
The GenLOT: generalized linear-phase lapped orthogonal transform

Developmental Brain Research, 4 (1982) 91-101 91 Elsevier Biomedical Press
Changes in Number and Distribution of Orthogonal Arrays during Postnatal Muscle Development
C. SUE HUDSON, BEVERLY K. DYAS and JOHN E. RASH
Department of Pathology, V.A. Medical Center, anJ ( B.K.D.) Department of Neurology, University of Maryland School of Medicine, 10 S. Pine Street, Baltimore, MD 21201, and (J.E.R.) Department of Anatomy
Colorado State University, Ft. Collins, CO 80523 (U.S.A.)
(Accepted October 12th, 1981)
Key words: orthogonal arrays - - muscle development - - sarcolemrna - - fiber type - - innervation pattern - - freeze-fracture
Quantitative freeze-fracture electron microscopy was utilized to study the changes in number and distribution of orthogonal arrays (aggregates of 6-7 nm particles) of the sarcolemmas of the fast twitch extensor digitorum longus (EDL) and slow twitch soleus muscles during the first few weeks of postnatal development in the rat. In the adult rat, orthogonal arrays are present in high densities in the fast twitch type II fibers but only in low densities in slow twitch type I fibers. In this study, the changing histochemical profiles of fiber types in the EDL and soleus muscles were also determined for the first month of postnatal development and correlated with the changing number and distribution of orthogonal arrays during the same time frame.
At day 3 postpartum, EDL and soleus fibers possessed few orthogonal arrays. The developing EDL fibers rapidly acquired additional orthogonal arrays until the approximate adult number and distribution were attained at postpartum day 25. In contrast, the slow twitch soleus fibers rapidly acquired orthogonal arrays and type IIA fibers until day 35 when both were in excess of adult values. Subsequently, the number of arrays and type IIA fibers declined to normal adult ranges. We suggest that the patterns of de- velopment of orthogonal arrays and fiber types are different in the EDL and soleus because the types of innervating motor units are different in the two muscles. The EDL is innervated almost entirely by fast motor units throughout early development and ma- turity. The soleus, however, is initially innervated by a more heterogeneous population of motor neurons. Thus, during the period of polyneuronal innervation which occurs normally during the first few weeks of postnatal development, many individual soleus fibers may possess simultaneous innervation by axons from different motor neuron types. These dual influences may be responsible for the irregular pattern of development of orthogonal arrays and type IIA fibers in developing soleus fibers. Later, as the adult pattern of monosynaptic innervation is developed, expected adult values of orthogonal arrays and fiber types are attained.
INTRODUCTION
Freeze-f rac ture analysis o f p l a sma membranes of
many cell types has revealed the presence o f or tho-
gonal aggregates or square ar rays o f 6-7 nm intra-
m e m b r a n o u s particles. These structures have been
repor ted in var ious cell types including hepatocytes
(ref. 31), gastric par ie ta l cells 4, t rachea 2s, k idney
collect ing tubules 27, astrocytes13,sa,34, ependymal
cells 5, satelli te cells o f spinal ganglia14, 35, neurogl ia
o f the h y p o t h a l o m o - n e u r o p h o p h y s i a l system, and
skeletal muscle22, 86. A l though the precise funct ion
o f o r thogona l a r rays has not been established, it has
recent ly been suggested tha t the ar rays may funct ion
in the regula t ion o f cer tain ion concent ra t ions in
interst i t ia l fluids 21. Cer ta in o ther facts abou t these
intr iguing i n t r amembranous part icles are known.
Several repor ts have es tabl ished that the o r thogona l
a r rays apparen t ly possess a character is t ic densi ty in
each given m e m b r a n e and that a r rays are labi le in
given membranes under var ious metabol ic condi-
tions15A7, 34. F o r instance, freeze-fracture replicas o f
m a m m a l i a n muscle sa rco lemmas reveal high densi-
ties of o r thogona l a r rays in fast twitch muscle fibers
but only low densities in slow twitch fibers 17. How-
ever, the slow twitch soleus muscle acquires a high
densi ty o f o r thogona l a r rays and undergoes a l tera-
t ions in cer tain his tochemical and e lect rophysiologi-
cal character is t ics after exper imenta l cross-reinner-
va t ion with the phasic nerve f rom the fast twitch
E D L musc le lL Thus, neurona l influences may regu-
late o r thogona l a r ray densities and several histo-
O165-3806/82/0000-0000f$02.75 (~ l~.l~,~.,,i,~,- rt;,~,~.,~;oot D . . . .

92
chemical and electrophysiological properties in a- dult muscle fibers ~5. To test this hypothesis in younger animals, we chose to study the number and distribution of orthogonal arrays during the first few weeks of postnatal muscle fiber development in the rat hind limb.
The experiments reported here were designed to compare the number and distribution of orthogonal arrays in developing fast twitch EDL and slow twitch soleus fibers. Specifically, the two muscles were analyzed to determine the pattern of addition of orthogonal arrays, and the presence of arrays in relation to the presence of type II (fast twitch) fibers.
MATERIALS AND METHODS
Tissue preparation for freeze-fracture Albino rats of 3-130 days postpartum (2-3 ani-
mals per developmental age) were anesthetized and fixed by whole body perfusion through the left ventricle with oxygenated 2 . 5 ~ gtutaraldehyde in rat Ringer's solution. The EDL and soleus muscles were removed, and stained for cholinesterase activi- ty to visualize the neuromuscular junctions~9, 36. In the normal adult EDL, the numerical density of square arrays varies as a function of distance from the neuromuscular junction 16. Since many of the developing muscle fibers analyzed in the present study had a total length of approximately l ram, samples from animals 25 days and younger necessa- rily included as much as the entire muscle, while samples from animals over 25 days of age were composed of fiber bundles dissected 0.5-1.0 mm from stained neuromuscular junctions.
