
Biotechnological valorisation of raw glycerol discharged after bio-diesel (fatty acid methyl esters)...
14
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/223733740 Biotechnological valorisation of raw glycerol discharged after bio-diesel (fatty acid methyl esters) manufacturing process... Article in Biomass and Bioenergy · January 2008 DOI: 10.1016/j.biombioe.2007.06.007 CITATIONS 254 READS 559 8 authors, including: Some of the authors of this publication are also working on these related projects: bioactive compounds View project Characterization of phosphatidic acid phosphatase activity in the oleaginous yeast Yarrowia lipolytica View project Seraphim Papanikolaou Agricultural University of Athens 128 PUBLICATIONS 4,894 CITATIONS SEE PROFILE Isabelle Chevalot French National Centre for Scientific Research 60 PUBLICATIONS 1,508 CITATIONS SEE PROFILE Ivan Marc French National Centre for Scientific Research 78 PUBLICATIONS 1,673 CITATIONS SEE PROFILE George Aggelis University of Patras 121 PUBLICATIONS 4,989 CITATIONS SEE PROFILE All content following this page was uploaded by George Aggelis on 21 January 2015. The user has requested enhancement of the downloaded file. All in-text references underlined in blue are added to the original document and are linked to publications on ResearchGate, letting you access and read them immediately.
Transcript of Biotechnological valorisation of raw glycerol discharged after bio-diesel (fatty acid methyl esters)...

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/223733740
Biotechnologicalvalorisationofrawglyceroldischargedafterbio-diesel(fattyacidmethylesters)manufacturingprocess...
ArticleinBiomassandBioenergy·January2008
DOI:10.1016/j.biombioe.2007.06.007
CITATIONS
254
READS
559
8authors,including:
Someoftheauthorsofthispublicationarealsoworkingontheserelatedprojects:
bioactivecompoundsViewproject
CharacterizationofphosphatidicacidphosphataseactivityintheoleaginousyeastYarrowia
lipolyticaViewproject
SeraphimPapanikolaou
AgriculturalUniversityofAthens
128PUBLICATIONS4,894CITATIONS
SEEPROFILE
IsabelleChevalot
FrenchNationalCentreforScientificResearch
60PUBLICATIONS1,508CITATIONS
SEEPROFILE
IvanMarc
FrenchNationalCentreforScientificResearch
78PUBLICATIONS1,673CITATIONS
SEEPROFILE
GeorgeAggelis
UniversityofPatras
121PUBLICATIONS4,989CITATIONS
SEEPROFILE
AllcontentfollowingthispagewasuploadedbyGeorgeAggelison21January2015.
Theuserhasrequestedenhancementofthedownloadedfile.Allin-textreferencesunderlinedinblueareaddedtotheoriginaldocumentandarelinkedtopublicationsonResearchGate,lettingyouaccessandreadthemimmediately.

This article was published in an Elsevier journal. The attached copyis furnished to the author for non-commercial research and
education use, including for instruction at the author’s institution,sharing with colleagues and providing to institution administration.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Available at www.sciencedirect.com
http://www.elsevier.com/locate/biombioe
Biotechnological valorisation of raw glycerol dischargedafter bio-diesel (fatty acid methyl esters) manufacturingprocess: Production of 1,3-propanediol, citric acid andsingle cell oil
Seraphim Papanikolaoua,b,�, Stylianos Fakasb,c, Michel Ficka, Isabelle Chevalota,Maria Galiotou-Panayotoub, Michael Komaitisb, Ivan Marca, George Aggelisc
aLaboratoire des Sciences du Genie Chimique, CNRS, ENSIC/ENSAIA, UPR 6811, 2 Avenue de la Foret de Haye,
54505 Vandœuvre-les-Nancy, FrancebDepartment of Food Science & Technology, Agricultural University of Athens, 75 Iera Odos, 11855 Athens, GreececDivision of Genetics, Cell and Development Biology, Department of Biology, University of Patras, 26504 Patras, Greece
a r t i c l e i n f o
Article history:
Received 18 December 2006
Received in revised form
12 June 2007
Accepted 25 June 2007
Available online 21 August 2007
Keywords:
Clostridium butyricum
Yarrowia lipolytica
Mortierella isabellina
Bio-diesel
Citric acid
Fatty acid methyl esters
Single-cell oil
1,3-Propanediol
Raw glycerol
a b s t r a c t
Raw glycerol, byproduct from bio-diesel production process, is used as carbon substrate in
several biotechnological applications. Using Clostridium butyricum F2b, 47.1 g L�1 of
1,3-propanediol was produced in batch anaerobic cultures while substrate uptake rate
(rS, expressed in g L�1 h�1) increased with increase in glycerol concentration in the medium.
In continuous cultures, microbial behaviour was studied in transitory states after addition
of 1,3-propanediol in the chemostat vessel. Microbial growth was not affected by the high
1,3-propanediol (which was added in the chemostat vessel) concentration, while butyric
and acetic acids concentrations were increased. In a two-stage continuous culture,
43.5 g L�1 of 1,3-propanediol was produced with a total volumetric productivity of
1.33 g L�1 h�1.
Yarrowia lipolytica ACA-DC 50109 was grown in nitrogen-limited aerobic cultures on raw
glycerol and it exhibited remarkable biomass production even at high glycerol concentra-
tion media, while rS decreased with increase in glycerol concentration. Citric acid was
produced after nitrogen depletion in the medium, with the highest quantity of 62.5 g L�1,
and yield on glycerol consumed was 0.56 g g�1. Fatty acid analysis of total cellular lipids
showed that glycerol concentration increase in the growth medium somehow increased
the cellular unsaturated fatty acids content of lipids.
Mortierella isabellina ATHUM 2935 exhibited satisfactory growth in nitrogen-limited
aerobic cultures with raw glycerol used as sole substrate. When high initial glycerol
quantities were employed (e.g. 100 g L�1), 4.4 g L�1 of lipid were accumulated corresponding
to around 51% (wt/wt) of lipid in dry weight. rS constantly decreased with increase in
glycerol concentration in the medium, and in all cases notable glycerol quantities remained
unconsumed in the medium.
& 2007 Elsevier Ltd. All rights reserved.
ARTICLE IN PRESS
0961-9534/$ - see front matter & 2007 Elsevier Ltd. All rights reserved.doi:10.1016/j.biombioe.2007.06.007
�Corresponding author. Laboratory of Food Microbiology & Biotechnology, Agricultural University of Athens, Athens, Greece.Tel./fax: +30 210 5294700.
E-mail address: [email protected] (S. Papanikolaou).
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 1

Author's personal copy
1. Introduction
Bio-diesel fuels defined as fatty acid methyl esters derived
from various renewable lipid resources (e.g. rapeseed oil,
soybean oil, palm oil, etc.) constitute an alternative type of
fuel for various types of diesel engines and heating systems
[1]. With the obligatory application of bio-fuels in a large
commercial scale in Europe, enormous quantities of glycerol
deposit in the market are likely to be generated in the near
future [1], while glycerol-containing residue is also produced
in significant amounts from fat saponification and alcoholic
beverage fabrication units [2,3]. Conversion, thus, of glycerol
to various high-value added products attracts much interest.
The main technique of biotechnological valorisation of
glycerol is related to its biotransformation into 1,3-propane-
diol, a substance of notable importance for the textile and
chemical industry [4,5]. This conversion is carried out by a
number of prokaryotic cells belonging to the family Enter-
obacteriaceae and to the genus Clostridium [4,6]. Glycerol has
also been alternatively utilised as sole substrate by various
types of microorganisms for production of other metabolites
such as ethanol and hydrogen [7] or biomass and a-amylase
[8]. Organic acids have also been produced by strains using
glycerol [5,9], but citric acid in spite of being one of the most
important metabolites produced via biotechnological meth-
ods in industrial scale [10], has been rarely produced using
glycerol as starting material [11]. Microbial lipids (also called
single-cell oils, SCO) present a potential industrial interest
due to capacity of various microorganisms to store lipids with
unusual composition or structure [12,13]. In spite of the
significant number of studies dealing with the production of
SCO by various microorganisms grown on a plethora of
carbon sources and culture configurations, data dealing with
growth of oleaginous microorganisms on (raw) glycerol are
quite limited [14,15]. Through utilisation of raw glycerol
discharged from bio-diesel units for the production of lipid,
we could recycle the by-product, decreasing significantly the
production cost of the whole process.
Although utilisation of raw glycerol in the fermentation
medium without prior purification offers a remarkable
advantage against the traditional use of pure glycerol as
substrate, only few reports have appeared in the literature on
the use of this substrate as sole carbon source [1–3,11,16–18].
