
Detection and Mitigation of Botnet Infiltration using Intelligent Swarm Networks
Upload
univ-rennes1Category
view
2download
0
www.elsevier.com/locate/catena
Catena 54 (2003) 637–649
Changes of pore morphology, infiltration and
earthworm community in a loamy soil under
different agricultural managements
Mathieu Lamandea, Vincent Hallairea,*, Pierre Curmia,b,Guenola Peresc, Daniel Cluzeauc
a INRA-Centre de Rennes, Unite Sol et Agronomie Rennes Quimper, 65, route de Saint-Brieuc,
35042 Rennes, FrancebFrench Institute of Pondicherry, 11, Saint-Louis Street, PB 33, Pondicherry 605001, India
cCNRS, UMR 6553 ‘‘Ecobio’’—Laboratoire d’Ecologie du Sol et de Biologie des Populations,
Station Biologique, 35380 Paimpont, France
Abstract
Earthworm activity produces changes at different scales of soil porosity, including the
mesoporosity (between 1.000 and 30 Am eq. dia.) where both water retention and near-saturated
infiltration take place. At this scale, the structural changes are poorly described in temperate
agricultural systems, so we do not yet fully understand how these changes occur. The present study
was conducted to determine the relationships between the morphology of the mesopores, which is
mainly affected by earthworm activity, and the hydrodynamic behaviour (near-saturated infiltration)
of topsoil under different agricultural managements inducing a large range of earthworm
populations.
Investigations were carried out at the soil surface in three fields under different management
practices giving rise to three different earthworm populations: a continuous maize field where pig
slurry was applied, a rye-grass/maize rotation (3/1 year, respectively) also with pig slurry, and an old
pasture sown with white clover and rye-grass.
Pore space was quantified using a morphological approach and 2D image analysis.
Undisturbed soil samples were impregnated with polyester resin containing fluorescent pigment.
The images were taken under UV light, yielding a spatial resolution of 42 Am pixel� 1. Pores were
classified according to their size (which is a function of their area) and their shape. Hydraulic
conductivity K(h) was measured using a disc infiltrometer at four water potentials: � 0.05, � 0.2,
� 0.6, and � 1.5 kPa. The abundance and ecological categories groups of earthworms were also
investigated.
0341-8162/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/S0341-8162(03)00114-0
* Corresponding author. Tel.: +33-223-485-429; fax: +33-223-485-430.
E-mail address: [email protected] (V. Hallaire).

M. Lamande et al. / Catena 54 (2003) 637–649638
Continuous soil tillage causes a decrease in both abundance and functional diversity (cf. maize
compared with old pasture) when soil tillage every 4 years causes only a decrease in abundance (cf.
rotation compared with old pasture). There were no relationships between total porosity and effective
porosity at h =� 0.05 kPa. Image analysis was useful in distinguishing the functional difference
between the three managements. Fewer roots and anecic earthworms resulted in fewer effective
tubular voids under maize. There were fewer packing voids in the old pasture due to cattle trampling.
Greater hydraulic conductivity in the pasture phase of rotation may arise from a greater functional
diversity than in the maize and absence of cattle trampling compared with the pasture. We point to
some significant differences between the three types of agricultural management.
A better understanding is required of the influence of agricultural management systems on pore
morphology. This study provides a new methodology in which we consider the earthworm activity as
well as community in order to assess the effects of agricultural management on soil structure and
water movement.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Soil structure; Hydraulic conductivity; Image analysis; Cropping systems; Earthworms
1. Introduction
Understanding the role of fauna on soil physical properties is important in developing
sustainable agricultural managements, and during the last decade, important efforts were
devoted to the description and quantification of the direct and indirect effects of soil
invertebrates on the major processes of the soil, in particular, the formation and
conservation of the physical structure. In temperate regions, the earthworms in term of
biomass constitute the principal component of the total faunal biomass (Lee, 1985).
Earthworm populations are affected by agricultural management (Binet et al., 1997;
Paoletti et al., 1998; Chan, 2001), and earthworms have a large influence on soil physical
properties through their burrowing and casting activities. Also known as ‘‘ecosystem
engineers’’ (Jones et al., 1994), earthworms produce structural features at three different
scales of soil porosity. Much work deals with the characterisation of burrow networks
created by earthworm species (Capowiez et al., 1998; Jegou et al., 1999). In relation to
macropore space (>1 mm), burrow networks act as preferential flow paths (Bouche and
Al-Addan, 1997; Trojan and Linden, 1998). At a smaller scale, earthworms may change
the pore space between mineral and organic particles, i.e. the microporosity, and the
stability of soil structure (Shipitalo and Protz, 1989; Blanchart et al., 1993; Chauvel et al.,
1999). Packing voids within cast deposits control soil mesoporosity, in which large
amounts of water and solutes are transported and retained. Because these structural
features have been little studied in temperate agricultural systems, we are here primarily
concerned with the mesoscale aspects of pore morphology.
