
Characterization of Bacillus cereus Isolates Associated with Fatal Pneumonias: Strains Are Closely...
10
10.1128/JCM.00561-06. 2006, 44(9):3352. DOI: J. Clin. Microbiol. J. Jackson Helma, Lawrence O. Ticknor, Richard T. Okinaka and Paul Wilkins, Swati B. Avashia, Rahsaan Drumgoole, Charles H. P. Marston, Barun K. De, Tanja Popovic, David Sue, Patricia Alex R. Hoffmaster, Karen K. Hill, Jay E. Gee, Chung K. Virulence Genes B. anthracis and Harbor Bacillus anthracis Are Closely Related to Associated with Fatal Pneumonias: Strains Isolates Bacillus cereus Characterization of http://jcm.asm.org/content/44/9/3352 Updated information and services can be found at: These include: SUPPLEMENTAL MATERIAL Supplemental material REFERENCES http://jcm.asm.org/content/44/9/3352#ref-list-1 at: This article cites 24 articles, 14 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on March 17, 2014 by guest http://jcm.asm.org/ Downloaded from on March 17, 2014 by guest http://jcm.asm.org/ Downloaded from
Transcript of Characterization of Bacillus cereus Isolates Associated with Fatal Pneumonias: Strains Are Closely...

10.1128/JCM.00561-06.
2006, 44(9):3352. DOI:J. Clin. Microbiol. J. JacksonHelma, Lawrence O. Ticknor, Richard T. Okinaka and PaulWilkins, Swati B. Avashia, Rahsaan Drumgoole, Charles H.
P.Marston, Barun K. De, Tanja Popovic, David Sue, Patricia Alex R. Hoffmaster, Karen K. Hill, Jay E. Gee, Chung K.
Virulence GenesB. anthracisand Harbor Bacillus anthracisAre Closely Related to
Associated with Fatal Pneumonias: Strains IsolatesBacillus cereusCharacterization of
http://jcm.asm.org/content/44/9/3352Updated information and services can be found at:
These include:
SUPPLEMENTAL MATERIAL Supplemental material
REFERENCEShttp://jcm.asm.org/content/44/9/3352#ref-list-1at:
This article cites 24 articles, 14 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from
on M
arch 17, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

JOURNAL OF CLINICAL MICROBIOLOGY, Sept. 2006, p. 3352–3360 Vol. 44, No. 90095-1137/06/$08.00�0 doi:10.1128/JCM.00561-06
Characterization of Bacillus cereus Isolates Associated with Fatal Pneumonias:Strains Are Closely Related to Bacillus anthracis and
Harbor B. anthracis Virulence Genes†Alex R. Hoffmaster,1* Karen K. Hill,4 Jay E. Gee,1 Chung K. Marston,1 Barun K. De,1 Tanja Popovic,2
David Sue,1‡ Patricia P. Wilkins,1 Swati B. Avashia,3,4§ Rahsaan Drumgoole,4 Charles H. Helma,5Lawrence O. Ticknor,6 Richard T. Okinaka,5 and Paul J. Jackson7
Epidemiologic Investigations Laboratory, Meningitis and Special Pathogens Branch, MS G34,1 and Office of the Director2 andEpidemic Intelligence Service,3 MS E92, Centers for Disease Control and Prevention, 1600 Clifton Road, Atlanta,
Georgia 30333; Infectious Disease Epidemiology and Surveillance Division, T-801, Texas Department ofState Health Services, Austin, Texas 787564; Bioscience5 and Decision Applications6 Divisions,Los Alamos National Laboratory, Los Alamos, New Mexico 87545; and Defense Biology Division,
Lawrence Livermore National Laboratory, Livermore, California 945517
Received 15 March 2006/Returned for modification 19 June 2006/Accepted 10 July 2006
Bacillus cereus is ubiquitous in nature, and while most isolates appear to be harmless, some are associatedwith food-borne illnesses, periodontal diseases, and other more serious infections. In one such infection, B.cereus G9241 was identified as the causative agent of a severe pneumonia in a Louisiana welder in 1994. Thisisolate was found to harbor most of the B. anthracis virulence plasmid pXO1 (13). Here we report thecharacterization of two clinical and one environmental B. cereus isolate collected during an investigation of twofatal pneumonia cases in Texas metal workers. Molecular subtyping revealed that the two cases were notcaused by the same strain. However, one of the three isolates was indistinguishable from B. cereus G9241. PCRanalysis demonstrated that both clinical isolates contained B. anthracis pXO1 toxin genes. One clinical isolateand the environmental isolate collected from that victim’s worksite contained the cap A, B, and C genesrequired for capsule biosynthesis in B. anthracis. Both clinical isolates expressed a capsule; however, neitherwas composed of poly-D-glutamic acid. Although most B. cereus isolates are not opportunistic pathogens andonly a limited number cause food-borne illnesses, these results demonstrate that some B. cereus strains cancause severe and even fatal infections in patients who appear to be otherwise healthy.
Bacillus cereus is ubiquitous in nature and a common causeof emetic and diarrheal food poisoning. Most B. cereus isolatesappear to be harmless, but some are considered opportunisticpathogens. In immunocompromised individuals or patients re-covering from surgery, B. cereus can cause a variety of infec-tions, including endophthalmitis, bacteremia, septicemia, en-docarditis, salpingitis, cutaneous infections, pneumonia, andmeningitis (19). Some strains are known to cause periodontaldisease (8). Recently there have been reports of severe andsometimes fatal cases of pneumonia caused by B. cereus inapparently healthy welders (13, 20). The severity of these caseswas unusual for B. cereus infections, and the patients wereneither immunocompromised nor had any known underlyingconditions causing susceptibility to these infections.
B. cereus G9241, which was associated with severe pneumo-nia in a welder from Louisiana in 1994, has been well charac-terized, and its genome has been sequenced and analyzed (13).
Genomic analysis and multilocus sequence typing (MLST) ofthis isolate revealed it to be closely related to B. anthracis.Several methods have shown that B. cereus isolates closelyrelated to B. anthracis tend to be of clinical rather than envi-ronmental origin (9, 10, 11). However, it is very uncommon forB. cereus isolates, even those that are closely related to B.anthracis, to carry B. anthracis virulence plasmids (23). B.cereus G9241 carries an almost complete pXO1 plasmid, des-ignated pBCXO1. This isolate also harbors a 218-kb circularplasmid (pBC218) and a cryptic bacteriophage (pBClin29)(13). Another unique feature of G9241 relative to other B.cereus isolates is the presence of a capsule. However, thiscapsule is not composed of D-glutamyl polypeptides and is notencoded by the B. anthracis cap genes normally located on theB. anthracis pXO2 plasmid. Instead, it has been hypothesizedto be a polysaccharide and be encoded by a putative polysac-charide capsule biosynthetic operon located on pBC218 (13).While the presence of these plasmids in an isolate that causessevere disease similar to inhalation anthrax is intriguing, theirroles, if any, in the virulence of the isolate or the presentationof disease has not yet been determined.