ARTICLE IN PRESS
Fig. 1 – Pathways involved in glycerol breakdown by Clostridium butyricum, Yarrowia lipolytica and Mortierella isabellina.
TAGs: triacylglycerols; 3-HPA: 3-hydroxypropionaldehyde; (a)–(c) systems transporting pyruvic acid from cytosol to
mitochondrion and inversely; (d) system transporting citric and malic acid from cytosol to mitochondrion and inversely; ACL:
ATP-citrate lyase; FAS: fatty acid synthetase; ICDH: iso-citrate dehydrogenase; MDc: malate dehydrogenase (cytoplasmic);
MDm: malate dehydrogenase (mitochondrial); ME: NADPH+-malic enzyme; PD: pyruvate dehydrogenase; CS: citrate synthase;
ICL: iso-citrate lyase; GK: glycerol kinase; GDHt: glycerol dehydratase; GDH: glycerol dehydrogenase; 3-P-GDH: 3-P-glycerol
dehydrogenase; DHAk: di-hydroxyacetone kinase; PDOR: 1,3-propanediol oxidoreductase; FD: ferredoxine oxido-reductase.
Pathways described in [4,13].
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 1 61

Author's personal copy
Recent investigations showed that promising results can be
achieved with Clostridium butyricum F2b, which produced
considerable quantities of 1,3-propanediol on raw glycerol in
continuous mode [19]. Furthermore, Yarrowia lipolytica ACA-
DC 50109 [LGAM S(7)1] presented accumulation of citric acid
in the medium when grown on raw glycerol in nitrogen-
limited flask cultures [20]. Additionally, the mould Mortierella
isabellina ATHUM 2935 produced huge quantities of lipid when
cultivated on high-glucose nitrogen-limited media [21]. The
aim of the present study is to elucidate further the possibi-
lities of raw glycerol valorisation through different metabolic
pathways using the above-mentioned microorganisms as
model systems. These microorganisms were chosen as in
the case of C. butyricum, glycerol undergoes assimilation that
involves different pathways compared with the one of the
eukaryotic cells (DHA regulon against traditional phosphor-
ylation of glycerol to 3-P-glycerol), while the production of
SCO or citric acid is a non-growth-coupled process occurring
after nitrogen exhaustion from the medium in contrast to
biosynthesis of 1,3-propanediol [4,10,13]. Pathways involving
glycerol breakdown and biosynthesis of metabolites by
C. butyricum, Y. lipolytica and M. isabellina are shown in Fig. 1.
Bio-kinetic considerations are considered and discussed.
2. Materials and methods
2.1. Microorganisms, growth media and cultivationconditions
C. butyricum F2b, Y. lipolytica ACA-DC 50109 and M. isabellina
ATHUM 2935 were used. Strains were maintained as pre-
viously described [19–21]. C. butyricum preculturing and
bioreactor culturing were performed in the medium described
by Homann et al. [22]. Batch and single-stage continuous
cultures were carried out in a 2-L bioreactor (Biolafitte) as
described in Papanikolaou et al. [19]. Anaerobic conditions
were maintained by self-generated anaerobiosis. Two-stage
culture was performed with the aid of two 2-L bioreactors
(Biolafitte, France), one filled with 0.5 of medium and the
other with 1.3 L (agitation 200 rpm, incubation temperature
T ¼ 33 1C, pH ¼ 7.070.1, and flow rate is 5575 mL h�1). In
continuous cultures, steady state was obtained after contin-
uous flow of at least 5 working volumes of the medium
through the vessel.
Salt composition of the medium used for Y. lipolytica
cultures is described in Papanikolaou et al. [20], while that
for M. isabellina cultures is as in Fakas et al. [23]. Ammonium
sulphate and yeast extract, 0.5 g L�1 each, were used as
nitrogen sources. Experiments were performed in 250-mL
conical flasks, containing 5071 mL of growth medium
inoculated with 1 mL of exponential preculture (in Y. lipolytica
1 mL contains 1–3�108 cells, in M. isabellina 1 mL contains
1–3�105 spores; in both cases initial biomass inoculums
concentration was 0.2070.05 g L�1) as described in [20] in
T ¼ 28 1C. In yeast cultures, in order to prevent excessive pH
drop, highly buffered media were employed (utilisation of
KH2PO4 and Na2HPO4 at concentrations 12 g L�1 of each) while
additionally a volume (500–600mL) of KOH at 5 M was
periodically and aseptically added in order to maintain a
medium pH value greater than 5.0. All yeast or mould cultures
were carried out at initial pH ¼ 6.170.2. The carbon source
used was raw glycerol issued from bio-diesel (fatty acid
methyl esters) production unit [Agro-chemical industry
EPILOR-R&D, Compiegne, France—glycerol content 65%
(wt/wt)]. Impurities in the industrial glycerol solution were
potassium and sodium salts [4–5% (wt/wt)], methanol
[3% (wt/wt)], heavy metals and lignin [1% (wt/wt)] other
organic materials [0.5% (wt/wt)] and water [26% (wt/wt)]. For
all strains, control experiments were done using pure glycerol
[95% (wt/wt)].
2.2. Quantitative determinations and chemical analyses
Biomass, glycerol and products concentration were estimated
as previously described [19,20]. Carbon recovery (Rc) was
calculated in C. butyricum cultures as indicated elsewhere
[4,24]. Total cellular lipid was extracted with a mixture of
chloroform and methanol 2:1 (v/v), converted to methyl esters
and analysed by GLC as described earlier in [23]. Methods for
determining ammonium concentration, pH, dissolved oxygen
(DO) concentration and specific rate of oxygen uptake
(qO2Fmg mg�1 h�1) in the shaker cultures were as previously
described in [21].
2.3. Notation
Ac: acetic acid (g L�1), CAS 64-19-7; But: butyric acid (g L�1),
CAS 107-92-6; Cit: citric acid (g L�1), CAS 77-92-7; GLA: g-linolenic acid, CAS 506-26-3; L: microbial total lipid (g L�1); PD:
1,3-propanediol (g L�1), CAS 504-63-2; S: glycerol (g L�1), CAS
56-81-5; X: biomass (g L�1); YX/S: global biomass yield (g of
biomass per g of glycerol consumed); Yp/S: product yield (g of
product—p per g of glycerol consumed, where p was Ac, But,
PD, Cit, L); Pp: volumetric productivity (g of product—p per L
per h); Subscripts 0, r and max indicate the initial, remaining
and maximum quantity, respectively, of the components, in
the kinetics performed.
3. Results
3.1. Cultures of C. butyricum F2b
Anaerobic batch cultures of C. butyricum were carried out with
raw glycerol utilised as the sole substrate, at initial concen-
trations 39 and 90 g L�1, and satisfactory biomass production
was observed (Xmax ¼ 1.2–2.6 g L�1). mmax evaluated at the
early exponential phase was 0.3670.02 h�1 with lower values
obtained, at high initial glycerol concentration culture
(S0 ¼ 90 g L�1). Substrate uptake rate (rS), expressed as rS ¼
�DS=Dt and estimated during exponential growth phase,
showed some increase when initial substrate quantity in
the growth medium increased (at S0 ¼ 39 g L�1 rS was
2.3 g L�1 h�1 while at S0 ¼ 90 g L�1 rS was 3.4 g L�1 h�1), while
at the end of cultures insignificant S quantities remained
unconsumed. The fermentation kinetics at S0 ¼ 90 g L�1 is
shown in Fig. 2. YX/S presented a slightly lower value for
the cultivation of S0 ¼ 90 g L�1 compared with S0 ¼ 39 g L�1
(0.026 against 0.030 g g�1). PD was the principal metabolic
ARTICLE IN PRESS
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 162

Author's personal copy
product with a maximum concentration of 47.1 g L�1. Yields
YPD/S, YAc/S and YBut/S presented almost equivalent values
regardless of S0 concentration (values 0.53, 0.028 and
0.09 g g�1, respectively).
In order to investigate the effect of PD addition upon cell
growth and secretion of metabolites, a continuous culture
was carried out (dilution rate D imposed 0.0470.005 h�1,
S0 ¼ 80 g L�1), and when steady state was attained (at steady
state PD ¼ 41.2, X ¼ 1.5, Sr ¼ 5.5, Ac ¼ 1.0 and But ¼ 9.9 g L�1), a
43 g pulse of PD was carried out in the fermentation medium.
