of the variance in EPC success in a multiple linear regression (Table 3). However, interference alone explained 57% of the varia- tion in EPC success and remained significant when wins and attempts were held constant, while wins and at- tempts were not significant when the other two variables were held constant, indicating that interference is the

68
7 '
_~ 6-
I 4
o
3
t_ 2- 2~ ,,.t
( a ) 1 9 8 8
0 ~ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Rank order of males
1,89
!
2
i
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29
Rank order of males
Fig. 6a, b. Distributions of EPCs obtained by males in the lower mating arena in 1988 and 1989
Table 3. Number of successful EPCs achieved by males in relation to three behavioural variables in a multiple linear regression analy- sis
Independent variable t P =
Number of copulations interfered with 3.40 0.0016 Number of EPC attempts 1.60 0.12 Number of fights won 0.81 0.42
Multiple r z = 0.623, F = 20.9, df= 3.38, P < 0.0001
recorded behavior most closely associated with EPC suc- cess. Although interference accounted for most of the variance in EPC success, it was not a direct cause of success because only 2 (6%) out of 34 EPCs were ob- tained when males replaced the disrupted male.
In 1988, the number of wins was also correlated with EPC success (r=0.77, d f=33 , P<0.0001), as was the number of EPC attempts (r=0.76, d f=35 , P<0.0001). Interference was not comprehensively recorded in 1988.
The means of these three variables are compared in Table 4 for successful males (those which obtained at
Table 4. Differences in behaviour of 15 males who obtained at least one EPC and 27 males who did not obtain EPCs
Behaviour Obtained Mean SD z a P= EPC(s)
No. of copulations No 1.1 1.5 2.1 0.055 interfered with Yes 3.3 3.6
No. of fights won No 8.5 9.9 2.2 0.022 Yes 20.3 24.9
No. of EPC attempts No 5.9 6.9 3.0 0.026 Yes 14.0 10.1
a Mann-Whitney U-tests
least one EPC) and unsuccessful males. Successful males interfered with a mean of three copulation attempts compared with one for unsuccessful males, averaged 20 wins compared with 9 and made more attempts, 14 compared with 6.
Thirty-eight males engaged in a mean of 26 fights (range 6 122) in which the outcome was a win or a loss. The correlation of EPC success and the percentage of fights won was not significant (r = 0.28, P < 0.09).
I found no relationship between EPC success and either of two measures of mating arena attendance (Ta- ble 2). Males did not achieve more EPCs by attending on more days (r=0.20, d f=41 , P=0 .21) or by arriving earlier (r=0.14, d f=39 , P=0.38) .
Body size. I found no relationships between body size and EPC success. There were no significant (Pearson's) correlations between EPC success and any of the 11 morphological measurements defined in the Methods section (sample sizes ranged from 31-39). The relation- ship was not significant when all 11 variables were placed in a linear multiple regression (multiple r 2= 0.24, F=0.51 , d f = 11, 29, P=0.87) .
Age. There were no significant correlations between age and any of the following four variables: EPC success (r=0.33, d f = 14, P=0.23) , EPC attempts (r=0.38, d f = 14, P=0.17) , wins (r=0.39, d f = 17, P=0 .11) and inter- ferences (r=0.30, d f=17 , P=0.22) . The mean age of nine unsuccessful males was 14.2 years (SD = 5.9), which did not differ significantly from 15.8 (SD=3.6) years for four successful males (two-tailed t-test, t = 0.47, P = 0.64).
