Application of Waste Glycerol as a Draw Solution for Forward ...

BIOTECHNOLOGICAL PRODUCTS AND PROCESS ENGINEERING
Development of recombinant Klebsiella pneumoniae ΔdhaTstrain for the co-production of 3-hydroxypropionic acidand 1,3-propanediol from glycerol
Somasundar Ashok & Subramanian Mohan Raj &Chelladurai Rathnasingh & Sunghoon Park
Received: 12 October 2010 /Revised: 19 January 2011 /Accepted: 20 January 2011 /Published online: 20 February 2011# Springer-Verlag 2011
Abstract Klebsiella pneumoniae converts glycerol to thespecialty chemical 1,3-propanediol (1,3-PDO), which is usedfor the production of polytrimethylene terepthalate (PTT). Inthis study, an NAD+-dependent gamma-glutamyl-gamma-aminobutyraldehyde dehydrogenase (PuuC) of K. pneumo-niae DSM 2026, which oxidizes 3-hydroxypropionaldehydeto a platform chemical 3-hydroxypropionic acid (3-HP), wascloned and overexpressed in K. pneumoniae DSM 2026 forthe co-production of 3-HP and 1,3-PDO from glycerol. Inaddition, the gene dhaT, encoding NADH-dependent 1,3-propanediol oxidoreductase (1,3-PDOR), was deleted fromthe chromosome for the balanced production of 3-HP and 1,3-PDO. The recombinant K. pneumoniae ΔdhaT, expressingpuuC, produced 3.6 g 3-HP and 3.0 g 1,3-PDO per liter withan average yield of 81% on glycerol carbon in shakeflask culture under microaerobic conditions. When a fed-batch culture was carried out under microaerobic con-ditions at pH 7.0 in a 5-l bioreactor, the recombinant K.pneumoniae ΔdhaT (puuC) strain produced 16.0 g 3-HPand 16.8 g 1,3-PDO per liter with a cumulative yield of51% on glycerol carbon in 24 h. The production of 1,3-PDO in the dhaT-deletion mutant was attributed to theexpression of NAD(P)H-dependent hypothetical oxidore-
ductase. This study demonstrates the feasibility of obtain-ing two commercially valuable chemicals, 3-HP and 1,3-PDO, at a significant scale.
Keywords 3-hydroxypropionic acid . 1,3-propanediol .
Glycerol . Co-production . Gamma-glutamyl-gamma-aminobutyraldehyde dehydrogenase .Klebsiellapneumoniae
Introduction
The platform chemical 3-hydroxypropionic acid (3-HP;C3H6O3, MW 90.08), also known as hydracrylic acid, has awide range of industrial applications for the production ofmany commercially valuable chemicals (Paster et al. 2003;Global Renewable Chemicals Market 2009-2014). Recent-ly, we have developed several recombinant Escherichia colistrains for the production of 3-HP from glycerol byexpressing B12-dependent glycerol dehydratase and variousaldehyde dehydrogenases (Raj et al. 2008; Jo et al. 2008;Rathnasingh et al. 2009; Raj et al. 2009). Among them, therecombinant strain SH-BGK1 produced 3-HP at a maxi-mum of 39 g/l with an average yield of 35% (Rathnasinghet al. 2009). However, the requirement of the exogenoussupply of high-cost coenzyme B12 for the culture mediumwas a serious obstacle to the commercial application of theprocess. In addition, the yield of 3-HP on glycerol from theabove recombinant strains was not satisfactory. A potentialstrategy to solve the aforementioned problems is to useKlebsiella pneumoniae strain as a host and produce 3-HPand 1,3-propanediol (1,3-PDO) simultaneously from glyc-erol. K. pneumoniae does not require an exogenous supply
S. Ashok : S. M. Raj : C. Rathnasingh : S. Park (*)Department of Chemical and Biomolecular Engineering,International Affairs, Pusan National University,Busan 609-735, South Koreae-mail: [email protected]
S. M. RajDepartment of Biotechnology and Chemical Engineering,Kalasalingam University,Krishnankoil, TN 626-190, India
Appl Microbiol Biotechnol (2011) 90:1253–1265DOI 10.1007/s00253-011-3148-z

of coenzyme B12 for the glycerol dehydratase reaction, as itnaturally synthesizes B12. This process would yield anotherimportant specialty chemical, 1,3-PDO, and thus produc-tion of 3-HP along with 1,3-PDO is expected to increasethe carbon yield on glycerol.
In K. pneumoniae, glycerol is metabolized in twodifferent routes, namely (1) the oxidative route, leading topyruvate via the Embden–Meyerhof–Parnas (EMP) path-way, and (2) the reductive route, leading to 1,3-PDOformation (Fig. 1). In the oxidative route, glycerol isdissimilated to dihydroxyacetone phosphate (DHAP)through three different pathways depending on the electronacceptors available. Under aerobic conditions the glpK–glpD pathway is utilized, while under anaerobic conditions,the glpK–glpACB respiratory pathway (in the presence offumarate or nitrate) or dhaD/gldA–dhaK/dhaKLM fermen-tative pathway (in the absence of exogenous electronacceptor) becomes dominant (Lin 1976; Johnson and Lin1987; Ahrens et al. 1998; Blankschien et al. 2010). In thereductive route, a B12-dependent glycerol dehydratase(DhaB) converts glycerol to 3-hydroxypropionaldehyde(3-HPA), which is subsequently reduced to 1,3-PDO by atleast two 1,3-propanediol oxidoreductases (1,3-PDORs)such as DhaT (NADH-PDOR) and NADPH-dependenthypothetical oxidoreductase (NADPH-HOR) (Wang et al.2003; Zhao et al. 2009; Seo et al. 2010). The physiologicalrole of DhaT is primarily to regenerate NAD+ for DhaD/GldA to dissimilate glycerol through EMP pathway forenergy production, while NADPH-HOR is known to beexpressed in response to elevated levels of aldehydes,including 3-HPA, in order to degrade the toxic aldehydes(Wang et al. 2003; Perez et al. 2008; Seo et al. 2010).
Our recent studies have demonstrated that the geneproduct of puuC of K. pneumoniae, an NAD+-dependentgamma-glutamyl-gamma-aminobutyraldehyde dehydroge-nase, efficiently converts 3-HPA to 3-HP (Raj et al.2010). However, the wildtype K. pneumoniae DSM 2026did not produce 3-HP at a significant level during glycerolfermentation, since the native expression of this gene in K.pneumoniae is low. It is expected that if puuC is overex-pressed, K. pneumoniae can produce 3-HP along with 1,3-PDO, but it will result in increased amounts of reducingequivalents (NADH). The consequence of this effect willresult in enhanced production of 1,3-PDO rather than 3-HP.An effective way to achieve the balanced production ofboth compounds is to disrupt dhaT and utilize the NADPH-HOR. This study aimed to develop recombinant K.pneumoniae for the co-production of 3-HP and 1,3-PDOat a high glycerol conversion yield. To this end, wedeveloped recombinant K. pneumoniae strains by over-expressing puuC and/or disrupting dhaT genes, and theirperformance was evaluated in shake flask and bioreactorconditions. This report concludes that recombinant K.
pneumoniae mutant strains have a potential for the co-production of both compounds at a high titer and yield.
