Charged liposome affects the translation and folding steps of in vitro expression of green...
6
This article was originally published in a journal published by Elsevier, and the attached copy is provided by Elsevier for the author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues that you know, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at: http://www.elsevier.com/locate/permissionusematerial
Transcript of Charged liposome affects the translation and folding steps of in vitro expression of green...

This article was originally published in a journal published by Elsevier, and the attached copy is provided by Elsevier for the
author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use including without
limitation use in instruction at your institution, sending it to specific colleagues that you know, and providing a copy to your institution’s
administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access,
or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission
may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial

Author's Personal Copy
Journal of Bioscience and BioengineeringVOL. 108 No. 5, 450–454, 2009
www.elsevier.com/locate/jbiosc
Charged liposome affects the translation and folding steps of in vitro expression ofgreen fluorescent protein
Hiroshi Umakoshi, Keishi Suga, Huong Thi Bui, Masato Nishida, Toshinori Shimanouchi, and Ryoichi Kuboi⁎
⁎ CorrespondE-mail add
1389-1723/$doi:10.1016/j
Department of Chemical Science and Engineering, Graduate School of Engineering Science, Osaka University 1-3 Machikaneyama-cho,Toyonaka, Osaka 560-8531, Japan
Received 9 March 2009; accepted 19 May 2009
The role of the charged liposome on the in vitro expression of green fluorescent protein (GFP) was investigated, focusingon its elemental steps such as transcription, translation and folding. The total GFP expression was enhanced to 145% when aneutral liposome (POPC: 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocoline) was added externally to a cell-free translationsystem. On the contrary, the addition of the charged liposome composed of POPC with anionic 1-palmitoyl-2-oleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)] (POPG) or cationic stearyl amine (SA) inhibited the total GFP expression, depending onthe surface charge density of liposome. In transcription, the RNA synthesis was enhanced regardless of the variation of thesurface charge, indicating that transcription was enhanced due to the stabilization of RNA structure by its hydrophobicinteraction with liposome. Translation was inhibited by cationic liposome although it was enhanced by anionic liposome andneutral liposome. On the other hand, the folding was not inhibited in the presence of neutral liposome, whereas anionicliposome and cationic liposome inhibited the folding in proportion to the their surface charges, suggesting that the total GFPexpression was controlled by a charged liposome in the translation step and folding step.
© 2009, The Society for Biotechnology, Japan. All rights reserved.
[Key words: Liposome; Green fluorescent protein; In vitro gene expression; Transcription; Translation; Folding]
A biomembrane of the cell acts as a physical barrier that separatesthe biomolecules within a cell structure from the external environ-ment. In addition, the biomembrane protects inner compartmentsfrom environmental stresses, such as heat stress, pH changes, andoxidative stress. Since the biomembranes are composed of a variety ofamphiphiles, such as phospholipids, cholesterol, and so on, they oftenshow some surface properties, i.e. membrane fluidity, raft (micro-domain), surface charge densities, etc., which can be related to theinteraction with biomolecules; proteins (enzymes), DNAs, and RNAs.Liposome, a model biomembrane, has been reported to induce avariety of potential functions under the stress conditions. For example,it has been shown that the liposome reactivates fragmented super-oxide dismutase (SOD) under an oxidative condition in the presenceof Cu2+ and Zn2+ (1). Liposome modified with Mn–PhP exerts bothSOD activity and peroxidase activity (2), and negatively-chargedliposome refolds lysozyme effectively like a molecular chaperone (3).
A central dogma in molecular biology shows that the geneexpression is constructed in at least three sequential steps, such astranscription, translation, and folding. In the case of cell-free proteinexpression system, the DNA plasmid is first transcribed into mRNA byT7 RNA polymerase, and then translated to unfolded polypeptide onribosome (4). The unfolded peptide of green fluorescent protein (GFP)is automatically folded to mature protein by its own backbone
ing author. Tel./fax: +81 6 6850 6285.ress: [email protected] (R. Kuboi).
- see front matter © 2009, The Society for Biotechnology, Japan. All.jbiosc.2009.05.012
structure (5). In spite of much information from the viewpoint of thegenome and proteome being gradually accumulated, the role of themodel biomembrane on the gene expression has not been clarifiedyet. There have been several reports relating to gene expressionwithina model biomembrane (liposome) (6–10), although the liposomewasutilized just as a physical barrier. We have already reported that theliposome membrane itself can affect the gene expression in theEscherichia coli cell-free translation system (11). The total GFPexpression was enhanced in the presence of a zwitterionic (non-charged or neutral) liposome (POPC/cholesterol), suggesting that theliposome can affect the step of protein synthesis, including theelementary steps such as transcription, translation, and folding.
In the present study, the potential functions of charged liposomeon the total GFP expression were investigated, focusing on theelementary steps (transcription, translation, and folding), in order togain a deeper understanding of the role of the cell membrane. Thetotal GFP expression was performed in the presence of chargedliposomes with different compositions: neutral POPC, anionic POPC/POPG, and cationic POPC/SA.
