
Molecular Dynamics Simulations of the TEM-1 β-Lactamase Complexed with Cephalothin
Upload
independentCategory
view
1download
0
834
Clinical and Bacteriologic Epidemiology of Extended-Spectrum b-Lactamase–Producing Strains of Klebsiella pneumoniae in a Medical Intensive Care Unit
Dominique Decre, Bertrand Gachot, From Service de Microbiologie, Service de Reanimation des MaladiesInfectieuses, and Unite d’Hygiene et de Lutte contre l’InfectionJean Christophe Lucet, Guillaume Arlet,
Nosocomiale, Hopital Bichat-Claude Bernard, and Service deEugenie Bergogne-Berezin, and Bernard RegnierMicrobiologie, Hopital Saint-Louis, Paris, France
The epidemiology of extended-spectrum b-lactamase (ESBL)–producing strains of Klebsiellapneumoniae was studied over a 16-month period in a medical intensive care unit (ICU). A controlprogram involving enhanced isolation procedures, surveillance cultures at admission and then at 1-week intervals, and selective digestive decontamination (SDD) was instituted. Phenotypic and geno-typic markers (plasmid content and DNA macrorestriction polymorphism determined by pulsed-field gel electrophoresis) were used to compare 138 strains of ESBL-producing K. pneumoniae. Theincidence of colonization and/or infection with ESBL producers was 11.9%. ESBL-producingK. pneumoniae strains were isolated from 64 of 65 patients. Fifty-five cases were considered acquiredin the ICU, while nine cases were imported. Forty-five infections occurred in 32 patients; 20 infectionsinvolved the urinary tract. SDD failed to reduce the incidence of acquisition of ESBL-producingK. pneumoniae. Combined use of markers was necessary to achieve accurate differentiation ofstrains. A single epidemic clone (SHV-4 b-lactamase–producing K. pneumoniae) was the cause of85% of the ICU-acquired cases. Sporadic occurrence of SHV-5, TEM-3, SHV-2, and SHV-3 produc-ers accounted only for a few cases.
Transferable resistance to extended-spectrum cephalosporins epidemiological studies of ESBL-producing strains indicated acombination of epidemic modes of dissemination includingand monobactams in members of the Enterobacteriaceae family
has emerged and spread in many French hospitals since 1984 strains, self-transferable plasmids, and gene transposition [1,2, 4, 5, 9, 15, 16, 21, 27, 28].[1–11]. A large number of extended-spectrum b-lactamases
(ESBLs) derived from TEM-1, TEM-2, and SHV enzymes by In our hospital, ESBL-producing strains have been spreadingsince 1989, particularly in the ICUs. In February 1992, a controlone or more amino acid substitutions have been reported [11–
14]. The genes encoding the production of ESBLs are generally program for ESBLs was instituted that utilized increased barrierprecautions for patients colonized and/or infected with ESBLlocated on large transferable plasmids with molecular sizes
ranging from 85 to 275 kb [12, 15]. Many ESBL-producing producers as well as clinical and bacteriologic surveillance ofall patients admitted to the three medical or surgical ICUs.strains are also resistant to aminoglycosides, sulfonamides, tri-
methoprim, chloramphenicol, and tetracyclines as well as to To elucidate the mechanisms involved in patient-to-patienttransmission and to identify epidemic strains, we conducted anfluoroquinolones [5, 8, 15, 16]. ESBL production has been
found mainly in Klebsiella pneumoniae strains but has also epidemiologic study of strains of ESBL-producing K. pneu-moniae that were isolated in one ICU during a 16-month period.been described in other Enterobacteriaceae, including Esche-
richia coli, Proteus mirabilis, Salmonella, Enterobacter, Citro- Biotypes, antimicrobial susceptibility patterns, b-lactamasecharacteristics (isoelectric point [pI]), plasmid profiles, andbacter, and Serratia [5, 6, 10, 13, 16–19].
Outbreaks of nosocomial infections due to ESBL-producing DNA macrorestriction polymorphism determined by pulsed-field gel electrophoresis (PFGE) were used as epidemiologicalstrains are being increasingly reported in intensive care units
(ICUs) and chronic care facilities [1–3, 6, 7, 20–24]. The markers. In addition, we compared a new faster method forgenotype determination that involved PCR-mediated finger-reservoir for these strains appears to be the gastrointestinal
tract of patients [25] with patient-to-patient contamination oc- printing with primers based on repetitive extragenic palin-dromic (REP) DNA sequences with DNA macrorestrictioncurring via the hands of health care providers [26]. Previousanalysis.
Received 25 November 1997; revised 8 June 1998. MethodsThis work was presented in part at the 94th annual meeting of the American
Society for Microbiology held in Las Vegas on 23–27 May 1994. Clinical and Bacteriologic Surveillance ProgramReprints or correspondence: Dr. Dominique Decre, Service de Microbiolo-
gie, Hopital Saint-Antoine, 184 rue du Faubourg Saint-Antoine, 75571 Paris The Bichat-Claude Bernard University Hospital (Paris) is aCedex 12, France. 1,200-bed institution that admits ú32,000 patients each year.Clinical Infectious Diseases 1998;27:834–44 The Infectious Diseases ICU has 18 beds and admits 400–450q 1998 by the Infectious Diseases Society of America. All rights reserved.1058–4838/98/2704–0030$03.00 patients per year, of whom about one-half are referred from
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

835CID 1998;27 (October) Epidemiology of ESBL-Producing K. pneumoniae
other hospitals, 10%–15% are from the emergency department, method with use of Mueller-Hinton medium (Sanofi Diagnos-tics Pasteur, Marnes-la-Coquette, France). All ESBL producersand 30%–35% are from other departments in the hospital.
Bacteriologic surveillance included collection of rectal swabs detected by means of a positive disk synergy test were consid-ered as resistant to all extended-spectrum cephalosporins [32].and urine specimens at admission and then at weekly intervals.
Specimens were inoculated on Drigalski agar containing These strains were first subdivided according to their patternof resistance to b-lactam agents (CAZ or CTX phenotype),0.5 mg of cefotaxime/L [29]. Other specimens were obtained
for clinical reasons when it was considered necessary by the which was based on inhibition zone diameters for cefotaximeand ceftazidime as previously described [14]; the strains weremedical staff.
ESBL-producing strains were detected based on the results then further classified based on results for the other antibiotics.The CAZ phenotype was defined as a higher level of resistanceof double-disk diffusion testing of the synergistic effect be-
tween clavulanic acid/amoxicillin and cefotaxime, ceftazidime, to ceftazidime than to cefotaxime (inhibition zone diameter:õ12 mm, ceftazidime and aztreonam; ú18 mm, cefotaxime),or aztreonam [6, 30, 31].
On the basis of the results of surveillance cultures and clini- and the CTX phenotype was defined as similar inhibition zonediameters (15–25 mm) for cefotaxime and ceftazidime. Strainscal surveillance data (e.g., geographic origin of the patient,
date of admission, location and transfer within the unit, and were classified as susceptible, intermediately resistant, or resis-tant to the other antibiotics tested according to the recommen-previous admissions), each new ESBL-producing strain–posi-
tive case was classified as referred to or acquired in the unit. dations of the Antibiotic Susceptibility Testing Committee ofthe French Society for Microbiology [33]. The following antibi-Referred cases were those that met any of the following criteria:
ESBL-producing strain isolated at admission; ESBL-producing otics were tested: the aminoglycosides neomycin, kanamycin,gentamicin, tobramycin, and amikacin; the fluoroquinolonesstrain isolated from an infected site within 72 hours after ICU
admission; and/or history of colonization and/or infection with nalidixic acid, pefloxacin, and ciprofloxacin; chloramphenicol;tetracycline; sulfonamides; and trimethoprim.an ESBL-producing strain.
