
Cholesterol Organization in Membranes at Low Concentrations: Effects of Curvature Stress and...
13
Cholesterol Organization in Membranes at Low Concentrations: Effects of Curvature Stress and Membrane Thickness R. Rukmini, Satinder S. Rawat, Samares C. Biswas, and Amitabha Chattopadhyay Centre for Cellular & Molecular Biology, Hyderabad 500 007, India ABSTRACT Cholesterol is often found distributed nonrandomly in domains in biological and model membranes and has been reported to be distributed heterogeneously among various intracellular membranes. Although a large body of literature exists on the organization of cholesterol in plasma membranes or membranes with high cholesterol content, very little is known about organization of cholesterol in membranes containing low amounts of cholesterol. Using a fluorescent cholesterol analog (25-[N-[(7-nitrobenz-2-oxa-1,3-diazol-4-yl)-methyl]amino]-27-norcholesterol, or NBD-cholesterol), we have previously shown that cholesterol may exhibit local organization even at very low concentrations in membranes, which could possibly be attributable to transbilayer tail-to-tail dimers. This is supported by similar observations reported by other groups using cholesterol or dehydroergosterol, a naturally occurring fluorescent cholesterol analog which closely mimics cholesterol. In this paper, we have tested the basic features of cholesterol organization in membranes at low concentrations using spectral features of dehydroergosterol. More importantly, we have investigated the role of membrane surface curvature and thickness on transbilayer dimer arrangement of cholesterol using NBD-cholesterol. We find that dimerization is not favored in membranes with high curvature. However, cholesterol dimers are observed again if the curvature stress is relieved. Further, we have monitored the effect of membrane thickness on the dimerization process. Our results show that the dimerization process is stringently controlled by a narrow window of membrane thickness. Interestingly, this type of local organization of NBD-cholesterol at low concentrations is also observed in sphingomyelin-containing membranes. These results could be significant in membranes that have very low cholesterol content, such as the endoplasmic reticulum and the inner mito- chondrial membrane, and in trafficking and sorting of cellular cholesterol. INTRODUCTION Cholesterol is a major constituent of eukaryotic membranes and plays a crucial role in membrane organization, dynamics, function and sorting (Yeagle, 1985; Liscum and Underwood, 1995; Schroeder et al., 1996; Fielding and Fielding, 1997; Liscum and Munn, 1999; Simons and Ikonen, 2000). The role of cholesterol in signal transduction is being increasingly rec- ognized (Incardona and Eaton, 2000). It has recently been implicated in embryonic development in Drosophila (Porter et al., 1996), and programmed cell death (Maccarrone et al., 1998). It also serves as a precursor for the synthesis of steroid hormones, bile acids, and lipoproteins. Cellular levels of cho- lesterol and its compartmentalization into different organelles are stringently controlled, and defects in its trafficking pathway have been shown to give rise to metabolic storage diseases such as Niemann-Pick type C (Mukherjee and Maxfield, 1999). Cholesterol is often found distributed nonrandomly in domains in biological and model membranes (Liscum and Underwood, 1995; Mukherjee and Chattopadhyay, 1996; Schroeder et al., 1995, 1996; Simons and Ikonen, 1997, 2000; Xu and London, 2000) and has been shown to preferentially interact with a subset of membrane lipids (McMullen and McElhaney, 1996) or proteins (Gimpl et al., 1997). Indeed, structurally and kinetically distinct cholesterol-rich and -poor domains have been detected in cellular membranes (Schroeder et al., 1991). Dynamic clustering of cholesterol (along with other membrane components such as sphingolipids) has given rise to the idea of specialized microdomains or “rafts” in the cell membrane that might play crucial roles in cellular func- tions such as signal transduction, adhesion, motility, and sort- ing and trafficking of membrane components (Simons and Ikonen, 1997, 2000). In addition, cholesterol has been reported to be distributed heterogeneously among various intracellular membranes. For example, along the biosynthetic route, the lowest cholesterol concentration is found in the membranes of the endoplasmic reticulum (ER), which, interestingly, is the site of cholesterol biosynthesis (Bretscher and Munro, 1993). Cholesterol concentration increases progressively along the cis-, medial-, and trans-Golgi stacks (Bretscher and Munro, 1993), whereas the highest concentration (;90% of the total cellular cholesterol) is found in the plasma membrane (Lange et al., 1989). Although a large body of literature exists on the organization of cholesterol in plasma membranes or membranes with high cholesterol content, very little is known about its organiza- tion in the membrane when the cholesterol content is very low (, 5 mol %), similar to what is found in ER and inner mitochondrial membranes (Lange et al., 1999). We have previously shown, using a fluorescent analog of cholesterol Received for publication 21 April 2000 and in final form 12 June 2001. Address reprint requests to Amitabha Chattopadhyay, Centre for Cellular & Molecular Biology, Uppal Road, Hyderabad 500 007, India. Tel.: 91-40-7172241; Fax: 91-40-7171195; E-mail: [email protected]. Dr. Rawai’s present address is Room 211, Building 469, Laboratory of Experimental and Computational Biology, NCI-FCRDC, National Insti- tutes of Health, Frederick, MD 21702-1201. Dr. Biswas’ present address is Institut National de la Recherche Agronomique, Unite ´ de Biochimie et Technologie des Proteines, Rue de la Ge ´raudie `re, BP 71627, 44316 Nantes Cedex 03, France © 2001 by the Biophysical Society 0006-3495/01/10/2122/13 $2.00 2122 Biophysical Journal Volume 81 October 2001 2122–2134
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Cholesterol Organization in Membranes at Low Concentrations: Effects of Curvature Stress and...

Cholesterol Organization in Membranes at Low Concentrations:Effects of Curvature Stress and Membrane Thickness
R. Rukmini, Satinder S. Rawat, Samares C. Biswas, and Amitabha ChattopadhyayCentre for Cellular & Molecular Biology, Hyderabad 500 007, India
ABSTRACT Cholesterol is often found distributed nonrandomly in domains in biological and model membranes and hasbeen reported to be distributed heterogeneously among various intracellular membranes. Although a large body of literatureexists on the organization of cholesterol in plasma membranes or membranes with high cholesterol content, very little isknown about organization of cholesterol in membranes containing low amounts of cholesterol. Using a fluorescent cholesterolanalog (25-[N-[(7-nitrobenz-2-oxa-1,3-diazol-4-yl)-methyl]amino]-27-norcholesterol, or NBD-cholesterol), we have previouslyshown that cholesterol may exhibit local organization even at very low concentrations in membranes, which could possiblybe attributable to transbilayer tail-to-tail dimers. This is supported by similar observations reported by other groups usingcholesterol or dehydroergosterol, a naturally occurring fluorescent cholesterol analog which closely mimics cholesterol. In thispaper, we have tested the basic features of cholesterol organization in membranes at low concentrations using spectralfeatures of dehydroergosterol. More importantly, we have investigated the role of membrane surface curvature and thicknesson transbilayer dimer arrangement of cholesterol using NBD-cholesterol. We find that dimerization is not favored inmembranes with high curvature. However, cholesterol dimers are observed again if the curvature stress is relieved. Further,we have monitored the effect of membrane thickness on the dimerization process. Our results show that the dimerizationprocess is stringently controlled by a narrow window of membrane thickness. Interestingly, this type of local organization ofNBD-cholesterol at low concentrations is also observed in sphingomyelin-containing membranes. These results could besignificant in membranes that have very low cholesterol content, such as the endoplasmic reticulum and the inner mito-chondrial membrane, and in trafficking and sorting of cellular cholesterol.
INTRODUCTION
Cholesterol is a major constituent of eukaryotic membranesand plays a crucial role in membrane organization, dynamics,function and sorting (Yeagle, 1985; Liscum and Underwood,1995; Schroeder et al., 1996; Fielding and Fielding, 1997;Liscum and Munn, 1999; Simons and Ikonen, 2000). The roleof cholesterol in signal transduction is being increasingly rec-ognized (Incardona and Eaton, 2000). It has recently beenimplicated in embryonic development inDrosophila(Porter etal., 1996), and programmed cell death (Maccarrone et al.,1998). It also serves as a precursor for the synthesis of steroidhormones, bile acids, and lipoproteins. Cellular levels of cho-lesterol and its compartmentalization into different organellesare stringently controlled, and defects in its trafficking pathwayhave been shown to give rise to metabolic storage diseasessuch as Niemann-Pick type C (Mukherjee and Maxfield,1999). Cholesterol is often found distributed nonrandomly indomains in biological and model membranes (Liscum andUnderwood, 1995; Mukherjee and Chattopadhyay, 1996;
Schroeder et al., 1995, 1996; Simons and Ikonen, 1997, 2000;Xu and London, 2000) and has been shown to preferentiallyinteract with a subset of membrane lipids (McMullen andMcElhaney, 1996) or proteins (Gimpl et al., 1997). Indeed,structurally and kinetically distinct cholesterol-rich and -poordomains have been detected in cellular membranes (Schroederet al., 1991). Dynamic clustering of cholesterol (along withother membrane components such as sphingolipids) has givenrise to the idea of specialized microdomains or “rafts” in thecell membrane that might play crucial roles in cellular func-tions such as signal transduction, adhesion, motility, and sort-ing and trafficking of membrane components (Simons andIkonen, 1997, 2000). In addition, cholesterol has been reportedto be distributed heterogeneously among various intracellularmembranes. For example, along the biosynthetic route, thelowest cholesterol concentration is found in the membranes ofthe endoplasmic reticulum (ER), which, interestingly, is thesite of cholesterol biosynthesis (Bretscher and Munro, 1993).Cholesterol concentration increases progressively along thecis-, medial-, andtrans-Golgi stacks (Bretscher and Munro,1993), whereas the highest concentration (;90% of the totalcellular cholesterol) is found in the plasma membrane(Lange et al., 1989).
Although a large body of literature exists on the organizationof cholesterol in plasma membranes or membranes withhighcholesterol content, very little is known about its organiza-tion in the membrane when the cholesterol content is verylow (, 5 mol %), similar to what is found in ER and innermitochondrial membranes (Lange et al., 1999). We havepreviously shown, using a fluorescent analog of cholesterol
Received for publication 21 April 2000 and in final form 12 June 2001.
Address reprint requests to Amitabha Chattopadhyay, Centre for Cellular& Molecular Biology, Uppal Road, Hyderabad 500 007, India. Tel.:91-40-7172241; Fax: 91-40-7171195; E-mail: [email protected].
Dr. Rawai’s present address is Room 211, Building 469, Laboratory ofExperimental and Computational Biology, NCI-FCRDC, National Insti-tutes of Health, Frederick, MD 21702-1201.