After fixation, dissected muscle fiber bundles (0.5 mm on a side) were infiltrated with 30 ~ glycerol, frozen on gold specimen supports in Freon 12 main- tained at its freezing point (--150 °C), cleaved in a Balzers 360M freeze-etch device, replicated with platinum using an electron beam gun, and coated with carbon. Replica thickness was monitored and regulated by a quartz crystal thin film monitor. Specimens were thawed in rat Ringer's solution, the tissues digested with Clorox bleach and/or chromic acid solutions and the replicas picked up o n 200 or 400 mesh grids for EM analysis: Details of our methods have been described previously 2~'z6.
All EM samples were viewed on a Siemens Elmis-
kop 101 or JEOL 100B electron microscope. A randomly chosen portion of the sarcolemma of each replicated muscle fiber was photographed at an original magnification of 20.000 and printed al a final magnification of 50.000 or 60.000 Quan- titative data concerning the percentage of fibers possessing orthogonal arrays and the density (~f square arrays per #m 2 represent data collected from at least 2 animals per developmental age. We analy- zed 15-25 EDL fibers and 15-25 soleus fibers or approximately 180-300 um z total sarcolemmonal area per muscle type per dcvclopmental age. To establish the number of orthogonal arrays per square micrometer, all arrays wcrc circled with a felt tip pen. An appropriate acetate sheet calibrated to indicate square micrometers at 50.000 or 60.000
was placed over each nhcrograph and the ortho- gonal arrays appearing m each grid square represen- ting 1 #m 2 area were counted. Although this method of tabulation is somewhat different than the avera- ging method of Ellisman and co-workers ~s.~7 ̀direct comparison of data obtained b.v the two techniques using adult rat myofibers, revealed no major discre- pancies.
Tissue preparation_for histoehemical anatysi, s To assess postnatal fiber type differentiation be-
tween day 3 and day 130. both EDL and soleus muscles were characterized by routine analysis of myosin ATPase activity and NADH-TR (nicotina- mide adenine dinucleotide tetrazolium reductase) stain. Two to three postpartum rats per develop- mental age studied were anesthetized, and the EDL and soleus muscles were removed, surrounded by cryostat embedding medium, frozen in ~sopentane cooled to --170 °C in liquid nitrogen, and sectioned (10/zm thickness) at - -30 ~C in a Harris cryostat. Consecutive serial sections were examined after stai- ning for N A D H -TR and calcium activated myosin ATPase activity 19. Type I fibers stain darkly with the ATPase procedure (preincubated at pH 4.35) and have slow contraction times while type lI fibers stain lightly and have fast contraction times 6,a~". Type II fibers can be further categorized into type IIA fibers which are oxidative glycolytic, type lIB fibers which are glycolytic, and type ]IC fibers which are tess welt-characterized physiologically and biochemical- ly 6,:~. Type IIC fiber are considered undifferentiated

93
fibers which decrease in number with age. Although fiber typing is a controversial issue, particularly in developing muscle 2°, the procedure is used for this study as a convenient indicator of fiber types. An explanation for the ambiguity has been provided by Gauthier and co-workers who demonstrated the presence of both fast and slow isoenzymes of myosin in single myofibers during early postnatal develop- ment TM.
Sections were viewed and photographed with Polaroid Type 665 Positive-Negative film using a Nikon SKE microscope. Individual fibers were cha- racterized as types I, IIA, lIB, or IIC utilizing the 3 different staining methods. The relative frequency of each fiber type was determined in each muscle sample.
RESULTS
Membrane ultrastructure
Examination of freeze-fracture replicas of sarco- lemmas of selected ages of EDL and soleus muscles revealed changing numbers and distributions of orthogonal arrays throughout early development. A micrograph of a day 3 EDL fiber (Fig. la) reveals a low density of arrays. In comparison, the sarcolem- ma of a day 25 EDL fiber possesses a noticeably
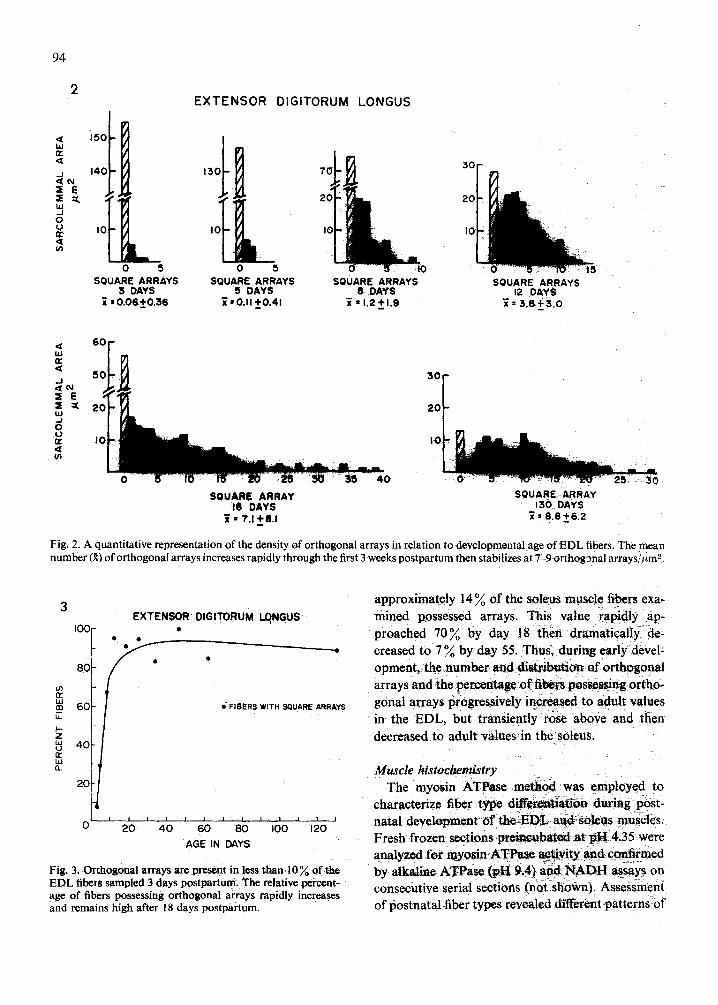
higher density of the macromolecular aggregates (Fig. lb). In the EDL, the density of orthogonal arrays per square micrometer of sarcolemmal area increased systematically during maturation. The quantified data appear in Fig. 2. At day 3, orthogo- nal arrays were present at a low density of 0.1//zmL This value rapidly increased to 9//~m 2 by day 25 and remained at this approximate level through day 130. Similarly, the percentage of fibers with significant numbers of orthogonal arrays rapidly increased from approximately 7 % at day 3 to approximately 90 ~ at day 18 (Fig. 3). The proportion of array-con- taining fibers remained high (above 83 %) from day 18 through day 130 wherein 90 ~ of the fibers analy- zed possessed orthogonal arrays.
