
Zoledronic Acid Induces Autophagic Cell Death in Human Prostate Cancer Cells

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=kaup20
Download by: [ Johns Hopkins University] Date: 15 September 2015, At: 08:31
Autophagy
ISSN: 1554-8627 (Print) 1554-8635 (Online) Journal homepage: http://www.tandfonline.com/loi/kaup20
Characterization of the ATG8-conjugation systemin 2 Plasmodium species with special focus on theliver stage
Bamini Jayabalasingham, Christiane Voss, Karen Ehrenman, Julia D Romano,Maria E Smith, David A Fidock, Juergen Bosch & Isabelle Coppens
To cite this article: Bamini Jayabalasingham, Christiane Voss, Karen Ehrenman, JuliaD Romano, Maria E Smith, David A Fidock, Juergen Bosch & Isabelle Coppens (2014)Characterization of the ATG8-conjugation system in 2 Plasmodium species with special focuson the liver stage, Autophagy, 10:2, 269-284, DOI: 10.4161/auto.27166
To link to this article: http://dx.doi.org/10.4161/auto.27166
View supplementary material Published online: 12 Dec 2013.
Submit your article to this journal Article views: 261
View related articles View Crossmark data
Citing articles: 2 View citing articles

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
Basic ReseaRch PaPeR
www.landesbioscience.com autophagy 269
autophagy 10:2, 269–284; February 2014; © 2014 Landes Bioscience
Basic ReseaRch PaPeR
*Correspondence to: Isabelle Coppens; Email: [email protected]: 01/29/2013; Revised: 11/02/2013; Accepted: 11/12/2013http://dx.doi.org/10.4161/auto.27166
Characterization of the ATG8-conjugation system in 2 Plasmodium species with special focus
on the liver stagePossible linkage between the apicoplastic
and autophagic systems?Bamini Jayabalasingham,1,2,† christiane Voss,1,† Karen ehrenman,1 Julia D Romano,1 Maria e smith,1 David a Fidock,2,3
Juergen Bosch,1,4 and isabelle coppens1,*
1Department of Molecular Microbiology and immunology; Malaria Research institute; Johns hopkins University Bloomberg school of Public health; Baltimore, MD Usa; 2Department of Microbiology & immunology; Division of infectious Diseases, Medical sciences; columbia University Medical center; New York, NY Usa;
3Department of Medicine Division of infectious Diseases, Medical sciences; columbia University Medical center; New York, NY Usa; 4Department of Biochemistry and Molecular Biology; Johns hopkins University Bloomberg school of Public health; Baltimore, MD Usa
†These authors contributed equally to this work.
Keywords: malaria, Plasmodium liver forms, cellular differentiation, autophagy-related genes, ATG8, apicoplast
Abbreviations: ACP, acyl carrier protein; ATG, autophagy-related (gene); ATG, autophagy-related (protein); GABARAP, GABA(A) receptor-associated protein; GFP, green fluorescent protein; IEM, immunoelectron microscopy; IFA, immunofluorescence
assays; LC3, microtubule-associated protein 1 light chain 3; mCherry, mCherry red fluorescent protein; ORF, open reading frame; PbATG3, Plasmodium berghei ATG3; PbATG7, Plasmodium berghei ATG7; PbATG8, Plasmodium berghei ATG8; PGK,
phosphoglycerate kinase; PDM, product of the differences from the mean; PE, phosphatidylethanolamine; PfATG8, Plasmodium falciparum ATG8; p.i., post-infection; prApe1, precursor form of aminopeptidase I; PtdIns3P, phosphatidylinositol 3-phosphate;
PV, parasitophorous vacuole
Plasmodium parasites successfully colonize different habitats within mammals and mosquitoes, and adaptation to various environments is accompanied by changes in their organelle composition and size. Previously, we observed that during hepatocyte infection, Plasmodium discards organelles involved in invasion and expands those implicated in biosynthetic pathways. We hypothesized that this process is regulated by autophagy. Plasmodium spp. possess a rudimentary set of known autophagy-related proteins that includes the ortholog of yeast atg8. in this study, we analyzed the activity of the aTG8-conjugation pathway over the course of the lifecycle of Plasmodium falciparum and during the liver stage of Plasmodium berghei. We engineered a transgenic P. falciparum strain expressing mcherry-PfaTG8. These transgenic parasites expressed mcherry-PfaTG8 in human hepatocytes and erythrocytes, and in the midgut and salivary glands of Anopheles mosquitoes. in all observed stages, mcherry-PfaTG8 was localized to tubular structures. Our eM and colocalization studies done in P. berghei showed the association of PbaTG8 on the limiting membranes of the endosymbiont-derived plastid-like organelle known as the apicoplast. interestingly, during parasite replication in hepatocytes, the association of PbaTG8 with the apicoplast increases as this organelle expands in size. PbATG3, PbATG7 and PbATG8 are cotranscribed in all parasitic stages. Molecular analysis of PbaTG8 and PbaTG3 revealed a novel mechanism of interaction compared with that observed for other orthologs. This is further supported by the inability of Plasmodium aTG8 to functionally complement atg8Δ yeast or localize to autophagosomes in starved mammalian cells. altogether, these data suggests a unique role for the aTG8-conjugation system in Plasmodium parasites.
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
270 autophagy Volume 10 issue 2
Introduction
Plasmodium parasites encounter diverse conditions as they cycle between their vertebrate host and mosquito vector. Successful colonization of different habitats by these parasites is accompanied by drastic transformations in which Plasmodium change their morphology, organellar content, and metabolism. For example, in the mammalian host hepatocyte, the elongated and motile sporozoite injected by mosquitoes converts into a round trophozoite form, which prepares the parasite for the generation of thousands of infective erythrocyte-invasive forms.1 We previously observed that soon after liver cell invasion, the sporozoite undergoes a spectacular shape change accompanied by major interior remodeling, resulting in the elimination of parasite organelles specialized for motility and invasion.2,3 At the completion of metamorphosis, mature trophozoites have retained organelles involved in biosynthesis, e.g., the endoplasmic reticulum (ER), the relict plastid or apicoplast, and the mitochondrion, in addition to the nucleus.4-6 Replication of the parasite occurs via the process of schizogony, during which the parasite nucleus divides multiple times and the ER, apicoplast, and mitochondrion expand considerably in size. The end of schizogony marks a phase of biogenesis of organelles that are critical for the production of the infectious merozoites that will invade erythrocytes. To date, the cellular and molecular events involved in parasite metamorphosis, e.g., organelle elimination and expansion remain largely unexplored.
Autophagy controls the quality of the eukaryotic cytoplasm through the cell-autonomous provision of nutrients by cytosol digestion and removal of damaged organelles.7 This process involves the enclosure of a large portion of the cytoplasm within a nascent cup-shaped membrane structure termed the phagophore which is subsequently molded into a double-membrane structure called the autophagosome. The outer membrane of the autophagosome fuses with a lysosome to become an autolysosome, and the luminal cargoes of autolysosomes are then rapidly degraded. Alternatively, some autophagosomes can be directed to the plasma membrane and fuse with this membrane, thereby promoting the release of their content into the environment for exophagy.8,9 A potential role of autophagy during the differentiation of protozoan parasites is emerging.10-13 A notable example has been illustrated for Trypanosoma brucei differentiation, in which a rapid turnover of glycosomes is facilitated by selective autophagy.14,15 The autophagic machinery is also activated during the differentiation of Leishmania major and Trypanosoma cruzi into infective metacyclic forms, and during the encystation of Entamoeba histolytica and Acanthamoeba.16-20
Plasmodium parasites contain a limited number of known autophagy-related genes compared with the large repertoire of orthologs in yeast and higher eukaryotes.10-12,21-23 It remains an open question whether the malaria parasites have a primitive form of autophagy or if they contain specialized autophagy-related genes that are absent from yeast and metazoan genomes. Among Atg proteins, the genome of Plasmodium falciparum and Plasmodium berghei encodes orthologs of the 4 components of the
ATG8-conjugation pathway (ATG4, ATG3, ATG7, and ATG8). Based on bioinformatics analyses, the Plasmodium orthologs of ATG8 and ATG3 share ~68% and ~48% similarity with the yeast molecules, respectively. In contrast, the Plasmodium orthologs of ATG4 and ATG7 contain large unique regions, and their similarity to yeast orthologs is limited to the catalytic sites. In many organisms, the Atg8-ubiquitin-like system mediates membrane fusion between vesicles and the phagophore, contributing to its enlargement.24 Our studies focus on the role of this system in Plasmodium spp.
Previous works on autophagy in P. falciparum and P. berghei have focused on the apicoplast localization of the ATG8 orthologs.22,23 In this study, we demonstrated the unique molecular features of the protein and present for the first time, the spatial and temporal distribution of ATG8-containing organelles in intrahepatic parasites in both P. falciparum and P. berghei. Our findings highlight the potential of autophagy machinery as an emerging target for malaria therapy.
Results
ATG8, ATG3, and ATG7 of P. berghei have conserved and unique features
As the most widely used rodent malaria parasite, P. berghei is a valuable model organism for studying human malaria, particularly with regards to the liver stage. In order to analyze the contribution of the Atg8-conjugation system to the development of Plasmodium liver forms, we cloned ATG8, ATG3, and ATG7 from P. berghei named PbATG8, PbATG3, and PbATG7 respectively. The ORF of the 3 P. berghei ATG genes was amplified by RT-PCR from blood-stage parasite mRNA. The single exon of PbATG8 located on contig PB000658.01.0, is 375 nt long and encodes a 124 amino acid (aa) protein with a predicted mol wt of 14.7 kDa (Figs. S1 and S2). The PbATG3 transcript consists of 11 exons and the PbATG3 gene spans ~2.2 kb in the PB_RP3272 contig. The mature PbATG3 transcript is 858 nt long, and translation of the ORF predicts a 33.7 kDa protein (Fig. S3). Of note, the ORF of PbATG3 obtained by sequencing differed significantly from the ORF predicted in PlasmoDB (Gene ID PBANKA_041570). Several attempt to specifically amplify potential splice variants of the PbATG3 transcript (including the transcript predicted in PlasmoDB) were unsuccessful, suggesting that the mRNA sequence prediction presented in PlasmoDB is incorrect. The PbATG7 gene spans ~3.3 kb, which is transcribed to generate a 3,258 nt transcript after excision of a single intron of 93 bp and encodes a 1,054 aa protein. Cloned PbATG7 obtained by sequencing was identical to the ORF annotated in PlasmoDB (Gene ID PBANKA_092220; Fig. S4A and S4B)
Yeast Atg8 and its mammalian functional orthologs (e.g., MAP1LC3A/B-LC3 [microtubule-associated protein 1A/1B-light chain 3], GABARAP (GABA [A] receptor-associated protein), and GABARAPL2/GATE-16 (GABA[A] receptor-associated protein-like 2) are small proteins that consist of a ubiquitin fold at the C terminus and 2 N-terminal α-helices.25 The α-helices are tethered to the ubiquitin core by hydrogen
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com autophagy 271
bonds and salt bridges and the amino acids involved in maintenance of these structural constraints are also present in the ATG8 orthologs of P. berghei, P. yoelii, and P. falciparum.26-28 The residues that constitute 2 hydrophobic patches in GABARAPL2 and GABARAP are conserved among the Plasmodium ATG8 orthologs. In contrast, the amino acid residues present on the other face of these proteins exhibit low conservation, suggesting a reduction in evolutionary constraints. It is hypothesized that these residues define the specificity of each of the ATG8 orthologs. To this point, ATG8 in all Plasmodium species share a unique inserted loop consisting of 9 amino acids between α-helix 3 and β-sheet 3. Such an insert is also present in ATG8 from other protozoa (e.g., Trypanosoma cruzi, Toxoplasma gondii, and Entamoeba; Fig. S2). A functionally important residue conserved among all ATG8 orthologs is a C-terminal Gly. In most orthologs of ATG8, this Gly is followed by additional amino acids that are cleaved by ATG4. In this respect, the Plasmodium ATG8 sequences contain a C-terminal Gly that is not followed by residues, as also observed for Toxoplasma gondii and Caenorhabditis elegans ATG8 orthologs (Fig. S2).
