
The betagamma-crystallin superfamily contains a universal motif for binding calcium

R E S EA RCH L E T T E R
Characterization of tetA-like gene encoding for a majorfacilitator superfamily efflux pump in Streptococcus
thermophilus
Stefania Arioli1, Simone Guglielmetti1, Stefano Amalfitano2, Carlo Viti3, Emmanuela Marchi3,Francesca Decorosi3, Luciana Giovannetti3 & Diego Mora1
1Department of Food Environmental and Nutritional Sciences (DeFENS), University of Milan, Milan, Italy; 2Istituto di Ricerca sulle Acque (IRSA-
CNR), Monterotondo, Rome, Italy; and 3Dipartimento di Scienze delle Produzioni Agroalimentari e dell’Ambiente – sezione di Microbiologia,
University of Florence, Florence, Italy
Correspondence: Stefania Arioli,
Department of Food Sciences and Nutrition
(DeFENS), University of Milan, via Mangiagalli
25, 20133 Milan, Italy.
Tel.: 0039 02 503 19133;
fax: 0039 02 503 19138;
e-mail: [email protected]
Received 5 March 2014; revised 7 April
2014; accepted 22 April 2014. Final version
published online 12 May 2014.
DOI: 10.1111/1574-6968.12449
Editor: Mark Enright
Keywords
antibiotic resistance; lactic acid bacteria; MFS
efflux pump; Streptococcus thermophilus.
Abstract
Efflux pumps are membrane proteins involved in the active extrusion of a wide
range of structurally dissimilar substrates from cells. A multidrug efflux pump
named TetA belonging to the major facilitator superfamily (MFS) of transport-
ers was identified in the Streptococcus thermophilus DSM 20617T genome. The
tetA-like gene was found in the genomes of a number of S. thermophilus strains
sequenced to date and in Streptococcus macedonicus ACA-DC 198, suggesting a
possible horizontal gene transfer event between these two Streptococcus species,
which are both adapted to the milk environment. Flow cytometry (single-cell)
analysis revealed bistable TetA activity in the S. thermophilus population, and
tetA-like gene over-expression resulted in a reduced susceptibility to ethidium
bromide, tetracycline, and other toxic compounds even when the efflux pump
was over-expressed in a strain naturally lacking tetA-like gene.
Introduction
Both bacterial and eukaryotic cells are usually equipped
with various types of membrane transport systems that
are involved in a number of vital functions. Indeed, such
carriers transport nutrients and ions into the cell, excrete
waste products and toxic compounds from the cell, and
are involved in the maintenance of cellular homeostasis
(Paulsen et al., 1996; Simm et al., 2012). Moreover,
several of these transport/efflux systems have been
demonstrated to play an important role in bacterial and
eukaryotic cells by conferring resistance to antimicrobial
molecules. Export systems that can manage a wide range
of structurally dissimilar substrates are defined as multi-
drug exporters or multidrug resistance (MDR) efflux
pumps (Paulsen et al., 1996; Floyd et al., 2010). Although
genes encoding selective drug efflux pumps are found on
transferable genetic elements or plasmids, those encoding
multidrug efflux pumps are generally located on the bac-
terial chromosome (Putman et al., 2000). Multidrug
transporters are considered to play a significant role in
the resistance of bacteria to antimicrobial compounds (Li
& Nikaido, 2004; Poole, 2007; Simm et al., 2012), and
these systems are energy-dependent membrane proteins
able to extrude a wide array of structurally and
functionally dissimilar compounds. On the basis of
bioenergetic and structural features, multidrug transport-
ers can be divided into two major groups: (1) primary
active ABC transporters, which utilize the free energy of
ATP binding/hydrolysis to efflux toxic substrates; and (2)
secondary active transporters, which derive energy to
mediate drug extrusion from a coupled exchange with H+
and/or Na+ ions (van Veen & Konings, 1997; Putman
et al., 2000; Wang & van Veen, 2012). Approximately
25% of all known membrane transport proteins in
prokaryotes belong to the major facilitator superfamily
(MFS; Saier et al., 1999), the largest and most diverse
superfamily of secondary active transporters (Reddy et al.,
FEMS Microbiol Lett 355 (2014) 61–70 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
MIC
ROBI
OLO
GY
LET
TER
S

2012), comprising 58 families involved in the symport,
antiport, or uniport of various substrate (Floyd et al.,
2010). Streptococcus thermophilus is a lactic acid bacterium
that is widely used for the preparation of several dairy
products, such as fermented milk, yogurt, and cheese
(Hols et al., 2005). Approximately 200 putative mem-
brane transporters have been identified on the basis of
S. thermophilus genome sequencing and according to the
TransportDB database (Ren et al., 2007), with 54% of the
drug efflux pumps belonging to the ABC superfamily and
23% belonging each to the multidrug/oligosaccharide/
polysaccharide superfamily and MFS (Lorca et al., 2007).
In this study, we focused our attention on the tetA-like
gene of S. thermophilus DSM 20617T, encoding a putative
MFS efflux pump potentially involved in reduced suscep-
tibility to cytotoxic compounds. To understand the role
and activity of TetA in S. thermophilus DSM 20617T, we
over-expressed the tetA-like gene and evaluated its effects
on the sensitivity of this bacterium to several antimicro-
bial compounds. Moreover, the effect of tetA-like over-
expression on tetracycline (TET) and ethidium bromide
(EB) sensitivity was evaluated in a strain naturally lacking
this efflux pump.
Materials and methods
Bacterial strains, media, and growth conditions
Wild-type S. thermophilus DSM 20617T and MIMSt24
were maintained in M17 broth (Difco Laboratories,
Detroit, MI) supplemented with 20 g L�1 lactose at 37 °C. The recombinant strains MIM27 and MIM28 (tetA
over-expressing recombinant strain) and MIM20 and
MIM21 (harboring the empty vector pMIT5) were culti-
vated in the same medium with chloramphenicol at a
final concentration of 4 lg mL�1. Recombinant strains
MIM20 and MIM27 were derived from strain DSM
20617T, and recombinant strains MIM21 and MIM28
were derived from strain MIMSt24. Escherichia coli cells
harboring plasmid pNZ8048 (de Ruyter et al., 1996),
pMIT5, or pMIT5-tetA were routinely maintained under
aerobic conditions in Luria broth at 37 °C supplemented
with 25 lg mL�1 chloramphenicol.
