
Characterization of lactic acid bacteria strains on the basis of neutral volatile compounds produced...
15
Characterization of lactic acid bacteria strains on the basis of neutral volatile compounds produced in whey G. Mauriello, L. Moio, G. Moschetti, P. Piombino, F. Addeo and S. Coppola Dipartimento di Scienza degli Alimenti, Universita` degli Studi di Napoli ‘Federico II’, Naples, Italy 617/11/00: received 1 November 2000, revised 2 February 2001 and accepted 7 February 2001 1 G. MAURIELLO, L. MOIO, G. MOSCHETTI, P. PIOMBINO, F. ADDEO AND S. COPPOLA. 2001. Aims: Seventy-eight strains of lactic acid bacteria belonging to five genera and showing six different phenotype combinations of Lac (lactose fermentation), Prt (proteolytic activity) and Cit (citrate degradation) characters were investigated for their main flavouring properties with the aim to detect variability among and within the groups. Methods and Results: High resolution gas chromatography–mass spectrometry analysis of neutral volatile compounds produced in whey showed that, considering both neo-formation compounds and substances quantified in the whey cultures at different concentrations in comparison to the extract from sterile whey, the groups of lactococci, enterococci, thermophilic streptococci and mesophilic lactobacilli produced a higher number of volatiles than thermophilic lactobacilli and leuconostocs. Applying principal component analysis (PCA) to the results, enterococci, mesophilic lactobacilli and thermophilic streptococci showed a broad diversity, while lactococci included rather similar strains as well as strains with special flavouring properties. Applying PCA to thermophilic streptococci and enterococci, to lactococci and enterococci, to lactococci and thermophilic streptococci, or to mesophilic and thermophilic lactobacilli, the strains gathered consistently with their systematic position. Conclusions: The study evidenced strains producing some volatile compounds responsible for food flavouring. Flavouring properties were variable among the systematic groups and in some cases different within the same bacterial group. Significance and Impact of the Study: The potential of the findings is discussed with reference to the development of flavouring adjuncts for the dairy industry. INTRODUCTION Lactic acid bacteria (LAB) differ considerably in morpho- logical, physiological and functional properties (Wijtzes et al. 1997). While lactic acid is the major end-product of their metabolism, diacetyl, acetoin, 2,3-butanediol, acetate, ethanol, formate, CO 2 and many others are also produced (Cogan 1995). These substances, important in flavour perception and texture of many fermented foods, are produced in different ratios according to species and/or strain, with the consequence that the composition of the LAB community can greatly affect product quality. Biotyping of LAB isolates, within studies on microbial biodiversity occurring in fermented foods, is today widely performed by molecular as well as biochemical proce- dures, which generally give poor information about flavouring abilities and cannot therefore be considered as exhaustive in defining the technological properties of a strain. Flavour molecules can be produced during growth by biosynthesis (de novo synthesis) and bioconversion (precur- sor biotransformation). Most volatile flavour compounds are produced during the stationary growth phase as secondary metabolites (terpenes, esters, ketones, lactones, alcohols and aldehydes) (Belin et al. 1992). In this paper, 78 LAB strains were studied to focus upon significant differences among volatile compounds produced in standard culture conditions. The possibility of using Correspondence to: Salvatore Coppola, Dipartimento di Scienza degli Alimenti, Sezione di Microbiologia, Universita`di Napoli ¢Federico II’, 80055 Portici, Italy (e-mail: [email protected]). ª 2001 The Society for Applied Microbiology Journal of Applied Microbiology 2001, 90, 928–942
Transcript of Characterization of lactic acid bacteria strains on the basis of neutral volatile compounds produced...

Characterization of lactic acid bacteria strains on the basisof neutral volatile compounds produced in whey
G. Mauriello, L. Moio, G. Moschetti, P. Piombino, F. Addeo and S. CoppolaDipartimento di Scienza degli Alimenti, UniversitaÁ degli Studi di Napoli `Federico II', Naples, Italy
617/11/00: received 1 November 2000, revised 2 February 2001 and accepted 7 February 200111
G. MAURIELLO, L . MOIO, G. MOSCHETTI , P . P IOMBINO, F . ADDEO AND S. COPPOLA. 2001.
Aims: Seventy-eight strains of lactic acid bacteria belonging to ®ve genera and showing six
different phenotype combinations of Lac (lactose fermentation), Prt (proteolytic activity) and
Cit (citrate degradation) characters were investigated for their main ¯avouring properties with
the aim to detect variability among and within the groups.
Methods and Results: High resolution gas chromatography±mass spectrometry analysis of
neutral volatile compounds produced in whey showed that, considering both neo-formation
compounds and substances quanti®ed in the whey cultures at different concentrations in
comparison to the extract from sterile whey, the groups of lactococci, enterococci,
thermophilic streptococci and mesophilic lactobacilli produced a higher number of volatiles
than thermophilic lactobacilli and leuconostocs. Applying principal component analysis
(PCA) to the results, enterococci, mesophilic lactobacilli and thermophilic streptococci
showed a broad diversity, while lactococci included rather similar strains as well as strains
with special ¯avouring properties. Applying PCA to thermophilic streptococci and
enterococci, to lactococci and enterococci, to lactococci and thermophilic streptococci, or to
mesophilic and thermophilic lactobacilli, the strains gathered consistently with their
systematic position.
Conclusions: The study evidenced strains producing some volatile compounds responsible
for food ¯avouring. Flavouring properties were variable among the systematic groups and in
some cases different within the same bacterial group.
Signi®cance and Impact of the Study: The potential of the ®ndings is discussed with
reference to the development of ¯avouring adjuncts for the dairy industry.
INTRODUCTION
Lactic acid bacteria (LAB) differ considerably in morpho-
logical, physiological and functional properties (Wijtzes
et al. 1997). While lactic acid is the major end-product of
their metabolism, diacetyl, acetoin, 2,3-butanediol, acetate,
ethanol, formate, CO2 and many others are also produced
(Cogan 1995). These substances, important in ¯avour
perception and texture of many fermented foods, are
produced in different ratios according to species and/or
strain, with the consequence that the composition of the
LAB community can greatly affect product quality.
Biotyping of LAB isolates, within studies on microbial
biodiversity occurring in fermented foods, is today widely
performed by molecular as well as biochemical proce-
dures, which generally give poor information about
¯avouring abilities and cannot therefore be considered as
exhaustive in de®ning the technological properties of a
strain.
Flavour molecules can be produced during growth by
biosynthesis (de novo synthesis) and bioconversion (precur-
sor biotransformation). Most volatile ¯avour compounds are
produced during the stationary growth phase as secondary
metabolites (terpenes, esters, ketones, lactones, alcohols and
aldehydes) (Belin et al. 1992).
In this paper, 78 LAB strains were studied to focus upon
signi®cant differences among volatile compounds produced
in standard culture conditions. The possibility of using
Correspondence to: Salvatore Coppola, Dipartimento di Scienza degli Alimenti,
Sezione di Microbiologia, UniversitaÁ di Napoli ¢Federico II', 80055 Portici,
Italy (e-mail: [email protected]).
ã 2001 The Society for Applied Microbiology
Journal of Applied Microbiology 2001, 90, 928±942

¯avouring properties as a marker trait of a single strain or
LAB groups was also investigated.
MATERIALS AND METHODS
Strains and culture conditions
Seventy-eight LAB strains were used in this study
(Table 1). Twenty-four lactococci, 14 thermophilic lactoba-
cilli, 10 enterococci, two leuconostocs and two thermophilic
streptococci were from natural whey cultures (NWC) used
as starter for the traditional manufacture of water buffalo
Mozzarella cheese (Coppola et al. 1988); eight mesophilic
lactobacilli and four leuconostocs were from pizza doughs
(PD) (Coppola et al. 1996); four thermophilic streptococci
were from pasteurized milk (PM). Lactococci, enterococci
and thermophilic streptococci were identi®ed by physio-
logical and biochemical techniques (Schleifer et al. 1991;
Teuber et al. 1991; Devriese and Pot 1995) and by
ampli®cation of the 16S±23S rDNA spacer region (Mosch-
etti et al. 1998). They were differentiated at strain level by
DNA ®ngerprinting (Moschetti et al. 1993) and randomly
ampli®ed polymorphic DNA (Moschetti et al. 1998). Lac-
tobacilli and leuconostocs were identi®ed according to
Hammes et al. (199122 ) and Villani et al. (1997), respectively.
Five type strains of leuconostocs and the lactococcal strains
DSM4644 (lactose/citrate plasmid carrier) and DSM4367
(protease plasmid carrier) were from DSM (Deutsche
Table 1 Strains and relative phenotype with respect to Lac, Prt and Cit characters
Phenotype
Microbial strains Lac Prt Cit
Lactococcus lactis subsp. lactis NWC:60, 109, 119, 125, 148, 167, 169, 170, 174, 175, 176, 186, DSM4644 + + +
Lactococcus lactis subsp. lactis BU2-60 ± ± ±
Lactobacillus plantarum PDM41
Lactobacillus brevis PDM207
Lactobacillus delbrueckii subsp. lactis NWC:400, 408, 413
Lactobacillus delbrueckii subsp. delbrueckii NWC475
Lactobacillus helveticus NWC:18, 412,
Lactobacillus acidophilus NWC417
Lactococcus lactis subsp. lactis NWC:0148, 120, 128, 146, 147, 154, 155, 156, 163 + ± +
Lactobacillus plantarum PD:E5, E7
Lactobacillus paracasei PD:T228, G11
Leuconostoc mesenteroides subsp. mesenteroides DSM20343, NWC543
Leuconostoc mesenteroides subsp. dextranicum DSM20484
Leuconostoc citreum DSM20188, PDA29
Leuconostoc carnosum PDLA17
Leuconostoc lactis DSM20202
Enterococcus faecalis NWC:95, 214, 222, 224
Lactococcus lactis subsp. lactis NWC:61, 126, 177 + ± ±
Enterococcus faecalis NWC:70, 76, 93, 94, 211
Lactobacillus paracasei PDE1
Lactobacillus plantarum PDT34
Lactobacillus delbrueckii subsp. delbrueckii NWC:425, 482
Lactobacillus helveticus NWC485,
Lactobacillus acidophilus NWC:447, 470
Lactobacillus delbrueckii subsp. lactis NWC:83, 455
Leuconostoc mesenteroides subsp. mesenteroides NWC532,
Leuconostoc mesenteroides subsp. dextranicun PDA7
Streptococcus thermophilus77 : PM:L1, LP25, NWC286, NCIMBNCDO822
Lactococcus lactis subsp. cremoris DSM4367 + + ±
Enterococcus faecalis NWC92
Streptococcus thermophilus88 PM:LP30, LP45, CNRZ302, NWC8C2
Leuconostoc mesenteroides subsp. cremoris DSM20200
Leuconostoc citreum PDA21 ± ± +
NWC: natural whey culture; PD: pizza dough; PM: pasteurized milk; DSM: Deutsche Sammlung von Mikroorganismen; NCIMB: National
Collection of Industrial and Marine Bacteria; CNRZ: Centre National de Recherches Zootechniques.