PbATG3 shares 50%, 48%, and 54% similarity with the Atg3 orthologs in yeast, human and A. thaliana, respectively. A potential catalytic Cys, shared by all orthologs, has been identified at position 256 (Fig. S3). Yeast Atg3 has 2 regions of interest: the “handle region,” which mediates the binding between Atg3 and Atg8, and the “flexible region,” which is implicated in the enzyme’s conjugation activity and binding to Atg7.29 The presence of these features predicts that the malaria protein is a functional enzyme. Interestingly, PbATG3 shares greater structural similarity with the H. sapiens and A. thaliana orthologs as all 3 either have a truncated “handle region” or lack this region completely, and they share a long extension of the “flexible region.”
The PbATG7 protein shares 34% and 32% similarity with its yeast and human orthologs, respectively (Fig. S4B). Studies in Saccharomyces cerevisiae show that Atg7 binds to Atg8 via its C terminus and to Atg3 via the N terminus.30 According to Noda and colleagues, the N-terminal domain (amino acids 1 to 288) consists of 6 α-helices and 15 β-strands and has no significant similarity to other E1 enzymes. On the other hand, the C-terminal (adenylation) domain of Atg7 (amino acids 294 to 572) is conserved among E1 enzymes and consists out of 7 α-helices and 10 β-strands. The adenylation domain of ScAtg7 exhibits high similarity with the PbATG7. The catalytic cysteine, at position 507 and 533 in the C-terminal domain of ScAtg7 and human ATG7 respectively, is also found in PbATG7 at position 913 within the conserved motif DxCTVTRPGG.
PbATG8 and PbATG3 interact in yeast 2-hybrid assaysOur previous pull-down assays demonstrated that PfATG8
and PfATG3 interact with each other.31 To provide further
functional insights into the mechanism of this physical interaction, we next investigated this interaction using the yeast 2-hybrid system. The Gal4 BD-PbATG8 and Gal4 AD-PbATG3 fusion proteins were coexpressed in AH109 cells and grown on plates selective for interaction. These cells were able to grow on Leu,Trp,Ade,His-deficient media demonstrating
Figure 1. Yeast 2-hybrid assay demonstrating interaction between PbaTG pro-teins. ah109 cells coexpressing the Gal4 aD/Gal4 BD fusion proteins as indicated were streaked on -Leu,Trp and -Leu, Trp,his,ade media to assay protein-protein interaction. (+), positive control for interaction, ah109 cotransformed with plasmids pTD1-1 and pVa3-1 (for expression of the interacting proteins sV40 T-antigen and TP53/p53); (−), negative control for interaction, ah109 cotrans-formed with plasmids pLaM 5′-1 and pTD1-1 (for expression of the noninteracting proteins sV40 T-antigen and Lamin c). PbaTG31 to 260 and PbaTG31 to 249 refer to 2 truncated versions of PbaTG3 that lack the handle region and catalytic cysteine, respectively. FsPbaTG8 refers to the N-terminal frameshift mutant of PbaTG8 which does not express PbaTG8.
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
272 autophagy Volume 10 issue 2
the interaction between PbATG3 and PbATG8 (Fig. 1). We confirmed that this growth on media selective for interaction was not the result of autoactivation of the promoter (i.e., a false-positive result) by introducing a frameshift mutation at the N terminus of PbATG8 which would prevent expression of PbATG8 as part of the Gal4 BD. Expression of Gal4 BD-PbATG8 alone failed to support growth on Leu,Trp,Ade,His-deficient media (data not shown). To identify the region of PbATG3 that promotes interaction, we generated 2 truncated mutants of PbATG3: one lacking the 25 aa at the C terminus (PbATG31 to
260) which constitute the putative “handle region,”32 and another one lacking 36 aa at the C terminus (PbATG31 to 249), consisting of both the “handle region” and the catalytic Cys256. Surprisingly, both of these truncated mutants of PbATG3 were able to interact with PbATG8. This suggests that Cys256 and the “handle region” of PbATG3 are unnecessary for interaction with PbATG8. Finally, we tested the potential binding property of the “flexible region” of PbATG3 (amino acids 83 to 173; PbATG383 to 173), which shares very low conservation of residues with other Atg3 orthologs. While the “flexible region” of PbATG3 interacts with PbATG8, its removal from the full-length protein did not abrogate binding (data not shown). There was no interaction detected between the N-terminal frameshift mutant of PbATG8 and PbATG31 to 260, PbATG31 to 249, and PbATG383 to 173. These results demonstrate that, unlike the interaction between Atg3 and Atg8 in yeast, PbATG3 interaction with PbATG8 is mediated by multiple sites on the PbATG3 protein, in agreement with our previous biophysical findings for PfATG8 and PfATG3 interactions.31
PbATG3, PbATG7, and PbATG8 are constitutively transcribed throughout the parasite lifecycle
We next examined the transcriptional profiles of PbATG3, PbATG7, and PbATG8 during parasite development in the mammalian host and the mosquito vector by RT-PCR. Data show that the PbATG3, PbATG7, and PbATG8 transcripts were detected in hepatic stage parasites assayed from day 1 to day 3, (corresponding to the duration of parasite intrahepatic development), mixed blood
Figure 2. Transcriptional profiles of PbATGg8, PbATG3, and PbATG7 in P. berghei mammalian and insect stages. expression in liver stage parasites 23, 47, and 71 h p.i., mixed RBc stage parasites, midgut oocysts collected 16 d after bloodmeal, and salivary glands sporozoites was assayed by RT-PcR. To verify the absence of genomic DNa contamination, RT-PcR reactions were set up in duplicate with (+) and without (−) reverse transcriptase (RT). Blood stage and salivary gland RT-PcR show 2 independent experiments. GaPDh, α-tubulin1 (αTub1) and γ-tubulin (γTub) were used as controls.
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com autophagy 273
stage parasites, midgut oocysts, and salivary glands sporozoites (Fig. 2). We then performed quantitative real-time PCR to compare the relative transcriptional levels of PbATG3, PbATG7, and PbATG8 in sporozoites and mammalian stages (Fig. 3). Among the 3 transcripts, we found that PbATG3 was more abundant in sporozoites and liver stages day 1and 2 p.i. (Fig. 3A). In blood-stage parasites, PbATG3 and PbATG8 were equally abundant and expressed at higher levels than PbATG7. All 3 autophagy transcripts were expressed at a higher level than the housekeeping gene α-tubulin1 in liver stage parasites but at a lower level than αtubulin1 in blood forms (Fig. 3B). In sporozoites, PbATG3 and PbATG8 and not PbATG7 are expressed in higher levels than α-tubulin1.
These data suggest that the 3 components of the PbATG8-conjugation system are transcriptionally regulated throughout parasite lifecycle.
ATG8 is expressed in P. berghei and is membrane-associatedTo characterize the biochemical properties of PbATG8,
analyses were done using protein isolated from mixed blood stage parasites because the very low levels of liver stage infectivity in vitro (~1 to 3%) limits the collection of parasite material sufficient for western blots from intrahepatic forms. Three independently generated antibodies against PbATG8 revealed the presence of a unique band at the expected size of 13 kDa when lysates were separated on 4–15% acrylamide gels (Fig. 4A, a). Conjugation of Atg8 to a phospholipid, usually phosphatidylethanolamine (PE) is indicative of active autophagy in several systems.33 Soluble Atg8 and the PE-conjugated form can be separated by 6 M SDS-PAGE. In contrast to yeast ScAtg8, which can be detected in its native and lipidated forms, we observed only a single conformation of PbATG8 by 6 M urea SDS-PAGE in 3 independent experiments (Fig. 4A, b). This suggests that PbATG8 is
primarily present in either the conjugated or nonconjugated form in the parasite.
We next investigated whether PbATG8 is membrane-associated by assessing the solubilization properties of the protein. Lysates of asynchronous P. berghei blood forms were fractionated by differential centrifugation. A low-speed spin pellet containing PbATG8 was treated with either urea or Triton X-100. The solubilized and nonsolubilized proteins were then separated by a high-speed centrifugation step (Fig. 4B). PbATG8 could only be solubilized by treatment with detergent, suggesting that PbATG8 is most likely membrane-associated.
Plasmodium ATG8 does not interact with the mammalian or yeast autophagy machineries
To assess the functional competence of the Plasmodium ATG8 ortholog, we investigated the ability of PfATG8 to complement autophagy in mammalian and yeast systems. COS cells were cotransfected with expression plasmids for mCherry-PfATG8 and GFP-LC3 to determine whether PfATG8 could participate in autophagy in mammalian cells. The cotransfected cells were incubated under normal growth conditions, or conditions that stimulate autophagy, e.g., chloroquine treatment or amino acid deprivation. The expression of the 2 fusion proteins was verified
Figure 3. Quantification of the PbATG3, PbATG7 and PbATG8 transcripts in P. ber-ghei sporozoites and mammalian stage parasites. Quantitative PcR done using with 100 ng cDNa from various stages of the lifecycle as indicated. (A) Transcript abundance (cT values) of PbATG3, PbATG7, PbATG8, and α-tubulin1 (αTub1). (B) expression levels (ΔcT values) of PbATG3, PbATG7, and PbATG8 normal-ized to P. berghei α-tubulin1 (αTub1) for the indicated stages. Data are means ± sD of 1 representative experiment done with 3 independent samples.
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
274 autophagy Volume 10 issue 2
by immunoblotting (Fig. S5A). Fluorescence microscopic observations reveal that under normal growth conditions, GFP-LC3 was mainly cytosolic like mCherry-PfATG8 (Fig. S5B). Upon chloroquine treatment or starvation, GFP-LC3 was associated with large autophagosomal structures while mCherry-PfATG8 remained cytoplasmic.