Vector construction
Plasmid pNZ8048 is a broad-spectrum host shuttle vector
that possesses the nisin-inducible nisA promoter and a
chloramphenicol resistance gene as the selection marker
(de Ruyter et al., 1996). The nisA promoter of pNZ8048
was replaced by a strong phage T5 promoter and lac
operator (Loimaranta et al., 1998; Guglielmetti et al.,
2008), generating pMIT5, as follows: the T5 promoter
was amplified using pCSS945 DNA as the template
(Arioli et al., 2010) with primers T5-BglF (50-TCTAGGAAAGATCTCAACGGAGGACTAGCGTA-30) and T5-
NcoR (50-CCTTGGAACCATGGTCATCATGGATCCCCTCAT-30), which contain the restriction sites of BglII and
NcoR, respectively (underlined). tetA-like gene (EMBL
accession number HG424699) was identified in the
whole-genome sequence of S. thermophilus DSM 20617T.
Pure genomic DNA was extracted and purified using the
UltraCleanTM Microbial DNA Isolation Kit (MO BIO,
CA). tetA was amplified using the primers tetA-BamHI-F
(50-TCTAGGAAGGATCCATTAAATATAATTTTTTCATTATCT-30) and tetA-PstI- (50-CCTTGGAACTGCAGTTAAATATAATTTTTAATAAAGTAATTT-30), which contain
BamHI and PstI restriction sites, respectively (under-
lined). tetA was then ligated to pMIT5 to generate
pMIT5-tetA. The ligation mixture was introduced into
One ShotR MatchTM-T1R Chemically competent E. coli
cells (Invitrogen) by heat shock (30 s at 42 °C). The
transformed cells were plated on Luria-agar containing
chloramphenicol (25 lg mL�1).
Streptococcus thermophilus DSM 20617T or MIMSt24
electrocompetent cells were prepared and transformed
with either pMIT5 or pMIT5-tetA, as previously described
(Arioli et al., 2007). The recombinant strains were
selected by plating on M17 lactose (20 g L�1) agar con-
taining 4 lg mL�1 of chloramphenicol after incubation
of 48 h at 37 °C under anaerobic conditions.
Genomic comparison among S. thermophilus and
Streptococcus macedonicus strains was carried out using
rapid annotation using subsystem technology (RAST) Ser-
ver (Aziz et al., 2008) and The SEED VIEWER version 2.0
(Overbeeck et al., 2005).
tetA transcriptional analysis
A transcriptional analysis of tetA-like gene was performed
by RT-qPCR in the S. thermophilus DSM 20617T wild-
type strain and in the recombinant tetA over-expressing
MIM27 strain following a previously described protocol
(Arioli et al., 2010). Total RNA was extracted using the
AurumTM Total RNA Fatty and Fibrous Tissue Kit
(Bio-Rad) from cells collected at an OD600 of 0.5 from
cultures grown in M17 broth with 20 g L�1 lactose in the
presence or absence of 0.125 lg mL�1 of chlorhexidine
(CH) or 0.5 or 0.125 lg mL�1 EB. The concentration
and purity of the purified RNA was analyzed using a
Take3TM Multi-Volume Plate (BioTek). cDNA was syn-
thesized using the iScriptTM cDNA synthesis kit (Bio-Rad)
with random primers. The resultant cDNA was subjected
to qRT-PCR; a control PCR of the same samples without
reverse transcriptase was included to assure the absence
of DNA contamination. The primers used are the
FEMS Microbiol Lett 355 (2014) 61–70ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
62 S. Arioli et al.

following: tetAF 50-tgggattgcagcagcatttgaaag-30 and tetAR
50-ccacctataccactaccgataatc-30 for tetA-like amplification;
ureCF 50-gctgacggtagtaatgcaacaa-30 and ureCR 50-gtgtgtcttaattcctgctgca-30 for ureC gene amplification;
murEF 50-tcaataggtccgatgtggtctgg-30 and murER 50-ccgtatgatggctgaagctgtatc-30 for murE amplification. The
results were normalized using murE and ureC, coding for
UDP-N-acetylmuramoylalanyl-D-glutamate-L-lysine ligase
and urease a-subunit, respectively, as reference genes. The
specificity of the primers was tested with melting curves
during amplification and by 1% agarose gels. For each
condition, we performed the relative quantification of
gene expression in triplicate with the cDNA synthesized
from two independent RNA samples. The PCR reactions
were performed as previously described (Arioli et al.,
2010) using a CFX96 thermocycler (Bio-Rad Laboratories,
Segrate, Italy). The data are expressed as the normalized
expression (DDCT) � standard error of the mean.
EB efflux assay
EB is a common efflux pump substrate that intercalates
with double-stranded nucleic acids in the cytoplasm,
thereby determining a fluorescence increase with the
proper excitation wavelength; as a substrate for a variety
of membrane pumps, EB is extruded across the mem-
brane, thus decreasing the overall measurable fluores-
cence (Lubelski et al., 2007; Czechowska & van der
Meer, 2012). For the EB efflux assay, strains MIM20
and MIM27 were grown until the mid-log phase (OD600
of 0.5). The cells were washed twice and resuspended in
sterile PBS (pH 7) in presence of 0.5 lg mL�1 EB with
and without two commonly employed efflux pump
inhibitors (EPIs): 0.1 mM of the protonophore carbonyl
cyanide m-chlorophenylhydrazone (CCCP) to disrupt
the electrochemical gradient across the membrane, thus
removing the proton gradient (Floyd et al., 2010); or
2 lM of reserpine (Schindler et al., 2013), a known
MDR EPI (Frempong-Manso et al., 2009), both pur-
chased from Sigma-Aldrich (Italy). This approach effec-
tively allowed the loading of the cells with EB after an
incubation time of 10 min at 37 °C; the cells were then
pelleted at 15 000 g for 1 min and resuspended in PBS.