Measures for preventing patient-to-patient contamination b-Lactamase analysis. Analytical isoelectric focusing wasperformed on a polyacrylamide gel (Pharmacia-Biotech, Orsay,that were instituted as part of the control program for ESBLs
consisted of isolation precautions and enhanced hygiene proce- France) in the 3.5 to 9.5 range with crude cell-free sonic ex-tracts. b-Lactamase activities were detected by using the chro-dures, including labeling of all cases of colonization and/or
infection with ESBL-producing strains and use of aseptic hand mogenic nitrocefin test [34]. The following enzymes were usedas standards: TEM-1 (pIP1100; pI, 5.4), TEM-2 (RP4; pI, 5.6),washing procedures and gloves by health care providers. Health
care personnel also wore gowns when caring for carriers. Selec- TEM-3 (pCFF04; pI, 6.3), SHV-1 (pIP173; pI, 7.7), SHV-3(pUD18; pI, 7.0), and SHV-4 (pUD21; pI, 7.8).tive digestive decontamination (SDD) with erythromycin (1 g
b.i.d.) and polymyxin E (6 million units b.i.d.) was also used; Plasmid analysis. Plasmid DNA from isolates of ESBL-producing K. pneumoniae was extracted after alkaline lysisall patients admitted to the unit during the first 7 months of
the study (period 1) received this treatment (prophylactic SDD), [35]. Plasmids were visualized under ultraviolet light afterstaining of the gel with ethidium bromide and analyzed compar-and then during the next 5 months (period 2), only colonized
and/or infected patients received this treatment (curative SDD). atively with the following reference plasmids: RP4 (54 kb),pCFF04 (85 kb), and pIP173 (126.8 kb). A supercoiled DNASDD was not used during the last 5 months of the study period.
Urinary tract colonization or infection was treated with cefo- ladder (2–16 kb) (GIBCO–BRL Life Technologies, Cergy-Pontoise, France) was used as the reference for small plasmids.tetan or with clavulanic acid/amoxicillin. Infections were de-
fined as previously reported elsewhere [29]. Genomic fingerprinting by PFGE. PFGE was performedas described previously [36]. The following restriction enzymeswere tested: XbaI, SpeI, and NotI (Boerhinger Mannheim, Mey-
Microbiologylan, France). Restriction fragments were separated by PFGE in1% agarose gel (seakem Gold; FMC Bioproducts, Rockland,Bacterial strains. Strains of ESBL-producing K. pneumo-
niae that were isolated both from clinical and surveillance ME) in 0.51 TBE (Tris-borate-EDTA) buffer (pH 8.0) at 147Cwith use of the Pulsaphor System (Pharmacia-Biotech, Orsay,specimens were studied. All strains isolated from November
1992 to June 1993 were studied prospectively. All available France). Running conditions were 200 V for 18 hours withpulse times of 10 to 30 seconds for XbaI and 300 V for 15isolates (several isolates per patient that were previously frozen
on the basis of their phenotypic characteristics) from patients hours with pulse times of 9 seconds for SpeI or NotI. Gelswere stained with ethidium bromide and photographed underwho were colonized and/or infected before November 1992
were analyzed retrospectively. ultraviolet light. Polymerized l DNA (Bio-Rad S.A., Ivry surSeine, France) was used as the molecular size marker.Biotypes. Biochemical tests were performed by using the
API 20E System (bioMerieux, Marcy l’Etoile, France). Band patterns were visually compared and classified as indis-tinguishable (clonal), closely related (clonal variants, three orAntibiotic susceptibility patterns. Susceptibility to antimi-
crobial agents was tested by using the standard agar diffusion less band differences), possibly related (four to six band differ-
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from
836 Decre et al. CID 1998;27 (October)
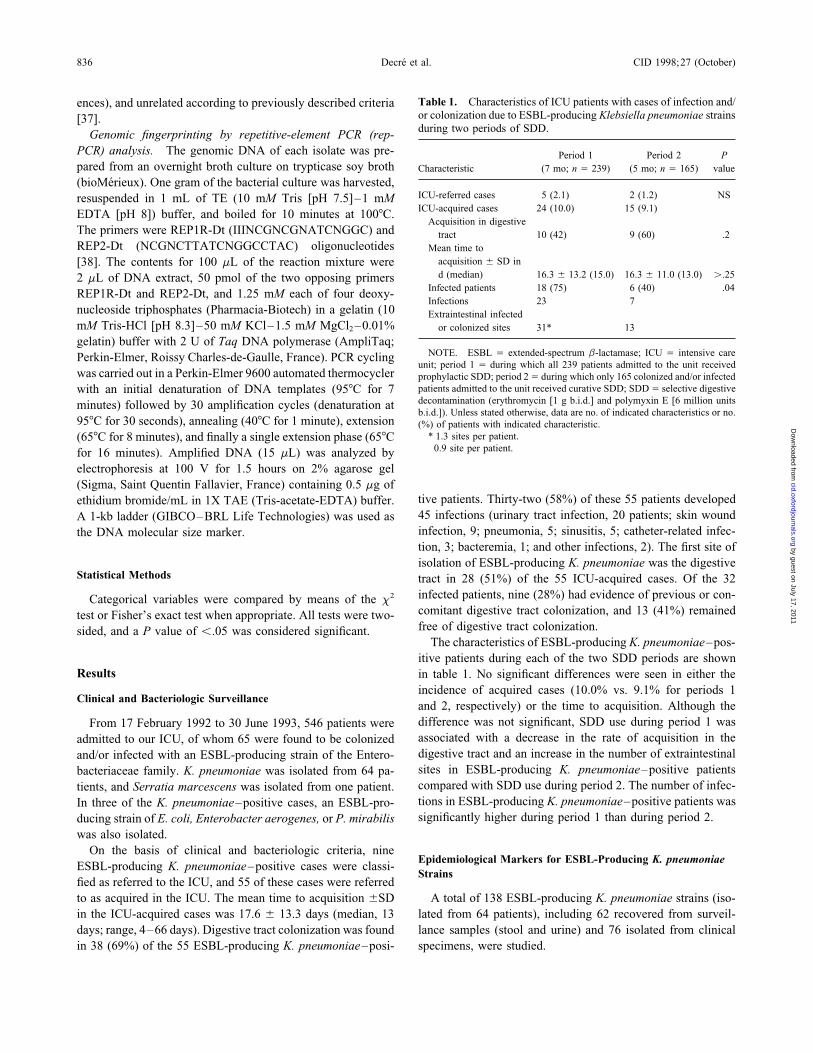
Table 1. Characteristics of ICU patients with cases of infection and/ences), and unrelated according to previously described criteriaor colonization due to ESBL-producing Klebsiella pneumoniae strains[37].during two periods of SDD.Genomic fingerprinting by repetitive-element PCR (rep-
PCR) analysis. The genomic DNA of each isolate was pre- Period 1 Period 2 Ppared from an overnight broth culture on trypticase soy broth Characteristic (7 mo; n Å 239) (5 mo; n Å 165) value
(bioMerieux). One gram of the bacterial culture was harvested,ICU-referred cases 5 (2.1) 2 (1.2) NSresuspended in 1 mL of TE (10 mM Tris [pH 7.5]–1 mMICU-acquired cases 24 (10.0) 15 (9.1)EDTA [pH 8]) buffer, and boiled for 10 minutes at 1007C.