NEUTRAL VOLATILE COMPOUNDS PRODUCED BY LACTIC ACID BACTERIA 929
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
930 G. MAURIELLO ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942

Sammlung von Mikroorganismen und Zellkulturen GmbH,
Braunschweig, Germany). Moreover, free-plasmid Lacto-coccus lactis subsp. lactis BU2-60 was kindly provided from
Neve et al. (1984), Streptococcus thermophilus NCDO822
and CNRZ302 were from NCIMB (National Collection of
Industrial and Marine Bacteria, Aberdeen, UK) and CNRZ
(Centre National de Recherches Zootechniques, Jouyen-
Josas, France), respectively. Lactobacilli and leuconostocs
were routinely grown in MRS broth (Oxoid Ltd, London,
England); enterococci, streptococci and lactococci in M17
broth (Oxoid).
Whey powder (Lactoserum doux, Unilait International,
Paris, France) was reconstituted with deionized water up to
4% solid content and autoclaved for 5 min at 121°C.
Lac, Prt and Cit phenotype determination
All the strains were tested on Lactic Agar (Elliker et al.1956) containing 0á004% bromocresol purple as pH
indicator, on Transparent Milk Citrate Agar (Brown and
Howe 1922) and on Citrate Agar (Kempler and Mckay
1980) for Lac, Prt and Cit phenotype attribution,
respectively.
Table 2 Volatile compounds isolated in
whey culture extracts of different strains of
lactic acid bacteriaPeak no.
Retention
time (min) Compound Odour*
1 2á391 3-hydroxy-2-butanone buttery
2 2á736 2-methyl-1-butanol fused oil, whiskey
3 3á715 3-methyl butanal malt
4 4á722 furfural sweet, woody, almond,
fragrant, baked bread
5 5á566 ethylbenzene
6 8á724 n.i.
7 8á931 3-hydroxy-methyl butanoate fruity
8 10á714 phenol stable
9 11á945 n.i.
10 12á716 3-hydroxy-4,4-dimethylbutyrolactone nut
11 13á001 2-hydroxy benzaldehyde pungent, phenolic, spicy,
almond, burning taste
12 14á785 n.i.
13 14á963 methyl-2-furoate fruity, mushroom
14 15á054 n.i.
15 15á257 n.i.
16 16á001 n.i.
17 16á449 maltol malt, toasted
18 23á756 n.i.
19 24á115 naphtalene camphoraceous
20 24á959 hydrocarbon
21 25á339 hydrocarbon
22 25á946 hydrocarbon
23 27á229 n.i.
24 27á484 n.i.
25 31á228 n.i.
26 34á403 dodecane
27 35á022 2,4-diterbutilphenol
28 35á355 4-ethoxy ethylbenzoate fruity
29 36á246 tetradecane
30 38á601 hexadecane
n.i. not identi®ed.
*Odour described in: Anon. (1998); Arctander (1969).
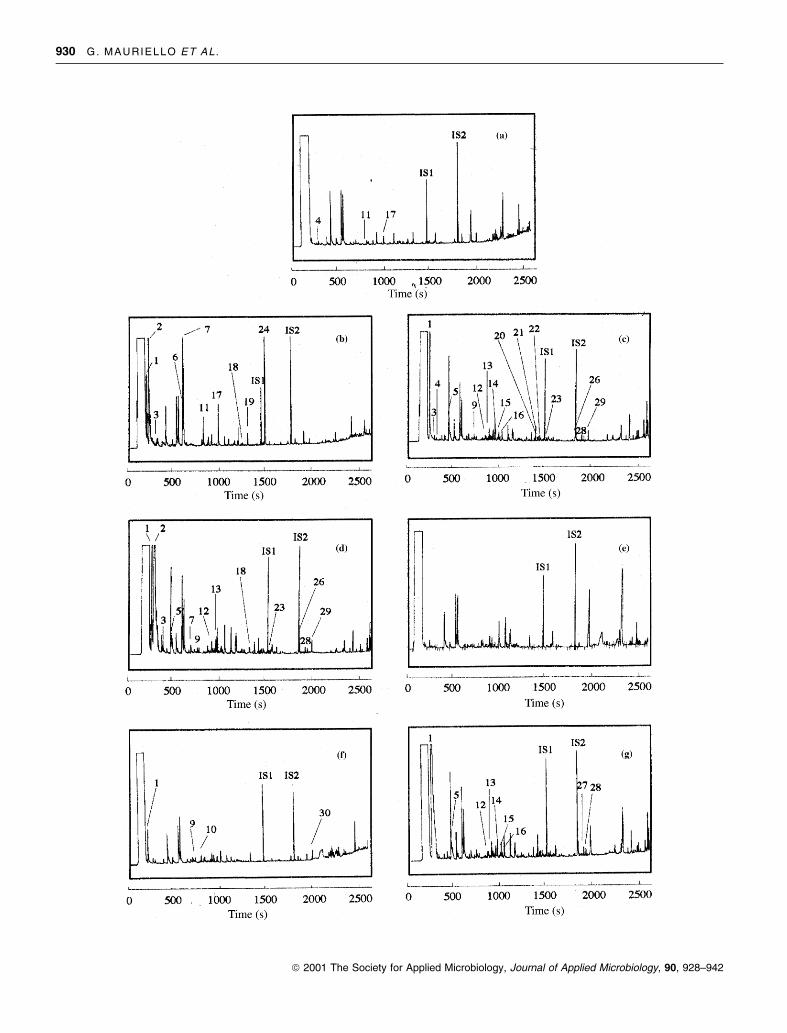
Fig. 1 Whey extracts chromatograms of representative strains of each
microbial group.(a) Control; (b) Lactococcus lactis subsp. lactis 119; (c)
Enterococcus faecalis 211; (d) Streptococcus thermophilus L1; (e) Leuco-
nostoc mesenteroides subsp. mesenteroides 532; (f) Lactobacillus delbrueckii
subsp. delbrueckii 482; (g) Lactobacillus paracasei T228.IS1: methyl
decanoate as ®rst internal standard; IS2: isopropyl decanoate as second
internal standard
b
NEUTRAL VOLATILE COMPOUNDS PRODUCED BY LACTIC ACID BACTERIA 931
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942

Tab
le3
Con
cen
trat
ion
(pp
m)
ofn
eutr
alvo
lati
leco
mp
oun
ds
pro
du
ced
inw
hey
by
lact
ococ
cal
stra
ins
Pea
k
Lac
toco
ccal
stra
ins
n°
6061
109
119
120
125
126
128
146
147
148
154
155
156
10á
030
�0á
009
0á01
2�
0á00
60á
024
�0á
006
0á08
1�
0á04
20á
117
�0á
018
0á59
7�
0á14
30á
038
�0á
005
0á01
6�
0á00
40á2
07�
0á05
10á
032
�0á
007
0á80
9�
0á18
44á
614
�1á
277
4á81
4�
0á39
14á
244
�0á
441
2±
±0á
017
�0á
005
0á06
9�
0á02
30á
009
�0á
002
0á04
9�
0á00
5±
±0á1
07�
0á02
6±
±±
±±
3±
±±
±±
±±
±±
±±
±±
±
50á
005
�0á
0±
0á00
5�
0á0
±±
±0á
014
�0á
002
±±
0á02
1�
0á00
6±
±±
±
6±
±±
0á05
4�
0á01
1±
0á01
3�
0á00
4±
±0á0
51�
0á00
5±
0á00
4�
0á00
1±
±±
70á
005
�0á
0±
0á02
2�
0á00
80á
471
�0á
110
0á05
0�
0á01
00á
122
�0á
024
±±
0á4
41�
0á04
6±
±±
±±
8±
±±
±±
±0á
004
�0á
0±
±0á
006
�0á
0±
±±
±
110á
006
�0á
002
0á00
8�
0á00
20á
039
�0á
014
0á04
6�
0á01
50á
069
�0á
010
0á05
7�
0á01
20á
007
�0á
001
0á01
0�
0á00
20á0
45�
0á00
70á
005
�0á
001
0á00
8�
0á00
3±
0á00
5�
0á0
0á00
6�
0á00
1
170á
013
�0á
003
0á02
3�
0á00
90á
039
�0á
008
0á05
6�
0á02
50á
060
�0á
007
0á06
3�
0á01
30á
027
�0á
010
0á03
2�
0á00
80á0
90�
0á01
30á
040
�0á
010
0á03
1�
0á01
50á
023
�0á
009
0á02
2�
0á00
50á
034
�0á
003
18±
±0á
005
�0á
00á
008
�0á
001
0á01
3�
0á00
30á
005
�0á
001
0á00
9�
0á00
3±
0á0
06�
0á00
10á
017
�0á
003
0á00
7�
0á0
0á00
7�
0á0
0á00
3�
0á00
30á
010
�0á
002
19±
±±
0á05
6�
0á02
2±
0á01
3�
0á00
3±
0á01
0�
0á00
1±
±0á
012
�0á
002
±±
±
240á
004
�0á
0±
0á00
5�
0á0
0á05
2�
0á01
8±
0á01
8�
0á00
2±
±±
±±
±±
±
Pea
k
Lac
toco
ccal
stra
ins
n°
163
167
169
170
174
175
176
177
186
0148
BU
2±60
DS
M43
67D
SM
4644
10á
535
�0á
085
0á06
�0á
002
0á05
8�
0á00
90á
162
�0á
046
0á06
6�
0á03
60á
021
�0á
006
0á03
3�
0á00
60á
186
�0á
068
0á0
6�
0á02
1á59
6�
0á15
60á
009
�0á
00á
048
�0á
0112
á75
�7á
601
2±
±±
±±
±±
±±
±±
±±
3±
±±
±±
0á01
0�
0á0
±±
0á0
06�
0á0
±±
±±
50á
004
�0á
0±
±±
0á00
5�
0á00
1±
±0á
013
�0á
005
±±
0á02
2�
0á00
20á
006
�0á
001
±
6±
±±
±±
0á00
5�
0á0
±±
±0á
005
�0á
001
±±
±
7±
±±
±±
±±
±±
±±
±±
80á
004
�0á
0±
±±
0á00
5�
0á00
1±
±0á
004
�0á
0±
±±
±±
110á
006
�0á
001
0á00
9�
0á00
20á
005
�0á
002
0á00
4�
0á00
40á
007
�0á
002
0á01
0�
0á00
40á
010
�0á
006
0á01
2�
0á00
30á0
08�
0á00
20á
012
�0á
003
0á01
1�
0á00
20á
009
�0á
003
0á00
7�
0á00
3
170á
032
�0á
007
0á02
8�
0á01
00á
022
�0á
007
0á02
1�
0á00
50á
028
�0á
008
0á01
8�
0á00
30á
023
�0á
007
0á04
4�
0á01
20á0
18�
0á00
40á
033
�0á
006
0á03
4�
0á00
30á
021
�0á
005
0á01
3�
0á00
8
180á
013
�0á
003
0á00
7�
0á00
10á
001
�0á
00á
006
�0á
00á
009
�0á
003
0á00
6�
0á00
20á
004
�0á
00á
013
�0á
002
0á0
03�
0á0
0á00
8�
0á00
10á
017
�0á
002
0á01
2�
0á00
10á
004
�0á
001
190á
009
�0á
001
±0á
007
�0á
003
0á00
5�
0á00
1±
±±
±±
±±
±±
24±
±±
±±
±±
±±
±±
±±
Val
ues
are
mea
ns
ofth
ree
exp
erim
ents
�st
and
ard
dev
iati
on;
±:
abse
nce
ofp
eak.