In October and November 2003, two fatal cases of B. cereuspneumonia occurred in metal workers (a welder and a mulleroperator) at different locations in Texas. A detailed report ofthese cases and the epidemiologic investigation is in prepara-tion (S. B. Avashia, submitted for publication). In this report,we describe the initial molecular genetic characterization of
* Corresponding author. Mailing address: 1600 Clifton Road, MSG34, Atlanta, GA 30333. Phone: (404) 639-0852. Fax: (404) 639-3023.E-mail: [email protected].
† Supplemental material for this article may be found at http://jcm.asm.org/.
‡ Present address: Department of Microbiology and Immunology,Emory University School of Medicine, 3004 Rollins Research Center,Atlanta, GA 30322.
§ Present address: Internal Medicine/Pediatrics, Seton KozmetskyClinic, 3706 South First Street Austin, TX 78704.
3352
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from

two clinical and one environmental B. cereus isolate from theinvestigation of these two fatal cases. We also describe howthese isolates are related to each other, to B. anthracis, and toother previously characterized B. cereus and B. thuringiensisisolates.
MATERIALS AND METHODS
Origin of B. cereus isolates. The two clinical B. cereus isolates described in thispaper were collected from two fatal pneumonia cases that occurred in Texasmetal workers within a 3-week period in 2003. The first isolate, B. cereus03BB102, was collected from a 39-year-old white male welder, while the secondisolate, B. cereus 03BB87, was cultured from a 56-year-old black male mulleroperator. Both B. cereus isolates were cultured from blood. In an effort toidentify the source of the infections, environmental samples were cultured forBacillus spp. and isolates were screened by PCR for the presence of pXO1 orpXO2. This screen resulted in the identification of a single B. cereus isolate fromsettled dust at the welder’s worksite; the isolate, 03BB108, was PCR positive forthe pXO2 cap genes and thus further characterized in this study. A summary ofthe strains and their common identifiers used throughout the paper is includedin Table 1. The B. cereus, B. anthracis, and B. thuringiensis isolates used forcomparison in phylogenetic studies were provided by the U.S. Department ofHomeland Security Microbial Strain Archive maintained at Los Alamos NationalLaboratory, Brigham Young University, and the Centers for Disease Control andPrevention.
Biochemical and phenotypic characterization. The B. cereus isolates werecharacterized by standard microbiological methods (19). Motility was deter-mined by microscopic observation of wet mounts of cells grown in heart infusionbroth. Testing to determine susceptibility to gamma phage (3) was performed byadding 5 �l of gamma phage (3.8 � 108 PFU/ml) on the first and secondquadrants of isolation streaks on Trypticase soy agar plates containing 5%(vol/vol) sheep blood (Becton Dickinson Microbiology Systems, Cockeysville,MD) and incubating at 37°C overnight. The B. anthracis-specific, two-componentdirect fluorescent assay (DFA) for the detection of the B. anthracis cell wallantigen and capsule antigen was performed as previously described by De et al.(6). Capsule staining of cells was performed on cells incubated at 30 and 37°C inheart infusion broth supplemented with 0.8% sodium bicarbonate and 50%horse serum as used for the capsule DFA (6). Capsules were visualized usingIndia ink (REMEL, Lenexa, KS) and a 100� oil immersion objective as de-scribed previously (13).
DNA isolation and purification. Five milliliters of nutrient broth was inocu-lated with bacteria from a single colony of each isolate, and each culture wasincubated overnight with shaking at 28°C. Four colonies from each isolate wereused to produce four different cultures. Bacterial cells were harvested by cen-trifugation at 1,000 � g for 15 min. The resulting pellets were subjected to threefreeze-thaw cycles. DNA was isolated from the disrupted cells using a QIAamptissue kit (QIAGEN, Inc., Valencia, CA) following the protocol provided by the
manufacturer. The quantity and quality of the isolated DNA were determined bygel electrophoresis of a small amount of the sample through a 1.0% agarose geldissolved in a solution containing 10 mM Tris borate (pH 8.3) and 1 mM EDTAfor 1 h at 80 V (26). Gels were stained for 20 min with a solution containing 1 �gethidium bromide/ml, destained in distilled water, and visualized and photo-graphed under UV light. Known concentrations of DNA molecular-weight stan-dards were compared to the isolated bacterial DNA to determine the size andapproximate concentration of the bacterial DNA.
PCR amplification of DNA. DNA oligomers that functioned as PCR primersto amplify representative B. anthracis gene fragments are listed in Table S1 in thesupplemental material. Primers designed to amplify a portion of all bacterial 16SrRNA genes were used as positive PCR controls. Initial PCR contained 10 mMTris-HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.001% (wt/vol) gelatin, 0.2 mMof each dNTP, 20 pmol of each primer, 2.5 U of AmpliTaq DNA polymerase,and approximately 1 ng template DNA in a 100-�l total reaction volume.PerkinElmer reagents (Foster City, CA) were used for all reactions. TemplateDNA was initially denatured by heating at 94°C for 2 min. This was followed by35 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min, andprimer extension at 72°C for 1 min. Incubation for 5 min at 72°C followed tocomplete the extension. PCR was conducted in a PerkinElmer GeneAmp PCRsystem 9600 thermal cycler. Control reactions containing B. anthracis 91-429C-2DNA demonstrated whether each PCR primer set functioned as expected toamplify the appropriate B. anthracis plasmid or genomic amplicon.
Analysis of PCR amplicons. PCR amplicons were analyzed by electrophoresisthrough a 3.0% agarose gel dissolved in a solution containing 10 mM Tris borate(pH 8.3) and 1 mM EDTA. Electrophoresis was for 1 h at 80 V. Gels werestained for 20 min with a solution containing 1 �g of ethidium bromide/ml,destained in distilled water, and then visualized and photographed under UVlight. Images were captured electronically using a Stratagene Eagle Eye II stillvideo system (Stratagene).
Protective antigen (PA) gene sequencing and multilocus sequence typing. ThepagA gene was amplified and sequenced as described previously (12, 24). How-ever, additional primers amplifying a 1,076-nucleotide fragment between pXO1nucleotide 134,396 and 135,472 (see Table S1 in the supplemental material) wereused in an attempt to amplify the 5� end of pagA in B. cereus 03BB102 becausethis gene failed to amplify using previously described primers. MLST was basedon the analysis of seven housekeeping gene partial sequences (glpF, gmk, ilvD,pta, pur, pycA, and tpi) and is described online at http://pubmlst.org/bcereus (25).Phylogenetic and molecular evolutionary analyses were conducted using MEGA,version 2.1 (18).