The evolution of biomass, 1,3-propanediol, substrate and
organic acids, followed until 25 h after the injection and
illustrated in Figs. 3(a and b), shows that despite the
significant quantity of PD found in the chemostat vessel
(84.2 g L�1), biomass concentration remained almost constant
(X ¼ 1.570.1 g L�1), obviously without inhibition exerted by
1,3-propanediol. Likewise, increment in PD in the vessel
resulted in a small decrease in glycerol consumption, and
acetic acid and butyric acid concentrations increased. When
the excess of PD was washed out from the bioreactor, S, Ac
and But concentrations tended to reach the values of steady-
state conditions (Fig. 3b). Furthermore, it appears that when
PD concentration increased, YX/S showed the tendency to
decrease despite constancy of biomass concentration in the
transitory stage, while when PD excess was discharged from
ARTICLE IN PRESS
0
2
4
6
8
0
0
X(g L-1)
X, A
c, B
ut (g
L-1
)
12
10
Time (h)
5040302010S
, P
D (
g L
-1)
100
80
60
40
20
S (g L-1)
PD (g L-1)But (g L-1)
Ac (g L-1)
Fig. 2 – Kinetics of biomass (X, g L�1), 1,3-propanediol (PD,
g L�1), acetic acid (Ac, g L�1) and butyric acid (But, g L�1)
production and glycerol (S, g L�1) consumption of Clostridium
butyricum during growth in batch bioreactor experiment.
Culture conditions: S0 ¼ 90.2 g L�1; pH ¼ 7.070.1; incubation
temperature T ¼ 33 1C. Kinetics were conducted in duplicate
by using different inocula.
0
1
2
3
0
0 5
0
5
0
1
2
0
X p
roduced (
g L
-1)
2.5
1.5
0.5
Time after transition (h)
3025201510-5
0 5
Time after transition (h)
3025201510-5
0 5
Time after transition (h)
3025201510-5
100
PD
(g L
-1)
80
60
40
20
Pulse of 1,3-propanediol
PD (g L-1) X (g L-1)
S r
em
ain
ing, B
ut pro
duced (
g L
-1) 20
15
10
Ac p
roduced (
g L
-1)1.5
0.5
Ac (g L-1)But (g L-1)
S (g L-1)
YX
/S, Y
Ac/S
(g g
-1)
0.05
0.04
0.03
0.02
0.01
0
YB
ut/S (
g g
-1)
0.2
0.15
0.1
0.05
YBut/S (g g-1) YX/S (g g-1)
YAc/S (g g-1)
Fig. 3 – Addition of 1,3-propanediol at steady-state
conditions in single-stage anaerobic continuous culture of
raw glycerol by Clostridium butyricum. Evolution of
1,3-propanediol and biomass concentrations (a), glycerol,
acetic acid and butyric acid concentrations (b) and biomass,
acetic acid and butyric acid yields on glycerol consumed (c).
Culture conditions: S0 ¼ 80 g L�1; pH ¼ 7.070.1;
D ¼ 0.0470.005 h�1; incubation temperature T ¼ 33 1C; pulse
of 43 g of PD at t ¼ 0.
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 1 63

Author's personal copy
the chemostat vessel, YX/S values tended to be the ones of the
steady state. Likewise, yields YAc/S and YBut/S clearly increased
with PD quantity increase, decreasing afterwards when PD
concentration gradually decreased (Fig. 3c). It should be noted
that similar biochemical behaviour has been observed when
lower 1,3-propanediol concentrations were added at steady-
state conditions into the chemostat vessel (added PD quan-
tities of 10, 20 or 39 g—data not presented).
A two-stage culture was also carried out (S0 ¼ 90 g L�1, flow
rate at 5575 mL h�1). The first stage of the culture (active
volume 0.5 L) presented a higher dilution rate (D ¼ 0.11 h�1),
are hence somewhat increased 1,3-propanediol volumetric
productivity could have been achieved. The second stage
(active volume 1.3 L), presenting a lower dilution rate
(D ¼ 0.04 h�1), was mainly used to further increase the
product concentration. It was also desirable to study the
biochemical behaviour of the microorganism when continu-
ously significant quantities of metabolites (mainly PD) were
injected into the second-stage vessel. The obtained result for
both stages is shown in Table 1. Indeed, for the first stage of
the culture satisfactory PD quantity and productivity were
obtained (PD ¼ 32.5 g L�1, PPD ¼ 3.58 g L�1 h�1), whereas
around 28% (wt/wt) of the substrate remained. In the second
stage, almost all of the remaining glycerol was assimilated
and a significant final 1,3-propanediol concentration
(43.5 g L�1) was achieved, but X quantity was significantly
lower as compared with the first one. Furthermore, in the
second stage of the culture a remarkably lower carbon
recovery was observed and a lower YPD/S was obtained.
Moreover, acetic acid was produced in higher amounts and
butyric acid in lower ones compared with the first stage.
3.2. Cultures of Y. lipolytica ACA-DC 50109
Aerobic nitrogen-limited flask cultures of Y. lipolytica were
carried out with raw glycerol utilised as sole substrate.
Literature suggests that enhancement of Cit production by
yeast strains occurs only in nitrogen-limited media in which
pH is maintained greater than 4.5 (see for instance Rymowicz
et al. [11]), while the strain under investigation produced low
quantities of organic acids in media that did not present a
satisfactory buffer capacity and medium pH dropped rapidly
to levels below 3.5 [20]. Therefore, in order to quantify the
effect of glycerol concentration upon growth and citric acid
production by the microorganism, cultures in a controlled
mode concerning pH evolution in the medium were done
(use of highly buffered media and daily correction of the
medium pH so that pH value was maintained within 5.0 and
6.2—see Section 2). Nitrogen concentration in the medium
remained constant [initial (NH4)2SO4 and yeast extract at
0.5 g L�1] while that of initial glycerol varied (S0 ranged
between 20 and 164 g L�1). The obtained result of cultures is
summarised in Table 2. In all cases and despite differentia-
tions in S0 value (which in some cases presented notably high
values, e.g., 90 and 164 g L�1), no lag phase was observed,
while mmax, as evaluated at the early exponential growth
phase, obtained the value of 0.1870.03 h�1, with the lower
values observed for the high S0 concentration cultures
presumably due to inhibition exerted by the high substrate
concentration. Up to 50710 h after inoculation, the micro-
organism consumed almost all of the quantity of the available
extracellular nitrogen of the growth medium, and at that
period, glycerol consumption led mainly to the creation of
biomass, since insignificant quantities of organic acids
(e.g. citric acid up to 0.5–1.0 g L�1) were produced. Then, when
nitrogen became limiting, X presented a slight further
increase before reaching the kinetics plateau, which was in
the same magnitude regardless of S0 quantity (Table 2). It is
noted that S0 concentration increment decreased rS, while at
high initial substrate amount media, significant glycerol
quantities remained unconsumed at the culture medium
even after long incubation periods (when S0 ¼ 164 g L�1, Sr
was 52.5 g L�1 at t ¼ 600 h—see Table 2). From mmax and rS
values, it may be assumed that at high initial glycerol
concentration media (specifically at S0 ¼ 164 g L�1) slight
inhibition of the microbial growth was observed, although
Xmax concentration was unaffected by the significant S0
concentration in the medium (concentration within the range
of 7.0–7.9 g L�1 in all trials—see Table 2). However, remarkable
citric acid quantities were accumulated in the growth
medium mainly at high S0 concentration media (when
S0 ¼ 164 g L�1, Citmax ¼ 62.5 g L�1), while yield YCit/S signifi-
cantly increased with glycerol concentration increase in the
culture medium (highest YCit/S value 0.56 g g�1) and maximum
ARTICLE IN PRESS
Table 1 – Growth parameters, conversion yields and carbon recoveries in the first and second stage of the culture at steadystate, during an anaerobic two-stage continuous fermentation of raw glycerol by Clostridium butyricum F2b
D(h�1)
X(g L�1)
S(g L�1)
PD(g L�1)
Ac(g L�1)
But(g L�1)
Rc (% wt/wt)
YPD/S
(g g�1)YBut/S
(g g�1)YAc/S
(g g�1)PPD
(g L�1 h�1)
First stage of the culture
0.11 2.2 25.4 32.5 2.0 7.8 95 0.50 0.12 0.031 3.58
Second stage of the culture
0.04 1.4 0.5 43.5 3.4 9.6 76 0.44 0.07 0.056 1.74
Global fermentation
1.4 0.5 43.5 3.4 9.6 88 0.49 0.11 0.038 1.33
Culture conditions: S0 ¼ 90 g L�1; flow rate 5575 mL h�1; pH ¼ 7.070.1; incubation temperature T ¼ 33 1C.