The effects of earthworm activity and agricultural management on soil physical
properties have been studied, but only in terms of preferential flow paths (Ehlers, 1975;
Sveistrup et al., 1997). Hallaire and Curmi (1994) and Kribaa et al. (2001) showed the
main role of morphology in linking the effective porosity with movement of water and
solutes. The effect of agricultural management systems on soil physical properties has

M. Lamande et al. / Catena 54 (2003) 637–649 639
often been quantified with near-saturated infiltration measurements (Ankeny et al., 1990;
Meek et al., 1992; Azevedo et al., 1998; Heddadj and Gascuel-Odoux, 1999; Angulo-
Jaramillo et al., 2000).
Our objective here is to evaluate the effect of various agricultural managements and
natural earthworm populations on the physical properties of topsoil. We looked for
relationships between the morphology of the mesopores and the rate of near-saturated
infiltration. We measured in situ hydraulic conductivity at multiple water potentials near
saturation with tension infiltrometers and characterised the morphology of a large part of
mesopores by image analysis of a soil affected by both agricultural treatments and
earthworm activity.
2. Materials and methods
2.1. The experimental design
The experiment was carried out at the experimental station of the Lycee Agricole de
Kerbernez, in western Brittany, France (latitude 47j57VN, longitude 4j8VW). Agriculture
in this region consists mainly of intensive milk production characterised by rotations of
maize and pasture. The climate is of temperate oceanic type, with an average annual
precipitation of 1200 mm and a mean annual temperature of 11.4 jC. The soil is a Humic
Cambisol (FAO) of loamy texture with a high concentration of organic matter in the first
30 cm (Table 1) developed on granitic saprolite.
The trial comprised three plots, each 9 m wide and 16 m long, that were managed as
follows: (i) continuous maize treated with pig slurry for 22 years, (ii) a pasture phase (1st
year) of a rye-grass/maize rotation (3/1 year) also with pig slurry for 22 years, (iii) old
pasture sown with white clover and rye-grass maintained over a period of 9 years. Physical
measurements and soil sampling were performed from soil surface on the most represen-
tative zone in the topsoil, determined by mapping the structural features of the given
ploughed soil horizon (Manichon and Roger-Estrade, 1990; Curmi et al., 1996) just before
maize seeding (March) after 6 months without tillage.
2.2. Earthworm community
Natural earthworm community was extracted in each field using the formaldehyde
method on 1 m2 (Bouche, 1972; Cluzeau et al., 1999); after three sprayings of
Table 1
Soil characteristics of the top soil in the three studied fields
< 2 Am(%)
2–20
Am (%)
20–50
Am (%)
50–200
Am (%)
200–2000
Am (%)
Organic matter
content (%)
pHwater
Maize 16.1 21.4 20.4 13.2 28.9 4.18 6.20
Rotation 17.7 19.6 23.6 13.1 26.6 4.43 5.65
Old pasture 17.0 17.5 17.5 18.0 30.0 4.20 5.90

M. Lamande et al. / Catena 54 (2003) 637–649640
formaldehyde solution (10 l per spraying with different concentrations: 0.25%, 0.25%,
0,4%), earthworms were collected at the soil surface. Three replicates were performed in
each field surrounding the sites chosen for physical measurement and soil sampling.
Earthworm communities were characterised by their abundance (number collected per
m2) and their ecological group. This last parameter is based on earthworm morphology
and behaviour (localisation in soil, feeding behaviour), and corresponds to three
ecological groups (Lee, 1959; Bouche, 1972, 1977) whose burrow systems were
described (Kretzschmar and Aries, 1990; Lee and Foster, 1991; Lavelle, 1997): epigeic
(range, 1–2.5 mm in diameter, live and feed above the soil surface, create no or few
burrows), anecic (range, 4–8 mm in diameter, feed at the ground surface, live in
semipermanent burrows, more or less vertical and opened to the soil surface), and
endogeic (range, 2–4.5 mm in diameter, ingeste soil, dig extensive systems of temporary
burrows that they immediately refill with their casts, the burrows are mostly subhorizontal
oriented and very ramified through the soil but rarely open to the surface). In order to link
earthworm activity to soil properties, the earthworm communities were also characterised
by their functional diversity that combines the ecological group and growth stage (juvenile,
adult) (Peres et al., 1998); six functional classes are defined.