Dr. Biswas’ present address is Institut National de la RechercheAgronomique, Unite´ de Biochimie et Technologie des Proteines, Rue de laGeraudiere, BP 71627, 44316 Nantes Cedex 03, France
© 2001 by the Biophysical Society
0006-3495/01/10/2122/13 $2.00
2122 Biophysical Journal Volume 81 October 2001 2122–2134

(25-[N-[(7-nitrobenz-2-oxa-1,3-diazol-4-yl)-methyl]amino]-27-norcholesterol, or NBD-cholesterol) in which the flexi-ble acyl chain of cholesterol is labeled with the NBD group,that cholesterol may exhibit local organization even at verylow concentrations in membranes. Our results indicated thepossible presence of transbilayer tail-to-tail dimers of cho-lesterol in such membranes (Mukherjee and Chattopadhyay,1996). This is supported by similar observations reported byother groups obtained under comparable cholesterol con-centrations (Harris et al., 1995; Loura and Prieto, 1997).
Fluorescent sterols offer a powerful approach for moni-toring cholesterol organization and dynamics in membranesbecause of their high sensitivity, time resolution, and mul-tiplicity of measurable parameters (Schroeder, 1984; Gre-chishnikova et al., 1999). Sterols covalently linked to ex-trinsic fluorophores are commonly used for such studies.The advantage with this approach is that one has a choice ofthe fluorescent label to be used and, therefore, specificprobes with appropriate characteristics can be designed forspecific applications. A widely used extrinsic fluorophore inbiophysical, biochemical, and cell biological studies is theNBD group (Chattopadhyay, 1990). NBD-labeled lipids areextensively used as fluorescent analogs of native lipids inbiological and model membranes to study a variety ofprocesses (Kobayashi and Pagano, 1988; Chattopadhyay,1990; Mazeres et al., 1996; Liao et al., 1997; Chattopadhyayand Mukherjee, 1999).
The NBD moiety possesses some of the most desirableproperties for serving as an excellent probe for bothspectroscopic and microscopic applications. It is veryweakly fluorescent in water. Upon transfer to a hydro-phobic medium, it fluoresces brightly in the visible rangeand exhibits a high degree of environmental sensitivity(Chattopadhyay and London, 1988; Lin and Struve, 1991;Chattopadhyay and Mukherjee, 1993; Fery-Forgues etal., 1993; Mukherjee et al., 1994). Fluorescence lifetimeof the NBD group is extremely sensitive to the environ-mental polarity (Lin and Struve, 1991; Rawat and Chat-topadhyay, 1999). It is relatively photostable, and lipidslabeled with the NBD group mimic endogenous lipids instudies of intracellular lipid transport (van Meer et al.,1987; Koval and Pagano, 1990).
A detailed characterization of the spectroscopic and ion-ization properties as well as the specific location of NBD-cholesterol in membranes has earlier been carried out byone of us (Chattopadhyay and London, 1987, 1988). Inmodel membrane systems, the NBD group of NBD-choles-terol has been found to be localized deep in the hydrocarbonregion of the membrane, approximately 5–6 Å from thecenter of the bilayer (Chattopadhyay and London, 1987,1988; Mukherjee and Chattopadhyay, 1996). Unlike theNBD-labeled phospholipids, the NBD group in NBD-cho-lesterol does not loop back to the membrane interface de-spite its polar nature, probably because of the stereochem-ical rigidity of the sterol ring (Chattopadhyay, 1990). This
unique orientation of NBD-cholesterol offers a convenienthandle for localization of a polar and potentially reactivegroup deep inside the membrane. The unique position ofthis probe in the membrane has been previously exploited ina fluorescence resonance energy transfer study of the spatialrelationships of specific sites on chloroplast coupling factorto the membrane surface in reconstituted vesicles (Mitra andHammes, 1990). A closely related analog of this probe inwhich the NBD group is attached to a shorter alkyl chain(known as fluoresterol) has recently been shown to faith-fully mimic the biochemistry and cell biology of intestinalcholesterol absorption (Sparrow et al., 1999). When givenorally to hamsters, it was taken up into plasma, and theuptake was blocked by compounds that block cholesterolabsorption. In addition, fluoresterol was shown to be ester-ified both in vivo and in vitro (in cultured cells), indicatingthat it was able to traffic to the ER and act as a substrate foracyl CoA cholesterol acyltransferase, the enzyme that es-terifies native cholesterol. Thus, NBD-labeled cholesterolanalogs represent a useful class of molecules that mimicnative cholesterol properties in vitro and in vivo.
In this paper, we have investigated the role of membranesurface curvature and thickness on transbilayer dimerizationoccurring at low concentrations of cholesterol using NBD-cholesterol as a fluorescent cholesterol analog. Our resultsshow that the transbilayer dimer arrangement of cholesterolobserved at low concentrations in membranes (Harris et al.,1995; Mukherjee and Chattopadhyay, 1996; Loura and Pri-eto, 1997) is sensitive to the membrane surface curvatureand that cholesterol dimerization is not favored in mem-branes with high surface curvature. However, if the curva-ture stress is relieved by regenerating large vesicles, cho-lesterol dimers are observed again indicating the reversiblemodulation of the monomer/dimer equilibrium by mem-brane surface curvature. Further, we have examined theeffect of membrane thickness on the dimerization processusing NBD-cholesterol fluorescence. Our results indicatethat cholesterol dimers are formed in dipalmitoyl-sn-glyc-ero-3-phosphocholine (DPPC) and dimyristoyl-sn-glycero-3-phosphocholine (DMPC) membranes in the gel phase, butnot in distearoyl-sn-glycero-3-phosphocholine (DSPC) anddiarachidoyl-sn-glycero-3-phosphocholine (DAPC) mem-branes which are thicker membranes because of longer fattyacyl chains. Likewise, cholesterol dimers are not detected inthe relatively thin dilauroyl-sn-glycero-3-phosphocholine(DLPC) membranes. Thus, it seems that the dimerizationprocess is stringently controlled by a narrow window ofmembrane thickness. We also show that some of the basicfeatures of cholesterol organization in membranes at lowconcentrations obtained using NBD-cholesterol are inagreement with concentration-dependent changes observedin spectral features (peak intensity ratios) of dehydroergos-terol (D5,7,9(11)22-ergostatetraen-3b-ol, or DHE), a naturallyoccurring fluorescent cholesterol analog which closelymimics cholesterol.
Cholesterol Organization at Low Concentrations 2123
Biophysical Journal 81(4) 2122–2134

MATERIALS AND METHODS
Materials
DPPC, DMPC, and DHE were obtained from Sigma Chemical Co. (St.Louis, MO). DLPC, DSPC, DAPC, and brain sphingomyelin were fromAvanti Polar Lipids (Birmingham, AL). NBD-cholesterol was from Mo-lecular Probes (Eugene, OR). Lipids were checked for purity by thin-layerchromatography on precoated silica gel plates (Sigma) in chloroform/methanol/water (65:35:5, v/v) and were found to give only one spot in allcases with a phosphate-sensitive spray and on subsequent charring (Ditt-mer and Lester, 1964). Thin-layer chromatography of NBD-cholesterolwas done using the same solvents but in a slightly different proportion(65:35:4, v/v), and it was found to be pure when detected by its color orfluorescence (Mukherjee and Chattopadhyay, 1996). The concentrations ofthe phospholipids were determined by phosphate assay subsequent to totaldigestion by perchloric acid (McClare, 1971). DMPC was used as aninternal standard to assess lipid digestion. The concentration of NBD-cholesterol and DHE in methanolic stock solutions were calculated fromtheir respective molar extinction coefficients (e) of 22,000 M21 cm21 at484 nm (Chattopadhyay and Mukherjee, 1999) and 10,500 M21 cm21 at324 nm (Smutzer et al., 1986). All other chemicals used were of the highestpurity available. Solvents used were of spectroscopic grade. Water waspurified through a Millipore (Bedford, MA) Milli-Q system and usedthroughout.
Sample preparations
Experiments in which the membrane surface curvature was changed weredone with different types of vesicles of DPPC containing 2.0 mol %NBD-cholesterol. Samples containing 640 nmol (1280 nmol for time-resolved fluorescence measurements) of DPPC and 12.8 nmol (25.6 nmolfor time-resolved fluorescence measurements) of NBD-cholesterol weretaken together in methanol, a few drops of chloroform were added andmixed well, and dried under a stream of nitrogen while being warmedgently (;35°C). After the lipids were dried further under a high vacuumfor at least 3 h, they were hydrated (swelled) by adding 1.5 ml of 10 mMsodium phosphate, 150 mM sodium chloride, pH 7.2 buffer, and vortexedfor 3 min to disperse the lipid and form homogeneous multilamellarvesicles. The buffer was always maintained at a temperature well above thephase transition temperature of DPPC (i.e.,.41°C) as the vesicles weremade. Large unilamellar vesicles (LUVs) of 100-nm diameter were thenprepared by the extrusion technique using an Avestin Liposofast Extruder(Ottawa, Ontario, Canada) as previously described (MacDonald et al.,1991). Briefly, the multilamellar vesicles of DPPC were freeze-thawed fivetimes using liquid nitrogen to ensure solute equilibration between trappedand bulk solutions, and then extruded through polycarbonate filters (porediameter of 100 nm) mounted in the extruder fitted with Hamilton syringes(Hamilton Company, Reno, NV). The samples were subjected to 17 passesthrough the polycarbonate filter to give the final LUV suspension. Sampleswere kept overnight in dark at room temperature for equilibration beforemaking measurements. For experiments involving DHE, LUVs of DPPCcontaining various concentrations of DHE were prepared the same way.For experiments in which sphingomyelin was used, LUVs of DPPC/sphingomyelin (70:30, mol/mol) containing various concentrations ofNBD-cholesterol were also prepared this way.
Small unilamellar vesicles (SUVs) were generated from LUVs (afterfluorescence and/or absorbance measurements were made) by sonicationtill clarity using a Branson model 250 sonifier (Branson Ultrasonics,Dansbury, CT) fitted with a microtip. The sonicated samples were centri-fuged at 15,000 rpm for 15 min to remove any titanium particle shed fromthe microtip during sonication. Samples were kept overnight in dark atroom temperature for equilibration before making measurements.
To prepare regenerated LUVs (RLUV) from SUVs, the samples werefreeze-thawed five times using liquid nitrogen and then extruded through
polycarbonate filters (pore diameter of 100 nm) mounted in the AvestinLiposofast Extruder as previously described. Samples were kept overnightin dark at room temperature for equilibration before making measurements.Background samples were prepared the same way except that the fluoro-phore was not added to them. All the measurements were performed at23°C to ensure that DPPC is in the gel phase with the exception of theexperiments described in Figs. 10 and 11 in which the measurementsinvolving DPPC in the fluid phases were performed at 55°C.
All experiments in which the membrane thickness was changed weredone using LUVs made from 640 nmol of a phospholipid (DLPC, DMPC,DPPC, DSPC, or DAPC) containing 0.1 to 2.0% (mol/mol) NBD-choles-terol prepared by the extrusion procedure as described above. The temper-ature of the buffer during swelling of the dried lipid mixtures was alwaysmaintained above the corresponding phase transition temperature of thespecific lipid being used (Koynova and Caffrey, 1998) as the vesicles weremade. The phospholipids used were of varying chain lengths: DLPC(12:0), DMPC (14:0), DPPC (16:0), DSPC (18:0), and DAPC (20:0).Samples were kept overnight in dark at room temperature for equilibrationbefore making measurements.