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 164
Author's personal copy
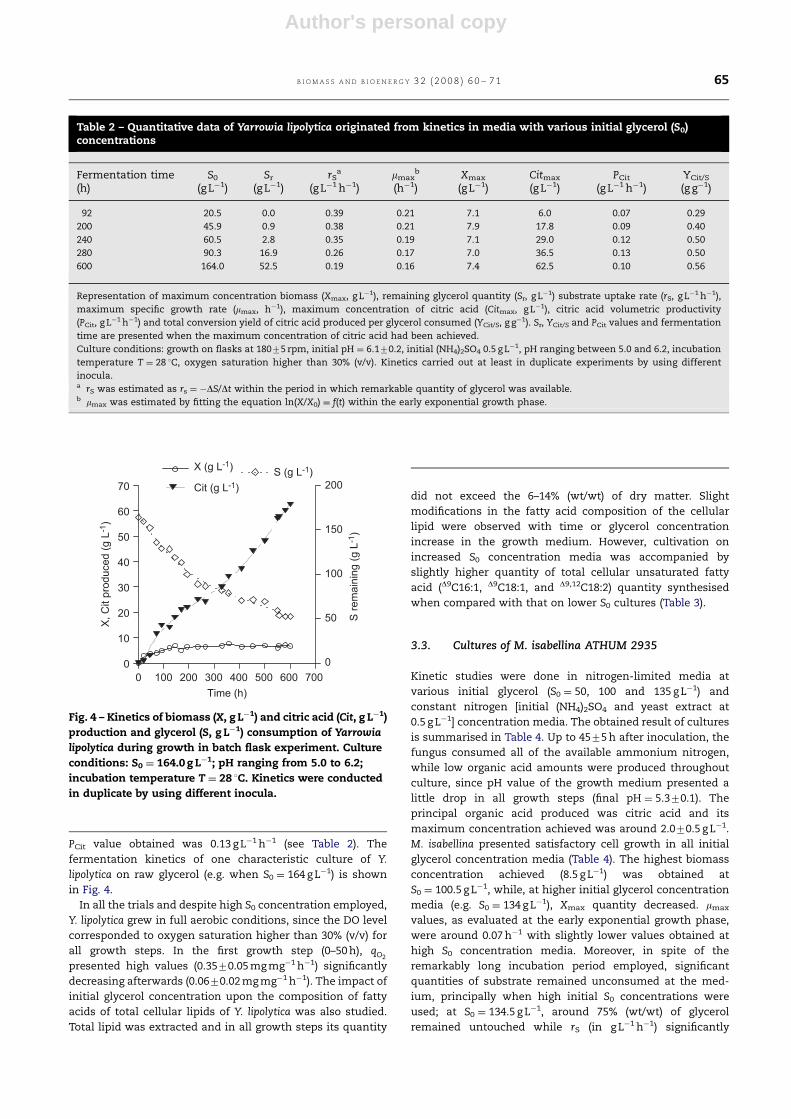
PCit value obtained was 0.13 g L�1 h�1 (see Table 2). The
fermentation kinetics of one characteristic culture of Y.
lipolytica on raw glycerol (e.g. when S0 ¼ 164 g L�1) is shown
in Fig. 4.
In all the trials and despite high S0 concentration employed,
Y. lipolytica grew in full aerobic conditions, since the DO level
corresponded to oxygen saturation higher than 30% (v/v) for
all growth steps. In the first growth step (0–50 h), qO2
presented high values (0.3570.05 mg mg�1 h�1) significantly
decreasing afterwards (0.0670.02 mg mg�1 h�1). The impact of
initial glycerol concentration upon the composition of fatty
acids of total cellular lipids of Y. lipolytica was also studied.
Total lipid was extracted and in all growth steps its quantity
did not exceed the 6–14% (wt/wt) of dry matter. Slight
modifications in the fatty acid composition of the cellular
lipid were observed with time or glycerol concentration
increase in the growth medium. However, cultivation on
increased S0 concentration media was accompanied by
slightly higher quantity of total cellular unsaturated fatty
acid (D9C16:1, D9C18:1, and D9,12C18:2) quantity synthesised
when compared with that on lower S0 cultures (Table 3).
3.3. Cultures of M. isabellina ATHUM 2935
Kinetic studies were done in nitrogen-limited media at
various initial glycerol (S0 ¼ 50, 100 and 135 g L�1) and
constant nitrogen [initial (NH4)2SO4 and yeast extract at
0.5 g L�1] concentration media. The obtained result of cultures
is summarised in Table 4. Up to 4575 h after inoculation, the
fungus consumed all of the available ammonium nitrogen,
while low organic acid amounts were produced throughout
culture, since pH value of the growth medium presented a
little drop in all growth steps (final pH ¼ 5.370.1). The
principal organic acid produced was citric acid and its
maximum concentration achieved was around 2.070.5 g L�1.
M. isabellina presented satisfactory cell growth in all initial
glycerol concentration media (Table 4). The highest biomass
concentration achieved (8.5 g L�1) was obtained at
S0 ¼ 100.5 g L�1, while, at higher initial glycerol concentration
media (e.g. S0 ¼ 134 g L�1), Xmax quantity decreased. mmax
values, as evaluated at the early exponential growth phase,
were around 0.07 h�1 with slightly lower values obtained at
high S0 concentration media. Moreover, in spite of the
remarkably long incubation period employed, significant
quantities of substrate remained unconsumed at the med-
ium, principally when high initial S0 concentrations were
used; at S0 ¼ 134.5 g L�1, around 75% (wt/wt) of glycerol
remained untouched while rS (in g L�1 h�1) significantly
ARTICLE IN PRESS
Table 2 – Quantitative data of Yarrowia lipolytica originated from kinetics in media with various initial glycerol (S0)concentrations
Fermentation time(h)
S0
(g L�1)Sr
(g L�1)rS
a
(g L�1 h�1)mmax
b
(h�1)Xmax
(g L�1)Citmax
(g L�1)PCit
(g L�1 h�1)YCit/S
(g g�1)
92 20.5 0.0 0.39 0.21 7.1 6.0 0.07 0.29
200 45.9 0.9 0.38 0.21 7.9 17.8 0.09 0.40
240 60.5 2.8 0.35 0.19 7.1 29.0 0.12 0.50
280 90.3 16.9 0.26 0.17 7.0 36.5 0.13 0.50
600 164.0 52.5 0.19 0.16 7.4 62.5 0.10 0.56
Representation of maximum concentration biomass (Xmax, g L�1), remaining glycerol quantity (Sr, g L�1) substrate uptake rate (rS, g L�1 h�1),
maximum specific growth rate (mmax, h�1), maximum concentration of citric acid (Citmax, g L�1), citric acid volumetric productivity
(PCit, g L�1 h�1) and total conversion yield of citric acid produced per glycerol consumed (YCit/S, g g�1). Sr, YCit/S and PCit values and fermentation
time are presented when the maximum concentration of citric acid had been achieved.
Culture conditions: growth on flasks at 18075 rpm, initial pH ¼ 6.170.2, initial (NH4)2SO4 0.5 g L�1, pH ranging between 5.0 and 6.2, incubation
temperature T ¼ 28 1C, oxygen saturation higher than 30% (v/v). Kinetics carried out at least in duplicate experiments by using different
inocula.a rS was estimated as rs ¼ �DS/Dt within the period in which remarkable quantity of glycerol was available.b mmax was estimated by fitting the equation ln(X/X0) ¼ f(t) within the early exponential growth phase.
0 0
0
X, C
it p
roduced (
g L
-1)
70
60
50
40
30
20
10
Time (h)
700600500400300200100
S r
em
ain
ing (
g L
-1)
200
150
100
50
X (g L-1)
Cit (g L-1)
S (g L-1)
Fig. 4 – Kinetics of biomass (X, g L�1) and citric acid (Cit, g L�1)
production and glycerol (S, g L�1) consumption of Yarrowia
lipolytica during growth in batch flask experiment. Culture
conditions: S0 ¼ 164.0 g L�1; pH ranging from 5.0 to 6.2;
incubation temperature T ¼ 28 1C. Kinetics were conducted
in duplicate by using different inocula.