The most successful male in both years was C88, who at the minimum age of 20 years in 1988 obtained seven EPCs, and five in 1989. C88 also made the largest number of EPC attempts in both years, 41 and 39, com- pared with means of 6 and 8. Old age did not ensure EPC success, however. The oldest male was at least 24 years old in 1989 but made only 11 EPC attempts, all unsuccessful. The two other males with minimum ages exceeding 20 years also performed unexceptionally, making only 3 and 2 attempts, respectively, and obtain- ing no EPCs. On the other hand, the second most suc- cessful male obtained 5 EPCs at the intermediate known

Table 5. Performance of paired (P) and unpaired (U) males
Behaviour Paired n M e a n SD z a P= status
Number of EPCs obtained
Number of copulations interfered with
Number of fights won
Number of EPC attempts
Percentage of fights won b
No. of days attendance in mating arenas
Mean arrival time (min)
P 34 0.6 1.1 0.1 0.91 U 8 0.9 1.7
P 43 1.7 2.1 0.5 0.63 U I0 1.8 3.7
P 43 11.3 12.1 1.5 0.12 U 10 12.7 28.4
P 34 9.3 7.8 1.8 0.07 U 8 6.8 13.3
P 32 57.0 29.6 0.8 0.40 U 6 44.7 32.7
P 43 24.7 7.5 1.2 0.25 U 10 27.7 7.2
P 40 396 16 1.2 0.21 U 10 388 16
a Mann-Whitney U-tests b Equals the number of wins/wins+losses; the mean number of fights per male was 21, ranging from 6 to 120. Percentages of fights were arcsine transformed
age of 13 years, implying that old age is not a prerequi- site for high EPC success.
Paired status. There was no evidence that paired males performed differently in competition over EPCs than unpaired males. The frequencies of the three behaviors associated with EPC success (wins, interferences and at- tempts) were similar for paired and unpaired males, and there was no significant difference in EPC success be- tween the two classes (Table 5). Paired and unpaired males also visited the arenas on a similar number of days, 25 and 28, respectively.
Relative E P C success
If active female choice influences male EPC success, then males with certain traits should achieve a greater relative
Table 6. Relative EPC s u c c e s s a ' b of males correlated with behav- ioural variables
Behaviour r P = Number of males
Number of copulation interferences 0.226 0.25 28 Number of fights won 0.147 0.45 28 Number of EPC attempts 0.046 0.82 28 Percentage u of fights won 0.083 0.67 28 Number of days attended -0.018 0.93 28 Mean arrival time c (rain) --0.129 0.52 27
a Relative EPC success is number of EPCs obtained/number of attempts. Males with fewer than 4 attempts were excluded. The number of attempts ranged from 4 to 39 with a mean of 13 (SD = 8.5) b Arcsine transformed c Mean arrival times calculated for individuals with a minimum of 10 days of attendance
69
EPC success. Relative success was defined as the number of successful EPCs males achieved per attempt. None of the three behavioral variables which correlated with absolute EPC success correlated with relative success. Males did not increase their relative success by interfer- ing, winning fights or making attempts (Table 6). The three behaviors together were also not correlated with relative success in a multiple linear regression (multiple r 2 =0.08, F=0 .66 , d f = 3 , 24, P=0.58) . There were also no relationships between relative EPC success and arena attendance, as measured by number of days and arrival time (Table 6), or with any of the 11 measurements of body size (sample sizes ranged from 21 to 24).
Discussion
The mating arena as a lek
Typical lek features include aggressive male-male compe- tition for matings (Beehler and Foster 1988), a highly male-biased operational sex ratio (Emlen and Oring 1977) and a skewed distribution of male mating success (Bradbury and Gibson 1983; Appolonio etal. 1989; Pruett-Jones and Pruett-Jones 1990; Gratson etal. 1991). For EPCs during the laying period, the razorbill mating arena exhibited all of these features. Females sought EPCs in the arenas where there were no resources other than males. Males who obtained EPCs were never observed contributing parental care to the offspring of males they had potentially cuckolded [in over 600 male feedings in 2 years (71 nest-years)]; males never fed at another nest (R.H. Wagner, unpublished data) and therefore did not provide any reproductive contribution other than gametes. Males fought aggressively and fre- quently over EPCs, timing their attempts to the probable fertilizable periods of females. At that time, the mating arena sex ratio became highly skewed, and, as in most leks, a few males obtained the majority of matings.