MATERIALS AND METHODS
Materials 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphocoline (POPC) and 1-pal-mitoyl-2-oleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)] (POPG) were purchased fromAvanti Polar Lipids (Alabaster, AL). A Rapid Translation System RTS 100 E. coli HY Kit(RTS-Kit) and recombinant GFP (1 mg/ml) were purchased from Roche Diagnostics(Indianapolis, IN). DNase I and the SV Total RNA Isolation Systemwere purchased fromPromega (Madison, WI). Other chemicals of commercially guaranteed reagent grade
rights reserved.
Author's Personal Copy
TABLE 1. Surface charge density and membrane fluidity of charged liposomes.
Liposome composition Surface charge densitya
[C/nm2]Membrane fluidity,
1/Pb [−]
POPC/POPG 70/30 −0.42 5.72±0.36POPC/POPG 93/7 −0.098 5.74±0.79POPC 0 s 5.58±0.42POPC/SA 88/12 0.182 5.34±0.33POPC/SA 70/30 0.532 4.23±0.07
a The surface charge density was calculated assuming unilamellarity, spherical shape,a bilayer thickness of 3.7 nm, and a mean head group area of POPC, POPG, and SA were0.72 nm2, 0.72 nm2, 0.20 nm2, respectively (14,17), and considering that the phosphategroup of POPG and SA are fully ionized at pH8.0 (15,16).
b The membrane fluidity was evaluated from reciprocal polarity of DPH (1/P).Polarization of DPH (Ex=360 nm, Em=430 nm) was calculated using the followingequation. P= IO � I8ð Þ= IO + I8ð Þ
GENE EXPRESSION VIA MEMBRANE INTERACTION 451VOL. 108, 2009
were purchased from Wako Pure Chemicals (Osaka, Japan), and were used withoutfurther purification.
Liposome preparation A solution of POPC, POPC/POPG, and POPC/SA (0–30 mol%) in chloroformwas dried in a round-bottom flask by rotary evaporation underreduced pressure. The lipid films obtained were dissolved in chloroform twice, and thesolvent was evaporated. The lipid film was kept under a high vacuum for at least 3 h,and then hydrated with 0.1 M Tris–HCl buffer (pH 8.0) at room temperature to formmultilamellar vesicles (MLVs). The vesicle suspension was frozen in a refrigerator(−80 °C) and thawed at 37 °C to enhance the transformation of small vesicles intolarger MLVs. This freeze–thaw cycle was performed five times. MLVs were used toprepare the small unilamellar vesicles (SUVs) by extruding the MLV suspension 11times through two layers of polycarbonate membranes with mean pore diameters of100 nm using an extruding device (Liposofast; Avestin Inc., Ottawa, Canada).
In vitro green fluorescent protein (GFP) expression with liposome TheRapid Translation System (RTS-Kit), an in vitro translation system based on E. colilysate, was used for cell-free protein synthesis (4). The pIVEX2.3d, containing the GFPgene, was used as an expression vector. The reaction solution (9.6 μl) contained thefollowing components: 2 μl E. coli lysate, 2 μl reaction mix, 2.4 μl amino acids solution,0.2 μl methionine solution,1 μl reconstitution buffer, and 0.1 μg of circular DNA templatein 2 μl nuclease-free water. The reaction was initiated by mixing the reaction solutionswith or without liposome (final lipid concentration is 1.17 mM) at 30 °C for 6 h and waskept at 4 °C for 24 h. The amount of GFP synthesized using RTS-Kit was evaluated by thefluorescence of GFP. The total GFP expression in the presence or absence of chargedliposomewas evaluated based on the GFP fluorescence (Ex=395 nm, Em=509 nm) byusing the fluorescence spectrophotometer (FP-6500; JASCO, Tokyo, Japan) with a slitwidth of 5 nm at excitation and emission of light path.
Evaluation of membrane fluidity of charged liposome To measure the outermembrane fluidity, a fluorescent probe, 1,3,5-hexatriene (DPH), was added to theliposome solution (molar ratio; liposome/DPH 250/1). The fluorescence polarization ofDPH (Ex=360 nm, Em=430 nm) measured after 1 h incubation at 30 °C, by using thefluorescence spectrophotometer (FP-6500; JASCO, Tokyo, Japan). The sample wasexcited with vertically polarized light (360 nm), and emission intensities (430 nm)both parallel (IO) and perpendicular (I⊥) to excited light were recorded. Then, thepolarization of DPH was calculated based on the following equation (P).
P= IO � I8ð Þ= IO + I8ð Þ
The membrane fluidity was evaluated based on the reciprocal of polarization (1/P).Total RNA isolation: Evaluation of transcription mRNAwas synthesized using
an RTS-Kit without amino acids by incubation for 3 h at 30 °C in the presence or absenceof liposome. Synthesized RNA was purified with an SV Total RNA Isolation System(Promega) (12). The reaction solution containing RNA was diluted in lysis buffer, andthen RNA was denatured for 3 min at 70 °C. The denatured RNA in solution wasseparated by a centrifuge (14,000 ×g) for 10 min, and then the upper phase was addedto a spin column assembly. RNA binding to spin column was eluted in nuclease-freewater. The purity of isolated RNA was determined by the absorbance ratio of A260/A280
(13), and isolated RNA was analyzed by UV absorbance A260 by using a UVspectrophotometer (UV-160A; SHIMADZU, Kyoto, Japan). The effect of chargedliposome was evaluated by comparing it with the control value (without liposome).