Absorption measurements
The absorption spectra were obtained using a Hitachi U-2000 UV-visibleabsorption spectrophotometer (Hitachi, Ltd., Tokyo, Japan). Quartz cu-vettes with a path length of 1 cm were used. Background samples withoutthe fluorophore were used for baseline corrections.
Steady-state fluorescence measurements
Steady-state fluorescence measurements were performed with a HitachiF-4010 spectrofluorometer using 1-cm path length quartz cuvettes. Exci-tation and emission slits with a nominal bandpass of 5 nm were used for allmeasurements. The excitation wavelength used was 484 nm in all cases.For recording excitation spectra, emission wavelength was kept fixed at539 nm. Background intensities of samples in which NBD-cholesterol wasomitted were subtracted from each sample spectrum to cancel out anycontribution because of the solvent Raman peak and other scatteringartifacts. All experiments were done with multiple sets of samples.
Time-resolved fluorescence measurements
Fluorescence lifetimes were calculated from time-resolved fluorescenceintensity decays using a Photon Technology International (London, On-tario, Canada) LS-100 luminescence spectrophotometer in the time-corre-lated single photon counting mode. This machine uses a thyratron-gatednanosecond flash lamp filled with nitrogen as the plasma gas (166 1inches of mercury vacuum) and is run at 22–25 kHz. Lamp profiles weremeasured at the excitation wavelength using colloidal silica (Ludox; Du-pont, Wilmington, DE) as the scatterer. To optimize the signal to noiseratio, 5000 photon counts were collected in the peak channel. The excita-tion wavelength used was 337 nm which corresponds to a peak in thespectral output of the nitrogen flash lamp. Emission wavelength was set at522 or 539 nm. All experiments were performed using excitation andemission slits with a nominal bandpass of 4 nm or less. The sample and thescatterer were alternated after every 10% acquisition to ensure compensa-tion for shape and timing drifts occurring during the period of datacollection. The data stored in a multichannel analyzer was routinely trans-ferred to an IBM PC for analysis. Intensity decay curves so obtained werefitted as a sum of exponential terms:
F~t! 5 Oi
ai exp~2t/ti! (1)
2124 Rukmini et al.
Biophysical Journal 81(4) 2122–2134
whereai is a preexponential factor representing the fractional contributionto the time-resolved decay of the component with a lifetimeti. The decayparameters were recovered using a nonlinear least-squares iterative fittingprocedure based on the Marquardt algorithm (Bevington, 1969). Theprogram also includes statistical and plotting subroutine packages(O’Connor and Phillips, 1984). The goodness of the fit of a given set ofobserved data and the chosen function was evaluated by the reducedx2
ratio, the weighted residuals (Lampert et al., 1983), and the autocorrelationfunction of the weighted residuals (Grinvald and Steinberg, 1974). A fitwas considered acceptable when plots of the weighted residuals and theautocorrelation function showed random deviation about zero with a min-imum x2 value not more than 1.5. Mean (average) lifetimes^t& forbiexponential decays of fluorescence were calculated from the decay timesand preexponential factors using the following equation (Lakowicz, 1983):
^t& 5a1t1
2 1 a2t22
a1t1 1 a2t2(2)
RESULTS
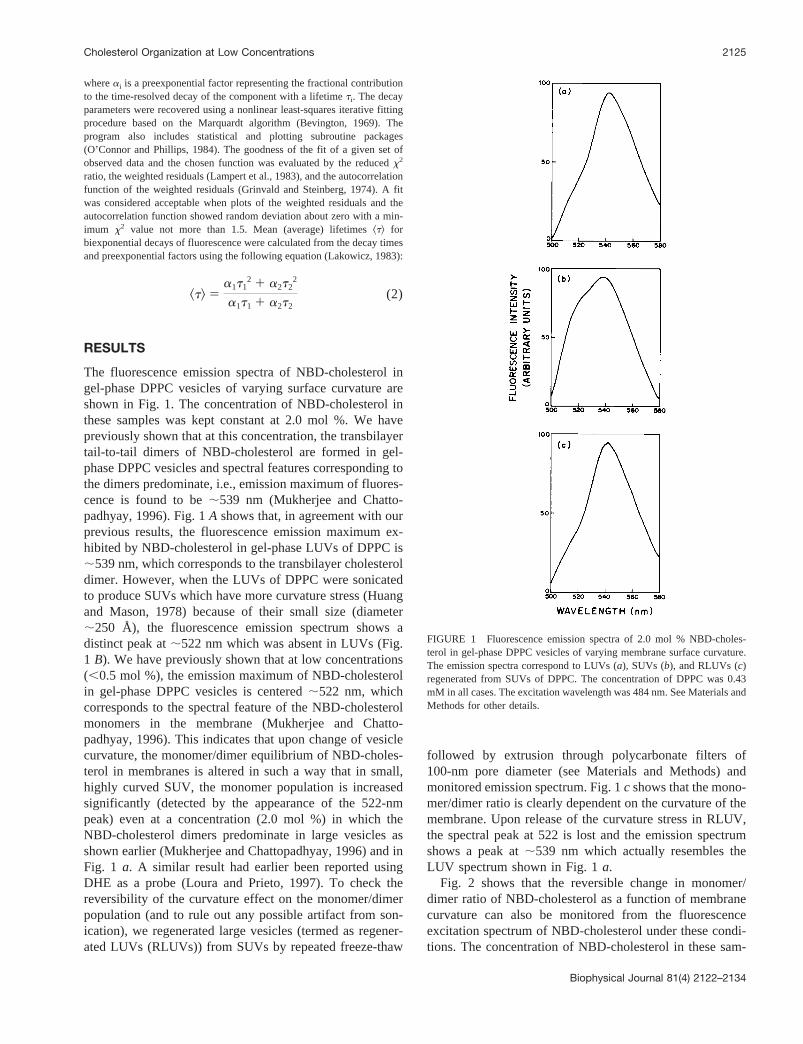
The fluorescence emission spectra of NBD-cholesterol ingel-phase DPPC vesicles of varying surface curvature areshown in Fig. 1. The concentration of NBD-cholesterol inthese samples was kept constant at 2.0 mol %. We havepreviously shown that at this concentration, the transbilayertail-to-tail dimers of NBD-cholesterol are formed in gel-phase DPPC vesicles and spectral features corresponding tothe dimers predominate, i.e., emission maximum of fluores-cence is found to be;539 nm (Mukherjee and Chatto-padhyay, 1996). Fig. 1A shows that, in agreement with ourprevious results, the fluorescence emission maximum ex-hibited by NBD-cholesterol in gel-phase LUVs of DPPC is;539 nm, which corresponds to the transbilayer cholesteroldimer. However, when the LUVs of DPPC were sonicatedto produce SUVs which have more curvature stress (Huangand Mason, 1978) because of their small size (diameter;250 Å), the fluorescence emission spectrum shows adistinct peak at;522 nm which was absent in LUVs (Fig.1 B). We have previously shown that at low concentrations(,0.5 mol %), the emission maximum of NBD-cholesterolin gel-phase DPPC vesicles is centered;522 nm, whichcorresponds to the spectral feature of the NBD-cholesterolmonomers in the membrane (Mukherjee and Chatto-padhyay, 1996). This indicates that upon change of vesiclecurvature, the monomer/dimer equilibrium of NBD-choles-terol in membranes is altered in such a way that in small,highly curved SUV, the monomer population is increasedsignificantly (detected by the appearance of the 522-nmpeak) even at a concentration (2.0 mol %) in which theNBD-cholesterol dimers predominate in large vesicles asshown earlier (Mukherjee and Chattopadhyay, 1996) and inFig. 1 a. A similar result had earlier been reported usingDHE as a probe (Loura and Prieto, 1997). To check thereversibility of the curvature effect on the monomer/dimerpopulation (and to rule out any possible artifact from son-ication), we regenerated large vesicles (termed as regener-ated LUVs (RLUVs)) from SUVs by repeated freeze-thaw
followed by extrusion through polycarbonate filters of100-nm pore diameter (see Materials and Methods) andmonitored emission spectrum. Fig. 1c shows that the mono-mer/dimer ratio is clearly dependent on the curvature of themembrane. Upon release of the curvature stress in RLUV,the spectral peak at 522 is lost and the emission spectrumshows a peak at;539 nm which actually resembles theLUV spectrum shown in Fig. 1a.
Fig. 2 shows that the reversible change in monomer/dimer ratio of NBD-cholesterol as a function of membranecurvature can also be monitored from the fluorescenceexcitation spectrum of NBD-cholesterol under these condi-tions. The concentration of NBD-cholesterol in these sam-
FIGURE 1 Fluorescence emission spectra of 2.0 mol % NBD-choles-terol in gel-phase DPPC vesicles of varying membrane surface curvature.The emission spectra correspond to LUVs (a), SUVs (b), and RLUVs (c)regenerated from SUVs of DPPC. The concentration of DPPC was 0.43mM in all cases. The excitation wavelength was 484 nm. See Materials andMethods for other details.