Materials and methods
Materials
The genomic DNA isolation kit and pGEM-T vector werepurchased from Promega (Madison, WI, USA). The high-fidelity pfx polymerase was purchased from Invitrogen(Seoul, Korea). The restriction and DNA-modifyingenzymes and the pUC19 plasmid were obtained from NewEngland Bio-Labs (Beverly, MA, USA). Miniprep and DNAgel extraction kits were purchased from Qiagen (Mannheim,Germany). Primers were synthesized by Cosmotech Co. Ltd.(Seoul, Korea). 3-HP was purchased from Tokyo KaseiKogyo Co. Ltd, Tokyo, Japan (TCI America). Since 3-HPAis not available commercially, it was synthesized chemicallyfrom acrolein using the method described by Hall and Stern(1950). Yeast extract (Cat. 212750) was obtained from Difco(Becton Dickinson; Franklin Lakes, NJ, USA). Glycerol andall other chemicals and enzymes, unless otherwise indicated,were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cloning and overexpression of puuC gene in K.pneumoniae
The bacterial strains, plasmids, and primers used in this studyare presented in Table 1. E. coli DH5 alpha was used forplasmid cloning and maintenance, while K. pneumoniaeserved as host for protein expression. LB medium was usedfor the routine genetic engineering and culture maintenancework. Kanamycin at 50 μg/mL was added to the culturemedium where indicated. Since K. pneumoniae is resistant toampicillin, the original version of the pUC19 plasmid cannotbe used for screening and plasmid maintenance. Therefore,the gene sequence carrying kanamycin resistance wasamplified from the pKD4 plasmid, which was obtained fromthe Coli Genetic Stock Center, USA, and cloned between theHindIII and NdeI restriction sites of the pUC19 plasmid toobtain a modified version of the pUC19 plasmid. The puuCgene (GenBank: YP_001334683), encoding an NAD+-dependent gamma-glutamyl-gamma-aminobutyraldehydedehydrogenase, was amplified from the genomic DNA ofK. pneumoniae DSM 2026 using the primers given inTable 1 and cloned in the pUC19 vector by replacing β-galactosidase (lacZ alpha) gene to be transcribed by the lacZpromoter. The resulting plasmid is referred to as pLB0066.Details about the construction of the plasmid are shown inFig. 2. To develop the recombinant which can co-produce 3-HP and 1,3-PDO, the plasmid pLB0066 was transferred intoK. pneumoniae DSM 2026 by the electroporation method at
1254 Appl Microbiol Biotechnol (2011) 90:1253–1265

18 kV/cm with 25 μF and 200 Ω (Severine et al. 1995).Immediately after electroporation, 1 mL of Brain HeartInfusion medium was added to the cuvette, and the cellswere incubated for 1 h at 37°C and 200 rpm in an orbitalincubator shaker (VS-8480 SF, Vision Scientific, Co. Ltd.Korea) before plating. The resulting transformant is referredto as KpPuuC, which was resistant to kanamycin.
Development of K. pneumoniae ΔdhaT mutant strain
The dhaT gene encodes an NADH-dependent 1,3-propane-diol oxidoreductase, which converts 3-HPA to 1,3-PDO. Toachieve a balanced production of 3-HP and 1,3-PDO, thegene dhaT was mutated. To develop the K. pneumoniaeΔdhaT mutant, an in-frame tagged deletion approach was
ATP
DHAP
G-3-P
Glycerol
DHA
dhaKLM
PEP
Pyr
QH2
glpKATP/PEP
ADP/Pyr
3-HPA
NADPH-HOR
Co-B12
puuC
H2O
Glycerol
glpD
QH2
glpABC
Pyruvate Lactate
PEP
NAD+
NADH
ATP
2,3-butanediol
CO2
NAD+
NADH
NAD+
NADH
ATPCO2
mgsA
gloA, gloB
lldD, ldhA, dldpflBybiW
GSH
GSH
pykA,pyF
ppC
dhaB
dhaTdhaD
gldA
Methylglyoxal
CO2H2
FHL
Acetyl-CoA Ethanol
FormateADP+Pi
ATP
TCA
adhE, adhP
CoASH
CO2
PDHC
budB, budC
Acetate
2 NADH
2 NAD
poxBQH2+CO2
3 NADH, FADH, ATP, 2 CO2
Succinate
2 NADH
2 NAD+
mdH, fumABC,frdABCD
OAA
dhaK
NAD+
NADHNAD+
NADH NADP+
NADPH
3-HP
1,3-PDO
ATP
ADP
Biomass
F
NA
DH
-HO
R
NAD+
NADH
Fig. 1 The major glycerol-utilizing metabolic pathways of recombi-nant Klebsiella pneumoniae strain(s) under microaerobic condition.The genes encoding relevant enzymes are given in italics: dhaBglycerol dehydratase; dhaT 1,3-propanediol oxidoreductase; NADH-HOR NADH-dependent hypothetical oxidoreductase (indicated ingray arrow, physiological function not clear); NADPH-HORNADPH-dependent hypothetical oxidoreductase; puuC gamma-glutamyl-gamma-aminobutyraldehyde dehydrogenase; dhaD glyceroldehydrogenase; gldA putative glycerol dehydrogenase; dhaK dihy-droxyacetone kinase I; dhaKLM dihydroxyacetone kinase II; glpKglycerol kinase; glpD glycerol-3-phosphate dehydrogenase (aerobic);glpABC glycerol-3-phosphate dehydrogenase (anaerobic); msgA,methylglyoxal synthase; gloA glyoxalse I; glob hydroxyacylgluta-
thione hydrolase; ppC phosphoenolpyruvate carboxylase; mdH malatedehydrogenase; fumABC fumarase (aerobic and anaerobic); frdABCDfumarate reductase; pykA pyruvate kinase; pykF pyruvate kinase; budBacetolactate synthease; budC acetoin reductase; pflB pyruvate formatelyase; ybiW putatative formate acetyltransferanse; PDHC pyruvatedehydrogenase complex; lldD L-lactate dehydrogenase (reversible);ldhA and dld D-lactate dehydrogenases; poxB pyruvate dehydroge-nase; pta phosphoacetyl transferase; eutD phosphotransacetylase;ackA acetate kinase; adhE bifunctional alcohol dehydrogenase; adhPputative alcohol dehydrogenase. The genetic modification is illustratedby the thick arrow, and the broken lines indicate the multiple steps inthe native metabolic pathways
Appl Microbiol Biotechnol (2011) 90:1253–1265 1255

carried out according to the method described by Link et al.(1997), with a slight modification. Briefly, PCR amplifica-tion of ∼500 bp of upstream (fragment A) and ∼500 bp ofdownstream (fragment B) regions of dhaT gene was carried
out using the primers given in Table 1. These twofragments were ligated using overlapping PCR methods togenerate an engineered deletion fraction AB, and thisfraction was cloned into the pGEMT vector. The
Table 1 Bacterial strains, plasmids, and primers used in this study
Strain and plasmid Description Source
Strains
K. pneumoniae DSMZ 2026 (GSB0001) Source for puuC gene and expression host DSM, Germany
E. coli DH5 alpha (GSB0005) Cloning host KCCM, Korea
KpPuuC (GSB0098) K. pneumoniae DSMZ 2026 harboring puuCon the pUC19 plasmid
This study
Kp ΔdhaT (GSB0162) K. pneumoniae DSMZ 2026 ΔdhaT mutant strain This study
KpC ΔdhaT (GSB0163) K. pneumoniae DSMZ 2026 ΔdhaT mutant strainharboring puuC on the pUC19 plasmid
This study
Plasmids
pLB0065 pUC19—lacZα; cloning vector; Ampr New England Biolabs
pLB0066 pUC19—lac promoter; puuC; Kmr This study
pLB0089 pGEMT//ΔdhaT—plasmid with ΔdhaT deletionfragment for sequence confirmation
This study
pLB0100 pKOV/ΔdhaT—plasmid used to developK. pneumoniae DSMZ 2026 ΔdhaT mutant strain
This study
pLB0102 pKOV—temperature sensitive pSC101; Sac B; Cmr Harvard University
pLB0111 pKD4—template for kanamycin-resistant cassette CGSCa
Primers (forward, F; reverse, R) Sequence (5′ to 3′) Restriction enzymes
puuC, F CAC ACA GGA AAC AGC TAT GAA TTT TCA GCA CC
puuC, R AAG CTT TCA AGA CTC CAG GGC AAT CCA GAT HindIII
Lac promoter, F CTG GCA CGA CAG GTT TCC CGA CTG G
Lac promoter, R GGT GCT GAA AAT TCA TAG CTG TTT CCT GTG TG
Kmr, F AAG CTT TCA CGC TGC CGC AAG C HindIII
Kmr, R CAT ATG GGG GTG GGC GAA GAA CTC CAG C NdeI
Upstream region of dhaT, F GGA TCC GGT GAC AAA ATA ACG CTG AC BamHI
Upstream region of dhaT, R CGT TGC CTT TAC GCG GGT TTA AGT TAA CGT TTG GCA CC
Downstream region of dhaT, F GGT GCC AAA CGT TAA CTT AAA CCC GCG TAA AGG CAA CG
Downstream region of dhaT, R GCG GCC GCG TAG CAG ACG CCC AGT TG NotI
a Coli Genetic Stock Center, USA
KmRP
0.79 kb
Hind III Nde I
Chromosomal DNA from Klebsiella pneumoniae DSM 2026
pKD4 pKD4
pUC19
1.31 kb
KmRPLac Z
puuC
PCR
1.48 kb
pUC19
Ligation
puuC
Ligation
PCR
Fig. 2 Construction of the plas-mid pUC19/puuC (pLB0066).The gene puuC was amplifiedfrom the chromosomal DNA ofKlebsiella pneumoniae DSMZ2026 and cloned into the pUC19vector by replacing the β-galactosidase (lacZ alpha) genedownstream of the lacZpromoter. The gene sequence forkanamycin resistance (KmR)was amplified from pKD4plasmid and cloned betweenHindIII and NdeI restrictionsites of the pUC19 plasmid
1256 Appl Microbiol Biotechnol (2011) 90:1253–1265

sequence-confirmed AB fraction was sub-cloned into thepKOV vector between the BamHI and NotI restriction sitesto develop the pKOV ΔdhaT plasmid; this plasmid wasused to knock out the dhaT gene from the chromosomalDNA of K. pneumoniae by double recombination. ThedhaT mutant strain was screened by PCR and confirmed bysequencing. To develop the recombinant ΔdhaT which canco-produce 3-HP and 1,3-PDO, the plasmid pLB0066 wastransformed into K. pneumoniae ΔdhaT, and the resultingmutant is referred to as KpCΔdhaT.