Acquisition in digestiveThe primers were REP1R-Dt (IIINCGNCGNATCNGGC) and tract 10 (42) 9 (60) .2REP2-Dt (NCGNCTTATCNGGCCTAC) oligonucleotides Mean time to
acquisition { SD in[38]. The contents for 100 mL of the reaction mixture wered (median) 16.3 { 13.2 (15.0) 16.3 { 11.0 (13.0) ú.252 mL of DNA extract, 50 pmol of the two opposing primers
Infected patients 18 (75) 6 (40) .04REP1R-Dt and REP2-Dt, and 1.25 mM each of four deoxy-Infections 23 7
nucleoside triphosphates (Pharmacia-Biotech) in a gelatin (10 Extraintestinal infectedmM Tris-HCl [pH 8.3]–50 mM KCl–1.5 mM MgCl2 –0.01% or colonized sites 31* 13†
gelatin) buffer with 2 U of Taq DNA polymerase (AmpliTaq;NOTE. ESBL Å extended-spectrum b-lactamase; ICU Å intensive carePerkin-Elmer, Roissy Charles-de-Gaulle, France). PCR cycling
unit; period 1 Å during which all 239 patients admitted to the unit receivedwas carried out in a Perkin-Elmer 9600 automated thermocycler prophylactic SDD; period 2Å during which only 165 colonized and/or infected
patients admitted to the unit received curative SDD; SDDÅ selective digestivewith an initial denaturation of DNA templates (957C for 7decontamination (erythromycin [1 g b.i.d.] and polymyxin E [6 million unitsminutes) followed by 30 amplification cycles (denaturation atb.i.d.]). Unless stated otherwise, data are no. of indicated characteristics or no.
957C for 30 seconds), annealing (407C for 1 minute), extension (%) of patients with indicated characteristic.* 1.3 sites per patient.(657C for 8 minutes), and finally a single extension phase (657C† 0.9 site per patient.for 16 minutes). Amplified DNA (15 mL) was analyzed by
electrophoresis at 100 V for 1.5 hours on 2% agarose gel(Sigma, Saint Quentin Fallavier, France) containing 0.5 mg of
tive patients. Thirty-two (58%) of these 55 patients developedethidium bromide/mL in 1X TAE (Tris-acetate-EDTA) buffer.45 infections (urinary tract infection, 20 patients; skin woundA 1-kb ladder (GIBCO–BRL Life Technologies) was used asinfection, 9; pneumonia, 5; sinusitis, 5; catheter-related infec-the DNA molecular size marker.tion, 3; bacteremia, 1; and other infections, 2). The first site ofisolation of ESBL-producing K. pneumoniae was the digestive
Statistical Methods tract in 28 (51%) of the 55 ICU-acquired cases. Of the 32infected patients, nine (28%) had evidence of previous or con-Categorical variables were compared by means of the x2
comitant digestive tract colonization, and 13 (41%) remainedtest or Fisher’s exact test when appropriate. All tests were two-free of digestive tract colonization.sided, and a P value of õ.05 was considered significant.
The characteristics of ESBL-producing K. pneumoniae–pos-itive patients during each of the two SDD periods are shown
Results in table 1. No significant differences were seen in either theincidence of acquired cases (10.0% vs. 9.1% for periods 1
Clinical and Bacteriologic Surveillanceand 2, respectively) or the time to acquisition. Although thedifference was not significant, SDD use during period 1 wasFrom 17 February 1992 to 30 June 1993, 546 patients wereassociated with a decrease in the rate of acquisition in theadmitted to our ICU, of whom 65 were found to be colonizeddigestive tract and an increase in the number of extraintestinaland/or infected with an ESBL-producing strain of the Entero-sites in ESBL-producing K. pneumoniae–positive patientsbacteriaceae family. K. pneumoniae was isolated from 64 pa-compared with SDD use during period 2. The number of infec-tients, and Serratia marcescens was isolated from one patient.tions in ESBL-producing K. pneumoniae–positive patients wasIn three of the K. pneumoniae–positive cases, an ESBL-pro-significantly higher during period 1 than during period 2.ducing strain of E. coli, Enterobacter aerogenes, or P. mirabilis
was also isolated.On the basis of clinical and bacteriologic criteria, nine
Epidemiological Markers for ESBL-Producing K. pneumoniaeESBL-producing K. pneumoniae–positive cases were classi-
Strainsfied as referred to the ICU, and 55 of these cases were referredto as acquired in the ICU. The mean time to acquisition {SD A total of 138 ESBL-producing K. pneumoniae strains (iso-
lated from 64 patients), including 62 recovered from surveil-in the ICU-acquired cases was 17.6 { 13.3 days (median, 13days; range, 4–66 days). Digestive tract colonization was found lance samples (stool and urine) and 76 isolated from clinical
specimens, were studied.in 38 (69%) of the 55 ESBL-producing K. pneumoniae–posi-
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

837CID 1998;27 (October) Epidemiology of ESBL-Producing K. pneumoniae
Table 2. Antibiotic resistance patterns for 138 strains of ESBL-producing Klebsiella pneumoniae isolated from infected and/or colonizedpatients in an intensive care unit.
Resistance markerPhenotype Phenotype(N/n), pI (N/n) Nm Gm Tm Amik NA Cpfx Chl Tet Sul
CAZ (114/52)7.8 I (79/35) R S R R R S R R R7.8 II (19/8) S S R R R S R S R7.8 III (4/2) R S R R R R R S R7.8 IV (6/4) R S R R R R I R R7.8 V (2/1) S S R R S S S R R8.2 VI (2/1) S S R R R R R S R8.2 VII (2/1) R S R R R R S R R
CTX (24/13)6.3 II (7/4) S S R R R S R S R6.3 III (3/1) R S R R R R R S R6.3 VIII (9/3) S S R R R R I R R6.3 IX (1/1) R R R R R S R R R7.6 X (1/1) S S S S S S S R R7.6 XI (1/1) S S S S S S R R R7.6 XII (1/1) S S R R S S R S S7.0 XIII (1/1) R R R R R R I R R
NOTE. Amik Å amikacin; CAZ phenotype Å smaller inhibition zone diameters around ceftazidime and aztreonam disks than around cefotaxime disk;ChlÅ chloramphenicol; CpfxÅ ciprofloxacin; CTX phenotypeÅ similar inhibition zone diameters for ceftazidime, aztreonam, and cefotaxime; ESBLÅ extended-spectrum b-lactamase; Gm Å gentamicin; I Å intermediately resistant; n Å number of patients; N Å number of strains; NA Å nalidixic acid; Nm Å neomycin;pI Å isoelectric point; R Å resistant; S Å susceptible; Sul Å sulfonamides; Tet Å tetracycline; Tm Å tobramycin.
