
Characterization of a Cu/Zn Superoxide dismutase-encoding gene region in Drosophila willistoni
14
Molecular Microbiology (2003) 47 (6), 1681–1694 © 2003 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 200347 616811694 Original Article Cn var. gatti sod1S. D. Narasipura et al. Accepted 26 November, 2002. *For correspondence. E-mail [email protected]; Tel. ( + 1) 518 474 7563; Fax ( + 1) 518 486 7971. Characterization of Cu,Zn superoxide dismutase ( SOD1 ) gene knock-out mutant of Cryptococcus neoformans var . gattii : role in biology and virulence Srinivas D. Narasipura, 1 Jeffrey G. Ault, 2 Melissa J. Behr, 3 Vishnu Chaturvedi 1,4 and Sudha Chaturvedi 1 * 1 Mycology Laboratory, 2 Electron Microscopy Core, 3 Anatomic and Pathology Laboratory, Wadsworth Center, New York State Department of Health, 120 New Scotland Ave., Albany, NY 12208-2002, USA. 4 Department of Biomedical Sciences, School of Public Health, State University of New York, Albany, NY, USA. Summary The pathogenic yeast Cryptococcus neoformans ( Cn ) var. gattii causes meningoencephalitis in healthy indi- viduals, unlike the better known Cn varieties grubii and neoformans , which are common in immunocom- promised individuals. The virulence determinants and mechanisms of host predilection are poorly defined for var. gattii . The present study focused on the char- acterization of a Cu,Zn superoxide dismutase ( SOD1 ) gene knock-out mutant constructed by developing a DNA transformation system. The sod1 mutant was highly sensitive to the redox cycling agent menadi- one, and showed fragmentation of the large vacuole in the cytoplasm, but no other defects were seen in growth, capsule synthesis, mating, sporulation, sta- tionary phase survival or auxotrophies for sulphur- containing amino acids. The sod1 mutant was mark- edly attenuated in virulence in a mouse model, and it was significantly susceptible to in vitro killing by human neutrophils (PMNs). The deletion of SOD1 also resulted in defects in the expression of a number of virulence factors, i.e. laccase, urease and phospholi- pase. Complementation of the sod1 mutant with SOD1 resulted in recovery of virulence factor expres- sion and menadione resistance, and in restoration of virulence. Overall, these results suggest that the anti- oxidant function of Cu,Zn SOD is critical for the patho- genesis of the fungus, but is dispensable in its saprobic life. This report constitutes the first instance in which superoxide dismutase has been directly implicated in the virulence of a fungal pathogen. Introduction Cryptococcus neoformans ( Cn ) var. gattii is an encapsu- lated pathogenic yeast found in tropical climates, most commonly on Eucalyptus trees. Cn var. gattii differs from two other well-known Cn varieties, grubii and neoformans , in phenotypic characters, natural habitat, epidemiology, clinical manifestations of disease and response to antifun- gal therapy (Kwon-Chung and Bennett, 1992; Fisher et al ., 1993; Speed and Dunt, 1995; Casadevall and Per- fect, 1998; Chen et al ., 2000; Sorrell, 2001). Cn var. gattii predominantly infects immunocompetent individuals, whereas varieties grubii and neoformans are common in immunocompromised individuals. The mechanism of dif- ferences in host predilection remain largely unknown, except for two experimental studies that reported that Cn var. gattii inhibits phagocyte response, whereas the other two varieties are readily killed by phagocyte oxidants (Church and Washburn, 1992; Dong and Murphy, 1995). It was hypothesized that this differential phagocyte response could result from innate differences among the antioxidants of these varieties. Support for this observa- tion comes from our earlier work on comparative analysis of Cu,Zn superoxide dismutase (SOD; encoded by SOD1 ) from three Cn varieties, which revealed significant differ- ences in the physical, biochemical, molecular and struc- tural properties of this enzyme (Hamilton and Holdom, 1997; Chaturvedi et al ., 2001). These results are impor- tant, as SOD is one of the essential elements of the primary antioxidant defence system. SOD protects cells from the toxic effects of reactive oxygen intermediates (ROIs) by dismutation of superoxide radicals (O 2 – ) into hydrogen peroxide (H 2 O 2 ) and oxygen (O 2 ), thereby pre- venting O 2 – -mediated reduction of iron and subsequent generation of highly toxic hydroxyl radicals (OH • ; Miller and Britigan, 1997). Cu,Zn SOD has been extensively investigated in the eukaryotic model organism Saccharomyces cerevisiae . Yeast lacking Cu,Zn SOD shows severe growth defects, auxotrophies for sulphur-containing amino acids, a higher rate of spontaneous mutation under aerobic conditions and rapid loss of viability under nutrient deprivation (Gralla
Transcript of Characterization of a Cu/Zn Superoxide dismutase-encoding gene region in Drosophila willistoni

Molecular Microbiology (2003)
47
(6) 1681ndash1694
copy 2003 Blackwell Publishing Ltd
Blackwell Science LtdOxford UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd 200347
616811694
Original Article
Cn var gatti sod1S D Narasipura et al
Accepted 26 November 2002 For correspondence E-mailschaturvwadsworthorg Tel (
+
1) 518 474 7563 Fax (
+
1) 518 4867971
Characterization of CuZn superoxide dismutase (
SOD1
) gene knock-out mutant of
Cryptococcus neoformans
var
gattii
role in biology and virulence
Srinivas D Narasipura
1
Jeffrey G Ault
2
Melissa J Behr
3
Vishnu Chaturvedi
14
andSudha Chaturvedi
1
1
Mycology Laboratory
2
Electron Microscopy Core
3
Anatomic and
Pathology Laboratory Wadsworth Center New York State Department of Health 120 New Scotland Ave Albany NY 12208-2002 USA
4
Department of Biomedical Sciences School of Public Health State University of New York Albany NY USA
Summary
The pathogenic yeast
Cryptococcus neoformans
(
Cn
)var
gattii
causes meningoencephalitis in healthy indi-viduals unlike the better known
Cn
varieties
grubii
and
neoformans
which are common in immunocom-promised individuals The virulence determinants andmechanisms of host predilection are poorly definedfor var
gattii
The present study focused on the char-acterization of a CuZn superoxide dismutase (
SOD1
)gene knock-out mutant constructed by developing aDNA transformation system The
sod1
mutant washighly sensitive to the redox cycling agent menadi-one and showed fragmentation of the large vacuolein the cytoplasm but no other defects were seen ingrowth capsule synthesis mating sporulation sta-tionary phase survival or auxotrophies for sulphur-containing amino acids The
sod1
mutant was mark-edly attenuated in virulence in a mouse model and itwas significantly susceptible to
in vitro
killing byhuman neutrophils (PMNs) The deletion of
SOD1
alsoresulted in defects in the expression of a number ofvirulence factors ie laccase urease and phospholi-pase Complementation of the
sod1
mutant with
SOD1
resulted in recovery of virulence factor expres-sion and menadione resistance and in restoration ofvirulence Overall these results suggest that the anti-oxidant function of CuZn SOD is critical for the patho-genesis of the fungus but is dispensable in itssaprobic life This report constitutes the first instance
in which superoxide dismutase has been directlyimplicated in the virulence of a fungal pathogen
Introduction
Cryptococcus neoformans
(
Cn
) var
gattii
is an encapsu-lated pathogenic yeast found in tropical climates mostcommonly on
Eucalyptus
trees
Cn
var
gattii
differs fromtwo other well-known
Cn
varieties
grubii
and
neoformans
in phenotypic characters natural habitat epidemiologyclinical manifestations of disease and response to antifun-gal therapy (Kwon-Chung and Bennett 1992 Fisher
et al
1993 Speed and Dunt 1995 Casadevall and Per-fect 1998 Chen
et al
2000 Sorrell 2001)
Cn
var
gattii
predominantly infects immunocompetent individualswhereas varieties
grubii
and
neoformans
are common inimmunocompromised individuals The mechanism of dif-ferences in host predilection remain largely unknownexcept for two experimental studies that reported that
Cn
var
gattii
inhibits phagocyte response whereas the othertwo varieties are readily killed by phagocyte oxidants(Church and Washburn 1992 Dong and Murphy 1995)It was hypothesized that this differential phagocyteresponse could result from innate differences among theantioxidants of these varieties Support for this observa-tion comes from our earlier work on comparative analysisof CuZn superoxide dismutase (SOD encoded by
SOD1
)from three
Cn
varieties which revealed significant differ-ences in the physical biochemical molecular and struc-tural properties of this enzyme (Hamilton and Holdom1997 Chaturvedi
et al
2001) These results are impor-tant as SOD is one of the essential elements of theprimary antioxidant defence system SOD protects cellsfrom the toxic effects of reactive oxygen intermediates(ROIs) by dismutation of superoxide radicals (O
2ndash
) intohydrogen peroxide (H
2
O
2
) and oxygen (O
2
) thereby pre-venting O
2ndash
-mediated reduction of iron and subsequentgeneration of highly toxic hydroxyl radicals (OH
bull
Millerand Britigan 1997)
CuZn SOD has been extensively investigated in theeukaryotic model organism
Saccharomyces cerevisiae
Yeast lacking CuZn SOD shows severe growth defectsauxotrophies for sulphur-containing amino acids a higherrate of spontaneous mutation under aerobic conditionsand rapid loss of viability under nutrient deprivation (Gralla
Used Mac Distiller 50x Job Options
This report was created automatically with help of the Adobe Acrobat Distiller addition Distiller Secrets v105 from IMPRESSED GmbH13You can download this startup file for Distiller versions 405 and 50x for free from httpwwwimpressedde1313GENERAL ----------------------------------------13File Options13 Compatibility PDF 1213 Optimize For Fast Web View Yes13 Embed Thumbnails Yes13 Auto-Rotate Pages No13 Distill From Page 113 Distill To Page All Pages13 Binding Left13 Resolution [ 600 600 ] dpi13 Paper Size [ 595 842 ] Point1313COMPRESSION ----------------------------------------13Color Images13 Downsampling Yes13 Downsample Type Average Downsampling13 Downsample Resolution 120 dpi13 Downsampling For Images Above 180 dpi13 Compression Yes13 Compression Type JPEG13 JPEG Quality Medium13 Bits Per Pixel As Original Bit13Grayscale Images13 Downsampling Yes13 Downsample Type Average Downsampling13 Downsample Resolution 120 dpi13 Downsampling For Images Above 180 dpi13 Compression Yes13 Compression Type JPEG13 JPEG Quality Medium13 Bits Per Pixel As Original Bit13Monochrome Images13 Downsampling Yes13 Downsample Type Average Downsampling13 Downsample Resolution 600 dpi13 Downsampling For Images Above 900 dpi13 Compression Yes13 Compression Type ZIP13 Anti-Alias To Gray No1313 Compress Text and Line Art Yes1313FONTS ----------------------------------------13 Embed All Fonts Yes13 Subset Embedded Fonts Yes13 Subset When Percent Of Characters Used is Less 100 13 When Embedding Fails Warn and Continue13Embedding13 Always Embed [ Courier-BoldOblique Helvetica-BoldOblique Courier Helvetica-Bold Times-Bold Courier-Bold Helvetica Times-BoldItalic Times-Roman ZapfDingbats Times-Italic Helvetica-Oblique Courier-Oblique Symbol ]13 Never Embed [ ]1313COLOR ----------------------------------------13Color Management Policies13 Color Conversion Strategy Leave Color Unchanged13 Intent Default13Device-Dependent Data13 Preserve Overprint Settings Yes13 Preserve Under Color Removal and Black Generation No13 Transfer Functions Preserve13 Preserve Halftone Information Yes1313ADVANCED ----------------------------------------13Options13 Use Prologueps and Epilogueps No13 Allow PostScript File To Override Job Options Yes13 Preserve Level 2 copypage Semantics Yes13 Save Portable Job Ticket Inside PDF File Yes13 Illustrator Overprint Mode No13 Convert Gradients To Smooth Shades No13 ASCII Format No13Document Structuring Conventions (DSC)13 Process DSC Comments Yes13 Log DSC Warnings No13 Resize Page and Center Artwork for EPS Files No13 Preserve EPS Information From DSC Yes13 Preserve OPI Comments No13 Preserve Document Information From DSC Yes1313OTHERS ----------------------------------------13 Distiller Core Version 500013 Use ZIP Compression Yes13 Deactivate Optimization No13 Image Memory 524288 Byte13 Anti-Alias Color Images No13 Anti-Alias Grayscale Images No13 Convert Images (lt 257 Colors) To Indexed Color Space Yes13 sRGB ICC Profile sRGB IEC61966-211313END OF REPORT ----------------------------------------1313IMPRESSED GmbH13Bahrenfelder Chaussee 491322761 Hamburg Germany13Tel +49 40 897189-013Fax +49 40 897189-7113Email infoimpressedde13Web wwwimpressedde
Adobe Acrobat Distiller 50x Job Option File
ltlt13 ColorSettingsFile ()13 LockDistillerParams false13 DetectBlends false13 DoThumbnails true13 AntiAliasMonoImages false13 MonoImageDownsampleType Average13 GrayImageDownsampleType Average13 MaxSubsetPct 10013 MonoImageFilter FlateEncode13 ColorImageDownsampleThreshold 1513 GrayImageFilter DCTEncode13 ColorConversionStrategy LeaveColorUnchanged13 CalGrayProfile (None)13 ColorImageResolution 12013 UsePrologue false13 MonoImageResolution 60013 ColorImageDepth -113 sRGBProfile (sRGB IEC61966-21)13 PreserveOverprintSettings true13 CompatibilityLevel 1213 UCRandBGInfo Remove13 EmitDSCWarnings false13 CreateJobTicket true13 DownsampleMonoImages true13 DownsampleColorImages true13 MonoImageDict ltlt K -1 gtgt13 ColorImageDownsampleType Average13 GrayImageDict ltlt VSamples [ 2 1 1 2 ] HSamples [ 2 1 1 2 ] Blend 1 QFactor 076 ColorTransform 1 gtgt13 CalCMYKProfile (US Web Coated (SWOP) v2)13 ParseDSCComments true13 PreserveEPSInfo true13 MonoImageDepth 113 AutoFilterGrayImages false13 SubsetFonts true13 GrayACSImageDict ltlt HSamples [ 2 1 1 2 ] VSamples [ 2 1 1 2 ] Blend 1 QFactor 09 gtgt13 ColorImageFilter DCTEncode13 AutoRotatePages None13 PreserveCopyPage true13 EncodeMonoImages true13 ASCII85EncodePages false13 PreserveOPIComments false13 NeverEmbed [ ]13 ColorImageDict ltlt VSamples [ 2 1 1 2 ] HSamples [ 2 1 1 2 ] Blend 1 QFactor 076 ColorTransform 1 gtgt13 AntiAliasGrayImages false13 GrayImageDepth -113 CannotEmbedFontPolicy Warning13 EndPage -113 TransferFunctionInfo Preserve13 CalRGBProfile (sRGB IEC61966-21)13 EncodeColorImages true13 EncodeGrayImages true13 ColorACSImageDict ltlt HSamples [ 2 1 1 2 ] VSamples [ 2 1 1 2 ] Blend 1 QFactor 09 gtgt13 Optimize true13 ParseDSCCommentsForDocInfo true13 GrayImageDownsampleThreshold 1513 MonoImageDownsampleThreshold 1513 AutoPositionEPSFiles false13 GrayImageResolution 12013 AutoFilterColorImages false13 AlwaysEmbed [ Courier-BoldOblique Helvetica-BoldOblique Courier Helvetica-Bold Times-Bold Courier-Bold Helvetica Times-BoldItalic Times-Roman ZapfDingbats Times-Italic Helvetica-Oblique Courier-Oblique Symbol ]13 ImageMemory 52428813 OPM 013 DefaultRenderingIntent Default13 EmbedAllFonts true13 StartPage 113 DownsampleGrayImages true13 AntiAliasColorImages false13 ConvertImagesToIndexed true13 PreserveHalftoneInfo true13 CompressPages true13 Binding Left13gtgt setdistillerparams13ltlt13 PageSize [ 5760 7920 ]13 HWResolution [ 600 600 ]13gtgt setpagedevice
1682
S D Narasipura
et al
copy 2003 Blackwell Publishing Ltd
Molecular Microbiology
47
1681ndash1694
and Valentine 1991 Liu
et al
1992 Longo
et al
1996)CuZn SOD has also been isolated from other fungiincluding
Neurospora crassa
Schizosaccharomycespombe
Candida albicans
and
Aspergillus fumigatus
(Goscin and Fridovich 1972 Misra and Fridovich 1972Hwang
et al
1999 Mutoh
et al
2002) The associationof
A fumigatus
CuZn SOD with the cell wall and itssecretion into the medium suggest that this enzyme mayprotect cells from externally generated free radicals(Hamilton
et al
1996) Significant inhibition in humanneutrophil (PMN)-induced killing of
Cn
by exogenousaddition of SOD also suggests a protective role for thisenzyme (Chaturvedi
et al