PuuC expression and gel electrophoresis
The recombinant strains harboring the gene puuC on thepUC19 plasmid were grown in LB medium with kanamy-cin at 50 μg/mL, and the cells were induced at ∼0.6 OD600
with 1.0 mM isopropyl β-D-1-thiogalactopyranoside(IPTG). The cells were harvested 3 h after induction andcentrifuged at 15,000×g for 5 min. The cell pellet waswashed twice with 100 mM potassium phosphate buffer(pH 7.0) and resuspended in the same buffer. The cells weredisrupted using a French Pressure Cell (FA-078A, ThermoElectron Corp.; Waltham, MA) at 1,250 psi. The cell lysatewas centrifuged at 30,000×g for 30 min, and the superna-tant was used for SDS-PAGE and enzyme activity analysis.Protein expression was examined by subjecting the cell-freesoluble extract to SDS-PAGE under denaturing conditions(Laemmli 1970). Coomassie Brilliant Blue was used tostain the proteins. The recombinant protein band intensitieson SDS-PAGE were determined using Scion image analyz-ing software (Scion Corp., Frederick, MD).
Determination of enzyme activities
DhaB activity in the crude cell extract was determinedaccording to the method previously described by Raj et al.(2008). DhaD and DhaT activities were determined accord-ing to the methods described by Ahrens et al. (1998). TheDhaT activity was measured using the reverse reaction(NAD+-dependent conversion of 1,3-PDO to 3-HPA), andthe physiological activity was determined using theconversion factor 0.83/0.21=3.95, according to Ahrens etal. (1998). NADPH-HOR activity was determined by themethod described by Perez et al. (2008). PuuC activity wasdetermined by measuring the reduction of NAD+ to NADHat 340 nm using the method described by Raj et al. (2010)with slight modifications. Briefly, the reaction mixturecontaining 100 mM potassium phosphate buffer (pH 8.0),1 mM DTT, and an appropriate amount of enzyme extractwas incubated at 37°C for 5 min, and the reaction wasinitiated by the addition of 2.0 mM 3-HPA and 2.0 mMNAD+. The amount of NADH formed was determined usinga molar extinction coefficient (Δε340) of 6.22×10
3 M−1cm−1.
One unit of PuuC activity was defined as the amount ofenzyme required to reduce 1 μmol of NAD+ to NADH in1 min. All enzyme activities were measured with crude cellextracts, which were prepared from the cells grown undermicroaerobic conditions (see below for the culture con-ditions) and actively producing 3-HP and/or 1,3-PDO.
Shake flask cultivation of recombinant KpPuuCand KpCΔdhaT
All K. pneumoniae strains were cultured in 250 mL non-baffled Erlenmeyer flasks with 50 mL working volume at37°C in an orbital incubator shaker. M9 medium was used,which contained the following components per liter ofdeionized water: MgSO4·7H2O, 0.25 g; NaCl, 1.0 g;NH4Cl, 1.0 g; yeast extract, 1.0 g; and glycerol, 100 mmol.The medium was supplemented with 100 mM potassiumphosphate buffer (pH 7.0) and kanamycin at 50 mg l−1.Three different culture conditions—aerobic, microaerobic,and anaerobic—were used to examine the performance ofthe recombinant strains in shake flasks by varying theagitation speed in the orbital incubator shaker. For aerobicand microaerobic conditions, the flasks were plugged withan oxygen-permeable, silicone-foam stopper. For anaerobiccultures, a screw cap was used, and the head space of theflask was replaced with argon gas before cultivation.Agitation speed was 200 rpm for aerobic culture, 30 rpmfor microaerobic culture, and 100 rpm for anaerobic culture.The cells were induced at 0.6±0.05 OD600 with 1.0 mMIPTG. Samples were withdrawn periodically to determinethe cell mass, residual substrate, and metabolites.
Bioreactor cultivation of recombinant KpCΔdhaT
The recombinant KpCΔdhaT (GSB0163) was cultured in a 5-l capacity Biotron-LiFlus GM bioreactor (Biotron, Seoul,Korea) with a 1-l initial working volume. The same M9medium as in the flask cultures, containing 100 mM glycerol,100 mM potassium phosphate buffer (pH 7.0), and 50 mg L−1
kanamycin, was used. The bioreactor cultivation was carriedout at 37°C in a fed-batch mode by feeding concentratedglycerol (10 M) at 48.6 mM h−1 from 6 to 24 h. The pH wasmaintained at 7.0±0.1 with 2.5 N NaOH and 2.5 N HCl. Thecells were induced at an OD600 of 0.69 with 1.0 mM IPTG.The agitation speed was set at 150 rpm with 1.0 VVM air, andDO was maintained <0.2%, except for the initial ∼2 h.Samples were withdrawn periodically to determine cell mass,residual substrate, and metabolites concentration.
Analytical methods
Cell concentrations were measured in a 10-mm-path-lengthcuvette using a double-beam spectrophotometer (Lambda
Appl Microbiol Biotechnol (2011) 90:1253–1265 1257
20, PerkinElmer; Norwalk, CT, USA). One unit ofabsorbance at 600 nm corresponded to 0.31 g dried cellmass per liter. Protein concentrations in the cell-free extractwere determined by the method of (Bradford 1976) on amicrotiter plate reader (1420, Wallac Victor2; PerkinElmer)using bovine serum albumin as a standard. Concentrationsof glycerol, 3-HP, and other organic acids were determinedby HPLC using the method described by Raj et al. (2008)with a slight modification. Briefly, the supernatants,obtained by centrifugation of the culture samples at10,000×g for 10 min, were filtered through the Tuffryn-membrane (Acrodisc, Pall Life Sciences) and elutedthrough a 300×7.8 mm Aminex HPX-87H (Bio-Rad,USA) column at 65°C using 1.0 mM H2SO4.