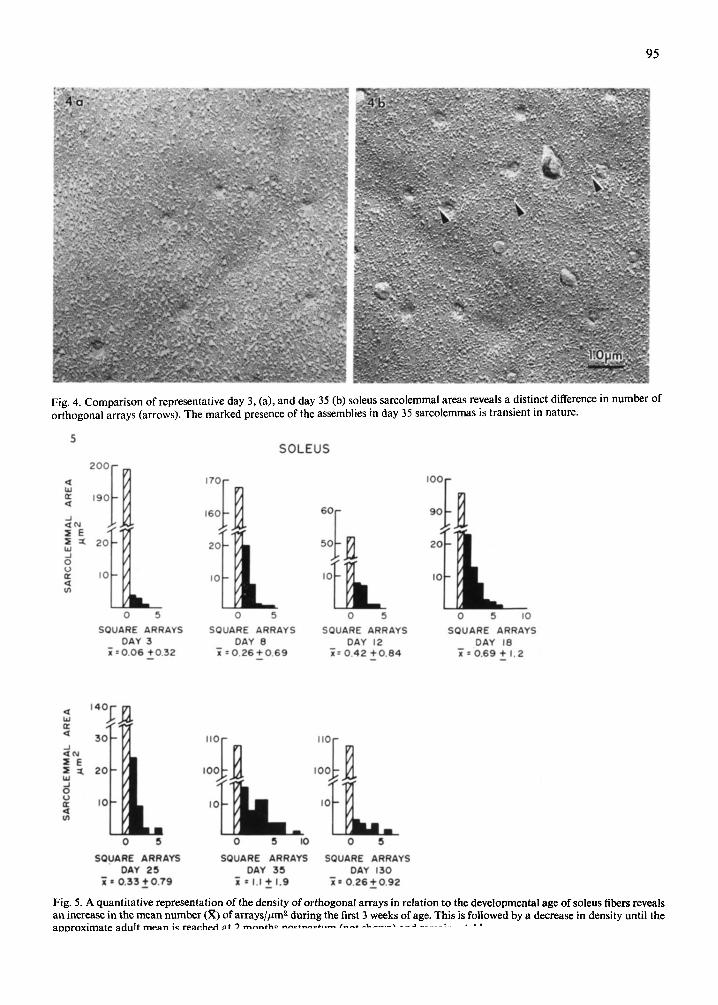
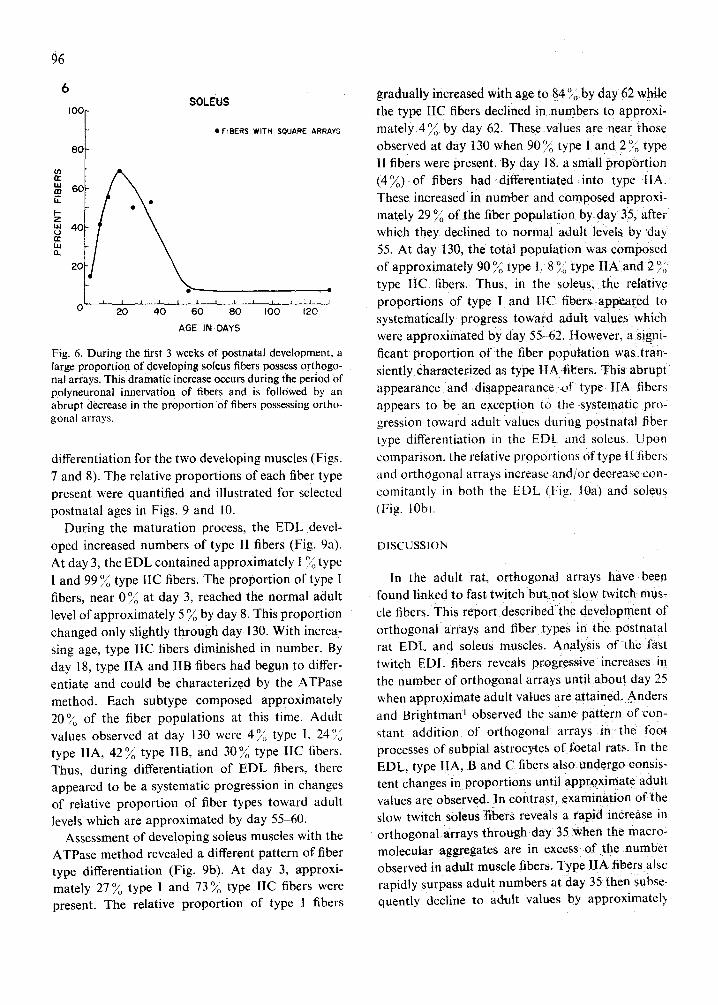
Analysis of data collected from developing soleus sarcolemmal preparations revealed fluctuations in the distribution and density of orthogonal arrays. Fig. 4a and b illustrate a low density of arrays in a 3 day soleus fiber and a somewhat higher density in a day 35 fiber. Upon quantification of the data, the increased density was found to be only transient in nature (Fig. 5). The relative number of orthogonal arrays increased from 0.1//tm 2 at day 3 to 1.0//zm 2 at day 35 then decreased to 0.3//~m z by day 130. A similar transient increase was apparent in the distri- bution of orthogonal arrays (Fig. 6). At day 3
Fig. 1. Representative replicas of P-face images of(a) day 3 and (b) day 25 EDL sarcolemmal membranes. The difference in density of orthogonal arrays (arrows) is obvious. Specific numerical data are presented in Fig. 2.