Cholesterol Organization at Low Concentrations 2125
Biophysical Journal 81(4) 2122–2134
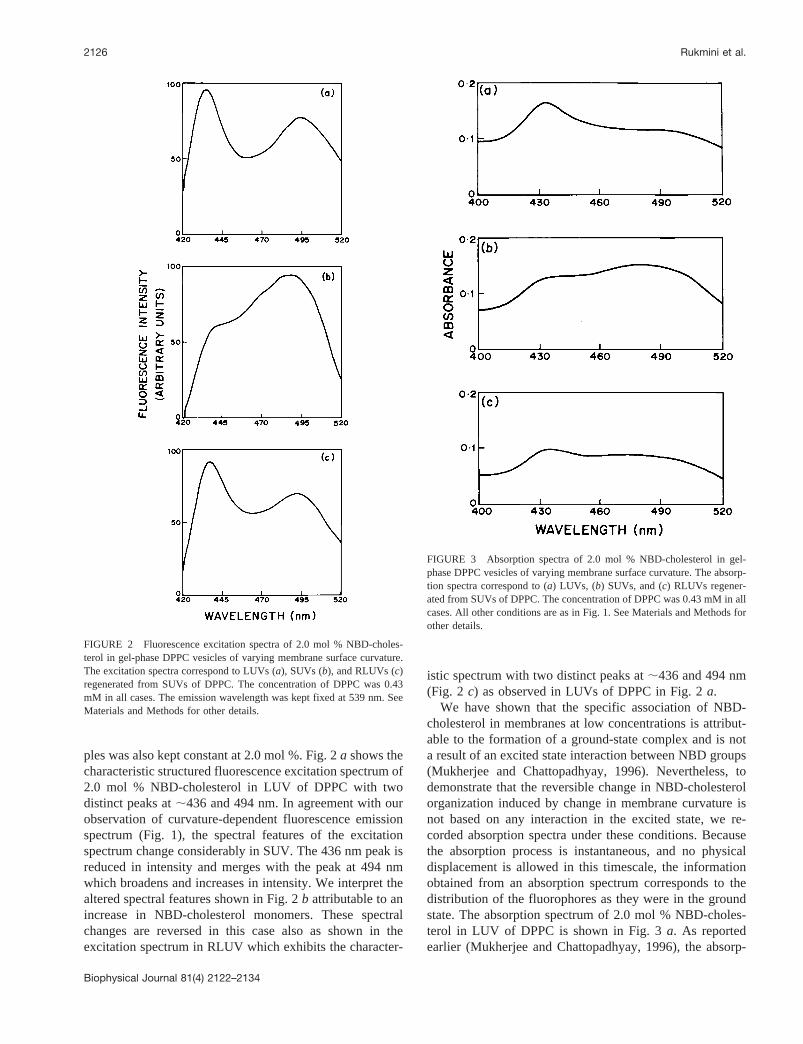
ples was also kept constant at 2.0 mol %. Fig. 2a shows thecharacteristic structured fluorescence excitation spectrum of2.0 mol % NBD-cholesterol in LUV of DPPC with twodistinct peaks at;436 and 494 nm. In agreement with ourobservation of curvature-dependent fluorescence emissionspectrum (Fig. 1), the spectral features of the excitationspectrum change considerably in SUV. The 436 nm peak isreduced in intensity and merges with the peak at 494 nmwhich broadens and increases in intensity. We interpret thealtered spectral features shown in Fig. 2b attributable to anincrease in NBD-cholesterol monomers. These spectralchanges are reversed in this case also as shown in theexcitation spectrum in RLUV which exhibits the character-
istic spectrum with two distinct peaks at;436 and 494 nm(Fig. 2 c) as observed in LUVs of DPPC in Fig. 2a.
We have shown that the specific association of NBD-cholesterol in membranes at low concentrations is attribut-able to the formation of a ground-state complex and is nota result of an excited state interaction between NBD groups(Mukherjee and Chattopadhyay, 1996). Nevertheless, todemonstrate that the reversible change in NBD-cholesterolorganization induced by change in membrane curvature isnot based on any interaction in the excited state, we re-corded absorption spectra under these conditions. Becausethe absorption process is instantaneous, and no physicaldisplacement is allowed in this timescale, the informationobtained from an absorption spectrum corresponds to thedistribution of the fluorophores as they were in the groundstate. The absorption spectrum of 2.0 mol % NBD-choles-terol in LUV of DPPC is shown in Fig. 3a. As reportedearlier (Mukherjee and Chattopadhyay, 1996), the absorp-
FIGURE 2 Fluorescence excitation spectra of 2.0 mol % NBD-choles-terol in gel-phase DPPC vesicles of varying membrane surface curvature.The excitation spectra correspond to LUVs (a), SUVs (b), and RLUVs (c)regenerated from SUVs of DPPC. The concentration of DPPC was 0.43mM in all cases. The emission wavelength was kept fixed at 539 nm. SeeMaterials and Methods for other details.
FIGURE 3 Absorption spectra of 2.0 mol % NBD-cholesterol in gel-phase DPPC vesicles of varying membrane surface curvature. The absorp-tion spectra correspond to (a) LUVs, (b) SUVs, and (c) RLUVs regener-ated from SUVs of DPPC. The concentration of DPPC was 0.43 mM in allcases. All other conditions are as in Fig. 1. See Materials and Methods forother details.
2126 Rukmini et al.
Biophysical Journal 81(4) 2122–2134
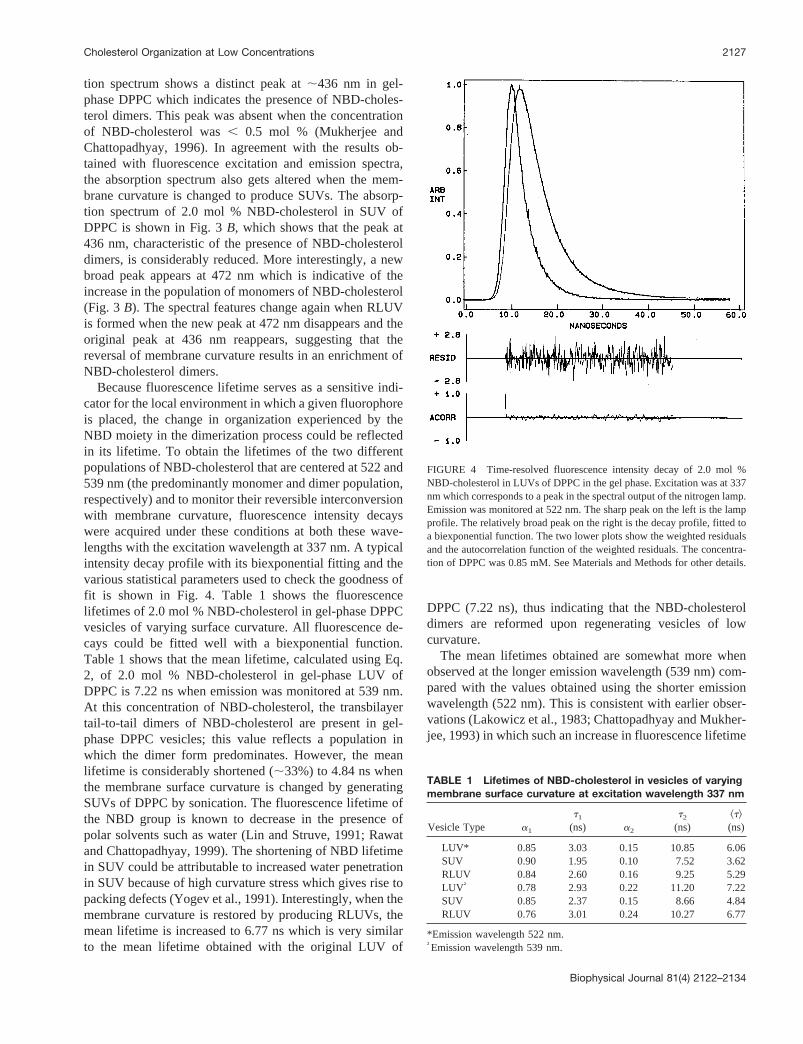
tion spectrum shows a distinct peak at;436 nm in gel-phase DPPC which indicates the presence of NBD-choles-terol dimers. This peak was absent when the concentrationof NBD-cholesterol was, 0.5 mol % (Mukherjee andChattopadhyay, 1996). In agreement with the results ob-tained with fluorescence excitation and emission spectra,the absorption spectrum also gets altered when the mem-brane curvature is changed to produce SUVs. The absorp-tion spectrum of 2.0 mol % NBD-cholesterol in SUV ofDPPC is shown in Fig. 3B, which shows that the peak at436 nm, characteristic of the presence of NBD-cholesteroldimers, is considerably reduced. More interestingly, a newbroad peak appears at 472 nm which is indicative of theincrease in the population of monomers of NBD-cholesterol(Fig. 3 B). The spectral features change again when RLUVis formed when the new peak at 472 nm disappears and theoriginal peak at 436 nm reappears, suggesting that thereversal of membrane curvature results in an enrichment ofNBD-cholesterol dimers.
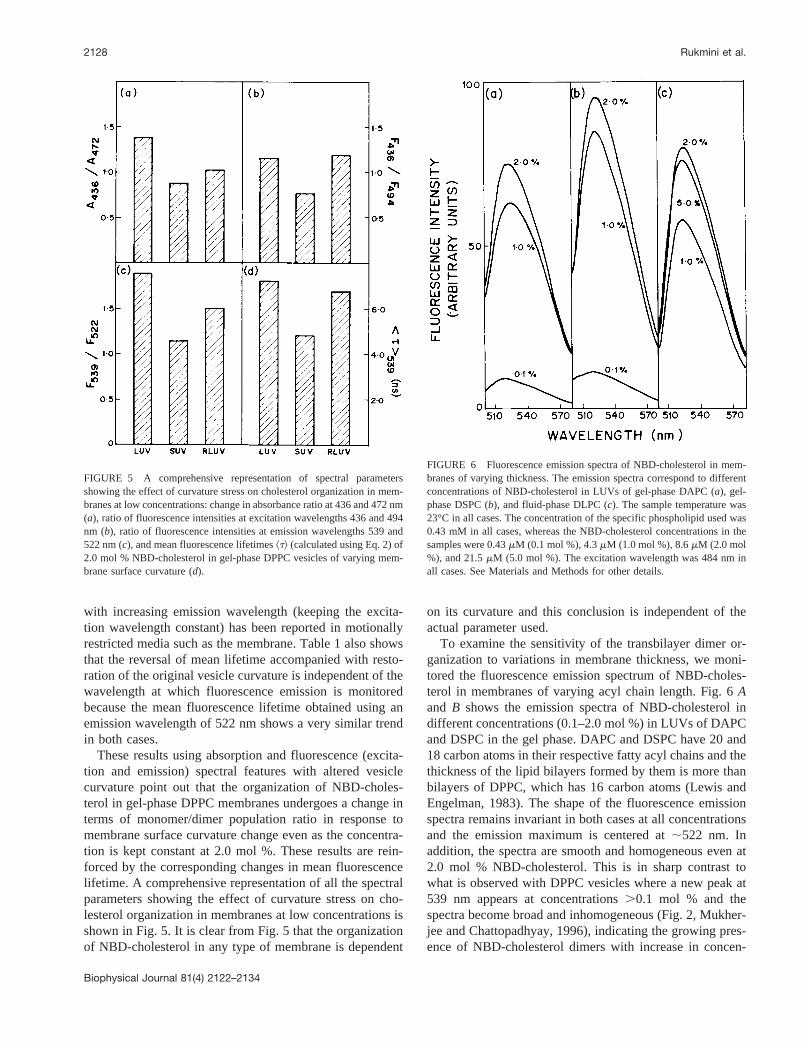
Because fluorescence lifetime serves as a sensitive indi-cator for the local environment in which a given fluorophoreis placed, the change in organization experienced by theNBD moiety in the dimerization process could be reflectedin its lifetime. To obtain the lifetimes of the two differentpopulations of NBD-cholesterol that are centered at 522 and539 nm (the predominantly monomer and dimer population,respectively) and to monitor their reversible interconversionwith membrane curvature, fluorescence intensity decayswere acquired under these conditions at both these wave-lengths with the excitation wavelength at 337 nm. A typicalintensity decay profile with its biexponential fitting and thevarious statistical parameters used to check the goodness offit is shown in Fig. 4. Table 1 shows the fluorescencelifetimes of 2.0 mol % NBD-cholesterol in gel-phase DPPCvesicles of varying surface curvature. All fluorescence de-cays could be fitted well with a biexponential function.Table 1 shows that the mean lifetime, calculated using Eq.2, of 2.0 mol % NBD-cholesterol in gel-phase LUV ofDPPC is 7.22 ns when emission was monitored at 539 nm.At this concentration of NBD-cholesterol, the transbilayertail-to-tail dimers of NBD-cholesterol are present in gel-phase DPPC vesicles; this value reflects a population inwhich the dimer form predominates. However, the meanlifetime is considerably shortened (;33%) to 4.84 ns whenthe membrane surface curvature is changed by generatingSUVs of DPPC by sonication. The fluorescence lifetime ofthe NBD group is known to decrease in the presence ofpolar solvents such as water (Lin and Struve, 1991; Rawatand Chattopadhyay, 1999). The shortening of NBD lifetimein SUV could be attributable to increased water penetrationin SUV because of high curvature stress which gives rise topacking defects (Yogev et al., 1991). Interestingly, when themembrane curvature is restored by producing RLUVs, themean lifetime is increased to 6.77 ns which is very similarto the mean lifetime obtained with the original LUV of
DPPC (7.22 ns), thus indicating that the NBD-cholesteroldimers are reformed upon regenerating vesicles of lowcurvature.