Results
Protein expression and enzyme activities of wild-typeand recombinant K. pneumoniae strains
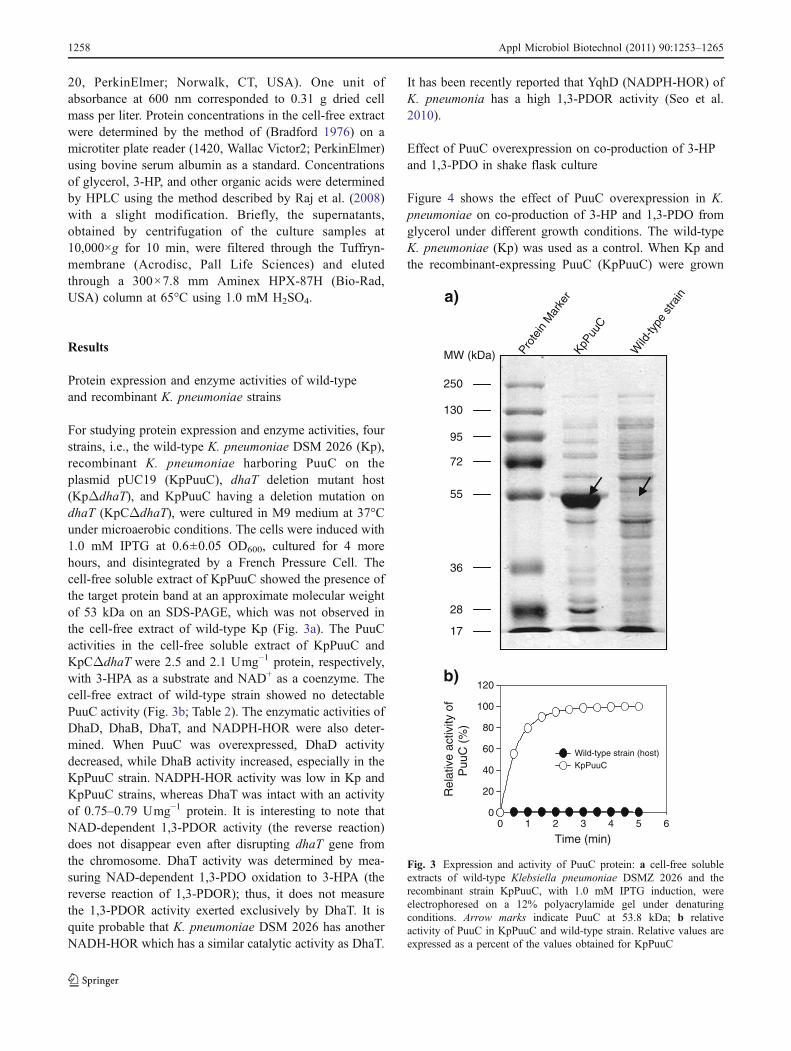
For studying protein expression and enzyme activities, fourstrains, i.e., the wild-type K. pneumoniae DSM 2026 (Kp),recombinant K. pneumoniae harboring PuuC on theplasmid pUC19 (KpPuuC), dhaT deletion mutant host(KpΔdhaT), and KpPuuC having a deletion mutation ondhaT (KpCΔdhaT), were cultured in M9 medium at 37°Cunder microaerobic conditions. The cells were induced with1.0 mM IPTG at 0.6±0.05 OD600, cultured for 4 morehours, and disintegrated by a French Pressure Cell. Thecell-free soluble extract of KpPuuC showed the presence ofthe target protein band at an approximate molecular weightof 53 kDa on an SDS-PAGE, which was not observed inthe cell-free extract of wild-type Kp (Fig. 3a). The PuuCactivities in the cell-free soluble extract of KpPuuC andKpCΔdhaT were 2.5 and 2.1 Umg−1 protein, respectively,with 3-HPA as a substrate and NAD+ as a coenzyme. Thecell-free extract of wild-type strain showed no detectablePuuC activity (Fig. 3b; Table 2). The enzymatic activities ofDhaD, DhaB, DhaT, and NADPH-HOR were also deter-mined. When PuuC was overexpressed, DhaD activitydecreased, while DhaB activity increased, especially in theKpPuuC strain. NADPH-HOR activity was low in Kp andKpPuuC strains, whereas DhaT was intact with an activityof 0.75–0.79 Umg−1 protein. It is interesting to note thatNAD-dependent 1,3-PDOR activity (the reverse reaction)does not disappear even after disrupting dhaT gene fromthe chromosome. DhaT activity was determined by mea-suring NAD-dependent 1,3-PDO oxidation to 3-HPA (thereverse reaction of 1,3-PDOR); thus, it does not measurethe 1,3-PDOR activity exerted exclusively by DhaT. It isquite probable that K. pneumoniae DSM 2026 has anotherNADH-HOR which has a similar catalytic activity as DhaT.
It has been recently reported that YqhD (NADPH-HOR) ofK. pneumonia has a high 1,3-PDOR activity (Seo et al.2010).
Effect of PuuC overexpression on co-production of 3-HPand 1,3-PDO in shake flask culture
Figure 4 shows the effect of PuuC overexpression in K.pneumoniae on co-production of 3-HP and 1,3-PDO fromglycerol under different growth conditions. The wild-typeK. pneumoniae (Kp) was used as a control. When Kp andthe recombinant-expressing PuuC (KpPuuC) were grown
Time (min)0 1 2 3 4 5 6
Rel
ativ
e ac
tivity
of
Puu
C (
%)
0
20
40
60
80
100
120
Wild-type strain (host)KpPuuC
250
130
95
72
55
36
28
17
KpPu
uC
Prot
ein
Mar
kera)
Wild
-type
stra
in
MW (kDa)
b)
Fig. 3 Expression and activity of PuuC protein: a cell-free solubleextracts of wild-type Klebsiella pneumoniae DSMZ 2026 and therecombinant strain KpPuuC, with 1.0 mM IPTG induction, wereelectrophoresed on a 12% polyacrylamide gel under denaturingconditions. Arrow marks indicate PuuC at 53.8 kDa; b relativeactivity of PuuC in KpPuuC and wild-type strain. Relative values areexpressed as a percent of the values obtained for KpPuuC
1258 Appl Microbiol Biotechnol (2011) 90:1253–1265

under microaerobic growth conditions (Fig. 4a, b), onlyKpPuuC produced a substantial amount of 3-HP, along with1,3-PDO. The final 3-HP titer at 24 h in KpPuuC was13 mM, while that of 1,3-PDO was 23 mM. The over-expression of PuuC affected cell growth and glycerolconsumption. In the KpPuuC strain, cell growth andglycerol consumption were delayed, and their rates were
low as compared to the wild-type counterpart. Themaximum specific growth rate (μmax) of the wild-type Kp,estimated between 0 and 9 h, was 0.57 h−1, while it was0.33 h−1 in the recombinant KpPuuC. The specific glycerolconsumption rate of the wild-type, between 0 and 9 h, wasestimated to be 72 mmol g−1 cdw h−1, but it was52 mmol g−1 cdw h−1 when PuuC was overexpressed.