Time-chase translation of GFP peptide: Evaluation of translation The time-chase translation of GFP was achieved in order to investigate the effect of chargedliposome on the translation step of GFP synthesis. mRNAwas first transcribed according
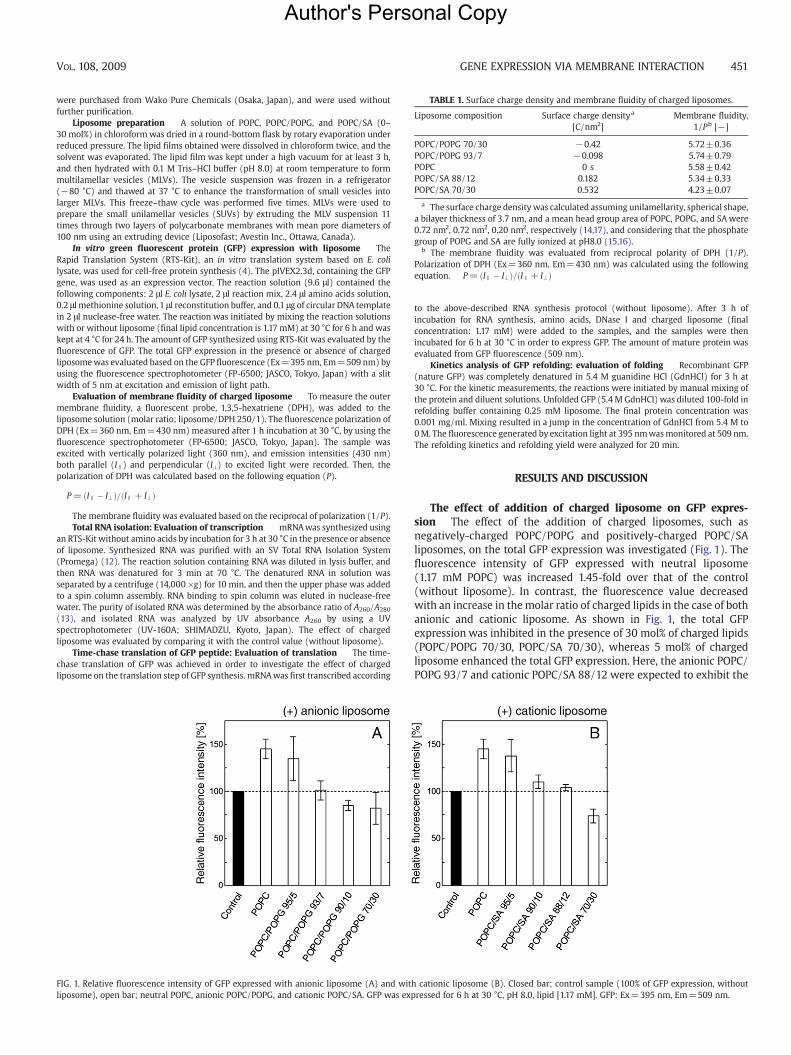
FIG. 1. Relative fluorescence intensity of GFP expressed with anionic liposome (A) and witliposome), open bar; neutral POPC, anionic POPC/POPG, and cationic POPC/SA. GFP was exp
to the above-described RNA synthesis protocol (without liposome). After 3 h ofincubation for RNA synthesis, amino acids, DNase I and charged liposome (finalconcentration: 1.17 mM) were added to the samples, and the samples were thenincubated for 6 h at 30 °C in order to express GFP. The amount of mature protein wasevaluated from GFP fluorescence (509 nm).
Kinetics analysis of GFP refolding: evaluation of folding Recombinant GFP(nature GFP) was completely denatured in 5.4 M guanidine HCl (GdnHCl) for 3 h at30 °C. For the kinetic measurements, the reactions were initiated by manual mixing ofthe protein and diluent solutions. Unfolded GFP (5.4 M GdnHCl) was diluted 100-fold inrefolding buffer containing 0.25 mM liposome. The final protein concentration was0.001 mg/ml. Mixing resulted in a jump in the concentration of GdnHCl from 5.4 M to0M. The fluorescence generated by excitation light at 395 nmwasmonitored at 509 nm.The refolding kinetics and refolding yield were analyzed for 20 min.
RESULTS AND DISCUSSION
The effect of addition of charged liposome on GFP expres-sion The effect of the addition of charged liposomes, such asnegatively-charged POPC/POPG and positively-charged POPC/SAliposomes, on the total GFP expression was investigated (Fig. 1). Thefluorescence intensity of GFP expressed with neutral liposome(1.17 mM POPC) was increased 1.45-fold over that of the control(without liposome). In contrast, the fluorescence value decreasedwith an increase in the molar ratio of charged lipids in the case of bothanionic and cationic liposome. As shown in Fig. 1, the total GFPexpression was inhibited in the presence of 30 mol% of charged lipids(POPC/POPG 70/30, POPC/SA 70/30), whereas 5 mol% of chargedliposome enhanced the total GFP expression. Here, the anionic POPC/POPG 93/7 and cationic POPC/SA 88/12 were expected to exhibit the
h cationic liposome (B). Closed bar; control sample (100% of GFP expression, withoutressed for 6 h at 30 °C, pH 8.0, lipid [1.17 mM]. GFP: Ex=395 nm, Em=509 nm.