1996) However none of thedata described above provide direct evidence for any rolefor CuZn SOD in fungal virulence
Virulence determinants of
Cn
are currently the subjectof intensive investigation in
Cn
varieties
neoformans
and
grubii
Several important virulence traits were recognizedearly by biochemical and classical genetics includingpolysaccharide capsule laccase production and ability togrow at 37
infin
C With the availability of DNA transformationsystems since 1992 many more factors contributing tovirulence in these two varieties have been identified ie
NMT1
CNA1
GPA1
URE1
and phospholipase (Lodge
et al
1994 Alspaugh
et al
1997 Odom
et al
1997 Cox
et al
2000 2001) These molecular pathogenesis studieshave also revealed previously unknown differences in vir-ulence for var
grubii
and var
neoformans
For examplevar
grubii
transcription factor
STE12
a
was shown to beessential for haploid fruiting but not necessary for matingand virulence whereas
STE12
a
in var
neoformans
wasfound to be essential for virulence and haploid fruiting butdispensable for mating (Yue
et al
1999 Chang
et al
2000) Similarly calcineurin was essential for cation resis-tance in var
grubii
although it was not essential for cationresistance in
Cn
var
neoformans
suggesting a divergentrole for this gene in these two varieties (Cruz
et al
2000)There are no studies yet on mechanisms of virulence in
Cn
var
gattii
presumably because an efficient DNA trans-formation and gene disruption system has not yet beendescribed The present study describes the developmentof a transformation system and the successful disruptionof CuZn SOD (
SOD1
) in
Cn
var
gattii
Results
Disruption and reintroduction of the
SOD1
gene in
Cn
var
gattii
The
SOD1
gene was disrupted by replacing 907 bp of the957 bp open reading frame (ORF) of the gene with the
URA5
selectable marker which resulted in the CuZnSOD null mutant
sod1URA5
(Fig 1A) Our initialattempts to use the
URA5
selectable marker from
Cn
var
neoformans
for transformation of var
gattii
were unsuc-
cessful The comparative analysis of
URA5 from Cn varneoformans and var gattii revealed that this gene is ordf9divergent at the intragenic region and 30 divergent atthe promoter region These results suggested that URA5from Cn var neoformans might not be functional in vargattii Therefore to increase the transformation efficiencyas well as the homologous recombination we used URA5and SOD1 from Cn var gattii type strain NIH 444 A NotI-linearized sod1URA5 disruption allele was transformedinto a ura5 strain of var gattii Sixty transformants wereobtained on synthetic medium lacking uracil Thesetransformants were analysed initially for a presumptive no-growth phenotype on YPD menadione broth This screen-ing procedure yielded four suspected sod1 clones Nextgenomic DNA was isolated from these clones and analy-sed by polymerase chain reaction (PCR) for the absenceof the 500 bp fragment of the SOD1 ORF using primerpair V523 and V524 (details in Experimental procedures)If the sod1URA5 allele has been integrated at the homol-ogous site then the primer pair V523 and V524 shouldnot give the 500 bp product however if it is integrated atthe ectopic site then the genomic DNA should give one500 bp product of SOD1 wild type Out of four isolatestwo lacked the SOD1 wild-type locus as revealed by theabsence of 500 bp amplicons (Fig 1B) Southern hybrid-ization confirmed that the SOD1 wild-type locus had beenreplaced by the sod1URA5 disruption allele by homolo-gous recombination as no signal for SOD1 was detectedin the mutant strain when it was probed with the 500 bpfragment of SOD1 used from the 907 bp deleted regionof wild-type SOD1 (Fig 1C) The percentage homologousrecombination achieved was 34 (two positive out of 60transformants)
Next we generated a reconstituted strain in which awild-type copy of SOD1 with its own promoter was reinte-grated in the genome To make a reconstituted strain wereverted the ura+ phenotype by plating the sod1URA5mutant onto medium with 5-fluoroorotic acid (5-FOA) toselect for urandash cells This revertant was used as the recip-ient for transformation with a pCR61 plasmid containingfull-length SOD1 (28 kb) and URA5 selectable markerPCR analysis confirmed the restoration of the wild-typegene in these two reconstituted strains Interestinglyrestriction digestion of genomic DNA with XbaI and BstXIproduced an identical band size to that of the wild-typestrain which suggested homologous integration of SOD1to the sod1URA5 locus These reconstituted strains weredesignated as sod1 + SOD1-A and B
To examine the expression of the SOD1 gene we per-formed reverse transcription (RT)-PCR on total RNA iso-lated from SOD1 wild type the sod1 mutant and thesod1 + SOD1 reconstituted strains In contrast to theSOD1 wild-type strain in which a 400 bp amplicon wasreadily detected no message was detected in the sod1
Cn var gatti sod1 1683
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant strain This result supports the conclusion thatSOD1 has been functionally deleted in the sod1 mutantstrain After reintroduction of the SOD1 wild-type geneby transformation SOD1 expression was restored in thesod1 + SOD1 strains A and B (Fig 1D) Further geno-typic and phenotypic analyses confirmed that bothreconstituted strains exhibited identical properties there-fore results from sod1 + SOD1-A are shown in all otherexperiments
Characterization of Cn var gattii sod1 phenotypes
To address whether CuZn SOD is essential for maintain-ing normal growth of Cn in laboratory medium we deter-mined the doubling time of exponentially dividing SOD1wild type sod1 mutant and sod1 + SOD1 reconstitutedstrains in YPD broth at both 30infinC and 37infinC The doublingtime of the sod1 mutant was slightly longer at 30infinC and37infinC (33 plusmn 02 h and 34 plusmn 01 h) than that of the SOD1
Fig 1 Disruption and reconstitution of Cn var gattii SOD1 geneA A 28 kb fragment containing the SOD1 gene (solid thick line and dashed line outside 28 kb) indicates the genomic locus based on restriction enzyme digestion The lsquoprobersquo indicates the 500 bp sequence within the ORF used for PCR Southern hybridization and RT-PCR to confirm gene replacementB PCR for SOD1 ORF Primers designed from the SOD1 ORF (indicated by an arrow in A) amplified a 500 bp product from genomic DNA of SOD1 wild-type and sod1 + SOD1 reconstituted strains but not from genomic DNA of the sod1 mutant strain Negative control is PCR without genomic DNAC Southern blot of genomic DNA from the SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains Genomic DNA was digested with XbaI and BstXI and probed with the 500 bp PCR product depicted in (A) No hybridization signal was detected in the sod1 mutant whereas both SOD1 wild-type and sod1 + SOD1 (A and B) reconstituted strains produced asymp8 kb 4 kb and 08 kb bandsD RT-PCR to examine the expression of the SOD1 gene Total RNA was isolated reverse transcribed to cDNA and amplified with primers directed to SOD1 or actin RT-PCR products were fractionated by electrophoresis in a 1 agarose gel and stained with ethidium bromide Positive control is RT-PCR with RNA provided in the kit to confirm the efficacy of the kit and negative control is RT-PCR in the absence of RNA
A
B C D
1684 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
wild type (28 plusmn 01 h and 29 plusmn 02 h) or the sod1 + SOD1reconstituted strain (27 plusmn 03 h and 28 plusmn 01 h) How-ever these differences in growth between the sod1 mutantand SOD1 wild type were not statistically significant(P = 04) These results indicate that CuZn SOD is notessential for Cn growth Similarly CuZn SOD was seennot to be required for Cn growth under nutrient depriva-tion as no difference in growth between the sod1 mutantand the SOD1 wild type was observed when they wereassessed in terms of colony-forming units (cfu) of cellscultured for 15 days in water (Fig 2) An initial drop in cfunoted after 3 days of incubation in water might result froma shift from a very rich medium such as YPD to water andthis drop in cfu was consistent among SOD1 wild-type
sod1 mutant and sod1 + SOD1 reconstituted strains Alsothe sod1 mutant did not show auxotrophies for the sul-phur-containing amino acids lysine and methionine as thegrowth (OD600) determined for the sod1 mutant in com-plete CSM medium or in CSM lacking either lysine ormethionine was similar (data not shown)
It was also interesting to note that CuZn SOD was notrequired for Cn mating The sod1 mutant produced abun-dant filaments and basidiospores within 7 days at 24infinCsimilar to the behaviour of SOD1 wild type when mixedwith var neoformans MATa type strain NIH430 (data notshown) Such successful mating reactions between varneoformans and var gattii tester strains have been reportedin the literature (Kwon-Chung et al 1982) We did notisolate basidiospores from these crossings to test for theviability of the progeny However neither SOD1 wild-typenor sod1 mutant strains produced any basidiospores andfilaments when mixed with var gattii MATa type strainNIH191 Although it is not known why SOD1 wild type andthe sod1 mutant were unable to mate with tester strainNIH191 one possibility is that NIH191 might no longer bemating competent
Sod1 mutant is highly sensitive to menadione
The cytosolic localization of CuZn SOD implies that itsmain function is to protect cells against reactive oxygenintermediates (ROI Jamieson 1998) It is also reasonableto suspect that this localization may also enhance theefficiency of Cn in coping with oxidative stresses in thehost To address this question we grew Cn strains in thepresence of menadione (a redox cycling agent thatgenerates superoxide ions in the cytosol) to an initialOD600 of 01 The sod1 mutant was extremely sensitive tomenadione and its growth was completely inhibited at3 mg ml-1 at 30infinC This effect was more pronounced at37infinC as even a 1 mg ml-1 concentration of menadionewas enough to inhibit the sod1 mutant completely
Fig 2 sod1 mutant is viable in stationary phase culture Cells were allowed to grow in flasks for 48 h in YPD broth washed three times with sterile distilled water and reincubated in water Viability was monitored on the indicated days by plating aliquots onto YPD plates and cfu were counted The experiment was repeated twice with sim-ilar results and a representative experiment is shown
A
Fig 3 sod1 mutant is highly sensitive to mena-dione Cells were grown in YPD broth contain-ing menadione (0ndash3 mg ml-1) at initial OD600 of 01 under shaking conditions (180 rpm) The data shown were acquired 16 h after incuba-tion Experiments were repeated three times with similar results a representative experiment is shown
Cn var gatti sod1 1685
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
whereas the SOD1 wild-type strain was relatively resis-tant Introduction of the SOD1 gene restored the menadi-one resistance of the sod1 mutant to the wild-type levelThese results strongly suggest that cytosolic CuZn SODprotects Cn against oxidative stress (Fig 3)
Sod1 mutant shows fragmented vacuoles
Ultrastructural examination of the sod1 mutant revealedfragmentation of its large vacuole into aberrant shapes(Fig 4) In general the wild-type strain had two kinds ofvacuoles one large loosely packed vacuole (VL) and twoto six small densely packed vacuoles (VS) It was notedthat VL in sod1 mutant was fragmented as the internalcontents of the fragmented vacuole appeared to be similarto that of the VL seen in SOD1 wild type The analysis of30 individual electron microscopic (EM) sections revealedthat gt75 of the sod1 mutant cells had fragmented VL(12 plusmn 46 VL per cell) compared with one intact VL inSOD1 wild type (P lt 00001) Reconstitution of the sod1mutant with SOD1 wild type restored vacuolar morphol-ogy suggesting that CuZn SOD is essential for vacuolarintegrity and possibly for vacuolar function
Sod1 is defective in the production of several virulence factors
We next examined whether SOD1 deletion in Cn influ-ences other known virulence factors The virulence traitstested included production of melanin laccase ureasephospholipase B polysaccharide capsule and mannitolThe sod1 mutant exhibited a marked defect in melaniza-tion (Fig 5A) Melanin production was restored in asod1 + SOD1 reconstituted strain As laccase is the soleenzyme responsible for melanin production in Cn (Salas
et al 1996) laccase activity was measured from theequal number of sod1 mutant and SOD1 wild-type cellsas described previously (Zhang et al 1999) SOD1 dele-tion resulted in significant reduction in laccase activity inthe sod1 mutant compared with SOD1 wild type (P lt 001Fig 5B) Similarly the sod1 mutant was severely defectivein the production of phospholipase B on egg yolk agarand in the production of urease on urea agar media(Fig 5C and D) Reconstitution of the sod1 mutant withSOD1 wild type restored all the virulence-associated phe-notypes to a large extent On the other hand SOD1 dele-tion had no influence on capsule synthesis as revealedby India ink stains of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains grown in low-ironmedium for 3 days (Fig 6A) Interestingly the sod1mutant produced significantly more mannitol (P lt 005) inthe medium than the SOD1 wild-type strain (Fig 6B)
Sod1 mutant is attenuated in virulence in the mouse meningoencephalitis model
We tested whether CuZn SOD is required for virulenceof Cn var gattii in a mouse meningoencephalitis modelAs shown in Fig 7 after injection of 1 yen 106 yeast cellsmedian survival of mice inoculated with the SOD1 wild-type strain was 6 days whereas the median survival ofmice inoculated with the sod1 mutant was 23 days(P lt 00001) Reconstitution of the SOD1 gene into thesod1 mutant restored virulence with a median survival ofmice injected with sod1 + SOD1 of 9 days (P lt 00003)These results suggested that CuZn SOD plays an impor-tant role in Cn var gattii pathogenesis
Next we examined the in vivo tissue reaction inducedby Cn strains in infected mice As Cn has a tropism for
Fig 4 sod1 mutant shows fragmented vacuoles Cn cells were grown in YPD broth overnight at 30infinC and then processed for electron microscopic analysis as described in Experimental procedures N nucleus VS small vacuole VL large vacuole arrow points to fragmentation of large vacuole in the sod1 mutant Bar corresponds to 120 nm
1686 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
brain we collected brain tissues from infected mice 7 daysafter infection SOD1 wild type induced severe infectionwith the formation of 17 plusmn 70 lesions per brain section(each brain section comprised rostral cortex caudal cor-tex mid-brain and cerebellum with brainstem) comparedwith the sod1 mutant with 20 plusmn 12 lesions per brain sec-tion (P lt 00001) Infection with the sod1 + SOD1 recon-stituted strain was intermediate in severity resulting in anaverage of 8 plusmn 40 lesions per brain section The numberof yeasts per lesion for SOD1 wild-type sod1 mutant and
sod1 + SOD1 reconstituted strains ranged from 2 to 54 1to 3 and 1 to 37 respectively (Fig 8) These results sug-gested that the sod1 mutant is defective in brain coloni-zation and persistence No lesions were found in sectionsfrom control brain (Fig 8)
Fig 5 Defective production of virulence factors in sod1 mutantA Cells were grown in YPD broth overnight then washed three times with PBS and a 5 ml suspension of 107 cells ml-1 was placed on Niger seed agar for the determination of melanin synthesis and incu-bated at 30infinC for 4 daysB Phenoloxidase activity from an equal number of glucose-starved cells was determined by measuring the oxidation of the diphenolic substrate ABTS (IU of activity = 001 A420 absorbance unit in 30 min) The results are the mean plusmn SD of three individual experiments Aster-isk denotes P pound 005 compared with SOD1 wild typeC and D An aliquot of 5 ml of Cn cells (described in detail in A) were placed on Christensenrsquos agar for the determination of urease synthe-sis and on egg yolk agar for the determination of phospholipase synthesis