Atg8 orthologs from other protozoan organisms including Leishmania major (ATG8A, ATG8B and ATG8C) or Trypansosma cruzi (TcATG8.1) are able to complement an atg8Δ strain of S. cerevisiae, indicating functional conservation of these Atg8 orthologs.17,34 We examined whether PbATG8 could likewise functionally complement an atg8Δ yeast strain. To assay complementation of autophagy, PbATG8 was expressed under the control of the PGK (phosphoglycerate kinase) promoter and processing of the precursor form of aminopeptidase I (prApe1) was monitored by western blotting.35,36 As observed for the mammalian system, complementation was not observed upon induction of starvation (Fig. S6A). This inability was not due to a lack of protein expression as PbATG8 expression in yeast was confirmed. PbATG8 differs from its yeast ortholog at several amino acid residues that are relevant for Atg8 conjugation to PE. We tested if gain-of-function could be induced by introducing specific mutations in PbATG8. Three mutations were introduced in the PbATG8 sequence: Phe48 (a residue involved in Atg8 lipidation in yeast) was mutated to tyrosine; Tyr85 (a residue implicated in binding Atg4 in yeast) was mutated to phenylalanine; and a C-terminal arginine was added at the protein (which is necessary for Atg4 recognition
in yeast). These PbATG8 mutants were tested for their ability to rescue the autophagy defect in atg8-deleted yeast. However, like wild-type PbATG8, all the mutants failed to complement the atg8Δ strain (Fig. S6A). Even the combination of the 3 mutations in PbATG8 failed to rescue the defect in yeast. The possibility of indirect effects of the promoter was also eliminated as yeast could express PbATG8 under the PGK1 and the ATG8 promoter at the same level (data not shown). Together, these data indicate that structural differences in the P. berghei protein prevent it from interacting with the autophagy machinery of S. cerevisiae. We investigated whether this lack of complementation was due to an inability of PbATG8 to directly interact with yeast Atg3 as would be required for functional autophagy to occur. Indeed, yeast-2 hybrid assays demonstrated that yeast Atg3 did not interact with PbAtg8 (assays done with Gal4 BD-PbATG8 and Gal4 AD-ScAtg3 fusion proteins coexpressed in yeast strain AH109 and grown on media selective for interaction; Fig. S6B). This suggests that ATG8 from either P. falciparum or P. berghei cannot functionally substitute for LC3 or ScAtg8 due to incompatibilities between the parasite and mammalian or yeast autophagic machineries.
At the onset of Plasmodium development in liver cells, PbATG8 localizes to punctate structures that align along the apicoplast
The temporal and spatial regulation of PbATG8 expression in P. berghei-infected hepatic cells was analyzed by immunofluoresence assays (IFA) using anti-PbATG8 antibodies at various times post-infection (p.i.). On different z-sections
Figure 4. expression of PbaTG8 and association with membranes. (A, a) immunoblots of parasite lysates prepared from GFP-expressing P. berghei blood forms, separated by sDs-PaGe and probed with antibodies against a conserved peptide of Lc3 (1), and rat (2) or mouse (3) antibodies against full-length PbaTG8 reveal the presence of a major band at 15 kDa. (A, b) immunoblots from lysates of yeast (left) and mixed Plasmodium blood forms (right) separated on a 13% acrylamide gel containing 6 M urea. Wild-type (WT) yeast and mutant yeast strains deleted of atg8 (Δ8) or atg4 (Δ4) were grown in normal growth media (Normal), then in medium lacking amino acids for 1 h to induce mild starvation (starvation). No atg8 is visible in Δ8 and scatg8 fails to be conjugated to Pe in the absence of atg4. (B) immunoblots of parasite lysates prepared from asynchronous P. berghei blood forms. a low speed pellet of parasites was treated with urea or Triton-X 100 and separated into 100,000 g pellet (P) and supernatant (s) before immunoblotting using anti-PbaTG8 antibody.
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015
©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com autophagy 275
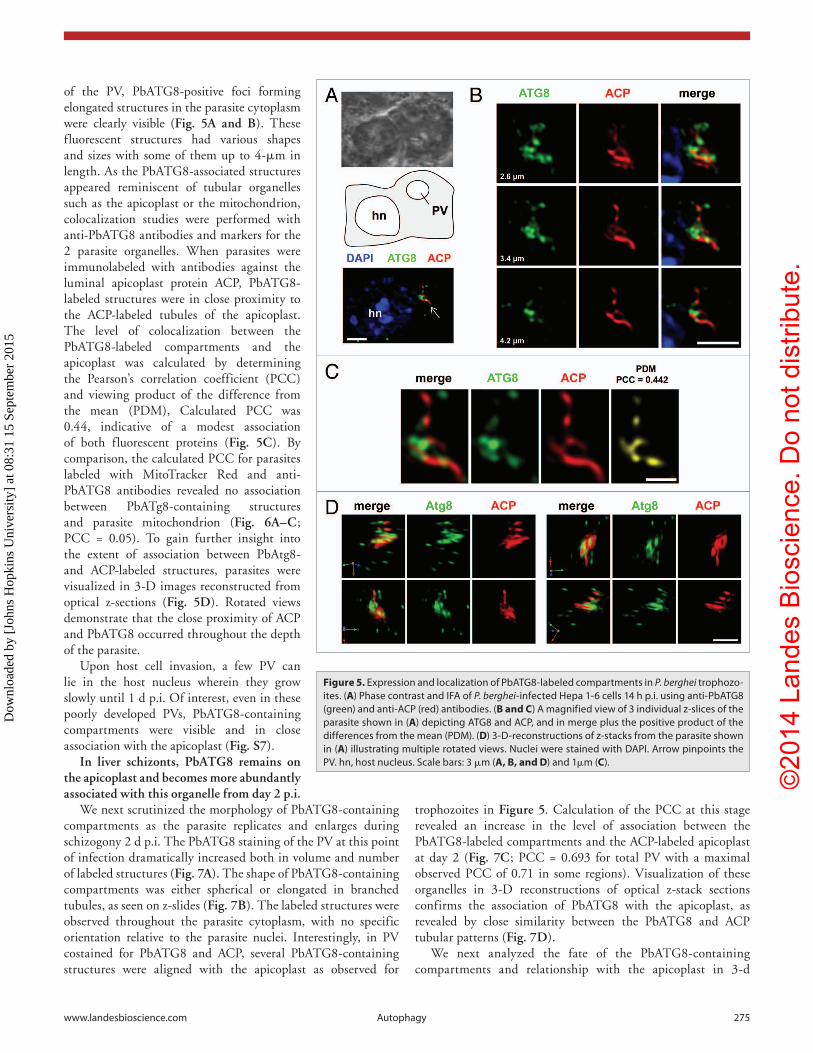
of the PV, PbATG8-positive foci forming elongated structures in the parasite cytoplasm were clearly visible (Fig. 5A and B). These fluorescent structures had various shapes and sizes with some of them up to 4-μm in length. As the PbATG8-associated structures appeared reminiscent of tubular organelles such as the apicoplast or the mitochondrion, colocalization studies were performed with anti-PbATG8 antibodies and markers for the 2 parasite organelles. When parasites were immunolabeled with antibodies against the luminal apicoplast protein ACP, PbATG8-labeled structures were in close proximity to the ACP-labeled tubules of the apicoplast. The level of colocalization between the PbATG8-labeled compartments and the apicoplast was calculated by determining the Pearson’s correlation coefficient (PCC) and viewing product of the difference from the mean (PDM), Calculated PCC was 0.44, indicative of a modest association of both fluorescent proteins (Fig. 5C). By comparison, the calculated PCC for parasites labeled with MitoTracker Red and anti-PbATG8 antibodies revealed no association between PbATg8-containing structures and parasite mitochondrion (Fig. 6A–C; PCC = 0.05). To gain further insight into the extent of association between PbAtg8- and ACP-labeled structures, parasites were visualized in 3-D images reconstructed from optical z-sections (Fig. 5D). Rotated views demonstrate that the close proximity of ACP and PbATG8 occurred throughout the depth of the parasite.
Upon host cell invasion, a few PV can lie in the host nucleus wherein they grow slowly until 1 d p.i. Of interest, even in these poorly developed PVs, PbATG8-containing compartments were visible and in close association with the apicoplast (Fig. S7).
In liver schizonts, PbATG8 remains on the apicoplast and becomes more abundantly associated with this organelle from day 2 p.i.
We next scrutinized the morphology of PbATG8-containing compartments as the parasite replicates and enlarges during schizogony 2 d p.i. The PbATG8 staining of the PV at this point of infection dramatically increased both in volume and number of labeled structures (Fig. 7A). The shape of PbATG8-containing compartments was either spherical or elongated in branched tubules, as seen on z-slides (Fig. 7B). The labeled structures were observed throughout the parasite cytoplasm, with no specific orientation relative to the parasite nuclei. Interestingly, in PV costained for PbATG8 and ACP, several PbATG8-containing structures were aligned with the apicoplast as observed for
trophozoites in Figure 5. Calculation of the PCC at this stage revealed an increase in the level of association between the PbATG8-labeled compartments and the ACP-labeled apicoplast at day 2 (Fig. 7C; PCC = 0.693 for total PV with a maximal observed PCC of 0.71 in some regions). Visualization of these organelles in 3-D reconstructions of optical z-stack sections confirms the association of PbATG8 with the apicoplast, as revealed by close similarity between the PbATG8 and ACP tubular patterns (Fig. 7D).
We next analyzed the fate of the PbATG8-containing compartments and relationship with the apicoplast in 3-d
Figure 5. expression and localization of PbaTG8-labeled compartments in P. berghei trophozo-ites. (A) Phase contrast and iFa of P. berghei-infected hepa 1-6 cells 14 h p.i. using anti-PbaTG8 (green) and anti-acP (red) antibodies. (B and C) a magnified view of 3 individual z-slices of the parasite shown in (A) depicting aTG8 and acP, and in merge plus the positive product of the differences from the mean (PDM). (D) 3-D-reconstructions of z-stacks from the parasite shown in (A) illustrating multiple rotated views. Nuclei were stained with DaPi. arrow pinpoints the PV. hn, host nucleus. scale bars: 3 μm (A, B, and D) and 1μm (C).
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
276 autophagy Volume 10 issue 2
old schizonts prior to the formation of hepatic merozoites. By that time, the apicoplast has formed a large and convoluted tubular network. PbATG8-containing compartments were also enlarged, forming a thicker tubular arrangement (Fig. 8A and B). A similar degree of superposition of the PbATG8-labeled tubules and ACP was observed at day 3 as on day 2 p.i., as determined by calculating PCC values (Fig. 8B; PCC = 0.655). In addition, 3-D views of schizonts at day 3 illustrate that the profiles of PbATG8-labeled compartments and the ACP-labeled region of the apicoplast were quite similar on each rotated view of the PV (Fig. 8C). In these images, the green fluorescent staining for PbATG8 seems to intersperse with the red fluorescent signal for ACP, in one continuous structure, suggesting that these proteins occupy different structures
regions of the same organelle (Fig. 7D and E).