To induce efflux activity, an energy source was applied
to the cells (10 g L�1 lactose, w/v, and urea 1 mM; Ari-
oli et al., 2010), thereby initiating cellular metabolism
and creating the electrochemical gradient across the
membrane that is necessary for efflux pump activity
(Floyd et al., 2010). The EB efflux was assessed at the
single-cell level by flow cytometry (Apogee A50-micro;
Apogee Flow System, UK) and by applying a double-
staining protocol with SYBR Green I (Molecular
Probes�; Invitrogen) and EB (Czechowska & van der
Meer, 2012). Under 488-nm laser beam excitation, the
SYBR signals were registered in the green channel (520/
10 nm), whereas the EB signals were registered in the
red channel (> 610 nm). A compensation factor (4%)
was calculated for a control sample and thus applied to
reduce the contribution of SYBR Green I to the red
channel. The HISTOGRAM software (version 2.6) was used
to plot the density diagrams (color plots and histo-
grams) of the log-transformed fluorescence signals of
each single cell.
Minimum inhibitory concentration and growth
fitness evaluations
The effect of tetA over-expression on the growth kinetic
parameters of MIM20 and MIM27 was determined using a
semi-automated microdilution method performed in a
384-well microtiter plate, as reported by Arioli et al.
(2013), in the absence (positive control) and presence of
six different concentrations of EB, CH, or TET. The effects
of each of the six concentrations of the above-mentioned
toxic compounds were tested in M17 broth in the presence
of two different concentrations of lactose (5 and 20 g L�1).
The strains were inoculated in quadruplicate at a final inoc-
ulum concentration of 105 bacteria mL�1 (with the bacte-
ria from cultures grown overnight to the stationary phase).
The bacterial cell concentration of the overnight culture
was determined microscopically with a Neubauer improved
counting chamber (Marienfeld GmbH, Lauda-K€Onigsho-
fen, Germany). To ensure anaerobic incubation, each well
was covered with 20 lL of sterile mineral oil (Sigma-
Aldrich). The microbial growth was monitored with a spec-
trophotometer (MicroWave RS2; Biotek) programmed for
145 readings (OD600) every 10 min for 24 h at 37 °C. Atthe end of the incubation, the lag time (h) was calculated
using the software GEN5 (Biotek) and reported as the mean
of four independent measurements � standard deviation.
Phenotype microarray assay
Phenotype microarray (PM) technology uses tetrazolium
violet reduction as an indicator of active metabolism; the
reduction of the dye causes the formation of a purple
color, which is recorded by a CCD camera every 15 min
to provide quantitative and kinetic information regarding
the response of the cells in PM plates (Bochner et al.,
2001). Strains MIM20 and MIM27 were tested by the PM
approach for 240 different compounds potentially exert-
ing toxic effects on bacterial cells; each was included at
four concentrations. To perform the PM experiments, the
strains were grown overnight at 37 °C on BUG agar (Bio-
log, Hayward, CA) supplemented with chloramphenicol
4 lg mL�1, and the cells were then collected with a sterile
FEMS Microbiol Lett 355 (2014) 61–70 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
TetA MFS efflux pump in Streptococcus thermophilus 63

cotton swab and suspended in 15-mL inoculation solu-
tion (IF-0; Biolog). The cell density was adjusted to 81%
transmittance using a Biolog turbidimeter. The PM11–PM20 inoculations were performed as described by Deco-
rosi et al. (2011), with the supplementation of chloram-
phenicol 2 lg mL�1. All the PM plates were incubated at
37 °C in an Omnilog Reader (Biolog) and monitored
automatically every 15 min for 72 h for color develop-
ment. Compounds giving bad kinetics over the four wells
(i.e., because the color development depended on the
increasing concentration of the compounds instead of the
real metabolic activity) were excluded from further analy-
sis. To identify the phenotypes gained or lost by MIM27
compared to MIM20, the kinetic curves obtained were
compared for the ‘Darea’ parameter using OMNILOG-PM
software (Biolog Inc) (release OM_PM_109M). Area
under the curve is expressed in arbitrary OmniLog Units
(AOU). We calculated the percentage of increase/decrease
of area of the MIM27 strain compared to MIM20. We
considered as significant all the compounds showing
Darea ≥ 100% in at least one of the four wells. We also
exploited the selection filter of the software, which high-
lights all the wells in which the chosen parameter exceeds
the setup limit. The distance limit for Darea parameter
was the setup at 15 000 AOU. We thus included also the
compounds selected by the software, giving an increase/
decrease of area values of at least 50%.
Results and discussion
Identification, sequence analysis, and tetA
distribution in S. thermophilus
Although MFS-type efflux pumps have been extensively
described, particularly for pathogenic bacteria, this is the
first characterization of this transport system in S. ther-
mophilus, a lactic acid bacterium. A genome analysis of
S. thermophilus DSM 20617T revealed that the 1170-
nucleotide-long tetA-like gene encodes a putative protein
of 389 amino acids. The tetA-like gene was found in the
whole genome of certain strains of S. thermophilus
(CNRZ1066, LMD-9, JIM 8232, and MN-ZLW-002) and
in S. macedonicus ACA-DC 198. In Fig. 1a is reported the
schematic organization of schematic organization of tetA-
like genomic locus as deduced by genomic comparison of
tetA-like gene from S. thermophilus DSM20617, and
S. thermophilus LMD9 and LMG18311, and S. macedoni-
cus ACA-DC198 genomes. Moreover, the nucleotide and
amino acid identities among the two species were found
to be 100% and 99%, respectively. The presence of the
IS1191 (Fig. 1a) could support the hypothesis of a possi-
ble horizontal gene transfer (HGT) event between these
two Streptococcus species adapted to the same milk envi-
ronment (Bolotin et al., 2004; Papadimitriou et al.,
2012).
A blast analysis of the deduced TetA amino acid
sequence revealed high identity with the multidrug trans-
porters of other Gram-positive bacteria, with highly con-
served motifs characteristic of the MFS family (Pao et al.,
1998). A more detailed amino acid sequence analysis
allowed the identification of a conserved structural and
functional domain TetA(P) (ExPxxxxxDxxxRK) (Fig. 1b),
which was previously characterized in the TET-resistant
proteins TetA(P) and Tet(40) of the anaerobes Clostrid-
ium perfringens and Clostridium saccharolyticum (Bannam
& Rood, 1999; Kazimierczak et al., 2008).