Biotypes above. All isolates produced a b-lactamase with a pI of 7.7,which was probably related to the SHV-1 chromosomal b-Six biotypes were observed among the 138 strains tested.lactamase of K. pneumoniae. Of 52 strains with the CAZ phe-Only 19% of the strains exhibited the common K. pneumoniaenotype, 50 produced a b-lactamase with a pI of 7.8, and twobiotype (urease activity, lysine decarboxylase activity, and su-produced a b-lactamase with a pI of Ç8.2. Of 13 strains withcrose fermentation); 61.5% had weak or negative urease activ-the CTX phenotype, nine produced a b-lactamase with a pI ofity, and 7% were negative for both urease activity and sucrose6.3 (usually accompanied by a band at pI 5.4), three producedfermentation.a b-lactamase with a pI of 7.6, and one produced a b-lactamasewith a pI of 7.0. These results were consistent with the produc-Antibiotic Resistance Patternstion of the following ESBLs: SHV-4, SHV-5, TEM-3 (CTX-Of the 138 strains, 114 (83%) isolated from 52 patients1), SHV-2, and SHV-3.(80%) exhibited the CAZ phenotype. The aminoglycoside re-
sistance pattern for all the strains with the CAZ phenotypethat were susceptible only to gentamicin was consistent with
Plasmid Analysisresistance mediated by 6*-N-aminoglycoside-acetyltransferase;resistance to neomycin was also present in 74.5% of strains. Plasmid content was determined for the 138 strains; thisAll strains with the CAZ phenotype were resistant to sulfon- analysis demonstrated 20 plasmid profiles, including one toamides and trimethoprim. Other resistance markers allowed five plasmids ranging from Ç20 to ú150 kb (figure 1). Theindividualization of five different phenotypes. Twenty-four predominant profile was Cp1, which corresponded to the pres-strains (17%) isolated from 13 patients (20%) had the CTX ence of two large plasmids of similar molecular sizephenotype; 87.5% also exhibited 6*-N-aminoglycoside-acetyl- (ú150 kb); this profile was found for 79 strains (isolated fromtransferase–mediated resistance. 32 patients). Two variants, Cp1a and Cp1b profiles, each
A total of 13 phenotypes were individualized, among which included an additional plasmid of smaller molecular sizephenotypes II and III were seen in strains with both the CAZ (õ50 kb), and these profiles were found for nine and 10 strainsand the CTX phenotypes (table 2). (isolated from five and six patients), respectively. The Cp1,
Cp1a, and Cp1b profiles shared similar cryptic plasmid con-b-Lactamase Analysis tents.
All other plasmid contents were seen in strains isolated fromElectrofocusing was performed on extracts from 65 strains,including representatives of all resistance phenotypes defined only a few patients (at least three). In a few cases, several
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

838 Decre et al. CID 1998;27 (October)
Figure 1. Plasmid profiles forisolates of extended-spectrum b-lactamase-producing Klebsiellapneumoniae recovered from in-fected and/or colonized patients inan intensive care unit. Lanes G, Q,and Y: RP4 (54 kb) and pIP173(126.8 kb); lanes H, R, and X:pCFF04 (85 kb) and supercoiledDNA ladder (2–16 kb [GIBCO-BRL Life Technologies, Cergy-Pontoise, France]); lane A: Cp1;lane B: Cp2; lane C: Cp3; lane D:Cp4; lane E: Cp5; lane F: Cp6;lane I: Cp7; lane J: Cp8; lane K:Cp9; lane L: Cp10; lane M: Cp11;lane N: Cp12; lane O: Cp13; laneP: Cp14; lane S: Cp1; lane T:Cp15; lane U: Cp16; lane V: Cp17;lane W: Cp18; lane Z: Cp1; laneAA: Cp1a; and lane AB: Cp1b. KbÅ kilobase.
plasmid contents shared a similar large plasmid but differed (e.g., P1 and P1a profiles and P1 and P1b profiles in strainsisolated from one and two patients, respectively); only oneregarding their cryptic plasmid patterns (e.g., the Cp3, Cp17,
Cp18 group and the Cp15, Cp16 group). Cp15 and the Cp1, patient was colonized with strains exhibiting markedly differentpatterns (P2 profile and then P3 profile).Cp1a, Cp1b group shared the same cryptic plasmid pattern but
differed regarding their large plasmid. The temporal distribution of the ESBL-producing K. pneu-moniae strains isolated from the 64 ICU patients according tothe PFGE profiles of the strains is shown in figure 3. This
Genomic Fingerprinting by PFGE distribution revealed the occurrence of dissemination of anepidemic strain over the 16-month period.Of the restriction endonucleases tested, only XbaI and SpeI
were found to generate PFGE profiles that were easy to analyze.Genomic Fingerprinting by Repetitive-Element PCR AnalysisXbaI digestion allowed differentiation of 19 profiles (figure 2),in Comparison with That by PFGEof which 14 (P1 to P14) were markedly different. In contrast,
six profiles (P1, P1a, P1b, P1c, P1d, and P1e) had very similar Repetitive-element PCR analysis of 36 ESBL-producing K.band patterns (three or less band differences) with both XbaI pneumoniae isolates exhibiting the 19 profiles individualizedand SpeI. by means of PFGE was performed, and rep-PCR analysis dis-
The P1, P1a, P1b, and P1c profiles differed mainly regarding tinguished 14 different profiles (R1 to R14). The R1 and R1aa single high molecular weight band: P1a and P1b differed profiles were very similar; the R1a profile had only one addi-from P1 by the absence of a band with a molecular size of tional amplified fragment (õ0.2 kb) (figure 4).Ç400 kb and 450 kb, respectively, and P1c included an addi- When compared with DNA macrorestriction polymorphismtional band with a molecular size of Ç480 kb. The P1e and determined by PFGE, the 14 different PFGE profiles (P1 toP1d profiles differed by three bands with molecular sizes that P14) corresponded to different rep-PCR fingerprinting patternsranged from 150 to 200 kb; they were closely similar to P1, (R1 to R14). Strains isolated from different patients that hadwith a few differences in bands with molecular sizes that ranged the same PFGE profile also had the same rep-PCR profile.from 100 to 250 kb. Reproducible profiles were obtained in Finally, the 18 strains with almost identical PFGE patterns (P1,independent experiments (data not shown). P1a, P1b, P1c, P1d, and P1e) also had very similar rep-PCR
The P1 profile was found in 75 strains isolated from 32 fingerprinting patterns, which could be subdivided only intopatients; taken together, the P1, P1a, P1b, and P1c profiles two profiles—namely, R1 (corresponding to P1, P1a, P1b, andaccounted for 98 strains (isolated from 43 patients). The P3, P1c profiles) and R1a (corresponding to P1e and P1d profiles).P4, and P7 profiles were found in strains isolated from six,two, and two patients, respectively, and each of the other pro-
Comparison of Phenotypic and Genotypic Markersfiles was observed in one or several strains recovered from asingle patient. Multiple isolates recovered from different body Antibiotic resistance patterns are compared with genotypic
markers (plasmid contents and PFGE profiles) in figure 5. Insites in a given patient had identical or closely related patterns
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

839CID 1998;27 (October) Epidemiology of ESBL-Producing K. pneumoniae
Figure 2. Profiles produced bypulsed-field gel electrophoresis ofXbaI macrorestriction fragments ofextended-spectrum b-lactamase-producing Klebsiella pneumoniaeisolates from 64 infected and/orcolonized patients in an intensivecare unit. Lanes A and U: molecu-lar size marker (l DNA concate-mer; Bio-Rad S. A., Ivry sur Seine,France); lane B: genotype P1; laneC: genotype P1a; lane D: genotypeP1b; lane E: genotype P1c; laneF: genotype P1d; lane G: genotypeP1e; lane H: genotype P2; lane I:genotype P3; lane J: genotype P4;lane K: genotype P5; lane L: geno-type P6; lane M: genotype P7; laneN: genotype P8; lane O: genotypeP9; lane P: genotype P10; lane Q:genotype P11; lane R: genotypeP12; lane S: genotype P13; andlane T: genotype P14. Kb Å kilo-base.