932 G. MAURIELLO ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
Tab
le4
Con
cen
trat
ion
(pp
m)
ofn
eutr
alvo
lati
leco
mp
oun
ds
pro
du
ced
inw
hey
by
ente
roco
ccal
stra
ins
Pea
kE
nte
roco
ccal
stra
ins
no.
7076
9293
9495
211
214
222
224
10á
090
�0á
006
0á0
60�
0á0
050á
076
�0á
006
0á11
0�
0á00
20á
197
�0á
035
0á81
8�
0á08
10á
125
�0á
013
0á15
5�
0á01
60á
338
�0á
035
0á60
0�
0á06
6
3±
0á0
07�
0á0
0á00
8�
0á0
0á00
7�
0á0
0á00
7�
0á00
10á
003
�0á
001
0á00
6�
0á0
0á00
5�
0á0
0á00
4�
0á0
0á00
7�
0á0
50á
022
�0á
001
0á0
25�
0á0
020á
021
�0á
002
0á03
1�
0á0
0á04
0�
0á00
70á
019
�0á
002
0á02
8�
0á00
30á
033
�0á
003
0á11
9�
0á01
20á
033
�0á
004
60á
006
�0á
00á0
07�
0á0
0á00
7�
0á0
0á00
7�
0á0
0á00
7�
0á00
10á
007
�0á
00á
010
�0á
001
0á00
7�
0á0
0á00
5�
0á0
0á00
6�
0á0
90á
003
�0á
00á0
06�
0á0
0á00
6�
0á0
0á00
5�
0á0
0á00
4�
0á0
0á00
5�
0á0
0á00
4�
0á0
0á00
6�
0á0
0á00
5�
0á0
0á00
6�
0á0
120á
017
�0á
001
0á0
24�
0á0
020á
024
�0á
002
0á02
3�
0á0
0á02
2�
0á00
40á
023
�0á
002
0á02
2�
0á00
20á
023
�0á
002
0á02
1�
0á00
20á
024
�0á
003
130á
010
�0á
00á0
15�
0á0
010á
014
�0á
001
0á01
3�
0á0
0á01
6�
0á00
30á
013
�0á
001
0á02
1�
0á00
20á
017
�0á
002
0á01
5�
0á00
20á
018
�0á
001
140á
008
�0á
00á0
10�
0á0
0á00
9�
0á0
0á00
9�
0á0
0á00
4�
0á0
0á00
6�
0á0
0á00
7�
0á0
0á00
5�
0á0
0á00
8�
0á0
0á00
5�
0á0
150á
012
�0á
00á0
14�
0á0
010á
014
�0á
001
0á01
4�
0á0
0á01
0�
0á00
10á
014
�0á
001
0á01
2�
0á00
10á
010
�0á
00á
012
�0á
001
0á01
0�
0á00
1
160á
007
�0á
00á0
08�
0á0
0á00
8�
0á0
0á00
9�
0á0
0á00
4�
0á0
0á00
8�
0á0
0á00
7�
0á0
0á00
4�
0á0
0á00
6�
0á0
0á00
4�
0á0
180á
022
�0á
001
0á0
32�
0á0
020á
024
�0á
002
0á03
2�
0á0
0á02
7�
0á00
50á
020
�0á
002
0á02
4�
0á00
20á
033
�0á
003
0á02
7�
0á00
30á
040
�0á
004
200á
021
�0á
001
0á0
24�
0á0
020á
024
�0á
002
0á02
5�
0á0
0á02
2�
0á00
40á
024
�0á
002
0á02
1�
0á00
20á
022
�0á
002
0á02
0�
0á00
20á
021
�0á
002
210á
004
�0á
00á0
06�
0á0
010á
005
�0á
001
0á00
5�
0á0
0á00
4�
0á0
0á00
6�
0á00
30á
004
�0á
00á
005
�0á
00á
005
�0á
00á
005
�0á
002
220á
005
�0á
00á0
08�
0á0
020á
006
�0á
00á
006
�0á
001
0á00
6�
0á0
0á00
7�
0á00
20á
006
�0á
001
0á00
7�
0á00
20á
007
�0á
00á
007
�0á
0
230á
008
�0á
00á0
09�
0á0
0á00
7�
0á0
0á01
0�
0á0
0á01
1�
0á00
20á
007
�0á
00á
008
�0á
001
0á00
9�
0á0
0á00
8�
0á0
0á00
8�
0á0
260á
013
�0á
00á0
15�
0á0
010á
015
�0á
001
0á01
5�
0á0
0á01
5�
0á00
30á
015
�0á
001
0á01
3�
0á00
10á
014
�0á
001
0á01
4�
0á00
10á
014
�0á
001
27±
0á0
03�
0á0
0á00
3�
0á0
0á00
2�
0á0
0á00
2�
0á0
0á00
3�
0á0
±0á
002
�0á
00á
003
�0á
00á
002
�0á
0
280á
010
�0á
00á0
12�
0á0
0á01
1�
0á0
0á01
2�
0á0
0á01
1�
0á00
20á
014
�0á
001
0á01
0�
0á0
0á01
0�
0á00
10á
011
�0á
001
0á01
4�
0á00
2
290á
008
�0á
00á0
09�
0á0
0á00
9�
0á0
0á00
9�
0á0
0á00
9�
0á00
20á
009
�0á
00á
008
�0á
00á
008
�0á
00á
008
�0á
00á
008
�0á
0
30±
±±
0á00
4�
0á0
0á00
8�
0á0
±±
±±
±
Val
ues
are
mea
ns
ofth
ree
exp
erim
ents
�st
and
ard
dev
iati
on;
±:
abse
nce
ofp
eak.
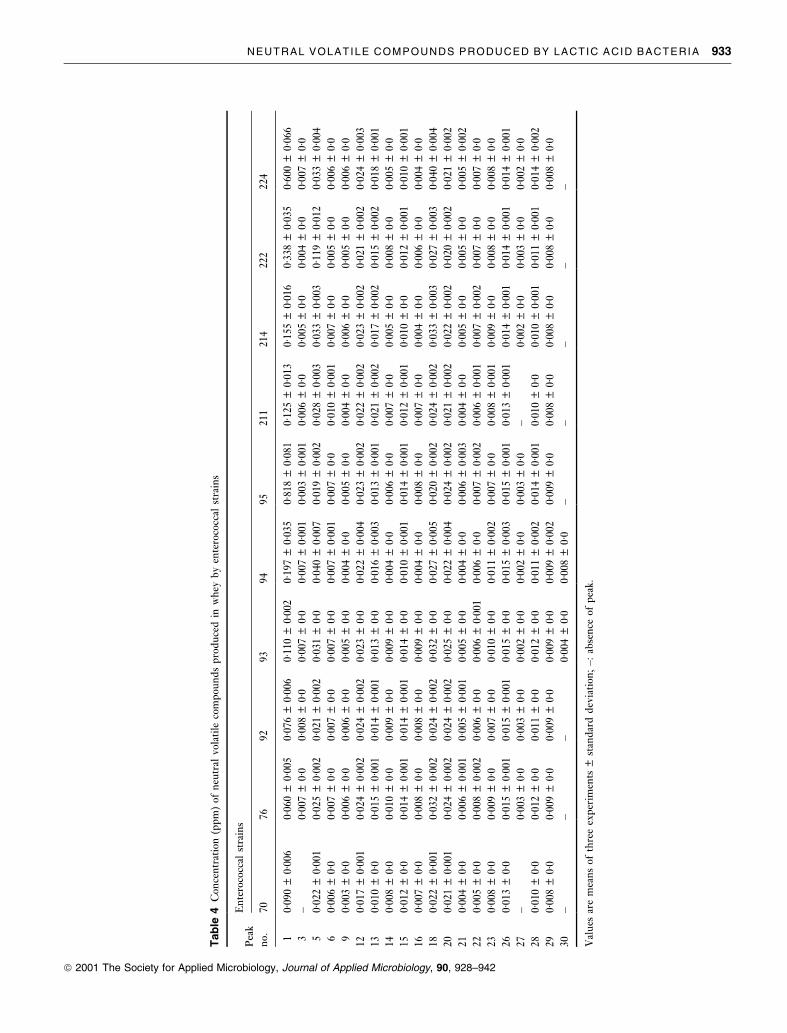
NEUTRAL VOLATILE COMPOUNDS PRODUCED BY LACTIC ACID BACTERIA 933
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
Whey culture preparation
Overnight cultures of each strain were used to prepare 10 ml
of whey subculture; 150 ml of whey were inoculated at 2%
with whey subculture and incubated for 18 h at 30°C and
40°C for mesophilic and thermophilic cultures, respectively.