94
2 EXTENSOR DIGITORUM
=
, ,
o 5 o 5 SQUARE ARRAYS SQUARE ARRAYS
3 DAYS 5 DAYS = 0.06 ±0 .36 ~ • 0.I I ~0.41
LONGUS
SQUARE ARRAYS 8 DAYS
~ • 1.2+1.9
:50
20
I0
SQUARE ARRAYS t2 DAYS
~= 3,8 + ~ . o
I
< 6 0 UJ rr (Z
5 0 . J
:S :~ 2 0 tzJ ,.J o r,, I0 Ul
3C
2(
I1
0 5 10 15 1'0 ~ 3 0 35 4 0
SQUARE ARRAY SQUARE ARRAY 18 DAYS 130 DAYS • 7.1+8.1 ~= 8 . 8 ~ 6 . 2
D
Fig. 2. A quantitative representation of the density of orthogonal arrays in relation to developmental age of EDL fibers. The mean number (~) of orthogonal arrays increases rapidly through the first 3 weeks postpartum then stabilizes at 7-9 orthogonal arTays//~m '~.
3
{11 m t,
Z
r ~ LU I 1
EXTENSOR DIGITORUM LQNGUS
100180 ~ ' ~ ' e • •
°°[ 40
20
_/
/ FIBERS WITH SQUARE ARRAYS
I I I / I I I I I I I l 0 20 40 60 80 I00 120
AGE IN DAYS
Fig. 3. Orthogonal arrays are present in less than 10% of tt~ EDL fibers sampled 3 days postpartum. The relative percent- age of fibers possessing orthogonal arrays rapidly increases and remains high after 18 days postpartum.
approximately 14 70 of the soteus mtLsete fibers exa- mined possessed arrays. This value rapidly ap- proached 7070 by day 18 then dramatically de- creased to 7 ~o by day 55. Thus, during early devel- opment, the number and distribution of orthogonal arrays and the percentase o f fibers,possessing ortho- gonal arrays progressively increased to adult values in the EDL, but transiently rose above and then decreased to adult values in the soteus.
Muscle histochemistry The myosin ATPase method was employed to
characterize fiber type differentiation dur ing post- natal development of t h e E D L ~ s o l e u s muscles. Fresh frozen sections ~ u b ~ t e d a t pt~I 4.35~were analyzed for myos iaATPaae ~a~tiyit~ a~._d:coniirmed by alkaline ATPase (pH 9A) and ~NADH assays on consecutive serial sections (nOt rshown). Assessment of postnatal fiber types revealed different patterns o f
95
Fig. 4. Comparison of representative day 3, (a), and day 35 (b) soleus sarcolemmal areas reveals a distinct difference in number of orthogonal arrays (arrows). The marked presence of the assemblies in day 35 sarcolemmas is transient in nature.
5 S O L E U S
200 - i ,~o- ' ~
~o i it ~ 160 60
L ~ I0 IO I
0 5 0 5 0 5 0 5 I0
SQUARE ARRAYS SQUARE ARRAYS SQUARE ARRAYS SQUARE ARRAYS DAY 3 DAY 8 DAY 12 DAY 18
~--0.06 +0.32 ~ : 0 .26+0.69 3-- 0.42 +0.84 "~" = 0.69 + 1.2
o °F L I I I
0 5 0 5 I0 0 5
SQUARE ARRAYS SQUARE ARRAYS SQUARE ARRAYS DAY 25 DAY 35 DAY 130
; = 0 .33 t"0 .79 ; = I.I __ 1.9 ; = 0.26_+0.92
Fig. 5. A quantitative representation of the density of orthogonal arrays in relation to the developmental age of soleus fibers reveals an increase in the mean number (R) of arrays/#m z during the first 3 weeks of age. This is followed by a decrease in density until the aooroximate adult mean is rearhad at 9 m a n f h ~ n ~ ¢ f n a ~ t , , ~ / ' - - - - * ^ I L . . . . . X - ~ J . . . . r , • •
96
6
I00 f
(/3 n.- W m 6 0 LL
I.- Z Lq 40
w (1.
2C
SOLEU$
• FIBERS WITH SQUARE ARRAYS
\\\ \
i i i i i ~ i 20 40
?
0 I00 12o AGE IN DAYS
Fig. 6. During the first 3 weeks of postnatal development, a large proportion of developing soleus fibers possess orthogo- nal arrays. This dramatic increase occurs during tile period of polyneuronal innervation of fibers and is followed by an abrupt decrease in the proportion of fibers possessing ortho- gonal arrays.