2.3. Physical measurements
Hydraulic conductivity K(h) was measured using a disc tension infiltrometer with an
80-mm-diameter base, which determined tension at the soil surface as described in Ankeny
et al. (1990, 1991). Steady-state infiltration rates were measured at four soil water
potentials h: � 0.05, � 0.2, � 0.6, and � 1.5 kPa. Flow was measured from � 1.5 to
� 0.05 kPa. The disc of the infiltrometer was positioned on the undisturbed surface
covered with a thin layer of sand to obtain a flat surface in the soil with maize. We gently
removed the upper root zone in the rotation and old pasture fields in order to place the disc
of the infiltrometer (respectively, 2 and 3 cm from the surface). The flow was measured at
a given potential for about 1 h to reach steady state. We used methylene blue in water (0.4
g l� 1) to dye the effective porosity at water potential h =� 0.05 kPa. We estimated the
unsaturated hydraulic conductivity curve at several tensions by computing multiple supply
potentials with the same disc, as proposed by Reynolds and Elrick (1991) and Ankeny et
al. (1991), assuming Wooding’s solution for three-dimensional axisymmetric infiltration
(Wooding, 1968). We estimated total porosity from measurements at the same sites by
weighing cylindrical samples having a volume of 250 cm3 (four replicates) assuming a
solid density of 2.65 g cm� 3.
2.4. Image analysis of macropore space
Pore space descriptions were made using undisturbed soil blocks (10� 10� 8 cm)
taken vertically beneath the locations of the infiltration measurements. Soil samples were
dried and impregnated with a polyester resin containing fluorescent dye (Ringrose-Voase,
1996). The blocks were then cut in four horizontal polished sections (7� 7 cm) at four
depths (1, 3, 5, and 7 cm). For each section, four areas (2.2� 3.1 cm) were analysed using
OPTIMAS software with a spatial resolution of 42 Am pixel� 1. We chose a spatial
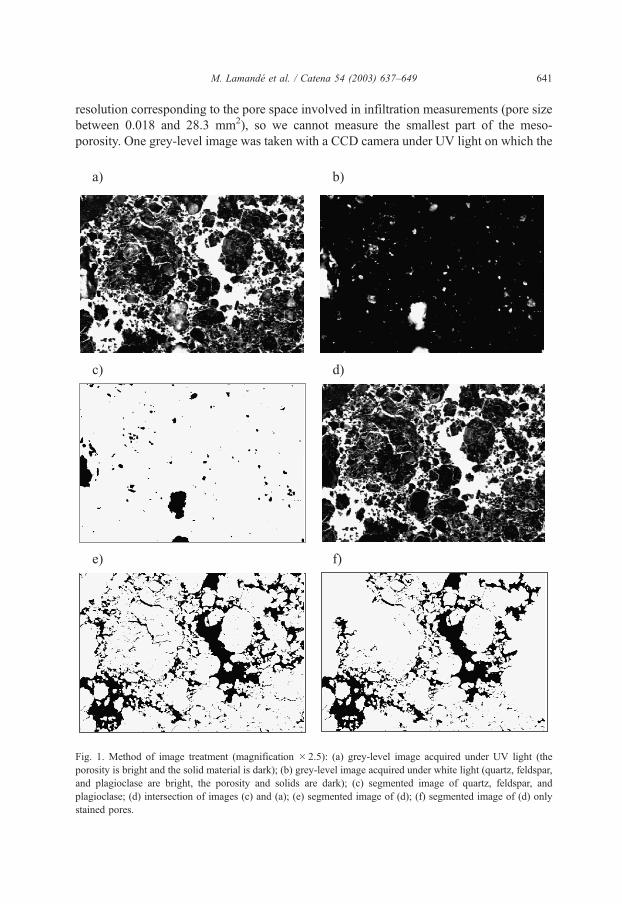
M. Lamande et al. / Catena 54 (2003) 637–649 641
resolution corresponding to the pore space involved in infiltration measurements (pore size
between 0.018 and 28.3 mm2), so we cannot measure the smallest part of the meso-
porosity. One grey-level image was taken with a CCD camera under UV light on which the
Fig. 1. Method of image treatment (magnification � 2.5): (a) grey-level image acquired under UV light (the
porosity is bright and the solid material is dark); (b) grey-level image acquired under white light (quartz, feldspar,
and plagioclase are bright, the porosity and solids are dark); (c) segmented image of quartz, feldspar, and
plagioclase; (d) intersection of images (c) and (a); (e) segmented image of (d); (f) segmented image of (d) only
stained pores.

Table 2
Pores classification according to size and shape
Shape classes Size classes (10� 6 m2)
1 2 3 4
[0.018; 0.031] [0.032; 0.196] [0.197; 1.77] >1.77
Tubular void Is< 38 T1 T2 T3 T4
Crack Isa[38; 89] C1 C2 C3 C4
Packing void Is>89 P1 P2 P3 P4
Is: elongation index.
M. Lamande et al. / Catena 54 (2003) 637–649642
solid phase appears dark and the porosity bright. Because some mineral fragments (quartz,
feldspar) appear bright under UV light, another grey-level image was taken under white
light on which the porosity is dark and coarse fragments are bright. We segmented and
inverted the image with coarse fragments and masked the coarse fragments on the grey-
level UV image. We then segmented the grey-level UV image into a binary image and
removed the porosity unaffected by methylene blue using a hand-made mask (Fig. 1). The
stained pores were classified from the final binary image according to their size and shape.