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 1 65
Author's personal copy
decreased with glycerol concentration increase in the med-
ium (Table 4). From the decrease of mmax, rS and principally
Xmax values when S0 concentration increased, it may be
assumed that noticeable substrate inhibition was exerted
mainly at significantly high S0 quantity media (this was
obvious at S0 ¼ 134 g L�1). In contrast, in the culture with
S0 ¼ 100.5 g L�1, despite slight decrease of mmax value, rS was
comparable with the trial with S0 ¼ 50.5 g L�1, while biomass
production and lipogenesis were significant; Xmax was
8.5 g L�1 with 51.7% of lipids (wt/wt in dry matter) accumu-
lated inside the fungal mycelia (quantity corresponding to
4.4 g L�1), while lipid yield (YL/S) obtained the value of
0.0770.01 g g�1 in all trials (Table 4). Finally, total biomass
yield (YX/S) presented the value of 0.1570.01 g g�1, with
the lower values obtained at high S0 concentration media
(data not presented).
In all the trials, M. isabellina grew in full aerobic conditions,
since the DO level corresponded to oxygen saturation higher
than 70% (v/v) for all growth steps. As previously (Y. lipolytica
fermentation), in the first growth step (0–45 h), in which
ammonium nitrogen was found in the medium, qO2presented
elevated values (around 0.4070.05 mg mg�1 h�1) and then the
respiratory activity of the strain significantly decreased
(qO2values around 0.0870.02 mg mg�1 h�1). The fermentation
kinetics of M. isabellina when S0 ¼ 100.5 g L�1 is shown in
Fig. 5.
In M. isabellina, the principal cellular fatty acids were oleic
(D9C18:1), linoleic (D9,12C18:2) and palmitic (C16:0) acid and
concentrations of these remained almost constant during
growth. Cellular fatty acids produced in lower quantities were
stearic acid (C18:0), palimitoleic acid (D9C16:1) and g-linolenic
acid (D6,9,12C18:3—GLA) (Table 5). In the first growth step, GLA
ARTICLE IN PRESS
Table 3 – Fatty acid composition of lipid produced by Yarrowia lipolytica (%, wt/wt), when this microorganism was grown onraw glycerol
Fermentation time C16:0 D9C16:1 C18:0 D9C18:1 D9,12C18:2 D9C16:1/C16:0
D9C18:1/C18:0
D9,12C18:2/D9C18:1
Growth on raw glycerol, S0 ¼ 45.9 g L�1
Late exponential
(50–70 h)
15.7 7.1 12.5 40.1 22.2 0.45 3.21 0.55
Mid stationary
(120–140 h)
13.1 8.1 9.9 45.1 19.2 0.62 4.55 0.42
Late stationary
(220–250 h)
14.0 8.0 11.0 44.9 20.2 0.57 4.08 0.45
Growth on raw glycerol, S0 ¼ 164.0 g L�1
Late exponential
(50–70 h)
15.9 8.9 7.9 46.9 20.0 0.55 5.93 0.42
Mid stationary
(120–140 h)
12.1 13.9 5.9 44.1 21.8 1.14 7.47 0.49
Late stationary
(220–250 h)
16.9 15.5 5.2 42.9 19.4 0.92 8.25 0.45
Culture conditions as in Table 2.
Table 4 – Quantitative data of Mortierella isabellina originated from kinetics in media with various initial glycerol (S0)concentrations
Fermentation time(h)
S0
(g L�1)Sr
(g L�1)rS
a
(g L�1 h�1)mmax
b
(h�1)Xmax
(g L�1)Lmax
(g L�1)Lipid (%, wt/
wt)YL/S
(g g�1)
310 50.5 9.8 0.13 0.08 6.5 2.4 36.9 0.06
420 100.5 45.1 0.13 0.07 8.5 4.4 51.7 0.08
380 135.1 99.5 0.09 0.07 5.0 2.5 50.0 0.07
Representation of maximum concentration biomass (Xmax, g L�1), remaining glycerol quantity (Sr, g L�1) substrate uptake rate (rS, g L�1 h�1),
maximum specific growth rate (mmax, h�1), maximum concentration of cellular lipid (Lmax, g L�1), corresponding lipid in dry weight (%, wt/wt)
and total conversion yield of lipid produced per glycerol consumed (YL/S, g g�1).
Sr and YL/S values and fermentation time are presented when the maximum concentration of lipid had been achieved. Culture conditions:
growth on flasks at 18075 rpm, initial pH ¼ 6.170.2, initial (NH4)2SO4 0.5 g L�1, pH ranging between 5.4 and 6.0, incubation temperature
T ¼ 28 1C, oxygen saturation higher than 70% (v/v). Kinetics carried out at least in duplicate experiments by using different inocula.a rS was estimated as rs ¼ �DS/Dt throughout culture.b mmax was estimated by fitting the equation ln(X/X0) ¼ f(t) within the early exponential growth phase.
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 166

Author's personal copy
content was elevated, while later, during lipid accumulation,
GLA concentration somewhat decreased, and then remained
constant. Similar total cellular fatty acid profiles were
observed for all fermentations of M. isabellina, regardless of
the concentration of glycerol employed in the culture
medium. GLAmax concentration was achieved at S0 ¼
100.5 g L�1 and corresponded to 120 mg L�1.
3.4. Comparison between growth on pure and rawglycerol
In order to investigate if the impurities of the industrial
feedstock could affect microbial growth and production of
metabolites, control experiments for all microorganisms
(batch cultures) were realised using pure glycerol as sole
carbon source and they were compared with the results
obtained when raw glycerol was used as substrate. The
cultures showed similar growth and production of metabo-
lites for all microorganisms used (C. butyricum, Y. lipolytica, M.
isabellina, data not presented). Therefore, it may be consid-
ered that the impurities of the industrial feedstock (salts,
methanol, etc.) do not have any impact on the metabolism of
the studied strains.
4. Discussion
Batch cultures of C. butyricum F2b were accompanied by PDmax
production of 47.1 g L�1, a quantity comparable with literature
values for batch and fed-batch fermentations of Clostridium
sp. or Enterobacteria cultured on glycerol used as sole
substrate [2,3,16–18,22,24–26]. Though, in some cases utilisa-
tion of C. butyricum mutants in fed-batch fermentations
resulted in significant PD quantities, of more than 70 g L�1
[27], while, the highest final PD concentrations reported in the
literature were 80.1–87.7 g L�1, obtained in fed-batch cultures
of a newly isolated Clostridium sp. strain on pure or raw
glycerol [1]. In the present study, YPD/S remained constant
despite differentiations in the S0 concentration in the
medium. Generally, the fermentation of glycerol by Clostri-
dium sp. strains presents a higher YPD/S than that conducted
by the Klebsiella sp. since in the later case the metabolism is
directed towards the synthesis of more end-fermentation
products compared with the former one [6]. With Klebsiella
sp., YPD/S can be substantially increased when differentiations
in the culture conditions are employed such as utilisation of
glycerol in significant excess or culture carried out under
micro-aerobic conditions or differentiations employed in
the extracellular oxido-reduction potential of culture
[25,26,28,29]. The spectrum of end-fermentation products in
K. pneumoniae cultures can also be critically influenced by the
‘‘purity’’ of the substrate used (use of ‘‘raw’’ or ‘‘pure’’ glycerol)
or the addition of organic compounds (e.g. fumarate) into the
medium [18,30]. In contrast, C. butyricum cultures are carried
out only in strict anaerobiosis, with distribution between final
concentrations of PD, Ac and But being strain dependent
[6,16].
In the present study, in continuous culture and steady-state
conditions, PD pulses were carried out. It was shown that at
increased PD concentrations at transitory stage, although X
concentration remained practically constant, yield YX/S de-
creased. Simultaneously, slight decrease of S uptake, and
increment of Ac and But concentrations and yields were
observed. The above findings suggest that even at very high
PD concentrations (e.g. 84.2 g L�1 into the chemostat vessel),
growth was not inhibited, while carbon flow was mainly
channelled towards the synthesis of organic acids instead of
biomass, resulting, thus, in carbon losses through the
reaction of decarboxylation of pyruvic acid (see reactions in
Fig. 1). Similarly, continuous cultures at constant D and
ARTICLE IN PRESS
0
2
4
6
8
0
0
L (g L-1)100
X, L p
roduced (
g L
-1)
Time (h)
500400300200100
10
12
S r
em
ain
ing (
g L
-1)
120
80
60
40
20
S (g L-1)X (g L-1)
Fig. 5 – Kinetics of biomass (X, g L�1) and lipid (L, g L�1)
production and glycerol (S, g L�1) consumption of Mortierella
isabellina during growth in batch flask experiment. Culture
conditions: S0 ¼ 100.5 g L�1; pH ranging from 6.1 to 5.2;
incubation temperature T ¼ 28 1C. Kinetics were conducted
in duplicate by using different inocula.