most cases, plasmid contents and PFGE profiles agreed closely. dissemination of such strains in our hospital since 1989 promptedinstitution of a control program for ESBLs and an epidemiologi-A b-lactamase with a pI of 7.8 that was consistent with SHV-4cal study of ESBL-producing strains that was aimed at elucidatingwas the most frequent type observed. Most of SHV-4 producersthe mechanisms of patient-to-patient transmission.exhibited one of the three very similar plasmid contents Cp1,
Over a 16-month study period in our ICU, the incidence ofCp1a, and Cp1b and consistently corresponded to the fourcolonization and/or infection due to ESBL-producing strainsclosely related PFGE profiles P1, P1a, P1b, and P1c. Thesewas 11.9% (65 of 546 patients). As previously reported,four profiles probably corresponded to four variants of the sameK. pneumoniae was the most frequent species isolated (64clonal strain. Strains with the PFGE profile P3 had one of theof 65 patients) [10, 39]. Clinical and bacteriologic data sug-three plasmid contents Cp3, Cp17, and Cp18, which shared thegested that most cases (86%) were acquired in the unit andsame large plasmid.that only 14% of cases were referred to the unit. TheseThe markers correlated well for all strains related to SHV-findings suggest that enhanced isolation measures may be5, SHV-2, and SHV-3 b-lactamases. Similar close correlationswarranted for patients hospitalized in ICUs of both univer-were observed for the strains related to TEM-3 b-lactamase,sity and general hospitals.with the exception of those exhibiting the P1e or P1d profile.
Prophylactic SDD was reported to be effective in control-These strains had PFGE profiles closely similar to the predomi-ling epidemic multidrug-resistant gram-negative bacilli innant P1 profile corresponding to SHV-4 b-lactamase, althoughICUs [40]. Different combinations have been used. The mostthe b-lactamase that they produced was different.widely used combinations of nonabsorbable antibiotics in-clude aminoglycoside, older quinolones, and polymyxin E.In our hospital, ESBL-producing Enterobacteriaceae wereDiscussionresistant to most aminoglycosides and quinolones; therefore,
Transferable resistance to third-generation cephalosporins and these compounds were avoided, and the choice for SDD wasmonobactam agents in Enterobacteriaceae has been reported polymyxin E and erythromycin. To prevent translocation insince 1983 and is now found worldwide. ESBL-producing strains a hematology unit, erythromycin base was used to reducehave been widely described in outbreaks of nosocomial infec- the aerobic gram-negative flora of the gastrointestinal tract
[41]. Our use of erythromycin was based on these data andtions, particularly in ICUs [1, 3, 5, 6, 10, 21, 22]. Continuing
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

840 Decre et al. CID 1998;27 (October)
Figure 3. Temporal distribution of extended-spectrum b-lactamase-producing Klebsiella pneumoniae isolated from infected and/or colonizedpatients in an intensive care unit according to their profiles determined by pulsed-field gel electrophoresis (PFGE).
on the previously reported high concentrations of erythromy- Enterobacteriaceae [43]. The incidence of gastrointestinal tractcolonization decreased nonsignificantly during routine SDDcin in stools [42].
In our study, prophylactic SDD failed to significantly reduce use (period 1). However, the proportion of ESBL-producingK. pneumoniae–positive patients who were infected was higherthe incidence of acquisition of ESBL-producing strains. These
data are in keeping with another study reporting failure of during period 1 than during period 2, as was the number ofextraintestinal sites that were colonized and/or infected. Rea-erythromycin to prevent colonization with multidrug-resistant
Figure 4. Repetitive-element(rep)-PCR fingerprints generatedby REP1R-Dt and REP2-Dt prim-ers of extended-spectrum b-lacta-mase-producing Klebsiella pneu-moniae isolates from infected and/or colonized patients in an inten-sive care unit. Lanes A, M, N, andZ: molecular size marker (1-kbDNA ladder; GIBCO-BRL LifeTechnologies, Cergy-Pontoise,France); lanes B, C, D, E, F, G,and H: R1; lanes I and J: R1a; laneK: R2; lane L: R3; lane O: R4;lane P: R5; lane Q: R6; lane R:R7; lane S: R8; lane T: R9; laneU: R10; lane V: R11; lane W: R12;lane X: R13; and lane Y: R14. KbÅ kilobase.
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

841CID 1998;27 (October) Epidemiology of ESBL-Producing K. pneumoniae
Figure 5. Differentiation of 138strains of extended-spectrum b-lactamase-producing Klebsiellapneumoniae isolated from 64 in-fected and/or colonized patients inan intensive care unit on the basisof phenotypic (isoelectric point[pI] and resistance [R] phenotype)and genotypic (plasmid contentand pulsed-field gel electrophore-sis [PFGE] profile) markers. CAZphenotype Å smaller inhibitionzone diameters around ceftazidimeand aztreonam disks than aroundcefotaxime disk; CTX phenotypeÅ similar inhibition zone diame-ters for ceftazidime, aztreonam,and cefotaxime.
sons for these findings are unclear; the risk of infections during reservoirs for ESBL-producing K. pneumoniae strains. Ineffec-tive SDD, used as either prophylaxis or a curative regimen,period 1 could have been higher. Unfortunately, underlying
diseases and invasive procedures were not recorded over the was stopped after 1 year.Implementation of enhanced isolation procedures actuallystudy period.
SDD seemed effective for intestinal decontamination but helped to reduce the incidence of colonization and infection dueto ESBL-producing strains. An observational study of isolationfailed to prevent the occurrence of ICU-acquired cases, particu-
larly at extraintestinal sites. We postulated that other sites, such precautions was performed in February 1993. Breaks in contactisolation procedures were observed especially when a patient’sas the oropharynx, colonized wounds, or urine, were alternative
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

842 Decre et al. CID 1998;27 (October)
care was disrupted by other requests. Nursing care was there- sis of both large and cryptic plasmids showed a number of clearlydifferent profiles, and the results were in agreement with thosefore reorganized with subsequent control of the outbreak. Pre-
liminary results of this study have been presented [44] and will of other molecular typing methods with the exception of the P3profile and the P1e, P1d group. Other investigators also reportedbe reported elsewhere.