Pore size was measured from surface area on the binary image. Four size classes were
determined, corresponding to the effective pore size at the water potentials used for
infiltration measurement: from 0.018 to 0.031, from 0.032 to 0.196, from 0.197 to 1.77,
and up to 1.77 mm2. Pore shape was measured using the elongation index Is (perimeter2/
area), and three shape classes were determined to distinguish between tubular voids,
cracks, and packing voids (Hallaire and Curmi, 1994). The thresholds used for the size and
shape classes are given in Table 2.
3. Results and discussion
3.1. Earthworm community
Earthworm abundances are significantly different in the three treatments (Student’s t-
test, p = 0.05) (Fig. 2): the highest abundance of earthworms is found in the old pasture and
lowest in the maize; the increase of anthropic constraints is associated with a decrease in
earthworm abundance and species diversity. In the old pasture, the community is
dominated by endogeic earthworms (48%) and especially by Aporrectodea caliginosa
(Savigny, 1826) (juveniles and adults) and Allolobophora c. chlorotica (Savigny, 1826)
(adults). Anecic earthworms are also found (42%), especially Lumbricus friendi (Cognetti,
1904) (juveniles and adults). The low abundance of epigeic species in the old pasture
(10%) compared to the pasture phase of rotation (52%) may be explained by cattle
trampling (Cluzeau et al., 1992). Although land management conditions associated with
maize culture (tillage, pesticide use, and low organic matter return) affect all the
earthworm communities, the changes mainly concern the anecic (6%) and epigeic species
(4%); tillage may affect the largest individuals (anecic adults and juveniles, and endogeic
adults), while the soil cover may affect the epigeic species. In fact, the community under

Fig. 2. Earthworm communities in the studied three fields: (a) continuous maize, (b) pasture phase of rotation, (c)
old pasture.
M. Lamande et al. / Catena 54 (2003) 637–649 643
maize is dominated by endogeic species (90%), especially by A. caliginosa observed
mainly at the juvenile stage, while both juvenile and adult anecic are almost totally absent.
Pasture after maize especially favours the epigeic species (52%), which are dominated by
Lumbricus castaneus (Savigny, 1826) and Lumbricus rubellus castenoides (Bouche, 1972)
(observed at the adult stage), as well as anecic species (33%) dominated by L. friendi. This
specific structure could be explained by (1) the high rate of reproduction of the epigeic
species, (2) the maintenance of some earthworm species via the cocoons during the maize
phase, (3) the restauration of the anecic species due to better environmental conditions
(grass cover, no more tillage), and (4) the recolonisation of the site by exogenic
earthworms.
Thus, functional diversity is higher in the two pasture soils than under maize. Anecic
(juvenile and adult) and endogeic (juvenile and adult) species are present in the two
pastures, but there are more individuals of the three ecological groups in the old pasture
than under the rotation system.
3.2. Bulk density
The Student’s t-test on the mean at the 95% confidence interval shows that bulk density
is not significantly different between maize (1.21 g cm� 3) and the pasture phase of rotation