Author's Personal Copy
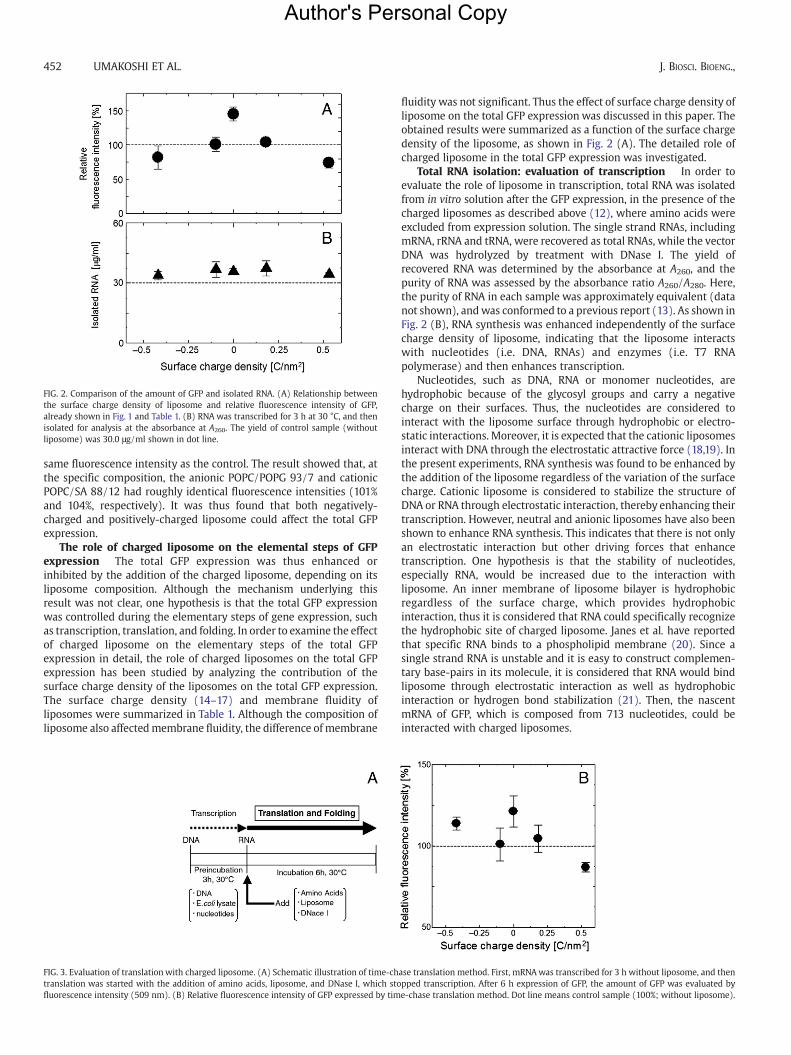
FIG. 2. Comparison of the amount of GFP and isolated RNA. (A) Relationship betweenthe surface charge density of liposome and relative fluorescence intensity of GFP,already shown in Fig. 1 and Table 1. (B) RNA was transcribed for 3 h at 30 °C, and thenisolated for analysis at the absorbance at A260. The yield of control sample (withoutliposome) was 30.0 μg/ml shown in dot line.
452 UMAKOSHI ET AL. J. BIOSCI. BIOENG.,
same fluorescence intensity as the control. The result showed that, atthe specific composition, the anionic POPC/POPG 93/7 and cationicPOPC/SA 88/12 had roughly identical fluorescence intensities (101%and 104%, respectively). It was thus found that both negatively-charged and positively-charged liposome could affect the total GFPexpression.
The role of charged liposome on the elemental steps of GFPexpression The total GFP expression was thus enhanced orinhibited by the addition of the charged liposome, depending on itsliposome composition. Although the mechanism underlying thisresult was not clear, one hypothesis is that the total GFP expressionwas controlled during the elementary steps of gene expression, suchas transcription, translation, and folding. In order to examine the effectof charged liposome on the elementary steps of the total GFPexpression in detail, the role of charged liposomes on the total GFPexpression has been studied by analyzing the contribution of thesurface charge density of the liposomes on the total GFP expression.The surface charge density (14–17) and membrane fluidity ofliposomes were summarized in Table 1. Although the composition ofliposome also affectedmembrane fluidity, the difference of membrane
FIG. 3. Evaluation of translationwith charged liposome. (A) Schematic illustration of time-chtranslation was started with the addition of amino acids, liposome, and DNase I, which stofluorescence intensity (509 nm). (B) Relative fluorescence intensity of GFP expressed by tim
fluidity was not significant. Thus the effect of surface charge density ofliposome on the total GFP expression was discussed in this paper. Theobtained results were summarized as a function of the surface chargedensity of the liposome, as shown in Fig. 2 (A). The detailed role ofcharged liposome in the total GFP expression was investigated.