We combined phenotypic and genotypic methods to eluci- good agreement between results of plasmid content analysis andchromosomal typing methods [47, 48]. In contrast, plasmid con-date the transmission of ESBL-producing K. pneumoniae
strains in our ICU. Most strains produced SHV-4 b-lactamase, tent analysis failed to efficiently differentiate strains with similari-ties regarding large and/or cryptic plasmids.which is one of the most frequent types reported in France [1,
5, 12–14, 45]. The high frequency of negative or weak urease Of the chromosomal genotypic methods, analysis of restric-tion fragment length polymorphisms of the rRNA regions (ribo-activity of such strains in our study is also consistent with
earlier data [1, 5]. Most strains displayed homogeneous geno- typing) was reported to effectively differentiate types of ESBL-producing K. pneumoniae [2].typic markers, thus suggesting dissemination of an epidemic
strain and its subclonal variants. Our findings indicate that PFGE of chromosomal DNA restriction polymorphism wasalso used for epidemiological studies and was reported to bethis single epidemic clone and its subcloncal variants were
responsible for as many as 85% of our ICU-acquired cases. This superior over other genotypic methods for typing various bacte-rial genera including Klebsiella [46, 47, 52]. Our study confirmedfinding is consistent with previous reports that dissemination
of SHV-4 b-lactamase–producing organisms in France was the usefulness of this method since a high degree of DNA poly-morphism was observed for all clinically unrelated strains. Inascribable mainly to a single strain [45].
Although transfer of SHV-4 b-lactamase has been reported contrast, the cluster of strains with profiles from the P1 groupexhibited only minor genomic differences. Similar findings wereto be readily achieved in vitro [4, 45], no other SHV-4 b-
lactamase–producing Enterobacteriaceae were identified dur- reported for outbreaks in which clones exhibiting closely relatedpatterns were recovered [47, 53]. Analysis of the other markersing the study. Other sporadic cases were related to strains
producing various ESBL types such as SHV-5, SHV-2, and used in our study confirmed that strains with four P1 profiles(P1, P1a, P1b, and P1c) were probably related since all of themSHV-3. TEM-3 b-lactamase–producing strains, which have
been described in several outbreaks [2, 3, 9, 28, 46], were produced SHV-4 b-lactamase and demonstrated one of the threevariant plasmid contents Cp1, Cp1a, and Cp1b. In contrast, theisolated from only nine patients and accounted for only a few
secondary cases. strains exhibiting the P1e and P1d profiles produced TEM-3 b-lactamase and shared a large plasmid.Multiple isolates recovered from different body sites in a
given patient were typically either identical or members of the These findings are not conflicting, since transfer of plasmidsencoding for various types of b-lactamases may occur amongsame clonal population, except those from one patient who
was successively colonized with two different strains. genetically unrelated strains. In addition, it is known that twoplasmids belonging to the same incompatibility group cannotPrevious studies of the spread of ESBL-producing K. pneu-
moniae used various typing methods, including determination be present in the same strain. The incompatibility groups ofplasmids encoding for TEM-3 b-lactamase have been reportedof phenotypes and, more recently, determination of genotypic
markers [2, 29, 46, 47]. Our investigation confirmed that pheno- for 85-kb plasmids, such as pCFF04, but not for plasmids withhigher molecular weights (ú150 kb) [5, 15]. We believe thattypic characteristics have little discriminating power. For
instance, an uncommon biotype (weak or negative urease activ- the P1, P1a, P1b, P1c, P1d, and P1e profiles indicated prove-nance from the same clonal population.ity) was observed in 83% of the SHV-4 b-lactamase–produc-
ing strains but was also found in 30% of the TEM-3 b-lacta- A new faster genotypic method with use of primer sets de-signed to amplify chromosomal regions lying between repeti-mase–producing strains.
Since most strains displayed resistance to multiple antimicro- tive multicopy elements, which are widely dispersed in eubac-terial genomes, was recently used for epidemiological studiesbials, antibiotic resistance profiles were also unsatisfactory for
differentiating isolates, except when unusual patterns (such as of bacterial strains [38, 54–56]. We used 38-bp REP elementsas primers. Rep-PCR fingerprinting of 36 selected strainssusceptibility to all aminoglycosides, quinolones, or sulfon-
amides) were exhibited. showed good agreement with DNA macrorestriction analysisby PFGE: all epidemiologically unrelated strains that werePlasmid analysis is an easy rapid method that was previously
used to differentiate strains of K. pneumoniae [1, 2, 20, 48–50]. found to be different by PFGE also exhibited distinct rep-PCR profiles. However, rep-PCR analysis was less accurateDetermination of plasmid contents is crucial to the analysis of
plasmid dissemination among genetically unrelated strains [51]. for differentiating strains belonging to the same subclonal pop-ulation. Similar data were found for other gram-negative bacte-However, plasmid contents are not reliable for determining the
spread of strains: genetically unrelated strains may harbor the ria with use of enterobacterial repetitive intergenic consensusprimers [57]. Rep-PCR fingerprinting, which does not requiresame plasmids [1, 2], and because plasmids are not a stable part
of the bacterial genome, related strains may lose plasmids [49]. conventional DNA extraction, is suitable for rapid screeningof strains during epidemiological investigations.In our study, all the isolates tested had detectable plasmids. Analy-
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

843CID 1998;27 (October) Epidemiology of ESBL-Producing K. pneumoniae
siella pneumoniae isolates in a pediatric hospital. J Clin Microbiol 1993;The molecular findings supported the validity of our clinical31:179–84.criteria for differentiation between ICU-acquired cases and
3. Brun-Buisson C, Legrand P, Philippon A, Montravers F, Ansquer M,ICU-referred cases: most of the strains from the nine cases Duval J. Transferable enzymatic resistance to third-generation cephalo-classified as ICU-referred exhibited different b-lactamase types sporins during a nosocomial outbreak of multi-resistant Klebsiella pneu-
moniae. Lancet 1987;2:302–6.and highly different genomic profiles. It is interesting that an4. Bure A, Legrand P, Arlet G, Jarlier V, Paul G, Philippon A. Disseminationidentical profile (P3) was identified in two ICU-referred cases
in five French hospitals of Klebsiella pneumoniae serotype K25 harbour-that occurred at different times and in different places. Thising a new transferable resistance to third-generation cephalosporins and
finding is further evidence that interhospital spread of SHV-4 aztreonam. Eur J Clin Microbiol Infect Dis 1988;7:780–2.b-lactamase–producing K. pneumoniae strains is occurring. 5. De Champs C, Sirot D, Chanal C, Poupart MC, Dumas MP, Sirot J.
Concomitant dissemination of three extended-spectrum b-lactamasesClinical data agreed with typing data in 51 of the 55 ICU-among different Enterobacteriaceae isolated in a French hospital. J Anti-acquired cases (which occurred in 50 patients); of the strainsmicrob Chemother 1991;27:441–7.
from these patients, most isolates (from 43 patients) corresponded6. Jarlier V, Nicolas MH, Fournier G, Philippon A. Extended-broad-spectrum
to the predominant clonal population (P1) producing SHV-4 b- b-lactamases conferring transferable resistance to newer b-lactamagents in Enterobacteriaceae: hospital prevalence and susceptibility pat-lactamase. Five cases (in five patients) classified as ICU-acquiredterns. Rev Infect Dis 1988;10:867–78.on the basis of our clinical criteria were probably referred to the
7. Philippon A, Ben Redjeb S, Fournier G, Ben Hassen A. Epidemiology ofICU since their strains displayed different molecular patterns (P2,extended-spectrum b-lactamases. Infection 1989;17:347–51.