Table 5 – Fatty acid composition of lipid produced by Mortierella isabellina (%, wt/wt), when this microorganism was grownon raw glycerol
Fermentation time C16:0 D9C16:1 C18:0 D9C18:1 D9,12C18:2 D6,9,12C18:3
Late exponential (40–50 h) 20.1 1.3 5.8 46.6 18.0 8.3
Mid stationary (130–150 h) 21.2 2.8 5.2 52.0 13.5 3.8
Late stationary (220–250 h) 22.0 4.2 4.3 50.0 16.7 3.7
Culture conditions as in Table 4.
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 1 67

Author's personal copy
various inlet concentrations of the employed strain showed
that when S0 concentration increased, decrease in YX/S and Rc
values was observed. It was assumed that at high S0
concentrations (when the 1,3-propanediol pathway was
somehow saturated), carbon flow towards biomass and PD
synthesis decreased and the percentage of NAD+ regenerated
via the pathway of butyric acid biosynthesis increased,
resulting, hence, in carbon losses [19]. In contrast, in C.
butyricum DSM 5431 and its product-tolerant mutants, with
increase of inlet glycerol and thus 1,3-propanediol produced
concentration, although the yield YPD/S and the ratio of GDH
versus GDHt remained constant in the range of substrate
concentrations tested, a net increase of the NADH2/NAD+
ratio was observed, suggesting insufficient electron flow
through the reductive branch of the pathway of glycerol
catabolism, while the pathway of Ac biosynthesis was
favoured instead of that of But due to high intracellular
concentrations of acetyl-CoA detected, and that suggest
bottlenecks in the enzymes (e.g. aceto-acetyl-CoA thiolase,
butyrate kinase) leading to the formation of butyric acid [31].
In a two-stage culture carried out, large PD quantities
(43.5 g L�1) comparable with the highest values reported in the
literature for single- or two-stage 1,3-propanediol production
processes [28,32,33] were produced. The second stage of the
culture showed lower YPD/S, Rc and X values compared with
those of the first stage. This is probably due to incomplete
reduction of 3-HPA (the only metabolic intermediate of PD
biosynthesis pathway) to 1,3-propanediol and accumulation
of this toxic intermediate in the growth medium. This
intermediate product was not measured, and, possibly, for
this reason in the second stage low Rc values were observed.
Furthermore, in the second stage a metabolic shift favouring
the synthesis of acetic acid against that of butyric acid was
observed (see Table 1). It may be assumed, hence, that in the
above conditions, in the second stage of the culture a
bottleneck for the carbon flux through the butyrate biosynth-
esis occurred, resulting, thus, in shifting of the cellular
metabolism towards the biosynthesis of the acetic acid.
Y. lipolytica ACA-DC 50109 presented significant cell growth
with slight inhibition when raw glycerol was utilised as the
sole carbon source, although in some cases extremely high
initial glycerol quantities (S0 ¼ 164 g L�1) were used. Incre-
ment of glycerol concentration in the medium resulted in
significant decrease of rS, while mainly at high-substrate
concentration media, notable quantities of glycerol remained
unconsumed even though the fermentation time was
remarkably high. In M. isabellina ATHUM 2935 cultures,
biomass production declined at relatively high S0 concentra-
tions in the medium (e.g. at 134 g L�1) due to substrate
inhibition, while in all cultures significant quantities of
glycerol remained unconsumed in spite of the fact that the
fermentations were carried out for long incubation periods
(e.g. 300–400 h). In the literature, data dealing with the
cultivation of Y. lipolytica or lipid-accumulating strains on
glycerol used as sole substrate are rather equivocal. The
recombinant Y. lipolytica CX161-1B strain was batch cultured
on various S0 concentration media and significant growth
with similar mmax values was observed, regardless of S0
concentration, while no substrate inhibition was observed
even when S0 quantity was 150 g L�1 and all of the available
glycerol quantity was consumed [8]. The lipid-accumulating
C. curvata ATCC 20509 strain showed almost a 2-fold decrease
of Xmax value, when glycerol concentration increased from 32
to 128 g L�1 in flask nitrogen-limited cultures, while further S0
quantity increase, almost ceased microbial growth due to
inhibition exerted in high initial glycerol concentrations [14].
However, in fed-batch stirred tank bioreactor experiments,
the above microorganism consumed 300 g L�1 of glycerol
within 55 h of culture (rS ¼ 5.5 g L�1 h�1), producing around
120 g L�1 of biomass that contained 25% (wt/wt) of lipid [14],
while with a specified pumped external loop bioreactor the
respective values were 100 g L�1 and 32% (wt/wt), within the
same fermentation time [34]. In contrast, the oleaginous
Cunninghamella echinulata CCRC 31840 presented decreased
microbial growth and accumulation of lipid when glycerol
was used as substrate [35], and similar results were obtained
for other oleaginous Mucorales [36]. Additionally, fed-batch
nitrogen-limited cultures of the thiamine auxotroph
Y. lipolytica VKM-374/4 strain growing on glycerol, showed
increased substrate uptake rate despite S0 amounts used [9],
while citric-acid-producing Y. lipolytica mutants cultivated on
raw glycerol at extremely high S0 quantities (i.e. 200 g L�1)
presented efficient cell growth (X ¼ 16.5–26.5 g L�1) and com-
plete substrate consumption within 80–140 h without obvious
inhibition observed (rS from 1.4 to 2.5 g L�1 h�1—values
significantly higher compared to the ones obtained in the
present study) [11].
In the lipid- or citric acid-producing microorganisms that
are cultivated on glycerol used as substrate, glycerol, which
enters inside the microbial cell by facilitated diffusion, is
firstly converted to 3-P-glycerol and then to 3-P-dihydroxya-
cetone, reactions catalysed by GK and 3-P-GDH, thus entering
the second branch of the glucolytic pathway to yield pyruvate.
Subsequent metabolic steps to lipid or citric acid biosynthesis
are the same as for glucose (see Fig. 1). However, a portion of
glycerol must be channelled to hexose synthesis and then to
pentose synthesis, which are necessary for nucleic acid
biosynthesis and NADPH2 generation. Hexose synthesis is
carried out through gluconeogenesis that in the case of
glycerol involves the reversal of carbon flow from
3-P-glyceraldehyde. In the present study, both M. isabellina
and Y. lipolytica consumed with reduced rates glycerol mainly
at high S0 concentration media, since significant quantities of
this substrate remained in the medium despite high fermen-
tation duration. Considering for the case of M. isabellina that
huge quantities of biomass and fat (36 and 18 g L�1, respec-
tively) were produced at high initial glucose media
(e.g. 100 g L�1) with glucose almost completely consumed
[21], the reason for the slow assimilation of glycerol and the
lower production of biomass and lipid compared with the
glucose fermentation can be poor regulation of the enzymes
involved in the primary metabolic steps of glycerol assimila-
tion (GK, 3-P-GDH), or decreased activity of gluconeogenic
enzymes, or, finally slow activity of the NADP+-malic enzyme
(ME) that can severely curtail substrate uptake and accumula-
tion of fat [13]. In contrast, in glucose fermentation of
Y. lipolytica, despite notable growth and citric acid produc-
tion at high initial substrate concentration media
(i.e. 150 g L�1), as in the case of glycerol fermentation, glucose
remained unconsumed in significant quantities at the end of
ARTICLE IN PRESS
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 168

Author's personal copy
growth [37]. The fact, hence, that non-negligible glycerol
amounts remain unconsumed in the medium after long
incubation periods, seems not related with the regulatory
mechanisms of glycerol assimilation or gluconeogenesis
process but with the fact of being a strain-dependant event.
Other Y. lipolytica strains cultured on various initial glucose
carbon-excess batch cultures (glucose variations from 50 to
150 g L�1), consumed almost all available glucose quantity
[38,39], while in our laboratory, growth of newly isolated or
genetically engineered Y. lipolytica strains on glucose or raw
glycerol at various concentrations and carbon-excess condi-
tions, showed indeed strain-dependent discrepancies on
substrate uptake rate and complete exhaustion of the carbon
source from the medium (Papanikolaou, unpublished data).