The mean lifetimes obtained are somewhat more whenobserved at the longer emission wavelength (539 nm) com-pared with the values obtained using the shorter emissionwavelength (522 nm). This is consistent with earlier obser-vations (Lakowicz et al., 1983; Chattopadhyay and Mukher-jee, 1993) in which such an increase in fluorescence lifetime
FIGURE 4 Time-resolved fluorescence intensity decay of 2.0 mol %NBD-cholesterol in LUVs of DPPC in the gel phase. Excitation was at 337nm which corresponds to a peak in the spectral output of the nitrogen lamp.Emission was monitored at 522 nm. The sharp peak on the left is the lampprofile. The relatively broad peak on the right is the decay profile, fitted toa biexponential function. The two lower plots show the weighted residualsand the autocorrelation function of the weighted residuals. The concentra-tion of DPPC was 0.85 mM. See Materials and Methods for other details.
TABLE 1 Lifetimes of NBD-cholesterol in vesicles of varyingmembrane surface curvature at excitation wavelength 337 nm
Vesicle Type a1
t1
(ns) a2
t2
(ns)^t&(ns)
LUV* 0.85 3.03 0.15 10.85 6.06SUV 0.90 1.95 0.10 7.52 3.62RLUV 0.84 2.60 0.16 9.25 5.29LUV† 0.78 2.93 0.22 11.20 7.22SUV 0.85 2.37 0.15 8.66 4.84RLUV 0.76 3.01 0.24 10.27 6.77
*Emission wavelength 522 nm.†Emission wavelength 539 nm.
Cholesterol Organization at Low Concentrations 2127
Biophysical Journal 81(4) 2122–2134
with increasing emission wavelength (keeping the excita-tion wavelength constant) has been reported in motionallyrestricted media such as the membrane. Table 1 also showsthat the reversal of mean lifetime accompanied with resto-ration of the original vesicle curvature is independent of thewavelength at which fluorescence emission is monitoredbecause the mean fluorescence lifetime obtained using anemission wavelength of 522 nm shows a very similar trendin both cases.
These results using absorption and fluorescence (excita-tion and emission) spectral features with altered vesiclecurvature point out that the organization of NBD-choles-terol in gel-phase DPPC membranes undergoes a change interms of monomer/dimer population ratio in response tomembrane surface curvature change even as the concentra-tion is kept constant at 2.0 mol %. These results are rein-forced by the corresponding changes in mean fluorescencelifetime. A comprehensive representation of all the spectralparameters showing the effect of curvature stress on cho-lesterol organization in membranes at low concentrations isshown in Fig. 5. It is clear from Fig. 5 that the organizationof NBD-cholesterol in any type of membrane is dependent
on its curvature and this conclusion is independent of theactual parameter used.
To examine the sensitivity of the transbilayer dimer or-ganization to variations in membrane thickness, we moni-tored the fluorescence emission spectrum of NBD-choles-terol in membranes of varying acyl chain length. Fig. 6Aand B shows the emission spectra of NBD-cholesterol indifferent concentrations (0.1–2.0 mol %) in LUVs of DAPCand DSPC in the gel phase. DAPC and DSPC have 20 and18 carbon atoms in their respective fatty acyl chains and thethickness of the lipid bilayers formed by them is more thanbilayers of DPPC, which has 16 carbon atoms (Lewis andEngelman, 1983). The shape of the fluorescence emissionspectra remains invariant in both cases at all concentrationsand the emission maximum is centered at;522 nm. Inaddition, the spectra are smooth and homogeneous even at2.0 mol % NBD-cholesterol. This is in sharp contrast towhat is observed with DPPC vesicles where a new peak at539 nm appears at concentrations.0.1 mol % and thespectra become broad and inhomogeneous (Fig. 2, Mukher-jee and Chattopadhyay, 1996), indicating the growing pres-ence of NBD-cholesterol dimers with increase in concen-
FIGURE 5 A comprehensive representation of spectral parametersshowing the effect of curvature stress on cholesterol organization in mem-branes at low concentrations: change in absorbance ratio at 436 and 472 nm(a), ratio of fluorescence intensities at excitation wavelengths 436 and 494nm (b), ratio of fluorescence intensities at emission wavelengths 539 and522 nm (c), and mean fluorescence lifetimes^t& (calculated using Eq. 2) of2.0 mol % NBD-cholesterol in gel-phase DPPC vesicles of varying mem-brane surface curvature (d).
FIGURE 6 Fluorescence emission spectra of NBD-cholesterol in mem-branes of varying thickness. The emission spectra correspond to differentconcentrations of NBD-cholesterol in LUVs of gel-phase DAPC (a), gel-phase DSPC (b), and fluid-phase DLPC (c). The sample temperature was23°C in all cases. The concentration of the specific phospholipid used was0.43 mM in all cases, whereas the NBD-cholesterol concentrations in thesamples were 0.43mM (0.1 mol %), 4.3mM (1.0 mol %), 8.6mM (2.0 mol%), and 21.5mM (5.0 mol %). The excitation wavelength was 484 nm inall cases. See Materials and Methods for other details.
2128 Rukmini et al.
Biophysical Journal 81(4) 2122–2134
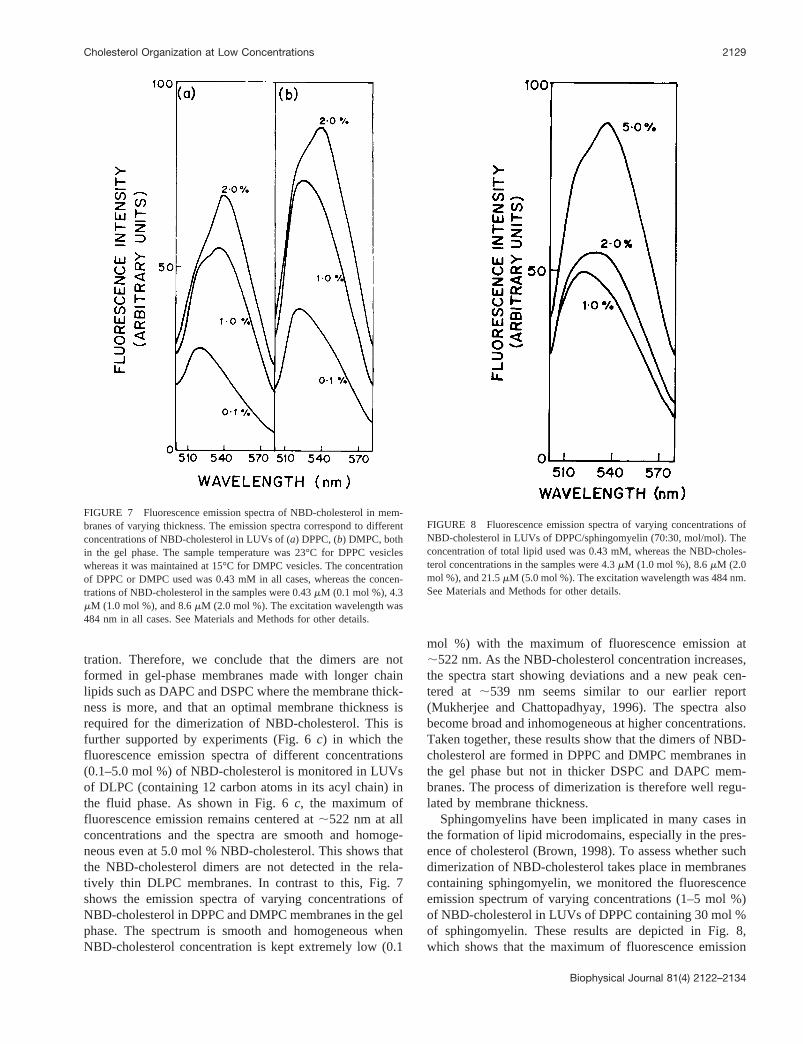
tration. Therefore, we conclude that the dimers are notformed in gel-phase membranes made with longer chainlipids such as DAPC and DSPC where the membrane thick-ness is more, and that an optimal membrane thickness isrequired for the dimerization of NBD-cholesterol. This isfurther supported by experiments (Fig. 6c) in which thefluorescence emission spectra of different concentrations(0.1–5.0 mol %) of NBD-cholesterol is monitored in LUVsof DLPC (containing 12 carbon atoms in its acyl chain) inthe fluid phase. As shown in Fig. 6c, the maximum offluorescence emission remains centered at;522 nm at allconcentrations and the spectra are smooth and homoge-neous even at 5.0 mol % NBD-cholesterol. This shows thatthe NBD-cholesterol dimers are not detected in the rela-tively thin DLPC membranes. In contrast to this, Fig. 7shows the emission spectra of varying concentrations ofNBD-cholesterol in DPPC and DMPC membranes in the gelphase. The spectrum is smooth and homogeneous whenNBD-cholesterol concentration is kept extremely low (0.1
mol %) with the maximum of fluorescence emission at;522 nm. As the NBD-cholesterol concentration increases,the spectra start showing deviations and a new peak cen-tered at ;539 nm seems similar to our earlier report(Mukherjee and Chattopadhyay, 1996). The spectra alsobecome broad and inhomogeneous at higher concentrations.Taken together, these results show that the dimers of NBD-cholesterol are formed in DPPC and DMPC membranes inthe gel phase but not in thicker DSPC and DAPC mem-branes. The process of dimerization is therefore well regu-lated by membrane thickness.