(1.26 g cm� 3) (Fig. 3). We expected a higher bulk density with rotation than maize due to
‘‘natural’’ compaction and the lack of tillage. The biological activity is more intense with
the rotation and could account for the better aggregation and porosity. Bulk density is
significantly higher in the old pasture (1.43 g cm� 3) (Fig. 3). We observed a compacted
layer on the top of the soil profile in the old pasture that was attributed to cattle trampling.
3.3. Near-saturated infiltration
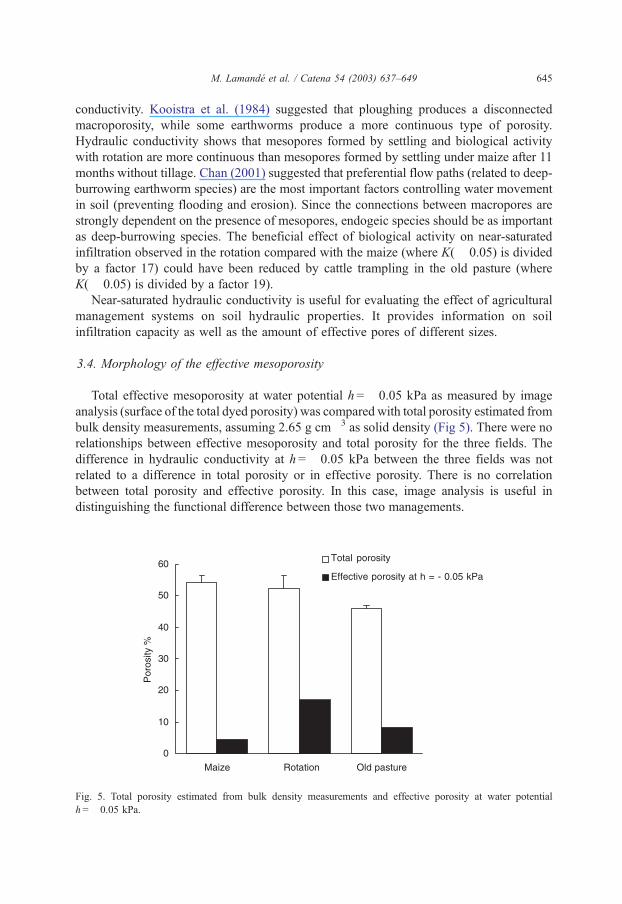
The results of near-saturated infiltration measurements are presented in Fig. 4. We
estimate a high hydraulic conductivity in the rotation fields at the two water potentials
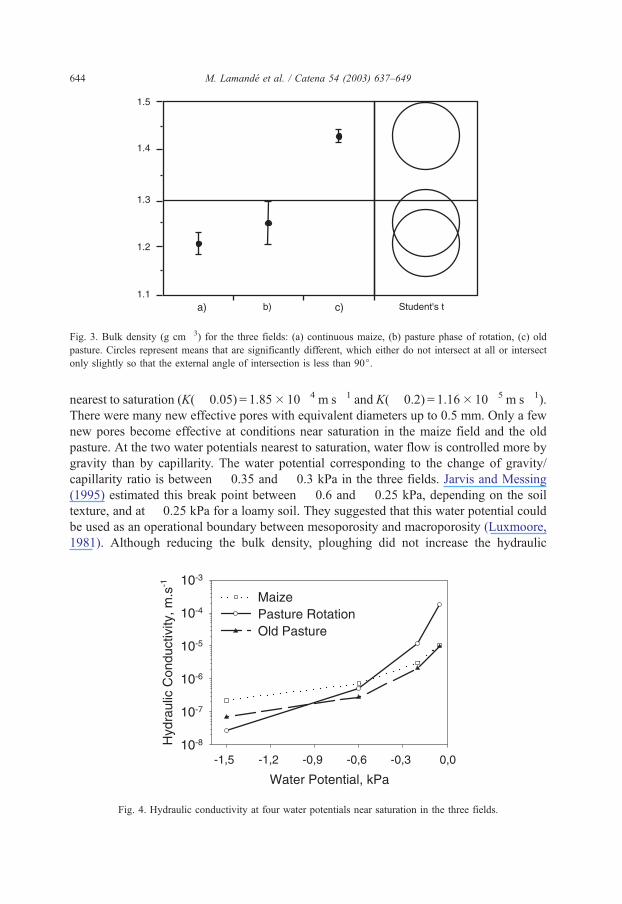
Fig. 3. Bulk density (g cm� 3) for the three fields: (a) continuous maize, (b) pasture phase of rotation, (c) old
pasture. Circles represent means that are significantly different, which either do not intersect at all or intersect
only slightly so that the external angle of intersection is less than 90j.
M. Lamande et al. / Catena 54 (2003) 637–649644
nearest to saturation (K(� 0.05) = 1.85� 10� 4 m s� 1 and K(� 0.2) = 1.16� 10� 5 m s� 1).
There were many new effective pores with equivalent diameters up to 0.5 mm. Only a few
new pores become effective at conditions near saturation in the maize field and the old
pasture. At the two water potentials nearest to saturation, water flow is controlled more by
gravity than by capillarity. The water potential corresponding to the change of gravity/
capillarity ratio is between � 0.35 and � 0.3 kPa in the three fields. Jarvis and Messing
(1995) estimated this break point between � 0.6 and � 0.25 kPa, depending on the soil
texture, and at � 0.25 kPa for a loamy soil. They suggested that this water potential could
be used as an operational boundary between mesoporosity and macroporosity (Luxmoore,
1981). Although reducing the bulk density, ploughing did not increase the hydraulic
Fig. 4. Hydraulic conductivity at four water potentials near saturation in the three fields.
M. Lamande et al. / Catena 54 (2003) 637–649 645
conductivity. Kooistra et al. (1984) suggested that ploughing produces a disconnected
macroporosity, while some earthworms produce a more continuous type of porosity.
Hydraulic conductivity shows that mesopores formed by settling and biological activity
with rotation are more continuous than mesopores formed by settling under maize after 11
months without tillage. Chan (2001) suggested that preferential flow paths (related to deep-
burrowing earthworm species) are the most important factors controlling water movement
in soil (preventing flooding and erosion). Since the connections between macropores are
strongly dependent on the presence of mesopores, endogeic species should be as important
as deep-burrowing species. The beneficial effect of biological activity on near-saturated
infiltration observed in the rotation compared with the maize (where K(� 0.05) is divided
by a factor 17) could have been reduced by cattle trampling in the old pasture (where
K(� 0.05) is divided by a factor 19).
Near-saturated hydraulic conductivity is useful for evaluating the effect of agricultural
management systems on soil hydraulic properties. It provides information on soil
infiltration capacity as well as the amount of effective pores of different sizes.