PbATG8 is preferentially distributed on the outermost membranes of the apicoplast and neighboring tubules and vesicles of this organelle
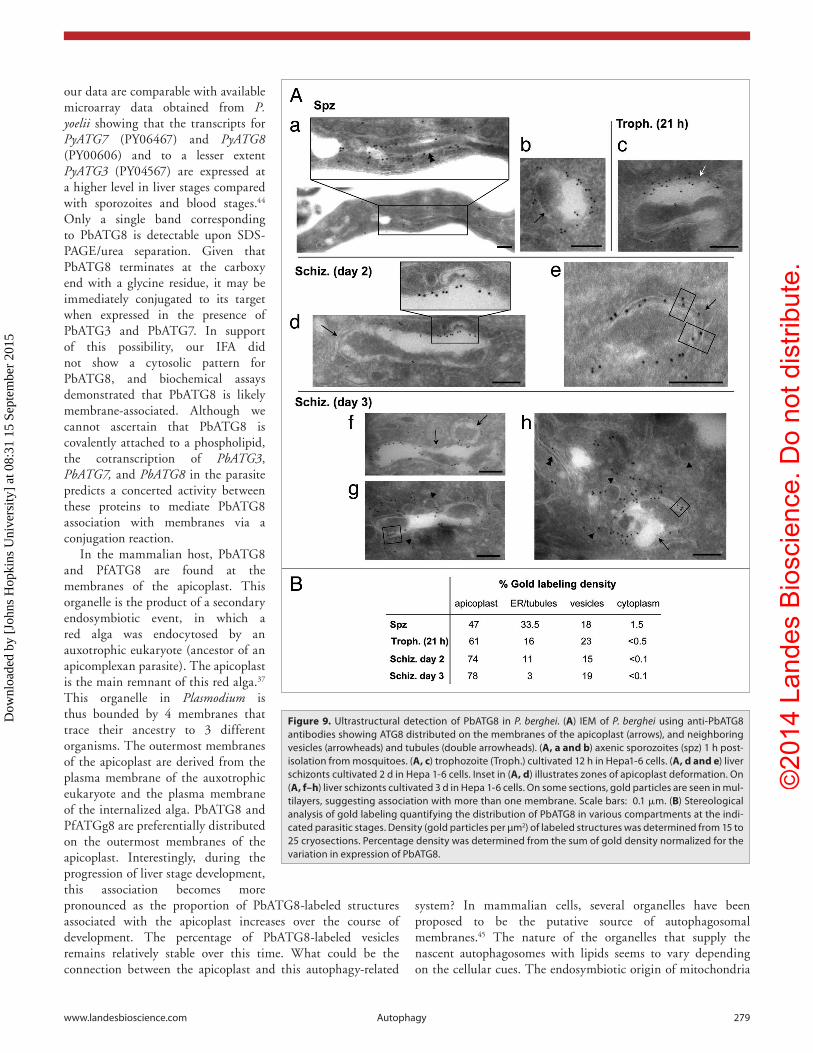
The apicoplast is a 4-membrane bounded plastid derived from an ancestral secondary endosymbiotic event.37 To provide more details about the fine localization of PbATG8 in P. berghei, we undertook immunoelectron microscopy (IEM) analyses using anti-PbATG8 antibodies at different parasitic stages. At the ultrastructural level, the apicoplast is easily identifiable due to its 4 membranes, electron dense genome, and bowtie shape in transverse sections. Sporozoites cultivated in axenic conditions to induce differentiation into trophozoites were analyzed by IEM. Immunogold staining in these trophozoites was associated with long tubules in the parasite (Fig. 9A, a) and on the contour of the apicoplast (Fig. 9A, b). Interestingly, we found that gold particles were more confined to one side of the apicoplast in many of the sections viewed. The apicoplastic network within intracellular trophozoites 21 h p.i. (Fig. 9A, c) and schizonts 2 d p.i. (Fig. 9A, d) was also labeled with anti-PbATG8 antibodies at the periphery. This apicoplastic labeling appeared to be constrained to specific regions of the membranes. These IEM observations may explain our IFA observations of a discontinuous pattern for PbATG8 on the apicoplast and its partial shift relative to the luminal ACP.
High magnification of day 2 schizonts revealed the association of PbATG8 on at least 2 of the 4 membranes of the apicoplast, identifiable as the outermost
membranes of this organelle (Fig. 9A, e). As the apicoplastic network expands, more gold particles were observed in different regions of the apicoplast as seen on schizonts at day 3 (Fig. 9A, f). Few tubules and several vesicles in close vicinity of the apicoplast were also peripherally labeled (Fig. 9A, g and h). IEM data were quantified by stereological analysis measuring the percentage of gold particle density within sporozoites, trophozoites, and schizonts (Fig. 9B), revealing a progressive increase of immunogold staining on the apicoplast as the parasite develops in liver cells.
Overall, our IEM studies corroborate the association of PbATG8 with membranes of the apicoplast and confirm our biochemical data suggesting that PbATG8 is a membrane-bound protein (Fig. 4B). Our IEM analysis also reveals the presence of a
Figure 6. Localization PbaTG8-labeled structures relative to the mitochondrion in P. berghei in liver schizonts. (A) Phase contrast and fluorescence staining of P. berghei-infected hepa 1-6 cells 51 h p.i. using anti-PbaTG8 antibodies (green) and mito-Tracker (mitoT in red). (B and C) a magnified view of 3 individual z-slices of the parasite shown in (A) and inset images showing the aTG8 and mitoT staining. calculated PDM value reveals no association between PbaTG8-labeled structures and the mitochondrion. Nuclei were stained with DaPi. arrow pinpoints the PV. hn, host nucleus. scale bars: 7 μm (A), 6 µm (B and C), and 2 µm (C).D
ownl
oade
d by
[Jo
hns
Hop
kins
Uni
vers
ity]
at 0
8:31
15
Sept
embe
r 20
15

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com autophagy 277
persistent population of PbATG8-labeled vesicles in the parasite. Although EM data cannot inform the nature of the interaction between the peri-apicoplastic vesicles and the apicoplast (i.e., docking, fusion, and/or budding), we hypothesize a dynamic interface between the apicoplast and PbATG8-labeled vesicles.
PfATG8 localizes to tubular structures throughout the lifecycle of P. falciparum
We next wanted to inspect the expression and localization of ATG8 throughout the parasite life cycle. A previous study reported the distribution of PfATG8 on the apicoplast of Plasmodium bloodforms.22 We engineered a parasite strain of P. falciparum expressing mCherry-PfATG8 to assess the morphology of the PfATG8-containing compartments in mosquito and mammalian stages. P. falciparum oocysts identified in the midgut by anti-capsule antibody were strongly positive for mCherry, indicative of PfATG8 expression (Fig. S8A). As seen in z-sections (Fig. S8A, a) and rotated views (Fig. S8A, b) of the oocysts, the pattern of the mCherry-PfATG8 was extended and tubular in shape, consistent with the morphology of the apicoplast. Sporozoites within salivary glands were also fluorescently labeled for mCherry-PfATG8, and the signal was largely punctate (Fig. S8B). Human hepatocytes were then infected with mCherryPfATG8-expressing sporozoites for 2 d (Fig. S9A). The parasite that was identified by HSP70 staining expressed mCherry-PfATG8 on tubular structures, with a similar pattern as that observed in P. berghei-infected hepatocytes using anti-PbATG8 antibodies (Fig. S9A). Finally, infected red blood cells were inspected and fluorescent staining was observed in punctate and tubular structures in the parasite cytoplasm (Fig. S9B, a). Our IEM analyses using anti-mCherry antibodies on merozoites, and early and late schizonts reveal the association of PfATG8 with the membranes of the apicoplast (Fig. 9B, b).
Altogether, these data indicate that ATG8 localized to tubular structures in all stages of P. falciparum, and definitively to the apicoplast membranes in blood forms.
Discussion
Besides its role as an emergency mechanism during periods of nutrient deficiency, autophagy is also employed during a variety of cellular processes, including the differentiation of mammalian cells.38 Recently, the contribution of autophagy to the differentiation of protozoan parasites that cycle between an insect and a vertebrate has been documented.17,20 During the malaria life cycle, Plasmodium parasites differentiate rapidly within new host environments as they initiate the process of replication.2,39-41 We previously showed that in hepatocytes, P. berghei and Plasmodium yoelii sporozoites undergo significant
Figure 7. expression and localization of PbaTG8-labeled compartments in P. berghei liver schizonts at day 2 p.i. (A) Phase contrast and iFa of P. berghei-infected hepa 1-6 cells 49 h p.i. using anti-PbaTG8 (green) and anti-acP (red) antibodies. (B and C) a magnified view of 3 individual z-slices of the parasite shown in (A), depicting aTG8 and acP, and in merge plus the positive PDM. (D) 3-D-reconstructions of z-stacks from the parasite shown in (A) illustrating multiple rotated views. Nuclei were stained with DaPi. arrow pinpoints the PV. hn, host nucleus. scale bars: 7.8 µm (A), 5 μm (B and C), and 4.5 µm (D).
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
278 autophagy Volume 10 issue 2
phenotypic changes associated with the clustering and clearance of organelles that are obsolete for replication of the parasites in the liver.2 In this paper, as a first step to explore the potential contribution of the parasite autophagy machinery to Plasmodium development, we have analyzed the expression profiles and characteristics of the components of the ATG8-conjugation system in Plasmodium during its development in the mammalian host and the Anopheles vector.
We previously showed the relatively close structural similarity of PfATG8 with mammalian LC3.31 While the surface-exposed residues located on one side of all Atg8 functional orthologs
are highly conserved, many of the surface-exposed residues located on the opposite side of the protein exhibit low conservation, even among orthologs expressed in humans, which suggests that this side mediates interactions that are unique to different Atg8 orthologs. Plasmodium ATG8 orthologs share a unique inserted loop consisting of 9 amino acids between α-helix 3 and β-sheet 3, suggesting that such binding partners exist. The N-terminal glycine of Atg8 forms a thioester bond with the catalytic cysteine of Atg3 in order to be conjugated to PE.42 The C-terminal “handle region” of Atg3 has been shown to mediate the interaction between Atg3 and Atg8.43 Interestingly, the interaction between PbATG3 and PbATG8 differs somewhat from that in yeast in that it does not seem to be solely dependent on any single region of Atg3. Indeed, the “handle region” and the catalytic cysteine of PbATG3 are not essential for its interaction with PbATG8. Additionally, the “flexible region” of PbATG3 can interact with PbATG8 but is not essential for this interaction. Compared with yeast Atg3, in which the C terminus forms a long α-helix, the C terminus of PbATG3 is shorter and is not predicted to form such secondary structure, similar to mammalian and A. thaliana ATG3.43 Thus, PbATG3 interacts with PbATG8 via numerous interfaces and this interaction may more closely mimic that of the mammalian and plant homologs. The inability of
PbATG8 to complement atg8Δ yeast or interact with yeast Atg3 is consistent with this observation. The lack of recognition of PfATG8 by the mammalian cell autophagy machinery suggests that the interaction between Atg8 and Atg3 has evolved in an organism-specific manner.