AFLP analysis of DNA samples. Amplified fragment length polymorphism(AFLP) analysis was accomplished as previously described (11, 14, 15, 26).Briefly, 100 ng of DNA was digested with the restriction endonucleases EcoRIand MseI and the resulting fragments were ligated to double-stranded adapters.The digested and ligated DNA was then amplified by PCR using EcoRI andMseI �0/�0 primers (5�-GTAGACTGCGTACCAATTC-3� and 5�-GACGATGAGTCCTGAGTAA-3�, respectively). The � 0/�0 PCR product was analyzedby agarose gel electrophoresis to determine the size range of amplified frag-
TABLE 1. Phenotypic, antigenic and pagA sequence analysis of strains
Species/strain Source Yearisolated Reference Hemolysis Motility Gamma-
phageb
DFA result for:Capsule
stain
pagA (PA)sequence
typegB. anthraciscell wallc
B. anthraciscapsulec
B. anthracis ASC69 CAMRa 1957 Nonhemolytic Nonmotile Susceptible Positive Positive Positived 6B. cereus G9241 La. welder 1994 13 Hemolytic Motile Resistant Positive Negative Positivee 9h
B. cereus 03BB87 Lubbock, Tex.,Muller operator
2003 This study Hemolytic Motile Resistant Positive Negative Positive f 9
B. cereus 03BB102 San Antonio,Tex., welder
2003 This study Hemolytic Motile Resistant Positive Negative Positivef Notdeterminedi
B. cereus 03BB108 San Antonio, Tex.,environmental
2003 This study Hemolytic Motile Resistant Positive Negative Negative Not present
a CAMR, Center for Applied Microbiology and Research, Porton Down, United Kingdom.b Gamma phage susceptibility is measured by the capacity of the gamma bacteriophage to lyse cultures. The titer of the bacteriophage stock used was 3.8 � 108
PFU/ml.c B. anthracis-specific DFAs as described by De et al. (6).d Poly-�-D-glutamic acid capsule.e Not a poly-�-D-glutamic acid capsule. Hypothesized to be polysaccharide based on genomic analysis (13).f Not a poly-�-D-glutamic acid capsule. Capsule expression was observed by adjusting incubation conditions.g Based on the method of Price et al. (24).h pagA on plasmid pBCXO1 (ORFC00004); not the more divergent copy on pBC218 (13).i Attempts to amplify the extreme 5� end of the gene were not successful and resulted in an incomplete sequence (the first 15 bases were not sequenced).
VOL. 44, 2006 BACILLUS CEREUS ASSOCIATED WITH FATAL PNEUMONIAS 3353
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from

ments. Three microliters of each product was used as a template in subsequentselective amplifications using the �1/�1 primer combination of 6-carboxyfluo-rescein-labeled EcoRI-C (5�-GTAGACTGCGTACCAATTCC-3�) and MseI-G(5�-GACGATGAGTCCTGAGTAAG-3�). Selective amplifications were per-formed in 20-�l reaction mixtures. The resulting products (0.5 to 1.0 �l) weremixed with a solution containing a mixture of DNA size standards (GeneScan-500 [Applied Biosystems Inc., Foster City, CA] and MapMarker 500 [Bio-Ventures, Inc., Murfreesboro, TN]) labeled with N,N,N,N-tetramethyl-6-car-boxyrhodamine. Following a 2-min heat denaturation at 90°C, the reactions wereloaded onto 5% Long Ranger DNA sequencing gels (Cambrex Bio Science,Rockland, ME) and visualized on an ABI 377 automated fluorescent sequencer.To make sure that the DNA assayed in these experiments was representative ofeach of the isolates, four individual colonies from each isolate were inoculatedinto medium and DNA purified. The DNA preparations were then analyzedusing AFLP. Forty DNA fragments generated from the AFLP experiments wereused to represent each of the DNA preparations. These DNA preparations fromindividual colonies of the same isolate were indistinguishable from each other.Therefore, only one DNA preparation was used in the other experiments. Eachset of reactions also contained an AFLP reaction using B. anthracis Vollum DNAas an experimental control. The inclusion of such a reaction in each analysis setallowed a comparison of results from earlier archived analysis sets run at differ-ent times or on different gels. GeneScan analysis software (Applied Biosystems,Inc.) was used to determine the lengths of the sample fragments by comparisonsto the DNA fragment length size standards included with each sample.
To minimize gel electrophoresis artifacts, each labeling reaction was run intriplicate. Samples were loaded on three different gels in a random order. AFLPdata analysis was performed as previously described by Ticknor et al. (26).Sample fragments of between 100 and 500 bp, with fluorescence above 50arbitrary units in all three runs on the ABI sequencer, were used in the analysis.
Similarities among samples were determined by the Jaccard coefficient. The 40tallest peaks for each sample fingerprint were used to calculate the Jaccardcoefficient among samples. Dendrograms were produced by using the similaritymatrix of Jaccard coefficients and the unweighted pair-group mean averagemethod (F. J. Rohlf, NTSYS-PC numerical taxonomy and multivariate analysissystem, version 1.8; Exeter Software, Setauket, NY). All statistical data manip-ulations were performed using codes developed in S-Plus (Data Analysis Prod-ucts Division, MathSoft, Seattle, WA).
RESULTS
Phenotypic analysis. The B. cereus isolates were hemolytic,motile, and resistant to lysis by gamma-phage and had pheno-types typical of the species (Table 1). Each of the three isolateshad distinct colony morphologies. B. cereus 03BB102 (isolatedfrom the Texas welder) produced small, convex, smooth colo-nies, demonstrating a greater zone of hemolysis on sheepblood agar than that of B. cereus 03BB87 (isolated from theTexas muller operator). This latter isolate produced large, flat,tan, granular colonies. The B. cereus 03BB108 (collected fromthe welder’s worksite) was intermediate in size, umbonate(having a raised center), tan, and granular.
The two-component B. anthracis-specific DFA (6) was per-formed on all three isolates and produced positive results forthe cell wall antigen but was negative for the B. anthraciscapsule antigen (poly-�-D-glutamic acid) when cells were incu-bated under several conditions, including conditions used forB. anthracis capsule expression. B. cereus clinical isolates aresometimes positive for the cell wall antigen. B. cereus G9241,isolated from a Louisiana welder (13), also tested positive forthis cell wall antigen. However, unlike the results obtained forB. cereus G9241, India ink staining of the two Texas clinicalisolates did not initially reveal any type of capsule when grownon sheep blood agar at 37°C in ambient atmosphere due to avery small percentage of the cells being encapsulated (Table1). Adjusting the medium and temperature resulted in in-creased capsule formation, but not to the level exhibited by B.
cereus G9241. A capsule was not detected on the environmen-tal isolate 03BB108.