Colonies as leks
Similar conditions experienced by razorbills in their mat- ing arenas may exist for some other monogamous birds within their colonies, suggesting that lekking for EPCs may occur in other species but is difficult to detect be- cause males compete for territories as well as EPCs. In the common murre, the sex ratio also becomes male- biased when females depart to sea on a prelaying exodus, and this change has also been associated with both the date and a marked increase in EPC frequency (Birkhead et al. 1985; Hatchwell 1988). Murres in both studies also showed a skew in the number of EPCs attempted by males, and in Hatchwell's study, interference was asso- ciated with EPC success (although unlike in razorbills, male murres obtained EPCs as a direct result of disrupt- ing and then replacing the mounting male). Female northern fulmars, some of whom seek EPCs, also depart to sea prior to egg-laying, resulting in a skewed sex ratio on the colony (Hatch 1987). Even in species in which

70
there is no change in the actual sex ratio during female fertility, the operational sex ratio may become male- biased as females complete egg-laying, but males may continue to produce sperm.
Male-male competition or female choice ?
Even if females preferred certain males, it is possible that they would be unable to accept EPCs from them because of two limitations: (a) females did not accept EPCs if their mates were present (Wagner 1991 a), and (b) females may have been unable to accept insemina- tions during mountings which were disrupted. However, males performed only 12% (n= 129) of EPC attempts in the presence of the female's mate. Excluding those attempts, as well as attempts which were disrupted, pre- laying females were capable of accepting a large majority (approximately 80%) of potentially fertilizing EPC op- portunities.
The frequencies of three behaviors (interference, at- tempts and wins) were strongly correlated with EPC suc- cess and with each other. All three were closely associat- ed with the male effort to obtain EPCs. It is unclear why interference explained most of the variance in EPC success because males rarely obtained EPCs as a direct result of interfering. Possibly interference might best be viewed as part of the same behavior as wins because these behaviors could be two aspects of the same strate- gy: to deprive other males of access to extra-pair fe- males. Since males won fights by displacing other males, they often succeeded in preventing their competitors from mounting and inseminating females. Such a strate- gy might also be regarded as a behavioral aspect of sperm competition because by preventing other males from inseminating a given female, a male may increase his own chances of fertilizing her eggs (Birkhead et al. 1987). Even if the interfering male had not himself inse- minated the female, the potential may exist for him to gain an insemination in the future, at which time his sperm would not be competing against the sperm of the disrupted male.
The close relationship between wins, interference and attempts suggests that successful males were those who were both persistent and aggressive, because males who attempted many EPCs also fought and interfered often. Persistence in attempting EPCs may be regarded as an aspect of male-male competition, because males often had to win fights to gain access to females. The lack of correlation between the percentage of fights males won and EPC success is consistent with the interpreta- tion that persistence is important. It is logical that the absolute rather than the relative number of fights males won is a good indicator of access to females. A male who won 7 out of 10 fights for example, would have the same percentage as a male who won 70 out of 100, but the latter male would encounter females many more times. If females do not select among males for EPCs then the raw number of opportunities that males create should be more important than the percentage of times they win.
I found no statistical evidence that females had exer- cised active choice in accepting EPCs from males. The frequency with which males interfered, won fights and attempted EPCs apparently did not attract females, be- cause they were not correlated with relative EPC success. If females were attracted to certain males because of their demonstrated competitive ability, then they should have favored those males with greater relative mating success. The lack of correlations between relative EPC success and any of these three behaviors, as well as with numerous male behavioral and morphological variables, argues against the occurrence of female choice. The ab- sence of such relationships, in contrast to the three vari- ables which significantly correlated with absolute suc- cess, implies that EPC success may have been determined primarily or entirely through male-male competition.
Identifying the benefit(s) of EPCs to females could help predict whether it is advantageous for females to exercise choice. Analyses of potential benefits which fe- male razorbills might accrue from EPCs raised the possi- bility that females may pursue EPCs to obtain insurance against their mates' infertility (Wagner 1992). This expla- nation for female promiscuity is compatible with the apparent lack of female choice because females are not required to discriminate among male qualities to obtain insurance, but only to distinguish other males from their mates. This parsimonious explanation is consonant with the view of Beehler and Foster (1988) that female de- cision-making rules are simple and largely determined by male-male competition. If females seek EPCs to ob- tain °' good genes" (Wittenberger 1979; Hamilton 1990), then we might expect them to favor successful males with higher relative success. However, by allowing males to compete for matings, females may be inseminated by the best male without having to discriminate (Cox and Le Boeuf 1977; Montgomerie and Thornhill 1989). Evidence in razorbills that females may incite male com- petition was suggested from observations that female visitations to the arenas often resulted in intermale ag- gression (Wagner 1991c).