PbATG3, PbATG7, and PbATG8 are cotranscribed in all Plasmodium stages, reflecting the importance of expression of these components of the Atg8-conjugation pathway in mammals and insects. Our data showed that PbATG3, PbATG7, and PbATG8 are upregulated in parasites upon liver infection (day 1), then downregulated in blood forms parasites. Interestingly,
Figure 8. expression and localization of Pbatg8-associated compartments in P. berghei late liver schizonts at day 3 p.i. (A) Phase contrast and iFa of P. berghei-infected hepa 1-6 cells 72 h p.i. using anti-PbaTG8 (green) and anti-acP (red) antibodies. (B) a magnified view of 3 individual z-slices of the parasite shown in (A), depicting aTG8 and acP, and in merge plus the positive PDM. (C) 3-D-reconstructions of z-stacks from the parasite shown in (A) illustrating multiple rotated views. (D) inset images from the PV shown in (B and E) its rotated views. Nuclei were stained with DaPi. arrow pinpoints the PV. hn, host nucleus. scale bars: 8 μm (A), 4 μm (B and C), and 1 μm (D and E).
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015
©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com autophagy 279
our data are comparable with available microarray data obtained from P. yoelii showing that the transcripts for PyATG7 (PY06467) and PyATG8 (PY00606) and to a lesser extent PyATG3 (PY04567) are expressed at a higher level in liver stages compared with sporozoites and blood stages.44 Only a single band corresponding to PbATG8 is detectable upon SDS-PAGE/urea separation. Given that PbATG8 terminates at the carboxy end with a glycine residue, it may be immediately conjugated to its target when expressed in the presence of PbATG3 and PbATG7. In support of this possibility, our IFA did not show a cytosolic pattern for PbATG8, and biochemical assays demonstrated that PbATG8 is likely membrane-associated. Although we cannot ascertain that PbATG8 is covalently attached to a phospholipid, the cotranscription of PbATG3, PbATG7, and PbATG8 in the parasite predicts a concerted activity between these proteins to mediate PbATG8 association with membranes via a conjugation reaction.
In the mammalian host, PbATG8 and PfATG8 are found at the membranes of the apicoplast. This organelle is the product of a secondary endosymbiotic event, in which a red alga was endocytosed by an auxotrophic eukaryote (ancestor of an apicomplexan parasite). The apicoplast is the main remnant of this red alga.37 This organelle in Plasmodium is thus bounded by 4 membranes that trace their ancestry to 3 different organisms. The outermost membranes of the apicoplast are derived from the plasma membrane of the auxotrophic eukaryote and the plasma membrane of the internalized alga. PbATG8 and PfATGg8 are preferentially distributed on the outermost membranes of the apicoplast. Interestingly, during the progression of liver stage development, this association becomes more pronounced as the proportion of PbATG8-labeled structures associated with the apicoplast increases over the course of development. The percentage of PbATG8-labeled vesicles remains relatively stable over this time. What could be the connection between the apicoplast and this autophagy-related
Figure 9. Ultrastructural detection of PbaTG8 in P. berghei. (A) ieM of P. berghei using anti-PbaTG8 antibodies showing aTG8 distributed on the membranes of the apicoplast (arrows), and neighboring vesicles (arrowheads) and tubules (double arrowheads). (A, a and b) axenic sporozoites (spz) 1 h post-isolation from mosquitoes. (A, c) trophozoite (Troph.) cultivated 12 h in hepa1-6 cells. (A, d and e) liver schizonts cultivated 2 d in hepa 1-6 cells. inset in (A, d) illustrates zones of apicoplast deformation. On (A, f–h) liver schizonts cultivated 3 d in hepa 1-6 cells. On some sections, gold particles are seen in mul-tilayers, suggesting association with more than one membrane. scale bars: 0.1 μm. (B) stereological analysis of gold labeling quantifying the distribution of PbaTG8 in various compartments at the indi-cated parasitic stages. Density (gold particles per µm2) of labeled structures was determined from 15 to 25 cryosections. Percentage density was determined from the sum of gold density normalized for the variation in expression of PbaTG8.
system? In mammalian cells, several organelles have been proposed to be the putative source of autophagosomal membranes.45 The nature of the organelles that supply the nascent autophagosomes with lipids seems to vary depending on the cellular cues. The endosymbiotic origin of mitochondria
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
280 autophagy Volume 10 issue 2
and the apicoplast contributes to the unique lipid composition of these organelles. Mitochondria have been proposed to be one of the lipid sources for autophagosome formation as this organelle contains PE that can be covalently linked to the members of the LC3 protein family.46 In mammalian cells, changes in mitochondrial activity (e.g., dysfunctional mitochondria characterized by loss of ATP production) can trigger a process of autophagy to preserve minimal cell functions.47 Mitochondria react to the depletion of nutrients by actively influencing their own fate, either promoting their extensive tubulation and increase in size or activating mitophagy. Like the mitochondrion, the apicoplast is an important cellular site for the synthesis of a plethora of essential metabolites and factors, including lipids, isoprenoids, heme, and amino acids.48 A reduction of metabolites produced by the apicoplast may also be strongly perceived as a stress signal to activate an autophagic process in Plasmodium, or the apicoplast may be itself an autophagy substrate to ensure quality control of its products.
Interestingly, the outermost membranes of the apicoplast are also enriched in phosphatidylinositol 3-phosphate (PtdIns3P),49 a phospholipid that marks autophagic structures in mammalian cells. In mammalian cells, PtdIns3P acts as a membrane-bound localized signal controlling membrane dynamics by specifically recruiting cytosolic protein effectors that mediate membrane deformation, expansion, and vesicle transport.50 The molecular link between PtdIns3P effectors and the Atg8-conjugation system is still unclear. However, it is known that local production of PtdIns3P on the phagophore membrane promotes the negative curvature of this membrane and controls autophagosome size.51 PtdIns3P is the product of the class III PtdIns 3-kinase, PIK3C3/VPS34, and this enzyme localizes to the phagophore.52 Ablation of Vps34 prevents the formation of autophagosomes.53 The Plasmodium genome encoded one predicted Vps34-type enzyme containing the characteristic domains of class III PtdIns 3-kinases (e.g., a calcium/lipid-binding C2 domain, a PtdIns 3-kinase family accessory domain and a C-terminal PtdIns 3- and PtdIns 4-kinase catalytic domain). Surprisingly, PtdIns3K (the P. falciparum PIK3C3 ortholog) has been shown to localize at the food vacuole and host plasma membrane during the blood stage.53 It will be interesting to examine whether Plasmodium PtdIns3K participates in the formation of autophagosomes budding off from the membrane of the apicoplast as described for mitochondria.
Alternatively, the apicoplast may not be the organelle provider of lipids to the autophagosomes but rather it may receive lipids or membranes via the Atg8-labeled vesicles that distribute around the apicoplast. Like the mitochondrion, the apicoplast is a highly dynamic organelle, with morphologies ranging from small roundish elements (in merozoites) to larger interconnected networks (in schizonts) for which large amounts of lipids are needed. The apicoplast may expand in size through a lipid transfer mechanism from an adjacent organelle, perhaps facilitated by ATG8. In support to this hypothesis, it is known that 2 mammalian Atg8 functional orthologs, GABARAP and GABARAPL2 participate in vesicular transport from the ER to the Golgi, and within the Golgi apparatus.54 Even the function
of LC3 does not seem to be limited to autophagy as the protein is able to bind fibronectin RNA, enhancing its translation.55,56 In C. elegans, the function of the Atg8 ortholog, LGG-1, is associated with autophagy while the role of LGG-2 remains unknown.57 Given the identification of only one Atg8 ortholog in Plasmodium species, it is possible that this single protein performs multiple functions within the parasite. It remains to be determined whether the parasite exploits the ATG8-conjugation pathway to mediate the elimination of unwanted organelles via autophagosomes derived from the membranes of the apicoplast and/or to expand the size of existing organelles, like the apicoplast.
Materials and Methods
Reagents and antibodiesAntibodies used for immunolocalization included: rat or
rabbit anti-PbACP (diluted at 1:1000 for IFA) generously provided by Dr S Prigge (Johns Hopkins University), mouse anti-PfCp380 (diluted at 1:1000 for IFA) as a generous gift from Dr A Ghosh (Johns Hopkins University), anti-mCherry (diluted at 1:10 for IEM; Clontech dsred polyclonal antibodies; 632496) and rat anti-PbATG8 (diluted at 1:200 and 1:20 for IFA and IEM respectively; this study). Anti-IgG antibodies conjugated to Alexa Fluor 350, 488 or 594 obtained from Invitrogen were used at a dilution of 1:1000–1:3000. For western blotting, rabbit anti-LC3 peptide antibodies from Dr LD Sibley (Washington University), mouse anti-GFP (clontech anti-GFP monoclonal antibody; 623280) and rabbit anti-mCherry were used at a dilution of 1:1000 and the anti-Ape1 antibody 1532 from Dr DJ Klionsky (University of Michigan) at a dilution of 1:5000. The MitoTracker Red CMXRos used at 1 μM was from Invitrogen (M-7512). Primers were synthesized by Integrated DNA Technologies and their sequences are provided in Table S1.
Mammalian cell linesMouse Hepa 1-6 cells (ATCC CRL-1830) used to host
P. berghei parasites and COS-7 cells (ATCC CCL-70) for transfection studies were obtained from ATCC. Cells were grown as monolayers at 37 °C in an atmosphere of 5% CO
2
in α-MEM (Gibco, 32561037), supplemented with 10% FBS, 2 mM L-glutamine and penicillin/streptomycin (100 U/ml per 100 µg/ml).
MiceFive- to 8- week old female Swiss-Webster were purchased
from Taconic. All animal procedures were approved by the Institutional Animal Care and Use Committee of the Johns Hopkins University following the National Institutes of Health guidelines for animal housing and care.
Parasite strainsThe P. berghei ANKA wild-type line was passaged in
Anopheles stephensi mosquitoes blood-fed on infected Swiss CD-1/ICR mice as described.3 To generate a P. falciparum line expressing mCherry-ATG8, the mCherry gene was fused to PfATG8 by fusion PCR using the primer pairs 139 and 140 for mCherry and 141 and 142 for PfATG8. The full construct, generated using primers 139 and 142, was then cloned into the
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com autophagy 281
AvrII/AflII sites of the vector pLN-ENR-GFP, obtained from the MR4 (Manassas, VA, plasmid number MRA-846, deposited by Dr DA Fidock from Columbia University), to replace ENR-GFP. This plasmid, designated pLN-mCherry-PfATG8 was transfected into Dd2 and NF54 parasites harboring the attB recombinant locus along with the plasmid pInt (Manassas; catalog numbers MRA-847 and MRA-843, deposited by Dr DA Fidock from Columbia University). Transfectants were selected with G418 (125 μg/ml) and blasticidin (2 μg/ml) for 6 d and then maintained continuously under blasticidin pressure. Fluorescent parasites were visible 30 d post-transfection.