An uninoculated batch of whey was used as control.
Extraction of neutral volatile compounds
The volatile constituents of whey cultures were isolated as
described by Moio et al. (1993) for milk aqueous distillates.
Whey cultures were centrifuged for 20 min at 9000 rev
min±1 and 100 ml of supernatant ¯uid were maintained
in ice bath and neutralized with 0á2 mol l±1 NaOH until
pH 9á00 to salify free fatty acid. After adding of
0á1032 ppm of methyl decanoate and 0á1044 ppm of
isopropyl decanoate as internal standards, the whey,
containing free neutral volatile compounds and salts of
fatty acids, were extracted three times with freshly distilled
dichloromethane (50 ml) with vigorous agitation (magnet-
ically stirred). The whey/CH2Cl2 emulsion formed during
stirring was separated from the aqueous layer and frozen
at )30°C (Moio and Etievant 1995). The ¯ask was then
allowed to reach room temperature, and the CH2Cl2solution, progressively separated from the remaining whey,
was dried over anhydrous MgSO4 and reduced to 0á5 ml
under a stream of nitrogen.
High resolution gas chromatography (HRGC)
HRGC analysis was carried out with a Hewlett Packard
series II 5890 gas chromatograph, equipped with a split-
splitless injector, a ¯ame ionization detector, and an HP-5
fused silica capillary column (30 m ´ 0á32 mm ID; ®lm
thickness, 1 lm) (Moio et al. 2000). The operating condi-
tions were as follows: the temperature was programmed to
rise from 40°C to 210°C at a rate of 3°C min±1. The injector
and detector temperatures were set at 250°C and 255°C,
respectively. The hydrogen carrier velocity was 37 cm s±1.
Quantitative measurements of each component were ob-
tained from the peak areas of the components relative to the
internal standards, assuming that the extraction ef®ciency
and the GC response were identical for all compounds
separated.
High resolution gas chromatography±massspectrometry (HRGC±MS)
The electron impact mass spectra were obtained with an
HP 5970 quadrupole mass spectrometer coupled with an
HP 5890 gas chromatograph equipped with a DB-5
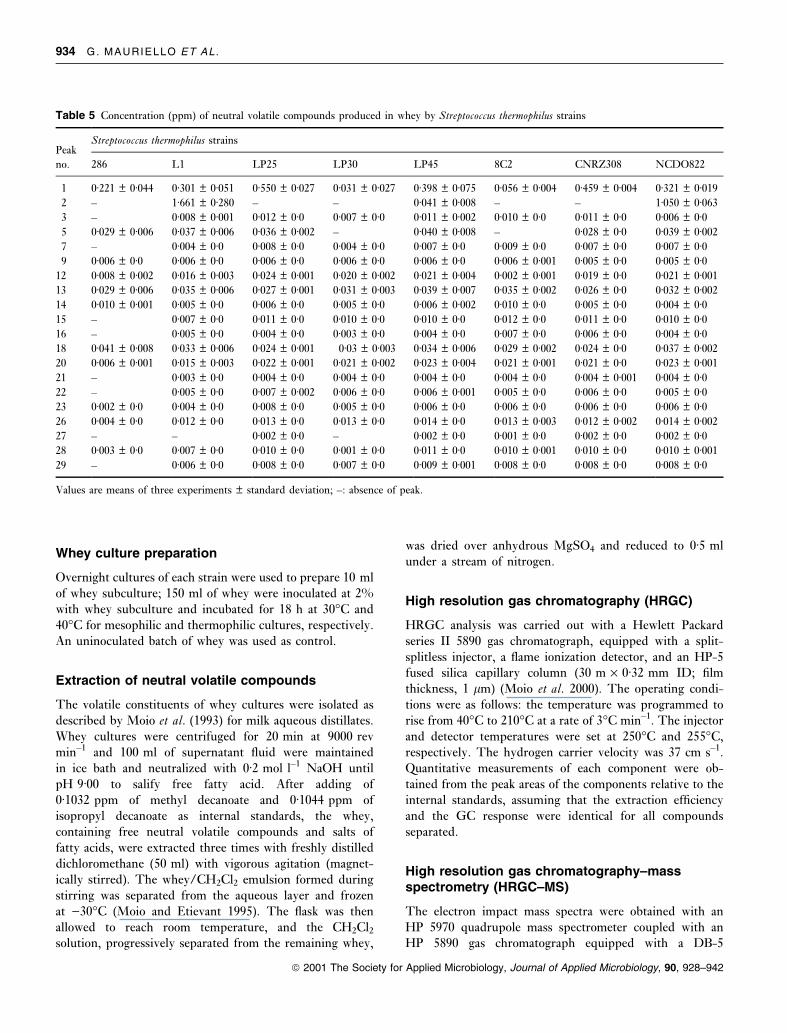
Table 5 Concentration (ppm) of neutral volatile compounds produced in whey by Streptococcus thermophilus strains
PeakStreptococcus thermophilus strains
no. 286 L1 LP25 LP30 LP45 8C2 CNRZ308 NCDO822
1 0á221 � 0á044 0á301 � 0á051 0á550 � 0á027 0á031 � 0á027 0á398 � 0á075 0á056 � 0á004 0á459 � 0á004 0á321 � 0á019
2 ± 1á661 � 0á280 ± ± 0á041 � 0á008 ± ± 1á050 � 0á063
3 ± 0á008 � 0á001 0á012 � 0á0 0á007 � 0á0 0á011 � 0á002 0á010 � 0á0 0á011 � 0á0 0á006 � 0á05 0á029 � 0á006 0á037 � 0á006 0á036 � 0á002 ± 0á040 � 0á008 ± 0á028 � 0á0 0á039 � 0á002
7 ± 0á004 � 0á0 0á008 � 0á0 0á004 � 0á0 0á007 � 0á0 0á009 � 0á0 0á007 � 0á0 0á007 � 0á09 0á006 � 0á0 0á006 � 0á0 0á006 � 0á0 0á006 � 0á0 0á006 � 0á0 0á006 � 0á001 0á005 � 0á0 0á005 � 0á0
12 0á008 � 0á002 0á016 � 0á003 0á024 � 0á001 0á020 � 0á002 0á021 � 0á004 0á002 � 0á001 0á019 � 0á0 0á021 � 0á001
13 0á029 � 0á006 0á035 � 0á006 0á027 � 0á001 0á031 � 0á003 0á039 � 0á007 0á035 � 0á002 0á026 � 0á0 0á032 � 0á002
14 0á010 � 0á001 0á005 � 0á0 0á006 � 0á0 0á005 � 0á0 0á006 � 0á002 0á010 � 0á0 0á005 � 0á0 0á004 � 0á015 ± 0á007 � 0á0 0á011 � 0á0 0á010 � 0á0 0á010 � 0á0 0á012 � 0á0 0á011 � 0á0 0á010 � 0á016 ± 0á005 � 0á0 0á004 � 0á0 0á003 � 0á0 0á004 � 0á0 0á007 � 0á0 0á006 � 0á0 0á004 � 0á018 0á041 � 0á008 0á033 � 0á006 0á024 � 0á001 0á03 � 0á003 0á034 � 0á006 0á029 � 0á002 0á024 � 0á0 0á037 � 0á002
20 0á006 � 0á001 0á015 � 0á003 0á022 � 0á001 0á021 � 0á002 0á023 � 0á004 0á021 � 0á001 0á021 � 0á0 0á023 � 0á001
21 ± 0á003 � 0á0 0á004 � 0á0 0á004 � 0á0 0á004 � 0á0 0á004 � 0á0 0á004 � 0á001 0á004 � 0á022 ± 0á005 � 0á0 0á007 � 0á002 0á006 � 0á0 0á006 � 0á001 0á005 � 0á0 0á006 � 0á0 0á005 � 0á023 0á002 � 0á0 0á004 � 0á0 0á008 � 0á0 0á005 � 0á0 0á006 � 0á0 0á006 � 0á0 0á006 � 0á0 0á006 � 0á026 0á004 � 0á0 0á012 � 0á0 0á013 � 0á0 0á013 � 0á0 0á014 � 0á0 0á013 � 0á003 0á012 � 0á002 0á014 � 0á002
27 ± ± 0á002 � 0á0 ± 0á002 � 0á0 0á001 � 0á0 0á002 � 0á0 0á002 � 0á028 0á003 � 0á0 0á007 � 0á0 0á010 � 0á0 0á001 � 0á0 0á011 � 0á0 0á010 � 0á001 0á010 � 0á0 0á010 � 0á001
29 ± 0á006 � 0á0 0á008 � 0á0 0á007 � 0á0 0á009 � 0á001 0á008 � 0á0 0á008 � 0á0 0á008 � 0á0
Values are means of three experiments � standard deviation; ±: absence of peak.
934 G. MAURIELLO ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942

capillary column (30 m ´ 0á32 mm ID; ®lm thickness,
1 lm) connected directly to the ion source of the mass-
spectrometer according to the operating procedure des-
cribed for HRGC. The operating HRGC conditions were
the same as those described for HRGC. The mass spectra
were determined at 70 eV while the ion source and the
interface were held at 150°C and 280°C, respectively (Moio
et al. 2000). Acquisition and processing of mass spectra
were carried out by means of a computer and the
compounds were identi®ed with the aid of mass spectral
data bases (33 Nover de Brauw et al. 1987).