Because the tetA-like gene has not been detected in the
genome of all S. thermophilus strains sequenced thus far,
a PCR survey of the tetA-like gene was performed in a
collection of 100 strains isolated from different sources,
revealing the presence of the gene in 34% of the strains
(Table S1). Interestingly, only one of 20 Georgian strains
screened for the tetA-like gene was positive in the PCR
analysis. It is worth noting that all Georgian strains were
isolated from several homemade yogurt samples prepared
without any strain selection or characterization, whereas
industrial strains are usually selected for their robustness,
rapid growth, and acidification of milk.
Single-cell analysis of EB cell efflux
To investigate the role and activity of this putative efflux
pump, we constructed S. thermophilus DSM 20617T rec-
ombinants over-expressing the tetA gene. To this end, we
cloned the gene under the control of the T5 promoter,
thus allowing the constitutive expression of tetA. The
effect of tetA over-expression on the ability of the cell to
pump out EB was evaluated by analyzing the kinetics of
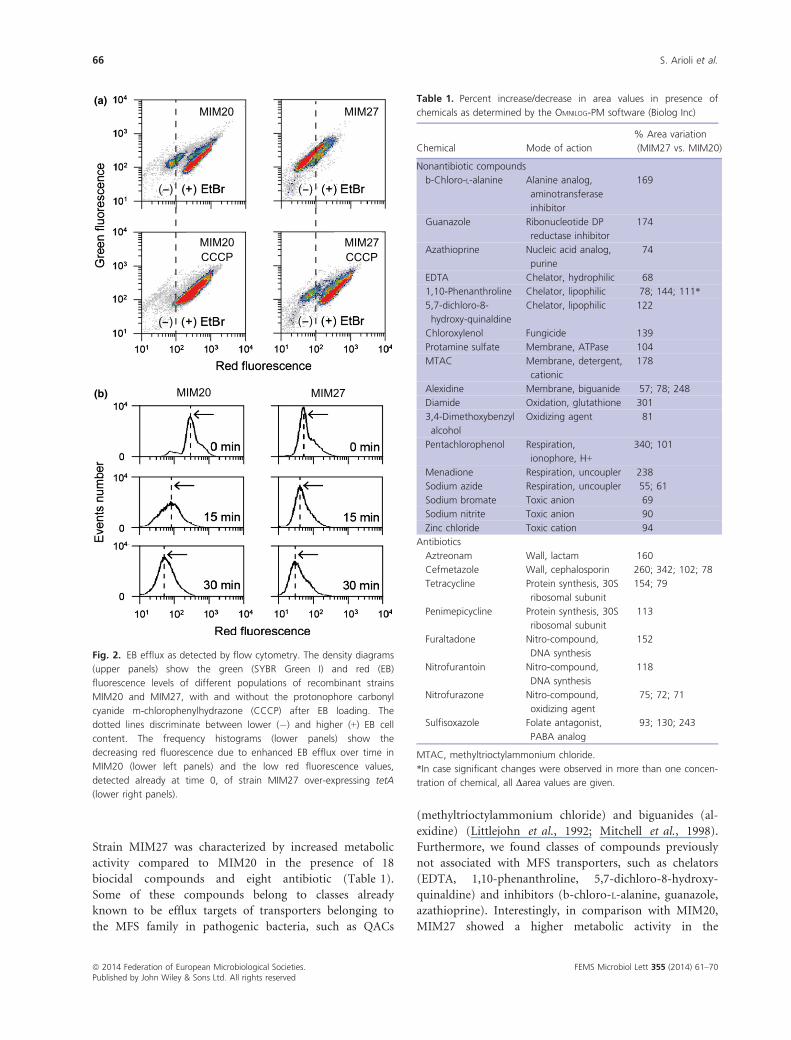
EB efflux at single-cell level by flow cytometry.
The EB efflux assay was performed with an EB concen-
tration that did not in any way affect cell viability, thus
allowing the detection of the extrusion activity of the
efflux pump system (Paix~ao et al., 2009). The tetA over-
expressing mutant MIM27 showed a more efficient EB
efflux compared to that of MIM20. In fact, the results
revealed a higher decrease in fluorescence after 30 min at
37 °C in MIM27 compared to the MIM20 control cells,
suggesting an active extrusion of EB from the cells over-
expressing tetA (Fig. 2b). Following the addition of the
protonophore CCCP (Fig. S1) or reserpine (data not
shown), the fluorescence of both the control and mutant
cells after 30 min was higher compared to the control
without EPIs due to the collapse of the proton gradient
across the membrane. This phenomenon could provide
indirect evidence for an H+-dependent activity of TetA,
as previously indicated for the characterization of LmrS,
FEMS Microbiol Lett 355 (2014) 61–70ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
64 S. Arioli et al.

an MFS multidrug efflux pump from Staphylococcus
aureus (Floyd et al., 2010). Interestingly, flow cytometry
revealed that the MIM27 population is extremely
homogeneous, whereas the MIM20 is composed of two
subpopulations identified on the basis of the ability to
efflux EB. The two populations observed for MIM20 were
also identified in the wild-type DSM 20617T (Fig. S2)
strain and in MIM27 when the cells were treated with
CCCP (Fig. 2), thus suggesting that TetA activity is only
partially blocked by the presence of the inhibitor in the
overall p3opulation over-expressing tetA. However, in the
absence of CCCP, the presence of two MIM20 subpopu-
lations underlined a potential transcriptional bistability of
tetA, a phenomenon observed for other phenotypes in
other bacterial species (Veening et al., 2008).
Substrate spectrum and transcriptional
analysis
Strains MIM20 and MIM27 were analyzed for chemical
sensitivity to 240 compounds, each at four concentrations
(PM11–PM20), using the BIOLOG Phenotype Microarray.