3.4. Morphology of the effective mesoporosity
Total effective mesoporosity at water potential h =� 0.05 kPa as measured by image
analysis (surface of the total dyed porosity) was compared with total porosity estimated from
bulk density measurements, assuming 2.65 g cm� 3 as solid density (Fig 5). There were no
relationships between effective mesoporosity and total porosity for the three fields. The
difference in hydraulic conductivity at h =� 0.05 kPa between the three fields was not
related to a difference in total porosity or in effective porosity. There is no correlation
between total porosity and effective porosity. In this case, image analysis is useful in
distinguishing the functional difference between those two managements.
Fig. 5. Total porosity estimated from bulk density measurements and effective porosity at water potential
h=� 0.05 kPa.
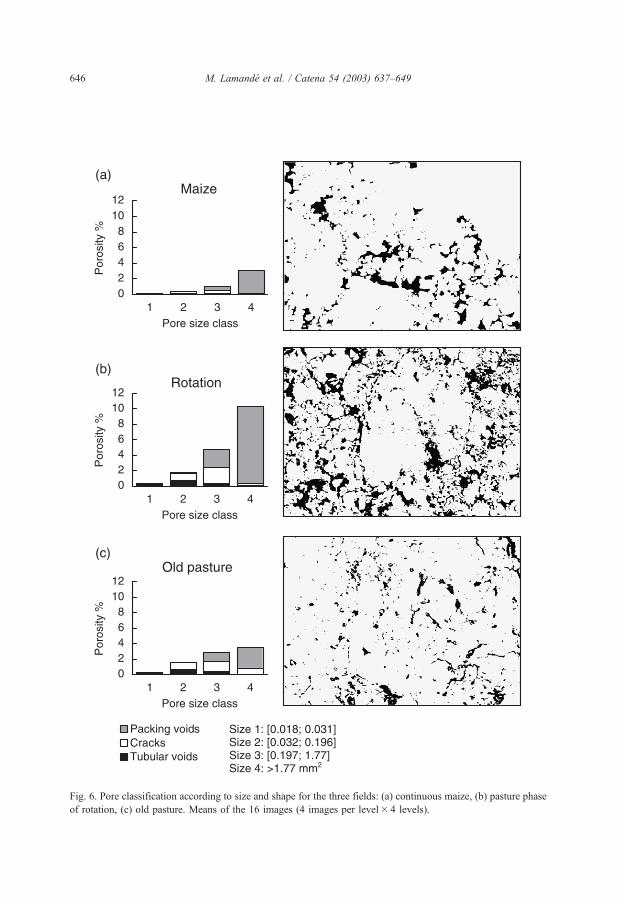
Fig. 6. Pore classification according to size and shape for the three fields: (a) continuous maize, (b) pasture phase
of rotation, (c) old pasture. Means of the 16 images (4 images per level� 4 levels).
M. Lamande et al. / Catena 54 (2003) 637–649646

M. Lamande et al. / Catena 54 (2003) 637–649 647
Effective pore size distribution increases sharply from size 1 to size 4 in the three
treatments (0.018 mm2 to more than 1.77 mm2, corresponding to efficient pores size at the
water potentials used for infiltration measurement) (Fig. 6). This increase was mainly due to
packing voids. Packing voids decrease in the old pasture compared with the rotation and
partly transformed into cracks due to cattle trampling (Fig. 6). There are more tubular voids
in the old pasture and in the rotation than in maize. Tubular voids are almost negligible in the
maize, where biological activity (roots and fauna) is lower. Although very few anecic
earthworms are present in the maize, they are responsible for most of the tubular voids larger
than 2mm in horizontal 2D sections. Such anecic species live in permanent vertical burrows.
The distribution of effective pores coincides well with the hydraulic conductivity curve.
Packing voids seem to have a major role in water flow at conditions near saturation (size
classes 3 and 4). At lower potentials, water flow is controlled by tubular voids and cracks.
The effective porosity at water potential h =� 0.05 kPa was two times less in maize than in
the old pasture, while the hydraulic conductivity was similar. In maize, most of the water
flow occurs through the pores of equivalent diameter smaller than 200 Am. These pores are
not all quantified by image analysis (42 Am pixel� 1 resolution) but are taken into account in
infiltration measurements at water potential h =� 1.5 kPa. These pores correspond to the
porosity observed within earthworm casts, but theymay correspond also to porosity between
enchytraeid casts (Dawod and FitzPatrick, 1993). In cultivated soil at low level of earthworm
densities, the enchytraeids can play an important role in creating a stable soil structure and
porosity (Topoliantz et al., 2000). But the high pH value in maize (6,2) is not favourable to
enchytraeid development (Gorny, 1984).