Statistical analysis
Statistical treatment of data, by cluster analysis (CA) and
principal component analysis (PCA), was performed using
Systat software for Macintosh ver. 5.2.1. PCA was per-
formed using Principal Components procedure of Factor
function on data matrix, with two PCs and without
rotational factor. CA was carried out by the Pearson
Correlation with the Corr procedure and then relationships
were established using Single Linkage Method by the
Cluster procedure.
RESULTS AND DISCUSSION
Lac, Prt and Cit phenotype
Glycolysis, proteolysis and citrate degradation are consid-
ered the principal metabolic pathways of LAB involved in
the formation of aroma and ¯avour compounds in
fermented products (Law 1981). On the basis of lactose
fermentation, degradation of caseins and citrate meta-
bolism, the strains were grouped into six phenotypes
(Table 1). Only lactococcal strains (about half of those
tested) showed the Lac+Prt+Cit+ phenotype. The
Lac±Prt±Cit± phenotype was shown by the plasmid-free
strain BU2-60, as expected, and by nine strains belonging
to species of Lactobacillus. As already reported (Cogan
1995), citrate is not metabolized by thermophilic cultures,
so in this study thermophilic lactobacilli and streptococci
showed the Cit± phenotype.
HRGC±MS analysis
On comparing volatile components detected in whey culture
extracts qualitative and quantitative differences could be
evidenced. All microbial groups produced new compounds
and the gas-chromatogram complexity is higher than the
control. Furfural (peak 4), 2-hydroxy benzaldehyde (peak
11) and maltol (peak 17) were already present in the control
chromatogram as shown in Fig. 1 where results are reported
from representative strains.
Tab
le6
Con
cen
trat
ion
(pp
m)
ofn
eutr
alvo
lati
leco
mp
oun
ds
pro
du
ced
by
Leu
cono
stoc
spp
.st
rain
s
Pea
kL
euco
nost
ocst
rain
s
no.
532
543
A7
A21
A29
LA
17L
N2
LN
3L
N13
LN
14L
N16
10á
002
�0á
00á
002
�0á
00á
007
�0á
002
0á00
3�
0á0
0á00
7�
0á00
20á
059
�0á
016
0á04
4�
0á01
00á
004
�0á
001
0á00
1�
0á0
±0á
009
�0á
002
4±
±±
±±
0á00
9�
0á00
2±
±±
±±
8±
±±
±±
±0á
035
�0á
006
±±
±±
9±
±±
0á00
7�
0á00
20á
004
�0á
0±
±0á
017
�0á
004
0á01
6�
0á0
±±
180á
020
�0á
008
0á02
6�
0á00
80á
026
�0á
009
±±
0á02
3�
0á00
6±
±±
0á01
3�
0á00
30á
029
�0á
012
25±
0á00
6�
0á00
1±
±±
0á00
4�
0á0
0á00
5�
0á0
±±
0á00
5�
0á0
±
Val
ues
are
mea
ns
ofth
ree
exp
erim
ents
�st
and
ard
dev
iati
on;
±:
abse
nce
ofp
eak.
NEUTRAL VOLATILE COMPOUNDS PRODUCED BY LACTIC ACID BACTERIA 935
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
Pea
kT
her
mop
hil
icla
ctob
acil
list
rain
s
no.
470
475
482
485
10á
064
�0á
010
±0á
038
�0á
003
0á13
9�
0á02
9
5±
±±
±
7±
±±
±
90á
007
�0á
00á
004
�0á
00á
007
�0á
00á
007
�0á
0
10±
±0á
013
�0á
02±
150á
004
�0á
00á
005
�0á
00á
014
�0á
001
0á00
6�
0á0
180á
012
�0á
001
0á00
4�
0á0
0á00
6�
0á0
0á00
8�
0á0
300á
020
�0á
002
±0á
017
�0á
010á
008
�0á
0
Val
ues
are
mea
ns
ofth
ree
exp
erim
ents
�st
and
ard
dev
iati
on;
±:
abse
nce
ofp
eak.
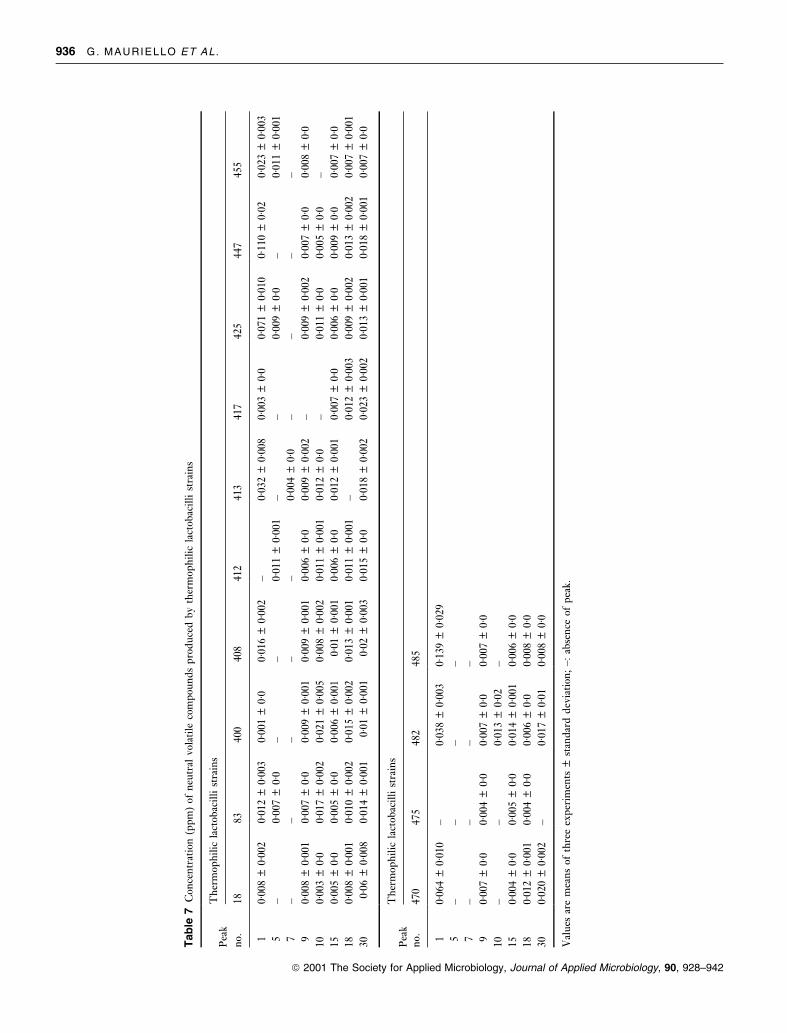
Tab
le7
Con
cen
trat
ion
(pp
m)
ofn
eutr
alvo
lati
leco
mp
oun
ds
pro
du
ced
by
ther
mop
hil
icla
ctob
acil
list
rain
s
Pea
kT
her
mop
hil
icla
ctob
acil
list
rain
s
no.
1883
400
408
412
413
417
425
447
455
10á
008
�0á
002
0á01
2�
0á00
30á
001
�0á
00á
016
�0á
002
±0á
032
�0á
008
0á00
3�
0á0
0á07
1�
0á01
00á
110
�0á
020á
023
�0á
003
5±
0á00
7�
0á0
±±
0á01
1�
0á00
1±
±0á
009
�0á
0±
0á01
1�
0á00
1
7±
±±
±±
0á00
4�
0á0
±±
±±
90á
008
�0á
001
0á00
7�
0á0
0á00
9�
0á00
10á
009
�0á
001
0á00
6�
0á0
0á00
9�
0á00
2±
0á00
9�
0á00
20á
007
�0á
00á
008
�0á
0
100á
003
�0á
00á
017
�0á
002
0á02
1�
0á00
50á
008
�0á
002
0á01
1�
0á00
10á
012
�0á
0±
0á01
1�
0á0
0á00
5�
0á0
±
150á
005
�0á
00á
005
�0á
00á
006
�0á
001
0á01
�0á
001
0á00
6�
0á0
0á01
2�
0á00
10á
007
�0á
00á
006
�0á
00á
009
�0á
00á
007
�0á
0
180á
008
�0á
001
0á01
0�
0á00
20á
015
�0á
002
0á01
3�
0á00
10á
011
�0á
001
±0á
012
�0á
003
0á00
9�
0á00
20á
013
�0á
002
0á00
7�
0á00
1
300á
06�
0á00
80á
014
�0á
001
0á01
�0á
001
0á02
�0á
003
0á01
5�
0á0
0á01
8�
0á00
20á
023
�0á
002
0á01
3�
0á00
10á
018
�0á
001
0á00
7�
0á0
936 G. MAURIELLO ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
The identity and aroma description of all volatile com-
pounds detected are given in Table 2. Tables 3±8 show, for
each microbial group, the concentration of neutral volatile
compounds isolated from each whey culture, undetected in
the control and/or quanti®ed at different concentration
levels in comparison to the control. The quantity of each
component was calculated with respect to the internal
standards, assuming that the extraction ef®ciency and the
GC response were identical for all compounds separated.
Considering all the substances referable to the strains of
each group, lactococci, enterococci, thermophilic strepto-
cocci and mesophilic lactobacilli showed 12, 20, 20 and 20
compounds, respectively. Thermophilic lactobacilli and
leuconostocs produced a small number of new substances
(eight and six volatile compounds, respectively). Only two
compounds, 3-hydroxy-2-butanone (acetoin, peak 1) and
the unidenti®ed peak 18 were found as produced by all
bacterial groups. The ®rst, together with diacetyl, plays an
important role in the aroma of dairy products and its level
is strain-dependent. Only six strains produced more than
1 ppm of 3-hydroxy-2-butanone. A high level (12á75 ppm)
was produced by the strain Lactococcus lactis subsp. lactis4644 according to its plasmid content. The strain Lacto-bacillus paracasei T228 produced 2á74 ppm of 3-hydroxy-
2-butanone, while lactococcal strains 154, 155 and 156
produced 4á61, 4á81 and 4á24 ppm, respectively. Finally,
the leuconostocs produced low concentrations of acetoin
probably due to the low acidity reached in the whey
cultures of these microorganisms. The citrate transport
system is known to be induced by natural acidi®cation of
the medium during growth and by a shift to media
buffered at acidic pHs (Garcia-Quintans et al. 1998).