A
B
C
D
Fig 6 Capsule and mannitol synthesis in sod1 mutantA Cn cells were grown in low-iron medium for 3 days at 30infinC Cells were mixed with India ink and the capsule which excludes the ink particles was photographed Final magnification yen400B Extracellular mannitol produced from Cn cells was determined by a colorimetric method with absorbance at 412 nm The results are the mean plusmn SD of three individual experiments Asterisk denotes P lt005 compared with SOD1 wild type
A
B
A
B
Fig 7 Virulence of sod1 mutant in a mouse model SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains were grown in YPD broth for 16 h a suspension of 1 yen 106 cells was then injected intravenously into 6-week-old male BALBc mice which were observed daily until moribund
Cn var gatti sod1 1687
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Fig 8 Defective brain colonization and persistence of sod1 mutant Three mice were infected with 1 times 105 Cn cells intravenously then euthanized on day 7 and the brain sections were stained with Mayers mucicarmineA and B Sections from SOD1 wild-type infection reveal a large lesion (arrow) filled with fluid and SOD1 wild-type cells in the thalamus Organisms were also present in smaller numbers in the lateral ventricles and meninges (double arrow) (A at 20times) Higher magnification of the large cyst revealed numerous variable sized SOD1 wild-type cells some with budding (B at 200times)C and D A section from the sod1 mutant infection shows a very small cyst (arrow) (C at 20times) and higher magnification reveals a cyst with one budding sod1 mutant cell (D at 200times)E and F A section from the sod1 + SOD1 reconstituted strain infection reveals one large (arrow) and a few small cysts (double arrow) (E at 20times) and higher magnification of the large cyst reveals many Cn cells (F at 200times)
A
B C D
1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

1682
S D Narasipura
et al
copy 2003 Blackwell Publishing Ltd
Molecular Microbiology
47
1681ndash1694
and Valentine 1991 Liu
et al
1992 Longo
et al
1996)CuZn SOD has also been isolated from other fungiincluding
Neurospora crassa
Schizosaccharomycespombe
Candida albicans
and
Aspergillus fumigatus
(Goscin and Fridovich 1972 Misra and Fridovich 1972Hwang
et al
1999 Mutoh
et al
2002) The associationof
A fumigatus
CuZn SOD with the cell wall and itssecretion into the medium suggest that this enzyme mayprotect cells from externally generated free radicals(Hamilton
et al
1996) Significant inhibition in humanneutrophil (PMN)-induced killing of
Cn
by exogenousaddition of SOD also suggests a protective role for thisenzyme (Chaturvedi
et al
1996) However none of thedata described above provide direct evidence for any rolefor CuZn SOD in fungal virulence
Virulence determinants of
Cn
are currently the subjectof intensive investigation in
Cn
varieties
neoformans
and
grubii
Several important virulence traits were recognizedearly by biochemical and classical genetics includingpolysaccharide capsule laccase production and ability togrow at 37
infin
C With the availability of DNA transformationsystems since 1992 many more factors contributing tovirulence in these two varieties have been identified ie
NMT1
CNA1
GPA1
URE1
and phospholipase (Lodge
et al
1994 Alspaugh
et al
1997 Odom
et al
1997 Cox
et al
2000 2001) These molecular pathogenesis studieshave also revealed previously unknown differences in vir-ulence for var
grubii
and var
neoformans
For examplevar
grubii
transcription factor
STE12
a
was shown to beessential for haploid fruiting but not necessary for matingand virulence whereas
STE12
a
in var
neoformans
wasfound to be essential for virulence and haploid fruiting butdispensable for mating (Yue
et al
1999 Chang
et al
2000) Similarly calcineurin was essential for cation resis-tance in var
grubii
although it was not essential for cationresistance in
Cn
var
neoformans
suggesting a divergentrole for this gene in these two varieties (Cruz
et al
2000)There are no studies yet on mechanisms of virulence in
Cn
var
gattii
presumably because an efficient DNA trans-formation and gene disruption system has not yet beendescribed The present study describes the developmentof a transformation system and the successful disruptionof CuZn SOD (
SOD1
) in
Cn
var
gattii
Results
Disruption and reintroduction of the
SOD1
gene in
Cn
var
gattii
The
SOD1
gene was disrupted by replacing 907 bp of the957 bp open reading frame (ORF) of the gene with the
URA5
selectable marker which resulted in the CuZnSOD null mutant
sod1URA5
(Fig 1A) Our initialattempts to use the
URA5
selectable marker from
Cn
var
neoformans
for transformation of var
gattii
were unsuc-
cessful The comparative analysis of
URA5 from Cn varneoformans and var gattii revealed that this gene is ordf9divergent at the intragenic region and 30 divergent atthe promoter region These results suggested that URA5from Cn var neoformans might not be functional in vargattii Therefore to increase the transformation efficiencyas well as the homologous recombination we used URA5and SOD1 from Cn var gattii type strain NIH 444 A NotI-linearized sod1URA5 disruption allele was transformedinto a ura5 strain of var gattii Sixty transformants wereobtained on synthetic medium lacking uracil Thesetransformants were analysed initially for a presumptive no-growth phenotype on YPD menadione broth This screen-ing procedure yielded four suspected sod1 clones Nextgenomic DNA was isolated from these clones and analy-sed by polymerase chain reaction (PCR) for the absenceof the 500 bp fragment of the SOD1 ORF using primerpair V523 and V524 (details in Experimental procedures)If the sod1URA5 allele has been integrated at the homol-ogous site then the primer pair V523 and V524 shouldnot give the 500 bp product however if it is integrated atthe ectopic site then the genomic DNA should give one500 bp product of SOD1 wild type Out of four isolatestwo lacked the SOD1 wild-type locus as revealed by theabsence of 500 bp amplicons (Fig 1B) Southern hybrid-ization confirmed that the SOD1 wild-type locus had beenreplaced by the sod1URA5 disruption allele by homolo-gous recombination as no signal for SOD1 was detectedin the mutant strain when it was probed with the 500 bpfragment of SOD1 used from the 907 bp deleted regionof wild-type SOD1 (Fig 1C) The percentage homologousrecombination achieved was 34 (two positive out of 60transformants)
Next we generated a reconstituted strain in which awild-type copy of SOD1 with its own promoter was reinte-grated in the genome To make a reconstituted strain wereverted the ura+ phenotype by plating the sod1URA5mutant onto medium with 5-fluoroorotic acid (5-FOA) toselect for urandash cells This revertant was used as the recip-ient for transformation with a pCR61 plasmid containingfull-length SOD1 (28 kb) and URA5 selectable markerPCR analysis confirmed the restoration of the wild-typegene in these two reconstituted strains Interestinglyrestriction digestion of genomic DNA with XbaI and BstXIproduced an identical band size to that of the wild-typestrain which suggested homologous integration of SOD1to the sod1URA5 locus These reconstituted strains weredesignated as sod1 + SOD1-A and B
To examine the expression of the SOD1 gene we per-formed reverse transcription (RT)-PCR on total RNA iso-lated from SOD1 wild type the sod1 mutant and thesod1 + SOD1 reconstituted strains In contrast to theSOD1 wild-type strain in which a 400 bp amplicon wasreadily detected no message was detected in the sod1
Cn var gatti sod1 1683
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant strain This result supports the conclusion thatSOD1 has been functionally deleted in the sod1 mutantstrain After reintroduction of the SOD1 wild-type geneby transformation SOD1 expression was restored in thesod1 + SOD1 strains A and B (Fig 1D) Further geno-typic and phenotypic analyses confirmed that bothreconstituted strains exhibited identical properties there-fore results from sod1 + SOD1-A are shown in all otherexperiments
Characterization of Cn var gattii sod1 phenotypes
To address whether CuZn SOD is essential for maintain-ing normal growth of Cn in laboratory medium we deter-mined the doubling time of exponentially dividing SOD1wild type sod1 mutant and sod1 + SOD1 reconstitutedstrains in YPD broth at both 30infinC and 37infinC The doublingtime of the sod1 mutant was slightly longer at 30infinC and37infinC (33 plusmn 02 h and 34 plusmn 01 h) than that of the SOD1
Fig 1 Disruption and reconstitution of Cn var gattii SOD1 geneA A 28 kb fragment containing the SOD1 gene (solid thick line and dashed line outside 28 kb) indicates the genomic locus based on restriction enzyme digestion The lsquoprobersquo indicates the 500 bp sequence within the ORF used for PCR Southern hybridization and RT-PCR to confirm gene replacementB PCR for SOD1 ORF Primers designed from the SOD1 ORF (indicated by an arrow in A) amplified a 500 bp product from genomic DNA of SOD1 wild-type and sod1 + SOD1 reconstituted strains but not from genomic DNA of the sod1 mutant strain Negative control is PCR without genomic DNAC Southern blot of genomic DNA from the SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains Genomic DNA was digested with XbaI and BstXI and probed with the 500 bp PCR product depicted in (A) No hybridization signal was detected in the sod1 mutant whereas both SOD1 wild-type and sod1 + SOD1 (A and B) reconstituted strains produced asymp8 kb 4 kb and 08 kb bandsD RT-PCR to examine the expression of the SOD1 gene Total RNA was isolated reverse transcribed to cDNA and amplified with primers directed to SOD1 or actin RT-PCR products were fractionated by electrophoresis in a 1 agarose gel and stained with ethidium bromide Positive control is RT-PCR with RNA provided in the kit to confirm the efficacy of the kit and negative control is RT-PCR in the absence of RNA
A
B C D
1684 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
wild type (28 plusmn 01 h and 29 plusmn 02 h) or the sod1 + SOD1reconstituted strain (27 plusmn 03 h and 28 plusmn 01 h) How-ever these differences in growth between the sod1 mutantand SOD1 wild type were not statistically significant(P = 04) These results indicate that CuZn SOD is notessential for Cn growth Similarly CuZn SOD was seennot to be required for Cn growth under nutrient depriva-tion as no difference in growth between the sod1 mutantand the SOD1 wild type was observed when they wereassessed in terms of colony-forming units (cfu) of cellscultured for 15 days in water (Fig 2) An initial drop in cfunoted after 3 days of incubation in water might result froma shift from a very rich medium such as YPD to water andthis drop in cfu was consistent among SOD1 wild-type
sod1 mutant and sod1 + SOD1 reconstituted strains Alsothe sod1 mutant did not show auxotrophies for the sul-phur-containing amino acids lysine and methionine as thegrowth (OD600) determined for the sod1 mutant in com-plete CSM medium or in CSM lacking either lysine ormethionine was similar (data not shown)
It was also interesting to note that CuZn SOD was notrequired for Cn mating The sod1 mutant produced abun-dant filaments and basidiospores within 7 days at 24infinCsimilar to the behaviour of SOD1 wild type when mixedwith var neoformans MATa type strain NIH430 (data notshown) Such successful mating reactions between varneoformans and var gattii tester strains have been reportedin the literature (Kwon-Chung et al 1982) We did notisolate basidiospores from these crossings to test for theviability of the progeny However neither SOD1 wild-typenor sod1 mutant strains produced any basidiospores andfilaments when mixed with var gattii MATa type strainNIH191 Although it is not known why SOD1 wild type andthe sod1 mutant were unable to mate with tester strainNIH191 one possibility is that NIH191 might no longer bemating competent
Sod1 mutant is highly sensitive to menadione
The cytosolic localization of CuZn SOD implies that itsmain function is to protect cells against reactive oxygenintermediates (ROI Jamieson 1998) It is also reasonableto suspect that this localization may also enhance theefficiency of Cn in coping with oxidative stresses in thehost To address this question we grew Cn strains in thepresence of menadione (a redox cycling agent thatgenerates superoxide ions in the cytosol) to an initialOD600 of 01 The sod1 mutant was extremely sensitive tomenadione and its growth was completely inhibited at3 mg ml-1 at 30infinC This effect was more pronounced at37infinC as even a 1 mg ml-1 concentration of menadionewas enough to inhibit the sod1 mutant completely
Fig 2 sod1 mutant is viable in stationary phase culture Cells were allowed to grow in flasks for 48 h in YPD broth washed three times with sterile distilled water and reincubated in water Viability was monitored on the indicated days by plating aliquots onto YPD plates and cfu were counted The experiment was repeated twice with sim-ilar results and a representative experiment is shown
A
Fig 3 sod1 mutant is highly sensitive to mena-dione Cells were grown in YPD broth contain-ing menadione (0ndash3 mg ml-1) at initial OD600 of 01 under shaking conditions (180 rpm) The data shown were acquired 16 h after incuba-tion Experiments were repeated three times with similar results a representative experiment is shown
Cn var gatti sod1 1685
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
whereas the SOD1 wild-type strain was relatively resis-tant Introduction of the SOD1 gene restored the menadi-one resistance of the sod1 mutant to the wild-type levelThese results strongly suggest that cytosolic CuZn SODprotects Cn against oxidative stress (Fig 3)
Sod1 mutant shows fragmented vacuoles
Ultrastructural examination of the sod1 mutant revealedfragmentation of its large vacuole into aberrant shapes(Fig 4) In general the wild-type strain had two kinds ofvacuoles one large loosely packed vacuole (VL) and twoto six small densely packed vacuoles (VS) It was notedthat VL in sod1 mutant was fragmented as the internalcontents of the fragmented vacuole appeared to be similarto that of the VL seen in SOD1 wild type The analysis of30 individual electron microscopic (EM) sections revealedthat gt75 of the sod1 mutant cells had fragmented VL(12 plusmn 46 VL per cell) compared with one intact VL inSOD1 wild type (P lt 00001) Reconstitution of the sod1mutant with SOD1 wild type restored vacuolar morphol-ogy suggesting that CuZn SOD is essential for vacuolarintegrity and possibly for vacuolar function
Sod1 is defective in the production of several virulence factors
We next examined whether SOD1 deletion in Cn influ-ences other known virulence factors The virulence traitstested included production of melanin laccase ureasephospholipase B polysaccharide capsule and mannitolThe sod1 mutant exhibited a marked defect in melaniza-tion (Fig 5A) Melanin production was restored in asod1 + SOD1 reconstituted strain As laccase is the soleenzyme responsible for melanin production in Cn (Salas
et al 1996) laccase activity was measured from theequal number of sod1 mutant and SOD1 wild-type cellsas described previously (Zhang et al 1999) SOD1 dele-tion resulted in significant reduction in laccase activity inthe sod1 mutant compared with SOD1 wild type (P lt 001Fig 5B) Similarly the sod1 mutant was severely defectivein the production of phospholipase B on egg yolk agarand in the production of urease on urea agar media(Fig 5C and D) Reconstitution of the sod1 mutant withSOD1 wild type restored all the virulence-associated phe-notypes to a large extent On the other hand SOD1 dele-tion had no influence on capsule synthesis as revealedby India ink stains of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains grown in low-ironmedium for 3 days (Fig 6A) Interestingly the sod1mutant produced significantly more mannitol (P lt 005) inthe medium than the SOD1 wild-type strain (Fig 6B)