Plasmid PCR analysis. PCR analysis was performed usingprimers designed to amplify sequences from pXO1, pXO2, andpBC218. Table S1 in the supplemental material shows the
TABLE 2. Results of PCR analysis using primers that amplifyportions of different open reading frames found on
B. anthracis plasmids pXO1 and pXO2a
Targeted ORF
Isolate identification for
B. cereus03BB102
(central Tex.clinical)
B. cereus03BB108
(central Tex.environmental)
B. cereus03BB87
(north Tex.clinical)
B.cereusG9241
pXO1-01 �� �� � �pXO1-02 �� �� �� ��pXO1-03 �� �� �� ��pXO1-04 �� �� �� ��pXO1-05 �� �� �� ��pXO1-12 � � �� ��pXO1-18 � � �� ��pXO1-30 � � �� ��pXO1-37 � � �� ��pXO1-43 �� �� �� ��pXO1-52 � �� �� ��pXO1-61 �� �� �� ��pXO1-74 �� �� �� ��pXO1-84 � � �� ��pXO1-89 � � �� ��pXO1-90 �� � �� ��pXO1-91 �� � �� ��pXO1-92 �� � �� ��pXO1-93 �� � �� ��pXO1-94 �� �� �� ��pXO1-95 �� � �� ��pXO1-97 �� �� �� ��pXO1-106 �� � �� ��pXO1-107 (lef) �� � �� ��pXO1-108 �� � �� ��pX01-110 (pagA) �� � �� ��pXO1-115 � � �� ��pXO1-120 �� � �� ��pX01-122 (cya) �� � �� ��pXO1-130 �� � �� ��pXO1-135 � � � �pXO1-143 � � � �pX02-10 � � � �pX02-24 � � � �pX02-32 � � � �pX02-39 � � � �pX02-47 � � � �pX02-53 � � � �pX02-56 (capA) �� �� � �pX02-57 (capC) �� �� � �pX02-58 (capB) �� �� � �pX02-63 � � � �pX02-68 � � � �pX02-73 � � � �pX02-79/80 � � � �P3MOD, PC5B �� �� �� ��Chromosomal targetsb � � � �
a �, no visible amplicon; ��, the amplicon was present and was the expectedlength relative to amplicons produced using B. anthracis Vollum DNA as tem-plate. ORF numbers are those assigned by Okinaka, et al. (22). All primer setsamplified the expected amplicon, based on the length of the amplicon, when B.anthracis 91-429C-2 was included as template in the reactions.
b Reactions using nine different PCR primer sets that amplify chromosomalsequences thought to be highly specific to B. anthracis did not produce anyamplicons of the expected size and sequence.
3354 HOFFMASTER ET AL. J. CLIN. MICROBIOL.
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from
sequences and targets for a battery of PCR primers used toassay for the presence of different pXO1, pXO2, and pBC218open reading frames (ORFs). Table 2 shows the results of PCRassays to detect pXO1 and pXO2 sequences. All primers gen-erated the expected amplicon when the control strain (B.anthracis 91-429C-2) DNA was used as the template.
The results obtained for B. cereus 03BB87 were identical tothose obtained for B. cereus G9241, which was collected fromthe Louisiana welder almost 10 years earlier. Both were eithermissing pXO1-ORF 1, pXO1-ORF 135, and pXO1-ORF 143or the sequences in the priming sites within these ORFs weresufficiently different so that the primers used did not amplify aproduct. Sequences complementary to these primer sequenceswere identical in the four B. anthracis isolates so far sequenced.A search of the G9241 plasmid pBC218 did not show homologyto any of these primer sequences. In contrast, neither B. cereus03BB87 nor B. cereus G9241 contained sequences with sufficienthomology to any of the pXO2 sequences tested to allow amplifi-cation of the different pXO2-specific ORF fragments.
B. cereus 03BB102 and 03BB108, isolated from the Texaswelder and his worksite, respectively, did not contain se-quences of sufficient homology to generate PCR amplicons
from many of the pXO1 ORFs tested (Table 2). The amplifi-cation profile of B. cereus 03BB108 was a subset of that for B.cereus 03BB102. This latter isolate contained pXO1 ORFs thatwere not present in B. cereus 03BB108. A possible explanationof this result is a loss of DNA from B. cereus 03BB102 toproduce the profile seen in B. cereus 03BB108, but the patternof ORFs missing from B. cereus 03BB108 relative to B. cereus03BB102 is complex relative to the order of these ORFs in theB. anthracis pXO1 plasmid. While B. cereus 03BB102 testedpositive for several genes located within the pXO1 pathoge-nicity island (22), including the three toxin genes (pagA, lef,and cya), the environmental isolate (B. cereus 03BB108) wasmissing the majority of the pathogenicity island ORFs.
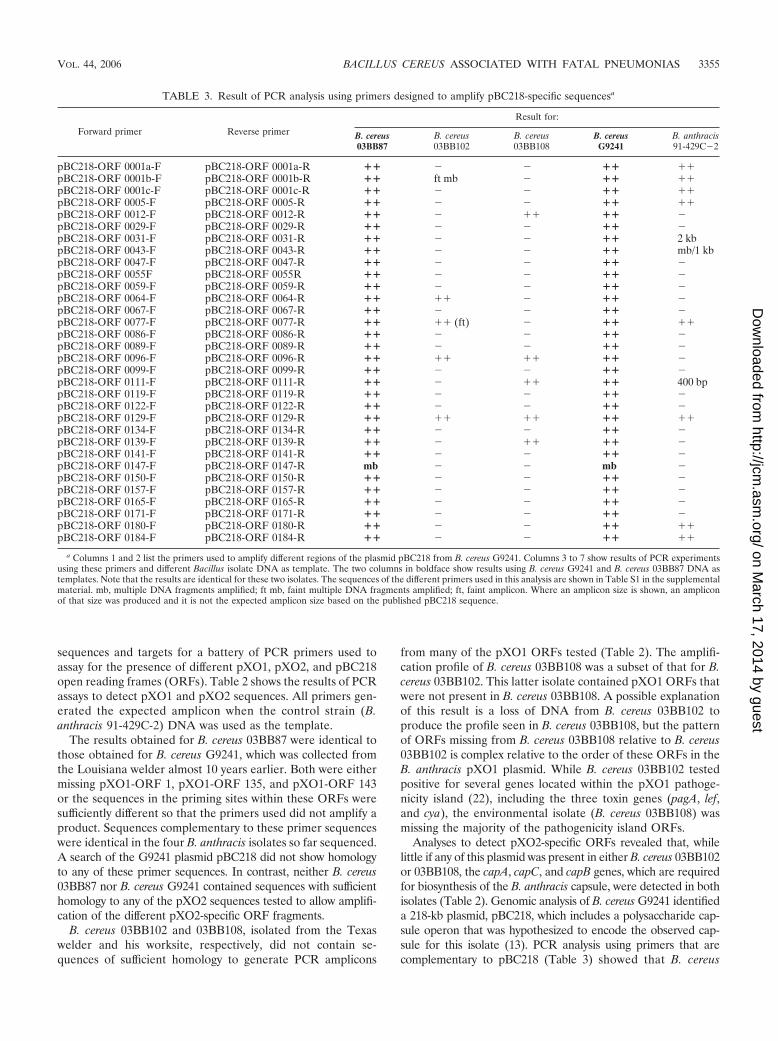
Analyses to detect pXO2-specific ORFs revealed that, whilelittle if any of this plasmid was present in either B. cereus 03BB102or 03BB108, the capA, capC, and capB genes, which are requiredfor biosynthesis of the B. anthracis capsule, were detected in bothisolates (Table 2). Genomic analysis of B. cereus G9241 identifieda 218-kb plasmid, pBC218, which includes a polysaccharide cap-sule operon that was hypothesized to encode the observed cap-sule for this isolate (13). PCR analysis using primers that arecomplementary to pBC218 (Table 3) showed that B. cereus