Therefore low acid-producing microorganisms are respon-
sible for low acetoin production. However, it is well known
that leuconostocs, cocultured with acidifying microrgan-
isms, are able to produce high quantities of acetoin during
their metabolism (Levata-Jovanovic and Sandine 1996a,
1997)44 . Acetoin contributes, together with diacetyl, to the
butter note (Cogan 1995). Due to the procedure carried
out, we were unable to evaluate the diacetyl level since it
was coeluted with dichloromethane used as extraction
solvent.
Naphthalene (peak 19) and the unidenti®ed compound
relative to peak 24 seem to be produced only by lactococci.
Another unidenti®ed compound (peak 25) was found only in
leuconostoc whey cultures.
Thermophilic streptococci and leuconostocs produced
2-methyl-1-butanol (peak 2) and phenol (peak 8), respect-
ively, while lactococcal strains produced both substances.
2-Methyl-1-butanol generally arises from deamination
and subsequent decarboxylation of isoleucine according to
Ehrlich's pathway (Anon. 1995). The isoleucine is probably
already present in the whey since there is no apparent
correlation between 2-methyl-1-butanol production and
Table 8 Concentration (ppm) of neutral volatile compounds produced by mesophilic lactobacilli strains
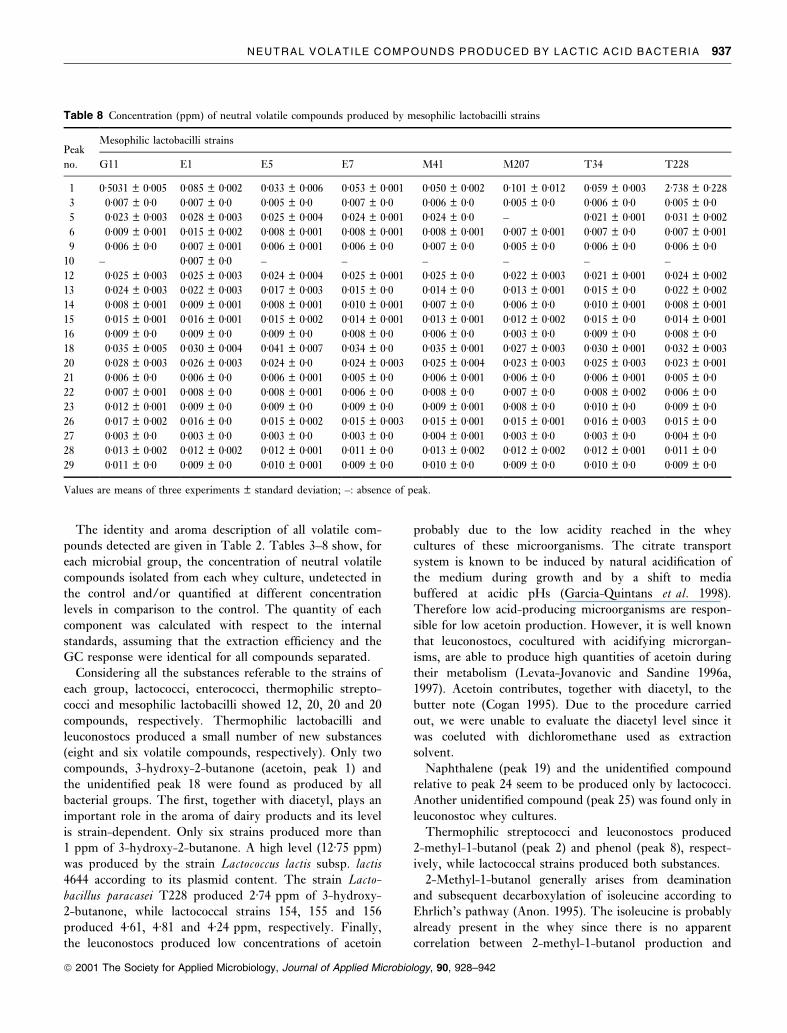
PeakMesophilic lactobacilli strains
no. G11 E1 E5 E7 M41 M207 T34 T228
1 0á5031 � 0á005 0á085 � 0á002 0á033 � 0á006 0á053 � 0á001 0á050 � 0á002 0á101 � 0á012 0á059 � 0á003 2á738 � 0á228
3 0á007 � 0á0 0á007 � 0á0 0á005 � 0á0 0á007 � 0á0 0á006 � 0á0 0á005 � 0á0 0á006 � 0á0 0á005 � 0á05 0á023 � 0á003 0á028 � 0á003 0á025 � 0á004 0á024 � 0á001 0á024 � 0á0 ± 0á021 � 0á001 0á031 � 0á002
6 0á009 � 0á001 0á015 � 0á002 0á008 � 0á001 0á008 � 0á001 0á008 � 0á001 0á007 � 0á001 0á007 � 0á0 0á007 � 0á001
9 0á006 � 0á0 0á007 � 0á001 0á006 � 0á001 0á006 � 0á0 0á007 � 0á0 0á005 � 0á0 0á006 � 0á0 0á006 � 0á010 ± 0á007 � 0á0 ± ± ± ± ± ±
12 0á025 � 0á003 0á025 � 0á003 0á024 � 0á004 0á025 � 0á001 0á025 � 0á0 0á022 � 0á003 0á021 � 0á001 0á024 � 0á002
13 0á024 � 0á003 0á022 � 0á003 0á017 � 0á003 0á015 � 0á0 0á014 � 0á0 0á013 � 0á001 0á015 � 0á0 0á022 � 0á002
14 0á008 � 0á001 0á009 � 0á001 0á008 � 0á001 0á010 � 0á001 0á007 � 0á0 0á006 � 0á0 0á010 � 0á001 0á008 � 0á001
15 0á015 � 0á001 0á016 � 0á001 0á015 � 0á002 0á014 � 0á001 0á013 � 0á001 0á012 � 0á002 0á015 � 0á0 0á014 � 0á001
16 0á009 � 0á0 0á009 � 0á0 0á009 � 0á0 0á008 � 0á0 0á006 � 0á0 0á003 � 0á0 0á009 � 0á0 0á008 � 0á018 0á035 � 0á005 0á030 � 0á004 0á041 � 0á007 0á034 � 0á0 0á035 � 0á001 0á027 � 0á003 0á030 � 0á001 0á032 � 0á003
20 0á028 � 0á003 0á026 � 0á003 0á024 � 0á0 0á024 � 0á003 0á025 � 0á004 0á023 � 0á003 0á025 � 0á003 0á023 � 0á001
21 0á006 � 0á0 0á006 � 0á0 0á006 � 0á001 0á005 � 0á0 0á006 � 0á001 0á006 � 0á0 0á006 � 0á001 0á005 � 0á022 0á007 � 0á001 0á008 � 0á0 0á008 � 0á001 0á006 � 0á0 0á008 � 0á0 0á007 � 0á0 0á008 � 0á002 0á006 � 0á023 0á012 � 0á001 0á009 � 0á0 0á009 � 0á0 0á009 � 0á0 0á009 � 0á001 0á008 � 0á0 0á010 � 0á0 0á009 � 0á026 0á017 � 0á002 0á016 � 0á0 0á015 � 0á002 0á015 � 0á003 0á015 � 0á001 0á015 � 0á001 0á016 � 0á003 0á015 � 0á027 0á003 � 0á0 0á003 � 0á0 0á003 � 0á0 0á003 � 0á0 0á004 � 0á001 0á003 � 0á0 0á003 � 0á0 0á004 � 0á028 0á013 � 0á002 0á012 � 0á002 0á012 � 0á001 0á011 � 0á0 0á013 � 0á002 0á012 � 0á002 0á012 � 0á001 0á011 � 0á029 0á011 � 0á0 0á009 � 0á0 0á010 � 0á001 0á009 � 0á0 0á010 � 0á0 0á009 � 0á0 0á010 � 0á0 0á009 � 0á0
Values are means of three experiments � standard deviation; ±: absence of peak.
NEUTRAL VOLATILE COMPOUNDS PRODUCED BY LACTIC ACID BACTERIA 937
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
Prt+ phenotype. Phenol, often responsible for important off-
¯avours due to its unpleasant stable odour, is produced only
by few lactococcal strains. Only whey cultures of thermo-
philic lactobacilli and of the mesophilic strain Lactobacillusparacasei E1 contain 3-hydroxy-4,4-dimethyl-butyrolactone
(pantolacton), a compound characterized by a typical nut
odour.
3-Hydroxy-methyl butanoate (peak 7), methyl-2-furoate
(peak 13), several hydrocarbons (peaks 20, 21 and 22),
dodecane (peak 26), 4-ethoxy-ethyl benzoate (peak 28),
tetradecane (peak 29) and ®ve unidenti®ed compounds
(peaks 6, 12, 14, 16 and 23) were produced by lactobacilli,
both mesophilic and thermophilic, as well as by enterococci.
Among these, 3-hydroxy-methyl butanoate, methyl-2-furo-
ate and 4-ethoxy-ethyl benzoate are odour-active com-
pounds and responsible for a fruit-like aroma (Arctander
1969).