Sod1 mutant is attenuated in virulence in the mouse meningoencephalitis model
We tested whether CuZn SOD is required for virulenceof Cn var gattii in a mouse meningoencephalitis modelAs shown in Fig 7 after injection of 1 yen 106 yeast cellsmedian survival of mice inoculated with the SOD1 wild-type strain was 6 days whereas the median survival ofmice inoculated with the sod1 mutant was 23 days(P lt 00001) Reconstitution of the SOD1 gene into thesod1 mutant restored virulence with a median survival ofmice injected with sod1 + SOD1 of 9 days (P lt 00003)These results suggested that CuZn SOD plays an impor-tant role in Cn var gattii pathogenesis
Next we examined the in vivo tissue reaction inducedby Cn strains in infected mice As Cn has a tropism for
Fig 4 sod1 mutant shows fragmented vacuoles Cn cells were grown in YPD broth overnight at 30infinC and then processed for electron microscopic analysis as described in Experimental procedures N nucleus VS small vacuole VL large vacuole arrow points to fragmentation of large vacuole in the sod1 mutant Bar corresponds to 120 nm
1686 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
brain we collected brain tissues from infected mice 7 daysafter infection SOD1 wild type induced severe infectionwith the formation of 17 plusmn 70 lesions per brain section(each brain section comprised rostral cortex caudal cor-tex mid-brain and cerebellum with brainstem) comparedwith the sod1 mutant with 20 plusmn 12 lesions per brain sec-tion (P lt 00001) Infection with the sod1 + SOD1 recon-stituted strain was intermediate in severity resulting in anaverage of 8 plusmn 40 lesions per brain section The numberof yeasts per lesion for SOD1 wild-type sod1 mutant and
sod1 + SOD1 reconstituted strains ranged from 2 to 54 1to 3 and 1 to 37 respectively (Fig 8) These results sug-gested that the sod1 mutant is defective in brain coloni-zation and persistence No lesions were found in sectionsfrom control brain (Fig 8)
Fig 5 Defective production of virulence factors in sod1 mutantA Cells were grown in YPD broth overnight then washed three times with PBS and a 5 ml suspension of 107 cells ml-1 was placed on Niger seed agar for the determination of melanin synthesis and incu-bated at 30infinC for 4 daysB Phenoloxidase activity from an equal number of glucose-starved cells was determined by measuring the oxidation of the diphenolic substrate ABTS (IU of activity = 001 A420 absorbance unit in 30 min) The results are the mean plusmn SD of three individual experiments Aster-isk denotes P pound 005 compared with SOD1 wild typeC and D An aliquot of 5 ml of Cn cells (described in detail in A) were placed on Christensenrsquos agar for the determination of urease synthe-sis and on egg yolk agar for the determination of phospholipase synthesis
A
B
C
D
Fig 6 Capsule and mannitol synthesis in sod1 mutantA Cn cells were grown in low-iron medium for 3 days at 30infinC Cells were mixed with India ink and the capsule which excludes the ink particles was photographed Final magnification yen400B Extracellular mannitol produced from Cn cells was determined by a colorimetric method with absorbance at 412 nm The results are the mean plusmn SD of three individual experiments Asterisk denotes P lt005 compared with SOD1 wild type
A
B
A
B
Fig 7 Virulence of sod1 mutant in a mouse model SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains were grown in YPD broth for 16 h a suspension of 1 yen 106 cells was then injected intravenously into 6-week-old male BALBc mice which were observed daily until moribund
Cn var gatti sod1 1687
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Fig 8 Defective brain colonization and persistence of sod1 mutant Three mice were infected with 1 times 105 Cn cells intravenously then euthanized on day 7 and the brain sections were stained with Mayers mucicarmineA and B Sections from SOD1 wild-type infection reveal a large lesion (arrow) filled with fluid and SOD1 wild-type cells in the thalamus Organisms were also present in smaller numbers in the lateral ventricles and meninges (double arrow) (A at 20times) Higher magnification of the large cyst revealed numerous variable sized SOD1 wild-type cells some with budding (B at 200times)C and D A section from the sod1 mutant infection shows a very small cyst (arrow) (C at 20times) and higher magnification reveals a cyst with one budding sod1 mutant cell (D at 200times)E and F A section from the sod1 + SOD1 reconstituted strain infection reveals one large (arrow) and a few small cysts (double arrow) (E at 20times) and higher magnification of the large cyst reveals many Cn cells (F at 200times)
A
B C D
1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

Cn var gatti sod1 1683
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant strain This result supports the conclusion thatSOD1 has been functionally deleted in the sod1 mutantstrain After reintroduction of the SOD1 wild-type geneby transformation SOD1 expression was restored in thesod1 + SOD1 strains A and B (Fig 1D) Further geno-typic and phenotypic analyses confirmed that bothreconstituted strains exhibited identical properties there-fore results from sod1 + SOD1-A are shown in all otherexperiments
Characterization of Cn var gattii sod1 phenotypes
To address whether CuZn SOD is essential for maintain-ing normal growth of Cn in laboratory medium we deter-mined the doubling time of exponentially dividing SOD1wild type sod1 mutant and sod1 + SOD1 reconstitutedstrains in YPD broth at both 30infinC and 37infinC The doublingtime of the sod1 mutant was slightly longer at 30infinC and37infinC (33 plusmn 02 h and 34 plusmn 01 h) than that of the SOD1
Fig 1 Disruption and reconstitution of Cn var gattii SOD1 geneA A 28 kb fragment containing the SOD1 gene (solid thick line and dashed line outside 28 kb) indicates the genomic locus based on restriction enzyme digestion The lsquoprobersquo indicates the 500 bp sequence within the ORF used for PCR Southern hybridization and RT-PCR to confirm gene replacementB PCR for SOD1 ORF Primers designed from the SOD1 ORF (indicated by an arrow in A) amplified a 500 bp product from genomic DNA of SOD1 wild-type and sod1 + SOD1 reconstituted strains but not from genomic DNA of the sod1 mutant strain Negative control is PCR without genomic DNAC Southern blot of genomic DNA from the SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains Genomic DNA was digested with XbaI and BstXI and probed with the 500 bp PCR product depicted in (A) No hybridization signal was detected in the sod1 mutant whereas both SOD1 wild-type and sod1 + SOD1 (A and B) reconstituted strains produced asymp8 kb 4 kb and 08 kb bandsD RT-PCR to examine the expression of the SOD1 gene Total RNA was isolated reverse transcribed to cDNA and amplified with primers directed to SOD1 or actin RT-PCR products were fractionated by electrophoresis in a 1 agarose gel and stained with ethidium bromide Positive control is RT-PCR with RNA provided in the kit to confirm the efficacy of the kit and negative control is RT-PCR in the absence of RNA
A
B C D
1684 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
wild type (28 plusmn 01 h and 29 plusmn 02 h) or the sod1 + SOD1reconstituted strain (27 plusmn 03 h and 28 plusmn 01 h) How-ever these differences in growth between the sod1 mutantand SOD1 wild type were not statistically significant(P = 04) These results indicate that CuZn SOD is notessential for Cn growth Similarly CuZn SOD was seennot to be required for Cn growth under nutrient depriva-tion as no difference in growth between the sod1 mutantand the SOD1 wild type was observed when they wereassessed in terms of colony-forming units (cfu) of cellscultured for 15 days in water (Fig 2) An initial drop in cfunoted after 3 days of incubation in water might result froma shift from a very rich medium such as YPD to water andthis drop in cfu was consistent among SOD1 wild-type
sod1 mutant and sod1 + SOD1 reconstituted strains Alsothe sod1 mutant did not show auxotrophies for the sul-phur-containing amino acids lysine and methionine as thegrowth (OD600) determined for the sod1 mutant in com-plete CSM medium or in CSM lacking either lysine ormethionine was similar (data not shown)
It was also interesting to note that CuZn SOD was notrequired for Cn mating The sod1 mutant produced abun-dant filaments and basidiospores within 7 days at 24infinCsimilar to the behaviour of SOD1 wild type when mixedwith var neoformans MATa type strain NIH430 (data notshown) Such successful mating reactions between varneoformans and var gattii tester strains have been reportedin the literature (Kwon-Chung et al 1982) We did notisolate basidiospores from these crossings to test for theviability of the progeny However neither SOD1 wild-typenor sod1 mutant strains produced any basidiospores andfilaments when mixed with var gattii MATa type strainNIH191 Although it is not known why SOD1 wild type andthe sod1 mutant were unable to mate with tester strainNIH191 one possibility is that NIH191 might no longer bemating competent
Sod1 mutant is highly sensitive to menadione
The cytosolic localization of CuZn SOD implies that itsmain function is to protect cells against reactive oxygenintermediates (ROI Jamieson 1998) It is also reasonableto suspect that this localization may also enhance theefficiency of Cn in coping with oxidative stresses in thehost To address this question we grew Cn strains in thepresence of menadione (a redox cycling agent thatgenerates superoxide ions in the cytosol) to an initialOD600 of 01 The sod1 mutant was extremely sensitive tomenadione and its growth was completely inhibited at3 mg ml-1 at 30infinC This effect was more pronounced at37infinC as even a 1 mg ml-1 concentration of menadionewas enough to inhibit the sod1 mutant completely
Fig 2 sod1 mutant is viable in stationary phase culture Cells were allowed to grow in flasks for 48 h in YPD broth washed three times with sterile distilled water and reincubated in water Viability was monitored on the indicated days by plating aliquots onto YPD plates and cfu were counted The experiment was repeated twice with sim-ilar results and a representative experiment is shown
A
Fig 3 sod1 mutant is highly sensitive to mena-dione Cells were grown in YPD broth contain-ing menadione (0ndash3 mg ml-1) at initial OD600 of 01 under shaking conditions (180 rpm) The data shown were acquired 16 h after incuba-tion Experiments were repeated three times with similar results a representative experiment is shown
Cn var gatti sod1 1685
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
whereas the SOD1 wild-type strain was relatively resis-tant Introduction of the SOD1 gene restored the menadi-one resistance of the sod1 mutant to the wild-type levelThese results strongly suggest that cytosolic CuZn SODprotects Cn against oxidative stress (Fig 3)
Sod1 mutant shows fragmented vacuoles
Ultrastructural examination of the sod1 mutant revealedfragmentation of its large vacuole into aberrant shapes(Fig 4) In general the wild-type strain had two kinds ofvacuoles one large loosely packed vacuole (VL) and twoto six small densely packed vacuoles (VS) It was notedthat VL in sod1 mutant was fragmented as the internalcontents of the fragmented vacuole appeared to be similarto that of the VL seen in SOD1 wild type The analysis of30 individual electron microscopic (EM) sections revealedthat gt75 of the sod1 mutant cells had fragmented VL(12 plusmn 46 VL per cell) compared with one intact VL inSOD1 wild type (P lt 00001) Reconstitution of the sod1mutant with SOD1 wild type restored vacuolar morphol-ogy suggesting that CuZn SOD is essential for vacuolarintegrity and possibly for vacuolar function
Sod1 is defective in the production of several virulence factors
We next examined whether SOD1 deletion in Cn influ-ences other known virulence factors The virulence traitstested included production of melanin laccase ureasephospholipase B polysaccharide capsule and mannitolThe sod1 mutant exhibited a marked defect in melaniza-tion (Fig 5A) Melanin production was restored in asod1 + SOD1 reconstituted strain As laccase is the soleenzyme responsible for melanin production in Cn (Salas
et al 1996) laccase activity was measured from theequal number of sod1 mutant and SOD1 wild-type cellsas described previously (Zhang et al 1999) SOD1 dele-tion resulted in significant reduction in laccase activity inthe sod1 mutant compared with SOD1 wild type (P lt 001Fig 5B) Similarly the sod1 mutant was severely defectivein the production of phospholipase B on egg yolk agarand in the production of urease on urea agar media(Fig 5C and D) Reconstitution of the sod1 mutant withSOD1 wild type restored all the virulence-associated phe-notypes to a large extent On the other hand SOD1 dele-tion had no influence on capsule synthesis as revealedby India ink stains of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains grown in low-ironmedium for 3 days (Fig 6A) Interestingly the sod1mutant produced significantly more mannitol (P lt 005) inthe medium than the SOD1 wild-type strain (Fig 6B)
Sod1 mutant is attenuated in virulence in the mouse meningoencephalitis model
We tested whether CuZn SOD is required for virulenceof Cn var gattii in a mouse meningoencephalitis modelAs shown in Fig 7 after injection of 1 yen 106 yeast cellsmedian survival of mice inoculated with the SOD1 wild-type strain was 6 days whereas the median survival ofmice inoculated with the sod1 mutant was 23 days(P lt 00001) Reconstitution of the SOD1 gene into thesod1 mutant restored virulence with a median survival ofmice injected with sod1 + SOD1 of 9 days (P lt 00003)These results suggested that CuZn SOD plays an impor-tant role in Cn var gattii pathogenesis
Next we examined the in vivo tissue reaction inducedby Cn strains in infected mice As Cn has a tropism for
Fig 4 sod1 mutant shows fragmented vacuoles Cn cells were grown in YPD broth overnight at 30infinC and then processed for electron microscopic analysis as described in Experimental procedures N nucleus VS small vacuole VL large vacuole arrow points to fragmentation of large vacuole in the sod1 mutant Bar corresponds to 120 nm
1686 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
brain we collected brain tissues from infected mice 7 daysafter infection SOD1 wild type induced severe infectionwith the formation of 17 plusmn 70 lesions per brain section(each brain section comprised rostral cortex caudal cor-tex mid-brain and cerebellum with brainstem) comparedwith the sod1 mutant with 20 plusmn 12 lesions per brain sec-tion (P lt 00001) Infection with the sod1 + SOD1 recon-stituted strain was intermediate in severity resulting in anaverage of 8 plusmn 40 lesions per brain section The numberof yeasts per lesion for SOD1 wild-type sod1 mutant and
sod1 + SOD1 reconstituted strains ranged from 2 to 54 1to 3 and 1 to 37 respectively (Fig 8) These results sug-gested that the sod1 mutant is defective in brain coloni-zation and persistence No lesions were found in sectionsfrom control brain (Fig 8)
Fig 5 Defective production of virulence factors in sod1 mutantA Cells were grown in YPD broth overnight then washed three times with PBS and a 5 ml suspension of 107 cells ml-1 was placed on Niger seed agar for the determination of melanin synthesis and incu-bated at 30infinC for 4 daysB Phenoloxidase activity from an equal number of glucose-starved cells was determined by measuring the oxidation of the diphenolic substrate ABTS (IU of activity = 001 A420 absorbance unit in 30 min) The results are the mean plusmn SD of three individual experiments Aster-isk denotes P pound 005 compared with SOD1 wild typeC and D An aliquot of 5 ml of Cn cells (described in detail in A) were placed on Christensenrsquos agar for the determination of urease synthe-sis and on egg yolk agar for the determination of phospholipase synthesis