P4, P5, P8, and P12, respectively). Rectal screening at admission 8. Sirot D, Sirot J, Morand A, et al. Transferable resistance to third-generationto the ICU was negative for three of these five patients, but these cephalosporins in clinical isolates of Klebsiella pneumoniae. Identifica-
tion of CTX-1, a novel b-lactamase. J Antimicrob Chemother 1987;20:tests were positive within 7 days after admission. The sensitivity323–34.of the screening method (ú102 cfu/g) [25] may explain these
9. Sirot J, Chanal C, Petit A, Sirot D, Labia R, Gerbaud G. Klebsiella pneu-observations. In addition, prophylactic SDD may explain the moniae and other Enterobacteriaceae producing novel plasmid-mediateddelayed positive results of rectal screening (days 11 and 30, b-lactamases markedly active against third-generation cephalosporins:
epidemiologic studies. Rev Infect Dis 1988;10:850–9.respectively) for the two remaining patients.10. Sirot D, Goldstein F, Soussy CJ, et al. Resistance to cefotaxime and otherIn conclusion, the molecular findings of this study were
b-lactams in members of the Enterobacteriaceae: a 3-year survey inconsistent with the ICU-acquired origin of most cases andFrance. Antimicrob Agents Chemother 1992;36:1677–81.
showed that an epidemic clone and its subclonal variants pro- 11. Sirot D. Extended-spectrum b-lactamases. J Antimicrob Chemother 1995;ducing SHV-4 b-lactamase was responsible for 85% of ICU- 36(suppl A):19–34.
12. Jacoby GA, Medeiros AA. More extended-spectrum b-lactamases. Antimi-acquired cases. Other sporadic cases were related to strainscrob Agents Chemother 1991;35:1697–704.producing various ESBL types such as SHV-5, SHV-2, SHV-
13. Philippon A, Labia R, Jacoby GA. Extended-spectrum b-lactamases. Anti-3, and TEM-3; a few of these strains were responsible for a microb Agents Chemother 1989;33:1131–6.limited number of secondary cases. 14. Philippon A, Arlet G, Lagrange PH. Origin and impact of plasmid-medi-
ated extended-spectrum beta-lactamases. Eur J Clin Microbiol InfectWe found no evidence that persistence of the dominant SHV-Dis 1994;13(suppl 1):S17–9.4 b-lactamase–producing clone was related to virulence factors
15. Jacoby GA, Sutton L. Properties of plasmids responsible for productiondescribed in K. pneumoniae [58]. None of 10 strains testedof extended-spectrum b-lactamases. Antimicrob Agents Chemother
produced aerobactin or mucoid factor (V. Vernet, unpublished 1991;35:164–9.data). However, plasmid-mediated adhesive factor was de- 16. Petit A, Gerbaud G, Sirot D, Courvalin P, Sirot J. Molecular epidemiology
of TEM-3 (CTX-1) b-lactamase. Antimicrob Agents Chemother 1990;scribed in ESBL-producing K. pneumoniae strains [59] and34:219–24.more recently in K. pneumoniae producing SHV-4 b-lactamase
17. Goldstein FW, Pean Y, Rosato A, Gertner J, Gutmann L, the Vigil’Roc[60], which may explain clonal dissemination. Study Group. Characterisation of ceftriaxone-resistant Enterobacteria-
Although phenotypic characteristics discriminated poorly be- ceae: a multicentre study in 26 French hospitals. J Antimicrob Chemo-ther 1993;32:595–603.tween strains, various genotypic methods distinguished epidemic
18. Hammani A, Arlet G, Ben Redjeb S, et al. Nosocomial outbreak of acutestrains from unrelated isolates. Combined use of several genotypicgastroenteritis in a neonatal intensive care unit caused by multiply drug
methods accurately differentiated between closely related strainsresistant Salmonella wien producing SHV-2 b-lactamase. Eur J Clin
and was helpful for evaluating interstrain genetic relatedness. We Microbiol Infect Dis 1991;10:641–6.conclude that genotypic methods are accurate for analyzing dis- 19. Mariotte S, Nordmann P, Nicolas MH. Extended-spectrum b-lactamase
in Proteus mirabilis. J Antimicrob Chemother 1994;33:925–35.semination of epidemic strains over extended periods.20. Bauerfeind A, Rosenthal E, Eberlein E, Holley M, Schweighart S. Spread
of Klebsiella pneumoniae producing SHV-5 b-lactamase among hospi-talized patients. Infection 1993;21:18–22.
21. Johnson AP, Weinbren MJ, Ayling-Smith B, Du Bois SGB, George RC.ReferencesOutbreak of infection in two UK hospitals caused by a strain of Kleb-
1. Arlet G, Sanson-le-Pors MJ, Rouveau M, et al. Outbreak of nosocomial siella pneumoniae resistant to cefotaxime and ceftazidime. J Hosp Infectinfections due to Klebsiella pneumoniae producing SHV-4 b-lactamase. 1992;20:97–103.Eur J Clin Microbiol 1990;9:179–84. 22. Meyer KS, Urban C, Eagan JA, Berger BJ, Rahal JJ. Nosocomial outbreak
2. Bingen E, Desjardin P, Arlet G, et al. Molecular epidemiology of plasmid of Klebsiella pneumoniae resistant to late-generation cephalosporins.Ann Intern Med 1993;119:353–8.spread among extended broad-spectrum b-lactamase-producing Kleb-
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from

844 Decre et al. CID 1998;27 (October)
23. Naumovski L, Quinn JP, Miyashiro D, et al. Outbreak of ceftazidime 44. Lucet JC, Fichelle A, Decre D, et al. Control of a prolonged outbreak ofextended-spectrum b-lactamase-producing Enterobacteriaceae in a univer-resistance due to a novel extended-spectrum b-lactamase in isolates of
cancer patients. Antimicrob Agents Chemother 1992;36:1991–6. sity hospital [abstract no. J108]. In: Program and abstracts of the 36thInterscience Conference on Antimicrobial Agents and Chemotherapy (New24. Rice LB, Willey SH, Papanicolaou GA, et al. Outbreak of ceftazidime resis-
tance caused by extended-spectrum b-lactamases at a Massachusetts Orleans). Washington, DC: American Society for Microbiology, 1996:238.45. Arlet G, Rouveau M, Casin I, Bouvet PJM, Lagrange PH, Philippon A.chronic care facility. Antimicrob Agents Chemother 1990;34:2193–9.
25. De Champ C, Sauvant MP, Chanal CM, et al. Prospective survey of Molecular epidemiology of Klebsiella pneumoniae strains that produceSHV-4 b-lactamase and which were isolated in 14 French hospitals. Jcolonization and infections caused by expanded-spectrum b-lactamase-
producing members of the family Enterobacteriaceae in an intensive Clin Microbiol 1994;32:2553–8.46. Gouby A, Neuwirth C, Bourg G, et al. Epidemiological study by pulsed-care unit. J Clin Microbiol 1989;12:2887–90.
26. Casewell MW, Phillips I. Hands as a route of transmission for Klebsiella field gel electrophoresis of an outbreak of extended-spectrum b-lacta-mase-producing Klebsiella pneumoniae in a geriatric hospital. J Clinspecies. BMJ 1977;2:1315–7.
27. Chanal CM, Sirot DL, Petit A, et al. Multiplicity of TEM-derived b- Microbiol 1994;32:301–5.47. Gori A, Espinasse F, Deplano A, Nonhoff C, Nicolas MH, Struelens M.lactamases from Klebsiella pneumoniae strains isolated at the same
hospital and relationship between the responsible plasmids. Antimicrob Comparison of pulsed-field gel electrophoresis and randomly amplifiedDNA polymorphism analysis for typing extended-spectrum-b-lactamase-Agents Chemother 1989;33:1915–20.