In conclusion, the striking resemblance of razorbill mating arenas to leks, combined with the similarity of behaviors performed in the arenas by razorbills with those performed by other colonial species in their colo- nies, suggests that colonies of other monogamous birds may also function as leks in which males compete for EPCs. In examining whether leks are °'hidden'' within colonies, it may pay to scrutinize male aggression to determine which male behaviors correlate with EPC suc- cess, although it will be difficult to separate the effects of male behaviors on EPC, mate-guarding and territorial defence when they all occur in the same place. For male razorbills, the mode of sexual selection responsible for most of the variation in EPC success is male-male com- petition. The degree to which this variation translated into variance in male fitness will require the use of genet- ic techniques to determine the paternity of offspring of females who engaged in EPC.
Acknowledgements. I thank Dr. Christopher M. Perrins for super- vising my doctoral studies of which this paper is a part. Nigel

71
Dougherty, Tom Helowell and Daniel Owen assisted in trapping and banding razorbills; Nigel has entered razorbill folklore for his crucial role in the capture of C88. I am grateful to the Dyfed Wildlife Trust for permitting me to work on Skomer Island. Finan- cial support was received from a British government overseas tea- search studentship, and Sigma Xi, and from the Smithsonian Insti- tution during revisions of the manuscript. This paper has benefitted from constructive comments on various versions from T.R. Birk- head, P.A. Gowaty, W.D. Hamilton, H.W. Power, I. Rowley, R.R. Warner and D.F. Westneat.
References
Appolonio M, Festa-Bianchet M, Mari F (1989) Correlates of co- pulatory success in a fallow deer lek. Behav Ecol Sociobiol 25:89 97
Beecher MD, Beecher IM (1979) Sociobiology of bank swallows: reproductive strategy of the male. Science 205:1282-1285
Beehler BM, Foster MS (1988) Hotshots, hotspots, and female preference in the organization of lek mating systems. Am Nat 131:203 219
Birkhead TR (1978) Behavioural adaptations to high density nest- ing in the common guillemot Uria aalge. Anita Behav 26 : 321- 331
Birkhead TR, Johnson SD, Nettleship DN (1985) Extra-pair mat- ings and mate guarding in the common mute Uria aalge. Anita Behav 33:608-619
Birkhead TR, Atkin L, Moller AP (1987) Copulation behaviour of birds. Behaviour 101:101 138
Blakey JK (1989) Biochemical and population genetics in the great tit (Parus major). D. Phil. thesis, University of Oxford
Bradbury JW, Gibson R (1983) Leks and mate choice. In: Bateson P (ed) Mate choice. Cambridge University Press, London, pp 109-138
Brooker MG, Rowley I, Adams M, Baverstock PR (1990) Promis- cuity : an inbreeding avoidance mechanism in a socially monog- amous species? Behav Ecol Sociobiol 26:191 199
Burger J, Gochfeld M (1990) The black skimmer, social dynamics of a colonial species. Columbia University Press, New York
Cox CR, Boeuf BJ Le (1977) Female incitation of male competi- tion: a mechanism of sexual selection. Am Nat 111:317-335
Darwin C (1871) The descent of man and suxal selection in relation to sex. Murray, London
Draulans D (1988) The importance of heronries for mate attrac- tion. Ardea 76:187-192
Emlen ST, Oring LW (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197:215-223
Frederick P (1987) Extrapair copulations in the mating system of white ibis (Eudocimus albus). Behaviour 100:170 201
Gibbs HL, Weatherhead P J, Boag PT, White BN, Tabak LM, Hoysak DJ (1990) Realized reproductive success of polygynous red-winged blackbirds revealed by DNA markers. Science 250:1394-1397