Total RNA isolation: evaluation of transcription In order toevaluate the role of liposome in transcription, total RNA was isolatedfrom in vitro solution after the GFP expression, in the presence of thecharged liposomes as described above (12), where amino acids wereexcluded from expression solution. The single strand RNAs, includingmRNA, rRNA and tRNA, were recovered as total RNAs, while the vectorDNA was hydrolyzed by treatment with DNase I. The yield ofrecovered RNA was determined by the absorbance at A260, and thepurity of RNA was assessed by the absorbance ratio A260/A280. Here,the purity of RNA in each sample was approximately equivalent (datanot shown), andwas conformed to a previous report (13). As shown inFig. 2 (B), RNA synthesis was enhanced independently of the surfacecharge density of liposome, indicating that the liposome interactswith nucleotides (i.e. DNA, RNAs) and enzymes (i.e. T7 RNApolymerase) and then enhances transcription.
Nucleotides, such as DNA, RNA or monomer nucleotides, arehydrophobic because of the glycosyl groups and carry a negativecharge on their surfaces. Thus, the nucleotides are considered tointeract with the liposome surface through hydrophobic or electro-static interactions. Moreover, it is expected that the cationic liposomesinteract with DNA through the electrostatic attractive force (18,19). Inthe present experiments, RNA synthesis was found to be enhanced bythe addition of the liposome regardless of the variation of the surfacecharge. Cationic liposome is considered to stabilize the structure ofDNA or RNA through electrostatic interaction, thereby enhancing theirtranscription. However, neutral and anionic liposomes have also beenshown to enhance RNA synthesis. This indicates that there is not onlyan electrostatic interaction but other driving forces that enhancetranscription. One hypothesis is that the stability of nucleotides,especially RNA, would be increased due to the interaction withliposome. An inner membrane of liposome bilayer is hydrophobicregardless of the surface charge, which provides hydrophobicinteraction, thus it is considered that RNA could specifically recognizethe hydrophobic site of charged liposome. Janes et al. have reportedthat specific RNA binds to a phospholipid membrane (20). Since asingle strand RNA is unstable and it is easy to construct complemen-tary base-pairs in its molecule, it is considered that RNA would bindliposome through electrostatic interaction as well as hydrophobicinteraction or hydrogen bond stabilization (21). Then, the nascentmRNA of GFP, which is composed from 713 nucleotides, could beinteracted with charged liposomes.
ase translation method. First, mRNAwas transcribed for 3 h without liposome, and thenpped transcription. After 6 h expression of GFP, the amount of GFP was evaluated bye-chase translation method. Dot line means control sample (100%; without liposome).
Author's Personal Copy
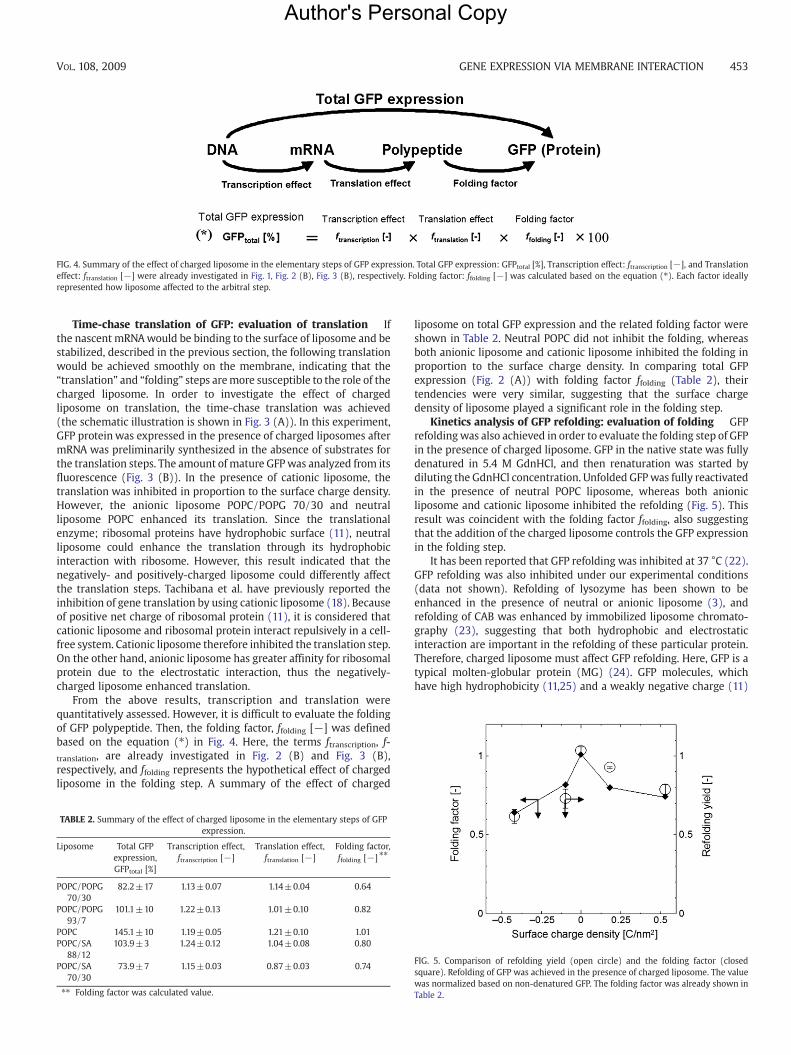
FIG. 4. Summary of the effect of charged liposome in the elementary steps of GFP expression. Total GFP expression: GFPtotal [%], Transcription effect: ftranscription [−], and Translationeffect: ftranslation [−] were already investigated in Fig. 1, Fig. 2 (B), Fig. 3 (B), respectively. Folding factor: ffolding [−] was calculated based on the equation (⁎). Each factor ideallyrepresented how liposome affected to the arbitral step.