Table 2 DhaD, DhaB, PuuC, DhaT/NADH-HOR, and NADPH-HOR activities in wild-type Kp, KpPuuC, KpΔdhaT, and KpCΔdhaT crude cellextracts
Strain Specific activity (U/mg protein)
DhaDa DhaBb PuuCc DhaT/NADH-HORd NADPH-HORe
Wild-type Kp 5.9 6.8 0 0.75 0.07
KpPuuC 3.4 20.8 2.5 0.79 0.03
KpΔdhaT 5.2 8.7 0 0.28 0.22
KpCΔdhaT 3.7 10.2 2.1 0.71 0.17
a DhaD (glycerol dehydrogenase) activity was determined using glycerol as a substrate and NAD+ as a cofactor, according to Ahrens et al. (1998)b DhaB (glycerol dehydratase) activity was determined using glycerol as a substrate and coenzyme B12, according to Raj et al. (2008)c PuuC (gamma-glutamyl-gamma-aminobutyraldehyde dehydrogenase) activity was determined using 3-HPA and NAD+ as substrate and cofactor,respectively. The activity indicated here is the forward reaction (NAD+ -dependent oxidation of 3-HPA to 3-HP), according to Raj et al. (2008)d For determining DhaT or NADH-HOR (NADH-dependent hypothetical oxidoreductase) activities, 1,3-PDO and NAD+ were used as substrate andcofactor, respectively. The activity indicated here is the reverse reaction (NAD+ -dependent oxidation of 1,3-PDO to 3-HPA) of DhaT or NADH-HOR,according to Ahrens et al. (1998)e For determining NADPH-HOR (NADPH-dependent hypothetical oxidoreductase) activity, 3-HPA and NADPH were used as substrate and cofactor,respectively. The activity indicated here is the forward reaction (NADPH-dependent reduction of 3-HPA to 1,3-PDO) of NADPH-HOR, according to Perezet al. (2008)
Time (h)
0 5 10 15 20 25
Gly
cero
l (m
mol
l-1 )
Gly
cero
l (m
mol
l-1 )
0
20
40
60
80
100
Cel
l dry
mas
s (g
l-1)
0.00
0.50
1.00
1.50
2.00
CD
W (
gl-1
)
0.25
0.75
1.25
1.75
Time (h)
0 5 10 15 20 253-
HP
& 1
, 3-P
DO
(m
mol
l-1 )
3-H
P &
1, 3
-PD
O (
mm
ol l-
1 )0
5
10
15
20
25
30a) b)
Time (h)
0 5 10 15 20 25
0
20
40
60
80
100
Cel
l dry
mas
s (g
l-1)
0.0
0.5
1.0
1.5
2.0
Time (h)
0 5 10 15 20 25
0
5
10
15
20
25
30c) d)
Fig. 4 Time-course profile ofwild-type Kp strain and therecombinant KpPuuC on cellgrowth, glycerol consumption,and co-production of 3-HP and1,3-PDO under microaerobicconditions are indicated as a andb, respectively. Similarly, thetime-course profile of recombi-nant KpPuuC under aerobic andanaerobic conditions are pre-sented in c and d, respectively.Symbols: semi-filled circle celldry mass, close triangle residualglycerol, close circle 3-HP, opencircle 1,3-PDO
Appl Microbiol Biotechnol (2011) 90:1253–1265 1259

However, final cell density was almost the same as 0.71±0.01 gL−1.
To further explore the potential of the KpPuuC strain, thestrain was cultured under aerobic (Fig. 4c) and anaerobic(Fig. 4d) conditions. 3-HP production was not high undereither condition, but the production of 1,3-PDO was as highas 26 mM under anaerobic conditions (Fig. 4d), which wasabout the same as that under microaerobic conditions.Although cell growth was fast under aerobic conditions, theproduction of both 3-HP and 1-3-PDO was very low.Similar experiments were also carried out with a wild-typeKp strain under aerobic and anaerobic conditions (data notshown). The fermentation profiles were almost the same asthose of the recombinant counterpart, except that the lagperiod in cell growth was not observed, and the time toreach the termination of fermentation was shortened by 3 to4 h, accordingly. These results indicate that, when PuuC isoverexpressed, K. pneumoniae can efficiently produce 3-HP along with 1,3-PDO under microaerobic condition.However, 1,3-PDO production was more as expected.
Therefore, the residual metabolites and carbon materialbalance were analyzed for microaerobic culture samplestaken at 24 h, and the results are shown in Table 3. Ethanol,lactate, and acetate were major by-products for both strains.Ethanol was produced at 46.5 mM in the Kp strain, and itwas decreased to 27.0 mM in the KpPuuC strain. On theother hand, when PuuC was overexpressed, acetate andlactate were increased to 12.2 from 6.4 mM and 18.2 from
4.0 mM, respectively. Carbon metabolite analysis showed acarbon recovery of ∼72.5% for the wild-type strain and>95% for the KpPuuC strain. The loss of carbon in thecarbon balance was attributed to the evolution of CO2,which was not measured in this study.
Effect of dhaT deletion on co-production of 3-HP and 1,3-PDO in shake flask culture
The strain KpPuuC co-produced 3-HP and 1,3-PDO.However, the production of 1,3-PDO was higher than thatof 3-HP due to higher carbon flux toward 1,3-PDO at the 3-HPA node (Fig. 4b). The wild-type K. pneumoniae strain isknown to have at least two 1,3-PDOR isozymes, DhaT andYqhD (NADPH-HOR). In order to produce both com-pounds in an equal ratio, the dhaT gene was deleted fromthe chromosome of K. pneumoniae while leaving yqhDintact. Figure 5 shows the performance of the recombinantstrain KpCΔdhaT on co-production of 3-HP and 1,3-PDOfrom glycerol under different culture conditions. StrainKpΔdhaT, the host strain without PuuC, was used as acontrol. As shown in Fig. 5a, under microaerobic con-ditions, the KpΔdhaT strain produced 1,3-PDO even afterdisrupting the gene dhaT, indicating the presence of other1,3-PDOR-like enzymes. When PuuC was overexpressed inthe dhaT mutant strain, the recombinant strain KpCΔdhaTproduced both compounds in equal amounts. The titers ofboth compounds were as high as 40 mM (Fig. 5b), which is
Table 3 Carbon distribution for the microaerobic metabolism of glycerol by the wild-type Kp and recombinant strains in shake flask conditions
Substrate and metabolites Wild-type Kp (GSB0001) KpPuuC (GSB0162) KpCΔdhaT (GSB0163)
mmol l−1 C-mmol l−1 C (%) mmol l−1 C-mmol l−1 C (%) mmol l−1 C-mmol l−1 C (%)
Substrate
Glycerol 93.18 279.54 100.00 93.89 281.67 100.00 99.79 299.25 100.00
Biomass 0.68a 28.02b 10.02 0.67a 27.66b 9.82 0.46a 18.56b 6.20
Metabolites
1,3-PDO 18.69 56.07 20.06 23.24 69.72 24.75 40.47 121.41 40.57
3-HP 0.80 2.40 0.86 12.77 38.31 13.60 40.02 120.06 40.12
Acetate 6.43 12.86 4.60 12.17 24.34 8.64 5.83 11.66 3.90
Lactate 3.95 11.85 4.24 18.20 54.60 19.38 2.75 8.25 2.76
Ethanol 46.46 92.92 33.24 27.02 54.04 19.19 11.85 23.70 7.92
Total products 204.12 – 268.67 – 303.64 –
Carbon recovery %c 72.47 95.39 101.47
The data correspond to the 24-h sample for the culture shown in Figs. 4 and 5
The calculation did not consider CO2 in the liquid and head space
The standard deviation of the measurements was less than 5% for the substrate and the metabolitesa Grams per literb An average molecular weight of 23.86, which corresponds to an average cell with a molecular formula of CH1.74 O0.44 N0.22. The average ash content of3.6% was deduced from the actual cell dry massc (Total carbon in biomass and metabolites)/(Total carbon in glycerol)×100
1260 Appl Microbiol Biotechnol (2011) 90:1253–1265

∼2-fold higher than the values obtained from its parentstrain KpPuuC.
Figure 5 also shows the results on aerobic and anaerobiccultivation of KpCΔdhaT (Fig. 5c, d). Under bothconditions, the production of 3-HP and 1,3-PDO was nothigh compared to that of microaerobic conditions. It isnoticed that under aerobic conditions, a long lag in cellgrowth and reduced glycerol consumption were observedtill 12 h. Although glycerol consumption was started after12 h, production of both 3-HP and 1,3-PDO was low.Under anaerobic condition, cell growth and production ofboth 3-HP and 1,3-PDO were negligible. Similar experi-ments were also conducted for the host KpΔdhaT strainunder aerobic and anaerobic conditions (data not shown).Except for the fact that a much longer lag period wasobserved, the overall cultivation trends were similar tothose of the wild-type Kp strain that were previouslyexplained.
The carbon distribution for KpCΔdhaT was analyzed at24 h (Table 3). In KpCΔdhaT, glycerol assimilationthrough DhaB was greatly improved as compared to Kpor KpPuuC strains; almost 81% of glycerol was directedthrough DhaB. In comparison, glycerol assimilation via theoxidative pathway was only 19.3%, which includes theamount of carbon to cell mass (6.2%). Ethanol, acetate, andlactate were major by-products of glycerol fermentation, asin the Kp and KpPuuC strains. However, their concen-
trations were significantly decreased. Also, it was noticedthat the carbon recovery with KpCΔdhaT is ∼100%,indicating that the production of CO2 is negligible.