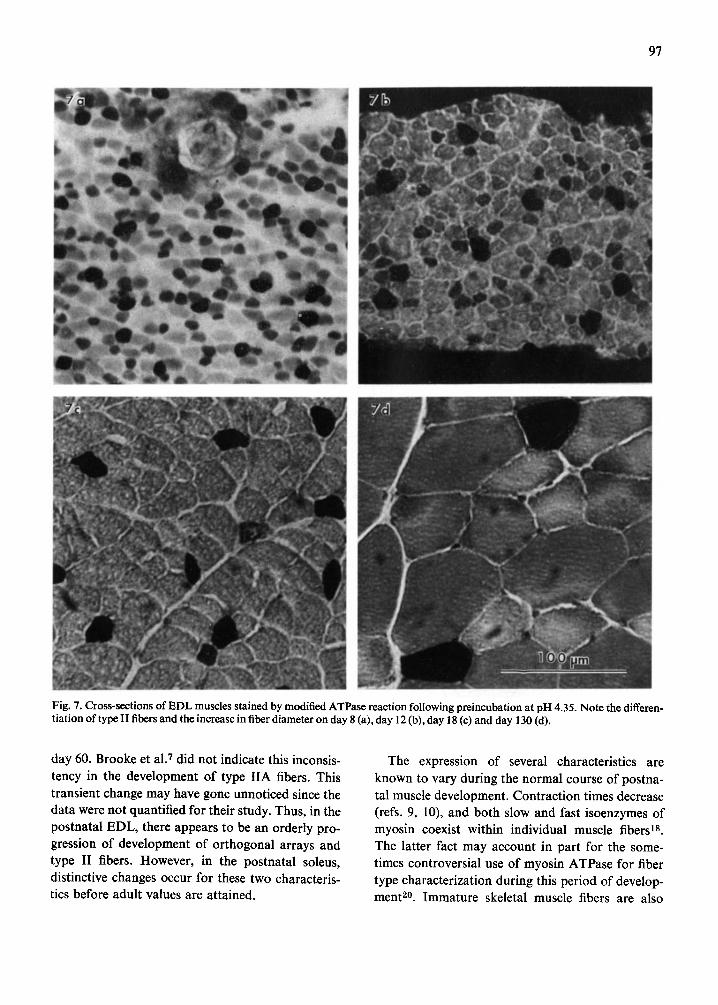
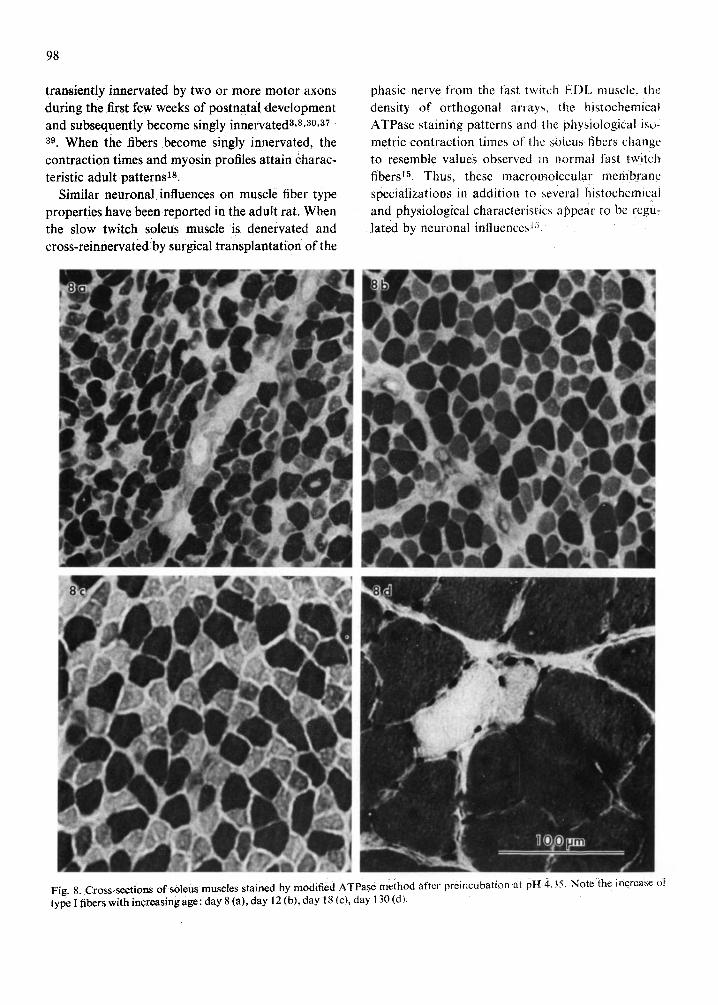
differentiation for the two developing muscles (Figs. 7 and 8). The relative proportions of each fiber type present were quantified and illustrated for selected postnatal ages in Figs. 9 and 10.
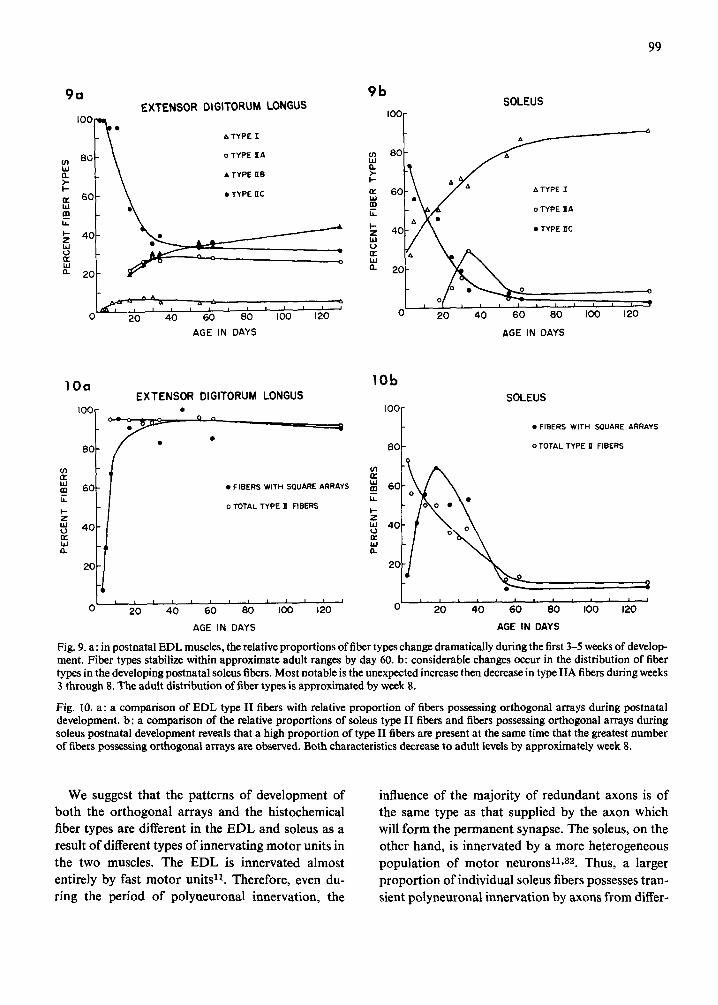
During the maturation process, the EDL devel- oped increased numbers of type II fibers (Fig. 9a). At day 3, the EDL contained approximately 1% type 1 and 99 % type I1C fibers. The proportion of type I fibers, near 0 % at day 3. reached the normal adult level of approximately 5 ~ by day 8. This proportion changed only slightly through day 130. With increa- sing age, ~type I IC fibers diminished in number. By day 18. type IIA and liB fibers had begun to differ- entiate and could be characterized by the ATPase method. Each subtype composed approximately 2 0 ~ of the fiber populations at this time. Adult values observed at day 130 were 4°/,', type I. 24';~, type IIA. 4 2 ~ type liB. and 30~, q type IIC fibers. Thus. during differentiation of EDL fibers, there appeared to be a systematic progression in changes of relative proportion of fiber types toward adult levels which are approximated by day 55-60.