GENE EXPRESSION VIA MEMBRANE INTERACTION 453VOL. 108, 2009
Time-chase translation of GFP: evaluation of translation Ifthe nascent mRNAwould be binding to the surface of liposome and bestabilized, described in the previous section, the following translationwould be achieved smoothly on the membrane, indicating that the“translation” and “folding” steps aremore susceptible to the role of thecharged liposome. In order to investigate the effect of chargedliposome on translation, the time-chase translation was achieved(the schematic illustration is shown in Fig. 3 (A)). In this experiment,GFP protein was expressed in the presence of charged liposomes aftermRNA was preliminarily synthesized in the absence of substrates forthe translation steps. The amount of mature GFPwas analyzed from itsfluorescence (Fig. 3 (B)). In the presence of cationic liposome, thetranslation was inhibited in proportion to the surface charge density.However, the anionic liposome POPC/POPG 70/30 and neutralliposome POPC enhanced its translation. Since the translationalenzyme; ribosomal proteins have hydrophobic surface (11), neutralliposome could enhance the translation through its hydrophobicinteraction with ribosome. However, this result indicated that thenegatively- and positively-charged liposome could differently affectthe translation steps. Tachibana et al. have previously reported theinhibition of gene translation by using cationic liposome (18). Becauseof positive net charge of ribosomal protein (11), it is considered thatcationic liposome and ribosomal protein interact repulsively in a cell-free system. Cationic liposome therefore inhibited the translation step.On the other hand, anionic liposome has greater affinity for ribosomalprotein due to the electrostatic interaction, thus the negatively-charged liposome enhanced translation.
From the above results, transcription and translation werequantitatively assessed. However, it is difficult to evaluate the foldingof GFP polypeptide. Then, the folding factor, ffolding [−] was definedbased on the equation (⁎) in Fig. 4. Here, the terms ftranscription, f-translation, are already investigated in Fig. 2 (B) and Fig. 3 (B),respectively, and ffolding represents the hypothetical effect of chargedliposome in the folding step. A summary of the effect of charged
TABLE 2. Summary of the effect of charged liposome in the elementary steps of GFPexpression.
Liposome Total GFPexpression,GFPtotal [%]
Transcription effect,ftranscription [−]
Translation effect,ftranslation [−]
Folding factor,ffolding [−] ⁎⁎
POPC/POPG70/30
82.2±17 1.13±0.07 1.14±0.04 0.64
POPC/POPG93/7
101.1±10 1.22±0.13 1.01±0.10 0.82
POPC 145.1±10 1.19±0.05 1.21±0.10 1.01POPC/SA
88/12103.9±3 1.24±0.12 1.04±0.08 0.80
POPC/SA70/30
73.9±7 1.15±0.03 0.87±0.03 0.74
⁎⁎ Folding factor was calculated value.
liposome on total GFP expression and the related folding factor wereshown in Table 2. Neutral POPC did not inhibit the folding, whereasboth anionic liposome and cationic liposome inhibited the folding inproportion to the surface charge density. In comparing total GFPexpression (Fig. 2 (A)) with folding factor ffolding (Table 2), theirtendencies were very similar, suggesting that the surface chargedensity of liposome played a significant role in the folding step.
Kinetics analysis of GFP refolding: evaluation of folding GFPrefoldingwas also achieved in order to evaluate the folding step of GFPin the presence of charged liposome. GFP in the native state was fullydenatured in 5.4 M GdnHCl, and then renaturation was started bydiluting the GdnHCl concentration. Unfolded GFPwas fully reactivatedin the presence of neutral POPC liposome, whereas both anionicliposome and cationic liposome inhibited the refolding (Fig. 5). Thisresult was coincident with the folding factor ffolding, also suggestingthat the addition of the charged liposome controls the GFP expressionin the folding step.