Bioreactor cultivation of the strain KpCΔdhaT
In flask experiments, the strain KpCΔdhaT exhibited thehighest 3-HP and 1,3-PDO yield, 81% on glycerol carbon.However, at the end of 24 h of cultivation, glycerol wasdepleted and the culture pH decreased below 6.0. In orderto further investigate the potential of KpCΔdhaT, the strainwas cultured at pH 7.0±0.1 in a 5-l bioreactor with aworking volume of 1 l (Fig. 6). The initial glycerolconcentration was 100 mM, and concentrated glycerol(10 M) was added at 48.6 mmol−1 L h−1 between 6 and24 h. In 24 h of the bioreactor culture, KpCΔdhaTproduced 178 mmol L−1 3-HP and 221 mmol L−1 1,3-PDO, which are about 5.5-fold and 4.5-fold higher,respectively, than the titers obtained from the flask culture.The biomass reached a maximum of 1.58 gL−1 at 18 h butdecreased to 1.07 gL−1 at 24 h. Total glycerol consumptionat 24 h was 783 mmol L−1. The μmax, estimated between 0and 6 h was 0.58 h−1. The qmax of 1,3-PDO and 3-HP,estimated between 3 and 24 h, were 26.4 and21.2 mmol g−1 cdw h−1, respectively.
Figure 6b shows the time-course profile of metaboliteproduction. Unlike flask results, lactate was observed to be the
Time (h)
0
20
40
60
80
100
0.0
0.5
1.0
1.5
2.0
Time (h)0 5 10 15 20 250 5 10 15 20 25
0 5 10 15 20 250 5 10 15 20 25
3-H
P &
1,3
-PD
O (
mm
ol l-
1 )
0
10
20
30
40
(C)
Time (h)
Gly
cero
l (m
mol
l-1 )
Gly
cero
l (m
mol
l-1 )
0
20
40
60
80
100
Cel
l dry
mas
s (g
l-1 )
Cel
l dry
mas
s (g
l-1 )
0.0
0.5
1.0
1.5
2.0a)
Time (h)
3-H
P &
1,3
-PD
O (
mm
ol l-
1 )
0
10
20
30
40b)
d)c)
Fig. 5 Time-course profile ofKpΔdhaT and the recombinantKpCΔdhaT mutants on cellgrowth, glycerol consumption,and co-production of 3-HP and1,3-PDO under microaerobicgrowth conditions are indicatedas a and b, respectively.Similarly, the time-courseprofile of recombinantKpCΔdhaT under aerobic andanaerobic conditions arepresented as c and d, respec-tively. Symbols: semi-filledcircle cell dry mass, closetriangle residual glycerol, closecircle 3-HP, and open circle1,3-PDO
Appl Microbiol Biotechnol (2011) 90:1253–1265 1261
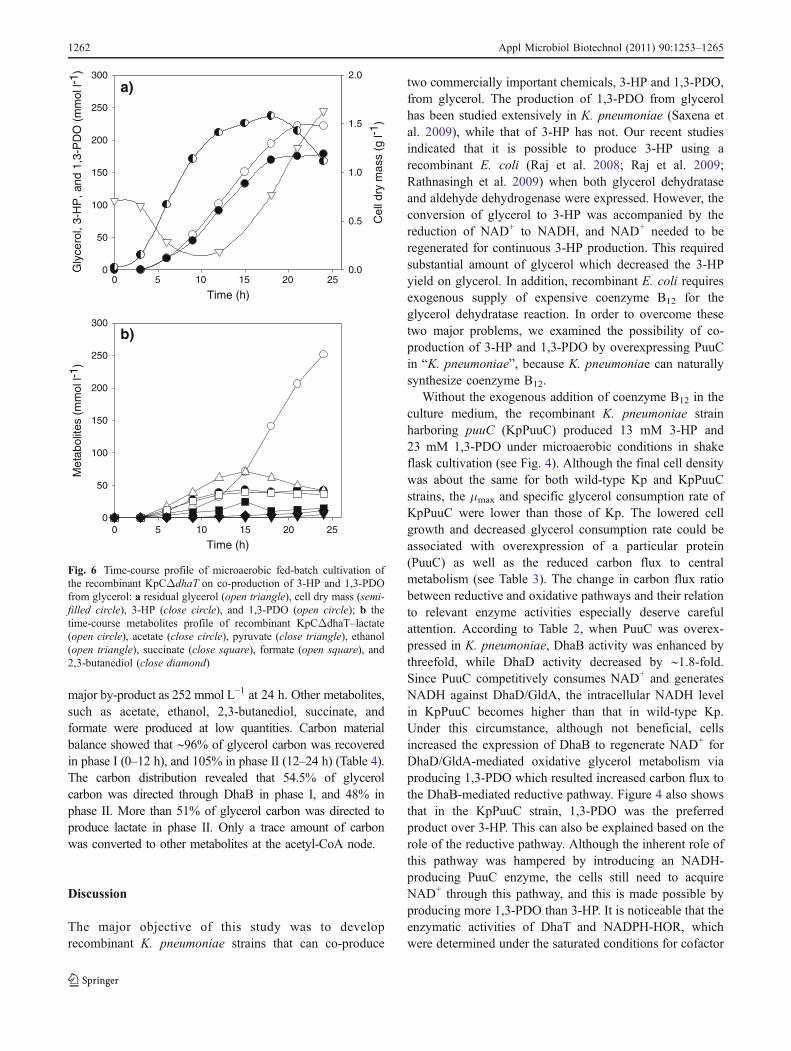
major by-product as 252 mmol L−1 at 24 h. Other metabolites,such as acetate, ethanol, 2,3-butanediol, succinate, andformate were produced at low quantities. Carbon materialbalance showed that ∼96% of glycerol carbon was recoveredin phase I (0–12 h), and 105% in phase II (12–24 h) (Table 4).The carbon distribution revealed that 54.5% of glycerolcarbon was directed through DhaB in phase I, and 48% inphase II. More than 51% of glycerol carbon was directed toproduce lactate in phase II. Only a trace amount of carbonwas converted to other metabolites at the acetyl-CoA node.
Discussion
The major objective of this study was to developrecombinant K. pneumoniae strains that can co-produce
two commercially important chemicals, 3-HP and 1,3-PDO,from glycerol. The production of 1,3-PDO from glycerolhas been studied extensively in K. pneumoniae (Saxena etal. 2009), while that of 3-HP has not. Our recent studiesindicated that it is possible to produce 3-HP using arecombinant E. coli (Raj et al. 2008; Raj et al. 2009;Rathnasingh et al. 2009) when both glycerol dehydrataseand aldehyde dehydrogenase were expressed. However, theconversion of glycerol to 3-HP was accompanied by thereduction of NAD+ to NADH, and NAD+ needed to beregenerated for continuous 3-HP production. This requiredsubstantial amount of glycerol which decreased the 3-HPyield on glycerol. In addition, recombinant E. coli requiresexogenous supply of expensive coenzyme B12 for theglycerol dehydratase reaction. In order to overcome thesetwo major problems, we examined the possibility of co-production of 3-HP and 1,3-PDO by overexpressing PuuCin “K. pneumoniae”, because K. pneumoniae can naturallysynthesize coenzyme B12.