A
B
C
D
Fig 6 Capsule and mannitol synthesis in sod1 mutantA Cn cells were grown in low-iron medium for 3 days at 30infinC Cells were mixed with India ink and the capsule which excludes the ink particles was photographed Final magnification yen400B Extracellular mannitol produced from Cn cells was determined by a colorimetric method with absorbance at 412 nm The results are the mean plusmn SD of three individual experiments Asterisk denotes P lt005 compared with SOD1 wild type
A
B
A
B
Fig 7 Virulence of sod1 mutant in a mouse model SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains were grown in YPD broth for 16 h a suspension of 1 yen 106 cells was then injected intravenously into 6-week-old male BALBc mice which were observed daily until moribund
Cn var gatti sod1 1687
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Fig 8 Defective brain colonization and persistence of sod1 mutant Three mice were infected with 1 times 105 Cn cells intravenously then euthanized on day 7 and the brain sections were stained with Mayers mucicarmineA and B Sections from SOD1 wild-type infection reveal a large lesion (arrow) filled with fluid and SOD1 wild-type cells in the thalamus Organisms were also present in smaller numbers in the lateral ventricles and meninges (double arrow) (A at 20times) Higher magnification of the large cyst revealed numerous variable sized SOD1 wild-type cells some with budding (B at 200times)C and D A section from the sod1 mutant infection shows a very small cyst (arrow) (C at 20times) and higher magnification reveals a cyst with one budding sod1 mutant cell (D at 200times)E and F A section from the sod1 + SOD1 reconstituted strain infection reveals one large (arrow) and a few small cysts (double arrow) (E at 20times) and higher magnification of the large cyst reveals many Cn cells (F at 200times)
A
B C D
1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

1684 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
wild type (28 plusmn 01 h and 29 plusmn 02 h) or the sod1 + SOD1reconstituted strain (27 plusmn 03 h and 28 plusmn 01 h) How-ever these differences in growth between the sod1 mutantand SOD1 wild type were not statistically significant(P = 04) These results indicate that CuZn SOD is notessential for Cn growth Similarly CuZn SOD was seennot to be required for Cn growth under nutrient depriva-tion as no difference in growth between the sod1 mutantand the SOD1 wild type was observed when they wereassessed in terms of colony-forming units (cfu) of cellscultured for 15 days in water (Fig 2) An initial drop in cfunoted after 3 days of incubation in water might result froma shift from a very rich medium such as YPD to water andthis drop in cfu was consistent among SOD1 wild-type
sod1 mutant and sod1 + SOD1 reconstituted strains Alsothe sod1 mutant did not show auxotrophies for the sul-phur-containing amino acids lysine and methionine as thegrowth (OD600) determined for the sod1 mutant in com-plete CSM medium or in CSM lacking either lysine ormethionine was similar (data not shown)
It was also interesting to note that CuZn SOD was notrequired for Cn mating The sod1 mutant produced abun-dant filaments and basidiospores within 7 days at 24infinCsimilar to the behaviour of SOD1 wild type when mixedwith var neoformans MATa type strain NIH430 (data notshown) Such successful mating reactions between varneoformans and var gattii tester strains have been reportedin the literature (Kwon-Chung et al 1982) We did notisolate basidiospores from these crossings to test for theviability of the progeny However neither SOD1 wild-typenor sod1 mutant strains produced any basidiospores andfilaments when mixed with var gattii MATa type strainNIH191 Although it is not known why SOD1 wild type andthe sod1 mutant were unable to mate with tester strainNIH191 one possibility is that NIH191 might no longer bemating competent
Sod1 mutant is highly sensitive to menadione
The cytosolic localization of CuZn SOD implies that itsmain function is to protect cells against reactive oxygenintermediates (ROI Jamieson 1998) It is also reasonableto suspect that this localization may also enhance theefficiency of Cn in coping with oxidative stresses in thehost To address this question we grew Cn strains in thepresence of menadione (a redox cycling agent thatgenerates superoxide ions in the cytosol) to an initialOD600 of 01 The sod1 mutant was extremely sensitive tomenadione and its growth was completely inhibited at3 mg ml-1 at 30infinC This effect was more pronounced at37infinC as even a 1 mg ml-1 concentration of menadionewas enough to inhibit the sod1 mutant completely
Fig 2 sod1 mutant is viable in stationary phase culture Cells were allowed to grow in flasks for 48 h in YPD broth washed three times with sterile distilled water and reincubated in water Viability was monitored on the indicated days by plating aliquots onto YPD plates and cfu were counted The experiment was repeated twice with sim-ilar results and a representative experiment is shown
A
Fig 3 sod1 mutant is highly sensitive to mena-dione Cells were grown in YPD broth contain-ing menadione (0ndash3 mg ml-1) at initial OD600 of 01 under shaking conditions (180 rpm) The data shown were acquired 16 h after incuba-tion Experiments were repeated three times with similar results a representative experiment is shown
Cn var gatti sod1 1685
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
whereas the SOD1 wild-type strain was relatively resis-tant Introduction of the SOD1 gene restored the menadi-one resistance of the sod1 mutant to the wild-type levelThese results strongly suggest that cytosolic CuZn SODprotects Cn against oxidative stress (Fig 3)
Sod1 mutant shows fragmented vacuoles
Ultrastructural examination of the sod1 mutant revealedfragmentation of its large vacuole into aberrant shapes(Fig 4) In general the wild-type strain had two kinds ofvacuoles one large loosely packed vacuole (VL) and twoto six small densely packed vacuoles (VS) It was notedthat VL in sod1 mutant was fragmented as the internalcontents of the fragmented vacuole appeared to be similarto that of the VL seen in SOD1 wild type The analysis of30 individual electron microscopic (EM) sections revealedthat gt75 of the sod1 mutant cells had fragmented VL(12 plusmn 46 VL per cell) compared with one intact VL inSOD1 wild type (P lt 00001) Reconstitution of the sod1mutant with SOD1 wild type restored vacuolar morphol-ogy suggesting that CuZn SOD is essential for vacuolarintegrity and possibly for vacuolar function
Sod1 is defective in the production of several virulence factors
We next examined whether SOD1 deletion in Cn influ-ences other known virulence factors The virulence traitstested included production of melanin laccase ureasephospholipase B polysaccharide capsule and mannitolThe sod1 mutant exhibited a marked defect in melaniza-tion (Fig 5A) Melanin production was restored in asod1 + SOD1 reconstituted strain As laccase is the soleenzyme responsible for melanin production in Cn (Salas
et al 1996) laccase activity was measured from theequal number of sod1 mutant and SOD1 wild-type cellsas described previously (Zhang et al 1999) SOD1 dele-tion resulted in significant reduction in laccase activity inthe sod1 mutant compared with SOD1 wild type (P lt 001Fig 5B) Similarly the sod1 mutant was severely defectivein the production of phospholipase B on egg yolk agarand in the production of urease on urea agar media(Fig 5C and D) Reconstitution of the sod1 mutant withSOD1 wild type restored all the virulence-associated phe-notypes to a large extent On the other hand SOD1 dele-tion had no influence on capsule synthesis as revealedby India ink stains of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains grown in low-ironmedium for 3 days (Fig 6A) Interestingly the sod1mutant produced significantly more mannitol (P lt 005) inthe medium than the SOD1 wild-type strain (Fig 6B)
Sod1 mutant is attenuated in virulence in the mouse meningoencephalitis model
We tested whether CuZn SOD is required for virulenceof Cn var gattii in a mouse meningoencephalitis modelAs shown in Fig 7 after injection of 1 yen 106 yeast cellsmedian survival of mice inoculated with the SOD1 wild-type strain was 6 days whereas the median survival ofmice inoculated with the sod1 mutant was 23 days(P lt 00001) Reconstitution of the SOD1 gene into thesod1 mutant restored virulence with a median survival ofmice injected with sod1 + SOD1 of 9 days (P lt 00003)These results suggested that CuZn SOD plays an impor-tant role in Cn var gattii pathogenesis
Next we examined the in vivo tissue reaction inducedby Cn strains in infected mice As Cn has a tropism for
Fig 4 sod1 mutant shows fragmented vacuoles Cn cells were grown in YPD broth overnight at 30infinC and then processed for electron microscopic analysis as described in Experimental procedures N nucleus VS small vacuole VL large vacuole arrow points to fragmentation of large vacuole in the sod1 mutant Bar corresponds to 120 nm
1686 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
brain we collected brain tissues from infected mice 7 daysafter infection SOD1 wild type induced severe infectionwith the formation of 17 plusmn 70 lesions per brain section(each brain section comprised rostral cortex caudal cor-tex mid-brain and cerebellum with brainstem) comparedwith the sod1 mutant with 20 plusmn 12 lesions per brain sec-tion (P lt 00001) Infection with the sod1 + SOD1 recon-stituted strain was intermediate in severity resulting in anaverage of 8 plusmn 40 lesions per brain section The numberof yeasts per lesion for SOD1 wild-type sod1 mutant and
sod1 + SOD1 reconstituted strains ranged from 2 to 54 1to 3 and 1 to 37 respectively (Fig 8) These results sug-gested that the sod1 mutant is defective in brain coloni-zation and persistence No lesions were found in sectionsfrom control brain (Fig 8)
Fig 5 Defective production of virulence factors in sod1 mutantA Cells were grown in YPD broth overnight then washed three times with PBS and a 5 ml suspension of 107 cells ml-1 was placed on Niger seed agar for the determination of melanin synthesis and incu-bated at 30infinC for 4 daysB Phenoloxidase activity from an equal number of glucose-starved cells was determined by measuring the oxidation of the diphenolic substrate ABTS (IU of activity = 001 A420 absorbance unit in 30 min) The results are the mean plusmn SD of three individual experiments Aster-isk denotes P pound 005 compared with SOD1 wild typeC and D An aliquot of 5 ml of Cn cells (described in detail in A) were placed on Christensenrsquos agar for the determination of urease synthe-sis and on egg yolk agar for the determination of phospholipase synthesis
A
B
C
D
Fig 6 Capsule and mannitol synthesis in sod1 mutantA Cn cells were grown in low-iron medium for 3 days at 30infinC Cells were mixed with India ink and the capsule which excludes the ink particles was photographed Final magnification yen400B Extracellular mannitol produced from Cn cells was determined by a colorimetric method with absorbance at 412 nm The results are the mean plusmn SD of three individual experiments Asterisk denotes P lt005 compared with SOD1 wild type
A
B
A
B
Fig 7 Virulence of sod1 mutant in a mouse model SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains were grown in YPD broth for 16 h a suspension of 1 yen 106 cells was then injected intravenously into 6-week-old male BALBc mice which were observed daily until moribund
Cn var gatti sod1 1687
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Fig 8 Defective brain colonization and persistence of sod1 mutant Three mice were infected with 1 times 105 Cn cells intravenously then euthanized on day 7 and the brain sections were stained with Mayers mucicarmineA and B Sections from SOD1 wild-type infection reveal a large lesion (arrow) filled with fluid and SOD1 wild-type cells in the thalamus Organisms were also present in smaller numbers in the lateral ventricles and meninges (double arrow) (A at 20times) Higher magnification of the large cyst revealed numerous variable sized SOD1 wild-type cells some with budding (B at 200times)C and D A section from the sod1 mutant infection shows a very small cyst (arrow) (C at 20times) and higher magnification reveals a cyst with one budding sod1 mutant cell (D at 200times)E and F A section from the sod1 + SOD1 reconstituted strain infection reveals one large (arrow) and a few small cysts (double arrow) (E at 20times) and higher magnification of the large cyst reveals many Cn cells (F at 200times)
A
B C D
1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

Cn var gatti sod1 1685
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
whereas the SOD1 wild-type strain was relatively resis-tant Introduction of the SOD1 gene restored the menadi-one resistance of the sod1 mutant to the wild-type levelThese results strongly suggest that cytosolic CuZn SODprotects Cn against oxidative stress (Fig 3)
Sod1 mutant shows fragmented vacuoles
Ultrastructural examination of the sod1 mutant revealedfragmentation of its large vacuole into aberrant shapes(Fig 4) In general the wild-type strain had two kinds ofvacuoles one large loosely packed vacuole (VL) and twoto six small densely packed vacuoles (VS) It was notedthat VL in sod1 mutant was fragmented as the internalcontents of the fragmented vacuole appeared to be similarto that of the VL seen in SOD1 wild type The analysis of30 individual electron microscopic (EM) sections revealedthat gt75 of the sod1 mutant cells had fragmented VL(12 plusmn 46 VL per cell) compared with one intact VL inSOD1 wild type (P lt 00001) Reconstitution of the sod1mutant with SOD1 wild type restored vacuolar morphol-ogy suggesting that CuZn SOD is essential for vacuolarintegrity and possibly for vacuolar function
Sod1 is defective in the production of several virulence factors
We next examined whether SOD1 deletion in Cn influ-ences other known virulence factors The virulence traitstested included production of melanin laccase ureasephospholipase B polysaccharide capsule and mannitolThe sod1 mutant exhibited a marked defect in melaniza-tion (Fig 5A) Melanin production was restored in asod1 + SOD1 reconstituted strain As laccase is the soleenzyme responsible for melanin production in Cn (Salas
et al 1996) laccase activity was measured from theequal number of sod1 mutant and SOD1 wild-type cellsas described previously (Zhang et al 1999) SOD1 dele-tion resulted in significant reduction in laccase activity inthe sod1 mutant compared with SOD1 wild type (P lt 001Fig 5B) Similarly the sod1 mutant was severely defectivein the production of phospholipase B on egg yolk agarand in the production of urease on urea agar media(Fig 5C and D) Reconstitution of the sod1 mutant withSOD1 wild type restored all the virulence-associated phe-notypes to a large extent On the other hand SOD1 dele-tion had no influence on capsule synthesis as revealedby India ink stains of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains grown in low-ironmedium for 3 days (Fig 6A) Interestingly the sod1mutant produced significantly more mannitol (P lt 005) inthe medium than the SOD1 wild-type strain (Fig 6B)
Sod1 mutant is attenuated in virulence in the mouse meningoencephalitis model
We tested whether CuZn SOD is required for virulenceof Cn var gattii in a mouse meningoencephalitis modelAs shown in Fig 7 after injection of 1 yen 106 yeast cellsmedian survival of mice inoculated with the SOD1 wild-type strain was 6 days whereas the median survival ofmice inoculated with the sod1 mutant was 23 days(P lt 00001) Reconstitution of the SOD1 gene into thesod1 mutant restored virulence with a median survival ofmice injected with sod1 + SOD1 of 9 days (P lt 00003)These results suggested that CuZn SOD plays an impor-tant role in Cn var gattii pathogenesis