It has been reported that GFP refolding was inhibited at 37 °C (22).GFP refolding was also inhibited under our experimental conditions(data not shown). Refolding of lysozyme has been shown to beenhanced in the presence of neutral or anionic liposome (3), andrefolding of CAB was enhanced by immobilized liposome chromato-graphy (23), suggesting that both hydrophobic and electrostaticinteraction are important in the refolding of these particular protein.Therefore, charged liposome must affect GFP refolding. Here, GFP is atypical molten-globular protein (MG) (24). GFP molecules, whichhave high hydrophobicity (11,25) and a weakly negative charge (11)
FIG. 5. Comparison of refolding yield (open circle) and the folding factor (closedsquare). Refolding of GFP was achieved in the presence of charged liposome. The valuewas normalized based on non-denatured GFP. The folding factor was already shown inTable 2.

Author's Personal Copy
454 UMAKOSHI ET AL. J. BIOSCI. BIOENG.,
on their surface, tend to aggregate together (26). In the presence ofanionic liposome, protein-membrane distance was kept high by theelectrostatic repulsion, which induced the aggregation of MGintermediates. In contrast, MG intermediate was considered tointeract strongly with cationic liposome. Here, we previously reportedthat the interaction between positively-charged lysozyme and anionicliposome induced membrane fusion, which inhibited the refolding oflysozyme (3). By applying the membrane fusion model to GFP andcationic liposome, it was indicated that cationic liposome inhibits GFPrefolding due to membrane fusion. Although the refolding of GFP wasjust a bit of the folding step of nascent GFP polypeptide, chargedliposome exerted a drastic effect to the conformational change of GFP.It was indicating that the effect of charged liposome on GFP expressionwas significant in the folding step.
In conclusion, it was found that the total GFP expression wascontrolled by the addition of charged liposome. The analysis of theeffect of liposome on the elementary steps revealed that the reactionsin transcription, translation and folding steps could be differentlycontrolled on the surface of liposome. Transcription was enhancedwith liposome independent of the surface charge density. RNAbinding to the liposome surface was stabilized. Translation wasenhanced by anionic liposome and neutral liposome, though it wasinhibited by cationic liposome. Neutral liposome enhanced thetranslation through its hydrophobic interaction with ribosome.Ribosomal protein, which also has a positive net charge on its surface,binds to anionic liposome and translates mRNA to polypeptide on thesurface of the membrane. However cationic liposome induceselectrostatic repulsion, resulting in the inhibition of translation. GFPrefolding was not inhibited by the neutral liposome, possibly, becauseof the hydrophobic interaction to assist the GFP folding. On the otherhand, anionic liposome and cationic liposome inhibited GFP refolding,inducing aggregation of the MG intermediate and membrane fusion,respectively. We investigated that the total GFP expression wasinhibited in proportion to the surface charge density of liposome;similarly, the refolding of GFP was inhibited. Therefore, the effect ofcharged liposome on GFP expression was significant in the foldingstep. However, it should be noted that transcription was enhancedindependently of the surface charge, and translationwas enhanced byanionic liposome. Therefore it was concluded that charged liposomecontrolled the translation and folding steps of the total GFPexpression. Liposome acts as like a bioreactor, the reactions in theelementary steps such as transcription, translation and folding wereachieved on the surface of liposome. This would be one important roleof a biomembrane that controls gene expression.
ACKNOWLEDGMENTS
The fundamental concept was supported by the Research Group of“Membrane Stress Biotechnology” and “Engineering Science ofLiposome”. It was partly supported by a Grant-in-Aid for ScientificResearch (No. 15206089, 16686046, 16760635, 17656268, 19656203,19656220, and 20360350) from the Ministry of Education, Science,Sports, and Culture of Japan, a grant from the 21st Century COEprogram “Creation of Integrated EcoChemistry”, the Global COEprogram “Bio-Environmental Chemistry” of the Japan Society for thePromotion of Science (JSPS) and the JSPS-VAST Core UniversityProgram. The author also thanks the useful discussion with andcomments from Prof. T. Tsuchido, Dr. Y. Matsumura, and Dr. J.Sakamoto (Graduate School of Chemistry, Materials, and Bioengineer-ing, Kansai University, Suita, Japan). The authors are grateful to theResearch Center for Solar Energy Chemistry and the Gas hydrateAnalyzing System of Osaka University. One of the authors (H.T. Bui)
also acknowledges the financial support of JSPS and ministry ofeducation and training in Vietnam (MOET).
References
1. Tuan, L. Q., Umakoshi, H., Shimanouchi, T., and Kuboi, R.: Liposome-recruitedactivity of oxidized and fragmented superoxide dismutase, Langmuir, 24, 350–354(2008).
2. Umakoshi, H., Morimoto, K., Ohama, Y., Nagami, H., Shimanouchi, T., and Kuboi,R.: Liposome modified with Mn–porphyrin complex can simultaneously induceantioxidative enzyme-like activity of both superoxide dismutase and peroxidase,Langmuir, 24, 4451–4455 (2008).
3. Kuboi, R., Mawatari, T., and Yoshimoto, M.: Oxidative refolding of lysozymeassisted by negatively charged liposomes: Relationship with lysozyme-mediatedfusion of liposomes, J. Biosci. Bioeng., 90, 14–19 (2000).
4. Spirin, A. S., Baranov, V. I., Ryabova, L. A., Ovodov, S. Y., and Alakhov, Y. B.: Acontinuous cell-free translation system capable of producing polypeptides in highyield, Science, 242, 1162–1164 (1988).