In the present investigation, in Y. lipolytica cultures citric
acid was synthesised in significant quantities (Citmax ¼
62.5 g L�1), comparable with values reported in the literature
(values within 30 and 80 g L�1) [38–43]. It should be noted that
very high concentrations of citric acid (Citmax ¼ 120–135 g L�1)
have been achieved by mutant Y. lipolytica strains cultivated
on ethanol [44], crude lipids [45] or raw glycerol [11] in batch,
fed-batch or continuous operations. Likewise, YCit/S signifi-
cantly increased with glycerol concentration increment in the
medium due to ‘‘metabolic overflow’’ at carbon-excess con-
ditions [10,40]. Additionally, the highest yield YCit/S presented
the value of 0.56 g g�1 which was satisfactory and comparable
with the values reported in the literature for various Candida
sp. strains cultured on sugar-based media (yields from 0.50 to
0.77 g g�1 [38–40,43]). YCit/S is significantly dependent on the
fermented carbon source utilised, since substances with
higher carbon and reductance degree compared with glycerol,
can lead to higher conversion yields; for instance, growth of Y.
lipolytica strains on ethanol was accompanied by a citric acid
yield of 0.87 g g�1 [44], while growth on hydrophobic sub-
strates can be accompanied by conversion yields, which may
be close to or even higher than 1.0 g g�1 [41,45].
Although Y. lipolytica ACA-DC 50109 was reported capable to
produce huge quantities of lipid during growth on raw
glycerol in continuous bioreactor nitrogen-limited cultures
[15], in flask experiments the microorganism did not accu-
mulate notable lipid quantities. Cellular lipids showed slightly
higher quantity of unsaturated fatty acids with high initial
glycerol media. The higher ‘‘fluidity’’ on the membrane,
hence, could reflect the increased osmotic pressure of the
medium due to high glycerol concentration.
M. isabellina ATHUM 2935 cultivated on raw glycerol utilised
as the sole substrate in nitrogen-limited experiments pro-
duced relatively satisfactory quantities of microbial lipid
[up to 4.4 g L�1, around 50% (wt/wt) of lipid in dry weight].
This is an interesting result, given that glycerol is not
considered as an adequate substrate for lipid production for
various oleaginous Zygomycetes [12,35,36]. In contrast, sub-
stances favouring production of SCO from oleaginous Zygo-
mycetes in higher quantities compared with the present
study (e.g. quantities higher than 8 g L�1) are various sugars
and polysaccharides [12,35,36,46] or fatty materials [47–49]. In
general, a scarce number of investigations have been carried out
dealing with the production of lipid using glycerol as the sole
substrate, with YL/S values 0.08–0.15 g g�1 (values comparable
with the present study) [14,15,34]. Additionally, the fatty acid
profile of the microbial oil produced was largely unaffected by
initial concentration of glycerol, while GLA concentration in the
produced lipid decreased as a function of the fermentation time
(in accordance with Chen and Chang [35] and Chen and Liu [46]).
It is noted that in oleaginous Zygomycetes, GLA resides mostly
in the TAG fraction that comprises the edible portion of the lipid
[23]. GLAmax concentration of 120 mg L�1 was obtained, which is
2.5–10-fold lower compared with that obtained in various
Mucorales strains. The same strain (ATHUM 2935) produced
around 800 mg L�1 of GLA during growth on high-glucose media
[21]. GLAmax quantity of Mucor spp. strains was 370mg L�1 of
medium during growth on mixtures of fats and glucose [47].
A Mucor racemosus strain cultivated on olive oil produced huge
quantities of lipids (13.4 g L�1) with GLA produced at 725 mg L�1
[49]. C. echinulata CCRC 31840 produced 964 mg L�1 of GLA after
optimisation of the growth conditions [35] while additional
optimisation of the inoculation led to a production of
1350 mg L�1, after 5 days of culture [46].
In conclusion, raw glycerol, by-product discharged after
bio-diesel production process, was an adequate substrate for
the growth of C. butyricum F2b, Y. lipolytica ACA-DC 50109 and
M. isabellina ATHUM 2935. High value-added metabolic
products (1,3-propanediol, citric acid and SCO) in remarkable
quantities and satisfactory conversion yields were produced.
As for the economic significance of the results obtained from
the present study, it should be stressed that the utilisation of
bio-diesel (fatty acid methyl esters) in continuously increas-
ing quantities has dramatically decreased the price of
glycerol, resulting in the necessity of discovery of various
integrated bioprocesses of valorisation of this residue
(‘‘biorefinery approach’’). By 2007, it is expected that there
will be an over-capacity of more than 600,000 metric tonnes of
glycerol residue produced, based on the bio-diesel industry
growth in Europe. Currently, in various countries of Western
Europe (e.g. Germany), crude glycerine water derived from
various bio-diesel plants is treated as a typical ‘‘industrial
waste water’’ (with a cost of 0$ per kg—it is, hence, a waste
material) being used directly for biogas production. The
current cost of 1,3-propanediol is estimated to be 1.5–2.0$
per kg while that of citric acid is 0.8–1.2$ per kg. As for plant
lipids that contain g-linolenic acid and are currently com-
mercialised [in general, it is the case of the oil deriving from
the plant Oenothera biennis called also Evening primrose oil,
that contains GLA in concentrations 8–10% (wt/wt)], their cost
is estimated to be 45–50$ per kg. Therefore, although our
experiments have been carried out in small-scale operations
and scale-up studies are envisaged in future investigations of
our research teams, the significantly low (or even zero) cost or
raw glycerol and the (relatively) high price of the metabolites
produced, can potentially result in the viability of the
proposed approaches dealing with the biotransformation of
raw glycerol into 1,3-propanediol, citric acid and SCO contain-
ing g-linolenic acid.
Acknowledgments
Financial support was provided by: (a) the project entitled
‘‘Bio-diesel production from agro-industrial by-products’’
funded by the Greek Fuel Company DRACOIL SA; (b) the
ARTICLE IN PRESS
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 1 69

Author's personal copy
bilateral project between Greece and Slovak Republic entitled
‘‘Biotechnological production of bioactive lipids from
agro-industrial by-products’’.
R E F E R E N C E S
[1] Hirschmann S, Baganz K, Koschik I, Vorlop KD. Developmentof an integrated bioconversion process for the production of1,3-propanediol from raw glycerol waters. LandbauforschungVolkenrode 2005;55:261–7.
[2] Barbirato F, Himmi EH, Conte T, Bories A. 1,3-Propanediolproduction by fermentation: an interesting way to valorizeglycerin from the ester and ethanol industries. IndustrialCrops and Products 1998;7:281–9.
[3] Himmi EH, Bories A, Barbirato F. Nutrient requirements forglycerol conversion to 1,3-propanediol by Clostridium butyr-icum. Bioresource Technology 1999;67:123–8.
[4] Zeng AP, Biebl H. Bulk chemicals from biotechnology: thecase of 1,3-propanediol production and the new trends.Advances in Biochemical Engineering and Biotechnology2002;74:239–59.
[5] Lee SY, Hong SH, Lee SH, Park SJ. Fermentative production ofchemicals that can be used for polymer synthesis. Macro-molecular Bioscience 2004;4:157–64.
[6] Biebl H, Menzel K, Zeng AP, Deckwer WD. Microbial produc-tion of 1,3-propanediol. Applied Microbiology and Biotech-nology 1999;52:289–97.
[7] Ito T, Nakashimada Y, Senba K, Matsui T, Nishio N. Hydrogenand ethanol production from glycerol-containing wastesdischarged after biodiesel manufacturing process. Journal ofBioscience and Bioengineering 2005;100:260–5.
[8] Kim JW, Park TJ, Ryu DDY, Kim JY. High cell density culture ofYarrowia lipolytica using a one-step feeding process. Biotech-nology Progress 2000;16:657–60.
[9] Morgunov I, Kamzolova S, Perevoznikova O, Shishkanova N,Finogenova T. Pyruvic acid production by a thiamineauxotroph of Yarrowia lipolytica. Process Biochemistry2004;39:1469–74.
[10] Soccol CR, Vandenberghe LPS, Rondriques C, Pandey A. Newperspectives for citric acid production and application. FoodTechnology and Biotechnology 2006;44:141–9.
[11] Rymowicz W, Rywinska A, Zarowska B, Juszczyk P. Citric acidproduction from raw glycerol by acetate mutants of Yarrowialipolytica. Chemical Papers 2006;60:391–4.
[12] Certık M. Fermentation physiology and regulation of micro-bial polyunsaturated fatty acid biosynthesis. Research inAdvanced Bioscience and Bioengineering 2000;1:45–64.
[13] Ratledge C. Regulation of lipid accumulation in oleaginousmicro-organisms. Biochemical Society Transactions2002;30:1047–50.
[14] Meesters PAEP, Huijberts GNM, Eggink G. High-cell-densitycultivation of the lipid accumulating yeast Cryptococcuscurvatus using glycerol as a carbon source. Applied Micro-biology and Biotechnology 1996;45:575–9.
[15] Papanikolaou S, Aggelis G. Lipid production by Yarrowialipolytica growing on industrial glycerol in a single-stagecontinuous culture. Bioresource Technology 2002;82:43–9.