TABLE 3. Result of PCR analysis using primers designed to amplify pBC218-specific sequencesa
Forward primer Reverse primer
Result for:
B. cereus03BB87
B. cereus03BB102
B. cereus03BB108
B. cereusG9241
B. anthracis91-429C�2
pBC218-ORF 0001a-F pBC218-ORF 0001a-R �� � � �� ��pBC218-ORF 0001b-F pBC218-ORF 0001b-R �� ft mb � �� ��pBC218-ORF 0001c-F pBC218-ORF 0001c-R �� � � �� ��pBC218-ORF 0005-F pBC218-ORF 0005-R �� � � �� ��pBC218-ORF 0012-F pBC218-ORF 0012-R �� � �� �� �pBC218-ORF 0029-F pBC218-ORF 0029-R �� � � �� �pBC218-ORF 0031-F pBC218-ORF 0031-R �� � � �� 2 kbpBC218-ORF 0043-F pBC218-ORF 0043-R �� � � �� mb/1 kbpBC218-ORF 0047-F pBC218-ORF 0047-R �� � � �� �pBC218-ORF 0055F pBC218-ORF 0055R �� � � �� �pBC218-ORF 0059-F pBC218-ORF 0059-R �� � � �� �pBC218-ORF 0064-F pBC218-ORF 0064-R �� �� � �� �pBC218-ORF 0067-F pBC218-ORF 0067-R �� � � �� �pBC218-ORF 0077-F pBC218-ORF 0077-R �� �� (ft) � �� ��pBC218-ORF 0086-F pBC218-ORF 0086-R �� � � �� �pBC218-ORF 0089-F pBC218-ORF 0089-R �� � � �� �pBC218-ORF 0096-F pBC218-ORF 0096-R �� �� �� �� �pBC218-ORF 0099-F pBC218-ORF 0099-R �� � � �� �pBC218-ORF 0111-F pBC218-ORF 0111-R �� � �� �� 400 bppBC218-ORF 0119-F pBC218-ORF 0119-R �� � � �� �pBC218-ORF 0122-F pBC218-ORF 0122-R �� � � �� �pBC218-ORF 0129-F pBC218-ORF 0129-R �� �� �� �� ��pBC218-ORF 0134-F pBC218-ORF 0134-R �� � � �� �pBC218-ORF 0139-F pBC218-ORF 0139-R �� � �� �� �pBC218-ORF 0141-F pBC218-ORF 0141-R �� � � �� �pBC218-ORF 0147-F pBC218-ORF 0147-R mb � � mb �pBC218-ORF 0150-F pBC218-ORF 0150-R �� � � �� �pBC218-ORF 0157-F pBC218-ORF 0157-R �� � � �� �pBC218-ORF 0165-F pBC218-ORF 0165-R �� � � �� �pBC218-ORF 0171-F pBC218-ORF 0171-R �� � � �� �pBC218-ORF 0180-F pBC218-ORF 0180-R �� � � �� ��pBC218-ORF 0184-F pBC218-ORF 0184-R �� � � �� ��
a Columns 1 and 2 list the primers used to amplify different regions of the plasmid pBC218 from B. cereus G9241. Columns 3 to 7 show results of PCR experimentsusing these primers and different Bacillus isolate DNA as template. The two columns in boldface show results using B. cereus G9241 and B. cereus 03BB87 DNA astemplates. Note that the results are identical for these two isolates. The sequences of the different primers used in this analysis are shown in Table S1 in the supplementalmaterial. mb, multiple DNA fragments amplified; ft mb, faint multiple DNA fragments amplified; ft, faint amplicon. Where an amplicon size is shown, an ampliconof that size was produced and it is not the expected amplicon size based on the published pBC218 sequence.
VOL. 44, 2006 BACILLUS CEREUS ASSOCIATED WITH FATAL PNEUMONIAS 3355
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from

03BB87 has an identical PCR profile to B. cereus G9241 for32 different pBC218 markers scattered around the plasmid,strongly suggesting the presence of pBC218 or a very similarplasmid in B. cereus 03BB87. In particular, B. cereus 03BB87possessed the putative polysaccharide polymerase gene (ORFpBC218-0073) thought to participate in the synthesis of a poly-saccharide capsule, while B. cereus 03BB102 and 03BB108 didnot possess this gene.
PCR analysis using primers that amplify sequences dispersed
throughout the B. anthracis genome and shown to be very specificfor B. anthracis chromosomal sequences showed that none ofthese sequences were present in B. cereus 03BB87, 03BB102,03BB108, or G9241. (Table 2). In contrast, these sequences arepresent in all 79 B. anthracis isolates tested to date.
pagA sequencing. The B. anthracis protective antigen genepagA was detected by PCR in B. cereus 03BB87 and 03BB102.To determine similarities between the B. anthracis pagA gene,G9241 pagA, and the pagA sequences from these isolates, we
FIG. 1. AFLP-based phylogeny of different B. anthracis, B. cereus, and B. thuringiensis isolates, including the Texas and Louisiana B. cereusisolates. A comparison of AFLP profiles generated from B. cereus G9241 and the three Texas isolates, B. cereus 03BB87, 03BB102, and 03BB108,to profiles from a variety of B. anthracis, B. cereus, and B. thuringiensis isolates placed all four isolates into the phylogenetic branch containing allknown B. anthracis isolates and many characterized pathogenic and toxigenic B. cereus and B. thuringiensis isolates. Arrows show that B. cereus03BB87 and B. cereus G9241, clinical isolates from the Texas muller operator and Louisiana welder, respectively, are very similar or identical andthat B. cereus 03BB102, the clinical isolate from the Texas welder is closely related to, but clearly distinct from, B. cereus 03BB108 collected fromthe environment of this individual’s workplace. Bc, B. cereus; Bt, B. thuringiensis.