5. Zimmer, M.: Green fluorescent protein (GFP): Applications, structure, andphotophysical behavior, Chem. Rev., 102, 759–781 (2002).
6. Oberholzer, T., Neirhaus, K. H., and Luisi, P. L.: Protein expression in liposomes,Biochem. Biophys. Res. Commun., 261, 238–241 (1999).
7. Nomura, S. M., Tsumoto, K., Hamada, T., Akiyoshi, K., Nakatani, Y., andYoshikawa, K.: Gene expression within cell-sized lipid vesicles, Chembiochem, 4,1172–1175 (2003).
8. Fischer, A., Franco, A., and Oberholzer, T.: Giant vesicles as microreactors forenzymatic mRNA synthesis, Chembiochem, 3, 409–417 (2002).
9. Ishikawa, K., Sato, K., Shima, Y., Urabe, I., and Yomo, T.: Expression of a cascadinggenetic network within liposomes, FEBS Lett., 576, 387–390 (2004).
10. Yu, W., Sato, K., Wakabayashi, M., Nakaishi, T., Ko-Mitamura, E. P., Shima, Y.,Urabe, I., and Yomo, T.: Synthesis of functional protein in liposome, J. Biosci.Bioeng., 92, 590–593 (2001).
11. Bui, H. T., Umakoshi, H., Ngo, K. X., Nishida, M., Shimanouchi, T., and Kuboi, R.:Liposome membrane itself can affect gene expression in the Escherichia coli cell-free translation system, Langmuir, 24, 10537–10542 (2008).
12. Otto, P., Kephart, D., Bitner, R., Huber, S., and Volkerding, K.: Separate isolation ofgenomic DNA and total RNA from single samples using the SV total RNA isolationsystem, Promega Notes, 69, 19 (1998).
13. Kobs, G.: Isolation of RNA from plant, yeast, and bacteria, Promega Notes, 68, 28(1998).
14. Taran, V. D., Wick, R., and Walde, P.: A 1H nuclear magnetic resonance method forinvestigating the phospholipase D-catalyzed hydrolysis of phosphatidylcholine inliposomes, Anal. Biochem., 240, 37–47 (1996).
15. Tocanne, J. F. and Teissie, J.: Ionization of phospholipids and phospholipid-supported interfacial lateral diffusion of protons in membrane model systems,Biochim. Biophys. Acta, 1031, 111–142 (1990).
16. Ptak, M., Eqret-Charlie, M., Sanson, A., and Bouloussa, O.: A NMR study of theionization of fatty acids, fatty amines and N-acylamino acids incorporated inphosphatidylcholine vesicles, Biochim. Biophys. Acta, 600, 387–397 (1980).
17. Korner, D., Benita, S., Albrecht, G., and Baszkin, A.: Surface properties of mixedphospholipid-stearylamine monolayers and their interaction with a non-ionicsurfactant (poloxamer), Colloids Surf., B Biointerfaces, 3, 101–109 (1994).
18. Tachibana, R., Harashima, H., Ishida, T., Shinohara, Y., Hino, M., Terada, H., Baba,Y., and Kiwada, H.: Effect of cationic liposomes in an in vitro transcription andtranslation system, Biol. Pharm. Bull., 25, 529–531 (2002).
19. Monnard, P. A., Oberholzer, T., and Luisi, P. L.: Entrapment of nucleic acids inliposomes, Biochim. Biophys. Acta, 1329, 39–50 (1997).
20. Janas, T., Janas, T., and Yarus, M.: Specific RNA binding to ordered phospholipidbilayers, Nucleic Acids Res., 34, 2128–2136 (2006).
21. Vlassov, A. and Yarus, M.: Interaction of RNA with phospholipid membranes, Mol.Biol., 36, 389–393 (2002).
22. Fukuda, H., Arai, M., and Kuwajima, K.: Folding of green fluorescent protein andthe Cycle3 mutant, Biochemistry, 39, 12025–12032 (2000).
23. Yoshimoto, M., Kuboi, R., Yang, Q., and Miyake, J.: Immobilized liposomechromatography for studies of protein–membrane interactions and refolding ofdenatured bovine carbonic anhydrase, J. Chromatogr., B, 712, 59–71 (1998).
24. Enoki, S., Saeki, K., Maki, K., and Kuwajima, K.: Acid denaturation and refolding ofgreen fluorescent protein, Biochemistry, 43, 14238–14248 (2004).
25. Abraham, D. J., Leo, A., and Leo, J.: Extension of the fragment method to calculateamino acid zwitterion and side chain partition coefficients, Proteins, 2, 130–152(1987).
26. Enoki, S., Maki, K., Inobe, T., Takahashi, K., Kamagata, K., Oroguchi, T., Nakatani,H., Tomoyori, K., and Kuwajima, K.: The equilibrium unfolding intermediateobserved at pH 4 and its relationship with the kinetic folding intermediates ingreen fluorescent protein, J. Mol. Biol., 361, 969–982 (2006).




