[16] Petitdemange E, Durr C, Abbad-Andaloussi S, Raval G. Fermen-tation of raw glycerol to 1,3-propanediol by new strains ofClostridium butyricum. Journal of Industrial Microbiology 2005;15.
[17] Gonzalez-Pajuelo M, Andrade JC, Vasconcelos I. Production of1,3-propanediol by Clostridium butyricum VPI 3266 using asynthetic medium and raw glycerol. Journal of IndustrialMicrobiology and Biotechnology 2004;31:442–6.
[18] Mu Y, Zhang D, Teng H, Wang W, Xiu ZL. Microbial productionof 1,3-propanediol by Klebsiella pneumoniae using crude
glycerol from bio-diesel preparation. Biotechnology Letters2006;28:1755–9.
[19] Papanikolaou S, Fick M, Aggelis G. The effect of raw glycerolconcentration on the production of 1,3-propanediol byClostridium butyricum. Journal of Chemical Technology andBiotechnology 2004;79:1189–96.
[20] Papanikolaou S, Muniglia L, Chevalot I, Aggelis G, Marc I.Yarrowia lipolytica as a potential producer of citric acid fromraw glycerol. Journal of Applied Microbiology 2002;92:737–44.
[21] Papanikolaou S, Komaitis M, Aggelis G. Single cell oil (SCO)production by Mortierella isabellina grown on high-sugarcontent media. Bioresource Technology 2004;95:287–91.
[22] Homann T, Tag C, Biebl H, Decker WD, Schink B. Fermenta-tion of glycerol to 1-3 propanediol by Klebsiella and Citrobacterstrains. Applied Microbiology and Biotechnology1990;33:121–6.
[23] Fakas S, Papanikolaou S, Galiotou-Panayotou M, Komaitis M,Aggelis G. Lipids of Cunninghamella echinulata with emphasisto g-linolenic acid distribution among lipid classes. AppliedMicrobiology and Biotechnology 2006;73:676–83.
[24] Chen Z, Xiu ZL, Wang JF, Zhang D, Xu P. Stoichiometricanalysis and experimental investigation of glycerol biocon-version to 1,3-propanediol by Klebsiella pneumoniae undermicroaerobic conditions. Enzyme and Microbial Technology2003;33:386–94.
[25] Chen X, Zhang DJ, Qi WT, Gao SJ, Xiu ZL, Xu P. Microbial fed-batch production of 1,3-propanediol by Klebsiella pneumoniaeunder micro-aerobic conditions. Applied Microbiology andBiotechnology 2003;63:143–6.
[26] Du C, Yan H, Zhang Y, Li Y, Cao Z. Use of oxidoreductionpotential as an indicator to regulate 1,3-propanediol fer-mentation by Klebsiella pneumoniae. Applied Microbiology andBiotechnology 2006;69:554–63.
[27] Reimann A, Biebl H. Production of 1,3-propanediol byClostridium butyricum DSM 5431 and product tolerant mu-tants in fed-batch cultures: feeding strategy for glycerol andammonium. Biotechnology Letters 1996;18:827–32.
[28] Menzel K, Zeng AP, Deckwer WD. High concentration andproductivity of 1,3-propanediol from continuous fermenta-tion of glycerol by Klebsiella pneumoniae. Enzyme and Micro-bial Technology 1997;20:82–6.
[29] Menzel K, Zeng AP, Deckwer WD. Enzymatic evidence for aninvolvement of pyruvate dehydrogenase in the anaerobicglycerol metabolism of Klebsiella pneumoniae. Journal ofBiotechnology 1997;56:135–42.
[30] Lin R, Liu H, Hao J, Cheng K, Liu D. Enhancement of 1,3-propanediol production by Klebsiella pneumoniae with fuma-rate addition. Biotechnology Letters 2005;27:1755–9.
[31] Reimann A, Abbad-Andaloussi S, Biebl H, Petitdemange H.1,3-Propanediol formation with product tolerant mutants ofClostridium butyricum DSM 5431 in continuous culture:productivity, carbon and electron flow. Journal of AppliedMicrobiology 1998;84:1125–30.
[32] Boenigk R, Bowien S, Gottschalk G. Fermentation of glycerolin continuous cultures of Citrobacter freundii. Applied Micro-biology and Biotechnology 1993;38:453–7.
[33] Xiu ZL, Song BH, Wang ZT, Sun LH, Feng EM, Zeng AP.Optimization of dissimilation of glycerol to 1,3-propanediolby Klebsiella pneumoniae in one- and two-stage anaerobiccultures. Biochemical Engineering Journal 2004;19:189–97.
[34] Meesters PAEP, van der Wal H, Weusthuis R, Eggink G.Cultivation of the oleaginous yeast Cryptococcus curvatus ina new reactor with improved mixing and mass transfercharacteristics (Super-s). Biotechnology Techniques1996;10:277–82.
[35] Chen HC, Chang CC. Production of g-linolenic acid by thefungus Cunninghamella echinulata CCRC 31840. BiotechnologyProgress 1996;12:338–41.
ARTICLE IN PRESS
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 170

Author's personal copy
[36] Sajbidor J, Certık M, Dobronova S. Influence of differentcarbon sources on growth, lipid content and fatty acidcomposition in four strains belonging to Mucorales. Bio-technology Letters 1988;10:347–50.
[37] Papanikolaou S, Galiotou-Panayotou M, Chevalot I, KomaitisM, Marc I, Aggelis G. Influence of glucose and saturated free-fatty acid mixtures on citric acid and lipid production byYarrowia lipolytica. Current Microbiology 2006;52:134–42.
[38] Rane KD, Sims KA. Production of citric acid by Candidalipolytica Y 1095: effect of glucose concentration on yield andproductivity. Enzyme and Microbial Technology1993;15:646–51.
[39] Moresi C. Effect of glucose concentration on citric acidproduction by Yarrowia lipolytica—kinetics of the tropho-phase, citrate lag phase and idiophase. Journal of ChemicalTechnology and Biotechnology 1994;60:387–95.
[40] Anastassiadis S, Aivasidis A, Wandrey C. Citric acid produc-tion by Candida strains under intracellular nitrogen limita-tion. Applied Microbiology and Biotechnology 2002;60:81–7.
[41] Crolla A, Kennedy KJ. In-line mixing for production of citricacid by Candida lipolytica grown on n-paraffins. Journal ofChemical Technology and Biotechnology 2004;79:720–8.
[42] Anastassiadis S, Rehm HJ. Continuous citric acid secretion bya high specific pH dependent active transport system in yeastCandida oleophila ATCC 20177. Electronic Journal of Biotech-nology 2005;8:146–61.
[43] Rymowicz W, Cibis E. Optimization of citric acid productionfrom glucose syrup by Yarrowia lipolytica using responsesurface methodology. Electronic Journal of Polish Agricul-tural Universities 2006;9:#20.
[44] Kamzolova S, Shishkanova N, Morgunov I, Finogenova T.Oxygen requirements for growth and citric acid productionof Yarrowia lipolytica. FEMS Yeast Research 2003;3:217–22.
[45] Kamzolova S, Morgunov I, Aurich A, Perevoznikova S,Shiskanova N, Stottmeister U, et al. Lipase secretion andcitric acid production in Yarrowia lipolytica yeast grown onanimal and vegetable fat. Food Technology and Biotechnol-ogy 2005;43:113–22.
[46] Chen HC, Liu TM. Inoculum effects on the production of g-linolenic acid by the shake culture of Cunninghamella echinulataCCRC 31840. Enzyme and Microbial Technology 1997;21:137–42.
[47] Certık M, Balteszova L, Sajbidor J. Lipid formation and g-linolenic acid production by Mucorales fungi grown onsunflower oil. Letters in Applied Microbiology 1997;25:101–5.
[48] Mantzouridou F, Tsimidou M, Roukas T. Performance ofcrude olive pomace oil and soybean oil during carotenoidproduction by Blakeslea trispora in submerged fermentation.Journal of Agricultural and Food Chemistry 2006;54:2575–81.
[49] Szczesna-Antczak M, Antczak T, Piotrowicz-Wasiak M,Rzyska M, Binkowska N, Bielecki S. Relationships betweenlipases and lipids in mycelia of two Mucor strains. Enzymeand Microbial Technology 2006;39:1214–22.
ARTICLE IN PRESS
B I O M A S S A N D B I O E N E R G Y 3 2 ( 2 0 0 8 ) 6 0 – 7 1 71




