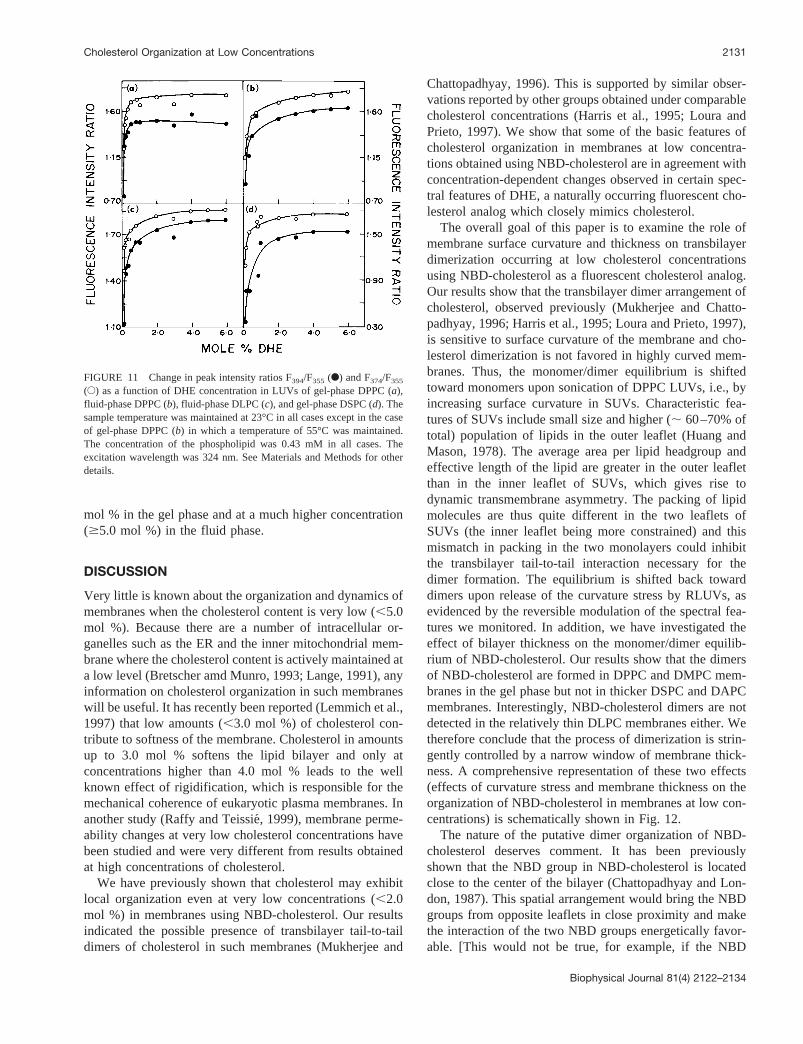
Sphingomyelins have been implicated in many cases inthe formation of lipid microdomains, especially in the pres-ence of cholesterol (Brown, 1998). To assess whether suchdimerization of NBD-cholesterol takes place in membranescontaining sphingomyelin, we monitored the fluorescenceemission spectrum of varying concentrations (1–5 mol %)of NBD-cholesterol in LUVs of DPPC containing 30 mol %of sphingomyelin. These results are depicted in Fig. 8,which shows that the maximum of fluorescence emission
FIGURE 7 Fluorescence emission spectra of NBD-cholesterol in mem-branes of varying thickness. The emission spectra correspond to differentconcentrations of NBD-cholesterol in LUVs of (a) DPPC, (b) DMPC, bothin the gel phase. The sample temperature was 23°C for DPPC vesicleswhereas it was maintained at 15°C for DMPC vesicles. The concentrationof DPPC or DMPC used was 0.43 mM in all cases, whereas the concen-trations of NBD-cholesterol in the samples were 0.43mM (0.1 mol %), 4.3mM (1.0 mol %), and 8.6mM (2.0 mol %). The excitation wavelength was484 nm in all cases. See Materials and Methods for other details.
FIGURE 8 Fluorescence emission spectra of varying concentrations ofNBD-cholesterol in LUVs of DPPC/sphingomyelin (70:30, mol/mol). Theconcentration of total lipid used was 0.43 mM, whereas the NBD-choles-terol concentrations in the samples were 4.3mM (1.0 mol %), 8.6mM (2.0mol %), and 21.5mM (5.0 mol %). The excitation wavelength was 484 nm.See Materials and Methods for other details.
Cholesterol Organization at Low Concentrations 2129
Biophysical Journal 81(4) 2122–2134

spectrum is centered at;522 nm when the concentration ofNBD-cholesterol is 1 mol %. However, the emission spectraat NBD-cholesterol concentrations.1 mol % display a shiftin emission maximum with an emission maximum of 539nm at 5 mol %, similar to what is observed with DPPCvesicles alone. This shows that the process of dimerizationalso takes place in sphingomyelin-containing membranes.
If the local organization exhibited by cholesterol in mem-branes of low cholesterol content is a general phenomenon,it should not be dependent on the probe chosen. To examinethis issue, we performed experiments in which we moni-tored an environment-sensitive spectral feature (peak inten-sity ratio) of DHE as a function of concentration. DHE is anaturally occurring fluorescent cholesterol analog which isfound in yeast and differs from cholesterol in having threeadditional double bonds and a methyl group (Fig. 9). Anumber of studies have shown that DHE faithfully mimicsnatural cholesterol in biophysical, biochemical, and cellbiological studies (Schroeder et al., 1991, 1995).
The fluorescence emission spectra of DHE in DPPCvesicles in fluid and gel phase are shown in Fig. 10. Acharacteristic feature of the structured emission spectra arethe maxima at 355, 374, and 394 nm, denoted as I, II, andIII, respectively. This type of structured vibronic band in-tensities are known to be sensitive to environment. This hasbeen effectively used for elucidating microenvironments ofthe hydrophobic fluorophore pyrene whose emission spec-trum shows similar vibronic fine structure (Kalyansundaramand Thomas, 1977). In particular, the ratio of emissionmaxima was shown to be sensitive to aggregation state ofthe fluorophore and environmental polarity (Kalyan-sundaram and Thomas, 1977). We used this sensitive spec-tral feature of DHE to monitor its change in local organi-zation at low concentrations in membranes. Fig. 11 showsthe change in peak intensity ratios (F394/F355 and F374/F355)
as a function of DHE concentration in LUV of gel and fluidphase DPPC (Fig. 11,a andb) and in fluid phase DLPC, andgel-phase DSPC (Fig. 11,c andd, respectively). The fluo-rescence intensity ratio initially increases with increasingDHE concentration in all cases and then attains a steadyvalue. We attribute this change in slope to the change inlocal organization of DHE. Fig. 11 shows that for gel-phaseDPPC membranes (a), the intensity ratio changes sharplyand assumes a steady value at DHE concentrations$1.0mol %. In contrast, in case of fluid-phase DPPC, fluid-phaseDLPC, or gel-phase DSPC vesicles, the change in slope ismuch more gradual and is observed at a much higherconcentration. The intensity ratio does not seem to assumea constant value even at higher concentrations in thesecases. This indicates that the change in local organization ofDHE occurs at a low concentration in gel-phase DPPCvesicles but not in the other cases. This is in excellentagreement with Fig. 6 and with our earlier observation usingNBD-cholesterol (Figs. 10 and 11, Mukherjee and Chatto-padhyay, 1996) where the appearance of a new fluorescenceor absorbance peak was observed at NBD-cholesterol$1.0
FIGURE 9 Chemical structure of DHE. The structure of cholesterol isshown in the inset for comparison.
FIGURE 10 Fluorescence emission spectra of 2.0 mol % DHE in LUVsof DPPC in fluid phase (a) and in gel phase (b). The sample temperaturewas maintained at 55°C for the fluid-phase experiment whereas the gel-phase experiment was performed at 23°C. The emission maxima at 355,374, and 394 nm are marked as I, II, and III, respectively. The concentra-tion of DPPC was 0.43 mM in both cases. The excitation wavelength was324 nm. See Materials and Methods for other details.
2130 Rukmini et al.
Biophysical Journal 81(4) 2122–2134
mol % in the gel phase and at a much higher concentration($5.0 mol %) in the fluid phase.
DISCUSSION
Very little is known about the organization and dynamics ofmembranes when the cholesterol content is very low (,5.0mol %). Because there are a number of intracellular or-ganelles such as the ER and the inner mitochondrial mem-brane where the cholesterol content is actively maintained ata low level (Bretscher amd Munro, 1993; Lange, 1991), anyinformation on cholesterol organization in such membraneswill be useful. It has recently been reported (Lemmich et al.,1997) that low amounts (,3.0 mol %) of cholesterol con-tribute to softness of the membrane. Cholesterol in amountsup to 3.0 mol % softens the lipid bilayer and only atconcentrations higher than 4.0 mol % leads to the wellknown effect of rigidification, which is responsible for themechanical coherence of eukaryotic plasma membranes. Inanother study (Raffy and Teissie´, 1999), membrane perme-ability changes at very low cholesterol concentrations havebeen studied and were very different from results obtainedat high concentrations of cholesterol.
We have previously shown that cholesterol may exhibitlocal organization even at very low concentrations (,2.0mol %) in membranes using NBD-cholesterol. Our resultsindicated the possible presence of transbilayer tail-to-taildimers of cholesterol in such membranes (Mukherjee and
Chattopadhyay, 1996). This is supported by similar obser-vations reported by other groups obtained under comparablecholesterol concentrations (Harris et al., 1995; Loura andPrieto, 1997). We show that some of the basic features ofcholesterol organization in membranes at low concentra-tions obtained using NBD-cholesterol are in agreement withconcentration-dependent changes observed in certain spec-tral features of DHE, a naturally occurring fluorescent cho-lesterol analog which closely mimics cholesterol.
The overall goal of this paper is to examine the role ofmembrane surface curvature and thickness on transbilayerdimerization occurring at low cholesterol concentrationsusing NBD-cholesterol as a fluorescent cholesterol analog.Our results show that the transbilayer dimer arrangement ofcholesterol, observed previously (Mukherjee and Chatto-padhyay, 1996; Harris et al., 1995; Loura and Prieto, 1997),is sensitive to surface curvature of the membrane and cho-lesterol dimerization is not favored in highly curved mem-branes. Thus, the monomer/dimer equilibrium is shiftedtoward monomers upon sonication of DPPC LUVs, i.e., byincreasing surface curvature in SUVs. Characteristic fea-tures of SUVs include small size and higher (; 60–70% oftotal) population of lipids in the outer leaflet (Huang andMason, 1978). The average area per lipid headgroup andeffective length of the lipid are greater in the outer leafletthan in the inner leaflet of SUVs, which gives rise todynamic transmembrane asymmetry. The packing of lipidmolecules are thus quite different in the two leaflets ofSUVs (the inner leaflet being more constrained) and thismismatch in packing in the two monolayers could inhibitthe transbilayer tail-to-tail interaction necessary for thedimer formation. The equilibrium is shifted back towarddimers upon release of the curvature stress by RLUVs, asevidenced by the reversible modulation of the spectral fea-tures we monitored. In addition, we have investigated theeffect of bilayer thickness on the monomer/dimer equilib-rium of NBD-cholesterol. Our results show that the dimersof NBD-cholesterol are formed in DPPC and DMPC mem-branes in the gel phase but not in thicker DSPC and DAPCmembranes. Interestingly, NBD-cholesterol dimers are notdetected in the relatively thin DLPC membranes either. Wetherefore conclude that the process of dimerization is strin-gently controlled by a narrow window of membrane thick-ness. A comprehensive representation of these two effects(effects of curvature stress and membrane thickness on theorganization of NBD-cholesterol in membranes at low con-centrations) is schematically shown in Fig. 12.