Without the exogenous addition of coenzyme B12 in theculture medium, the recombinant K. pneumoniae strainharboring puuC (KpPuuC) produced 13 mM 3-HP and23 mM 1,3-PDO under microaerobic conditions in shakeflask cultivation (see Fig. 4). Although the final cell densitywas about the same for both wild-type Kp and KpPuuCstrains, the μmax and specific glycerol consumption rate ofKpPuuC were lower than those of Kp. The lowered cellgrowth and decreased glycerol consumption rate could beassociated with overexpression of a particular protein(PuuC) as well as the reduced carbon flux to centralmetabolism (see Table 3). The change in carbon flux ratiobetween reductive and oxidative pathways and their relationto relevant enzyme activities especially deserve carefulattention. According to Table 2, when PuuC was overex-pressed in K. pneumoniae, DhaB activity was enhanced bythreefold, while DhaD activity decreased by ∼1.8-fold.Since PuuC competitively consumes NAD+ and generatesNADH against DhaD/GldA, the intracellular NADH levelin KpPuuC becomes higher than that in wild-type Kp.Under this circumstance, although not beneficial, cellsincreased the expression of DhaB to regenerate NAD+ forDhaD/GldA-mediated oxidative glycerol metabolism viaproducing 1,3-PDO which resulted increased carbon flux tothe DhaB-mediated reductive pathway. Figure 4 also showsthat in the KpPuuC strain, 1,3-PDO was the preferredproduct over 3-HP. This can also be explained based on therole of the reductive pathway. Although the inherent role ofthis pathway was hampered by introducing an NADH-producing PuuC enzyme, the cells still need to acquireNAD+ through this pathway, and this is made possible byproducing more 1,3-PDO than 3-HP. It is noticeable that theenzymatic activities of DhaT and NADPH-HOR, whichwere determined under the saturated conditions for cofactor
Time (h)
Met
abol
ites
(mm
ol l-
1 )
0
50
100
150
200
250
300
Time (h)0 5 10 15 20 25
0 5 10 15 20 25
Gly
cero
l, 3-
HP
, and
1,3
-PD
O (
mm
ol l-
1 )
0
50
100
150
200
250
300
Cel
l dry
mas
s (g
l-1 )
0.0
0.5
1.0
1.5
2.0a)
b)
Fig. 6 Time-course profile of microaerobic fed-batch cultivation ofthe recombinant KpCΔdhaT on co-production of 3-HP and 1,3-PDOfrom glycerol: a residual glycerol (open triangle), cell dry mass (semi-filled circle), 3-HP (close circle), and 1,3-PDO (open circle); b thetime-course metabolites profile of recombinant KpCΔdhaT–lactate(open circle), acetate (close circle), pyruvate (close triangle), ethanol(open triangle), succinate (close square), formate (open square), and2,3-butanediol (close diamond)
1262 Appl Microbiol Biotechnol (2011) 90:1253–1265

and substrate, are relatively lower than that of PuuC. Thisindicates that the physiological concentrations of cofactorsand substrates may have been far from saturation levels,and the in vivo reaction rates may have not beenproportional to the enzyme activities determined in vitro.
Since more 1,3-PDO was produced by the KpPuuCstrain, it would be natural to decrease the 1,3-PDORactivity in order to obtain balanced production of 3-HPand 1,3-PDO. Two 1,3-PDORs, DhaT and NADPH-HOR(YqhD), have been well documented thus far, and either onecould be disrupted (Emptage et al. 2001; Wang et al. 2003;Seo et al. 2010). In this study, dhaT, rather than the gene forNADPH-HOR, was disrupted, since the physiological roleof DhaT is directly linked to NAD+ regeneration for theoxidative pathway and thus more 1,3-PDO is expected tobe produced if dhaT is left intact in the puuC over-expressing strain. As shown in Fig. 5b, equal amounts of 3-HP and 1,3-PDO were obtained when disrupting dhaT. Inaddition, in KpCΔdhaT, the overall carbon flux towards 3-HP and 1,3-PDO was increased significantly. Althoughfurther investigation is required, one possible explanationfor the increased carbon flux through DhaB is that the highcellular NADH level (because of the NAD+-dependentPuuC activity and the lack of DhaT to reoxidize NADH)
inhibits the reaction by NAD+-dependent DhaD that isrequired for the oxidative glycerol metabolism. The failureof the KpCΔdhaT strain to grow under anaerobic conditionsuggests that the strain has a high ATP demand whichcannot be fulfilled under anaerobic condition, since NADH,produced by PuuC and DhaD, should be converted toNADPH which can be utilized by NADPH-HOR.
It is also interesting to note that the dhaT deletionmutants express a high NAD+-dependent 1,3-PDOR activitywhen 1,3-PDO was used as a substrate (reverse reaction). Asshown in Table 2, especially in KpCΔdhaT, the unknownNAD+-dependent 1,3-PDOR activity was 0.71 Umg−1
protein, which is close to the NAD+-dependent 1,3-PDORactivity (reverse reaction) in the wild-type strain expressingDhaT. This suggests that there must be more NAD+-dependent 1,3-PDORs in K. pneumoniae in addition toDhaT, similar to the results observed previously (Wang et al.2003; Seo et al. 2009; Seo et al. 2010). Wang et al. (2003)have reported that the presence of another putative 1,3-PDOR (alkylhydroperoxide dehydratase), in addition toDhaT and HOR, in K. pneumoniae. However, it is not clearwhether the enzyme reported by Wang et al. (2003) is theone expressed in the KpCΔdhaT strain. Also, it is not clearwhich enzyme, either the putative NAD+-dependent 1,3-
Table 4 Carbon distribution for the microaerobic metabolism of glycerol by the recombinant KpCΔdhaT in bioreactor conditions
Substrate and metabolites Molecular formulae Cultivation time
Phase I (0–12 h) Phase II (12–24 h)
mmol l−1 C-mmol l−1 C (%) mmol l−1 C-mmol l−1 C (%)
Substrate
Glycerol C3H5(OH)3 353.59 1,060.77 100.00 429.4 1,288.20 100.00
Biomass CH1.74 O0.44 N0.22 1.36a 55.22b 5.21 0a 0.00b 0.00
Metabolites
1,3-PDO C3H8O2 101.51 304.53 28.71 119.68 359.04 27.87
3-HP C3H6O3 91.12 273.36 25.77 87.1 261.30 20.28
Acetate C2H4O2 38.76 77.52 7.31 3.41 6.82 0.53
Lactate C3H6O3 32.11 96.33 9.08 219.53 658.59 51.12
Ethanol C2H5OH 61.96 123.92 11.68 0 0.00 0.00
2,3-butanediol C4H10O2 1.59 6.36 0.60 9.19 36.76 2.85
Succinate C4H6O4 11.27 45.08 4.25 3.05 12.20 0.95
Formate CH2O2 35.94 35.94 3.39 0.21 0.21 0.02
Pyruvate C3H4O3 0.04 0.12 0.01 5.07 15.21 1.18
Total products 1,018.38 1,350.13
Carbon recovery %c 96.00 104.81
The calculation did not consider CO2 in the liquid and head space
The standard deviation of the measurements was less than 5% for the substrate and the metabolitesa Grams per literb An average molecular weight of 23.86, which corresponds to an average cell with a molecular formula of CH1.74 O0.44 N0.22. The average ash content of3.6% was deduced from the actual cell dry mass (Stephanopoulos et al. 1998)c (Total carbon in biomass and metabolites)/(Total carbon in glycerol)×100
Appl Microbiol Biotechnol (2011) 90:1253–1265 1263

PDOR or NADPH-HOR, played a major role inproducing 1,3-PDO in the KpCΔdhaT strain. Detailedstudies are thus required to understand the role ofadditional putative NAD+-dependent 1,3-PDOR.
Given the success in co-producing 3-HP and 1,3-PDOwithimproved titers and yield, we examined the potential of thestrain KpCΔdhaT under bioreactor conditions. Glycerol wasadded in a fed-batch mode, and pH was maintained at 7.0.The results revealed that this strain had a potential toproduce both compounds ≥16 gL−1 each. However, thecumulative carbon yield on glycerol decreased to 51% due toexcessive production of lactate. Production of large amountsof lactate at 220 mM, especially in the second phase (12–24 h), could be associated with the formation of methyl-glyoxal (MG). DHAP accumulation is known to activatelactate production through MG by GSH-dependent GloAand GloB. MG is a potent electrophile, and its accumulationin subcellular concentrations leads to growth inhibition andcell death due to modification of proteins and DNA(MacLean et al. 1998). Therefore, growth inhibition andproduction of lactate could be associated with accumulationof MG (Ferguson et al. 1998; Booth 2005; http://www.ecosal.org). Low levels of other metabolites from the acetyl-CoA branch could also be linked with the strong inhibitionof pyruvate formate lyase (PFL) by high concentrations ofDHAP (Melchiorsen et al. 2002). The inhibition of PFL,which converts pyruvate to acetyl-CoA and formate underanaerobic conditions, could have also contributed to lactateproduction from pyruvate which disposes of excess reducingequivalents.