3356 HOFFMASTER ET AL. J. CLIN. MICROBIOL.
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from

attempted to amplify and sequence pagA from each of theTexas clinical isolates. The pagA sequence in B. cereus 03BB87was identical to the pagA sequence from B. cereus G9241 (13)and has been assigned the designation of PA genotype 9 (Table1). Attempts to amplify the extreme 5� end of pagA from B.cereus 03BB102 were not successful, despite using additionalprimers. We were therefore able to generate only a partialpagA sequence that was missing the first 15 bases relative to thepublished B. anthracis pagA sequence (see reference 24 andother GenBank entries for this gene). A comparison of thispartial sequence to the pagA gene sequence amplified from B.cereus 03BB87 and G9241 revealed two additional mutations atpositions 1862 and 1898, which further distinguishes them fromthe other two isolates and previously sequenced B. anthracisisolates. These two mutations result in serine-to-asparagineand lysine-to-threonine changes, respectively, and are locatedin the N-terminal 20-kDa PA fragment (PA20) that is cleavedfrom PA63 (63-kDa C-terminal fragment) by a class of furinproteases (21).
Molecular subtyping. All three Texas isolates and the Lou-isiana isolate (G9241) were analyzed using AFLP to under-stand their genetic relationship to one another, to B. an-thracis, and to other previously characterized B. cereus andB. thuringiensis isolates. AFLP is a multilocus samplingmethod that relies on the generation of a fingerprint orsignature that is created by restriction enzyme digestion ofthe entire genome and any extrachromosomal material thatmight be present. After digestion with restriction enzymesand further selective PCR amplification, the AFLP finger-print for an isolate was represented by approximately 40fragments between 100 and 500 bp in length that werepresent in all three replicates of each isolate. These finger-
print fragment sizes were then used to generate a phyloge-netic tree that illustrates the genetic relationships amongthe isolates analyzed. The fingerprints generated from thethree Texas isolates were compared to a fingerprint gener-ated from total DNA isolated from the B. anthracis Vollumstrain that is included in every AFLP analysis as a control.They were also compared to fingerprints generated for abattery of B. cereus and B. thuringiensis isolates that areknown to be closely related to B. anthracis. Figure 1 showsthat, relative to the majority of B. cereus and B. thuringiensisisolates studied, all three Texas isolates (B. cereus 03BB87,03BB102, and 03BB108) and the Louisiana isolate (B. cereusG9241) are very closely related to B. anthracis (11, 26).However, they are clearly distinct from this pathogen. Theymapped to a branch of the phylogenetic tree that includesmany of the known pathogenic and toxigenic B. cereus andB. thuringiensis isolates so far characterized (11), while mostB. cereus and B. thuringiensis isolates map to other branchesof the tree. AFLP analysis could not differentiate B. cereus03BB87, isolated from the Texas muller operator, from B.cereus G9241, isolated from the Louisiana welder in 1994. Incontrast, B. cereus 03BB102 is phylogenetically significantlydifferent from these two clinical isolates. Its AFLP fragmentprofile was similar to, but distinct from, the profile gener-ated for B. cereus 03BB108, the isolate collected from theworking environment of the B. cereus 03BB102-infected vic-tim (Texas welder). Figure 2 shows a direct comparison of amajor portion of the AFLP DNA fragment profiles for thetwo clinical isolates and that of G9241. A direct comparisonof the AFLP profiles of B. cereus G9241, isolated in 1994from a Louisiana welder, and B. cereus 03BB87, isolated in2003 from a Texas muller operator (Fig 2A), showed that,
FIG. 2. Portions of the AFLP profiles generated for B. cereus 03BB87, collected from the Texas muller operator (panel A, black profile), B.cereus G9241, collected from the Louisiana welder (panel A, red profile), and B. cereus 03BB102, collected from the Texas welder (panel B, blackprofile). Approximately two thirds of the fragments used in the analysis are shown for illustrative purposes.
VOL. 44, 2006 BACILLUS CEREUS ASSOCIATED WITH FATAL PNEUMONIAS 3357
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from

within the resolution of the method, the isolates were vir-tually identical. In contrast, Fig. 2B shows the B. cereus03BB102 profile. There were distinct differences betweenthis profile and the two profiles shown in Fig. 2A. The AFLPprofile of B. cereus 03BB102 from the Texas welder, wassimilar to but distinct from the profile for B. cereus03BB108, which was collected from the welder’s work envi-ronment (Fig. 1).
In addition to AFLP, we applied a MLST scheme that wasoriginally used to subtype B. cereus G9241 (13) to subtypethe three Texas isolates. This method compares partial nu-cleotide sequences of seven housekeeping gene sequences(glpF, gmk, ilvD, pta, pur, pycA, and tpi) (25). B. cereus03BB87 and G9241 were indistinguishable by this MLSTmethod (Fig. 3) (MLST type 78). B. cereus 03BB102 and03BB108 were closely related to each other and more closelyrelated to B. anthracis than were B. cereus 03BB87 andG9241, in agreement with the AFLP analyses (compare re-sults shown in Fig. 1 and 3). B. cereus 03BB102 and 03BB108were also closely related to another clinical B. cereus isolate(R_3039/03) in the MLST database (MLST genotype 75),originally collected in England (Fig. 3).
DISCUSSION
B. cereus isolates do not frequently cause pneumonia orother serious diseases. However, there are several reports ofsevere and often fatal pneumonia in welders caused by B.cereus (1, 2, 14, 20). The genome sequence of B. cereus G9241,
isolated from a Louisiana welder, shares significant homologywith B. anthracis genome sequences and harbors almost theentire pXO1 anthrax virulence plasmid. Here we report thecharacterization of three B. cereus isolates associated with twofatal pneumonia cases that occurred in 2003 within a 3-weekperiod in Texas. The two victims were from geographicallydistinct locations in Texas, but both were metal workers (awelder and a muller operator). Although it is highly unusualfor two fatal cases of B. cereus pneumonia to occur in Texas,both in metal workers and within a few weeks of each other,there was no identified epidemiological link between the cases(S. B. Avashia, submitted for publication). The molecularmethods employed here to characterize the isolates also showthat the two Texas clinical isolates are clearly distinct from oneanother and thus support the epidemiological assessment.However, B. cereus 03BB87, isolated from the muller operator,is indistinguishable from B. cereus G9241, isolated in 1994 froma Louisiana welder, by all molecular methods applied to theisolates in this study. The only difference detected betweenthese two isolates was variation in the degree of capsuleexpression. PCR analysis suggests that B. cereus 03BB87,like B. cereus G9241, carries the pBC218 operon, which isspeculated to be responsible for capsule formation in thelatter isolate (13). Further studies of these two isolates willbe required to understand differences in capsule expressionin these pathogens.