Next we examined the in vivo tissue reaction inducedby Cn strains in infected mice As Cn has a tropism for
Fig 4 sod1 mutant shows fragmented vacuoles Cn cells were grown in YPD broth overnight at 30infinC and then processed for electron microscopic analysis as described in Experimental procedures N nucleus VS small vacuole VL large vacuole arrow points to fragmentation of large vacuole in the sod1 mutant Bar corresponds to 120 nm
1686 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
brain we collected brain tissues from infected mice 7 daysafter infection SOD1 wild type induced severe infectionwith the formation of 17 plusmn 70 lesions per brain section(each brain section comprised rostral cortex caudal cor-tex mid-brain and cerebellum with brainstem) comparedwith the sod1 mutant with 20 plusmn 12 lesions per brain sec-tion (P lt 00001) Infection with the sod1 + SOD1 recon-stituted strain was intermediate in severity resulting in anaverage of 8 plusmn 40 lesions per brain section The numberof yeasts per lesion for SOD1 wild-type sod1 mutant and
sod1 + SOD1 reconstituted strains ranged from 2 to 54 1to 3 and 1 to 37 respectively (Fig 8) These results sug-gested that the sod1 mutant is defective in brain coloni-zation and persistence No lesions were found in sectionsfrom control brain (Fig 8)
Fig 5 Defective production of virulence factors in sod1 mutantA Cells were grown in YPD broth overnight then washed three times with PBS and a 5 ml suspension of 107 cells ml-1 was placed on Niger seed agar for the determination of melanin synthesis and incu-bated at 30infinC for 4 daysB Phenoloxidase activity from an equal number of glucose-starved cells was determined by measuring the oxidation of the diphenolic substrate ABTS (IU of activity = 001 A420 absorbance unit in 30 min) The results are the mean plusmn SD of three individual experiments Aster-isk denotes P pound 005 compared with SOD1 wild typeC and D An aliquot of 5 ml of Cn cells (described in detail in A) were placed on Christensenrsquos agar for the determination of urease synthe-sis and on egg yolk agar for the determination of phospholipase synthesis
A
B
C
D
Fig 6 Capsule and mannitol synthesis in sod1 mutantA Cn cells were grown in low-iron medium for 3 days at 30infinC Cells were mixed with India ink and the capsule which excludes the ink particles was photographed Final magnification yen400B Extracellular mannitol produced from Cn cells was determined by a colorimetric method with absorbance at 412 nm The results are the mean plusmn SD of three individual experiments Asterisk denotes P lt005 compared with SOD1 wild type
A
B
A
B
Fig 7 Virulence of sod1 mutant in a mouse model SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains were grown in YPD broth for 16 h a suspension of 1 yen 106 cells was then injected intravenously into 6-week-old male BALBc mice which were observed daily until moribund
Cn var gatti sod1 1687
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Fig 8 Defective brain colonization and persistence of sod1 mutant Three mice were infected with 1 times 105 Cn cells intravenously then euthanized on day 7 and the brain sections were stained with Mayers mucicarmineA and B Sections from SOD1 wild-type infection reveal a large lesion (arrow) filled with fluid and SOD1 wild-type cells in the thalamus Organisms were also present in smaller numbers in the lateral ventricles and meninges (double arrow) (A at 20times) Higher magnification of the large cyst revealed numerous variable sized SOD1 wild-type cells some with budding (B at 200times)C and D A section from the sod1 mutant infection shows a very small cyst (arrow) (C at 20times) and higher magnification reveals a cyst with one budding sod1 mutant cell (D at 200times)E and F A section from the sod1 + SOD1 reconstituted strain infection reveals one large (arrow) and a few small cysts (double arrow) (E at 20times) and higher magnification of the large cyst reveals many Cn cells (F at 200times)
A
B C D
1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

1686 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
brain we collected brain tissues from infected mice 7 daysafter infection SOD1 wild type induced severe infectionwith the formation of 17 plusmn 70 lesions per brain section(each brain section comprised rostral cortex caudal cor-tex mid-brain and cerebellum with brainstem) comparedwith the sod1 mutant with 20 plusmn 12 lesions per brain sec-tion (P lt 00001) Infection with the sod1 + SOD1 recon-stituted strain was intermediate in severity resulting in anaverage of 8 plusmn 40 lesions per brain section The numberof yeasts per lesion for SOD1 wild-type sod1 mutant and
sod1 + SOD1 reconstituted strains ranged from 2 to 54 1to 3 and 1 to 37 respectively (Fig 8) These results sug-gested that the sod1 mutant is defective in brain coloni-zation and persistence No lesions were found in sectionsfrom control brain (Fig 8)
Fig 5 Defective production of virulence factors in sod1 mutantA Cells were grown in YPD broth overnight then washed three times with PBS and a 5 ml suspension of 107 cells ml-1 was placed on Niger seed agar for the determination of melanin synthesis and incu-bated at 30infinC for 4 daysB Phenoloxidase activity from an equal number of glucose-starved cells was determined by measuring the oxidation of the diphenolic substrate ABTS (IU of activity = 001 A420 absorbance unit in 30 min) The results are the mean plusmn SD of three individual experiments Aster-isk denotes P pound 005 compared with SOD1 wild typeC and D An aliquot of 5 ml of Cn cells (described in detail in A) were placed on Christensenrsquos agar for the determination of urease synthe-sis and on egg yolk agar for the determination of phospholipase synthesis
A
B
C
D
Fig 6 Capsule and mannitol synthesis in sod1 mutantA Cn cells were grown in low-iron medium for 3 days at 30infinC Cells were mixed with India ink and the capsule which excludes the ink particles was photographed Final magnification yen400B Extracellular mannitol produced from Cn cells was determined by a colorimetric method with absorbance at 412 nm The results are the mean plusmn SD of three individual experiments Asterisk denotes P lt005 compared with SOD1 wild type
A
B
A
B
Fig 7 Virulence of sod1 mutant in a mouse model SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strains were grown in YPD broth for 16 h a suspension of 1 yen 106 cells was then injected intravenously into 6-week-old male BALBc mice which were observed daily until moribund
Cn var gatti sod1 1687
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Fig 8 Defective brain colonization and persistence of sod1 mutant Three mice were infected with 1 times 105 Cn cells intravenously then euthanized on day 7 and the brain sections were stained with Mayers mucicarmineA and B Sections from SOD1 wild-type infection reveal a large lesion (arrow) filled with fluid and SOD1 wild-type cells in the thalamus Organisms were also present in smaller numbers in the lateral ventricles and meninges (double arrow) (A at 20times) Higher magnification of the large cyst revealed numerous variable sized SOD1 wild-type cells some with budding (B at 200times)C and D A section from the sod1 mutant infection shows a very small cyst (arrow) (C at 20times) and higher magnification reveals a cyst with one budding sod1 mutant cell (D at 200times)E and F A section from the sod1 + SOD1 reconstituted strain infection reveals one large (arrow) and a few small cysts (double arrow) (E at 20times) and higher magnification of the large cyst reveals many Cn cells (F at 200times)
A
B C D
1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240
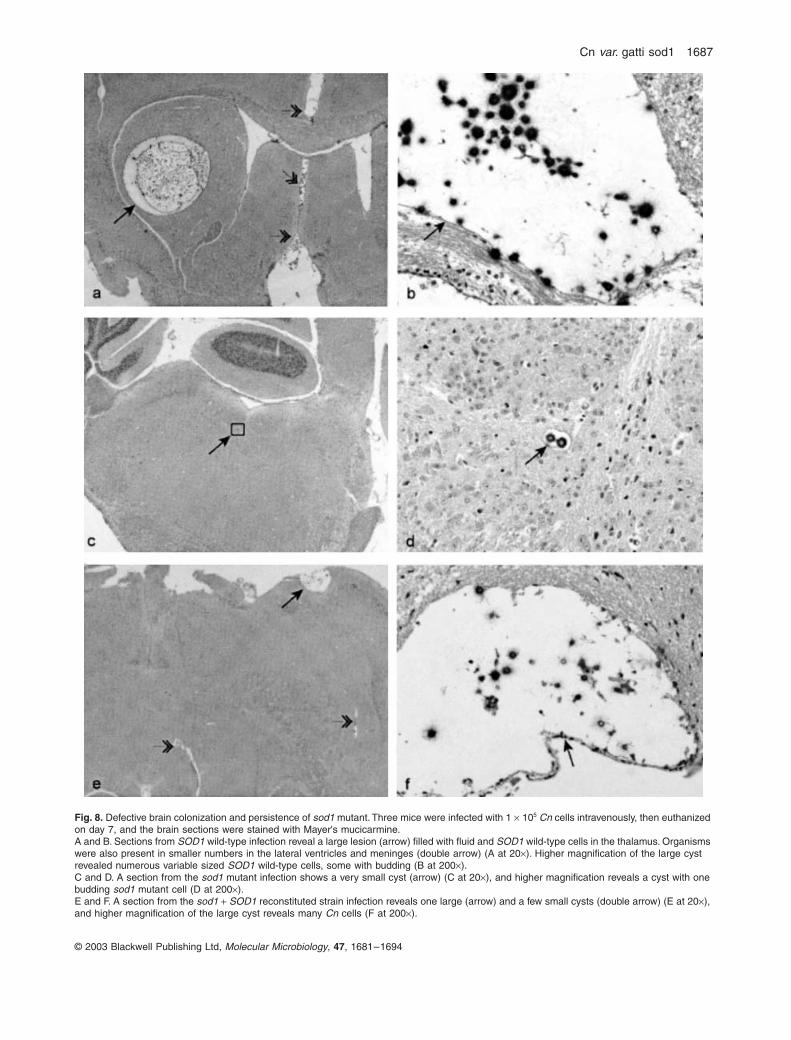
Cn var gatti sod1 1687
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Fig 8 Defective brain colonization and persistence of sod1 mutant Three mice were infected with 1 times 105 Cn cells intravenously then euthanized on day 7 and the brain sections were stained with Mayers mucicarmineA and B Sections from SOD1 wild-type infection reveal a large lesion (arrow) filled with fluid and SOD1 wild-type cells in the thalamus Organisms were also present in smaller numbers in the lateral ventricles and meninges (double arrow) (A at 20times) Higher magnification of the large cyst revealed numerous variable sized SOD1 wild-type cells some with budding (B at 200times)C and D A section from the sod1 mutant infection shows a very small cyst (arrow) (C at 20times) and higher magnification reveals a cyst with one budding sod1 mutant cell (D at 200times)E and F A section from the sod1 + SOD1 reconstituted strain infection reveals one large (arrow) and a few small cysts (double arrow) (E at 20times) and higher magnification of the large cyst reveals many Cn cells (F at 200times)
A
B C D
1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

1688 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Sod1 mutant is susceptible to PMN killing
Human PMNs were incubated with opsonized SOD1 wild-type sod1 mutant and sod1 + SOD1 reconstituted strainsin an effector-to-target ratio of 101 for 4 h and then Cnwere cultured on YPD medium to determine survivabilityThe sod1 mutant was significantly more susceptible to theantifungal activity of PMN than SOD1 wild type (percent-age cells killed 5100 plusmn 420 versus 2300 plusmn 230P lt 005) The exogenous addition of the O2
ndash inhibitorSOD inhibited PMN-induced killing of the sod1 mutantwhereas killing was minimal for SOD1 wild-type andsod1 + SOD1 reconstituted strains These results confirmthat CuZn SOD protects against PMN-mediated killing ofCn var gattii (Fig 9) It was interesting to note that overallPMN-induced killing of Cn var gattii wild type was quitelow 2300 plusmn 230 compared with the 80ndash85 killing ofCn var grubii reported earlier (Chaturvedi et al 1996)Increasing the PMN-to-Cn ratio from 101 to 1001 did notenhance killing of Cn var gattii (data not shown)
Discussion
The successful construction of a gene deletion mutantsod1 constitutes the first instance of the disruption of afunctional gene in Cn var gattii These results suggestthat the absence of the SOD1 gene in Cn var gattii (i)abrogates protection against oxidative killing by humanPMN and by the redox cycling agent menadione (ii)
causes reduced expression of the well-known virulencefactors laccase urease and phospholipase and (iii)causes attenuation of virulence in a mouse model of cryp-tococcal meningoencephalitis Reintroduction of the wild-type SOD1 gene complemented these defects and largelyrestored virulence Thus SOD1 is required for the primarypathogen Cn var gattii to express a number of othervirulence factors especially the metalloproteins for itssurvival under high oxidative stress conditions and for itsvirulence in an infected host This is believed to be thefirst report in which superoxide dismutase has been impli-cated directly in the virulence of a fungal pathogen
The SOD1 gene has been well characterized in a modeleukaryotic organism S cerevisiae in which it is requiredfor mating aerobic growth biosynthesis of sulphur-con-taining amino acids and long-term survival under nutrientdeprivation (Bilinski et al 1985 Gralla and Valentine1991 Liu et al 1992 Longo et al 1996) In generalSODs are important in maintaining an optimal intracellularredox environment in the cytoplasm that is needed toprevent lipid peroxidation DNA cross-linking and cys-teinendashcysteine bonding of essential enzymes (Powiset al 1995) In a Cn sod1 mutant the functional proper-ties of laccase urease and phospholipase were abro-gated Superoxide radicals are known to oxidize ironndashsulphur cluster proteins liberating iron (Keyer and Imlay1996) Thus CuZn SOD-deficient Cn may containincreased pools of reactive iron as has been shown forSOD1-deficient S cerevisiae and Escherichia coli(Liochev and Fridovich 1994 Keyer and Imlay 1996)This free iron may react with hydrogen peroxide andpossibly other reactive oxygen species to generatehydroxyl radical which is highly toxic for biomoleculesAnother possibility is that a SOD-induced reductive envi-ronment in the cytosol may be critical for the metal ioninsertion in metalloenzymes Support for this possibilitycomes from another study in which SOD was shown toprotect the FendashZn active centre of calcineurin in a mam-malian system (Wang et al 1996) Reintroduction of theCn SOD1 wild-type gene restored laccase urease andphospholipase functions to a large extent in the sod1mutant thus confirming the role of Cn CuZn SOD insustaining the function of other metalloenzymes It wasinteresting to note that SOD1 deletion in Cn did not influ-ence the synthesis of capsular proteins both the sod1mutant and the SOD1 wild type produced a large capsulewhen grown in low-iron medium On the other hand man-nitol synthesis was upregulated in the sod1 mutant com-pared with the wild-type parent strain This is notsurprising considering the fact that CuZn SOD being animportant antioxidant plays a crucial role not only in elim-inating O2
ndash but also in preventing the subsequent genera-tion of highly toxic OH Under the influence of distal ROIs(perhaps found throughout the cytosol of the sod1
Fig 9 sod1 susceptibility to human PMN killing Human PMNs at an effector-to-target ratio of 101 were incubated with opsonized Cn cells (5 yen 103) for 4 h at 37infinC in 5 CO2-95 air The percent Cn killed was determined from cfu counts as described in Experimental pro-cedures Results are mean plusmn SEM from PMNs of three individual donors Asterisk denotes P pound 005 compared with SOD1 wild type To determine whether exogenous SOD protects the sod1 mutant from PMN killing we preincubated PMNs with 10 mg ml-1 SOD for 1 h followed by incubation with opsonized Cn cells as shown above
Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

Cn var gatti sod1 1689
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
mutant) the mannitol upregulation may be a strategy forcoping with the loss of CuZn SOD by the sod1 mutantThere is also ample indirect evidence that mannitol pro-tects Cn by scavenging distal ROIs such as OHbull and HOCl(Chaturvedi et al 1996)
Recently a vesicular (H+)-ATPase proton pump(Vph1p) has been identified in Cn var grubii and disrup-tion of the vph1 gene resulted in a growth defect at 37infinCand also defects in the expression of many virulencefactors including the metalloenzymes laccase and ureaseThese results provide the first evidence for the critical roleof vacuoles in Cn survival and pathogenesis (Ericksonet al 2001) We have observed the fragmentation of largevacuoles (VL) into aberrant shapes in a sod1 mutant(12 plusmn 46 VL per cell) which might also be the cause ofdefective expression of many secreted virulence factorsVacuolar fragmentations were also observed in the Scerevisiae sod1 mutant Vacuoles play a critical role in ironhomeostasis in S cerevisiae Subcellular fractionation ofiron-loaded cells suggests that the vacuole is the majorsite of iron sequestration (Raguzzi et al 1988) Under theinfluence of increased superoxide levels vacuolar ironpools may be converted to a more accessible and activeFe2+ state that can catalyse OHbull production and can dam-age macromolecules in the immediate vicinity of the vac-uoles It is also possible that the primary damage occursto cytosolic components resulting in defective vacuolarstructure and function Future experiments may confirmthese possibilities
A second explanation for the attenuation of virulence inthe sod1 mutant could be its enhanced susceptibility toROIs This possibility is suggested by the (i) increasedkilling of the sod1 mutant by human PMN and (ii)enhanced sensitivity to the redox cycling agent menadi-one PMN killing of the sod1 mutant was significantlyabrogated by pretreatment with exogenous SOD whichindicated that the O2