In summary, we demonstrated that the strainK. pneumoniaeΔdhaT mutant overexpressing PuuC has a potential to co-produce two commercially valuable products, 3-HP and 1,3-PDO. In flask scale cultivation, both compounds wereproduced at 40 mmol L−1 each, with the cumulative carbonyield of 81% on glycerol under microaerobic conditions.When a fed-batch process was carried out in a bioreactorconditions, the strain KpCΔdhaT produced both 3-HP and1,3-PDO at 16.0±0.5 gL–1 each, with a cumulative carbonyield of 51% on glycerol. In bioreactor cultivation, a majorportion of glycerol was converted to lactate, and the growthwas inhibited after 18 h, presumably due to intracellularaccumulation of DHAP and MG. This suggests that furthermetabolic engineering work is required to regulate themetabolic shift in order to improve the titer and yield of bothproducts for commercial applications.
Acknowledgments This work was financially supported by theKorean Ministry of Commerce, Industry, and Energy (Grant No.10028380-2006-11), and Korean Ministry of Education, Science andTechnology through Advanced Biomass R&D Centre (ABC; GrantNo. 2010-0029799), KAIST, Korea. Dr. S. M. Raj is grateful to theBrain Korea 21 program, Pusan National University for providingfinancial assistance.
References
Ahrens K, Menzel K, Zeng A, Deckwer W (1998) Kinetic, dynamic,and pathway studies of glycerol metabolism by Klebsiellapneumoniae in anaerobic continuous culture: III. Enzymes andfluxes of glycerol dissimilation and 1, 3-propanediol formation.Biotechnol Bioeng 59(5):544–552
Blankschien MD, Clomburg JM, Gonzalez R (2010) Metabolicengineering of Escherichia coli for the production of succinatefrom glycerol. Metab Eng 12(5):409–419
Booth IR (2005) Glycerol and methylglyoxal metabolism. In:Neidhardt FC, Curtiss RIII, Ingraham JL, Lin ECC, Low KB,Magasanik B, Reznikoff WS, Riley M, Schaechter M, UmbargerHE (eds) Escherichia coli and salmonella: cellular and molecularbiology (web edition). ASM, Washington, Available at http://www.ecosal.org
Bradford MM (1976) A rapid and sensitive for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Emptage M, Haynie S, Laffend L, Pucci J, Whited G (2001) Processfor the biological production of 1,3-propanediol with high titer.E. I. du Pont de Nemours and Co. and Genencor International,Inc. U.S. patent WO 01/12833 A2
Ferguson GP, Totemeyer S, MacLean MJ, Booth IR (1998) Methyl-glyoxal production in bacteria: suicide or survival. Arch Micro-biol 170:209–218
Global Renewable Chemicals Market (2009–2014). Available athttp://www.marketsandmarkets.com/Market-Reports/renewable-chemicals-82.html
Hall RH, Stern ES (1950) Acid-catalysed hydration of acrylaldehyde:kinetics of the reaction and isolation of β-hydroxypropionaldehyde.J Chem Soc 490–498. Available at http://www.brdisolutions.com/pdfs/BioProductsOpportunitiesReportFinal.pdf
Jo JE, Raj SM, Rathnasingh C, Selvakumar E, Jung WC, Park SH(2008) Cloning, expression, and characterization of an aldehydedehydrogenase from Escherichia coli K-12 that utilizes 3-hydroxypropionaldehyde as a substrate. Appl Microbiol Bio-technol 81:51–60
Johnson EA, Lin ECC (1987) Klebsiella pneumoniae 1, 3-propanediol: NAD+ oxidoreductase. J Bacteriol 169:2050–2054
Laemmli UK (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227:680–685
Lin EC (1976) Glycerol dissimilation and its regulation in bacteria.Annu Rev Microbiol 23:573–580
Link AJ, Phillips D, Church GM (1997) Methods for generatingprecise deletions and insertions in the genome of wild-typeEscherichia coli: application to open reading frame characteriza-tion. J Bacteriol 179:6228–6237
MacLean MJ, Ness LS, Ferguson GP, Booth IR (1998) The role ofglyoxalase-I in the detoxification of methylglyoxal and in theactivation of the KefB K+ efflux system in Escherichia coli. MolMicrobiol 27(3):563–571
Melchiorsen CR, Jokumsen KV, Villadsen J, Israelsen H, Arnau J(2002) The level of pyruvate-formate lyase controls the shiftfrom homolactic to mixed-acid product formation in Lactococcuslactis. Appl Microbiol Biotechnol 58:338–344
Paster M, Pellegrino JL, Carole TM (2003) Industrial bioproducts:today and tomorrow. US DOE report, July, pp 48–49
Perez JM, Arenas FA, Pradenas GA, Sandoval JM, Vasquez CC (2008)Escherichia coli YqhD exhibits aldehyde reductase activity andprotects from the harmful effect of lipid peroxidation-derivedaldehydes. J Biol Chem 283(12):7346–7353
Raj SM, Rathnasingh C, Jo JE, Park SH (2008) Production of 3-hydroxypropionic acid from glycerol by a novel recombinantEscherichia coli BL21 strain. Process Biochem 43:1440–1446
1264 Appl Microbiol Biotechnol (2011) 90:1253–1265

Raj SM, Rathnasingh C, Jung WC, Park SH (2009) Effect of processparameters on 3-hydroxypropionic acid production from glycerolusing a recombinant Escherichia coli. Appl Microbiol Biotechnol84:649–657
Raj SM, Rathnasingh C, Jung WC, Selvakumar E, Park SH (2010) Anovel NAD+-dependent aldehyde dehydrogenase encoded by thepuuC gene of Klebsiella pneumoniae DSM 2026 that utilizes 3-hydroxypropionaldehyde as a substrate. Biotechnol BioprocessEng 15(1):131–138
Rathnasingh C, Raj SM, Jo JE, Park SH (2009) Development andevaluation of efficient recombinant Escherichia coli strains forthe production of 3-hydroxypropionic acid from glycerol.Biotechnol Bioeng 104:729–739
Saxena RK, Anand P, Saran S, Isar J (2009) Microbial production of1, 3-propanediol: recent developments and emerging opportuni-ties. Biotechnol Adv 27(6):895–913
Seo MY, Seo JW, Heo SY, Baek JO, Rairakhwada D, Oh BR, Seo PS,Choi MG, Kim CH (2009) Elimination of by-product formationduring production of 1, 3-propanediol in Klebsiella pneumoniaeby inactivation of glycerol oxidative pathway. Appl MicrobiolBiotechnol 84:527–534
Seo JW, Seo MY, Oh BR, Heo SY, Baek JO, Rairakhwada D, Luo LH,Hong WK, Kim CH (2010) Identification and utilization of a 1,3-propanediol oxidoreductase isoenzyme for production of 1, 3-propanediol from glycerol in Klebsiella pneumoniae. ApplMicrobiol Biotechnol 85:659–666
Severine FF, Bernard J, Christiane F (1995) Transformation of wild-type Klebsiella pneumoniae with plasmid DNA by electro-poration. J Microbiol Methods 24:49–54
Stephanopoulos GN, Aristidou AA, Nielsen J (1998) Metabolicengineering: principles and methodologies. Academic Press,San Diego, California
Wang W, Sun J, Hartlep M, Deckwer WD, Zeng AP (2003)Combined use of proteomic analysis and enzyme activityassays for metabolic pathway analysis of glycerol fermen-tation by Klebsiella pneumoniae. Biotechnol Bioeng 83:525–536
Zhao L, Zheng Y, Xingyuan M, Wei D (2009) Effects of over-expression of glycerol dehydrogenase and 1, 3-propanedioloxidoreductase on bioconversion of glycerol into 1, 3-propandediol by Klebsiella pneumoniae under microaerobicconditions. Bioprocess Biosys Eng 32:313–320
Appl Microbiol Biotechnol (2011) 90:1253–1265 1265
Copyright © 2022 FDOKUMEN