Plasmodium cultivation and infectionSporozoites were isolated from disrupted salivary glands
of infected A. stephensi mosquitoes and further purified as described6 before incubation in the presence of Hepa 1-6 for various times. Blood stage parasites were obtained from anesthetized infected mice with a parasitemia above 3% by cardiac puncture. Blood was collected into syringes coated with a stock solution of 25,000 U/ml of heparin to prevent clotting and washed with PBS prior to saponin lysis. P. falciparum were maintained in blood cultures as described.58 Mosquito and liver stage infections with P. falciparum expressing mCherry-PfATG8 were performed by Sanaria.
Cloning of autophagy-related genes in P. bergheiPbATG8 was cloned using primers 89 and 90, and PbATG3
using primers 40 and 41. PbATG8 and PbATG3 genes were cloned from cDNA generated from 1μg of blood stage parasite RNA (purified as described below) using the ABI high-capacity cDNA Reverse Transcription Kit and random hexamers. PCR was done using 1 μl cDNA and recombinant Taq DNA polymerase (Invitrogen, 10342-053) in 1× PCR buffer containing 20 mM TRIS-HCl (pH 8.4), 50 mM KCl, 1.5 mM MgCl
2 and the following conditions: 94 °C for 2 min
followed by 35 cycles of melting at 94 °C for 30 s, annealing for 30 s at the temperature described below for each primer set, elongation at 72 °C for 1 min, and a final elongation at 72 °C for 7 min following the completion of the 35 cycles. Primers 89 and 90, annealed at 50 °C were used to detect PbATG8 ; primers 40 and 41, annealed at 45 °C were used to detect PbATG3. For amplification of PbATG3, DMSO was added to the PCR reaction at a final concentration of 5%. PbATG7 was cloned in 2 steps. The primer sets PR12/PR23 and PR20/PR14 were used to amplify 2 halves of the gene independently from cDNA generated form blood stage parasite RNA. Both RT PCR products were used in equal amounts as template for a standard fusion PCR reaction (using Phusion High Fidelity from New England Biolabs, NEB, M0530L) to generate PbATG7 using the primer set PR12/PR14.
Transcriptional analysisRNA was purified from midgut oocysts, sporozoites,
Plasmodium infected-hepatocytes and blood stage parasites using the RNeasy RNA purification kit (Qiagen, 74104). One microgram of total RNA was used to generate cDNA as a template for subsequent RT-PCR (Invitrogen, 10928-034). Two sets of RT-PCR were performed for each gene. As a negative control, a second sample of −RT cDNA was also generated by
omission of the reverse transcriptase and used for PCR. Primers PR45 and PR46 were used to detect PbATG3 (transcript expected size: 512 bp). Primers PR9 and PR15, and PR43 and PR44 were used to detect PbATG7 (transcript expected sizes: 440 bp and 504 bp). For PbATG8 detection, primers PR47 and PR48 were used (transcript expected size: 376 bp). As controls, GADPH (PR51 and PR52; transcript expected size: 509 bp), γ-tubulin (PR38 and PR39; transcript size: 501 bp), or α-tubulin1 (PR32 and PR33; transcript expected size: 511 bp) were selected. For quantitative real-time PCR, total RNA from sporozoites, liver, and blood forms was isolated using the RNeasy Mini Kit (Qiagen, 74104) prior to reverse transcription of 1 μg of RNA each using with the SuperScript III First-Strand Synthesis System (Invitrogen, 18080-051) and oligo-dt primers. One hundred ng of each cDNA were amplified with the primer pairs qRT_3_f and qRT_3_r for PbATG3, qRT_7_f and qRT_7_r for PbATG7, qRT_8_f and qRT_8_r for PbATG8, and qRT_atub1_f and qRT_atub1_r for α-tubulin1 listed in Table S1. qPCR was run in the Applied Biosystems StepOnePlus Real-Time PCR apparatus. Transcript abundance was examined by calculating the C
T values
of PbATG3, PbATG7, PbATG8, and P. berghei α-tubulin1, and the relative expression of PbATG3, PbATG7, and PbATG8 vs. the housekeeping gene of P. berghei α-tubulin1 was determined by the ΔC
T values.59
Recombinant PbATG8 expression and affinity purificationFull-length PbATG8 was cloned from blood stage parasite
RNA by RT-PCR using the ABI high-capacity cDNA Reverse Transcription Kit and primers 23 and 31. The approximately 400 bp PCR product was then digested with BamHI and HindIII, and ligated into the PQE-30 vector (Qiagen, 33203) to generate an N-terminal 6-His tagged fusion protein. His-tagged PbATG8 was expressed in E. coli SG13009. Bacteria were grown in LB, and when cultures reached the O.D.
600 of 0.6, expression was
induced with 1 mM IPTG for 3 h at 37 °C. After centrifugation, bacteria were resuspended in buffer A, containing 100 mM NaCl, 10 mM imidazole and 20 mM phosphate (pH of 7.2) and lysed by sonication with 60 pulses at 50% duty cycle and an output of 50 followed by 1 min on ice. The lysate was then cleared by centrifugation at 1,100 g for 10 min and the pellet was resuspended in buffer A containing 6 M urea. Total lysate was loaded onto an HP Nickel Column (HiTrap chelating HP from GE Healthcare, 17-0408-01), washed with 6 volume of buffer A containing 20 mM imidizole and His-PbATG8 was eluted with 250 mM imidazole. Fractions were separated on a 15% acrylamide gel, Coomassie blue-stained and the band corresponding to His-PbATG8 was cut, destained, lyophilized, and used for immunization in rat and mice (Covance).
Subcellular fractionationWhole blood with asynchronous P. berghei was incubated in
0.05% saponin in PBS for 15 min at room temperature to isolate parasites from RBC. All the following steps were performed at 4 °C. Parasite pellets were lysed by glass beads in buffer containing 50 mM TRIS-HCl pH 7.5, 150 mM NaCl, 5 mM EDTA, 5 mM EGTA, 1 mM PMSF and 1× Complete Protease Inhibitor Cocktail (Roche, 04 693 116 001). Cell debris and intact erythrocytes were removed by centrifugation at
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
282 autophagy Volume 10 issue 2
800 g for 5 min. The supernatant was treated with either urea at 2 M or Triton X-100 at 2% (final concentrations) and samples were shaken for 1 h before centrifugation at 100,000 g for 1 h. Proteins were then precipitated with ice-cold TCA, and protein pellets were washed with acetone, resuspended in SDS-PAGE sample buffer, and analyzed by immunoblotting with anti-PbATG8 antibodies.
Western blottingThe specificity of anti-PbATG8 antibodies was verified by
western blot at a dilution of 1:1000 against blood stage parasite lysates. To prepare lysates, sporozoites were washed 2 times in PBS and blood stage parasites were isolated by saponin lysis and resuspended in buffer consisting of 50 mM TRIS-HCl pH 7.5, 150 mM NaCl, 5 mM EDTA, 5 mM EGTA, I mM PMSF and 1× Complete Protease Inhibitor Cocktail (Roche, 04 693 116 001). Laemmli buffer was added to a final concentration of 1×; samples were boiled for 5 min and centrifuged briefly prior to separation by SDS-PAGE on a 4–15% TRIS-HCl gradient gel. Proteins were then transferred to nitrocellulose and blocked with 2% dehydrated milk and 0.1% Tween-20 in PBS. Anti-Rat-HRP, diluted at 1:10,000 in blocking buffer, was used to detect bound antibody (GE Healthcare, NA935). HRP was visualized using ECL Plus. For separation of ATG8 from ATG8-PE a 13.5% acrylamide gel containing 6 M urea was prepared as described.28 Sample buffer and running buffer also contained 6 M urea. Western blotting was performed using standard procedures as described above.
Yeast 2-hybrid assaysFor yeast 2-hybrid assays, the plasmids pGBK-T7 (630489)
and pGAD-T7 (630442), and the AH109 strain of yeast (630444) were used (Clontech). Primers and restriction sites used to clone the PbATG3 and PbATG8 ORF and mutants into pGBK-T7 and pGAD-T7 are detailed in Table S1. Yeast transformation and media was prepared according to Matchmaker Gal4 Yeast Two-Hybrid Manual (Clontech).
Yeast complementation assaysFor yeast complementation assays, an atg8 knockout strain
(13104) from Open Biosystems was used. ScAtg8, PbATG8, and PbATG8 mutants were expressed in the vector pSM-7 (gift of Marie Hardwick, Johns Hopkins University). PbATG8 mutants were generated using the QuikChange Multi Site-Directed Mutagenesis Kit (Stratagene, 200513), and primers used are summarized in Table S1. To test if precursor Ape1 maturation varied depending on the promoter driving expression, we used the vector TK108 from Yoshinori Ohsumi (Okazaki National Research Institutes), in which ScAtg8 is expressed under the control of its endogenous promoter.28 To assay the maturation of prAPe1, yeast strains were grown overnight at 30 °C in SD-uracil. Cells were then diluted to A
600 of 0.2 and allowed
to double twice at 30 °C. Cells were washed once in distilled water and 5 ml of culture was then harvested and resuspended in 100 μl of lysis buffer as described above under western blotting. Acid-washed glass beads were added to 30% of the final sample volume and the samples were mixed by vortexing for 3 pulses of 1 min each separated by 1 min on ice. Finally, Laemmli buffer was added at 1× and samples were boiled for 5 min, and stored
at −80 °C. Samples were centrifuged briefly prior to loading on SDS-PAGE gel for separation. To induce starvation, cells were cultured in SD-uracil and washed in distilled water as described above and 5 ml of culture was resuspended in SD lacking all amino acids for 1 h at 30 °C. Lysates were prepared as described above. Premature and mature forms of Ape1 were detected by western blotting as described above using mouse anti-Ape1 antibody 1532. Anti-rabbit-HRP antibodies were used at 1:10,000 to detect bound antibody.
Expression of Plasmodium ATG8 in mammalian cellsmCherry-tagged PfATG8 was generated by fusion PCR by
amplifying mCherry using primers 139 and 140, and PfATG8 using primers 141 and 142. These PCR products were then used as a template for a subsequent PCR using primers 139 and 142 to generate the mCherry-PfATG8 fusion protein, which was cloned into the vector pcDNA3.1 (Invitrogen, V790-20) for expression in COS cells. COS cells were transfected with 1 μg plasmid DNA using lipofectamine (Invitrogen, 11668-027). GFP-LC3 (generous gift of J Lippincott-Schwartz, NIH) was similarly transfected into COS cells.