ndash was critical for PMN-induced killingThus Cn CuZn SOD may be important for in vivo survivalof the fungus Further support for this conclusion comesfrom the results of experimental infection in mice Themedian survival of mice infected with the sod1 mutant was23 days versus 6 days for mice infected with the SOD1wild-type parent strain Introduction of SOD1 wild type intothe sod1 mutant restored virulence with a median survivalof 9 days These results indicate that SOD1 deletion sig-nificantly delays mortality in experimental cryptococcosisA related experimental study on brain infection and colo-nization indicated that the sod1 mutant caused signifi-cantly smaller and fewer lesions compared with parentand reconstituted strains These results suggest that Cncolonization in the brain is greatly reduced by SOD1 dele-tion However we cannot determine from these experi-ments whether oxidative conditions prohibit the survival ofthe sod1 mutant before it can establish residency in the
brain tissues or whether it is more sensitive and falls preyto the oxidative burst of a host immune response whileresiding in the brain tissues Overall CuZn SOD seemsto be critical for the ability of Cn var gattii to survive andmultiply in the host It is important to recall that SOD-deficient mutants of Helicobacter pylori Campylobactercoli and Mycobacterium tuberculosis were also shown tobe hypersensitive to oxidative stress and defective in hostcolonization (Purdy et al 1999 Edwards et al 2001Seyler et al 2001)
It was noteworthy that SOD1 deletion in general had noinfluence on Cn var gattii mating sporulation growthstationary phase survival or sulphur metabolism Theseresults are in contrast to phenotypes observed in the Scerevisiae sod1 mutant in which the loss of CuZn SODwas associated with 80ndash90 growth inhibition matingand sporulation defects dramatic loss of viability undernutrient deprivation and a defect in sulphur-containingamino acid metabolism (Liu et al 1992 Longo et al1996) These differences between S cerevisiae sod1 andCn var gattii sod1 could result from the fact that the latterfungus is highly adapted as a human and animal patho-gen and contains additional antioxidant systems that areabsent in S cerevisiae There is ample evidence thatlaccase (the enzyme involved in melanin synthesis)serves as an antioxidant in Cn (Salas et al 1996)Although both laccase and melanin production was signif-icantly reduced in the sod1 mutant the minuscule amountof antioxidant remaining in the mutant might be sufficientto sustain life under saprobic conditions Cn also producelarge amounts of the hexitol mannitol in culture which wasshown indirectly to act as an antioxidant (Chaturvedi et al1996) Recently an atypical cytosolic manganese-con-taining superoxide dismutase (SOD3) has been identifiedin C albicans and Sod3p was shown functionally to com-plement the defects of the S cerevisiae sod1 but not sod2(MnSOD2) mutant (Lamarre et al 2001) The Sod3penzyme was reported to protect C albicans against nutri-ent-limiting conditions and during the induction of oxida-tive stress At this time the search of the Cn varneoformans and Cn var grubii EST databases (httpwwwgenomeoueducneohtml) using the amino acidsequence of C albicans SOD3 as a query produced asingle hit that showed ordf63 homology to MnSOD(SOD2) suggesting that the existing Cn database doesnot have a putative sequence for SOD3 However thisfinding must be interpreted with caution as Cn EST data-bases are not yet complete
An important technical advance in our study was thesuccessful disruption of a gene by homologous recombi-nation in Cn var gattii The frequency of homologousrecombination was 34 which was comparable with thatreported for Cn varieties grubii and neoformans To avoidmismatches during homologous recombination we used
1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

1690 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
the SOD1 genomic locus from Cn var gattii We demon-strated earlier a 20ndash29 difference in deduced aminoacids of SOD1 among the three Cn varieties (Chaturvediet al 2001) Strict sequence identity is required for high-efficiency homologous recombination in Cn (Davidsonet al 1999) To increase the transformation efficiency inCn var gattii we used the URA5 gene from the samevariety It is important to point out here that the URA5gene from var gattii is 91 identical to the URA5 genefrom var neoformans However when the upstreamregion containing the promoter sequence was comparedthere was only 71 identity between these two varietiesindicating a possibility of low transformation of var gattiiwith the var neoformans URA5 gene Our findings indi-cate that it should now be feasible to disrupt any non-essential gene in Cn var gattii
In conclusion we have shown that CuZn SOD is criticalfor the pathogenesis of Cn var gattii but is not requiredfor the saprobic life of the fungus The mechanism ofaction of CuZn SOD could be protection against oxidativekilling by phagocytes combined with a reduction in theactivity of a number of virulence-related factors
Experimental procedures
Strains media and plasmids
All strains and plasmids used in this study are listed inTable 1 Standard yeast media eg YPD (yeast extract pep-tone dextrose) and CSM (complete synthetic medium) withrelevant amino acid dropouts were made as described pre-viously (Guthrie and Fink 1991) Niger seed agar (Salkin1979) urea agar (McGinnis 1980) egg yolk agar (Chenet al 1997) and limited iron medium with EDTA and batho-
phenanthroline disulphonic acid (Pierini and Doering 2001)were used for the determination of melanin production ure-ase activity phospholipase B activity and capsule formationrespectively Cn var gattii transformation was done usingbiolistic DNA delivery as described previously (Toffaletti et al1993) For biolistic transformation ordf5 mg of linear DNA wasused and transformants were selected on CSM lackinguracil
Preparation of probes
Probes used for Southern hybridization were prepared fromrestriction fragments or PCR-amplified fragments asdescribed in the text Probes were labelled with 32P asdescribed previously (Ausubel et al 1998)
Isolation of ura5 auxotrophs and cloning of the URA5 gene
A ura5 auxotrophic derivative of NIH 444 of Cn var gattii wasisolated by selection for 5-FOA resistance as reported inanother study (Kwon-Chung et al 1992)
Our initial attempt to transform Cn var gattii with pCIP-3(2 kb URA5 gene from Cn var neoformans in Bluescriptvector Edman and Kwon-Chung 1990) was unsuccessfulTherefore the URA5 gene was PCR amplified from genomicDNA of NIH 444 from var gattii with primer sets V479 (5cent-GGCATGGGTGATATACGTTTG-3cent) and V480 (5cent-TAGATAC-GATGAAGATTGACAGCC-3cent) The resulting 14 kb URA5fragment was cloned into pCR21 to yield pCR31 andsequenced A BLAST search confirmed the closest match withthe Cn URA5 gene The GenBank accession number for thevar gattii URA5 sequence is AF536328 The alignment ofURA5 from var gattii with that of var neoformans showed92 and 71 identity at the intragene and promoter regionsrespectively Approximately 20 transformants mg-1 linear DNA(pCR31) were obtained on CSM minus uracil medium
Table 1 Strains and plasmids used in this study
Strains and plasmids Parent Genotype Source
NIH 430(ATCC 28958)
Wild-type MATa (serotype D) American Type CultureCollection (ATCC) Manassas VA
NIH 191(ATCC 32608)
Wild type MATa (serotype C) ATCC
NIH 444(ATCC 32609)
Wild type MATa (serotype B) ATCC
NIH 444 ura5 MATa ura5 This studyCnsod1 NIH 444 ura5 MATa ura5 sod1URA5 This studyCnsod1undash Cnsod1 MATa ura5 sod1ura5 This studyCnsod1 + SOD1 Cnsod1undash MATa ura5 sod1ura5 SOD1URA5 This studypCIP-3 pBluescript 2 kb fragment containing the URA5 gene
from Cn var neoformans in Bluescript vectorJ EdmanEdman and Kwon-Chung (1990)
pCR31 pCR21 14 kb PCR insert containing URA5 ofCn var gattiiin pCR21vector
This study
pCR41 pCR21 28 kb PCR insert containing the SOD1gene fromCn var gattii in pCR21 vector
This study
pCR51 pCR41 14 kb URA5 subcloned into ClaI of sod1in pCR 41vector (sod1 disruption cassette)
This study
pCR61 pCR41 14 kb PCR insert of URA5 gene subcloned into This studyNotI of PCR41 (SOD1 complementation cassette)
Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

Cn var gatti sod1 1691
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Isolation and disruption of the Cn var gattii SOD1 gene
Full-length SOD1 was PCR amplified from genomic DNAof NIH 444 using primer sets 5cent-TACGAGGAAGAGAAATGGGG-3cent (V483) and 5cent-ATGTTGATCTGTGAACGCCC-3cent (V484) The 28 kb SOD1 PCR product wascloned into pCR21 vector to yield pCR41 pCR41 vectorwas digested with ClaI which resulted in the removal of atotal of 907 bp of the 957 bp SOD1 ORF A 14 kb PCRfragment of the URA5 gene obtained from genomic DNA ofNIH 444 strain was subcloned into ClaI sites to yield pCR51as shown in Fig 1 A total of 5 mg of NotI-linearized pCR51(sod1URA5) disruption plasmid was precipitated on 06 mggold microcarrier beads (Bio-Rad) and biolistically trans-formed into the NIH 444 ura5 strain (Toffaletti et al 1993)Ura+ transformants were selected on CSM lacking uracilThese clones were initially analysed for no-growth pheno-types on YPD broth containing 3 mg ml-1 menadione (super-oxide-generating compound) Genomic DNA was preparedfrom suspected sod1 clones using the DNeasy Plant Mini kit(Qiagen) PCR was performed according to the manufac-turerrsquos directions (Life Technologies) with the following SOD1specific primers designed from the deleted region of thegene 5cent-TACCGGAAATGTTGACGGA (V-527) and 5cent-CACAACTCCGCCATCAAAV (V-524)
Southern hybridization and RT-PCR
Genomic DNA (10 mg) was digested with various restrictionenzymes and elecrophoresed in 08 agarose gel The DNAwas then transferred to nylon membranes (Boehringer Man-nheim) using standard protocols (Sambrook et al 1989)Hybridization washing and detection of hybridized bandswere performed according to the instructions from the man-ufacturer (Boehringer Mannheim)
For RT-PCR total RNA from the Cn var gattii wild-typeSOD1 strain the sod1 mutant strain and the sod1 + SOD1reconstituted strain grown in YPD broth was isolated usingthe RNeasy mini kit (Ambion) after the mechanical disruptionof cells with glass beads First-strand cDNA synthesis wasperformed with 05 mg of total RNA and oligo (dT) primers asper the protocol described by the Gene Amp RNA PCR kit(Perkin-Elmer) The second-strand cDNA synthesis wasperformed with the primer sets 5cent-GCTGTTGCTGTCCTAAAGGG (V-548) and 5cent-GGAGATACCAATGACACCGC-3cent (V549) yielding a 413 bp SOD1 PCR amplicon Theseprimers were designed from the two separate exonic regionsof SOD1 If there was any genomic DNA contamination inthe RNA mixture this primer set would yield 513 bp becauseof the presence of two introns of 50 bp each RT-PCR wasalso carried out with actin as a loading control The actinprimer sets were 5cent-CAGCTGGAAGGTAGACAAAGAGGC-3cent(V-649) and 5cent-CGCTATTCCTCCGTATCGATCTTGC-3cent (V-650) these were again designed from two exonic regions ofthe actin gene to yield a 545 bp product from RNA and a600 bp product from DNA resulting from the presence of oneintron of 51 bp RT-PCR of control RNA provided in the kitwas also tested to confirm the efficacy of the kit whichyielded an expected amplicon of 308 bp
Reintroduction of the wild-type Cn var gattii SOD1 gene
A sod1URA5 prototrophic revertant sod1ura5 wasobtained by counterselection using 5-FOA medium (Kwon-Chung et al 1992) This strain was transformed biolisticallywith BamHI-linearized pCR61 plasmid and clones wereselected on CSM lacking uracil The growth rates at 30infinC ofthe 100 randomly selected reconstituted strains were com-pared with the wild-type strain in YPD broth containing3 mg ml-1 menadione Five of these strains with a more orless similar growth pattern to the wild-type strain were chosenfor further analysis by PCR Southern hybridization and RT-PCR Southern analysis confirmed that in two of the recon-stituted strains the sod1URA5 locus was replaced by SOD1by homologous integration These reconstituted strains weredesignated sod1 + SOD1 A and B
Characterization of growth and survival of the sod1 mutant
In vitro growth rates of SOD1 wild type sod1 mutant andsod1 + SOD1 reconstituted strains were determined by grow-ing them in YPD broth at 30infinC and 37infinC at 180 rpm withan initial OD600 of 01 Growth was followed up to 32 h bymonitoring the turbidity every 4 h Cn generation time wasdetermined by removing an aliquot every 1 h from the expo-nential phase of growth and counting cells in a haemocytom-eter For the long-term stationary phase survival experimentCn cells grown for 48 h in YPD broth were removed washedwith sterile distilled water and resuspended in water andincubation with shaking was continued for another 15 daysCell viability was determined by plating aliquots from culturesonto YPD agar and colony-forming units (cfu) were counted
Sensitivity to oxidative conditions
The sensitivity of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains to oxidative stress wasdetermined as described previously (Gralla and Valentine1991) Cn cultures were inoculated at OD600 = 01 in YPDbroth containing menadione ranging from 0 to 3 mg ml-1Growth was measured at both 30infinC and 37infinC 16 h afterincubation by monitoring the turbidity at OD600
Ultrastructure characterization
For ultrastructural analysis of Cn by electron microscopystrains were grown for 16 h before harvesting Cells werepelleted washed in phosphate-buffered saline (PBS pH 74)and fixed with 2 paraformaldehyde and 02 gluteralde-hyde in 01 M PBS for 3 h at room temperature Cells werewashed three times with PBS and frozen using a BalzersHPM010 then freeze-substituted in 1 osmium in acetonefor 72 h at -90infinC followed by 48 h incubation at -60infinC Cellswere slowly brought to room temperature washed twice in100 acetone for 1 h each infiltrated with 11 acetonendasharaldite epon and polymerized overnight at 65infinC Semi-thick(80ndash120 nm) sections were cut and stained with uranyl ace-tate and lead and examined using a Zeiss 910 electronmicroscope
1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

1692 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
Virulence factor expression
Laccase and melanin production The activity of laccasethe rate-limiting enzyme in melanin production wasassessed by measuring the oxidation of 22cent-azinobis (3-ethylbenzthiazolin-6-sulphonate) (ABTS) as describedpreviously (Zhang et al 1999) SOD1 wild-type sod1mutant and sod1 + SOD1 reconstituted strains were incu-bated in minimal asparagine medium with 01 glucosefor 16 h with shaking at 30infinC Cells were pelleted washedtwice with water resuspended in minimal asparaginemedium without glucose and incubated with shaking for5 h at 30infinC Cells were pelleted and then resuspended in01 M sodium acetate buffer (pH 50) at 2 yen 108 cells ml-1 The oxidation of ABTS was assessed spectrophotomet-rically by measuring the A420 of the supernatant of the cellsuspension 30 min after the addition of ABTS One unitof enzyme activity was defined as 001 absorbance unitsat 30 min Melanin production by the SOD1 wild-typesod1 mutant and sod1 + SOD1 reconstituted strains wasassayed on Niger seed agar A 5 ml suspension of 107
cells ml-1 from each strain was placed on the medium andincubated at 30infinC for 4 days
Urease and phospholipase B production Urease and phos-pholipase B activities of SOD1 wild-type sod1 mutant andsod1 + SOD1 reconstituted strains were tested in Chris-tensenrsquos urea agar and egg yolk agar media as reported inthe literature (Chen et al 1997 Cox et al 2000)