Gratson MW, Gratson GK, Bergerud AT (1991) Male dominance and copulation disruption do not explain variance in male mat- ing success on sharp-tailed grouse (Tympanuchus phasianellus) leks. Behaviour 118 : 187 213
Hamilton WD (1990) Mate choice near or far. Am Zool 30: 341- 352
Hatch SA (1987) Copulation and mate guarding in the northern fulmar. Auk 104:450-461
Hatchwell BJ (1988) Intraspecific variation in extra-pair copulation and mate defence in common guillemots Uria aalge. Behaviour 107:157 185
H6glund J, Lundberg A (1987) Sexual selection in a monomorphic lek-breeding bird: correlates of male mating success in the great snipa Gallinago media. Behav Ecol Sociobiol 21:211-216
Hudson PJ (1985) Population parameters for the Atlantic Alcidae. In: Nettleship DN, Birkhead TR (eds) The Atlantic Alcidae. Academic Press, London, pp 233-263
Lloyd CS, Perrins CM (1977) Survival and age of first breeding in the razorbill Alca torda. Bird Banding 48 : 239-252
MacRoberts MH (1973) Extra-marital courting in lesser black- backed and herring gulls. Z Tierpsychol 32:62-74
McDonald DB (1989) Correlates of male mating success in a lekk- ing bird with male-male cooperation. Anita Behav 37:1007- 1022
Mead CJ, Clark JA (1988) Report on bird ringing in Britain and Ireland for 1987. Ringing and Migration 9:169-186
Moller AP (1985) Mixed reproductive strategy and mate guarding in a semi-colonial passerine, the swallow Hirundo rustica. Behav Ecol Sociobiol 17:401-408
Moiler AP (1988) Female choice selects for male sexual tail orna- ments in the monogamous swallow. Nature 332: 640-642
Montgomerie R, Thornhill R (1989) Fertility advertisement in birds: a means of inciting male-male competition? Ethology 81 : 209 220
Morton ES, Forman L, Braun M (1990) Extrapair fertilizations and the evolution of colonial breeding in purple martins. Auk 107:275-283
Pierotti R (1981) Male and female parental roles in the western gull under different environmental conditions. Auk 98 : 532-549
Pruett-Jones SG, Pruett-Jones MA (1990) Sexual selection through female choice in Lawe's parotia, a lek-mating bird of paradise. Evolution 44:486-501
Sherman PW, Morton ML (1988) Extra-pair fertilizations in moun- tain white-crowned sparrows. Behav Ecol Sociobio122 : 413-420
Smith SM (1988) Extra-pair copulations in black-capped chicka- dees: the role of the female. Behaviour 107:15 23
Trivers RL (1972) Parental investment and sexual selection. In: Campbell BG (ed) Sexual selection and the descent of man 1871-1971. Aldine Pres, Chicago, pp 136-179
Wagner RH (1991 a) Evidence that female razorbills control extra- pair copulations. Behaviour 118 : 157 169
Wagner RH (1991b) The role of extra-pair copulation in razorbill mating strategies. D Phil thesis, University of Oxford
Wagner RH (1991c) The use of extra-pair copulations for mate appraisal by razorbills. Behav Ecol 2:198-203
Wagner RH (1992) The pursuit of extra-pair copulations by mo- nogamous female razorbills: how do females benefit? Behav Ecol Sociobiol 29:455-464
Westneat DF (1990) Genetic parentage in the indigo bunting: a study using DNA fingerprinting. Behav Ecol Sociobiol 27:67- 76
Westneat DF, Sherman PW, Morton ML (1990) The ecology and evolution of extrapair copulations in birds. In: Power DM (ed) Current ornithology, vol 7. Plenum Press, New York, pp 331 369
Wittenberger JF (1979) The evolution of mating systems in birds and mammals. In: Marler P, Vandenbergh P (eds) Handbook of behavioural neurobiology, vol 3. Social behaviour and com- munication. Plenum Press, New York, pp 271-349
Copyright © 2022 FDOKUMEN




