TetA(20617) Streptococcus thermophilus … 53 I L F E F P S G M L S D N W S R K 69… 389aa
TetA(408) Clostridium perfringens … 56 F I F E I P T G I V A D V Y S R K 72… 409aa
TetA(40) Clostridium saccharolyticum … 57 F L F E M P T G V V A D L Y S R R 73… 406aa
TetA(B) Escherichia coli … 55 V I F A P W L G K M S D R F G R R 71… 401aa
TetA(B)Motif G x x x x R x G R R
TetA(P)Motif E x P x x x x x D x x x R K
(a)
(b)
Ster_1914tetA-like
Ster_1913 mccB-like IS1191-trIS1191-tr
Ster_1919
Stu1947Stu1940 Stu1942
S. thermophilus LMD9/DSM20617
S. thermophilus LMG18311
SMA_1914tetA-like mccB-like
IS1191
S. macedonicus ACA-DC 198
Fig. 1. (a) Schematic organization of tetA-like genomic locus as deduced by genomic comparison of tetA-like gene from S. thermophilus
DSM20617, and S. thermophilus LMD9 and LMG18311, and Streptococcus macedonicus ACA-DC 198 genomes (accession numbers CP000419,
CP000023, HE613569). SMA_1914, putative transposase; SMA_1917, tetA-like gene; SMA_1918, mccB, putative bacteriocin biosynthesis gene,
Ster_1913 and its orthologs stu1940, coding for a putative adenylosuccinate synthase; ster_1914, coding for a putative DNA topology
modulation protein; IS1191, coding for IS256 transposase family; IS1191-tr, IS1191 truncated gene; Ster_1919 and its orthologs Stu1947, XRE
family transcript regulator; Stu1942, putative protein with unknown function; Genes identified by black arrows code for putative transposases. S.
thermophilus genes located upstream and the downstream the tetA-like and mccB-like genes are identified by light blue arrows. (b) Alignment of
the regions containing the TetA(P) motif. The prototype TET efflux protein TetA(B) and the previously identified TetA(B) motif are shown. The first
column lists the names of the protein included, followed by the host organism. The name of the first and last residues included is given, as well
as the total length of the protein.
FEMS Microbiol Lett 355 (2014) 61–70 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
TetA MFS efflux pump in Streptococcus thermophilus 65
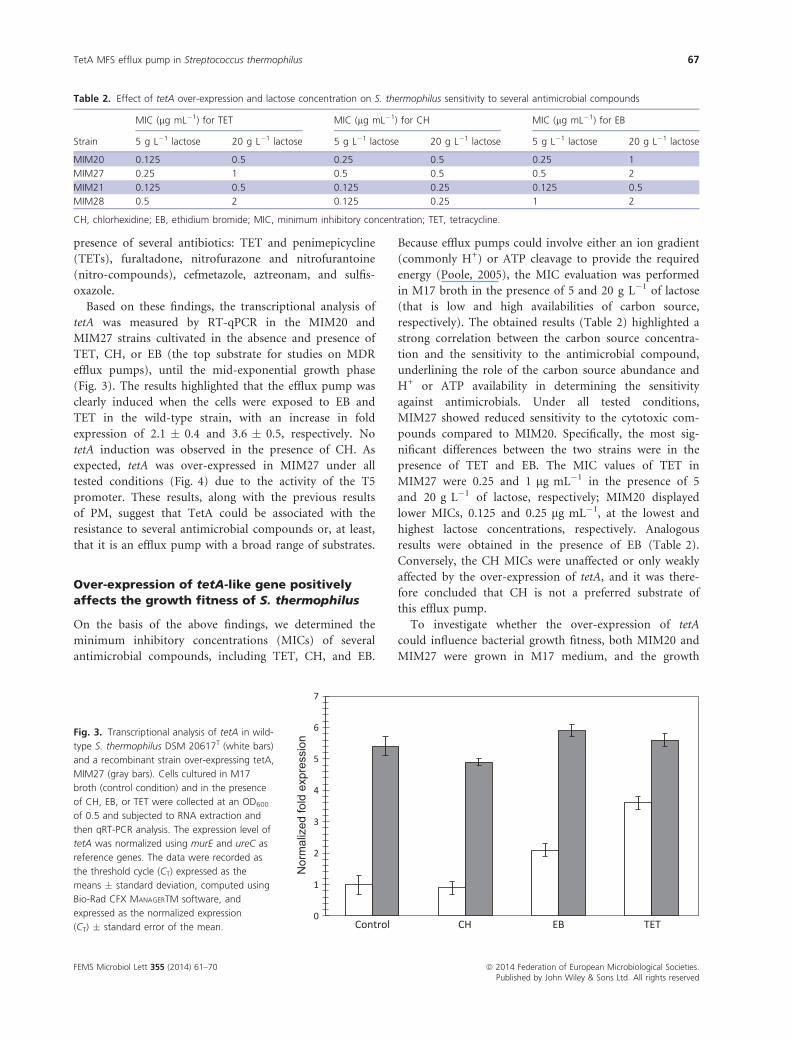
Strain MIM27 was characterized by increased metabolic
activity compared to MIM20 in the presence of 18
biocidal compounds and eight antibiotic (Table 1).
Some of these compounds belong to classes already
known to be efflux targets of transporters belonging to
the MFS family in pathogenic bacteria, such as QACs
(methyltrioctylammonium chloride) and biguanides (al-
exidine) (Littlejohn et al., 1992; Mitchell et al., 1998).
Furthermore, we found classes of compounds previously
not associated with MFS transporters, such as chelators
(EDTA, 1,10-phenanthroline, 5,7-dichloro-8-hydroxy-
quinaldine) and inhibitors (b-chloro-L-alanine, guanazole,
azathioprine). Interestingly, in comparison with MIM20,
MIM27 showed a higher metabolic activity in the
MIM20CCCP
MIM27CCCP
MIM20 MIM27
MIM20 MIM27
(a)
(b)
Fig. 2. EB efflux as detected by flow cytometry. The density diagrams
(upper panels) show the green (SYBR Green I) and red (EB)
fluorescence levels of different populations of recombinant strains
MIM20 and MIM27, with and without the protonophore carbonyl
cyanide m-chlorophenylhydrazone (CCCP) after EB loading. The
dotted lines discriminate between lower (�) and higher (+) EB cell
content. The frequency histograms (lower panels) show the
decreasing red fluorescence due to enhanced EB efflux over time in
MIM20 (lower left panels) and the low red fluorescence values,
detected already at time 0, of strain MIM27 over-expressing tetA
(lower right panels).