4. Conclusion
The types of agricultural management and earthworm community both induce changes
in the structural features and physical properties of soil. To propose sustainable agricultural
management systems, we need to improve our understanding of the processes controlling
these changes. In this study, we compare pore morphology, infiltration rate, and earthworm
community under three different agricultural managements. We show it is necessary to
consider the functional diversity of earthworms as well as their abundance. The packing
voids are important in controlling water flow and retention, especially when preferential
flow paths such as earthworm burrows or cracks are disconnected by tillage. The present
study provides a new methodology that may be used to assess the effects of agricultural
managements on soil structure and water movement. In particular, we consider the type of
earthworm community as a factor influencing the changes in soil physical properties.
References
Angulo-Jaramillo, R., Vandervaere, J.-P., Roulier, S., Thony, J.-L., Gaudet, J.-P., Vauclin, M., 2000. Fields
measurement of soil surface hydraulic properties by disc and ring infiltrometers: a review and recent develop-
ments. Soil and Tillage Research 55, 1–29.
Ankeny, M.D., Kaspar, T.C., Horton, R., 1990. Characterization of tillage and traffic effects on unconfined
infiltration measurements. Soil Science Society of America Journal 54, 837–840.

M. Lamande et al. / Catena 54 (2003) 637–649648
Ankeny, M.D., Ahmed, M., Kaspar, T.C., Horton, R., 1991. Simple field method for determining unsatured
hydraulic conductivity. Soil Science Society of America Journal 55, 467–470.
Azevedo, A.S., Kanwar, R.S., Horton, R., 1998. Effect of cultivation on hydraulic properties of an Iowa soil
using tension infiltrometers. Soil Science 163, 22–29.
Binet, F., Hallaire, V., Curmi, P., 1997. Agricultural practices and the spatial distribution of earthworms in maize
fields. Relationships between earthworm abundance, maize plants and soil compaction. Soil Biology &
Biochemistry 29, 577–583.
Blanchart, E., Bruand, A., Lavelle, P., 1993. The physical structure of casts of Millosonia anomala (Oligochaeta:
Megascolecidae) in shrub savanna soils (Cote d’Ivoire). Geoderma 56, 119–132.
Bouche, M.B., 1972. Lombriciens de France. Ecologie et systematique. INRA, Paris.
Bouche, M.B., 1977. Strategies Lombriciennes. In: Lohm, U., Persson, T. (Eds.), Soil Organisms as Components
of Ecosystems. Proc. 6th Int. Coll. Soil Zool. Ecol. Bull., Stockholm, pp. 122–132.
Bouche, M.B., Al-Addan, F., 1997. Earthworms, water infiltration and soil stability: some new assessments. Soil
Biology & Biochemistry 29, 441–452.
Capowiez, Y., Pierret, A., Daniel, O., Monestiez, P., Kretzschmar, A., 1998. 3D skeleton reconstructions of
natural earthworm burrow systems using CAT scan images of soil cores. Biology and Fertility of Soils 27,
51–59.
Chan, K.Y., 2001. An overview of some tillage impacts on earthworm population abundance and diversity:
implications for functioning in soils. Soil and Tillage Research 57, 179–191.
Chauvel, A., Grimaldi, M., Barros, E., Blanchart, E., Desjardins, T., Sarrazin, M., Lavelle, P., 1999. An Ama-
zonian earthworm compacts more than a bulldozer. Nature 398, 32–33.
Cluzeau, D., Binet, F., Vertes, F., Simon, J.C., Riviere, J.-M., Trehen, P., 1992. Effects of intensive cattle
trampling on soil-plant-earthworms system in two grassland types. Soil Biology & Biochemistry 24,
1661–1665.
Cluzeau, D., Cannavacciulo, M., Peres, G., 1999. Indicateurs macrobiologiques des sols: les lombriciens—
Methode d’echantillonnage dans les agrosystemes temperes. In: Institut Technique du Vin. 12eme Colloque
Viticole et Œnologique, Paris, pp. 25–35.
Cognetti, L., 1904. Lombricidi dei Pirenei. Bollettino dei Musei di Zoologia ed Anatomıa Comparata Torino 19
(476), 1–14.
Curmi, P., Merot, P., Roger-Estrade, J., Caneill, J., 1996. Use of environmental isotopes for field study of water
infiltration in the ploughed soil layer. Geoderma 72, 203–217.
Dawod, V., FitzPatrick, E.A., 1993. Some population sizes and effects of the Enchytraeide (Oligochaeta) on soil
structure in a selection of Scottish soils. Geoderma 56, 173–178.
Ehlers, W., 1975. Observations on earthworm channels and infiltration on tilled and untilled loess soil. Soil
Science 119, 242–249.
Gorny, M., 1984. Studies on the relationship between enchytraeides and earthworms. In: Szegi, J. (Ed.), Soil
Biology and Conservation of the Biosphere. Akademiai Kiado, Budapest, pp. 769–776.
Hallaire, V., Curmi, P., 1994. Image analysis of pore space morphology in soil sections, in relation to water
movement. In: Ringrose-Voase, A.J., Humphreys, G.S. (Eds.), Soil Micromorphology: Studies in Manage-
ment and Genesis. Developments in Soil Science, vol. 22. Elsevier, Amsterdam, pp. 559–567.