Capsule induction All strains assayed for capsule produc-tion were incubated in YPD medium at 30infinC for 48 h andsubsequently inoculated into 10 ml of low-iron medium After72 h of shaking (250 rpm) at 30infinC the cells were mixedwith standard India ink and photographed at 400yen magnifica-tion on an Olympus AX70 compound microscope equippedwith an Olympus PM-C350 digital camera
Mannitol production Extracellular mannitol produced by Cnwas determined by a published calorimetric method withsome modifications (Sanchez 1998) Cn cells were grownovernight in YPD broth to mid-log phase at 30infinC at 180 rpmApproximately 2 yen 108 cells were withdrawn and pelleted at5000 rpm for 2 min The supernatant (100 ml) was used forthe measurement of extracellular mannitol and we thenback-calculated for total extracellular mannitol secreted by2 yen 108 cells Fresh YPD broth was used as a blank Theassay was performed by mixing 100 ml of sample with 500 mlof 500 mM formate (pH 30) and 300 ml of 5 mM sodiumperiodate in a 15 ml microcentrifuge tube The mixture wasallowed to stand at room temperature for 15 s and then300 ml of a solution consisting of 100 mM acetyl acetone2000 mM acetate and 20 mM sodium thiosulphate wasadded This mixture was heated in a boiling water bath for2 min and then cooled to room temperature and absorbancewas read at 412 nm D-Mannitol (Sigma) was used for prep-aration of the standard curve Each experiment was per-formed in triplicate on two separate occasions
Mating and haploid filamentation assays
Cn var gattii SOD1 wild-type sod1 mutant and sod1 + SOD1reconstituted MATa strains and NIH 430 and NIH 191 MATastrains were grown on YPD agar for 48 h at 30infinC co-culturedon V8 mating medium and incubated at room temperature inthe dark for 2ndash5 days Haploid filamentation was assayed byincubating spotted suspension of cells on filament agar at24infinC for up to 4 weeks (Wickes et al 1996 Yue et al 1999)
Pathogenicity test
The pathogenic potential of the sod1 mutant strain comparedwith that of SOD1 wild-type and sod1 + SOD1 reconstitutedstrains was tested in a mouse model of meningoencephalitisBriefly Cn strains were grown in YPD broth for 16 h washedand resuspended in sterile saline Cells were counted bytrypan blue exclusion using a haemocytometer and by cfuon YPD agar Six male BALBc mice (ordf 6 weeks old 15ndash20 g)in each group were injected with a suspension of 106 cellsintravenously The injected animals were observed for anyovert sign of illness and all sick animals were promptly sac-rificed by CO2 inhalation to minimize pain and suffering Braintissues from dead mice were cultured on Niger seed agar forCn recovery
Additionally the histological lesions in the brain wereexamined in detail from mice infected with one log less Cncells (1 yen 105) as described for the survival curve experi-ments except that all infected animals were euthanized byday 7 (Chaturvedi et al 2002) Whole brain from two micefrom each group was fixed entire in 10 neutral-bufferedformalin along with one control brain Four transverse brainslices containing rostral cortex caudal cortex mid-brain andcerebellum with brainstem were processed into paraffinblocks using routine histological methods The tissue sec-tions were stained with haematoxylin and eosin (HampE) andMayerrsquos mucicarmine (Luna 1992) to stain the capsule brightmagenta Lesions were counted in six brain sections offour slices each from two brain tissues The values wererepresented as mean number of lesions plusmn SD per brainsection
Neutrophil (PMN) fungicidal activity
PMNs were isolated from the peripheral blood of normalhuman volunteers by Ficoll-Paque (Pharmacia LKB Biotech-nology) centrifugation PMNs containing red blood cells(RBCs) were harvested from the bottom of the tube mixedwith 50 ml of lysing solution (08 NH4CL 037 disodiumEDTA) and incubated at room temperature for 10 min to lyseRBCs PMNs were washed twice in RPMI (tissue culturemedium) and adjusted to the appropriate concentration in thesame medium PMN-induced killing of Cn var gattii SOD1wild-type sod1 mutant and sod1 + SOD1 reconstitutedstrains was determined as described earlier (Chaturvediet al 1996) Aliquots (100 ml) of PMNs 100 ml of viable Cncells (5 yen 103) and 20 ml of pooled human serum (PHS) wereadded to the wells of a 96-well tissue culture plate and incu-bated at 37infinC in 5 CO2-95 air After incubation for 4 hthe plates were centrifuged at 2000 rpm for 10 min and thesupernatants were carefully aspirated through 27-gauge nee-dles The effector cells were lysed by the addition of 100 ml
Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

Cn var gatti sod1 1693
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
of 005 Triton X-100 and the Cn cells were serially dilutedand plated on YPD agar The YPD plates were incubated at30infinC for 2ndash3 days for quantification of cfu Equivalentamounts of Cn grown in the culture medium without PMNswere treated identically with Triton X-100 and cfu wererecorded as a control The results were expressed as per-centage of Cn killed = 1ndash(cfu experimentcfu control) yen 100To determine the significance of CuZn SOD in PMN fungi-cidal activity we preincubated PMNs for 1 h with SODenzyme (10 mg ml-1) The controls included PMNs treatedwith SOD inactivated by autoclaving at 121infinC for 15 min
Statistical analysis
Statistical analysis of the data was performed using INSTAT
software for MacIntosh Studentrsquos t-test was used for com-parison of two groups Results were considered significant ata P-value lt005
Acknowledgements
We thank Adriana Verschoor for editorial comments We alsothank three anonymous reviewers for their valuable com-ments Nucleotide sequencing was performed at the Molec-ular Genetics Core Wadsworth Center The study wassupported financially in part by NIH grants RO1- A48462(SC) and R29-AI41968 (VC)
References
Alspaugh JA Perfect JR and Heitman J (1997) Crypto-coccus neoformans mating and virulence are regulated bythe G-protein a subunit GPA1 and cAMP Genes Dev 113206ndash3217
Ausubel FM Brent R Kingston RE Moore DDSeidman JG Smith JA and Struhl K (1998) CurrentProtocols in Molecular Biology New York John Wiley ampSons pp13111ndash13112
Bilinski T Krawiec Z Liczmanski A and Litwinska J(1985) Is hydroxyl radical generated by the Fenton reactionin vivo Biochem Biophys Res Commun 130 533ndash539
Casadevall A and Perfect JR (1998) Cryptococcus neo-formans Washington DC American Society for Microbiol-ogy Press pp1ndash531
Chang YC Wickes BL Miller GF Penoyer LA andKwon-Chung KJ (2000) Cryptococcus neoformansSTE12alpha regulates virulence but is not essential formating J Exp Med 191 871ndash882
Chaturvedi S Hamilton AJ Hobby P Zhu G LowryCV and Chaturvedi V (2001) Molecular cloning phylo-genetic analysis and three-dimensional modeling of CuZnsuperoxide dismutase (CnSOD1) from three varieties ofCryptococcus neoformans Gene 268 41ndash51
Chaturvedi V Wong B and Newman SL (1996) Oxidativekilling of Cryptococcus neoformans by human neutrophilsEvidence that fungal mannitol protects by scavenging reac-tive oxygen intermediates J Immunol 156 3836ndash3840
Chaturvedi V Fan J Stein B Behr MJ SamsonoffWA Wickes BL and Chaturvedi S (2002) Moleculargenetic analyses of mating pheromones reveals intervari-ety matinghybridization in Cryptococcus neoformansInfect Immun 70 5225ndash5235
Chen SC Muller AM Zhou JZ Wright LC and SorrellTC (1997) Phospholipase activity in Cryptococcus neo-formans a new virulence factor J Infect Dis 175 414ndash420
Chen S Sorrell T Nimmo G Speed B Currie B EllisD et al (2000) Epidemiology and host- and variety-depen-dent characteristics of infection due to Cryptococcus neo-formans in Australia and New Zealand AustralasianCryptococcal Study Group Clin Infect Dis 31 499ndash508
Church D and Washburn R (1992) Comparison of viru-lence among serotypes of Cryptococcus neoformans in amurine inhalation model In Abstracts of the General Meet-ing Washington DC American Society for MicrobiologyPress Abstract B-328 p 80
Cox GM Mukherjee J Cole GT Casadevall A andPerfect JR (2000) Urease as a virulence factor in exper-imental cryptococcosis Infect Immun 68 443ndash448
Cox GM McDade HC Chen SC Tucker SC Got-tfredsson M Wright LC et al (2001) Extracellular phos-pholipase activity is a virulence factor for Cryptococcusneoformans Mol Microbiol 39 166ndash175
Cruz MC Sia RA Olson M Cox GM and HeitmanJ (2000) Comparison of the roles of calcineurin inphysiology and virulence in serotype D and serotype Astrains of Cryptococcus neoformans Infect Immun 68982ndash985
Davidson RC Cruz MC Sia RAL Allen B AlspaughJA and Heitman J (1999) Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neo-formans Fungal Genet Biol 29 38ndash48
Dong ZM and Murphy JW (1995) Effects of two varietiesof Cryptococcus neoformans cells and culture filtrate anti-gens on neutrophil locomotion Infect Immun 63 2632ndash2644
Edman JC and Kwon-Chung KJ (1990) Isolation of theURA5 gene from Cryptococcus neoformans var neofor-mans and its use as a selectable marker for transformationMol Cell Biol 10 4538ndash4544
Edwards KM Cynamon MH Voladri RK Hager CCDeStefano MS Tham KT et al (2001) Iron-cofactoredsuperoxide dismutase inhibits host responses to Mycobac-terium tuberculosis Am J Respir Crit Care Med 164 2213ndash2219
Erickson T Liu L Gueyikian A Zhu X Gibbons J andWilliamson PR (2001) Multiple virulence factors of Cryp-tococcus neoformans are dependent on VPH1 Mol Micro-biol 42 1121ndash1131
Fisher D Burrow J Lo D and Currie B (1993) Crypto-coccus neoformans tropical northern Australia predomi-nantly variant gattii with good outcomes Aust NZ J Med23 678ndash682
Goscin SA and Fridovich I (1972) The purification andproperties of superoxide dismutase from Saccharomycescerevisiae Biochim Biophys Acta 289 276ndash283
Gralla E and Valentine JS (1991) Null mutants of Sac-charomyces cerevisiae CuZn superoxide dismutase char-acterization of spontaneous mutation rates J Bacteriol173 5918ndash5920
Guthrie C and Fink GR (1991) Guide to Yeast Geneticsand Molecular Biology In Methods in Enzymology SanDiego CA Academic Press
Hamilton AJ and Holdom MD (1997) Biochemical com-parison of the CuZn superoxide dismutases of Cryptococ-
1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240

1694 S D Narasipura et al
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1681ndash1694
cus neoformans var neoformans and Cryptococcusneoformans var gatti Infect Immun 65 488ndash494
Hamilton AJ Holdom MD and Jeavons L (1996)Expression of the CuZn superoxide dismutase of Aspergil-lus fumigatus as determined by immunochemistry andimmunoelectron microscopy FEMS Immunol Med Micro-biol 14 95ndash102
Hwang C Rhie SG Kim ST Kim YR Huh WKBaek YU and Kang SO (1999) Copper- and zinc-containing superoxide dismutase and its gene from Can-dida albicans Biochim Biophys Acta 1427 245ndash255
Jamieson DJ (1998) Oxidative stress responses of theyeast Saccharomyces cerevisiae Yeast 14 1511ndash1527
Keyer K and Imlay JA (1996) Superoxide acceleratesDNA damage by elevating free-iron levels Proc Natl AcadSci USA 93 13635ndash13640
Kwon-Chung KJ and Bennett JA (1992) Medical Mycol-ogy Philadelphia Lea amp Febiger
Kwon-Chung KJ Bennett JE and Rhodes JC (1982)Taxonomic studies on Filobasidiella species and theiranamorphs Antonie van Leeuwenhoek 48 25ndash38
Kwon-Chung KJ Varma A Edman JC and BennettJE (1992) Selection of ura5 and ura3 mutants from thetwo varieties of Cryptococcus neoformans on 5-fluorooroticacid medium J Med Vet Mycol 30 61ndash69
Lamarre C LeMay JD Deslauriers N and BourbonnaisY (2001) Candida albicans expresses an unusual cyto-plasmic manganese-containing superoxide dismutase(SOD3 gene product) upon the entry and during the sta-tionary phase J Biol Chem 276 43784ndash43791
Liochev SI and Fridovich I (1994) The role of O2bull - in
the production of HObull in vitro and in vivo Free Rad BiolMed 16 29ndash33
Liu X Elashvill FI Gralla EB Valentine J LapinskasP and Culotta V (1992) Yeast lacking superoxide dismu-tase isolation of a genetic suppressors J Biol Chem 26718298ndash18303
Lodge JK Jackson-Machelski E Toffaletti DL PerfectJR and Gordon JI (1994) Targeted gene replacementdemonstrates that myristoyl-CoA protein N-myristoyltrans-ferase is essential for viability of Cryptococcus neofor-mans Proc Natl Acad Sci USA 91 12008ndash12012
Longo VD Gralla EB and Valentine JS (1996) Super-oxide dismutase activity is essential for stationary phasesurvival in Saccharomyces cerevisiae J Biol Chem 27112275ndash12280
Luna LG (1992) Histopathologic Methods and Color Atlasof Special Stains and Tissue Artifacts Downers Grove ILJohnson Printers
McGinnis MR (1980) Laboratory Handbook of MedicalMycology New York Academic Press
Miller RA and Britigan BE (1997) Role of oxidants inmicrobial pathophysiology Clin Microbiol Rev 10 1ndash18
Misra HP and Fridovich I (1972) The purification andproperties of superoxide dismutase from Neurosporacrassa J Biol Chem 247 3410ndash3414
Mutoh NC Nakagawa W and Yamada K (2002) Char-acterization of CuZn-superoxide dismutase-deficientmutant of fission yeast Schizosaccharomyces pombe CurrGenet 41 82ndash88
Odom A Muir S Lim E Toffaletti DL Perfect J and
Heitman J (1997) Calcineurin is required for virulence ofCryptococcus neoformans EMBO J 16 2576ndash2589
Pierini LM and Doering TL (2001) Spatial and temporalsequence of capsule construction in Cryptococcus neofor-mans Mol Microbiol 41 105ndash115
Powis G Briehl M and Oblong J (1995) Redox signalingand the control of cell growth and death Pharmacol Ther68 149ndash173
Purdy D Cawthraw S Dickinson JH Newell DGand Park SF (1999) Generation of a superoxide dismu-tase (SOD)-deficient mutant of Campylobacter coli evi-dence for the significance of SOD in Campylobactersurvival and colonization Appl Environ Microbiol 652540ndash2546
Raguzzi F Lesuisse E and Crichton RR (1988) Ironstorage in Saccharomyces cerevisiae FEBS Lett 231253ndash258
Salas SD Bennett JE Kwon-Chung KJ Perfect JRand Williamson PR (1996) Effect of the laccase geneCNLAC1 on virulence of Cryptococcus neoformans J ExpMed 184 377ndash386
Salkin IF (1979) Further simplification of Guizotia abyssin-ica seed medium for identification of Cryptococcus neofor-mans and Cryptococcus bacillispora Can J Microbiol 251116ndash1118
Sambrook J Fritsch EF and Maniatis T (1989) Analysisof genomic DNA by Southern Hybridization In MolecularCloning A Laboratory Manual Vol 1 2nd edn Cold SpringHarbor NY Cold Spring Harbor Laboratory Press 931ndash945
Sanchez J (1998) Colorimetric assay of alditols in complexbiological samples J Agric Food Chem 46 157ndash160
Seyler RW Jr Olson JW and Maier RJ (2001) Super-oxide dismutase-deficient mutants of Helicobacter pyloriare hypersensitive to oxidative stress and defective in hostcolonization Infect Immun 69 4034ndash4040
Sorrell TC (2001) Cryptococcus neoformans variety gattiiMed Mycol 39 155ndash168
Speed B and Dunt D (1995) Clinical and host differencesbetween infections with two varieties of Cryptococcus neo-formans Clin Infect Dis 21 28ndash34
Toffaletti D Rude TH Johnston SA Durack DT andPerfect JR (1993) Gene transfer in Cryptococcus neofor-mans by use of biolistic delivery of DNA J Bacteriol 1751405ndash1411
Wang X Culaotta VC and Kliee C (1996) Superoxidedismutase protects calcineurin from inactivation Nature383 434ndash437
Wickes BL Mayorga ME Edman U and Edman JC(1996) Dimorphism and haploid fruiting in Cryptococcusneoformans Association with the a-mating type Proc NatlAcad Sci USA 93 7327ndash7331
Yue CL Cavallo M Alspaugh JA Wang P Cox GMPerfect JR and Heitman J (1999) The STE12 alphahomolog is required for haploid filamentation but largelydispensable for mating and virulence in Cryptococcus neo-formans Genetics 153 1601ndash1615
Zhang S Varma A and Williamson PR (1999) The Cryp-tococcus neoformans uses lsquomammalianrsquo enhancer sites inthe regulation of the virulence gene CNLAC1 Gene 227231ndash240




