Table 1. Percent increase/decrease in area values in presence of
chemicals as determined by the OMNILOG-PM software (Biolog Inc)
Chemical Mode of action
% Area variation
(MIM27 vs. MIM20)
Nonantibiotic compounds
b-Chloro-L-alanine Alanine analog,
aminotransferase
inhibitor
169
Guanazole Ribonucleotide DP
reductase inhibitor
174
Azathioprine Nucleic acid analog,
purine
74
EDTA Chelator, hydrophilic 68
1,10-Phenanthroline Chelator, lipophilic 78; 144; 111*
5,7-dichloro-8-
hydroxy-quinaldine
Chelator, lipophilic 122
Chloroxylenol Fungicide 139
Protamine sulfate Membrane, ATPase 104
MTAC Membrane, detergent,
cationic
178
Alexidine Membrane, biguanide 57; 78; 248
Diamide Oxidation, glutathione 301
3,4-Dimethoxybenzyl
alcohol
Oxidizing agent 81
Pentachlorophenol Respiration,
ionophore, H+
340; 101
Menadione Respiration, uncoupler 238
Sodium azide Respiration, uncoupler 55; 61
Sodium bromate Toxic anion 69
Sodium nitrite Toxic anion 90
Zinc chloride Toxic cation 94
Antibiotics
Aztreonam Wall, lactam 160
Cefmetazole Wall, cephalosporin 260; 342; 102; 78
Tetracycline Protein synthesis, 30S
ribosomal subunit
154; 79
Penimepicycline Protein synthesis, 30S
ribosomal subunit
113
Furaltadone Nitro-compound,
DNA synthesis
152
Nitrofurantoin Nitro-compound,
DNA synthesis
118
Nitrofurazone Nitro-compound,
oxidizing agent
75; 72; 71
Sulfisoxazole Folate antagonist,
PABA analog
93; 130; 243
MTAC, methyltrioctylammonium chloride.
*In case significant changes were observed in more than one concen-
tration of chemical, all Darea values are given.
FEMS Microbiol Lett 355 (2014) 61–70ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
66 S. Arioli et al.
presence of several antibiotics: TET and penimepicycline
(TETs), furaltadone, nitrofurazone and nitrofurantoine
(nitro-compounds), cefmetazole, aztreonam, and sulfis-
oxazole.
Based on these findings, the transcriptional analysis of
tetA was measured by RT-qPCR in the MIM20 and
MIM27 strains cultivated in the absence and presence of
TET, CH, or EB (the top substrate for studies on MDR
efflux pumps), until the mid-exponential growth phase
(Fig. 3). The results highlighted that the efflux pump was
clearly induced when the cells were exposed to EB and
TET in the wild-type strain, with an increase in fold
expression of 2.1 � 0.4 and 3.6 � 0.5, respectively. No
tetA induction was observed in the presence of CH. As
expected, tetA was over-expressed in MIM27 under all
tested conditions (Fig. 4) due to the activity of the T5
promoter. These results, along with the previous results
of PM, suggest that TetA could be associated with the
resistance to several antimicrobial compounds or, at least,
that it is an efflux pump with a broad range of substrates.
Over-expression of tetA-like gene positively
affects the growth fitness of S. thermophilus
On the basis of the above findings, we determined the
minimum inhibitory concentrations (MICs) of several
antimicrobial compounds, including TET, CH, and EB.
Because efflux pumps could involve either an ion gradient
(commonly H+) or ATP cleavage to provide the required
energy (Poole, 2005), the MIC evaluation was performed
in M17 broth in the presence of 5 and 20 g L�1 of lactose
(that is low and high availabilities of carbon source,
respectively). The obtained results (Table 2) highlighted a
strong correlation between the carbon source concentra-
tion and the sensitivity to the antimicrobial compound,
underlining the role of the carbon source abundance and
H+ or ATP availability in determining the sensitivity
against antimicrobials. Under all tested conditions,
MIM27 showed reduced sensitivity to the cytotoxic com-
pounds compared to MIM20. Specifically, the most sig-
nificant differences between the two strains were in the
presence of TET and EB. The MIC values of TET in
MIM27 were 0.25 and 1 lg mL�1 in the presence of 5
and 20 g L�1 of lactose, respectively; MIM20 displayed
lower MICs, 0.125 and 0.25 lg mL�1, at the lowest and
highest lactose concentrations, respectively. Analogous
results were obtained in the presence of EB (Table 2).
Conversely, the CH MICs were unaffected or only weakly
affected by the over-expression of tetA, and it was there-
fore concluded that CH is not a preferred substrate of
this efflux pump.
To investigate whether the over-expression of tetA
could influence bacterial growth fitness, both MIM20 and
MIM27 were grown in M17 medium, and the growth
Table 2. Effect of tetA over-expression and lactose concentration on S. thermophilus sensitivity to several antimicrobial compounds
Strain
MIC (lg mL�1) for TET MIC (lg mL�1) for CH MIC (lg mL�1) for EB
5 g L�1 lactose 20 g L�1 lactose 5 g L�1 lactose 20 g L�1 lactose 5 g L�1 lactose 20 g L�1 lactose
MIM20 0.125 0.5 0.25 0.5 0.25 1
MIM27 0.25 1 0.5 0.5 0.5 2
MIM21 0.125 0.5 0.125 0.25 0.125 0.5
MIM28 0.5 2 0.125 0.25 1 2
CH, chlorhexidine; EB, ethidium bromide; MIC, minimum inhibitory concentration; TET, tetracycline.
0Control CH EB TET
1
2
3
4
5
6
7
Nor
mal
ized
fold
exp
ress
ionFig. 3. Transcriptional analysis of tetA in wild-
type S. thermophilus DSM 20617T (white bars)
and a recombinant strain over-expressing tetA,
MIM27 (gray bars). Cells cultured in M17
broth (control condition) and in the presence
of CH, EB, or TET were collected at an OD600
of 0.5 and subjected to RNA extraction and
then qRT-PCR analysis. The expression level of
tetA was normalized using murE and ureC as
reference genes. The data were recorded as
the threshold cycle (CT) expressed as the
means � standard deviation, computed using
Bio-Rad CFX MANAGERTM software, and
expressed as the normalized expression
(CT) � standard error of the mean.