Immunofluorescence assays (IFA)IFA on parasites or Plasmodium-infected cells were performed
as previously described.60 Briefly, cells were fixed in a solution consisting of 4% paraformaldehyde and 0.02% gluteraldehyde in PBS for 15 min, washed with PBS, permeabilized with 0.3% Triton X-100 for 5 min and washed 3 times with PBS. Samples were then blocked with 3% BSA dissolved in PBS for 45 min and probed with primary antibodies diluted in blocking buffer for 1 to 2 h. Samples were then washed 3 times and probed with secondary antibodies diluted in blocking buffer for 45 min. Coverslips were mounted onto glass microscope slides using ProLong Gold anti-fade mounting solution (Invitrogen, P36930) with or without DAPI. For P. falciparum-infected midgut staining, samples were blocked in 4% BSA/PBS overnight, permeabilized with 0.05% Triton X-100/PBS for 1 h, washed with PBS, and incubated overnight in rabbit anti-PbCap380 antibodies diluted into 4% BSA/PBS for IFA as described.61 All images were viewed with a Nikon Plan Apo 100× objective using a Nikon Eclipse 90i microscope (Nikon Instruments Inc.) and pictures were taken using a Hamamatsu ORCA-ER camera (Hamamatsu Photonics) and Volocity software. Images (a z-stack per field) were processed using iterative restoration (confidence limit 98% and iteration limit 25) and further processing was done using Adobe Photoshop software (Adobe Systems Inc).
Electron microscopyFor IEM, axenic sporozoites or intracellular parasites were
fixed in 4% paraformaldehyde (Electron Microscopy Sciences, 15710) in 0.25 M HEPES (ph7.4) for 1 h at RT, then in 8% PFA in the same buffer overnight at 4 °C. They were infiltrated, frozen, and sectioned as previously described.62 The sections of P. berghei and P. falciparum were immunolabeled with antibodies against PbATG8 or mCherry, respectively, then with mouse anti-IgG antibodies conjugated directly to 10 nm protein A-gold particles before examination with a Philips CM120 electron microscope (Philips Electronics) under 80-kV. Stereological
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
www.landesbioscience.com autophagy 283
analysis of gold labeling to quantify the distribution of PbATG8 gold particles was performed as described.63
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
The authors are grateful to DJ Klionsky and J Lippincott-Schwartz for their helpful discussions on autophagy. We thank the generous providers of strains, antibodies and plasmids, and Usha Nair at the University of Michigan for her assistance in the analysis of PbATG8 conformation. We also wish to thank Dabeiba Bernal-Rubio, Sandra Gonzalez and Jean Nonan from New York University School of Medicine and Chris Kizito from
the JHMRI Insectary for their excellent technical support in the rearing and infection of the mosquitoes, and the competent technical staffs from the Microscopy Facility at Yale University and Johns Hopkins University. We also thank Kim Lee Sim and Stephen Hoffman, Sanaria Inc. (Rockville, MD) for providing infected mosquito samples. This research was supported by the Johns Hopkins Malaria Research Institute and the Bloomberg Family Foundation. DF gratefully acknowledges support from the NIH (R01 AI085584). BJ was a recipient of a predoctoral MRI fellowship and CV is a recipient of a postdoctoral MRI fellowship.
Supplemental Materials
Supplemental materials may be found here:www.landesbioscience.com/journals/autophagy/article/27166
References1. Prudêncio M, Rodriguez A, Mota MM. The silent
path to thousands of merozoites: the Plasmodium liver stage. Nat Rev Microbiol 2006; 4:849-56; PMID:17041632; http://dx.doi.org/10.1038/nrmicro1529
2. Jayabalasingham B, Bano N, Coppens I. Metamorphosis of the malaria parasite in the liver is associated with organelle clearance. Cell Res 2010; 20:1043-59; PMID:20567259; http://dx.doi.org/10.1038/cr.2010.88
3. Kaiser K, Camargo N, Kappe SH. Transformation of sporozoites into early exoerythrocytic malaria parasites does not require host cells. J Exp Med 2003; 197:1045-50; PMID:12707302; http://dx.doi.org/10.1084/jem.20022100
4. Meis JF, Verhave JP, Jap PH, Meuwissen JH. Transformation of sporozoites of Plasmodium berghei into exoerythrocytic forms in the liver of its mammalian host. Cell Tissue Res 1985; 241:353-60; PMID:3896506; http://dx.doi.org/10.1007/BF00217180
5. Meis JF, Verhave JP, Jap PH, Sinden RE, Meuwissen JH. Malaria parasites--discovery of the early liver form. Nature 1983a; 302:424-6; PMID:6339945; http://dx.doi.org/10.1038/302424a0
6. Meis JF, Verhave JP, Jap PH, Sinden RE, Meuwissen JH. Ultrastructural observations on the infection of rat liver by Plasmodium berghei sporozoites in vivo. J Protozool 1983b; 30:361-6; PMID:6355454; http://dx.doi.org/10.1111/j.1550-7408.1983.tb02931.x
7. Mizushima N, Komatsu M. Autophagy: renovation of cells and tissues. Cell 2011; 147:728-41; PMID:22078875; http://dx.doi.org/10.1016/j.cell.2011.10.026
8. Abrahamsen H, Stenmark H. Protein secretion: unconventional exit by exophagy. Curr Biol 2010; 20:R415-8; PMID:20462486; http://dx.doi.org/10.1016/j.cub.2010.03.011
9. Ushio H, Ueno T, Kojima Y, Komatsu M, Tanaka S, Yamamoto A, Ichimura Y, Ezaki J, Nishida K, Komazawa-Sakon S, et al. Crucial role for autophagy in degranulation of mast cells. J Allergy Clin Immunol 2011; 127:1267-76, e6; PMID:21333342; http://dx.doi.org/10.1016/j.jaci.2010.12.1078
10. Coppens I. Metamorphoses of malaria: the role of autophagy in parasite differentiation. Essays Biochem 2011; 51:127-36; PMID:22023446
11. Duszenko M, Ginger ML, Brennand A, Gualdrón-López M, Colombo MI, Coombs GH, Coppens I, Jayabalasingham B, Langsley G, de Castro SL, et al. Autophagy in protists. Autophagy 2011; 7:127-58; PMID:20962583; http://dx.doi.org/10.4161/auto.7.2.13310
12. Brennand A, Gualdrón-López M, Coppens I, Rigden DJ, Ginger ML, Michels PA. Autophagy in parasitic protists: unique features and drug targets. Mol Biochem Parasitol 2011; 177:83-99; PMID:21315770; http://dx.doi.org/10.1016/j.molbiopara.2011.02.003
13. Besteiro S, Brooks CF, Striepen B, Dubremetz JF. Autophagy protein Atg3 is essential for maintaining mitochondrial integrity and for normal intracellular development of Toxoplasma gondii tachyzoites. PLoS Pathog 2011; 7:e1002416; PMID:22144900; http://dx.doi.org/10.1371/journal.ppat.1002416
14. Herman M, Pérez-Morga D, Schtickzelle N, Michels PA. Turnover of glycosomes during life-cycle differentiation of Trypanosoma brucei. Autophagy 2008; 4:294-308; PMID:18365344
15. Koopmann R, Muhammad K, Perbandt M, Betzel C, Duszenko M. Trypanosoma brucei ATG8: structural insights into autophagic-like mechanisms in protozoa. Autophagy 2009; 5:1085-91; PMID:19736525; http://dx.doi.org/10.4161/auto.5.8.9611
16. Besteiro S, Williams RA, Morrison LS, Coombs GH, Mottram JC. Endosome sorting and autophagy are essential for differentiation and virulence of Leishmania major. J Biol Chem 2006; 281:11384-96; PMID:16497676; http://dx.doi.org/10.1074/jbc.M512307200
17. Alvarez VE, Kosec G, Sant’Anna C, Turk V, Cazzulo JJ, Turk B. Autophagy is involved in nutritional stress response and differentiation in Trypanosoma cruzi. J Biol Chem 2008b; 283:3454-64; PMID:18039653; http://dx.doi.org/10.1074/jbc.M708474200
18. Picazarri K, Nakada-Tsukui K, Nozaki T. Autophagy during proliferation and encystation in the protozoan parasite Entamoeba invadens. Infect Immun 2008; 76:278-88; PMID:17923513; http://dx.doi.org/10.1128/IAI.00636-07
19. Moon EK, Chung DI, Hong YC, Kong HH. Autophagy protein 8 mediating autophagosome in encysting Acanthamoeba. Mol Biochem Parasitol 2009; 168:43-8; PMID:19560492; http://dx.doi.org/10.1016/j.molbiopara.2009.06.005
20. Williams RA, Smith TK, Cull B, Mottram JC, Coombs GH. ATG5 is essential for ATG8-dependent autophagy and mitochondrial homeostasis in Leishmania major. PLoS Pathog 2012; 8:e1002695; PMID:22615560; http://dx.doi.org/10.1371/journal.ppat.1002695
21. Rigden DJ, Michels PA, Ginger ML. Autophagy in protists: Examples of secondary loss, lineage-specific innovations, and the conundrum of remodeling a single mitochondrion. Autophagy 2009; 5:784-94; PMID:19483474
22. Kitamura K, Kishi-Itakura C, Tsuboi T, Sato S, Kita K, Ohta N, Mizushima N. Autophagy-related Atg8 localizes to the apicoplast of the human malaria parasite Plasmodium falciparum. PLoS One 2012; 7:e42977; PMID:22900071; http://dx.doi.org/10.1371/journal.pone.0042977
23. Eickel N, Kaiser G, Prado M, Burda PC, Roelli M, Stanway RR, Heussler VT. Features of autophagic cell death in Plasmodium liver-stage parasites. Autophagy 2013; 9:568-80; PMID:23388496; http://dx.doi.org/10.4161/auto.23689
24. Mizushima N. The pleiotropic role of autophagy: from protein metabolism to bactericide. Cell Death Differ 2005; 12(Suppl 2):1535-41; PMID:16247501; http://dx.doi.org/10.1038/sj.cdd.4401728
25. Xie Z, Nair U, Klionsky DJ. Atg8 controls phagophore expansion during autophagosome formation. Mol Biol Cell 2008; 19:3290-8; PMID:18508918; http://dx.doi.org/10.1091/mbc.E07-12-1292