Assessment of developing soleus muscles with the ATPase method revealed a different pattern of fiber type differentiation (Fig. 9b). At day 3. approxi- mately 27 ~ type 1 and 73 ~ type IIC fibers were present. The relative proportion of type ! fibers
gradually increased with age to 84 % by day 62 while the type IIC fibers declined in numbers to approxi- mately 4 7 / b y day 62. These values are near those observed at day 130 when 90% type I and 2 ')~; type ii fibers were present. By day 18. a small proportion {4%) of fibers had differentiated i n to type tlA. These increased in number and composed approxi- mately 29 % of the fiber population by day 35, after which they declined to normal adult levels by day 55. At day 130, the total population was composed
o , o of approximately 90 ~ type 1, /; type I1A and 2/,, type IIC fibers. Thus. in the soleus, the relative proportions of type I and IlC fibers appeared to systematically progress toward adult values which were approximated by day 55-62. However, a signi- ficant proportion of the fiber population was tran- siently characterized as type t lA fii~ers. This abrupt appearance and disappearance of type IIA fibers appears to be an exception to the systematic pro- gression toward adult values during postnatal fiber type differentiation in the EDL and soleus. Upon comparison, the relative proportions of type il fibers and orthogonal arrays increase and or decrease con- comitantly in both the EDL (Fig, lOa) and soleus
( Fig. lObl
DISCUSSION
In the adult rat. orthogonal arrays have been found linked to fast twitch but not slow twitch mus- cle fibers. This report described the development of orthogonal arrays and fiber types in the postnatal rat E DL and soleus muscles. Analysis of the fast twitch EDL fibers reveals progressive increases in the number of orthogonal arrays until about day 25 when approximate adult values are attained. Anders and Brightman 1 observed the same pattern of con- stant addition of orthogonal arrays in the foot processes of subpial astrocytes of foetal rats. In the EDL. type IIA, B and C fibers also undergo consis- tent changes in proportions until approximate adult values are observed. In contrast, examination of the slow twitch soleus fibers reveals a rapid increase in orthogonal arrays through day 35 when the macro- molecular aggregates are in excess of the number observed in adult muscle fibers. Type IIA fibers alsc rapidly surpass adult numbers at day 35 then subse. quently decline to adult values by approximateb
97
Fig. 7. Cross-sections of EDL muscles stained by modified ATPase reaction following preincubation at pH 4.35. Note the differen- tiation of type II fibers and the increase in fiber diameter on day 8 (a), day 12 (b), day 18 (c) and day 130 (d).
day 60. Brooke et al. 7 did not indicate this inconsis- tency in the development of type IIA fibers. This transient change may have gone unnoticed since the data were not quantified for their study. Thus, in the postnatal EDL, there appears to be an orderly pro- gression of development of orthogonal arrays and type II fibers. However, in the postnatal soleus, distinctive changes occur for these two characteris- tics before adult values are attained.
The expression of several characteristics are known to vary during the normal course of postna- tal muscle development. Contraction times decrease (refs. 9, 10), and both slow and fast isoenzymes of myosin coexist within individual muscle fibers is. The latter fact may account in part for the some- times controversial use of myosin ATPase for fiber type characterization during this period of develop- ment 2°. Immature skeletal muscle fibers are also
98
transiently innervated by two or more motor axons during the first few weeks of postnatal development and subsequently become singly innervated a,s,a0,a7- a9 When the fibers become singly innervated, the contraction times and myosin profiles attain charac- teristic adult patterns is.
Similar neuronal influences on muscle fiber type properties have been reported in the adult rat. When the slow twitch soleus muscle is denervated and cross-reinnervated by surgical transplantation of the
phasic nerve from the fast twttch EDL muscle, the density of orthogonal array-, the histochemical ATPase staining patterns and the physiological iso- metric contraction times of the ~,oleus tibers change to resemble values observed in i~ormal fast twitch fibers 15. Thus. these macroalolecular membrane specializations in addition to several histochemical and physiological characteristics appear to be regu- lated by neuronal influences a'~'
99
9a
IOO
¢n 8G bJ O. >- I-- ¢: 60 LU m U-
I-- Z bJ U (F.. bJ 0.
E X T E N S O R D IG ITORUM LONGUS
& TYPE 'I.
o TYPE I A
• TYPE HB
• TYPE t i c
4 0
0 2 0 4 0 6 0 8 I 0 0
A G E IN D A Y S
o
i i
120
9b IOC
tn 8C i , i Q. 3- F-
n. 60 bJ (D
E t - z 40 td 0 E LU
a. 20
SOLEUS
? 0 I00 120
~ YPE I
YPE I A
YPE XC t
20 40 6 0 80
AGE IN DAYS
lOa
I00,
80
W m 60 E F- Z tu 4 0 r r bJ O.