FEMS Microbiol Lett 355 (2014) 61–70 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
TetA MFS efflux pump in Streptococcus thermophilus 67

kinetic parameters of MIM20 and the recombinant
mutant MIM27 were compared in the absence and pres-
ence of EB (0.5 lg mL�1), CH (0.25 lg mL�1), or TET
(0.125 lg mL�1), supplemented at subinhibiting concen-
trations. The presence of the antimicrobial compounds
resulted in an increased lag phase duration in MIM20
compared to MIM27. MIM20 showed a delay of 2.2, 7.6,
and 1.59 h in the presence of TET, CH, and EB, respec-
tively. The more extensive lag time duration and the
absence of tetA gene induction (Fig. 3) in the presence of
CH confirmed that TetA has not the same efficacy to
counteract the effect of CH on cell growth as showed for
TET or EB.
Effect of tetA over-expression in a tetA-
negative strain of S. thermophilus
As the tetA-like gene is present in only 34% of S. thermo-
philus strains screened to date, we attempted to over-
express tetA in the S. thermophilus MIMST24 strain,
which is classified as tetA negative. MIMSt24 electrocom-
petent cells were transformed with pMI-T5 or pMIT5-
tetA to obtain strains MIM21 (tetA negative) and MIM28
(over-expressing tetA). We then analyzed the MIC values
of both mutant strains to evaluate the effect of the pres-
ence of the efflux pump TetA. Analogously to the above
results for MIM20 and MIM27, the MICs values of
MIM21 and MIM28 were strongly affected by the lactose
concentration (Table 2). As observed for DSM 20617, the
over-expression of tetA did not appear to affect CH sensi-
tivity in the MIMST24 genetic background.
Conclusion
This study presents the first characterization of an
efflux pump, TetA, in the dairy species S. thermophilus.
The results obtained underline the activity of this efflux
system in the detoxification of the bacteria from several
cytotoxic compounds, including TET and EB. In a
more general view, efflux pumps can accommodate a
variety of substrates, including nutrients, metabolites,
and toxic compounds that bacteria encounter in their
surrounding environment (Martinez et al., 2009; Oliv-
ares et al., 2012). Moreover, the diffusion of this gene
within this species and its potential for transfer to
other species via HGT remains to be clarified to estab-
lish the safety of tetA-like gene harboring strains
according to EFSA recommendations (EFSA, 2012).
Streptococcus thermophilus is perceived as food-adapted
microorganisms that evolved from a commensal ances-
tor through loss-of-function events counterbalanced by
the acquisition of traits essential for the adaptation to
a milk environment. Therefore, the impact of genes
exchange by lateral gene transfer events occurring
among milk-adapted bacterial species should be relevant
on both the physiology and the safety of this dairy
species.
Acknowledgements
This work was supported by European Community FP7
project KBBE-227258 (BIOHYPO), which is a research
project aimed at evaluating the impact of biocide use on
the generation of antibiotic resistance. The funders had
no role in the study design, data collection, and analysis,
decision to publish, or preparation of the manuscript.
Stefania Arioli was supported by a postdoc grant ‘Dote
Ricerca’: FSE, Regione Lombardia. The authors thank
Sacco-Clerici srl for providing the S. thermophilus strains.
The authors have no conflict of interest to declare. SA
thanks Alessandro Mora for his valuable advice.
References
Arioli S, Monnet C, Guglielmetti S, Parini C, De Noni I,
Hogenboom J, Halami PM & Mora D (2007) Aspartate
biosynthesis is essential for the growth of Streptococcus
thermophilus in milk, and aspartate availability modulates
the level of urease activity. Appl Environ Microbiol 73:
5789–5796.Arioli S, Ragg E, Scaglioni L et al. (2010) Alkalizing reactions
streamline cellular metabolism in acidogenic
microorganisms. PLoS ONE 5: e1520.
Arioli S, Elli M, Ricci G & Mora D (2013) Assessment of the
susceptibility of lactic acid bacteria to biocides. Int J Food
Microbiol 163: 1–5.Aziz RK, Bartels D, Best AA et al. (2008) The RAST Server:
rapid annotations using subsystems technology. BMC
Genomics 9: 75.
Bannam TL & Rood JI (1999) Identification of structural
and functional domains of the tetracycline efflux protein
TetA(P) from Clostrisium perfringens. Microbiology 145:
2947–2955.Bochner BR, Gadzinski P & Panomitros E (2001) Phenotype
microarrays for high-throughput phenotypic testing and
assay of gene function. Genome Res 11: 1246–1255.Bolotin A, Quinquis B, Renault P et al. (2004) Complete
sequence and comparative genome analysis of the dairy
bacterium Streptococcus thermophilus. Nat Biotechnol 22:
1554–1558.Czechowska K & van der Meer JR (2012) Reversible and
irreversible pollutant-induced bacterial cellular stress effects
measured by ethidium bromide uptake and efflux. Environ
Sci Technol 46: 1201–1208.de Ruyter PG, Kuipers OP & de Vos W (1996) Controlled
gene expression systems for Lactococcus lactis with the food-
grade inducer nisin. Appl Environ Microbiol 62:
3662–3667.
FEMS Microbiol Lett 355 (2014) 61–70ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