B. cereus 03BB102 and 03BB108, from the Texas welderand his workplace, respectively, are distinctly different from
FIG. 3. Multiple-locus sequence typing analysis of B. cereus 03BB87, B. cereus G9241, B. cereus 03BB102, and B. cereus 03BB108 in comparisonto other Bacillus spp. isolates. The tree was generated using neighbor-joining Kimura two-parameter software (MEGA, version 2.1). The numbersat the end of each branch refer to the sequence type as defined by Priest, et al. (25). The Texas isolates and G9241 are shown with bold arrowsindicating their respective sequence types. Strains labeled “B. cereus/B. thuringiensis” were flagellar (H) antigen serotyped (commonly used to typeB. thuringiensis); however, these strains have not been shown to produce parasporal crystal proteins and, thus, cannot be definitively differentiatedfrom B. cereus.
3358 HOFFMASTER ET AL. J. CLIN. MICROBIOL.
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from

B. cereus 03BB87, collected from the Texas muller operator,based on AFLP and MLST analyses (Fig. 1, 2, and 3). Thesetwo isolates are similar to, but distinct from, one another.AFLP analysis places them in a small subbranch of the phylo-genetic tree (Fig. 1). MLST results (Fig. 3) are consistent withthis finding, showing the two isolates to be similar but dis-tinct. PCR analysis (Table 2) also reveals differences in theirpXO1 gene profiles. The isolates also manifest differentcolony morphologies. It is clear from these results that B.cereus 03BB108 was not derived from B. cereus 03BB102 bya simple loss of plasmid DNA material since the two isolatesare genetically distinct from one another beyond this level.A comparison of AFLP and MLST profiles for B. cereus03BB102 and 03BB108 to those generated from B. anthracisand other B. cereus and B. thuringiensis isolates (Fig. 1 and3) demonstrates that, while these isolates may contain fewersequences homologous to B. anthracis pXO1 and pXO2sequences, they are phylogenetically more similar to B. an-thracis than are B. cereus 03BB87 and G9241. They are alsoclosely related to two other pathogenic Bacillus isolates (B.thuringiensis 97-27 and B. cereus E33L) and two toxigenic B.cereus isolates (D17 and 3a) (Fig. 1).
The presence of the capA, B, and C genes in B. cereus03BB102 and 03BB108 suggests that these genes may play arole in virulence as they do in B. anthracis. However, we didnot observe the expression of the poly-�-D-glutamic acidcapsule in these isolates under in vitro conditions where it isnormally expressed in B. anthracis and we do not yet knowwhether the entire genes are present or under what condi-tions they might be expressed. The presence of the cap genesequences in the absence of any other detectable pXO2sequences is unique relative to other isolates so far charac-terized. Most of the differences in the detection of pXO1genes between these isolates are within the pathogenicityisland (22).
Despite the fact that B. anthracis toxin genes are found inseveral of the B. cereus isolates associated with severe pneu-monia in metal workers, we do not know whether these genesare appropriately expressed or whether any expressed toxinsare functional or play a role in virulence. At least one suchisolate, described by Miller et al. (20), did not contain the pagAgene yet still caused a fatal infection in a previously healthyindividual (13). However, relative to the general population,professional exposure to a variety of environmental factorsmay place welders at additional risk of acquiring such infec-tions. Reports suggest welders experience higher frequenciesof pulmonary infections with increased severity and duration(1, 2). The majority of data suggest that while welders may notbe at increased risk for infections overall, certain conditionsand practices may impact susceptibility to pneumonias (1, 4, 7).It has also been suggested that increased exposure to thesetypes of B. cereus isolates and other respiratory pathogens mayplay a role in increased pneumonia in foundry workers, includ-ing welders (5, 17).
All of the isolates clustered in a branch of the AFLP-based phylogenetic tree occupied by B. anthracis and severalother pathogenic B. cereus and B. thuringiensis isolates so farcharacterized (Fig. 1). It is also intriguing that they werecollected in a relatively small geographic area of the UnitedStates (Texas and Louisiana) and that this area coincides
with regions where anthrax occurs naturally in herbivores. Ifthese isolates acquired these plasmids or plasmid sequencesvia horizontal transfer from B. anthracis in the environment,it is tempting to speculate that such isolates may be re-stricted to or at least be more common in areas whereanthrax is endemic.
B. cereus is ubiquitous in the environment, which can makeit difficult to link clinical cases to their environmental sources.Although the source of the clinical isolate was not identifiedfor either case, the isolation of 03BB108 from the welder’sworksite illustrates that closely related B. cereus isolates werepresent. Increased sampling and analysis of isolates may haveallowed for the detection of isolates in the environment thatmatched the clinical isolate. It is also possible that the sourceof infection was present in material to which both victims wereoccupationally exposed because they shared similar work en-vironments. This scenario would be more likely if the samestrain infected both Texas victims. However, B. cereus 03BB87,isolated from the Texas muller operator, is very similar oridentical to B. cereus G9241, isolated from the Louisianawelder 10 years earlier. Therefore, the possibility of a commonsource is very intriguing, even though these cases were sepa-rated temporally by 10 years.
Taken together with previous reports, these results clearlydemonstrate the need for increased awareness of the potentialfor B. cereus to cause serious systemic infections, even in pa-tients who appear to be otherwise healthy. They also demon-strate the need for a better understanding of environmentaland occupational risk factors that may increase the suscepti-bility of individuals to infection by B. cereus.
ACKNOWLEDGMENTS
The work described in the manuscript (review number UCRL-JRNL-219753) was partially performed under the auspices of the U.S.Department of Energy by Los Alamos National Laboratory undercontract W-7405-ENG-36 and Lawrence Livermore National Labora-tory under contract W-7405-ENG-48.
Funding for a portion of this work was provided by the United StatesDepartment of Homeland Security.
REFERENCES
1. Antonini, J. M., A. B. Lewi, J. R. Roberts, and D. A. Whaley. 2003. Pulmo-nary effects of welding fumes: review of worker and experimental animalstudies. Am. J. Ind. Med. 43:350–360.
2. Antonini, J. M., M. D. Taylor, L. Millecchia, A. R. Bebout, and J. R. Roberts.2004. Suppression in lung defense responses after bacterial infection in ratspretreated with different welding fumes. Toxicol. Appl. Pharmacol. 200:206–218.
3. Brown, E. R., and W. B. Cherry. 1955. Specific identification of Bacillusanthracis by means of a variant bacteriophage. J. Infect. Dis. 96:34–39.
4. Coggon, D., H. Inskip, P. Winter, and B. Pannett. 1994. Lobar pneumonia:an occupational disease in welders. Lancet 344:4–5.