The nature of the putative dimer organization of NBD-cholesterol deserves comment. It has been previouslyshown that the NBD group in NBD-cholesterol is locatedclose to the center of the bilayer (Chattopadhyay and Lon-don, 1987). This spatial arrangement would bring the NBDgroups from opposite leaflets in close proximity and makethe interaction of the two NBD groups energetically favor-able. [This would not be true, for example, if the NBD
FIGURE 11 Change in peak intensity ratios F394/F355 (F) and F374/F355
(E) as a function of DHE concentration in LUVs of gel-phase DPPC (a),fluid-phase DPPC (b), fluid-phase DLPC (c), and gel-phase DSPC (d). Thesample temperature was maintained at 23°C in all cases except in the caseof gel-phase DPPC (b) in which a temperature of 55°C was maintained.The concentration of the phospholipid was 0.43 mM in all cases. Theexcitation wavelength was 324 nm. See Materials and Methods for otherdetails.
Cholesterol Organization at Low Concentrations 2131
Biophysical Journal 81(4) 2122–2134

group looped up in the bilayer as is often seen with NBD-labeled phospholipids with NBD groups placed at the end ofthe fatty acyl chain (Chattopadhyay and London, 1988).]Fluorophores, when placed close to each other without anyspecific pattern (spatial arrangement) of organization, oftengive rise to self-quenching (Chattopadhyay et al., 1992). Infact, concentration-dependent self-quenching of the NBDgroup is well characterized and constitutes the basis of anumber of assays in membrane biology because this type ofquenching is very sensitive to local surface density of lipidsin the membrane (Nichols and Pagano, 1981; Chatto-padhyay, 1990). It is interesting that the association of theNBD groups from the opposing leaflets in this case does notgive rise to self-quenching, but, instead, gives rise to a newpeak and other well defined spectral features in the fluores-cence and absorption spectra (Figs. 1–3). The unique spec-tral features imply that the two NBD groups are not onlyclose in space but have a definite spatial orientation whichwould have been lacking otherwise. That this is a ground-state phenomenon is supported by the concomitant changesin absorption spectrum with changes in fluorescence spec-tra. An important factor contributing to the stability of theNBD-cholesterol dimers is aromatic-aromatic interactionbetween the NBD rings. However, the possibility of othertypes of association cannot be ruled out from the presentdata. Moreover, at this point it is not clear whether thedimers can lead to further aggregation. The formation ofsuch transmembrane dimers could represent the first step inthe ultimate separation of a pure cholesterol domain.
The transbilayer tail-to-tail cholesterol organization couldhave a role in diseased states. The importance of cholesteroltransbilayer domains in atherogenesis was recently demon-strated by Tulenko et al. (1998). In this study, small anglex-ray diffraction was used to examine arterial smooth mus-cle cell plasma membranes isolated from control and cho-lesterol-fed (2.0%) atherosclerotic rabbits. Beyond 9 weeksof cholesterol feeding, x-ray diffraction patterns demon-strated a lateral immiscible cholesterol domain with a bi-layer thickness of 34 Å coexisting with the liquid crystalline(fluid) lipid bilayer. The membrane thickness of 34 Å cor-responds to a tail-to-tail arrangement of cholesterol dimersas the length of an individual cholesterol molecule is 17 Å(Craven, 1976). This points out the possible involvement ofthe transbilayer tail-to-tail organization of cholesterol inatherogenesis.
Such cholesterol domains found in membranes of lowcholesterol content (such as the ER or the inner mitochon-drial membrane) could have important functional implica-tions in intracellular sorting and trafficking. The low cho-lesterol content of the ER membrane has been attributed toits need to be readily deformable in order for newly syn-thesized membrane proteins to be folded and assembledproperly (Bretscher and Munro, 1993). It has been sug-gested that the specific gradient of cholesterol concentrationfound in Golgi and ER facilitates protein sorting in thesecretory pathway via cholesterol’s ability to modify themembrane thickness (Pelham and Munro, 1993). Membranesoftening caused by low levels of cholesterol (Lemmich etal., 1997) may play a role in the vesicle budding processassociated with membrane traffic in Golgi and ER. Choles-terol is removed from ER, its site of synthesis, by using themembrane traffic pathway where membranes move fromthe ER to the plasma membrane through the Golgi appara-tus. Cholesterol is known to modulate lipid-protein interac-tions by increasing the thickness of the lipid bilayer. Forexample, it has been shown that 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine vesicles have a bilayer thicknessof ;26 Å which increases to 30 Å in the presence of 30 mol% cholesterol (Nezil and Bloom, 1992). In fact, there isevidence that a gradient of cholesterol exists in Golgi andthe membrane coming from the Golgi apparatus is believedto be thicker and to have more cholesterol than the mem-brane entering from the ER. Because our results show thatcholesterol dimerization is controlled by membrane thick-ness, this could imply that there is a gradient of cholesteroldimers across the Golgi apparatus.
It is noteworthy that many important cellular traffickingevents (such as vesiculation, tubulation, pinching off ofendocytic vesicles, and fusion of exocytic vesicles) areaccompanied by sharp changes in membrane curvature(Zimmerberg, 2000). The overall process of membrane traf-ficking and turnover is influenced by membrane curvature,e.g., vesicles budding from ER or Golgi may have moresurface curvature than their parent membrane. Curvature-
FIGURE 12 A schematic diagram of the membrane bilayer showing theeffects of curvature stress and membrane thickness on the organization ofNBD-cholesterol in membranes at low concentrations. (a) The transbilayertail-to-tail dimers of NBD-cholesterol proposed earlier (Mukherjee andChattopadhyay, 1996). (b) NBD-cholesterol monomers in membranes withhigh surface curvature such as SUVs. (c) NBD-cholesterol monomers inrelatively thick membranes formed by phospholipids having longer fattyacyl chains. (d) NBD-cholesterol monomers in relatively thin membranesformed by phospholipids having shorter fatty acyl chains. See text for otherdetails.
2132 Rukmini et al.
Biophysical Journal 81(4) 2122–2134

induced membrane domains have recently been reported tobe responsible for differential sorting and trafficking of lipidanalogs in CHO cells (Mukherjee et al., 1999). Some of thevesicles involved in cellular traffic, e.g., coated vesiclesused in exocytosis, are quite small (;500 Å diameter,comparable with SUV dimensions) and are highly curved(Aridor and Balch, 1996). Certain regions of sorting endo-somes experience high curvature stress (Mukherjee et al.,1999). The endosomal vesicles also experience low (acidic)pH (Mukherjee et al., 1997). Interestingly, it has recentlybeen reported that pH has a strong effect on the membranecurvature (Lee et al., 1999).
In summary, we have investigated the role of membranesurface curvature and thickness on transbilayer dimerizationoccurring at low concentrations of cholesterol using NBD-cholesterol as a fluorescent cholesterol analog. Our resultsshow that the transbilayer dimer arrangement of cholesterolpreviously observed at low concentrations in membranes issensitive to the membrane surface curvature and is strin-gently controlled by a narrow window of membrane thick-ness. These results are significant in the context of choles-terol organization in membranes at low concentrations inparticular, and in the dynamics and regulation of cellularcholesterol, in general.
We thank Y. S. S. V. Prasad and G. G. Kingi for technical help. This workwas supported by the Council of Scientific and Industrial Research, Gov-ernment of India. S.S.R. thanks the Council of Scientific and IndustrialResearch for the award of a Senior Research Fellowship. S.C.B. thanks theDepartment of Biotechnology, Government of India, for the award of apostdoctoral fellowship. We thank Dr. K. G. Harikumar for criticallyreading the manuscript.
REFERENCES
Aridor, M., and W. E. Balch. 1996. Principles of selective transport: coatcomplexes hold the key.Trends Cell Biol.6:315–320.
Bevington, P. R. 1969. Data Reduction and Error Analysis for the PhysicalSciences. McGraw-Hill, New York.
Bretscher, M. S., and S. Munro. 1993. Cholesterol and the Golgi apparatus.Science.61:1280–1281.
Brown, R. E. 1998. Sphingolipid organization in biomembranes: whatphysical studies of model membranes reveal.J. Cell Sci.111:1–9.
Chattopadhyay, A. 1990. Chemistry and biology ofN-(7-nitrobenz-2-oxa-1, 3-diazol-4-yl)-labeled lipids: fluorescent probes of biological andmodel membranes.Chem. Phys. Lipids.53:1–15.
Chattopadhyay, A., and E. London. 1987. Parallax method for directmeasurement of membrane penetration depth utilizing fluorescencequenching by spin-labeled phospholipids.Biochemistry.26:39–45.
Chattopadhyay, A., and E. London. 1988. Spectroscopic and ionizationproperties ofN-(7-nitrobenz-2-oxa-1, 3-diazol-4-yl)-labeled lipids inmodel membranes.Biochim. Biophys. Acta.938:24–34.
Chattopadhyay, A., and S. Mukherjee. 1993. Fluorophore environments inmembrane-bound probes: a red edge excitation shift study.Biochemis-try. 32:3804–3811.
Chattopadhyay, A., and S. Mukherjee. 1999. Red edge excitation shift of adeeply embedded membrane probe: implications in water penetration inthe bilayer.J. Phys. Chem. B.103:8180–8185.
Chattopadhyay, A., S. S. Komath, and B. Raman. 1992. Aggregation oflasalocid A in membranes: a fluorescence study.Biochim. Biophys. Acta.1104:147–150.
Craven, B. M. 1976. Crystal structure of cholesterol monohydrate.Nature.260:727–729.
Dittmer, J. C., and R. L. Lester. 1964. A simple, specific spray for thedetection of phospholipids on thin-layer chromatograms.J. Lipid Res.5:126–127.
Fielding, C. J., and P. E. Fielding. 1997. Intracellular cholesterol transport.J. Lipid Res.38:1503–1520.
Fery-Forgues, S., J.-P. Fayet, and A. Lopez. 1993. Drastic changes in thefluorescence properties of NBD probes with the polarity of the medium:involvement of a TICT state?J. Photochem. Photobiol. A.70:229–243.
Gimpl, G., K. Burger, and F. Fahrenholz. 1997. Cholesterol as modulatorof receptor function.Biochemistry.36:10959–10974.