Heddadj, D., Gascuel-Odoux, C., 1999. Topographic and seasonal variations of unsatured hydraulic conductivity
as measured by tension disc infiltrometers at the field scale. European Journal of Soil Science 50, 275–283.
Jarvis, N.J., Messing, I., 1995. Near-saturated hydraulic conductivity in soils of contrasting texture measured by
tension infiltrometers. Soil Science Society of America Journal 59, 27–34.
Jegou, D., Hallaire, V., Cluzeau, D., Trehen, P., 1999. Characterisation of the burrow system of the earthworms
using X-ray computed tomography and image analysis. Biology and Fertility of Soils 29, 314–318.
Jones, C.G., Lawton, J.H., Shachak, M., 1994. Organisms as ecosystem engineers. Oikos 69, 373–386.
Kooistra, M.J., Bouma, J., Boersma, O.H., Jager, A., 1984. Physical and morphological characterization of
undisturbed and disturbed ploughpans in a sandy loam soil. Soil and Tillage Research 4, 405–417.
Kretzschmar, A., Aries, F., 1990. 3D images of natural and experimental earthworm burrow systems. Revue
d’Ecologie et de Biologie du Sol 27, 407–414.
Kribaa, M., Hallaire, V., Curmi, P., Lahmar, R., 2001. Effect of various cultivation methods on the structure and
hydraulic properties of a soil in a semi-arid climate. Soil and Tillage Research 60, 43–53.

M. Lamande et al. / Catena 54 (2003) 637–649 649
Lavelle, P., 1997. Faunal activities and soil processes: adaptative strategies that determine ecosystem function.
Advances in Ecological Research 27, 93–122.
Lee, K.E., 1959. The earthworms fauna of New Zealand. New Zealand Department of Scientific and Industrial
Research Bulletin, 130–382.
Lee, K.E., 1985. Earthworm. Their Ecology and Relationships with Soil and Land Use. Academic Press Aus-
tralia, Sydney, p. 411.
Lee, K.E., Foster, R.C., 1991. Soil fauna and soil structure. Australian Journal of Soil Research 29, 745–775.
Luxmoore, R.J., 1981. Micro-, meso-, macroporosity in soil. Soil Science Society of America Journal 45,
671–673.
Manichon, H., Roger-Estrade, J., 1990. Caracterisation de l’etat structural et etude de son evolution a court et
moyen terme sous l’action des systemes de cultures. In: Picard, D., Combe, L. (Eds.), Un Point sur Les
Systemes de Culture. INRA, Paris, pp. 27–55.
Meek, B.D., Rechel, E.R., Carter, L.M., DeTar, W.R., Urie, A.L., 1992. Infiltration rate of a sandy loam soil:
effects of traffic, tillage, and plant roots. Soil Science Society of America Journal 56, 908–913.
Paoletti, M.G., Sommaggio, D., Favretto, M.R., Petruzzelli, G., Pezzarossa, B., Barbafieri, M., 1998. Earthworms
as useful indicators of agroecosystem sustainability in orchards and vineyards with different inputs. Applied
Soil Ecology 10, 137–150.
Peres, G., Cluzeau, D., Curmi, P., Hallaire, V., 1998. Earthworms activity and soil structure changes due to
organic enrichments in vineyard systems. Biology and Fertility of Soils 27, 417–424.
Reynolds, W.D., Elrick, D.E., 1991. Determination of hydraulic conductivity using a tension infiltrometer. Soil
Science Society of America Journal 55, 633–639.
Ringrose-Voase, A.J., 1996. Measurement of soil macropore geometry by image analysis of sections through
impregnated soil. Plant and Soil 183, 27–47.
Savigny, J.C., 1826. Analyse des trav. acad. roy. sci. pendant l’annee 1821, partie physique. In: Cuvier, Memoires
de l’Academie Royale des Sciences de l’Institut de France 5, 176–184.
Shipitalo, M.J., Protz, R., 1989. Chemistry and micromorphology of aggregation in earthworm casts. Geoderma
45, 357–374.
Sveistrup, T.E., Haraldsen, T.K., Engelstad, F., 1997. Earthworm channels in cultivated clayed and loamy
Norwegian soils. Soil and Tillage Research 43, 251–262.
Topoliantz, S., Ponge, J.F., Viaux, P., 2000. Earthworm and enchytraeid activity under different arable farming
systems, as exemplified by biogenic structures. Plant and Soil 225, 39–51.
Trojan, M.D., Linden, D.R., 1998. Macroporosity and hydraulic properties of earthworm-affected soils as influ-
enced by tillage and residue management. Soil Science Society of American Journal 62, 1687–1692.
Wooding, R.A., 1968. Steady infiltration from a shallow circular pond. Water Resources Research 4, 1259–1273.
Copyright © 2022 FDOKUMEN




