28. Kitzis MD, Billot-Klein D, Goldstein FW, et al. Dissemination of the producing Klebsiella pneumoniae. J Clin Microbiol 1996;34:2448–53.48. Eisen D, Russel EG, Tymms M, Roper EJ, Grayson ML, Tunidge J.novel plasmid-mediated b-lactamase CTX-1, which confers resistance
to broad-spectrum cephalosporins, and its inhibition by b-lactamase Random amplified polymorphic DNA and plasmid analysis used ininvestigation of an outbreak of multiresistant Klebsiella pneumoniae. Jinhibitors. Antimicrob Agents Chemother 1988;32:9–14.
29. Lucet JC, Chevret S, Decre D, et al. Outbreak of multiply resistant Entero- Clin Microbiol 1995;33:713–7.49. Thompson W, Romance L, Bialkowska-Hobrazanska H, Rennie RP, Ash-bacteriaceae in an intensive care unit: epidemiology and risk factors for
acquisition. Clin Infect Dis 1996;22:430–6. ton F, Nicolle LE. Klebsiella pneumoniae infection on a rehabilitationunit: comparison of epidemiologic typing methods. Infect Control Hosp30. Legrand P, Fournier G, Bure A, et al. Detection and distribution of ex-
tended-broad-spectrum b-lactamases in Enterobacteriaceae in four Epidemiol 1993;14:203–10.50. Wallia S, Madhavan T, Reuman P, Tewari R, Duckwork D. PlasmidFrench hospitals. Eur J Clin Microbiol Infect Dis 1989;8:527–9.
31. Swenson JM, Hinler JA, Peterson LR. Special tests for detecting antibacte- profiles and klebocin in epidemiologic studies of infections by Klebsiellapneumoniae. Eur J Clin Microbiol Infect Dis 1988;7:279–84.rial resistance. In: Murray PR, Baron EJ, Pfaller MA, Tenover FC,
Yolken RH, eds. Manual of clinical microbiology. 6th ed. Washington, 51. Mayer L. Use of plasmid profile in epidemiologic surveillance of diseaseoutbreaks and in tracing the transmission of antibiotic resistance. ClinDC: American Society for Microbiology, 1995:1356–67.
32. Livermore DM. b-Lactamases in laboratory and clinical resistance. Clin Microbiol Rev 1988;1:228–43.52. Goering RV. Molecular epidemiology of nosocomial infection: analysisMicrobiol Rev 1995;8:557–84.
33. Acar J, Chardon P, Choutet P, et al. Comite Francais de l’Antibiogramme of chromosomal restriction fragment patterns by pulsed-field gel electro-phoresis. Infect Control Hosp Epidemiol 1993;14:595–600.de la Societe Francaise de Microbiologie. Communique 1996. Pathol
Biol (Paris) 1996;44(8):I–VIII. 53. Kersulyte D, Struelens MJ, Deplano A, Berg DE. Comparison of arbi-trarily-primed PCR and macrorestriction (pulsed-field gel electrophore-34. Matthew M, Harris AM, Marshall MG, Ross GW. The use of analytical
isoelectric focusing for detection and identification of b-lactamases. J sis) typing of Pseudomonas aeruginosa strains from cystic fibrosis pa-tients. J Clin Microbiol 1995;33:2216–9.Gen Microbiol 1975;88:169–78.
35. Kado CI, Liu ST. Rapid procedure for detection and isolation of large and 54. Georghiou PR, Hamill RJ, Wright CE, et al. Molecular epidemiologyof infections due to Enterobacter aerogenes: identification of hospitalsmall plasmids. J Bacteriol 1981;145:1365–73.
36. Smith CL, Cantor CR. Purification, specific fragmentation, and separation outbreak–associated strains by molecular techniques. Clin Infect Dis1995;20:84–94.of large DNA molecules. Methods Enzymol 1987;155:449–67.
37. Tenover F, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA 55. Reboli AC, Houton ED, Monteforte JS, Wood CA, Hamill RJ. Discrimina-tion of epidemic and sporadic isolates of Acinetobacter baumannii byrestriction patterns produced by pulsed-field gel electrophoresis: criteria
for bacterial strain typing. J Clin Microbiol 1995;33:2233–9. repetitive element PCR-mediated DNA fingerprinting. J Clin Microbiol1994;32:2635–40.38. Versalovic J, Koeuth T, Lupski JR. Distribution of repetitive DNA se-
quences in eubacteria and application to fingerprinting of bacterial ge- 56. Woods CR, Versalovic J, Koeuth T, Lupski JR. Analysis of relationshipsamong isolates of Citrobacter diversus by using DNA fingerprints gen-nome. Nucleic Acid Res 1991;19:6823–31.
39. Liu PYF, Gur D, Hall LMC, Livermore DM. Survey of the prevalence of erated by repetitive sequence–based primers in the polymerase chainreaction. J Clin Microbiol 1992;30:2921–9.b-lactamases amongst 1000 Gram-negative bacilli isolated consecu-
tively at the Royal London Hospital. J Antimicrob Chemother 1992; 57. Struelens MJ, Carlier E, Maes N, Serruys E, Quint WGV, van Belkum A.Nosocomial colonization and infection with multiresistant Acinetobacter30:429–47.
40. Brun-Buisson C, Legrand P, Rauss F, et al. Intestinal decontamination for baumannii: outbreak delineation using DNA macrorestriction analysisand PCR-fingerprinting. J Hosp Infect 1993;25:15–32.control of nosocomial multiresistant gram-negative bacilli. Ann Intern
Med 1989;110:873–81. 58. Vernet V, Madoulet C, Chippaux C, Philippon A. Incidence of two viru-lence factors (aerobactin and mucoid phenotype) among 190 clinical41. Andremont A, Sancho-Garnier H, Tancrede C. Epidemiology of intestinal
colonization by members of the family Enterobacteriaceae highly resis- isolates of Klebsiella pneumoniae producing extended-spectrum b-lac-tamases. FEMS Microbiol Lett 1992;96:1–6.tant to erythromycin in hematology-oncology unit. Antimicrob Agents
Chemother 1986;29:1104–7. 59. Darfeuille-Michaud A, Jallat C, Aubel D, et al. R-Plasmid-encoded adhesinfactor in strains responsible for human nosocomial infections. Infect42. Andremont A, Gerbaud G, Courvalin P. Plasmid-mediated high-level resis-
tance to erythromycin in Escherichia coli. Antimicrob Agents Chemo- Immun 1992;60:44–55.60. Di Martino P, Livrelli V, Sirot D, Joly B, Darfeuille-Michaud A. A newther 1986;29:515–8.
43. De Champs C, Guelon DP, Garnier RM, et al. Selective digestive decon- fimbrial antigen harbored by CAZ-5/SHV-4-producing Klebsiella pneu-moniae strains involved in nosocomial infections. Infect Immun 1996;tamination by erythromycin-base in a polyvalent intensive care unit.
Intensive Care Medicine 1993;19:191–6. 64:2266–73.
/ 9C57$$oc14 09-15-98 19:13:29 cida UC: CID
by guest on July 17, 2011cid.oxfordjournals.org
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