Lactococci, enterococci, thermophilic streptococci and
mesophilic lactobacilli produced 3-methyl butanale, an
odourant belonging to the aldehyde class. This aldehyde
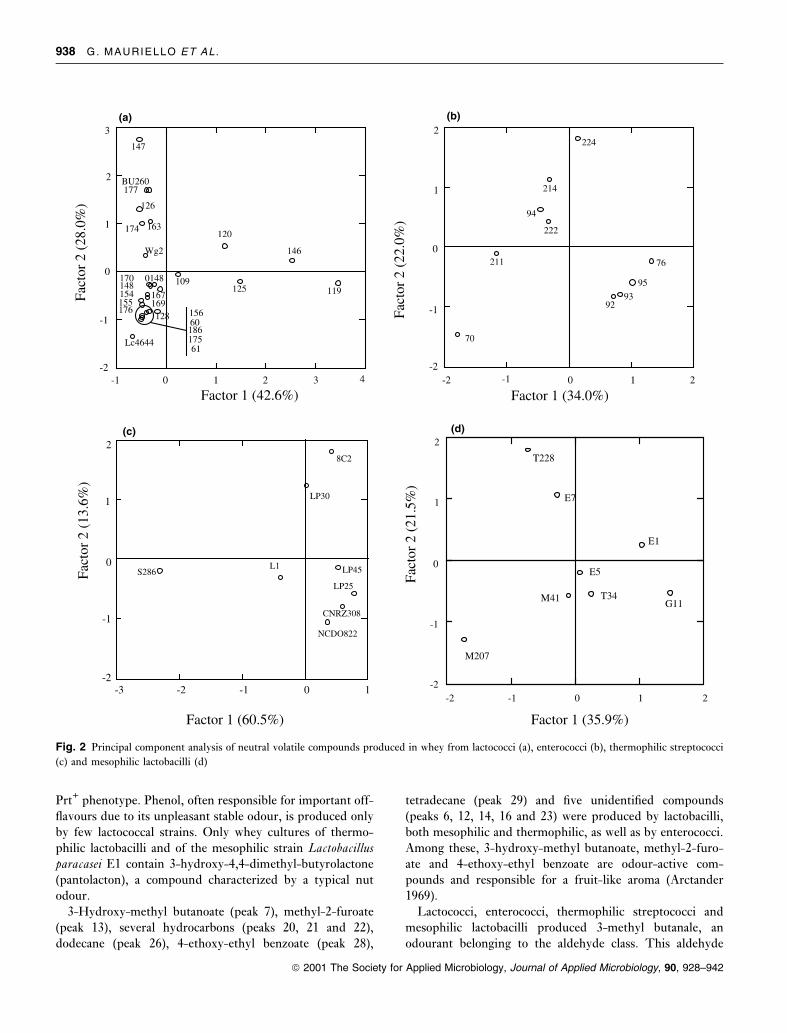
Fig. 2 Principal component analysis of neutral volatile compounds produced in whey from lactococci (a), enterococci (b), thermophilic streptococci
(c) and mesophilic lactobacilli (d)
938 G. MAURIELLO ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
was identi®ed in water buffalo Mozzarella cheese as
responsible for a typical odour of malt (Moio et al. 1993).
Finally, the concentration of furfural (peak 4), an
odourant characterized by a typical baker bread aroma and
arising from carbohydrate dehydration and cyclization
(Anon. 1995), seems to change only as a consequence of
the growth of the LA17 leuconostoc strain. However, the
presence of this odourant is unlikely to be linked to
microbial metabolism but it develops during whey powder
preparation.
Principal component analysis (PCA)
PCA was performed by comparing strains of single
microbial groups (Fig. 2) and strains of different microbial
groups (Fig. 3). As shown in Fig. 2, only for lactococci,
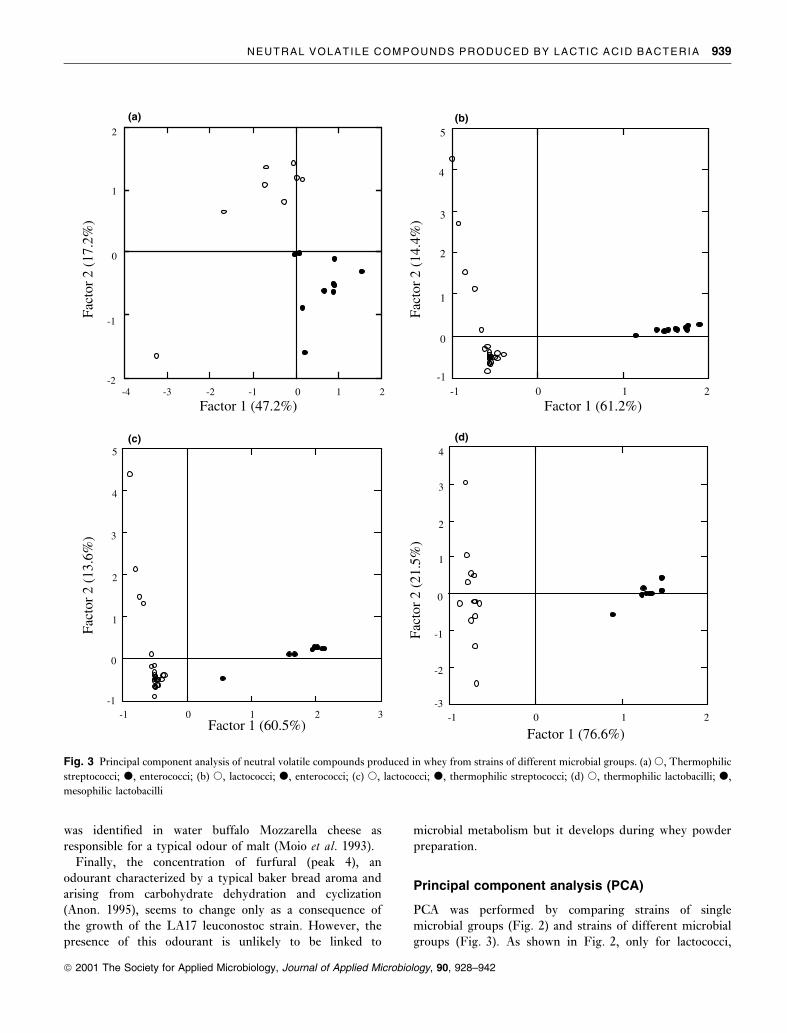
Fig. 3 Principal component analysis of neutral volatile compounds produced in whey from strains of different microbial groups. (a) s, Thermophilic
streptococci; d, enterococci; (b) s, lactococci; d, enterococci; (c) s, lactococci; d, thermophilic streptococci; (d) s, thermophilic lactobacilli; d,
mesophilic lactobacilli
NEUTRAL VOLATILE COMPOUNDS PRODUCED BY LACTIC ACID BACTERIA 939
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942
940 G. MAURIELLO ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942

enterococci, thermophilic streptococci and mesophilic lac-
tobacilli was it possible to obtain high percentages of total
variance on two components. In fact, leuconostocs and
thermophilic lactobacilli accounted for 40% and 44% of
total variance, respectively (data not shown), probably due
to the small number of substances produced by these
microorganisms.
The score plot of volatile compounds produced by
lactococci is shown in Fig. 2a; the total variance accounted
for was 62á6% with low discrimination among the strains.
Strains 109, 119, 120, 125 and 146 were well separated in
the right half of plane. In particular, strains 109, 119 and
125, placed in the lower part, produced the higher number
of volatile compounds. Strains 120 and 146, placed in the
upper part, showed a similar GC-pattern, but the latter
produced high quantities of 3-hydroxy-methyl butanoate
(lacking the GC-pattern of strain 120) and 2-methyl-
1-butanol. The citrate plasmid bearing strain DSM4644 was
well separated in the left lower part, where a single group
containing 14 lactococcal strains was also placed. However,
in this part all high-level acetoin-producing strains
(DSM4644, 148, 154, 155, 156 and 0148) were found. In
the left upper part seven strains were found, among which
the protease plasmid carrying strain DSM4367.
Enterococci, as well as mesophilic lactobacilli, showed
similar GC-patterns but the quantitative analysis allowed
reliable discrimination among the strains (Fig. 2b, d).
Thermophilic streptococci (Fig. 2c) were well separated
into eight single-strain groups according to their different
GC-patterns.
PCA results comparing strains of different groups are
reported in Fig. 3. In all the cases considered, there was
good separation between the groups, allowing each strain to
be properly ascribed to the appropriate group.
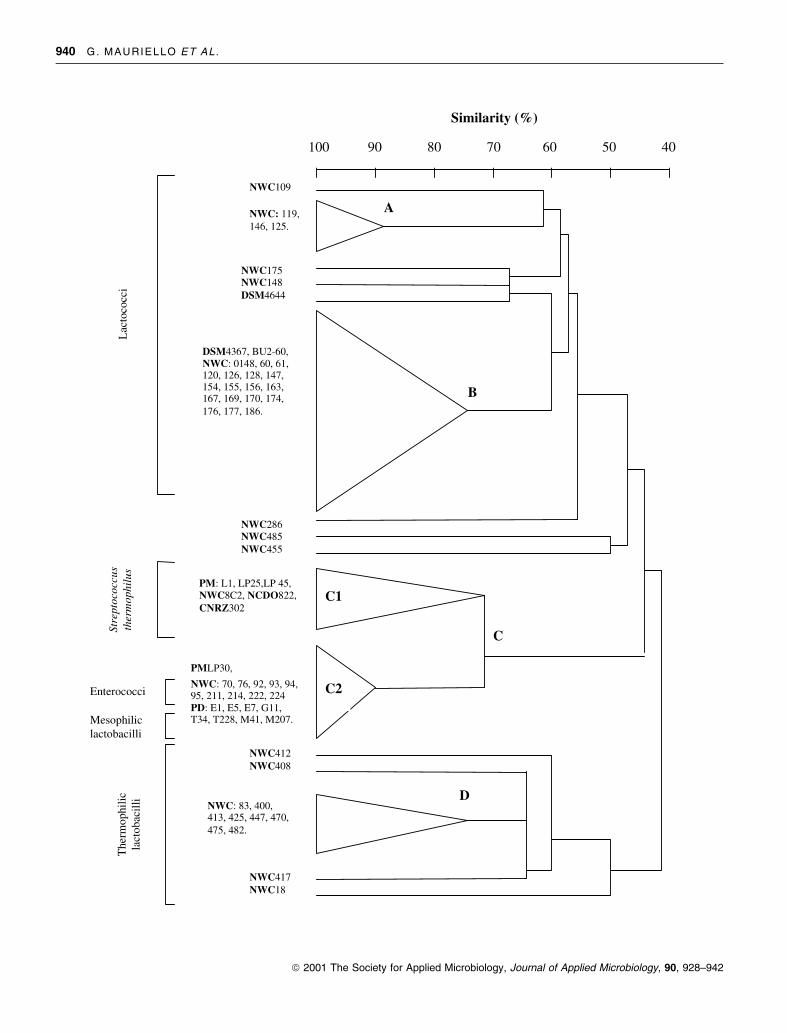
Cluster analysis (CA)
Results of HRGC were used for hierarchical cluster analysis.