68 S. Arioli et al.

Decorosi F, Santopolo L, Mora D, Viti C & Giovannetti L
(2011) The improvement of a phenotype microarray
protocol for the chemical sensitivity analysis of Streptococcus
thermophilus. J Microbiol Methods 86: 258–261.EFSA (2012) Guidance of the assessment of bacterial
susceptibility to antimicrobials of human and veterinary
importance. EFSA J 10: 1–48.Floyd JL, Smith KP, Kumar SH, Floyd JT & Varela MF (2010)
Lmrs is a multidrug efflux pump of the major facilitator
superfamily from Staphylococcus aureus. Antimicrob Agents
Chemother 54: 5406–5412.Frempong-Manso E, Raygada JL, DeMarco CE, Seo SM & Kaatz
GW (2009) Inability of a reserpine-based screen to identify
strains over-expressing efflux pump genes in clinical isolates of
Staphylococcus aureus. Int J Antimicrob Agents 33: 360–363.Guglielmetti S, Ciranna A, Mora D, Parini C & Karp M (2008)
Construction, characterization and exemplificative
application of bioluminescent Bifidobacterium longum biovar
longum. Int J Food Microbiol 124: 285–290.Hols P, Hancy F, Fontaine L et al. (2005) New insights in the
molecular biology and physiology of Streptococcus
thermophilus revealed by comparative genomics. FEMS
Microbiol Rev 29: 435–463.Kazimierczak KA, Rincon MT, Patterson AJ, Martin JC, Young
P, Flint HJ & Scott KP (2008) A new tetracycline efflux
gene, tet(40), is located in tandem with tet(O/32/O) in a
human gut firmicute bacterium and in metagenomic library
clones. Antimicrob Agents Chemother 52: 4001–4009.Li XZ & Nikaido H (2004) Efflux-mediated drug resistance in
bacteria. Drugs 64: 159–204.Littlejohn TG, Paulsen IT, Gillespie MT, Tennent JM, Midgley
M, Jones IG, Purewal AS & Skurray RA (1992) Substrate
specificity and energetics of antiseptic and disinfectant
resistance in Staphylococcus aureus. FEMS Microbiol Lett 74:
259–265.Loimaranta V, Tenuovo J, Koivisto L & Karp M (1998)
Generation of bioluminescent Streptococcus mutans and its
usage in rapid analysis of the efficacy of antimicrobial
compounds. Antimicrob Agents Chemother 42: 1906–1910.Lorca GL, Barabote RD, Zlotopolski V et al. (2007) Transport
capabilities of eleven gram-positive bacteria: comparative
genomic analyses. Biochim Biophys Acta 1768: 1342–1366.Lubelski J, Kinigs WN & Driessen AJ (2007) Distribution of
ABC-type transporters contributing to multidrug resistance
in bacteria. Microbiol Mol Biol Rev 71: 463–476.Martinez JL, Sabchez MB, Martinez-Solano L, Hernandez A,
Garmendia L, Fajardo A & Alvarez-Ortega C (2009) Functional
role of bacterial multidrug efflux pumps in microbial natural
eco system. FEMS Microbiol Rev 33: 430–449.Mitchell BA, Brown MH & Skurray RA (1998) QacA
multidrug efflux pump from Staphylococcus aureus:
comparative analysis of resistance to diamidines,
biguanidines, and guanylhydrazones. Antimicrob Agents
Chemother 42: 475–477.Olivares J, Alvarez-Ortega C, Linares J, Rojo F, Kohler T &
Martinez J (2012) Overproduction of the multidrug efflux
pump MexEF-OprN does not impair Pseudomonas
aeruginosa fitness in competition tests, but produces specific
changes in bacterial regulatory networks. Environ Microbiol
14: 1968–1981.Overbeeck R, Begley T, Butler RM et al. (2005) The
subsystems approach to genome annotation and its use in
the project to annotate 1000 genomes. Nucleic Acids Res 33:
5691–5702.Paix~ao L, Rodrigues L, Couto I, Martins M, Fernandes P, de
Carvalho C, Monteiro G, Sansonetty F, Amaral L & Viveiros
M (2009) Fluorometric determination of ethidium bromide
efflux kinetics in Escherichia coli. J Biol Eng 3: 18.
Pao S, Paulsen I & Saier M (1998) Major facilitator
superfamily. Microbiol Mol Biol Rev 62: 1–34.Papadimitriou K, Ferreira S, Papandreou NC, Mavrogonatou
E, Supply P, Pot B & Tsakalidou E (2012) Complete
genome sequence of the dairy isolate S. macedonicus ACA-
DC 198. J Bacteriol 194: 1838.
Paulsen IT, Brown MH & Skurray RA (1996) Proton-
dependent multidrug efflux system. Microbiol Rev 60: 575–608.
Poole K (2005) Efflux-mediated antimicrobial resistance. J
Antimicrob Chemother 56: 20–51.Poole K (2007) Efflux pumps as antimicrobial resistance
mechanisms. Ann Med 39: 162–176.Putman M, Van Veen HW & Konings WN (2000) Molecular
properties of bacterial multidrug transporters. Microbiol Mol
Biol Rev 64: 672–693.Reddy VS, Dhlykov MA, Castillo R, Sun EI & Saier MH Jr
(2012) The major facilitator superfamily (MFS) revisited.
FEBS J 279: 2022–2035.Ren Q, Chen K & Paulsen IT (2007) TransportDB: a
comprehensive database resource for cytoplasmic membrane
transport systems and outer membrane channels. Nucleic
Acids Res 35: D274–D279.Saier MH Jr, Beatty JT, Goffeau A, Harley KT & Heijne WH
(1999) The major facilitator superfamily. J Mol Microbiol
Biotechnol 1: 257–279.Schindler BD, Patel D, Seo S & Kaatz GW (2013) Mutagenesis
and modeling to predict structural and functional
characteristics of the Staphylococcus aureus MepA multidrug
efflux pump. J Bacteriol 195: 523–533.Simm R, Voros A, Ekman JV et al. (2012) BC4707 is a major
facilitator superfamily multidrug resistance transport protein
from Bacillus cereus implicated in fluoroquinolone tolerance.
PLoS ONE 7: e36720.
van Veen HW & Konings WN (1997) Multidrug transporters
from bacteria to man: similarities in structure and function.
Semin Cancer Biol 8: 183–191.Veening JW, Smits WK & Kuipers OP (2008) Bistability,
epigenetics, and bet-hedging in bacteria. Annu Rev Microbiol
62: 193–210.Wang V & van Veen HW (2012) Basic residues R260 and
K357 affect the conformational dynamics of the major
facilitator superfamily multidrug transporter LmrP. PLoS
ONE 7: e38715.
FEMS Microbiol Lett 355 (2014) 61–70 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
TetA MFS efflux pump in Streptococcus thermophilus 69

Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Fig. S1. Ethidium bromide (EB) efflux as detected by
flow cytometry. The density diagrams show the green
(SYBR Green I) and red (EB) fluorescence levels of
MIM20 and MIM27 after 30 min of EB efflux, with and
without the protonophore carbonyl cyanide m-chloro-
phenylhydrazone (CCCP) after EB loading.
Fig. S2. Ethidium bromide (EB) efflux as detected by
flow cytometry. Density diagrams showing the green
(SYBR Green I) and red (EB) fluorescence levels of the
different populations (1 and 2) of wild-type DSM 20617T.
Table S1. LiMIM of S.thermophilus MIMrains of differ-
ent origin screened for the presence of tetA-like gene.
FEMS Microbiol Lett 355 (2014) 61–70ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
70 S. Arioli et al.
Copyright © 2022 FDOKUMEN




