20
E X T E N S O R DIGITORUM LONGUS
° °
• FIBERS WITH SQUARE ARRAYS
o TOTAL TYPE 11 FIBERS
2'0 ~ ~0 = ~0 ' ' ' ' ' ' 4 6 80 I00 12O
AGE IN DAYS
lOb SOLEUS
tO0 f ~ . . . . ~ • FIBERS WITH SOUARE ARRAYS
~0~- o TOTAL TYPE D" FIBERS
,'t,'
~ 60 h •
I-- Z
" ' 4 0
W
2O
' 0 ' 2 eO I00 120
AGE IN DAYS
Fig. 9. a: in postnatal EDL muscles, the relative proportions of fiber types change dramatically during the first 3-5 weeks of develop- ment. Fiber types stabilize within approximate adult ranges by day 60. b: considerable changes occur in the distribution of fiber types in the developing postnatal soleus fibers. Most notable is the unexpected increase then decrease in type IIA fibers during weeks 3 through 8. The adult distribution of fiber types is approximated by week 8.
Fig. 10. a: a comparison of EDL type II fibers with relative proportion of fibers possessing orthogonal arrays during postnatal development, b: a comparison of the relative proportions of soleus type II fibers and fibers possessing orthogonal arrays during soleus postnatal development reveals that a high proportion of type II fibers are present at the same time that the greatest number of fibers possessing orthogonal arrays are observed. Both characteristics decrease to adult levels by approximately week 8.
We suggest that the patterns of development of both the orthogonal arrays and the histochemical fiber types are different in the EDL and soleus as a result of different types of innervating motor units in the two muscles. The EDL is innervated almost entirely by fast motor units 11. Therefore, even du- ring the period of polyneuronal innervation, the
influence of the majority of redundant axons is of the same type as that supplied by the axon which will form the permanent synapse. The soleus, on the other hand, is innervated by a more heterogeneous population of motor neurons zl,32. Thus, a larger proportion of individual soleus fibers possesses tran- sient polyneuronal innervation by axons from differ-

100
ent motor neuron types. This could result in the
s imul taneous influence of more than one type of
rr~otor neuron on individual muscle fibers. We sug-
gest that variat ions in or thogonat array numbers
and in histochemical staining pat terns observed in
the soleus dur ing the first few weeks of postnatal
development are due to influences supplied by differ-
ent motor neuron types as a result of polyneuronal
innervat ion. Fur thermore . in the period following
the loss of polyneuronal innerva t ion and the esta-
b l ishment of monosynapt ic innervat ion, the soleus
muscle cont inues to adjust to the growth of the
animal 3z. Dur ing this prolonged period, type 1I fiber
units decrease from 33 to 10°~ while type i fiber
units increase from 67 to 90 ~a2. Thus, the period of
changing neuronal influence on individual soleus
fibers is correlated with the gradual decrease in
orthogonat arrays and the comirmed alterations ,~I
histochemical and physiologicai fiber type proper-
ties.
ACKNOWLEI)GEMENTS
We are grateful to Dr. Lloyd Guth for his helpful
suggestions dur ing the preparat ion of this mantl-
script and to Mr. R. C Wierwillc for his technical
assistance. Some of the work included in this report
was performed in 1978-79 while Drs. Hudson and
Rash were members of the Depar tment of Pharma-
cology and Experimental Therapeutics at the Uni-
versity of Mary land School of Medicine in Balti-
more. MD.
This work was supported by N S F G r a n t BNS-78-
12317 (C.S.H.) and a Muscular Dystrophy Associa-
t ion grant- in-aid (J.E.R.).
REFERENCES
l Anders, J. J. and Brightman, M. W., Assemblies of parti- cles in the cell membranes of developing, mature and reactive astrocytes, J. NeurocytoL, 8 (1979) 777-795.
2 Anders, J. J. and Pagmanelli, D. M., The protein nature and arrangement of intramembranous partiele assemblies in normal and reactive astrocytes, Anat. Rec., 193 (1979) 470 (Abstr.).
3 Bennett, M. R. and Pettigrew, A. G., The formation of synapses in reinnervated and cross-reinnervated striated muscle during development. J. Physiol. (Load.), 241 (1974) 547-573.
4 Bordi, C. and Perrelet, A., Orthogonal arrays of particles in plasma membranes of the gastric parietal cell, Anat. Rec., 192 (1978) 297-304.
5 Brightman, M, W., Prescott, L. and Reese, T. S., Intercel- lular junctions of special ependyma. In K. M. Knigge, D. E. Scott, H. Kobayaski and S. Ishii (Eds.), Brain-Endo- crine Interaction. IL The Ventricular System, Karger, Basel, 1975, pp. 146-165.
6 Brooke. M. H. and Kaiser, K. K., Muscle fiber types: how many and what kind?, Arch. NeuroL, 23 (1970) 369-379.
7 Brooke, M. H., Williamson, E. and Kaiser, K. K., The behavior of the four principle muscle fiber types in the developing rat and in reinnervated muscle, Arch. NeuroL. 25 (1971) 360-366.
8 Brown, M. C., Jansen. J. K. S. and Van Essen, D., Poly- neuronal innervation of skeletal muscle in new-born rats and its elimination during maturation, J. Physiol. (Lond.), 261 (1976) 387-422.
9 Buller, A. J., Eccles, J. C. and Eccles, R. M., Differentia- tion of fast and slow muscles in the cat hind limb, J. Phy- siol. (Lond.), 150 (1960) 399-416.
10 Close, R., Dynamic properties of fast and slow skeletal muscles of the rat during development, J. PhysioL (Load.), 173 (1964) 74-95.
11 Close, R., Properties of motor units in fast and slow mus-
cles of the rat, J. Physiol. (Land.), 193 {1967) 45 55. 12 Close, R., Dynamic properties of mammalian skeletal
muscles, Physiol. Rev., 52 (1972) 129-197. 13 Dermietzel, R., Junctions in the central nervous system
of the cat. Ill. Gap junctions and membrane-associated orthogonal particle complexes (MOPC) in astrocytic membranes. Cell Tiss. Res., 149 (1974) 121-135.