As well as for PCA of the substances produced by strains
of single groups, in this analysis leuconostocs were not
included. For strain clustering purposes, concentrations
of peak ³ 0á01 ppm were coded as 1, concentrations
< 0á01 ppm were coded as 0.
Figure 4 depicts the dendrogram showing the relation-
ships among lactococci, enterococci, thermophilic strepto-
cocci, thermophilic and mesophilic lactobacilli. It was
possible to differentiate almost all the strains tested,
except for a few strains which clustered at the 100%
similarity level (data not shown). At the 71% similarity
level 15 clusters were differentiated: four main multistrain
clusters and 11 single-strain clusters. Two of the main
clusters included 23 lactococcal strains (three in cluster A
and 20 in cluster B). Cluster C included all the
enterococci, all the mesophilic lactobacilli and seven of
the eight studied thermophilic streptococci. However, in
this cluster we can discriminate a subcluster (cluster C1)
at 71% similarity level including six thermophilic strep-
tococci and a subcluster (cluster C2) at 90% similarity
level including enterococci, mesophilic lactobacilli and the
LP30 Streptococcus thermophilus strain. Cluster D included
eight strains of thermophilic lactobacilli; 12 of the 14
studied strains of this group clustered at about 50%
similarity level.
CONCLUSION
By studying Lac, Prt and Cit phenotype characterization of
78 strains of LAB, partially expected information is
achieved about their metabolic properties, lacking direct
evidence for ability to produce ¯avouring compounds.
However, the occurrence in the same habitat of strains
belonging to the same species with different phenotypes
could be observed.
By identifying the neutral volatile compounds occurring
in the whey cultures by HRGC±MS analysis, the
¯avouring potential of each strain could be de®ned,
evidencing that thermophilic streptococci, enterococci and
mesophilic lactobacilli produce a higher number of sub-
stances than the other LAB groups. Quantitative analysis
of results performed by PCA revealed that enterococci,
mesophilic lactobacilli and thermophilic streptococci are
characterized by a certain diversity, while the group of
lactococci include rather similar strains as well as strains
with special properties. Interestingly, by applying PCA to
groups such as thermophilic streptococci and enterococci
as well as mesophilic and thermophilic lactobacilli, the
strains clustered consistently with their systematic posi-
tion.
The production of neutral volatile compounds in
standard cultural conditions has therefore good potential
for characterizing LAB, allowing also preliminary screen-
ing of strains on the basis of ¯avouring capability.
However, the development of cultures to employ as
¯avouring adjuncts, undoubtedly requires experiments and
analyses at the dairy scale with reference to speci®c target
products.
Fig. 4 Dendrogram showing relationships among lactococci, entero-
cocci, thermophilic streptococci, thermophilic and mesophilic lacto-
bacilli on the basis of quantitative analysis of neutral volatile
compounds produced in whey. NWC: natural whey culture; PD: pizza
dough; PM: pasteurized milk; DSM: Deutsche Sammlung von
Mikroorganismen; NCIMB: National Collection of Industrial
and Marine Bacteria; CNRZ: Centre National de Recherches
Zootechniques
b
NEUTRAL VOLATILE COMPOUNDS PRODUCED BY LACTIC ACID BACTERIA 941
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942

ACKNOWLEDGEMENTS
This work was supported by a grant of MURST, Rome.
Authors thank Stefania Pesacane for HRGC analysis.
REFERENCES
Arctander, S. (ed.) (1969) Perfume and Flavour Chemicals Vol. I and II,
New Jersey: Montclair.
Anon. (1995) Fenaroli's Handbook of Flavour Ingredients Vol. I and II,
3rd edn. London: CRC Press.
Anon. (1998) Flavour and Fragrances International edn. Aldrich.
Belin, J.M., Bensoussan, M. and Serrano-Carreon, L. (1992) Microbial
biosynthesis: the production of food ¯avours. Trends in Food Science
and Technology 3, 11±14.
Brown, J.H. and Howe, P.E. (1922) Transparent milk as a bacterio-
logical medium. Journal of Bacteriology 7, 511±514.
Cogan, T.M. (1995) Flavour production by dairy starter cultures.
Journal of Applied Microbiology Symposium Supplement 79,
49S±64S.
Coppola, S., Parente, E., Dumontet, S. and La Peccerella, A. (1988)
The micro¯ora of natural whey cultures utilized as starter in the
manufacture of Mozzarella cheese from water-buffalo milk. Lait 68,
295±310.
Coppola, S., Pepe, O., Masi, P. and Sepe, M. (1996) Characterization
of leavened doughs for Pizza in Naples. Advances in Food Science 18,
160±162.
Devriese, L.A. and Pot, B. (1995) The genus Enterococcus. In The
Lactic Acid Bacteria, The Genera of Lactic Acid Bacteria Vol. II, ed.
Wood, B.J.B. and Holzapfel, W.H. pp. 327±367. London: Blackie
Academic.
Elliker, P.R., Anderson, A. and Hannesson, G. (1956) An agar culture
medium for lactic acid streptococci and lactobacilli. Journal of Dairy
Science 39, 1611±1612.
Garcia-Quintans, N., Christian, M., de Mendoza, D. and Lopez, P.
(1998) The citrate transport system of Lactococcus lactis subsp. lactis
biovar diacetylactis is induced by acid stress. Applied and Environ-
mental Microbiology 64, 850±857.
Hammes, W.P., Weiss, N. and Holzapfel, W. (1991) The Genera
Lactobacillus and Carnobacterium. In The Prokaryotes Vol. II, ed.
Balows, A., Truper, H.G., Dworkin, M., Harder, W. and Schleifer,
K.-H. pp. 1535±1594. New York: Springer-Verlag.
Kempler, G.M. and Mckay, L.L. (1980) Improved medium for
detection of citrate-fermenting Streptococcus lactis subsp. diacetylac-
tis. Applied and Environmental Microbiology 39, 926±927.
Law, B.A. (1981) The formation of aroma and ¯avour compounds in
fermented dairy products. Dairy Science Abstract 43, 143±154.
Levata-Jovanovic, M. and Sandine, W.E. (1997)55 A method to use
Leuconostoc mesenteroides subsp. cremoris 91404 to improve milk
fermentations. Journal of Dairy Science 80, 11±18.
Levata-Jovanovic, M. and Sandine, W.E. (1996b) Citrate utilization
and diacetyl production by various strains of Leuconostoc mesentero-
ides subsp. cremoris. Journal of Dairy Science 79, 1928±1935.
Moio, L. and Etievant, P. (1995) Ethyl anthranilate, ethyl cinnamate,
2,3-dihydrocinnamate and methyl anthranilate: four important
odorants identi®ed in Pinot noir wines of burgundy. American
Journal of Enology and Viticulture66 46, 392±398.
Moio, L., Dekimpe, J., Etievant, P. and Addeo, F. (1993) The neutral
volatile compounds of water buffalo milk. Italian Journal of Food
Science 5, 43±56.
Moio, L., Piombino, P. and Addeo, F. (2000) Odour-impact com-
pounds of Gorgonzola cheese. Journal of Dairy Research 67, 273±285.
Moschetti, G., Salzano, G., Villani, F., Pepe, O. and Coppola, S. (1993)
Biotyping strains of Streptococcus thermophilus, lactococci and entero-
cocci by DNA ®ngerprinting. In Biotechnology and Molecular Biology
of Lactic Acid Bacteria for Improvements of Foods and Feeds Quality
ed. Zamorani, A., Manachini, P.L., Bottazzi, V. and Coppola, S.
pp. 145±154. Rome: Istituto Poligra®co e Zecca dello Stato.
Moschetti, G., Blaiotta, G., Aponte, M., Villani, F., Deiana, P. and
Coppola, S. (1998) Random ampli®ed polymorphic DNA and
ampli®ed ribosomal DNA spacer polymorphism: powerful methods
to differentiate Streptococcus thermophilus strains. Journal of Applied
Microbiology 85, 25±36.
Neve, H., Geis, A. and Teuber, M. (1984) Conjugal transfer and
characterization of bacteriocin plasmids in Group N (lactic acid)
streptococci. Journal of Bacteriology 157, 833±838.
Nover de Brauw, M.C., Bouwman, J., Tas, A.C. and Lavos, G.F.
(1987) Compilation of mass spectra of volatile compounds in food.
Zeist: Food Analysis Institute, TNO-CIVO.
Schleifer, K.H., Ehrmann, M., Krusch, U. and Neve, H. (1991)
Revival of the species Streptococcus thermophilus (ex Orla-Jensen,
1919) nom. rev. Systematic and Applied Microbiology 14, 386±388.
Teuber, M., Geis, A. and Neve, H. (1991) The genus Lactococcus. In
The Prokaryotes Vol. II, ed. Balows, A., Truper, H.G., Dworkin, M.,
Harder, W. and Schleifer, K.-H. pp. 1482±1501. New York:
Springer-Verlag.
Villani, F., Moschetti, G., Blaiotta, G. and Coppola, S. (1997)
Characterization of strains of Leuconostoc mesenteroides by analysis
of soluble whole-cell protein pattern, DNA ®ngerprinting and
restriction of ribosomal DNA. Journal of Applied Microbiology 82,
578±588.
Wijtzes, T., Bruggeman, M.R., Nout, M.J.R. and Zwietering, M.H.
(1997) A computerised system for the identi®cation of lactic acid
bacteria. International Journal of Food Microbiology 38, 65±70.
942 G. MAURIELLO ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 928±942




