5. Cordes, L. G., E. W. Brink, P. J. Checko, A. Lentnek, R. W. Lyons, P. S.Hayes, T. C. Wu, D. G. Tharr, and D. W. Fraser. 1981. A cluster of Acineto-bacter pneumonia in foundry workers. Ann. Intern. Med. 95:688–693.
6. De, B. K., S. L. Bragg, G. N. Sanden, K. E Wilson, L. A. Diem, C. K. Marston,A. R. Hoffmaster, G. A. Barnett, R. S. Weyant, T. G. Abshire, J. W. Ezzell,and T. Popovic. 2002. A two-component direct fluorescent-antibody assay forrapid identification of Bacillus anthracis. Emerg. Infect. Dis. 8:1060–1065.
7. Doig, A. T., and P. J. Challen. 1964. Respiratory hazards in welding. Ann.Occup. Hyg. 111:223–231.
8. Helgason, E., D. A. Caugant, I. Olsen, and A.-B. Kolstø. 2000. Geneticstructure of population of Bacillus cereus and B. thuringiensis isolates asso-ciated with periodontitis and other human infections. J. Clin. Microbiol.38:1615–1622.
9. Helgason, E., O. A. Økstad, D. A. Caugant, H. A. Johansen, A. Fouet, M.Mock, I. Hegna, and A.-B. Kolstø. 2000. Bacillus anthracis, Bacillus cereus,and Bacillus thuringiensis—one species on the basis of genetic evidence.Appl. Environ. Microbiol. 66:2627–2630.
VOL. 44, 2006 BACILLUS CEREUS ASSOCIATED WITH FATAL PNEUMONIAS 3359
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from

10. Helgason, E., N. J. Tourasse, R. Meisal, D. A. Caugant, and A.-B. Kolsto.2004. Multilocus sequence typing scheme for bacteria of the Bacillus cereusgroup. Appl. Environ. Microbiol. 70:191–201.
11. Hill, K. K., L. O. Ticknor, R. T. Okinaka, M. Asay, H. Blair, K. A. Bliss, M.Laker, P. E. Pardington, A. P. Richardson, M. Tonks, D. J. Beecher, J. D.Kemp, A.-B. Kolsto, A. C. Lee Wong, P. Keim, and P. J. Jackson. 2004.Fluorescent amplified fragment length polymorphism analysis of Bacillusanthracis, Bacillus cereus, and Bacillus thuringiensis isolates. Appl. Environ.Microbiol. 70:1068–1080.
12. Hoffmaster, A. R., C. C. Fitzgerald, E. Ribot, L. W. Mayer, and T. Popovic.2002. Molecular subtyping of Bacillus anthracis and the 2001 bioterrorism-associated anthrax outbreak, United States. Emerg. Infect. Dis. 8:1111–1116.
13. Hoffmaster, A. R., J. Ravel, D. A. Rasko, G. D. Chapman, M. D. Chute, C. K.Marston, B. K. De, C. T. Sacchi, C. Fitzgerald, L. W. Mayer, M. C. J.Maiden, F. G. Priest, M. Barker, L. Jiang, R. Z. Cer, J. Rilsone, S. N.Peterson, R. S. Weyant, D. R. Galloway, T. D. Read, T. Popovic, and C. M.Fraser. 2004. Identification of anthrax toxin genes in a Bacillus cereus asso-ciated with an illness resembling inhalation anthrax. Proc. Natl. Acad. Sci.USA 101:8449–8454.
14. Jackson, P. J., K. K. Hill, M. T. Laker, L. O. Ticknor, and P. Keim. 1999.Genetic comparison of Bacillus anthracis and its close relatives using ampli-fied fragment length polymorphism and polymerase chain reaction analysis.J. Appl. Microbiol. 87:263–269.
15. Keim, P., A. Kalif, J. Schupp, K. Hill, S. E. Travis, K. Richmond, D. M.Adair, M. Hugh-Jones, C. R. Kuske, and P. Jackson. 1997. Molecular evo-lution and diversity in Bacillus anthracis as detected by amplified fragmentlength polymorphism markers. J. Bacteriol. 179:818–824.
16. Reference deleted.17. Kennedy, S. M. 1994. When is a disease occupational? Lancet 344:41–43.
18. Kumar, S., K. Tamura, I. B. Jakobsen, and M. Nei. 2001. MEGA2: molec-ular evolutionary genetics analysis software. Bioinformatics 17:1244–1255.
19. Logan, N. A., and P. C. Turnbull. 1999. Bacillus and recently derived genera,p. 357–369. In P. R. Murray (ed.), Manual of clinical microbiology. AmericanSociety for Microbiology, Washington, D.C.
20. Miller, J. M., J. G. Hair, M. Hebert, L. Hebert, F. J. Roberts, Jr., and R. S.Weyant. 1997. Fulminating bacteremia and pneumonia due to Bacilluscereus. J. Clin. Microbiol. 35:504–507.
21. Mock, M., and T. Mignot. 2003. Anthrax toxins and the host: a story ofintimacy. Cell. Microbiol. 5:15–23.
22. Okinaka, R. T., K. Cloud, O. Hampton, A. R. Hoffmaster, K. K. Hill, P.Keim, T. M. Koehler, G. Lamke, S. Kumano. J. Mahillon, D. Manter, Y.Martinez, D. Ricke, R. Svensson, and P. J. Jackson. 1999. Sequence andorganization of pXO1, the large Bacillus anthracis plasmid harboring theanthrax toxin genes. J. Bacteriol. 181:6509–6515.
23. Pannucci, J., R. T. Okinaka, R. Sabin, and C. R. Kuske. 2002. Bacillusanthracis pXO1 plasmid sequence conservation among closely related bac-terial species. J. Bacteriol. 184:134–141.
24. Price, L. B., M. Hugh-Jones, P. J. Jackson, and P. Keim. 1999. Geneticdiversity in the protective antigen gene of Bacillus anthracis. J. Bacteriol.181:2358–2362.
25. Priest, F. G., M. Barker, L. W. Baillie, E. C. Holmes, and M. C. Maiden.2004. Population structure and evolution of the Bacillus cereus group. J.Bacteriol. 186:7959–7970.
26. Ticknor, L. O., A.-B. Kolstø, K. K. Hill, P. Keim, M. T. Laker, M. Tonks, andP. J. Jackson. 2001. Fluorescent amplified fragment length polymorphismanalysis of Norwegian Bacillus cereus and Bacillus thuringiensis soil isolates.Appl. Environ. Microbiol. 67:4863–4873.
3360 HOFFMASTER ET AL. J. CLIN. MICROBIOL.
on March 17, 2014 by guest
http://jcm.asm
.org/D
ownloaded from




