26. Paz Y, Elazar Z, Fass D. Structure of GATE-16, membrane transport modulator and mammalian ortholog of autophagocytosis factor Aut7p. J Biol Chem 2000; 275:25445-50; PMID:10856287; http://dx.doi.org/10.1074/jbc.C000307200
27. Bavro VN, Sola M, Bracher A, Kneussel M, Betz H, Weissenhorn W. Crystal structure of the GABA(A)-receptor-associated protein, GABARAP. EMBO Rep 2002; 3:183-9; PMID:11818336; http://dx.doi.org/10.1093/embo-reports/kvf026
28. Kirisako T, Baba M, Ishihara N, Miyazawa K, Ohsumi M, Yoshimori T, Noda T, Ohsumi Y. Formation process of autophagosome is traced with Apg8/Aut7p in yeast. J Cell Biol 1999; 147:435-46; PMID:10525546; http://dx.doi.org/10.1083/jcb.147.2.435
29. Kirisako T, Ichimura Y, Okada H, Kabeya Y, Mizushima N, Yoshimori T, Ohsumi M, Takao T, Noda T, Ohsumi Y. The reversible modification regulates the membrane-binding state of Apg8/Aut7 essential for autophagy and the cytoplasm to vacuole targeting pathway. J Cell Biol 2000; 151:263-76; PMID:11038174; http://dx.doi.org/10.1083/jcb.151.2.263
30. Noda NN, Satoo K, Fujioka Y, Kumeta H, Ogura K, Nakatogawa H, Ohsumi Y, Inagaki F. Structural basis of Atg8 activation by a homodimeric E1, Atg7. Mol Cell 2011; 44:462-75; PMID:22055191; http://dx.doi.org/10.1016/j.molcel.2011.08.035
31. Hain AU, Weltzer RR, Hammond H, Jayabalasingham B, Dinglasan RR, Graham DR, Colquhoun DR, Coppens I, Bosch J. Structural characterization and inhibition of the Plasmodium Atg8-Atg3 interaction. J Struct Biol 2012; 180:551-62; PMID:22982544; http://dx.doi.org/10.1016/j.jsb.2012.09.001
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015

©20
14 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
284 autophagy Volume 10 issue 2
32. Yamada Y, Suzuki NN, Hanada T, Ichimura Y, Kumeta H, Fujioka Y, Ohsumi Y, Inagaki F. The crystal structure of Atg3, an autophagy-related ubiquitin carrier protein (E2) enzyme that mediates Atg8 lipidation. J Biol Chem 2007; 282:8036-43; PMID:17227760; http://dx.doi.org/10.1074/jbc.M611473200
33. Nakatogawa H, Ichimura Y, Ohsumi Y. Atg8, a ubiquitin-like protein required for autophagosome formation, mediates membrane tethering and hemifusion. Cell 2007; 130:165-78; PMID:17632063; http://dx.doi.org/10.1016/j.cell.2007.05.021
34. Williams RA, Woods KL, Juliano L, Mottram JC, Coombs GH. Characterization of unusual families of ATG8-like proteins and ATG12 in the protozoan parasite Leishmania major. Autophagy 2009; 5:159-72; PMID:19066473; http://dx.doi.org/10.4161/auto.5.2.7328
35. Klionsky DJ, Cueva R, Yaver DS. Aminopeptidase I of Saccharomyces cerevisiae is localized to the vacuole independent of the secretory pathway. J Cell Biol 1992; 119:287-99; PMID:1400574; http://dx.doi.org/10.1083/jcb.119.2.287
36. Seguí-Real B, Martinez M, Sandoval IV. Yeast aminopeptidase I is post-translationally sorted from the cytosol to the vacuole by a mechanism mediated by its bipartite N-terminal extension. EMBO J 1995; 14:5476-84; PMID:8521804
37. Striepen B. The apicoplast: a red alga in human parasites. Essays Biochem 2011; 51:111-25; PMID:22023445
38. Mizushima N. The pleiotropic role of autophagy: from protein metabolism to bactericide. Cell Death Differ 2005; 12(Suppl 2):1535-41; PMID:16247501; http://dx.doi.org/10.1038/sj.cdd.4401728
39. Hanssen E, Goldie KN, Tilley L. Ultrastructure of the Asexual Blood Stages of Plasmodium falciparum, In: Thomas Müller-Reichert, Editor(s). Methods in Cell Biology, Academic Press, 2010; 96:93-116.
40. Carter V, Nacer AM, Underhill A, Sinden RE, Hurd H. Minimum requirements for ookinete to oocyst transformation in Plasmodium. Int J Parasitol 2007; 37:1221-32; PMID:17482621; http://dx.doi.org/10.1016/j.ijpara.2007.03.005
41. Baton LA, Ranford-Cartwright LC. Spreading the seeds of million-murdering death: metamorphoses of malaria in the mosquito. Trends Parasitol 2005; 21:573-80; PMID:16236552; http://dx.doi.org/10.1016/j.pt.2005.09.012
42. Ichimura Y, Kirisako T, Takao T, Satomi Y, Shimonishi Y, Ishihara N, Mizushima N, Tanida I, Kominami E, Ohsumi M, et al. A ubiquitin-like system mediates protein lipidation. Nature 2000; 408:488-92; PMID:11100732; http://dx.doi.org/10.1038/35044114
43. Amar N, Lustig G, Ichimura Y, Ohsumi Y, Elazar Z. Two newly identified sites in the ubiquitin-like protein Atg8 are essential for autophagy. EMBO Rep 2006; 7:635-42; PMID:16680092
44. Tarun AS, Peng X, Dumpit RF, Ogata Y, Silva-Rivera H, Camargo N, Daly TM, Bergman LW, Kappe SH. A combined transcriptome and proteome survey of malaria parasite liver stages. Proc Natl Acad Sci U S A 2008; 105:305-10; PMID:18172196; http://dx.doi.org/10.1073/pnas.0710780104
45. Tooze SA, Yoshimori T. The origin of the autophagosomal membrane. Nat Cell Biol 2010; 12:831-5; PMID:20811355; http://dx.doi.org/10.1038/ncb0910-831
46. Hailey DW, Rambold AS, Satpute-Krishnan P, Mitra K, Sougrat R, Kim PK, Lippincott-Schwartz J. Mitochondria supply membranes for autophagosome biogenesis during starvation. Cell 2010; 141:656-67; PMID:20478256; http://dx.doi.org/10.1016/j.cell.2010.04.009
47. Schieke SM, Phillips D, McCoy JP Jr., Aponte AM, Shen RF, Balaban RS, Finkel T. The mammalian target of rapamycin (mTOR) pathway regulates mitochondrial oxygen consumption and oxidative capacity. J Biol Chem 2006; 281:27643-52; PMID:16847060; http://dx.doi.org/10.1074/jbc.M603536200
48. Seeber F, Soldati-Favre D. Metabolic pathways in the apicoplast of apicomplexa. Int Rev Cell Mol Biol 2010; 281:161-228; PMID:20460186; http://dx.doi.org/10.1016/S1937-6448(10)81005-6
49. Tawk L, Dubremetz JF, Montcourrier P, Chicanne G, Merezegue F, Richard V, Payrastre B, Meissner M, Vial HJ, Roy C, et al. Phosphatidylinositol 3-monophosphate is involved in toxoplasma apicoplast biogenesis. PLoS Pathog 2011; 7:e1001286; PMID:21379336; http://dx.doi.org/10.1371/journal.ppat.1001286
50. Raiborg C, Schink KO, Stenmark H. Class III phosphatidylinositol 3-kinase and its catalytic product PtdIns3P in regulation of endocytic membrane traffic. FEBS J 2013; 280:2730-42; PMID:23289851; http://dx.doi.org/10.1111/febs.12116
51. Burman C, Ktistakis NT. Regulation of autophagy by phosphatidylinositol 3-phosphate. FEBS Lett 2010; 584:1302-12; PMID:20074568; http://dx.doi.org/10.1016/j.febslet.2010.01.011
52. Funderburk SF, Wang QJ, Yue Z. The Beclin 1-VPS34 complex--at the crossroads of autophagy and beyond. Trends Cell Biol 2010; 20:355-62; PMID:20356743; http://dx.doi.org/10.1016/j.tcb.2010.03.002
53. Vaid A, Ranjan R, Smythe WA, Hoppe HC, Sharma P. PfPI3K, a phosphatidylinositol-3 kinase from Plasmodium falciparum, is exported to the host erythrocyte and is involved in hemoglobin trafficking. Blood 2010; 115:2500-7; PMID:20093402; http://dx.doi.org/10.1182/blood-2009-08-238972
54. Kabeya Y, Mizushima N, Yamamoto A, Oshitani-Okamoto S, Ohsumi Y, Yoshimori T. LC3, GABARAP and GATE16 localize to autophagosomal membrane depending on form-II formation. J Cell Sci 2004; 117:2805-12; PMID:15169837; http://dx.doi.org/10.1242/jcs.01131
55. Zhou B, Boudreau N, Coulber C, Hammarback J, Rabinovitch M. Microtubule-associated protein 1 light chain 3 is a fibronectin mRNA-binding protein linked to mRNA translation in lamb vascular smooth muscle cells. J Clin Invest 1997; 100:3070-82; PMID:9399954; http://dx.doi.org/10.1172/JCI119862
56. Cann GM, Guignabert C, Ying L, Deshpande N, Bekker JM, Wang L, Zhou B, Rabinovitch M. Developmental expression of LC3alpha and beta: absence of fibronectin or autophagy phenotype in LC3beta knockout mice. Dev Dyn 2008; 237:187-95; PMID:18069693; http://dx.doi.org/10.1002/dvdy.21392
57. Meléndez A, Tallóczy Z, Seaman M, Eskelinen EL, Hall DH, Levine B. Autophagy genes are essential for dauer development and life-span extension in C. elegans. Science 2003; 301:1387-91; PMID:12958363; http://dx.doi.org/10.1126/science.1087782
58. Trager W, Jensen JB. Human malaria parasites in continuous culture. Science 1976; 193:673-5; PMID:781840; http://dx.doi.org/10.1126/science.781840
59. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001; 25:402-8; PMID:11846609; http://dx.doi.org/10.1006/meth.2001.1262
60. Karsten V, Qi H, Beckers CJ, Joiner KA. Targeting the secretory pathway of Toxoplasma gondii. Methods 1997; 13:103-11; PMID:9405194; http://dx.doi.org/10.1006/meth.1997.0503
61. Srinivasan P, Fujioka H, Jacobs-Lorena M. PbCap380, a novel oocyst capsule protein, is essential for malaria parasite survival in the mosquito. Cell Microbiol 2008; 10:1304-12; PMID:18248630; http://dx.doi.org/10.1111/j.1462-5822.2008.01127.x
62. Quittnat F, Nishikawa Y, Stedman TT, Voelker DR, Choi JY, Zahn MM, Murphy RC, Barkley RM, Pypaert M, Joiner KA, et al. On the biogenesis of lipid bodies in ancient eukaryotes: synthesis of triacylglycerols by a Toxoplasma DGAT1-related enzyme. Mol Biochem Parasitol 2004; 138:107-22; PMID:15500922; http://dx.doi.org/10.1016/j.molbiopara.2004.08.004
63. Lucocq JM, Habermann A, Watt S, Backer JM, Mayhew TM, Griffiths G. A rapid method for assessing the distribution of gold labeling on thin sections. J Histochem Cytochem 2004; 52:991-1000; PMID:15258174; http://dx.doi.org/10.1369/jhc.3A6178.2004
Dow
nloa
ded
by [
John
s H
opki
ns U
nive
rsity
] at
08:
31 1
5 Se
ptem
ber
2015
Copyright © 2022 FDOKUMEN




