14 Elfvin, L.-G. and Forsman. C.. The ultrastructure of junctions between satellite cells in mammalian sympathe- tic ganglia as revealed by freeze-etching, J. ultrastruct. Res.. 63 {1978) 261--274.
15 Ellisman. M. H., Brooke, M. H., Kaiser, K. K. and Rash, J. E., Appearance in slow muscle sarcolemma of speciali- zations characteristic of fast muscle after reinnervation by a fast muscle nerve, Exp. Neurol., 58 (1978) 59-67.
f6 Ellisman, M. H. and Rash. J. E.. Studies of excitable membranes. I11. Freeze-fracture examination of the mem- brane specializations at the neuromuscular junction and in the non-junctional sarcolemma after denervation. Brain Res., 137 (1977) 197-206. Ellisman, M. H., Rash, J. E., Staehelin, L. A. and Porter. K. R., Studies of excitable membranes. 1 I. A comparison of specializations at neuromuscular junctions and non- junctional sarcotemmas of mammalian fast and slow twitch muscle fibers, J. Cell Biol., 68 (1976) 752-774.
18 Gauthier, G. F., Lowery, S. and Hobbs, A. W. Fast and slow myosin in developing muscte fibers. Nature (Load./, 274 (1978) 25-29.
19 Guth, L. and Samaha, F. J.. Qualitative differences be- tween actomyosin ATPase of slow and fast mammalian muscle, Exp Neurol.. 25 (1969) 138-152. Guth, L. and Samaha, F. J., Erroneous interpretations which may result from application of the 'myofibrilLar ATPase' histochemical procedure to developing muscle, Exp. Neurot., 34 (1972) 465-475.
2t Hatton, J. D. and Ellisman, M. l-L, The distribution of orthogonal arrays and lheir relationship to intercellular junctions m neuroglia of the freeze-fractured hypolhala-
17
20

mo-neuropophysial system, Cell Tiss. Res., 215 (1981) 309-323.
22 Heuser, J. E., Reese, T. S. and Landis, D. M. D., Func- tional changes in frog neuromuscular junctions studied with freeze fracture, J. Neurocytol., 3 (1974) 109-131.
23 Hudson, C. S., Dyas B., K. and Rash, J. E., Changes in the number and distribution of square arrays during post- natal rat fiber type differentiation, J. Cell BioL, 79 (1978) 37a.
24 Hudson, C. S., Rash, J. E. and Albuquerque, E. X., A thin section and freeze-fracture study of mammalian neuromuscular junction development, J. Cell Biol., 75 (1977) 116a.
25 Hudson, C. S., Rash, J. E. and Graham, W. F., Sample preparation for freeze-fracture. In J. E. Rash and C. S. Hudson (Eds.), Freeze-fracture: Methods, Artifacts, Inter- pretations, Raven Press, NY, 1979, pp. 1-10.
26 Hudson, C. S., Rash, J. E. and Shinowara, N. L., Freeze- fracture and freeze-etch methods. In J. Johnson (Ed.), Current Trends in Morphological Techniques, VoL II, CRC Press, West Palm Beach, Florida, 1981.
27 Humbert, F., Pricam, C., Perrelet, A. and Orci, L., Speci- fic plasma membrane differentiations in the cells of the kidney collecting tubule, J. ultrastruct. Res., 52 (1975) 13-20.
28 Inoue, S. and Hogg, J. C., Freeze-etch study of the tra- cheal epithelium of normal guinea pigs with particular reference to intercellular junctions, J. ultrastruct. Res., 61 (1977) 89-99.
29 Karnovsky, M. J. and Roots, L., A direct coloring thio- choline method for cholinesterases, J. Histochem. Cyto- chem., 12 (1964) 219-221.
30 Korneliussen, H. and Jansen, J. K. S., Morphological aspects of the elimination of polyneuronal innervation of
101
skeletal muscle fibers in newborn rats, J. NeurocytoL, 5 (1976) 591-604.
31 Kreutziger, G. O., Freeze-etching of intercellular junc- tions in mouse liver. In Proceedings of the 26th Meeting of the Electron Microscope Society of America, Claitors Publishing Division, Baton Rouge, 1968, pp. 234-235.
32 Kugelberg, E., Adaptive transformation of rat soleus motor units during growth. Histochemistry and contrac- tion speed, J. Neurol. Sci., 27 (1976) 269-289.
33 Landis, D. M. D. and Reese, T. S., Arrays of particles in freeze-fractured astrocytic membranes, J. Cell BioL, 60 (1974) 316-320.
34 Landis, D. M. D. and Reese, T. S., Astrocyte membrane structure: changes after circulatory arrest, J. Cell BioL, 88 (1981) 660-663.
35 Pannese, E., Luciano, L., Iurato, S. and Reale, E., Inter- cellular junctions and other membrane specializations in developing spinal ganglia: a freeze-fracture study, 3". ultrastruct. Res., 60 (1977) 169-180.
36 Rash, J. E. and Ellisman, M. H., Studies of excitable membranes. I. Macromolecular specializations of the neuromuscular junction and the nonjunctional sarco- lemma, J. Cell Biol., 63 0974) 567-586.
37 Redfern, P. A., Neuromuscular transmission in newborn rats, J. Physiol. (Lond.), 209 (1970) 701-709.
38 Riley, D. A., Multiple innervation of fiber types in the soleus muscles of postnatal rats, Exp. Neurok, 56 (1977) 400-409.
39 Riley, D. A., Spontaneous elimination of nerve terminals from the end-plates at skeletal myofibers, Brain Res., 134 (1977) 279-285.
40 Schmalbruch, H., 'Square arrays' in the sarcolemma of human skeletal muscle fibers, Nature (Lond.), 281 (1979) 145-146.
Copyright © 2022 FDOKUMEN




