Grechishnikova, IV, F. Bergstrom, L. B.-A. Johansson, R. E. Brown, andJ. G. Molotkovsky. 1999. New fluorescent cholesterol analogs as mem-brane probes.Biochim. Biophys. Acta.1420:189–202.
Grinvald, A., and I. Z. Steinberg. 1974. On the analysis of fluorescencedecay kinetics by the method of least-squares.Anal. Biochem.59:583–598.
Harris, J. S., D. E. Epps, S. R. Davio, and F. J. Kezdy. 1995. Evidence fortransbilayer, tail-to-tail cholesterol dimers in dipalmitoylglycerophos-phocholine liposomes.Biochemistry.34:3851–3857.
Huang, C., and J. T. Mason. 1978. Geometric packing constraints in eggphosphatidylcholine vesicles.Proc. Natl. Acad. Sci. U.S.A.75:308–310.
Incardona, J. P., and S. Eaton. 2000. Cholesterol in signal transduction.Curr. Opin. Cell Biol.12:193–203.
Kalyansundaram, K., and J. K. Thomas. 1977. Environmental effects onvibronic band intensities in pyrene monomer fluorescence and theirapplication in studies of micellar systems.J. Am. Chem. Soc.99:2039–2044.
Kobayashi, T., and R. E. Pagano. 1988. ATP-dependent fusion of lipo-somes with the Golgi apparatus of perforated cells.Cell. 55:797–805.
Koval, M., and R. E. Pagano. 1990. Sorting of an internalized plasmamembrane lipid between recycling and degradative pathways in normaland Niemann-Pick, type A fibroblasts.J. Cell. Biol.111:429–442.
Koynova, R., and M. Caffrey. 1998. Phases and phase transitions of thephosphatidylcholines.Biochim. Biophys. Acta.1376:91–145.
Lakowicz, J. R. 1983. Principles of Fluorescence Spectroscopy. PlenumPress, New York.
Lakowicz, J. R., D. R. Bevan, B. P. Maliwal, H. Cherek, and A. Balter.1983. Synthesis and characterization of a fluorescent probe of the phasetransition and dynamic properties of membranes.Biochemistry.22:5714–5722.
Lampert, R. A., L. A. Chewter, D. Phillips, D. V. O’Connor, A. J. Roberts,and S. R. Meech. 1983. Standards for nanosecond fluorescence decaytime measurements.Anal. Chem.55:68–73.
Lange, Y. 1991. Disposition of intracellular cholesterol in human fibro-blasts.J. Lipid Res.33:329–339.
Lange, Y., M. H. Swaisgood, B. V. Ramos, and T. L. Stack. 1989. Plasmamembranes contain half the phospholipid and 90% of the cholesterol andsphingomyelin in cultured human fibroblasts.J. Biol. Chem.264:3786–3793.
Lange, Y., J. Ye, M. Rigney, and T. L. Stack. 1999. Regulation of ERcholesterol by plasma membrane cholesterol.J. Lipid Res. 40:2264–2270.
Lee, J. B., P. G. Petrov, and H.-G. Do¨bereiner. 1999. Curvature of zwit-terionic membranes in transverse pH gradients.Langmuir. 15:8543–8546.
Lemmich, J., K. Mortensen, J. H. Ipsen, T. Hønger, R. Bauer, and O. G.Mouritsen. 1997. The effect of cholesterol in small amounts on lipid-bilayer softness in the region of the main phase transition.Eur. Bio-phys. J.25:293–304.
Lewis, B. A., and D. M. Engelman. 1983. Lipid bilayer thickness varieslinearly with acyl chain length in fluid phosphatidylcholine vesicles.J. Mol. Biol. 166:211–217.
Cholesterol Organization at Low Concentrations 2133
Biophysical Journal 81(4) 2122–2134

Liao, S., J. Lin, H. Do, and A. E. Johnson. 1997. Both lumenal andcytosolic gating of the aqueous ER translocon pore are regulated frominside the ribosome during membrane protein integration.Cell. 90:31–41.
Lin, S., and W. S. Struve. 1991. Time-resolved fluorescence of nitroben-zoxadiazole-aminohexanoic acid: effect of intermolecular hydrogen-bonding on non-radiative decay.Photochem. Photobiol.54:361–365.
Liscum, L., and K. W. Underwood. 1995. Intracellular cholesterol transportand compartmentalization.J. Biol. Chem.270:15443–15446.
Liscum, L., and N. J. Munn. 1999. Intracellular cholesterol transport.Biochim. Biophys. Acta.1438:19–37.
Loura, L. M. S., and M. Prieto. 1997. Dehydroergosterol structural orga-nization in aqueous medium and in a model system of membranes.Biophys. J.72:2226–2236.
Maccarrone, M., L. Bellincampi, G. Melino, and A. Finazzi Agro. 1998.Cholesterol, but not its esters, triggers programmed cell death in humanerythroleukemia K562 cells.Eur. J. Biochem.263:107–113.
MacDonald, R. C., R. I. MacDonald, B. P. Menco, K. Takeshita, N. K.Subbarao, and L. R. Hu. 1991. Small-volume extrusion apparatus forpreparation of large, unilamellar vesicles.Biochim. Biophys. Acta.1061:297–303.
Mazeres, S., V. Schram, J.-F. Tocanne, and A. Lopez. 1996. 7-Nitrobenz-2-oxa-1,3-diazole-4-yl-labeled phospholipids in lipid membranes: dif-ferences in fluorescence behavior.Biophys. J.71:327–335.
McClare, C. W. F. 1971. An accurate and convenient organic phosphorusassay.Anal. Biochem.39:527–530.
McMullen, T. P. W., and R. N. McElhaney. 1996. Physical studies ofcholesterol-phospholipid interactions.Curr. Opin. Colloid Interface Sci.1:83–90.
Mitra, B., and G. G. Hammes. 1990. Membrane-protein structural mappingof chloroplast coupling factor in asolectin vesicles.Biochemistry.29:9879–9884.
Mukherjee, S., and A. Chattopadhyay. 1996. Membrane organization atlow cholesterol concentrations: a study using 7-nitrobenz-2-oxa-1,3-diazol-4-yl-labeled cholesterol.Biochemistry.35:1311–1322.
Mukherjee, S., A. Chattopadhyay, A. Samanta, and T. Soujanya. 1994.Dipole moment change of NBD group upon excitation using solvato-chromic and quantum chemical approaches: implications in membraneresearch.J. Phys. Chem.98:2809–2812.
Mukherjee, S., R. N. Ghosh, and F. R. Maxfield. 1997. Endocytosis.Physiol. Rev.77:759–803.
Mukherjee, S., and F. R. Maxfield. 1999. Cholesterol: stuck in traffic.Nature Cell Biol.1:E37–E38.
Mukherjee, S., Soe, T. T., and F. R. Maxfield. 1999. Endocytic sorting oflipid analogs differing solely in the chemistry of their hydrophobic tails.J. Cell Biol. 144:1271–1284.
Nezil, F. A., and M. Bloom. 1992. Combined influence of cholesterol andsynthetic amphiphilic peptides upon bilayer thickness in model mem-branes.Biophys. J.61:1176–1183.
Nichols, J. W., and R. E. Pagano. 1981. Kinetics of soluble lipid monomerdiffusion between vesicles.Biochemistry.20:2783–2789.
O’Connor, D. V., and D. Phillips. 1984. Time-Correlated Single PhotonCounting. Academic Press, London. 180–189.
Pelham, H. R. B., and S. Munro. 1993. Sorting of membrane proteins in thesecretory pathway.Cell. 75:603–605.
Porter, J. A., K. E. Young, and P. A. Beachy. 1996. Cholesterol modifi-cation of hedgehog signalling proteins in animal development.Science.274:255–259.
Raffy, S., and J. Teissie´. 1999. Control of lipid membrane stability bycholesterol content.Biophys. J.76:2072–2080.
Rawat, S. S., and A. Chattopadhyay. 1999. Structural transition in themicellar assembly: a fluorescence study.J. Fluorescence.9:233–244.
Schroeder, F. 1984. Fluorescent sterols: probe molecules of membranestructure and function.Prog. Lipid Res.23:97–113.
Schroeder, F., A. A. Frolov, E. J. Murphy, B. P. Atshaves, J. R. Jefferson,L. Pu, W. G. Wood, W. B. Foxworth, and A. B. Kier. 1996. Recentadvances in membrane cholesterol domain dynamics and intracellularcholesterol trafficking.Proc. Soc. Exp. Biol. Med.213:150–177.
Schroeder, F., J. R. Jefferson, A. B. Kier, J. Knittel, T. J. Scallen, W. G.Wood, and I. Hapala. 1991. Membrane cholesterol dynamics: choles-terol domains and kinetic pools.Proc. Soc. Exp. Biol. Med.196:235–252.
Schroeder, F., J. K. Woodford, J. Kavecansky, W. G. Wood, and C. Joiner.1995. Cholesterol domains in biological membranes.Mol. Membr. Biol.12:113–119.
Simons, K., and E. Ikonen. 1997. Functional rafts in cell membranes.Nature.387:569–572.
Simons, K., and E. Ikonen. 2000. How cells handle cholesterol.Science.290:1721–1726.
Smutzer, G., B. F. Crawford, and P. L. Yeagle. 1986. Physical propertiesof the fluorescent sterol probe dehydroergosterol.Biochim. Biophys.Acta.862:361–371.
Sparrow, C. P., S. Patel, J. Baffic, Y.-S. Chao, M. Hernandez, M.-H. Lam,J. Montenegro, S. D. Wright, and P. A. Detmers. 1999. A fluorescentcholesterol analog traces cholesterol absorption in hamsters and is es-terified in vivo and in vitro.J. Lipid Res.40:1747–1757.
Tulenko, T. N., M. Chen, P. E. Mason, and R. P. Mason. 1998. Physicaleffects of cholesterol on arterial smooth muscle membranes: evidence ofimmiscible cholesterol domains and alterations in bilayer width duringatherogenesis.J. Lipid Res.39:947–956.
Van Meer, G., E. H. Stelzer, R. W. Wijnaendts-van-Resandt, and K.Simons. 1987. Sorting of sphingolipids in epithelial (Madin-Darby ca-nine kidney) cells.J. Cell. Biol.105:1623–1635.
Xu, X., and E. London. 2000. The effect of sterol structure on membranelipid domains reveals how cholesterol can induce lipid domain forma-tion. Biochemistry.39:843–849.
Yeagle, P. L. 1985. Cholesterol and the cell membrane.Biochim. Biophys.Acta.822:267–287.
Yogev, D., A. T. Todorov, and J. H. Fendlar. 1991. Fluorescence lifetimesof diphenylhexatriene in flat and bent bilayer lipid membranes.J. Phys.Chem.95:3892–3894.
Zimmerberg, J. 2000. Are the curves in all the right places?Traffic.1:366–368.
2134 Rukmini et al.
Biophysical Journal 81(4) 2122–2134




















